Embed Size (px)
Citation preview

The Saccharomyces cerevisiae COQ6 gene encodes a mitochondrial flavin
dependent monooxygenase required for coenzyme Q biosynthesis
Peter Gin, Adam Y. Hsu, Steven C. Rothman†, Tanya Jonassen, Peter T. Lee,
Alexander Tzagoloff§, and Catherine F. Clarke*
Department of Chemistry and Biochemistry and the Molecular Biology Institute,
University of California, Los Angeles, 90095
§Department of Biological Sciences, Columbia University, New York, NY 10027
†Present address: Molecular and Cellular Biology Stanley/Donner ASU,
University of California, Berkeley, 229 Stanley Hall #3206, Berkeley, CA 94720-3206
*Corresponding author: Catherine F. Clarke
Department of Chemistry and Biochemistry
University of California, Los Angeles
607 Charles E. Young Drive East
Los Angeles, CA 90095-1569
Tel: (310) 825-0771
Fax: (310) 206-5213
Email: [email protected]
Running Title: Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on April 29, 2003 as Manuscript M303234200 by guest on July 28, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
2
Summary
Coenzyme Q (Q) is a lipid that functions as an electron carrier in the mitochondrial respiratory
chain in eukaryotes. There are eight complementation groups of Q-deficient Saccharomyces
cerevisiae mutants, designated coq1 - coq8. Here we have isolated the COQ6 gene by functional
complementation and, in contrast to a previous report, find it is not an essential gene. coq6 mutants
are unable to grow on nonfermentable carbon sources and do not synthesize Q, but instead
accumulate the Q biosynthetic intermediate 3-hexaprenyl-4-hydroxybenzoic acid. The Coq6
polypeptide is imported into the mitochondria in a membrane potential-dependent manner. Coq6p
is a peripheral membrane protein that localizes to the matrix side of the inner mitochondrial
membrane. Based on sequence homology to known proteins, we suggest that COQ6 encodes a
flavin-dependent monooxygenase required for one or more steps in Q biosynthesis.
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
3
Introduction
Coenzyme Q (ubiquinone or Q) is a prenylated benzoquinone found in cell membranes and
functions in redox chemistry as both an oxidant (Q) and reductant (QH2) (1). Q is most commonly
associated with its role in the respiratory chain where it transports electrons from either Complex I
or Complex II to Complex III (2); however, Q serves multiple functions. Q has been demonstrated
to play a role in stabilizing the bc1 complex (3). Additionally, Q functions in the electron transport
chains of lysosomal and plasma membranes (4, 5), and QH2 acts as a chain breaking antioxidant of
lipid peroxyl radicals (6). In Escherichia coli, a high QH2:Q ratio is sensed by ArcB, a
transmembrane sensor kinase, that phosphorylates ArcA, activating operons involved in
fermentation and repressing those involved in respiration (7). In Caenorhabditis elegans, dietary Q
produces a shortened life span (8). This phenomenon has been attributed to the generation of
superoxide by Q–• (the Q semiquinone radical) generated during respiratory electron transport (9).
In humans, Q supplementation has been shown to be effective in treating patients with specific
respiratory chain defects (10) and to slow the progression of Parkinson’s Disease symptoms (11).
Cells normally acquire Q through de novo synthesis, and the length of the prenyl tail varies
amongst different organisms (12). Saccharomyces cerevisiae produce Q6, which has six isoprene
units, while E. coli synthesize Q8 and humans synthesize Q10 . Eight COQ genes have been
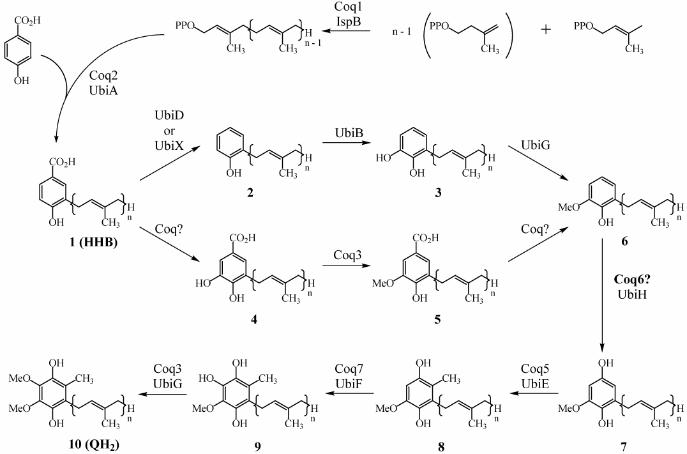
identified to be required for Q synthesis in S. cerevisiae (13, 14). Figure 1 shows the pathway for
Q biosynthesis in both prokaryotes and eukaryotes. In yeast, mutations in any of the eight COQ
genes results in cells which cannot synthesize Q and fail to grow on nonfermentable carbon
sources. Yeast coq3 – coq8 mutants each accumulate the same early intermediate in Q biosynthesis,
3-hexaprenyl-4-hydroxybenzoic acid (HHB or compound 1, Figure 1) (15).
In this work the yeast COQ6 gene has been isolated by functional complementation of a mutant
from the G63 (coq6) complementation group. Transformation of coq6 mutant strains with a
plasmid bearing wild type COQ6 restores Q biosynthesis and growth on non-fermentable carbon
sources. In contrast to a previous report (16), we find COQ6 to be a non-essential gene. Here we
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
4
show the Coq6 polypeptide is imported into mitochondria and is peripherally associated with the
inner membrane on the matrix side. Based on sequence analysis and alignment with other known
hydroxylases, the proposed function of Coq6 polypeptide is that of a flavin-dependent
monooxygenase, required for one or more steps in Q biosynthesis.
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
5
Experimental Procedures
Strains and growth media. The strains used in this study are listed in Table I. Growth media for
yeast were prepared as described (17) and included YPD (1% yeast extract, 2% peptone, 2%
dextrose), YPG (1% yeast extract, 2% peptone, 3% glycerol), YPGal (1% yeast extract, 2% peptone,
2% galactose, 0.1% dextrose), SDC (0.18% yeast nitrogen base without amino acids, 2% dextrose,
0.14% NaH2PO4, 0.5% (NH4)2SO4, and complete supplement of amino acids), SD-Leu (SDC
minus leucine), and SD-Ura (SDC minus uracil). The complete supplement was modified as
described (18). Semisynthetic lactate media was prepared as described (19). Media for sporulation
and tetrad analysis were prepared as described (17). Each component of growth media was
purchased from Difco, Fisher or Sigma. 2% agar was added for solid media.
Cloning of the COQ6 gene. The haploid strain SR128-3C containing the coq6-1 allele was
obtained from mating C128 and FY250 (Table 1). SR128-3C yeast were grown to early-log phase
in YPD medium and transformed (20) with the YCp50 centromeric plasmid library of S. cerevisiae
genomic DNA (21) containing the URA3 gene as a selectable marker. Transformants were selected
on SD-Ura plates, and after a three-day incubation at 30ºC were replicaplated onto YPG plates to
test for respiratory growth. Putative Q prototrophic transformants, able to grow on media
containing a nonfermentable carbon source, were further purified and tested for cosegregation of
uracil prototrophy and respiration competency following vegetative growth in rich media. Such
cosegregation was observed in two transformants, indicating these traits were plasmid linked. Yeast
plasmid DNA was recovered from one transformant (494SR) and was amplified in DH5α E. coli
(Life Technologies, Inc.). The plasmid p494SR contained an insert of 4.1-kb and transformation of
SR128-3C with p494SR restored growth on YPG media. A similar cloning procedure was also
performed with a multicopy expression library prepared from yeast genetic DNA in the vector
YEp24 (22) and resulted in the isolation of pG63/T1, which was found to contain a 2.8-kb segment
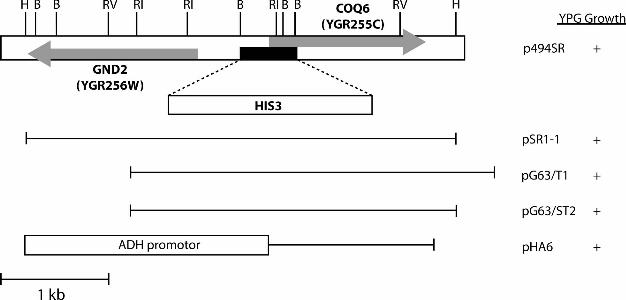
of DNA that overlapped with the insert present in p494SR (Figure 2). Seven other rescuing yeast
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
6
genomic DNA clones were similarly isolated from a recombinant pUV1-based plasmid library
(generous gift of Junichi Nikawa and Michael Wigler, Cold Spring Harbor Laboratory). Southern
analysis showed that all the clones contained overlapping DNA fragments (data not shown).
Subcloning and disruption of the COQ6 gene. A 3.8-kb HindIII fragment was isolated from
p494SR and was ligated to the HindIII site of pRS316 (23) to generate pSR1-1. The insert of
pSR1-1 was sequenced and found to contain two ORFs located on a segment of chromosome VII.
