Embed Size (px)
Citation preview

Cancer Therapy: Preclinical
The MCT4 Gene: A Novel, Potential Target forTherapy of Advanced Prostate CancerStephen Yiu Chuen Choi1,2, Hui Xue1,2, Rebecca Wu2, Ladan Fazli1, Dong Lin1,2,Colin C. Collins1, Martin E. Gleave1, Peter W. Gout2, and Yuzhuo Wang1,2
Abstract
Purpose: The management of castration-resistant prostatecancer (CRPC) is a major challenge in the clinic. Androgenreceptor signaling–directed strategies are not curative in CRPCtherapy, and new strategies targeting alternative, key cancerproperties are needed. Using reprogrammed glucose metabo-lism (aerobic glycolysis), cancer cells typically secrete excessiveamounts of lactic acid into their microenvironment, promotingcancer development, survival, and progression. Cellular lacticacid secretion is thought to be predominantly mediated byMCT4, a plasma membrane transporter protein. As such, theMCT4 gene provides a unique, potential therapeutic target forcancer.
Experimental Design: A tissue microarray of various Gleasongrade human prostate cancers was stained for MCT4 protein.Specific,MCT4-targeting antisense oligonucleotides (MCT4 ASO)were designed and candidate MCT4 ASOs checked for effects on
(i) MCT4 expression, lactic acid secretion/content, glucose con-sumption, glycolytic gene expression, and proliferation of humanCRPC cells and (ii) growth of PC-3 tumors in nude mice.
Results: ElevatedMCT4 expressionwas associatedwith humanCRPC and an earlier time to relapse. The treatment of PC-3,DU145, and C4-2 CRPC cultures with candidate MCT4 ASOs ledtomarked inhibition of MCT4 expression, lactic acid secretion, toincreased intracellular lactic acid levels, and markedly reducedaerobic glycolysis and cell proliferation. Treatment of PC-3tumor-bearing nude mice with the MCT4 ASOs markedly inhib-ited tumor growth without inducing major host toxicity.
Conclusions: MCT4-targeting ASOs that inhibit lactic acidsecretion may be useful for therapy of CRPC and othercancers, as they can interfere with reprogrammed energymetabolism of cancers, an emerging hallmark of cancer.Clin Cancer Res; 22(11); 2721–33. �2016 AACR.
IntroductionProstate cancer is the most commonly diagnosed noncuta-
neous cancer and a leading cause of cancer-related death forNorth American men (1). When the malignancy is confined tothe prostate, surgery and radiotherapy can be curative. However,many treated patients experience local cancer recurrence andmetastasis (2). Although androgen deprivation therapy (ADT),currently the treatment of choice for metastatic prostate cancer,can lead to remissions, tumors frequently return in a form that ishighly resistant to ADT and other therapies, that is, metastaticcastration–resistant prostate cancer (mCRPC). Although the effi-cacy of mCRPC treatment has recently been improved by usingmore powerful chemotherapeutics targeting the androgen recep-tor (AR) signaling axis, such as enzalutamide (3) and abiraterone(4), the overall survival of patients has only marginally increased(5–7). Moreover, it is thought that further improved versions of
such drugs can promote transdifferentiation of prostatic adeno-carcinoma to neuroendocrine prostate cancer, a subtype of thedisease that is currently incurable (8–10). There is, therefore, acritical need for new,more effective strategies for therapy of CRPCthat target different key properties of cancer.
There is increasing evidence that targeting reprogrammed ener-gy metabolism of cancers offers a unique approach for effectivetherapeutic intervention (11). For glucose utilization, cancer cells,as distinct from normal resting cells, in general have a preferencefor glycolysis coupled to lactic acid production, i.e., a processcalled aerobic glycolysis (theWarburg effect; ref. 12). This leads toelevated glucose consumption, a near-universal property of pri-mary and metastatic cancers. In addition, aberrant utilization ofglutamine, also leading to elevated lactic acid production, hasbeen observed to be highly common for cancers (13). Thesemetabolic energy pathways lead to increased lactic acid secretionby the cancer cells into their microenvironment, facilitatingmultiple oncogenic, lactate-stimulated processes, including tissueinvasion/metastasis, neoangiogenesis and responses to hypoxia(14–17); furthermore, lactic acid–induced acidification of thecancer cell microenvironment (to pH 6.0–6.5) can lead to thesuppression of local host anticancer immunity (14, 18). Thephenomenon of enhanced glucosemetabolismby cancers ismostcommonly exploited clinically by 18F-fluorodeoxyglucose PET(FDG-PET). Although this imaging technique is not generallyused for prostate cancer (19), there is evidence suggesting thatglucose metabolism of prostate cancer cells is increased by ARsignaling and progression to treatment resistance (20, 21). Assuch, targeting the aerobic glycolytic pathway could be effectivefor treating advanced prostate cancers.
1The Vancouver Prostate Centre, Vancouver General Hospital andDepartment of Urologic Sciences, The University of British Columbia,Vancouver, British Columbia, Canada. 2Department of ExperimentalTherapeutics, BC Cancer Research Centre,Vancouver, British Colum-bia, Canada.
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
Corresponding Author: Yuzhuo Wang, BC Cancer Agency, 675 W. 10th Ave.,Vancouver, BC V5Z 1L3, Canada. Phone: 604-675-8013; Fax: 160-4675-8019;E-mail: [email protected]
doi: 10.1158/1078-0432.CCR-15-1624
�2016 American Association for Cancer Research.
ClinicalCancerResearch
www.aacrjournals.org 2721
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624

The monocarboxylate transporter (MCT) family consists ofplasma membrane transporter proteins involved in the trans-port of lactic acid and other metabolic monocarboxylates. Inparticular, the cellular efflux of lactic acid/Hþ is thought to bepredominantly mediated by MCT4 (SLC16A3; ref. 22). Theexpression of MCT4 has been associated with highly glycolyticcells (17,23,24), and elevated expression of MCT4 in tumors isclinically relevant, as it has been associated with poor patientprognosis in multiple types of cancer (25–27), includingprostate cancer (28,29). Furthermore, elevated MCT4 expres-sion may be important in cancer–stroma interactions facili-tating prostate cancer progression (30). This information,together with the cancer growth–promoting ability of can-cer-generated lactic acid, suggests that the inhibition of theexpression or function of MCTs provides a promising thera-peutic strategy for a wide variety of neoplasms (31). Currently,small-molecule inhibitors of MCTs are available that specif-ically target MCT1 and MCT2 (e.g., AZD3965, AR-C155858;refs. 32,33), nonspecifically inhibit MCTs, including MCT1and MCT4 (e.g., a-cyano-4-hydroxycinnamate; refs. 34,35), orselectively inhibit lactic acid import via MCT4 (e.g., 7-amino-carboxycoumarins; ref. 36). However, a therapeutic strategyspecifically targeting MCT4-mediated efflux of lactic acid isstill lacking.
In this study, we established that the development of humanCRPC is associated with elevated expression of MCT4. Further-more, evidence was obtained that MCT4-targeting antisense oli-gonucleotides (ASO) inhibiting MCT4-mediated lactic acid secre-tion may be useful for the treatment of CRPC.
Materials and MethodsMaterials
Chemicals, solvents, and solutions were obtained from Sigma-Aldrich, unless otherwise indicated.
