Embed Size (px)
Citation preview

1
Title: Francisella like endosymbionts and Rickettsia species in local and imported 1
Hyalomma ticks. 2
Running title: Francisella and Rickettsia in Hyalomma ticks. 3
Tal Azagi1, Eyal Klement1, Gidon Perlman2, Yaniv Lustig3, Kosta Y. Mumcuoglu 4, Dmitry 4
Apanaskevich5, Yuval Gottlieb1* 5
1Koret School of Veterinary Medicine, The Robert H. Smith Faculty of Agriculture, Food 6
and Environment, The Hebrew University of Jerusalem, POB 12, Rehovot, 7610001, 7
Israel. 8
2Jerusalem Bird Observatory, The Society for the Protection of Nature in Israel, POB 9
3557, Jerusalem, Israel. 10
3Central Virology Laboratory, Ministry of Health, Sheba Medical Center, Ramat-Gan, 11
Israel. 12
4Parasitology Unit, Department of Microbiology and Molecular Genetics, The Kuvin 13
Center for the Study of Infectious and Tropical Diseases, The Hebrew University-14
Hadassah Medical School, Jerusalem, Israel. 15
5United States National Tick Collection, the James H. Oliver, Jr. Institute for Coastal Plain 16
Science, Georgia Southern University, Statesboro, GA 30460-8056, USA. 17
* Corresponding author: [email protected] 18
AEM Accepted Manuscript Posted Online 14 July 2017Appl. Environ. Microbiol. doi:10.1128/AEM.01302-17Copyright © 2017 American Society for Microbiology. All Rights Reserved.
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

2
Abstract 19
Hyalomma ticks (Acari: Ixodidae) are hosts for Francisella like endosymbionts (FLE) and 20
may serve as vectors of zoonotic disease agents. This study was aimed to provide initial 21
characterization of the interaction between Hyalomma and FLE and to determine the 22
prevalence of pathogenic Rickettsia in these ticks. Hyalomma marginatum, H. rufipes, 23
H. dromedarii, H. aegyptium and H. excavatum ticks, identified morphologically and 24
molecularly, were collected from different hosts and locations, representing the genus 25
distribution in Israel, as well as from migratory birds. High prevalence of FLE was found 26
in all Hyalomma species (90.6%), as well as efficient maternal transmission of FLE 27
(91.8%), and the localization of FLE in Malpighian tubules, ovaries and salivary glands in 28
H. marginatum. Furthermore, we demonstrated strong co-phylogeny between FLE and 29
their host species. Contrary to FLE, the prevalence of Rickettsia ranged from 2.4% to 30
81.3% and was significantly different between Hyalomma species, with higher 31
prevalence in ticks collected from migratory birds. Using ompA gene sequences, most of 32
the Rickettsia were similar to R. aeschlimannii, while a few were similar to R. africae of 33
the spotted fever group (SFG). Given their zoonotic importance, 249 ticks were tested 34
for Crimean Congo hemorrhagic fever virus infection and all were negative. The results 35
imply that Hyalomma and FLE have obligatory symbiotic interactions, and indicate on a 36
potential SFG Rickettsia zoonosis risk. Further understanding of the possible influence 37
of FLE on Hyalomma development as well as on its infection with Rickettsia pathogens 38
may lead to novel ways for control of tick borne zoonoses. 39
40
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

3
Importance: 41
This study shows that Francisella-like endosymbionts were ubiquitous in Hyalomma, 42
maternally transmitted, and co-speciated with their hosts. These findings imply that the 43
interaction between FLE and Hyalomma is of an obligatory nature. It provides an 44
example of an integrative taxonomy approach to simply differentiate among species 45
infesting the same host and identification of nymphal and larval stages to be used in 46
further studies. In addition, it shows the potential of imported Hyalomma ticks to vector 47
spotted fever group rickettsiae. The information gathered in this study can be further 48
implemented in the development of symbiont based disease control strategies for the 49
benefit of human health. 50
51
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

4
Introduction: 52
Hyalomma (Acari: Ixodidae) hard ticks include approximately 30 species and 53
several subspecies (1) to which taxonomic relationships are constantly being re-54
evaluated (2). Ticks of this genus infest a wide range of vertebrate hosts, and are 55
prevalent on migratory birds, which can be important in the dissemination of emerging 56
Hyalomma-borne zoonoses such as spotted fever group rickettsiae (SFGR) and Crimean 57
Congo hemorrhagic fever virus (CCHFV) (3-7). 58
Other than pathogenic agent infections, Hyalomma carry Francisella-like 59
endosymbionts (FLE) (8-10) that may be obligatory primary symbionts supporting the 60
restricted blood diet as in other ticks and hematophagous insects (11). Several 61
endosymbiotic bacteria in arthropod vectors were shown to interact with pathogens 62
and affect their host’s susceptibility to infection including in tick hosts (12-14). These 63
symbiotic interactions can lead to the development of novel vector-borne disease 64
control (15) as in the case of Dengue virus (www.eliminatedengue.com). The potential 65
symbiosis between FLE bacteria and Hyalomma ticks can serve as a target for novel 66
development of Hyalomma-borne disease control, however, there is no current study 67
testing the nature of FLE in these vectors or its interaction with potential pathogens 68
such as Rickettsia. 69
The genus Rickettsia is composed of several groups, including the Spotted Fever 70
Group (SFGR) found in ticks (16). SFGR include at least 15 species described as the 71
causative agents of rickettsioses mostly transmitted to humans by ticks of the genera 72
Dermacentor, Amblyomma, Rhipicephalus and Hyalomma (17). Israel is an endemic 73
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

5
region for Israeli spotted fever caused by R. conorii israelensis where fatal cases have 74
been reported (18, 19). SFGR such as R. felis, R. sibirica mongolitimonae, R. africae, R. 75
massiliae and R. aeschlimannii, were also found in various ticks species in Israel 76
including in Hyalomma (20-22). However, the distribution of the SFGR species among 77
the various Hyalomma species and those introduced to Israel was never investigated. 78
Hyalomma ticks are the main vector of CCHFV, the most widespread tick-borne 79
zoonosis worldwide that can cause a fatal hemorrhagic fever in humans (23). Despite 80
the existence of Hyalomma ticks in Israel, human and animal infection was never 81
demonstrated in the country. CCHFV has been isolated from H. marginatum and other 82
Hyalomma ticks collected from migratory birds in Morocco and Turkey (3, 4), and the 83
ability of H. rufipes ticks to become infected from birds inoculated with the virus has 84
been experimentally proven (24). Since Israel is situated at one of the largest migration 85
routes in the world, where about 4% of the estimated 5 billion birds migrating from 86
Africa to the Western Palearctic fly through every spring (25), it is suitable to identify 87
which Hyalomma species are introduced into the country and determine their infection 88
status, as was performed in other countries where yearly migratory events occur (26-89
28). 90
This study is aimed to characterize the Hyalomma species distribution on various 91
hosts in Israel and on birds migrating through Israel, to determine the role of FLE as a 92
potential primary symbiont in these ticks, and the prevalence of SFG Rickettsia and 93
CCHFV in endemic and imported Hyalomma species to Israel. The knowledge gained in 94
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

6
this study is expected to enable assessment of the zoonotic risk imposed by these ticks 95
and to open novel possibilities for their control. 96
97
Materials and Methods: 98
Tick collection: 99
Hyalomma ticks actively search for their hosts, therefore rather than flagging 100
vegetation, we removed the ticks directly from hosts in representative locations in 101
Israel. Ticks were collected from horses, camels, tortoises (Testudo graeca), and 102
migratory birds during 2011-2015 (Fig. 1). Ticks were morphologically identified using 103
taxonomic keys (1, 29-31) and were preserved in vials containing 100% ethanol and kept 104
at -20oC until further processing, except for ticks from tortoises which were preserved in 105
dry vials at -80oC until further use. 106
Nucleic acid extraction 107
Prior to nucleic acid extraction, each tick was washed in 3% sodium hypochlorite 108
and then with 70% ethanol and finally twice in sterile Dulbecco's phosphate buffer 109
saline solution (Biological Industries, Israel) in order to reduce external contaminants. 110
DNA and RNA were then extracted using either the RTP DNA/RNA virus mini kit or the 111
RTP pathogen kit (Stratec, Germany) according to the manufacturer protocols with few 112
modifications: following sample disinfection ticks were cut to four pieces on a sterile 113
Petri dish using a sterile scalpel blade and placed in the provided extraction tube with 114
lysis buffer, fully engorged ticks were extracted in two separate tubes. After lysis in the 115
pre-filled extraction tube (with lyophilized lysis components), the tubes were 116
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

