Embed Size (px)
Citation preview

The Olfactory Organ ModulatesGonadotropin-Releasing Hormone Typesand Nest-Building Behavior in theTilapia Oreochromis niloticus
Hiroshi Uchida,1,2 Satoshi Ogawa,1 Mina Harada,1 Masato Matushita,1
Munehico Iwata,2 Yasuo Sakuma,1 Ishwar S. Parhar1
1 Department of Physiology, Nippon Medical School, Tokyo 113-8602, Japan
2 Laboratory of Ecophysiology, Kitasato University, Sanriku, Iwate 022-0101, Japan
Received 18 August 2004; accepted 15 February 2005
ABSTRACT: Direct olfactory inputs to any of the
known gonadotropin-releasing hormone (GnRH) con-
taining neurons have not been demonstrated. Therefore,
the rationale of this study was to examine whether olfac-
tory inputs might in some way interact with the GnRH
system(s) to synchronize reproductive behaviors. In
order to establish this, we used anosmic mature male
tilapia to investigate changes in reproductive behaviors,
gonadal morphology, and GnRH1, GnRH2, and GnRH3
cellular morphology and change in GnRH mRNA levels
by real-time polymerase chain reaction. Bilateral
removal of the olfactory rosettes followed by occlusion
of the nasal cavity (ORX) inhibited nest-building behav-
ior, but had no effect on aggressive and sexual behaviors
or gonadal morphology. ORX failed to alter the mor-
phological features of GnRH1, GnRH2, and GnRH3
(cell number, size, GnRH optical density), but signifi-
cantly decreased copies of GnRH1 and GnRH2 mRNAs.
GnRH immunoreactive fibers were not evident in the
olfactory nerve and rosettes. DiI application to the olfac-
tory nerve labeled inputs primarily to the glomerular
layer of the olfactory bulbs and extrabulbar inputs to
the forebrain but not to GnRH neurons. These results
provide evidence that the olfactory rosette is crucial for
modulating nest-building behavior through second-
order olfactory pathways interacting with GnRH1 and
GnRH2 neuronal systems. ' 2005 Wiley Periodicals, Inc. J Neu-
robiol 65: 1–11, 2005
Keywords: terminal nerve; preoptic; midbrain; aggressive;
sexual behavior
INTRODUCTION
Fish do not possess a distinct vomeronasal organ
(Eisthen, 1992). Therefore, chemical cues in fish are
mediated primarily by the olfactory system express-
ing both the odorant and pheromone receptors that in
mammals are respectively segregated to the olfactory
epithelium–main olfactory bulb and the vomeronasal
organ–accessory olfactory bulb system (Dulka, 1993;
Bargmann, 1997; Westberry and Meredith, 2003).
Another neuronal system that has been proposed to
be chemosensory in nature in teleosts is the terminal
nerve (Demski and Northcutt, 1983); a bundle of
fibers and cells immunoreactive to gonadotropin-
releasing hormone (GnRH), which runs along the
olfactory nerve (Schwanzel-Fukuda and Silverman,
1980; Wirsig-Wiechmann et al., 2002) with cell
bodies located at the caudal-most part of the olfactory
Correspondence to: Ishwar S. Parhar ([email protected]).Contract grant sponsor: Ministry of Education, Culture, Sports,
Science, and Technology; contract grant numbers: 14580777 (I.S.P.)and 14370025 (Y.S.).
' 2005 Wiley Periodicals, Inc.Published online 7 July 2005 in Wiley InterScience (www.interscience.wiley.com).DOI 10.1002/neu.20156
1

bulbs (Parhar, 2002). Hence, there exists a possible
relationship between the olfactory chemosensory
inputs and the GnRH system(s), although the chemo-
sensory nature of the terminal nerve in teleosts
remains debatable (Fujita et al., 1991). In addition to
the terminal nerve GnRH (¼ GnRH3 in teleosts),
more recently derived teleosts possess two other
GnRH types in the brain: GnRH1 (preoptic area) and
GnRH2 (midbrain) whose precise functions remain
unclear (Parhar, 1997, 2002). Therefore, what
remains to be establish is whether olfactory inputs
might in some way interact with the GnRH system(s)
to synchronize reproductive behaviors.
In the present study, we examined the role of the
olfactory system in the control of reproductive behav-
iors and GnRH1, GnRH2, and GnRH3 types. For this
purpose, we used males of tilapia, Oreochromis nilot-cus, because they display distinctive reproductive
behaviors (nest-building and aggression). We surgi-
cally removed the olfactory rosettes and occluded the
nasal cavity, and observed reproductive behaviors
and gonadal morphology. Using immunocytochemis-
try and real-time quantitative reverse transcriptase
polymerase chain reaction (RT-Q-RT-PCR), we
examined morphological changes and mRNA levels
of GnRH1, GnRH2, and GnRH3 types.
MATERIALS AND METHODS
Animals
Experimental procedures were performed under the guide-
lines of the Animal Care Committee of Nippon Medical
School. Tilapia Oreochromis nilotcus were kept in a large
community tank (114 L) under controlled water tempera-
ture (27 6 18C) and photoperiods (10:14 h dark/light
cycle). Animals were fed, once per day, with commercial
fish pellets (Asakaze mini, Nippon-Haigoshiryo, Kanagawa,
Japan).
Sexually mature male fish that were aggressively
defending territories were selected for experiments. The
olfactory rosettes were heat lesioned for 15 min (electrode
diameter ¼ 0.7 mm; Radiofrequency lesion generator sys-
tem, RFG-4A, Radionics, Inc., MA, USA) and the resulting
dissociated cells and tissue were drawn using a Pasteur
capillary pipette (Corning Glass Works, Corning, NY,
USA). The nasal cavity was filled with dental cement
(Repairsin, GC Corporation, Tokyo, Japan) and sealed
using an adhesive (Aron� 201; Toua-gousei, Tokyo, Japan).
All operations were performed on fish anesthetized in
0.01% solution of tricaine methane-sulfonate (MS222;
Sigma, St. Louis, MO, USA). After the operation, fish were
kept individually in 40-L tanks for 7 days to recover. Fish
with bilaterally removed olfactory rosettes followed by
occlusion of the nasal cavity (ORX) and control fish with
intact olfactory rosettes were segregated into three experi-
mental groups.