Only one ORF, YGR255C, was also present on pG63/T1, and was designated COQ6 (deposited in
GenBank with accession number AF003698).
To construct a disruption allele of COQ6, a 2.9-kb HindIII fragment, containing about 350-bp of
the YEp24 sequence and 2.55-kb of yeast genomic DNA, was excised from pG63/T1 and inserted
into the HindIII site of YEp352 to generate the plasmid pG63/ST2. As shown in Figure 2, the coq6
deletion plasmid was constructed by replacing a 414-bp BglII fragment in pG63/ST2 with a 1.7-kb
BamHI fragment containing HIS3 (24). The resulting plasmid, pG63/ST3 was used to obtain a
4.0-kb EcoRI fragment with the disrupted gene. W303-1A, W303-1B and SR128-3C were each
transformed with 1 µg of DNA (25). Most of the histidine prototrophic clones issued from the
transformations were respiration-defective and were complemented by the ° test strains JM6 or
JM8, but not by the coq6-1 mutant strains, implying a genetic linkage between the coq6 ::HIS3
and the coq6-1 alleles. Nuclear DNAs from three independent transformants (one from each of the
three separate parental strains) were digested with EcoRI and EcoRV, separated on 1% agarose, and
transferred to a nitrocellulose membrane. The probe, a 1.1-kb EcoRI/EcoRV fragment within the
COQ6 ORF, recognizes a 1.1-kb fragment in the genomic DNA of the parental strain. The
genomic DNA of the mutant strains, (W303∆COQ6-1, αW303∆COQ6-1, and SR∆COQ6-1) each
contained a larger hybridizing species at approximately 3.15-kb, as expected for the disrupted allele.
To verify the allelism between coq6 and coq6-1 mutations, SR∆COQ6-1 was mated to a wild type
strain W303-1A to form diploid cells, which were then sporulated to produce meiotic progeny.
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
7
Spores from 14 tetrads showed 2:2 segregation for respiration competence and histidine
auxotrophy. In all cases, the spores competent to grow on glycerol media were histidine
auxotrophs, while the spores unable to grow on glycerol were histidine prototrophs, confirming the
allelism between the cloned COQ6 gene and the original coq6-1 mutation.
Complete deletion of the COQ6 ORF. A complete deletion of the COQ6 open reading frame was
performed with a PCR-targeting strategy (26). A 1.53-kb fragment containing LEU2 was
generated from the YEp13 vector (GenBank Acc# U03498) using a forward primer pPG6DLF 5’
ATAATTCTTAAAAGTGGAGCTAGTCTATTTCTATTTACATACCTCATTTTGTAATTTCGT
GTCG 3’ and a reverse primer pPG6DLR 5’
TCAAATTGGTCTTTCAGTGAACCTTGTATCGATTGACACAGAGGCAGAGGTCGCCTGA
CGCATA 3’. The 5’ end of the forward primer contained 45 nucleotides corresponding to -45 to -
1 upstream of the COQ6 ORF and at the 3’ end 19 nucleotides corresponding to 5579 to 5597 of
YEp13. The reverse primer similarly was corresponding to 45 nucleotides from +1485 to +1441
of the reverse strand of the COQ6 ORF followed by 19 residues from 7018 to 7000 of the reverse
strand of YEp13. A W303-1AB diploid was created by mating W3031A with W3031B. W3031A,
W3031B, CENPK.2-1C, and W303-1AB cells were transformed with 1 µg of the fragment using
the lithium-acetate method as described (27). Transformed cells were grown on SD-Leu plates at
30ºC for 2 days. Gene disruption was verified by PCR using forward primer Coq6SeqF1 5’
ACCTTTGCATTACAAGTGCAACGCTCTACC 3’ and reverse primer Coq6SeqR1 5’
GGTGACGCGTGTATCCGCCCGCTCTTTTGG 3’ and produced a product of 1.84-kb for both
the wild type and disrupted strains. The PCR product was then digested using the restriction
enzyme EcoRI, which digests the wild type product and produces two products of 0.44 and 1.4-kb,
while the EcoRI digestion fragments of the disrupted product are 1.2 and 0.64-kb. Tetrad analysis
was performed as described above.
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
8
In vivo radiolabeling of Q6-intermediates, lipid extraction, and analysis by HPLC. Yeast strains
SR128-3C and W303∆COQ6-1 were grown in 1 liter SDC medium supplemented with 0.65 µCi
of 4-[U-14C]hydroxybenzoic acid (365 Ci/mol), synthesized from L-[U-14C]tyrosine by alkali heat
fusion (28). Cells were harvested after 3 days of incubation at 30ºC with shaking (200 rpm, OD600
nm = 10), and lipids were extracted as described (29). Lipid extracts were concentrated, the volume
adjusted to 1 ml with hexane, and 0.10 ml aliquots (5.0 - 12.0 x 103 cpm) were analyzed by normal
phase HPLC employing a cyanopropyl column (Zorbax, 5 µm, 4.6 mm x 250 mm, MacMod
Analytical, Chadds Ford, PA) as described (30). The column was equilibrated with a mobile phase
composed of 98% solvent A (hexane) and 2% solvent B (isopropanol:hexane:water:methylene
chloride, 52:41:5:2) at a flow rate of 1ml/min. Ten minutes after sample injection, the percentage of
solvent B was increased linearly from 2% to 27% in 25 minutes (35 min from the start) and then
from 27% to 100% in 20 minutes (55 min after the start). In the next 5 minutes, the percentage of
solvent B was decreased linearly from 100% to 2% and remained at 2% for 30 minutes to
equilibrate the column before next sample injection. The radioactivity of one ml fractions was
measured by scintillation counting in 10 ml of BIOsafe nonaqueous scintillation cocktail (Research
Products International) with a Beckman scintillation counter (model LS-3133P). The efficiency of
14C detection was 90%.
Determination of Q6 content in lipid extracts by HPLC and electrochemical detection. Lipid
extraction and analysis were performed as described previously (31). Yeast strains W303-1A,
SR128-3C, W303∆COQ6-1, and W303∆COQ6-1:pSR1-1 (the coq6 null strain harboring a COQ6
containing plasmid), were grown at 30ºC with shaking (200 rpm) in 50 ml of YPGal to an OD600 nm
of about 4 or alternatively were grown to saturation (2 days). 750 ng of Q9 was added as an
internal standard to a yeast cell pellet (100 mg wet weight), and cells were lysed by vortexing with 1
g of glass beads in 0.35 ml water.
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
9
Mitochondrial import assay. An in vitro transcription template plasmid was constructed by
inserting the COQ6 ORF into the pRS426 vector downstream of the T7 promoter (32). The COQ6
ORF with SalI and NotI linkers at 5' and 3' ends respectively, was PCR-amplified with the template
pSR1-1, 5' primer JF3 (5’ ACGCACGCGTCGACATGTTCTTTTCAAAAGTTATGC 3’), and 3'
primer JF4-1 (5’ ATAAGAATGCGGCCGCTCTCATTTCTCATTTCCTCC 3’) using Vent DNA
polymerase (New England Biolabs). The PCR product was digested with SalI and NotI and
inserted into the corresponding SalI and NotI sites in pRS426. The resulting product, pT7Q6, was
linearized with XhoI and provided template DNA for in vitro transcription (Promega Ribomax
Large Scale RNA Production System). The resulting mRNA was then translated with Promega
Flexi Rabbit Reticulocyte Lysate System in the presence of [35S] methionine from Amersham Life
Sciences (1000 Ci/mmol, at final concentration of 0.75 µM). Both the mRNA and the [35S]-labeled
polypeptides were stored at –80°C. The isolation of mitochondria (from D273-10B/A1) and
import reaction conditions were performed according to Yaffe (33) as described previously (34).
Cell cultures (1 liter) were grown in semisynthetic lactate media to saturation density. Spheroplasts
were prepared and lysed by Dounce homogenization with a tight fitting pestle as described (19).
Purified mitochondria were isolated from a linear Nycodenz gradient as described (19). Each
import reaction contained 6 µl radiolabeled in vitro translated product and isolated mitochondria
(200 µg protein). Following the 30 min incubation at 30°C, the mitochondria were reisolated and
washed once as described (33). Proteinase K treatment after the import was performed by adding
proteinase K at final concentration of 50 µg/ml to resuspended mitochondria. The proteolytic
digest was allowed to proceed at 0°C for 20 minutes and was terminated by the addition of
phenylmethylsulfonylflouride (PMSF) to a final concentration of 1mM.
