Embed Size (px)
Citation preview

The occurrence and ecology of a marine hydrobiidmudsnail in the southern hemisphere: the KnysnaEstuary, South Africa
R. S. K. BarnesDepartment of Zoology, University of Cambridge, Cambridge CB23EJ, UK
Abstract
Earlier published records of the gastropod Hydrobia fromthewarmtemperate Knysna Lagoononthe Indian Oceancoastof SouthAfricaare inerror.Nevertheless, ananimalhitherto identi¢ed as an Assiminea (Assimineidae) or asa Tomichia (Pomatiopsidae) is in fact a Hydrobia s.l., andit does occur intertidally within the Knysna system.The same species also occurs in salt pans in the cooltemperate Great Berg Estuary on the Atlantic coast ofSouth Africa. This extends the distribution of con¢rmedspecies of the dominant coastal hydrobiid mudsnailsof the northern hemisphere into the southern hemi-sphere. H. knysnaensis (Krauss) occurs patchily at lowdensity near the head of the Knysna Estuary in watersof low salinity. This appears to be a suboptimal habitat,and it is suggested that Hydrobia is restricted to thiszone as a result of interference competition from pota-midid mudwhelks and ocypodid crabs. The prevalenceof potamidids and ocypodids in the tropics and in thetemperate southern hemisphere may account for therareness or absence of hydrobiids in these areas. Unlikeits equally intertidal relative, the widespread NorthAtlantic/Arctic H. ulvae, H. knysnaensis develops directlylike other Hydrobia.
Key words: competition, intertidal, lagoon, Ocypodidae,Potamididae, salinity
Re¤ sume¤
Les publications faisant e¤ tat de la pre¤ sence du gaste¤ ro-pode Hydrobia dans le lagon tempe¤ re¤ chaud de Knysna,
sur la co“ te sud-africaine de l’oce¤ an Indien sont errone¤ es.Ne¤ anmoins, un animal identi¢e¤ jusqu’ici comme unAssiminea (Assimineidae) ou comme unTomichia (Poma-tiopsidae), est en fait un Hydrobia s.l. et il appara|‡t surl’estran dans le syste' me de Knysna. La me“ me espe' ce serencontre aussi dans des cuvettes sauma“ tres dansl’Estuaire de Great Berg, tempe¤ re¤ frais, sur la co“ te atlan-tique d’Afrique du Sud. Ceci e¤ largit vers l’he¤ misphe' resud la distribution d’une espe' ce con¢rme¤ e des mollus-ques co“ tiers hydrobiinae dominants de l’he¤ misphe' renord. H. knysnaensis (Krauss) se trouve ca' et la' , en faiblequantite¤ , au fond de l’estuaire de Knysna, dans des eauxde faible salinite¤ . Ceci semble e“ tre un habitat sub-opti-mal, et on sugge' re qu’Hydrobia est con¢ne¤ dans cettezone suite a' la compe¤ tition avec les buccins potamilidaeet avec les crabes ocypodidae. La pre¤ valence de ces der-niers sous les tropiques et dans la partie tempe¤ re¤ e del’he¤ misphe' re sud pourrait expliquer la rarete¤ oul’absence d’hydrobiidae dans ces re¤ gions. Contrairementa' son parent H. ulvae, qui vit comme lui sur l’estran, tre' sre¤ pandu dans l’Atlantique nord et dans l’Arctique, H.knysnaensis se de¤ veloppe directement, comme les autresHydrobia.
Introduction
The known species of coastal mudsnails (the rissoaceansubfamily Hydrobiinae or Hydrobia s.l.) are con¢ned tothe northern hemisphere, extending from the Atlanticcoast of North America to the Caspian Sea and from theArcticOceanto the southernshores of theMediterranean(Wilke, Rola¤ n & Davis, 2000). There they are abundant,widespread and often key members of intertidal andlagoonal soft sediments, in some areas completely
# 2002 East AfricanWild Life Society, Afr. J. Ecol., 40, 289^294 289
Correspondence e-mail: [email protected]

dominating benthic biomass and energy transfer (e.g.Asmus & Asmus,1985).
