Embed Size (px)
Citation preview

UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Statistical methods for twins and families
Klaus K. Holst
Department of BiostatisticsUniversity of Copenhagen
November 10, 2013
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Welcome and practicalities
Statistical methods for twins and familiesThis course will describe particular statistical methods and their usefor multivariate data with a focus on how to quantify familialaggregation of specific diseases and quantitative and binomial traits.
- Thomas Scheike [email protected] Klaus Kähler Holst [email protected] Jacob Hjelmborg
http://www.biostat.ku.dk/~kkho/undervisning/family2013
Time plan
915–1030 Lecture & Practicals1045–1200 Lectures & Practicals1200–1300 Lunch1300–1415 Lectures & Practicals1445–1600 Lectures & Practicals
Course outlineNov 11 Continuous traitsNov 12 Binary traitsNov 18 SurvivalNov 19 Competing risks
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Welcome and practicalities
Statistical methods for twins and familiesThis course will describe particular statistical methods and their usefor multivariate data with a focus on how to quantify familialaggregation of specific diseases and quantitative and binomial traits.
- Thomas Scheike [email protected] Klaus Kähler Holst [email protected] Jacob Hjelmborg
http://www.biostat.ku.dk/~kkho/undervisning/family2013
Time plan
915–1030 Lecture & Practicals1045–1200 Lectures & Practicals1200–1300 Lunch1300–1415 Lectures & Practicals1445–1600 Lectures & Practicals
Course outlineNov 11 Continuous traitsNov 12 Binary traitsNov 18 SurvivalNov 19 Competing risks
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Overview
Plan for today
Basic concepts in quantitative geneticsFamily resemblance; kinship coefficientsTwin/Family studiesPath analysis; polygenic modelsMultivariate outcomes; Interactions
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Litterature
1 Methodology for Genetic Studies of Twins and Families byMichael C. Neale, Lon R. Cardon and Hermine H.M. Maes.
2 Genetics and Analysis of Quantitative Traits by Michael Lynchand Bruce Walsh. Sinauer Associates 1998.
3 Mathematical and Statistical Methods for Genetic Analysis byKenneth Lange. Springer-Verlag 2003.
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
LitteratureRijsdijk, F. and Sham, P (2002). Analytic approaches to twindata using structural equation models. Briefings inBioinformatics 3 (2) pp.119-133.
Dongen, J. et al (2012). The continuing value of twin studiesin the omics era. Nature Reviews Genetics 13, 640-653.10.1038/nrg3243
Witte JS, Carlin JB, Hopper JL. (1999). Likelihood-basedapproach to estimating twin concordance for dichotomoustraits. Genet.Epidemiol.16(3):290–304.
Scheike, T and Holst, K and Hjelmborg, J (2013), Estimatingheritability for cause specific mortality based on twin studies.Lifetime Data Analysis. DOI: 10.1007/s10985-013-9244-x.
Scheike, T and Holst, K and Hjelmborg, J (2013). Estimatingtwin concordance for bivariate competing risks twin data.Statistics in Medicine. DOI: 10.1002/sim.6016.
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Practicals
Computer practicals with R and the mets package.
R and the package should be installed on your laptops:
1 install.packages("mets",dependencies=TRUE)
1 library(mets)
Loading ’mets’ version 0.2.5
Other popular software for family studies: Mplus, (open)Mx, stataGLLAMM. . . . However, currently much better methods in mets fordealing with time-to-event endpoint (e.g. disease status in a cohortstudy).
