Embed Size (px)
Citation preview

1348 Journal of Lipid Research Volume 53, 2012
Copyright © 2012 by the American Society for Biochemistry and Molecular Biology, Inc.
This article is available online at http://www.jlr.org
Multiple studies have shown that oxidative modifi ca-tions of LDL (oxLDL) are a major factor in the develop-ment of atherosclerosis ( 1–6 ). The level of oxLDL increases dramatically with hypercholesterolemia in animal models of atherosclerosis ( 7, 8 ), and in humans ( 9, 10 ), it is found in atherosclerotic lesions ( 11 ). It is also well established that dyslipidemia-induced dysfunction of vascular en-dothelial cells is a critical step in the early stage of athero-sclerosis (e.g., Refs. 12–14 ) and a strong predictor of cardiovascular disease (CVD) development ( 15–17 ). The mechanisms, however, of dyslipidemia-induced endothe-lial dysfunction are still poorly understood. Our recent studies have shown that exposure to oxLDL in vitro or to plasma dyslipidemia in vivo signifi cantly increases the stiff-ness of aortic endothelial cells, which in turn is associated with an increase in endothelial contractility, enhanced an-giogenic potential, and sensitivity to shear stress ( 18–20 ). Furthermore, unexpectedly, our studies showed that dys-lipidemia-induced endothelial stiffening is caused not by cholesterol loading but by disruption of lipid packing of cholesterol-rich membrane domains in endothelial cells ( 18, 19, 21 ). Consistent with these observations, oxLDL-induced endothelial stiffness could be fully reversed by enriching the cells with cholesterol, even though oxLDL had no effect on the cholesterol content of endothelial membranes ( 19 ). These studies led us to the hypothesis that oxLDL induces endothelial dysfunction by inserting oxysterols into the plasma membrane, resulting in the dis-ruption of cholesterol-rich membrane domains and en-dothelial stiffening. The goal of this study, therefore, was to determine the impact of oxysterols on endothelial stiffness.
Abstract Endothelial dysfunction is a key step in athero-sclerosis development. Our recent studies suggested that oxLDL-induced increase in endothelial stiffness plays a ma-jor role in dyslipidemia-induced endothelial dysfunction. In this study, we identify oxysterols, as the major component of oxLDL, responsible for the increase in endothelial stiff-ness. Using Atomic Force Microscopy to measure endothe-lial elastic modulus, we show that endothelial stiffness increases with progressive oxidation of LDL and that the two lipid fractions that contribute to endothelial stiffening are oxysterols and oxidized phosphatidylcholines, with ox-ysterols having the dominant effect. Furthermore, endothe-lial elastic modulus increases as a linear function of oxysterol content of oxLDL. Specifi c oxysterols, however, have dif-ferential effects on endothelial stiffness with 7-ketocholes-terol and 7 � -hydroxycholesterol, the two major oxysterols in oxLDL, having the strongest effects. 27-hydroxycholes-terol, found in atherosclerotic lesions, also induces endothe-lial stiffening. For all oxysterols, endothelial stiffening is reversible by enriching the cells with cholesterol. ox-LDL-induced stiffening is accompanied by incorporation of oxysterols into endothelial cells. We fi nd signifi cant ac-cumulation of three oxysterols, 7 � -hydroxycholesterol, 7 � -hydroxycholesterol, and 7-ketocholesterol, in mouse aortas of dyslipidemic ApoE � / � mice at the early stage of athero-sclerosis. Remarkably, these are the same oxysterols we have identifi ed to induce endothelial stiffening. —Shentu, T. P., D. K. Singh, M-J. Oh, S. Sun, L. Sadaat, A. Makino, T. Mazzone, P. V. Subbaiah, M. Cho, and I. Levitan. The role of oxys-terols in control of endothelial stiffness. J. Lipid Res. 2012. 53: 1348–1358.
Supplementary key words cholesterol • atomic force microscopy • cell stiffness • oxidized phosphatidylcholine
The work was supported by National Institutes of Health Grants HL-073965 (to I. L.), HL-083298 (to I. L.), and HL-68585 (to P. V. S.); by American Heart Association predoctoral fellowship (to T. P. S.); and by Max Goldenberg Foun-dation Grant 10PRE2570025 (to T. P. S.). The Atomic Force Microscope was purchased with Defense University Research Instrument Program Grant N00014-04-1-0805 (to M. C.). Its contents are solely the responsibility of the authors and do not necessarily represent the offi cial views of the National Insti-tutes of Health.
Manuscript received 3 April 2012.
Published, JLR Papers in Press, April 11, 2012 DOI 10.1194/jlr.M027102
The role of oxysterols in control of endothelial stiffness
Tzu Pin Shentu , * ,† Dev K. Singh , * Myung-Jin Oh , * Shan Sun , † Laleh Sadaat , * Ayako Makino , § Theodore Mazzone , § Papasani V. Subbaiah , § Michael Cho , † and Irena Levitan 1, * ,†
Pulmonary, Critical Care, and Sleep Medicine* and Endocrinology, Diabetes and Metabolism, § Department of Medicine, University of Illinois , Chicago IL 60612; and Department of Bioengineering, † University of Illinois , Chicago IL 60607
Abbreviations: ApoE � / � , ApoE-defi cient; BAEC, bovine aortic en-dothelial cell; Chol/CE, cholesterol/cholesteryl ester; mmLDL , mini-mally oxidized LDL; oxLDL, oxidized LDL; LPC, lysophosphatidylcholine; M � CD, methyl- � -cyclodextrin; MDA, malondialdehyde; PC, phosphati-dylcholine; oxPC, oxidized PC; PE, phosphatidylethanolamine; SLO, soybean lipoxygenase; SM, sphingomyelin; TBARS, thiobarbituric acid-reactive substance; WT, wild-type.
1 To whom correspondence should be addressed. e-mail: [email protected]
The online version of this article (available at http://www.jlr.org) contains supplementary data in the form of three fi gures .
by guest, on April 6, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2012/04/19/jlr.M027102.DC1Supplemental Material can be found at:

oxLDL and endothelial biomechanics 1349
measuring OD at 234 nm, a method that was used in the previous studies ( 34, 35 ).
Total lipids of LDL or oxLDL extraction and separation of individual lipid fraction
Total lipids were extracted by the Bligh and Dyer procedure ( 36 ) and separated on a silica gel TLC plate (Alltech Associates, Inc.) with the solvent system of chloroform:methanol:water (65:25:4 v/v). A mixture of standard lysophosphatidylcholine (LPC), sphin-gomyelin (SM), phosphatidylcholine (PC), phosphatidyletha-nolamine (PE), and cholesterol/cholesteryl ester (Chol/CE) was run on a on the same plate in separate lane for identifi cation pur-poses. After chromatography, the sample lanes were covered with a glass plate, and the standard lane was exposed to iodine vapors. The region corresponding to individual fraction of lipid was scraped from each lane and dispersed in 500 � l of water, following by the lipid extraction described above. The concentrations of indi-vidual phospholipids were determined by phosphorus content ( 37, 38 ). Total cholesterol (Chol/CE) was obtained by Amplex® Red Cholesterol Assay Kit (Invitrogen, Carlsbad, CA).
Gas chromatography oxysterol analysis For sterol analysis, total lipids were extracted from LDL and
oxLDL after adding 10 � g 19-hydroxycholesterol (19HC) as an internal standard. Trimethylsilyl ether (TMS) derivatives of the sterols were prepared by incubating with 100 � l mixture of N , O -bis-(trimethylsilyl)-acetamide (BSA): n-decane (1:1) at 60°C for 1 h. The TMS ether derivatives of sterols were analyzed by gas chromatography (Shimadzu GC-17A) using a fused carbon-silica column (30 m × 0.25 mm id) coated with (5% phenyl meth-ylpolysiloxane (DB-5MS; fi lm thickness 1 PM) (J and W Scientifi c, Folsom, CA) as described previously ( 39 ). In brief, samples were injected using a split ratio of 1:10. The temperature was pro-grammed for 80–280°C at a rate of 30°C/min after initial hold of 1 min, and then maintained at 280°C for 15 min. The injector and detector temperatures were 280°C and 290°C, respectively.
Extraction of total lipids in BAECs After 1 h incubation with LDL or oxLDL (10 � g of protein/ml
serum free media), cells were washed three times with PBS and lysed using 50 mM Tris-HCl containing 0.15M NaCl and 0.1% Triton X-100. The total lipids were extracted by the Bligh and Dyer method, and the levels of sterols were determined to the internal standard (19HC, 10 � g). For the comparative analysis, values were normalized to LDL-treated cells.
Extraction of total lipids in aorta in ApoE-defi cient mice Aortas were harvested from ApoE-defi cient (ApoE � / � ) and
C57BL/6 wild-type (WT) mice at 10–12 weeks of age. All animals were fed low-fat chow diet. Aortas were dissected /isolated, and samples were put on the fi lter paper to absorb liquid, and then weighed and ground to a powder before extraction. The total lipids were extracted by the Bligh and Dyer method, and the lev-els of sterols were determined by gas chromatography as de-scribed in the previous section. All investigations conformed to the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health (NIH Publication No. 85-23, Revised 1985). The study was conducted in accordance with the guidelines established by the Institutional Animal Care and Use Committee of the University of Illinois at Chicago.