7
centrifuged and the supernatant was used for the remainder of the protocol. DNA and 117
RNA were quantified in a NanoDrop ND1000 spectrophotometer (NanoDrop 118
Technologies, Denmark) at A260/A280 and the samples were stored at -80oC until 119
further use. 120
For DNA extraction of individual eggs, these were placed separately in a 0.2 ml 121
micro-tube and crushed against the bottom of the tube with a sterile needle. Each 122
micro-tube was then filled with 12 µl of extraction buffer containing 1 mg/ml proteinase 123
K (Sigma Aldrich, USA) 0.01 m NaCl, 0.1 m EDTA, 0.01 m Tris-HCl (pH 8.0), 0.5% Nonidet 124
P-40 substitute (Sigma Aldrich, USA) (32), after a spin down of the tubes, they were 125
incubated for 15 minutes at 65oC followed by 10 minutes inactivation at 95oC. 126
Standard PCR and real time RT-PCR 127
All standard PCR amplifications were performed on a T-gradient 180 basic 128
thermocycler (Biometra, Germany) according to published protocols with suitable 129
primers (Table 1) in a final volume of 25µl, containing 2X GoTaq Green Master Mix 130
(Promega, USA), 0.2µM forward and reverse primers, gDNA template and nuclease free 131
water. Negative controls with no template were included in all reactions. Real time RT-132
PCR for CCHFV detection was performed according to (33). 133
Sequencing and analysis 134
PCR products were purified with ExoSAP (New England Biolabs, USA). Each 135
reaction contained 0.1µl Exonuclease I, 0.2µl Antartic phosphatase, 1µl Exonuclease I 136
buffer, 1 µl Antartic phosphatase buffer, 7.7µl nuclease free water and 10µl PCR 137
product, the reaction was incubated for 30 minutes at 37°C followed by 5 minutes 138
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

8
inactivation at 70oC. The purified samples were Sanger sequenced (Macrogen, Holland) 139
from both directions. All sequences were manually edited and aligned using BioEdit 140
version 7.2.5 (http://jwbrown.mbio.ncsu.edu/BioEdit/bioedit.html) (34). The sequences 141
were then compared to the GenBank database using the BLAST algorithm to confirm 142
their identity. 143
Restriction fragment length polymorphism (RFLP) analyses: 144
The species H. marginatum, H. excavatum and H. rufipes, which can be found on 145
the same host, were chosen for RFLP discrimination. After morphological identification 146
of all samples, and based on the Cytochrome c oxidase I (COI) consensus sequences 147
obtained from representative samples as described above, restriction enzymes were 148
chosen in silico using Webcutter 2.0 software (35) based on a distinct cleavage site (hpaI 149
for H. marginatum, resulting in 650bp fragment; draI for H. excavatum, resulting in 150
550pb fragment, and hindIII for H. rufipes, resulting in 600bp fragment). The restriction 151
reactions were performed on 25µl PCR template added directly to a mixture containing 152
1µl restriction enzyme, 3µl NEBuffer 2.1 (New England Biolabs, USA) for hindIII or 153
CutSmart Buffer (New England Biolabs, USA) for hpaI and draI, and 1µl Diethyl 154
pryrocarbonate treated double distilled water (DEPC-DDW). The mixture was incubated 155
for 60 min at 37°C, restricted versus unrestricted amplicons were visualized in a 1.5% 156
agarose gel. All analyses were performed on previously morphologically identified and 157
sequenced samples. Since the restricted fragments of each species differ in size, it 158
allowed for the simultaneous identification of samples with all enzymes in the same 159
reaction: 25µl PCR template, 0.66µl enzyme each, 4µl CutSmart Buffer and 9µl DEPC 160
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

9
DDW followed by 2 hours incubation at 37°C and 20 minutes of heat inactivation at 161
80oC. 162
Maternal transmission assay: 163
Seven fully engorged H. marginatum females were collected live from horses 164
(location 6, Fig.1), brought to the laboratory and kept separately in a chamber with 165
suitable conditions for laying eggs (36). The relative humidity of 75-85% was maintained 166
using Monopotassium phosphate (KH2PO4) diluted in DEPC DDW in a sterile Petri dish 167
under 23-25oC. During the following 21 days, five of the 7 ticks laid eggs and all were 168
transferred to -80oC until further processing. 169
Phylogenetic analyses: 170
For Hyalomma COI, PCR products from 10 H. marginatum, 10 H. rufipes, 10 H. 171
dromedarii, 9 H. aegyptium, 4 H. excavatum and 75 unidentified Hyalomma ticks from 172
migratory birds, were sequenced as described above. Consensus sequences were 173
manually determined for each species except for H. aegyptium which segregated into 174
two different consensus sequences. 175
For FLE, 4 sequences from each tick species, obtained as described, and partial 176
16S rRNA Francisella sequences: FLE of Amblyomma maculatum (AC#:LNCT01000002), 177
Francisella tularensis (AC#: CP017155.1) and F. philomiragia (AC#: NR_114925.1) were 178
used for analyses with MEGA7 Molecular Evolutionary Genetics Analysis version 7.0 for 179
bigger datasets (37). Sequence alignment was performed with MUSCLE (multiple 180
sequence comparison by log-expectation) (38) using default parameters, and 181
phylogenetic trees were inferred based on the Maximum Likelihood method with 1000 182
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

10
bootstraps. The distance between sequences was calculated based on the number of 183
base differences per site from between sequences using pairwise comparisons with 184
1000 bootstraps, this simple method was applied since most distance methods are 185
comparable and give similar estimates (39-41). 186
Co-phylogeny was tested with the tick COI tree and the FLE tree using the 187
Procrustes Approach to Co-phylogeny – PACo (42) in R version 3.3.1 (https://www.R-188
project.org/) with the vegan and ape packages (43, 44). 189
Fluorescence in-situ hybridization (FISH) 190
To target FLE in tick organs, the probe /Cy3/FLE was designed based on the FLE 191
16S rRNA partial gene sequence (Table 1). The specificity for FLE of the probe was 192
tested in silico using the ARB probe design tool against the SILVA database (45). 193
Individual female ticks were disinfected as described above and dissected inside a 194
droplet of sterile Dulbecco's PBS as described in (46). Salivary glands, ovaries and 195
Malpighian tubules were removed and placed in a droplet of sterile PBS. Pools of organs 196
were transferred into cell strainers (Corning, USA) and placed in 6 well plates filled with 197
FAA (5% Acetic acid, 5% formaldehyde, 90% ethanol) in a vacuum chamber for 1 hour. 198
After 24 hours at room temperature, the organs were dehydrated in increasing ethanol 199
concentrations: 50% for 10 minutes and then: 50%, 80% and 100% for 15 min each step, 200
finally the organs were left to dry at room temperature for 10 minutes. Staining was 201
performed on separate pools of salivary glands, ovaries and Malpighian tubules as 202
described before (46) with a few modifications: cell strainers were used to transfer the 203
organs from buffer to buffer. The probes /Cy3/FLE and /Cy5/Eub338 were diluted in the 204
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

