Embed Size (px)
Citation preview

Cellular and Molecular Neurobiology, Vol. 3, No. 2, I983
Review and Commentary
Theory of Electromechanical Effects in Nerve
David Gross, 1-3 Wendell S. Williams, 1'4 and John A. Connor 5'6
Received October 6, 1982; revised April 1, 1983; accepted April 14, 1983
KEY WORDS: mechanoreception; bioelectromechanics; membrane surface charge; theoretical model; electrostriction; piezoelectricity.
SUMMARY
1. The electromechanical transduction mechanisms operating in nerve mem- branes are considered theoretically.
2. For mechanical-to-electrical transduction (mechanical generator potentials), a model is proposed in which the surface charge on the membrane mediates stress-induced changes in the intramembrane electric field, thus opening transmem- brane ion conductance channels or reducing the ion selectivity of the membrane via leak conductance pathways.
3. For electrical-to-mechanical transduction (axon diameter change with excita- tion), an investigation into two well-known electrostatic properties of dielectrics, electrostriction and piezoelectricity, in the context of the nerve membrane is under- taken which predicts a few percent change in axon dimensions for voltage- and space-clamped axons.
INTRODUCTION
This paper is concerned with the transduction of mechanical to electrical energy, and vice versa, in nerve tissue. Such phenomena are well-known from the literature but are
Department of Physics, Bioengineering Program, University of Illinois, Urbana, Illinois 61801. 2 Present address: School of Applied and Engineering Physics, Clark Hall, Cornell University, Ithaca, New
York 14853. 3 To whom correspondence should be sent. a Also with the Department of Ceramic Engineering, University of Illinois, Urbana, Illinois 61801. 5 Department of Physiology and Biophysics, University of Illinois, Urbana, Illinois 61801. 6 Present address: Bell Telephone Labs, Department of Biophysics, Murray Hill, New Jersey 07974.
89
0272-4340/83/0600-0089503.00/0 © i983 Plenum Publishing Corporation

90 Gross, Williams, and Connor
poorly understood theoretically in terms of explicit physical models. Two such models are presented here, one describing the mechanical-to-electrical transduction phenome- non and the other describing the converse phenomenon.
Mechanica l - to -Elec t r ica l Transduct ion. As is obvious to any living thing that can see, hear, taste, feel, or smell, external stimuli can and do excite certain cells. Each of these phenomena is mediated by the transfer of energy (electromagnetic, chemical, or mechanical) from the external environment to a specialized receptor nerve ending which is capable of transducing one or more of these stimuli into electrical information which then can be transmitted along the nervous system to be processed in the brain. There are several specialized structures, termed meehanoreceptors, which perform this transduction for mechanical stimuli, e.g., the vertebrate Pacinian corpuscle, the crustacean stretch receptor, the cochlea, the lateral line organ in fish, insect leg hairs, cerci hairs, muscle spindles, joint angle detectors, and so forth. In addition to the specialized receptors, it is known that simple nonmyelinated axons mimic mechanore- eeptors when they are mechanically stimulated (Goldman, 1965; Julian and Goldman, 1962). Thus it appears that the eleetromechanical transduction process is not specific for specialized nerve structures but is in fact a general property of nonmyelinated axons .
It has been argued convincingly by Goldman that the eleetromechanical trans- duction process occurs when the axon membrane is stretched (Goldman, 1965). He notes that many specialized receptors either are directly stretched by the stimulus or are stretched by some mechanical modifying apparatus upon which the stimlus acts.
When a receptor or an axon is strained, the transmembrane potential decreases toward zero, that is, the cell depolarizes. This depolarization is graded, that is, it is proportional to the amount of strain applied to the system. The depolarization, if it is of sufficient size, can generate one or a series of action potentials in the part of the nerve cell immediately attached to the region which is depolarized. The effective resistance of the depolarized membrane decreases upon stretch, while the specific capacitance remains constant (Julian and Goldman, 1962). The change in membrane potential associated with the strain (and, in fact, any such potential change in any sensory receptor) is called a generator potential.
In such specialized structures as stretch receptors, depolarization is approxi- mately linearly related to strain (Obara, 1968). Several authors have investigated the effect of altered ionic composition of bathing solutions with quite interesting results. If sodium ion is removed or replaced by an impermeable organic cation (choline, for example), the generator potential is greatly reduced in the crustacean stretch receptor and the Pacinian corpuscle (Edwards et al., 1963; Diamond et a l., 1958). If procaine, a drug which decreases both sodium and potassium conductance, is added to the bathing solution, the generator potential is much reduced in the crustacean giant axon and the Pacinian corpuscle (Julian and Goldman, 1962; Diamond et al., 1958). The addition of TTX, which blocks Na + conduction for the normal sodium channel, has no effect on the generator potential in the crustacean stretch receptor (Obara, 1968; Loewenstein et al., 1963). Likewise, no change is seen for the addition of five times normal external calcium (Obara, 1968; Edwards et al., 1963), but the removal of Ca z+ reduces the generator potential in the stretch receptor (Edwards et al., 1963).
From the above results it appears that upon stretch the axon membrane becomes more permeable to Na + ion, which, according to the Goldman equation, causes

Theory of Nerve Electromechanics 91
depolarization. That the Na + permeability increase is different from that in the action potential is shown by the TTX data. The action of procaine on the generator potential, however, is similar to its action on the action potential.
Most authors agree that the permeability change of the transducing membrane to Na + ions is the transducing process. Goldman mentions several possible mediating mechanisms for this effect, among them piezoelectricity and streaming potential, but dismisses them as unlikely and opts for a direct mechanical modification of membrane permeability (Goldman, 1965).
Elec tr ica l - to-Mechanical Transduction. Several investigators have reported that swelling of axons results from electrical activity. Hill (1950) found an average increase in radius of 0.11 #m/104 stimuli for a 220 urn-diameter cuttlefish axon resulting from stimulation at 200/sec. Cohen and Keynes (1971) found a light- scattering change in excited crab nerve which they attributed to an increased axon volume. Iwasa et al. (1980) and Iwasa and Tasaki (1980) reported that single action potentials in squid and crab nerves produced a swelling of the fiber concomitant with the action potential. However, Hill et al. (1977) found a diameter decrease concomi- tant with the action potential in crayfish giant axon.
In addition to diameter changes in active axons, it has been found that axons shorten during activity. Both Bryant and Tobias (1955) and Stepanov (1968) demonstrated a contraction in whole nerve associated with action potential propaga- tion. Tasaki and Iwasa (1980) have shown that conduction of an action potential along a crab nerve is accompanied by a shortening of the nerve of 5-10 nm. They further found that an action potential causes a crab nerve to develop a force of -0.02 dyne when the nerve is at a constant length. Thus it appears that the mechanical response to electrical activity in axons is another general property of these systems.
It has been suggested that the mechanism behind these processes is osmotic in origin (Hill, 1950). Hill suggested that the exchange of Na + and K + during the action potential would produce swelling as observed since a solution of NaC1 has a slightly greater osmotic pressure than a solution of KC1 of the same strength. Cohen et al.
(1972) presented arguments that the influx of C1 might be a factor in the swelling of active axons. In addition to these osmotic swelling arguments, Iwasa et al. (1980) thought that the phenomenon was related to the swelling of polyelectrolyte gels and of charged membranes.
In the present paper, we propose and discuss mechanisms for both senses of electromechanical transduction of membranes from the point of view of electrostatics.
MECHANICAL-TO-ELECTRICAL TRANSDUCTION MODEL
Most biological membranes have a negative surface charge (McLaughlin, 1977; McLaughlin and Poo, 1981), although certain transformed cells appear to have a positive surface charge (Elul, 1967). Experiments on squid axon have indicated external surface charge values of - e / 1 2 0 to - e /325 coulomb/]~ 2 and internal surface charge values of - e / 7 3 0 to - e /1600 coulomb//~ 2 (Rojas and Atwater, 1968; Gilbert and Ehrenstein, 1969; McLaughlin, 1977). The presence of such a charged membrane- solution interface should produce a potential difference and an ionic concentration gradient from surface to bulk solution.

