Embed Size (px)
Citation preview

Eur. J. Biochem. 164,675-680 (1987) 0 FEBS 1987
The uncoupling protein from brown-adipose-tissue mitochondria Chymotrypsin-induced structural and functional modifications
Margarita FERNANDEZ, David G. NICHOLLS and Eduardo RIAL Department of Biochemistry, University of Dundee
(Received November 12, 1986) - EJB 86 1224
Mitoplasts prepared from brown adipose tissue mitochondria were treated with chymotrypsin and the fragments derived from the 32-kDa uncoupling protein identified by immunoblotting. Extensive proteolysis of the uncoupling protein occurred, the polypeptide pattern being affected by binding of the inhibitory nucleotide GDP. Chymotrypsin modifies the nucleotide binding site, lowering its affinity from 1.7 pM to 21 pM but without decreasing its binding capacity. Nucleotide bound to the modified site can still inhibit the permeation of H + and C1- through the protein. The ion conducting pathway itself is also sensitive to chymotrypsin, CI- and H + transport being partially inhibited in parallel. The ability of fatty acids to increase the Hf permeability of the protein is also inhibited in parallel with the basal H+ permeability. The results confirm that the transport of Hf and C1-, and the fatty acid regulation of H + permeation all share a common structural element within the 32- kDa protein.
The uncoupling protein from brown adipose tissue mitochondria catalyzes the regulated re-entry of protons ex- truded by the respiratory chain, thus dissipating the proton electrochemical gradient as heat (for review see [l , 21). While the uncoupling protein is structurally simple, consisting of a dimer of identical 32-kDa polypeptides (for review see [3]), complex relationships exist between the ligands which interact with it.
In addition to protons which are relevant physiologically (for a review see [l]), the uncoupling protein also transports anions such as Cl-, Br-, or NO; [4, 51. Competition studies demonstrate that protons and anions are transported by the same carrier [4, 61, although there are clear differences in the transport and regulatory mechanisms [7 - 111.
The ion transport can be regulated by two classes of ligands. Purine nucleoside di- or triphosphates bind to the carrier and inhibit the transport of both ions [5,12 - 141. Fatty acids increase proton permeation but leave anion transport unaffected [4, 71. The activating effect of fatty acids depends on the magnitude of the proton electrochemical potential: at low potentials (e.g. those generated during passive swelling) their actions are blocked by the inhibitory nucleotide, while under physiological conditions (high potential) fatty acids override the nucleotide inhibition [7]. Fatty acids have been proposed as the physiological regulators of the uncoupling protein in vivo [7, 15 - 171.
Recently, chemical modifications with tetranitromethane [I 1, 181, N-ethylmaleimide [18] or diazobenzenesulphonate [19] have been used as tools to investigate the relationship between nucleotide binding and the inhibition of chloride and proton transport. With these reagents it has been shown that the linkage between nucleotide binding and inhibition of ion transport can be broken, and that it is possible to affect
Correspondence to E. Rial, Department of Biochemistry, Univer- sity of Dundee, Dundee, Scotland DD1 4HN
transport of the two ions differentially. In the present paper we use proteolytic modification by chymotrypsin for the same ends. We show that the nucleotide binding and transport domains are both susceptible to chymotrypsin. Additionally, since protons and chloride show the same kinetics of inactiva- tion by the protease, the transport of the two species must share a common step.
MATERIALS AND METHODS Preparation of mitochondria and mitoplasts
Mitochondria were prepared, as previously described [20], from the brown adipose tissue of Syrian hamsters that had been cold-adapted (4°C) for at least two weeks. Mitoplasts were prepared by treating mitochondria with digitonin. Mitochondria ( 5 mg/ml) were incubated at 4"C, in a buffer containing 150 mM sucrose, 0.16 mg digitonin/mg mitochon- drial protein, 5 mM Tes (Na salt) pH 7.4. After 15 min the mitoplasts were centrifuged at 8500 x g for 10 min, resuspend- ed in 250 mM sucrose, 80 pM albumin, 5 mM Tes (Na salt) pH 7.4, and centrifuged again under the same conditions. The final pellet was resuspended in 250 mM sucrose, 5 mM Tes (Na salt), pH 7.4. Protein concentration was determined using the Biuret method, with albumin as standard.