Experiment 1 . ORX males (standard length, SL ¼ 14.4
6 0.9 cm; body weight, BW ¼ 76.46 13.7 g, n ¼ 5), intact
males (SL ¼ 13.8 6 1.0 cm, BW ¼ 77.1 6 6.0 g, n ¼ 6),
and ovariectomized prostaglandin injected females (SL
¼ 13.7 6 0.3 cm; BW ¼ 76.7 6 5.5 g) were kept for a 70-
dayssurvival period and observed for nest-building, aggres-
sive, and sexual behavior (see below for details about ovar-
iectomy and behavioral test). To minimize bias in behavio-
ral testing, each female or intact male had an equal opportu-
nity to be paired with a different ORX male. Each ORX
male was tested for sexual behavior with a female and
aggressive behavior with an intact male. Observations for
nest-building behavior were conducted daily, however,
observations for aggressive and sexual behaviors were con-
ducted on different days but at a particular time of the day
(1400–1700 h), which needed more time; therefore animals
were kept for a 70-day survival period. Because the olfac-
tory epithelium has the unique capacity to reconstitute near-
normal its population of neurons and non-neuronal cells
after near-complete destruction (Schwob et al., 1999), ORX
was performed three times (days 0, 30, and 45) during the
70-day survival period to ensure remnants of the olfactory
rosettes did not regenerate. At the end of the 70-day experi-
mental period, the brains were dissected out and immunor-
eacted with specific GnRH antibodies, and changes in cell
morphological features were analyzed. The fact that fish
actively building nests also aggressively defend their terri-
tories suggests nest-building and aggressive behaviors
might be regulated through a common neuronal pathway.
However, to our surprise the results in Experiment 1
showed a significant decrease in the number of ORX fish
making nests without change in their aggressive behavior,
which prompted us to conduct Experiments 2 and 3 to
examine if we could repeat these results; test for sexual
behavior was omitted. Therefore, Experiments 2 and 3 had
a short survival period (14 days).
Experiment 2. ORX males (SL ¼ 12.6 6 0.9 cm, BW
¼ 65.0 6 12.0 g, n ¼ 5) and intact males (SL ¼ 13.1 6 0.6
cm, BW ¼ 70.0 6 11.7 g, n ¼ 6) were kept for a 14-day
survival period and observed for nest-building and aggres-
sive behavior. The brains were immunoreacted with spe-
cific GnRH antibodies, and changes in GnRH1, GnRH2,
and GnRH3 cell morphological features were analyzed.
Experiment 3. ORX males (SL ¼ 12.6 6 0.9 cm, BW
¼ 61.2 6 12.2 g, n ¼ 6) and intact males (SL ¼ 13.1 6 0.7
cm, BW ¼ 63.6 6 6.9 g, n ¼ 5) were kept for a 14-day sur-
vival period and observed for nest-building and aggressive
behavior. The brains were analyzed for GnRH1, GnRH2,
and GnRH3 mRNA levels.
Behavioral Observations
Behavioral tests were conducted in twelve 40-L (liters)
observation aquaria each containing a gravel bed (�2 cm
2 Uchida et al.

thick). Observation aquaria (45 � 30 � 30 cm) containing
40 L of water were covered along the sides with opaque
sheets of paper to prevent visual contact between aquaria,
and arranged in two rows of six on a stand so that activity
in two aquaria could be recorded simultaneously by one
video camera. To observe aggressive behaviors in tilapia
niloticus (BW ¼ 50–70 g; SL ¼ 10–15 cm), 40–60-L tanks
are ideal (personal observations). Nest-building, aggressive,
and sexual behaviors were observed between 1400 and
1700 h, i.e., the time when aggressive behavior is at the
peak under natural daylight conditions in tilapia niloticus(unpublished observations) and tilapia mossambicus(Munro and Singh, 1987). To minimize bias in testing, each
female or intact male had an equal opportunity to be paired
with a different ORX male. Each ORX male was tested for
sexual behavior with a female or for aggressive behavior
with an intact male.
Nest-Building Behavior. The gravel bed in each tank
allowed the male fish to make nest. During nest-building,
the male fish picks up large amounts of gravels with the
mouth and deposits them away from the cleaned surface,
resulting in a clear depression in the gravel substrate near
the chosen spawning site. The nest is generally circular and
the glass bottom of the tanks becomes clearly visible. The
nest size was scored as 3 points (>15 cm), 2 points (5–
15 cm), 1 point (1–5 cm), or 0 point (no nest). The tanks
were observed for the presence of nest once a day (2 PM),
after which the nests were destroyed.
Aggressive Behavior. In Experiment 1, to avoid bias in
testing, aggressive behavior was conducted in the ORX
male’s tank and again on a different day in the intact male’s
tank; in Experiments 2 and 3, aggressive behavior was con-
ducted in an unfamiliar tank (neutral tank) to both the ORX
and the intact male. Behavioral observations were limited
to 15 min duration, during which the dominant male (hold-
ing spawning site) was pinkish/red in coloration and the
defeated fish changed its body coloration to dark pigmenta-
tion (‘‘zebra-like’’ pattern) and became motionless at the
surface of the water. For each fish, behavior was observed
on alternate days. N ¼ 5/6 observations/fish. Aggressive
behavior was scored on a 3-point scale: winner (2 points),
equal (1 point) and loser (0 point).
Male Sexual Behavior. Sexually mature females (SL
¼ 13.76 0.3 cm; BW ¼ 76.76 5.5 g) were ovariectomized
and kept for a week to recover. Prostaglandin F2� [10 �g/2 �L; a dose that should induce female spawning behavior
for at least 1 h postinjection in the goldfish (Stacey and
Kobayashi, 1996) and in a cichlid fish (Cole and Stacey,
1984) was injected intraperitoneally into ovariectomized
females 30 min before being exposed to ORX males. Male
sexual behavior such as nudging (‘‘gentle bites’’: male pla-
ces his mouth close to the female’s anal region and gently
nudges the female) or leading (the male swims ostenta-
tiously in front of the female, leading her to the spawning
site), courtship (the male quivers sideways and circles
around the female), and foraging behavior (the fish picks up
small amounts of gravel, which it ‘‘chews’’ awhile and
spits out again, appears searching for food) were video
recorded for 10 min, and the number of times each act was
performed was counted. Male sexual behavior was
observed only in Experiment 1.
Tissue Fixation
At the end of the survival period, male fish from Experi-
ments 1 and 2 were anesthetized by immersing in 0.01%
solution of tricaine methane-sulfonate (MS222; Sigma,
St. Louis, MO, USA) and sacrificed by rapid spinal transec-
tion. The brains and testis tissues were removed and fixed
in Bouin’s solution for 20 h at room temperature. The tis-
sues were then dehydrated through a graded series of etha-
nols, cleared in n-butanol, and embedded in Paraplast Plus
(Oxford Labware, St. Louis, MO, USA). Serial brain sec-
tions were cut in the coronal plane (15 �m) and processed
for GnRH1, GnRH2, and GnRH3 immunocytochemistry;
testis sections (5 �m) were processed for routine histology
using cresyl violet staining.
