Embed Size (px)
Citation preview

1553
INTRODUCTION
Muscle LIM protein (MLP) is a positive regulator of myogenicdifferentiation that was first identified in a molecular screensearching for genes that become transcriptionally upregulatedas a result of skeletal muscle denervation (Arber et al., 1994).MLP gene expression is restricted to striated muscle where itis first expressed to high levels at the onset of the differentiatedphenotype and remains expressed in both cardiac and skeletalmuscle of the adult (Arber et al., 1994). This expression patterncoincides with the expression of early muscle-specific genesand the initial events leading to functional sarcomereformation. Overexpression of MLP in myogenic cells leads toan enhancement of the differentiated phenotype whereasexpression of antisense MLP dramatically blocks myogenesis,suggesting that MLP is essential to striated muscle function(Arber et al., 1994).
MLP (also known as CRP3) belongs to a family of proteinsreferred to as the cysteine-rich proteins (CRPs; Louis et al.,1997). Each of the other CRP family members share a highdegree of identity with MLP but are expressed in non-striatedmuscle tissues such as smooth muscle (CRP1), vascular tissue(CRP2) and fibroblasts (CRP1 and CRP2; Louis et al., 1997).The CRPs are composed of two tandem LIM domains followedby a glycine-rich region (Weiskirchen et al., 1995; Arber et al.,
1994). LIM domains contain double zinc finger-like structuresthat conform to the amino acid consensus sequence [C-X2-C-X16-23-H-X2-C]-X2-[C-X2-C-X16-21-C-X2-C(D,H)] with eachfinger coordinating a single zinc ion through the characteristiccysteine and histidine residues (Michelsen et al., 1993). LIMmotifs are found in a variety of proteins that characteristicallyare associated with cell determination and differentiationevents (reviewed by Sánchez-García and Rabbitts, 1994;Dawid et al., 1998). The LIM domain has been shown to serveas a protein binding interface and thus the activity of a LIMdomain containing protein is often intimately linked to itscorresponding protein binding partner(s) (Schmeichel andBeckerle, 1994).
An interesting feature of MLP is that it exhibits a bimodalsubcellular distribution, initially present exclusively in nucleiof early differentiated muscle cells and later accumulating tohigh levels in the cytoplasm (Arber et al., 1994). In skeletalmuscle, nuclear MLP appears to positively regulatemyogenesis by binding to the MyoD family of basic helix-loop-helix (bHLH) myogenic regulatory factors through the N-terminal LIM domain (LIM1). The MLP:MyoD interactionenhances the binding of MyoD:E-protein complexes to E-boxDNA target sites, thereby increasing MyoD-mediated geneactivation (Kong et al., 1997). Cytoplasmic MLP also is capableof positively affecting myogenesis, presumably due to its ability
Journal of Cell Science 113, 1553-1564 (2000)Printed in Great Britain © The Company of Biologists Limited 2000JCS1324
Muscle LIM protein (MLP) is a striated muscle-specificfactor that enhances myogenic differentiation and is criticalto maintaining the structural integrity of the contractileapparatus. The ability of MLP to regulate myogenesisis particularly interesting since it exhibits multiplesubcellular localizations, being found in both nuclear andcytoplasmic compartments. Despite extensive biochemicalanalyses on MLP, the mechanism(s) by which it influencesthe myogenic program remains largely undefined. Tofurther examine the role of MLP as a positive myogenicregulator, a yeast two-hybrid screen was employed toidentify cytoplasmic-associated MLP binding partners.From this screen, the cytoskeletal protein βΙ-spectrin wasisolated. Protein interaction assays demonstrate that MLPand βΙ-spectrin associate with one another in vivo as wellas when tested under several in vitro binding conditions.
βΙ-spectrin binds specifically to MLP but not to the MLPrelated proteins CRP1 and CRP2 or to other LIM domaincontaining proteins. The MLP:β-spectrin interaction ismediated by the second LIM motif of MLP and by repeat7 of β-spectrin. Confocal microscopy studies also revealthat MLP co-localizes with β-spectrin at the sarcolemmaoverlying the Z- and M-lines of myofibrils in both cardiacand skeletal muscle tissue. Given that β-spectrin is a knowncostamere protein, we propose that sarcolemma-associatedMLP also serves as a key costamere protein, stabilizingthe association of the contractile apparatus with thesarcolemma by linking the β-spectrin network to the α-actinin crosslinked actin filaments of the myofibril.
Key words: Contractile apparatus, LIM domain, Muscle, Sarcomere,Spectrin
SUMMARY
The muscle regulatory and structural protein MLP is a cytoskeletal binding
partner of βΙ-spectrin
Matthew J. Flick and Stephen F. Konieczny*
Department of Biological Sciences, Purdue University, West Lafayette, IN 47907-1392, USA*Author for correspondence (e-mail: [email protected])
Accepted 22 February; published on WWW 6 April 2000

1554
to enhance downstream myogenic events while associated withthe actin-based cytoskeleton through its LIM2 domain (Arberand Caroni, 1996). In striated muscle, cytoplasmic MLPlocalizes not just to the actin-based cytoskeleton but to distinctregions along the contractile apparatus in a cross striationregister with the Z-line (Arber et al., 1997). A similarobservation has been made with the muscle LIM proteins inDrosophila. The five LIM domain protein Mlp84B localizes tothe periphery of muscle fibers at myofibril attachment sites inregions overlying and flanking Z-lines (Stronach et al., 1999). Aphysiological role for MLP has been revealed through thegeneration of MLP deficient mice which display a highlydisorganized cytoskeletal architecture (Arber et al., 1997). TheMLP null phenotype is characterized by severe myofibrildisorganization and decreased cell-cell contact culminating in acondition resembling dilated cardiomyopathy (Arber et al.,1997). Despite the initial observations regarding myofibrillocalization, identification of specific MLP binding partners andthe role of MLP in myofibril formation and sarcomeremaintenance remains largely undefined.
Here we report a novel finding demonstrating a direct linkbetween actin-associated cytoplasmic MLP and the membranebound cytoskeleton. Through a yeast two-hybrid screenemploying a cardiac cDNA library we have identified βI-spectrinas an essential MLP interacting molecule. A variety ofbiochemical protein binding assays indicate that the MLP:βΙ-spectrin interaction is specific and mediated through the LIM2and spectrin repeat 7 regions of MLP and βΙ-spectrin,respectively. Co-immunoprecipitation assays have revealed thatMLP:βΙ-spectrin interactions occur in vivo. Furthermore, MLPand β-spectrin co-localize in both skeletal and cardiac tissue atthe sarcolemma overlying Z- and M-lines. Interestingly, theMLP related proteins CRP1 and CRP2 do not bind β-spectrin,providing an important distinction between the individualCRP/MLP family members and demonstrating for the first timethat functional differences exist between CRP proteins indifferent cell types.
MATERIALS AND METHODS
Gene constructs and expression plasmidsCloning of the rat MLP cDNA and the generation of HA-epitope taggedMLP mammalian expression plasmids has been described previously(Kong et al., 1997). Note that all LIM domains for MLP/CRP constructsdescribed in this report are accompanied by the corresponding glycine-rich regions. A Gal4 DNA binding domain (DB)-MLP fusion constructwas generated by PCR amplification of the MLP cDNA using primershaving a NcoI site on the 5′ end and an EcoRI site on the 3′ end. Theresulting PCR product was cloned in-frame into the yeast expressionvector pAS2-1 (Clontech, Inc.) containing an upstream Gal4 DB. Asimilar strategy was used to generate a Gal4 DB-CRP1 fusion construct.Gal4 transcription activation domain (TAD)-paraxis and Gal4 TAD-MEF2A fusion constructs were cloned by identical techniques into theyeast expression plasmid pACT2 (Clontech, Inc.). The Gal4 DB-p53(pVA3-1), Gal4 TAD-SV40 large T antigen (pTD1-1) and the Gal4TAD-lamin C (pLAM5′-1) constructs were supplied by Clontech, Inc.with the MATCHMAKER Two-Hybrid System 2 kit.
Human β-spectrin cDNA constructs were generated by PCR usingthe previously described pcDNA βΙΣΙΙ plasmid as a template (Weed etal., 1996). β-Spectrin inserts encoding repeats 7-10 and repeats 8-10were generated using primers containing a 5′ KpnI site and an ATGinitiation codon and a 3′ EcoRI site. The resulting PCR products were
digested with KpnI and EcoRI and cloned into the pcDNA3 expressionplasmid to produce pcDNA3-β-spectrin 7-10 and β-spectrin 8-10constructs. β-Spectrin repeats 5-6 and repeats 6-7 were generated in asimilar fashion using primers containing a 5′ HindIII site and an ATGinitiation codon plus a 3′ KpnI site. The resulting PCR products weredigested and cloned into the pcDNA3 expression plasmid to producepcDNA3-β-spectrin 5-6 and β-spectrin 6-7 constructs. β-Spectrinprotein fragments containing the actin binding (AB) domain-repeat 1,repeats 1-5 and repeat 11-plekstrin homology (PH) domain weregenerated by PCR using primers containing a 5′ EcoRI site and a 3′XbaI site and cloned into pcDNA3.
The His-MLP, His-MLP-LIM1 and His-MLP-LIM2 bacterialexpression plasmids were generated as previously reported (Kong et al.,1997). The glutathione-S-transferase (GST)-MLP and GST-CRP2protein expression plasmids were produced by ligating the full-lengthMLP or CRP2 cDNA in-frame with GST into the SalI-NotI or SmaI-NotI sites of pGEX-4T2, respectively (Pharmacia Biotech Products).The GST-CRP1 and GST-LMO2 bacterial expression plasmids havebeen described previously (Jurata et al., 1996). The GST-P-LIMbacterial expression plasmid was kindly provided by Dr Simon Rhodes.GST-MLP-LIM1 and GST-MLP-LIM2 expression plasmids wereproduced by isolating the respective cDNAs from the pACT2 clones byNcoI/XhoI digestion followed by ligation into the vector pGSTag2 (giftfrom Dr O. Andrisani). β-Spectrin 7-10 myc-his and β-spectrin 8-10myc-his were generated by isolating the respective cDNAs from thepcDNA3 clones by KpnI/EcoRI digestion followed by ligation into thebacterial expression vector pTrcHis2C (Invitrogen). All constructs wereverified by DNA sequencing.