11
hybridization buffer (20mM Tris-HCl pH 8.0, 0.9 M NaCl, 35% (v/v) formamide, 0.01% 205
(w/v) Sodium dodecyl sulfate-SDS) to a concentration of 6.25 ng/µl. Later, DAPI, 4', 6'-206
diamidino-2-phenylindole, (Invitrogen, USA), staining was applied to label DNA. After 207
the procedure, samples were mounted on a Superfrost plus slide (Baor-Naor LTD, Israel) 208
with a drop of CitiFluorTM (EMS, USA). The whole procedure was also performed on 209
organs using only the probe /Cy5/Eub338, as well as no-probe, and antisense-EUB Probe 210
(Table 1) for fluorescence control. Samples were viewed under Leica’s CTR65000 211
confocal laser scanning microscope (Leica Microsystems, Germany). 212
Statistical analyses: 213
Pearson Chi-square tests were performed to ascertain differences between 214
groups, the analyses were performed using WinPEPI version 11.65 (47). 215
Results: 216
Ticks sampled locally 217
Out of 130 adult ticks collected (during 2012) from 29 camels, 99.3% were 218
Hyalomma dromedarii and 0.7% H. excavatum. Out of 151 adult ticks collected from 56 219
horses (during 2011 and 2014) 47.6% were H. marginatum and 52.4% H. excavatum. All 220
28 ticks collected from seven tortoises were identified as H. aegyptium. 221
Ticks sampled from migratory birds 222
One hundred and fifty six ticks were collected from 89 migratory birds over the 223
spring migration of 2014 and 2015 of which 150 ticks were of the genus Hyalomma 224
(96.1%, H. rufipes in 2014 and H. marginatum complex in 2015), and the remaining 225
samples (3.9%) were five Amblyomma spp. ticks and one Rhipicephalus guilhoni 226
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

12
(collected in 2014). Out of 81 ticks collected in 2014, one was identified as an adult tick, 227
12 were larvae and 68 were nymphs. Four of the nymphs were found with their molted 228
skins, meaning molting occurred while attached to the bird host. In 2015, 75 ticks were 229
collected, 14 were larvae and 61 were nymphs of which 5 were found with their molted 230
skins. Most ticks were at least semi-engorged. 231
Of all birds examined for ticks in 2014 and 2015, 19 (0.29%) and 4 (0.1%) 232
respectively, were found infested in the Jerusalem Bird Observatory (JBO), while 27 233
(0.47%) and 39 (0.59%) were found infested in Eilat. The average infestation prevalence 234
(0.22%, 23 out of 10,232 in JBO and 0.54%, 66 out of 12,190 in Eilat) differs significantly 235
between stations (p<0.001 ). Ticks at larval and nymphal stages were found on 17 bird 236
species, mostly of the order Passeriformes but also from the order Accipitriformes 237
(Supp. Table 1). The species Sylvia atricapilla (Eurasian blackcap) was the most 238
commonly infested bird (33.7%) in both years and prevalent in both locations, followed 239
by Iduna pallida (Eastern olivaceous warbler) and Sylvia curruca (Lesser whitethroat) 240
which amount to 16.8% and 13.4% of all infested birds respectively. 241
Hyalomma molecular identification by RFLP 242
In order to ease the identification of ticks found on the same host or those in 243
early developmental stages, we used a restriction enzyme analysis. The enzymatic 244
reactions correlated to the morphological taxonomic identification: The hpaI restriction 245
analysis was successful for 93% of H. marginatum samples, the enzyme draI successfully 246
cut all samples morphologically identified as H. excavatum and the hindIII restriction 247
analysis was effective for 96.6% of H. rufipes samples (Supp. Fig. 1). 248
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

13
Ticks of immature stages, collected from birds in 2015 were morphologically 249
identified with uncertainty regarding their species (H. marginatum or H. turanicum). 250
Using the restriction analysis 88% samples were cleaved and resulted in two fragment 251
patterns: 11 samples showed a 650bp fragment (hpaI - H. marginatum) and 55 samples 252
showed a 600bp fragment (hindIII- H. rufipes). 253
Hyalomma Phylogeny 254
The COI consensus sequences were compared to the GenBank and BOLD 255
databases (http://www.boldsystems.org/). The sequences of H. dromedarii, H. 256
marginatum, H. excavatum and H. rufipes matched sequences corresponding to the 257
same species with high similarities (>97%). The sequence H. aegyptium 1 was similar to 258
a sequence from Romania of the same species (GenBank AC#: JX394192.1, BOLD AC# 259
ACH4580) with a 99.5% identity match. The sequence H. aegyptium 2 was 99.04% 260
similar to an H. aegyptium sequence from Belgium (GenBank AC#: AF132821.1, BOLD 261
AC# AAX2350). According to a pairwise analysis of evolutionary divergence, the most 262
closely related sequences were H. aegyptium1 and H. aegyptium2 with 3.88% 263
divergence, followed by H. marginatum and H. rufipes with 4.07% divergence, the 264
average distance between species was 10.29% with a standard error of 0.01 (Table 2). 265
All sequences were deposited in Genbank: H. dromedarii AC# KY548842, H. 266
excavatum AC# KY548843, H. marginatum AC# KY548844, H. rufipes AC# KY548845, H. 267
aegyptium 1 AC# KY548846, H. aegyptium 2 AC# KY548847. A phylogenetic tree was 268
constructed and used for further analysis as described below. 269
Prevalence and taxonomy of FLE: 270
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

14
Of the 257 Hyalomma ticks screened for the presence of FLE (Supp. Table 2), 271
90.66% samples were found positive. The prevalence varied between tick species with 272
84.6% positive H. marginatum, 90.5% H. excavatum, 89.8% H. dromedarii, 100% H. 273
aegyptium and 90.4% H. rufipes. The difference in prevalence between species and life 274
stages was not statistically significant (p=0.996, p=0.931; respectively). Statistically 275
significant difference in prevalence between males and females was found only for H. 276
dromedarii (p<0.001) with 64.7% prevalence in females and 96.77% prevalence in males. 277
The consensus sequences obtained from H. marginatum, H. excavatum, H. 278
dromedarii, H. aegyptium and H. rufipes ranged from 688-702 bp. The FLE sequences of 279
H. marginatum, H. excavatum and H. aegyptium were closely related to the FLE of H. 280
truncatum (AC# JF290387.1) with 99% identity. The FLE sequence of H. dromedarii was 281
also similar to the H. truncatum sequence and to that of an Ornithodoros porcinus 282
symbiont (AC# AB001522.1) with 99% identity. There were no FLE sequences 283
homologue to the H. rufipes sequence in GenBank. All sequences were deposited in 284
GenBank (AC# KY469285-KY469289). 285
Co-phylogeny of FLE with Hyalomma hosts: 286
The phylogenetic tree inferred from FLE and other Francisella sequences (Fig. 287
2A), and the tree inferred from Hyalomma COI sequences (Fig. 2B) are shown with links 288
between corresponding host and FLE branches. The FLE sequences are clustered 289
together, resembling a monophyletic group separated from pathogenic Francisella 290
sequences (bootstrap value 97%). However, the inner splits in this monophyletic group 291
are poorly supported (53%, 54%) with little divergence between FLE sequences (0-1.6%). 292
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

15
The FLE sequences of H. marginatum and H. rufipes appear to be the closest since their 293
sequences were identical. 294
The FLE and Hyalomma phylogenies show obvious similarities (Fig. 2), the global 295
goodness-of-fit statistic obtained from the PACo analysis was significant (m 2 XY = 296
0.007355749, P =0.00094), which shows the apparent dependence of the symbiont 297
phylogeny on the host phylogeny is unlikely to be incidental. The agreement between 298
them can be visualized with a Procrustes superimposition plot (Supp Fig. 2). The bar plot 299
obtained through the PACo analysis (Fig. 3) provides a representation of the 300
contribution of each host-symbiont association to the global co-speciation fit, measured 301
by means of jackknife estimation of their respective squared residuals (e2i ) at a 95% 302
confidence interval. Links that represent a smaller fraction of the sum of squares are 303
more likely to be the result of co-speciation, meaning that the lower bars, showing the 304
link between H. rufipes and its FLE, H. aegyptium1 and 2 and their FLE, and the link 305
between H. dromedarii and its FLE or even H. marginatum and its FLE which are very 306
close to the median, represent links where co-evolution most likely occurred. The link 307
between H. excavatum and its FLE appear to contribute less to the co-evolutionary 308
congruence between the phylogenetic trees. 309
Maternal transmission of FLE: 310
All five egg laying H. marginatum females were positive for FLE. The samples 311
obtained from the DNA extraction of 110 single eggs (22 laid by each female: Fe1-Fe5) 312
were screened for FLE. From Fe1, 21/22 (95.4%) eggs were positive for FLE, as were 313
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