Adenylate kinase assay Adenylate kinase was measured by the method of
Sottocasa et al. [21], following NADP reduction in a medium containing ADP, hexokinase and glucose-6-phosphate dehy- drogenase.
Purification of the uncoupling protein and preparation of antibodies
Uncoupling protein was purified exactly as described by Lin and Klingenberg [13]. Its purity was checked using SDS-

676
PAGE. Antibodies were raised in rabbits by the following method. I00 pg of purified uncoupling protein with Freund's complete adjuvant were injected intramuscularly in the back legs in two consecutive weeks. The following week 100 pg of uncoupling protein with Freund's incomplete adjuvant were injected subcutaneously at several sites in the back. A booster
described [4] from the rate of light-scattering decrease. For proton permeability the medium contained 100 mM potassium acetate, 16 pM albumin, 1 mM azide, 10 pM rote- none, 10 mM Tes (Na salt) pH 7.0. To determine chloride permeability potassium acetate was substituted by KCl. In both cases swelling was initiated by 0.5 pM valinomycin.
was given by injecting 100 pg of pure uncoupling protein intravenously (ear vein) and the following week the rabbit was bled. The blood was left to clot overnight at 4 T , was centrifuged at 2500 x g for 30 min, the pellet discarded and the supernatant spun at the same speed for 15 min. In order to adsorb trace contaminating antibodies to other mitochondrial proteins, 500-pl aliquots of antiserum were incubated at 18 "C for 2 h with 50 p1 of sonicated hamster liver mitochondria (40 - 50 mgiml). The incubations were centrifuged at 11 000 x g for 2 min, and the immune serum used without further purification.
Reagents
Nitrocellulose filters were from Bio-Rad Laboratories Ltd (Watford, UK). Sheep-anti-(rabbit IgG) conjugated with per- oxidase was from Serotec Ltd (Blackthorn, UK). Adjuvants were from Difco Laboratories (Detroit, USA). Isotopes were from Amersham International (Amersham, UK). All other chemicals were from Sigma Chemical Co. (Poole, Dorset, UK).
RESULTS
Structure of the mitoplasts Treatment of mitoplasts and purified uncoupling protein with ch ymo trypsin
Mitoplasts (20 - 25 mg protein/ml) were incubated at 30°C in a medium containing 250 mM sucrose, 5 mM Tes (Na salt) pH 7.4, together with defined concentrations of a-chymotrypsin (treated with N-a-p-tosyl-L-lysine chloro- methane). To terminate proteolysis, 10 mg soy bean trypsin inhibitor [22] were added per mg chymotrypsin.
Purified uncoupling protein (0.4 mg protein/ml) was in- cubated at 30°C in a medium containing 20 mM Na2S04, 0.16 mM EDTA, 20 mM Mops pH 7.4, in the presence of 10 pg chymotrypsin/mg uncoupling protein, and the reaction stopped by addition of a 10-fold weight excess of trypsin inhibitor. All samples of purified uncoupling protein and mitoplasts for SDS-PAGE and immunoblotting were imme- diately diluted 1 : 1 with 0.25 M Tris, 20 mM dithiothreitol, 20% (v/v) glycerol, 4% SDS, pH 6.8 and heated to 100°C for 5 min.
Polyacrylamide gel electrophoresis and immunoblotting
The discontinuous gel system of Laemmli [23] was used in a vertical slab gel assembly. The stacking gel contained 5% acrylamide pH 6.8, and the gradient gel 12 - 15% acrylamide, pH 8.8. Proteins were transferred to nitrocellulose overnight (16 h at 250 mA). The transfer buffer contained 25 mM Tris/ C1, 190 mM glycine, 20% (v/v) methanol, pH 8.3. After trans- fer, filters were stained reversibly with Ponceau dye and photographed. Filters were blocked by incubation for 2 min with 0.2% Tween 20. The immune serum was used after dilu- tion 1/160 in 10 mM Tris/Cl, 150 mM NaC1, 0.05% sodium azide, 3% albumin. Bound antibody was detected by sheep- anti-(rabbit IgG) conjugated with peroxidase, developed with 4-chloronaphthol and photographed with Kodak technical pan film 241 5.
