Embed Size (px)
Citation preview

Biochem. J. (1971) 123, 805-814Printed in Great Britain
805
The Stimulatory Effects of Carbon Tetrachloride and other Halogeno-alkanes on Peroxidative Reactions in Rat Liver Fractions in vitro
GENERAL FEATURES OF THE SYSTEMS USED
By T. F. SLATER* AND B. C. SAWYERDepartment of Biochemical Pathology, UniversityCollegeHospital MedicalSchool, London W.C.1, U.K.
(Received 8 April 1971)
1. The general features of the reaction by which carbon tetrachloride stimulateslipid peroxidation have been elucidated in rat liver microsomal suspensions and inmixtures of microsomes plus cell sap. The production of lipid peroxides has beencorrelated with malonaldehyde production in the systems used. 2. The stimulationof malonaldehyde production by carbon tetrachloride requires a source of reducedNADP+ and is dependent on the extent of the endogenous peroxidation of themicrosomal membranes: if extensive endogenous peroxidation occurs duringincubation then no stimulation by carbon tetrachloride is apparent. 3. Thestimulation of malonaldehyde production by carbon tetrachloride has been shownto be proportional to the square root of the carbon tetrachloride concentration inthe incubation mixture. It is concluded that the stimulation of malonaldehydeproduction by carbon tetrachloride results frem an initiation process that is itselfdependent on the homolytic dissociation of carbon tetrachloride to free-radicalproducts. 4. The increased production of malonaldehyde due to carbon tetra-chloride is accompanied by a decreased activity of glucose 6-phosphatase in ratliver microsomal suspensions. 5. The relative activities of bromotrichloromethane,fluorotrichloromethane and chloroform have been evaluated in comparison withthe effects of carbon tetrachloride in increasing malonaldehyde production and indecreasing glucose 6-phosphatase activity. Bromotrichloromethane was more
effective, and fluorotrichloromethane and chloroform were less effective, thancarbon tetrachloride in producing these two effects. It is concluded that homolyticbond fission of the halogenomethanes is a requisite for the occurrence of the twoeffects observed in the endoplasmic reticulum.
The toxic action of carbon tetrachloride on ratliver, which is manifest as an extensive centrilo-bular necrosis, is dependent on the metabolism ofcarbon tetrachloride by an enzyme system located inthe endoplasmic reticulum (see Slater, 1966a;Recknagel, 1967). It has been suggested that onelikely route of the metabolism of carbon tetra-chloride involves the formation of free-radicalproducts (Hove, 1948; Butler, 1961; Wirtschafter& Cronyn, 1964; Recknagel & Ghoshal, 1966;Comporti, Saccocci & Dianzani, 1965; Slater,1966a).The formation of significant quantities of non-
associative radicals such as the trichloromethylradical, of transient existence but high chemicalreactivity, can be expected to result in severe dis-turbances to enzyme activities and organized struc-ture of the endoplasmic reticulum. In particular,
* Present address: Department of Biochemistry,Brunel University, Uxbridge, Middx., U.K.
26
the attack of free-radical intermediates on unsatur-ated lipids in intracellular membranes to result inlipidperoxidationhasbeen stressedby several groupsofworkers to be ofparticular relevance to the necro-genic action of carbon tetrachloride (Ghoshal &Recknagel, 1965; Comporti et al. 1965; Slater,1966a). Previous studies have shown that carbontetrachloride will stimulate lipid peroxidation invitro in suspensions of liver microsomes plus super-natant (Ghoshal & Recknagel, 1965; Comportiet al. 1965) and in microsomes alone provided that asource of NADPH is present (Slater, 1966b).In view of the possible relationship between lipid
peroxidation and the onset of necrosis (seeRecknagel, 1967) it is of obvious importance toinvestigate the mechanisms by which carbon tetra-chloride stimulates lipid peroxidation in vitro, andthe relationship of such results in vitro to the situa-tion in vivo. In this and following papers (Slater &Sawyer, 1971a,b) these points are considered.
Bioch. 1971, 123

T. F. SLATER AND B. C. SAWYER
METHODS
General procedures and enzyme a88ays. The rats usedwere albino females of about 120g body wt.; they were fedon a modified diet M.R.C. 41 (Oxo Ltd., London S.E.1,U.K.) and water ad libitum. The rats used were obtainedfrom four sources: (A) Twyford Laboratories Ltd.; (B)University College London; (C) Carworth (Europe) Ltd.,Alconbury, Hunts., U.K.; (D) A. J. Tuck and Son, Ray-leigh, Essex, U.K.; changes in the source of albino ratsused were a consequence of external factors outside our
control. Although the rate of endogenous peroxidationvaried in liver suspensions prepared from rats from dif-ferent sources, no significant differences were observedin the response to the addition of carbon tetrachloride toliver suspensions.
Rats were killed by cervical dislocation, and the liverswere quickly removed, weighed and cooled in ice-cold0.25M - sucrose. The suspensions were centrifuged at11 700g,,. for lOmin at 20C; the supernatant solutionwas carefully removed and reoentrifuged by using thesame conditions. The supernatant solution so obtainedwas used as 'the microsomes-plus-supernatant mixture'.In experiments with microsome suspensions, the micro-somes-plus-supernatant mixture described above was fur-ther centrifuged at 20C for 40min at 157000g,y. (Super-speed 50 centrifuge; MSE Ltd., Crawley, Sussex, U.K.).The microsomal pellet was rinsed until free from overlyingsupernatant with ioe-cold 0.15m-KCl and was resuspendedin ice-cold 0.15m-KCI such that ml of suspension con-
tained microsomes equivalent to g wet wt. of originalliver.A standard stock solution contained 83.5mm-KOCl;
37.2mM-tris-HCl buffer, pH8.0; 5.5mM-glucose 6-phos-phate (sodium salt); 0.245mm-NADP+ (sodium salt);l0mmi-acetamide; and 8.4 i.u. ofglucose 6-phosphate de-hydrogenase in a total volume of 32.4ml. Standard stocksolution (32.4ml) was mixed with either 4ml ofmicrosomesuspension or with 6ml of microsome-plus-supernatantsuspension to constitute the microsome-stock suspensionsor the microsomes-plus-supernatant-stock suspensionsthat were added immediately on preparation to thecentral oompartments of the Warburg flasks. In a fewinstances the tris buffer was replaced by phosphate buffer(Na2HPO4-NaH2PO4, 0.l1x, pH7.4) and the acetamidewas replaced by 5 mx-nicotinamide; no differences inexperimental behaviour were observed in such cases.