16
21/22 (95.4%) from Fe2, 19/22 (83.3%) from Fe3 and 20/22 (90.9%) from Fe4 and Fe5, 314
with an overall vertical transmission efficiency of 91.81%. 315
Localization of FLE in tick organs: 316
High densities of FLE can be seen in Malpighian tubules (Fig. 4A) with clusters 317
apparently surrounding the cell nuclei (Fig. 4B). FLE was also observed in the poles of 318
the oocytes (Fig. 4D,E) and strong signals of the FLE probe were recorded scattered on 319
salivary gland acini (Fig. 4G), where the bacteria appears to surround cell nuclei (Fig. 320
4H). Assays with no probes were performed as controls in order to rule out auto-321
fluorescence as the source of recorded signals (Fig. 4C, F, I). Images of antisense-EUB 322
control, FLE and of EUB338 probes can be viewed separately in supplementary figures 3-323
6. 324
Prevalence of SFGR and CCHFV: 325
Out of 280 ticks screened (Supp. Table 3), 114 (40.7%) were found positive for 326
SFGR. Prevalence of SFGR infection was distributed among the various Hyalomma 327
species as follows: 54.5% H. marginatum, 10% H. excavatum, 2.4% H. dromedarii, 19.2% 328
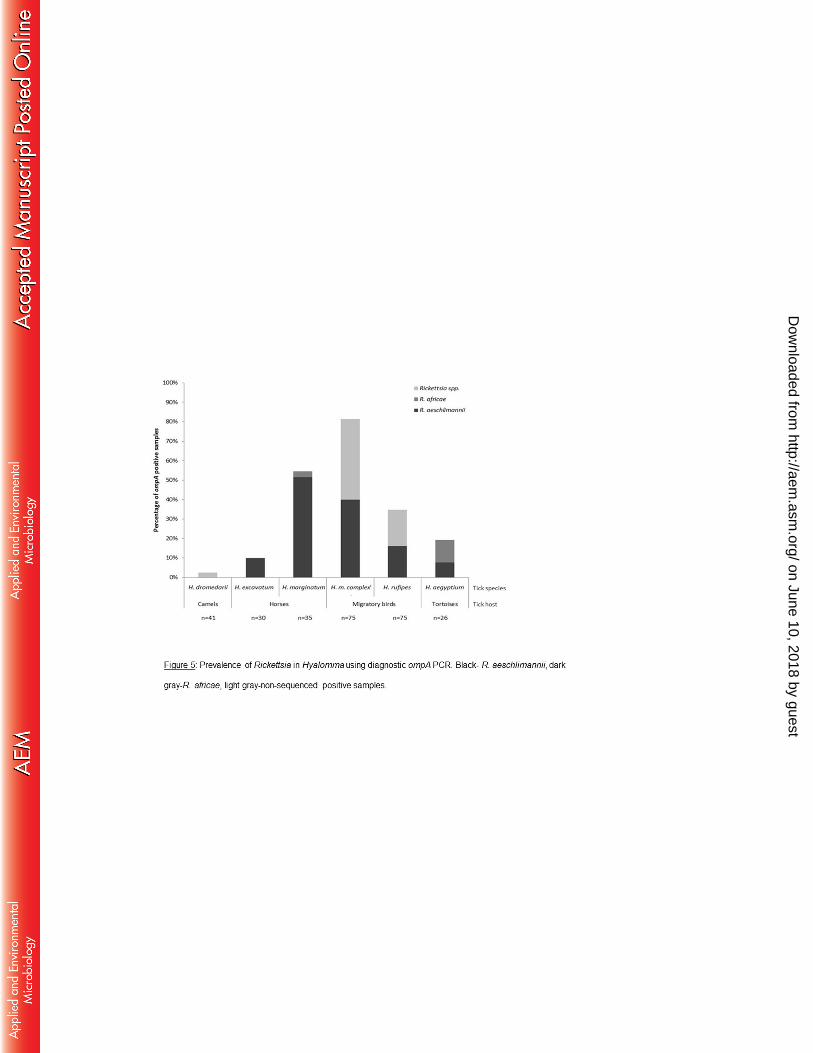
H. aegyptium, 34.6% H. rufipes and 81.3% H. marginatum complex were positive (Fig. 5). 329
The difference in the prevalence of Rickettsia was statistically significant among tick 330
species (Chi-square test DF=5; p<0.001) and tick hosts (Chi-square test DF=3; p<0.001), 331
but not between locations. Sixty eight of the positive samples were randomly chosen 332
and Sanger sequenced, these were identified as Spotted Fever Group Rickettsia (SFGR). 333
Of the 68 samples, 64 were identified as R. aeschlimannii (AC#: HQ335157.1) with 100% 334
identity match, and 4 as R. africae, (AC#: HQ335137.1) with 99% identity match (Fig. 5). 335
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

17
None of the 249 tick samples collected from migratory birds, camels and 336
tortoises in various regions in Israel (Supp. Table 4) were found positive for CCHFV. 337
Maternal transmission of SFGR: 338
Three of the five H. marginatum females screened for Rickettsia ompA gene 339
were found positive and maternal transmission of SFGR was demonstrated by screening 340
22 individual eggs from each positive female. The overall efficiency of maternal 341
transmission was 95.3%: one of the females transmitted Rickettsia to all of the offspring 342
analyzed while the other two females transmitted the bacterium to 21 (95.45%) and 19 343
(90.47%) of the eggs. In order to ascertain the veracity of the positive results from egg 344
samples, 4 of them were sequenced and identified as SFGR of the species R. 345
aeschlimannii, with a 99% identity match using BLAST algorithm in GenBank. 346
347
Discussion: 348
In this study we determined the phylogeny, prevalence and potential symbiont-349
host relationships of FLE in five different Hyalomma species either found on local hosts 350
or imported via migratory birds. Our results support the assumption that FLE are 351
obligatory symbionts of their Hyalomma hosts: 352
First, FLE was found to be highly prevalent across all Hyalomma species 353
screened, including H. marginatum whereas other studies reported lower FLE 354
prevalence (9, 48). This difference, however, might be explained by method sensitivity, 355
or the possibility that FLE have different importance in Hyalomma ticks from different 356
areas. In H. dromedarii we found higher FLE prevalence in males, which is in contrast to 357
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

18
other studies that reported prevalence of symbionts to be typically higher in females (8, 358
49-51). Sampling effort (17 females and 62 males) could explain this deviation, and a 359
quantitative approach may ascertain the accuracy of this observation. 360
Second, the 16S rRNA based phylogenetic analysis demonstrated that the FLE 361
from Hyalomma ticks are clustered together as a monophyletic group separated from 362
pathogenic Francisella in agreement with other FLE phylogeny analyses (9, 48, 52, 53), 363
and the phylogenetic congruence between FLE and their Hyalomma hosts is significant. 364
The sequences of FLE from H. marginatum and H. rufipes were identical as was shown 365
previously (10, 48). These two tick species, which are closely related, may also harbor 366
the same endosymbionts. Also, the distance between FLE from H. aegyptium and H. 367
marginatum corresponds to that shown in studies from Hungary, Ethiopia and Yemen 368
(9, 10, 48), indicating that FLE strains are comparable in different locations. Congruence 369
between host and symbiont phylogenies has been shown for many obligatory symbiosis 370
systems where the association is required to support the dietary needs of the host (54), 371
and was also shown for Rhipicephalus and Coxiella-like endosymbiont lineages (55). The 372
FLE sequences of Hyalomma are close to an FLE sequence from Amblyomma maculatum 373
which was hypothesized to have recently evolved from pathogenic Francisella (56). 374
These findings suggest that FLE in some or all Hyalomma species evolved in the same 375
manner, however, the congruency with their hosts’ phylogeny suggest the transition to 376
endosymbiosis occurred earlier in Hyalomma than in A. maculatum. In contrast, other 377
studies on FLE phylogeny concluded that the symbiont-host association is relatively 378
recent, although no specific analyses were applied to test the hypothesis in these 379
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

