Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CH&.MISTRY Vol. 254, No. 3, Issue of February 10, pp. 9X1-986, 1979 Printed m U S A.
The Chemistry and Biology of Thymosin I. ISOLATION, CHARACTERIZATION, AND BIOLOGICAL ACTIVITIES OF THYMOSIN (~1 AND POLYPEPTIDE
/?I FROM CALF THYMUS*
(Received for publication, June 2, 1978, and in revised form, September 5, 1978)
Teresa L. K. Low,+ Gary B. Thurman,+ Martha McAdoo,§ John McClure,+ Jeffrey L. Rossio,l Paul H. Naylor, 11 and Allan L. Goldstein+
From the Division of Biochemistry, Department of Human Biological Chemistry and Genetics, University of Texas Medical Brunch, Galveston, Texas 77550
A partially purified extract from thymus tissue termed thymosin Fraction 5 has been shown to recon- stitute immunological deficiencies resulting from the lack of thymic function in several animal models, as well as in humans with primary and secondary immu- nodeficiency diseases. Thymosin Fraction 5 consists of a family of polypeptides with molecular weights rang- ing from 1,000 to 15,000. Several of these polypeptides contribute individually to the biological activity of the parent compound. Two polypeptide components of thy- mosin Fraction 5, termed thymosin (~1 and polypeptide /II, have been characterized chemically and biologi- cally. Thymosin LYI is a highly acidic molecule composed of 28 amino acid residues. This polypeptide has potent biological activity and has been found to be 10 to 1000 times as active as thymosin Fraction 5 in one in vivo and several in vitro bioassay systems designed to mea- sure differentiation and function of thymus-dependent lymphocytes (T cells). Polypeptide PI, in contrast, is inactive in our bioassay systems, suggesting that it is not involved in thymic hormone action. Sequence anal- ysis and homology studies have indicated that poly- peptide /!!I, although present in Fraction 5, does not contribute to the biological activity of thymosin Frac- tion 5.
Investigations describing attempts to determine the nature of thymic factors date back over 150 years. However, until the early 1960’s the central role of the thymus gland in the ontogenesis and function of the immune system was not recognized (cf Ref. 1). In 1961, Miller (2) and Archer and Pierce (3) reported that neonatal thymectomy in the mouse and in the rabbit results in failure of normal postnatal growth and lymphoid tissue development and maturation. These studies demonstrated the essential nature of the thymus gland in the development of the lymphoid system. Since then,
* This research is supported in part by grants from the National Cancer Institute (CA16964 and CA20667), Hoffmann-La Roche, Inc., and the John A. Hartford Foundation, Inc. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 USC. Section 1734 solely to indicate this fact.
+ Present address, Department of Biochemistry, The George Wash- ington University Medical Center, Washington, D. C. 20037.
9 Present address, Marine Biomedical Institute, University of Texas Medical Branch, Galveston, Tex. 77550.
1 Present address, Department of Microbiology and Immunology, School of Medicine, Wright State University, Dayton, Ohio 45431.
11 Present address, Department of Experimental Therapeutics, Grace Cancer Drug Center, Roswell Park Memorial Institute, Buffalo, N. Y. 14263.
investigations involving thymic ablation and replacement by thymic grafts or extracts in several species have demonstrated a principal role for the thymus gland in the maturation, proliferation, and immunological competence of the lympho- cyte (4-7).
Although it is still not clear how the thymus gland exerts control over T cell’ development, it appears that a vital part of the process occurs via a hormonal mechanism (8). For the past several years much research has been directed toward the isolation and identification of the biologically active thymic factors or hormones which can act in lieu of an intact thymus in thymectomized animals or enhance immunological competence in normal animals (9-15). One of these partially purified thymic preparations termed thymosin Fraction 5 (12-15) has been shown to be a potent immunopotentiating agent. This thymosin fraction corrects some of the deficiencies resulting from lack of thymic function in a number of animal models (16-22), as well as in humans with primary immuno- deficiency diseases (23-26) and in immunosuppressed cancer patients (25, 27, 28).