Cell culturesHuman PC-3 and DU145 CRPC cells, human LNCaP prostate
cancer cells, and mouse TRAMPC2 prostate cancer cells werepurchased from the ATCC; C4-2 CRPC cells were obtained fromDr. Martin E. Gleave (Vancouver Prostate Centre, Vancouver,British Columbia, Canada). Human monolayer cultures weremaintained in RPMI1640 (GEHealthcare HyClone), supplemen-ted with 10% FBS (GE Healthcare HyClone), whereas TRAMPC2cells were maintained in DMEM (GE Healthcare HyClone),supplemented with 5% FBS. For cell counting, cells were trypsi-nized to form a single-cell suspension and counted using a TC20Automated Cell Counter (Bio-Rad). Cell viability was assessed byTrypan blue exclusion.
Human prostate cancer TMA construction and IHCTissue microarrays (TMA) were manually constructed, as pre-
viously described (37,38), using various Gleason grades–exhibit-ing prostate cancer specimens (n ¼ 342), obtained from theVancouver Prostate Centre Tissue Bank with written informedconsent of patients, clinical information, and institutional studyapproval. All specimens were obtained through radical prostatec-tomy, except CRPC samples that were obtained via transurethralresection of the prostate. Immunohistochemical staining wasconducted using a Ventana autostainer (model Discover XT;Ventana Medical Systems) with an enzyme-labeled biotin–strep-tavidin system and a solvent-resistant DAB Map Kit (VentanaMedical Systems). Staining intensity was scored by a trainedpathologist on a 4-point scale: 0 represents no staining on anytumor cells, 1 represents a faint or focal, questionably presentstain, 2 represents a stain of convincing intensity in a minority ofcells, and 3 represents a stain of convincing intensity in a majorityof cells.
AntibodiesThe following antibodies and conjugateswere used: rabbit anti-
MCT4 antibody (Santa Cruz Biotechnology; WB 1:4,000, IHC1:100), mouse anti-vinculin antibody (Sigma; WB 1:1,000), rab-bit anti-cleaved caspase-3 antibody (Cell Signaling Technology;IHC 1:50), mouse anti-Ki67 antibody (Dako; IHC 1:50), rat anti-CD31 antibody (Dianova; IHC 1:20), mouse anti-pan-T cellmarker CD3 antibody (Dako; IHC 1:50), biotinylated mouseanti-NK1.1 (Cedarlane; IHC 1:100), IRDye 800CW goat anti-mouse antibody (LI-COR Biosciences; WB 1:10,000), IRDye680RD goat anti-rabbit antibody (LI-COR Biosciences; WB1:10,000), biotinylated goat anti-rabbit antibody (Vector Labo-ratories; IHC 1:200), biotinylated goat anti-rat antibody (VectorLaboratories; IHC 1:200), and biotinylated goat anti-mouse anti-body (Vector Laboratories; IHC 1:200).
ASO design and selectionFirst-generation phosphorothioate-modified ASOs against
human MCT4 were rationally designed by selecting sequencescontaining favorable motifs while excluding unfavorable ones(39). Specificity of MCT4-targeting sequences, compared withhuman and mouse genes (at least 3/20 bases mismatched),was evaluated using BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Ten sequences (see Supplementary Table S1) dis-tributed throughout the length of the transcript with perfectcomplementarity to all human MCT4 transcript variants(NM_001042422.2, NM_001042423.2, NM_001206950.1,NM_001206951.1, NM_001206952.1, and NM_004207.3)were selected and synthesized by Eurofins MWG Operon. The
Translational Relevance
The management of castration-resistant prostate cancer(CRPC) is a major challenge in the clinic. Although CRPCtreatment efficacy has been improved by using more powerfuldrugs targeting the androgen receptor signaling axis, patientsurvival has only marginally increased, and further improvedversions of such drugs can lead to transdifferentiation of CRPCto currently incurable neuroendocrine prostate cancer. There-fore, new strategies targeting alternative, fundamental cancerproperties are critically needed. Increasing evidence indicatesthat reprogrammed energy metabolism of cancers offers aunique target. For glucose utilization, cancer cells generallyprefer glycolysis coupled to lactic acid production, that is,aerobic glycolysis. This pathway leads to increased lactic acidsecretion by cancers facilitating oncogenic processes, includingtissue invasion/metastasis and neoangiogenesis. The lacticacid secretion is predominantly mediated by MCT4, a plasmamembrane transporter protein. Here, we have shown, both invitro and in vivo, that MCT4-targeting antisense oligonucleo-tides (ASO) can inhibit MCT4 expression, lactic acid secretion,aerobic glycolysis, and cell proliferation of CRPC models. Assuch, MCT4-targeting ASOs may provide a novel therapeuticapproach for CRPC.
Choi et al.
Clin Cancer Res; 22(11) June 1, 2016 Clinical Cancer Research2722
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624
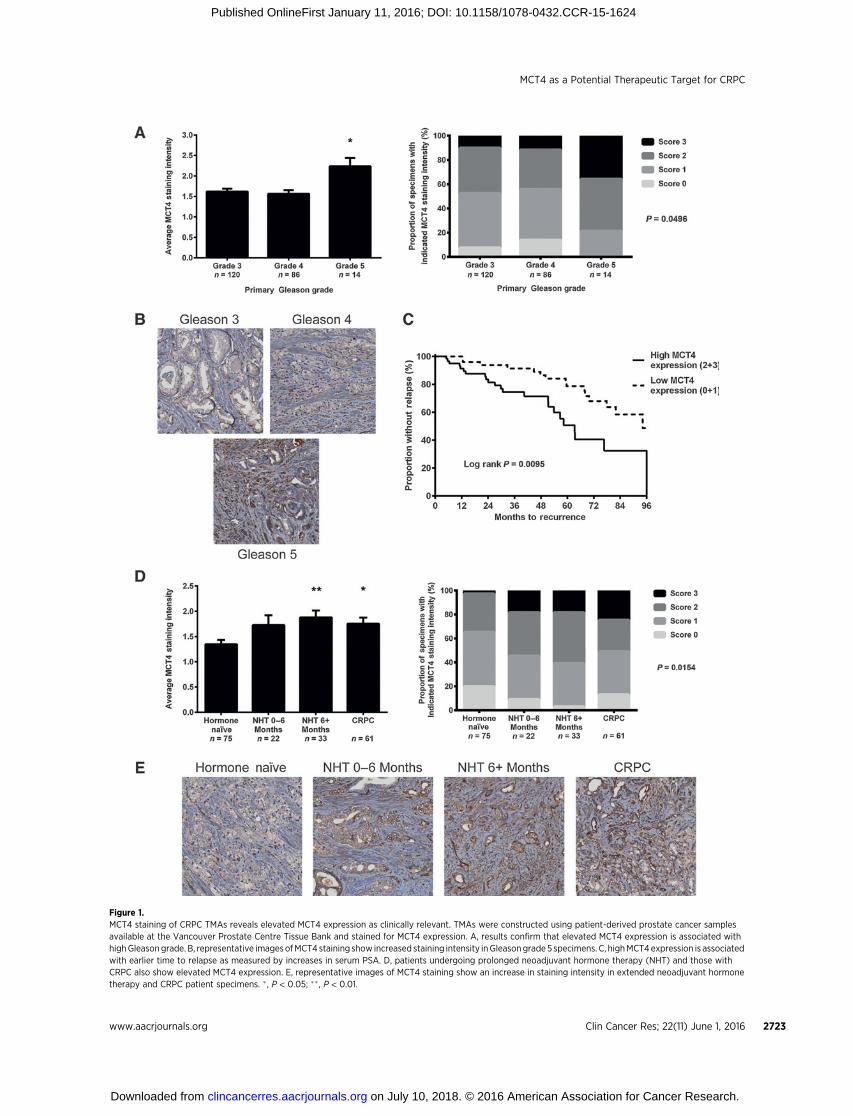
Figure 1.MCT4 staining of CRPC TMAs reveals elevated MCT4 expression as clinically relevant. TMAs were constructed using patient-derived prostate cancer samplesavailable at the Vancouver Prostate Centre Tissue Bank and stained for MCT4 expression. A, results confirm that elevated MCT4 expression is associated withhighGleasongrade. B, representative imagesofMCT4 staining show increased staining intensity inGleasongrade5 specimens. C, highMCT4expression is associatedwith earlier time to relapse as measured by increases in serum PSA. D, patients undergoing prolonged neoadjuvant hormone therapy (NHT) and those withCRPC also show elevated MCT4 expression. E, representative images of MCT4 staining show an increase in staining intensity in extended neoadjuvant hormonetherapy and CRPC patient specimens. �, P < 0.05; �� , P < 0.01.