Yeast two-hybrid assaysA human adult cardiac cDNA library cloned into the Gal4 TAD yeastexpression vector pGAD10 was obtained from Clontech, Inc. and thenamplified as recommended. Two-hybrid screens were carried out bytransformation of the library into the yeast strain Y190 that had beenpreviously transformed with pAS2-1-MLP according to themanufacturer’s protocol. Clones which sustained growth on syntheticmedium (sm) lacking leucine, tryptophan and histidine were transferredto Whatman filters and tested for β-galactosidase activity. Briefly, filterswere placed in liquid nitrogen to lyse the cells and then treated with astaining solution containing the β-galactosidase substrate X-galfollowed by incubation at 30°C. The library plasmid was recoveredfrom His+/LacZ+ colonies and used to transform the bacterial strainDH5. Identification of the library plasmid was based on a series ofrestriction enzyme digests which distinguished the pGAD10 libraryplasmid from the pAS2-1-MLP bait plasmid. All isolated clones weresequenced in order to determine their identity. In order to confirm andcharacterize specific yeast two-hybrid interactions, small scaletransformations into the Y190 yeast strain were performed by standardprocedures. Cotransformation of Gal4 DB and Gal4 TAD fusion proteinencoding plasmids were performed to determine the specificity of theobserved two-hybrid interaction from the original screens.Transformants were plated on sm leu−/trp− plates and assayed for β-galactosidase activity as described above. Liquid β-galactosidase assayswere performed as described using the substrate CPRG (Miller, 1972).Three separate reactions using three individual transformants wereanalyzed for each group.
His-tagged proteins, GST-tagged proteins and in vitrobinding assaysThe following plasmids were used to generate in vitro translatedproteins: pcDNA3-β-spectrin-AB-1, pcDNA3-β-spectrin 1-5,pcDNA3-β-spectrin 5-6, pcDNA3-β-spectrin 6.5-9.5, pcDNA3-β-spectrin 6-7, pcDNA3-β-spectrin 7-10, pcDNA3-β-spectrin 8-10,pcDNA3 β-spectrin 11-PH, pcDNA3-HA-MyoD (Kong et al., 1997)and pcDNA3-E47 (Kong et al., 1997). In vitro translations wereperformed using the TNT T7 Quick Coupled Transcription/TranslationSystem (Promega). For each protein, 4 µg of plasmid DNA was
M. J. Flick and S. F. Konieczny

1555MLP and βΙ-spectrin interactions
incubated with 40 µl of lysate in the presence of [35S]methionine(Amersham) for 1 hour at 30°C. Protein production was analyzed byresolving 1 µl of the programmed lysate on a SDS-PAGE gel followedby autoradiography. The bacterially expressed fusion proteins werepurified by standard procedures and then analyzed by SDS-PAGE andCoomassie blue staining.
For in vitro protein binding assays using 35S-labeled proteins, 1 µgof His-tagged or GST-tagged protein was incubated with 10 µl of invitro translated protein in 250 µl of binding buffer (10 mM Tris-HCl,pH 7.5, 150 mM NaCl, 0.1% Triton X-100, 50 µM ZnCl2, 1 mM PMSF,1 mM DTT) for 2 hours at 4°C. The His-tagged reactions also contained20 mM imidazole in the binding buffer. For in vitro protein bindingstudies using α-actinin, 1 µg of GST-tagged protein was incubated with1 µg of purified α-actinin (Sigma) in 250 µl of Hepes binding buffer(20 mM Hepes, pH 7.4, 10 mM NaCl, 0.1 mM EGTA and 0.1% β-mercaptoethanol) for 3 hours at room temperature. The bound proteinswere subjected to three 1 ml washes with binding buffer followed byelution from the beads by heating to 95°C in the presence of Laemmlibuffer. Proteins were separated by SDS-PAGE and transferred to PVDFmembranes for detection of α-actinin by western blot analysis using ananti-α-actinin polyclonal antibody (Sigma).
For the competition binding experiment, 10 µg of β-spectrin 7-10myc-his, β-spectrin 8-10 myc-his or α-actinin was added to the GST-MLP/35S-labeled β-spectrin (6.5-9.5) binding reactions. The boundproteins were subjected to three 1 ml washes with binding bufferfollowed by elution from the beads by heating at 95°C in the presenceof Laemmli buffer. Samples were subsequently analyzed by SDS-PAGEand autoradiography.
In vitro co-immunoprecipitationsIn order to label MLP and p48 proteins with 32P, the respective cDNAswere cloned into the vector pGSTag2 which incorporates thrombincleavage and PKA phosphorylation sites downstream of the GSTprotein. The GST fusion proteins were produced in bacteria and purifiedusing glutathione agarose beads. An in vitro kinase reaction wasperformed in which 1 µg of GST fusion protein bound to glutathioneagarose beads was incubated in PKA kinase buffer containing 5 unitsof PKA (New England Biolabs) in the presence of [γ-32P]ATP(Amersham) for 30 minutes at room temperature. The phosphorylatedGST fusion proteins were then washed three times with TBS to removeunreacted ATP. The phosphorylated MLP or p48 protein wassubsequently isolated by digestion of the fusion protein with bovinethrombin (Sigma) in thrombin cleavage buffer (50 mM Tris-HCl, pH7.5, 150 mM NaCl, 2.5 mM CaCl2, 0.1% β-mercaptoethanol). Fractionsof eluted proteins were pooled and then quantified. A comparison ofthe recovered proteins indicated that they each had similar specificactivities. For co-immunoprecipitation reactions, 5×105 cpm of labeledprotein was incubated with an anti-spectrin (α and β) monoclonalantibody (Sigma) in the presence or absence of spectrin protein (purifiedfrom red blood cells; Sigma) in binding buffer (20 mM Tris-HCl, pH7.5, 120 mM NaCl, 1 mM EDTA, 1 mM DTT, 0.1% Triton X-100, 1mM PMSF, 50 µM ZnCl2, 0.1% BSA) at 4°C for 12 hours. Protein-Aagarose was then added to the reactions followed by a 1 hour incubationat 4°C. The protein complexes were washed three times with bindingbuffer and resolved by SDS-PAGE electrophoresis. ImmunoprecipitatedMLP or p48 was visualized by autoradiography.
In vivo co-immunoprecipitationsHEK293 cells were transiently transfected with expression plasmids forMLP, flag epitope-tagged βΙΣΙΙ -spectrin or both using a Ca2+-phosphateprecipitation method. After 72 hours, cells were harvested in lysis/IPbuffer (20 mM Hepes, pH 7.4, 100 mM NaCl, 0.5% Triton X-100, 10%glycerol, 10 µg/ml leupeptin, 10 µg/ml aprotinin and 1 mM PMSF) andincubated on ice for 20 minutes. Lysates were then passed through a 20gauge needle and centrifuged at 14,000 rpm to remove insolublematerial. Immunoprecipiation was performed by incubating lysateswith anti-flag M2 monoclonal antibody (Sigma) for 3 hours followed
by the addition of Protein A-Sepharose beads and a second 2 hoursincubation. The protein complexes were washed two times withLysis/IP buffer. Immunoprecipitated proteins, as well as whole cellextracts of each group, were resolved by SDS-PAGE and transferred toPVDF membrane. Detection of flag epitope-tagged βΙΣΙΙ -spectrin orMLP was accomplished by western blots using an anti-flag M2monoclonal antibody or an affinity purified anti-MLP antibody,respectively.
Tissue sections, single teased fiber preparations andimmunohistochemistryFor immunohistochemistry of tissue sections, the EDL muscle and heartwere isolated from adult Sprague-Dawley rats and embedded inTissueTek (Sakura FineTek). Cryostat sections (5 µm) were generatedusing a Zeiss microtome cryostat set at −20°C and mounted ontoSuperfrost/Plus microscope slides. Sections were fixed in 4%paraformaldehyde (PFA)/PBS solution for 5 minutes and then washedwith PBS. For single teased fibers, flexor digitorum brevis muscle wasisolated from identical rats. Initial dissection opened up the tissue topermit fixation with 4% PFA while under slight tension. Single fibersor bundles of 4-5 fibers were gently removed from the muscle tissueand were fixed in PFA for an additional 15 minutes then washed withPBS. The sections or whole mount fibers were blocked andpermeabilized with a solution of PBS, 10% HS and 0.1% Triton X-100.MLP was detected using an affinity purified MLP polyclonal antibodygenerated against the last 15 amino acids of the rat protein followed byincubation with a Texas Red- or fluorescein-conjugated anti-rabbit IgGsecondary antibody (Vector Laboratories, Inc). β-Spectrin was detectedusing the VD4 monoclonal anti-β-spectrin antibody (gift of J. Morrow)or a commercial spectrin monoclonal antibody (Chemicon) inconjunction with a Texas Red- or fluorescein-conjugated anti-mouseIgG secondary antibody (Vector Laboratories, Inc). α-Actininlocalization was examined using a sarcomeric α-actinin specificmonoclonal antibody (Sigma) followed by a Texas Red- or fluorescein-conjugated anti-mouse IgG secondary antibody. All antibodies werediluted in a solution of PBS containing 0.1% BSA. Incubations ofprimary antibodies were performed at 4°C overnight while allsecondary antibodies were incubated at room temperature for 2 hours.Control experiments for each primary antibody were performed inwhich the primary antibody in question was left out of the stainingprotocol.
All samples were mounted with Fluorsave (Calbiochem) andcoverslipped. Samples were visualized using laser scanning confocalmicroscopy (Bio-Rad MRC-1024 Laser Scanning Confocal ImagingSystem using a Nikon Optiphot-2 Microscope). Images were capturedwith Lasersharp Imaging Software (Bio-Rad).