Thymosin Fraction 5 is heat-stable up to 80°C. It is com- posed of a group of polypeptides with molecular weights ranging from 1,000 to 15,000. The preparation has a very low content of non-protein material. There are detectable and identifiable amounts of carbohydrate.
A major effort in our continuing studies of thymosin is to understand the molecular events by which the thymus gland exerts control over T cell development. In order to understand the detailed mechanism(s) of how each thymosin polypeptide exerts control over the development and senescence of im- munological responses, we have undertaken the isolation and characterization of each polypeptide component in thymosin Fraction 5 and a detailed study of their individual biological functions.
Using a combination of ion exchange chromatography and gel filtration, we have isolated several polypeptide compo- nents from Fraction 5 for further characterization. In this report, we present the isolation, purification, and biological properties of two of the polypeptide components designated thymosin ai, and polypeptide PI.” In the accompanying paper (29) we present the detailed sequence analysis of these two polypeptides.
Thymosin a, has been demonstrated to be a potent immu- nologically active thymic polypeptide. It is 10 to 1,000 times
’ The abbreviations used are: T cell, thymus-dependent lympho- cyte; MIF, macrophage inhibitory factor; RIA, radioimmunoassay; LPS, lipopolysaccharide; MLR, mixed lymphocyte reaction; THF, thymic humoral factor; FTS, facteur thymique serique.
‘See “Methods” in the miniprint section for a description of the nomenclature used to identify these polypeptides.
981
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

982 Isolation and Biological Studies of Thymosin a1 and Polypeptide ,&
as active as the parent thymosin Fraction 5 when assayed in several biological models, including an in uivo mouse mitogen assay (30), an in vitro lymphokine assay measuring the pro- duction of macrophage inhibitor factor (MIF) (31), an in vitro induction of Lyt surface markers on putative T cell precursors (32), and an in vitro human E-rosette assay measuring the production of T cells (33). Thymosin (Ye is composed of 28 amino acid residues and has an isoelectric point of 4.2. A preliminary report on the primary structure of thymosin (~1 has been published (34).
Polypeptide /3x is composed of 74 amino acid residues with a molecular weight of 8451 and isoelectric point of 6.7. This polypeptide is the most predominant and intense band on polyacrylamide gel after isoelectric focusing of thymosin Frac- tion 5. Nevertheless, it does not exhibit any biological activity as tested in our bioassay systems. These observations suggest that /31, although present as a major component in thymosin Fraction 5, is not involved in thymic hormone action. The complete sequence of this polypeptide has revealed that this molecule is identical with ubiquitin (35) and a portion of a nuclear chromosomal protein A24 (36). The possible relation- ship among these proteins will be discussed in the accompa- nying paper (29).
EXPERIMENTAL PROCEDURES
The experimental procedures are described in the miniprint sec- tion.”
RESULTS
Preparation of Thymosin Fraction 5 and Fraction 5A
The yield of thymosin Fraction 5 from acetone powder (Fraction 3) is approximately 15% (based on Lowry assay (37)) and the yield of Fraction 5A about 13%. On an isoelectric focusing gel with pH range 3.5 to 9.5, Fraction 5A appears to contain fewer polypeptides which migrate in the (Y (very acidic) region in comparison to Fraction 5. Polypeptide ,& was found to be present in both Fractions 5 and 5A. However, the relative concentration of /31 in Fraction 5A is higher than in Fraction 5.
Isoelectric Focusing in Polyacrylamide Gels
We have found that staining procedures have a marked effect on the gel pattern of thymosin polypeptides. Some polypeptides which are stained weakly or not at all with one staining procedure (e.g. the procedure which includes sulfo- salicylic acid in the fixing solution) may be stained strongly with another procedure (e.g. procedure which includes 20% trichloroacetic acid in fiiing and staining). Therefore, it is important to use at least two alternative staining methods for each polypeptide preparation. Both thymosin (~1 and poly- peptide /?I stained well with procedures which included iso- propyl alcohol in the fixing solution.
