Embed Size (px)
Citation preview

Gene 552 (2014) 249–254
Contents lists available at ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r .com/ locate /gene
Methods paper
The role of phosphoenolpyruvate carboxykinase in neuronalsteroidogenesis under acute inflammation
Mohanraj Sadasivam, Balamurugan Ramatchandirin, Sivasangari Balakrishnan,Karthikeyan Selvaraj, Chidambaram Prahalathan ⁎Department of Biochemistry, Bharathidasan University, Tiruchirappalli 620 024, India
Abbreviations: PEPCK, phosphoenolpyruvate carboxykregulatory protein; 3β-HSD, 3β-hydroxysteroid dehhydroxysteroid dehydrogenase; LPS, lipopolysaccharide.⁎ Corresponding author.
E-mail address: [email protected] (C. Prahalathan
http://dx.doi.org/10.1016/j.gene.2014.09.0430378-1119/© 2014 Elsevier B.V. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 28 July 2014Received in revised form 10 September 2014Accepted 19 September 2014Available online 23 September 2014
Keywords:Bacterial lipopolysaccharidePhosphoenolpyruvate carboxykinaseBrainSteroidogenesisGlipizide
Phosphoenolpyruvate carboxykinase (PEPCK) is a key gluconeogenic enzyme found inmany tissues throughoutthe body including brain. In the present study, we have investigated the effect of bacterial lipopolysaccharide(LPS) on PEPCK and its role in neuronal steroidogenesis. Adult female albino rats were administered LPS(5 mg/kg body weight) to induce acute inflammation. LPS administration resulted in a significant increase ofPEPCK mRNA expression with concomitant increase in mRNA levels of steroidogenic acute regulatory (StAR)protein and other steroidogenic enzymes including 3β-hydroxysteroid dehydrogenase (3β-HSD), 17β-hydroxysteroid dehydrogenase (17β-HSD) and aromatase in brain tissue. Further, the inhibition of PEPCKexpression by glipizide significantly decreased the mRNA expression of steroidogenic proteins and concurrentlyincreased the mRNA levels of proinflammatory cytokines under LPS administration. The results of this studysuggest a novel finding that PEPCK may have an important role in neuronal steroidogenesis; which serves asan adaptive response under inflammation.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
The brain synthesizes significant amounts of sex steroids besidestestis and ovary. Steroid hormones in the brain exert important biolog-ical functions including neuroprotection, regulation of gene expression,neuronal survival, synaptic transmission, and neuronal and glial differ-entiation in many brain areas (Saldanha et al., 2009; Garcia-Segura,2008; Stoffel-Wagner et al., 1999). Bacterial lipopolysaccharide (LPS)is a major component of the cell wall of gram negative bacteria contrib-uting to the pathogenesis of bacterial infection, in particular to thosediseases affecting central nervous system including cerebral malariaand bacterial meningitis. Proinflammatory cytokines generated inresponse to LPS exposure inhibit steroidogenesis in many tissue typesincluding the ovary and testes (Magata et al., 2014; Reddy et al.,2006). In contrast to the ovary and testes, neurosteroid levels are in-creased in vivo after LPS treatment and result in the suppression ofTNF production in brain (Ghezzi et al., 2000). LPS increases the expres-sion and activity of 17β-hydroxysteroid dehydrogenase (17β-HSD) inbrain microglia (Gottfried-Blackmore et al., 2008). However, the
inase; StAR, steroidogenic acuteydrogenase; 17β-HSD, 17β-
).
mechanisms behind the increased production of neurosteroids in in-flammatory conditions are not well understood.
Phosphoenolpyruvate carboxykinase (PEPCK) is found in many tis-sues throughout the body and catalyzes the conversion of oxaloacetateto phosphoenolpyruvate in gluconeogenesis. Besides its role in gluconeo-genesis, PEPCK has a number of other physiological roles such asglyceroneogenesis, anaplerosis and cataplerosis (Hanson, 2009). Most re-cently, it has been demonstrated that PEPCK and glucose-6-phosphatase(Glc-6-Pase) are required for steroidogenesis in testicular Leydig cells(Ahn et al., 2012). Further, Sugita et al. (2002) have shown that PEPCKmRNA expression levels were increased in liver in LPS administeredrats. Interestingly, PEPCK is also expressed in the brain; however its phys-iological role in neurons is not well studied. With this background, thepresent work was designed to check whether PEPCK has any role in neu-ronal steroidogenesis under LPS induced acute infection.
2. Materials and methods
2.1. Chemicals
Lipopolysaccharide from Escherichia coli (serotype 055:B5)was pur-chased from Sigma Chemicals Company, Saint Louis, MO, USA. All of theother chemicals used were of analytical grade and were obtained fromSisco Research Laboratories Pvt. Ltd., Mumbai, India and HiMedia Labo-ratories Pvt. Ltd., Mumbai, India.

