Embed Size (px)
Citation preview

[CANCER RESEARCH 45, 3843-3849, August 1985]
Thermotolerance and Profile of Protein Synthesis in Murine Bone Marrow Cellsafter Heat Shock1
Nahid F. Mivechi2 and Gloria C. Li
Radiation Oncology Research Laboratory, CED-200, The University of California at San Francisco, San Francisco, California 94143
ABSTRACT
In this study, we first investigated the survival of colony-forming units, granulocyte and macrophage (CFU-GM), after a
single heat treatment. We then examined the induction, development, and decay of thermotolerance in CFU-GM. Finally, we
analyzed the profiles of protein synthesis in the total murine bonemarrow population during the development of thermotolerance.Several salient features emerged from our study: (a) granulocyte-
macrophage progenitors were very sensitive to heat as compared to other mammalian cell lines; (b) CFU-GM can developthermotolerance after both prolonged heating at 41 -42°C or anacute heat treatment at 43°C in vitro; (c) thermotolerance in
CFU-GM can be induced in vivo; the kinetics of development of
thermotolerance in vivo is similar to that in vitro; (d) in contrastto other cell lines where thermotolerance lasts for several days,tolerance acquired by CFU-GM disappeared within 24 h, regard
less of the temperature or duration of the initial heat treatment.The difference between the kinetics of systemic thermotoler
ance and thermotolerance in CFU-GM in the same animal modelshows that bone marrow stem cells, or at least CFU-GM, are
not the critical targets for systemic thermal death. When proteinsynthesis profiles of the heat-shocked bone marrow cells werecompared to those from nonheated controls by one- and two-
dimensional gel electrophoresis, the rates of synthesis of the M,70,000 and 87,000 proteins were shown to be enhanced duringthe development of thermotolerance. The enhanced rate of synthesis of these polypeptides lasted only 2-4 h and then returned
to the control value.
INTRODUCTION
The phenomenon of thermotolerance in mammalian cells hasbeen extensively documented in the literature, and thermotolerance is known to play an important role in modifying the responseof cells to fractionated hyperthermia (1-7). Many investigators
have examined the induction, development, and decay of thermotolerance in normal rodent tissues (8-11 ). A few studies have
been reported on the heat response of murine and human bonemarrow progenitors after a single heat treatment (12,13). However, there has been no data on the kinetics of induction anddevelopment of thermotolerance of bone marrow cells heatedeither in vivo or in vitro.
Studies on the thermal response of bone marrow cells aftersingle or multiple heat treatments have intrinsic biological interest. Bone marrow contains pluripotent stem cells. These stemcells have the ability to divide and give rise to differentiatedprogeny at some stage of their development. Recent evidence
1This work was supported by NIH Grant CA-31397.2To whom requests for reprints should be addressed.Received11/6/84; revised 3/4/85; accepted 4/20/85.
has shown that differentiation may involve HSPs3 (14-17). In
recent years, tissue culture techniques have been developed toselectively assay progenitor populations in each of the majorlines of blood cells. Using these assays, one can easily study theeffect of hyperthermia on progenitor cells in different blood celllineages. Such information may be of clinical value. If patientswhose bone marrow has been severely compromised by previous treatments such as chemotherapy are treated by hyperthermia, bone marrow suppression might become a treatment-limiting factor. Furthermore, when whole-body hyperthermia is
combined with chemotherapy, bone marrow toxicity may play adose-limiting role.
The molecular mechanism of thermotolerance is not wellunderstood, although there is considerable evidence that it maybe correlated with a preferential synthesis of some specificproteins, termed HSPs (18,19). The levels of certain HSPs werefound to correlate well with thermal sensitivity in many mammalian cell lines (18-21). Recent studies have shown that mouse
embryos (14) or sea urchin embryos (16) have a developmentalstage-specific response to heat shock, specifically at the levels
of HSP synthesis. This transition in the expression of HSP hasalso been reported for very early embryos of Drosophila melan-
ogaster (15). This may be a general property of early embryoniccells. It is also suggested that HSPs may be associated with celldifferentiation (14, 16, 17). Since bone marrow is a continuallyrenewing developmental system dependent on differentiation ofa stem cell population, it is of interest to examine whether heatshock can induce HSP synthesis in bone marrow cells. No dataare available on the protein synthesis profiles of bone marrowcells after heat shock. The relationship between the kinetics ofthermotolerance and the synthesis of HSPs in bone marrow cellsis also not known.
We report here on experiments designed to study the effectsof hyperthermia on the induction of thermotolerance in bonemarrow cells, specifically the precursors of monocytes, granulo-cytes, and macrophages (CFU-GM). We also examined the pro
files of protein synthesis in bone marrow cells after a primingheat treatment.
MATERIALS AND METHODS
Mice. Female C3H/MGH mice (8 to 12 weeks old) were housed 4 toa cage and given standard lab chow and acidified water ad libitum.