Plasmid construction of HA-tagged COQ6. Two yeast expression plasmids, one single copy and
one multicopy, were constructed to express the Coq6 polypeptide containing a carboxyl-terminal
peptide (MYPYDVPDYASLDGPMST) corresponding to the carboxy-terminus of the influenza
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
10
hemagglutinin (HA) viral protein, an epitope for the 12CA5 monoclonal antibody (35). The COQ6
ORF region without the stop codon was PCR-amplified using 5' primer JF3 (5’
ACGCACGCGTCGACATGTTCTTTTCAAAAGTTATGC 3’) and 3' primer JF4 (5’
ATAAGAATGCGGCCGCAGTTTCTCATTTCCTCCTAATGTG 3’). The PCR product was
directionally cloned into the multicopy pADCL vector (SalI and NotI at 5' and 3' respectively) (36),
to generate pHA6. pSHA6, a single copy version of COQ6-HA construct, was generated by
removing the 3.6-kb transcriptional cassette from pHA6 by BamH1 partial digestion and insertion
into BamHI site of the vector pRS316 (23). Yeast cells were transformed with pHA6, pSHA6 or
pSR1-1 (25). Transformants were selected for the presence of either the LEU2 (pHA6) or URA3
(pSHA6) gene on SD-Leu or SD-Ura plate media. Colonies obtained on the respective plate media
were subsequently replicaplated to YPG plate media.
Generation of antisera against Coq6p. The 1.44-kb COQ6 ORF was PCR-amplified with a
forward primer pPG6F 5’ GCGGATCCGATGTTCTTTTCAAAAGTTATGCTT 3’ and a reverse
primer pPG6R 5’ GCGGATCCTCATTTCTCATTTCCTCCTAATGTG 3’ and Vent DNA
Polymerase. The product was then digested with BamHI and inserted into PET15b (Novagen) at
the BamHI site to generate a fusion protein containing a 6-His tag at the N-terminus. The fusion
protein was overexpressed in the E. coli BL21(DE3) under induction by 1mM IPTG and was
purified over His-Bind resin (Novagen) and used to generate antiserum in rabbits (Cocalico).
Mitochondrial localization of Coq6p. Yeast cultures (W303-1A) were grown in YPGal to an
OD600 nm between 2 and 4, and mitochondria were isolated and purified as described above.
Mitoplasts were generated by hypotonic treatment of mitochondria (19). Mitochondria (1 mg
protein) were suspended in five volumes of 20mM HEPES-KOH, pH 7.4 and incubated on ice for
20 minutes. The mixture was then sedimented in a microcentrifuge for 10 min at 4°C to separate
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
11
the intermembrane space components (supernatant) and the mitoplasts (pellet). Mitoplasts were
then sonicated and centrifuged at 150,000 × g for 60 min at 4°C to generate matrix (supernatant)
and membrane (pellet) fractions. Alternatively, mitoplasts were alkaline extracted by incubating
with 0.1 M Na2CO3, pH 11.5 for 30 min on ice, followed by centrifugation at 150,000 × g for 60
min at 4°C to separate the integral membrane components (pellet) from the peripheral membrane
and matrix components (supernatant) (37). Proteinase K protection experiments were carried out
as described (38). Samples were analyzed by immunoblot.
Immunoblot analysis. Fractions were assayed for protein concentration by the bicinchoninic acid
assay (Pierce, Rockford, IL). Equal amounts of protein from the mitochondrial fractions of cells
were analyzed by electrophoresis on 12% Tris-Glycine gels and were subsequently transferred to
Hybond ECL Nitrocellulose (Amersham). Immunoblot analysis and treatment of membranes for
re-use with another antisera were performed as described by Amersham. An exception to the stated
protocol was the use of washing buffer: 1X PBS, 0.1% Tween-20. Primary antibodies were used at
the following concentrations: anti-Coq6p, 1:500; anti-Coq3p, 1:1000; anti-Coq4p, 1:1000; anti-
cytochrome b2, 1:1000; anti-Hsp60p, 1:10,000; anti-β subunit of F1-ATPase, 1:10,000; anti-
cytochrome c1, 1:1000; anti-OM45p, 1:1000; anti-Mas2p, 1:1000. Goat anti-rabbit secondary
antibodies conjugated to horseradish peroxidase (Calbiochem) were used at a 1:10,000 dilution.
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
12
Results
Isolation and in situ disruption of the COQ6 gene. The original mutant in the coq6
complementation group of yeast mutants (G63) was identified as a Q-deficient nuclear petite strain
(13). The respiratory deficiency was ascribed to a defect in Q because addition of exogenous Q2 or
Q6 to isolated mitochondria rescued NADH-cytochrome c reductase activity (39). SR128-3C, one
of the strains resulting from the cross between C128 (coq6-1) and FY251, had a very low reversion
rate and provided the biological tool for cloning the COQ6 gene by screening yeast genomic DNA
libraries as described under Experimental Procedures. Nine respiration-competent transformants
were obtained, and the representative plasmid p494SR was studied in detail. A 3.8-kb HindIII
fragment was isolated from p494SR and ligated to the HindIII site of pRS316, a centromeric vector,
to form pSR1-1 (Figure 2). Transformation of SR128-3C with pSR1-1 restored growth on media
containing glycerol. DNA sequence analysis revealed the insert to contain two complete ORFs,
identified as YGR256W (the GND2 gene) and YGR255C. The plasmids pG63/T1 and pG63/ST2
containing the complete YGR255C ORF and only a portion of the GND2 gene, each restored the
ability of SR128-3C to grow on YPG (Figure 2). Expression of the YGR255C ORF as a carboxy-
terminal HA-tagged fusion protein from the alcohol dehydrogenase promoter also rescued YPG
growth of SR128-3C.
The one-step gene replacement procedure (24) was used to obtain strains harboring disrupted
alleles of the COQ6 gene (W303∆COQ6-1, αW303∆COQ6-1 and SR∆COQ6-1). Analysis of
these strains (see Experimental Procedures) confirmed the allelism between the coq6 null mutants
and the original coq6-1 mutant. These data identify the YGR255C ORF as COQ6.
COQ6 is not an essential gene. It has been reported that a complete disruption of the COQ6 ORF
results in lethality as heterozygous knockouts failed to produce viable spores containing a coq6
disruption (16). However, all of the coq6 mutant strains used in this study were viable. A complete
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
13
ORF disruption was created to address this apparent discrepancy. Three haploid strains (W303-
1A, W3031B, and CEN.PK2-1C) were transformed with a PCR-generated disruption cassette and
all produced viable disruptants (W303∆COQ6-2, W303∆COQ6-2, and CEN∆COQ6-2) as
confirmed by PCR and restriction digest (data not shown). Similarly, a W303-1AB diploid was
created by mating W303-1A with W303-1B and was transformed using the same cassette. The
heterozygous COQ6/coq6 diploid strain was subjected to sporulation and tetrad dissection. Each
of the ten tetrads analyzed produced four viable spores on YPD plate media. The spores from each
tetrad showed a 2:2 segregation for respiratory competence and leucine auxotrophy. These results
indicate that the COQ6 gene is not essential for viability but is required for growth on
nonfermentable carbon sources.
Yeast coq6 null mutants lack Q6 and accumulate 3-hexaprenyl-4-hydroxybenzoic acid (HHB).
Growth of coq6-1 mutants in the presence of 4-[U-14C]hydroxybenzoic acid, the ring precursor in
Q biosynthesis, showed this mutant lacked Q6 and accumulated HHB (15). HHB is an early
intermediate in the Q biosynthetic pathway, and accumulates in mutant yeast strains harboring
deletions or disruptions in any one of the COQ3, COQ4, COQ5, COQ7 or COQ8/ABC1 genes
(15, 29, 40, 41). To characterize the defect in Q biosynthesis in a coq6 null mutant, both SR128-3C
(coq6-1) and W303∆COQ6-1 (coq6 ) were grown in the presence of 4-[U-14C]hydroxybenzoic
acid, and lipid extracts were analyzed by normal phase HPLC as described in Experimental
Procedures. Both strains lacked Q6 and accumulate a radioactive intermediate that comigrated with
HHB in fraction 26 (Figure 3). Neither the SR128-3C nor W303∆COQ6-1 was observed to
produce Q6 as analyzed by electrochemical detection, a method that can detect as little as 2 pmol Q6
per mg wet weight of yeast. Q6 levels of W303-1A and W303∆COQ6-1:pSR1-1 were 187 and
148 pmol / mg wet weight of yeast, respectively, when grown to log phase, and were 189 and 169
pmol / mg wet weight of yeast, respectively, when grown to stationary phase (data not shown).
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
14
Coq6p may act as a flavin dependent monooxygenase to catalyze quinone formation. The
predicted amino acid sequence of the COQ6 ORF revealed 21% and 24% sequence identity with
the respective E. coli UbiH and UbiF polypeptides (Figure 4). E. coli ubiH mutants lack Q8 and
accumulate compound 6, indicating UbiH is required for the monooxygenase step that catalyzes
quinone formation (Figure 1 and Ref. 42). The ubiH gene product was identified as a flavin-
dependent monooxygenase (43). The E. coli ubiF gene product has also been identified as a flavin-
dependent monooxygenase, with 31% amino acid sequence identity to UbiH (44). E. coli ubiF
mutants lack Q8, accumulate compound 8, and both ubiH and ubiF mutants fail to grow on media
containing succinate (Figure 1 and Ref 45). Coq6p, UbiH, UbiF, and other eukaryotic homologs of
Coq6 each contain three regions with amino acid sequence identities that are present in a large
family of flavin-dependent monooxygenases (Figure 5 and Ref 46). Region 1 contains an ADP-
binding fingerprint (47), Region 2 is implicated in the recognition of NAD(P)H and may also be
involved indirectly in binding the pyrophosphate moiety of FAD (48), while Region 3 contains a
consensus sequence for binding to the ribityl moiety of FAD (49). Based on the homology with
UbiH, and UbiF, and the presence of the conserved motifs found in other aromatic flavin-dependent
monooxygenases, it seems likely that Coq6p functions in one or more hydroxylation steps in Q
biosynthesis.