GDP binding and determination of ion permeabilities
For GDP-binding experiments, control and treated mitoplasts were diluted to 14mg/ml. GDP binding was performed as previously described [20] in a medium contain- ing 100 mM potassium acetate, 10 mM Tes (Na salt), 1 mM azide, 10 pM rotenone. 13H]GDP concentrations ranged over 0.25 - 8 pM. GDP-binding parameters were calculated using the Rothamsted Maximum Likelihood Programme [24]. Pro- ton and chloride permeabilities were determined as previously
To allow free access of the protease to the outer face of the inner mitochondrial membrane, it is essential to permeabilize the outer membrane. Mitoplasts were prepared using low concentrations of digitonin in order not to damage the integrity of the inner mitochondrial membrane. A reduced osmolarity of the medium was found to improve the yield. The mitoplast preparation was routinely characterized by assaying adenylate kinase release [98% f 0.7 (n = lo), range 95- 1001. The mitoplasts retained large areas of ruptured outer membrane, and kept a condensed matrix (data not shown).
A major problem encountered when performing chemical or proteolytic modification of mitochondrial transport pro- teins in situ is unspecific permeabilization of the inner mem- brane [ll]. The mitoplasts remained largely impermeable to K + (in the absence of valinomycin) swelling in KC1 or potassium acetate at only 1.3% (control) or 9% (50 pg chymotrypsin/mg for 15 min) of the rate observed after the addition of 0.5 pM valinomycin.
Proteolytic fragmentation of the isolated uncoupling protein
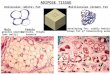
Fig. 1 shows a representative immunoblot of the isolated uncoupling protein after treatment with 10 pg chymotrypsin/ mg uncoupling protein. In the absence of GDP (lanes 5 - 8) there is a steady decrease in the intensity of the 32-kDa band with the appearance within 1 min of a characteristic fragmentation pattern. Bands in the range 27-14 kDa are most apparent at 1 min and then decline, while there is an increase in lower-molecular-mass fragments (10 - 11 kDa). In the presence of 500 pM GDP (lanes 2-4), the pattern of fragments at 1 min is identical, but in contrast to the above the further stages of proteolysis are strongly inhibited.
Proteolytic fragmentation of the in situ uncoupling protein
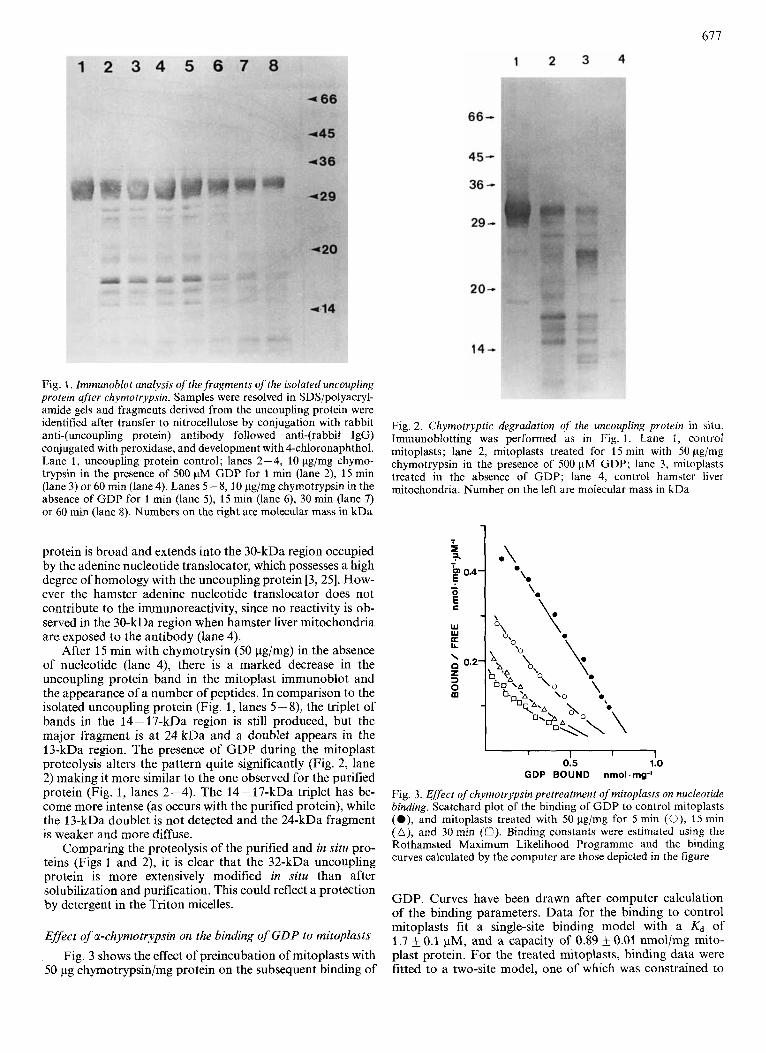