92 Gross, Williams, and Connor
As a first approximation, it is reasonable to treat biological membranes by Gouy-Chapman electrochemistry theory (c.f., McLaughlin, 1977). In this theory, one approximates the electric charge density in solution adjacent to a charged surface by
p(x) = F ~_. zsCi(x) = F ~ ziCi,= e -:`F'~(x)IR~ (1) i i
where x is the Cartesian coordinate perpendicular to the surface of the membrane, o is the charge density in the aqueous phase adjacent to the membrane, Ci.= is the concentration of ion species i in the bulk aqueous phase, and ¢ is the electrostatic potential at a distance x from the membrane surface. As seen in Eq. (1), the ions are assumed to obey Boltzmann statistics, and the energy of a given ion is assumed to be due only to its interaction with the electric field due to membrane plus attracted ions.
Inserting the charge density given by Eq. (1) into the Poisson equation of electrostatics leads to a differential equation for the surface electric potential ¢:
d25 dx 2 - -O(x) /~
(2)
E i
where e is the dielectric constant of the aqueous phase. To solve this equation, one makes the assumptions that e is not a function of position and that the electrolyte is symmetrical (i.e., I zil is equal for all i's). The latter assumption is not necessary for a closed analytic solution (Abraham-Shrauner, 1975), but it simplifies the mathematics. The former assumption is necessary in that the behavior of the dielectric constant of aqueous solutions is not completely understood, particularly in the region of charged interfaces (Bockris and Reddy, 1970). With these two assumptions, the solution of Eq. (2) becomes
¢ -~ ¢oe -"x (3)
if z¢0 is small compared to 2 R T / F ~ 50 mV. The constant K -~ is the Debye length, which is a familiar concept from the Debye-Htickel theory of dilute electrolytes. It is important to note that the surface potential ¢0 is dependent on the surface charge density, temperature, and solution composition.
Consider the effect of a charged membrane on the axon resting potential. First, at each charged surface a surface potential develops which is negative with respect to the bulk solution if the membrane faces are negatively charged. Second, the surface concentrations of anions are decreased from their bulk concentrations and the surface cation concentrations are enhanced by the Boltzmann factor [see Eq. (1)]. This alteration of ionic concentrations at the faces of the membrane can have a profound effect on the electric field in the membrane.
Consider the Goldman equation (Goldman, 1943):
~ _ PKCKx + PNaCNa,e + PclCcl,~ V m = In PKCK,a + PNaCNa,a + PclCcl,e . (4)
Since the concentrations appearing in Eq. (4) are concentrations at the external and

Theory of Nerve Electromechanics 93
axoplasmic faces of the membrane, they will reflect the above-mentioned concentra- tion alteration effect of the surface potential ~Po.
Surface potentials affect the potential difference from bulk extracellular fluid to bulk axoplasm in a second way. As shown in Fig. 1, the surface potentials themselves shift the bulk-bulk potential. The top illustration shows the membrane potential profile of an uncharged axon membrane as described by Eq. (4), that is, a Goldman membrane. The assumed ionic concentrations are those given by Hodgkin (1951) for squid giant axon, and the permeabilities are assumed to be PK:PN,:PcI::3.0:O.04:0.05. If, now, membrane surface charge is introduced on the inner and outer faces of the membrane, inner and outer surface potentials will develop between each of the
V : -70mV
Vm =-70 mV -- --T -- --
.,i
--"A . . . . 0
V = -80mY
VB =-70 mV
~ - i _ ~ ~ e . . . . . . 0
!AI &l
V = - 7 0 mV
Vm = -60mY
. . . . . . o
V
i/Ym !
distance Fig. 1. The effect of surface potentials on membrane poten- tial Vm and bulk-to-bulk potential V. (Top) Potential profile if no surface charge is present. V m = V. (Middle) Potential profile in the presence of surface charge, ignoring surface ion concentration changes. Vm is unchanged but Vchanges. (Bot- tom) Potential profile if surface ion concentration changes are included. Note that Vin the lower drawing is equal to Vm for the upper drawing, but Vm has decreased from top to bottom traces. See text for details.

94 Gross, Williams, and Connor
membrane faces and the bulk solutions, along with interfacial ion concentration profiles. The middle illustration in Fig. 1 shows the effect of these surface potentials alone without the ion concentration effect included. Note that the potential difference falling across the membrane itself is different from the bulk-to-bulk potential difference. Also note that the membrane potential difference (or, equivalently, the membrane ff~ field) is identical in these two cases. The lower illustration in Fig. 1 shows the complete effect of the surface charges, including concentration perturbation effects. Note that now the potential drop across the membrane proper is decreased compared to the case for the uncharged membrane surface in the top illustration.
From Fig. 1 it can be seen that the bulk-to-bulk potential V = ~Pe - ffa + Vm, where the lp's are extracellular and axoplasmic surface potentials and Vm is the potential dropping across the membrane proper. Quantitatively, this is, from Eq. (4) and the Boltzmann surface concentration enhancement factor,
RT In PKC~'ee-F~dRT + PNaCNa'ee-F+°/RT + PcICcl'aeU~"/~r (5) V m = ~ pKCK,ae -F~°/RT -~- pyaCNa,a e-F~k"/RT + PclCcl,ee F~°/RT"
Since CK.a ~ Ccl.e and PK >> Pc1, and ~c and ~b~ are both negative (since a¢ and a~ are negative), we may ignore the C1 terms in Eq. (5). This allows the great simplification to
Vm = ~, - ~b¢ + V~ lk , (6)
where Vb~ ~k is the membrane potential calculated in the Goldman Eq. (4), employing bulk solution values of ion concentrations. As can be seen from Eq. (6), when ~b~ is more negative than ~p,, as is apparently the case in squid axon, the electric field which is actually present in the membrane proper is less than that calculated using the Goldman equation with bulk solution concentrations.
Although Vm is altered by surface potentials, the measured bulk-to-bulk "mem- brane potential" V is not. This is easily seen from Eq. (6) and Fig. 1. Since the bulk-to-bulk potential difference is V = ~ - ~b~ + V~, it is readily apparent that the measured bulk-to-bulk potential difference V is equal to the potential difference calculated via the Goldman equation using bulk concentration values, V bu~k. Thus, the membrane plus ion double layers behave as a single membrane unit, as one might expect. One should note that this effect is valid if the ions near the surface of the membrane obey Boltzmann statistics and if the assumptions of the Goldman equation are valid. It is independent of the details of the electrochemistry near the surfaces of the membrane.
As mentioned in the introduction, electrical changes in axons undergoing mechanical stress have been seen experimentally. Such changes, according to Julian and Goldman (1962), appear to be caused primarily by a change in the permeability of the axon membrane to Na + ion. This was demonstrated by the fact that removal of Na + greatly reduced the electrical changes found in mechanically stimulated axons and mechanoreceptors. Also, partial control of membrane voltage during mechanical stimulation of lobster axon indicated that membrane conductance increases almost as much as it would in the absence of any voltage control. This observation led Goldman to conclude that the depolarization was chiefly an effect of the resistance decrease rather than a cause (Goldman, 1965). He further ruled out piezoelectricity and