RESULTS
Identification of potential protein partners for MLPTo identify novel protein binding partners for MLP, weperformed a yeast two-hybrid screen using an adult humancardiac cDNA library with a Gal4-MLP fusion protein as thebait. The cardiac cDNA library was chosen for these studiesdue to the high MLP gene expression levels detected early inembryonic cardiac development and the maintenance of MLPexpression throughout adulthood (Arber et al., 1994, 1997).From a yeast screen of 6×106 independent transformants, oneparticular cDNA clone, independently isolated two times,generated colonies that exhibited a predicted His+ and LacZ+
phenotype. Sequence analysis of the partial cDNA revealedthat it encodes amino acids 890-1290 of βI-spectrin.Identification of β-spectrin as a binding partner for MLP isconsistent with previous data showing that β-spectrin is found

1556
within the actin-based cytoskeleton and that cytoplasmic MLPassociates with cytoplasmic f-actin filaments (Tilney andDetmers, 1975; Arber et al., 1996).
The region of β-spectrin that was isolated spans the C-terminal half of spectrin repeat 6 through the N-terminal halfof repeat 10 (termed β-spectrin 6.5-9.5). To confirm thespecificity of the MLP:β-spectrin interaction, Gal4-MLP andGal4-β-spectrin yeast plasmids were tested for in vivointeraction with a variety of different proteins. As predictedfrom the initial screen, co-expression of MLP and β-spectringenerates high β-galactosidase activity whereas no β-galactosidase is detected when MLP is tested with a variety ofother TAD-associated proteins, including MEF2A, paraxis andthe SV40 large T antigen (Table 1). Similarly, β-spectrin doesnot interact with the control proteins lamin C or p53 (Table 1).
MLP and β-spectrin are co-expressed in striatedmuscleIn order to establish if the isolated spectrin cDNA is derivedfrom the β-spectrin gene known to be expressed in striatedmuscle, northern blot analyses were performed on mRNAisolated from human adult tissues. As shown in Fig. 1, a singleMLP transcript of approximately 1.25 kb is detectedexclusively in cardiac and skeletal muscle tissues, which is inagreement with previously published reports for the MLPand CLP (the human homologue of MLP) genes (Arber et al.,1994; Fung et al., 1995). No MLP expression is detected innonmuscle tissues (Fig. 1) nor in any smooth musclederivatives (data not shown). Hybridization of the samenorthern blot with the β-spectrin cDNA obtained from theyeast two-hybrid screen yielded two transcripts ofapproximately 11 kb and 8.5 kb that are present in both cardiacand skeletal muscle tissues (Fig. 1). With extended exposuresa faint 11 kb transcript also is detected in brain mRNA (datanot shown). To confirm that the observed β-spectrin geneexpression pattern represents the muscle-specific spectrinisoform, a second β-spectrin cDNA fragment corresponding tothe unique portion of the striated muscle βΙΣΙΙ spectrin geneproduct (nucleotides 3183-3815 of the human βΙΣΙΙ cDNA)was used as a probe (Winkelmann et al., 1990). As predicted,the βΙΣΙΙ -specific probe also recognizes two transcripts of 11kb and 8.5 kb that are found only in cardiac and skeletal muscletissues (Fig. 1). These hybridized transcripts are identical inboth size and intensity to those observed with the spectrinprobe derived from the yeast two-hybrid screen and confirmthat the MLP and βΙ-spectrin genes are co-expressed in cardiacand skeletal muscle cells.
MLP:β-spectrin interactions are specific andmediated through their respective LIM2 and spectrinrepeat 7 domainsIn an effort to further characterize the MLP and β-spectrin
interaction, an in vitro protein binding assay was performed inwhich 35S-labeled proteins were incubated with His-taggedMLP immobilized on Ni2+-agarose beads. Protein complexesthen were thoroughly washed to clear free protein and thebound proteins were eluted, resolved by SDS-PAGE anddetected by autoradiography. As predicted from our previousstudies (Kong et al., 1997), MLP binds to the skeletal musclebHLH transcription factor MyoD, but does not interact withnonmuscle specific bHLH factors such as E47 or with thecontrol Ni2+-agarose beads (Fig. 2A). Similarly, MLP binds tothe β-spectrin (6.5-9.5) fragment, supporting the initialobservations obtained with the yeast two-hybrid analyses (Fig.2A). The MLP:β-spectrin interaction also is observed in co-immunoprecipitation assays. In this instance, an anti-spectrinmonoclonal antibody is capable of immunoprecipitating MLPin the presence of full-length β-spectrin protein but not whenthe β-spectrin protein is left out of the reaction (Fig. 2B).Conversely, the unrelated protein p48 (Krapp et al., 1996) isnot immunoprecipitated under these conditions. Thus, MLP:β-spectrin interactions can be detected through yeast two-hybrid,in vitro protein binding and co-immunoprecipitation assays.
Since members of the CRP family share a high degree ofidentity, we next examined if other LIM domain containingproteins also possess an ability to bind β-spectrin or whetherLIM proteins in general exhibit unique target bindingspecificities. For these studies, in vitro protein binding assayswere utilized in which GST-LIM domain fusion proteinsimmobilized on glutathione-agarose beads were incubated withthe 35S-labeled β-spectrin (6.5-9.5) fragment. As expected,GST-MLP interacts with the β-spectrin protein, but neithercontrol GST beads, the unrelated LIM-only LMO2 protein(Boehm et al., 1991), the P-LIM protein containing a LIMhomeodomain motif (Bach et al., 1995) nor CRP1 or CRP2bind to β-spectrin (Fig. 2C). Similarly, the nuclear protein
M. J. Flick and S. F. Konieczny
Fig. 1. The MLP and β-spectrin genes are co-expressed in cardiacand skeletal muscle. Human tissue RNA blots were probed with therat MLP cDNA, the β-spectrin (6.5-9.5) fragment obtained from theyeast two-hybrid screen and a βIΣII-specific fragment representingthe muscle-specific isoform of βI-spectrin. The expected 11 kb and8.5 kb β-spectrin and 1.25 kb MLP transcripts are detectedexclusively in the striated muscle samples.
Table 1. Yeast two-hybrid interactionsGal4 DB construct Gal4 TAD construct β-Galactosidase activity
MLP β-Spectrin (6.5-9.5) YesMLP MEF2A NoMLP Paraxis NoMLP SV40 large T antigen NoLamin C β-Spectrin (6.5-9.5) Nop53 β-Spectrin (6.5-9.5) No
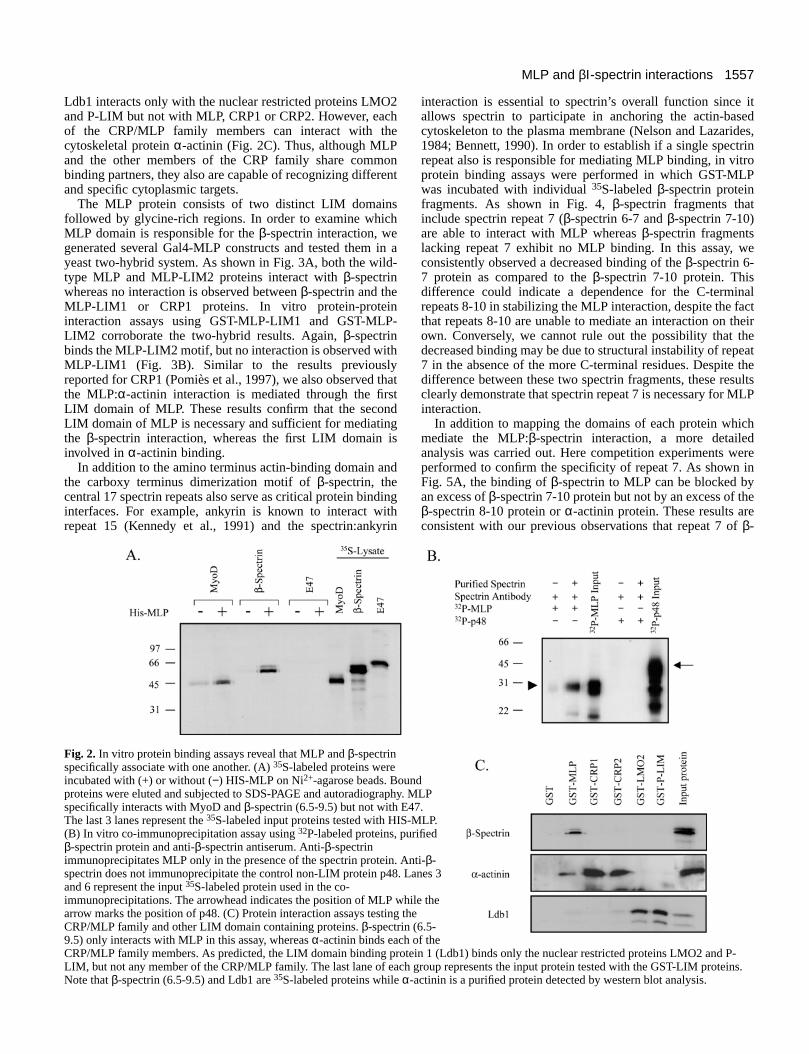
Ldb1 interacts only with the nuclear restricted proteins LMO2and P-LIM but not with MLP, CRP1 or CRP2. However, eachof the CRP/MLP family members can interact with thecytoskeletal protein α-actinin (Fig. 2C). Thus, although MLPand the other members of the CRP family share commonbinding partners, they also are capable of recognizing differentand specific cytoplasmic targets.
