Embed Size (px)
Citation preview

The Study of Genetic Diversity in Khazak Native Chicken Using
Whole Genome Sequencing
Elaheh Rostamzadeh Mahdabi
PhD Student, Department of Animal Science, Shahid Bahonar University of Kerman, Kerman,
Iran. E-mail: [email protected]
Masood Asadi Fozi
* Corresponding author. Professor, Department of Animal Science, Shahid Bahonar University
of Kerman, Kerman, Iran. E-mail: [email protected]
Ali Esmailizadeh
Professor, Department of Animal Science, Shahid Bahonar University of Kerman, Kerman, Iran.
E-mail: [email protected]
Ahmad Ayatollahi Mehrgardi
Associate Professor, Department of Animal Science, Shahid Bahonar University of Kerman,
Kerman, Iran. E-mail: [email protected]
Seyed Hojat Masoudzadeh
MSc. of animal breeding, Department of Animal Science, Shahid Bahonar University of Kerman,
Kerman, Iran. E-mail: [email protected]
Abstract Objective
Indigenous animals are considered as the national treasures and strategic sources of each country.
It is essential to preserve and reproduce them. Since no artificial selection has been employed in
Iranian native breeds, there is considerable diversity in their genome. Therefore, having detailed
information on the animals’ genome can be important for designing breeding plans in the future.
The aim of this study was to identify the functional effects of single nucleotide polymorphisms
and investigate the genetic diversity in the Khazak chicken ecotype using sequence information
of the whole genome.
Materials and methods
In this study, 17 Khazak chicken samples were sequenced using Next Generation Sequencing
(NGS) technique. After preprocessing short sequences using FastQC, Trimmomatic, BWA,

92
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
Picard software and the GATK program, single nucleotide polymorphisms were identified in
GATK program using the UnifiedGenotyper tool. Samtools software was applied to calculate
short reads coverage and the percentage of read alignment with the reference genome. The
filtering steps of SNPs were performed using Plink software. The genetic diversity within the
target population was calculated using the VCFtools program.
Results
The mean coverage depth for 17 chickens was 6.45. After preprocessing the short sequences, were
identified 15,981,176 and 10,552,735 SNPs before and after the quality control, respectively.
Functional annotation of the single nucleotide polymorphisms indicated that most of the SNPs
are located in the intron and intergenic regions and only 2.8% of them belong to the exon region.
The rate of Transition to transversion mutations (Ti / Tv) was 2.43 and the ratio of heterozygous to
homozygous SNPs was 3.3 in the Khazak birds, on average.
Conclusions
The higher number of heterozygous SNPs indicates that there is a high genetic diversity in the
Khazak ecotype. Like other Iranian native chickens, the ecotype has the genetic diversity that can
be considered a native laying ecotype in the breeding programs.
Keywords: Genetic diversity, single nucleotide polymorphisms, Khazak native chicken.
Paper Type: Research Paper.
Citation: Rostamzadeh Mahdabi E, Asadi Fozi M, Esmailizadeh A, Ayatollahi A, Masoudzadeh
H (2021) The study of genetic diversity in Khazak native chicken using whole genome
sequencing. Agricultural Biotechnology Journal 13 (3), 91-106.
Agricultural Biotechnology Journal 13 (3), 91-106. DOI: 10.22103/jab.2021.17202.1296
Received: July 20, 2021. Accepted: August 22, 2021.
Publisher: Faculty of Agriculture and Technology Institute of Plant
Production, Shahid Bahonar University of Kerman-Iranian
Biotechnology Society.
© the authors

یابی کل ژنومهای توالیادهمرغ بومی خزک با استفاده از دبررسی تنوع ژنتیکی در
مهدابی زادهالهه رستم
رايانامه:. ژنتيك و اصلاح نژاد دام، بخش علوم دامی، دانشكدة کشاورزی، دانشگاه شهيد باهنر کرمان، ايراندانشجوی دکتری
مسعود اسدی فوزی
[email protected] رايانامه: .دانشكدة کشاورزی، دانشگاه شهيد باهنر کرمان، ايراناستاد بخش علوم دامی، :نويسنده مسئول*
زادهعلی اسمعيلی
[email protected] رايانامه: .استاد بخش علوم دامی، دانشكدة کشاورزی، دانشگاه شهيد باهنر کرمان، ايران
مهرجردی احمد آيت الهی
[email protected] رايانامه: .دانشيار بخش علوم دامی، دانشكدة کشاورزی، دانشگاه شهيد باهنر کرمان، ايران
زادهسيد حجت مسعود
بخش علوم دامی، دانشكدة کشاورزی، دانشگاه شهيد باهنر کرمان، دانش آموخته مقطع کارشناسی ارشد ژنتيك و اصلاح دام،
[email protected] رايانامه: .ايران
31/05/1400تاريخ پذيرش: 29/04/1400تاريخ دريافت:
چکیده
رزش و ها از اشرروند. ح و و تكرير آنکشرروری موسررو میاير اسررتراتژيك هر حيوانات بومی به عنوان سرررمايه ملی و خ هدف:
ل تنوع قاب ،انجام نشده است ايران انتخا مصنوعی در مرغان نژاد بومیتاکنون باشد. با توجه به اينكه اهميت بسياری برخوردار می
های اصلاح نژادی انات برای برنامهتر ژنوم اين حيواين رو کسب اطلاعات و شناخت دقيقاز .شودها مشاهده میتوجهی در ژنوم آن
های تك نوکلئوتيدی و همچنين بنابراين، هدف از انجام اين توقيق، شناسايی و بررسی آثار عملكردی چندشكلی .باشدضروری می
باشد.مرغ بومی خزک با است اده از اطلاعات توالی کل ژنوم می 17بررسی تنوع ژنتيكی در

(1400 زمستان، 4، شماره 13مجله بیوتکنولوژی کشاورزی )دوره
94
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
ست اده از روش توالی 17 تعداد در اين پژوهش، :هاروشمواد و شد. بعد از ( توالیNGSيابی نسل بعد )نمونه مرغ خزک با ا يابی
و ابزارهای FastQC ،Trimmomatic ،BWA ،Picardافزارهای های کوتاه با است اده از نرمانجام مراحل پيش پردازش توالی
GATKدر برنامه UnifiedGenotyperاز ابزار دههای تك نوکلئوتيدی با اسرررت اچند شررركلی در نهايت، GATKبرنامه
است اده Samtoolsافزار با ژنوم مرجع از نرم هاها و درصد الاينمنت خوانشخوانش مواسبه ميزان پوشششناسايی شدند. جهت
انجام شررد، سررنو تنوع ژنتيكی درون Plinkهای تك نوکلئوتيدی با اسررت اده از نرم افزار . مراحل کنترل کي يت چندشرركلیشررد
مواسبه شد. VCFtoolsنظر با برنامه مورد جمعيت
شش ميانگين :نتایج ست آمد. 45/6خزک مرغ 17 برای هاتوالیعمق پو و 15،981،176های کوتاه، تعداد بعد از پردازش توالیبد
10،552،735 SNP شد سايی شنا شيه. نتايج به ترتيب قبل و بعد از مرحله کنترل کي يت های تك نويسی عملكردی چند شكلیحا
درصرررد آنها متعلق به ناحيه انزون 8/2ها در نواحی اينترون و بين ژنی قرار دارند و تنها SNPنوکلئوتيدی نشررران داد که اکرر
های هتروزيگوت به هموزيگوت به طور SNPنسبت و 43/2( Ti / Tvهای متقاطع )های انتقالی به جهشنسبت جهشباشند. می
.بدست آمد 3/3ميانگين در مرغان خزک
اين اکوتيپ مانند . باشدی تنوع ژنتيكی بالا در اکوتيپ خزک مینشان دهنده ،های هتروزيگوت بالاترSNPتعداد گیری:نتیجه
های اصلاح نژاد حائز نذار بومی در برنامهتواند به عنوان يك اکوتيپ تخمباشد که میتنوع میهای بومی ايران دارای ساير اکوتيپ
اهميت باشد.