Theory of Nerve Electromechanics 95
streaming potentials as possible transducing mechanisms since they would primarily effect a change in transmembrane voltage rather than conductance. However, as pointed out by Gross and Williams (1978), such mechanisms cannot so easily be discarded.
As an example, consider Eq. (6), which relates the potential drop across the membrane proper to the inner and outer surface potentials and the Goldman voltage calculated using bulk solution values of ion concentrations. The equation predicts that the electric field in the membrane proper, controlled by Vm, can change even if vb~ ~k does not (that is, even if any of the Pi's or ion concentrations do not change). It is conceivable that the increased ion permeability of the mechanically stimulated axon is not so different from that in an electrically stimulated axon if a mechanism which modifies surface potentials as a function of stretch is acting.
One such possible mechanism is a reduction in the surface charge density on the two surfaces of the axon due to increased surface area during stretch of the cell. As an illustration, consider Fig. 2. Suppose that the potential profile in the vicinity of the membrane before stretch is that shown in the upper illustration. If, now, the axon is stretched such that the inner and outer surface charge densities decrease, the potential V m falling across the membrane proper decreases. If the Vm-dependence of the ion permeabilities were turned off, the potential distribution near the membrane would be that shown in the middle illustration in Fig. 2. In this case, the bulk-bulk potential V remains unchanged from its prestretch value. However, in a real axon it is known that the Pi's are dependent on Vm such that they increase with depolarization. Thus, one would expect the bulk-bulk potential Vto decrease due to the V~-induced increase in membrane ion conductance, as shown in the bottom illustration in Fig. 2.
Note that surface electrochemistry theory demonstrates that surface potential ~p and surface charge density a are related (Grahame, 1947; McLaughlin, 1977). A variation in the surface charge density Ao- produces a variation in the surface potential AI~ = (01~/00")~O" -~ 0[(Ao')2]. The derivative 0~/&r is always positive, so a decrease in the magnitude of cr decreases the magnitude of ~b. If a cylindrical membrane is stretched by an amount Al, the charge density, assumed fixed to the membrane, decreases by an amount Aa ~ - a ( A l / l ) / 2 , where o- is the surface charge density before stretch. Such a decrease would decrease both @a and ~b~ and, thus, would affect Vm according to Eq. (6).
Consider first the effect of the decreased surface potentials alone--i.e., hold fixed the ion permeabilities PK and PNa such that V~ Ik is unchanged. In such a case the change in V m would be
Al AVm = A(~ba - ~be) -~ (~0 _ ~p0) Aao_ ~- (~o _ ~b0) 2-1' (7)
wherc the surface potentials in the right-hand portions of thc equation are of prestretch magnitude. Note that, as mentioncd earlier, the bulk-bulk potential V (which equals V~ Ik) is unaffected by the above AVm.
However, thc change in V, in Eq. (7) would couple to a change in membrane conductance by the normal conductance-voltage relationship known for cxcitable ceils. Thus, Vbm ulk would be affected as, then, the bulk-bulk potential V would. The quantitative predictions of this model are shown in Fig. 3. In this set of plots, the

96 Gross, Williams, and Connor
V : -70 mV
Vm=-eOmV
0 - = - _ - -- 0
/ Before Stretch
" - 0
V : - 70mY
Vm:-75 mV ,/ After Stretch
Pi's Fixed
V -" - 50mY
v =-ssmv / After Stretch
Pi's V m- Dependent
distand Fig. 2. The effect of axon stretch on surface potentials ~b, and ~ko, membrane potential Vm, and bulk-to-bulk potential V. (Top) Potential profiles for a hypothetical unstretcbed axon. (Middle) Potential profiles after stretch if ion permeabilities remain at their initial values. (Bottom) Potential profiles if permeabilities are allowed to adjust to the new Vm.
bulk-to-bulk potential change from rest, AV = V - Vrost, is plotted versus the longitudinal strain, AL/L, which is directly related to the depolarizing effect of reduced surface charge density as described by Eq. (7). The three curves correspond to three different values of (~po _ ~po), namely, 100 mV (solid line), 40 mV (dashed line), and 10 mV (dotted line).
The curves were generated by iteratively computing the total membrane potential Vm due to both the effects of reduced surface charge density and the resultant change in membrane potential V due to the altered permeabilities PK and Pya" The Vm- dependence of the permeabilities was taken from standard Hodgkin-Huxley theory (in the steady-state limit); a description of this theory as well as plots of the steady-state conductance parameters n, m, and h are given by Cole (1968). The bulk ion concentrations used in Eq. (4) were those for squid given by Hodgkin (1951) as quoted by Junge (1976). The permeability of the membrane to C1 was treated as negligible.
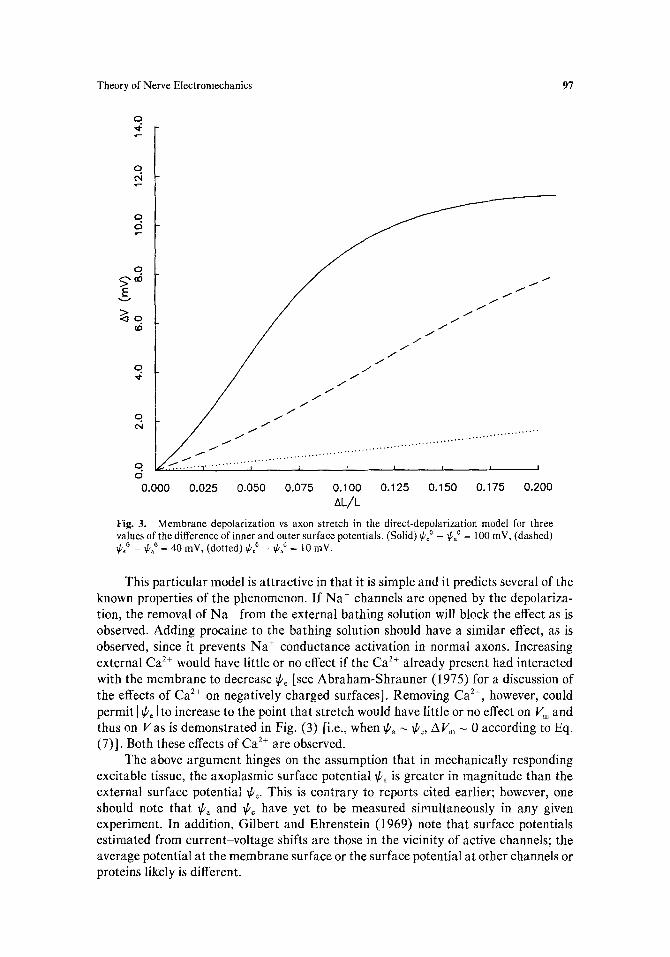
Theory of Nerve Electromechanics 97
O 4 ,r-
q ¢N
O le-
q ~ , o o /
t / /
o.,¢ / j / / / / ' w /
O / / ~ " / ~ ' f / /
. . . . . . . . . . . , . .......... , . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . , . ............ , ............ : ....... ,
O 0.000 0.025 0.050 0.075 0.100 0.125 0.150 0.175 0.200
AL/L Fig. 3. Membrane depolarization vs axon stretch in the direct-depolarization model for three values of the difference of inner and outer surface potentials. (Solid) ~p0 _ ~po0 = 100 mV, (dashed) ~boo _ ~o = 40 mY, (dotted) ~0 _ ~ao = 10 inV.
This particular model is attractive in that it is simple and it predicts several of the known properties of the phenomenon. If Na ÷ channels are opened by the depolariza- tion, the removal of Na ÷ from the external bathing solution will block the effect as is observed. Adding procaine to the bathing solution should have a similar effect, as is observed, since it prevents Na ÷ conductance activation in normal axons. Increasing external Ca 2+ would have little or no effect if the Ca 2+ already present had interacted with the membrane to decrease ~be [see Abraham-Shrauner (1975) for a discussion of the effects of Ca 2+ on negatively charged surfaces]. Removing Ca 2+, however, could permit I ~b~ I to increase to the point that stretch would have little or no effect on V~ and thus on Vas is demonstrated in Fig. (3) [i.e., when ~a ~ ff~, AVm ~ 0 according to Eq. (7)]. Both these effects of Ca 2+ are observed.
The above argument hinges on the assumption that in mechanically responding excitable tissue, the axoplasmic surface potential ~b a is greater in magnitude than the external surface potential ~b e. This is contrary to reports cited earlier; however, one should note that ~ba and ~bc have yet to be measured simultaneously in any given experiment. In addition, Gilbert and Ehrenstein (1969) note that surface potentials estimated from current-voltage shifts are those in the vicinity of active channels; the average potential at the membrane surface or the surface potential at other channels or proteins likely is different.