Inorganic pyrophosphatase activity in the incubationmixtures was determined as described by Slater &Sawyer (1969). Glucose 6-phosphatase activity was
determined by the method of Delaney & Slater (1970);incubations for both enzyme assays were for 30min at370C. Oxidized and reduced NADP were measured intissue extraots by the method of Slater, Sawyer & Strauli(1964).
Determination of malonaldehyde and lipid peroxide. Tostudy the effects of CC14 and other halogenoalkanes on theproduction of material giving a positive reaction in thethiobarbituric acid reaction (see below), 2.5ml of a micro-some-stock suspension, or of a microsomes-plus-super-natant-stock suspension, was placed in the central com-
partment of a Warburg flask. An appropriate quantity(l-10{sl) of the halogenoalkane in liquid paraffin was
added to the side arm of the flask with a Hamilton micro-
syringe to allow diffusion of the halogenoalkane duringincubation. The flask was sealed and incubated at 3700in the dark with gentle shaking (60 shakes/min). Incuba-tion times were usually 10-15min for microsome-stocksuspensions and 60 min for microsomes-plus-supernatant-stock suspensions; the actual incubation time used andother individual details are given for each experiment atthe appropriate position in the text.At the end of the incubation periods 1 ml samples of the
suspension were removed from the central compartmentsof the Warburg flasks, mixed with 2ml of 10% (w/v) tri-chloroacetic acid and stood in ice for approx. 15min. Theprecipitates were separated by centrifugation and 2mlsamples ofthe clear supernatant solution were mixed with2ml of aq. 0.67% thiobarbituric acid and heated in aboiling-water bath for 10min. The solutions were thencooled in ice for 5min, after which the E535 was determined.The results are expressed in terms of malonaldehyde pro-duction; a calibration curve was constructed by usingmalonaldehyde bis-diethyl acetal (Schuchardt ChemischeFabrik, Munchen, Germany) as a standard. The ES35 waslinear with increasing amounts of malonaldehyde added(range studied 0-25nmol). The molar extinction coeffi-cient determined with the standard sample was 1.49x10 1- mol - cm-,; this was used in calculating results ona molar basis. Because the stimulations ofmalonaldehydeproduction due to CC14 were small in terms of the absoluteamounts of malonaldehyde measured, incubations foreach point described in the Results section were performedin triplioate and sometimes in pentuplicate. Thiobarbit-uric acid determinations on the incubated mixtures werecarried out on each incubation mixture in duplicate; eachvalue described in the Results section is thus the mean ofsix to ten malonaldehyde determinations.
Gas-liquid chromatography of the thiobarbituric acidreaction mixture (Frinz & Cole, 1962) has indicated thepresence of several reaction products. This and otherevidence (Saslow, Corwin & Waravdekar, 1966) suggeststhat the material(s) measured in tissue extracts by thethiobarbituric acid reaction may be more complex thanmalonaldehyde alone. Bearing in mind such reservationsabout the precise identity of the component reacting withthiobarbituric aoid, we shall use 'malonaldehyde' syn-onymously in this and the following papers (Slater &Sawyer, 1971a,b) with 'material giving a positive reactionwith thiobarbituric acid'.
Malonaldehyde production that occurred in microsomesuspensions (or in microsomes-plus-supernatant suspen-sions) mixed with stock solution containing NADPH isdescribed as endogenous peroxidation to distinguish itfrom the additional peroxidation that results from incuba-tion in the presence of CC14. In a few instances experi-ments were performed with tissue suspensions in the ab-sence ofNADPH; this is noted at the relevant position inthe text.
Lipid peroxide was measured in samples of incubationmixture by the procedure of Swoboda & Lea (1958). Acalibration curve was prepared with a solution of laurylperoxide in acetic acid-chloroform (3:2, v/v); the E350of the final cadmium acetate aqueous phase was linearwith increasing amounts of lauryl peroxide over the range
When different concentrations of microsomes-plus-supernatant in 0.25M-suorose were incubated for 2h at
197180vl6

CARBON TETRACHLORIDE AND LIPID PEROXIDATION37°C there was a very good correlation between lipidperoxide concentration (measured iodometrically) andmalonaldehyde concentration (measured by the thio-barbituric acid reaction). The relationship was linear overthe corresponding ranges studied (malonaldehyde, 0-16nmol/ml of suspension; lipid peroxide, 0-400nmol/ml ofsuspension). It can be seen that malonaldehyde produc-tion is much lower on a molar basis than the correspondingchange in lipid peroxide. This is a well-recognized hazardof using malonaldehyde determinations to follow thecourse of lipid peroxidation and it is important to checkthe basic correspondence between the two parameters inthe particular system under study. Provided that theincubation time was kept constant then malonaldehydeconcentration was linearly related to the concentration oflipid peroxide in the systems studied here.
Determination of halogenoalkane8 in tis8ue suspensions.The concentrations of halogenoalkanes present in theincubation mixtures contained in the central compart-ments of Warburg flasks, and resulting from the diffusionof the halogenoalkanes from the side arms of the flasks,were determined by three methods: colorimetry, radio-isotope dilution, and g.l.c.
CC14 was determined colorimetrically by the method ofRecknagel & Litteria (1960). A calibration curve wasconstructed by using a CCl4-toluene mixture; the E530 ofthe final heated alkaline pyridine mixture was linear withincreasing amounts of CC14 added (range studied 0-3 ,umolof CCl4/ml of toluene).The concentration of CC14 in incubation mixtures was
also determined by radioisotope dilution. 14CC14 wasobtained from The Radiochemical Centre, Amersham,Bucks., U.K., and was accurately diluted with '2CC14.Samples (2,u1) of various dilutions of the labelled 14CC14solution were added to a mixture of 15 ml of scintillationfluid and 0.5 ml of microsome-plus-supernatant-stocksuspensions to provide a calibration curve. Microsomes-plus-supernatant-stock suspensions (3.0ml) were incub-ated for 60min at 37°C in the central compartments ofWarburg flasks containing known amounts of the 14CC14solutions in the side arms after which samples (0.5ml) ofthe suspensions in the central compartment were added to15ml of scintillation fluid. The scintillation fluid was thethixotropic mixture described by Bray (1960) and Gordon& Wolfe (1960). The radioactive content of the mixtureswas determined by using a Packard Tri-Carb spectrometeron the '4C-mode, set for 10000c.p.m. or 10min.
Halothane, bromotrichloromethane, fluorotrichloro -methane and chloroform were measured in tissue sus-pensions by g.l.c. by using a Pye model 104 gas chromato-graph with a 63Ni electron-capture detector.