Marrow Cell Preparation. Bone marrow cell suspensions were prepared by flushing the femur with MEM supplemented with 15% FCS.The cell suspension was then counted using a hemocytometer andcentrifuged at 1000 rpm for 12 min. The resulting pellet was resuspendedand diluted to approximately 1 x 106 cells/ml in MEM plus 15% PCS for
3The abbreviations used are: HSP, heat shock protein; HSP70, HSP87, heatshock proteins with molecular weights of 70,000 and 87,000, respectively; CFU-GM, colony-forming units, granulocyte and macrophage; SDS, sodium dodecylsulfate; MEM, minimal essential medium; «-MEM,enriched minimal essential medium; PCS,fetal calf serum.
CANCER RESEARCH VOL. 45 AUGUST-1985
3843
on May 18, 2020. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
THERMOTOLERANCE AND PROTEIN SYNTHESIS IN MURINE BONE MARROW CELLS
further experiments.In Vitro Heating. The cell suspensions in MEM supplemented with
15% PCS were heated in 15-ml centrifuge tubes in a precision temperature-controlled circulating water bath (Precision Scientific Group). The
pH of the medium was maintained at 7.4 during and after heating bypreflushing the cell suspension with 5% CO2 and 95% air before sealingthe tubes. After heating, the cell suspensions were centrifuged andresuspended in «-MEM before plating (see below).
In Vivo Heating. For in vivo heating at temperatures of 41 °C,the mice
were heated by half-body immersion in a circulating hot water bath as
described previously by Li et al. (22). For ¡nvivo heating at temperaturesgreater than 41 °C,the mice were anesthetized with sodium pentobarbital
(0.06 mg/g body weight) (Diamond Laboratories, Inc., Des Moines, lA),and only one leg of the animal was immersed in the water bath.
Temperature Measurement. The temperature of the bone marrowduring in vivo heating was monitored by inserting a thermal probe (BailyInstruments, Saddle Brook, NJ) in the femur of the anesthetized mouse.Rectal probes were used to monitor the body temperature.
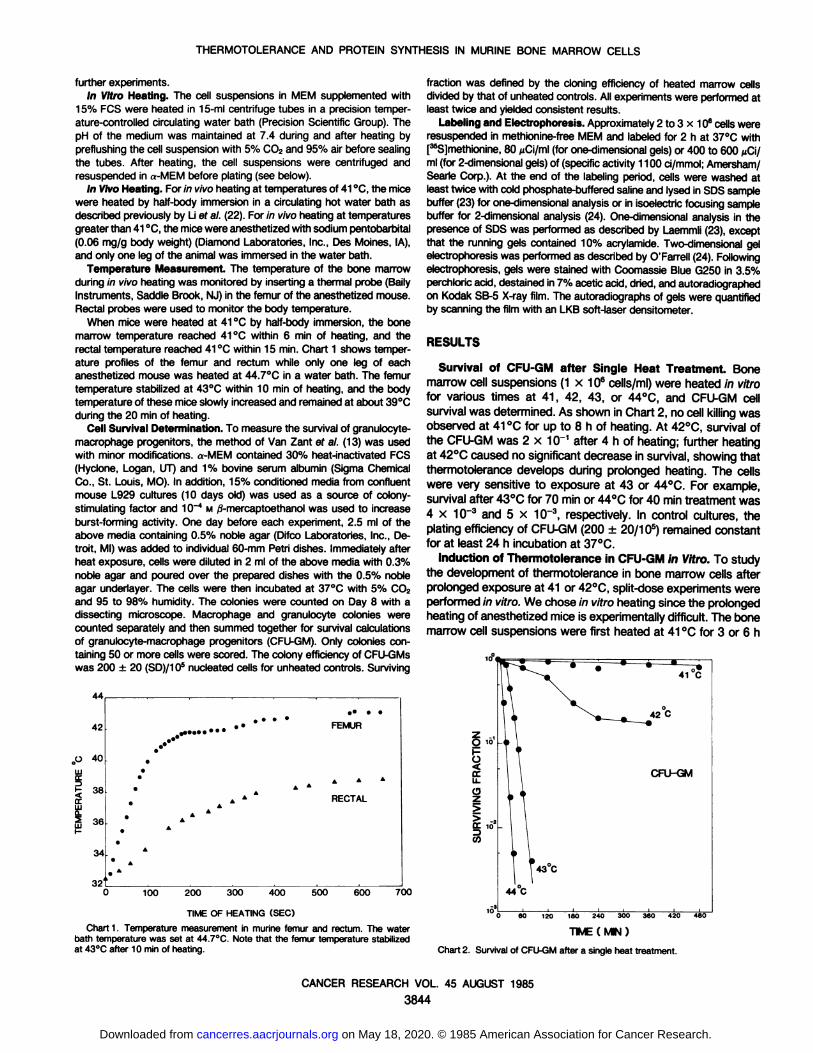
When mice were heated at 41 °Cby half-body immersion, the bonemarrow temperature reached 41 °Cwithin 6 min of heating, and therectal temperature reached 41 °Cwithin 15 min. Chart 1 shows temper
ature profiles of the femur and rectum while only one leg of eachanesthetized mouse was heated at 44.7°C in a water bath. The femurtemperature stabilized at 43°C within 10 min of heating, and the bodytemperature of these mice slowly increased and remained at about 39°C
during the 20 min of heating.Cell Survival Determination. To measure the survival of granulocyte-
macrophage progenitors, the method of Van Zant et al. (13) was usedwith minor modifications. a-MEM contained 30% heat-inactivated FCS
(Hyclone, Logan, UT) and 1% bovine serum albumin (Sigma ChemicalCo., St. Louis, MO). In addition, 15% conditioned media from confluentmouse L929 cultures (10 days old) was used as a source of colony-stimulating factor and 10"4 M ,8-mercaptoethanol was used to increase
burst-forming activity. One day before each experiment, 2.5 ml of the
above media containing 0.5% noble agar (Difco Laboratories, Inc., Detroit, Ml) was added to individual 60-mm Petri dishes. Immediately after