Nevertheless, the literature does contain occasionalrecords of ‘Hydrobia sp.’ from other temperate regions,although most of these seem tobe based onmisidenti¢ca-tions of pomatiopsid or other small gastropods with mor-phologically extremely similar, if not e¡ectivelyidentical, shells (T. Wilke, pers. com.). In South Africa,for example, such records hail from the cool temperateGreat Berg Estuary on the west coast (328460S; 188090E)(Kalejta & Hockey,1991) round to the19 km long,18 km2
warm temperate Knysna Lagoon on the south coast(348040S; 238030E) (Day,1981a).
Indeed, Day (1981a) in his ‘summary of current knowl-edge’ of the Knysna Lagoon states that Hydrobia sp. isone of the three abundant gastropods in that system,and Day’s sentence to that e¡ect is repeated verbatim byGrindley (1985) in his ‘synopsis of available information’on Knysna. The primary literature on the benthic faunaof Knysna (Day,Millard&Harrison,1952;Day,1967;Allan-son, Nettleton & DeVilliers, 2000), however, contains norecords of any hydrobiids, and neither do Day’s otherworks on South African estuaries and lagoons (Day,1969, 1981b), other than to repeat a record from Lange-baan Lagoon (Puttick,1977).
In1999, however, some specimens of a putative hydro-biid were obtained by the author and Dr V. E. McArthurnear the upper reaches of the Knysna Estuary (althoughnot in the Lagoon). These were sent to Dr T.Wilke of theDepartment of Malacology, Academyof Natural Sciences,Philadelphia, USA and he (pers. com.) (a) identi¢ed thematerial as Paludina knysnaensis Krauss, (b) con¢rmed itas belonging to the Hydrobiinae although it had pre-viously been assigned to eitherAssiminea (Assimineidae)orTomichia (Pomatiopsidae), and (c) on the basis of DNAsequence data suggested that amongst known species,it is closest phylogenetically to the common northernAtlantic species Hydrobia (Peringia) ulvae (Pennant). Thisis as yet the onlyknownHydrobia s.l. species in the south-ern hemisphere.
The 1999 survey data found the species in low abun-dance in a very limited section of the estuary and thisreport describes a subsequent investigation carried outinto the distribution, abundance, breeding strategy andgeneral ecology of H. knysnaensis, with special referenceto the Knysna system and to why hydrobiids might be sodominant in the northern hemisphere but seemingly soinsigni¢cant in the southern.
Methods
Intertidal mud£ats at Knysna were sampled in April andMay 2001 by means of a 0.076-m-diameter core tubeand/or a 0.01-m2 area,2-cm-deep scoop; subtidal regionswere sampled (in 1999) solely using the corer. Sampleswere passed through a1-mm-mesh diameter sieve. Mea-surement of egestion rate was carried out essentially asin Barnes (2001), and other methodology was asdescribed below.The Great Berg Estuary was also visitedin March 2001 to ascertain the nature of the ‘Hydrobiasp.’ reported from there.
Results
Habitat, distribution and abundance
The Knysna system can be divided into three zones(Largier, Attwood & Harcourt-Baldwin, 2000): the outer,middle and inner basins, of which the outer and middleare commonly referred to as ‘Knysna Lagoon’ whilst theinner is often termed ‘the estuary’ (Allanson, 2000)(Fig.1). In spite of intensive search, noHydrobiawas foundin either of the lagoonal basins, only in the inner basinor ‘estuary’.