GnRH Peptides: Immunocytochemistry
The brain sections were deparaffinized in xylene, rehy-
drated through graded ethanols to phosphate-buffered saline
(0.01 M PBS; pH 7.6), and incubated in 0.03% solution of
H2O2 in 20% methanol for 15 min in the dark. Primary anti-
serum (anti-mammalian GnRH, no. 635.5: 1:3500; anti-
chicken GnRH II, ISP II: 1:3000 in 0.01 M PBS) with 0.3%
Triton X-100 was applied to each section, and slides were
incubated in a closed moist chamber for 36 h at 48C. Thesections were then incubated in biotinylated anti-rabbit
immunoglobulin G (IgG) for 1 h and in avidin-biotinylated
horseradish peroxidase complex for 1 h (Vectastain ABC
Elite kit, Vector Laboratories, Burlingame, CA, USA). Sec-
tions were rinsed in 0.05 M Tris, pH 7.6 (Sigma) and devel-
oped with 0.05% 3,30 diaminobenzidine tetrahydrochloride
(Sigma) with 0.03% H2O2 in 0.05 M Tris buffer for 15 min.
To stop the reaction, sections were rinsed in tap water.
Dehydrated sections were cleared in xylene and coverslips
were applied with Permount (Fisher Chemical, NJ, USA).
The specificities of GnRH antisera used in this study have
been reported previously (Parhar et al., 1996).
GnRH Morphological Data Analysis
GnRH cell numbers, cell area, and cell optical density were
determined by digitizing images of the cells with a micro-
scope (X 400; Leica Microsystems, Model DM-RXA)
equipped with a CCD video camera (Model 1300-Y/HS;
Roper Scientific, Tucson, AZ, USA). Two-dimensional area
was calculated for all stained cells in the plane of the sec-
tion, whose perimeter and nucleus was discernible. Outlines
of stained cells were traced by moving a mouse-controlled
cursor along the digitized image to remove all cell proc-
esses. The cell size and optical density of staining were ana-
Olfactory Organ Modulates GnRH 3

lyzed using an image-processing program (Leica Quantimet
500). The imaging system was set to predetermined settings
on the microscope, so that the objective, the light intensity,
the openings of the condenser, and base diaphragms center-
ing the light for the condenser on the specimen and the con-
denser height were all used at constant settings. The optical
density of staining for each GnRH cell was corrected by
subtracting the background value. The average optical den-
sity of staining per cell was calculated from values taken
from all GnRH-immunoreactive cells in alternate sections
throughout the brain. Although semiquantitative, the esti-
mates of optical density were highly reproducible. To
reduce variability in immunocytochemical results, the con-
ditions of the immunocytochemical reactions were con-
trolled and kept homogeneous on all parameters. The
results were analyzed using Student’s t test.
GnRH mRNAs: Real-Time PCR
ORX male fish from Experiments 3 were anesthetized by
immersing in 0.01% solution of MS222 (Sigma) and sacri-
ficed by rapid spinal transaction at the end of the 14-day
survival period. The brains were removed and placed on
dry ice and stored at �808C. Total RNA was isolated from
whole brain of tilapia O. niloticus using Isogen (Nippon
Gene, Tokyo, Japan). First-strand cDNA was synthesized
from 5 �g of total RNA using Superscript III Reverse Tran-
scriptase (Invitrogen Corp., Carlsbad, CA, USA) and
50 pmol of oligo(dT)18–21 primer (Invitrogen) according to
the manufacturer’s instructions.
RT-Q-RT-PCR was performed in duplicate in 10 �Lreaction volumes consisting of 1X TaqMan Universal PCR
Master Mix (Applied Biosystems, Foster City, CA), 300
nM of primers (G1, G2, G4, G5, G7, G8, A1, A2; Table 1),
200 nM of hybridization probe (G3, G6, G9, A3; Table 1),
and 1/20 of a reverse transcribed sample cDNA or standard
cDNA using the ABI PRISM 7700 Sequence Detection
System (TaqMan PCR, PE Applied Biosystems). The PCR
conditions were 958C for 10 min, followed by 75 cycles at
958C for 15 sec, 608C for 1 min. Hybridization primers and
fluorogenic probes for RT-Q-RT-PCR were optimized
using the ABI PRISM Primer Express Software (Applied
Biosystems). The GenBank accession numbers of the
primers and probes of GnRH1, GnRH2, and GnRH3
types and �-actin in tilapia are as follows: GnRH1,
AB101665; GnRH2, AB101666; GnRH3, AB101667; �-actin, AB037865.
For quantification of transcripts by RT-Q-RT-PCR,
standard curves were generated following the method of
Fronhoffs et al. (2002). Briefly, cRNA of target genes
(GnRH1, GnRH2, GnRH3, and �-actin) were synthesized
by in vitro transcription. After linearization and removal of
the plasmid vector, the total cRNA was diluted in 20 �L of
RNase-free water and quantified using a spectrophotometer
at 260 nm (GeneQuant pro ‘‘S,’’ Amersham Pharmacia
Biotech, NJ, USA). Serial dilutions of cRNA of the target
genes (104–108 molecules), reverse-transcribed to cDNA,
were used to determine the threshold cycle at which the flu-
orescence intensity became delectable. Using an analysis
software (TaqMan PCR, ABI PRISM 7700 Sequence
Detection System, PE Applied Biosystems, Foster City,
CA, USA), a standard curve was generated by plotting the
threshold cycles against the serially diluted known copy
numbers of transcripts. The unknown copies of transcripts
in the samples were calculated from the standard curve
using the number of threshold cycles at which the tran-
scripts became detectable. Copy numbers of GnRH tran-
scripts were normalization with �-actin. Quantification data
are given as mean 6 SEM followed by Student’s t test.
Carboxycyanine Dye Application to theOlfactory Nerve
Sexually mature male and female tilapia (BW ¼ 45.56 3.3 g;
SL ¼ 11.4 6 0.2 cm; n ¼ 7) were anesthetized in 0.01%
solution of MS222 (Sigma). Carbocyanine dye 1,10-diocta-decyl-3,3,3,30-tetramethylindocarbocyanine perchlorate (DiI,
Molecular Probes, Inc., Eugene, OR) crystals were applied
to the right olfactory nerve and the nasal cavity was filled
with dental cement (GC Corporation) in order to keep the
DiI crystal from dislocating, and sealed using an adhesive
(Toua-gousei). After DiI application, animals were sacri-
Table 1 Sequence of GnRH1, GnRH2, and GnRH3 Primers and Fluorogenic Probes Used in TaqMan Quantitative PCR
Primer Sequence Code
Q-GnRH1-F 50-CTCGCAGGGACGGTGTTT-30 G1
Q-GnRH1-R 50-TCTTCCCTCCTGGGCTCAGT-30 G2
Q-GnRH1 Probe 50-CACAGGGCTGCTGTCAACACTGGTCATA-30 G3
Q-GnRH2-F 50-TGGTCCCATGGTTGGTATCC-30 G4
Q-GnRH2-R 50-CCCTGCTTCACACAGCTTAATCT-30 G5
Q-GnRH2 Probe 50-AAATCTCTGATGTCCCAAAGGAGTCCAGCT-30 G6
Q-GnRH3-F 50-TGCTGGCGTTGGTGGTT-30 G7
Q-GnRH3-R 50-CCTCAAGCTCTCCCACACTTCT-30 G8
Q-GnRH3 Probe 50-CAGCACTGGTCCTATGGATGGCTACC-30 G9
Q-�-actin F 50-CCTGACAGAGCGTGGCTACTC-30 A1
Q-�-actin R 50-TCTCTTTGATGTCACGCACGAT-30 A2
Q-�-actin Probe 50-TTCACCACCACAGCCGAGAGGGA-30 A3
All probes were labeled with 50 FAM reporter dye and 30TAMARA quencher dye.