MCT4 as a Potential Therapeutic Target for CRPC
www.aacrjournals.org Clin Cancer Res; 22(11) June 1, 2016 2723
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624

knockdown efficiencies of these ten ASOs were tested bydetermining target mRNA and protein expression 48 hoursafter transfection of cells using qPCR and Western blotting.Two candidate ASOs (#1 and #14) were selected for furtherstudies. Sequences: ASO #1, 50-TCCCATGGCCAGGAGGGTTG-30; ASO #14, 50-AGATGCAGAAGACCACGAGG-30; a publishednontargeting control ASO, 50-CCTTCCCTGAAGGTTCCTCC-30
(40,41).
ASO and siRNA transfectionCells were transfected in 6-well plates with ASOs at 100 nmol/L
for 48 hours (unless otherwise indicated) using Oligofectamine(Invitrogen) or withMCT4-targeting siRNAs and controls (Dhar-macon) at 50 nmol/L for 48 hours using Lipofectamine 2000(Invitrogen), following the manufacturer's instructions.
Quantitative PCRTotal RNA was isolated using the RNeasy Mini Kit (Qiagen
Inc.) and cDNA synthesized using the QuantiTect ReverseTranscription Kit (Qiagen). Primers (see Supplementary TableS1) were designed using Primer-BLAST (http://www.ncbi.nlm.nih.gov/tools/primer-blast/). qRT-PCR reactions using KAPASYBR FAST Universal (Kapa Biosystems) were performed intriplicate in a ViiA 7 Real-Time PCR System (Applied Biosys-tems). Target genes were normalized to a geometric average of 3internal reference genes (42).
Western blottingCells were harvested and lysed in RIPA buffer (50 mmol/L Tris
Cl pH 7.4, 150 mmol/L NaCl, 1% IGEPAL, 0.5% sodium deox-ycholate, and 0.1%SDS), supplementedwith a complete proteaseinhibitor cocktail (Roche). The protein concentration of the lysatewas determined by Pierce BCA Protein Assay (Thermo Scientific).The lysate was run on 8% SDS polyacrylamide gel (20 mg ofprotein per lane), and proteins were transferred onto PVDFmembrane (Millipore). The blot was blocked with the OdysseyBlocking Buffer (LI-COR Biosciences) and probed with anti-MCT4 antibody. Vinculin was used as a loading control. Follow-ingovernight incubation at 4�C, the primary antibodywas probedwith the corresponding secondary antibody and detected usingthe Odyssey Infrared Imaging System (LI-COR Biosciences) andImage Studio Version 3.1 (LI-COR Biosciences). Densitometryanalysis was done using ImageJ (NIH, Bethesda, MD).
Modified Boyden chamber assayThe migration and invasion potential of PC-3 cells following
treatment with MCT4 ASOs was investigated using Matrigel-coatedmodifiedBoyden chambers (BDBiosciences) as previouslydescribed (38). Briefly, ASO-treated cells were seeded into the topchamber at 50,000 live cells per well. The cells were then resus-pended after 48 hours using dissociation buffer (Trevigen) con-taining calcein AMS (12.5 mmol/L; Trevigen). The number ofmigrated/invaded cells in the lower chamber was determined byfluorescence measurement (485-nm excitation, 520-nm emis-sion) of the cell suspensions using the Infinite F500 fluorometer(Tecan).
Lactate and glucose determinationPC-3 cells transfected with ASOs for 48 hours were assessed
for lactate and glucose levels. Cells were incubated with freshmedia for 4 hours. A sample of the media was then taken anddeproteinized with 10K Spin Columns (BioVision) prior to
determination of lactate concentration using Lactate Assay Kit(BioVision) and glucose concentration using Glucose Assay Kit(BioVision). Intracellular lactate levels were determined bylysing ASO-transfected cells in MQH2O. Final concentrationswere determined by normalizing to the total number of livecells.
Treatment with MCT4 ASO of PC-3 tumor-bearing nude micePC-3 cells (106 cells in 1:1 HBSS:Matrigel) were injected sub-
cutaneously into both flanks of 24 male athymic nude mice(Simonsen Laboratories). Once the mean tumor volume hadreached approximately 100 mm3, mice were randomized intofour groups and treated with intraperitoneal injections of MCT4ASO#1, #14, control ASO, or vehicle (PBS) at 10mg/kg daily for 5days, followed by 2 days off treatment for a total of 15 days. Thehealth of the mice was monitored throughout the study bymeasuring body weights and checking for abnormal behavior,such as lethargy, lack of hydration, and additional signs ofweakness. Tumor size was measured twice weekly and tumorvolume calculated using the formula: volume¼ length�width�depth� 0.5236 (mm3). Mice were sacrificed 1 hour after the finaldose for tissue harvesting.
IHC of tumor tissueTumor tissue was formalin-fixed and paraffin-embedded
for immunohistochemical analysis. Tissues were sectioned,probed, and stained with DAB (Sigma) as previously described(43). For Ki-67 and cleaved caspase-3 staining, images of fiverandom fields at 400�magnification were taken per tumor andcells counted to determine the percentage of positively stainedcells. For MCT4, images of five random fields at 200� magni-fication were taken per tumor, and staining intensity wasassessed by percentage scoring, using the formula: intensity¼ (% area score 3) � 3 þ (% area score 2) � 2 þ (% area score1) � 1. The extent of immune cell aggregation was quantifiedfollowing CD31 staining by imaging the five most prominentregions of aggregates per tumor at 200� magnification anddetermining the percent area of the field they occupied. Theproportions of immune cells were evaluated as the area ofpositive staining normalized to the area occupied by immunecell aggregates in the same five prominent regions.
Statistical analysisAll pooled results are represented as mean � SEM. Statistical
analysis was performed using GraphPad Prism 6 (GraphPadSoftware, Inc). The Student t test was carried out to comparemeans between two groups. One-way ANOVA followed by thepost hocDunnett testwas used to comparemeansofmore than twogroups. Two-way ANOVA followed by post-hoc multiple compar-ison was applied to compare tumor growth. A contingency testwas done to compare staining intensity among patient cohorts onthe TMA. A log rank test was done to compare patient survivalcurves.c2 testswere done to correlateMCT4 expression levelswithvarious clinical parameters. Results with P < 0.05 were consideredstatistically significant and are indicated by �,P < 0.05; ��,P < 0.01;and ���, P < 0.001.