Fig. 2 shows the immunoblot pattern before and after proteolysis of the uncoupling protein in situ. Control mitoplasts (lane 1) show two faint bands in addition to the 32-kDa band corresponding to the uncoupling protein. The 18-kDa band is also seen in hamster liver mitochondria (lane 4) and so is not related to the uncoupling protein. The 25-kDa band corresponds to a similar band after proteolysis (lane 3) and may therefore be due to slight endogenous proteolysis. The major, 32-kDa, band corresponding to the uncoupling
677
1 2 3 4
66 - 45-
36 - 29 -.
20-
14-
Fig. 1 . Immunoblot analysis of the fragments of the isolated uncoupling protein after chymotrypsin. Samples were resolved in SDS/polyacryl- amide gels and fragments derived from the uncoupling protein were identified after transfer to nitrocellulose by conjugation with rabbit anti-(uncoupling protein) antibody followed anti-(rabbit IgG) conjugated with peroxidase, and development with 4-chloronaphthol. Lane 1, uncoupling protein control; lanes 2-4, 10 pg/mg chymo- trypsin in the presence of 500 pM GDP for 1 rnin (lane 2), 15 rnin (lane 3) or 60 min (lane 4). Lanes 5 - 8,lO kg/mg chymotrypsin in the absence of GDP for 1 min (lane 5 ) , 15 rnin (lane 6), 30 rnin (lane 7) or 60 min (lane 8). Numbers on the right are molecular mass in kDa
protein is broad and extends into the 30-kDa region occupied by the adenine nucleotide translocator, which possesses a high degree of homology with the uncoupling protein [3,25]. How- ever the hamster adenine nucleotide translocator does not contribute to the immunoreactivity, since no reactivity is ob- served in the 30-kDa region when hamster liver mitochondria are exposed to the antibody (lane 4).
After 15 min with chymotrysin (50 pg/mg) in the absence of nucleotide (lane 4), there is a marked decrease in the uncoupling protein band in the mitoplast immunoblot and the appearance of a number of peptides. In comparison to the isolated uncoupling protein (Fig. 1, lanes 5 - S), the triplet of bands in the 14-17-kDa region is still produced, but the major fragment is at 24 kDa and a doublet appears in the 13-kDa region. The presence of GDP during the mitoplast proteolysis alters the pattern quite significantly (Fig. 2, lane 2) making it more similar to the one observed for the purified protein (Fig. 1, lanes 2-4). The 14-17-kDa triplet has be- come more intense (as occurs with the purified protein), while the 13-kDa doublet is not detected and the 24-kDa fragment is weaker and more diffuse.
Comparing the proteolysis of the purified and in situ pro- teins (Figs 1 and 2), it is clear that the 32-kDa uncoupling protein is more extensively modified in situ than after solubilization and purification. This could reflect a protection by detergent in the Triton micelles.