Isolation of Thymosin a1 and Polypeptide p1
Thymosin Q, isolated from Fraction 5 according to the procedures outlined in Fig. 3S, migrates as a single band on analytical polyacrylamide gels at pH 8.3 or pH 2.9 according to the procedures described previously (15). It is shown as a major band with an isoelectric point of 4.2 on an~isoelectric
3 Experimental Procedures including Materials, Methods, and Figs. 1s through 48 and References are presented as a miniprint supple- ment immediately following this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Bethesda, Md. 20014. Request Document No. 78M-908, cite author(s), and include a check or’ money order for $2.10 per set of photocopies.
focusing slab gel of pH range 3 to 5 (Fig. 1). The yield of thymosin o1 from Fraction 5 is about 0.6%. Polypeptide ,& migrates as a single band on an isoelectric focusing slab gel of pH range 3.5 to 9.5 (LKB PAG,& with an isoelectric point of 6.7. When larger amounts (200 pg) of sample were loaded on the gel, a few minor bands with p1 around 6 were observed. The yield of polypeptide ,& from thymosin Fraction 5A is approximately 6.2%. This polypeptide was also isolated from Fraction 5 in a less pure form. The yield from Fraction 5 is estimated to be 1.5%.
The molecular weights of thymosin (~1 and polypeptide p1 were estimated to be 3350 and 9000, respectively, using a calibrated Sephadex G-75 column in 6 M guanidine hydro- chloride as described under “Methods” in the miniprint. These values are reasonably close to the results obtained from the established amino acid sequences as reported in the ac- companying paper (29), 3108 for thymosin (~1 and 8451 for polypeptide /3*.
The purification steps and total yields of each of the thy- mosin fractions and the purified thymosin polypeptides are summarized in Table I.
Biological Properties of Thymosin a1 and Polypeptide fi,
Summary of Bioassay Data-As summarized in Table II, seven different bioassay systems (mixed lymphocyte reactivity (MLR), production of macrophage inhibitory factor (MIF), E- rosette formation, antibody production, mitogen responsivity, Lyt surface marker induction, and induction of terminal deox- ynucleotidyltransferase) for measuring the biological activity of thymosin Fraction 5 and the purified thymosin polypeptides have been utilized. In these assays, the standard thymosin Fraction 5 is active in the range of 1 to 100 pg/ml. Under the same bioassay conditions, thymosin (Y~ is from 10 to 1000 times (or higher) as active as Fraction 5 in some assays (MIF, E- rosette, mitoaen. and Lvt surface marker induction). but not in others (MLR, antibody production,