Fig1 Sketch map of the Knysna estuarine system, showing thedistribution of H. knysnaensis. The outer and middle basins areoften collectively termed the Knysna Lagoon
290 R. S. K. Barnes
# 2002 East AfricanWild Life Society, Afr. J. Ecol., 40, 289^294
The shore in that region comprises a high level Juncusmarsh dominated by the gastropod Cerithidea decollata(Linnaeus) and the crab Sesarma catenata Ortmann, alow level Zostera meadow and/or beds of the burrowingprawns Upogebia or Callianassa, and between thema nar-row (often <2 m across), relatively steeply sloping bankof sandy mud with novegetation. It was only this narrowzone that supported any Hydrobia; none was found sub-tidally or in pools on the salt-marsh surface (cf. below).Mean overall abundance, based on ten transects downthe inhabited shores of the inner basin, was 202 m�2
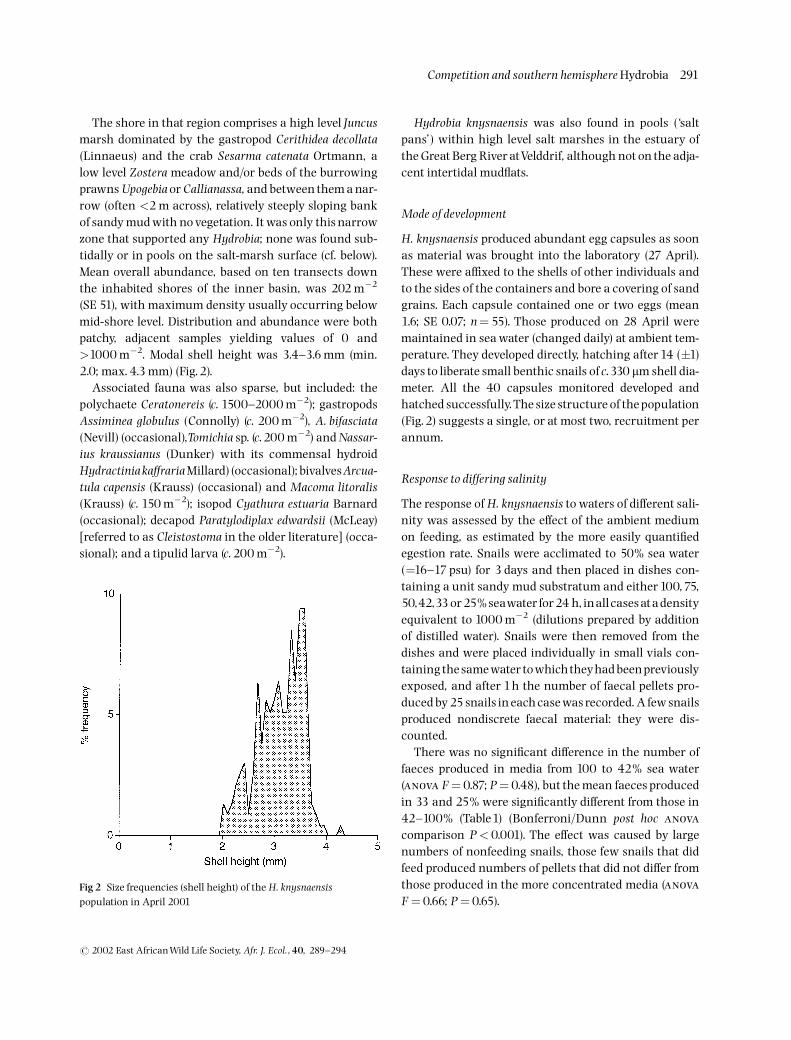
(SE 51), with maximum density usually occurring belowmid-shore level. Distribution and abundance were bothpatchy, adjacent samples yielding values of 0 and>1000 m�2. Modal shell height was 3.4^3.6 mm (min.2.0; max. 4.3 mm) (Fig.2).
Associated fauna was also sparse, but included: thepolychaete Ceratonereis (c. 1500^2000 m�2); gastropodsAssiminea globulus (Connolly) (c. 200 m�2), A. bifasciata(Nevill) (occasional),Tomichia sp. (c.200 m�2) andNassar-ius kraussianus (Dunker) with its commensal hydroidHydractiniaka¡rariaMillard) (occasional); bivalvesArcua-tula capensis (Krauss) (occasional) and Macoma litoralis(Krauss) (c. 150 m�2); isopod Cyathura estuaria Barnard(occasional); decapod Paratylodiplax edwardsii (McLeay)[referred to as Cleistostoma in the older literature] (occa-sional); and a tipulid larva (c. 200 m�2).