تنوع ژنتيكی، چند شكلی تك نوکلئوتيدی، مرغ بومی خزک.: هاکلیدواژه
پژوهشی.: نوع مقاله
بررسی تنوع ژنتيكی در (1400) زاده حجتمسعود الهی احمد،آيت ،زاده علیاسمعيلی، اسدی فوزی مسعود، زاده الههرستم استناد:
.106-91(، 3)13، مجله بيوتكنولوژی کشاورزی. يابی کل ژنوم های توالیت اده از دادهمرغ بومی خزک با اس
Publisher: Faculty of Agriculture and Technology Institute of Plant
Production, Shahid Bahonar University of Kerman-Iranian
Biotechnology Society.
© the authors
مقدمه
سال، با توجه به وجود تنوع ژنتيكی بسيار زياد و همچنين سازناری با مويط اطراف ميليون 5/3تكامل حيوانات اهلی طی
، شروع به دستكاری در تكامل نياهان و و با است اده از تنوع ژنتيكی موجود در حيوانات با توجه به نيازهايش است. انسان شكل نرفته
یهاها و مكانزمان ،مطالعات متعدد (Diamond 2002) شودناخته مینيز ش سازیکه اين فرآيند به عنوان اهلی است کرده هاآن

1400، زاده مهدابی و همکارانالهه رستم
95
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
یاز مرغ جنگل اهلیدهند که مرغ یآنها نشان م که اکرراند نزارش کردهی مرغ جنگلی هاشدن مرغ از نونهیاهل را برای یمختل
Sawai et al. 2010; Tixier-Boichard et al. 2011; Peters et) نرفته است نشات ايآس یقرمز در جنو و جنو شرق
al. 2015; Osman et al. 2016). ای بسيار کهن دارد. ايران طريق اين کشور تاريخچه ماکيان در ايران و انتشار آن از پرورش
ميلادی بود و از هند )دهلی( تا درياهای سياه و مديترانه نسترده 7تقريبا قرن قبل از ميلاد تا 5)پرشيا( يك امنراطوری بزرگ از قرن
و بعد از آن، در قرون . در آن زمان(Moazeni et al. 2016a; Mohammadifar and Mohammadabadi 2017) بود
ا برای حمل و نقل موصولات، از قبيل ماکيان از شرق به غر ، هم از طريق خشكی و هم از هوسطی ايران در مول تقاطع راه
های ها نيز توسعه و نسترش جمعيتطريق دريا قرار داشت. جنگ های زيادی در حوالی ايران و کشورهای همسايه در طی اين دوره
Nikbakhti etهای باستان تأييد کرده است )ا در ايران در زمانهای باستان شناسی حضور ماکيان رماکيان را تسهيل کرد. ح اری
al., 2009; Moazeni et al. 2016b بر اساس توقيقات .) West and Zhouهای يافت شده در ايران در سه منطقه استخوان
قبل از ميلاد و 3900تا 3800( )جنو شرقی ايران( به ترتيب متعلق به Tepe Yahyaاند: دو کشف در تنه يويی )وجود داشته
.Mohammadifar et alقبل از ميلاد ) 1000قبل از ميلاد و ديگری در تخت سليمان )شمال غربی ايران( متعلق به 1000
2014; Mohammadifar and Mohammadabadi 2018 .)باشند. های آسيايی میاکرر نژادهای بومی ايران از دسته مرغ
باشد. عوامل مويطی ی نسترده خود دارای آ و هوای بسيار متنوع و در نتيجه توت عوامل مويطی نونانون میکشور ايران در پهنه
به یبوم ینژادها .(Tavakolian 2000ها و تنوع حيوانات مناطق مختلف ايران شده است )مختلف موجب تشكيل انواع زيستگاه
يیروستا یهادر خانواده ینقش مهممرغ بومی باشند.ها دارای اهميت مییماريابر بو مقاومت در بری به شرايط مويطی سازنار ليدل
اقتصاد یبرا یبوم ینژادها تياهمدارد. و ايجاد اشتغال و افزايش درآمد روستاييان بالا تي يبا ک یوانيح نيبه عنوان منبع پروتئ
های بومی توليد تخم پايين و سرعت باشد. مرغمی اديز اريبس ايعمدتاً در آسو افتهيدر حال توسعه و توسعه یدر کشورها ييانروستا
یهاد. مرغنرا دار ینامناسب دامنرور یهاوهيو ش یطيسخت مو طيتومل شرا يیتوانا رشد کندی نسبت به نژادهای تجاری دارند، اما
ها یماريو مقاومت در برابر ب یرينرمس ی با مناطقسازنار، بالا با عملكرداصلاح مرغ یبرا یاصل یهامخزن ژنوم و ژن ینژاد بوم
سازنار و مناسب با توليد نژادهای توليد برای را زمينه تواندمی بومی ژنتيكی مرغان به عبارت ديگر منابع .(Padhi 2016) باشندمی
(.Hoffmann 2005) کند فراهم روستايی شرايط با
، يابیی، معروف به توالدهاينوکلئوت قيدق بيترت نييروش بدست آوردن اطلاعات در مورد ژنوم هر موجود زنده، تع نيترجامع
واريانتها ونيليباعث کشف م (NGS1)جديد نسل يابییتوالظهور .( .2015Venter et alباشد )می DNAکامل یدر توال
یتوان برایم DNA یاز نشانگرها .(Gheyas et al. 2015b) شده استن دارامهره ژنومو اثرات عملكردی آنها در یكيژنت
مانند مختل ی DNA هاینشانگر .(Boettcher et al. 2010) است اده کرد هاتيجمع نيدر داخل و ب یكيتنوع ژنت برآورد
1. Next Generation Sequencing

(1400 زمستان، 4، شماره 13مجله بیوتکنولوژی کشاورزی )دوره
96
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
RAPDs ، AFLPs اند )مرغ است اده شده یكيتنوع ژنت یابيارز یها براساتلايتكرويو مLynch & Milligan 1994;
Abebe et al. 2015.) که به طور خاص SNP ساختار تعيين دقيق و یكيتنوع ژنت برآورد یبرابه دليل تراکم ژنومی بالا، ها
(.Engelsma 2012; Strillacci et al. 2017) رندينیقرار م ادهمورد است تيجمع
شود. اين پرنده به عنوان های ديگر به ندرت يافت میباشد و در بعضی شهرستانخزک، بومی منطقه سيستان )زابل( میمرغ
شناخته میتيپ تخم سط وزن تخم 120مرغ توليدی آنها شود که ميانگين تخمنذار سال و متو شد. نرم می 15/45مرغ عدد در با
نرم 1578و 1245تيب ای کوتاه دارد. وزن بدن در مرغ و خروس اکوتيپ خزک به طور ميانگين به تراين مرغ جره کوچك با پاه
شد به طوریمی شد که اکرر خوراک مصرفی مرغ، که وزن پايين اين اکوتيپ میبا هت توليد تخم مصرف جتواند دارای اين مزيت با
صرفمی Deljoyشود )مرغ توليد میکند و در نتيوه با هزينه کمتری تخممی شود و خوراک کمتری را برای انرژی نگهداری م
Sarayan 2011; Tavakolian 2000.) توان سررت میابا توجه به اينكه انتخا مصررنوعی در مرغان نژاد بومی انجام نشررده
شاهده می شت که تنوع قابل توجهی در ژنوم آنها م سرمايه ح و آنها به عنوان شود. از اين رو لزوم انتظار دا های خاير ژنتيكی و
شناخت دقيقملی برای برنامه سب اطلاعات و صلاح نژادی، ک ضروری میهای ا سازد. هدف از انجام اين تر ژنوم اين حيوانات را
شكلیتوقيق، سی آثار عملكردی چند سايی و برر سی تنوع ژنتيكی شنا کوتيپ مرغ بومی در اهای تك نوکلئوتيدی و همچنين برر
خزک با است اده از اطلاعات توالی کل ژنوم بود.