heat exposure, cells were diluted in 2 ml of the above media with 0.3%noble agar and poured over the prepared dishes with the 0.5% nobleagar underlayer. The cells were then incubated at 37°C with 5% CO2
and 95 to 98% humidity. The colonies were counted on Day 8 with adissecting microscope. Macrophage and granulocyte colonies werecounted separately and then summed together for survival calculationsof granulocyte-macrophage progenitors (CFU-GM). Only colonies containing 50 or more cells were scored. The colony efficiency of CFU-GMswas 200 ±20 (SD)/105 nucleated cells for unheated controls. Surviving
4442„ü
40UH
38È1
363432«•
••.•••.
.•FEMURX"""*"••
• **A
•*»
ARECTAL'A«
.»•
A»
A•
*0
100 200 300 400 500 600 7(
fraction was defined by the cloning efficiency of heated marrow cellsdivided by that of unheated controls. All experiments were performed atleast twice and yielded consistent results.
Labeling and Electrophoresis. Approximately 2 to 3 x 106 cells wereresuspended in methionine-free MEM and labeled for 2 h at 37°Cwith[^SJmethionine, 80 pC\/m\ (for one-dimensional gels) or 400 to 600 nC\/
ml (for 2-dimensional gels) of (specific activity 1100 ci/mmol; Amersham/
Searle Corp.). At the end of the labeling period, cells were washed atleast twice with cold phosphate-buffered saline and lysed in SDS samplebuffer (23) for one-dimensional analysis or in isoelectric focusing samplebuffer for 2-dimensional analysis (24). One-dimensional analysis in the
presence of SDS was performed as described by Laemmli (23), exceptthat the running gels contained 10% acrylamide. Two-dimensional gelelectrophoresis was performed as described by O'Farrell (24). Following
electrophoresis, gels were stained with Coomassie Blue G250 in 3.5%perchloric acid, destained in 7% acetic acid, dried, and autoradiographedon Kodak SB-5 X-ray film. The autoradiographs of gels were quantifiedby scanning the film with an LKB soft-laser densitometer.
RESULTS
Survival of CFU-GM after Single Heat Treatment. Bonemarrow cell suspensions (1 x 106 cells/ml) were heated in vitrofor various times at 41, 42, 43, or 44°C, and CFU-GM cell
survival was determined. As shown in Chart 2, no cell killing wasobserved at 41 °Cfor up to 8 h of heating. At 42°C,survival ofthe CFU-GM was 2 x 10~1 after 4 h of heating; further heatingat 42°Ccaused no significant decrease in survival, showing that
thermotolerance develops during prolonged heating. The cellswere very sensitive to exposure at 43 or 44°C. For example,survival after 43°Cfor 70 min or 44°Cfor 40 min treatment was4 x 10~3 and 5 x 10~3, respectively. In control cultures, the
plating efficiency of CFU-GM (200 ±20/10s) remained constantfor at least 24 h incubation at 37°C.
Induction of Thermotolerance in CFU-GM in Vitro. To studythe development of thermotolerance in bone marrow cells afterprolonged exposure at 41 or 42°C,split-dose experiments were
performed in vitro. We chose in vitro heating since the prolongedheating of anesthetized mice is experimentally difficult. The bonemarrow cell suspensions were first heated at 41 °Cfor 3 or 6 h
TIME OF HEATING (SEC)
Chart 1. Temperature measurement in murine femur and rectum. The waterbath temperature was set at 44.7°C. Note that the femur temperature stabilizedat 43°Cafter 10 min of heating.
60 120 1
TIME ( MIN )
Chart 2. Survival of CFU-GMafter a single heat treatment.