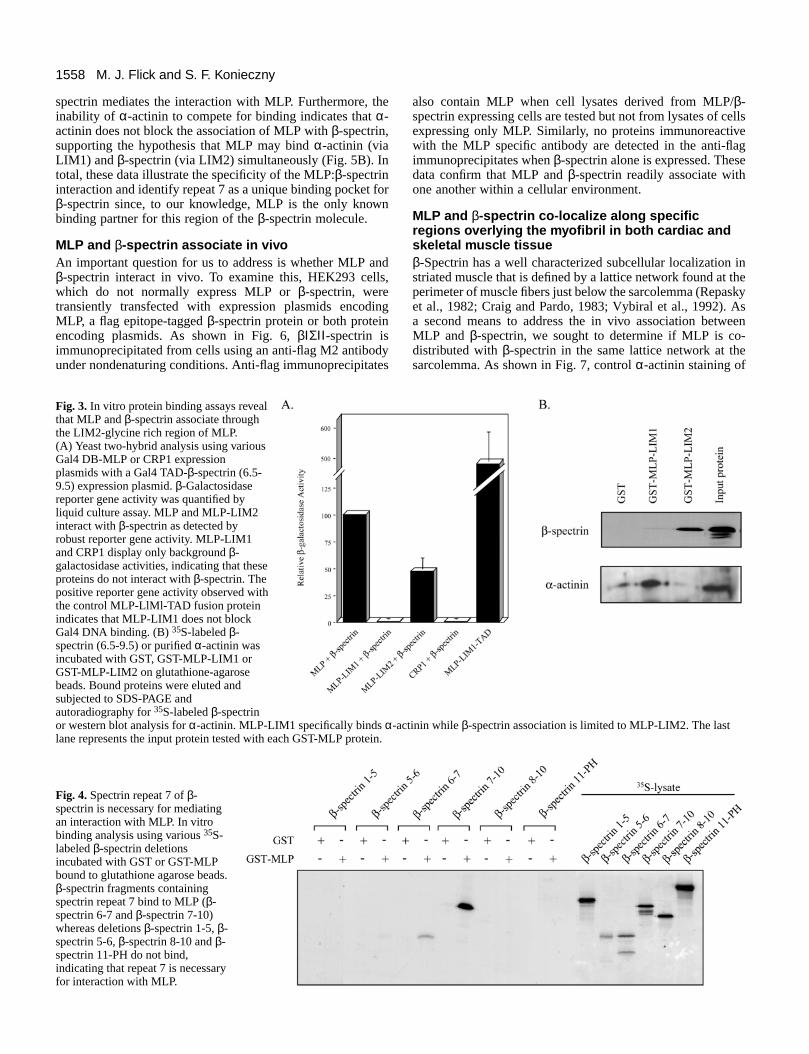
The MLP protein consists of two distinct LIM domainsfollowed by glycine-rich regions. In order to examine whichMLP domain is responsible for the β-spectrin interaction, wegenerated several Gal4-MLP constructs and tested them in ayeast two-hybrid system. As shown in Fig. 3A, both the wild-type MLP and MLP-LIM2 proteins interact with β-spectrinwhereas no interaction is observed between β-spectrin and theMLP-LIM1 or CRP1 proteins. In vitro protein-proteininteraction assays using GST-MLP-LIM1 and GST-MLP-LIM2 corroborate the two-hybrid results. Again, β-spectrinbinds the MLP-LIM2 motif, but no interaction is observed withMLP-LIM1 (Fig. 3B). Similar to the results previouslyreported for CRP1 (Pomiès et al., 1997), we also observed thatthe MLP:α-actinin interaction is mediated through the firstLIM domain of MLP. These results confirm that the secondLIM domain of MLP is necessary and sufficient for mediatingthe β-spectrin interaction, whereas the first LIM domain isinvolved in α-actinin binding.
In addition to the amino terminus actin-binding domain andthe carboxy terminus dimerization motif of β-spectrin, thecentral 17 spectrin repeats also serve as critical protein bindinginterfaces. For example, ankyrin is known to interact withrepeat 15 (Kennedy et al., 1991) and the spectrin:ankyrin
interaction is essential to spectrin’s overall function since itallows spectrin to participate in anchoring the actin-basedcytoskeleton to the plasma membrane (Nelson and Lazarides,1984; Bennett, 1990). In order to establish if a single spectrinrepeat also is responsible for mediating MLP binding, in vitroprotein binding assays were performed in which GST-MLPwas incubated with individual 35S-labeled β-spectrin proteinfragments. As shown in Fig. 4, β-spectrin fragments thatinclude spectrin repeat 7 (β-spectrin 6-7 and β-spectrin 7-10)are able to interact with MLP whereas β-spectrin fragmentslacking repeat 7 exhibit no MLP binding. In this assay, weconsistently observed a decreased binding of the β-spectrin 6-7 protein as compared to the β-spectrin 7-10 protein. Thisdifference could indicate a dependence for the C-terminalrepeats 8-10 in stabilizing the MLP interaction, despite the factthat repeats 8-10 are unable to mediate an interaction on theirown. Conversely, we cannot rule out the possibility that thedecreased binding may be due to structural instability of repeat7 in the absence of the more C-terminal residues. Despite thedifference between these two spectrin fragments, these resultsclearly demonstrate that spectrin repeat 7 is necessary for MLPinteraction.
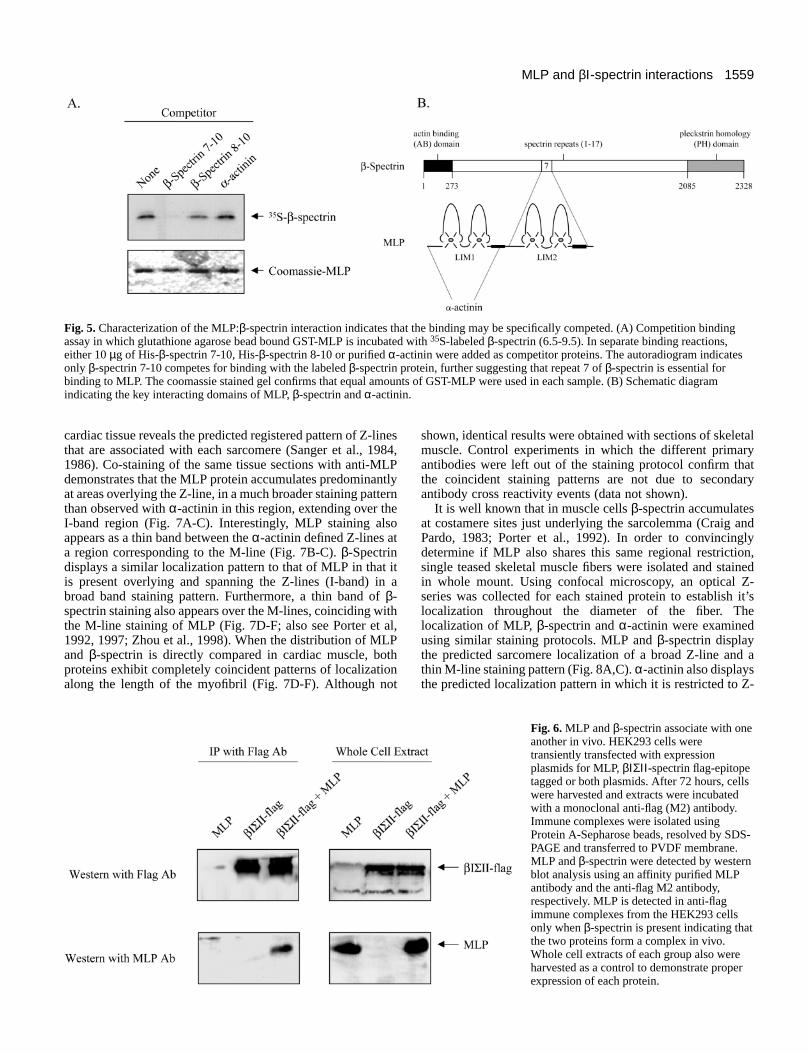
In addition to mapping the domains of each protein whichmediate the MLP:β-spectrin interaction, a more detailedanalysis was carried out. Here competition experiments wereperformed to confirm the specificity of repeat 7. As shown inFig. 5A, the binding of β-spectrin to MLP can be blocked byan excess of β-spectrin 7-10 protein but not by an excess of theβ-spectrin 8-10 protein or α-actinin protein. These results areconsistent with our previous observations that repeat 7 of β-
1557MLP and βΙ-spectrin interactions
Fig. 2. In vitro protein binding assays reveal that MLP and β-spectrinspecifically associate with one another. (A) 35S-labeled proteins wereincubated with (+) or without (−) HIS-MLP on Ni2+-agarose beads. Boundproteins were eluted and subjected to SDS-PAGE and autoradiography. MLPspecifically interacts with MyoD and β-spectrin (6.5-9.5) but not with E47.The last 3 lanes represent the 35S-labeled input proteins tested with HIS-MLP.(B) In vitro co-immunoprecipitation assay using 32P-labeled proteins, purifiedβ-spectrin protein and anti-β-spectrin antiserum. Anti-β-spectrinimmunoprecipitates MLP only in the presence of the spectrin protein. Anti-β-spectrin does not immunoprecipitate the control non-LIM protein p48. Lanes 3and 6 represent the input 35S-labeled protein used in the co-immunoprecipitations. The arrowhead indicates the position of MLP while thearrow marks the position of p48. (C) Protein interaction assays testing theCRP/MLP family and other LIM domain containing proteins. β-spectrin (6.5-9.5) only interacts with MLP in this assay, whereas α-actinin binds each of theCRP/MLP family members. As predicted, the LIM domain binding protein 1 (Ldb1) binds only the nuclear restricted proteins LMO2 and P-LIM, but not any member of the CRP/MLP family. The last lane of each group represents the input protein tested with the GST-LIM proteins.Note that β-spectrin (6.5-9.5) and Ldb1 are 35S-labeled proteins while α-actinin is a purified protein detected by western blot analysis.
1558
spectrin mediates the interaction with MLP. Furthermore, theinability of α-actinin to compete for binding indicates that α-actinin does not block the association of MLP with β-spectrin,supporting the hypothesis that MLP may bind α-actinin (viaLIM1) and β-spectrin (via LIM2) simultaneously (Fig. 5B). Intotal, these data illustrate the specificity of the MLP:β-spectrininteraction and identify repeat 7 as a unique binding pocket forβ-spectrin since, to our knowledge, MLP is the only knownbinding partner for this region of the β-spectrin molecule.
MLP and β-spectrin associate in vivoAn important question for us to address is whether MLP andβ-spectrin interact in vivo. To examine this, HEK293 cells,which do not normally express MLP or β-spectrin, weretransiently transfected with expression plasmids encodingMLP, a flag epitope-tagged β-spectrin protein or both proteinencoding plasmids. As shown in Fig. 6, βΙΣΙΙ -spectrin isimmunoprecipitated from cells using an anti-flag M2 antibodyunder nondenaturing conditions. Anti-flag immunoprecipitates
also contain MLP when cell lysates derived from MLP/β-spectrin expressing cells are tested but not from lysates of cellsexpressing only MLP. Similarly, no proteins immunoreactivewith the MLP specific antibody are detected in the anti-flagimmunoprecipitates when β-spectrin alone is expressed. Thesedata confirm that MLP and β-spectrin readily associate withone another within a cellular environment.