Table 1Primer sequences for PCR for each target gene.
Gene Primer
Steroidogenic acute regulatory (StAR)protein
F: 5′-AGCCAGCAGGAGAATGGAGAT-3′R: 5′-CACCTCCAGTCGGAACACCTT-3′
3β-HSD F: 5′-AACTGGTCTTCAGGTCACCAGAA-3′R: 5′-GTCCCCTGCACCTTGTTCA-3′
17β-HSD F: 5′-CCTTTGGCTTTGCCATGAGA-3′R: 5′-CAATCCATCCTGCTCCAACCT-3′
Aromatase F: 5′-TCCTCCTGATTCGGAATTGTG-3′R: 5′-GGCCCGATTCCCAGACA-3′
CREB-binding protein/E1A bindingprotein (CBP/p300)
F: 5′-TGCAGTCTGCTACTACTGCG-3′R: 5′-AAGAGCCGTATGCCAAGGTC-3′
PEPCK F: 5′-TCCCATTGGCTACGTCCCT-3′R: 5′-CAGCATTGTGCTTGCTGGTT-3′
Tumor necrosis factor alpha (TNF-α) F: 5′-TTCCTTACGGAACCCCCTCT-3′R: 5′-CCCGTAGGGCGATTACAGTC-3′
Interleukin-6 (IL-6) F: 5′-CACTTCACAAGTCGGAGGCT-3′R: 5′-AGAGCATTGGAAGTTGGGGT-3′
β-Actin F: 5′-TTCAACACCCCAGCCATGT-3′R: 5′-TGGTACGACCAGAGGCATACAG-3′
250 M. Sadasivam et al. / Gene 552 (2014) 249–254
2.2. Experimental animals
Adult female albino rats of Wistar strain weighing 220 ± 10 g(10–12 weeks old) were used in the study. The animals were main-tained under standard conditions of humidity, temperature (25 ±2 °C), and light (12 h light/12 h dark). The animals were housed inlarge spacious cages bedded with husk and fed with a standard ratpelleted diet and had free access to water. Experimental animals werehandled according to the university and institutional legislation, regu-lated by the Committee for the Purpose of Control and Supervision ofExperiments on Animals (CPCSEA), Ministry of Social Justice andEmpowerment, Government of India.
2.3. Study design
The animals were randomly divided into three groups of six ratseach. Saline treated animals served as controls (Group I). Group II(LPS) received intraperitoneal (i.p.) injections of LPS (dissolved in0.5 ml of sterile saline) at a dose of 5 mg/kg body weight to induceacute inflammation. Group III (LPS + glipizide) received a single intra-venous injection of glipizide (5 mg/kg bodyweight); 1 h prior to the ad-ministration of intraperitoneal injections of LPS (5 mg/kg bodyweight).The animals were fasted 12 h before commencement and also duringthe experimental period. The blood was collected and the brains wereexcised immediately after 24 h of treatment period.
2.4. Preparation of tissue homogenate
Briefly, the tissue was chopped and minced in 6–8 ml of 0.25 M su-crose/TKM buffer (50 mM Tris–HCl, 25 mM KCl and 5 mMMgCl2). Thetissue was then homogenized gently by using Potter-Elvehjem homog-enizer. The homogenate was centrifuged at 1500 rpm for 5 min. Thesupernatant was transferred and used as whole tissue homogenate forenzymatic analyses.
2.5. Estimation of protein
Protein content of the tissue fractions was estimated by the methodof Lowry et al. (1951). Briefly, 0.1 ml of the diluted tissue homogenatewas made up to 1 ml with water. To this, 4.5 ml of alkaline copper re-agent (2% Na2CO3 in 0.1 NNaOHwasmixedwith 0.5% CuSO4 containing1% sodium potassium tartrate in the ratio of 50:1) was added, mixedand allowed to stand at room temperature for 20 min. Later, 0.5 ml ofFolin–Ciocalteu reagent was added and shaken well. The blank andbovine serum albumin (BSA) standards were also treated in a similarmanner. The blue color complex formed was measured at 640 nmafter 15 min against the blank.