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
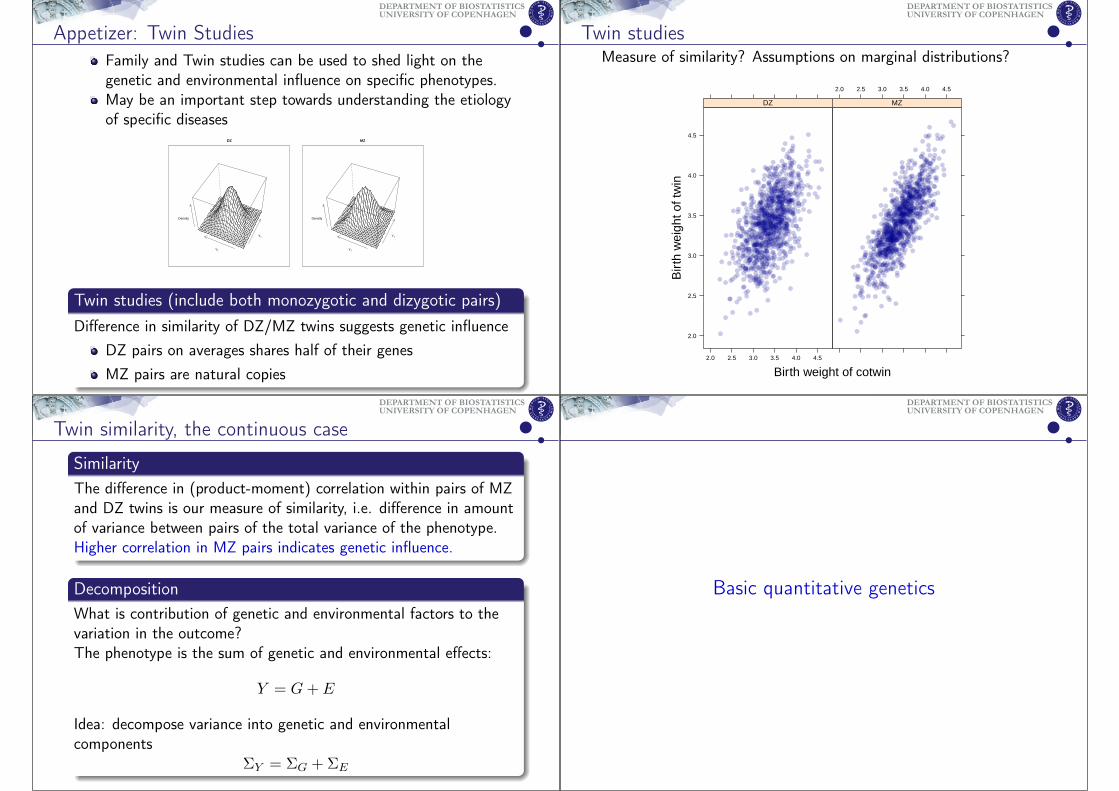
Appetizer: Twin StudiesFamily and Twin studies can be used to shed light on thegenetic and environmental influence on specific phenotypes.May be an important step towards understanding the etiologyof specific diseases
DZ
Y1
Y2
Density
MZ
Y1
Y2
Density
Twin studies (include both monozygotic and dizygotic pairs)Difference in similarity of DZ/MZ twins suggests genetic influence
DZ pairs on averages shares half of their genesMZ pairs are natural copies
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Twin studiesMeasure of similarity? Assumptions on marginal distributions?
Birth weight of cotwin
Bir
th w
eigh
t of t
win
2.0
2.5
3.0
3.5
4.0
4.5
2.0 2.5 3.0 3.5 4.0 4.5
DZ
2.0 2.5 3.0 3.5 4.0 4.5
MZ
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Twin similarity, the continuous case
SimilarityThe difference in (product-moment) correlation within pairs of MZand DZ twins is our measure of similarity, i.e. difference in amountof variance between pairs of the total variance of the phenotype.Higher correlation in MZ pairs indicates genetic influence.
DecompositionWhat is contribution of genetic and environmental factors to thevariation in the outcome?The phenotype is the sum of genetic and environmental effects:
Y = G+ E
Idea: decompose variance into genetic and environmentalcomponents
ΣY = ΣG + ΣE
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Basic quantitative genetics
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Basic quantitative-genetics
The phenotype or disease status is a compounding ofgenes (G) and environment (E)
Important step in understanding the phenotype oretiology of disease is to be able to quantify the role ofG and E (both may be unobserved!).
Research questionIs G (or E) present?To what magnitude?Which particular genotypes or environmentaleffects causes the disease or phenotype.
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Genetic inference
Two different path-ways to genetic inference:
1 Family-study + Linkage analysis −→ Candidategenes
2 Genome-Wide-Association Studies3 Family-based studies are the gold-standard
(hypothesis-driven) for quantification of familialrisk and heritability.
Idea based on familial resemblance: is trait more similaramong related than unrelated individuals?
Continuous trait: tighter correlation in closerrelatives.Disease: Risk of disease greater in case familiescompared to none-cases of general population
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Hierarchical research questions: causes of phenotype
Family aggregation?
Heritability? Environmental effects?
ChromosomesGenesGenotypeProteins/RNA
Identification
interactions?