CANCER RESEARCH VOL. 45 AUGUST 1985
3844
on May 18, 2020. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
THERMOTOLERANCE AND PROTEIN SYNTHESIS IN MURINE BONE MARROW CELLS
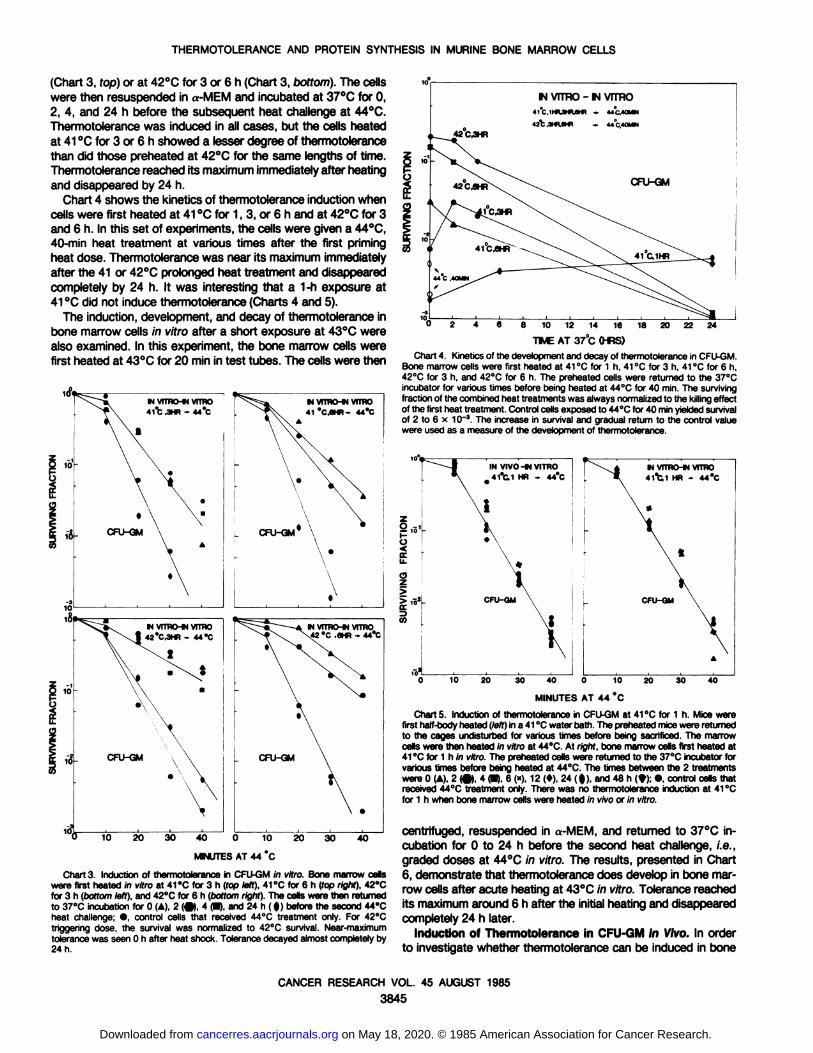
(Chart 3, fop) or at 42°Cfor 3 or 6 h (Chart 3, bottom). The cellswere then resuspended in a-MEM and incubated at 37°Cfor 0,2, 4, and 24 h before the subsequent heat challenge at 44°C.
Thermotolerance was induced in all cases, but the cells heatedat 41 °Cfor 3 or 6 h showed a lesser degree of thermotolerancethan did those preheated at 42°Cfor the same lengths of time.
Thermotolerance reached its maximum immediately after heatingand disappeared by 24 h.
Chart 4 shows the kinetics of thermotolerance induction whencells were first heated at 41 °Cfor 1, 3, or 6 h and at 42°Cfor 3and 6 h. In this set of experiments, the cells were given a 44°C,
40-min heat treatment at various times after the first priming
heat dose. Thermotolerance was near its maximum immediatelyafter the 41 or 42°Cprolonged heat treatment and disappeared
completely by 24 h. It was interesting that a 1-h exposure at41 °Cdid not induce thermotolerance (Charts 4 and 5).
The induction, development, and decay of thermotolerance inbone marrow cells in vitro after a short exposure at 43°Cwere
also examined. In this experiment, the bone marrow cells werefirst heated at 43°Cfor 20 min in test tubes. The cells were then
w vrmo-w VITRO42°C.3HR - 44 °C
N VITRO-*! VITRO42 °C.6HR - 44°C
10MINUTES AT 44 °C
20 30 40
Chart 3. Induction of thermotolerance in CFU-GM in vitro. Bone marrow cellswere first heated in vitro at 41 °Cfor 3 h (top left), 41 °Cfor 6 h (fop right). 42°Cfor 3 h (öoffomleft), and 42"C for 6 h (bottom right). The cells were then returnedto 37°Cincubation for 0 (A), 2 (•),4 (•),and 24 h ( |) before the second 44°Cheat challenge; •,control cells that received 44°C treatment only. For 42°Ctriggering dose, the survival was normalized to 42°C survival. Near-maximum
tolerance was seen 0 h after heat shock. Tolerance decayed almost completely by24 h.