2.6. Assay of steroidogenic enzymes
2.6.1. 3β-Hydroxysteroid dehydrogenase (3β-HSD)The activity of 3β-HSD was measured by the method of
Shivanandappa and Venkatesh (1997). The enzyme was assayed in 0.1M Tris–HCl buffer (pH 7.8) containing 500 μMNAD+, 0.5 ml of color re-agent (1% Tween 20 containing 0.08% iodonitrotetrazolium chloride)and the substrate (100 μM pregnenolone) in a total volume of 3 ml.The reaction was started by adding the enzyme (50 μl) and incubatedat 37 °C for 60 min. The reaction was stopped by the addition of 2 mlof phthalate buffer (2.55 g of potassium hydrogen phthalate dissolvedin a mixture of 51 ml 0.1 N HCl and 2.5 ml Tween 20; pH was adjustedto 3.0 and the volume made up to 250 ml). The turbidity was removedby centrifugation at 3000 rpm for 20 min and the supernatant was readat 490 nm in a spectrophotometer. For standard curve, aliquots ofgraded concentrations of freshly prepared NADH (0–150 nM) werereacted with 0.5 ml of color reagent (1% Tween 20 containing 0.08%iodonitrotetrazolium chloride and 0.02% phenazine methosulfate)
and after color formed, 2 ml of phthalate buffer was added and theabsorbance was read at 490 nm. The enzyme activity was calculatedfrom the standard curve of NADH and expressed as nmol of NADHformed/min/mg protein.
2.6.2. 17β-Hydroxysteroid dehydrogenase (17β-HSD)The activity of 17β-HSDwas determined by themethod as described
previously (Bergmeyer, 1974). 17β-HSD catalyzes the reversible reac-tion of androstenedione into testosterone using NADPH as a coenzyme.The activity is determined by the optical measurement of the rate ofconversion of NADPH to NADP. In brief, the reaction mixture contained100 μl of testicular supernatant, 200 μl of 0.5 μM NADPH, and 100 μl of0.8 μM androsten-3,17-dione in a final volume of 3 ml 100 μM phos-phate buffer solution (pH 7.4). The reaction was initiated by the addi-tion of the substrate and the decrease in absorbance of NADPH wasfollowed at 340 nm for 5 min at 20 s interval. The enzyme activity wasexpressed as nmol of NADPH oxidized/min/mg protein.
2.7. Estimation of blood glucose
Blood glucose levels were measured with OneTouch Select bloodglucose monitoring system (LifeScan, Inc., USA).
2.8. Analysis of the gene expression by reverse transcriptase polymerasechain reaction (RT-PCR)
Briefly, total RNA from whole brain was isolated according to theRNA isolation kit instructions (One step RNA Trizol Reagent; BiobasicInc., Canada). RT-PCR was performed with 4 μg of total RNA isolatedfrom tissue by using AMV-one step RT-PCR kit (GeNei, India). The spe-cific sets of primers for target genes are shown in Table 1. PCR amplifi-cation was carried out according to a protocol for the initialdenaturing step at 95 °C for 10 min; then 30 cycles at 95 °C for 1 min(denaturing), at 55 °C for 1 min (annealing) and 72 °C for 1.5 min (ex-tension); and a further extension at 72 °C for 10 min. To compare theamount of steady state mRNA, 5 μl of each PCR product was resolvedonto 1.4% agarose gel using TBE buffer. After electrophoresis, the gelswere viewed under UV light and digital images were captured onGelstan gel documentation system. The densitometric analyses werecarried out with lab image platform ver 2.1 software by Kapelan Bio-Imaging GmbH. The expression of each target gene was normalizedwith internal control and represented as a ratio.
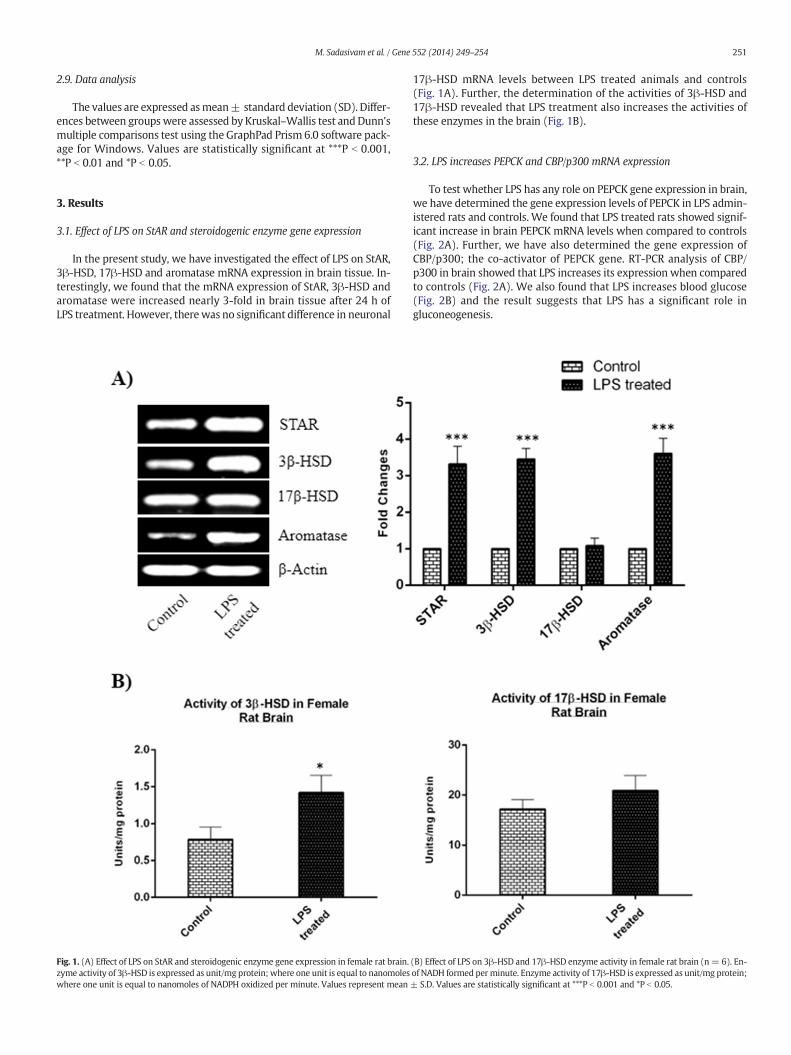
251M. Sadasivam et al. / Gene 552 (2014) 249–254
2.9. Data analysis
The values are expressed asmean± standard deviation (SD). Differ-ences between groupswere assessed by Kruskal–Wallis test andDunn'smultiple comparisons test using the GraphPad Prism 6.0 software pack-age for Windows. Values are statistically significant at ***P b 0.001,**P b 0.01 and *P b 0.05.
3. Results
3.1. Effect of LPS on StAR and steroidogenic enzyme gene expression
In the present study, we have investigated the effect of LPS on StAR,3β-HSD, 17β-HSD and aromatase mRNA expression in brain tissue. In-terestingly, we found that the mRNA expression of StAR, 3β-HSD andaromatase were increased nearly 3-fold in brain tissue after 24 h ofLPS treatment. However, therewas no significant difference in neuronal
Fig. 1. (A) Effect of LPS on StAR and steroidogenic enzyme gene expression in female rat brain.zyme activity of 3β-HSD is expressed as unit/mg protein; where one unit is equal to nanomoleswhere one unit is equal to nanomoles of NADPH oxidized per minute. Values represent mean
17β-HSD mRNA levels between LPS treated animals and controls(Fig. 1A). Further, the determination of the activities of 3β-HSD and17β-HSD revealed that LPS treatment also increases the activities ofthese enzymes in the brain (Fig. 1B).
3.2. LPS increases PEPCK and CBP/p300 mRNA expression
To test whether LPS has any role on PEPCK gene expression in brain,we have determined the gene expression levels of PEPCK in LPS admin-istered rats and controls. We found that LPS treated rats showed signif-icant increase in brain PEPCK mRNA levels when compared to controls(Fig. 2A). Further, we have also determined the gene expression ofCBP/p300; the co-activator of PEPCK gene. RT-PCR analysis of CBP/p300 in brain showed that LPS increases its expression when comparedto controls (Fig. 2A). We also found that LPS increases blood glucose(Fig. 2B) and the result suggests that LPS has a significant role ingluconeogenesis.
(B) Effect of LPS on 3β-HSD and 17β-HSD enzyme activity in female rat brain (n = 6). En-of NADH formed perminute. Enzyme activity of 17β-HSD is expressed as unit/mg protein;± S.D. Values are statistically significant at ***P b 0.001 and *P b 0.05.