RESULTSCoenzyme dependence and incubation time. The
stimulation of malonaldehyde production in livermicrosomes-stock suspensions by carbon tetra-chloride is dependent on a source of NADPH thatcannot be replaced by NADH (Table 1). Thestimulation of malonaldehyde production in micro-somes-stock suspensions increased linearly withthe time of incubation at 37°C up to approx. 15 minbut then decreased; with microsomes-plus-super-natant-stock suspensions the stimulation by
carbon tetrachloride increased linearly with in-cubation time up to at least 90min at 37°C (Fig. 1).Measurements of malonaldehyde concentrationduring the linear regions of the two curves in Fig. 1may thus be equated in a proportional manner withthe rate of production of malonaldehyde.
This conclusion is further favoured by the findingthat no significant destruction of added malon-aldehyde (0-lOnmol/ml of suspension) could bedetected in a microsomes-stock suspension duringan incubation period of 15 min at 370C.
-The decrease in the stimulation produced bycarbon tetrachloride in microsomes-stock sus-pensions with incubation times greater than 15 rmin(Fig. 1) possibly reflects the rapid destruction ofNADP++NADPH in the microsomes-stock sus-pension. There was a rapid destruction ofNADP + +NADPH during the incubations at 370C, as shownin Table 2 for microsomes-plus-supernatant-stocksuspensions. The stimulation of malonaldehydeproduction by carbon tetrachloride was increasedby high initial concentrations of NADP+ in thestock solution; there was a similar rise in the endo-genous production of malonaldehyde. Table 3shows the results obtained by using a microsome sus-pension prepared from female rats prestarved offood for 24 h before death; a similar correla-tion of stimulation with the concentration ofNADP+ was found by using microsomes isolatedfrom normally fed rats.No stimulation of malonaldehyde production by
carbon tetrachloride was observed with micro-somes-stock suspensions in which the microsomesample had first been heated under anaerobicconditions for 5min at 700C or 5min at 1000C.Similarly, no stimulation was observed when glucose6-phosphate was omitted from the standard stocksolution. These results indicate that the stimula-tory effect of carbon tetrachloride involves anenzymic interaction with the microsomes andrequires the presence of NADPH.The mean stimulation of malonaldehyde produc-
tion by carbon tetrachloride (2,u of mixture withliquid paraffin, 1:1, v/v, in the side arms ofWarburgflasks) in microsomes-stock suspensions afterincubation times of 10-15min, was 19+1% (meanof 64 separate experiments ±S.E.M.) expressed as apercentage of the endogenous peroxidation mea-sured in the absence of carbon tetrachloride. Incorresponding experiments with microsomes-plus-supernatant-stock suspensions and an incubationtime of 60min at 37°C, the mean stimulation inmalonaldehyde production due to carbon tetra-chloride was 64+13% (mean of 68 experiments±5.E.M.). The wide variability in the stimulationproduced by carbon tetrachloride in microsomes-plus-supernatant-stock suspensions is discussedbelow.
Vol. 123 8007
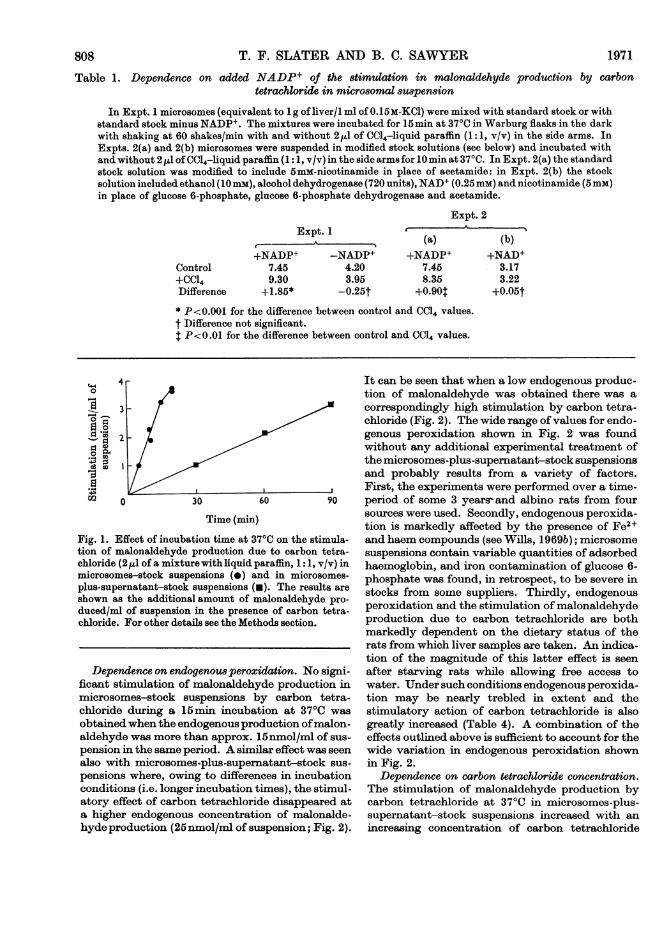
Table 1. Dependence on added NADP+ of the 8timulation in malonaldehyde production by carbontetrachloride in micro8omal 8uspen8ion
In Expt. 1 microsomes (equivalent to 1 g of liver/i ml of0.15M-KC1) were mixed with standard stock or withstandard stock minus NADP+. The mixtures were incubated for 15min at 370C in Warburg flasks in the darkwith shaking at 60 shakes/min with and without 2,pl of CC14-liquid paraffin (1:1, v/v) in the side arms. InExpts. 2(a) and 2(b) microsomes were suspended in modified stock solutions (see below) and incubated withand without 2,u of CC14-liquid paraffin (1:1, v/v) in the side arms for lOmin at 370C. In Expt. 2(a) the standardstock solution was modified to include 5mM-nicotinamide in place of acetamide: in Expt. 2(b) the stocksolution included ethanol (10 mm), alcohol dehydrogenase (720 units), NAD+ (0.25 mm) and nicotinamide (5 mM)in place of glucose 6-phosphate, glucose 6-phosphate dehydrogenase and acetamide.
Expt. 2Expt. 1
Control+CC14Difference
+NADP+7.459.30
+1.85*
-NADP+4.203.95
-0.25t
(a)+NADP+
7.458.35
+0.90t
(b)+NAD+
3.173.22
+0.05t* P<0.001 for the difference between control and CC14 values.t Difference not significant.t P<0.01 for the difference between control and CC14 values.
4
o
.0 v)mL*_ O
3
2
0 30 60
Time (min)
Fig. 1. Effect of incubation time at 370C on the stimula-tion of malonaldehyde production due to carbon tetra-chloride (2,u1 of a mixture with liquid paraffin, 1:1, v/v) inmicrosomes-stock suspensions (e) and in microsomes-plus-supernatant-stock suspensions (m). The results are
shown as the additional amount of malonaldehyde pro-
duced/ml of suspension in the presence of carbon tetra-chloride. For other details see the Methods section.