98 Gross, Williams, and Connor
One further difficulty with this model is the effect of TTX on the generator potential. Since TTX does not block the mechanically induced conductance increase, the normal voltage-controlled Na + channels involved in propagation of the action potential cannot be involved in this process. Thus one must postulate a different, voltage-sensitive, Na+-selective, TTX-insensitive channel. At present no evidence for such a channel exists.
A recent paper by Ganot et al. suggests a second possible mechanism by which stretch-altered surface charge density might transduce axon stretch to depolarization. A correlation between the reversal potentials of stretch-induced current and leakage current in Myxicola was reported; the implication is that the ionic pathways for these two phenomena might be related (Ganot et aI., 1981). Since K + permeability dominates the membrane potential of a resting excitable cell, if stretch could reduce that dominance, i.e., decrease the average ion selectivity of the cell membrane, then one would expect stretch to produce depolarization of the cell. Just such an effect can be produced by stretch-altered surface charge density.
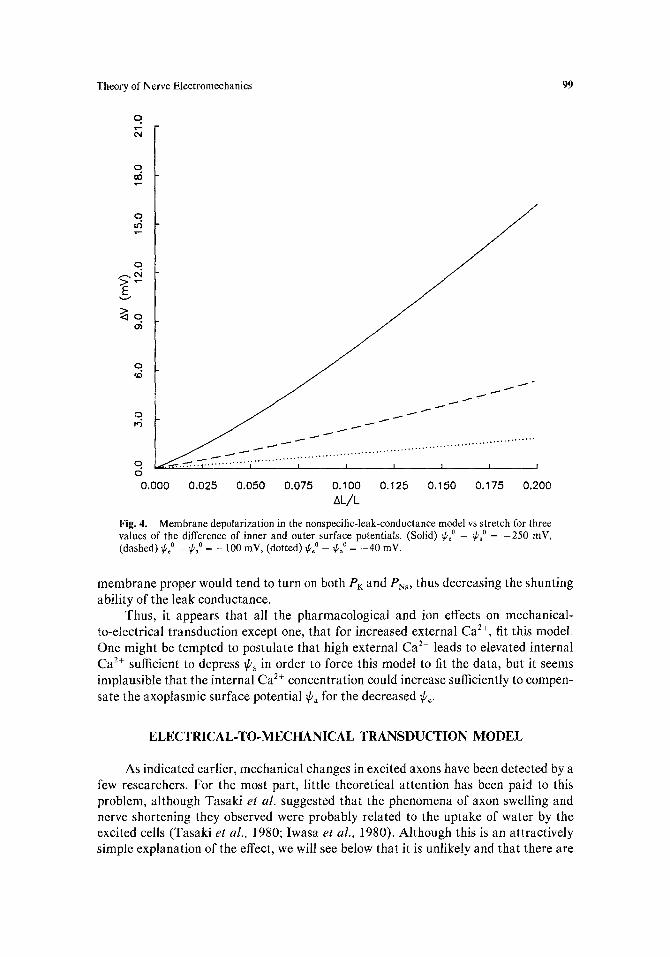
It is well known that PK and Pn, decrease when a cell is hyperpolarized. Thus, if a leak conductance which is independent of Vm is present, the cell will tend to become less ion selective and thereby, according to the Goldman equation [Eq. (4)], tend to depolarize. If inner and outer surface charge densities are such that I¢c [ > ]lPa I as has been suggested in the literature, then the effect of stretch is to hyperpolarize V~ as shown by Eq. (7). This hyperpolarization in the membrane proper then would tend to depress the Vm-sensitive PK and PN~, allowing the non-Vm-sensitive leak permeabilities to depolarize the bulk-bulk potential V. An iterative fit of this process, produced in the same way as was Fig. 3, is shown in Fig. 4.
In Fig. 4 the three curves correspond to (¢o _ Ca 0) of --250 mV (solid), -100 mV (dashed), and - 4 0 mV (dotted). As in Fig. 3, the V m dependence of P~: and PN, are modeled from steady-state Hodgkin-Huxley theory. In this case, however, constant "leakage" components are added to PK and PN, before V is calculated from the Goldman equation. The plots in Fig. 4 are for nonselective leak permeabilities, i.e., for PK,leak = PNa,leak" A similar fit with the leak permeabilities at the ratio PK,leak:PNa,leak of 14:1 gives similar curves which are reduced in magnitude from those in Fig. 4 by -20% (not shown).
As with the previous model, both the removal of external Na ÷ and the addition of procaine would greatly diminish the stretch-induced depolarization as is seen experi- mentally. Again, as is measured experimentally, the addition of TTX to the external medium would not noticeably affect the stretch-induced depolarization since the Vm-gated Na + channels do not mediate this depolarization. TTX, in fact, is predicted to enhance the effect by -5% [not shown].
The known effects of Ca 2+ on the stretch-induced depolarization do not fit this model, however. In increased external Ca 2+, the external surface charge is titrated, thus reducing [¢¢1. According to Eq. (7), as fie approaches ~b, in magnitude, the stretch-induced change in Vm is reduced. Thus, one would predict that increasing external Ca 2+ would reduce the mechanically induced depolarization. Likewise, decreased external Ca 2+ would reduce the effect of stretch on depolarization, but for another reason. The removal of external Ca 2+ would tend to increase the size of ¢o and thus, according to Fig. 1, would tend to depolarize V~,, This depolarization in the
Theory of Nerve Electromechanics 99
O
q
q u~
O ,~,ei
q ~O
t I
q i O
0.000 0.025 0.050 0.075 0.100 0.125 0.150 0.175 0.200 AL/L
Fig. 4. Membrane depolarization in the nonspecific-leak-conductance model vs stretch for three values of the difference of inner and outer surface potentials. (Solid) ~o _ ~ao = -250 mV, (dashed) ~poo _ ~pao = _ 100 mY, (dotted) ~o _ ~o = -40 mV.
membrane proper would tend to turn on both PK and PN,, thus decreasing the shunting ability of the leak conductance.
Thus, it appears that all the pharmacological and ion effects on mechanical- to-electrical transduction except one, that for increased external Ca 2+, fit this model. One might be tempted to postulate that high external Ca 2+ leads to elevated internal Ca 2~ sufficient to depress ~a in order to force this model to fit the data, but it seems implausible that the internal Ca 2+ concentration could increase sufficiently to compen- sate the axoplasmic surface potential ~ba for the decreased ~o.
ELECTRICAL-TO-MECHANICAL TRANSDUCTION MODEL
As indicated earlier, mechanical changes in excited axons have been detected by a few researchers. For the most part, little theoretical attention has been paid to this problem, although Tasaki et al. suggested that the phenomena of axon swelling and nerve shortening they observed were probably related to the uptake of water by the excited ceils (Tasaki et al., 1980; Iwasa et al., 1980). Although this is an attractively simple explanation of the effect, we will see below that it is unlikely and that there are

100 Gross, Williams, and Connor
other phenomena which could also produce such an effect and which, potentially, could be related to the molecular gating of ion channels.
Let us first consider the basis of the water-swelling hypothesis. It has long been known that polyelectrolyte gels swell in the presence of water due to the entrance of the solvent into the gel phase (Hermans, 1949). Water will enter and fill spaces between polymer chains if it is thermodynamically favorable to do so. Thus it is plausible that water entering the cell during excitation could cause the cell to swell.
However, consider the amount of water necessary to cause the cell radius to increase by an amount Aa during an action potential if the radius of the cell is "a" and the portion of the cell which is excited is of length "/." For a cylinder, a change in volume "V" is related to changes in radius and length by A V / V = 2Aa/a + Al/l. If one includes the elastic resistance of the nonexcited portion of the axon, then Al/ l = - ( a / 2 ) ( l / L ) A a / a , where ~ is Poisson's ratio (the ratio of radial contraction to longitudinal extension) and L is the full axon length. Since l - 1 mm, L > 1 cm, and 0 <
< 1/2, the largest magnitude of Al/ l is - 1 / 4 0 Aa/a. Thus, if Aa - 10 A (Iwasa and Tasaki, 1980) for squid axon (for which a - 250 urn), the volume of water which would be necessary to produce the swelling reported would be A V - 5 x 10 -15 m 3. This corresponds to 3 x 10 3 mol of water, or, since the flow was through -1.6 x 1 0 -6 m 2 of membrane, the flux of water necessary for such a swelling would be - 2 x 10 -4 mol/m 2. It is known that the fluxes of sodium and potassium for an action potential are ~3.7 x 10 -8 and - - 4 . 3 x 10 -8 mol/m 2, respectively (inward flux positive). Thus, it would appear that the necessary flux of water is about 4 orders of magnitude greater than the known fluxes of ions during the action potential. This result leads one to consider other possible explanations for the reported swelling.
Before we proceed, it should be mentioned that the swelling results reported by Iwasa and co-workers in single squid axons (Iwasa and Tasaki, 1980) and crab nerve (Tasaki et al., 1980; Iwasa et al., 1980) are in contradiction to the laser interferometric results of Hill et al., (1977), who measured a radial contraction of the axon when it was excited. Other less direct experiments indicated that nerve tissue swells slightly during activity, however (Hill, 1950; Cohen and Keynes, 1971; Cohen et al., 1972).
From the results of the previous section, one might be led to consider the putative converse role of membrane surface charge in the effect. However, this approach does not seem promising, as can quickly be seen.
In order for membrane surface charge to mediate the transduction process, electrical activity in the nerve cell must modify any of three parameters: (i) membrane surface charge, (ii) the surface-charge-generated E field at the axoplasmic and external surfaces of the cell, or (iii) the ion concentration profiles near the surfaces of the cell. That all three of these parameters are unchanged in an excited patch of axon membrane is shown by measurements of the voltage shift of ion permeability due to altered surface charge (i.e., Gilbert and Ehrenstein, 1969). In such experiments, the effect of surface charge can be fit to a model of static surface effects, unmodified by current flowing through the surface layers. In addition, modification of the last two parameters can be shown to perturb the static Gouy-Chapman double layer by <1% (D. Gross, unpublished results). Thus, other mechanisms for the transduction process must be considered.
The large magnitude of the electric field in the resting membrane (~107 V/m)