19
studies (10, 53), excluding Dermacentor reticulatus and its FLE that formed a separate 380
phylogenetic subgroup (57). As in the current study phylogenies of FLE and their 381
Hyalomma hosts were analyzed on a relatively large scale, the results of other studies 382
are not necessarily contradictory. 383
Third, we demonstrated efficient maternal transmission of FLE to offspring 384
(91.8%). In Dermacentor and Amblyomma ticks, transovarial transmission was tested in 385
two pools of 10 larvae from 16 ticks and revealed a high transmission rate of 95–100% 386
(58). Transmission may be even higher in Hyalomma, since individual eggs were tested 387
and detection of low bacterial concentrations by standard PCR might not be optimal. In 388
addition, we demonstrated FLE in the tick ovaries, suggesting a specific mechanism to 389
ensure transmission. High densities of FLE were found in the Malpighian tubules, organs 390
which have been shown to harbor main endosymbionts in ticks (8, 46). Interestingly, 391
ubiquitous sporadic clusters of FLE were found in the salivary glands which raise 392
questions regarding the pathogenic potential of FLE (8). Nevertheless, FLE and other 393
bacterial endosymbionts have been known to colonize salivary glands and their 394
presence in this organ does not ensure transmission to vertebrate hosts (59). Similar 395
observations have been made for other hard ticks and their obligatory symbionts (55, 396
46), suggesting a potential nutritional role of the symbionts in supplying B vitamins 397
missing in the tick blood meal (11, 60). 398
In this study, we also developed a RLFP identification method based on the 399
mitochondrial gene COI in order to discriminate 3 Hyalomma species (H. marginatum, H. 400
excavatum and H. rufipes) that can be found on the same host (horse), and screened the 401
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

20
ticks for the major zoonotic agents they may transmit, Rickettsia and CCHFV. For each 402
morphologically identified Hyalomma species, a single consensus sequence was 403
generated, except for H. aegyptium which is divided into two branches with a relatively 404
high intraspecific divergence (3.88%) (58). This difference could represent geographic 405
distribution, as was shown for the African H. rufipes (61), however, the sample size in 406
our study is too low to conclude the same. While the average inter-specific sequence 407
divergence between species in this study was 10.29%, only 4.07% divergence was found 408
between H. marginatum and H. rufipes. These two species were considered part of the 409
Hyalomma (Euhyalomma) marginatum Koch complex until recent years (2), and perhaps 410
the low divergence reflects recent species separation. The RFLP analysis was also useful 411
for the identification of immature stages of Hyalomma ticks, which is a limiting factor 412
for identification of ticks collected from migratory birds (3, 4, 27, 62). The main species 413
found on migratory birds were H. rufipes and H. marginatum. The first is common 414
through sub-Saharan Africa, while the second is common in North Africa, as well as 415
Southern Europe, and Asia Minor to western Iran (2). The bird species with higher tick 416
infestation in this study were S. atricapilla, I. pallida and S. curruca (33.7%, 16.8% and 417
13.4% respectively) known to feed on the ground and forage in low scrub (63), thus are 418
exposed to ectoparasites (64, 65). 419
The role of migratory birds in the dissemination of disease by importing 420
pathogen infected ticks over a broad geographic range has become evident (66). Thus 421
we tested the ticks for CCHFV and SFGR. While no virus was detected, as was the case in 422
similar surveys (27, 62, 64), Rickettsia was prevalent in H. rufipes and H. marginatum 423
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

21
complex ticks from birds (34.6% and 81.3% respectively). The sequenced SFGR samples 424
were identified as R. aeschlimannii, a prevalent tick borne SFGR in the African continent, 425
previously detected in ticks from migratory birds arriving to Europe (67, 68). Rickettsia 426
aeschlimannii is more common in northern Africa and its high prevalence might point to 427
the wintering location of the tick infested birds (67). The most commonly tick infested 428
bird species in this study, Sylvia atricapilla, may occasionally winter in the area (63), 429
suggesting that the majority of the ticks inspected originated from northern Africa. In 430
local hosts, the prevalence of R. aeschlimannii varied. It was low in H. dromedarii ticks 431
from camels (2.4%) and higher in H. excavatum and H. marginatum ticks from horses 432
(10% and 54.5%, respectively). This is in agreement with the low prevalence found 433
previously in H. dromedarii ticks from camels and in contrast to the prevalence in H. 434
excavatum ticks from a camel and a horse (0.38% and 3.92%, respectively) (20). 435
Rickettsia aeschlimannii was also detected in H. aegyptium ticks from tortoises, as has 436
been shown previously (69). The ability of R. aeschlimannii to be transovarially 437
transmitted from H. marginatum ticks to their offspring (70) with high efficacy as shown 438
here in individual eggs, strengthens the notion that H. marginatum may act as reservoirs 439
for the disease in Israel. Rickettsia africae was detected in a few local ticks from 440
northern Israel. Previous studies showed R. africae in Hyalomma ticks collected in 441
southern Israel, and the authors hypothesized that it was imported to Israel from tick 442
infesting camels arriving from Egypt (20, 22). Thus our findings may indicate that R. 443
africae propagated since its original introduction to the country via Hyalomma ticks. 444
The clinical symptoms for most spotted fever rickettsioses are nonspecific and may be 445
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

22
misdiagnosed (19). Spotted fever reported cases in Israel are currently minor, 6 and 7 446
reported cases in 2016 and 2017, respectively with no specific rickettsial agent diagnosis 447
(http://www.health.gov.il). Assuming this study presents the actual infection prevalence 448
of both H. marginatum and H. excavatum ticks from horses with which humans have 449
frequent and prolonged interactions, Hyalomma and SFGR control and surveillance 450
strategies should be considered. 451
Understanding the relationship between ticks and their bacterial endosymbionts 452
is a first step towards symbiont-based control plans (15). Non-pathogenic bacteria in 453
ticks may interact with pathogens and affect host’s susceptibility to infection (8), thus 454
the obligatory nature of the symbiosis between Hyalomma and FLE established in this 455
study and finding of high prevalence SFGR, can be further implemented in zoonotic 456
disease risk assessment as well as in the development of control strategies for the 457
benefit of human health. 458
459
Acknowledgements 460
We are grateful to Sharon Tirosh, Gabi Kleinerman, Jessica Rose, Noam Weiss 461
and Yael Lander for their help in tick collection. We thank Michael Ben-Yosef for 462
assistance with FISH analyses. We also thank Dr. Roger Hewson (Microbiology Services, 463
Health Protection Agency, UK) who provided the CCFHV positive control and together 464
with Prof. Iain Hay (University at Buffalo, The State University of New York) contributed 465
to CCHVF discussion. This study was funded by The Dutch Friends of the Hebrew 466
University (NVHU) grant to YG and EK. 467
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