هامواد و روش
ستخراج های خون مربوط به اکوتيپدر اين پژوهش نمونه شد. ا 17های خون از نمونه DNAخزک از وريد زير بال نرفته
شهيد باهنر کرمان شگاه شگاه ژنتيك بخش علوم دامی دان سانجام نرفت. توالیمرغ خزک در آزماي ت اده از روش يابی کل ژنوم با ا
يابی توسط های خروجی توالیدر کشور چين انجام شد. کي يت اوليه داده Hiseq 2000دستگاه ايلومينا توسط توالی يابی نسل بعد
های کوتاه با کي يت پايين توالی Trimmomaticو با اسررت اده از نرم افزار (Andrews 2010بررسرری شررد ) FastQC برنامه
يش يرا لی .2014Bolger et alشرررردنررد ) 2پ توا ي ی مرد ه جهررت مرغ (. جع مر نوم کوترراه نسررربررت برره ژ هررای
(ftp://ftp.ensembl.org/pub/release-/fasta/gallus_gallus/dna از الگوريتم )MEM در نرم افزارBWA اسرررت اده
ست اده از نرم هادازش دادهسازی و پر خيرهبه منظور عملكرد بهتر، راندمان .(Li & Durbin 2009شد ) ، خروجی مرحله قبل با ا
ها به خصوص در زمان سازی نمونههنگام آماده .(Li et al. 2009) ( تبديل شدBAM fileبه صورت فشرده ) Samtoolsافزار
ای پليمراز به وجود نجيرههای تكراری از يك نمونه يكسررران در طول واکنش زهای ژنومی، تعداد زيادی از توالیسررراخت کتابخانه
2. Trim

1400، زاده مهدابی و همکارانالهه رستم
97
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
های کوتاه تكرار (. به همين جهت اين توالی .2016Ebbert et alشررود )مربت کا می 3هایآيندکه باعث فراخوانی واريانتمی
پردازش ( حذف شدند. بعد از انجام مراحل پيش http://broadinstitute.github.io/picard) Picardشده با است اده از برنامه
های کوتاه و اطراف حذف و اضرررافه 4، جهت همردي ی مجددPicardو Samtools فايل ژنوم مرجع با اسرررت اده از نرم افزار
ها خوانش (. از آنجايی کهMcKenna et al. 2010اسررت اده شررد ) GATKکاليبراسررون مجدد نمره کي يت بازها از نرم افزار
شرررود. تعداد يابی میيك ناحيه ژنومی بيش از يك بار توالی ،يابی باشرررندند حاوی خطاهای توالیتوانو می معمولاً کوتاه هسرررتند
مواسبه ميزان پوششد. در مطالعه حاضر، جهت کنژنوم را مشخص میيابی توالی 5های همنوشان با يك ناحيه ژنومی عمقخوانش
صد خوانش يا عمق شد Samtoolsافزار ژنوم مرجع از نرم ها باخوانش همردي یها و در ست اده شكلیا های تك . در نهايت چند
فراخوانی شدند. GATKدر برنامه ی UnifiedGenotyper نوکلئوتيدی توسط ابزار
(Z, W)های جنسی کروموزوم SNPsهای اتوزومی بررسی شد و بنابراين درتوقيق حاضر، تنوع ژنتيكی واقع بر کروموزوم
به همين منظور های با کي يت پايين شناسايی و حذف شوند.حذف شدند. جهت آناليز بعدی، لازم است تا داده 6های پيوستهو نروه
های SNPو 05/0کمتر از 7های با فراوانی آللی کميا SNPدرصد ژنوتيپ نمشده، 10افراد با بيش از Plinkبا است اده از نرم افزار
. بعد ح و شدند 8تعيين ژنوتيپ درصد نرخ فراخوانی 90های با SNPبودند حذف شدند و 10-7آنها کمتر از value-Pی که آستانه
انجام SnpEffی عملكردی آنها با است اده از برنامه 9نويسیهای تك نوکلئوتيدی در اکوتيپ خزک، حاشيهاز شناسايی چند شكلی
در kb25با اندازه kb50های کشويی ن مطالعه، تنوع ژنتيكی درون جمعيت خزک در پنجره(. در ايCingolani et al. 2012شد )
(.Danecek et al. 2011) شد اسبهمو VCFtoolsبا است اده از نرم افزار در امتداد هر کروموزوم هر مرحله
نتایج و بحث