ResultsElevated MCT4 protein expression is associated with CRPC
A TMA composed of tissues from Gleason grade 3, 4, and 5human prostate cancers was stained for MCT4 protein. As shown
Choi et al.
Clin Cancer Res; 22(11) June 1, 2016 Clinical Cancer Research2724
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624

Figure 2.Efficacy screening ofMCT4-targeting ASOs using PC-3 cells in vitro reveals candidatesmarkedly inhibitingMCT4 expression and cell proliferation. MCT4ASOs#1 and#14 exert their in vitro effects in a dose-dependent manner, and the effects persist past 96 hours after transfection. A, siRNA silencing of MCT4 in PC-3 cells showedsignificant inhibition of cell proliferation, indicating that inhibition of MCT4 expression could have potential therapeutic efficacy. B, screening of tenMCT4-targetingASOs revealed varying inhibitory effects on cell proliferation, with sequences #1 and #14 showing the most profound inhibitions. C, MCT4 ASOs inducedvarious levels of MCT4 knockdown as measured by qPCR and Western blot, with ASOs #1 and #14 being the most effective. D, a strong correlation (P < 0.001) wasfound between theMCT4mRNAexpression and the resulting cell numbers, indicating that the inhibitory effects on cell proliferation are strongly related to decreasedMCT4expression (dotted lines represent the95%confidence interval). E, transfectionof 5 to 200nmol/L ofMCT4ASOs showed that the inhibitionof cell proliferationand expression of MCT4 is similarly dose dependent (IC50¼ 32 nmol/L for ASO#14 and IC50¼ 50 nmol/L for ASO#1). F, a time course experiment demonstrates thatthe inhibition of cell proliferation and MCT4 expression following MCT4 ASO transfections persists up to at least 96 hours post transfection. ��� , P < 0.001.
MCT4 as a Potential Therapeutic Target for CRPC
www.aacrjournals.org Clin Cancer Res; 22(11) June 1, 2016 2725
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624

Figure 3.MCT4 ASOs #1 and #14 are effective in vitro against human prostate cancer cells other than PC-3, but not against mouse prostate cancer cells. A, candidate MCT4ASOs are also able to inhibit C4-2 human prostate cancer cell proliferation and expression of MCT4 in a dose-dependent manner with IC50 values comparableto those observed with PC-3 cells. B, candidate MCT4 ASOs also inhibit DU145 human prostate cancer cell proliferation in a dose-dependent manner with IC50
values similar to those obtained with the other cell lines. The inhibition of cell proliferation is accompanied by a decrease in MCT4 expression. C, candidateMCT4 ASOs do not have any appreciable effects on TRAMPC2 mouse prostate cancer cells. They neither affect cell proliferation nor mouse MCT4 expression levelseven at the highest tested concentration of 200 nmol/L. This indicates that our candidate MCT4 ASOs are specific for human MCT4 and that the inhibitionof cell proliferation is a phenomenon related to MCT4 knockdown.
Choi et al.
Clin Cancer Res; 22(11) June 1, 2016 Clinical Cancer Research2726
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624

in Fig. 1A and B, Gleason grade 5 prostate cancers had signif-icantly increased MCT4 protein expression relative to Gleasongrade 3 and 4 specimens. Also, elevated MCT4 expression wasassociated with an earlier time to relapse from primary treat-ment, as measured by increases in serum prostate-specificantigen (PSA) levels, with the high MCT4–expressing cohorthaving a median time to relapse of 63.3 months versus 94.2months for the low MCT4–expressing cohort (Fig. 1C). Inaddition, high MCT4 expression was correlated with otherclinical characteristics associated with poor prognosis, such ashigher serum PSA levels at diagnosis and clinical T stage(Supplementary Table S2). Furthermore, elevated MCT4 pro-tein expression was found in tumors from patients subjected toprolonged neoadjuvant hormone therapy (>6 months) andCRPC patients (Fig. 1D and E), indicating that elevated expres-sion of MCT4 protein in prostate cancer is associated with thedevelopment of CRPC.
Knockdown of MCT4 inhibits PC-3 cell proliferationThe potential therapeutic efficacy of inhibiting MCT4 expres-
sion was investigated using MCT4-targeting siRNAs and ASOs.Human PC-3 CRPC cells were used as they present a distinctglycolytic metabolic profile, a property associated with MCT4expression (20). As shown in Fig. 2A, treatment of PC-3 cellswith MCT4 siRNA led to an inhibition of cell proliferation,suggesting potential therapeutic efficacy of an MCT4 knock-down approach. Accordingly, ASOs specifically targetinghuman MCT4 were designed. Screening of ten MCT4 ASOsrevealed varying capacities of inhibiting PC-3 cell proliferationand MCT4 expression, with ASOs #1 and #14 showing thegreatest potency (Fig. 2B and C). A strong correlation was foundbetween the reduced levels of MCT4mRNA in PC-3 cells treatedwith the various ASOs and the resulting cell numbers (Fig. 2D),indicating that the growth inhibition by the ASOs was directlyrelated to MCT4 knockdown.
Candidate MCT4 ASOs inhibit PC-3 cell proliferation in asustained and dose-dependent manner
The growth-inhibitory activities of MCT4 ASOs #1 and #14were further characterized. As shown in Fig. 2E, PC-3 cell prolif-eration was inhibited by both ASOs in a dose-dependentmanner,with ASO #14 being slightly more effective (IC50 ¼ 26 nmol/L)than ASO #1 (IC50 ¼ 50 nmol/L). The ASOs also reduced MCT4mRNA levels in a dose-dependent manner, mirroring their inhi-bition of cell proliferation (IC50 ASO #14 ¼ 32 nmol/L and IC50
ASO#1¼50nmol/L). As shown in Fig. 2F, bothASOsmaintainedthe inhibition of cell proliferation even at 96 hours post trans-fection. While MCT4 mRNA levels began to increase slightlystarting after 48 hours of transfection, MCT4 protein levelsremained low even at 96 hours.
Candidate MCT4 ASOs exert similar growth-inhibitory andMCT4 expression–inhibitory effects in other human prostatecancer cell lines and are specific for human MCT4
When theMCT4ASOs #1 and #14were transfected into humanC4-2 CRPC cells, they inhibited their proliferation with IC50
values comparable to those observed with PC-3 cells, i.e., ASO#1 ¼ 40 nmol/L and ASO #14 ¼ 27 nmol/L. Similarly, theyreduced the MCT4 expression with IC50 values of 50 nmol/L forASO#1 and 26 nmol/L for ASO#14 (Fig. 3A). Furthermore, when
the ASOs were transfected into human DU145 prostate cancercells, a similar inhibitory effect on cell proliferation and MCT4expression was observed with almost identical IC50 values (Fig.3B). Transfection of ASOs into LNCaP prostate cancer cells alsoshowed a similar inhibition of cell proliferation and MCT4expression (Supplementary Fig. S1), suggesting that the inhib-itory effect is more associated with a glycolytic phenotype thanandrogen receptor status. Importantly, transfection of MCT4ASOs #1 and #14 into mouse TRAMPC2 prostate cancer cellsdid not lead to a significant reduction in cell proliferation ormouse MCT4 expression (Fig. 3C). Taken together, the resultssuggest that these ASOs specifically target human MCT4 andthat their inhibitory effect on cell proliferation is a consequenceof MCT4 knockdown.