4 Uchida et al.
ficed by rapid spinal transection. The brains were kept
in situ in the brain cases, but the skulls were carefully
opened to give better access to the fixative. Brains were
exposed in 4% paraformaldehyde and kept in the dark at
428C for 2–12 weeks.
Before cryostat sectioning, the heads were cryoprotected
by overnight infiltration with 20% sucrose in phosphate
buffer (pH 7.6). The brains were excised, embedded in Tis-
sue Tek OCT Compound (Sakura Finetechnical, Tokyo,
Japan), and serial 25-�m sagittal sections were cut and thaw
mounted on gelatin-coated slides. The sections were air
dried, evaluated for DiI fluorescence either with or without
coverslipping with PBS, and further processed for GnRH1,
GnRH2, and GnRH3 fluorescence immunocytochemistry.
RESULTS
Behavior Observations
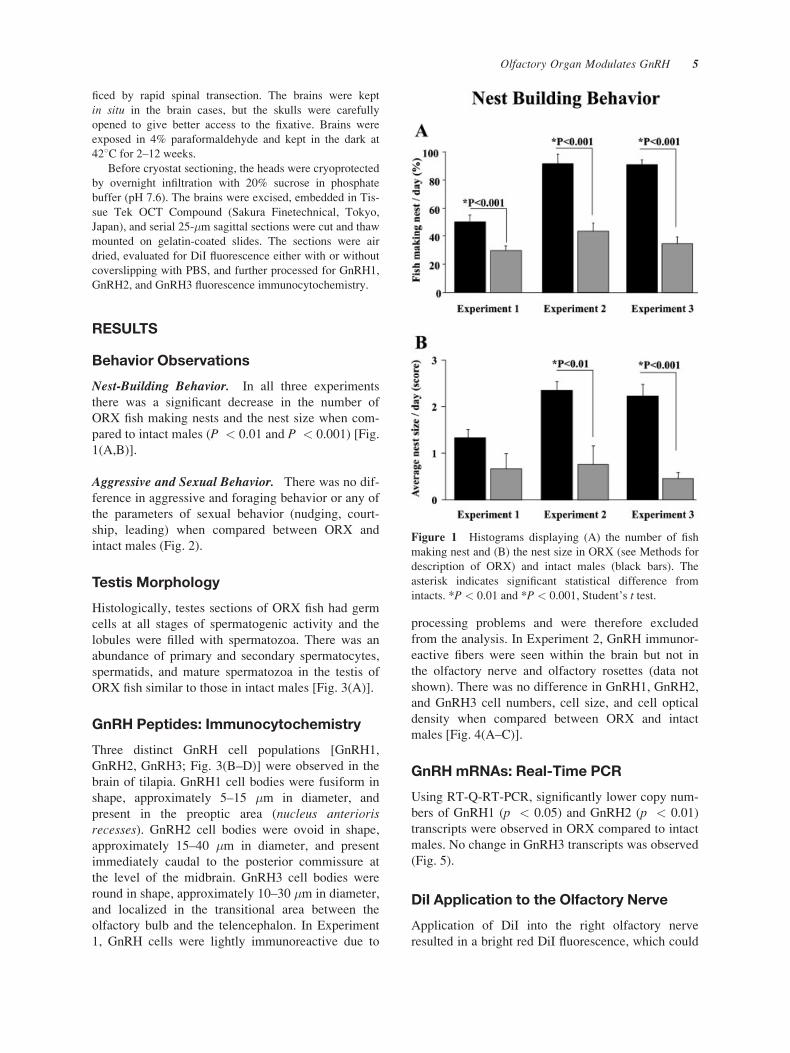
Nest-Building Behavior. In all three experiments
there was a significant decrease in the number of
ORX fish making nests and the nest size when com-
pared to intact males (P < 0.01 and P < 0.001) [Fig.
1(A,B)].
Aggressive and Sexual Behavior. There was no dif-
ference in aggressive and foraging behavior or any of
the parameters of sexual behavior (nudging, court-
ship, leading) when compared between ORX and
intact males (Fig. 2).
Testis Morphology
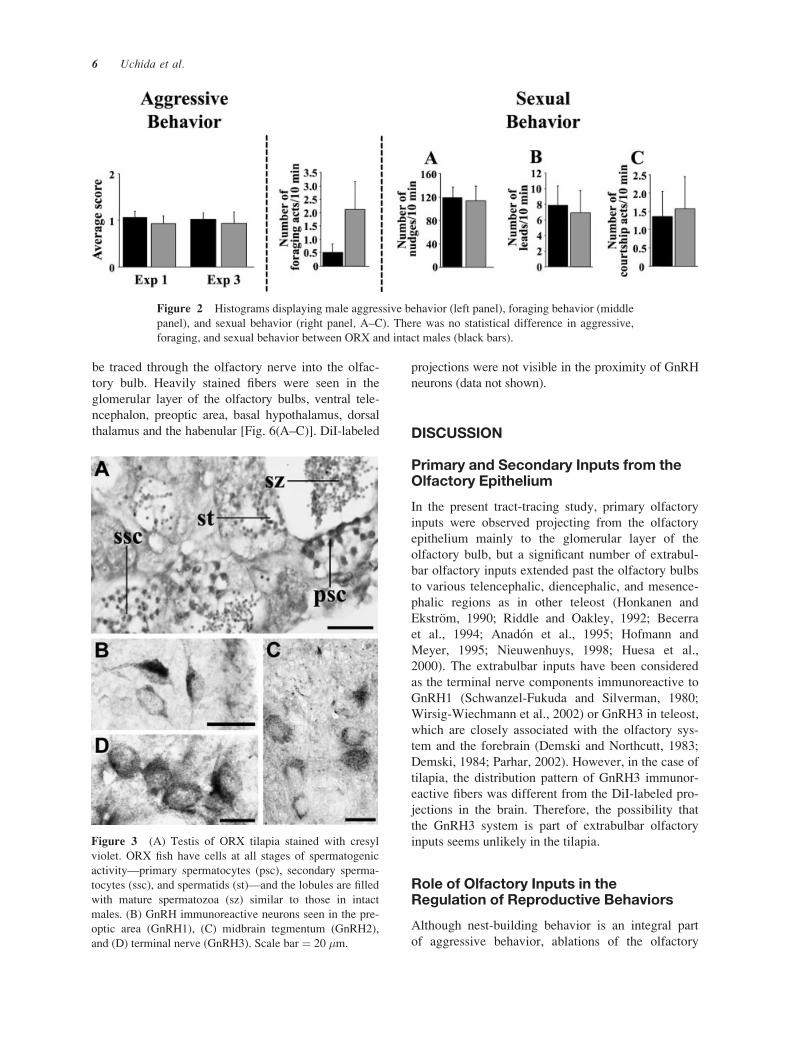
Histologically, testes sections of ORX fish had germ
cells at all stages of spermatogenic activity and the
lobules were filled with spermatozoa. There was an
abundance of primary and secondary spermatocytes,
spermatids, and mature spermatozoa in the testis of
ORX fish similar to those in intact males [Fig. 3(A)].