Theory of Nerve Electromechanics 101
and the large changes in the field during activity suggest that one or both of two classical dielectric mechanoelectric phenomena--piezoelectricity and electrostric- t ion--might play a role in the "converse effect" in nerve tissue. The possible contribution of these effects is considered in the next sections.
Electrostriction. The phenomenon of electrostriction is well-known to materials scientists concerned with the electromechanical properties of materials. The effect, as suggested by the name, is an electric field-induced deformation or stress in a dielectric. It is related to the minimization of total (electric plus elastic) energy in the material. As seen below, the effect is to be expected for all dielectric materials and the stress produced in the effect is proportional to the square of the imposed electric field.
It is well-known (Landau and Lifshitz, 1960; Jackson, 1975) that the presence of an electric field, whether in a material or in vacuo, produces a corresponding stress field. The form of the tensor describing this stress field, called the Maxwell stress tensor, is
t,, = - (E2 + E/)} - {6,EI 2 + 2 + E?)} (S)
62 - - 61 it2 = t2, = eE, E2 + E,E2,
2 • , ,
where
0 E
61 = Osl---~ (9)
0e 0e 32
0 5 ' 2 2 0 S 3 3 '
assuming the electric field is in the 1-direction (Stratton, 1941) and where s is the strain tensor in the dielectric of permitivity ~. Equations (8) and (9) apply to the case of a homogeneous, isotropic dielectric. As shown by Stratton (1941), the body force, or force per unit volume, in an isotropic dielectric in which an electric field is present is
E 2 0~ 1/~Xl [El2~l + (E22 + E32)62] FI = pE, 2 0 x , 2
k
o o } + ~x~ [E,E2(al - a2)] + 7x~ [E,E3(6, - 6 2 ) ] , ( 1 0 )
where p is the charge density in the dielectric, with the other two components of the body force obtained by cyclic advancement of the subscripts. For the case in which the dielectric is uncharged and homogenous, the body force reduces simply to
= (6, + 62) rE2. (11) 4
This expression presumes that the condition ~7 × /~ = _ ~ = 0 holds, which is true for axons in which the potential profile is V = V o f ( z ) / r , which is reasonable for resting or active axons, space clamped or not.

102 Gross, Williams, and Connor
Electrostriction in a dielectric tube was discussed in detail by Adams (1911 ). The calculation he presented may be applied to axons if the following assumptions are made: (i) the electric field in the membrane is radial only, (ii) the planar mechanical properties of the membrane may be represented by the two Lam6 elastic coefficients X and ~t, (iii) the cell is in elastic equilibrium, and (iv) there is no angular or length dependence in the problem. The first and last of these assumptions are clearly valid for a space-clamped axon; they are less satisfactory for a region of a cell in which an action potential is present. As the characteristic length across which potential changes occur axially is larger than the characteristic radial distance (the thickness of the membrane), only the radial electric field need be considered in this calculation] Thus, the axon responds to the local ff~ field only.
The second and third of these assumptions are somewhat more difficult to justify. In particular, the hydrodynamic damping due to the axoplasm and extracellular fluid close to the membrane surfaces could easily alter the time dependence of the mechanical response of the system. Thus, the calculations which follow are strictly true only for DC transmembrane potentials.
Adams considered a free, hollow dielectric cylinder which had a potential V applied from interior to exterior such that the electric field in the dielectric was radial. He calculated that the longitudinal strain for such a cylinder from electrostriction is
2d V 2 l 2d2E v (61 + 62)¢r, (12)
where l is the length of the cylinder, Ey = #(3X + 2#)/(X + U) is Young's modulus, and ~r = X/2(X + u) is Poisson's ratio. Kemble (1916) pointed out that Adams' assumption that end effects are negligible is not correct in the case of electrostriction since the decay of E at the ends of the cylinder produces a longitudinal force which does not diminish in relative significance with increasing cylinder length. An additional term due to these effects should be added to Eq. (12) to give
AI V 2 l 2dZE v [62 - (¢3, + ~2)cr]. (13)
This equation is the complete expression for electrostriction in a dielectric cylinder subject to the mentioned assumptions.
Thus, if the axon can be represented as a dielectric tube in contact with electrolyte, any change in membrane potential should be accompanied by a change in length of the axon if the cell is not fixed at the ends. (All of the above equations exploit the approximation that d<<a, which is true for axons down to 1 ~zm in diameter.)
To determine the amount of longitudinal strain 2xl/1 to be expected during a change in membrane potential, one needs to estimate the values of the parameters ¢51 and 62 in Eq. (13). Such an estimate is easy to derive if one makes the assumption that shear stress does not alter the dielectric constant. This condition is equivalent to equating 6~ and 62. This estimate is not unreasonable for a biomembrane since the interior of the membrane is very liquid-like (Singer and Nicholson, 1972; Webb, 1981)
7 The statement fails when the membrane potential goes to zero locally during an action potential. Thus, one might expect the following calculation to fail for the two times during an action potential when Vm ~ 0.