MLP and β-spectrin co-localize along specificregions overlying the myofibril in both cardiac andskeletal muscle tissueβ-Spectrin has a well characterized subcellular localization instriated muscle that is defined by a lattice network found at theperimeter of muscle fibers just below the sarcolemma (Repaskyet al., 1982; Craig and Pardo, 1983; Vybiral et al., 1992). Asa second means to address the in vivo association betweenMLP and β-spectrin, we sought to determine if MLP is co-distributed with β-spectrin in the same lattice network at thesarcolemma. As shown in Fig. 7, control α-actinin staining of
M. J. Flick and S. F. Konieczny
Fig. 4. Spectrin repeat 7 of β-spectrin is necessary for mediatingan interaction with MLP. In vitrobinding analysis using various 35S-labeled β-spectrin deletionsincubated with GST or GST-MLPbound to glutathione agarose beads.β-spectrin fragments containingspectrin repeat 7 bind to MLP (β-spectrin 6-7 and β-spectrin 7-10)whereas deletions β-spectrin 1-5, β-spectrin 5-6, β-spectrin 8-10 and β-spectrin 11-PH do not bind,indicating that repeat 7 is necessaryfor interaction with MLP.
Fig. 3. In vitro protein binding assays revealthat MLP and β-spectrin associate throughthe LIM2-glycine rich region of MLP.(A) Yeast two-hybrid analysis using variousGal4 DB-MLP or CRP1 expressionplasmids with a Gal4 TAD-β-spectrin (6.5-9.5) expression plasmid. β-Galactosidasereporter gene activity was quantified byliquid culture assay. MLP and MLP-LIM2interact with β-spectrin as detected byrobust reporter gene activity. MLP-LIM1and CRP1 display only background β-galactosidase activities, indicating that theseproteins do not interact with β-spectrin. Thepositive reporter gene activity observed withthe control MLP-LlMl-TAD fusion proteinindicates that MLP-LIM1 does not blockGal4 DNA binding. (B) 35S-labeled β-spectrin (6.5-9.5) or purified α-actinin wasincubated with GST, GST-MLP-LIM1 orGST-MLP-LIM2 on glutathione-agarosebeads. Bound proteins were eluted andsubjected to SDS-PAGE andautoradiography for 35S-labeled β-spectrinor western blot analysis for α-actinin. MLP-LIM1 specifically binds α-actinin while β-spectrin association is limited to MLP-LIM2. The lastlane represents the input protein tested with each GST-MLP protein.
1559MLP and βΙ-spectrin interactions
cardiac tissue reveals the predicted registered pattern of Z-linesthat are associated with each sarcomere (Sanger et al., 1984,1986). Co-staining of the same tissue sections with anti-MLPdemonstrates that the MLP protein accumulates predominantlyat areas overlying the Z-line, in a much broader staining patternthan observed with α-actinin in this region, extending over theI-band region (Fig. 7A-C). Interestingly, MLP staining alsoappears as a thin band between the α-actinin defined Z-lines ata region corresponding to the M-line (Fig. 7B-C). β-Spectrindisplays a similar localization pattern to that of MLP in that itis present overlying and spanning the Z-lines (I-band) in abroad band staining pattern. Furthermore, a thin band of β-spectrin staining also appears over the M-lines, coinciding withthe M-line staining of MLP (Fig. 7D-F; also see Porter et al,1992, 1997; Zhou et al., 1998). When the distribution of MLPand β-spectrin is directly compared in cardiac muscle, bothproteins exhibit completely coincident patterns of localizationalong the length of the myofibril (Fig. 7D-F). Although not
shown, identical results were obtained with sections of skeletalmuscle. Control experiments in which the different primaryantibodies were left out of the staining protocol confirm thatthe coincident staining patterns are not due to secondaryantibody cross reactivity events (data not shown).
It is well known that in muscle cells β-spectrin accumulatesat costamere sites just underlying the sarcolemma (Craig andPardo, 1983; Porter et al., 1992). In order to convincinglydetermine if MLP also shares this same regional restriction,single teased skeletal muscle fibers were isolated and stainedin whole mount. Using confocal microscopy, an optical Z-series was collected for each stained protein to establish it’slocalization throughout the diameter of the fiber. Thelocalization of MLP, β-spectrin and α-actinin were examinedusing similar staining protocols. MLP and β-spectrin displaythe predicted sarcomere localization of a broad Z-line and athin M-line staining pattern (Fig. 8A,C). α-actinin also displaysthe predicted localization pattern in which it is restricted to Z-
Fig. 5. Characterization of the MLP:β-spectrin interaction indicates that the binding may be specifically competed. (A) Competition bindingassay in which glutathione agarose bead bound GST-MLP is incubated with 35S-labeled β-spectrin (6.5-9.5). In separate binding reactions,either 10 µg of His-β-spectrin 7-10, His-β-spectrin 8-10 or purified α-actinin were added as competitor proteins. The autoradiogram indicatesonly β-spectrin 7-10 competes for binding with the labeled β-spectrin protein, further suggesting that repeat 7 of β-spectrin is essential forbinding to MLP. The coomassie stained gel confirms that equal amounts of GST-MLP were used in each sample. (B) Schematic diagramindicating the key interacting domains of MLP, β-spectrin and α-actinin.
Fig. 6. MLP and β-spectrin associate with oneanother in vivo. HEK293 cells weretransiently transfected with expressionplasmids for MLP, βΙΣΙΙ -spectrin flag-epitopetagged or both plasmids. After 72 hours, cellswere harvested and extracts were incubatedwith a monoclonal anti-flag (M2) antibody.Immune complexes were isolated usingProtein A-Sepharose beads, resolved by SDS-PAGE and transferred to PVDF membrane.MLP and β-spectrin were detected by westernblot analysis using an affinity purified MLPantibody and the anti-flag M2 antibody,respectively. MLP is detected in anti-flagimmune complexes from the HEK293 cellsonly when β-spectrin is present indicating thatthe two proteins form a complex in vivo.Whole cell extracts of each group also wereharvested as a control to demonstrate properexpression of each protein.
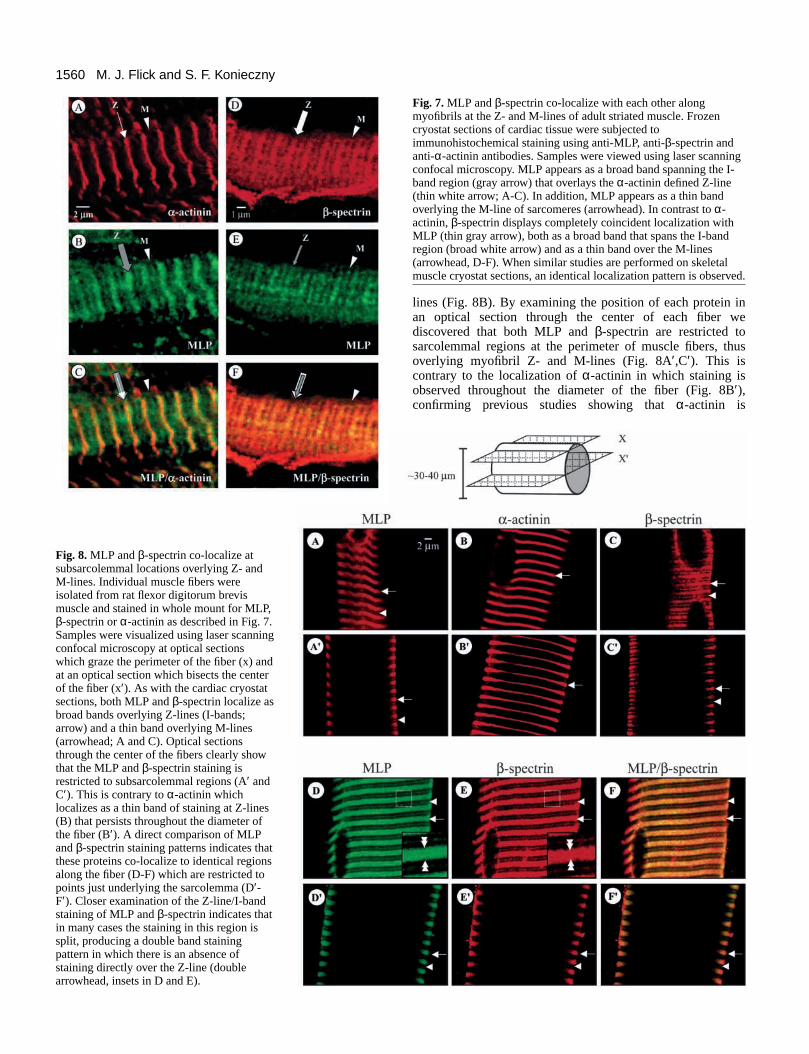
1560
lines (Fig. 8B). By examining the position of each protein inan optical section through the center of each fiber wediscovered that both MLP and β-spectrin are restricted tosarcolemmal regions at the perimeter of muscle fibers, thusoverlying myofibril Z- and M-lines (Fig. 8A′,C′). This iscontrary to the localization of α-actinin in which staining isobserved throughout the diameter of the fiber (Fig. 8B′),confirming previous studies showing that α-actinin is
M. J. Flick and S. F. Konieczny
Fig. 7. MLP and β-spectrin co-localize with each other alongmyofibrils at the Z- and M-lines of adult striated muscle. Frozencryostat sections of cardiac tissue were subjected toimmunohistochemical staining using anti-MLP, anti-β-spectrin andanti-α-actinin antibodies. Samples were viewed using laser scanningconfocal microscopy. MLP appears as a broad band spanning the I-band region (gray arrow) that overlays the α-actinin defined Z-line(thin white arrow; A-C). In addition, MLP appears as a thin bandoverlying the M-line of sarcomeres (arrowhead). In contrast to α-actinin, β-spectrin displays completely coincident localization withMLP (thin gray arrow), both as a broad band that spans the I-bandregion (broad white arrow) and as a thin band over the M-lines(arrowhead, D-F). When similar studies are performed on skeletalmuscle cryostat sections, an identical localization pattern is observed.