23
Bibliography 468
1. Nava S, Guglielmone AA, Mangold AJ. 2009. An overview of systematics and 469
evolution of ticks. Front Biosci Landmark Ed 14:2857–2877. 470
2. Apanaskevich DA, Horak IG. 2008. The genus Hyalomma Koch, 1844: V. re-471
evaluation of the taxonomic rank of taxa comprising the H. (Euhyalomma) 472
marginatum Koch complex of species (Acari: Ixodidae) with redescription of all 473
parasitic stages and notes on biology. Int J Acarol 34:13–42. 474
3. Palomar AM, Portillo A, Santibáñez P, Mazuelas D, Arizaga J, Crespo A, Gutiérrez Ó, 475
Cuadrado JF, Oteo JA. 2013. Crimean-Congo hemorrhagic fever virus in ticks from 476
migratory birds, Morocco. Emerg Infect Dis 19:260–263. 477
4. Leblebicioglu H, Eroglu C, Erciyas-Yavuz K, Hokelek M, Acici M, Yilmaz H. 2014. Role 478
of migratory birds in spreading Crimean-Congo hemorrhagic fever, Turkey. Emerg 479
Infect Dis 20:1331–4. 480
5. Ioannou I, Chochlakis D, Kasinis N, Anayiotos P, Lyssandrou A, Papadopoulos B, 481
Tselentis Y, Psaroulaki A. 2009. Carriage of Rickettsia spp., Coxiella burnetii and 482
Anaplasma spp. by endemic and migratory wild birds and their ectoparasites in 483
Cyprus. Clin Microbiol Infect 15 Suppl 2:158–160. 484
6. Hildebrandt A, Franke J, Meier F, Sachse S, Dorn W, Straube E. 2010. The potential 485
role of migratory birds in transmission cycles of Babesia spp., Anaplasma 486
phagocytophilum, and Rickettsia spp. Ticks Tick Borne Dis 1:105–107. 487
7. Movila A, Reye AL, Dubinina H V, Tolstenkov O, Toderas I, Hübschen JM, Muller CP, 488
Alekseev AN. 2011. Detection of Babesia sp. EU1 and members of spotted fever 489
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

24
group rickettsiae in ticks collected from migratory birds at Curonian Spit, North-490
Western Russia. Vector Borne Zoonotic Dis 11:89–91. 491
8. Ahantarig A, Trinachartvanit W, Baimai V, Grubhoffer L. 2013. Hard ticks and their 492
bacterial endosymbionts (or would be pathogens). Folia Microbiol (Praha) 58:419–493
428. 494
9. Ivanov IN, Mitkova N, Reye AL, Hübschen JM, Vatcheva-Dobrevska RS, Dobreva EG, 495
Kantardjiev TV, Muller CP. 2011. Detection of new Francisella-like tick 496
endosymbionts in Hyalomma spp. and Rhipicephalus spp. (Acari: Ixodidae) from 497
Bulgaria. Appl Environ Microbiol 77:5562–5565. 498
10. Szigeti A, Kreizinger Z, Hornok S, Abichu G, Gyuranecz M. 2014. Detection of 499
Francisella-like endosymbiont in Hyalomma rufipes from Ethiopia. Ticks Tick Borne 500
Dis 5:818-820. 501
11. Manzano-Marín A, Oceguera-Figueroa A, Latorre A, Jiménez-García LF, Moya A. 502
2015. Solving a bloody mess: B-vitamin independent metabolic convergence 503
among gammaproteobacterial obligate endosymbionts from blood-feeding 504
arthropods and the leech Haementeria officinalis. Genome Biol Evol 7:2871-2884. 505
12. Gottlieb Y, Zchori-Fein E, Mozes-Daube N, Kontsedalov S, Skaljac M, Brumin M, 506
Sobol I, Czosnek H, Vavre F, Fleury F, Ghanim M. 2010. The transmission efficiency 507
of tomato yellow leaf curl virus by the whitefly Bemisia tabaci is correlated with 508
the presence of a specific symbiotic bacterium species. J Virol 18:9310-7. 509
13. Weiss B, Aksoy S. 2011. Microbiome influences on insect host vector competence. 510
Trends Parasitol 27:514–522. 511
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

25
14. Gall CA, Reif KE, Scoles GA, Mason KL, Mousel M, Noh SM, Brayton KA. 2016. The 512
bacterial microbiome of Dermacentor andersoni ticks influences pathogen 513
susceptibility. ISME J 10:1846–1855. 514
15. Zindel R, Gottlieb Y, Aebi A. 2011. Arthropod symbioses: a neglected parameter in 515
pest- and disease-control programmes. J Appl Ecol 48:864–872. 516
16. Weinert LA, Werren JH, Aebi A, Stone GN, Jiggins FM. 2009. Evolution and 517
diversity of Rickettsia Bacteria. BMC Biol. 7:6. 518
17. Parola P, Paddock CD, Raoult D. 2005. Tick-borne rickettsioses around the world: 519
emerging diseases challenging old concepts. Clin Microbiol Rev 18:719–56. 520
18. Aharonowitz G, Koton S, Segal S, Anis E, Green MS. 1999. Epidemiological 521
Characteristics of Spotted Fever in Israel over 26 Years. Clin Infect Dis 29:1321–522
1322. 523
19. Weinberger M, Keysary A, Sandbank J, Zaidenstein R, Itzhaki A, Strenger C, Leitner 524
M, Paddock CD, Eremeeva ME. 2008. Fatal Rickettsia conorii subsp. israelensis 525
infection, Israel. Emerg Infect Dis 14:821–824. 526
20. Kleinerman G, Baneth G, Mumcuoglu KY, van Straten M, Berlin D, Apanaskevich 527
DA, Abdeen Z, Nasereddin A, Harrus S. 2013. Molecular detection of Rickettsia 528
africae, Rickettsia aeschlimannii, and Rickettsia sibirica mongolitimonae in camels 529
and Hyalomma spp. ticks from Israel. Vector borne zoonotic Dis 13:851–856. 530
21. Harrus S, Perlman-Avrahami A, Mumcuoglu KY, Morick D, Baneth G. 2011. 531
Molecular detection of Rickettsia massiliae, Rickettsia sibirica mongolitimonae and 532
Rickettsia conorii israelensis in ticks from Israel. Clin Microbiol Infect 17:176–180. 533
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

26
22. Waner T, Keysary A, Eremeeva ME, Din AB, Mumcuoglu KY, King R, Atiya-Nasagi Y. 534
2014. Rickettsia africae and Candidatus Rickettsia barbariae in ticks in Israel. Am J 535
Trop Med Hyg 90:920–922. 536
23. Bente DA, Forrester NL, Watts DM, McAuley AJ, Whitehouse CA, Bray M. 2013. 537
Crimean-Congo hemorrhagic fever: history, epidemiology, pathogenesis, clinical 538
syndrome and genetic diversity. Antiviral Res 100:159–189. 539
24. Zeller HG, Cornet JP, Camicas JL. 1994. Experimental transmission of Crimean-540
Congo hemorrhagic fever virus by West African wild ground-feeding birds to 541
Hyalomma marginatum rufipes ticks. Am J Trop Med Hyg 50:676–681. 542
25. Frumkin R, Pinshow B, Kleinhaus S. 1995. A review of bird migration over Israel. J 543
Ornithol 136:127–147. 544
26. Hoogstraal H, Kaiser MN, Traylor MA, Guindy E, Gaber S. 1963. Ticks (Ixodidae) on 545
birds migrating from Europe and Asia to Africa 1959-61. Bull World Health Organ 546
28:235–262. 547
27. Mancini F, Toma L, Ciervo A, Di Luca M, Faggioni G, Lista F, Rezza G. 2013. Virus 548
investigation in ticks from migratory birds in Italy. New Microbiol 36:433–4. 549
28. Jameson LJ, Morgan PJ, Medlock JM, Watola G, Vaux AGC. 2012. Importation of 550
Hyalomma marginatum, vector of Crimean-Congo haemorrhagic fever virus, into 551
the United Kingdom by migratory birds. Ticks Tick Borne Dis 3:95–99. 552
29. Apanaskevich DA. 2003. Differentiation of subspecies of the polymorphic species 553
Hyalomma marginatum (Acari: Ixodidae) based on immature stages. Parazitologiia 554
6:462-472. 555
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