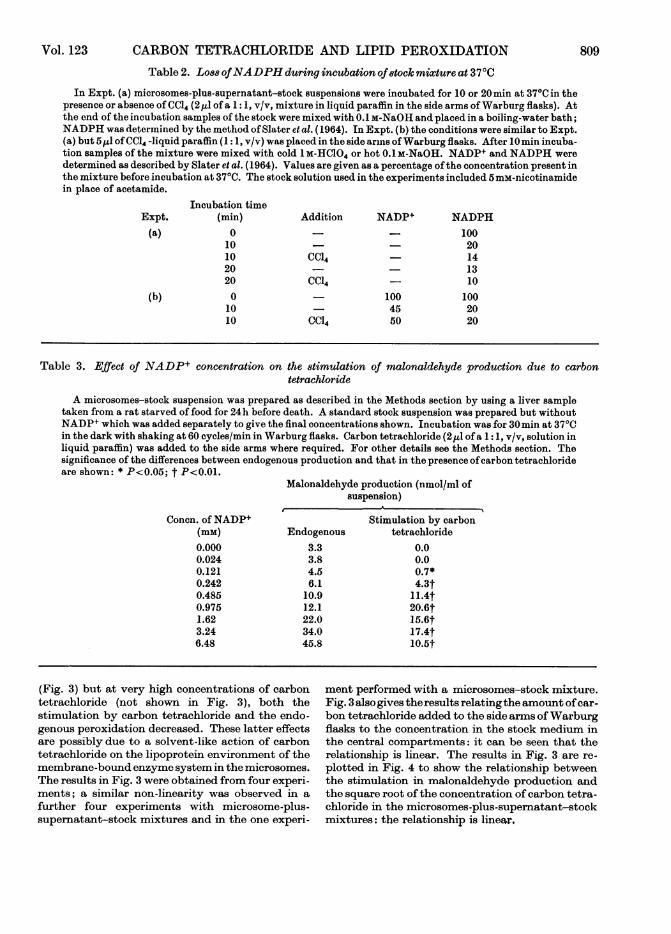
Dependence on endogenoua peroxidation. No signi-ficant stimulation of malonaldehyde production inmicrosomes-stock suspensions by carbon tetra-chloride during a 1imin incubation at 370C was
obtained when the endogenous production ofmalon-aldehyde was more than approx. 15nmol/ml of sus-
pension in the same period. A similar effect was seen
also with microsomes-plus-supematant-stock sus-
pensions where, owing to differences in incubationconditions (i.e. longer incubation times), the stimul-atory effect of carbon tetrachloride disappeared ata higher endogenous concentration of malonalde-hyde production (25 nmol/ml of suspension; Fig. 2).
It can be seen that when a low endogenous produc-tion of malonaldehyde was obtained there was acorrespondingly high stimulation by carbon tetra-chloride (Fig. 2). The wide range ofvalues for endo-genous peroxidation shown in Fig. 2 was foundwithout any additional experimental treatment ofthe microsomes-plus-supernatant-stock suspensionsand probably results from a variety of factors.First, the experiments were performed over a time-period of some 3 years-and albino rats from foursources were used. Secondly, endogenous peroxida-tion is markedly affected by the presence of Fe2+and haem compounds (see Wills, 1969b); microsomesuspensions contain variable quantities of adsorbedhaemoglobin, and iron contamination of glucose 6-phosphate was found, in retrospect, to be severe instocks from some suppliers. Thirdly, endogenousperoxidation and the stimulation of malonaldehydeproduction due to carbon tetrachloride are bothmarkedly dependent on the dietary status of therats from which liver samples are taken. An indica-tion of the magnitude of this latter effect is seenafter starving rats while allowing free access towater. Under such conditions endogenous peroxida-tion may be nearly trebled in extent and thestimulatory action of carbon tetrachloride is alsogreatly increased (Table 4). A combination of theeffects outlined above is sufficient to account for thewide variation in endogenous peroxidation shownin Fig. 2.
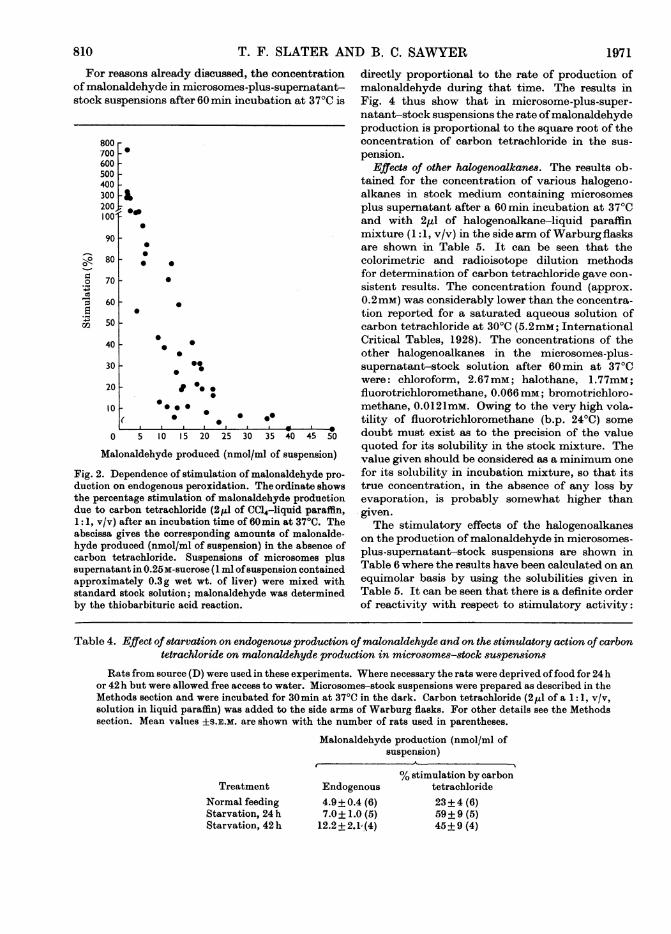
Dependence on carbon tetrachloride concentration.The stimulation of malonaldehyde production bycarbon tetrachloride at 370C in microsomes-plus-supernatant-stock suspensions increased with anincreasing concentration of carbon tetrachloride
I t
808 T. F. SLATER AND B. C. SAWYER 1971
CARBON TETRACHLORIDE AND LIPID PEROXIDATIONTable 2. L088 ofNADPHduring incubation of8tock mixture at 370C
In Expt. (a) microsomes-plus-supernatant-stock suspensions were incubated for 10 or 20min at 37°C in thepresence or absence of CCl4 (2,u of a 1:1, v/v, mixture in liquid paraffin in the side arms of Warburg flasks). Atthe end of the incubation samples of the stock were mixed with 0.1 M-NaOH and placed in a boiling-water bath;NADPH was determined by the method ofSlater et at. (1964). In Expt. (b) the conditions were similar to Expt.(a) but 5,ul ofCC14 -liquid paraffin (1:1, v/v) was placed in the side arms ofWarburg flasks. After lOmin incuba-tion samples of the mixture were mixed with cold 1 M-HC104 or hot 0.1 M-NaOH. NADP+ and NADPH weredetermined as described by Slater et al. (1964). Values are given as a percentage ofthe concentration present inthe mixture before incubation at 370C. The stock solution used in the experiments included 5 mM-nicotinamidein place of acetamide.