Fig. 8. MLP and β-spectrin co-localize atsubsarcolemmal locations overlying Z- andM-lines. Individual muscle fibers wereisolated from rat flexor digitorum brevismuscle and stained in whole mount for MLP,β-spectrin or α-actinin as described in Fig. 7.Samples were visualized using laser scanningconfocal microscopy at optical sectionswhich graze the perimeter of the fiber (x) andat an optical section which bisects the centerof the fiber (x′). As with the cardiac cryostatsections, both MLP and β-spectrin localize asbroad bands overlying Z-lines (I-bands;arrow) and a thin band overlying M-lines(arrowhead; A and C). Optical sectionsthrough the center of the fibers clearly showthat the MLP and β-spectrin staining isrestricted to subsarcolemmal regions (A′ andC′). This is contrary to α-actinin whichlocalizes as a thin band of staining at Z-lines(B) that persists throughout the diameter ofthe fiber (B′). A direct comparison of MLPand β-spectrin staining patterns indicates thatthese proteins co-localize to identical regionsalong the fiber (D-F) which are restricted topoints just underlying the sarcolemma (D′-F′). Closer examination of the Z-line/I-bandstaining of MLP and β-spectrin indicates thatin many cases the staining in this region issplit, producing a double band stainingpattern in which there is an absence ofstaining directly over the Z-line (doublearrowhead, insets in D and E).
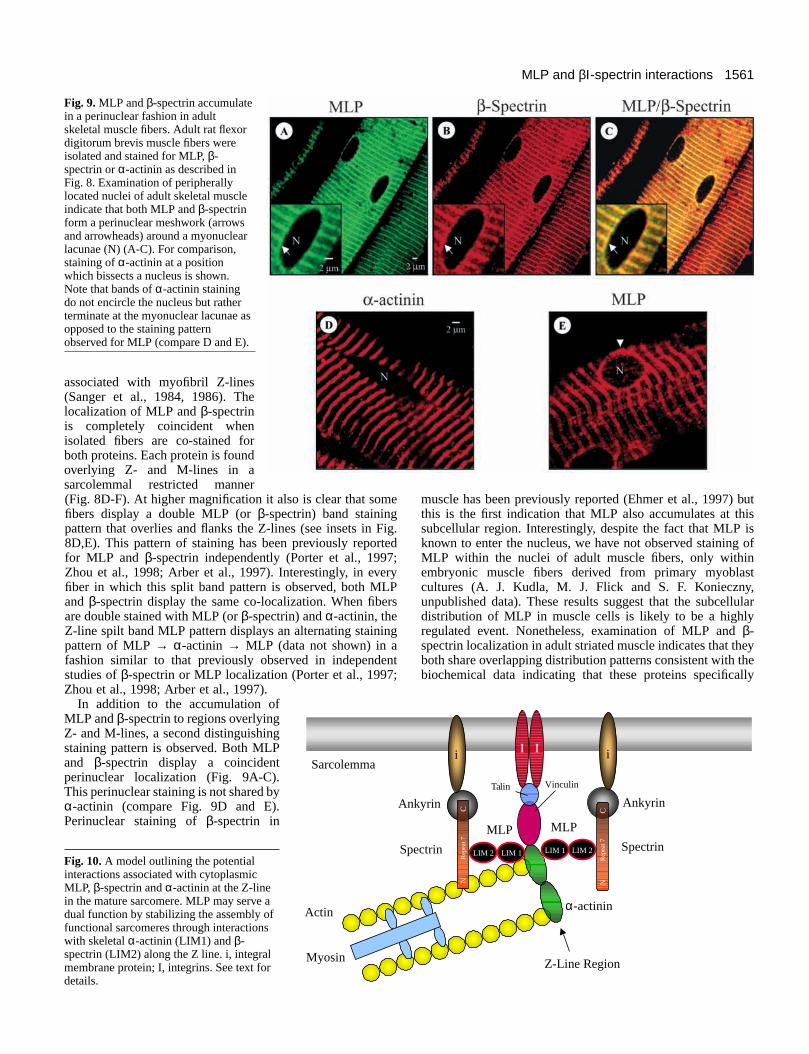
1561MLP and βΙ-spectrin interactions
associated with myofibril Z-lines(Sanger et al., 1984, 1986). Thelocalization of MLP and β-spectrinis completely coincident whenisolated fibers are co-stained forboth proteins. Each protein is foundoverlying Z- and M-lines in asarcolemmal restricted manner(Fig. 8D-F). At higher magnification it also is clear that somefibers display a double MLP (or β-spectrin) band stainingpattern that overlies and flanks the Z-lines (see insets in Fig.8D,E). This pattern of staining has been previously reportedfor MLP and β-spectrin independently (Porter et al., 1997;Zhou et al., 1998; Arber et al., 1997). Interestingly, in everyfiber in which this split band pattern is observed, both MLPand β-spectrin display the same co-localization. When fibersare double stained with MLP (or β-spectrin) and α-actinin, theZ-line spilt band MLP pattern displays an alternating stainingpattern of MLP → α-actinin → MLP (data not shown) in afashion similar to that previously observed in independentstudies of β-spectrin or MLP localization (Porter et al., 1997;Zhou et al., 1998; Arber et al., 1997).
In addition to the accumulation ofMLP and β-spectrin to regions overlyingZ- and M-lines, a second distinguishingstaining pattern is observed. Both MLPand β-spectrin display a coincidentperinuclear localization (Fig. 9A-C).This perinuclear staining is not shared byα-actinin (compare Fig. 9D and E).Perinuclear staining of β-spectrin in
muscle has been previously reported (Ehmer et al., 1997) butthis is the first indication that MLP also accumulates at thissubcellular region. Interestingly, despite the fact that MLP isknown to enter the nucleus, we have not observed staining ofMLP within the nuclei of adult muscle fibers, only withinembryonic muscle fibers derived from primary myoblastcultures (A. J. Kudla, M. J. Flick and S. F. Konieczny,unpublished data). These results suggest that the subcellulardistribution of MLP in muscle cells is likely to be a highlyregulated event. Nonetheless, examination of MLP and β-spectrin localization in adult striated muscle indicates that theyboth share overlapping distribution patterns consistent with thebiochemical data indicating that these proteins specifically
Fig. 9. MLP and β-spectrin accumulatein a perinuclear fashion in adultskeletal muscle fibers. Adult rat flexordigitorum brevis muscle fibers wereisolated and stained for MLP, β-spectrin or α-actinin as described inFig. 8. Examination of peripherallylocated nuclei of adult skeletal muscleindicate that both MLP and β-spectrinform a perinuclear meshwork (arrowsand arrowheads) around a myonuclearlacunae (N) (A-C). For comparison,staining of α-actinin at a positionwhich bissects a nucleus is shown.Note that bands of α-actinin stainingdo not encircle the nucleus but ratherterminate at the myonuclear lacunae asopposed to the staining patternobserved for MLP (compare D and E).
Talin Vinculin
Actin
Myosin
Sarcolemma
α-actinin
LIM 1LIM 2Spectrin
MLP
LIM 1 LIM 2 Spectrin
MLP
Z-Line Region
i I iI
Ankyrin Ankyrin
NC
Rep
eat 7
NC
Rep
eat 7
Fig. 10. A model outlining the potentialinteractions associated with cytoplasmicMLP, β-spectrin and α-actinin at the Z-linein the mature sarcomere. MLP may serve adual function by stabilizing the assembly offunctional sarcomeres through interactionswith skeletal α-actinin (LIM1) and β-spectrin (LIM2) along the Z line. i, integralmembrane protein; I, integrins. See text fordetails.

1562
form complexes with one another. The fact that both proteinsare restricted to the sarcolemma, coupled with the previousidentification of β-spectrin as a key costamere protein, suggeststhat MLP also serves a vital structural role at this location.
DISCUSSION
Previous studies have demonstrated that MLP is critical to theprocess of muscle differentiation at a variety of levels,including organization and maintenance of the contractileapparatus (Arber et al., 1994, 1997; Kong et al., 1997).Therefore, identification of β-spectrin, a key cytoskeletalprotein in striated muscle, as a protein target for MLP isconsistent with the available limited data regarding MLP’smechanistic roles. Through a variety of in vitro and in vivoassay systems, including yeast two-hybrid, in vitro proteinbinding and in vivo co-immunoprecipitation, we havedemonstrated that the MLP:β-spectrin interaction is direct.Furthermore, protein binding assays have revealed that theinteraction is specific and can be competed only by specificligands. Molecular dissection of the MLP:β-spectrininteraction indicates that it is mediated through the LIM2 motifof MLP and the spectrin repeat 7 of β-spectrin. Although β-spectrin has binding sites for many known partners (reviewedby Hartwig, 1995), identification of repeat 7 as the MLPinteraction domain reveals a novel binding interface for the β-spectrin molecule.
Identification of β-spectrin as a MLP LIM2-specific targetprotein represents the first protein partner defined for the C-terminal LIM motif of any CRP/MLP family member. We wereparticularly interested in determining if, like the α-actinin:LIM1 interaction, this property is shared between CRPfamily members or between LIM domain containing proteinsin general. Our results demonstrate that β-spectrin is a partnerunique to the LIM2 domain associated with MLP since theLIM-only proteins CRP1, CRP2, LMO2 and the LIM-homeodomain protein P-LIM do not bind β-spectrin. Althoughother proteins have been identified (i.e. α-actinin, zyxin) thatbind all three CRP family members, these proteins areexpressed in each of the cell types in which the CRP/MLPfamily members are found (Crawford et al., 1992; Sadler et al.,1992). This is in contrast to β-spectrin in which only MLP isco-expressed in striated muscle cells. Similarly, the bHLHMyoD family of skeletal muscle-specific proteins have beenfound to interact with MLP, but again CRP1 fails to bind thesemuscle-specific factors (Kong et al., 1997). The unique featureof the MLP target proteins (βΙ-spectrin, MyoD) is that theyexhibit a tissue specificity that is similar to MLP. Neither βΙ-spectrin nor the MyoD family of proteins is expressed in celltypes which express CRP1 or CRP2 (Winkelmann et al., 1990;Ludolph and Konieczny, 1995; Louis et al., 1997). These datasuggest that although the CRP/MLP family members exhibitcommon binding partners, they also utilize unique partners thatare found only in specific cell types.