Candidate MCT4 ASOs are able to inhibit glucose metabolismand tissue invasion/migration of CRPC cells in vitro
To further examine the effects of MCT4-targeting ASOs onprostate cancer cells, we measured their effects on lactic acidsecretion, intracellular lactate concentrations, and glucose con-sumption of PC-3 cells. Transfection of the cells with MCT4ASOs #1 and #14 led to a marked inhibition of lactic acidsecretion, a corresponding accumulation of intracellular lac-tate, and an extensive decrease in glucose consumption, mea-sured after 48 hours of transfection (Fig. 4A). Furthermore, asshown in Fig. 4B, treatment with the ASOs resulted in thedownregulation of various genes involved in glycolysis, i.e.,GAPDH, PGK1, PGAM1, and ENO1. In addition, the expressionof lactate dehydrogenase A was found to be depressed, indic-ative of a decrease in the conversion of pyruvate to lactic acid.Moreover, decreased expression was found for pyruvate dehy-drogenase kinase-1, an enzyme that shunts pyruvate away fromthe TCA cycle and promotes its conversion to lactic acid. Thus,the treatment with the MCT4 ASOs led to inhibition of aerobicglycolysis.
Treatment with MCT4 ASOs also inhibited the migration andtissue invasion of PC-3 cells in modified Boyden chambers(Supplementary Fig. S2), suggesting that lactic acid secretion asfacilitated by MCT4 could also play an important role in themetastatic process.
Growth of PC-3 xenografts in nudemice inhibited by treatmentwith MCT4 ASOs
Male athymic nude mice bearing subcutaneous PC-3 tumorswere treated with MCT4 ASOs #1 and #14 for a total of 15days. Both ASOs markedly inhibited the growth of the tumors(Fig. 5A) without inducing major host toxicity, as assessed bymonitoring animal weights (Supplementary Fig. S3) andbehavior. Immunohistochemical analysis revealed that theASO-induced inhibition of tumor growth was associated withan increase in cell apoptosis, as measured by cleaved caspase-3staining and a decrease in cell proliferation, as measured by Ki-67 staining (Fig. 5B). The decrease in tumor growth wasassociated with a decrease in MCT4 protein expression (Fig.5C), consistent with an antiproliferative effect generated byMCT4 knockdown.
Effects of MCT4 ASOs on immune cell aggregates in nude miceAs lactic acid–induced acidification of tumors has been linked
to the suppression of local host anticancer immunity (14), it wasof interest to determinewhether the treatment of the PC-3 tumor–
MCT4 as a Potential Therapeutic Target for CRPC
www.aacrjournals.org Clin Cancer Res; 22(11) June 1, 2016 2727
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624
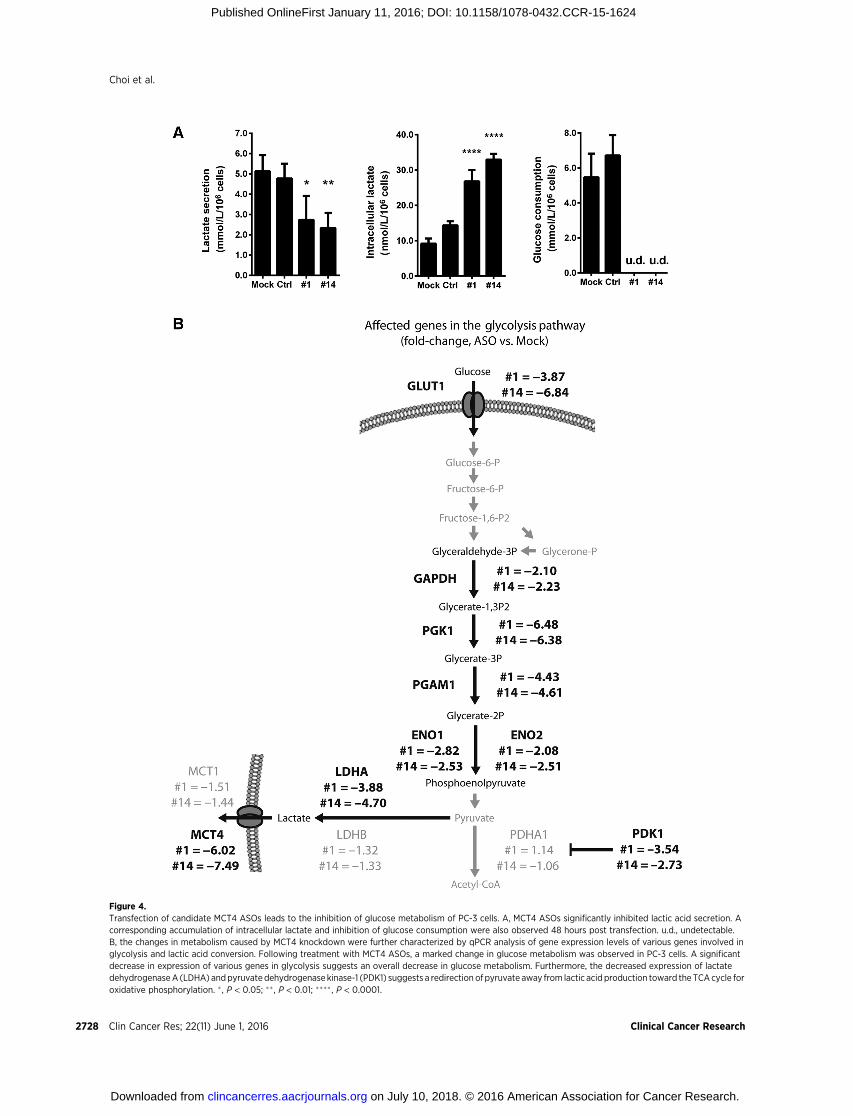
Figure 4.Transfection of candidate MCT4 ASOs leads to the inhibition of glucose metabolism of PC-3 cells. A, MCT4 ASOs significantly inhibited lactic acid secretion. Acorresponding accumulation of intracellular lactate and inhibition of glucose consumption were also observed 48 hours post transfection. u.d., undetectable.B, the changes in metabolism caused by MCT4 knockdown were further characterized by qPCR analysis of gene expression levels of various genes involved inglycolysis and lactic acid conversion. Following treatment with MCT4 ASOs, a marked change in glucose metabolism was observed in PC-3 cells. A significantdecrease in expression of various genes in glycolysis suggests an overall decrease in glucose metabolism. Furthermore, the decreased expression of lactatedehydrogenaseA (LDHA) andpyruvate dehydrogenase kinase-1 (PDK1) suggests a redirection of pyruvate away from lactic acid production toward the TCAcycle foroxidative phosphorylation. � , P < 0.05; �� , P < 0.01; ���� , P < 0.0001.
Clin Cancer Res; 22(11) June 1, 2016 Clinical Cancer Research2728
Choi et al.