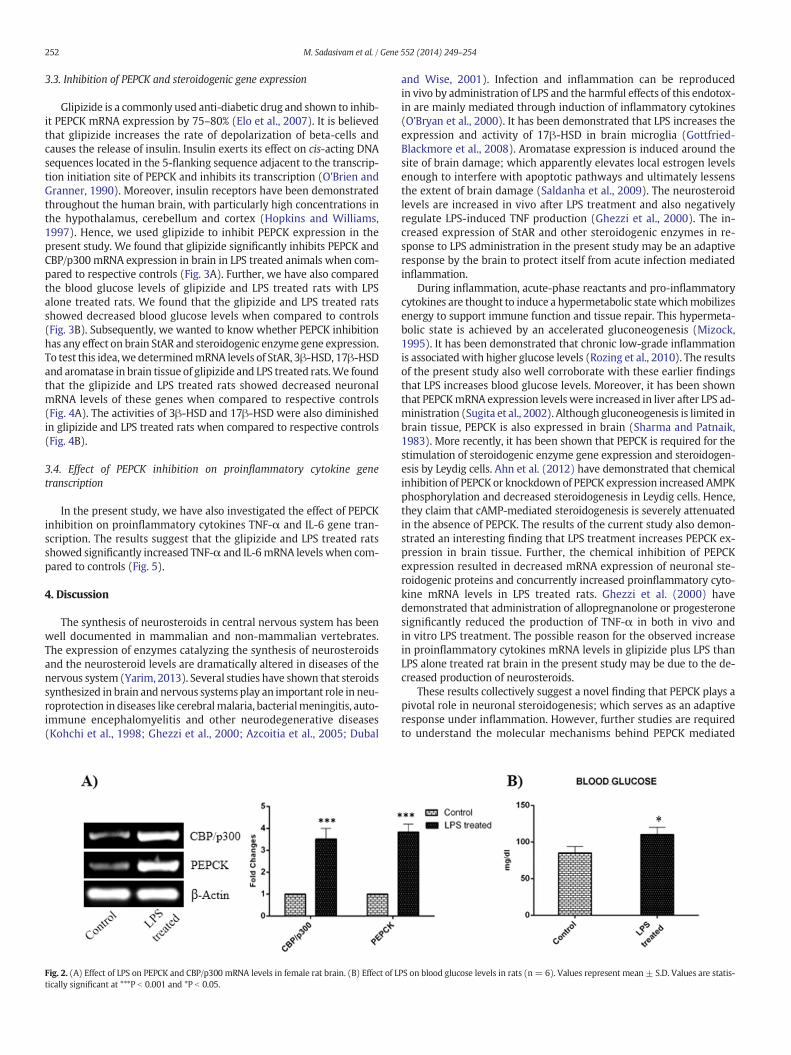
252 M. Sadasivam et al. / Gene 552 (2014) 249–254
3.3. Inhibition of PEPCK and steroidogenic gene expression
Glipizide is a commonly used anti-diabetic drug and shown to inhib-it PEPCK mRNA expression by 75–80% (Elo et al., 2007). It is believedthat glipizide increases the rate of depolarization of beta-cells andcauses the release of insulin. Insulin exerts its effect on cis-acting DNAsequences located in the 5-flanking sequence adjacent to the transcrip-tion initiation site of PEPCK and inhibits its transcription (O'Brien andGranner, 1990). Moreover, insulin receptors have been demonstratedthroughout the human brain, with particularly high concentrations inthe hypothalamus, cerebellum and cortex (Hopkins and Williams,1997). Hence, we used glipizide to inhibit PEPCK expression in thepresent study. We found that glipizide significantly inhibits PEPCK andCBP/p300mRNA expression in brain in LPS treated animals when com-pared to respective controls (Fig. 3A). Further, we have also comparedthe blood glucose levels of glipizide and LPS treated rats with LPSalone treated rats. We found that the glipizide and LPS treated ratsshowed decreased blood glucose levels when compared to controls(Fig. 3B). Subsequently, we wanted to know whether PEPCK inhibitionhas any effect on brain StAR and steroidogenic enzyme gene expression.To test this idea,we determinedmRNA levels of StAR, 3β-HSD, 17β-HSDand aromatase in brain tissue of glipizide and LPS treated rats.We foundthat the glipizide and LPS treated rats showed decreased neuronalmRNA levels of these genes when compared to respective controls(Fig. 4A). The activities of 3β-HSD and 17β-HSD were also diminishedin glipizide and LPS treated rats when compared to respective controls(Fig. 4B).
3.4. Effect of PEPCK inhibition on proinflammatory cytokine genetranscription
In the present study, we have also investigated the effect of PEPCKinhibition on proinflammatory cytokines TNF-α and IL-6 gene tran-scription. The results suggest that the glipizide and LPS treated ratsshowed significantly increased TNF-α and IL-6mRNA levels when com-pared to controls (Fig. 5).
4. Discussion
The synthesis of neurosteroids in central nervous system has beenwell documented in mammalian and non-mammalian vertebrates.The expression of enzymes catalyzing the synthesis of neurosteroidsand the neurosteroid levels are dramatically altered in diseases of thenervous system (Yarim, 2013). Several studies have shown that steroidssynthesized in brain and nervous systemsplay an important role in neu-roprotection in diseases like cerebralmalaria, bacterialmeningitis, auto-immune encephalomyelitis and other neurodegenerative diseases(Kohchi et al., 1998; Ghezzi et al., 2000; Azcoitia et al., 2005; Dubal
Fig. 2. (A) Effect of LPS on PEPCK and CBP/p300 mRNA levels in female rat brain. (B) Effect of Ltically significant at ***P b 0.001 and *P b 0.05.
and Wise, 2001). Infection and inflammation can be reproducedin vivo by administration of LPS and the harmful effects of this endotox-in are mainly mediated through induction of inflammatory cytokines(O'Bryan et al., 2000). It has been demonstrated that LPS increases theexpression and activity of 17β-HSD in brain microglia (Gottfried-Blackmore et al., 2008). Aromatase expression is induced around thesite of brain damage; which apparently elevates local estrogen levelsenough to interfere with apoptotic pathways and ultimately lessensthe extent of brain damage (Saldanha et al., 2009). The neurosteroidlevels are increased in vivo after LPS treatment and also negativelyregulate LPS-induced TNF production (Ghezzi et al., 2000). The in-creased expression of StAR and other steroidogenic enzymes in re-sponse to LPS administration in the present study may be an adaptiveresponse by the brain to protect itself from acute infection mediatedinflammation.