The biochemical data defining the MLP:β-spectrininteraction is strongly supported by immunohistochemicalstudies examining the distribution of MLP and β-spectrin instriated muscle. We have found that the cytoplasmicsubcellular localization of MLP is completely coincident withthe β-spectrin lattice along the myofibril in adult striated
muscle. In both cardiac and skeletal muscle, MLP and β-spectrin are observed as broad bands overlying Z-lines and thinbands overlying M-lines. As clearly demonstrated in skeletalfibers, MLP localization, like that of β-spectrin, is limited tothe sarcolemma. The co-distribution overlying the Z-lineregion of myofibrils is often disrupted, producing a pattern justflanking the α-actinin defined Z-lines, therefore extending overthe I-band. This staining pattern resembles that of a host ofother cytoskeletal proteins, including dystrophin and vinculin,whose localization is restricted to the sarcolemma (Craig andPardo, 1983; Pardo et al., 1983; Shear and Bloch, 1985; Porteret al., 1992; Straub et al., 1992). This subsarcolemmallocalization that overlies the I-band and flanks the Z-lines isknown as the costamere (Craig and Pardo, 1983; Pardo et al.,1983; Porter et al., 1992). Costameres are believed to serve ascritical links between the sarcolemma and the actin basedcontractile apparatus as sites of force transmission (Street,1983; Danowski et al., 1992). Based on its subsarcolemmallocalization and association with β-spectrin, we propose thatMLP serves, in part, as a costamere protein (Fig. 10).
The role of β-spectrin as a costamere protein may be definedby the multiple contacts it makes with the sarcolemma andassociated proteins as well as with the actin based cytoskeleton(reviewed by Berthier and Blaineau, 1997). Interestingly, thepredominant isoform of β-spectrin that is found in muscle(βΙΣΙΙ) incorporates a C-terminal plekstrin homology (PH)domain (Winkelmann et al., 1990; Porter et al., 1990, 1997;Zhou et al., 1998). PH domains bind phosphotidylinositols and,for β-spectrin, this domain is believed to be essential forstabilizing the association with the sarcolemma (Lombardo etal., 1994; Wang and Shaw, 1995). It would appear that β-spectrin serves as a direct link between the sarcolemma and thecontractile apparatus. Since MLP binds the actin crosslinkingprotein α-actinin through its LIM1 domain (Fig. 3B; also seePomiès et al., 1997) and β-spectrin through the LIM2 domain,we propose that costameric MLP functions as a scaffoldingfactor that links the α-actinin crosslinked actin filaments of themyofibril to the β-spectrin network (Fig. 10).
The scaffold protein model for MLP is strongly supportedby the phenotype of MLP deficient mice in which a highlydisorganized cytoskeletal architecture develops (Arber et al.,1997). MLP null mice are viable up through postnatal day oneat which time an increased work demand is placed on theanimal, particularly on the heart, and the integrity of themyofibril is no longer maintained (Arber et al., 1997).Although the myofibril arrangement is disrupted, the structureof the sarcomere remains intact, suggesting that the majordefect in MLP null mice involves a lack of costamere function(Arber et al., 1997). Furthermore, mutations or loss of otherknown costamere proteins result in quite similar phenotypicabnormalities. Perturbations of the subsarcolemmal proteindystrophin, which also associates with β-spectrin, haveprofound effects on muscle cytoarchitectural integrity (Porteret al., 1992; reviewed by Emery, 1990, 1993). Interestingly,loss of dystrophin function in cardiac muscle results in thesame myopathy, dilated cardiomyopathy, as does loss of MLP,suggesting that these two proteins may serve complementaryroles in maintaining muscle integrity (reviewed by Chen andChien, 1999; Chien, 1999). Others have shown that in mdxmice, which lack dystrophin, the organization of manycostamere and costamere associated proteins is disrupted
M. J. Flick and S. F. Konieczny

1563MLP and βΙ-spectrin interactions
(Williams and Bloch, 1999). These results again demonstratehow the loss of a single protein can have profound effects onthe overall arrangement of costamere members. Preliminaryanalysis of β-spectrin in MLP null muscle indicates a similarloss of costemere organization (unpublished results). Currentstudies are aimed at examining if this effect extends to othermembrane associated proteins. Linking membrane and actinassociated proteins may indeed be the physiological role of theCRP/MLP family. The smooth muscle protein CRP1 is foundassociated with the adhesion plaque protein zyxin (Sadler etal., 1992). Coupled with its ability to bind α-actinin, it has beensuggested that CRP1 may also participate in organizing andmaintaining protein complexes at sites of cell-substratumadhesion (reviewed by Beckerle et al., 1997). If this modelproves correct it will be completely analogous to the MLP/β-spectrin model proposed here.
A final consideration of the β-spectrin:MLP interaction isthe possible relationship between the β-spectrin:MLP complexand the role of MLP as a nuclear protein, which in skeletalmuscle is capable of affecting MyoD-mediated transcriptionalactivity (Kong et al., 1997). The mechanism of MLP nuclearlocalization, as well as the factors dictating its subcellularshuttling, are unknown. In skeletal muscle, β-spectrin forms ameshwork surrounding individual nuclei (Ehmer et al., 1997).In examining adult muscle, we observed perinuclear β-spectrinstaining that was accompanied by a similar MLP stainingpattern. In these adult tissues, true nuclear MLP localizationwas not detected. However, the lack of nuclear staining in adultmuscle preparations is not surprising since during late stagesof differentiation MLP is exclusively cytoplasmic (Arber et al.,1994). Nonetheless, there are clear functional distinctions thatcan be made between the regulatory pathways through whicha nuclear vs cytoplasmic MLP participates. It also is formallypossible that cytoplasmic and nuclear MLP share commonregulatory functions. For example, the cytoplasmic protein β-catenin has been found to bind the transcription factor LEF-1and play a role in transporting LEF-1 to the nucleus in responseto specific intracellular signals (Behrens et al., 1996). It isconceivable that early in striated muscle differentiation,nuclear vs cytoplasmic MLP localization may be dependent onthe downstream organization of the spectrin lattice.Alternatively, perinuclear localized MLP and β-spectrin mayactually represent the association of these proteins with othermembrane-based organelles. Erythroid and nonerythroidhomologs of β-spectrin have been observed associated with theGolgi membrane and other cytoplasmic vesicles in nonmusclecell types (Beck et al., 1994; Devarajan et al., 1997;Stankewich et al., 1998). Golgi spectrin is hypothesized tostabilize the Golgi membrane and play a role in the traffickingof a subset of proteins from the endoplasmic reticulum to theGolgi (Devarajan et al., 1997). MLP also may participate withspectrin in performing similar functions in muscle, but thisremains to be investigated. Current studies aimed atdetermining if additional factors participate in MLP-mediatedcytoskeletal organization and/or nuclear localization and atfurther elucidating how MLP:β-spectrin interactions play a rolein other key cellular processes.
We thank Jon Morrow for providing the βΙΣΙΙ -spectrin plasmid andfor the VD4 mouse monoclonal β-spectrin antibody, Mary Beckerleand Gordon Gill for providing CRP1, CRP2 and LMO2 expression
plasmids and Simon Rhodes for providing the P-LIM expressionplasmid. We also acknowledge the important technical contributionsof Thai Tran, Christopher Pin, Arthur Kudla and David Asai as wellas thank Christopher Pin and Arthur Kudla for providing criticalcomments on the manuscript. We particularly thank Don Ready forhis patience and unlimited assistance with confocal microscopy andfor his interpretation of the captured images. This work was supportedby grants to S.F.K. from the National Institutes of Health and theMuscular Dystrophy Association. M.J.F. was supported by a PurdueUniversity Andrew’s Fellowship and a NIH training grant.
REFERENCES
Arber, S., Halder, G. and Caroni, P. (1994). Muscle LIM protein, a novelessential regulator of myogenesis, promotes myogenic differentiation. Cell79, 221-231.
Arber, S. and Caroni, P. (1996). Specificity of single LIM motifs in targetingand LIM/LIM interactions in situ. Genes Dev. 10, 289-300.
Arber, S., Hunter, J. J., Ross, J. Jr, Hongo, M., Sansig, G., Borg, J.,Perriard, J.-C., Chien, K. R. and Caroni, P. (1997). MLP-deficient miceexhibit a disruption of cardiac cytoarchitectural organization, dilatedcardiomyopathy, and heart failure. Cell 88, 393-493.
Bach, I., Rhodes, S. J., Pearse, R. V. II, Heinzel, T., Gloss, B., Scully, K.M., Sawchenko, P. E. and Rosenfeld, M. G. (1995). P-Lim, a LIMhomeodomain factor, is expressed during pituitary organ and cellcommitment and synergizes with Pit-1. Proc. Nat. Acad. Sci. USA 92, 2720-2724.
Beck, K. A., Buchanan, J. A., Malhotra, V. and Nelson, W. J. (1994). Golgispectrin: identification of an erythroid β-spectrin homolog associated withthe Golgi complex. J. Cell Biol. 127, 707-723.
Beckerle, M. C. (1997). Zyxin: zinc fingers at sites of cell adhesion. BioEssays19, 949-957.
Behrens, J., von-Kries, J. P., Kuhl, M., Bruhn, L., Wedlich, D., Grosschedl,R. and Birchmeier, W. (1996). Functional interaction of β-catenin with thetranscription factor LEF-1. Nature 382, 638-642.
Bennett, V. (1990). Spectrin-based membrane skeleton: a multipotentialadapter between plasma membrane and cytoplasm. Physiol. Rev. 70, 1029-1065.
Berthier, C. and Blaineau, S. (1997). Supramolecular organization of thesubsarcolemma cytoskeleton of adult skeletal muscle fibers. A review. Biol.Cell 89, 413-434.
Boehm, T., Foroni, L., Kaneko, Y., Perutz, M. F. and Rabbitts, T. H. (1991).The rhombotin family of cystein-rich LIM-domain oncogenes: Distinctmembers are involved in T-cell translocations to human chromosomes 11p15and 11p13. Proc. Nat. Acad. Sci. USA 88, 4367-4371.