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624

Figure 5.MCT4 ASO–induced reduction of MCT4 expression in PC-3 tumor cells in vivo was associated with inhibition of PC-3 tumor growth, characterized by anincrease in apoptosis and inhibition of cell proliferation. Athymic nude mice bearing subcutaneous PC-3 tumors were treated with intraperitoneal injections ofMCT4 ASOs #1, #14, control ASO, or vehicle (PBS) at 10 mg/kg daily for 5 days followed by 2 days off treatment for a total of 15 days. A, treatmentwith the MCT4 ASOs significantly slowed down tumor growth. B, the decrease in tumor growth rates following treatment with MCT4 ASOs is attributable to anincrease in cell apoptosis and a decrease in cell proliferation. C, treatment with MCT4 ASOs decreased MCT4 expression in the tumor, as measured byimmunohistochemical staining. Representative images of tumors from each group show the presence of strong membrane staining in the control tumors thatis absent in the MCT4 ASO–treated tumors. � , P < 0.05; �� , P < 0.01.
www.aacrjournals.org Clin Cancer Res; 22(11) June 1, 2016 2729
MCT4 as a Potential Therapeutic Target for CRPC
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624

bearing nude mice with MCT4 ASOs had caused changes in thelocal host immune response of these mice, even though theirimmune reactivity was very limited. To that end, we quantifiedimmune cell aggregates that had extravasated from CD31þ bloodvessels, particularly in the tumor periphery. As shown in Fig. 6A,xenografts treated with the two MCT4 ASOs had significantlylarger immune cell aggregates compared with control tumors.Quantification of the natural killer (NK) cell population, thepredominant cytotoxic immune cell subtype in nude mice(44), revealed that the treatment with the MCT4 ASOs markedlyincreased the proportion of tumor-associated NK cells (Fig. 6B).Furthermore, the activation of NK cells is facilitated by CD3 (45),a molecule commonly regarded as a T-cell marker for its associ-ationwith the T-cell receptor complex. Its expression inNK cells isdetectable by IHC (46), and in view of the absence of T cells innude mice, can be used as an indicator of NK cell activation (47).As shown in Fig. 6C, CD3 staining revealed that the proportion ofactivated NK cells associated with the ASO-treated tumors hadalso increased.
DiscussionCancers have a common preference for reprogrammed
energy metabolism leading to upregulated glycolysis andexcessive lactic acid secretion, an emerging hallmark of cancer(48); the lactic acid secretion is thought to be predominantlymediated by the MCT4 plasma membrane transporter protein(22). This study indicates that the progression of prostatecancer to CRPC, a major challenge in the management of thedisease (1), is coupled to an increase in MCT4 expression.Elevated MCT4 protein expression in clinical prostate cancerspecimens was associated with increases in Gleason grade,prolonged treatment of patients with neoadjuvant hormonetherapy, CRPC, and early disease relapse (Fig. 1A–E), as wellas other clinicopathologic characteristics indicative of poorprognosis (Supplementary Table S2). These observations areconsistent with reports by others (28,29), indicating thatelevated MCT4 expression is clinically relevant and may playan important role in the late and more aggressive stages of thedisease. Thus, stimulation of cancer progression by elevatedexpression of MCT4 leading to increased lactic acid secretion isconsistent with the notion that lactic acid generated by cancerscan aid their progression by promoting a variety of oncogenicprocesses.
Support for potential use of MCT4-targeting ASOs for therapyof CRPC comes from our findings that the treatment with MCT4ASOs#1 and#14markedly reducedMCT4mRNAexpression (Fig.2C), lactic acid secretion, and glucose consumption by PC-3 cells(Fig. 4A) and inhibited in vitro proliferation of CRPC cell lines in aspecies-specific manner (Figs 2A–F and 3A–C). Furthermore,treatment of PC-3 tumor-bearing nude mice with the MCT4-targeting ASOs led to marked inhibition of the growth of thetumors (Fig. 5A), which was associated with increased apoptosis(Fig. 5B) and a reduction inMCT4protein levels (Fig. 5C). Aswell,the treatment with the ASOs did not induce major toxicity to thehosts (Supplementary Fig. S3).
The MCT4 ASOs produced in this study appear to be highlyspecific for human MCT4, as the specificities of their sequenceswere checked via BLAST analysis (see Materials and Methods)and as they had highly similar inhibitory effects on the MCT4expression and proliferation of human PC-3, DU145, C4-2,
and LNCaP prostate cancer cells (Figs 2A–F, 3A and B, andSupplementary Fig. S1) as distinct from mouse TRAMPC2prostate cancer cells, which were not significantly affected bythe ASOs (Fig. 3C). The marked MCT4 ASO–induced inhibitionof lactic acid secretion and corresponding increase in intracel-lular lactic acid concentration are therefore likely a direct resultof MCT4 knockdown (Fig. 4A). The ASO-induced inhibition ofglucose consumption (Fig. 4A) and downregulation of enzymesinvolved in aerobic glycolysis (Fig. 4B) appear to be secondaryeffects, probably reflecting a response of the cancer cells toreduce the potentially fatal increase in intracellular lactic acidcontent by shutting off the lactic acid supply. Furthermore,decreasing MCT4 expression also resulted in the reduction ofMatrigel invasion and in vitro migration of PC-3 cells (Supple-mentary Fig. S2), lending preliminary confirmation that lacticacid secretion may play an important role in local tissueinvasion and metastasis (15,16). Elucidation of the precisemechanisms by which the downregulation of MCT4 inhibitsproliferation, tissue invasion, and migration of prostate cancercells and other aspects of cancer biology, such as tumorigenesis,requires further studies. In particular, FDG-PET could be usedas a metabolic tracer to determine whether glucose uptake byprostate cancer cells in vivo is affected by treatment with MCT4ASOs. Alternatively, metabolomic approaches using stable iso-tope–labeled metabolites, such as 13C-glucose (49,50), may beused to more comprehensively assess alterations in glucosemetabolism following MCT4 knockdown.
Of interest are the findings that the treatment of the PC-3tumor–bearing nude mice with the MCT4 ASOs led to increasesin (i) the number of extravasated immune cell aggregates in thetumor cell microenvironment (Fig. 6A), (ii) the proportions ofhost NK cells (Fig. 6B), and (iii) the proportions of host NK cellsthat had been activated (Fig. 6C). They indicate that MCT4knockdown might lead to restoration of cancer-suppressed hostimmunity, a possibility that should be examined using immu-nocompetent hosts.
Elevated MCT4 expression has also been reported as clinicallyrelevant and functionally significant in other types of cancer(25,27). For example, MCT4-targeting siRNA has been reportedto be effective at inhibiting renal cell carcinoma growth (51), andMCT4 expression in small cell lung cancer was found to conferresistance to specific MCT1 inhibitors (32). The candidate MCT4ASOs developed in our laboratory could therefore be useful inmultiple clinical settings, in particular if their efficacies can beimproved by incorporating second-generation 20-methoxyethylmodifications in their backbone (52). In addition, treatmentefficacy could also be increased using a combination strategywith other therapeutics currently used for treating CRPC (suchas second-generation hormone therapies or docetaxel). Alterna-tively, combination with other modulators of glucose metabo-lism (such as metformin or mitochondrial inhibitors) mightresult in a more complete blockade of glucose utilization bycancer cells.
ConclusionsThe development of CRPC was found to be associated with
elevated expression of MCT4, a plasma membrane transporterprotein mediating lactic acid secretion by cancers. MCT4-targeting ASOs that inhibit lactic acid secretion may be usefulfor therapy of CRPC and other cancers characterized by
Choi et al.
Clin Cancer Res; 22(11) June 1, 2016 Clinical Cancer Research2730
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624

Figure 6.MCT4-targetingASOs increase extravasated immune cell aggregation and alter tumor-associated immune cell proportions in vivo. The potential immunomodulatoryproperties of MCT4 ASOs were investigated through immunohistochemical staining. A, immune aggregates are characterized by areas of small, circular,densely packed nuclei that are distinct from the surrounding tumor cells. Staining for CD31 reveals that these immune cells have extravasated and surround the bloodvessels in the tumor periphery. The immunomodulatory effects of treatment with MCT4 ASO are partially exerted through a significant increase in the extentof such immune cell aggregations. B, treatment with MCT4 ASO also significantly altered the composition of the immune cells present in the aggregates.Staining using the NK cell marker NK1.1 revealed that treatment with MCT4 ASO increased the proportion of NK cells associated with the tumor. C, becausenudemice lack T cells, CD3was used as amarker for activatedNK cells. Staining usingCD3 also revealed that treatmentwithMCT4ASOalso increased the proportionof activated NK cells associated with the tumor, suggesting stimulation of anticancer immunity. � , P < 0.05; �� , P < 0.01.