Effect of a-chymotrypsin on the binding of GDP to mitoplasts Fig. 3 shows the effect of preincubation of mitoplasts with
50 pg chymotrypsin/mg protein on the subsequent binding of
Fig. 2. Chymotryptic degradation of the uncoupling protein in situ. Immunoblotting was performed as in Fig. 1 . Lane 1, control mitoplasts; lane 2, mitoplasts treated for 15 rnin with 50 pg/mg chymotrypsin in the presence of 500 pM GDP; lane 3, mitoplasts treated in the absence of GDP; lane 4, control hamster liver mitochondria. Number on the left are molecular mass in kDa
1
I I I I
GDP BOUND nmol.mg-' 0.5 1.0
Fig. 3. Effect of chymotrypsin pretreatment of mitoplasts on nucleotide binding. Scatchard plot of the binding of GDP to control mitoplasts ( O ) , and mitoplasts treated with 50 pg/mg for 5 rnin (0), 15 rnin (A), and 30min (13). Binding constants were estimated using the Rothamsted Maximum Likelihood Programme and the binding curves calculated by the computer are those depicted in the figure
GDP. Curves have been drawn after computer calculation of the binding parameters. Data for the binding to control mitoplasts fit a single-site binding model with a Kd of 1.7 +_ 0.1 pM, and a capacity of 0.89 0.01 nmol/mg mito- plast protein. For the treated mitoplasts, binding data were fitted to a two-site model, one of which was constrained to
678
501 A
2001
1: A
1 1 50 100
PROTON - F A
30 60 TIME min
PROTON - F A
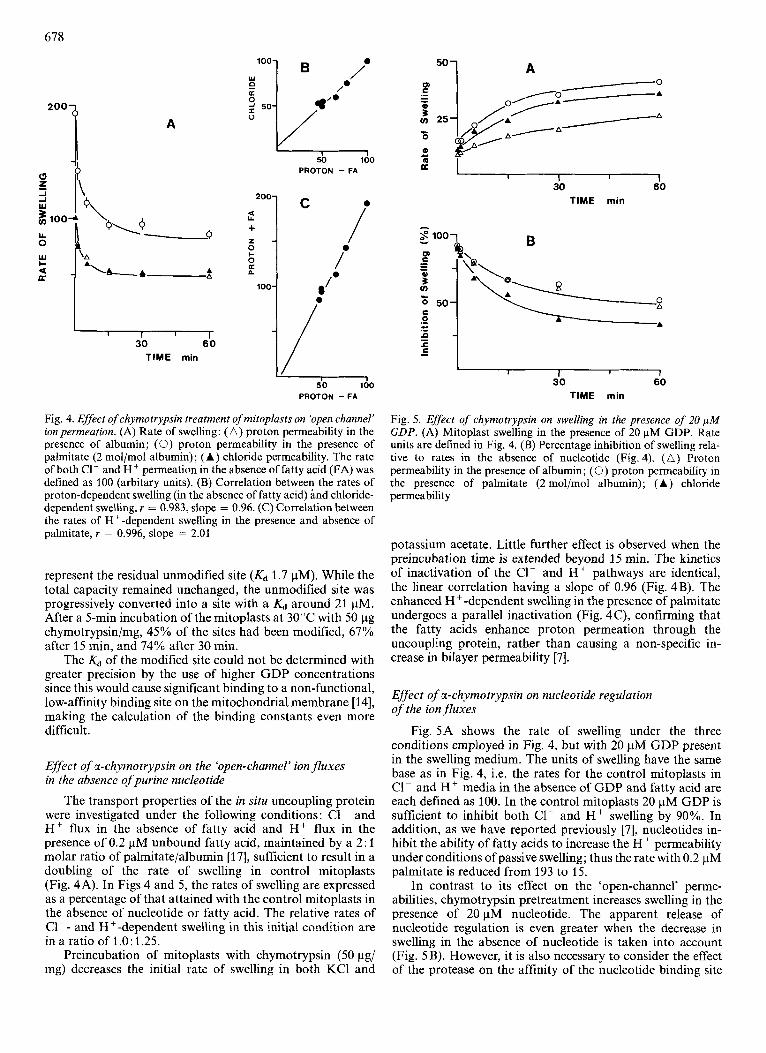
Fig. 4. Effect of chymotrypsin treatment of mitoplasts on 'open channel' ion permeation. (A) Rate of swelling: ( A ) proton permeability in the presence of albumin; (0) proton permeability in the presence of palmitate (2 rnol/mol albumin); (A) chloride permeability. The rate of both C1- and H + permeation in the absence of fatty acid (FA) was defined as 100 (arbitary units). (B) Correlation between the rates of proton-dependent swelling (in the absence of fatty acid) and chloride- dependent swelling, r = 0.983, slope = 0.96. (C) Correlation between the rates of H+-dependent swelling in the presence and absence of palmitate, r = 0.996, slope = 2.01
represent the residual unmodified site (Kd 1.7 pM). While the total capacity remained unchanged, the unmodified site was progressively converted into a site with a Kd around 21 pM. After a 5-min incubation of the mitoplasts at 30°C with 50 pg chymotrypsin/mg, 45% of the sites had been modified, 67% after 15 min, and 74% after 30 min.