Hydrobia knysnaensis was also found in pools (‘saltpans’) within high level salt marshes in the estuary ofthe Great Berg River atVelddrif, although not on the adja-cent intertidal mud£ats.
Mode of development
H. knysnaensis produced abundant egg capsules as soonas material was brought into the laboratory (27 April).These were a⁄xed to the shells of other individuals andto the sides of the containers and bore a covering of sandgrains. Each capsule contained one or two eggs (mean1.6; SE 0.07; n¼55). Those produced on 28 April weremaintained in sea water (changed daily) at ambient tem-perature. They developed directly, hatching after 14 (�1)days to liberate small benthic snails of c.330 mm shell dia-meter. All the 40 capsules monitored developed andhatched successfully.The size structure of thepopulation(Fig.2) suggests a single, or at most two, recruitment perannum.
Response to differing salinity
The response of H. knysnaensis to waters of di¡erent sali-nity was assessed by the e¡ect of the ambient mediumon feeding, as estimated by the more easily quanti¢edegestion rate. Snails were acclimated to 50% sea water(¼16^17 psu) for 3 days and then placed in dishes con-taining a unit sandy mud substratum and either100,75,50,42,33or25%seawater for24 h, inall casesatadensityequivalent to 1000 m�2 (dilutions prepared by additionof distilled water). Snails were then removed from thedishes and were placed individually in small vials con-taining the samewater towhichtheyhadbeenpreviouslyexposed, and after 1h the number of faecal pellets pro-duced by25 snails in eachcasewas recorded.A few snailsproduced nondiscrete faecal material: they were dis-counted.
There was no signi¢cant di¡erence in the number offaeces produced in media from 100 to 42% sea water(anova F¼0.87; P¼0.48), but the mean faeces producedin 33 and 25% were signi¢cantly di¡erent from those in42^100% (Table1) (Bonferroni/Dunn post hoc anova
comparison P<0.001). The e¡ect was caused by largenumbers of nonfeeding snails, those few snails that didfeed produced numbers of pellets that did not di¡er fromthose produced in the more concentrated media (anovaF¼0.66; P¼0.65).
Fig 2 Size frequencies (shell height) of the H. knysnaensispopulation in April 2001
# 2002 East AfricanWild Life Society, Afr. J. Ecol., 40, 289^294
Competition and southern hemisphere Hydrobia 291
Intraspecific effects
The e¡ect of populationdensityonH.knysnaensis feedingwas assessed in a similar manner to that of salinity, allexperiments being conducted in 100% sea water. Densi-ties equivalent to 350^7000 m�2 were tested (seeTable 2). Faecal productiondid not varysigni¢cantlywithpopulationdensityover the range350^3500 m�2 (anovaF¼1.14; P¼0.34) but egestion rate did decrease signi¢-cantly at the highest density (7000 m�2) (Bonferroni/Dunn post hoc anova comparison P<0.002).
Interspecific effects
The e¡ect of three sympatric organisms on faecal produc-tion of unit density of H. knysnaensis (850 m�2) in seawater was tested. These were (a) juveniles (12^15 mmshell height) of the potamidid snail Cerithidea decollatathat unlike the adults do not ascend vegetation duringlow tide but remain on the sediment surface (Cockroft &Forbes, 1981), (b) the small ocypodid crab Paratylodiplaxedwardsii, and (c) themudsnailA. bifasciata (Table 3).Den-sities of 170 and 350 m�2 of both C. decollata andP. edwardsii had a marked e¡ect, feeding being drasticallyreduced or stopping (Cerithidea: 170 m�2, anova F¼33,P<0.0001;350.m�2,anova F¼67,P<0.0001.Paratylo-diplax: 170 m�2, anova F¼55, P<0.0001; 350 m�2
anova F¼73, P<0.0001). Densities of 350 m�2 Assimi-nea had no e¡ect (anova F¼0.35, P¼0.56).