Mitochondrial import of Coq6p. The amino-terminal sequence predicted for Coq6p showed
characteristics of mitochondrial targeting sequences (14). The first 28 amino acid residues are
abundant in positively charged amino acid residues and devoid of acidic residues. Arrangement of
these 28 residues in a helical wheel shows the positively charged residues are located along one side
of the α-helix. The amino terminal region contains the sequence motif common in many
mitochondrial matrix proteins that are proteolytically cleaved twice once imported into matrix, as
characterized by an arginine at -10 , a hydrophobic residue (F, L or I) at -8, and serine, threonine or
glycine at -5 relative to the N-terminal residue of the proteolytically processed protein. To
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
15
determine whether Coq6p is a mitochondrial protein, an in vitro mitochondrial import assay of the
Coq6 polypeptide was performed. Upon incubation with mitochondria prepared from wild type
yeast, the in vitro translated Coq6p was cleaved, and the resulting mature form of the protein was
observed to have a mass of about 51kD, consistent with lysine 21 as the putative cleavage site
(Figure 6). This mature protein was protected and resistant to exogenous proteinase K treatment
(Lane 3, Figure 6). Disruption of mitochondrial membrane with detergent exposed the imported
and processed Coq6p, which became accessible to the proteinase K (Lane 4, Figure 6). The import
of Coq6p also required a mitochondrial membrane potential, as the addition of valinomycin
inhibited import of Coq6p (Lanes 5 and 6, Figure 6).
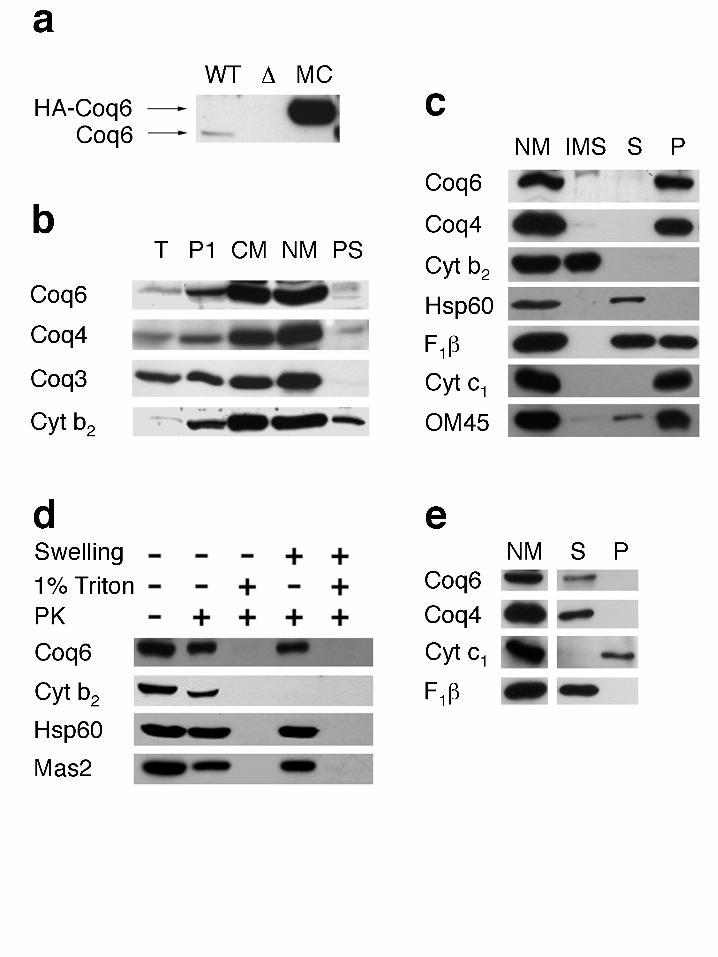
Mitochondrial localization of Coq6p. Our initial investigation of the subcellular localization of
Coq6p made use of a carboxyl-terminal fusion protein between Coq6p and the HA epitope peptide
(see Experimental Procedures). The Coq6-HA epitope fusion protein retained activity as assayed
by the ability of either the single- or multi-copy plasmid construct to rescue coq6 null mutant yeast
strains for growth on media containing a nonfermentable carbon source (YPG plates, data not
shown). However, subsequent subcellular and submitochondrial fractionation analysis revealed that
the Coq6-HA tagged construct was present in mitochondria as an insoluble aggregate because it
remained in the pellet following treatment with 1% Triton-X100 and 1 M NaCl. For this reason,
antibodies were generated to the Coq6 polypeptide.
A polyclonal antibody was generated in rabbit against a Coq6-His-tagged fusion protein. This
antibody recognized a polypeptide of 51 kD in wild type yeast cell extracts, and also recognized the
Coq6-HA tagged polypeptide that migrates at about 57 kD (Figure 7a). No polypeptide at these
molecular weights was detected in cell extracts of the coq6 deletion strain, indicating that the
antibody is specific for Coq6p. Western blot analysis of subcellular yeast fractions revealed that
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
16
Coq6p co-fractionated with the mitochondria along with Coq4p, Coq3p, and cytochrome b2 (Figure
7b). To determine if Coq6p is a membrane bound or soluble protein, purified mitochondria were
osmotically shocked to disrupt the outer membrane and release components of the intermembrane
space. Mitoplasts were then further disrupted by sonication to separate the remaining soluble and
membrane components. Figure 7c shows that Coq6p localizes to the membrane (pellet) fraction.
To determine if Coq6p resides in the inner or outer membrane, mitochondria were treated with
combinations of proteinase K, Triton X-100, and hypotonic swelling. Figure 7d shows that Coq6p
is degraded only when the detergent is present as is the case for two matrix proteins, Hsp60p and
Mas2p. Finally, to determine if Coq6p is a peripheral or integral protein in the inner mitochondrial
membrane, mitoplasts were treated with 0.1M sodium carbonate buffer, pH 11.5. Figure 7e shows
that following this treatment Coq6p is released into the supernatant, as are the β subunit of F1-
ATPase and Coq4p (two peripheral membrane proteins), whereas cytochrome c1 (an integral
membrane protein) is not.
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
17
Discussion
This work characterizes yeast coq6 mutants and the isolation of the COQ6 gene. COQ6 is
necessary for Q biosynthesis. In the original coq6 mutant and in both coq6 partial and complete
deletion mutants, Q6 is undetectable and as a result, these cells are rendered respiratory incompetent.
Based on the data presented here, COQ6 encodes a mitochondrial protein necessary for Q
biosynthesis. Similar to Coq2p, Coq3p, Coq4p, Coq5p, Coq7p, and Coq8p/Abc1p, the Coq6
polypeptide is imported into mitochondria (14). In agreement with submitochondrial localization
studies of Coq1p, Coq3p, Coq4p, Coq5p, and Coq7p, Coq6p is localized to the matrix side of the
inner membrane (18, 40, 50, and unpublished data).
S. cerevisiae is a facultative anaerobe capable of fermentation and aerobic respiration, and none of
the other COQ genes have been identified as being essential. Complete deletions of the COQ6
gene were introduced into both haploid and diploid strains of W303, and also into CEN.PK2-1C, a
haploid strain similar in genetic background to the diploid used by Fiori et al. (16). In contrast to
their results, all yeast strains were viable when COQ6 was deleted, indicating that none of the eight
COQ genes is essential. Q itself is non-essential for growth in this organism.
The function of Coq6p is still unknown, as are the functions of Coq4p and Coq8p. Based on
sequence homology to known flavin dependent monooxygenases, it is likely that Coq6p acts
similarly. Although Coq6p and UbiF share 24% sequence identity, it seems unlikely that Coq6p
functions like UbiF in the hydroxylation of compound 8 in yeast, because previous studies have
shown that yeast Coq7p is required for this hydroxylation step (29). The Coq7 polypeptide has
been identified as a di-iron carboxylate protein, a member of a monooxygenase family distinct from
UbiF (51). Although E. coli lack a homolog of Coq7p, homologs of yeast Coq7p from other
prokaryotic species rescue the E. coli ubiF hydroxylase mutants, indicating that these two distinct
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
18
types of monooxygenases each catalyze the same reaction in Q biosynthesis (51). Currently, two
of the uncharacterized steps in Q biosynthesis are hydroxylations and Coq6p may be responsible
for either or both of these steps. Coq6p has 21% identity to the E. coli UbiH, which converts
compound 6 to 7. Of course other hydroxylation substrates for Coq6p are also possible, including
the hydroxylation of HHB to compound 4 (Figure 1). Indeed, since coq6-1 and coq6 null mutants
both accumulate HHB (Figure 3), it seems attractive to postulate its action at this step. However, the
accumulation of HHB is not diagnostic of the blocked step, since so many other coq mutants also
accumulate HHB (15). Another possibility is that Coq6p catalyzes the oxidative decarboxylation of
compound 5 to 7. It has been demonstrated that 4-hydroxybenzoate 1-hydroxylase oxidatively
decarboxylates 4-hydroxybenzoic acid with the formation of hydroquinone (52). Salicylate
hydroxylase similarly oxidatively decarboxylates salicylate to catechol (53). However, in yeast this
is less likely given that previous work showed a Q-deficient yeast mutant accumulated compound 6
(54). Furthermore, that mutant was never sequenced nor is it available. Identification of the role of
Coq6p in the hydroxylation step(s) of Q biosynthesis will require development of in vitro assays
for these hydroxylation steps.