يابی . درصد نقشهماند باقی توالی کوتاه به ازای هر فرد ونيليم 64تعداد ميانگينطور ه، بهاداده شيرايو پ تي ياز کنترل ک پو
ميانگين باشد. ی راندمان خو همردي ی با ژنوم مرجع میدهندهدرصد بود که نشان 86/97ها با ژنوم مرجع به طور ميانگين اين توالی
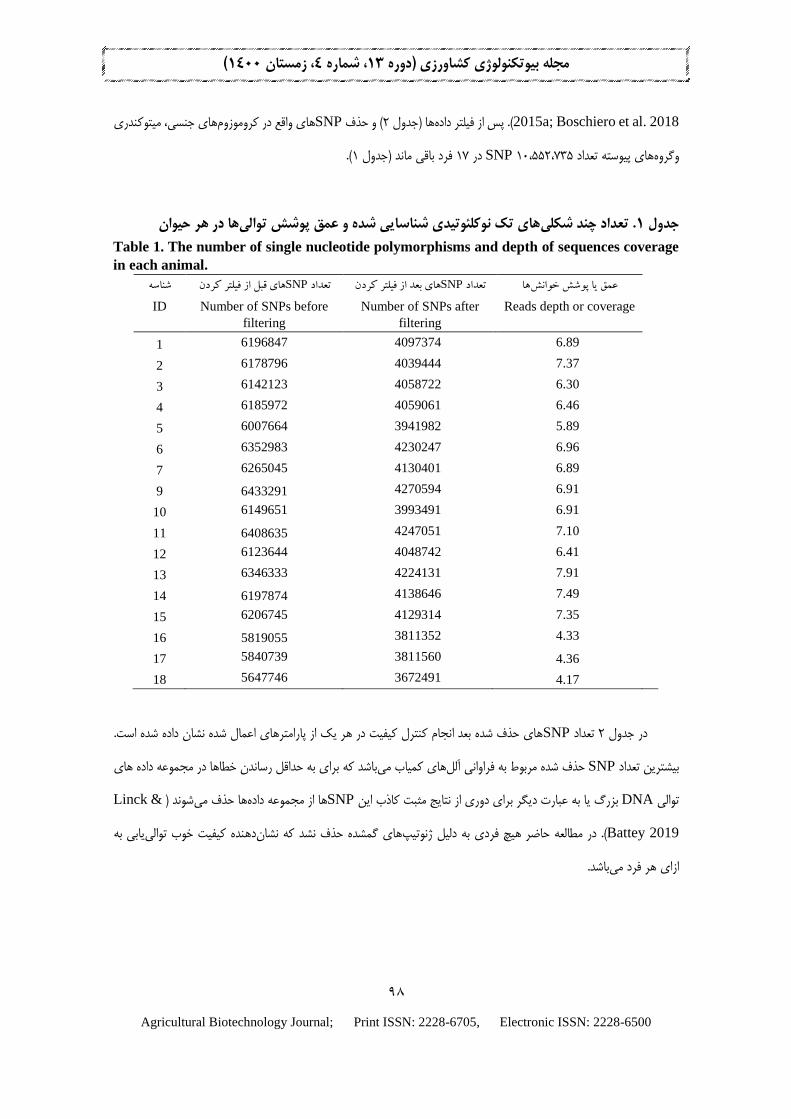
که با نتايج مطالعات شد یاز کل ژنوم فراخوان SNP 15،981،176. در ابتدا (1)جدول بود 45/6خزک فرد 17 برای هاتوالیپوشش
.Gheyas et alسازناری دارد )يابی کل ژنوم های تك نوکلئوتيدی در مرغ با است اده از توالیدر مورد شناسايی چند شكلینذشته
3. Variant calling
4. Realignment
5 Coverage
6. Linkage groups 7. Minor allele frequency
8. Genotyping rate
9. Annotation
(1400 زمستان، 4، شماره 13مجله بیوتکنولوژی کشاورزی )دوره
98
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
2015a; Boschiero et al. 2018 .)( و حذف 2ها )جدول داده لتريپو از فSNP های جنسی، ميتوکندری کروموزومهای واقع در
(.1فرد باقی ماند )جدول 17در SNP 10،552،735 های پيوسته تعدادونروه
ها در هر حیوانهای تک نوکلئوتیدی شناسایی شده و عمق پوشش توالی. تعداد چند شکلی1جدول
Table 1. The number of single nucleotide polymorphisms and depth of sequences coverage
in each animal. شناسه
ID
های قبل از فیلتر کردنSNPتعداد
Number of SNPs before
filtering
های بعد از فیلتر کردنSNPتعداد
Number of SNPs after
filtering
هاعمق یا پوشش خوانش
Reads depth or coverage
1 6196847 4097374 6.89
2 6178796 4039444 7.37
3 6142123 4058722 6.30
4 6185972 4059061 6.46
5 6007664 3941982 5.89
6 6352983 4230247 6.96
7 6265045 4130401 6.89
9 6433291 4270594 6.91
10 6149651 3993491 6.91
11 6408635 4247051 7.10
12 6123644 4048742 6.41
13 6346333 4224131 7.91
14 6197874 4138646 7.49
15 6206745 4129314 7.35
16 5819055 3811352 4.33
17 5840739 3811560 4.36
18 5647746 3672491 4.17
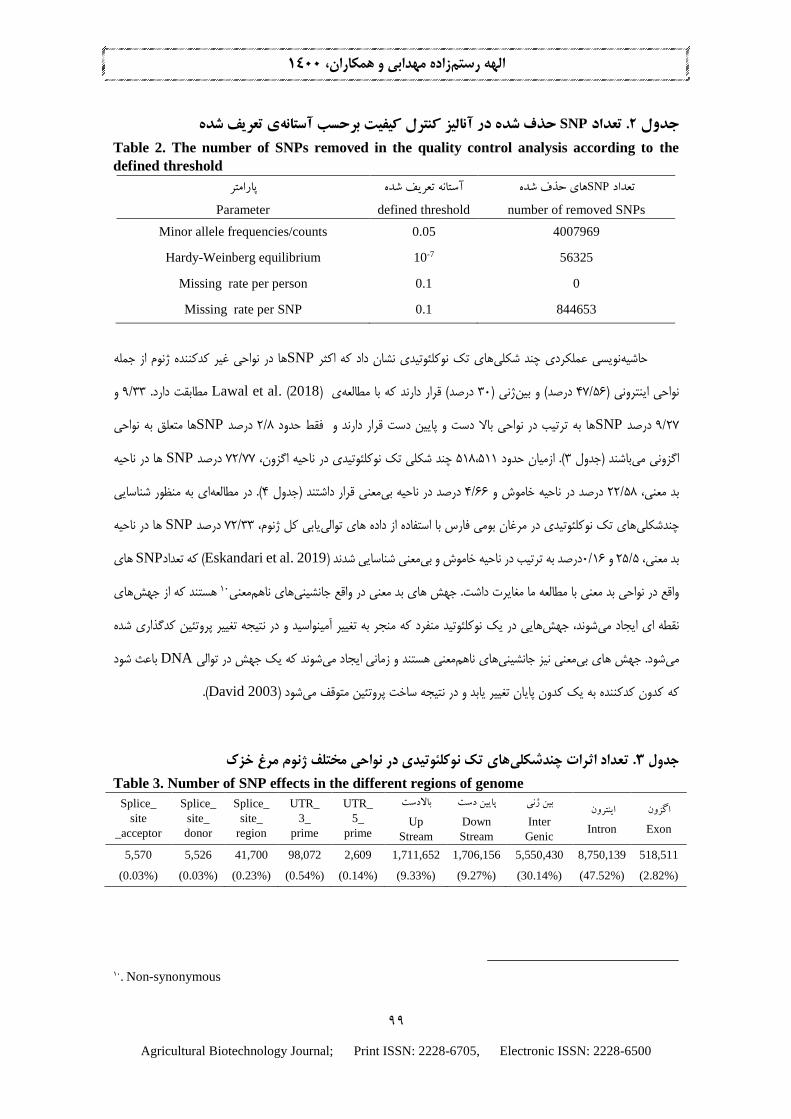
های حذف شده بعد انجام کنترل کي يت در هر يك از پارامترهای اعمال شده نشان داده شده است. SNPتعداد 2در جدول
یبه حداقل رساندن خطاها در مجموعه داده ها یبراباشد که های کميا میحذف شده مربوط به فراوانی آلل SNPبيشترين تعداد
& Linckشوند )ها حذف میها از مجموعه دادهSNPيا به عبارت ديگر برای دوری از نتايج مربت کا اين بزرگ DNA یتوال
Battey 2019) يابی به دهنده کي يت خو توالیهای نمشده حذف نشد که نشاندليل ژنوتيپ. در مطالعه حاضر هيچ فردی به
باشد.ازای هر فرد می
1400، زاده مهدابی و همکارانالهه رستم
99
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
ی تعریف شدهآستانهحذف شده در آنالیز کنترل کیفیت برحسب SNP. تعداد 2جدول
Table 2. The number of SNPs removed in the quality control analysis according to the
defined threshold پارامتر
Parameter
آستانه تعریف شده
defined threshold
های حذف شدهSNPتعداد
number of removed SNPs Minor allele frequencies/counts 0.05 4007969
Hardy-Weinberg equilibrium 10-7 56325
Missing rate per person 0.1 0
Missing rate per SNP 0.1 844653
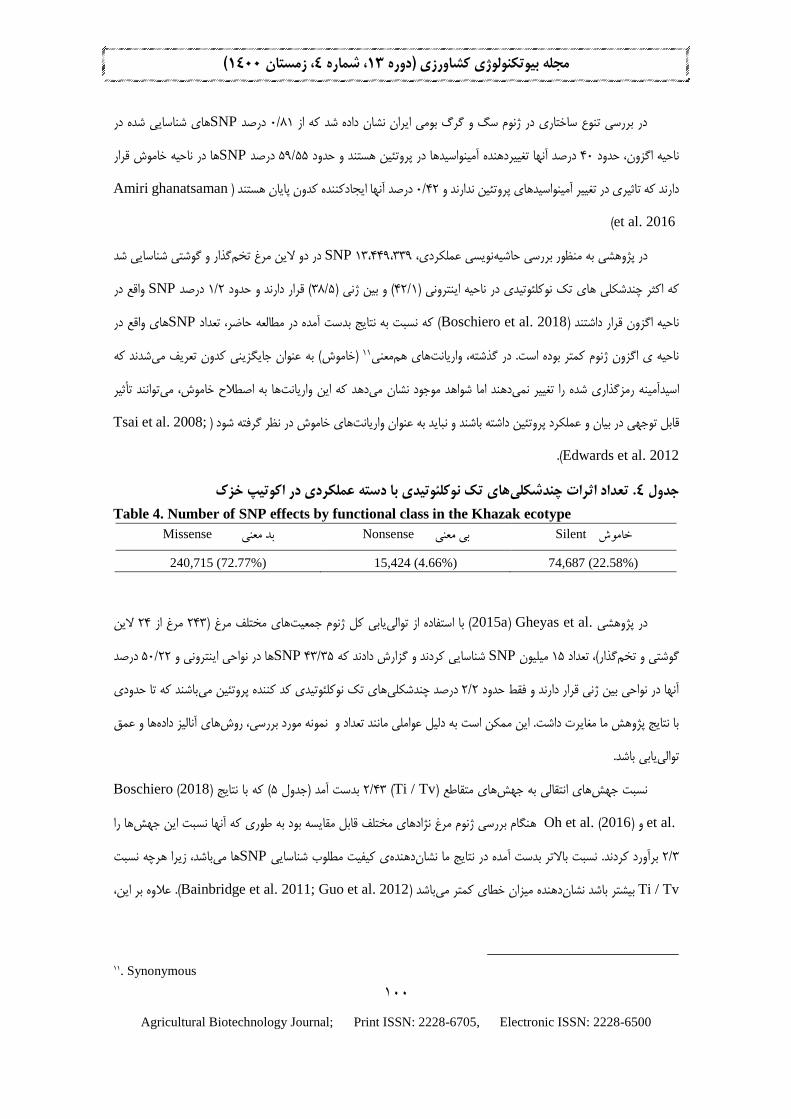
ها در نواحی غير کدکننده ژنوم از جمله SNPهای تك نوکلئوتيدی نشان داد که اکرر نويسی عملكردی چند شكلیحاشيه