Discussion
Like the dominant Hydrobia of the northern NorthAtlan-tic (Barnes, 1999), H. knysnaensis can withstand full-strength sea water, and the in£uence of salinity on itsfeeding rate suggests that media less concentrated than50%seawaterare likely tobe suboptimal. Its developmentwas also100% successful in sea water.Yet in the Knysnasystem, it appears con¢ned to the low salinity regionsnear the head of the estuary, where salinities averagesome 9^22 psu and may fall as low as 4 or 5 psu (Dayet al., 1952; Allanson, 2000). The salt pan habitat of H.knysnaensis in the Great Berg is also a fringe marine sys-tem.
In the tropics generally, and inwarm temperate south-ernAfrica,mud£atsaredominated inter aliabypotamididmudwhelks and ocypodid crabs (e.g. Macnae,1968), andKnysna is no exception. At Knysna,Paratylodiplax occurs
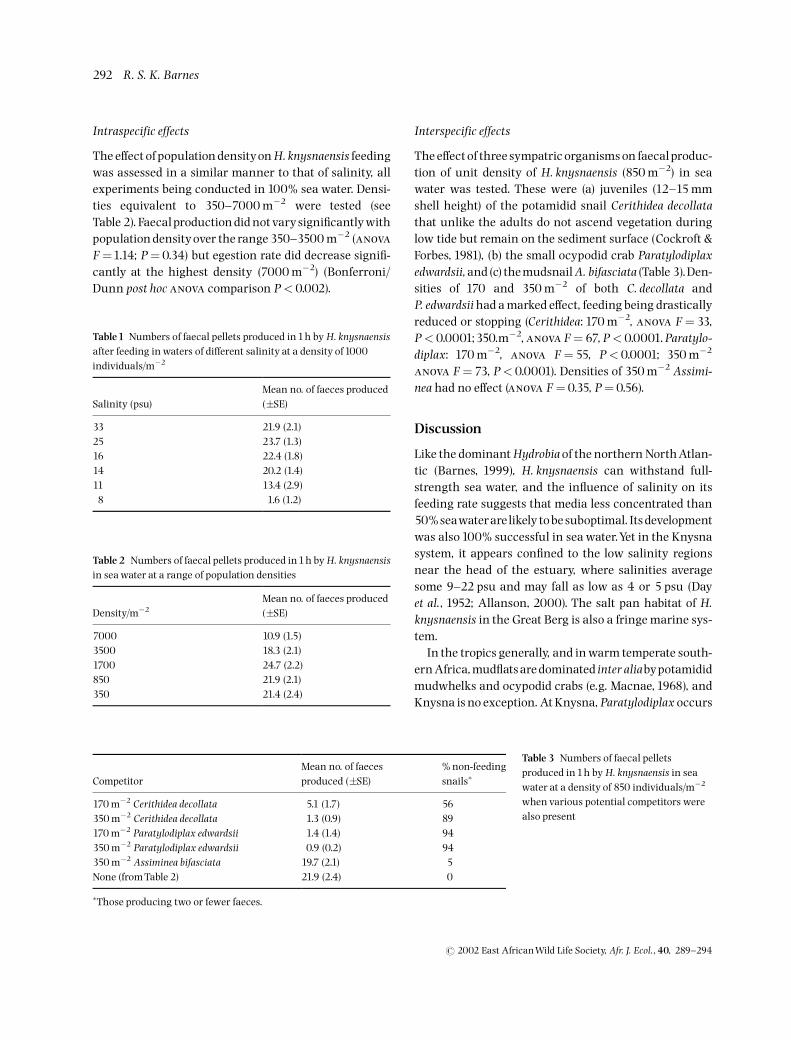
Table1 Numbers of faecal pellets produced in1h by H. knysnaensisafter feeding in waters of different salinity at a density of1000individuals/m�2
Salinity (psu)Mean no. of faeces produced(�SE)
33 21.9 (2.1)25 23.7 (1.3)16 22.4 (1.8)14 20.2 (1.4)11 13.4 (2.9)8 1.6 (1.2)
Table 2 Numbers of faecal pellets produced in1h by H. knysnaensisin sea water at a range of population densities
Density/m�2Mean no. of faeces produced(�SE)
7000 10.9 (1.5)3500 18.3 (2.1)1700 24.7 (2.2)850 21.9 (2.1)350 21.4 (2.4)
CompetitorMean no. of faecesproduced (�SE)
% non-feedingsnails�
170 m�2 Cerithidea decollata 5.1 (1.7) 56350 m�2 Cerithidea decollata 1.3 (0.9) 89170 m�2 Paratylodiplax edwardsii 1.4 (1.4) 94350 m�2 Paratylodiplax edwardsii 0.9 (0.2) 94350 m�2 Assiminea bifasciata 19.7 (2.1) 5None (fromTable 2) 21.9 (2.4) 0
�Those producing two or fewer faeces.