Incubation time(min)
01010202001010
Addition NADP+ NADPH
CC14
CC14
CC14
1004550
10020141310
1002020
Table 3. Effect of NADP+ concentration on the 8timulation of malonaldehyde production due to carbontetrachloride
A microsomes-stock suspension was prepared as described in the Methods section by using a liver sampletaken from a rat starved of food for 24h before death. A standard stock suspension was prepared but withoutNADP+ which was added separately to give the final concentrations shown. Incubation was for 30 min at 37°Cin the dark with shaking at 60 cycles/min in Warburg flasks. Carbon tetrachloride (2,u1 ofa 1: 1, v/v, solution inliquid paraffin) was added to the side arms where required. For other details see the Methods section. Thesignificance of the differences between endogenous production and that in the presence ofcarbon tetrachlorideare shown: * P<0.05; t P<0.01.
Malonaldehyde production (nmol/ml ofsuspension)
Concn. of NADP+(mM)0.0000.0240.1210.2420.4850.9751.623.246.48
Endogenous
3.33.84.56.1
10.912.122.034.045.8
Stimulation by carbontetrachloride
0.00.00.7*4.3t
11.4t20.6t15.6t17.4t10.5t
(Fig. 3) but at very high concentrations of carbontetrachloride (not shown in Fig. 3), both thestimulation by carbon tetrachloride and the endo-genous peroxidation decreased. These latter effectsare possibly due to a solvent-like action of carbontetrachloride on the lipoprotein environment of themembrane-bound enzyme system in the microsomes.The results in Fig. 3 were obtained from four experi-ments; a similar non-linearity was observed in a
further four experiments with microsome-plus-supernatant-stock mixtures and in the one experi-
ment performed with a microsomes-stock mixture.Fig. 3 also gives the results relatingthe amount ofcar-bon tetrachloride added to the side arms ofWarburgflasks to the concentration in the stock medium inthe central compartments: it can be seen that therelationship is linear. The results in Fig. 3 are re-
plotted in Fig. 4 to show the relationship betweenthe stimulation in malonaldehyde production andthe square root of the concentration ofcarbon tetra-chloride in the microsomes-plus-supernatant-stockmixtures: the relationship is linear.
Expt.(a)
(b)
Vol. 123 809
T. F. SLATER AND B. C. SAWYERFor reasons already discussed, the concentration
ofmalonaldehyde in microsomes-plus-supernatant-stock suspensions after 60 min incubation at 3700 is
800700600500400300200100'
90
800
04-D
ci
-c
Go
-0
AII 0*4
0
0S0
70 F
60
50
40
30
20
10
0
0
0
0
w0
0 0
0 %00
( * *. *( 0 0 @0
0 5 10 1 5 20 25 30 35 40 45 50
Malonaldehyde produced (nmol/ml of suspension)
Fig. 2. Dependence of stimulation of malonaldehyde pro-duction on endogenous peroxidation. The ordinate showsthe percentage stimulation of malonaldehyde productiondue to carbon tetrachloride (2,ul of CC14-liquid paraffin,1:1, v/v) after an incubation time of 60min at 370C. Theabscissa gives the corresponding amounts of malonalde-hyde produced (nmol/ml of suspension) in the absence ofcarbon tetrachloride. Suspensions of microsomes plussupernatant in 0.25 M-sucrose (1 ml ofsuspension containedapproximately 0.3g wet wt. of liver) were mixed withstandard stock solution; malonaldehyde was determinedby the thiobarbituric acid reaction.
directly proportional to the rate of production ofmalonaldehyde during that time. The results inFig. 4 thus show that in microsome-plus-super-natant-stock suspensions the rate ofmalonaldehydeproduction is proportional to the square root of theconcentration of carbon tetrachloride in the sus-pension.
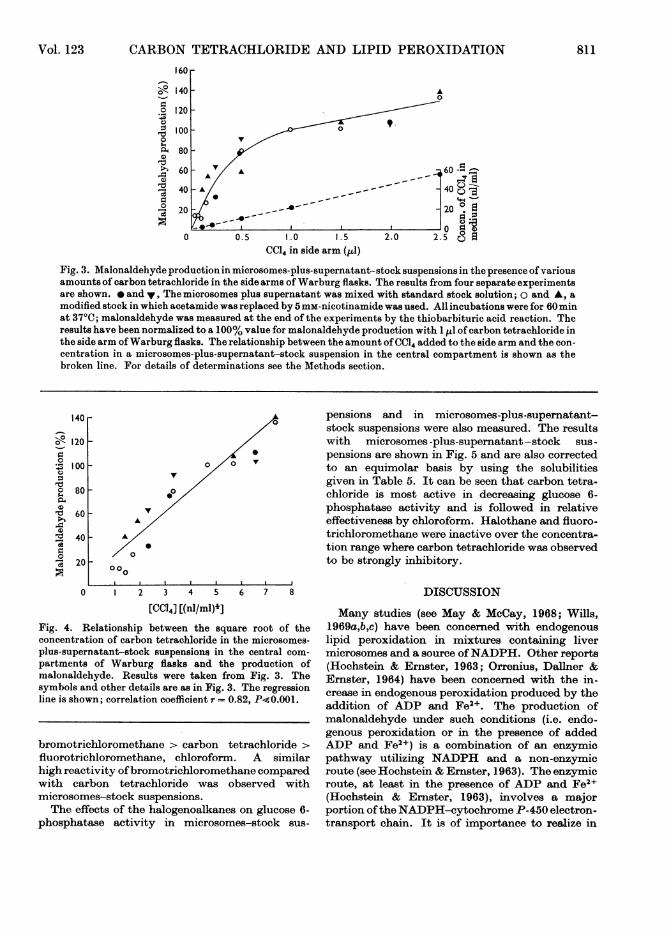