AFM microindentation The elastic modulus of individual endothelial cells was mea-
sured with a Novascan atomic force microscope (AFM; Novascan Technologies, Ames, IA) as described previously ( 19, 40 ). Briefl y,
OxLDL exists in multiple forms, characterized by differ-ent degrees of oxidation and different mixtures of bio-active components ( 3, 6, 22, 23 ). Specifi cally, LDL is progressively oxidized from a minimally oxidized form (mmLDL), which contains mostly oxidized phosphatidyl-cholines (oxPC), to extensively oxidized LDL, which is strongly enriched in oxysterols, as well in several other bio-active lipids, such as lysophospholipids and sphingolipids ( 3, 6, 23 ). Among the bioactive lipids in oxLDL, oxysterols have been shown to be involved at all stages of atheroscle-rosis development, and they play an important role in plaque formation ( 24, 25 ). In the early stage of atheroscle-rosis, oxysterols are involved in impairment of several en-dothelial functions, such as endothelial nitric-oxide synthase (eNOS) activation ( 26 ), reactive oxygen species (ROS) production ( 27 ), and regulation of the permeabil-ity barrier ( 28, 29 ). In the late stage of atherosclerosis, an increase in the levels of oxysterols was shown to contribute to apoptosis of vascular cells ( 29, 30 ) and degradation of extracellular matrix ( 31, 32 ), which led to instability of the plaque. In this study, we demonstrate that oxysterols play a major role in the increase in endothelial stiffness and identify specifi c oxysterols responsible for this effect.
MATERIALS AND METHODS
Cell culture and reagents Bovine aortic endothelial cells (BAEC; Cambrex East Ruther-
ford, NJ) were grown between passages 5 and 20 in DMEM (Invit-rogen, Carlsbad, CA) containing 10% fetal bovine serum (FBS; Sigma-Aldrich, St Louis, MO), 10 µg/ml penicillin, streptomycin, and kanamycin sulfate (Invitrogen). Cell cultures were main-tained in a humidifi ed incubator at 37°C, with 5% CO2 . Cells were split every 3–4 days. Methyl- � -cyclodextrin (M � CD) and cholesterol were purchased from Sigma Chemical, St. Louis, MO. M � CD saturated with cholesterol was prepared as described previously ( 33 ). 7-ketocholesterol, 7 � -hydroxycholesterol, 7 � -hydroxycholesterol, and 25-hydroxycholsterol were purchased from Steraloids, Inc. (Newport, RI). 27-hydroxycholesterol was purchased from Avanti Polar Lipids, Inc. (Alabaster, AL).
Isolation and oxidization of LDL Normal human plasma prepared in acid citrate dextrose was
purchased from the local blood bank (Life Source, Chicago). LDL was isolated by sequential centrifugation in KBr between the density range 1.019 to 1.063 g/ml � 1 . The preparation is dialyzed against three changes of 10 mM Tris/HCl buffer, pH 7.4, con-taining 0.15M NaCl and 1 mM EDTA at 4°C to remove KBr. To proceed to the copper oxidization, further dialysis against PBS (pH 7.4) was performed to reduce the EDTA presence. Copper sulfate was then added to LDL with a fi nal concentration of 25 � M. The oxidation was performed at 37°C for 2, 4, 8, 16, and 24 h and stopped by adding 1 mM EDTA. The content of thiobar-bituric acid-reactive substances (TBARS) in LDL and oxLDL was determined by using TBARS Assay Kit (ZeptoMetrix, Buffalo, NY) as expressed with malondialdehyde (MDA) equivalents (nanomoles of MDA per milligram of LDL protein). In addition, LDL was oxidized by the incubation with 5,000 units of soybean lipoxygenase type IB (SLO) overnight at 37°C, as described ear-lier by ( 34 ). The oxidation state of SLO-oxidated LDL was veri-fi ed by measuring the formation of conjugated dienes by
by guest, on April 6, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2012/04/19/jlr.M027102.DC1Supplemental Material can be found at:
1350 Journal of Lipid Research Volume 53, 2012
for capturing the membrane deformation with 180 s time inter-val. Negative pressure was applied to a pipette by a pneumatic transducer tester (BioTek Instruments,Winooski, VT).
Statistic analysis Statistical analysis was performed using a standard t -test assum-
ing two-tailed distributions with unequal variance.
RESULTS
Endothelial stiffening and degree of LDL oxidation To explore how different oxidization states of LDL af-
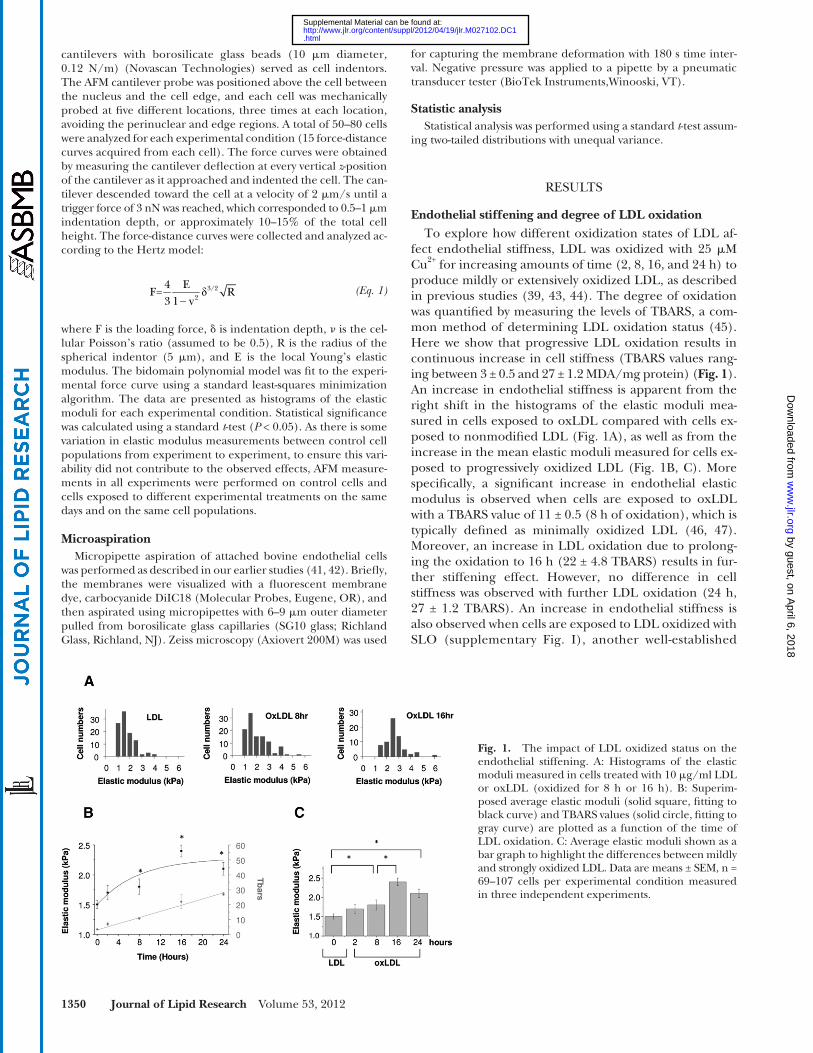
fect endothelial stiffness, LDL was oxidized with 25 � M Cu 2+ for increasing amounts of time (2, 8, 16, and 24 h) to produce mildly or extensively oxidized LDL, as described in previous studies ( 39, 43, 44 ). The degree of oxidation was quantifi ed by measuring the levels of TBARS, a com-mon method of determining LDL oxidation status ( 45 ). Here we show that progressive LDL oxidation results in continuous increase in cell stiffness (TBARS values rang-ing between 3 ± 0.5 and 27 ± 1.2 MDA/mg protein) ( Fig. 1 ). An increase in endothelial stiffness is apparent from the right shift in the histograms of the elastic moduli mea-sured in cells exposed to oxLDL compared with cells ex-posed to nonmodifi ed LDL ( Fig. 1A ), as well as from the increase in the mean elastic moduli measured for cells ex-posed to progressively oxidized LDL ( Fig. 1B, C ). More specifi cally, a signifi cant increase in endothelial elastic modulus is observed when cells are exposed to oxLDL with a TBARS value of 11 ± 0.5 (8 h of oxidation), which is typically defi ned as minimally oxidized LDL ( 46, 47 ). Moreover, an increase in LDL oxidation due to prolong-ing the oxidation to 16 h (22 ± 4.8 TBARS) results in fur-ther stiffening effect. However, no difference in cell stiffness was observed with further LDL oxidation (24 h, 27 ± 1.2 TBARS). An increase in endothelial stiffness is also observed when cells are exposed to LDL oxidized with SLO (supplementary Fig. I), another well-established
cantilevers with borosilicate glass beads (10 � m diameter, 0.12 N/m) (Novascan Technologies) served as cell indentors. The AFM cantilever probe was positioned above the cell between the nucleus and the cell edge, and each cell was mechanically probed at fi ve different locations, three times at each location, avoiding the perinuclear and edge regions. A total of 50–80 cells were analyzed for each experimental condition (15 force-distance curves acquired from each cell). The force curves were obtained by measuring the cantilever defl ection at every vertical z -position of the cantilever as it approached and indented the cell. The can-tilever descended toward the cell at a velocity of 2 � m/s until a trigger force of 3 nN was reached, which corresponded to 0.5–1 � m indentation depth, or approximately 10–15% of the total cell height. The force-distance curves were collected and analyzed ac-cording to the Hertz model :
3/22
4 EF= R
3 1 v (Eq. 1)
where F is the loading force, � is indentation depth, � is the cel-lular Poisson’s ratio (assumed to be 0.5), R is the radius of the spherical indentor (5 � m), and E is the local Young’s elastic modulus. The bidomain polynomial model was fi t to the experi-mental force curve using a standard least-squares minimization algorithm. The data are presented as histograms of the elastic moduli for each experimental condition. Statistical signifi cance was calculated using a standard t -test ( P < 0.05). As there is some variation in elastic modulus measurements between control cell populations from experiment to experiment, to ensure this vari-ability did not contribute to the observed effects, AFM measure-ments in all experiments were performed on control cells and cells exposed to different experimental treatments on the same days and on the same cell populations.