Table 3 Numbers of faecal pelletsproduced in1h by H. knysnaensis in seawater at a density of 850 individuals/m�2
when various potential competitors werealso present
# 2002 East AfricanWild Life Society, Afr. J. Ecol., 40, 289^294
292 R. S. K. Barnes

from thehighwater neap tide level downtoand below thelow water of springs (Day, 1981b), in densities in SouthAfricaof up to 600 m�2 (deVilliers & Hodgson,1999).Cer-ithidea, for which Knysna is the southern-most limit,occurs above high water neap, in densities during thissurvey of up to 200 m�2. Both these species at ¢eld den-sitycaused complete oralmost complete cessationof feed-ing in H. knysnaensis, almost certainly as a result ofinterference competition of the type demonstrated inHydrobia by Levinton (1979) and Levinton, Stewart &Dewitt (1985). Ocypodid crabs and potamididmudwhelksare not recorded from the Great Berg (Kalejta & Hockey,1991). Indeed, in no other reported locality doesHydrobiaoccur sympatrically with a potamidid or ocypodid andthis study provides evidence of why this might be so. H.knysnaensis seems to occur in the Knysna system in theonly place where these potential competitors are absent.In South Africa,Hydrobia is largely replaced byAssiminea(Branch & Branch, 1981): competitive interactionsbetween Assiminea and Cerithidea/Paratylodiplax couldtherefore repay investigation.
Although the intertidal marine/estuarineH.ulvaemaybe the North Atlantic species closest phylogenetically toH. knysnaensis, the developmental mode, egg capsules,lack of tendency to climb above the water level, body sizeand shell shape of the Knysna species are all unlike thesubgenus Peringia (Barnes, 1988) and are more closelyakin to species such as H. glyca (Servain) in the subgenusHydrobia (Barnes,1996; as ‘H. neglecta’). In any event, lar-viparity is clearly notauniversal feature of intertidal spe-cies of Hydrobia s.l.
The question of why the species had not apparentlybeen found by earlier surveys of Knysna remains ^ is itperhaps a recent introduction? The fact that its nearestrelative is H. ulvae from which it departs so signi¢cantlyin behaviour and ecology suggests that it cannot be arecent arrival. The stretch of the estuary in which itoccurs had been surveyed by Day et al. (1952) and byAllanson et al. (2000). Although Day et al. (1952) recor-ded a single ‘Assiminea sp.’ throughout the entire Knysnasystem, this situation had been re¢ned by the time ofDay’s (1967) report into the occurrence of an ‘Assiminea’in the Lagoon and an‘?Assiminea’ in the estuary or innerbasin. In Allanson et al. (2000), Day’s ‘Assiminea’ isrecorded as ‘A. globulus’and is shown to occur through-out the whole system especially at high tidal levels,whilst Day’s ‘?Assiminea’ is listed as ‘Assiminea sp.’ con-¢ned to mid tidal levels in the inner basin. The locality,
position on the shore and habitat of Day’s ‘?Assiminea’and Allanson et al.’s ‘Assiminea sp.’, are therefore, identicalto that of H. knysnaensis, and no unknown species ofAssiminea also occurs at the site. The various namescan all almost certainly safely be assumed to belong tothe one animal, especially because it was for a long timemistakenly regarded as an Assiminea. The abun-dance of the species reported by Day (1967) (<6 m�2)and Allanson et al. (2000) (up to 32 m�2) are lower thanthose reportedhere, but thismaysimply re£ect thepatchydistribution and general scarcity of H. knysnaensis. Itwould not appear from the results above that its lowpopulation density is a consequence of intraspeci¢ccompetition, although interspeci¢c competition fromthe occasional sympatric Paratylodiplax must be a strongpossibility.