و 33/9مطابقت دارد. .Lawal et al(2018) ی درصد( قرار دارند که با مطالعه 30ژنی )درصد( و بين 56/47نواحی اينترونی )
ها متعلق به نواحی SNPدرصد 8/2ها به ترتيب در نواحی بالا دست و پايين دست قرار دارند و فقط حدود SNPدرصد 27/9
ها در ناحيه SNPدرصد 77/72ناحيه انزون، چند شكلی تك نوکلئوتيدی در 518،511(. ازميان حدود 3باشند )جدول انزونی می
ای به منظور شناسايی (. در مطالعه4)جدول معنی قرار داشتنددرصد در ناحيه بی 66/4درصد در ناحيه خاموش و 58/22بد معنی،
ها در ناحيه SNPدرصد 33/72يابی کل ژنوم، های تك نوکلئوتيدی در مرغان بومی فارس با است اده از داده های توالیچندشكلی
های SNP ( که تعدادEskandari et al. 2019معنی شناسايی شدند )درصد به ترتيب در ناحيه خاموش و بی16/0و 5/25بد معنی،
هایکه از جهش هستند 10معنیناهم هایینيجانشدر واقع بد معنی های جهشواقع در نواحی بد معنی با مطالعه ما مغايرت داشت.
شده یکدنذار نيپروتئ رييتغ جهيو در نت دياسنويآمتغيير من رد که منجر به دينوکلئوت كيدر يیهاد، جهشنشویم جاديا ینقطه ا
شود باعث DNAشوند که يك جهش در توالی و زمانی ايجاد می هستند معنیناهم هایینيجانشمعنی نيز بی های جهش شود.یم
(. David 2003شود )کدون کدکننده به يك کدون پايان تغيير يابد و در نتيجه ساخت پروتئين متوقف میکه
های تک نوکلئوتیدی در نواحی مختلف ژنوم مرغ خزک. تعداد اثرات چندشکلی3جدول
Table 3. Number of SNP effects in the different regions of genome Splice_
site
_acceptor
Splice_
site_
donor
Splice_
site_
region
UTR_
3_
prime
UTR_
5_
prime
بالادست
Up
Stream
پایین دست
Down
Stream
بین ژنی
Inter
Genic
اینترون
Intron
اگزون
Exon
5,570 5,526 41,700 98,072 2,609 1,711,652 1,706,156 5,550,430 8,750,139 518,511
(0.03%) (0.03%) (0.23%) (0.54%) (0.14%) (9.33%) (9.27%) (30.14%) (47.52%) (2.82%)
10. Non-synonymous
(1400 زمستان، 4، شماره 13مجله بیوتکنولوژی کشاورزی )دوره
100
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
سايی شده در های شناSNPدرصد 81/0در بررسی تنوع ساختاری در ژنوم سگ و نرگ بومی ايران نشان داده شد که از
ها در ناحيه خاموش قرار SNPدرصد 55/59درصد آنها تغييردهنده آمينواسيدها در پروتئين هستند و حدود 40ناحيه انزون، حدود
Amiri ghanatsamanهستند ) درصد آنها ايجادکننده کدون پايان 42/0دارند که تاثيری در تغيير آمينواسيدهای پروتئين ندارند و
et al. 2016 )
نذار و نوشتی شناسايی شد در دو لاين مرغ تخم SNP 13،449،339نويسی عملكردی، به منظور بررسی حاشيه در پژوهشی
واقع در SNPدرصد 2/1( قرار دارند و حدود 5/38( و بين ژنی )1/42ناحيه اينترونی )های تك نوکلئوتيدی در که اکرر چندشكلی
های واقع در SNP( که نسبت به نتايج بدست آمده در مطالعه حاضر، تعداد Boschiero et al. 2018ناحيه انزون قرار داشتند )
کهد دنشیم فيکدون تعر ینيگزيبه عنوان جا)خاموش( 11یمعنهای همناحيه ی انزون ژنوم کمتر بوده است. در نذشته، واريانت
ريتوانند تأثیم ها به اصطلاح خاموش،واريانت نيکه ا دهدیشواهد موجود نشان ماما دهندینم رييشده را تغ یرمزنذار نهيآمدياس
;Tsai et al. 2008) در نظر نرفته شود های خاموشبه عنوان واريانت ديداشته باشند و نبا نيو عملكرد پروتئ انيدر ب یتوجه لقاب
Edwards et al. 2012.)