During inflammation, acute-phase reactants and pro-inflammatorycytokines are thought to induce a hypermetabolic statewhichmobilizesenergy to support immune function and tissue repair. This hypermeta-bolic state is achieved by an accelerated gluconeogenesis (Mizock,1995). It has been demonstrated that chronic low-grade inflammationis associated with higher glucose levels (Rozing et al., 2010). The resultsof the present study also well corroborate with these earlier findingsthat LPS increases blood glucose levels. Moreover, it has been shownthat PEPCKmRNA expression levels were increased in liver after LPS ad-ministration (Sugita et al., 2002). Although gluconeogenesis is limited inbrain tissue, PEPCK is also expressed in brain (Sharma and Patnaik,1983). More recently, it has been shown that PEPCK is required for thestimulation of steroidogenic enzyme gene expression and steroidogen-esis by Leydig cells. Ahn et al. (2012) have demonstrated that chemicalinhibition of PEPCK or knockdown of PEPCK expression increased AMPKphosphorylation and decreased steroidogenesis in Leydig cells. Hence,they claim that cAMP-mediated steroidogenesis is severely attenuatedin the absence of PEPCK. The results of the current study also demon-strated an interesting finding that LPS treatment increases PEPCK ex-pression in brain tissue. Further, the chemical inhibition of PEPCKexpression resulted in decreased mRNA expression of neuronal ste-roidogenic proteins and concurrently increased proinflammatory cyto-kine mRNA levels in LPS treated rats. Ghezzi et al. (2000) havedemonstrated that administration of allopregnanolone or progesteronesignificantly reduced the production of TNF-α in both in vivo andin vitro LPS treatment. The possible reason for the observed increasein proinflammatory cytokines mRNA levels in glipizide plus LPS thanLPS alone treated rat brain in the present study may be due to the de-creased production of neurosteroids.
These results collectively suggest a novel finding that PEPCK plays apivotal role in neuronal steroidogenesis; which serves as an adaptiveresponse under inflammation. However, further studies are requiredto understand the molecular mechanisms behind PEPCK mediated
PS on blood glucose levels in rats (n= 6). Values represent mean ± S.D. Values are statis-
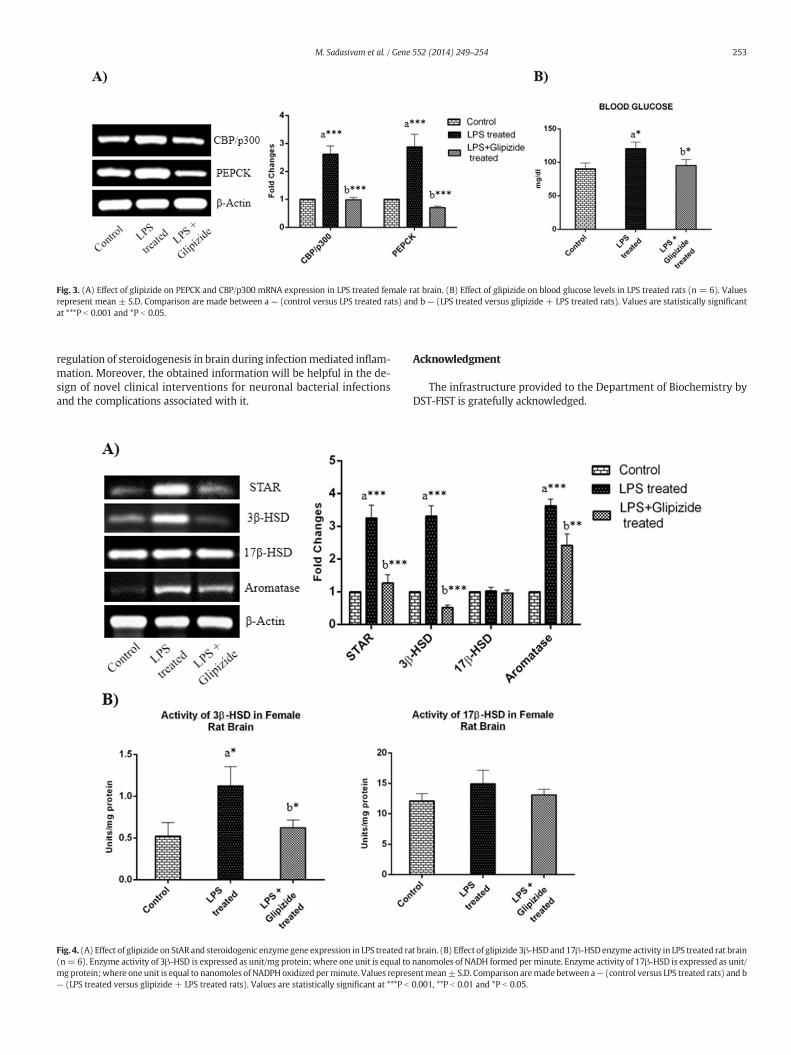
Fig. 3. (A) Effect of glipizide on PEPCK and CBP/p300 mRNA expression in LPS treated female rat brain. (B) Effect of glipizide on blood glucose levels in LPS treated rats (n = 6). Valuesrepresent mean ± S.D. Comparison are made between a — (control versus LPS treated rats) and b — (LPS treated versus glipizide + LPS treated rats). Values are statistically significantat ***P b 0.001 and *P b 0.05.