Blast analysis of Coq6p against other higher eukaryotic genomes revealed that only one homologue
existed for H. sapiens, M. musculus, and C. elegans indicating a conserved evolution for this
protein (Figure 4). For D. melanogaster, two matches were found but only one had a high score
and spanned the entire sequence. The homologs in each case retained the three conserved motifs
depicted in Figure 5.
Genetic data have suggested that a putative complex of these Coq polypetides is responsible for Q
biosynthesis because a deletion in any one of the eight COQ genes results in a profound decrease
in steady state protein levels for Coq3p, Coq4p, and Coq7p (34 and unpublished observations).
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
19
Furthermore, null mutations in coq3, coq4, coq5, coq7 or coq8/abc1 result in the accumulation of
HHB (15). Similarly, analyses of lipid extracts of coq6 mutants in the studies presented here show
these mutants also accumulate HHB. It has been observed that an E226K coq4 point mutant did
allow for the steady state expression of Coq3p and Coq7p (40). In addition, a G104D coq7 point
mutant allowed for the accumulation of 2-hexaprenyl-3-methyl-6-methoxy-1,4-benzoquinone
(demethoxy-Q, or compound 8, Figure 1) (29). It would appear that both the coq4 and coq7 amino
acid substitution mutants allow for the expression of correctly folded but inactive Coq4p and
Coq7p, respectively, and each may be interacting and stabilizing the steady state expression of the
other Coq polypeptides in a complex. The accumulation of HHB in both the coq6 mutants further
suggests that Coq6p similarly is required for stabilization of a complex. Additionally, the coq6-1
mutant has been found to be a nonsense mutation of the tyrosine at position 218 (Hsieh, E., and
Clarke, C., unpublished). This mutant would be unable to produce a stable polypeptide at full
length and would explain why we see an accumulation of HHB.
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
20
References
1. Brandt, U., and Trumpower, B. (1994) Crit. Rev. Biochem. Mol. Biol. 29, 165-197
2. Dutton, P. L., Ohnishi, T., Darrouzet, E., Leonard, M. A., Sharp, R. E., Gibney, B. R.,
Daldal, F., and Moser, C. C. (2000) in Coenzyme Q: Molecular Mechanisms in Health and
Disease (Kagan, V. E. and Quinn, P. J., eds) pp 65-82, CRC Press, Boca Raton, FL
3. Santos-Ocaña, C., Do, T. Q., Padilla, S., Navas, P., and Clarke, C. F. (2002) J. Biol. Chem.
277, 10973-10981
4. Gille, L., and Nohl, H. (2000) Arch. Biochem. Biophys. 375, 347-354
5. Santos-Ocana, C., Do, T. Q., Padilla, S., Navas, P., and Clarke, C. F. (2002) J. Biol. Chem.
277, 10973-10981
6. Kagan, V. E., Nohl, H., and Quinn, P. J. (1996) in Handbook of Antioxidants (Cadenas, E.,
and Packer, L., eds.), pp 157-201, Dekker, New York
7. Georgellis, D., Kwon, O., Lin, E.C. (2001) Science 292, 2314-2316
8. Larsen, P. L., and Clarke, C. F. (2002) Science 295, 120-123
9. Jonassen, T., Marbois, B.N., Faull, K.F., Clarke, C.F., Larsen, P.L. (2002) J Biol Chem 277,
45020-45027
10. Geromel, V., Darin, N., Chretien, D., Benit, P., DeLonlay, P., Rotig, A., Munnich, A., Rustin,
P. (2002) Mol Genet Metab 77, 21-30
11. Shults, C. W., Oakes, D., Kieburtz, K., Beal, M. F., Haas, R., Plumb, S., Juncos, J. L., Nutt,
J., Shoulson, I., Carter, J., Kompoliti, K., Perlmutter, J. S., Reich, S., Stern, M., Watts, R. L.,
Kurlan, R., Molho, E., Harrison, M., and Lew, M. (2002) Arch. Neurol. 59, 1541-1550
12. Meganathan R. (2001) FEMS Microbiol Lett 203, 131-139
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
21
13. Tzagoloff, A., and Dieckmann, C. L. (1990) Microbiol. Rev. 54, 211-225
14. Jonassen, T. and Clarke, C. F. “Genetic Analysis of Coenzyme Q Biosynthesis”
Coenzyme Q: Molecular Mechanisms in Health and Disease. (Kagan, V. E. and Quinn, P.