Whatever they are, Day’s (1981a) Hydrobia that hereported as being one of the three most common gastro-pods in the Lagoon, and his (1969) ‘Assiminea cf. ponson-byi’ that he described as ‘abundant in Knysna Lagoon onsandy mud at mid-tide’ are not H. knysnaensis. To whatthese names refer remaina mystery, although this surveydid record aTomichia sp. in the Lagoon that would ¢tDay’s(1969) description.
In the Great Berg Estuary, Kalejta & Hockey (1991)recorded‘Hydrobia sp.’as comprising84.5% of themud£atinvertebrates at their site B (with a mean density of over7,5500 m�2), but did not ¢nd Assiminea, whereas the visitreported here in March 2001 failed to ¢nd Hydrobia onthe mud£ats, although Assiminea was present.Whetheror not the H. knysnaensis of the Berg salt marshes is thesame animal as Kalejta & Hockey’s ‘Hydrobia sp.’ is there-fore, alsouncertain.But inanyevent, the rangeofH.knys-naensis does extend from the Great Berg Estuary in thenorth and west to ^ and probably beyond ^ Knysna inthe south and east, and the‘Hydrobia sp.’recorded byPut-tick (1977) from the Langebaan Lagoon and by Koop,Batty & McQuaid (1983), de Decker & Batty (1985) andBennett & Branch (1990) from the closed Bot River Estu-ary may well be of the same animal.
Acknowledgements
I am most grateful to Brian Allanson for allowing me touse the facilities of the Thesen & Co. Field Laboratory,Thesen’s Island, Knysna and for much other help, toCarol Barnes for copious invaluable ¢eld and laboratoryassistance, and to the British Ecological Society, the
# 2002 East AfricanWild Life Society, Afr. J. Ecol., 40, 289^294
Competition and southern hemisphere Hydrobia 293

University of Cambridge, and St. Catharine’s College for¢nancial support. The visit to the site in1999 was underthe aegis of the Department of Zoology, University of theWestern Cape, through the Royal Society of London(UK)/Foundation for Research Development (RSA) JointSupport Initiative.
References
Allanson, B.R. (2000) An introduction to the chemistry of thewater column of the Knysna River Estuary with particularreference to nutrients and suspended solids.Trans. roy. Soc. SouthAfrica 55,141^162.
Allanson, B.R., Nettleton, J. & DeVilliers, C. (2000)Benthic macrofaunal richness and diversity in the Knysnaestuary: a 50 year comparison.Trans. roy. Soc. South Africa 55,177^186.
Asmus, H. & Asmus, R. (1985) The importance of grazing food chainfor energy flow and production in three intertidal sand bottomcommunities of the northernWadden Sea. Helgol. Meeresunters.39, 273^301.
Barnes, R.S.K. (1988) On reproductive strategies in adjacentlagoonal and intertidal-marine populations of the gastropodHydrobia ulvae. J. Mar. Biol. Assoc. UK 68,365^375.
Barnes, R.S.K. (1996) Breeding, recruitment and survival in amixed intertidal population of the mudsnails Hydrobia ulvae andH. neglecta. J. Mar. Biol. Assoc. UK 76,1003^1012.
Barnes, R.S.K. (1999) What determines the distribution of coastalhydrobiid mudsnails within north-western Europe? Mar. Ecol.(Naples) 20,97^110.
Barnes, R.S.K. (2001) Interference competition in the intertidalmud snail Hydrobia ulvae: egestion rates revisited. J. Mar. Biol.Assoc. UK 81, 491^495.