GnRH Peptides: Immunocytochemistry
Three distinct GnRH cell populations [GnRH1,
GnRH2, GnRH3; Fig. 3(B–D)] were observed in the
brain of tilapia. GnRH1 cell bodies were fusiform in
shape, approximately 5–15 �m in diameter, and
present in the preoptic area (nucleus anteriorisrecesses). GnRH2 cell bodies were ovoid in shape,
approximately 15–40 �m in diameter, and present
immediately caudal to the posterior commissure at
the level of the midbrain. GnRH3 cell bodies were
round in shape, approximately 10–30 �m in diameter,
and localized in the transitional area between the
olfactory bulb and the telencephalon. In Experiment
1, GnRH cells were lightly immunoreactive due to
processing problems and were therefore excluded
from the analysis. In Experiment 2, GnRH immunor-
eactive fibers were seen within the brain but not in
the olfactory nerve and olfactory rosettes (data not
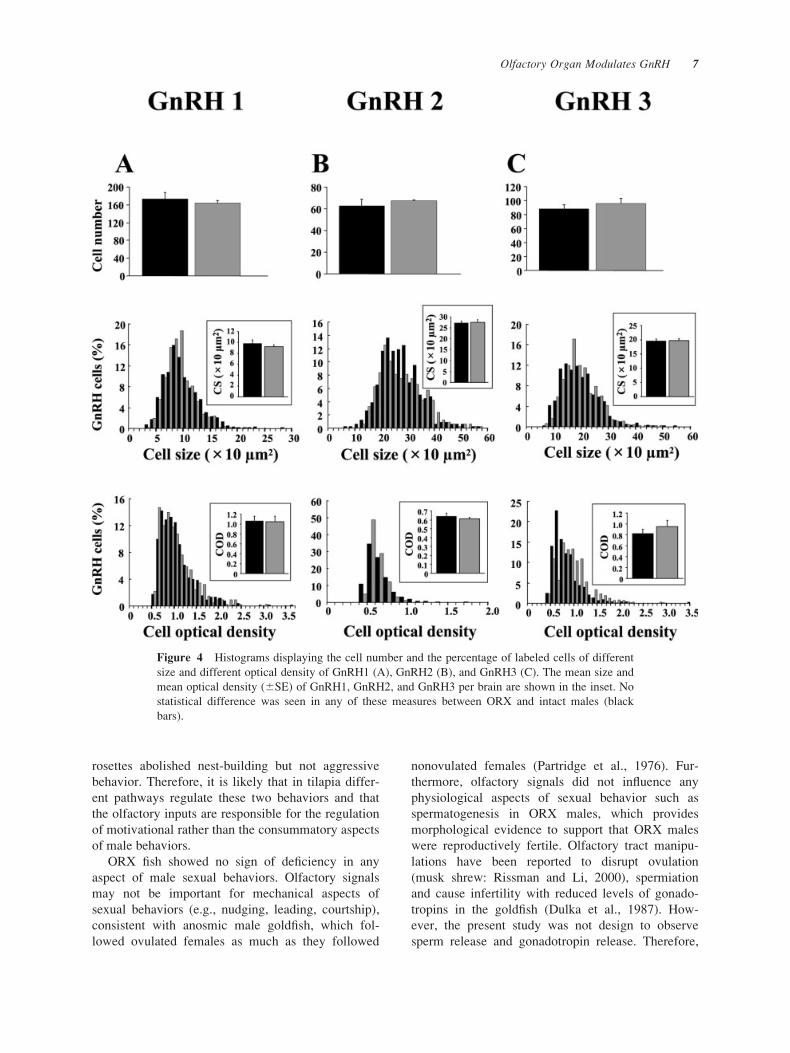
shown). There was no difference in GnRH1, GnRH2,
and GnRH3 cell numbers, cell size, and cell optical
density when compared between ORX and intact
males [Fig. 4(A–C)].
GnRH mRNAs: Real-Time PCR
Using RT-Q-RT-PCR, significantly lower copy num-
bers of GnRH1 (p < 0.05) and GnRH2 (p < 0.01)
transcripts were observed in ORX compared to intact
males. No change in GnRH3 transcripts was observed
(Fig. 5).
DiI Application to the Olfactory Nerve
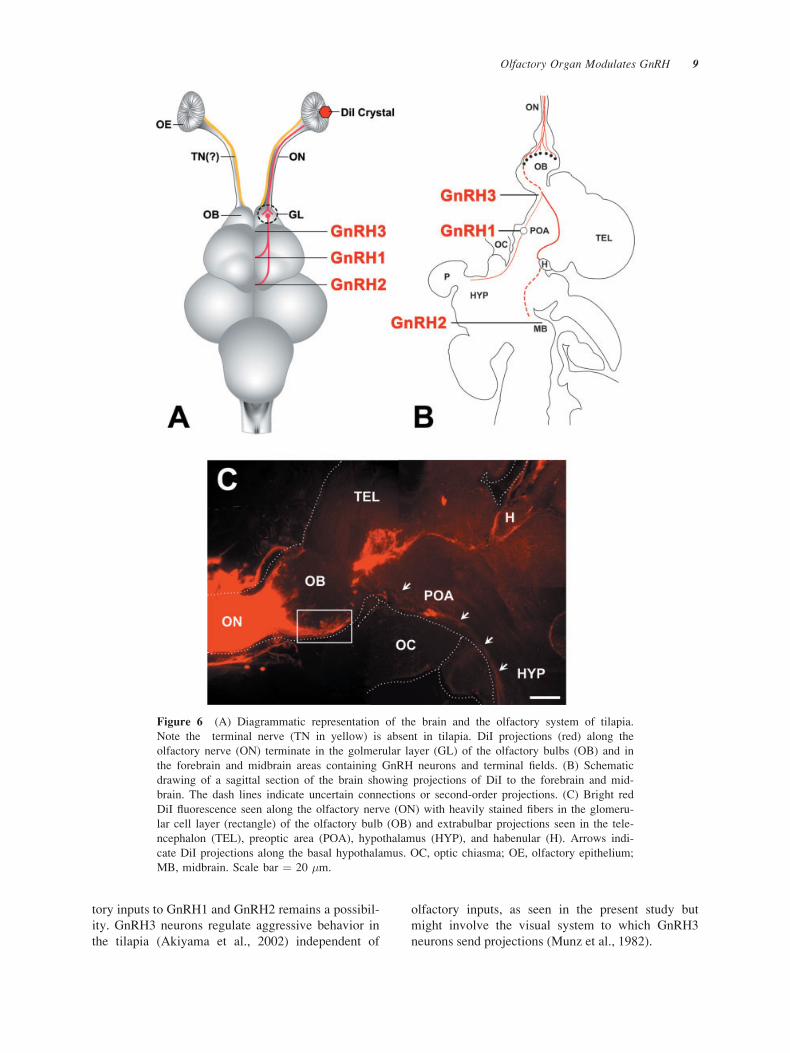
Application of DiI into the right olfactory nerve
resulted in a bright red DiI fluorescence, which could
Figure 1 Histograms displaying (A) the number of fish
making nest and (B) the nest size in ORX (see Methods for
description of ORX) and intact males (black bars). The
asterisk indicates significant statistical difference from
intacts. *P < 0.01 and *P < 0.001, Student’s t test.