The & of the modified site could not be determined with greater precision by the use of higher GDP concentrations since this would cause significant binding to a non-functional, low-affinity binding site on the mitochondria1 membrane [14], making the calculation of the binding constants even more difficult.
Effect of a-chymotrypsin on the 'open-channel' ion fluxes in the absence of purine nucleotide
The transport properties of the in situ uncoupling protein were investigated under the following conditions: C1- and Hf flux in the absence of fatty acid and Hf flux in the presence of 0.2 pM unbound fatty acid, maintained by a 2: 1 molar ratio of palmitate/albumin [17], sufficient to result in a doubling of the rate of swelling in control mitoplasts (Fig. 4A). In Figs 4 and 5, the rates of swelling are expressed as a percentage of that attained with the control mitoplasts in the absence of nucleotide or fatty acid. The relative rates of C1-- and H+-dependent swelling in this initial condition are in a ratio of 1.0: 1.25.
Preincubation of mitoplasts with chymotrypsin (50 pg/ mg) decreases the initial rate of swelling in both KCI and
I I 1 I I
30 60 TIME min
30 60 TIME min
Fig. 5. EfSect of chymotrypsin on swelling in the presence of 20 p M GDP. (A) Mitoplast swelling in the presence of 20 pM GDP. Rate units are defined in Fig. 4. (B) Percentage inhibition of swelling rela- tive to rates in the absence of nucleotide (Fig. 4). ( A ) Proton permeability in the presence of albumin; (0) proton permeability in the presence of palmitate (2 mol/mol albumin); (A) chloride permeability
potassium acetate. Little further effect is observed when the preincubation time is extended beyond 15 min. The kinetics of inactivation of the C1- and H+ pathways are identical, the linear correlation having a slope of 0.96 (Fig. 4B). The enhanced H+ -dependent swelling in the presence of palmitate undergoes a parallel inactivation (Fig. 4 C), confirming that the fatty acids enhance proton permeation through the uncoupling protein, rather than causing a non-specific in- crease in bilayer permeability [7].
Effect of a-chymotrypsin on nucleotide regulation of the ion fluxes
Fig. 5A shows the rate of swelling under the three conditions employed in Fig. 4, but with 20 pM GDP present in the swelling medium. The units of swelling have the same base as in Fig. 4, i.e. the rates for the control mitoplasts in C1- and Hf media in the absence of GDP and fatty acid are each defined as 100. In the control mitoplasts 20 pM GDP is sufficient to inhibit both C1- and H + swelling by 90%. In addition, as we have reported previously [7], nucleotides in- hibit the ability of fatty acids to increase the H+ permeability under conditions of passive swelling; thus the rate with 0.2 pM palmitate is reduced from 193 to 15.
In contrast to its effect on the 'open-channel' perme- abilities, chymotrypsin pretreatment increases swelling in the presence of 20 pM nucleotide. The apparent release of nucleotide regulation is even greater when the decrease in swelling in the absence of nucleotide is taken into account (Fig. 5B). However, it is also necessary to consider the effect of the protease on the affinity of the nucleotide binding site

679
50 100
?”’ SATURATION GDP BINDING
Fig. 6. Correlation between inhibition of swelling and saturation of nucleotide binding sites by 20 p M GDP during chymotrypsin treatment. Data from Figs 3 and 5. (A) Nucleotide binding vs inhibition of chloride permeation, r = 0.980, slope = 1.04. (B) Nucleotide binding vs inhibition of proton permeation (- fatty acid, FA), r = 0.996, slope = 1.02. (C) Nucleotide binding vs inhibition of proton permea- tion in the presence of palmitate, r = 0.996, slope = 1.03
(Fig. 3) . Using the binding data computed from Fig. 3, the saturation of the total (modified plus unmodified) binding sites by 20 pM GDP decreases in potassium acetate medium from 92% to 60% after 30 min of chymotrypsin exposure. This accounts rather precisely for the decrease in inhibition observed in this medium (Fig. 5 B). The chymotrypsin-modi- fied binding site is therefore still fully capable of regulating proton conductance. This is shown more clearly at higher GDP concentrations. 100 pM GDP saturates the control mitoplasts by 98% and causes a 95% inhibition of H + swelling. For the mitoplasts treated for 15 min with 50 pg/mg chymotrypsin, the expected saturation would be 88% and the inhibition observed is 81 %. The slight discrepancy between binding and inhibition in both cases can reasonably be accounted for by non-specific proton permeation other than through the uncoupling protein.