Bennett, B.A. & Branch, G.M. (1990) Relationships betweenproduction and consumption of prey species by resident fish inthe Bot, a cool temperate South African estuary. Estuar. Coast.Shelf Sci.31,139^175.
Branch, G. & Branch, M. (1981) The Living Shores of Southern Africa.Struik, CapeTown.
Cockroft,V.G. & Forbes, A.T. (1981) Tidal activity rhythms in themangrove snail Cerithidea decollata (Linn.). South Afr. J. Zool.16,5^9.
Day, J.H. (1967) The biology of Knysna Estuary, South Africa. In:Estuaries (Ed. G. H. Lauff). AAAS,Washington, DC.
Day, J.H. (1969) AGuide to Marine Life on South African Shores.Balkema, CapeTown.
Day, J.H. (1981a) Summaries of current knowledge of 43 estuaries insouthern Africa. In: Estuarine Ecology with Particular Reference toSouthern Africa (Ed. J. H. Day). Struik, CapeTown.
Day, J.H. (1981b) The estuarine fauna. In: Estuarine Ecology withParticular Reference to Southern Africa (Ed. J. H. Day). Struik, CapeTown.
Day, J.H., Millard, N.A.H. & Harrison, A.D. (1952) The ecology ofSouth African estuaries. Part III. Knysna: a clear open estuary.Trans. royal Soc. South Afr. 33,367^413.
de Decker, H.P. & Batty, R. (1985) The benthic macrofauna of theBot River Estuary, South Africa, with a note on its meiofauna.Trans. royal Soc. South Afr.45,379^396.
Grindley, J.R. (1985) Estuaries of the Cape. Report no.30: Knysna.National Research Institute for Oceanology, Stellenbosch.
Kalejta, B. & Hockey, P.A.R. (1991) Distribution, abundance andproductivity of benthic invertebrates at the Berg River estuary,South Africa. Estuar. Coast. Shelf Sci.33,175^191.
Koop, K., Batty, R. & McQuaid, C.M. (1983) The ecology of SouthAfrican estuaries. Part XII. the Bot River, a closed estuary in thesouth-western Cape. South Afr. J. Zool.18,1^10.
Largier, J.L., Attwood, C. & Harcourt-Baldwin, J.L. (2000) Thehydrographic character of the Knysna Estuary.Trans. roy. Soc.South Afr.55,107^122.
Levinton, J.S. (1979) The effect of density upon deposit-feedingpopulations: movement, feeding and floating of Hydrobia ventrosa(Gastropoda: Prosobranchia). Oecologia (Berlin) 43, 27^39.
Levinton, J.S., Stewart, S. & Dewitt,T.H. (1985) Field andlaboratory experiments on interference between Hydrobia totteniand Ilyanassa obsoleta (Gastropoda) and its possible relation toseasonal shifts in vertical mudflat zonation. Mar. Ecol. Prog. Ser.22,53^58.
Macnae,W. (1968) A general account of the fauna and flora ofmangrove swamps and forests in the Indo-West-Pacific region.Adv. Mar. Biol. 6,74^270.
Puttick, G.M. (1977) Spatial and temporal variations in intertidalanimal distributions at Langebaan Lagoon, South Africa.Trans.Royal Soc. South Afr.42, 403^440.
deVilliers, C. & Hodgson, A. (1999) Studies on estuarinemacroinvertebrates. The macrobenthos. In: Estuaries of SouthAfrica (Eds B. Allanson and D. Baird). Cambridge UniversityPress, Cambridge.
Wilke,T., Rola¤ n, E. & Davis, G.M. (2000) The mudsnail genusHydrobia s.s. in the northern Atlantic and westernMediterranean: a phylogenetic hypothesis. Mar. Biol. (Berlin) 137,827^833.
(Manuscript accepted 25 October 2001)
# 2002 East AfricanWild Life Society, Afr. J. Ecol., 40, 289^294
294 R. S. K. Barnes