Olfactory Organ Modulates GnRH 5
be traced through the olfactory nerve into the olfac-
tory bulb. Heavily stained fibers were seen in the
glomerular layer of the olfactory bulbs, ventral tele-
ncephalon, preoptic area, basal hypothalamus, dorsal
thalamus and the habenular [Fig. 6(A–C)]. DiI-labeled
projections were not visible in the proximity of GnRH
neurons (data not shown).
DISCUSSION
Primary and Secondary Inputs from theOlfactory Epithelium
In the present tract-tracing study, primary olfactory
inputs were observed projecting from the olfactory
epithelium mainly to the glomerular layer of the
olfactory bulb, but a significant number of extrabul-
bar olfactory inputs extended past the olfactory bulbs
to various telencephalic, diencephalic, and mesence-
phalic regions as in other teleost (Honkanen and
Ekstrom, 1990; Riddle and Oakley, 1992; Becerra
et al., 1994; Anadon et al., 1995; Hofmann and
Meyer, 1995; Nieuwenhuys, 1998; Huesa et al.,
2000). The extrabulbar inputs have been considered
as the terminal nerve components immunoreactive to
GnRH1 (Schwanzel-Fukuda and Silverman, 1980;
Wirsig-Wiechmann et al., 2002) or GnRH3 in teleost,
which are closely associated with the olfactory sys-
tem and the forebrain (Demski and Northcutt, 1983;
Demski, 1984; Parhar, 2002). However, in the case of
tilapia, the distribution pattern of GnRH3 immunor-
eactive fibers was different from the DiI-labeled pro-
jections in the brain. Therefore, the possibility that
the GnRH3 system is part of extrabulbar olfactory
inputs seems unlikely in the tilapia.
Role of Olfactory Inputs in theRegulation of Reproductive Behaviors
Although nest-building behavior is an integral part
of aggressive behavior, ablations of the olfactory
Figure 2 Histograms displaying male aggressive behavior (left panel), foraging behavior (middle
panel), and sexual behavior (right panel, A–C). There was no statistical difference in aggressive,
foraging, and sexual behavior between ORX and intact males (black bars).
Figure 3 (A) Testis of ORX tilapia stained with cresyl
violet. ORX fish have cells at all stages of spermatogenic
activity—primary spermatocytes (psc), secondary sperma-
tocytes (ssc), and spermatids (st)—and the lobules are filled
with mature spermatozoa (sz) similar to those in intact
males. (B) GnRH immunoreactive neurons seen in the pre-
optic area (GnRH1), (C) midbrain tegmentum (GnRH2),
and (D) terminal nerve (GnRH3). Scale bar ¼ 20 �m.
6 Uchida et al.
rosettes abolished nest-building but not aggressive
behavior. Therefore, it is likely that in tilapia differ-
ent pathways regulate these two behaviors and that
the olfactory inputs are responsible for the regulation
of motivational rather than the consummatory aspects
of male behaviors.
ORX fish showed no sign of deficiency in any
aspect of male sexual behaviors. Olfactory signals
may not be important for mechanical aspects of
sexual behaviors (e.g., nudging, leading, courtship),
consistent with anosmic male goldfish, which fol-
lowed ovulated females as much as they followed
nonovulated females (Partridge et al., 1976). Fur-
thermore, olfactory signals did not influence any
physiological aspects of sexual behavior such as
spermatogenesis in ORX males, which provides
morphological evidence to support that ORX males
were reproductively fertile. Olfactory tract manipu-
lations have been reported to disrupt ovulation
(musk shrew: Rissman and Li, 2000), spermiation
and cause infertility with reduced levels of gonado-
tropins in the goldfish (Dulka et al., 1987). How-
ever, the present study was not design to observe
sperm release and gonadotropin release. Therefore,
Figure 4 Histograms displaying the cell number and the percentage of labeled cells of different
size and different optical density of GnRH1 (A), GnRH2 (B), and GnRH3 (C). The mean size and
mean optical density (6SE) of GnRH1, GnRH2, and GnRH3 per brain are shown in the inset. No
statistical difference was seen in any of these measures between ORX and intact males (black
bars).
Olfactory Organ Modulates GnRH 7

there remains the possibility that different pathways
regulate different components of male sexual
behavior (Weltzien et al., 2003), which is supported
by the fact that anosmia has little effect on the
spawning success of female blue gourami that are
already in visual contact with males (Pollack et al.,
1978). Thus, it is likely that the olfactory system
together with other sensory systems (visual, tactile)
play an integrated role in the control of sexual
behaviors in male tilapia.
Role of Olfactory Inputs in theRegulation of GnRH Types
GnRH1, GnRH2, and GnRH3 immunoreactive effer-
ents were not detectable in the olfactory rosettes or
the olfactory nerve. On the contrary, olfactory inputs
from the olfactory epithelium were seen projecting to
the olfactory bulbs, ventral telencephalon, the pre-
optic area, basal hypothalamus, dorsal thalamus, and
the habenular. These brain regions are known to con-
tain GnRH cell bodies and/or GnRH terminal fields
(Silverman et al., 1994; Parhar, 1997). Since olfac-
tory projections were not visible in the proximity of
GnRH neurons, the possibility that primary olfactory
inputs project to GnRH terminal fields or second-
order olfactory pathways to GnRH neurons/terminal
fields could account for the down-regulation of
GnRH1 and GnRH2 transcripts levels in ORX fish.