Microaspiration Micropipette aspiration of attached bovine endothelial cells
was performed as described in our earlier studies ( 41, 42 ). Briefl y, the membranes were visualized with a fl uorescent membrane dye, carbocyanide DiIC18 (Molecular Probes, Eugene, OR), and then aspirated using micropipettes with 6–9 � m outer diameter pulled from borosilicate glass capillaries (SG10 glass; Richland Glass, Richland, NJ). Zeiss microscopy (Axiovert 200M) was used
Fig. 1. The impact of LDL oxidized status on the endothelial stiffening. A: Histograms of the elastic moduli measured in cells treated with 10 � g/ml LDL or oxLDL (oxidized for 8 h or 16 h). B: Superim-posed average elastic moduli (solid square, fi tting to black curve) and TBARS values (solid circle, fi tting to gray curve) are plotted as a function of the time of LDL oxidation. C: Average elastic moduli shown as a bar graph to highlight the differences between mildly and strongly oxidized LDL. Data are means ± SEM, n = 69–107 cells per experimental condition measured in three independent experiments.
by guest, on April 6, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2012/04/19/jlr.M027102.DC1Supplemental Material can be found at:
oxLDL and endothelial biomechanics 1351
method of LDL oxidation that results in mildly oxidized LDL ( 34, 35, 47 ). In the latter case, endothelial stiffness was assessed by measuring progressive membrane defor-mation into the pipette using microaspiration, an ap-proach that has been extensively used in our previous studies ( 18, 41 ). Thus, increase in endothelial stiffness is observed across the full range of LDL oxidation states ( Fig. 1B, C ).
Differential effects of oxLDL lipid fractions It is known that the lipid extracts of LDL and oxLDL
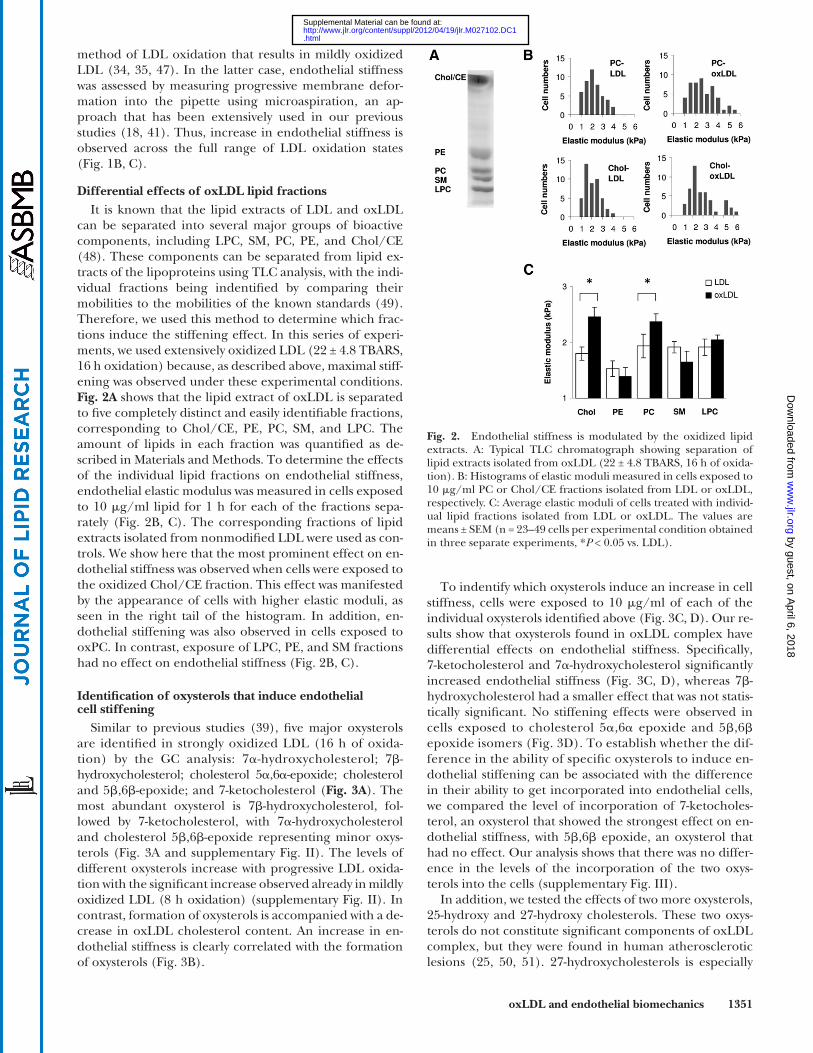
can be separated into several major groups of bioactive components, including LPC, SM, PC, PE, and Chol/CE ( 48 ). These components can be separated from lipid ex-tracts of the lipoproteins using TLC analysis, with the indi-vidual fractions being indentifi ed by comparing their mobilities to the mobilities of the known standards ( 49 ). Therefore, we used this method to determine which frac-tions induce the stiffening effect. In this series of experi-ments, we used extensively oxidized LDL (22 ± 4.8 TBARS, 16 h oxidation) because, as described above, maximal stiff-ening was observed under these experimental conditions. Fig. 2A shows that the lipid extract of oxLDL is separated to fi ve completely distinct and easily identifi able fractions, corresponding to Chol/CE, PE, PC, SM, and LPC. The amount of lipids in each fraction was quantifi ed as de-scribed in Materials and Methods. To determine the effects of the individual lipid fractions on endothelial stiffness, endothelial elastic modulus was measured in cells exposed to 10 � g/ml lipid for 1 h for each of the fractions sepa-rately ( Fig. 2B, C ). The corresponding fractions of lipid extracts isolated from nonmodifi ed LDL were used as con-trols. We show here that the most prominent effect on en-dothelial stiffness was observed when cells were exposed to the oxidized Chol/CE fraction. This effect was manifested by the appearance of cells with higher elastic moduli, as seen in the right tail of the histogram. In addition, en-dothelial stiffening was also observed in cells exposed to oxPC. In contrast, exposure of LPC, PE, and SM fractions had no effect on endothelial stiffness ( Fig. 2B, C ).
Identifi cation of oxysterols that induce endothelial cell stiffening
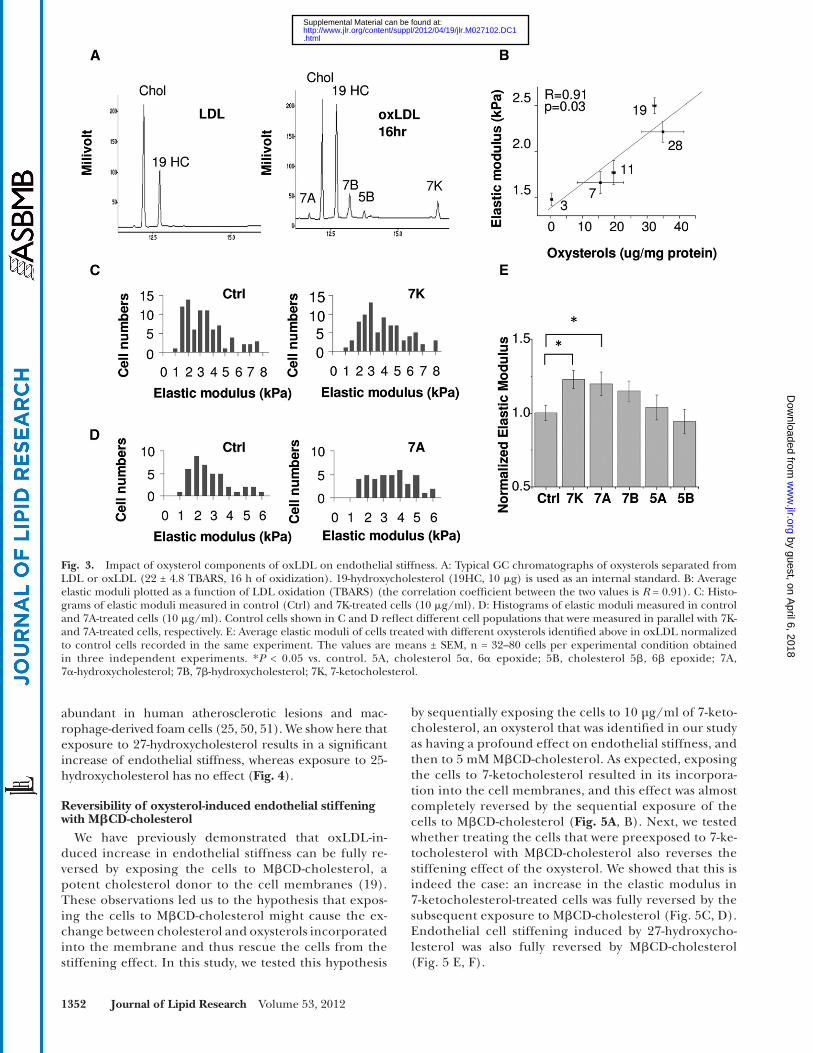
Similar to previous studies ( 39 ), fi ve major oxysterols are identifi ed in strongly oxidized LDL (16 h of oxida-tion) by the GC analysis: 7 � -hydroxycholesterol; 7 � -hydroxycholesterol; cholesterol 5 � ,6 � -epoxide; cholesterol and 5 � ,6 � -epoxide; and 7-ketocholesterol ( Fig. 3A ). The most abundant oxysterol is 7 � -hydroxycholesterol, fol-lowed by 7-ketocholesterol, with 7 � -hydroxycholesterol and cholesterol 5 � ,6 � -epoxide representing minor oxys-terols ( Fig. 3A and supplementary Fig. II). The levels of different oxysterols increase with progressive LDL oxida-tion with the signifi cant increase observed already in mildly oxidized LDL (8 h oxidation) (supplementary Fig. II). In contrast, formation of oxysterols is accompanied with a de-crease in oxLDL cholesterol content. An increase in en-dothelial stiffness is clearly correlated with the formation of oxysterols ( Fig. 3B ).