CATHODE (-) *
ANODE (i-1 N
I I
and terminal deoxy-
8.80
5.70
5.20
4.80
-I 4.60 &!&
4.40
4.20
4.05
3.85
3.60
Fr. 5 Thymosin txl
FIG. 1. Isoelectric focusing in polyacrylamide gel (narrow range pH 3 to 5). Thymosin (Y, is shown as a major band with a p1 of 4.2.
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from
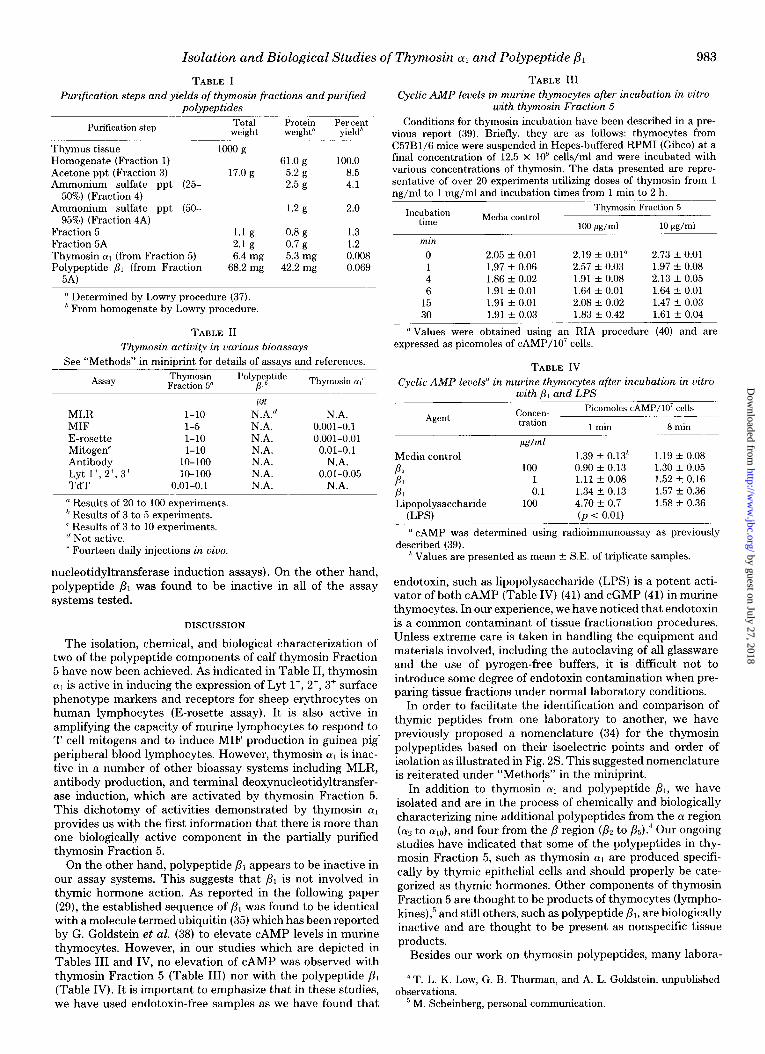
Isolation and Biological Studies of Thymosin a1 and Polypeptide ,L& 983
TABLE I Purification steps and yields of thymosin fractions and purified
polypeptides
Purification step Total weieht
Protein Per cent weieht” vield’
Thymus tissue Homogenate (Fraction 1) Acetone ppt (Fraction 3) Ammonium sulfate ppt (25-
50%) (Fraction 4) Ammonium sulfate ppt (50-
95%) (Fraction 4A) Fraction 5 Fraction 5A Thymosin (Y, (from Fraction 5) Polypeptide PI (from Fraction
5-4)
looog 61.0 g 100.0
17.0 g 5.2 g 8.5 2.5 g 4.1
1.2 g 2.0
1.1 g 0.8 g 1.3 2.1 g 0.7 g 1.2 6.4 mg 5.3 mg 0.008
68.2 mg 42.2 mg 0.069
” Determined by Lowry procedure (37). * From homogenate by Lowry procedure.
TABLE II Thymosin activity in various bioassays
See “Methods” in miniprint for details of assays and references. Thymosin Fraction 5” Thymosin o[,’
MLR l-10 MIF l-5 E-rosette l-10 Mitogen’ l-10 Antibody IO-100 Lyt 1+, 2+, 3+ 10-100 TdT 0.01-0.1
n Results of 20 to 106 experiments. ’ Results of 3 to 5 experiments. ’ Results of 3 to 10 experiments. ” Not active. ’ Fourteen daily injections in uiuo.
F%f? N.A.” N.A. N.A. 0.001-0.1 N.A. 0.001-0.01 N.A. 0.01-0.1 N.A. N.A. N.A. 0.01-0.05 N.A. N.A.
nucleotidyltransferase induction assays). On the other hand, polypeptide p1 was found to be inactive in all of the assay systems tested.
DISCUSSION
The isolation, chemical, and biological characterization of two of the polypeptide components of calf thymosin Fraction 5 have now been achieved. As indicated in Table II, thymosin (Y, is active in inducing the expression of Lyt l’, 2’, 3’ surface phenotype markers and receptors for sheep erythrocytes on human lymphocytes (E-rosette assay). It is also active in amplifying the capacity of murine lymphocytes to respond to T cell mitogens and to induce MIF production in guinea pig’ peripheral blood lymphocytes. However, thymosin (Ye is inac- tive in a number of other bioassay systems including MLR, antibody production, and terminal deoxynucleotidyltransfer- ase induction, which are activated by thymosin Fraction 5. This dichotomy of activities demonstrated by thymosin (~1 provides us with the first information that there is more than one biologically active component in the partially purified thymosin Fraction 5.