27
30. Apanaskevich DA, Horak IG. 2005. The genus Hyalomma Koch, 1844. II. Taxonomic 556
status of H. (Euhyalomma) anatolicum Koch, 1844 and H. (E.) excavatum Koch, 557
1844 (Acari: Ixodidae) with redescriptions of all stages. Acarina 13:181-197. 558
31. Apanaskevich DA, Schuster AL, Horak IG. 2008. The genus Hyalomma: VII. 559
Redescription of all parasitic stages of H. (Euhyalomma) dromedarii and H. (E.) 560
schulzei (Acari: Ixodidae). J Med Entomol 5:817-831. 561
32. Black WC, Nancy MD, Gary JP, James RN, Jennifer MP. 1992. Use of the random 562
amplified polymorphic DNA polymerase chain reaction (RAPD-PCR) to detect DNA 563
polymorphisms in aphids (Homoptera: Aphididae). Bull Entomol Res 82:151. 564
33. Wölfel R, Paweska JT, Petersen N, Grobbelaar AA, Leman PA, Hewson R, Georges-565
Courbot M-C, Papa A, Günther S, Drosten C. 2007. Virus detection and monitoring 566
of viral load in Crimean-Congo hemorrhagic fever virus patients. Emerg Infect Dis 567
13:1097–1100. 568
34. Hall T. 1999. BioEdit: a user-friendly biological sequence alignment editor and 569
analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 41:95-98. 570
35. Maarek YS, Jacovi M, Shtalhaim M, Ur S, Zernik D, Ben-Shaul IZ. 1997. WebCutter: a 571
system for dynamic and tailorable site mapping. Comput Networks ISDN Syst 572
29:1269–1279. 573
36. Sweatman GK. 1968. Temperature and humidity effects on the oviposition of 574
Hyalomma aegyptium ticks of different engorgement weights. J Med Entomol 575
5:429–439. 576
37. Kumar S, Stecher G, Tamura K. 2016. MEGA7: molecular evolutionary genetics 577
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

28
analysis version 7.0 for bigger datasets. Mol Biol Evol 33:1870–1874. 578
38. Edgar RC. 2004. MUSCLE: multiple sequence alignment with high accuracy and high 579
throughput. Nucleic Acids Res 32:1792–1797. 580
39. Jin L, Nei M. 1990. Limitations of the evolutionary parsimony method of 581
phylogenetic analysis. Mol Biol Evol 7:82–102. 582
40. Tateno Y, Takezaki N, Nei M. 1994. Relative efficiencies of the maximum-likelihood, 583
neighbor-joining, and maximum-parsimony methods when substitution rate varies 584
with site. Mol Biol Evol 11:261–277. 585
41. Mangold AJ, Bargues MD, Mas-Coma S. 1998. 18S rRNA gene sequences and 586
phylogenetic relationships of European hard-tick species (Acari: Ixodidae). 587
Parasitol Res 84:31–37. 588
42. Balbuena JA, Míguez-Lozano R, Blasco-Costa I. 2013. PACo: A novel procrustes 589
application to cophylogenetic analysis. PLoS One 8:e61048. 590
43. Oksanen J, Blanchet F, Kindt R, Legendre P, Minchin P, O’Hara R, Simpson G, 591
Solymos P, Stevens M, Wagner H. 2013. Vegan: Community Ecology Package. R 592
Package Version. 2.0-10. URL: cran.r-project.org/package=vegan. 593
44. Paradis E, Claude J, Strimmer K. 2004. APE: Analyses of Phylogenetics and Evolution 594
in R language. Bioinformatics 20:289–90. 595
45. Pruesse E, Quast C, Knittel K, Fuchs BM, Ludwig W, Peplies J, Glockner FO. 2007. 596
SILVA: a comprehensive online resource for quality checked and aligned ribosomal 597
RNA sequence data compatible with ARB. Nucleic Acids Res 35:7188–7196. 598
46. Lalzar I, Friedmann Y, Gottlieb Y. 2014. Tissue tropism and vertical transmission of 599
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

29
Coxiella in Rhipicephalus sanguineus and Rhipicephalus turanicus ticks. Environ 600
Microbiol 16:3657–3668. 601
47. Abramson JH. 2011. WINPEPI updated: computer programs for epidemiologists, 602
and their teaching potential. Epidemiol Perspect Innov 8:1. 603
48. Montagna M, Chouaia B, Pella F, Mariconti M, Pistone D, Fasola M, Epis S. 2012. 604
Screening for bacterial DNA in the hard tick Hyalomma marginatum (Ixodidae) 605
from Socotra Island (Yemen): detection of Francisella-like endosymbiont. J 606
Entomol Acarol Res 44:e13. 607
49. Lalzar I, Harrus S, Mumcuoglu KY, Gottlieb Y. 2012. Composition and seasonal 608
variation of Rhipicephalus turanicus and Rhipicephalus sanguineus bacterial 609
communities. Appl Environ Microbiol 78:4110–4116. 610
50. Wójcik-Fatla A, Zając V, Sawczyn A, Cisak E, Sroka J, Dutkiewicz J. 2015. Occurrence 611
of Francisella spp. in Dermacentor reticulatus and Ixodes ricinus ticks collected in 612
eastern Poland. Ticks Tick Borne Dis 6:253–257. 613
51. Dergousoff SJ, Chilton NB. 2012. Association of different genetic types of 614
Francisella-like organisms with the Rocky Mountain wood tick (Dermacentor 615
andersoni) and the American dog tick (Dermacentor variabilis) in localities near 616
their northern distributional limits. Appl Environ Microbiol 78:965–971. 617
52. Machado-Ferreira E, Piesman J, Zeidner NS, Soares CAG. 2009. Francisella-like 618
endosymbiont DNA and Francisella tularensis virulence-related genes in Brazilian 619
ticks (Acari: Ixodidae). J Med Entomol 46:369–374. 620
53. Scoles GA. 2004. Phylogenetic analysis of the Francisella-like endosymbionts of 621
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

30
Dermacentor ticks. J Med Entomol 41:277–286. 622
54. Zchori-Fein E, Bourtzis K. (Eds) 2011. Manipulative tenants: bacteria associated 623
with arthropods. CRC Press, Boca Raton. 624
55. Duron O, Binetruy F, Noël V, Cremaschi J, McCoy KD, Arnathau C, Plantard O, 625
Goolsby J, Pérez de León AA, Heylen DJA, Van Oosten AR, Gottlieb Y, Baneth G, 626
Guglielmone AA, Estrada-Peña A, Opara MN, Zenner L, Vavre F, Chevillon C. 2017. 627
Evolutionary changes in symbiont community structure in ticks. Mol Ecol, In press. 628
56. Gerhart JG, Moses AS, Raghavan R. 2016. A Francisella-like endosymbiont in the 629
Gulf Coast tick evolved from a mammalian pathogen. Sci Rep 6:33670. 630
57. Michelet L, Bonnet S, Madani N, Moutailler S. 2013. Discriminating Francisella 631
tularensis and Francisella-like endosymbionts in Dermacentor reticulatus ticks: 632
Evaluation of current molecular techniques. Vet Microbiol 163:399–403. 633
58. Baldridge GD, Scoles GA, Burkhardt NY, Schloeder B, Kurtti TJ, Munderloh UG. 634
2009. Transovarial transmission of Francisella-like endosymbionts and Anaplasma 635
phagocytophilum variants in Dermacentor albipictus (Acari: Ixodidae). J Med 636
Entomol 46:625–632. 637
59. Klyachko O, Stein BD, Grindle N, Clay K, Fuqua C. 2007. Localization and 638
visualization of a Coxiella-type symbiont within the lone star tick, Amblyomma 639
americanum. Appl Environ Microbiol 73:6584–6594. 640
60. Gottlieb Y, Lalzar I, Klasson L. 2015. Distinctive genome reduction rates revealed by 641
genomic analyses of two Coxiella-like endosymbionts in ticks. Genome Biol Evol 642
7:1779–1796. 643
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