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Basic quantitative-genetics (Vocabulary)
The phenotype is the observable characteristics ofan individual z = G+ E - quantitative trait.Genetic information encoding resides onchromosomes (strands of DNA). Most cellscontains 46 chromosomes (22 homologous pairs -autosomes - and 2 pairs of sex chromosomes XXand XY).Sequences along the chromosomes that encode forparticular proteins/RNAs: genes. Chromosomallocation: loci.Except for sex chromosomes there are two genesat every locus. The various forms of a gene ateach locus are called alleles.Gene loci that exhibit more than one allele arepolymorphic (then one subject to genetics).

UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Quantitative geneticsCentral goal: quantify association between phenotypic, Z, andgenotypic, G, values.Assume phenotype is sum of the total effects of all loci on the traitand an environmental effect/deviation E
Z = G+ E
In multi-locus trait G may be very complicated function of alleles!Squared correlation
ρ2(G,Z) =Cov(G,Z)2
{Var(G)Var(Z)}Assuming uncorrelated genes and environment we obtain the broadsense heritability:
H2 = ρ2(G,Z) =Var(G)
Var(Z)
OBS: Cov(G,E) 6= 0 may pull ρ(G,Z) in any direction
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Quantitative genetics
Key pointsNot all genes are transmitted
At autosomal loci offsprings gets one maternal and onepaternal allele
F A A A a M
O1 A A A a O2
Family resemblance (and clever design) can be used to estimategenotypic effects
Full siblings get one allele from common father and one allelefrom common mother.Monozygotic twins on average shares all genes
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Quantitative geneticsProblem: In general only the phenotype Z is directly observable.I.e. we can only directly estimate Var(Z).
Solution: Exploit that related individuals carry copies of many ofthe same alleles.
Example: Single locus with additive effectsFather Zf and mother Zm with offspring Zo.Single locus (autosomal genotype/diploid) with additive effect
Zf = gf + g′f + Ef
Zm = gm + g′m + Em
Zo = gf + gm + Eo
Mid-parent value:
Zmp =Zf + Zm
2
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Example: Single locus with additive effectsgm, gf effects of alleles inherited from mother and fatherg′m, g
′f effects of alleles not transmitted from mother and father
Ef , Em, Eo environmental effects
gf gm g′f g′m Ef Em Eo
gf Var(gf )
gm Cov(gm, gf ). . .
g′f...
. . .
g′m...
. . .
Ef...
. . .
Em...
. . .Eo Cov(E0, gf ) · · · · · · · · · · · · · · · Var(Eo)
18 terms in mid-parent-offspring covariance. . .Covariance

UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Example: Single locus with additive effectsAssumptions
1 Random mating, no selectionNo correlation between gene at specific locus in mother andfather. Genes inherited by offspring not correlated with theones that were not transmitted.
2 No gene-environment correlationIndividuals not assorted into environment based on theirgenetic attributes
3 Parents do not transmit their environmental effects to offspring
Cov(Zmp, Zo) = Cov(1
2(gf + gm), gf + gm) =
Var(gf ) + Var(gm)
2
As we only assume additive genetic effects we define
σ2A = Var(gf ) + Var(gm)
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Example: Single locus with additive effectsAssumptions
1 Random mating, no selectionNo correlation between gene at specific locus in mother andfather. Genes inherited by offspring not correlated with theones that were not transmitted.
2 No gene-environment correlationIndividuals not assorted into environment based on theirgenetic attributes
3 Parents do not transmit their environmental effects to offspring
Cov(Zmp, Zo) = Cov(1
2(gf + gm), gf + gm) =
Var(gf ) + Var(gm)
2
As we only assume additive genetic effects we define
σ2A = Var(gf ) + Var(gm)
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Example: Single locus with additive effectsKey-point: genetic component can be extracted from phenotypicresemblance between relativesRegression of of one relatives value on values of other familymember.
E(Zo | Zmp) = µ+ βZmp
Under random mating cov(Zm, Zf ) = 0. Assuming same variancein males and females
Var(Zmp) =1
2σ2Z
Least squares estimator gives the narrow-sense heritability
β =Cov(Zo, Zmp)
Var(Zmp)=σ2Aσ2Z
= h2
h2 := efficiency of response to selection (i.e. h2 = 0 means there isno tendency for offspring to resemble their parents - no evolutionarychange)
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
DominanceAdditive vs non-additive effects
3 possible genotypic values
Genotype
Genotypic value
Aa
(1 + k)d
AA
0
aa
2d
2d = E(Z | G = AA)−E(Z | G = aa)
k =
k ∈ (−1, 1), dominantk = 1, cmplt. dominantk = 0, cmplt. additivek < −1, underdominantk > 1, overdominant
Genotypic variance decomposition
AA aa Aa
AA/Aa aa
Additive
Overdominant
Dominant
AA Aa aa

UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Fishers decomposition of the genotypic value
We consider a phenotype Z genetically determined by a single locus
Let ak denote the kth allele with population frequency pkIn absence of environmental effect an individual with(maternal/paternal) genotype ak/al has constant phenotypevalue (or equivalently, genotypic value) µkl = µlk.We wish to approximate the genotypic value/phenotype valueby additive effect of the alleles
µkl = αk + αl + δkl
Without loss of generality we assume standardized phenotypeE(Z) =
∑k
∑l µklpkpl = 0
Here we assume the locus is in Hardy-Weinberg Equilibrium.