Fig. 2. Endothelial stiffness is modulated by the oxidized lipid extracts. A: Typical TLC chromatograph showing separation of lipid extracts isolated from oxLDL (22 ± 4.8 TBARS, 16 h of oxida-tion). B: Histograms of elastic moduli measured in cells exposed to 10 � g/ml PC or Chol/CE fractions isolated from LDL or oxLDL, respectively. C: Average elastic moduli of cells treated with individ-ual lipid fractions isolated from LDL or oxLDL. The values are means ± SEM (n = 23–49 cells per experimental condition obtained in three separate experiments, * P < 0.05 vs. LDL).
To indentify which oxysterols induce an increase in cell stiffness, cells were exposed to 10 � g/ml of each of the individual oxysterols identifi ed above ( Fig. 3C, D ). Our re-sults show that oxysterols found in oxLDL complex have differential effects on endothelial stiffness. Specifi cally, 7-ketocholesterol and 7 � -hydroxycholesterol signifi cantly increased endothelial stiffness ( Fig. 3C, D ), whereas 7 � -hydroxycholesterol had a smaller effect that was not statis-tically signifi cant. No stiffening effects were observed in cells exposed to cholesterol 5 � ,6 � epoxide and 5 � ,6 � epoxide isomers ( Fig. 3D ). To establish whether the dif-ference in the ability of specifi c oxysterols to induce en-dothelial stiffening can be associated with the difference in their ability to get incorporated into endothelial cells, we compared the level of incorporation of 7-ketocholes-terol, an oxysterol that showed the strongest effect on en-dothelial stiffness, with 5 � ,6 � epoxide, an oxysterol that had no effect. Our analysis shows that there was no differ-ence in the levels of the incorporation of the two oxys-terols into the cells (supplementary Fig. III).
In addition, we tested the effects of two more oxysterols, 25-hydroxy and 27-hydroxy cholesterols. These two oxys-terols do not constitute signifi cant components of oxLDL complex, but they were found in human atherosclerotic lesions ( 25, 50, 51 ). 27-hydroxycholesterols is especially
by guest, on April 6, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2012/04/19/jlr.M027102.DC1Supplemental Material can be found at:
1352 Journal of Lipid Research Volume 53, 2012
Fig. 3. Impact of oxysterol components of oxLDL on endothelial stiffness. A: Typical GC chromatographs of oxysterols separated from LDL or oxLDL (22 ± 4.8 TBARS, 16 h of oxidization). 19-hydroxycholesterol (19HC, 10 � g) is used as an internal standard. B: Average elastic moduli plotted as a function of LDL oxidation (TBARS) (the correlation coeffi cient between the two values is R = 0.91). C: Histo-grams of elastic moduli measured in control (Ctrl) and 7K-treated cells (10 � g/ml). D: Histograms of elastic moduli measured in control and 7A-treated cells (10 � g/ml). Control cells shown in C and D refl ect different cell populations that were measured in parallel with 7K-and 7A-treated cells, respectively. E: Average elastic moduli of cells treated with different oxysterols identifi ed above in oxLDL normalized to control cells recorded in the same experiment. The values are means ± SEM, n = 32–80 cells per experimental condition obtained in three independent experiments. * P < 0.05 vs. control. 5A, cholesterol 5 � , 6 � epoxide; 5B, cholesterol 5 � , 6 � epoxide; 7A, 7 � -hydroxycholesterol; 7B, 7 � -hydroxycholesterol; 7K, 7-ketocholesterol.
abundant in human atherosclerotic lesions and mac-rophage-derived foam cells ( 25, 50, 51 ). We show here that exposure to 27-hydroxycholesterol results in a signifi cant increase of endothelial stiffness, whereas exposure to 25-hydroxycholesterol has no effect ( Fig. 4 ).
Reversibility of oxysterol-induced endothelial stiffening with M � CD-cholesterol
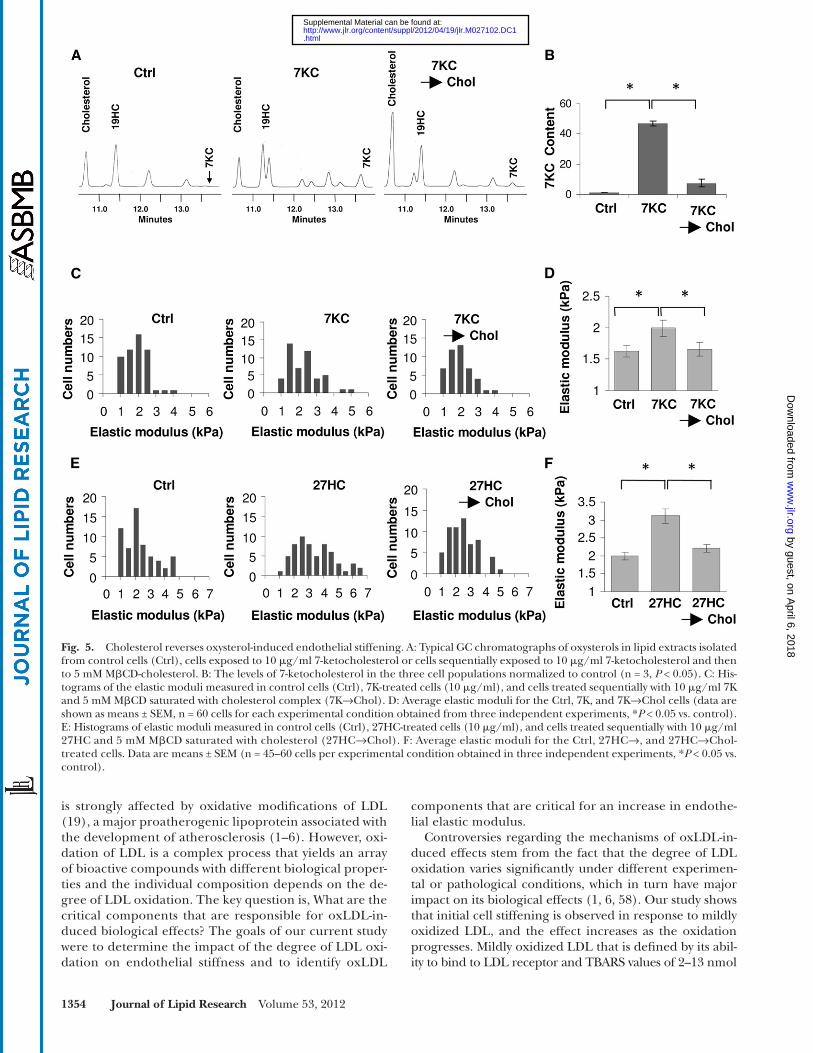
We have previously demonstrated that oxLDL-in-duced increase in endothelial stiffness can be fully re-versed by exposing the cells to M � CD-cholesterol, a potent cholesterol donor to the cell membranes ( 19 ). These observations led us to the hypothesis that expos-ing the cells to M � CD-cholesterol might cause the ex-change between cholesterol and oxysterols incorporated into the membrane and thus rescue the cells from the stiffening effect. In this study, we tested this hypothesis
by sequentially exposing the cells to 10 µg/ml of 7-keto-cholesterol, an oxysterol that was identifi ed in our study as having a profound effect on endothelial stiffness, and then to 5 mM M � CD-cholesterol. As expected, exposing the cells to 7-ketocholesterol resulted in its incorpora-tion into the cell membranes, and this effect was almost completely reversed by the sequential exposure of the cells to M � CD-cholesterol ( Fig. 5A , B ). Next, we tested whether treating the cells that were preexposed to 7-ke-tocholesterol with M � CD-cholesterol also reverses the stiffening effect of the oxysterol. We showed that this is indeed the case: an increase in the elastic modulus in 7-ketocholesterol-treated cells was fully reversed by the subsequent exposure to M � CD-cholesterol ( Fig. 5C, D ). Endothelial cell stiffening induced by 27-hydroxycho-lesterol was also fully reversed by M � CD-cholesterol ( Fig. 5 E, F ).
by guest, on April 6, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2012/04/19/jlr.M027102.DC1Supplemental Material can be found at:

oxLDL and endothelial biomechanics 1353
change in the level of membrane cholesterol in cells ex-posed to oxLDL.