J., eds.) (2000), 185-208, CRC Press, Boca Raton, FL
15. Poon, W. W., Do, T. Q., Marbois, B. N., and Clarke, C. F. (1997) Mol. Aspects Med. 18,
s121-s127
16. Fiori, A., Bianchi, M. M., Fabiani, L., Falcone, C., Francisci, S., Palleschi, C., Solimando, N.,
Uccelletti, D., and Frontali, L. (2000) Yeast 16, 377-386
17. Adams, A., Gottschling, D. E., Kaiser, C. A., and Stearns, T. (1997) Methods in Yeast
Genetics pp. 145-154, Cold Spring Harbor Laboratory Press, Plainview, NY
18. Poon, W. W., Barkovich, R. J., Hsu, A. Y., Frankel, A., Lee, P. T., Shepherd, J. N., Myles,
D. C., and Clarke, C. F. (1999) J. Biol. Chem. 274, 21665-21672
19. Glick, B. S., and Pon, L. A. (1995) Methods Enzymol. 260, 213-223
20. Marbois, B. N., Hsu, A., Pillai, R., Colicelli, J., and Clarke, C. F. (1994) Gene 138, 213-217
21. Rose, M. D., Novick, P., Thomas, J. H., Botstein, D., and Fink, G. R. (1987) Gene 60, 237-
243
22. Carlson, M., and Botstein, D. (1982) Cell 28, 145-154
23. Sikorski, R. S., and Hieter, P. (1989) Genetics 122, 19-27
24. Rothstein, R. J. (1983) Methods Enzymol. 101, 202-211
25. Elble, R. (1992) Biotechniques 13, 18-20
26. Baudin, A, Ozier-Kalogeropoulos, O., Denouel, A., Lacroute, F., and Cullin, C. (1993) Nucl.
Acids Res. 21, 3329-3330
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
22
27. Gietz, R., Schiestl, R., Willems, A., and Woods, R. (1995) Yeast 11, 355-360
28. Clarke, C. F., Williams, W. and Teruya, J. H. (1991) J. Biol. Chem. 266, 16636-16644
29. Marbois, B .N., and Clarke, C. F. (1996) J. Biol. Chem. 271, 2995-3004
30. Poon, W. W., Marbois, B. N., Faull, K. F. and Clarke, C. F. (1995) Arch. Biochem.
Biophys. 320, 305-314
31. Jonassen, T., and Clarke, C. F. (2000) J. Biol. Chem. 276, 18161-18168
32. Christianson, T. W., Sikorski, R. S., Dante, M., Shero, J. H., and Hieter, P. (1992) Gene
110, 119-122
33. Yaffe, M. P. (1991) Methods Enzymol. 194, 627-643
34. Hsu, A. Y., Poon, W. W., Shepherd, J. A., Myles, D. A., and Clarke, C. F. (1996)
Biochemistry 35, 9797-9806
35. Niman, H. L., Houghten, R. A., Walker, L. E., Reisfeld, R. A., Wilson, I. A., Hogle, J. M.,
and Lerner, R. A. (1983) Proc. Natl. Acad. Sci. USA 80, 4949-4953
36. Spain, B. H., Koo, D., Ramakrishnan, M., Dzudzor, B., and Colicelli, J. (1995) J. Biol.
Chem. 270, 25435-25444
37. Jarosch, E., Tuller, G., Daum, G., Waldherr, M., Voskova, A., and Schweyen, R. J. (1996) J.
Biol. Chem. 271, 17219-17225
38. Glick, B. S., Brandt, A., Cunningham, K., Muller, S., Hallberg, R. L., and Schatz, G. (1992)
Cell 69, 809–822
39. Tzagoloff, A., Akai, A., and Needleman, R. B. (1975) J. Biol. Chem. 250, 8228-8235
40. Belogrudov, G. I., Lee, P. T., Jonassen, T., Hsu, A. Y., Gin, P., and Clarke, C. F. (2001)
Arch. Biochem. Biophys. 392, 48-58
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
23
41. Do, T. Q., Hsu, A. Y., Jonassen, T., Lee, P. T., and Clarke, C. F. (2001) J. Biol. Chem. 276,
18161-18168
42. Young, I. G., Stroobant, P., MacDonald, C. G., and Gibson, F. (1973) J. Bacteriol. 114, 42-
52
43. Nakahigashi, K., Miyamoto, K., Nishimura, K., and Inokuchi, H. (1992) J. Bacteriol. 174,
7352-7359
44. Kwon, O., Kotsakis, A., and Meganathan, R. (2000) FEMS Microbiol. Lett. 186, 157-161
45. Young, I. G., McCann, L. M., Stroobant, P., and Gibson, F. (1971) J. Bacteriol. 105, 769-
778
46. Palfey, B. A., Ballou, D. P., and Massey, V. (1995) Oxygen activation by flavins and
pterins, In Active Oxygen in Biochemistry, (Valentine, J. S., Foote, C. S., Greenberg, A., and
Liebman, J. F., eds.) pp. 37-83, Blackie Academic & Professional Press, Glasgow, UK
47. Wierenga, R. K., Terpstra, P., and Hol, W. G. J. (1986) J. Mol. Biol. 187, 101-107
48. Eppink, M. H. M., Schreuder, H. A., and Van Berkel, W. J. H. (1997) Protein Sci. 6, 2454-
2458
49. Eggink, G., Engel, H., Vriend, G., Terpstra, P., and Witholt, B. (1990) J. Mol. Biol. 212,
135-142
50. Jonassen T., Proft M., Randez-Gil F., Schultz J. R., Marbois B. N., Entian K. D., and
Clarke C. F. (1998) J. Biol. Chem. 273, 3351-3357
51. Stenmark, P., Grunler, J., Mattsson, J., Sindelar, P. J., Nordlund, P., and Berthold, D. A.
(2001) J. Biol. Chem. 276, 33297-300
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
24
52. Eppink, M. H. M., Boeren, S. A., Vervoort, J., and Van Berkel, W. J. H. (1997) J. Bacteriol.
179, 6680-6687
53. Katagiri, M., Takemori, S., Suzuki, K., and Yasuda, H. (1966) J. Biol. Chem. 241, 5675-
5677
54. Goewert, R. R., Sippel, C. J., Grimm, M. F., and Olson, R. E. (1978) FEBS Lett. 87, 219-
221
55. Tzagoloff, A., Akai, A., and Foury, F. (1976) FEBS Lett. 65, 391-395
56. Proft, M., Kotter, P., Hedges, D., Bojunga, N., and Entian, K. D. (1995) EMBO J. 14, 6116-
6126
57. McEwen, J. E., Ko, C., Kloeckner-Gruissem, B., and Poyton, R .O. (1986) J. Biol. Chem.
261, 11872-11879
58. Wong, C. M., Dilworth, M. J., and Glen, A. R. (1994) Microbiology 140, 2775-2786
59. Kalin, M., Neujahr, H. Y., Weissmahr, R. N., Sejlitz, T., Johl, R., Fiechter, A., and Reiser,
J. (1992) J. Bacteriol. 174, 7112-7120
60. Suzuki, K., Mizuguchi, M., Ohnishi, K., and Itagaki, E. (1996) Biochim. Biophys. Acta
1275, 154-156
61. Orser, C. S., Lange, C. C., Xun, L., Zahrt, T. C., and Schneider, B. J. (1993) J. Bacteriol.
175, 411-416
62. Weijer, W. J., Hofsteenge, J., Beintema, J. J., Wierenga, R. K., and Drenth, J. (1983) Eur.
J. Biochem. 133, 109-118
63. DiMarco, A. A., Averhoff, B. A., Kim, E. E., and Ornston, L. N. (1993) Gene 125, 25-33
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
25
64. Filippini, S., Solinas, M. M., Breme, U., Schluter, M. B., Gabellini, D., Biamonti, G.,
Colombo, A. L., and Garofano, L. (1995) Micriobiology 141, 1007-1016
65. Nurk, A., Kasak, L., and Kivisaar, M. (1991) Gene 102, 13-18
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
26
Acknowledgements
This work was supported in part by National Institutes of Health Grant GM45952 (to C. Clarke)
and National Institutes of Health Grant HL2274 (to A. Tzagoloff).
1The abbreviations used are: Q, ubiquinone or coenzyme Q; Coq6p, the polypeptide encoded by
the COQ6 gene.
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
27
Figure Legends
Figure 1. The Q biosynthetic pathway in eukaryotes and prokaryotes. Coq1p (S.
cerevisiae) or IspB (E. coli) assemble the polyprenyldiphosphate tail with n=6 and n=8 isoprene
units, respectively. After formation of 3-polyprenyl-4-hydroxybenzoic acid (compound 1), by the
4-hydroxybenzoic acid:polyprenyltransferase (Coq2p or UbiA), the proposed biosynthetic
pathways for Q in eukaryotes and in prokaryotes is thought to diverge as shown. In yeast, n=6 and
compound 1 is 3-hexaprenyl-4-hydroxybenzoic acid (HHB). E. coli gene products are identified
as Ubi (and also include IspB); S. cerevisiae gene products are identified as Coq.
Figure 2. Restriction map of isolated yeast genomic DNA containing the COQ6 gene and
strategy of COQ6 deletion. The gray bars indicate the positions of the GND2 and COQ6 genes
present on p494SR. Each of the plasmids depicted was able to restore growth of coq6 yeast mutant
strains on YPG plate media as identified by the (+) sign. A deletion construct was prepared by
replacing a 414-bp BglII fragment (black bar) with the HIS3 gene on a BamHI fragment, as
described in Experimental Procedures. Restriction sites are symbolized by either one or two letters
and correspond to the following DNA restriction enzymes: B, BglII; H, HindIII; RI, EcoRI; RV,
EcoRV.
Figure 3. Yeast coq6 mutants lack Q and accumulate a polar intermediate that
corresponds to HHB. (a) Lipid extracts were prepared from SR128-3C (coq6-1 allele) or (b)
from W303∆COQ6-1 (coq6 deletion mutant) and separated by normal phase HPLC as described
in Experimental Procedures. Fractions were collected (1 ml) and 14C radioactivity was determined
by scintillation counting. A Q6 standard eluted in fraction 6.
Figure 4. Alignment of predicted yeast COQ6 amino acid sequence with two E. coli
homologs. The sequence of the yeast Coq6 polypeptide is shown in alignment with homologs in
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
28
C. elegans (GenBank Acc# NP_505415), H. sapiens (GenBank Acc# NP_057024), M. musculus
(GenBank Acc# XP_126972), D. melanogaster (GenBank Acc# NP_608934), E. coli UbiH
(GenBank Acc# P25534), and E. coli UbiF (GenBank Acc# P75728). Alignments were created on
DNASTAR™'s Megalign with the Clustal method and the PAM 250 residue weight table.
Identical amino acid residues are shaded and noted above the alignment. Yeast Coq6 amino acid
sequence shared identities of 29.0% (C. elegans), 28.6% (H. sapiens), 29.2% (M. musculus),
25.8% (D. melanogaster), 21.2% (E. coli UbiH), 24.3% (E. coli UbiF).
Figure 5. Comparison of regions of the predicted COQ6 amino acid sequence with
conserved sequences found in FAD binding aromatic hydroxylases. Alignments were
created on DNASTAR™'s Megalign with the Clustal method and the PAM 250 residue weight
table. The (∆) symbol represents amino acid residues A, I, L, V, M, or C that occur as part of the
ADP binding fingerprint of Region 1 (47). Region 2 is implicated in the recognition of NADH or
NADPH and is also involved indirectly in binding the pyrophosphate moiety of FAD (48). Region
3 contains a consensus for binding to the ribityl moiety of FAD (49). The aromatic hydroxylases
are designated as UbiH, 2-octaprenyl-6-methoxyphenol hydroxylase from Escherichia coli
(GenBank Acc# P25534) (43); PobARh, 4-hydroxybenzoate hydroxylase (PobA) from Rhizobium
leguminosarum (GenBank Acc# AAA73519) (58); PhyATc, phenol hydroxylase from
Trichosporon cutaneum (GenBank Acc# AAA34202) (59); ShPs, salicylate hydroxylase from
Pseudomonas putida (GenBank Acc# d1011754) (60); PcpB, pentachlorophenol 4-
monooxygenase(pcpB) from Flavobacterium sp. (GenBank Acc# AAF15368) (61); PobAPS, p-
hydroxybenzoate hydroxylase from Pseudomonas fluorescens (GenBank Acc# WHPSBF) (62);
PobAAc, p-hydroxybenzoate hydroxylase (pobA) from Acinetobacter calcoaceticus (GenBank
Acc# AAC37163) (63); DnrF, 11-aklavinone hydroxylase from Streptomyces peucetius (GenBank
Acc# AAC43342) (64); PheA, phenol monooxygenase from Pseudomonas sp (GenBank Acc#
AAC64901) (65).