On the other hand, polypeptide p1 appears to be inactive in our assay systems. This suggests that PI is not involved in thymic hormone action. As reported in the following paper (29), the established sequence of ,& was found to be identical with a molecule termed ubiquitin (35) which has been reported by G. Goldstein et al. (38) to elevate CAMP levels in murine thymocytes. However, in our studies which are depicted in Tables III and IV, no elevation of CAMP was observed with thymosin Fraction 5 (Table III) nor with the polypeptide PI (Table IV). It is important to emphasize that in these studies, we have used endotoxin-free samples as we have found that
TABLE III Cyclic AMP levels in murine thymocytes after incubation in vitro
with thymosin Fraction 5 Conditions for thymosin incubation have been described in a pre-
vious report (39). Briefly, they are as follows: thymocytes from C57B1/6 mice were suspended in Hepes-buffered RPM1 (Gibco) at a final concentration of 12.5 x 10” cells/ml and were incubated with various concentrations of thymosin. The data presented are repre- sentative of over 20 experiments utilizing doses of thymosin from 1 ns/ml to 1 mz/ml and incubation times from 1 min to 2 h.
Incubation time Media control
Thymosin Fraction 5
100 M/d 10 Wml min
0 2.05 -c 0.01 2.19 f 0.01” 2.73 t- 0.01 1 1.97 f 0.06 2.57 -I- 0.03 1.97 -c 0.08 4 1.86 + 0.02 1.91 f 0.08 2.13 + 0.05 6 1.91 + 0.01 1.64 f 0.01 1.64 f 0.01
15 1.91 * 0.01 2.08 -I- 0.02 1.47 f 0.03 30 1.91 f 0.03 1.83 + 0.42 1.61 + 0.04
“Values were obtained using an RIA procedure (40) and are expressed as picomoles of cAMP/lO’ cells.
TABLE IV Cyclic AMP levels” in murine thymocytes after incubation in vitro
with /I, hnd LPS Picomoles cAMP/lO’ cells
Agent COMXn- tration 1 min 8 min
Media control PI PI PI Lipopolysaccharide
(LPS)
!&ml
100 1 0.1
106
1.39 f 0.13h 0.90 -c 0.13 1.11 f 0.08 1.34 u 0.13 4.70 + 0.7 (p < 0.01)
1.19 +- 0.08 1.30 f 0.05 1.52 + 0.16 1.57 -c 0.36 1.58 f 0.36
’ CAMP was determined using radioimmunoassay as previously described (39).
’ Values are presented as mean +- S.E. of triplicate samples.
endotoxin, such as lipopolysaccharide (LPS) is a potent acti- vator of both CAMP (Table IV) (41) and cGMP (41) in murine thymocytes. In our experience, we have noticed that endotoxin is a common contaminant of tissue fractionation procedures. Unless extreme care is taken in handling the equipment and materials involved, including the autoclaving of all glassware and the use of pyrogen-free buffers, it is difficult not to introduce some degree of endotoxin contamination when pre- paring tissue fractions under normal laboratory conditions.
In order to facilitate the identification and comparison of thymic peptides from one laboratory to another, we have previously proposed a nomenclature (34) for the thymosin polypeptides based on their isoelectric points and order of isolation as illustrated in Fig. 2s. This suggested nomenclature is reiterated under “Methods” in the miniprint.
In addition to thymosin (Y] and polypeptide ,&, we have isolated and are in the process of chemically and biologically characterizing nine additional polypeptides from the (Y region ((Ye to aIo), and four from the p region (Pz to p~).~ Our ongoing studies have indicated that some of the polypeptides in thy- mosin Fraction 5, such as thymosin LY, are produced specifi- cally by thymic epithelial cells and should properly be cate- gorized as thymic hormones. Other components of thymosin Fraction 5 are thought to be products of thymocytes (lympho- kines),” and still others, such as polypeptide p,, are biologically inactive and are thought to be present as nonspecific tissue products.
Besides our work on thymosin polypeptides, many labora-
4 T. L. K. Low, G. B. Thurman, and A. L. Goldstein, unpublished observations.
’ M. Scheinberg, personal communication.