های تک نوکلئوتیدی با دسته عملکردی در اکوتیپ خزک. تعداد اثرات چندشکلی4جدول
Table 4. Number of SNP effects by functional class in the Khazak ecotype
Silent خاموش Nonsense بی معنی Missense بد معنی
240,715 (72.77%) 15,424 (4.66%) 74,687 (22.58%)
لاين 24رغ از م 243های مختلف مرغ )يابی کل ژنوم جمعيت( با است اده از توالی2015a) .Gheyas et alدر پژوهشی
درصد 22/50نترونی و ها در نواحی ايSNP 35/43شناسايی کردند و نزارش دادند که SNPميليون 15نذار(، تعداد نوشتی و تخم
باشند که تا حدودی ئين میهای تك نوکلئوتيدی کد کننده پروتدرصد چندشكلی 2/2قط حدود آنها در نواحی بين ژنی قرار دارند و ف
ها و عمق های آناليز دادهروش با نتايج پژوهش ما مغايرت داشت. اين ممكن است به دليل عواملی مانند تعداد و نمونه مورد بررسی،
يابی باشد.توالی
Boschiero(2018)که با نتايج (5)جدول بدست آمد 43/2( Ti / Tvهای متقاطع )جهشهای انتقالی به نسبت جهش
et al. ( 2016و) Oh et al. ها را هنگام بررسی ژنوم مرغ نژادهای مختلف قابل مقايسه بود به طوری که آنها نسبت اين جهش
هرچه نسبت باشد، زيراها میSNPی کي يت مطلو شناسايی دهندهنسبت بالاتر بدست آمده در نتايج ما نشان .برآورد کردند 3/2
Ti / Tv ميزان خطای کمتر میدهنده بيشتر باشد نشان( باشدBainbridge et al. 2011; Guo et al. 2012 ،علاوه بر اين .)
11. Synonymous

1400، زاده مهدابی و همکارانالهه رستم
101
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
های ناحيه مورد هدف )کل ژنوم، انزوم، ژنتواند در ژنوم نسبت به یم یواقع ديجد هایواريانتمورد انتظار از Ti / Tvنسبت
، هام. به عنوان مرال، در مورد انزو(Rosenberg et al. 2003) مت اوت باشد آن ناحيه GCو CpG یموتواخاص(، نونه و
يك ليبه دل Ti / Tvنسبت شيمنجر به افزا یدر مناطق انزون CpG یدهاينوکلئوتی شده در د لهيمت نيتوزيحضور س شيافزا
مشاهده شده است که نيهمچن. (Aslam et al. 2012) شودیم نيميبه تشده لهيمت نيتوزيو انتقال سدی آميناسيون ساده
ی از يافته کمتر عهمطال نيمشاهده شده در ا Ti / Tv. نسبت باشدمیمهرنان یاز ب شتريدر پرندنان و پستانداران ب GC یموتوا
(2012)Aslam et al. (2/2باشد )میبالاتر از انسان ی(. اما کم45/2)باشد در مورد بوقلمون می (DePristo et al. 2011.)
شود.میداده حيبالاتر در ژنوم پرندنان توض GCم و درصد با اندازه کوچكتر ژنو ادينسبت بالاتر به احتمال ز نيا
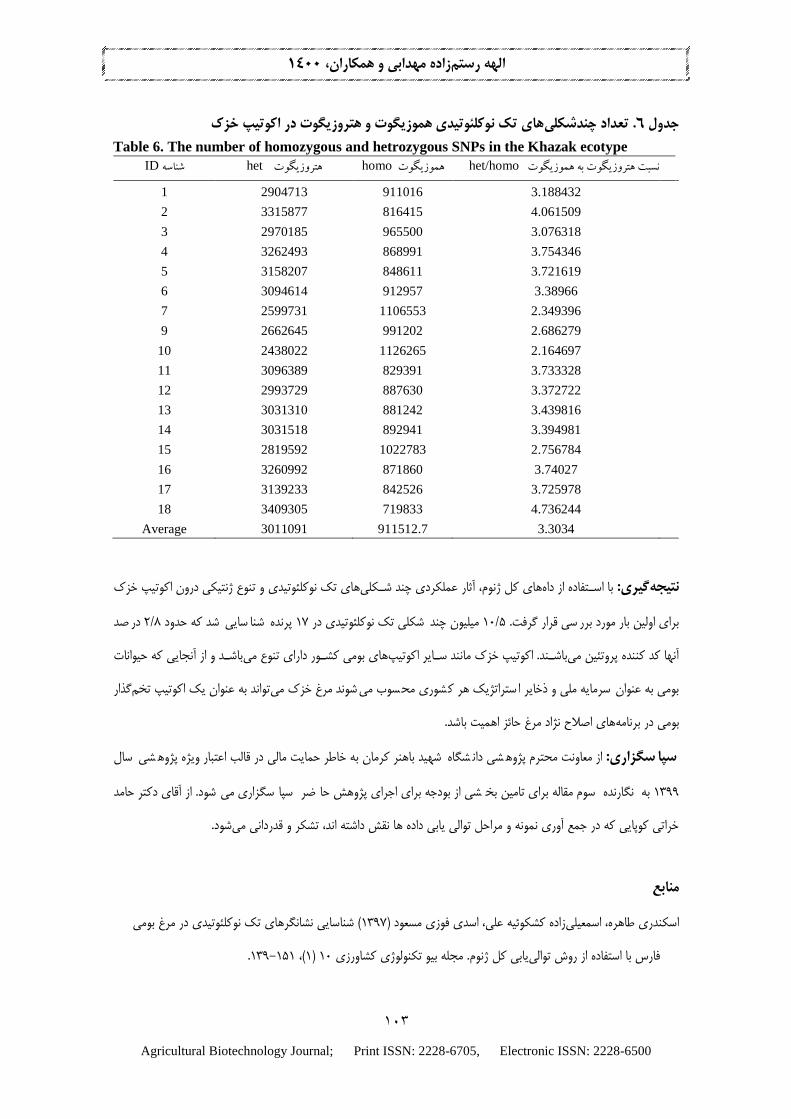
یبرا X6-10 یپوشش توال كيکه نشان داده شده است . باشدمی گوتيدشوارتر از هموز گوتيهتروز هایواريانت يیشناسا
از مطالعات نزارش داده اند که یحال، برخ ني. با اباشدمی ی( کافهتروزيگوتو گوتهموزي) مختلف هایواريانت از ٪99 صيتشخ
;Li et al. 2008; Sims et al. 2014باشد )می یضرور x 20بالاتر یپوشش گوتيهتروز یهاSNP ٪99 صيتشخ یبرا
Boschiero et al. 2018) . در اين مطالعه با است اده از پوشش توالیx6-7 های تك نوکلئوتيدی نشان داده شد که تعداد چندشكلی
و هتروزيگوت به ازای هر مرغ نشان داده شده های تك نوکلئوتيدی هموزيگوتتعداد چند شكلی 6هتروزيگوت بيشتر بود. در جدول
های مرندی و فارس های هتروزيگوت بيشتر از هموزيگوت بود که با مطالعات پيشين در اکوتيپSNPتعداد پرنده 17است. در همه
ب مربوط به هتروزيگوت به ترتي و کمترين واريانت نبيشتري .(Eskandari et al. 2019; Akbari et al. 2020مطابقت دارد )
و 18دهنده تنوع بيشتر در مرغ با شناسه اين امر احتمالا نشان ;باشد( می2،438،022) 10( و 3،409،305) 18های با شناسه مرغ
30/3خزک های هتروزيگوت به هموزيگوت در اکوتيپ SNPباشد. به طور ميانگين نسبت می 10همخونی بيشتر در مرغ با شناسه
برآورد شد.