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
29
Figure 6. In vitro mitochondrial import and proteolytic processing of Coq6. Lane 1: 10%
of the radiolabeled in vitro translation product used in each import reaction. Lane 2: in vitro
translation product incubated with isolated mitochondria. Lane 3: Same as Lane 2 but including a
post-import proteinase K treatment. Lane 4: Same as Lane 3 except Triton X-100 was added to
disrupt the mitochondria before proteinase K treatment. Lane 5 and Lane 6: in vitro translation
product was incubated with mitochondria in the presence of the uncoupler valinomycin (10µg/ml
final concentration). After the incubation, the reisolated mitochondria were either treated with
proteinase K as in Lane 5 or were directly analyzed as in Lane 6. The samples were mixed with
sample buffer and heated at 100°C for 5 minutes prior to analysis by SDS polyacrylamide
electrophoresis (12% polyacrylamide) and autoradiography. P and M indicate the positions of the
precursor form and the mature form of Coq6p respectively.
Figure7. Coq6p is peripherally associated with the inner membrane of mitochondria. (a)
The antiserum generated against Coq6p and its specificity was tested against the wild type strain
W303-1A (WT), the coq6 deletion strain W303-1A∆COQ6-1 (∆), and the multicopy HA-tagged
rescued strain W303-1A∆COQ6-1:pHA6-1 (MC). (b) Whole cell yeast was homogenized and
separated by differential centrifugation into nuclear (P1), crude mitochondrial (CM), Nycodenz
gradient purified mitochondrial (NM), and post mitochondrial supernatant fractions (PS). (c)
Nycodenz purified mitochondria (NM) were subjected to hypotonic swelling, centrifuged to
separate intermembrane space proteins in the supernatant (IMS). The pellet was then sonicated and
centrifuged to release a soluble matrix protein fraction in the supernatant (S) and membrane protein
fraction pellet (P). (d) Intact mitochondria or mitoplasts generated by hypotonic swelling were
treated with proteinase K (100 µg/ml) for 30 min with or without 1% Triton X-100. (e) Mitoplasts
were incubated with 0.1 M Na2CO3, pH 11.5 on ice 30 min. Centrifugation produced soluble (S)
and insoluble (P) fractions, which were compared against Nycodenz purified mitochondria (NM).
All samples were separated by SDS-PAGE and analyzed by immunoblotting as described in
experimental procedures. Antiserum was used against Coq6p and compared to results of antisera
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
30
against outer membrane protein OM45p, intermembrane space protein cytochrome b2 (Cyt b2),
integral inner membrane protein cytochrome c1 (Cyt c1), peripheral inner membrane proteins
Coq3p, Coq4p, and the β subunit of F1-ATPase (F1β), and matrix proteins Hsp60 and Mas2.
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6 is a Flavin-Dependent Monooxygenase in Q Biosynthesis
31
TABLE I
Genotypes and sources of S. cerevisiae strains
Strain Genotype Source or Ref.
D273-10B/A1 α, met6 55
W303-1Α a, ade2-2, his3-11,15, leu2-3-115, trp1-1, ura3-1 R. Rothsteina
W303-1Β α, ade2-2, his3-11,15, leu2-3-115, trp1-1, ura3-1 R. Rothsteina
W303-1AB a/α, ade2-2/ade2-2, his3-11,15/his3-11,15, leu2-3-
115/leu2-3-115, trp1-/ trp1-1, ura3-1/ura3-1
This study
CEN.PK2-1C a, ura3, his3, leu2, trp1 56
FY251 a, ura3-52, his3 200, leu2 1, trp1 63 F. Winstonb
JM6 a, his4, ρ° 57
JM8 α, ade1, ρ° 57
C128 α, coq6-1, met 13
SR128-1A a, coq6-1, leu2 1 C128 x FY251
SR128-3C α, coq6-1, his3 200, trp1 63, ura3-52 C128 x FY251
W303∆COQ6-1 W303-1A, coq6::HIS3 This study
αW303∆COQ6-1 W303-1B, coq6::HIS3 This study
SR∆COQ6-1 SR128-3C, coq6::HIS3 This study
W303∆COQ6-2 W303-1A, coq6::LEU2 This study
αW303∆COQ6-2 W303-1B, coq6::LEU2 This study
W303-1AB∆COQ6-2 W303, coq6::LEU2 This study
CEN∆COQ6-2 CEN.PK2-1C, coq6::LEU2 This study
W303∆COQ6-1:pSR1-1 W303-1A, coq6::HIS3:pSR1-1 This studya Dr. Rodney Rothstein, Department of Human Genetics, Columbia Universityb Dr. Fred Winston, C. Dollard and S. Ricapero-Hovasse, Department of Genetics, Harvard
University
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

- - - - - - - - - - - - - - - - - - - - - M F F S K V M L T R R I L V R G L A T A K S S A P K L T D V L I V G G G P1 S. cerevisiae- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - M K L P G G T I I C A R N A S S Y Y D T V I V G G G M1 C. elegans- - - - - - - - - - - - - M A A R L V S R C G A V R A A P H S G P L A V L A Q V V R R S T D T V Y D V V V S G G G L1 H. sapiens- - - - - M A A R I G S M A G L L C V R W W S S A Q L A A R G G P L V A S Q R W A G S S A D T V Y D V V V S G G G L1 M. musculusM L G V L R I Q G A L A S A G Q A R L L S V R L L A S K S T T D M T T N R G E S T Q S T S T E H F D I I I G G G G L1 D. melanogaster- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - M S V I I V G G G M1 E. coli UbiH- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - M T N Q P T E I A I V G G G M1 E. coli UbiF
A G L T L A A S I K N S P Q L K D L K T T L V D M V D L K D K L S D F Y N S P P D Y F T N R I V S V T P R S I H F L38 S. cerevisiaeV G N A M A C S L G A N K S F Q S K S V L L L D A G R S P S L A S F - - - K P G A P F N N R V V A T S P T S I D T F28 C. elegansV G A A M A C A L G Y D I H F H D K K I L L L E A G - - P K K V L E - - - K L S E T Y S N R V S S I S P G S A T L L46 H. sapiensV G S A M A C A L G H D I H F H D K K I L L L E A G - - P K K A L E - - - K L S E T Y S N R V S S I S P G S T T L L54 M. musculusV G T T L A A A L A K N S T L A D K K V L L L E G A - - P E F R G F - - - N P T G P Y Q N R V S A I N H N S I E L F59 D. melanogasterA G A T L A L A I S R L S H - G A L P V H L I E A T A P E S H - A H - - - - - - P G F D G R A I A L A A G T C Q Q L11 E. coli UbiHV G G A L A L G - - - L A Q - H G F T V T V I E H A E P A P F V A D - - - - - - S Q P D V R I S A I S A A S V S L L16 E. coli UbiF
E N N A G - A T L M H D R I Q - S Y D G L Y V T D G C S K A T L D L A R D S M L - C M I E - I I N I Q A S L Y N R I96 S. cerevisiaeK K L G V W D Q I N S H R T K - K V N R L F V F D S C S T S E I E F E R G Q - Q - E E V A F I I E N D L I V G S L Y83 C. elegansS S F G A W D H I C N M R Y R - A F R R M Q V W D A C S E A L I M F D K D N L D - - D M G Y I V E N D V I M H A L T99 H. sapiensS S F G A W D H I C N M R C K - A F R R M Q V W D S C S E A L I M F D R D N L D - - D M G Y I V E N D V I M Y A L T107 M. musculusK S I D A W K H I E S A R Y K - P V K Q M Q V W E S N T D A L I Q F Q H D N F A - S D V A C I I E N D L I L D A V Y112 D. melanogasterA R I G V W Q S L A D - - C A T A I T T V H V S D R G H A G F V T L A A E D Y Q L A A L G Q V V E L H N V G Q R L F61 E. coli UbiHK G L G V W D A V Q A M R C H - P Y R R L E T W E W - E T A H V V F D A A E L K L P L L G Y M V E N T V L Q Q A L W64 E. coli UbiF