31
61. Cangi N, Horak IG, Apanaskevich DA, Matthee S, das Neves LCBG, Estrada-Peña A, 644
Matthee CA. 2013. The influence of interspecific competition and host preference 645
on the phylogeography of two African ixodid tick species. PLoS One 8:e76930. 646
62. England ME, Phipps P, Medlock JM, Atkinson PM, Atkinson B, Hewson R, Gale P. 647
2016. Hyalomma ticks on northward migrating birds in southern Spain: 648
Implications for the risk of entry of Crimean-Congo haemorrhagic fever virus to 649
Great Britain. J Vector Ecol 41:128–134. 650
63. del Hoyo J, Elliott A, Sargatal J, Cabot J. 1992. Handbook of the birds of the world. 651
Lynx Edicions. 652
64. Klaus C, Gethmann J, Hoffmann B, Ziegler U, Heller M, Beer M. 2016. Tick 653
infestation in birds and prevalence of pathogens in ticks collected from different 654
places in Germany. Parasitol Res 115:2729–2740. 655
65. Pietzsch ME, Mitchell R, Jameson LJ, Morgan C, Medlock JM, Collins D, Chamberlain 656
JC, Gould EA, Hewson R, Taylor MA, Leach S. 2008. Preliminary evaluation of exotic 657
tick species and exotic pathogens imported on migratory birds into the British 658
Isles. Vet Parasitol 155:328–332. 659
66. Hasle G. 2013. Transport of ixodid ticks and tick-borne pathogens by migratory 660
birds. Front Cell Infect Microbiol 3:48. 661
67. Wallménius K, Barboutis C, Fransson T, Jaenson TGT, Lindgren P-E, Nyström F, 662
Olsen B, Salaneck E, Nilsson K. 2014. Spotted fever Rickettsia species in Hyalomma 663
and Ixodes ticks infesting migratory birds in the European Mediterranean area. 664
Parasit Vectors 7:318. 665
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

32
68. Toma L, Mancini F, Di Luca M, Cecere JG, Bianchi R, Khoury C, Quarchioni E, Manzia 666
F, Rezza G, Ciervo A. 2014. Detection of microbial agents in ticks collected from 667
migratory birds in central Italy. Vector Borne Zoonotic Dis 14:199–205. 668
69. Bitam I, Kernif T, Harrat Z, Parola P, Raoult D. 2009. First detection of Rickettsia 669
aeschlimannii in Hyalomma aegyptium from Algeria. Clin Microbiol Infect 15:253–670
254. 671
70. Matsumoto K, Parola P, Brouqui P, Raoult D. 2004. Rickettsia aeschlimannii in 672
Hyalomma ticks from Corsica. Eur J Clin Microbiol Infect Dis 23:732–734 673
71. Atkinson B, Chamberlain J, Logue CH, Cook N, Bruce, C, Dowall SD, Hewson R. 2012. 674
Development of a real-time RT-PCR assay for the detection of Crimean-Congo 675
hemorrhagic fever virus. Vector Borne Zoonotic Dis 12:786–793 676
72. Fournier, P.-E., Roux, V., and Raoult, D. (1998). Phylogenetic analysis of spotted 677
fever group rickettsiae by study of the outer surface protein rOmpA. Int. J. Syst. 678
Bacteriol. 48, 839–849. 679
73. Lv J, Wu S, Zhang Y, Chen Y, Feng C, Yuan X, Jia G, Deng J, Wang C, Wang Q, et al. 680
2014. Assessment of four DNA fragments (COI , 16S rDNA , ITS2 , 12S rDNA) for 681
species identification of the Ixodida (Acari : Ixodida ). Parasit Vectors 7:9 682
74. Amann RI, Binder BJ, Olson RJ, Chisholm, SW, Devereux R, Stahl, DA. 1990. 683
Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for 684
analyzing mixed microbial populations. Appl Environ Microbiol 56:1919-1925. 685
686
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

33
Tables 687
Table 1: List of primers and probes used in this study. 688
Primer name Platform Specificity Sequence (5′-3′) Product
Size (bp)
Reference and notes (*)
F- CCHF S1 RT-PCR CCHF virus S
segment
TCTCAAAGAAACACGTGCC 122 71
R- CCHF S122 CCTTTTTGAACTCTTCAAACC
Probe (FAM)ACTCAAGGKAACACTGTGGGCGTA
AG(BHQ1)
NC-Fran16S-F PCR FLE 16S rRNA
gene
CAACATTCTGGACCGAT 373 52
*Used for screening, 40
cycles for egg samples and
35 cycles for others
NC-Fran16S-R TGCGGGACTTAACCCAACAT
NC-Fran16S-F FLE 16S rRNA
gene +
universal
primer 16S
rRNA
CAACATTCTGGACCGAT 728 52
*Used for phylogeny with
R1494 with same conditions
as screening the PCR
R1494 CTACGGCTACCTTGTTACGA 49
Rr190.70F Rickettsial
ompA-
encoding
gene
ATGGCGAATATTTCTCCAAAA 630 72
*Extension for 1 min. at 72°C Rr190.701R GTTCCGTTAATGGCAGCATCT
TY-J-1449 5’ region of
cytochrome
oxidase I
(COI)
AATTTACAGTTTATCGCCT 860 73
*Used to obtain consensus
sequences
C1-N-2312 CATACAATAAAGCCTAATA
HCO2064 5’ region of
cytochrome
oxidase I
(COI)
GGTGGGCTCATACAATAAATCC 860 73
*Used for restriction
analyses HCO1215 GCCATTTTACCGCGATGA
/Cy5/Eub338 FISH Bacterial
16rRNA
GCTGCCTCCCGTAGGAGT 74
/Cy3/anti-
sense Eub338
No bacteria
detection
ACT CCT ACG GGA GGC AGC 74
/Cy3/FLE FLE 16rRNA ACTCCAACAGCTAGTACTCA This study
689
690
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

34
Table 2: Matrix of mt cytochrome oxidase I gene sequence divergence between species 691
based on the percentage of unshared nucleotides on pairwise comparisons (lower left). 692
Standard deviations are on the upper right side of the table (gray). 693
Species H. dromedarii H. excavatum H. marginatum H. rufipes H. aegyptium 1 H. aegyptium 2
H. dromedarii 0.01 0.01 0.01 0.01 0.01
H. excavatum 14.73 0.01 0.01 0.01 0.01
H. marginatum 12.60 7.75 0.01 0.01 0.01
H. rufipes 12.60 7.95 4.07 0.01 0.01
H. aegyptium 1 12.02 10.47 9.11 8.91 0.01
H. aegyptium 2 12.21 11.24 10.08 10.47 3.88
694
695
696
697 on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

35
Figure legends: 698
Figure 1: Collection sites and hosts of all Hyalomma ticks in this study. 699
700
Figure 2: Phylogenetic trees based on the Maximum likelihood method with 1000 701
bootstraps. To the left (A): 16S rRNA of FLE from Hyalomma species (this study), 702
Amblyomma maculatum (AC#:LNCT01000002), Francisella tularensis (AC#: CP017155.1) 703
and Francisella philomiragia (AC#: NR_114925.1) and corresponding Hyalomma host 704
species COI gene to the right (B). 705
706
Figure 3: Bar plot showing the contribution of each host-symbiont link to the 707
Procrustean fit, the bars represent jackknifed squared residuals, the error bars 708
represent the upper 95% confidence intervals and the dashed line represents the 709
median squared residual value. 710
711
Figure 4: FLE within Hyalomma marginatum females Malpighian tubules (A -C), ovaries 712
(D-F) and salivary glands (G-I), using whole mount organ FISH viewed under a Confocal 713
microscope. Red- FLE specific probe; blue- direct DNA staining using DAPI; 714
yellow/orange -EUB338 probe for general bacteria simultaneously labeled with FLE 715
probes. 716
FLE within Hyalomma sp. females Malpighian tubules (A -C), ovaries (D-F) and salivary 717
glands (G-I), using whole mount organ FISH viewed under a Confocal microscope. Red- 718
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

36
FLE specific probe; blue- direct DNA staining using DAPI; yellow/orange -EUB338 probe 719
for general bacteria simultaneously labeled with FLE probes. 720
721
Figure 5: Prevalence of Rickettsia in Hyalomma using diagnostic ompA PCR. Black- R. 722
aeschlimannii, dark gray-R. africae, light gray-non-sequenced positive samples. 723
on June 10, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from