Incorporation of oxysterols in aorta tissue extracts of ApoE � / � mice
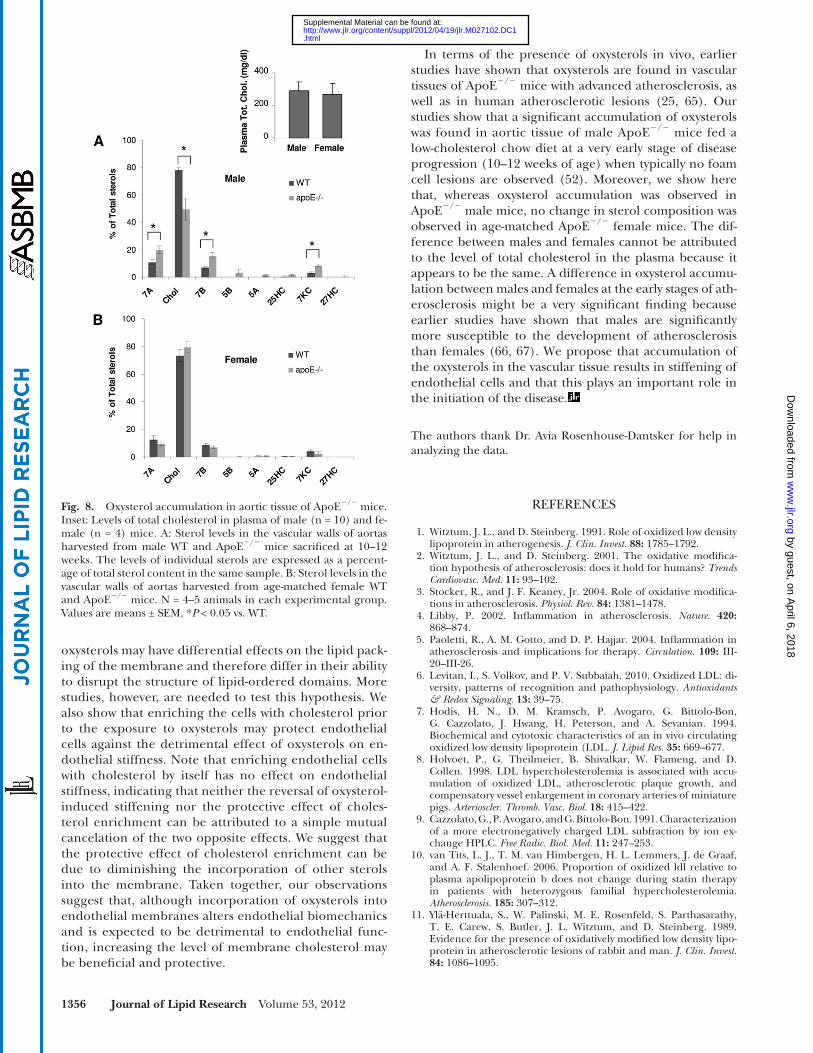
Finally, we addressed the question whether dyslipidemia in vivo also results in the accumulation of oxysterols in the vascular tissues of mouse aortas at the early stage of athero-sclerosis development in ApoE � / � mice, a well-established model for atherosclerosis ( 6, 52, 53 ). Earlier studies have shown that ApoE � / � mice fed low-fat diet can be used as a model for the early stage of atherosclerosis because, al-though these mice are dyslipidemic, development of ath-erosclerosis is signifi cantly delayed, with lesions contain-ing foam cells forming only after 15 weeks and advanced lesions appearing after 20 weeks ( 52 ). In our study, there-fore, the levels of oxysterols were measured in aortas har-vested from 10- to 12-week-old ApoE � / � mice fed a low-fat diet. Sex- and age-matched WT mice of the same genetic background (C57BL/6) were used as controls. The rela-tive amounts of each of the oxysterols, as well as choles-terol, normalized to the total level of tissue sterols are shown in Fig. 8 . Our study shows that in male ApoE � / � mice, there is a signifi cant increase in the two oxysterols that were identifi ed in this study to induce endothelial stiffening: 7-ketocholesterol and 7 � -hydroxycholesterol. There was also some increase in 7- � hydroxycholesterol. Interestingly, although the levels of oxysterols increase, the level of cholesterol does not change signifi cantly, and therefore, its relative contribution to total tissue sterols de-creases, as shown in Fig. 8A . Note, however, that in con-trast to male mice, no change in oxysterol composition was observed in age-matched female mice ( Fig. 8B ). No difference was observed in the levels of plasma cholesterol between males and females (Fig. 8, inset).
DISCUSSION
A growing number of studies suggest that endothelial biomechanical properties play major roles in endothe-lial function ( 20, 42, 54–57 ). The factors that control en-dothelial biomechanics under normal and pathological conditions, however, are still poorly understood. Our re-cent studies have shown that endothelial elastic modulus
Cholesterol enrichment protects against oxysterol-induced endothelial stiffening
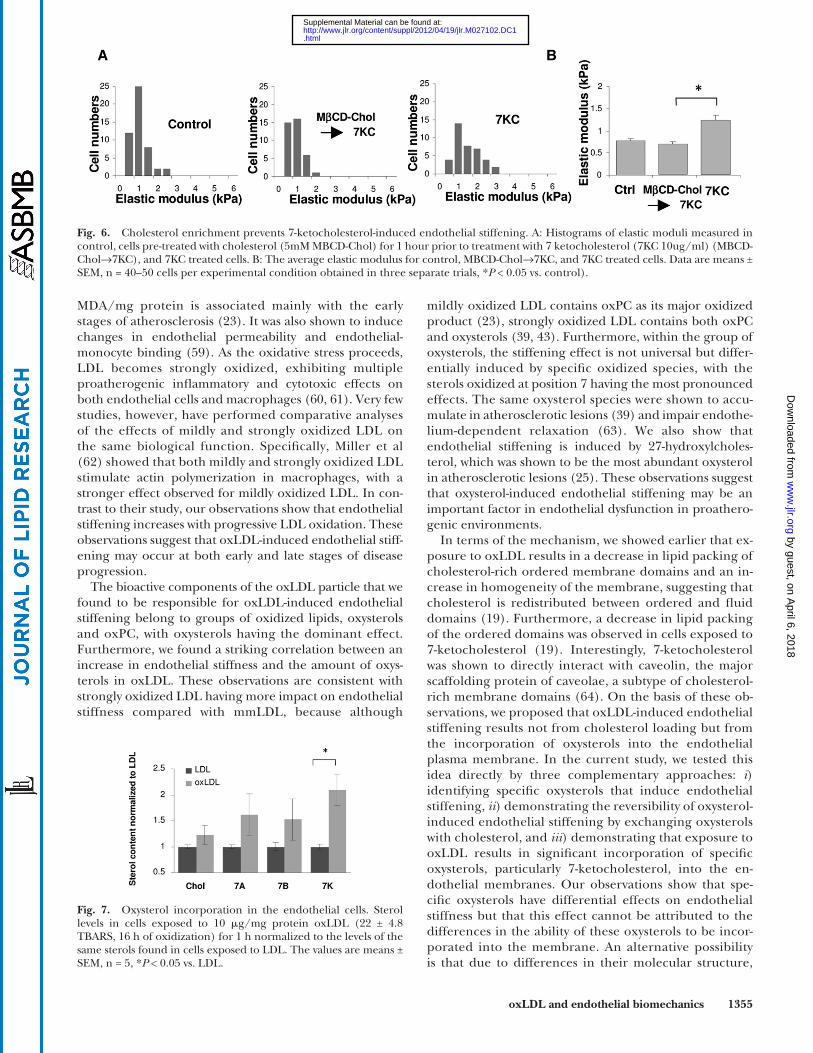
To evaluate further the impact of membrane choles-terol on oxysterol-induced endothelial stiffening, we tested whether cholesterol enrichment prior to the exposure to oxysterols alters the effect of oxysterols on endothelial stiffness. To address this question, cells were fi rst exposed to M � CD-cholesterol, using a standard protocol that re-sults in enriching the cells with cholesterol, and then ex-posed to 7-ketocholesterol. Consistent with the experiments described above, exposure to 7-ketocholesterol alone re-sulted in a signifi cant increase in endothelial elastic modu-lus, demonstrating the robustness and the reproducibility of this effect. However, no stiffening was observed when cells were exposed to 7-ketocholesterol after being choles-terol enriched ( Fig. 6 ). These observations indicate that an increase in membrane cholesterol may have a protective effect against oxysterol-induced endothelial stiffening.
Incorporation of oxysterols into endothelial membranes Next, we addressed the question of whether signifi cant
amount of oxidized lipids is incorporated into the en-dothelial membranes. This question was addressed by esti-mating the levels of different oxysterols in total membrane lysates of endothelial cells exposed to oxLDL or to non-modifi ed LDL used as a control. Endothelial cells grown to confl uence were exposed to 10 µg/ml of extensively oxidized LDL (16 h oxidation, 22 ± 4.8 TBARS) or to non-modifi ed LDL for 1 h, then cells were washed with PBS to remove oxLDL/LDL particles and lysed for the prepara-tion of lipid extracts. The levels of oxysterols were deter-mined in the whole-cell lipid extracts by the GC analysis. This analysis showed that exposure to oxLDL resulted in a highly signifi cant increase in the level of 7-ketocholesterol in lipid extracts of endothelial cells, as is demonstrated by the normalized ratio of 7-ketocholesterol level in cells treated with oxLDL compared with cells treated with nonmodifi ed LDL ( Fig. 7 ). There was also some increase in the levels of 7 � - and 7 � -hydroxycholesterol, but these in-creases were not statistically signifi cant. The difference in the basal levels of the three oxysterols may be the reason that exposure to oxLDL resulted in a signifi cant increase in 7-ketocholesterol but not in 7 � - and 7 � -hydroxycholesterols. Consistent with our earlier studies ( 18 ), we found no
Fig. 4. Impact of oxysterols abundant in atherosclerotic lesions but not in oxLDL. A: Histograms of elastic moduli measured in control cells (Ctrl) and in cells treated with 10 � g/ml 27-hydroxycholesterol (27HC) or 25-hydroxycholesterol (25HC). B: Average elastic moduli for the same experimental cell populations. Data are means ± SEM (n = 58 cells per experimental condition obtained in three independent experiments, * P < 0.05 vs. control).
by guest, on April 6, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2012/04/19/jlr.M027102.DC1Supplemental Material can be found at:
1354 Journal of Lipid Research Volume 53, 2012
components that are critical for an increase in endothe-lial elastic modulus.