However, ORX significantly reduced copies of
GnRH1 and GnRH2 transcripts but failed to influence
GnRH1 and GnRH2 cell numbers, cell size, and cell
optical density. These unexpected paradoxical find-
ings between transcripts and peptide levels are prob-
ably due to the sensitivities of the techniques; RT-Q-
RT-PCR being more sensitive to detect small changes
in mRNA but cell optical density, a semiquantitative
method, does not detect small changes in GnRH pep-
tide.
The regulation of GnRH3 is probably independ-
ent of the olfactory organ, since destruction of the
olfactory rosettes did not change GnRH3 mRNA
and peptide levels. This came as a surprise, since
the terminal nerve (¼GnRH3 in teleosts) is associ-
ated with the olfactory organ (Parhar, 2002) and
removal of the olfactory rosettes would damage the
GnRH3 system. Since adult tilapia lacked GnRH
fibers along the olfactory nerve/rosettes, GnRH3
neurons in tilapia may not be the terminal nerve
ganglia by definition (Wirsig-Wiechmann et al.,
2002); rather, it is the nucleus olfactoretinalis
(Szabo et al., 1991) that has projections to the ret-
ina (Munz et al., 1982) and play an integrated role
with the visual system. In addition, the activity of
the GnRH3 system does not change when exposed
to odorants or chemical stimuli (Bullock and North-
cutt, 1984); instead, the olfactory system (Fujita
et al., 1991; Huesa et al. 2000) and/or a population
of neurons, present in the vicinity of GnRH3 neu-
rons, might be chemosensory in teleost (Laberge
and Hara, 2003).
Role of GnRH Neurons inReproductive Behaviors
GnRH1 neurons in the forebrain (Pfaff, 1973) or
administration of GnRH1 into the midbrain elicit sex-
ual behavior in female rats (Sakuma and Pfaff, 1980).
More recently, GnRH2 has been reported in the mid-
brain of many vertebrate species, which is more
potent than GnRH1 in the regulation of gonadotro-
pins and reproductive behaviors in the goldfish and in
other vertebrates (Parhar, 2002; Millar, 2003; Volkoff
and Peter, 1999; Rissman, 1996). In the present study,
ablations of olfactory rosettes inhibited GnRH1 and
GnRH2 transcripts and nest-building behavior possi-
bly through primary or second-order olfactory path-
ways to GnRH neurons/terminal fields that have been
implicated in reproductive behaviors in vertebrates
(Silverman et al., 1994; Rissman, 1996; Parhar,
1997). Interestingly, chronic social stress (Ogawa
et al., 2003) or ablations of olfactory rosettes down-
regulate GnRH1 and GnRH2 in tilapia, which suggest
that olfactory inputs and the hypothalamic–pituitary–
adrenal axis might be part of a common integrated
circuitry that regulates GnRH1 and GnRH2 neuronal
system. In another recently derived teleost, the dwarf
gourami, lesions of GnRH3 neurons inhibited nest-
building behavior (Yamamoto et al., 1997); whether
these lesions include destruction of secondary olfac-
Figure 5 Histograms displaying copies of GnRH1,
GnRH2, and GnRH3 transcripts. The asterisk indicates sig-
nificant statistical difference in GnRH1 and GnRH2 tran-
scripts between ORX and intact males (black bars). Stu-
dent’s t test *P < 0.01 and *P < 0.05.
8 Uchida et al.
tory inputs to GnRH1 and GnRH2 remains a possibil-
ity. GnRH3 neurons regulate aggressive behavior in
the tilapia (Akiyama et al., 2002) independent of
olfactory inputs, as seen in the present study but
might involve the visual system to which GnRH3
neurons send projections (Munz et al., 1982).
Figure 6 (A) Diagrammatic representation of the brain and the olfactory system of tilapia.
Note the terminal nerve (TN in yellow) is absent in tilapia. DiI projections (red) along the
olfactory nerve (ON) terminate in the golmerular layer (GL) of the olfactory bulbs (OB) and in
the forebrain and midbrain areas containing GnRH neurons and terminal fields. (B) Schematic
drawing of a sagittal section of the brain showing projections of DiI to the forebrain and mid-
brain. The dash lines indicate uncertain connections or second-order projections. (C) Bright red
DiI fluorescence seen along the olfactory nerve (ON) with heavily stained fibers in the glomeru-
lar cell layer (rectangle) of the olfactory bulb (OB) and extrabulbar projections seen in the tele-
ncephalon (TEL), preoptic area (POA), hypothalamus (HYP), and habenular (H). Arrows indi-
cate DiI projections along the basal hypothalamus. OC, optic chiasma; OE, olfactory epithelium;
MB, midbrain. Scale bar ¼ 20 �m.
Olfactory Organ Modulates GnRH 9

The Leica Image Analysis System was purchased with a
grant from the Japanese Ministry of Education and Science
for Independent Colleges and Universities (2000).
REFERENCES
Akiyama G, Kato S, Soga T, Tamano K, Kawai T, Sakuma
Y, Parhar IS. 2002. Social stress controls terminal nerve
GnRH neurons. Society for Behavioral Neuroscience,
Amherst, MA, USA, Abstract 179.
Anadon R, Manso MJ, Rodriguez-Moldes I, Becerra M.
1995. Neurons of the olfactory organ projecting to the
caudal telencephalon and hypothalamus: a carbocyanine-
dye labelling study in the brown trout (Teleostei). Neuro-
sci Lett 191:157–160.
Bargmann CI. 1997. Olfactory receptors, vomeronasal
receptors, and the organization of olfactory information.
Cell 90:585–587.
Becerra M, Manso MJ, Rodriguez-Moldes I, Anadon R.
1994. Primary olfactory fibres project to the ventral tele-
ncephalon and preoptic region in trout (Salmo trutta): adevelopmental immunocytochemical study. J Comp Neu-
rol 342:131–143.
Bullock TH, Northcutt RG. 1984. Nervus terminalis in dog-
fish (Squalus acanthias, Elasmobranchii) carries tonic
efferent impulses. Neurosci Lett 44:155–160.
Cole KS, Stacey NE. 1984. Prostaglandin induction of
spawning behavior in Cichlasoma bimaculatum (Pisces
cichlidae). Horm Behav 18:235–248.
Demski LS. 1984. The evolution of neuroanatomical sub-
strates of reproductive behavior: sex steroid and LHRH-
specific pathways including the terminal nerve. Am Zool
24:809–830.
Demski LS, Northcutt RG. 1983. The terminal nerve: a new
chemosensory system in vertebrates? Science 220:435–
437.
Dulka JG. 1993. Sex pheromone systems in goldfish: com-
parisons to vomeronasal systems in tetrapods. Brain
Behav Evol 42:265–280.