MCT4 as a Potential Therapeutic Target for CRPC
www.aacrjournals.org Clin Cancer Res; 22(11) June 1, 2016 2731
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624

excessive lactic acid secretion resulting from reprogrammedcellular energy (glucose) metabolism, an emerging hallmarkof cancer.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: S.Y.C. Choi, C.C. Collins, M.E. Gleave, Y. WangDevelopment of methodology: S.Y.C. Choi, H. XueAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): S.Y.C. Choi, H. Xue, R. Wu, C.C. Collins, M.E. GleaveAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): S.Y.C. Choi, D. Lin, C.C. Collins, M.E. GleaveWriting, review, and/or revision of the manuscript: S.Y.C. Choi, C.C. Collins,M.E. Gleave, P.W. Gout, Y. WangAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): R. Wu, D. Lin
Study supervision: C.C. Collins, Y. WangOther (pathology): L. Fazli
AcknowledgmentsThe authors thank James Killam, Fang Zhang, and Francesco Crea for their
support during various scientific discussions and Estelle Li, Xindy Dong, JoyWang, and Kendra Fu for their technical assistance.
Grant SupportThis study was supported by The Urology Foundation for LAST Project (to Y.
Wang), CIHR Master's Award (Frederick Banting and Charles Best CanadaGraduate Scholarships; to S.Y.C. Choi).
The costs of publication of this articlewere defrayed inpart by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received July 8, 2015; revised November 30, 2015; accepted December 6,2015; published OnlineFirst January 11, 2016.
References1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin
2015;65:5–29.2. Albertsen PC, Hanley JA, Fine J. 20-year outcomes following conserva-
tive management of clinically localized prostate cancer. JAMA 2005;293:2095–101.
3. Ning YM, Pierce W, Maher VE, Karuri S, Tang SH, Chiu HJ, et al.Enzalutamide for treatment of patients with metastatic castration-resis-tant prostate cancer who have previously received docetaxel: U.S. Foodand Drug Administration drug approval summary. Clin Cancer Res2013;19:6067–73.
4. de Bono JS, Logothetis CJ, Molina A, Fizazi K, North S, Chu L, et al.Abiraterone and increased survival in metastatic prostate cancer. N Engl JMed 2011;364:1995–2005.
5. Loriot Y, Bianchini D, Ileana E, Sandhu S, Patrikidou A, Pezaro C, et al.Antitumour activity of abiraterone acetate against metastatic castration-resistant prostate cancer progressing after docetaxel and enzalutamide(MDV3100). Ann Oncol 2013;24:1807–12.
6. Badrising S, van der Noort V, van Oort IM, van den Berg HP, Los M,Hamberg P, et al. Clinical activity and tolerability of enzalutamide(MDV3100) inpatientswithmetastatic, castration-resistant prostate cancerwho progress after docetaxel and abiraterone treatment. Cancer 2014;120:968–75.
7. Tannock IF, de Wit R, Berry WR, Horti J, Pluzanska A, Chi KN, et al.Docetaxel plus prednisone or mitoxantrone plus prednisone for advancedprostate cancer. N Engl J Med 2004;351:1502–12.
8. Yuan TC, Veeramani S, Lin MF. Neuroendocrine-like prostate cancer cells:neuroendocrine transdifferentiation of prostate adenocarcinoma cells.Endocr Relat Cancer 2007;14:531–47.
9. Nadal R, SchweizerM, KryvenkoON, Epstein JI, EisenbergerMA. Small cellcarcinoma of the prostate. Nat Rev Urol 2014;11:213–9.
10. Beltran H, Rickman DS, Park K, Chae SS, Sboner A, MacDonald TY, et al.Molecular characterization of neuroendocrine prostate cancer and identi-fication of new drug targets. Cancer Discov 2011;1:487–95.
11. Zhang Y, Yang JM. Altered energy metabolism in cancer: a uniqueopportunity for therapeutic intervention. Cancer Biol Ther 2013;14:81–9.
12. Warburg O. On respiratory impairment in cancer cells. Science 1956;124:269–70.
13. Koochekpour S, Majumdar S, Azabdaftari G, Attwood K, Scioneaux R,Subramani D, et al. Serum glutamate levels correlate with gleason scoreand glutamate blockade decreases proliferation, migration, and inva-sion and induces apoptosis in prostate cancer cells. Clin Cancer Res2012;18:5888–901.
14. Choi SY, Collins CC, Gout PW, Wang Y. Cancer-generated lactic acid: aregulatory, immunosuppressive metabolite? J Pathol 2013;230:350–5.
15. Choi SY, Lin D, Gout PW, Collins CC, Xu Y, Wang Y. Lessons from patient-derived xenografts for better in vitromodeling of human cancer. Adv DrugDeliv Rev 2014;79–80:222–37.
16. Doherty JR, Cleveland JL. Targeting lactate metabolism for cancer thera-peutics. J Clin Invest 2013;123:3685–92.
17. Ullah MS, Davies AJ, Halestrap AP. The plasma membrane lactatetransporter MCT4, but not MCT1, is up-regulated by hypoxiathrough a HIF-1alpha-dependent mechanism. J Biol Chem 2006;281:9030–7.
18. Parks SK, Chiche J, Pouyss�egur J. Disrupting proton dynamics andenergy metabolism for cancer therapy. Nat Rev Cancer 2013;13:611–23.
19. Jadvar H. Molecular imaging of prostate cancer with 18F-fluorodeoxyglu-cose PET. Nat Rev Urol 2009;6:317–23.
20. Vaz CV, Alves MG, Marques R, Moreira PI, Oliveira PF, Maia CJ, et al.Androgen-responsive and nonresponsive prostate cancer cells present adistinct glycolytic metabolism profile. Int J Biochem Cell Biol 2012;44:2077–84.
21. Tennakoon JB, Shi Y, Han JJ, Tsouko E, White MA, Burns AR, et al.Androgens regulate prostate cancer cell growth via an AMPK-PGC-1a-mediated metabolic switch. Oncogene 2014;33:5251–61.
22. Dimmer KS, Friedrich B, Lang F, Deitmer JW, Br€oer S. The low-affinitymonocarboxylate transporter MCT4 is adapted to the export of lactate inhighly glycolytic cells. Biochem J 2000;350:219–27.
23. Halestrap AP. The SLC16 gene family - structure, role and regulation inhealth and disease. Mol Aspects Med 2013;34:337–49.
24. Manning Fox JE, Meredith D, Halestrap AP. Characterisation of humanmonocarboxylate transporter 4 substantiates its role in lactic acid effluxfrom skeletal muscle. J Physiol 2000;529:285–93.
25. Fisel P, Kruck S, Winter S, Bedke J, Hennenlotter J, Nies AT, et al. DNAmethylation of the SLC16A3 promoter regulates expression of the humanlactate transporter MCT4 in renal cancer with consequences for clinicaloutcome. Clin Cancer Res 2013;19:5170–81.