* 44C.40MN42 C ¿m.eHB -. 44°C40MN
8 10 12 14 16TIME AT 37°C(HRS)
24
Chart 4. Kinetics of the development and decay of thermotolerance in CFU-GM.Bone marrow cells were first heated at 41 °Cfor 1 h, 41 °Cfor 3 h, 41 °Cfor 6 h,42°Cfor 3 h, and 42°Cfor 6 h. The preheated cells were returned to the 37°Cincubator for various times before being heated at 44°Cfor 40 min. The surviving
fraction of the combined heat treatments was always normalized to the killing effectof the first heat treatment. Control cells exposed to 44°Cfor 40 min yielded survivalof 2 to 6 x 10~3. The increase in survival and gradual return to the control value
were used as a measure of the development of thermotolerance.
M VTTRO-tJ VTTRO41°C1 HR - 44°C
20 30 40 0 10
MINUTES AT 44 °C30 40
Charts. Induction of thermotolerance in CFU-GM at 41 °Cfor 1 h. Mice werefirst half-body heated (left) in a 41 °Cwater bath. The preheated mice were returned
to the cages undisturbed for various times before being sacrificed. The marrowcells were then heated m vitro at 44°C.At right, bone marrow cells first heated at41 °Cfor 1 h in vitro. The preheated cells were returned to the 37°Cincubator forvarious times before being heated at 44°C.The times between the 2 treatments
were 0 (A), 2 (•)4 (•),6 (*), 12 (»).24 ( f), and 48 h (9); •control cells thatreceived 44°Ctreatment only. There was no thermotolerance induction at 41 "C
for 1 h when bone marrow cells were heated in vivo or in vitro.
centrifugea, resuspended in a-MEM, and returned to 37°Cin
cubation for 0 to 24 h before the second heat challenge, i.e.,graded doses at 44°Cin vitro. The results, presented in Chart
6, demonstrate that thermotolerance does develop in bone marrow cells after acute heating at 43°Cin vitro. Tolerance reached
its maximum around 6 h after the initial heating and disappearedcompletely 24 h later.
Induction of Thermotolerance in CFU-GM in Vivo. In order
to investigate whether thermotolerance can be induced in bone
CANCER RESEARCH VOL. 45 AUGUST 1985
3845
on May 18, 2020. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
THERMOTOLERANCE AND PROTEIN SYNTHESIS IN MURINE BONE MARROW CELLS
«fr
4O 0 10MWUTES AT 44 °C
Chart 6. Induction of thermotolerance in CFU-GM at 43°C.Bone marrow cellswere first heatedat 43°Cfor 20 min in vivo (left).The preheatedmice were returnedto the cages undisturbed for various times before being sacrificed. The marrowcells were then heated in vitro at 44°C.At right, bone marrow cells were firstheated at 43°Cfor 20 min in wfro. The preheated cells were incubated at 37°Cforvarious times before being heated at 44°C.The times between the 2 treatmentswere 0 (A), 4 (•),6 (»),12 (*), 24 (|), and 48 h (*). Controls were cells thatreceived 44°Ctreatment only. •,control; O, control anesthetized mice; A, 4-hanesthetized mice. The kinetics of thermotolerance induction and decay weresimilar when bone marrow cells were heated in vivo or in vitro.
marrow cells in vivo after a short exposure at 43°C,the following
experiments were performed. The femurs of anesthetized micewere first heated at 43°Cfor 20 min in situ. Some of the mice
were sacrificed immediately while others were returned to theircages for various times up to 24 h before being sacrificed. Themarrow cell suspensions were then prepared from these preheated mice and challenged by graded heat treatments at 44°C
in vitro. The results are presented in Chart 6. Our data showedthat CFU-GM developed resistance in vivo to subsequent heat
treatments. Thermotolerance reached its maximum 4 to 12 hafter the first heat dose and again disappeared completely by 24h (Charts 6 and 7). The kinetics of development of thermotolerance in vivo are similar to that seen in vitro.
Since we found that 41 °C,1-h heating of bone marrow cells
in vitro did not induce thermotolerance, we performed experiments to examine the induction and development of thermotolerance after 41 °C,1-h heating in vivo. Mice were first heatedat 41 °Cby immersion in hot water baths. Some of the mice
were sacrificed immediately, and bone marrow cells were collected. The other mice were returned to their cages undisturbedfor 2,4,6,12,24, and 48 h before being sacrificed. Bone marrowcell suspensions were then prepared. These cell suspensionswere challenged by graded doses at 44°Cin vitro. As shown in
Chart 5, thermotolerance did not develop in vivo either immediately after the 41 °C,1-h heat treatment or at later times (up to
48 h). Thus, similar results were observed whether the experiments were carried out in vitro or in vivo.
Protein Synthesis Profiles in Murine Bone Marrow Cellsafter Heat Shock. Studies on the kinetics of protein synthesisin murine bone marrow cells after a priming heat treatment wereperformed in parallel to the CFU-GM survival studies. A fraction
of the bone marrow cells from the above treatment groups usedfor survival studies were used for the labeling experiments.