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Hardy-Weinberg Equilibrium (HWE)
Hardy-Weinberg EquilibriumHardy-Weinberg Equilibrium (HWE) describes the relation betweenallele and genotype frequencies under assumptions of
Random matingInfinite population (no drift)No selection/mutation/migration
In the case of a single autosomal locus with alleles A and a, withpopulation frequencies pA and pa = 1− PA, and under the abovegiven assumptions the three possible genotypes, aa, AA, Aa,occurs asymptotically with frequencies p2a, p
2A, 2papA.
AA Aa aa
Genotypic value 0 d(1 + k) 2dFrequency p2A 2pApa p2a
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Hardy-Weinberg Equilibrium (HWE)
~A a
|A AA (p2) Aa (pq)a Aa (pq) aa (q2)
Likelihood Ratio Test for HWEhypothesis may be constructedfrom observed Nij and expectedcell counts N̂ij
Q = −22∑
i=1
2∑
j=i
Nij log(N̂ij/Nij)0.0 0.2 0.4 0.6 0.8 1.0
0.0
0.2
0.4
0.6
0.8
1.0
Allele frequency
Gen
otypefrequen
cy
AAaaAa
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Fishers decomposition of the genotypic value
We consider a phenotype Z genetically determined by a single locus
Let ak denote the kth allele with population frequency pkIn absence of environmental effect an individual with(maternal/paternal) genotype ak/al has constant phenotypevalue (or equivalently, genotypic value) µkl = µlk.We wish to approximate the genotypic value/phenotype valueby additive effect of the alleles
µkl = αk + αl + δkl
Without loss of generality we assume standardized phenotypeE(Z) =
∑k
∑l µklpkpl = 0
Here we assume the locus is in Hardy-Weinberg Equilibrium.
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Fishers decomposition of the genotypic valueLet X1, X2 be independent random variables (maternal/paternalallele). We wish to determine functions h1, h2 that minimizes
arg minf,g E({Z − f(X1)− g(X2)}2)
under condition E(h1(X1)) = E(h2(X2)) = 0.Now rewriting as
Var(Z − h1(X1)− h2(X2)) = Var(Z) + Var(h1(X1)) + Var(h2(X2))
− 2Cov(Z, h1(X1))− 2Cov(Z, h2(X2))
= Var(Z − h1(X1)) + Var(Z − h2(D2))
− Var(Z)
solution is given by
hi(Xi) = E(Z | Xi)
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Fishers decomposition of the genotypic valueWe define the average additive allele effects
αk = E(Z | Xi = ak) =∑
l
µklpl
and orthogonal on that the dominant deviation
δkl = Z − E(Z | X1 = ak)− E(Z | X2 = al)
Genetic variancesAs a consequence a natural definition of additive genetic variance is
σ2A = 2Var[E(Z | Xi)] = 2∑
k
α2kpk
and dominant genetic variance
σ2D = Var[Z − E(Z | X1)]− E(Z | X2)] =∑
k,l
δ2klpkpl
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Fishers decomposition of the genotypic valueAlternative derivation, 2 alleles. . .Wish to decompose genotypic value (assume environmental effecthas mean zero) into additive and residual (dominant dev.)component
µkl = µG + αkl + δkl = µG + αk + αl + δkl
= µG + [α1N1 + α2N2] + δkl
where N1 is the number of A-allele (0,1, or 2). µG is simply themean genotypic value
µG =∑
k,l
µklpkpl
and hence δkl, αkl describes deviations from the mean.Re-parametrization leads to simple linear regression expression:
Gkl = [µG + 2α1] + (α2 − α1)N2 + δkl.