0037/0 ± 0014/0ميزان تنوع ژنتيكی درون اکوتيپ خزک VCFtoolsدر بررسی تنوع ژنتيكی با است اده از نرم افزار
تنوع ژنتيكی در نژادهای مختلف مرغ را بررسی کردند و نشان دادند که بالاترين .Lawal et al(2018برآورد شد. در پژوهشی )
نيبالاتر لانكايسرمرغان و در نژادهای مرغ بومی، ( π= 0052/0 )باشد ميزان تنوع نوکلئوتيدی ژنومی درون نژاد مرغ جنگلی قرمز می
جارسو یوپيو ات (= π 0039/0ی )عربستان سعودمرغان باشند و تنوع ژنتيكی در ميان( را دارا می= 0046/0πی )ديتنوع نوکلئوت
(0036/0π = .تا حدودی با نتايج ما مطابقت داشت )
های از نوع انتقالی و متقاطع در ژنوم SNP. تعداد 5جدول
Table 5. SNP number of transition and transversion in genome
های انتقالیتعداد جهش
The number of transition mutations
های متقاطعتعداد جهش
The number of transversons
mutations
های انتقالی به متقاطعنسبت جهش
Transition/ transversion ratio
58,258,716 23,921,271 2.43

(1400 زمستان، 4، شماره 13مجله بیوتکنولوژی کشاورزی )دوره
102
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
1400، زاده مهدابی و همکارانالهه رستم
103
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
اکوتیپ خزک های تک نوکلئوتیدی هموزیگوت و هتروزیگوت درچندشکلی. تعداد 6جدول
Table 6. The number of homozygous and hetrozygous SNPs in the Khazak ecotype
het/homo نسبت هتروزیگوت به هموزیگوت homo هموزیگوت het هتروزیگوت ID شناسه
1 2904713 911016 3.188432
2 3315877 816415 4.061509
3 2970185 965500 3.076318
4 3262493 868991 3.754346
5 3158207 848611 3.721619
6 3094614 912957 3.38966
7 2599731 1106553 2.349396
9 2662645 991202 2.686279
10 2438022 1126265 2.164697
11 3096389 829391 3.733328
12 2993729 887630 3.372722
13 3031310 881242 3.439816
14 3031518 892941 3.394981
15 2819592 1022783 2.756784
16 3260992 871860 3.74027
17 3139233 842526 3.725978
18 3409305 719833 4.736244
Average 3011091 911512.7 3.3034
و تنوع ژنتيكی درون اکوتيپ خزک های تك نوکلئوتيدیآثار عملكردی چند شرركلیهای کل ژنوم، با اسررت اده از داه: گیرینتیجه
سی قرار نرفت. شكلی تك نوکلئوتيدی ميليون 5/10برای اولين بار مورد برر شد که حدود 17در چند سايی شنا صد 8/2پرنده در
باشررد و از آنجايی که حيوانات های بومی کشررور دارای تنوع میک مانند سرراير اکوتيپاکوتيپ خز باشررند.آنها کد کننده پروتئين می
ستراتژيك هر کشوری موسو می سرمايه ملی و خاير ا نذار تواند به عنوان يك اکوتيپ تخمشوند مرغ خزک میبومی به عنوان
های اصلاح نژاد مرغ حائز اهميت باشد. بومی در برنامه
سگ ش زاری:سپا ش یاز معاونت موترم پژوه سال یمال يتبه خاطر حماشهيد باهنر کرمان گاهدان شی در قالب اعتبار ويژه پژوه
شی از بودجه برای به 1399 سوم مقاله برای تامين بخ سگزار یاجرا نگارنده سنا ضر حامد دکتر یاز آقا .شودیم یپژوهش حا
.شودیم یتشكر و قدردانخراتی کوپايی که در جمع آوری نمونه و مراحل توالی يابی داده ها نقش داشته اند،
منابع
بومی غ مری در تك نوکلئوتيدی شناسايی نشانگرها( 1397زاده کشكوئيه علی، اسدی فوزی مسعود )اسكندری طاهره، اسمعيلی
. 139-151(، 1) 10. مجله بيو تكنولوژی کشاورزی منوژيابی کل لیاتوده از روش ست اابا رس فا

(1400 زمستان، 4، شماره 13مجله بیوتکنولوژی کشاورزی )دوره
104
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
( شناسايی تنوع ژنوم در 1399زاده کشكوئيه علی، اميری قنات سامان زينب، آيت اللهی مهجردی احمد )ول، اسمعيلیاکبری رس
. 176-161(، 1) 12يابی کل ژنوم. مجله بيو تكنولوژی کشاورزی مرغ مرندی با است اده از روش توالی
( بررسی تنوع ساختاری ژنگان سگ و نرگ 1395زاده کشكوئيه علی، اسدی فوزی مسعود )اميری قنات سامان زينب، اسمعيلی
. 277-271(، 2) 47يابی کل ژنوم. مجله علوم دامی ايران بومی ايران با روش توالی
201-169 .راني، ا رانيا یعلوم دام قاتيتوق بومی ايران. موسسه رطيودام و نتيكیژ خاير نگرشی بر (1378) اد،جو توکليان
.تعيين ص ات کي ی تخم مرغ در مرغ بومی خزک. پايان نامه کارشناسی ارشد، دانشكده کشاورزی، 1390دلجوی سرايان ج،
.42-28 .ابلزدانشگاه
های فنوتينی بر ارزش TGFB3( تاثير ژن 1392مومدی فر آمنه، فقيه ايمانی سيد علی، مومد آبادی مومد رضا، س لايی مومد )
. 136-125(، 4) 5و ارثی ص ات وزن بدن در مرغ بومی فارس. مجله بيوتكنولوژی کشاورزی
بررسی تنوع ژنتيكی مرغان بومی استان خراسان ( 1388نيكبختی مهدی، ميرزايی حميدرضا، افشاريان شانديز مجيد و همكاران )
.25-19(، 2)1های علوم دامی ايران پژوهش نشانگرهای ريزماهواره. رضوی با است اده
References
Abebe AS, Mikko S, Johansson AM (2015) Genetic diversity of five local Swedish chicken
breeds detected by microsatellite markers. PLoS One 4, 1-13.
Akbary R, Esmaeelizadeh A, Amiri Ghanatsaman Z, Ayetollahi Mehrjerdi A (2020)
Identification of genome diversity in marandi chicken using whole genome sequencing
method. J Agric Biotech 12,161-176 (In Persian).
Amiri Ghanatsaman Z, Esmailizadeh Koshkoiyeh A, Asadi Fozi M (2016) Study of structural
diversity of genome Iranian native dog and wolf with the method whole genome
sequencing. Iran J Anim Sci 47, 271-277 (In Persian).
Andrews S (2010) FastQC: a quality control tool for high throughput sequence data. Babraham
Bioinformatics, Babraham Institute, Cambridge, United Kingdom.
Aslam ML, Bastiaansen JW, Elferink MG et al. (2012) Whole genome SNP discovery and
analysis of genetic diversity in Turkey (Meleagris gallopavo). BMC genomics 13, 1-14.
Bainbridge MN, Wang M, Wu Y et al. (2011) Targeted enrichment beyond the consensus coding
DNA sequence exome reveals exons with higher variant densities. Genome Biol 12, 1-
12.
Boettcher P, Tixier‐Boichard M, Toro M et al. (2010) Objectives, criteria and methods for using
molecular genetic data in priority setting for conservation of animal genetic resources.
Anim Genetics 41, 64-77.

1400، زاده مهدابی و همکارانالهه رستم
105
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
Bolger AM, Lohse M, Usadel B (2014) Trimmomatic: a flexible trimmer for Illumina sequence
data. Bioinformatics 30, 2114-2120.
Boschiero C, Moreira GCM, Gheyas AA et al. (2018) Genome-wide characterization of genetic
variants and putative regions under selection in meat and egg-type chicken lines. BMC
Genomics 19, 83.