Theory of Nerve Electromechanics 103
and liquids support no shearing. With the above assumption, one can immediately express the variation of the dielectric constant in terms of strain:
& & & Ae = ASII ÷ - - AS2 2 -~ - - OSII 0S22 0S33 AS33' (1 4)
= ~A(S l l ÷ $22 ÷ $33) = ~A0,
where 0 is the trace of the strain tensor, known as the cubical dilatation. Equation (14) immediately leads to a relation for the electrostriction parameter 6, namely,
AO Op m Apm/AO = --Pro 0Orn, (15)
where Pm is the mass density. From the Clausius-Mossotti law 8 (Stratton, 1941), the relation of dielectric constant K = e/% to density in gases, liquids, and even some solids is, to a good approximation,
3Apm K = 1 + 1 - A p m (16)
where A is a constant determined by the nature of the dielectric. This relation leads immediately to
& E0 (K - 1)(K + 2). (17) Pm 0p---2 =
Combining Eqs. (13), (15), and (17), the expression for longitudinal strain becomes
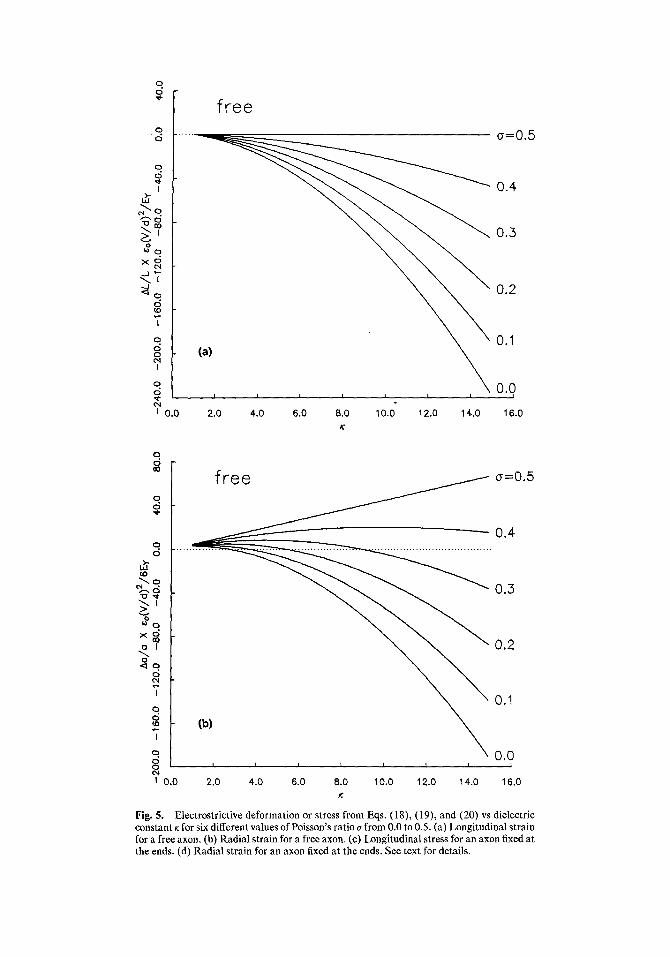
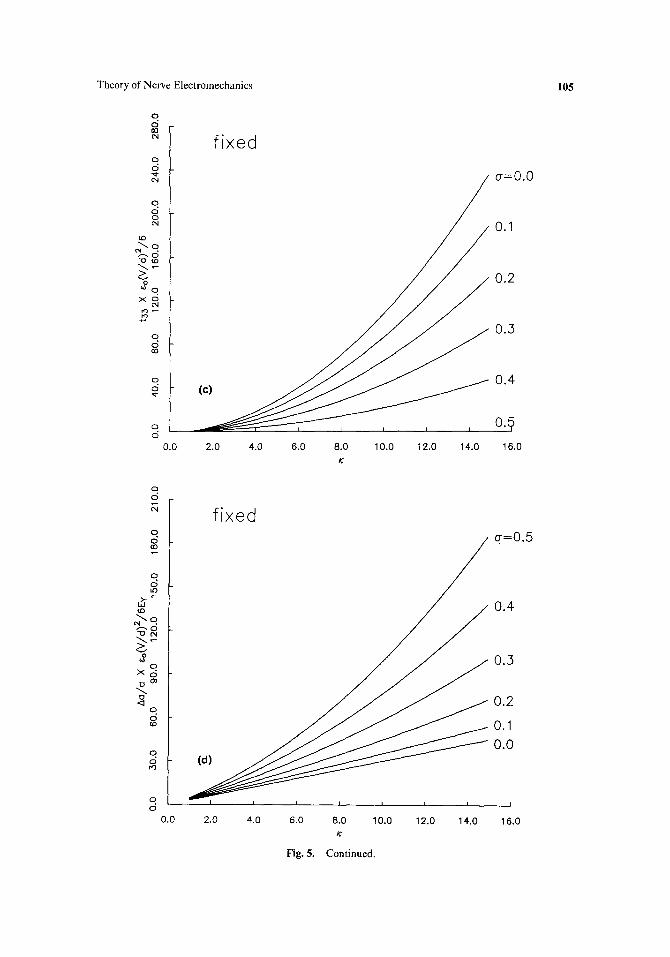
~l ~o ( V ) 2 I 6Ev d (K+ 2 ) (K- 1 ) ( 2 a - 1). (18)
It is possible to derive the longitudinal stress and radial displacement of an axon clamped at the ends as well as the radial displacement for the free axon for which Eq. (18) describes the longitudinal displacement. The expected radial displacement for the free axon is
Aa Eo ( V ) 2 a 6Er -d [3K(1 +cr) + (K+2)(K-- 1)(2~-- l)]. (19)
For an axon clamped at the end such that A l / l = 0, the longitudinal stress and radial displacement are
% [VVf , t33 =-~[~ ) ttx + 2)(K - 1)(1 - 2~)],
(20)
a 6e,~ d [3~(1+o)+2~ ~(~+2)(K-1)] .
The expressions in brackets in Eqs. (18), (19), and (20) are plotted in Figs. 5a-d. The
8 In the presence of permanent (orientation) polarization (see the next section), this equation is no longer valid.
O c5
O ~5
I
~ g
. d -m R
O to
7 o O
O O
O ¢q I
o O
tN i 0.0
free
o=0,5
0.4
0.3
0.2
0.1 (a
, , , , 0 . 0
2.0 4.0 6.0 8.0 10,0 12.0 14.0 16.0
q O
tO
~a
D I
< I Q
d c l
I
q 0 (b)
free G=0.5
0.4
0.3
0.2
0.1
I
0.0 0 I I I L L I I I e q
I 0,0 2,0 4.0 6.0 8.0 10.0 12.0 14.0 16.0
Fig. 5. Electrostrictive deformation or stress from Eqs. (18), (19), and (20) vs dielectric constant ~ for six different values of Poisson's ratio a from 0.0 to 0.5. (a) Longitudinal strain for a free axon. (b) Radial strain for a free axon. (c) Longitudinal stress for an axon fixed at the ends. (d) Radial strain for an axon fixed at the ends. See text for details.
Theory of Nerve Electromechanics 105
O
00 cN
q o
c',l
q O o t'N
to
v
~o X o
t"q
4-,
o ,:5 00
q O
O O
0.0
fixed
o-=0.0
0.1
0.2
.3
0,4 ( e )
, , 0.5
2 . 0 4 . 0 6 . 0 8 . 0 1 0 . 0 1 2 . 0 1 4 . 0 1 6 . 0
/("
O
q O G O
q o tO
tO ~--.q %.o
,2 O
x c ~
o <:1
q O
o O r~
o d
fixed
0.4
0.3
0.2
0.1
0.0
I I I I I I I I
0.0
o-=0.5
2.0 4.0 6.0 8.0 10.0 12.0 14.0 16.0
Fig. 5. Continued.

106 Gross, Williams, and Connor
plots are for the range of dielectric constants from 1 to 15. Families of six curves corresponding to Poisson's ratios of 0 to 0.5 in 0.1 steps are shown on each graph. For most materials, a is approximately 1/3, although highly incompressible materials such as rubber have a - 0.5.
The curves in Fig. 5 show that, for a free axon, a longitudinal contraction is generated as the magnitude of Vincreases, while the radial dimensional change may be of either sign depending on the values of K and ~ for the membrane. For a fixed axon, radial expansion and longitudinal tension can be expected.
To calculate the displacement through which the axon surface should move during an action potential, it is necessary to know the elastic and dielectric constants of the membrane. White (1974) gives values of Ev in the range of 10 s N t / m 2 for artificial membranes, while Hochmuth et al. (1973) give the value of ~103 N t / m 2 for the red-cell membrane. Evans and Skalak (1979) quote a value of 2 × 104 N t / m 2 for the unfertilized sea urchin egg. If one takes Ey = 105 N t / m 2, a = 0.3, and K = 5, the estimated radial displacement for a 180-#m-diameter nerve axon undergoing a change in bulk-to-bulk potential from - 6 0 to + 40 mV is a 2-urn increase if the ends of the excited portion of the axon are assumed fixed and the cell is space clamped. One must note that this result overestimates the radial displacement during an action potential for two reasons. First, the hydrodynamic damping of the axon due to extracellular, intracellular, and membrane viscosity would enter as a time-dependent factor. The present calculation is for steady-state displacements only. Second, it is clear that an axon is not a free, hollow cylinder, incompressible across its thickness as was assumed above. It is well-known that cytoplasmic structural proteins form an intracellular matrix which could strongly modify the elastic properties of an axon [cf. the case of spectrin in erythrocytes (Hochmuth, 1982; Coakley et al., 1978)]. In addition, the glycocalix coat of the axon membrane as well as the Schwann cells surrounding it could prove to be critical in the elastic behavior of the cell. Thus, the rather large value for radial displacement calculated above, compared to the measured 1 to 10 ~ displace- ments, is perhaps not unreasonable. An increase in the effective stiffness of the axon membrane to 109 N t / m 2 would reduce the expected displacement to 2 ~.
It should be mentioned that in applying Eqs. (18), (19), and (20) one must assume some variation of the potential V. For the above calculations, a change in V from - 60 to +40 mV during the action potential was assumed. In addition, it was assumed that the parabolic dependence of the four equations was symmetric about V = 0. In measurements of birefringence changes during squid axon activity, Cohen and co-workers found an optical change proportional to the square of potential which they thought was similar to the Kerr effect (Cohen, 1973; Cohen et al., 1971); this change was offset from V = 0 by about + 60 inV. If a similar offset is included in the above prediction, the calculated displacement reverses sign but is of the same magnitude.
Piezoelectr ici ty . Since there is reasonable evidence of permanently polarized molecules in excitable tissue and in modified artificial bilayers (Armstrong and Bezanilla, 1973; Yantorno et al., 1982), one is encouraged to investigate the possibility of a piezoelectric effect in axons since permanent polarization in materials leads to both a pyro- and a piezoelectric effect. As a simple example, consider a permanent radial polarization ~. If the membrane is stretched, the density of the polarization decreases inversely proportionally to the area increase in the membrane. Such a