Effects of other halogenoalkanes. The results ob-tained for the concentration of various halogeno-alkanes in stock medium containing microsomesplus supernatant after a 60 min incubation at 370Cand with 2,1j of halogenoalkane-liquid paraffinmixture (1 : 1, v/v) in the side arm of Warburg flasksare shown in Table 5. It can be seen that thecolorimetric and radioisotope dilution methodsfor determination of carbon tetrachloride gave con-sistent results. The concentration found (approx.0.2mM) was considerably lower than the concentra-tion reported for a saturated aqueous solution ofcarbon tetrachloride at 30°C (5.2mM; InternationalCritical Tables, 1928). The concentrations of theother halogenoalkanes in the microsomes-plus-supernatant-stock solution after 60 min at 370Cwere: chloroform, 2.67mM; halothane, 1.77mM;fluorotrichloromethane, 0.066mM; bromotrichloro-methane, 0.0121mM. Owing to the very high vola-tility of fluorotrichloromethane (b.p. 240C) somedoubt must exist as to the precision of the valuequoted for its solubility in the stock mixture. Thevalue given should be considered as a minimum onefor its solubility in incubation mixture, so that itstrue concentration, in the absence of any loss byevaporation, is probably somewhat higher thangiven.The stimulatory effects of the halogenoalkanes
on the production ofmalonaldehyde in microsomes-plus-supernatant-stock suspensions are shown inTable 6 where the results have been calculated on anequimolar basis by using the solubilities given inTable 5. It can be seen that there is a definite orderof reactivity with respect to stimulatory activity:
Table 4. Effect of starvation on endogenous production ofmalonaldehyde and on the stimulatory action of carbontetrachloride on malonaldehyde production in microsomes-stock su8pensions
Rats from source (D) were used in these experiments. Where necessary the rats were deprived offood for 24hor 42 h but were allowed free access to water. Microsomes-stock suspensions were prepared as described in theMethods section and were incubated for 30min at 370C in the dark. Carbon tetrachloride (2,1I of a 1:1, v/v,solution in liquid paraffin) was added to the side arms of Warburg flasks. For other details see the Methodssection. Mean values ±s.E.M. are shown with the number of rats used in parentheses.
Malonaldehyde production (nmol/ml ofsuspension)
TreatmentNormal feedingStarvation, 24 hStarvation, 42 h
Endogenous
4.9±0.4 (6)7.0± 1.0 (5)
12.2 + 2.1(4)
% stimulation by carbontetrachloride23i4 (6)59±9 (5)45+9 (4)
810 1971
a
CARBON TETRACHLORIDE AND LIPID PEROXIDATION
0-~
0.5 1.0 1.5CC]4 in side arm (,ul)
60.-_ 40 v g
-
20 °'~~~~;o
0 22.0 2.5 c0
Fig. 3. Malonaldehyde production in microsomes-plus-supernatant--stock suspensions in tlle presence of variousamounts of carbon tetrachloride in the side arms of Warburg flasks. The results from four separate experimentsare shown. * and y, The microsomes plus supernatant was mixed with standard stock solution; o and A, amodified stock in which acetamide was replaced by 5 mM-nicotinamide was used. All incubations were for 60minat 37°C; malonaldehyde was measured at the end of the experiments by the thiobarbituric acid reaction. Theresults have been normalized to a 100% value for malonaldehyde production with 1 ,ul ofcarbon tetrachloride inthe side arm ofWarburg flasks. The relationship between the amount of CC14 added to the side arm and the con-centration in a microsomes-plus-supernatant-stock suspension in the central compartment is shown as thebroken line. For details of determinations see the Methods section.
0
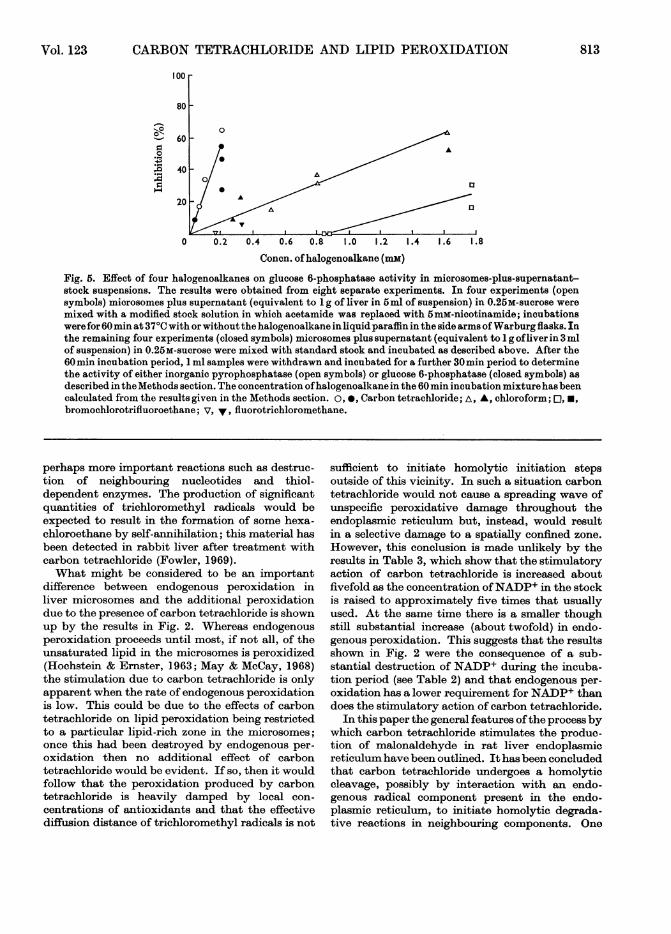
pensions and in microsomes-plus-supernatant-stock suspensions were also measured. The resultswith microsomes-plus-supernatant-stock sus-pensions are shown in Fig. 5 and are also correctedto an equimolar basis by using the solubilitiesgiven in Table 5. It can be seen that carbon tetra-chloride is most active in decreasing glucose 6-phosphatase activity and is followed in relativeeffectiveness by chloroform. Halothane and fluoro-trichloromethane were inactive over the concentra-tion range where carbon tetrachloride was observedto be strongly inhibitory.