Dulka JG, Stacey NE, Sorensen PW, Van der Kraak GJ,
Marchant TA. 1987. A sex pheromone system in gold-
fish: is the nervus terminalis involved? Ann NY Acad Sci
519:411–420.
Eisthen HL. 1992. Phylogeny of the vomeronasal system
and of receptor cell types in the olfactory and vomero-
nasal epithelia of vertebrates. Micro Res Tech 23:
1–21.
Fronhoffs S, Totzke G, Stier S, Wernert N, Rothe M, Brun-
ing T, Koch B, Sachinidis A, Vetter H, Ko Y. 2002.
A method for the rapid construction of cRNA stan-
dard curves in quantitative real-time reverse transcrip-
tion polymerase chain reaction. Mol Cell Probes 16:
99–110.
Fujita I, Sorensen PW, Stacey NE, Hara TJ. 1991. The
olfactory system, not the terminal nerve, functions as the
primary chemosensory pathway mediating responses to
sex pheromones in male goldfish. Brain Behav Evol
38:313–321.
Hofmann MH, Meyer DL. 1995. The extrabulbar olfactory
pathway: primary olfactory fibers bypassing the olfactory
bulb in bony fishes? Brain Behav Evol 46:378–388.
Honkanen T, Ekstrom P. 1990. An immunocytochemical
study of the olfactory projections in the three-spined
stickleback, Gasterosteus aculeatus, L. J Comp Neurol
292:65–72.
Huesa G, Anadon R, Yanez J. 2000. Olfactory projections
in a chondrostean fish, Acipenser baeri: an experimental
study. J Comp Neurol 428:145–158.
Laberge F, Hara TJ. 2003. Non-oscillatory discharges of an
F-prostaglandin responsive neuron population in the
olfactory bulb-telencephalon transition area in lake
whitefish. Neuroscience 116:1089–1095.
Millar RP. 2003. GnRH II and type II GnRH receptors.
Trends Endocrinol Metab 14:35–43.
Munro AD, Singh I. 1987. Diurnal changes in the territorial
behavior of the tilapia Oreochromis (¼Sarotheradon)mossambicus. J Fish Biol 30:459–464.
Munz H, Claas B, Stumpf WE, Jennes L. 1982. Centrifugal
innervation of the retina by luteinizing hormone-releasing
hormone (LHRH)-immunoreactive telencephalic neurons
in teleostean fishes. Cell Tissue Res 222:313–323.
Nieuwenhuys R. 1998. Chondrostean fishes. In: Nieuwen-
huys R, Ten Donkelaar HJ, Nicholson C, editors. The
Central Nervous System of Vertebrates, Vol. 1. Berlin:
Springer, p 701–758.
Ogawa S, Soga T, Sakuma Y, Parhar IS. 2003. Modulation
of GnRH subtypes by social stress and aggressive behav-
ior. Fish Physiol Biochem 28:49–50.
Parhar IS. 1997. GnRH in tilapia: three genes, three origins
and their roles. In: Parhar IS, Sakuma Y, editors. GnRH
neurons: gene to behavior. Tokyo: Brain Shuppan, p 99–
122.
Parhar IS. 2002. Cell migration and evolutionary signifi-
cance of GnRH subtypes. Prog Brain Res 141:3–17.
Parhar IS, Pfaff DW, Schwanzel-Fukuda M. 1996. Gonado-
tropin-releasing hormone gene expression in teleosts.
Mol Brain Res 41:216–227.
Partridge BL, Liley NR, Stacey NE. 1976. The role of pher-
omones in the sexual behavior of the goldfish. Anim
Behav 24:291–299.
Pfaff DW. 1973. Luteinizing hormone-releasing factor
potentiates lordosis behavior in hypophysectomized
ovariectomized female rats. Science 182:1148–1149.
Pollack EL, Becker LR, Haynes K. 1978. Sensory control
of mating in the blue gourami, Trichogaster trichopterus(Pisces, Belontiidae). Behav Biol 22:92–103.
Riddle DR, Oakley B. 1992. Immunocytochemical identifi-
cation of primary olfactory afferents in rainbow trout.
J Comp Neurol 324:575–589.
Rissman EF. 1996. Behavioral regulation of gonadotropin-
releasing hormone. Biol Reprod 54:413–419.
Rissman EF, Li X. 2000. Olfactory bulbectomy blocks mat-
ing-induced ovulation in musk shrews (Suncus murinus).Biol Reprod 62:1052–1058.
Sakuma Y, Pfaff DW. 1980. LH-RH in the mesencephalic
central grey can potentiate lordosis reflex of female rats.
Nature 283:566–567.
10 Uchida et al.

Schwanzel-Fukuda M, Silverman AJ. 1980. The nervus ter-
minalis of the guinea pig: a new luteinizing hormone-
releasing hormone (LHRH) neuronal system. J Comp
Neurol 191:213–225.
Schwob JE, Youngentob SL, Ring G, Iwema CL,
Mezza RC. 1999. Reinnervation of the rat olfactory
bulb after methyl bromide-induced lesion: timing and
extent of reinnervation. J Comp Neurol 412:439–
457.
Silverman AJ, Livne I, Witkin JW. 1994. The gonadotro-
pin-releasing hormone (GnRH) neuronal systems: immu-
nocytochemistry and in situ hybridization. In: Knobil E,
Neill JE, editors. Physiology of Reproduction, 2nd ed.
New York: Raven Press, p 1683–1709.
Stacey N, Kobayshi M. 1996. Androgen induction of male
sexual behaviors in female goldfish. Horm Behav 30:
434–445.
Szabo T, Blahser S, Denizot J-P, Ravaille-Veron M. 1991.
The olfactoretinalis system ¼ terminal nerve? Neuro-
report 2:73–76.
Volkoff H, Peter RE. 1999. Actions of two forms of gona-
dotropin releasing hormone and a GnRH antagonist on
spawning behavior of the goldfish Carassius auratus.Gen Comp Endocrinol 116:347–355.
Weltzien FA, Hoglund E, Hamadani el H, Døving KB.
2003. Does the lateral bundle of the medial olfactory
tract mediate reproductive behavior in male crucian
carp? Chem Senses 28:293–300.
Westberry J, Meredith M. 2003. The influence of chemo-
sensory input and gonadotropin releasing hormone on
mating behavior circuits in male hamsters. Brain Res
974:1–16.
Wirsig-Wiechmann CR, Wiechmann AF, Eisthen HL.
2002. What defines the nervus terminalis? Neurochem-
ical, development, and anatomical criteria. Prog Brain
Res 141:45–58.
Yamamoto N, Oka Y, Kawashima S. 1997. Lesions of gona-
dotropin-releasing hormone-immunoreactive terminal
nerve cells: effects on the reproductive behavior of male
dwarf gouramis. Neuroendocrinology 65:403–412.
Olfactory Organ Modulates GnRH 11





