The autoradiograph of SDS-polyacrylamide slab gels of
12 18 24 30 36 42"PMEAT 37°C(HRS)
48
Chart 7. Kineticsof the developmentand decay of thermotolerancein CFU-GM.Bone marrow cells were first heated at 43°Cfor 20 min in vivo. The preheatedmice were returned to the cages undisturbed for various times before beingsacrified. The marrow cells were then heated in vitro at 44°Cfor 40 min. Controlcells exposed to 44°Cfor 40 min yielded survival of 2 x 10~3.The increase in
survival and gradual return to the control value were used as a measure of thedevelopmentof thermotolerance.
[^Slmethionine-labeled proteins from control and heated (43°C
for 20 min) bone marrow cells are presented in Fig. 1. Thedensitometer tracings from the autoradiograph are presented inChart 8. It is clearly demonstrated that immediately after a 43°C,20-min heat treatment, the synthesis of M, 68,000 to 70,000
proteins was greatly enhanced when compared to the nonheatedcontrols. The synthesis of these proteins returned to the controlrate within 6 h after heating. In examining the protein patterns ofcontrol and heated bone marrow cells by 2-dimensional gel
electrophoresis, we can identify at least 2 major polypeptides inthe M, 68,000 to 70,000 region in both control and heated groups(Fig. 2, Spots a and b). These proteins were synthesized in theunheated controls, but their rates of synthesis were greatlyenhanced after heat shock. In addition, a third protein in the M,70,000 region which was not present in the control was significantly induced in the heated group (Fig. 2, Spot c).
Fig. 3 shows the kinetics of protein synthesis after prolongedtreatment at 41 and 42°C.The autoradiographs show that the
rate of synthesis of proteins with molecular weights of 68,000 to70,000 (HSP 70) and 87,000 (HSP 87) was significantly enhancedat 0 or 2 h after heat shock. The synthesis of HSPs returned tothe control rate by 24 h. No significant amount of HSP synthesiswas seen when cells were labeled at 0, 2, or 24 h after heatingat 41 °Cfor 6 h. In Table 1, the rates of M, 70,000 HSP synthesis
at various times after heat shock are presented. The data wasobtained by dividing the magnitude of the HSP peak by themagnitude of the actin peak on densitometer tracings obtainedfrom autoradiographs of one-dimensional SDS gels.
DISCUSSION
Several salient features emerge from the study of the thermalresponse of CFU-GM after single or split-dose heat treatments.Consistent with the results of Van Zant ef a/. (13), granulocyte-
CANCER RESEARCH VOL. 45 AUGUST 1985
3846
on May 18, 2020. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
THERMOTOLERANCE AND PROTEIN SYNTHESIS IN MURINE BONE MARROW CELLS
1Fig. 1. Autoradiograph of SDS-polyacrylamide slab gel of [^SJmethionine-la-
beled proteins of murine bone marrow cells after 43°C,20-min heat shock. Theenhancedsynthesis of M, 68,000 to 70,000 and 87,000 HSPs in cells from heatedbone marrow is evident. C,, unheated controls; C2, unheated controls from anesthetized mice; 0, 6, 24, recovery time (h) in vitro after 43°C,20-min heating andbefore labeling; C0, heat-shocked Chinese hamster fibroblasts HA-1 cells forcomparison. Molecular weights (x10~3)are shown at left. A, actin (M, 43,000).
macrophage progenitors were very sensitive to heat as compared to other mammalian cell lines (25). Also, CFU-GM candevelop thermotolerance after prolonged heating at 41-42°C.
This is in contrast with human bone marrow cell colony-forming
units which do not show thermotolerance with prolonged heatingat 41 °C(12). Thermotolerance also develops with acute heattreatment at 43°Cin vitro. In addition, in contrast to other cell
lines where thermotolerance lasts for several days (3, 9, 20, 21),tolerance acquired in CFU-GM disappeared almost completely
within 24 h, regardless of the temperature or duration of theinitial heat treatment. There may be an inherent limitation of CFU-
GM in maintaining this transient resistance to subsequent heatchallenge, but the cause of this phenomenon is not known atpresent. Furthermore, thermotolerance in CFU-GM can be in
duced in vivo. The kinetics of development of thermotolerancein vivo is similar to that in vitro. One interesting finding, in contrastto other cell lines, was that if marrow cells were preheated at41 °Cfor 1 h either in vivo or in vitro, thermotolerance did not
develop either in vivo or in vitro. We want to emphasize that theeffect of heat on the cell differentiation pathway cannot bedetermined by the colony-formation technique.
CONTROL
HEATEDO HR
A HEATED6 HR
87 70 43 87 70
MOLECULAR WEIGHT
43
Charts. Densitometer tracing of autoradiograph shown in Fig. 2. Molecularweights (x10'3) are shown at bottom.A, actin (M,43,000); arrows, M, 70,000 HSP
family and M, 87,000 HSP.