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
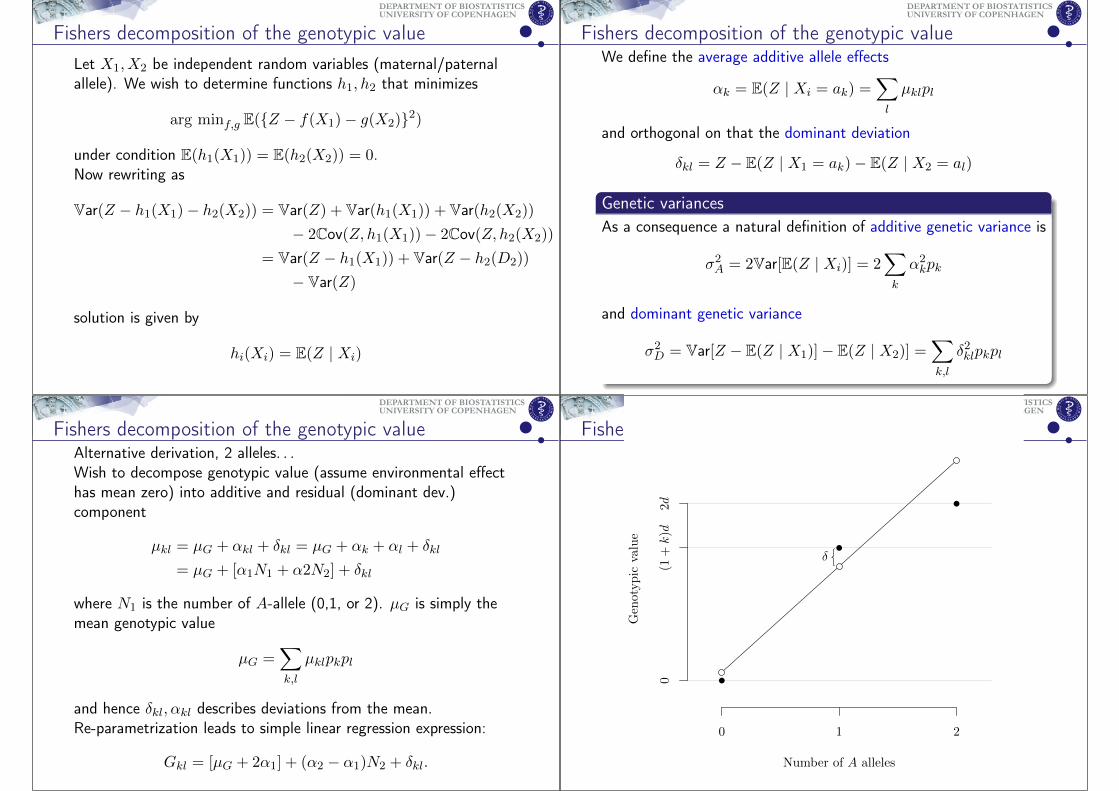
Fishers decomposition of the genotypic value
Number of A alleles
Genotypic
value
0 1 2
0(1
+k)d
2d
δ

UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Fishers decomposition of the genotypic value
Assume N2 ⊥⊥ δij we can use least squares estimator to obtainestimates of intercept and slope β = α2 − α1. The OLS estimator
can be expressed [Lynch&Walsh 98] in gene coefficients Dominance
average effects if allele substitution:
β =Cov(G,N2)
Var(N2)= d[1 + k(p1 − p2)]
Parametrization choice leads to
α1E(N1) + α2E(N2) = 2α1p1 + 2α2p2 = 0 =⇒α2 = p1β and α1 = −p2β
where we have used E(Nk)/2 = pk, p1 + p2 = 1.
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Fishers decomposition of the genotypic value
Indep. between prediction and residual leads to
σ2G = Var(A1 +A2 +D) = σ2A + σ2D
Ak = α11{Xk=a1} + α21{Xk=a2}, D =∑
k,l
δkl1{Xi=ak}1{Xj=al}
Genetic variances, biallelic case
σ2A = 2(α21p1 + α2
2p2)
= 2β2p1p2(p1 + p2) = 2p1p2d2[1 + k(p1 − p2)]2
σ2D = (2p1p2dk)2
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Multiple Loci
If the trait is inherited from n loci in linkage equilibrium andassuming no epistasis, we obtain
Genetic variance n loci
σ2G =
n∑
i=1
σ2Ai+
n∑
i=1
σ2Di
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
LinkageLinkage causes additional covariance terms, e.g.