253M. Sadasivam et al. / Gene 552 (2014) 249–254
regulation of steroidogenesis in brain during infectionmediated inflam-mation. Moreover, the obtained information will be helpful in the de-sign of novel clinical interventions for neuronal bacterial infectionsand the complications associated with it.
Fig. 4. (A) Effect of glipizide on StAR and steroidogenic enzyme gene expression in LPS treated ra(n= 6). Enzyme activity of 3β-HSD is expressed as unit/mg protein; where one unit is equal tomg protein;where one unit is equal to nanomoles of NADPHoxidized perminute. Values repres— (LPS treated versus glipizide + LPS treated rats). Values are statistically significant at ***P b
Acknowledgment
The infrastructure provided to the Department of Biochemistry byDST-FIST is gratefully acknowledged.
t brain. (B) Effect of glipizide 3β-HSD and17β-HSDenzyme activity in LPS treated rat brainnanomoles of NADH formed per minute. Enzyme activity of 17β-HSD is expressed as unit/entmean± S.D. Comparison aremade between a— (control versus LPS treated rats) and b0.001, **P b 0.01 and *P b 0.05.

Fig. 5. Effect of glipizide on proinflammatory cytokine mRNA levels in LPS treated rat brain (n= 6). Values representmean± S.D. Comparison aremade between a— (Control versus LPStreated rats) and b — (LPS treated versus glipizide + LPS treated rats). Values are statistically significant at ***P b 0.001 and *P b 0.05.
254 M. Sadasivam et al. / Gene 552 (2014) 249–254
References
Ahn, S.W., Gang, G.T., Tadi, S., Nedumaran, B., Kim, Y.D., Park, J.H., Kweon, G.R., Koo, S.H.,Lee, K., Ahn, R.S., Yim, Y.H., Lee, C.H., Harris, R.A., Choi, H.S., 2012. Phosphoenolpyr-uvate carboxykinase and glucose-6-phosphatase are required for steroidogenesis intesticular Leydig cells. J. Biol. Chem. 287, 41875–41887.
Azcoitia, I., Perez-Martin, M., Salazar, V., Castillo, C., Ariznavarreta, C., Garcia-Segura, L.M.,Tresguerres, J.A., 2005. Growth hormone prevents neuronal loss in the aged rat hip-pocampus. Neurobiol. Aging 26, 697–703.
Bergmeyer, H.U., 1974. Methods of enzymatic analysis. Steroid Dehydrogenase. AcademicPress, New York, p. 476.
Dubal, D.B., Wise, P.M., 2001. Neuroprotective effects of estradiol in middle-aged femalerats. Endocrinology 142, 43–48.
Elo, B., Villano, C.M., Govorko, D., White, L.A., 2007. Larval zebrafish as a model for glucosemetabolism: expression of phosphoenolpyruvate carboxykinase as a marker for ex-posure to anti-diabetic compounds. J. Mol. Endocrinol. 38, 433–440.
Garcia-Segura, L.M., 2008. Aromatase in the brain: not just for reproduction anymore. J.Neuroendocrinol. 20, 705–712.
Ghezzi, P., Santo, E.D., Sacco, S., Foddi, C., Barbaccia, M.L., Mennini, T., 2000. Neurosteroidlevels are increased in vivo after LPS treatment and negatively regulate LPS-inducedTNF production. Eur. Cytokine Netw. 11, 464–469.