Table 1Rateof M, 70,000 HSPsynthesis
TreatmentControl43°C
for 20 min + 37"C forOh
6h41°Cfor3h
+ 37°Cfor
Oh2h
4h42°C
for 3 h + 37°Cfor
Oh2h4hHSP-actin0.201.54
0.1590.29
0.270.141.86
1.431.11
When the protein synthesis profiles of heat shocked bonemarrow cells were compared to those from unheated controls,the synthesis of M, 68,000 to 70,000 and 87,000 HSP wassignificantly enhanced. The protein profiles were further examined by 2-dimensional gel electrophoresis of total cellular extract
of control and heated bone marrow cells. In the M, 70,000 region,we can identify at least 2 major polypeptides whose expressionis enhanced by heat treatment. Those proteins were also synthesized in smaller amounts in the unheated cells. The heat-
induced enhancement of HSP synthesis lasted only 2 to 4 h afterheat shock and returned to the control value shortly afterward.We want to emphasize the fact that when the cellular proteinswere examined, we were using cells from the total bone marrowpopulation for the analysis. The colony-forming efficiency forCFU-GM is only around 0.2%. Thus, we cannot be certain thatthe heat-induced enhanced HSP synthesis in total marrow pop
ulation accurately represents the profiles of protein synthesis ofgranulocyte-macrophage progenitors. In other words, the enhanced HSP synthesis may not directly reflect the CFU-GM
CANCER RESEARCH VOL. 45 AUGUST 1985
3847
on May 18, 2020. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

THERMOTOLERANCE AND PROTEIN SYNTHESIS IN MURINE BONE MARROW CELLS
survival during the development of thermotolerance.It has been shown that a conditioning whole-body heat dose
(41°Cfor 40 min or 41.8°Cfor 1 h), by itself nonlethal, can give
substantial protection to C3H mice or rats, respectively, againsta subsequent heat treatment (22, 26). The heat-induced protec
tion is transient in nature. It reaches a maximum by 6 to 24 h formice and 48 h for rats following the conditioning dose and decaysby approximately 60% 72 h later for mice and returns to controlvalue by 120 h for rats. Even though these data do not provideinformation on the cause of cell death following whole-body
hyperthermia, they certainly suggest that the response of acomplex organism to heat can be modified by previous heatexposure.
In the present study, we found that heating at 41 °Cfor 60 min
did not induce thermotolerance in CFU-GM either in vivo or invitro. Heating bone marrow cells for longer intervals at 41 °C(41 °Cfor 3 to 6 h) or for short intervals at a higher temperature(43°Cfor 20 min) did induce thermotolerance, but the acquired
tolerance disappeared by 24 h. Thus, the kinetics of the development and decay of thermotolerance observed in the CFU-GM
is very different from that found for animal survival (22). Giventhe disparity between the kinetics of systemic thermotolerance(22) and the thermotolerance of CFU-GM in the same system, itwould appear that bone marrow stem cells (or at least CFU-GM)
are not critical for systemic thermotolerance. In other words,bone marrow most probably is not the critical target which isresponsible for systemic thermal death.
REFERENCES
1. Gemer, E. W. Thermotolerance.In: F. K. Storm (ed.), Hyperthermia¡nCancerTherapy, pp. 141-162. Boston: G. K. Hall & Co., 1983.
2. Gemer, E. W., and Schneider, M. J. Induced thermal resistance in HeLa cells.Nature (Lond.),256; 500-502, 1975.
3. Hahn, G. M., and U, G. C. Thermotolerance and heat shock proteins inmammaliancells. Radiât.Res., 92: 452-457,1982.
4. Henle, K. J., and Dethlefsen, L. A. Heat tractionation and thermotolerance: areview. Cancer Res., 38: 1843-1851,1978.
5. Li, G. C., and Hahn, G. M. A proposed operational model of thermotolerancebased on effects of nutrients and the initial treatment temperature. CancerRes., 40. 4501-4508,1980.
6. Martinez, A. A., Meshorer, A., Meyer, J. L., Hahn, G. M., and Fajardo, L. F.
Thermal sensitivity and thermotolerance in normal porcine tissues. CancerRes., 43: 2072-2075,1983.
7. Raaphorst, G. P., and Azzam, E. I. Hyperthermia and thermal tolerance innormal and ataxia telangiectasia human cell strains. Cancer Res., 43: 2618-2621,1983.
8. Henle, K. J. Thermotolerance in the murine jejunum. J. Nati. Cancer Inst., 68:1033-1036,1982.
9. Hume, S. P., and Marigold,J. C. L. Transient, heat-inducedthermal resistancein the small intestine of mouse. Radiât.Res., 82: 526-535,1980.
10. Law, M. P. Induced thermal resistance in the mouse ear: the relationshipbetween heating time and temperature. Int. J. Radiât.Biol., 35: 481-485,1979.
11. Rice, L. C., Urano, M., and Mäher,J. The kinetics of thermotolerance in themouse foot. Radiât.Res., 89: 291-297,1982.
12. Bromer, R. H., Mitchell, J. B., and Soares, N. Response of human hemato-poietic precursor cells (CFUc)to hyperthermia and radiation. Cancer Res., 42:1261-1265,1982.