σ2A = 2
n∑
k=1
α2kpkpl + 2
n∑
k=1
n∑
l=k+1
αkαkDkl
whereDkl = pkl − pkpl
is the linkage disequilibrium coefficient

UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
EpistasisEpistasisIn the multi-loci case interactions between loci may occur. Variancedecomposition can still be obtained by
σ2G =∑
m,n
σ2AnDm
= σ2A + σ2D + σ2AA + σ2AD + σ2DD
+ σ2AAA + σ2AAD + σ2ADD + σ2DDD + · · ·
n number of additive effects and m the number of dominant effects
Cov(Gi, Gj) =∑
m,n
(2Φij)n∆m
7ijσ2AnDm
= 2Φijσ2A + ∆7ijσ
2D
+ 4Φ2ijσ
2AA + ∆2
7ijσ2DD + 2Φij∆7ijσ
2AD + · · ·
Parent-offspring
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Familial resemblance
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Familial resemblance
The fundamental property we exploit is that phenotypic dependenceamong relatives can be used to estimate the variances of thecomponents of genotypic effect. Fisher 1918
To further develop this we need to be able to quantify degree ofrelatedness between individuals. . .
“No, but I would to save two brothers or eight cousins.”
Reply from JBS Haldane when asked if he would give his life tosave a drowning brother1
1Mathematical Models of Social Evolution : A Guide for the Perplexed(2007) by Richard McElreath and Robert Boyd, p. 82.
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Family resemblanceConcept of relatives should be understood in the framework of agiven pedigree structure
2 1 4 3 6 5
8 7 9 10 7 11 12 14 13
15 16 17 18 19
with independence at base
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Familial resemblanceWe define relatedness via concepts of identical alleles. . .
Identical allelesTwo genes are identical by state (i.b.s.) if they represent the sameallele.The genes are identical by descent (i.b.d.) if the alleles are directdescendants of the same gene carried by a common ancestralindividual.
A A A a
A A A a
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
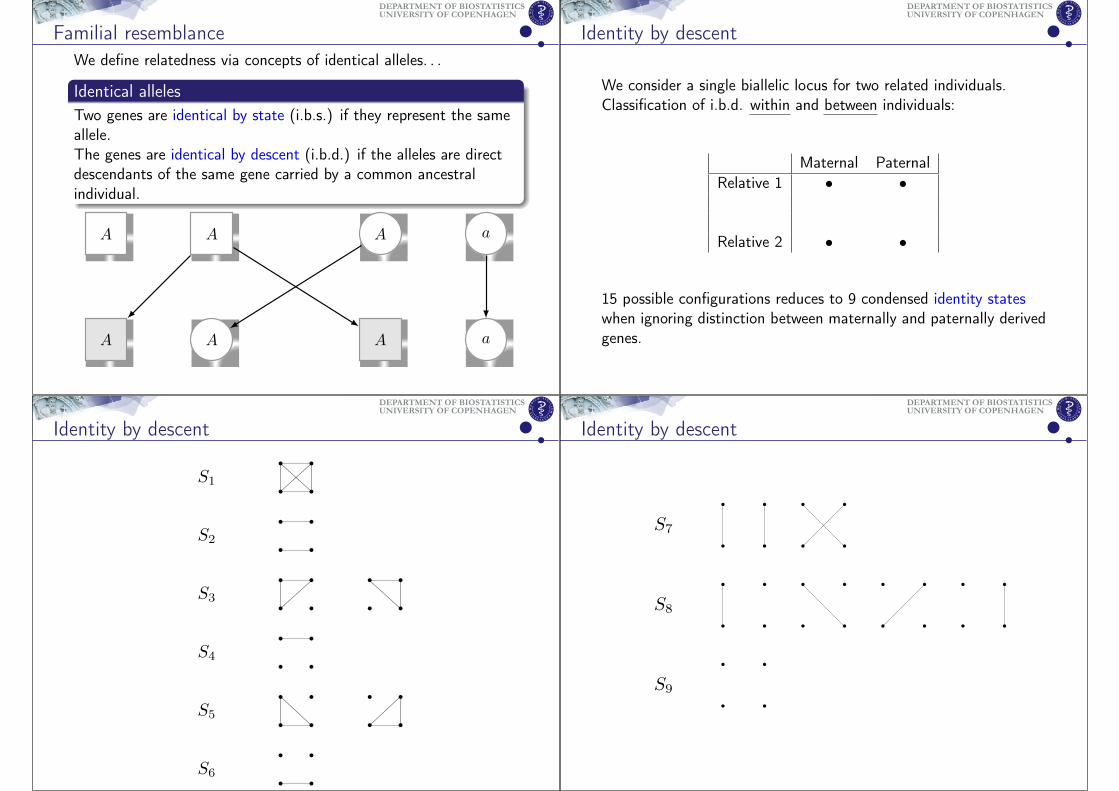
Identity by descent
We consider a single biallelic locus for two related individuals.Classification of i.b.d. within and between individuals:
Maternal PaternalRelative 1 • •
Relative 2 • •
15 possible configurations reduces to 9 condensed identity stateswhen ignoring distinction between maternally and paternally derivedgenes.