Fig. 5B shows that 20 pM GDP inhibits C1- swelling less than H+ swelling. However Lin and Klingenberg [13] have reported that C1- is a weak inhibitor of GDP binding to the uncoupling protein. Under our conditions, we have observed that the Kd for GDP binding to control mitoplasts increases from 1.7 f 0.1 pM in potassium acetate to 2.9 f 0.1 pM in KCl medium. This, together with a proportionate increase in the Kd of the modified site, accounts rather exactly for the observed rates without the need to postulate ‘decoupling’ in KC1 (Fig. 6A).
Therefore, the apparent decrease in the effectiveness of the nucleotide for the inhibition of ion permeation is not due
to the decoupling of nucleotide binding from the actual trans- port, as has been described in the case of the chemical modification of the uncoupling protein by tetranitromethane, N-ethylmaleimide, or diazobenzenesulfonate [ l l , 18, 191, but is a reflection of the lowered affinity of the binding site.
DISCUSSION Chymotrypsin readily attacks the uncoupling protein in
mitoplasts to (a) modify the nucleotide binding site, by lowering its affinity, and (b) to inactive the ‘open channel’ permeability of the protein to both C1- and H+. These modifications appear to be independent, since the decrease in ‘open channel’ conductance is largely complete within 15 min (Fig. 4A), whereas GDP inhibition continues to decrease over a 60-min period (Fig. 5 B). Chymotrypsin does not (a) destroy nucleotide binding sites, (b) enable C1- and H + permeation to be dissociated, (c) block the ability of fatty acids to activate those uncoupling proteins which can still conduct ions or (d) ‘decouple’ GDP binding from the regulation of ion permea- tion. Chymotrypsin modification therefore differs from chemical modification of the uncoupling protein by tetranitro- methane, N-ethylmaleimide, or diazobenzenesulfonate [l 1, 18, 191 where there is clear evidence that nucleotide can bind without stoichiometric inhibition of ion permeation.
It is not possible to correlate these functional changes with particular cleavages of the polypeptide since the techniques used do not allow a quantification of the fragments. However, since the fragments from the in situ protein differ in the pres- ence and absence of GDP (Fig. 2), GDP binding must either directly protect a potential cleavage point, or induce a conformational change affecting the accessibility of other susceptible peptide bonds.
Recently, French et al. [26] have presented the effect of treating intact mitochondria and isolated uncoupling protein with trypsin and chymotrypsin. In each case they identified only two fragments, of 21 -22 kDa and 10- 11 kDa, consis- tent with a single cleavage site for either protease. Although these authors used a fivefold higher protease concentration than in the present study, it is difficult to understand how the uncoupling protein could be modified in situ without the removal of the outer mitochondria1 membrane.
The parallel inactivation of the C1- and H+ permeabilities is of particular interest. A number of recent reports have emphasized differences that exist between the two: the lack of activation by fatty acids of CI- transport [4, 71, the failure to detect C1- transport by the reconstituted uncoupling protein [lo], and their differential sensitivity to nucleotide following chemical modification of the uncoupling protein [ l l , 191. These indications have led to the proposal that the two ions are transported by two independent pathways within the uncoupling protein [8, 9, 111, and even by two different pro- teins [9, 101. The present results are consistent with our early data demonstrating that the two ions compete for the same carrier [4, 61. Furthermore our results demonstrate once more that the ability of fatty acids to increase proton permeability in brown fat mitochondria is dependent upon the presence of functional uncoupling protein [17, 20, 271.