Gottfried-Blackmore, A., Sierra, A., Jellinck, P.H., McEwen, B.S., Bulloch, K., 2008. Brain mi-croglia express steroid-converting enzymes in the mouse. J. Steroid Biochem. Mol.Biol. 109, 96–107.
Hanson, R.W., 2009. Thematic minireview series: a perspective on the biology of phos-phoenolpyruvate carboxykinase 55 years after its discovery. J. Biol. Chem. 284,27021–27023.
Hopkins, D.F., Williams, G., 1997. Insulin receptors are widely distributed in human brainand bind human and porcine insulin with equal affinity. Diabet. Med. 14, 1044–1050.
Kohchi, C., Ukena, K., Tsutsui, K., 1998. Age- and region-specific expressions of the mes-senger RNAs encoding for steroidogenic enzymes p450scc, P450c17 and 3beta-HSDin the postnatal rat brain. Brain Res. 801, 233–238.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurement with theFolin phenol reagent. J. Biol. Chem. 193, 265–275.
Magata, F., Horiuchi, M., Echizenya, R., Miura, R., Chiba, S., Matsui, M., Miyamoto, A.,Kobayashi, Y., Shimizu, T., 2014. Lipopolysaccharide in ovarian follicular fluid
influences the steroid production in large follicles of dairy cows. Anim. Reprod. Sci.144, 6–13.
Mizock, B.A., 1995. Alterations in carbohydrate metabolism during stress: a review of theliterature. Am. J. Med. 98, 75–84.
O'Brien, R.M., Granner, D.K., 1990. PEPCK gene as model of inhibitory effects of insulin ongene transcription. Diabetes Care 13, 327–339.
O'Bryan, M.K., Schlatt, S., Phillips, D.J., de Kretser, D.M., Hedger, M.P., 2000. Bacteriallipopolysaccharide-induced inflammation compromises testicular function at multi-ple levels in vivo. Endocrinology 141, 238–246.
Reddy, M.M., Mahipal, S.V., Subhashini, J., Reddy, M.C., Roy, K.R., Reddy, G.V., Reddy, P.R.,Reddanna, P., 2006. Bacterial lipopolysaccharide-induced oxidative stress in the im-pairment of steroidogenesis and spermatogenesis in rats. Reprod. Toxicol. 22,493–500.
Rozing, M.P., Westendorp, R.G., de Craen, A.J., Frölich, M., de Goeij, M.C., Heijmans, B.T.,Beekman, M., Wijsman, C.A., Mooijaart, S.P., Blauw, G.J., Slagboom, P.E., vanHeemst, D., Leiden Longevity Study Group, 2010. Favorable glucose tolerance andlower prevalence of metabolic syndrome in offspring without diabetes mellitus ofnonagenarian siblings: the Leiden longevity study. J. Am. Geriatr. Soc. 58, 564–569.
Saldanha, C.J., Duncan, K.A., Walters, B.J., 2009. Neuroprotective actions of brain aroma-tase. Front. Neuroendocrinol. 30, 106–118.
Sharma, R., Patnaik, S.K., 1983. Induction of phosphoenolpyruvate carboxykinase by hy-drocortisone in rat liver and brain as a function of age. Biochem. Int. 7, 535–540.
Shivanandappa, T., Venkatesh, S., 1997. A colorimetric assay method for 3beta-hydroxy-delta5-steroid dehydrogenase. Anal. Biochem. 254, 57–61.
Stoffel-Wagner, B., Watzka, M., Steckelbroeck, S., Schramm, J., Bidlingmaier, J.F.,Klingmüller, D., 1999. Expression of 17beta-hydroxysteroid dehydrogenase types 1,2, 3 and 4 in the human temporal lobe. J. Endocrinol. 160, 119–126.
Sugita, H., Kaneki, M., Tokunaga, E., Sugita, M., Koike, C., Yasuhara, S., Tompkins, R.G.,Martyn, J.A., 2002. Inducible nitric oxide synthase plays a role in LPS-induced hyper-glycemia and insulin resistance. Am. J. Physiol. Endocrinol. Metab. 282, E386–E394.
Yarim, Gül Fatma, 2013. A new strategy in treatment of neurodegenerative diseases:neurosteroids. Ank. Univ. Vet. Fak. Derg. 79–83.
![For Research Use Only PCK2 Polyclonal antibody · Background Information PCK2(phosphoenolpyruvate carboxykinase [GTP], mitochondrial) is also named as PEPCK2, PEPCK-M and belongs](https://img.pdfslide.net/doc/110x75/60b24c18c6049f6cff2e0b4c/for-research-use-only-pck2-polyclonal-antibody-background-information-pck2phosphoenolpyruvate.jpg)




