Controversies regarding the mechanisms of oxLDL-in-duced effects stem from the fact that the degree of LDL oxidation varies signifi cantly under different experimen-tal or pathological conditions, which in turn have major impact on its biological effects ( 1, 6, 58 ). Our study shows that initial cell stiffening is observed in response to mildly oxidized LDL, and the effect increases as the oxidation progresses. Mildly oxidized LDL that is defi ned by its abil-ity to bind to LDL receptor and TBARS values of 2–13 nmol
is strongly affected by oxidative modifi cations of LDL ( 19 ), a major proatherogenic lipoprotein associated with the development of atherosclerosis ( 1–6 ). However, oxi-dation of LDL is a complex process that yields an array of bioactive compounds with different biological proper-ties and the individual composition depends on the de-gree of LDL oxidation. The key question is, What are the critical components that are responsible for oxLDL-in-duced biological effects? The goals of our current study were to determine the impact of the degree of LDL oxi-dation on endothelial stiffness and to identify oxLDL
Fig. 5. Cholesterol reverses oxysterol -induced endothelial stiffening. A: Typical GC chromatographs of oxysterols in lipid extracts isolated from control cells (Ctrl), cells exposed to 10 � g/ml 7-ketocholesterol or cells sequentially exposed to 10 � g/ml 7-ketocholesterol and then to 5 mM M � CD-cholesterol. B: The levels of 7-ketocholesterol in the three cell populations normalized to control (n = 3, P < 0.05). C: His-tograms of the elastic moduli measured in control cells (Ctrl), 7K-treated cells (10 � g/ml), and cells treated sequentially with 10 � g/ml 7K and 5 mM M � CD saturated with cholesterol complex (7K→Chol). D: Average elastic moduli for the Ctrl, 7K, and 7K→Chol cells (data are shown as means ± SEM, n = 60 cells for each experimental condition obtained from three independent experiments, * P < 0.05 vs. control). E: Histograms of elastic moduli measured in control cells (Ctrl), 27HC-treated cells (10 � g/ml), and cells treated sequentially with 10 � g/ml 27HC and 5 mM M � CD saturated with cholesterol (27HC→Chol). F: Average elastic moduli for the Ctrl, 27HC→, and 27HC→Chol-treated cells. Data are means ± SEM (n = 45–60 cells per experimental condition obtained in three independent experiments, * P < 0.05 vs. control).
by guest, on April 6, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2012/04/19/jlr.M027102.DC1Supplemental Material can be found at:
oxLDL and endothelial biomechanics 1355
mildly oxidized LDL contains oxPC as its major oxidized product ( 23 ), strongly oxidized LDL contains both oxPC and oxysterols ( 39, 43 ). Furthermore, within the group of oxysterols, the stiffening effect is not universal but differ-entially induced by specifi c oxidized species, with the sterols oxidized at position 7 having the most pronounced effects. The same oxysterol species were shown to accu-mulate in atherosclerotic lesions ( 39 ) and impair endothe-lium-dependent relaxation ( 63 ). We also show that endothelial stiffening is induced by 27-hydroxylcholes-terol, which was shown to be the most abundant oxysterol in atherosclerotic lesions ( 25 ). These observations suggest that oxysterol-induced endothelial stiffening may be an important factor in endothelial dysfunction in proathero-genic environments.
In terms of the mechanism, we showed earlier that ex-posure to oxLDL results in a decrease in lipid packing of cholesterol-rich ordered membrane domains and an in-crease in homogeneity of the membrane, suggesting that cholesterol is redistributed between ordered and fl uid domains ( 19 ). Furthermore, a decrease in lipid packing of the ordered domains was observed in cells exposed to 7-ketocholesterol ( 19 ). Interestingly, 7-ketocholesterol was shown to directly interact with caveolin, the major scaffolding protein of caveolae, a subtype of cholesterol-rich membrane domains ( 64 ). On the basis of these ob-servations, we proposed that oxLDL-induced endothelial stiffening results not from cholesterol loading but from the incorporation of oxysterols into the endothelial plasma membrane. In the current study, we tested this idea directly by three complementary approaches: i ) identifying specifi c oxysterols that induce endothelial stiffening, ii ) demonstrating the reversibility of oxysterol-induced endothelial stiffening by exchanging oxysterols with cholesterol, and iii ) demonstrating that exposure to oxLDL results in signifi cant incorporation of specifi c oxysterols, particularly 7-ketocholesterol, into the en-dothelial membranes. Our observations show that spe-cifi c oxysterols have differential effects on endothelial stiffness but that this effect cannot be attributed to the differences in the ability of these oxysterols to be incor-porated into the membrane. An alternative possibility is that due to differences in their molecular structure,
MDA/mg protein is associated mainly with the early stages of atherosclerosis ( 23 ). It was also shown to induce changes in endothelial permeability and endothelial-monocyte binding ( 59 ). As the oxidative stress proceeds, LDL becomes strongly oxidized, exhibiting multiple proatherogenic infl ammatory and cytotoxic effects on both endothelial cells and macrophages ( 60, 61 ). Very few studies, however, have performed comparative analyses of the effects of mildly and strongly oxidized LDL on the same biological function. Specifi cally, Miller et al ( 62 ) showed that both mildly and strongly oxidized LDL stimulate actin polymerization in macrophages, with a stronger effect observed for mildly oxidized LDL. In con-trast to their study, our observations show that endothelial stiffening increases with progressive LDL oxidation. These observations suggest that oxLDL-induced endothelial stiff-ening may occur at both early and late stages of disease progression.
The bioactive components of the oxLDL particle that we found to be responsible for oxLDL-induced endothelial stiffening belong to groups of oxidized lipids, oxysterols and oxPC, with oxysterols having the dominant effect. Furthermore, we found a striking correlation between an increase in endothelial stiffness and the amount of oxys-terols in oxLDL. These observations are consistent with strongly oxidized LDL having more impact on endothelial stiffness compared with mmLDL, because although
Fig. 6. Cholesterol enrichment prevents 7-ketocholesterol-induced endothelial stiffening. A: Histograms of elastic moduli measured in control, cells pre-treated with cholesterol (5mM MBCD-Chol) for 1 hour prior to treatment with 7 ketocholesterol (7KC 10ug/ml) (MBCD-Chol→7KC), and 7KC treated cells. B: The average elastic modulus for control, MBCD-Chol→7KC, and 7KC treated cells. Data are means ± SEM, n = 40–50 cells per experimental condition obtained in three separate trials, * P < 0.05 vs. control).
Fig. 7. Oxysterol incorporation in the endothelial cells. Sterol levels in cells exposed to 10 � g/mg protein oxLDL (22 ± 4.8 TBARS, 16 h of oxidization) for 1 h normalized to the levels of the same sterols found in cells exposed to LDL. The values are means ± SEM, n = 5, * P < 0.05 vs. LDL.
by guest, on April 6, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2012/04/19/jlr.M027102.DC1Supplemental Material can be found at:
1356 Journal of Lipid Research Volume 53, 2012
In terms of the presence of oxysterols in vivo, earlier studies have shown that oxysterols are found in vascular tissues of ApoE � / � mice with advanced atherosclerosis, as well as in human atherosclerotic lesions ( 25, 65 ). Our studies show that a signifi cant accumulation of oxysterols was found in aortic tissue of male ApoE � / � mice fed a low-cholesterol chow diet at a very early stage of disease progression (10–12 weeks of age) when typically no foam cell lesions are observed ( 52 ). Moreover, we show here that, whereas oxysterol accumulation was observed in ApoE � / � male mice, no change in sterol composition was observed in age-matched ApoE � / � female mice. The dif-ference between males and females cannot be attributed to the level of total cholesterol in the plasma because it appears to be the same. A difference in oxysterol accumu-lation between males and females at the early stages of ath-erosclerosis might be a very signifi cant fi nding because earlier studies have shown that males are signifi cantly more susceptible to the development of atherosclerosis than females ( 66, 67 ). We propose that accumulation of the oxysterols in the vascular tissue results in stiffening of endothelial cells and that this plays an important role in the initiation of the disease.
The authors thank Dr. Avia Rosenhouse-Dantsker for help in analyzing the data.
REFERENCES
1 . Witztum , J. L. , and D. Steinberg . 1991 . Role of oxidized low density lipoprotein in atherogenesis. J. Clin. Invest. 88 : 1785 – 1792 .
2 . Witztum , J. L. , and D. Steinberg . 2001 . The oxidative modifi ca-tion hypothesis of atherosclerosis: does it hold for humans? Trends Cardiovasc. Med. 11 : 93 –102.
3 . Stocker, R., and J. F. Keaney , Jr . 2004. Role of oxidative modifi ca-tions in atherosclerosis. Physiol. Rev. 84 : 1381 – 1478 .
4 . Libby , P. 2002 . Infl ammation in atherosclerosis. Nature . 420 : 868 – 874 .
5 . Paoletti, R. , A. M. Gotto , and D. P. Hajjar . 2004. Infl ammation in atherosclerosis and implications for therapy. Circulation . 109 : III-20 – III-26 .