0 2 3 4 5 6 7 8[CC14] [(nl/ml)+]
Fig. 4. Relationship between the square root of theconcentration of carbon tetrachloride in the microsomes-plus-supernatant-stock suspensions in the central com-partments of Warburg flasks and the production ofmalonaldehyde. Results were taken from Fig. 3. Thesymbols and other details are as in Fig. 3. The regressionline is shown; correlation coefficient r = 0.82, P.<0.001.
bromotrichloromethane > carbon tetrachloride >
fluorotrichloromethane, chloroform. A similarhigh reactivity ofbromotrichloromethane comparedwith carbon tetrachloride was observed withmicrosomes-stock suspensions.The effects of the halogenoalkanes on glucose 6-
phosphatase activity in microsomes-stock sus-
DISCUSSION
Many studies (see May & McCay, 1968; Wills,1969a,b,c) have been concerned with endogenouslipid peroxidation in mixtures containing livermicrosomes and a source ofNADPH. Other reports(Hochstein & Ernster, 1963; Orrenius, Dallner &Ernster, 1964) have been concerned with the in-crease in endogenous peroxidation produced by theaddition of ADP and Fe2+. The production ofmalonaldehyde under such conditions (i.e. endo-genous peroxidation or in the presence of addedADP and Fe2+) is a combination of an enzymicpathway utilizing NADPH and a non-enzymicroute (see Hochstein & Ernster, 1963). The enzymicroute, at least in the presence of ADP and Fe2+(Hochstein & Ernster, 1963), involves a majorportion ofthe NADPH-cytochrome P-450 electron-transport chain. It is of importance to realize in
Vol. 123160
>0!0
0-4
0
S-
$:1
Q
811
140 r
120 I
l00o
80 F
60 F
oo-1~0'0
0-4
'S-cc
S
0
IC
v
P./
40 F
20
vA
0
000

Table 5. Concentrations of halogenoalkanes in microsoMes-plu8-supernatant-standard stock suspensions(2.5ml) in the central compartments of Warburgflasks
The side arms of the flasks contained 1 ,ul of the halogenoalkane as a 1:1 (v/v) solution in liquid paraffin.Incubation time was 60min at 370C. Details of the experimental procedures used are given in the Methodssection.
HalogenoalkaneCCI4
CEC013CFCl3CBrCl3Halothane
Method of determinationColorimetricRadioisotope dilutionG.l.c.G.l.c.G.l.c.G.l.c.
Conen. in stocksolution (yg/ml)
3134266
92.4
350
Table 6. Comparative effects of halogenoalkanes onmalonaldehyde production in rat liver microsoMes-
plus-supernatant-stock suspensionsIncubations were for 60min at 370C in the dark. The
results are expressed relative to the stimulatory action of21,l of a carbon tetrachloride-liquid paraffin solution(1:1, v/v) in the side arms of Warburg flasks and arecorrected to equimolar concentrations by using the resultsofTable 5. Mean values are given iS.E.M. with the numberof experiments in parentheses. For other details see theMethods section.
HalogenoalkaneCC14CHC13CFC13CBrCl3CF3-CHClBr
Stimulatory effect onmalonaldehyde production
1007+2 (7)
34 (2)3650+ 200 (4)
5+2 (5)
advance of the discussion here, and in followingpapers (Slater & Sawyer, 1971a,b), that the stimula-tion of malonaldehyde production due to carbontetrachloride probably involves a shorter segmentof the NADPH-cytochrome P-450 chain than doesthe endogenous peroxidative route(s) dependent on
NADPH. The two peroxidative pathways (endo-genous peroxidation and the stimulation due tocarbon tetrachloride) can, in fact, be varied inde-pendently of each other; evidence for this state-ment is given in the following paper (Slater &
Sawyer, 1971a).Stimulation of malonaldehyde production by carbon
tetrachloride. The process of lipid peroxidation thatis responsible for the production of malonaldehydein tissue suspensions is a complex reaction involvinga free-radical initiation stage. The stimulation ofmalonaldehyde production by carbon tetrachloridecould represent the chemical involvement of carbontetrachloride as an initiator of lipid peroxidation(this would entail the metabolism of carbon tetra-chloride to free-radical intermediates) or an un-
specific lipophilic action of carbon tetrachloride onthe membranes of the endoplasmic reticulum. Thelatter possibility is unlikely since other halogeno-alkanes with physical properties similar to carbontetrachloride (e.g. chloroform, fluorotrichloro-methane), but which undergo homolysis far lessreadily, are much less active in stimulating malon-aldehyde production (see Table 6). Conversely,bromotrichloromethane, which enters homolyticreaction pathways more readily than carbon tetra-chloride, is more active in stimulating the produc-tion of malonaldehyde in microsomes-plus-super-natant-stock suspensions. These results on therelative reactivities of the halogenomethanes notonly rule out the possibility of the stimulatoryactions on malonaldehyde production being un-specific in character but strongly indicate that ahomolysis of the halogenomethane is necessary forits stimulatory action. The latter indication is sub-stantiated, at least for carbon tetrachloride, by theresults in Figs. 3 and 4. It was found that the rate ofmalonaldehyde production due to the presence ofcarbon tetrachloride was proportional to the squareroot of the concentration of carbon tetrachloride inthe suspension. These results are strong evidencethat initiation of the lipid-peroxidative reaction isby a free-radical product of the homolysis of carbontetrachloride (for a general discussion of this pointin relation to polymerization reactions see Walling,1957). The most likely mechanism for the hom-olysis of carbon tetrachloride in the relatively non-polar environment of the membranes of theendoplasmic reticulum is through an interactionbetween carbon tetrachloride and some endogenous'frozen radical centre' in the membrane. A processof electron capture (Gregory, 1966) would therebyresult in the formation of the highly reactivetrichloromethyl radical:
CCl4+e- - CCl3 +Cl-
the trichloromethyl radical then initiating per-oxidation in neighbouring lipid as well as other and
812 T. F. SLATER AND B. C. SAWYER 1971
CARBON TETRACHLORIDE AND LIPID PEROXIDATION
600-
640 °A
20 , , i,,, ,
0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8
Conen. ofhalogenoalkane (mM)
Fig. 5. Effect of four halogenoalkanes on glucose 6-phosphatase activity in microsomes-plus-supernatant-stock suspensions. The results were obtained from eight separate experiments. In four experiments (opensymbols) microsomes plus supernatant (equivalent to 1 g of liver in 5ml of suspension) in 0.25M-sucrose weremixed with a modified stock solution in which acetamide was replaced with 5mM-nicotinamide; incubationswere for 60min at 37°C with or without the halogenoalkane in liquid paraffin in the side arms ofWarburg flasks. Inthe remaining four experiments (closed symbols) microsomes plus supernatant (equivalent to 1 g ofliver in 3 mlof suspension) in 0.25M-sucrose were mixed with standard stock and incubated as described above. After the60min incubation period, 1 ml samples were withdrawn and incubated for a further 30min period to determinethe activity of either inorganic pyrophosphatase (open symbols) or glucose 6-phosphatase (closed symbols) asdescribed in theMethods section. The concentration ofhalogenoalkane in the 60 min incubation mixturehasbeencalculated from the results given in the Methods section. o, *, Carbon tetrachloride; A, A, chloroform; O,*,bromochlorotrifluoroethane; v, V, fluorotrichloromethane.