This work was supported by a grant from the Medical Research Council. M. F. was the recipient of a fellowship from FEBS and a grant from the Spanish Ministry of Education and Science. We are very grateful to Dr M. Kerr for his help and advice on the immunologi- cal techniques. The technical assistance of Mr Craig Adam, the help of Mr Gordon Milne with the electron microscopy, and Ms Alice Harrison with the photography are gratefully acknowledged.

REFERENCES 1. Nicholls, D. G. & Locke, R. M. (1984) Physiol. Rev. 64, 1-64. 2. Trayhurn, P. & Nicholls, D. G. (eds) (1986) Brown adipose tissue,
3. Klingenberg, M. (1985) Proc. N Y Acad. Sci. 456,279-288. 4. Nicholls, D. G. & Lindberg, 0. (1973) Eur. J. Biochem. 37, 523-
530. 5. Nicholls, D. G. & Heaton, G. (1978) in Proton and calciumpumps
(Azzone, G. F., Avron, M., Metcalfe, J. C., Quagliariello, E. & Silliprandi, N., eds) pp. 309- 318, Elsevier, Amsterdam.
6. Nicholls, D. G. (1974) Eur. J. Biochem. 49, 585-593. 7. Rial, E., Poustie, E. & Nicholls, D. G. (1983) Ew. J . Biochem.
8. Nicholls, D. G., Snelling, R. & Rial, E. (1984) Biochem. Soc.
9. Kopeck$, J., Guerrieri, F., Jeiek, P., Drahota, Z. & HouStEk, J.
10. Klingenberg, M. & Winkler, E. (1985) EMBO J. 4, 3087-3092. 11. Rial, E. & Nicholls, D. G. (1986) Eur. J. Biochem. 161,689-694. 12. Nicholls, D. G., Cannon, B., Grav, H. J. & Lindberg, 0. (1974)
in Dynamics of energy-transducing membranes (Ernster, L., Estabrook, R. & Slater, E. C., eds) pp. 529-537, Elsevier, Amsterdam.
13. Lin, C.4. & Klingenberg, M. (1982) Biochemistry 21, 2950- 2956.
Edward Arnold, London.
137,197-203.
Trans. 12,388 - 390.
(1984) FEBSLett. 170, 186-190.
14. Rial, E. & Nicholls, D. G. (1983) FEBS Lett. 161, 284-288. 15. Locke, R. M., Rial, E., Scott, I. D. & Nicholls, D. G. (1982) Eur.
16. Strieleman, P. J., Schalinske, K. L. & Shrago, E. (1985) J . Bid.
17. Cunningham, S . A,, Wiesinger, H. & Nicholls, D. G. (1986) Eur.
18. Rial, E. & Nicholls, D. G. (1986) FEBS Lett. 198, 29-32. 19. Kopecky, J., Jeiek, P., Drahota, Z. & HouStEk, J. (1986) EBEC
20. Rial, E. & Nicholls, D. G. (1984) Biochem. J . 222, 685-693. 21. Sottocasa, G. L., Kuylenstierna, B., Ernster, L. & Bergstrand, A.
22. Ozawa, K. & Laskowski, M. Jr (1966) J. Biol. Chem. 241,3955-
23. Laemmli, U. K. (1970) Nature (Lond.) 227, 680-685. 24. Ross, G. J. S. (1978) in Numerical software - Needs and
availability (Jacobs, D. A. H., ed.) pp. 203-323, Academic Press, New York.
25. Aquila, H., Link, T. A. & Klingenberg, M. (1985) EMBO J . 4,
26. French, R. R., Peachey, T. J. & York, D. A. (1986) EBEC Rep.
27. Locke, R. M., Rial, E. & Nicholls, D. G. (1982) Eur. J . Biochem.
J . Biochem. 129, 373 - 380.
Chem. 260,13402-13405.
J. Biochem. 157,415-420.
Rep. 4, 73.
(1967) Methods Enzymol. 10, 448-460.
3961.
2369 -2376.
4, 383.
129, 381 -387.