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

984 Isolation and Biological Studies of Thymosin (~1 and Polypeptide ,&I
tories have documented the isolation of thymic extracts with various biological properties (cfi Ref. 42). The best character- ized of these thymic factors are thymopoietin (ll), thymic humoral factor (THF) (9), and facteur thymique serique (FTS) (43,44). The amino acid sequence of thymosin a1 (29) was found to bear no homology to either thymopoietin or FTS. Although the sequence of THF has not been reported, from its amino acid composition (9), it appears that THF differs from thymosin aI. However, thymosin (~1, THF and FTS have several functional properties in common, including the capacity to reconstitute immunological functions in vitro and in viva in animal models. Thymopoietin has been found to induce the expression of surface markers characteristics of T cells (45) and to induce symptoms of myasthenia gravis in rodents (II).
One of the most important contributions of the purified thymic peptides such as thymosin (Y, to the field of immunol- ogy will be their utilization in helping to decipher the early steps in the differentiation of T cell maturation. Of particular importance are the demonstrations (32) that thymosin Frac- tion 5 induces the maturation of three populations of T cells as characterized by the Lyt phenotypes, namely Lyt If, Lyt 2+, 3+ and Lyt lc, 2’, 3+, whereas thymosin 01, induces specif- ically the expression of Lyt I’, 2+, 3+ surface markers. This would suggest that thymosin LY, affects the very early stages of T cell development. Of additional interest are the recent observations (46) that two other purified thymosin polypep- tides termed thymosin ,& and fi4 specifically induce the expres- sion of terminal deoxynucleotidyltransferase in albumin gra- dient-separated bone marrow stem cells. Furthermore, the studies of S. Horowitz” and Ahmed et al. (32) show that another polypeptide isolated from the (Y region termed thy- mosin IY~ induces the expression of suppressor T cells. These
” S. Horowitz, personal communication.
studies support the hypothesis that there exists a family of thymosin polypeptides that act on different subsets of T cells which are required to induce the maturation of these various classes of T cells (47).
Clinically, several of the purified thymosin polypeptides should have a major role in restoring immune function and augmenting specific lymphocyte functions in children with hypothymic function and in adults with a variety of T cell disorders including cancer and autoimmune disease. Ongoing Phase I and Phase II clinical trials with thymosin in patients with primary immunodeficiency diseases (23-26), and in im- munosuppressed cancer patients (27,28) support this conten- tion. Most recently, the first Phase II randomized trial of thymosin has been completed in patients with nonresectable small cell carcinoma of the lungs in conjunction with intensive chemotherapy7 (48). In this study, it has been found that the mean survival increased from 243 days in the group receiving chemotherapy alone to over 450 days in the group receiving thymosin Fraction 5 twice a week for the first 6 weeks of the chemotherapy induction period. The completion of other Phase II clinical trials with thymosin Fraction 5 presently underway should provide a clearer understanding of the ca- pacity of thymosin Fraction 5 to potentiate and reconstitute immunity and should provide the foundation for initiation of studies designed to determine the clinical role of the purified thymosin polypeptides in dealing with diseases associated with thymic malfunction and immune imbalance.
Acknowledgments-We wish to acknowledge Janet Anderson, Bar- bara Simms, Janelle Hatcher, Milly Chin, Paul Geibel, and James Oliver for their technical assistance.
REFE:RENCES
The references are found on pp. 985 and 9%.
’ P. Chretien, personal communication.
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

Isolation and Biological Studies of Thymosin a1 and Polypeptide PI
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

986 Isolation and Biological Studies of Thymosin a1 and Polypeptide /.%
Ahwx ,+, - -mlyrmm d, - 1
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from

GoldsteinT L Low, G B Thurman, M McAdoo, J McClure, J L Rossio, P H Naylor and A L
thymus.biological activities of thymosin alpha1 and polypeptide beta1 from calf The chemistry and biology of thymosin. I. Isolation, characterization, and
1979, 254:981-986.J. Biol. Chem.
http://www.jbc.org/content/254/3/981Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/254/3/981.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on July 27, 2018http://w
ww
.jbc.org/D
ownloaded from





