6 . Levitan, I. , S. Volkov , and P. V. Subbaiah . 2010. Oxidized LDL: di-versity, patterns of recognition and pathophysiology. Antioxidants & Redox Signaling . 13 : 39 – 75 .
7 . Hodis , H. N. , D. M. Kramsch , P. Avogaro , G. Bittolo-Bon , G. Cazzolato , J. Hwang , H. Peterson , and A. Sevanian . 1994 . Biochemical and cytotoxic characteristics of an in vivo circulating oxidized low density lipoprotein (LDL . J. Lipid Res. 35 : 669 – 677 .
8 . Holvoet, P. , G. Theilmeier , B. Shivalkar , W. Flameng , and D. Collen . 1998. LDL hypercholesterolemia is associated with accu-mulation of oxidized LDL, atherosclerotic plaque growth, and compensatory vessel enlargement in coronary arteries of miniature pigs. Arterioscler. Thromb. Vasc. Biol. 18 : 415 – 422 .
9 . Cazzolato, G. , P. Avogaro , and G. Bittolo-Bon . 1991. Characterization of a more electronegatively charged LDL subfraction by ion ex-change HPLC. Free Radic. Biol. Med. 11 : 247 – 253 .
10 . van Tits , L. J. , T. M. van Himbergen , H. L. Lemmers , J. de Graaf , and A. F. Stalenhoef . 2006 . Proportion of oxidized ldl relative to plasma apolipoprotein b does not change during statin therapy in patients with heterozygous familial hypercholesterolemia. Atherosclerosis . 185 : 307 – 312 .
11 . Ylä-Herttuala , S. , W. Palinski , M. E. Rosenfeld , S. Parthasarathy , T. E. Carew , S. Butler , J. L. Witztum , and D. Steinberg . 1989 . Evidence for the presence of oxidatively modifi ed low density lipo-protein in atherosclerotic lesions of rabbit and man. J. Clin. Invest. 84 : 1086 – 1095 .
oxysterols may have differential effects on the lipid pack-ing of the membrane and therefore differ in their ability to disrupt the structure of lipid-ordered domains. More studies, however, are needed to test this hypothesis. We also show that enriching the cells with cholesterol prior to the exposure to oxysterols may protect endothelial cells against the detrimental effect of oxysterols on en-dothelial stiffness. Note that enriching endothelial cells with cholesterol by itself has no effect on endothelial stiffness, indicating that neither the reversal of oxysterol-induced stiffening nor the protective effect of choles-terol enrichment can be attributed to a simple mutual cancelation of the two opposite effects. We suggest that the protective effect of cholesterol enrichment can be due to diminishing the incorporation of other sterols into the membrane. Taken together, our observations suggest that, although incorporation of oxysterols into endothelial membranes alters endothelial biomechanics and is expected to be detrimental to endothelial func-tion, increasing the level of membrane cholesterol may be benefi cial and protective.
Fig. 8. Oxysterol accumulation in aortic tissue of ApoE � / � mice. Inset: Levels of total cholesterol in plasma of male (n = 10) and fe-male (n = 4) mice. A: Sterol levels in the vascular walls of aortas harvested from male WT and ApoE � / � mice sacrifi ced at 10–12 weeks. The levels of individual sterols are expressed as a percent-age of total sterol content in the same sample. B: Sterol levels in the vascular walls of aortas harvested from age-matched female WT and ApoE � / � mice. N = 4–5 animals in each experimental group. Values are means ± SEM, * P < 0.05 vs. WT.
by guest, on April 6, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2012/04/19/jlr.M027102.DC1Supplemental Material can be found at:

oxLDL and endothelial biomechanics 1357
35 . Subbaiah, P. V. , V. S. Subramanian , and K. Wang . 1999. Novel phys-iological function of sphingomyelin in plasma. J. Biol. Chem. 274 : 36409 – 36414 .
36 . Bligh , E. G. , and W. J. Dyer . 1959 . A rapid method of total lipid extractment and purifi cation. Can. J. Biochem. Physiol. 37 : 911 – 917 .
37 . Allen , R. J. 1940 . The estimation of phosphorus. Biochem. J. 34 : 858 – 865 .
38 . Subbaiah , P. V. , D. Sircar , R. S. Lankalapalli , and R. Bittman . 2009 . Effect of double bond geometry in sphingosine base on the an-tioxidant function of sphingomyelin. Arch. Biochem. Biophys. 481 : 72 – 79 .
39 . Brown , A. J. , R. T. Dean , and W. Jessup . 1996 . Free and esterifi ed oxysterol: formation during copper-oxidation of low density lipo-protein and uptake by macrophages. J. Lipid Res. 37 : 320 – 335 .
40 . Titushkin , I. , and M. Cho . 2007 . Modulation of cellular mechan-ics during osteogenic differentiation of human mesenchymal stem cells. Biophys. J. 93 : 3693 – 3702 .
41 . Byfi eld , F. J. , H. Aranda-Aspinoza , V. G. Romanenko , G. H. Rothblat , and I. Levitan . 2004 . Cholesterol depletion increases membrane stiffness of aortic endothelial cells. Biophys. J. 87 : 3336 – 3343 .
42 . Byfi eld , F. J. , B. D. Hoffman , V. G. Romanenko , Y. Fang , J. C. Crocker , and I. Levitan . 2006 . Evidence for the role of cell stiffness in modulation of volume-regulated anion channels. Acta Physiol. (Oxf.) . 187 : 285 – 294 .
43 . Lenz, M. L. , H. Hughes , J. R. Mitchell , D. P. Via , J. R. Guyton , A. A. Taylor , A. M. Gotto , Jr., and C. V. Smith. 1990. Lipid hydroperoxy and hydroxy derivatives in copper-catalyzed oxidation of low den-sity lipoprotein. J. Lipid Res. 31 : 1043 – 1050 .
44 . Dzeletovic , S. , A. Babiker , E. Lund , and U. Diczfalusy . 1995 . Time course of oxysterol formation during in vitro oxidation of low den-sity lipoprotein. Chem. Phys. Lipids . 78 : 119 – 128 .
45 . Schuh, J. , F. F. Gordon , and R. H. Haschemeyer . 1978. Oxygen-mediated heterogeneity of apo-low-density lipoprotein. Proc. Natl. Acad. Sci. USA . 75 : 3173 – 3177 .
46 . Thorne , S. A. , S. E. Abbot , P. G. Winyard , D. R. Blake , and P. G. Mills . 1996 . Extent of oxidative modifi cation of low density lipo-protein determines the degree of cytotoxicity to human coronary artery cells. Heart . 75 : 11 – 16 .
47 . Sigari, F. , C. Lee , J. L. Witztum , and P. D. Reaven . 1997. Fibroblasts that overexpress 15-Lipoxygenase generate bioactive and minimally modifi ed LDL. Arterioscler. Thromb. Vasc. Biol. 17 : 3639 – 3645 .
48 . Bloom , R. J. , and J. C. Elwood . 1981 . Quantitation of lipid profi les from isolated serum lipoproteins using small volumes of human serum. Clin. Biochem. 14 : 119 – 125 .
49 . Cuzner , M. L. , and A. N. Davison . 1967 . Quantitative thin layer chromatography of lipids. J. Chromatogr. 27 : 388 – 397 .
50 . Björkhem , I. , O. Andersson , U. Diczfalusy , B. Sevastik , R. J. Xiu , C. Duan , and E. Lund . 1994 . Atherosclerosis and sterol 27-hydroxylase: evidence for a role of this enzyme in elimination of cholesterol from human macrophages. Proc. Natl. Acad. Sci. USA . 91 : 8592 – 8596 .
51 . Crisby, M. , J. Nilsson , V. Kostulas , I. Bjökhem , and U. Diczfalusy . 1997. Localization of sterol 27-hydroxylase immuno-reactivity in hu-man atherosclerotic plaques. Biochim. Biophys. Acta . 1344 : 278 – 285 .
52 . Nakashima, Y. , A. S. Plump , E. W. Raines , J. L. Breslow , and R. Ross . 1994. ApoE-defi cient mice develop lesions of all phases of atherosclerosis throughout the arterial tree. Arterioscler. Thromb. 14 : 133 – 140 .
53 . Breslow, J. L . 1996. Mouse models of atherosclerosis. Science . 272 : 685 – 688 .
54 . Chen, C. S. , M. Mrksich , S. Huang , G. M. Whitesides , and D. E. Ingber . 1997. Geometric control of cell life and death. Science . 276 : 1425 – 1428 .
55 . Sato , M. , K. Nagayama , N. Kataoka , M. Sasaki , and K. Hane . 2000 . Local mechanical properties measured by atomic force micros-copy for cultured bovine endothelial cells exposed to shear stress. J. Biomech. 33 : 127 – 135 .
56 . Sieminski , A. L. , R. P. Hebbel , and K. J. Gooch . 2004 . The rela-tive magnitudes of endothelial force generation and matrix stiff-ness modulate capillary morphogenesis in vitro. Exp. Cell Res. 297 : 574 – 584 .
57 . Oberleithner , H. , C. Riethmuller , H. Schillers , G. A. MacGregor , H. E. de Wardener , and M. Hausberg . 2007 . Plasma sodium stiffens vascular endothelium and reduces nitric oxide release. Proc. Natl. Acad. Sci. USA . 104 : 16281 – 16286 .
58 . Jialal , I. , and S. Devaraj . 1996 . The role of oxidized low density lipo-protein in atherogenesis. J. Nutr. 126 : 1053S – 1057S .
12 . Shimokawa , H. 1999 . Primary endothelial dysfunction: atheroscle-rosis. J. Mol. Cell. Cardiol. 31 : 23 – 37 .