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Identity by descent
S1
S2
S3
S4
S5
S6
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Identity by descent
S7
S8
S9

UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Kinship and identity coefficients
Condensed coefficients of identityThe kinship coefficient between relative i and j is the probability,Φij , that an allele selected randomly from i and an allele selectedrandomly from j are i.b.d.The condensed coefficient of identity, is the probability of observingstate k for two relatives i and j
∆k = P(Sk)
Φij = ∆1ij +1
2(∆3ij + ∆5ij + ∆7ij) +
1
4∆8ij
Assuming random mating in a large population, we only have toconsider the last three states (no inbreeding)
∆1 = ∆2 = ∆3 = ∆4 = ∆5 = ∆6 = 0
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Kinship and identity coefficients
Calculate the kinship and condensed coefficient of identity for
1 Parent-offspring2 Full siblings3 Monozygotic twin4 Half-siblings
under the assumption of no inbreeding.
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Kinship and identity coefficients
∆7 ∆8 ∆9 Φ
Parent-offspring 0 1 0 14
Grandparent-grandchild 0 12
12
18
Great grandparent-great grandchild 0 14
34
116
Half siblings 0 12
12
18
Full siblings, DZ twins 14
12
14
14
Uncle(aunt)-nephew(neice) 0 12
12
18
First cousins 0 14
34
116
Double first cousins 116
616
916
18
Second cousins 0 116
1516
164
MZ twins/Clones 1 0 0 12
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Covariance between relativesPhenotype could be decomposed into additive and dominant effects
Genotypic variance decomposition under previously specified assumptions
E(XiXj) = ∆7ij
∑
k
∑
l
(αk + αl + δkl)2pkpl
+ ∆8ij
∑
k
∑
l
∑
m
(αk + αl + δkl)(αk + αm + δkm)pkplpm
+ ∆9ij
∑
k
∑
l
(αk + αl + δkl)(αm + αn + δmn)pkplpmpn
Using∑pk = 1,
∑k αkpk =
∑k δkpk = 0, αk =
∑l µklpl and that
σ2A = 2∑
k α2kpk, σ
2D = 2
∑k δ
2klpkpl we obtain
Decompostion of covariance between non-inbred relatives
Cov(Zi, Zj) = 2Φijσ2A + ∆7ijσ
2D

UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Covariance between relativesProof in the biallelic case:
Zi = µG+A1i+A2i+Di, (A1+A2) ⊥⊥ D,E(A1) = E(A2) = E(D) = 0
The two genes may be i.b.d. with probability Φij in which case
E(AkiAkj) = E(A2k) = 0.5σ2A
If not i.b.d. they are independent
E(AkiAkj) = [E(Ak)]2 = 0
same arguments for other combinations leads to
E[(Aki +Ali)(Akj +Alj)] = 4ΦijE(A2k) = 2Φijσ
2A
Similarly,E(DiDj) = E(D2)
when both genes are i.b.d. occurring with probability ∆7ij and zerootherwise.
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Heritability
Narrow-sense heritability
h2 =σ2A
Var(Z)
Broad-sense heritability
H2 =σ2G
Var(Z)
assuming no-epistastis and gene-environment effects
H2 =σ2A + σ2DVar(Z)
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Dermal Ridges
We will use the mets::dermalridges data set in the practicals
Sarah B. Holt (1952). Genetics of dermal ridges: bilateralasymmetry in finger ridge-counts. Annals of Eugenics 17 (1),pp.211-231. DOI: 10.1111/j.1469-1809.1952.tb02513.x
Data consists of 50 families with ridge counts in left and right handfor mother, father and each child. Additional 18 pairs of MZ twinsare available.
Predominantly additive genetic determined trait
Heritability, gender differences, symmetry. . .