S Q Y D S K K D S I D I I D N T K V V N I K H S D P - - - N D P L S W P L V T L S N G E V Y K T R L L V G A D G F N150 S. cerevisiaeE K L A E Y K N - V D V K T G A K V E D C S I P N A L E N M A T - - - - - I K L E N G D V I E T S L L I G A D G V N138 C. elegansK Q L E A V S D R V T V L Y R S K A I R Y T W P C P F P M A D S S P W V H I T L G D G S T F Q T K L L I G A D G H N154 H. sapiensK Q L E A V A D R V K V L Y E S K A V G Y S W P G A F S M A D S S P W V H I T L G D G S T L Q T K L L I G A D G H K162 M. musculus- A L A K E S P N V E I L N K A R - I Q C V R - - - L P R D S N S N H S E L Q L E D G R N F S C D L L I G A D G A N168 D. melanogasterA L L R K - A P G V T L H C P D R V A N V A R T Q S H - - - - - - - - V E V T L E S G E T L T G R V L V A A D G T H117 E. coli UbiHQ A L E A - H P K V T L R V P T S L I A L H R D N D L - - - - - - - - Q E L E L K G G E V I R A K L V I G A D G A N120 E. coli UbiF
S P T R R F S Q I P S R G W M - - - Y N A Y G V V A S M K L E Y P P F K L R - G W Q R F L P T G P I A H L P M P E N205 S. cerevisiaeS K V R H A S N L D Y T T F N - - - Y N Q H G L V A I V N I E T A N G K N E T A W Q R F T T L G P V A L L P L S D T190 C. elegansS G V R Q A V G I Q N V S W N - - - Y D Q S A V V A T L H L S E A T - E N N V A W Q R F L P S G P I A L L P L S D T212 H. sapiensS G V R Q A A G I Q N V S W K - - - Y D Q S A V V A T L H L S E A T - E N N V A W Q R F L P S G P I A L L P L S D T220 M. musculusS V V R K E M N V D V F S L N - - - Y D R M G L V A T L E L G E D A C D N S V A W Q R F L P N G P V A L L P L T D R221 D. melanogasterS A L A T A C G V D - - - W Q Q E P Y E Q L A V I A N V A T S V A H - - E G R A F E R F T Q H G P L A M L P M S D G166 E. coli UbiHS Q V R Q M A G I G V H A W Q - - - Y A Q S C M L I S V Q C E N D P - - G D S T W Q Q F T P D G P R A F L P L F D N169 E. coli UbiF
N A T L V W S S S E R L S R L L L S L P P E S F T A L I N A A F V L E D A D M N Y Y Y R T L E D G S M D T D K L I E259 S. cerevisiaeV S G L T W S T S P E E A Q R L K Q L P S D Q F V D E L N S A L F S Q N N Q I P L - - - - - - - - - - - V N Q T I F245 C. elegansL S S L V W S T S H E H A A E L V S M D E E K F V D A V N S A F W S D A D H T D F - - - - - - - - - - - I D T A G A266 H. sapiensL S S L V W S T S H E H A A E L V S M D E E E F V D A I N S A F W S D V H H T D F - - - - - - - - - - - V D S A S A274 M. musculusL S S L V W S T T N E Q A K M L Q A L P P T E F V D A L N E A F C R Q Y P R V E L - - - - - - - - - - - A D K A V Q276 D. melanogasterR C S L V W C H P L E R R E E V L S W S D E K F C R E L Q S A F G W R L G K I T - - - - - - - - - - - - - - - - - -219 E. coli UbiHW A S L V W Y D S P A R I R Q L Q N M N M A Q L Q V E I A K H F P S R L G Y V T - - - - - - - - - - - - - - - - - -222 E. coli UbiF
D I K F R T E E I Y A T L K D E S D I D E I Y P P R V V S I I D K T R A R F P L K L T H A D R Y C T D R V A L V G D317 S. cerevisiaeA L N - R - M N P F R T E T F G R K A E G T T P P H V I T V Q D K S R A S F P L G F G N A H S Y I T T R C A L I G D292 C. elegansM L Q - Y P V S L L K P T K V S A R - Q - - L P P S V P W V D A K S R V L F P L G L G H A A E Y V R P R V A L I G D313 H. sapiensM V R - H A V A L L K P T K V S A R - Q - - L P P S I A K V D A K S R A L F P L G L G H A A E Y V R P R V A L I G D321 M. musculusA L N - - - - S L F G H N G S Q H Q V Q - - Y P P R V C G V L D K S R A T F P L G F L H A S S Y V C N G A A L V G D323 D. melanogaster- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - H A G K R S A Y P L A L T H A A R S I T H R T V L V G N259 E. coli UbiH- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - P L A A - G A F P L T R R H A L Q Y V Q P G L A L V G D262 E. coli UbiF
A A H T T H P L A G Q G L N M G Q T D V H G L V Y A L E K A M E R G L D I G S S L S L E P F W A E R Y P S N N V L L375 S. cerevisiaeA A H R M H P L A G Q G V N L G W S D V Q I L D K V L G D A V R E G A D I G S I T Y L R E Y D S A A Q K H N L P V M348 C. elegansA A H R V H P L A G Q G V N M G F G D I S S L A H H L S T A A F N G K D L G S V S H L T G Y E T E R Q R H N T A L L367 H. sapiensA A H R I H P L A G Q G V N M G F G D I S S L V H H L S T A A F N G K D L G S M S H L T G Y E T D R Q R H N T A L L375 M. musculusA A H R V H P L A G Q G V N L G F S D V R Y L V E S L A A G A Y A G F K L G D K Q H L I K Y E R K C L A K N V P I M375 D. melanogasterA A Q T L H P I A G Q G F N L G M R D V M S L A E T L T Q A Q E R G E D M G D Y G V L C R Y Q Q R R Q S D R E A T I287 E. coli UbiHA A H T I H P L A G Q G V N L G Y R D V D A L I D V L V N A R S Y G E A W A S Y P V L K R Y Q M R R M A D N F I M Q289 E. coli UbiF
G M A D K L F K L Y H T N F P P V V A L R T F G L N L T N K I G P V K N M I I D - T L G G N E K 433 S. cerevisiaeV S V D L L N R L Y R T D A P A I V A A R A F G L N A F N S L G P V K N F L M N - Y L S A H - - R 406 C. elegansA A T D L L K R L Y S T S A S P L V L L R T W G L Q A T N A V S P L K E Q I M - - A F A S - - - K 425 H. sapiensA A T D L L K R L Y S T S A T P L V L L R T W G L Q A T N A V S P L K E Q I M - - A F A S - - - K 433 M. musculusL G V H G L H T L Y A T Q F S P V V L L R S L G L Q L T Q N L P P V K N L F M R - G A M G - - - Q 433 D. melanogasterG V T D S L V H L F A N R W A P L V V G R N I G L M T M E L F T P A R D V L A Q R T L G W - V A R 345 E. coli UbiHS G M D L F Y A G F S N N L P P L R F V R N L G L M A A E - - - - R A G V L K R Q A L K Y A L G L 347 E. coli UbiF
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Coq6UbiHPobAPhyATcShPsPcpBPobAPsPobAAcDnrFPheA
Rh
R L L V G A D G F N S P T R195R V L V A A D G T H S A L A156D F I A G C D G F H G A S R153K Y V I G C D G G H S W V R219D V A I A A D G I K S S M R154R W V I G A D G V R S R V R167D Y I A G C D G F H G I S R153D F I A G C D G Y H G V C R155R Y L V A A D G P R S A I R173K Y L I G A D G A R S K V A201
V L I V G G G P A G L T L A A S I K N S P30V I I V G G G M A G A T L A L A I S R L S3V A I I G S G P S G L L L G Q L L T E A G5V L I V G A G P A G L M A A R V L S E Y V10V A V I G G G I A G T A L A L G L S K S S8V L I V G G G P T G L I A A N E L L R R G17V A I I G A G P S G L L L G Q L L H K A G5V A I I G S G P A G L L L G Q L L Y K A G8V L V V G G G L G G L S T A L F L A R R G10V L I V G S G P A G S S A A M F L S T Q G38
Coq6UbiHPobAPhyATcShPsPcpBPobAPsPobAAcDnrFPheA
Rh
D R V A L V G D A A H T T H P L A G Q G L N M G Q T D V H G L V Y A L367H R T V L V G N A A Q T L H P I A G Q G F N L G M R D V M S L A E T L279G R L F L V G D A A H I V P P T G A K G L N L A A S D V H Y L F S G L279E R V F I A G D A C H T H S P K A G Q G M N T S M M D T Y N L G W K L350G R V A L I G D A A H A M L P H Q G A G A G Q G L E D A Y F M A E L L305G N V F L A G D A A H C H S P S G G S G M N V G M Q D A F N L G W K I291G R L F L A G D A A H I V P P T G A K G L N L A A S D V S T L Y R L L279G K L F L A G D A A H I V P P T G A K G L N L A A S D I A Y L S S A L281G P V L L V G D A A K V T P P T G G M G G N T A I G H G F D V A W K L301G R V C C A G D A I H K H P P S H G L G S N T S I Q D S Y N L C W K L322
Coq6UbiHPobAPhyATcShPsPcpBPobAPsPobAAcDnrFPheA
Rh
Region 1: ADP Binding Fingerprint
Region 2: NAD(P)H / FAD Recognition
Region 3: Ribityl Binding Motif
∆ ∆ ∆ ∆
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from

Tzagoloff and Catherine F. ClarkePeter Gin, Adam Y. Hsu, Steven C. Rothman, Tanya Jonassen, Peter T. Lee, Alexander
monooxygenase required for coenzyme Q biosynthesisThe saccharomyces cerevisiae COQ6 gene encodes a mitochondrial flavin dependent
published online April 29, 2003J. Biol. Chem.
10.1074/jbc.M303234200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on July 28, 2018http://w
ww
.jbc.org/D
ownloaded from