13. Van Zant, G., Flentje, D., and Flentje, M. The effect of hyperthermia onhemopoietic progenitor cells of the mouse. Radiât.Res., 95:142-149,1983.
14. Bensaude, 0., Babinet, C., Morange, M., and Jacob, F. Heat shock proteins,first major products of zygotic gene activity in mouse embryo. Nature (Lond.),305:331-333,1983.
15. Savoini, A., Micali, F., Marzari, R., De Cristini, F., and Graziosi, G. Lowvariability of the protein species synthesized by Drosophila melanogasterembryos. WilhelmRoux' Arch. Entwicklungsmech.Org., 790:161-167,1981.
16. Sconzo, G., Roccheri, M. C., Di Carlo, M., Di Bernardo, M. G., and Giudice,G. Synthesis of heat shock proteins in dissociated sea urchin embryoniccells.Cell Differ., 72:317-320,1983.
17. Wittig, S., Hensse, S., Keitel, C., Eisner, C., and Wittig, B. Heat shock geneexpression is regulated during teratocarcinoma cell differentiation and earlyembryonic development. Dev. Biol., 95: 507-514,1983.
18. Landry, J., Bemier, D., Chretien, P., Nicole, L. M., Tanguay, R. M., andMarceau, N. Synthesis and degradation of heat shock proteins during development and decay of thermotolerance.Cancer Res., 42: 2457-2461,1982.
19. Li, G. C., and Werb, Z. Correlation between synthesis of heat shock proteinsand development of thermotolerance in Chinese hamster fibroblasts. Proc.Nati. Acad. Sci. USA, 79: 3219-3222,1982.
20. Li, G. C. Elevated levels of 70,000 dalton heat shock proteins in transientlythermotolerant Chinesehamster fibroblasts and in their stable heat resistantvariants. Int. J. Radiât.Oncol.Biol. Phys., 77:165-177, 1985.
21. Li, G. C., and Mak, J. Y. Induction of heat shock protein synthesis in murinetumors during the developmentof thermotolerance. Cancer Res., 45: 3816-3824, 1985.
22. Li, G. C., Mayer, J. L., Mak, J. Y., and Hahn, G. M. Heat-induced protectionof mice against thermal death. Cancer Res.,43: 5758-5760,1983.
23. Laemmli,U.K. Cleavageof structural proteins during the assemblyof the headof bacteriophageT4. Nature (Lond.),227: 680-685, 1970.
24. O'Farrell, P. H. High resolution twc-<iimensionalelectrophoresisof proteins. J.Biol. Chem., 250: 4007-4021,1975.
25. Raaphorst, G. P., Romano, S. L., Mitchell, J. B., Bedford, J. S., and Dewey,W. C. Intrinsicdifferences in heat and/or x-ray sensitivity of seven mammaliancell lines cultured and treated under identical conditions. Cancer Res., 39:396-401,1979.
26. Weshler, Z., Kapp, D. S., Lord, P. F., and Hayes, T. Developmentand decayof systemic thermotolerance in rats. Cancer Res., 44: 1347-1351, 1984.
CANCER RESEARCH VOL. 45 AUGUST 1985
3848
on May 18, 2020. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
THERMOTOLERANCE AND PROTEIN SYNTHESIS IN MURINE BONE MARROW CELLS
CONTROL HEATED
41C-3h_
41 °C
-6h-
42 °C
i—3h110-87 *-
70«-
A »-•4* -~ —>•—•—
Co 0 2 4 24 0 2 24 0 2 24
Fig. 2. Autoradiographof a 2-dimensionalgel showing the increasedsynthesis of the M, 68,000 and 70,000 HSPs in murinebone marrow cells after 43°Cheat shock.Bone marrow cells from control and heated (43°Cfor 20 min) groups were labeled with ["SJmethionine for 2 h after heating. Total cellular extracts were analyzed by¡soelectricfocusing in the first dimension (from right to left) and SDS-polyacrylamidegel electrophoresis in the second dimension (from top to bottom). A, actin; arrows,M, 70,000 HSP family in heated and unheated cells.
Fig.3. Autoradiographs of SDS-polyacrylamideslab gels of [MS]methionine-labeledmurine bone marrow cells after prolonged heating at 41 or 42°C.The enhancedsynthesis of M, 68,000 and 70,000 HSPs is evident. C0,unheatedcontrols; 0 to 24, recovery time (h) in vitro after 41°Cfor 3 h, 41°Cfor 6 h, or 42°Cfor 3 h and beforelabeling(heating time is indicated at the fop). Molecular weights (xlO'3) are shown at /eft.A, actin (M, 43,000).
CANCER RESEARCH VOL. 45 AUGUST 1985
3849
on May 18, 2020. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1985;45:3843-3849. Cancer Res Nahid F. Mivechi and Gloria C. Li Bone Marrow Cells after Heat ShockThermotolerance and Profile of Protein Synthesis in Murine
Updated version
http://cancerres.aacrjournals.org/content/45/8/3843
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/45/8/3843To request permission to re-use all or part of this article, use this link
on May 18, 2020. © 1985 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from