26. Ohno A, Yorita K,HaruyamaY, KondoK, Kato A,Ohtomo T, et al. Aberrantexpression of monocarbohydrate transporter 4 (MCT4) in tumor cellspredicts an unfavorable outcome in patients with hepatocellular carcino-ma. Liver Int 2014;34:942–52.
27. Lisanti MP, Sotgia F, Pestell RG, Howell A, Martinez-Outschoorn UE.Stromal glycolysis andMCT4 are hallmarks ofDCISprogression to invasivebreast cancer. Cell Cycle 2013;12:2935–6.
28. P�ertega-Gomes N, Vizcaíno JR, Miranda-Goncalves V, Pinheiro C, Silva J,Pereira H, et al. Monocarboxylate transporter 4 (MCT4) and CD147overexpression is associated with poor prognosis in prostate cancer. BMCCancer 2011;11:312.
29. Hao J, Chen H, Madigan MC, Cozzi PJ, Beretov J, Xiao W, et al. Co-expression of CD147 (EMMPRIN), CD44v3–10, MDR1 andmonocarbox-ylate transporters is associated with prostate cancer drug resistance andprogression. Br J Cancer 2010;103:1008–18.
30. Sanit�a P, Capulli M, Teti A, Galatioto GP, Vicentini C, Chiarugi P, et al.Tumor-stroma metabolic relationship based on lactate shuttle can sustainprostate cancer progression. BMC Cancer 2014;14:154.
Choi et al.
Clin Cancer Res; 22(11) June 1, 2016 Clinical Cancer Research2732
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624

31. Marchiq I, Le Floch R, RouxD, SimonMP, Pouyssegur J. Genetic disruptionof lactate/Hþ Symporters (MCTs) and their subunit CD147/BASIGINsensitizes glycolytic tumor cells to phenformin. Cancer Res 2015;75:171–80.
32. Polan´ski R, Hodgkinson CL, Fusi A, Nonaka D, Priest L, Kelly P, et al.Activity of the monocarboxylate transporter 1 inhibitor AZD3965 in smallcell lung cancer. Clin Cancer Res 2014;20:926–37.
33. Ovens MJ, Davies AJ, Wilson MC, Murray CM, Halestrap AP. AR-C155858is a potent inhibitor of monocarboxylate transporters MCT1 and MCT2that binds to an intracellular site involving transmembrane helices 7–10.Biochem J 2010;425:523–30.
34. Sonveaux P, V�egran F, Schroeder T, Wergin MC, Verrax J, Rabbani ZN, et al.Targeting lactate-fueled respiration selectively kills hypoxic tumor cells inmice. J Clin Invest 2008;118:3930–42.
35. Colen CB, Shen Y, Ghoddoussi F, Yu P, Francis TB, Koch BJ, et al. Metabolictargeting of lactate efflux by malignant glioma inhibits invasiveness andinduces necrosis: an in vivo study. Neoplasia 2011;13:620–32.
36. Draoui N, Schicke O, Seront E, Bouzin C, Sonveaux P, Riant O, et al.Antitumor activity of 7-aminocarboxycoumarin derivatives, a new class ofpotent inhibitors of lactate influx but not efflux. Mol Cancer Ther2014;13:1410–8.
37. Thomas C, Zoubeidi A, Kuruma H, Fazli L, Lamoureux F, Beraldi E, et al.Transcription factor Stat5 knockdown enhances androgen receptor degra-dation and delays castration-resistant prostate cancer progression in vivo.Mol Cancer Ther 2011;10:347–59.
38. Chiang YT, Wang K, Fazli L, Qi RZ, Gleave ME, Collins CC, et al.GATA2 as a potential metastasis-driving gene in prostate cancer.Oncotarget 2014;5:451–61.
39. Matveeva OV, Tsodikov AD, Giddings M, Freier SM, Wyatt JR, SpiridonovAN, et al. Identification of sequence motifs in oligonucleotides whosepresence is correlated with antisense activity. Nucleic Acids Res2000;28:2862–5.
40. Mullick AE, FuW, GrahamMJ, Lee RG, Witchell D, Bell TA, et al. Antisenseoligonucleotide reduction of apoB-ameliorated atherosclerosis in LDLreceptor-deficient mice. J Lipid Res 2011;52:885–96.
41. Samuel VT, Choi CS, Phillips TG, Romanelli AJ, Geisler JG, Bhanot S, et al.Targeting foxo1 in mice using antisense oligonucleotide improves hepaticand peripheral insulin action. Diabetes 2006;55:2042–50.
42. Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A,et al. Accurate normalization of real-time quantitative RT-PCR data bygeometric averaging of multiple internal control genes. Genome Biol2002;3:RESEARCH0034.
43. Wang Y, XueH, Cutz JC, Bayani J,Mawji NR, ChenWG, et al. An orthotopicmetastatic prostate cancer model in SCIDmice via grafting of a transplant-able human prostate tumor line. Lab Invest 2005;85:1392–404.
44. Shultz LD, Ishikawa F, Greiner DL. Humanized mice in translationalbiomedical research. Nat Rev Immunol 2007;7:118–30.
45. Koch J, Steinle A, Watzl C, Mandelboim O. Activating natural cytotoxicityreceptors of natural killer cells in cancer and infection. Trends Immunol2013;34:182–91.
46. Morice WG. The immunophenotypic attributes of NK cells and NK-celllineage lymphoproliferative disorders. Am J Clin Pathol 2007;127:881–6.
47. Lanier LL, Chang C, Spits H, Phillips JH. Expression of cytoplasmic CD3epsilon proteins in activatedhumanadult natural killer (NK) cells andCD3gamma, delta, epsilon complexes in fetal NK cells. Implications for therelationship of NK and T lymphocytes. J Immunol 1992;149:1876–80.
48. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell2011;144:646–74.
49. Gravel SP, Hulea L, Toban N, Birman E, Blouin MJ, Zakikhani M, et al.Serine deprivation enhances antineoplastic activity of biguanides. CancerRes 2014;74:7521–33.
50. Gravel SP, Andrzejewski S, Avizonis D, St-Pierre J. Stable isotope traceranalysis in isolated mitochondria from mammalian systems. Metabolites2014;4:166–83.
51. GerlingerM, SantosCR, Spencer-DeneB,Martinez P, EndesfelderD, BurrellRA, et al. Genome-wide RNA interference analysis of renal carcinomasurvival regulators identifies MCT4 as a Warburg effect metabolic target.J Pathol 2012;227:146–56.
52. Dias N, Stein CA. Antisense oligonucleotides: basic concepts and mechan-isms. Mol Cancer Ther 2002;1:347–55.
www.aacrjournals.org Clin Cancer Res; 22(11) June 1, 2016 2733
MCT4 as a Potential Therapeutic Target for CRPC
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624

2016;22:2721-2733. Published OnlineFirst January 11, 2016.Clin Cancer Res Stephen Yiu Chuen Choi, Hui Xue, Rebecca Wu, et al. Prostate Cancer
Gene: A Novel, Potential Target for Therapy of AdvancedMCT4The
Updated version
10.1158/1078-0432.CCR-15-1624doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2016/01/09/1078-0432.CCR-15-1624.DC1
Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/22/11/2721.full#ref-list-1
This article cites 52 articles, 17 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/22/11/2721.full#related-urls
This article has been cited by 1 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/22/11/2721To request permission to re-use all or part of this article, use this link
on July 10, 2018. © 2016 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst January 11, 2016; DOI: 10.1158/1078-0432.CCR-15-1624





