Theory of Nerve Electromechanics 107
variation of spontaneous or permanent polarization as a function of strain (or stress) is one aspect of piezoelectricity (Cady, 1946). Some materials, e.g., quartz, do not have a spontaneous polarization but still become polarized when stressed.
In general, the relation between induced polarization and strain in a piezoelectric material can be described by a third-rank tensor equation
Pi = eijxsjk, (21)
where Pi is the polarization induced by the strains sjk (note that repeated indices imply a summation over those repeated). If, for example, the above hypothetical permanent polarization were present in the axon membrane, the piezoelectric e-tensor would have three non zero elements, ell I = el2 2 = ei3 3 = - p . As well as producing an electric polarization when strained, a piezoelectric body will exhibit strain when an electric field is applied across it. The tensor relation for this (coverse) effect is
sjk = d~ykEi, (22)
where dijk = egz,~Cikl,,, the tensor cjktm relating stress and strain by sjk = --Cjkzmttm. Thus, strain directly proportional to the electric field is produced. This response is to be contrasted with electrostriction, in which the strain produced by changes in the electric field is proportional to the s q u a r e of the field.
To estimate the magnitude of the strain induced by changes in the membrane electric field, an estimate of the permanent polarization is needed. Since "gating" currents can be construed to be due to the relaxation of permanent polarization associated with the Na + channel, one can use the published values of gating current to estimate ft. In the measurements of gating currents, a charge movement of -5 x 10-~6 coul/#m 2 is seen during the opening of the Na + channel (Armstrong and Bezanilla, 1973; Rojas and Keynes, 1975). If one assumes that this charge movement is due to the relaxation of a permanent dipole density, then the magnitude of that density would be of the order of magnitude of the charge movement, or ~5 x 10 -4 coul/m 2. As the measured gating current is outward when the "gate" opens, the permanent polariza- tion is directed radially inward. This is thus equal to the piezoelectric e111, era, and ei33 coefficients, leading to d~11, d122, and dl3 3 coefficients of - I x 10 -9 m / V if Ev -105 N t / m 2 and o- -0.25.
This value is uncertain, however, for at least three reasons. First, the current transient measured during the opening of the Na + channel may be just the rapid phase of a polarization change, so much of the gating polarization may have gone undetected. Second, there may be polarization associated with the Na + channel which is not related to the turn-on of the channel and thus would not be measured in such a gating current experiment. Third, there may be other regions of permanent polarization associated with the K + channel (Bezanilla et al. , 1982) or other macromolecules in the membrane which could contribute to the (average) permanent polarization which affects the piezoelectric coefficients.
However, if one employs the value of the piezoelectric d coefficient calculated above, Eq. (22) predicts that for a 100-mV action potential, the radial deformation for a 500-#m-diameter axon with elastic properties mentioned above will be 6 ~tm, which is an expansion about three times larger than that predicted above for electrostriction. The same caveats about time dependence and modification of the cell mechanical

108 Gross, Williams, and Connor
properties hold here also--thus, one must consider both electrostriction and piezoelec- tricity with equal weight.
One should note that the above calculation presumed that the permanent polarization in the membrane was uniformly distributed. In fact, one would expect that the majority of the polarization is associated with ion channels and other membrane proteins. In such a case, the analysis presented above is not accurate since the presumed piezoelectric proteins would be effectively mechanically decoupled from the liquid-like lipid phase of the membrane due to the large difference in the elastic stiffness of the two phases. Thus, a piezoelectric deformation of the channels might be quite large, while the deformation of the remainder of the membrane would be small. It is easily seen that piezoelectricity could in principle play an important role in the conformational changes of an ion channel in an excitable cell, while the cell itself might display overall only a (relatively) small effect.
In addition to electrical-to-mechanical transduction, piezoelectricity as described above could be responsible for the direct (mechanical-to-electrical) transduction phenomenon seen in axons [see Eq. (21)]. However, the above discussion of the converse effect and particularly the cautions mentioned in the previous paragraph discourage one from drawing conclusions until more is known about the molecular- level structure of the axon membrane.
DISCUSSION
Mechanial-to-Electrical Transduction. The surface charge transduction mod- els described above predict the observed depolarization of axon or mechanoreceptor membranes when they are stretched. There is agreement between the models and the reported experiments, as described above. However, two particular pieces of experi- mental data appear to conflict with the first (direct depolarization) model.
First, the reported lack of TTX sensitivity of the mechanically induced depolar- ization indicates that the "normal" Na + channel is not the pore which is activated to carry Na + ions during the process. Second, although Na + ion has been shown to be the most important ion in the process, the above model predicts that both K + and Na + permeability should be increased by the internal stress-induced depolarizing field.
Both of these apparent inconsistencies can be illuminating in explaining some recent results of stretching squid axon (Wells and Goldman, 1981). In these experiments, two components of depolarization were found: one which was rapid and unaffected by TTX; the other which was slower and was abolished by TTX or exchange of choline for Na + in the external bathing solution. In addition, a slow hyperpolarization always followed the stretch-induced depolarization. One might interpret the slower response to stretch as due to the above surface charge model since the sensitivity to TTX and depolarization agree with the hypothesis that the "normal" Na + and K + channels are responding to an intramembrane stretch-induced depolariz- ing E field.
The second surface-charge model, that for the process of enhanced effectiveness of leakage pathways, is somewhat more difficult to bring into agreement with experiments. Although this model fits the TTX-insensitivity found in most experi-

Theory of Nerve Electromechanics 109
ments, the predicted behavior in elevated external Ca z+ is not found experimentally. In addition, one would not predict the large stretch-induced increase in ion permability as is observed.
It should be apparent that the best test of these models as well as any others is a voltage-clamp study of a stretched axon. Until such time as a definitive study is presented, one cannot definitively accept or rule out the above models.
Some totally different kinds of mechanisms for the phenomenon could also be imagined. For example, a transmembrane streaming potential resulting from a hydrostatic pressure change in a mechanically deformed cell forcing Na + through a nonion-selective channel might be a significant effect. Streaming potentials have been measured for ion flow produced by osmotic pressure differences across channels in artificial membranes (Miller, 1982); the possibility that mechanically induced flow is responsible for the generator potential needs to be explored. The streaming potential Miller measures in a sarcoplasmic reticulum K + channel, 1.1 mV/osmolal, implies that a transmembrane pressure gradient of 250 atmospheres is required for a 10-mV potential change. Clearly, this is unreasonable for axon transduction. However, the implied zeta potential, or effective electrokinetic surface potential, is 7 mV, very small by many standards. A channel with a larger zeta potential would be a more likely transducer.
One could also imagine another method of transduction--simple mechanical deformation of ion channels. Although this would seem to be the simplest and, thus, most aesthetically pleasing model, it also is difficult to support. Since ion pathways are likely through the interior regions of protein complexes, the mechanical coupling between the liquid-like lipid phase and the protein molecule itself is difficult to accept. One tends to believe that proteins would be fairly stiff; thus, the mechanism by which large stresses could be transmitted to these proteins is difficult to imagine.
ElectricaLto-Mechanical Transduction. The calculation presented above for the electrostrictive/piezoelectric deformation of an excited axon considered to axon membrane to be both homogeneous and isotropic, which clearly is not the case for a real membrane. In addtition, the hydrodynamic properties of the aqueous phases surrounding the membrane were ignored, i.e., the calculation was done for a time- independent deformation of the membrane. Again, this approximation might tend to weaken the argument. Nevertheless, the calculation presented indicates that the electrostrictive or piezoelectric effects might be of sufficient magnitude in nerve to consider them in more detail as possible candidates for the physical transduction mechanism.
Once again, it should be clear that an experimental study of the mechanical deformation of a voltage-clamped axon could discriminate between one of these two mechanisms (due to their Vm 2 or Vm I dependence) or a different, osmotic mechanism (due to its dependence on transmembrane current) in the effect.
ACKNOWLEDGMENTS
This work was supported by the University of Illinois Research Board and by the Veterans Administration through the Helen Hayes Hospital, West Haverstraw, New