perhaps more important reactions such as destruc-tion of neighbouring nucleotides and thiol-dependent enzymes. The production of significantquantities of trichloromethyl radicals would beexpected to result in the formation of some hexa-chloroethane by self-annihilation; this material hasbeen detected in rabbit liver after treatment withcarbon tetrachloride (Fowler, 1969).What might be considered to be an important
difference between endogenous peroxidation inliver microsomes and the additional peroxidationdue to the presence of carbon tetrachloride is shownup by the results in Fig. 2. Whereas endogenousperoxidation proceeds until most, if not all, of theunsaturated lipid in the microsomes is peroxidized(Hochstein & Ernster, 1963; May & McCay, 1968)the stimulation due to carbon tetrachloride is onlyapparent when the rate of endogenous peroxidationis low. This could be due to the effects of carbontetrachloride on lipid peroxidation being restrictedto a particular lipid-rich zone in the microsomes;once this had been destroyed by endogenous per-oxidation then no additional effect of carbontetrachloride would be evident. If so, then it wouldfollow that the peroxidation produced by carbontetrachloride is heavily damped by local con-centrations of antioxidants and that the effectivediffusion distance of trichloromethyl radicals is not
sufficient to initiate homolytic initiation stepsoutside of this vicinity. In such a situation carbontetrachloride would not cause a spreading wave ofunspecific peroxidative damage throughout theendoplasmic reticulum but, instead, would resultin a selective damage to a spatially confined zone.However, this conclusion is made unlikely by theresults in Table 3, which show that the stimulatoryaction of carbon tetrachloride is increased aboutfivefold as the concentration ofNADP+ in the stockis raised to approximately five times that usuallyused. At the same time there is a smaller thoughstill substantial increase (about twofold) in endo-genous peroxidation. This suggests that the resultsshown in Fig. 2 were the consequence of a sub-stantial destruction of NADP+ during the incuba-tion period (see Table 2) and that endogenous per-oxidation has a lower requirement for NADP+ thandoes the stimulatory action of carbon tetrachloride.
In this paper the general features ofthe process bywhich carbon tetrachloride stimulates the produc-tion of malonaldehyde in rat liver endoplasmicreticulum have been outlined. It has been concludedthat carbon tetrachloride undergoes a homolyticcleavage, possibly by interaction with an endo-genous radical component present in the endo-plasmic reticulum, to initiate homolytic degrada-tive reactions in neighbouring components. One
Vol. 123 813

814 T. F. SLATER AND B. C. SAWYER 1971
such degradative reaction is lipid peroxidation thatis capable of ready demonstration by the measure-ment of malonaldehyde production. As such,malonaldehyde production is an indication ofhomo-lytic reactions involving carbon tetrachloride, ofwhich lipid peroxidation is only one amongseveral potentially degradative reactions that wouldensue. The relative importance of such reactionsto the necrogenic action of carbon tetrachloridein vivo is not known but remains a problem of con-siderable interest. It is of obvious importance toidentify the site ofinteraction between carbon tetra-chloride and the endoplasmic reticulum that isresponsible for the homolytic consequences, and toinvestigate the effects of a number of free-radicalscavengers on the deleterious homolytic reactionsin vitro and in vivo. These studies are reported inthe following papers (Slater & Sawyer, 1971a,b).We are grateful to the Medical Research Council for
financial assistance. The sample oftrichlorofluoromethanewas generously given by I.C.I. Ltd., PharmaceuticalsDivision; Dr D. S. Corrigan and Mr M. Rackham of I.C.I.Ltd., Pharmaceuticals Division, very kindly carried outthe gas-liquid-chromatographic analyses for us.
REFERENCESBray, G. A. (1960). Analyt. Biochem. 1, 279.Butler, T. C. (1961). J. Pharmac. exp. Ther. 134,311.Comporti, M., Saccocci, C. & Dianzani, M. U. (1965).
Enzymologia, 29, 185.Delaney, V. B. & Slater, T. F. (1970). Biochem. J. 116,
299.Fowler, J. S. L. (1969). Br. J. Pharmac. Chemother. 37,
733.
Franz, J. & Cole, B. T. (1962). Arche Biochem. Biophy8.96, 382.
Ghoshal, A. K. & Recknagel, R. 0. (1965). Life Sci. 4,1521.
Gordon, C. F. & Wolfe, A. L. (1960). Analyt. Chem. 32,574.
Gregory, N. L. (1966). Nature, Lond., 212, 1460.Hochstein, P. & Ernster, L. (1963). Biochem. biophys.
Res. Commun. 12, 388.Hove, E. L. (1948). Archs Biochem. 17,467.May, H. E. & McCay, P. B. (1968). J. biol. Chem. 243,2296.Orrenius, S., Dallner, G. & Ernster, L. (1964). Biochem.
biophy8. Res. Commun. 14, 329.Recknagel, R. 0. (1967). Pharmac. Rev. 19, 145.Recknagel, R. 0. & Ghoshal, A. K. (1966). Lab. Invest.
15, 132.Recknagel, R. 0. & Litteria, M. (1960). Am. J. Path. 36,
521.Saslow, L. D., Corwin, L. M. & Waravdekar, V. S. (1966).
Arch. Biochem. Biophys. 114, 61.Slater, T. F. (1966a). Nature, Lond., 209, 36.Slater, T. F. (1966b). Biochem. J. 101, 16P.Slater, T. P. & Sawyer, B. C. (1969). Biochem. J. 111,
317.Slater, T. F. & Sawyer, B. C. (1971a). Biochem. J. 123, 815.Slater, T. F. & Sawyer, B. C. (1971b). Biochem. J. 123,823.Slater, T. F., Sawyer, B. C. & StrAuli, U. D. (1964). Arch.
int. Phy8iol. Biochim. 72, 427.Swoboda, P. A. T. & Lea, C. H. (1958). Chemy Ind. p.
1090.Walling, C. H. (1957). Free Radicals in Solution. NewYork: John Wiley and Sons Inc.
Wills, E. D. (1969a). Biochem. J. 113, 315.Wills, E. D. (1969b). Biochem. J. 113, 325.Wills, E. D. (1969c). Biochem. J. 113, 333.Wirtschafter, Z. T. & Cronyn, M. W. (1964). Environ.
Hlth, 9, 186.





