Google Image Search: Dermal Ridges
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
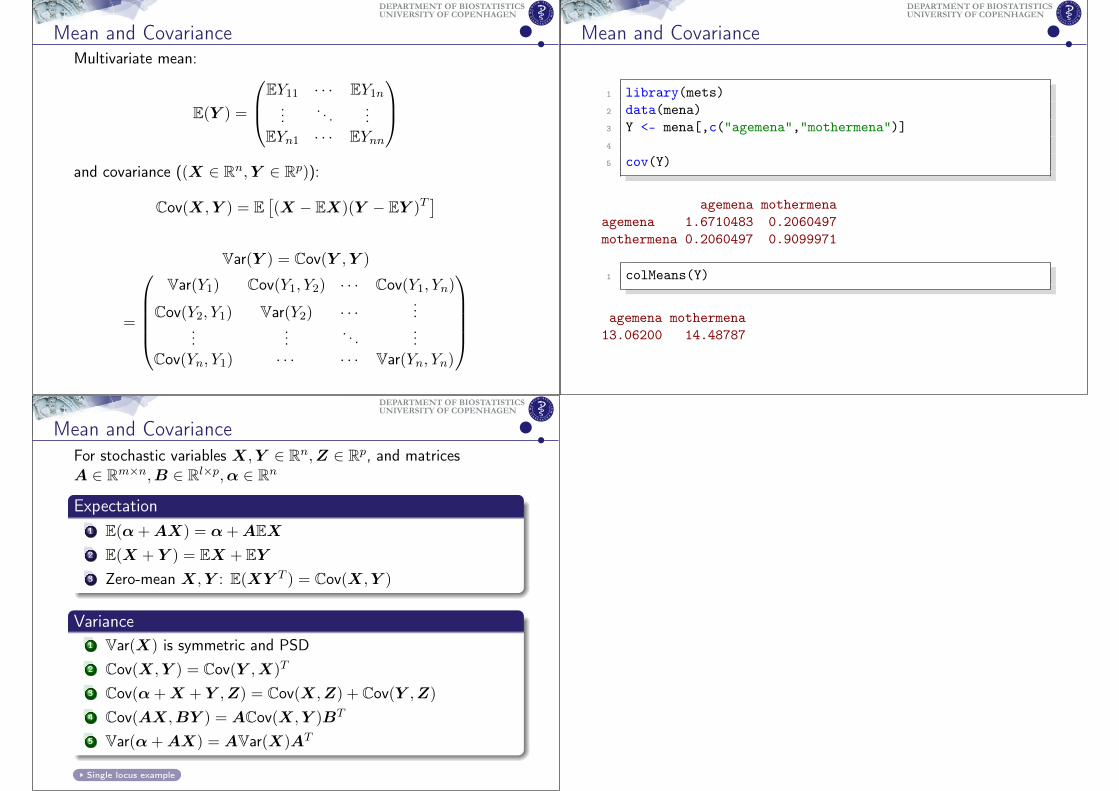
Mean and CovarianceMultivariate mean:
E(Y ) =
EY11 · · · EY1n...
. . ....
EYn1 · · · EYnn
and covariance ((X ∈ Rn,Y ∈ Rp)):
Cov(X,Y ) = E[(X − EX)(Y − EY )T
]
Var(Y ) = Cov(Y ,Y )
=
Var(Y1) Cov(Y1, Y2) · · · Cov(Y1, Yn)
Cov(Y2, Y1) Var(Y2) · · · ......
.... . .
...Cov(Yn, Y1) · · · · · · Var(Yn, Yn)
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Mean and Covariance
1 library(mets)2 data(mena)3 Y <- mena[,c("agemena","mothermena")]4
5 cov(Y)
agemena mothermenaagemena 1.6710483 0.2060497mothermena 0.2060497 0.9099971
1 colMeans(Y)
agemena mothermena13.06200 14.48787
UNIVERSITY OF COPENHAGENDEPARTMENT OF BIOSTATISTICS
Mean and CovarianceFor stochastic variables X,Y ∈ Rn,Z ∈ Rp, and matricesA ∈ Rm×n,B ∈ Rl×p,α ∈ Rn
Expectation1 E(α+AX) = α+AEX2 E(X + Y ) = EX + EY3 Zero-mean X,Y : E(XY T ) = Cov(X,Y )
Variance1 Var(X) is symmetric and PSD2 Cov(X,Y ) = Cov(Y ,X)T
3 Cov(α+X + Y ,Z) = Cov(X,Z) + Cov(Y ,Z)
4 Cov(AX,BY ) = ACov(X,Y )BT
5 Var(α+AX) = AVar(X)AT
Single locus example