Cingolani P, Platts A, Wang LL et al. (2012) A program for annotating and predicting the effects
of single nucleotide polymorphisms, SnpEff :SNPs in the genome of Drosophila
melanogaster strain w1118; iso-2; iso-3. Fly 6, 80-92.
Danecek P, Auton A, Abecasis G et al. (2011) The variant call format and VCFtools.
Bioinformatics 27, 2156-2158.
Deljoy Sarayan J, (2011). Determination of Egg Qualitative Traits in Khazak Native Chicken.
Master thesis, Zabol University. pp. 28-42 (In Persion).
DePristo MA, Banks E, Poplin R et al. (2011) A framework for variation discovery and
genotyping using next-generation DNA sequencing data. Nature genetics 43, 491-498.
Diamond J (2002) Evolution, consequences and future of plant and animal domestication. Nature
418, 700-707.
Ebbert MT, Wadsworth ME, Staley LA et al. (2016) Evaluating the necessity of PCR duplicate
removal from next-generation sequencing data and a comparison of approaches. BMC
Bioinformatics 17, 239.Edwards NC, Hing ZA, Perry A et al. (2012) Characterization of
coding synonymous and non-synonymous variants in ADAMTS13 using ex vivo and in
silico approaches. PLoS One 7, e38864.
Engelsma KA (2012) Use of SNP markers to conserve genome-wide genetic diversity in
livestock. PhD thesis, Wageningen University. pp. 12-20.
Eskandari T, Esmailizadeh AK, Mohammadabadi MR, Sohrabi S (2018) Identification of single
nucleotide polymorphisms in Fars native chicken using whole genome sequencing data.
Agric Biotechnol J 10, 139-151 (In Persian).
Gheyas AA, Boschiero C, Eory L et al. (2015a) Functional classification of 15 million SNPs
detected from diverse chicken populations. DNA Res 22, 205-217.
Gheyas AA, Boschiero C, Eory L et al. (2015b) Functional classification of 15 million SNPs
detected from diverse chicken populations. DNA Res 22, 205-217.
Guo Y, Li J, Li C-I et al. (2012) The effect of strand bias in Illumina short-read sequencing data.
BMC genomics 13, 1-11.
Hoffmann I (2005) Research and investment in poultry genetic resources–challenges and options
for sustainable use. Worlds Poult Sci J 61, 57-70.

(1400 زمستان، 4، شماره 13مجله بیوتکنولوژی کشاورزی )دوره
106
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
Lawal RA, Al-Atiyat RM, Aljumaah RS et al. (2018) Whole-genome resequencing of red
junglefowl and indigenous village chicken reveal new insights on the genome dynamics
of the species. Front Genet 9, 264.
Li H, Durbin R (2009) Fast and accurate short read alignment with Burrows–Wheeler transform.
Bioinformatics 25, 1754-1760.
Li H, Handsaker B, Wysoker A et al. (2009) The sequence alignment/map format and SAMtools.
Bioinformatics 25, 2078-2079.
Li H, Ruan J, Durbin R (2008) Mapping short DNA sequencing reads and calling variants using
mapping quality scores. Genome Res 18, 1851-1858.
Linck E, Battey C (2019) Minor allele frequency thresholds strongly affect population structure
inference with genomic data sets. Mol Ecol Resour 19, 639-647.
Lynch M, Milligan BG (1994) Analysis of population genetic structure with RAPD markers. Mol
Ecol 3, 91-99.
McKenna A, Hanna M, Banks E et al. (2010) The Genome Analysis Toolkit: a MapReduce
framework for analyzing next-generation DNA sequencing data. Genome Res 20, 1297-
1303.
Moazeni S, Mohammadabadi MR, Sadeghi M et al. (2016a) Association between UCP Gene
Polymorphisms and Growth, Brreeding Value of Growth and Reproductive Traits in
Mazandaran Indigenous Chicken. Open J. Anim. Sci 6, 1-8.
Moazeni SM, Mohammadabadi MR, Sadeghi M et al. (2016b) Association of the melanocortin-
3(MC3R) receptor gene with growth and reproductive traits in Mazandaran indigenous
chicken. J. Livest. Sci. Technol 4, 51-56.
Mohammadifar A, Faghih Imani SA, Mohammadabadi MR, Soflaei M (2014) The effect of
TGFb3 gene on phenotypic and breeding values of body weight traits in Fars native fowls.
Agric. Biotechnol J 5, 125-136 (In Persian).
Mohammadifar A, Mohammadabadi MR (2018) Melanocortin-3 receptor (mc3r) gene association
with growth and egg production traits in Fars indigenous chicken. Malays Appl Biol 47,
85–90.
Mohammadifar A, Mohammadabadi MR )2017( The Effect of Uncoupling Protein
Polymorphisms on Growth, Breeding Value of Growth and Reproductive Traits in the
Fars Indigenous Chicken. Iran J Appl Anim Sci 7, 679-685.
Nikbakhti M, Mirzaee HR, Afsharian Shandiz M, et al. (2009) Analyses of genetic variation in
Khorasan indigenous chicken breed by using of microsatellite markers. Iran J Anim Sci
Res 1 (2), 19-25 (In Persian).

1400، زاده مهدابی و همکارانالهه رستم
107
Agricultural Biotechnology Journal; Print ISSN: 2228-6705, Electronic ISSN: 2228-6500
Oh D, Son B, Mun S et al. (2016) Whole genome re-sequencing of three domesticated chicken
breeds. Zool Sci 33, 73-77.
Osman SA-M, Yonezawa T, Nishibori M (2016) Origin and genetic diversity of Egyptian native
chickens based on complete sequence of mitochondrial DNA D-loop region. Poult Sci
95, 1248-1256.
Padhi MK (2016) Importance of indigenous breeds of chicken for rural economy and their
improvements for higher production performance. Scientifica 2016.
Peters J, Lebrasseur O, Best J et al. (2015) Questioning new answers regarding Holocene chicken
domestication in China. Proc Natl Acad Sci 112, E2415-E2415.
Rosenberg MS, Subramanian S, Kumar S (2003) Patterns of transitional mutation biases within
and among mammalian genomes. Mol Biol Evol 20, 988-993.
Sawai H, Kim HL, Kuno K et al. (2010) The origin and genetic variation of domestic chickens
with special reference to junglefowls Gallus g. gallus and G. varius. PLoS One 5, e10639.
Sims D, Sudbery I, Ilott NE et al. (2014) Sequencing depth and coverage: key considerations in
genomic analyses. Nat. Rev. Genet 15, 121.
Strillacci M, Cozzi M, Gorla E et al. (2017) Genomic and genetic variability of six chicken
populations using single nucleotide polymorphism and copy number variants as markers.
Anim 11, 737-745.
Tavakolian J ( 2000) An introduction to genetic resorces of native farm animals in Iran. Animal
Science Research Center of Iran, Iran. pp. 169-201 (In Persion).
Tixier-Boichard M, Bed’hom B, Rognon X (2011) Chicken domestication: from archeology to
genomics. C. R. Biol 334, 197-204.
Tsai C-J, Sauna ZE, Kimchi-Sarfaty C et al. (2008) Synonymous mutations and ribosome stalling
can lead to altered folding pathways and distinct minima. J. Mol. Biol 383, 281-291.
Venter JC, Smith HO, Adams MD (2015) The sequence of the human genome. Clin. Chem 61,
1207-1208.





