13 . Kinlay , S. , P. Libby , and P. Ganz . 2001 . Endothelial function and coronary artery disease. Curr. Opin. Lipidol. 12 : 383 – 389 .
14 . Sima , A. V. , C. S. Stancu , and M. Simionescu . 2009 . Vascular en-dothelium in atherosclerosis. Cell Tissue Res. 335 : 191 -203.
15 . Gonzalez, M. A., and A. P. Selwyn . 2003. Endothelial function, in-fl ammation, and prognosis in cardiovascular disease. Am. J. Med. 115(Suppl. 8A): 99S – 106S .
16 . Heitzer, T. , T. Schlinzig , K. Krohn , T. Meinertz , and T. Munzel . 2001. Endothelial dysfunction, oxidative stress, and risk of cardio-vascular events in patients with coronary artery disease. Circulation . 104 : 2673 – 2678 .
17 . Schächinger , V. , M. B. Britten , and A. M. Zeiher . 2000 . Prognostic impact of coronary vasodilator dysfunction on adverse long-term outcome of coronary disease. Circulation . 101 : 1899 – 1906 .
18 . Byfi eld, F. J. , S. Tikku , G. H. Rothblat , K. J. Gooch , and I. Levitan . 2006. OxLDL increases endothelial stiffness, force generation and network formation. J. Lipid Res. 47 : 715 – 723 .
19 . Shentu, T. P. , I. Titushkin , D. K. Singh , K. J. Gooch , P. V. Subbaiah , M. Cho , and I. Levitan . 2010. oxLDL-induced decrease in lipid or-der of membrane domains is inversely correlated with endothelial stiffness and network formation. Am. J. Physiol. Cell Physiol. 299 : C218 –C 229 .
20 . Kowalsky, G. B. , F. J. Byfi eld , and I. Levitan . 2008. oxLDL facilitates fl ow-induced realignment of aortic endothelial cells. Am. J. Physiol. Cell. Physiol. 295 : C332 –C 340 .
21 . Levitan , I. , and T. P. Shentu . 2011 . Impact of oxLDL on cholesterol-rich membrane rafts. J. Lipids . 2011 : 730209 .
22 . Esterbauer , H. , J. Gebicki , H. Puhl , and G. Jürgens . 1992 . The role of lipid peroxidation and antioxidants in oxidative modifi cation of LDL. Free Radic. Biol. Med. 13 : 341 –390.
23 . Itabe, H. , M. Mori , Y. Fujimoto , Y. Higashi , and T. Takano . 2003. Minimally modifi ed LDL is an oxidized LDL enriched with oxi-dized phosphatidylcholines. Journal of Biochemistry . 134 : 459 – 465 .
24 . Poli , G. , B. Sottero , S. Gargiulo , and G. Leonarduzzi . 2009 . Cholesterol oxidation products in the vascular remodeling due to atheroslcerosis. Mol. Aspects Med. 30 : 180 – 189 .
25 . Brown , A. J. , and W. Jessup . 1999 . Oxysterols and atherosclerosis. Atherosclerosis . 142 : 1 – 28 .
26 . Deckert, V. , L. Persegol , L. Viens , G. Lizard , A. Athias , C. Lallemant , P. Gambert , and L. Lagrost. 1997. Inhibitors of arterial relaxation among components of human oxidized low-density lipoproteins: cholesterol derivatives oxidized in position 7 are potent inhibitors of endothelium-dependent relaxation. Circulation . 95 : 723 – 731 .
27 . Leonarduzzi, G. , B. Vizio , B. Sottero , V. Verde , P. Gamba , C. Mascia , E. Chiarpotto , G. Poli , and F. Biasi . 2006. Early involvement of ROS overproduction in apoptosis induced by 7-ketocholesterol. Antioxid. Redox Signal. 8 : 375 – 380 .
28 . Ramasamy, S. , G. A. Boissonneault , and B. Hennig . 1992. Oxysterol-induced endothelial cell dysfunction in culture. J. Am. Col. Nutr. 11 : 532 – 538 .
29 . Lizard , G. , M. Moisant , C. Cordelet , S. Monier , P. Gambert , and L. Lagrost . 1997 . Induction of similar features of apoptosis in human and bovine vascular endothelial cells treated by 7-ketocholesterol. J. Pathol. 183 : 330 – 338 .
30 . Panini , S. R. , and M. S. Sinensky . 2001 . Mechanisms of oxysterol-induced apoptosis. Curr. Opin. Lipidol. 12 : 529 – 533 .
31 . Moreau, M. , I. Brocheriou , L. Petit , E. Ninio , M. J. Chapman , and M. Rouis . 1999. Interleukin-8 mediates downregulation of tissue in-hibitor of metalloproteinase-1 expression in cholesterol-loaded hu-man macrophages. Relevance to stability of atherosclerotic plaque. Circulation . 99 : 420 – 426 .
32 . Gargiulo , S. , B. Sottero , P. Gamba , E. Chiarpotto , G. Poli , and G. Leonarduzzi . 2011 . Plaque oxysterols induce unbalanced up-regu-lation of matrix metalloproteinase-9 in macrophagic cells through redox-sensitive signaling pathways: Implications regarding the vulnerability of atherosclerotic lesions. Free Radic. Biol. Med. 51 : 844 – 855 .
33 . Levitan , I. , A. E. Christian , T. N. Tulenko , and G. H. Rothblat . 2000 . Membrane cholesterol content modulates activation of vol-ume-regulated anion current (VRAC) in bovine endothelial cells. J. Gen. Physiol. 115 : 405 – 416 .
34 . Cathcart, M. K. , A. K. McNally , and G. M. Chisolm . 1991. Lipoxygenase-mediated transformation of human low density lipoprotein to an oxidized and cytotoxic complex. J. Lipid Res. 32 : 63 – 70 .
by guest, on April 6, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2012/04/19/jlr.M027102.DC1Supplemental Material can be found at:

1358 Journal of Lipid Research Volume 53, 2012
59 . Berliner , J. A. , M. C. Territo , A. Sevanian , S. Ramin , J. A. Kim , B. Bamshad , M. Esterson , and A. M. Fogelman . 1990 . Minimally modi-fi ed low density lipoprotein stimulates monocyte endothelial inter-actions. J. Clin. Invest. 85 : 1260 – 1266 .
60 . Colles, S. M. , J. M. Maxson , S. G. Carlson , and G. M. Chisolm . 2001. Oxidized LDL-induced injury and apoptosis in atheroscle-rosis. Potential roles for oxysterols. Trends Cardiovasc. Med. 11 : 131 – 138 .
61 . Hakamata , H. , A. Miyazaki , M. Sakai , H. Matsuda , H. Suzuki , T. Kodama , and S. Horiuchi . 1998 . Isolation of macrophage-like cell mutants resistant to the cytotoxic effect of oxidized low density lipo-protein. J. Lipid Res. 39 : 482 – 494 .
62 . Miller, Y. I. , D. S. Worrall , C. D. Funk , J. R. Feramisco , and J. L. Witztum . 2003. Actin polymerization in macrophages in response to oxidized LDL and apoptotic cells: role of 12/15-lipoxygenase and phosphoinositide 3-kinase. Mol. Biol. Cell . 14 : 4196 – 4206 .
63 . Wong , W. T. , C. H. Ng , S. Y. Tsang , Y. Huang , and Z. Y. Chen . 2011 . Relative contribution of individual oxidized components in ox-LDL to inhibition on endothelium-dependent relaxation in rat aorta. Nutr. Metab. Cardiovasc. Dis. 21 : 157 – 164 .
64 . Sleer , L. S. , A. J. Brown , and E. R. Stanley . 2001 . Interaction of caveolin with 7-ketocholesterol. Atherosclerosis . 159 : 49 – 55 .
65 . Vaya , J. , M. Aviram , S. Mahmood , T. Hayek , E. Grenadir , A. Hoffman , and S. Milo . 2001 . Selective distribution of oxysterols in atherosclerotic lesions and human plasma lipoproteins. Free Radic. Res. 34 : 485 – 497 .
66 . Pereira , T. M. , B. V. Nogueira , L. C. Lima , M. L. Porto , J. A. Arruda , E. C. Vasquez , and S. S. Meyrelles . 2010 . Cardiac and vascular changes in elderly atherosclerotic mice: the infl uence of gender. Lipids Health Dis. 9 : 87 .
67 . Vitale , C. , M. E. Mendelsohn , and G. M. Rosano . 2009 . Gender differences in the cardiovascular effect of sex hormones. Nat. Rev. Cardiol. 6 : 532 – 542 .
by guest, on April 6, 2018
ww
w.jlr.org
Dow
nloaded from
.html http://www.jlr.org/content/suppl/2012/04/19/jlr.M027102.DC1Supplemental Material can be found at:

![Markers of subclinical atherosclerosis in patients with ... · impaired endothelial function[16, 17], and increased arterial stiffness[18] in patients with aortic valve sclerosis](https://img.pdfslide.net/doc/110x75/5fae2d024f851c134b38d58b/markers-of-subclinical-atherosclerosis-in-patients-with-impaired-endothelial.jpg)















![Free Radical Biology and Medicine · Oxysterols and their downstream metabolites, including choles- ... atherosclerosis [9,10], and neurodegenerative disease [11]. Oxysterols can](https://img.pdfslide.net/doc/110x75/5e5c3862ee70ba0b5c06ca26/free-radical-biology-and-medicine-oxysterols-and-their-downstream-metabolites-including.jpg)