110 Gross, Williams, and Connor
Y o r k 10993. D G rece ived fe l lowship suppor t f r o m the E x x o n E d u c a t i o n F o u n d a t i o n
du r ing par t of the t i m e this work was in progress .
R E F E R E N C E S
Abraham-Shrauner, B. (1975). Generalized Gouy-Chapman potential of charged phospolipid membranes with divalent cations. J. Math. Biol. 2:333-339.
Adams, E. P. (1911). On electrostriction. Phil. Mag. 22:889-900. Armstrong, C. M., and Bezanilla, F. (1973). Currents related to movement of the gating particles of the
sodium channels. Nature 242:459-461. Bezanilla, F., White, M. M., and Taylor, R. E. (1982). Gating currents associated with potassium channel
activation. Nature 296:657-659. Bockris, J. O'M, and Reddy, A. K. N. (1970), Modern Electrochemistry, Plenum Press, New York. Bryant, S. H., and Tobias, J. M. (1955). Optical and mechanical concomitants of activity in carcinius nerve.
J. Cell Comp. Physiol. 46:71-95. Cady, W. G. (1946). Piezoelectricity, McGraw-Hill, New York. Coakley, W. T., Bater, A. J., and Deeley, J. O. T. (1978). Vesicle production on heated and stressed
erythrocytes. Biochim. Biophys. Acta 512:318-330. Cohen, L. B. (1973). Changes in neuron structure during action potential propagation and synaptie
transmission. Physiol. Rev. 53:373-418. Cohen, L. B., and Keynes, R. D. (1971). Changes in light scattering associated with the action potential in
crab nerves. J. Physiol. (Lond.) 212:259-275. Cohen, L. B., Hille, B., Keynes, R. D., Landowne, D., and Rojas, E. (1971). Analysis of the potential-
dependent changes in optical retardation in the squid giant axon. J. Physiol. (Lond.) 218:205-237. Cohen, L. B., Keynes, R. D., and Landowne, D. (1972). Changes in axon light scattering that accompany the
action potential: Current dependent components. J. Physiol. (Lond.) 224:727-752. Cole, K. S. (1968). Membranes, Ions, and Impulses, University of California Press, Berkeley. Diamond, J., Gray, J. A. B., and Inman, D. R. (1958). The relation between receptor potentials and the
concentration of sodium ions. J. Physiol. (Lond.) 142:362-364. Edwards, C., Terzuolo, C. A., and Washizu, Y. (1963). The effect of changes of the ionic environment upon
an isolated crustacean sensory neuron. J. Neurophysiol. 26:948-957. Elul, R. (1967). Fixed charge in the cell membrane. J. Physiol. (Lond.) 189:351-356. Evans, E. A., and Skalak, R. (1979). Mechanics and thermodynamics of biomembranes. Crit. Rev. Bioeng.
3:181-418. Ganot, G., Wong, B. S., Binstock, L., and Ehrenstein, G. (1981). Reversal potentials corresponding to
mechanical stimulation and leakage current in Myxicola giant axons. Biochim. Biophys. Acta. 649:487-491.
Gilbert, D. C., and Ehrenstein, G. (1969). Effect of divalent cations on potassium conductance of squid axons: Determination of surface charge. Biophys. J. 9:447-463.
Goldman, D. E. (1943). Potential, impedance, and rectification in membranes. J. Gen. Physiol. 27:37-60. Goldman, D. E. (1965). The transducer action of mechanoreceptor membranes. Symp. Quant. Biol.
30:59-68. Grahame, D. C. (1947). The electrical double layer and the theory of electrocapillarity. Chem. Rev.
41:441-501. Gross, D. J., and Williams, W. S. (1978). Piezoelectric theory of electromechanical effects in nerve. Paper
TU-AM-FI 5 given at the 22nd Annual Meeting of the Biophysical Society, Washington, D.C. Hermans, P. G. (1949). Gels. In Colloid Science, Vol. II (Kryut, H. R., Ed.), Elsevier, New York, pp.
483-651. Hill, B. C., Schubert, E. D., Nokes, M. A., and Michelson, R. P. (1977). Laser interferometer measurement
of changes in crayfish axon diameter concurrent with action potential. Science 196:426-428. Hill, D. K. (1950). The volume change resulting from stimulation of a giant nerve fiber. J. Physiol. (Lond.)
111:304-327. Hochmuth, R. M. (1982). Solid and liquid behavior of red cell membrane. Ann. Rev. Biophys. Bioeng.
11:43-55. Hoehmuth, R. M., Mohandas, N., and Blackshear, P. L., Jr. (1973). Measurement of the elastic modulus for
red cell membrane using a fluid mechanical technique. Biophys. J. 13:747-762. Hodgkin, A. L. (1951). The ionic basis of electrical activity in nerve and muscle. Biol. Rev. 26:339-409. Iwasa, K., and Tasaki, L (1980). Mechanical changes in squid giant axon associated with production of
action potentials. Bioehem. Biophys. Res. Comm. 95:1328-1331.

Theory of Nerve Electromechanics 111
lwasa, K., Tasaki, I., and Gibbons, R. C. (1980). Swelling of nerve fibers associated with action potentials. Science 210:338-339.
Jackson, J. D. (1975). Classical Electrodynamics, John Wiley and Sons, New York. Julian, F. J., and Goldman, D. E. (1962). The effects of mechanical stimulation on some electrical properties
of axons. J. Gen. Physiol. 46:297-313. Junge, D. (1976). Nerve and Muscle Excitation, Sinauer Associates, Sunderland, Mass, Kemble, E. C. (1916). Notes on the end effect in the electrostriction of cylindrical condensers. Phys. Rev.
7:614-624. Landau, L. D., and Lifshitz, E. M. (1960). Electrodynamics of Continuous Media, Pergamon Press,
Oxford. Loewenstein, W. R., Terzuolo, C. A., and Washizu, Y. (1963). Separation of transduction and impulse-
generating processes in sensory receptors. Science 142:1180. McLaughlin, S. L. (1977). Electrostatic potentials at membrane-solution interfaces. Curt. Top. Memb.
Tr ansp. 9:71-144. McLaughlin, S., and Poo, M. (1981). The role of electro-osmosis in the electric-field-induced movement of
charged macromolecules on the surfaces of cells. Biophys. J. 34:85-93. Miller, C. (1982). Coupling of water and ion fluxes in a K÷-selective channel of sarcoplasmic reticulum.
Biophys. J. 38:227-230. Obara, S. (1968). Effects of some organic cations on generator potential of crayfish stretch receptor. J. Gen.
Physiol. 52:363-386. Rojas, E., and Atwater, I. (1968). An experimental approach to determine membrane charges in squid giant
axons. J. Gen. Physiol. 51:131s-135s. Rojas, E., and Keynes, R. D. (1975). On the relation between displacement currents and activation of the
sodium conductance in the squid giant axon. Phil. Trans. Roy. Soc. (Lond.) B 270:459-482. Singer, S. J., and Nicholson, G. L. (1972). The fluid mosaic model of the structure of cell membranes.
Science 175:720-731. Stepanov, K. A. (1968). Shortening of the frog sciatic nerve on stimulation. Biophysics 13:634-636. Stratton, J. A. (1941 ). Electromagnetic Theory, McGraw-Hill, New York. Tasaki, I., and lwasa, K. (1980). Shortening of nerve fibers associated with propagated nerve impulse.
Biochem. Biophys. Res. Comm. 94:716-720. Tasaki, I., Iwasa, K., and Gibbons, R. C. (1980). Mechanical changes in crab nerve fibers during action
potential. Japan. J. Physiol. 30:897-905. Webb, W. W. (1981). Luminescence measurements of macromolecular mobility. Ann. N.Y. Acad. Sci.
366:300-314. Wells, J. B., and Goldman, D. E. (1981). Membrane potential changes during stretch in squid giant axons,
Biophys. J. 33:91 a. White, S. H. (1974). Comments on "Electrical breakdown of bimotecular lipid membranes as an
electromechanical instability." Biophys. J. 14:155-158. Yantorno, R. E., Takashima, S., and Mueller, P. (1982). Dipole moment of alametfiiein as related to
voltage-dependent conductance in lipid bilayers. Biophys. J. 38:105-110.





