Chen, J. and Chien, K. R. (1999). Complexity and simplicity: monogenicdisorders and complex cardiomyopathies. J. Clin. Invest. 103, 1483-1485.
Chien, K. R. (1999). Stress pathways and heart failure. Cell 98, 555-558. Craig, S. W. and Pardo, J. V. (1983). Gamma actin, spectrin, and intermediate
filament proteins colocalize with vinculin at costameres, myofibril-to-sarcolemma attachment sites. Cell Motil. 3, 449-462.
Crawford, A. W., Michelsen, J. W. and Beckerle, M. C. (1992). Aninteraction between zyxin and α-actinin. J. Cell Biol. 116, 1381-1393.
Danowski, B. A., Imanaka-Yoshia, K., Sanger, J. M. and Sanger, J. W.(1992). Costameres are sites of force transmission to the substratum in adultrat cardiomyoctes. J. Cell Biol. 118, 1411-1420.
Dawid, I. B., Breen, J. J. and Toyama, R. (1998). LIM domains: multipleroles as adapters and functional modifiers in protein interactions. TrendsGenet. 14, 156-162.
Devarajan, P., Stabach, P. R., DeMatteis, M. A. and Morrow, J. S. (1997).Na, K-ATPase transport from endoplasmic reticulum to Golgi requires theGolgi spectrin-ankyrin G119 skeleton in Madin Darby canine kidney cells.Proc. Nat. Acad. Sci. USA 94, 10711-10716.
Ehmer, S., Herrmann, R., Bittner, R. and Voit, T. (1997). Spatial distributionof β-spectrin in normal and dystrophic human skeletal muscle. ActaNeuropathol. 94, 240-246.
Emery, A. E. H. (1990). Dystrophin function. Lancet (N. Am. edn). 335, 1289. Emery, A. E. H. (1993). Duchenne Muscular Dystrophy. Oxford University
Press, Oxford, UK. 392 pp. Fung, Y. W., Wang, R. X., Heng, H. H. Q. and Liew, C. C. (1995). Mapping
of a human LIM protein (CLP) to human chromosome 11p15.1 byfluorescence in situ hybridization. Genomics 28, 602-603.

1564
Hartwig, J. H. (1995). Actin-binding proteins. 1:spectrin super family. ProteinProfile. 2, 703-800.
Jurata, L. W., Kenny, D. A. and Gill, G. N. (1996). Nuclear LIM interactor,a rhombotin and LIM homeodomain interacting protein, is expressed earlyin neuronal development. Proc. Nat. Acad. Sci. USA 93, 11693-11698.
Kennedy, S., Warren, B., Forget, B. G. and Morrow, J. S. (1991). Ankyrinbinds to the 15th repetitive unit of erythroid and nonerythroid β-spectrin. J.Cell Biol. 115, 267-277.
Kong, Y., Flick, M., Kudla, A. J. and Konieczny, S. F. (1997). Muscle LIMprotein promotes myogenesis by enhancing the activity of MyoD. Mol. Cell.Biol. 17, 4750-4760.
Krapp, A., Knöfler, M., Frutiger, S., Hughes, G. J., Hagenbüchle, O. andWellauer, P. K. (1996). The p48 DNA-binding subunit of transcriptionfactor PTF1 is a new exocrine pancreas-specific basic helix-loop-helixprotein. EMBO J. 15, 4317-4329.
Lombardo, C. R., Weed, S. A., Kennedy, S. P., Forget, B. G. and Morrow,J. S. (1994). Beta II-spectrin and beta I epsilon 2-spectrin (muscle) containNH2- and COOH-terminal membrane association domains (MAD1 andMAD2). J. Biol. Chem. 269, 29212-29219.
Louis, H. A., Pino, D., Schmeichel, K. L., Pomiés, P. and Beckerle, M. C.(1997). Comparison of three members of the cystine-rich protein familyreveals functional conservation and divergent patterns of gene expression.J. Biol. Chem. 272, 27484-27491.
Ludolph, D. C. and Konieczny, S. F. (1995). Transcription factor families:muscling in on the myogenic program. FASEB J. 9, 1595-1604.
Michelsen, J. W., Schmeichel, K. L., Beckerle, M. C. and Winge, D. R.(1993). The LIM motif defines a specific zinc binding protein domain. Proc.Nat. Acad. Sci. USA 90, 4404-4408.
Miller, J. H. (1972). Experiments in Molecular Genetics. Cold Spring HarborLaboratory Press, Cold Spring Harbor, NY.
Nelson, W. J. and Lazarides, E. (1984). Goblin (ankyrin) in striated muscle:identification of the potential membrane receptor for erythorid spectrin inmuscle cells. Proc. Nat. Acad. Sci. USA 81, 3292-3296.
Pardo, J. V., Siciliano, J. D. and Craig, S. W. (1983). A vinculin containingcortical lattice in skeletal muscle: transverse lattice elements (‘costameres’)mark sites of attachment between myofibrils and sarcolemma. Proc. Nat.Acad. Sci. USA 80, 1008-1012.
Pomiès, P., Louis, H. A. and Beckerle, M. C. (1997). CRP1, a LIM domainprotein implicated in muscle differentiation, interacts with α-actinin. J. CellBiol. 139, 157-168.
Porter, G. A., Krikorian, J. C., Winkelmann, J. G., Morrow, J. S. andBloch, R. J. (1990). Multiple isoforms of β-spectrin in adult and neonatalrat skeletal muscle. J. Cell Biol. 111, 428a.
Porter, G. A., Dmytrenko, G. M., Winkelmann, J. C. and Bloch, R. J.(1992). Dystrophin colocalizes with β-spectrin in distinct subsarcolemmaldomains in mammalian skeletal muscle. J. Cell Biol. 117, 997-1005.
Porter, G. A., Resneck, W. G., Schner, M. G., Porter, N. C., Fowler, V. A.and Bloch, R. J. (1997). Two populations of β-spectrin in mammalianskeletal muscle. Cell. Motil. Cytoskel. 37, 7-19.
Repasky, E. A., Granger, B. L. and Lazarides, E. (1982). Widespreadoccurrence of avian spectrin in nonerythroid cells. Cell 29, 821-833.
Sadler, I., Crawford, A. W., Michelsen, J. W. and Beckerle, M. C. (1992).
Zyxin and cCRP: two interactive LIM domain proteins associated with thecytoskeleton. J. Cell Biol. 119, 1573-1587.
Sánchez-Garciá, I. and Rabbitts, T. H. (1994). The LIM domain: a newstructural motif found in zinc-finger-like proteins. Trends Genet. 10, 315-320.
Sanger, J. M., Mittal, B., Pochapin, M. B. and Sanger, J. W. (1986).Myofibrillogenesis in living cells microinjected with fluorescently labeledalpha-actinin. J. Cell Biol. 102, 2053-2066.
Sanger, J. W., Mittal, B. and Sanger, J. M. (1984). Formation of myofibrilsin spreading chick cardiac myocytes. Cell Motil. 4, 405-416.
Schmeichel, K. L. and Beckerle, M. C. (1994). The LIM domain is a modularprotein-binding interface. Cell 79, 211-219.
Shear, C. R. and Bloch, R. J. (1985). Vinculin in subsarcolemmal densitiesin chicken skeletal muscle: localization and relationship to intracellular andextracellular structures. J. Cell Biol. 113, 1133-1144.
Stankewich, M. C., Tse, W. T., Peters, L. L., Ch’ng, Y., John, K. M.,Stabach, P. R., Devarajan, P., Morrow, J. S. and Lux, S. E. (1998). Awidely expressed βIII spectrin associated with Golgi and cytoplasmicvesicles. Proc. Nat. Acad. Sci. USA 95, 14158-14163.
Straub, V., Bittner, R. E., Leger, J. J. and Voit, T. (1992). Direct visualizationof the dystrophin network on skeletal muscle fiber membrane. J. Cell Biol.119, 1183-1191.
Street, S. F. (1983). Lateral transmission of tension in frog myofibers:myofibrillar network and transverse cytoskeletal connections are possibletransmitters. J. Cell. Physiol. 114, 346-364.
Stronach, B. E., Renfranz, P. J., Lilly, B. and Beckerle, M. C. (1999).Muscle LIM proteins are associated with muscle sarcomeres and requiredMEF2 for their expression during Drosophila myogenesis. Mol. Biol. Cell10, 2329-2342.
Tilney, L. and Detmers, P. (1975). Actin in erythrocyte ghosts and itsassociation with spectrin. J. Cell Biol. 66, 508-520.
Vybiral, T., Winkelmann, J. C., Roberts, R., Joe, E.-H., Casey, D. L.,Williams, J. K. and Epstein, H. F. (1992). Human cardiac and skeletalmuscle spectrins: differential expression and localization. Cell Motil.Cytoskel. 21, 293-304.
Wang, D. S. and Shaw, G. (1995). The association of the c-terminal regionof beta I sigma II spectrin to brain membranes is mediated by a PH domain,does not require membrane proteins, and coincides with a inositol-1,4,5triphosphate binding site. Biochem. Biophys. Res. Commun. 94, 608-615.
Weed, S. A., Stabach, P. R., Oyer, C. E., Gallagher, P. G. and Morrow, J.S. (1996). The lethal hemolytic mutation in βΙΣΙΙ spectrin providence yieldsa null phenotype in neonatal skeletal muscle. Lab. Invest. 74, 1117-1129.
Weiskirchen, R., Pino, J. D., Macalma, T., Bister, K. and Beckerle, M. C.(1995). The cysteine-rich protein family of highly related LIM domainproteins. J. Biol. Chem. 270, 28946-28954.
Williams, M. W. and Bloch, R. J. (1999). Extensive but coordinatedreorganization of the membrane skeleton in myofibers of dystrophic (mdx)mice. J. Cell Biol. 144, 1259-1270.
Winkelmann, J. C., Costa, F. F., Linzie, B. L. and Forget, B. G. (1990). βspectrin in human skeletal muscle. J. Biol. Chem. 265, 20449-20454.
Zhou, D., Ursitti, J. A. and Bloch, R. J. (1998). Developmental expressionof spectrins in rat skeletal muscle. Mol. Biol. Cell 9, 47-61.
M. J. Flick and S. F. Konieczny