Embed Size (px)
Citation preview

Title Site-independent allometric relationships for estimating above-ground weights of mangroves( 本文(Fulltext) )
Author(s) POUNGPARN, Sasitorn
Citation [Tropics] vol.[12] no.[2] p.[147]-[158]
Issue Date 2003-03-01
Rights The Japan Society of Tropical Ecology (日本熱帯生態学会)
Version 出版社版 (publisher version) postprint
URL http://hdl.handle.net/20.500.12099/31933
※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。

TROPICS Vol. 12 (2)
Site-independent allometric relationshipsfor estimating above-ground weights ofmangroves
Sasitom POUNGPARNI)', Akira KOMIYAMA", Pipat PATANAPONPAIPOON3), Vipak JINTANA",Tanuwong SANGTIEAN", Paisam TANAPERMPOOL", Somsak PIRIYAYOTA", Chatree MAKNUAL",and Shogo KATO"
I) United Graduate School of Agricultural Science, Gifu University, Gifu 501-1193, Japan"Faculty of Agriculture, Gifu University, Gifu 501-1193,Japan3) Faculty of Science, Chulalongkorn University, Bangkok 10330, Thailand" Faculty of Forestry, Kasetsart University, Bangkok 10900, Thailand') Royal Forest Department, Bangkok 10900, Thailand
2002
'Corresponding author: c/o Akira Komiyama, Laboratory of Forest Ecology, Faculty of Agriculture, Gifu University, Yanagido,Gifu 501-1193, Japan
ABSTRACT Site-independent allometric relationships for above-ground weight were studied for three groups ofmangroves: Rhizophora, Bruguiera, and other species. A hundred and one tree samples were collected from fiveforest sites where a major difference is geographical locality. For stem weight, a site-independent allometricrelationship using the variable DBH'H or DROiH (D"", = stem diameter at 30 cm above the highest prop root) wasattained for each group, because the specific gravity of wood and overall stem shape are identical among the trees ofthe different sites. Although some mangroves showed difference in specific gravity of wood among different sites,this difference was not large enough to effect the site segregation of the allometric relationship for stem weight.The allometric relationships for the branch and leaf weights of all studied mangroves differed by site when we usedDBH'H or DROiH as an independent variable. However, when we used the stem diameter at the height of the lowestbranch DB as an independent variable, the difference in allometric relationships for both branch and leaf weightsamong sites became smaller. We discussed the application of the Pipe Model (Shinozaki et ai. 1964) for establishingsite-independent allometric relationships common to a variety of geographically distinct mangroves.
Key words: mangrove forest, site-independent allometric relationship, biomass estimation, pipe Model
INTRODUCTION
Forest biomass is estimated often by the allometric method (Ogawa and Kira, 1977; Brown et ai., 1989;Haase et at., 1995; Ter-Mikaelian et at., 1997). However, a practical problem of this method is that
allometric relationships are easily segregated by forest sites (Shinozaki et ai., 1964a). Such segregation
by forest site creates a tremendous amount of field work on the researchers, because weighing the tree
bodies at each site is practically impossible. Therefore, a site-independent allometric relationship

148S. POUNGPARN, A. KOMIYAMA, P. PATANAPONPAIPOON. V. JINTANA.
T. SANGTIEAN, P. TANAPERMPOOL, S. PIRIYAYOTA, C. MAKNuAL & S. KATO
common to the related forest sites is desired.
Site-independent allometric relationships have been established for estimating the biomass in
some terrestrial forests (Crow, 1978; Brown et al., 1989; Eamus et al., 2000; Ketterings et al., 2001), but
in only a few mangrove forests (Komiyama et al., 1988b). Most allometric relationships have been
practical for their respective studied sites (Suzuki et at., 1983; Tarnai, 1986; Clough et al., 1997;
Fromard et al., 1998; Komiyama et al., 2000). An approach for establishing site-independent allometric
relationships was shown by Ketterings et at. (2001). According to them, first, the allometric
relationships for each site are to be obtained, and then these relationships are compared statistically. If
no differences are found, data from all the sites are pooled and used to determine a site-independent
allometric relationship.
Shinozaki et al. (l964a, b) have proposed a sophisticated theory on tree form, known as the Pipe
Model, which can eliminate the segregation of allometric relationships to estimate leaf and branch
amount. They assumed a tree body to be the mass of unit pipes that carry leaves and branches.
According to the Pipe Model, the leaf and branch weights within a whole crown are proportional to the
cross-sectional area of the stem at base of the crown.
In this study, we examined whether allometric relationships for above-ground weights of
mangroves are site-independent within a genus composed of several species and within mangroves
whose wood specific gravity is relatively identical. Site-independency of the Pipe Model was also
tested.
STUDY SITE AND METHODS
Two secondary mangrove forests were studied to obtain data sets on tree weights during July-August
2000 and July-August 2001. These forests are managed and controlled by Mangrove Operation Center
No.1 and 2 of the Royal Forest Department of Thailand.
One of the mangrove forests studied is located at Pang-nga Bay (8° 20' N, 98° 36' E), Southern
Thailand facing to the Andaman Sea (Fig. 1). This area had been used for tin mining about 15 years
before our study. Four study plots of 10 m x 10 m were established. The average tree density of 2,560
stemlha (DBH > 4.0 em), and the total stem basal area of 22.15 m2/ha were recorded. These plots were
dominated by Rhizophora mucronata, Bruguiera cylindrica, Xylocarpus granatum, and Sonneratia alba.
Fig.I. The study sties in two secondary mangrove forests
at Pang-nga Bay and the Trat River estuary.

Site-independent allometric relationshipsfor estimating above-ground weights of mangroves 149
Adjacent to these four plots, two study plots of 10 m x 10 m were separately set and defined as the
open-forest site, because the canopy was relatively open and the total stem basal area (8.61 m2/ha) was
relatively low. The open-forest site was dominated by X. granatum and S. alba. The exact reason why
this open forest existed is unknown, however, intense soil disturbance from the tin mining is
considered to be a casual factor of causing the open-forest site.
The other secondary mangrove forest is located on the Trat River estuary (12° 12' N, 102° 33' E),
Eastern Thailand, along the Gulf of Thailand (Fig. 1). A plot of 40 m x no m was established. The tree
density, 1,525 stemlha (DBH > 4.0 em), was lower than that of the study site at Pang-nga Bay.
Sonneratia caseolaris and Avicennia alba co-dominated at sea fringe, while R. apiculata, R. mucronata,
and B. gymnorrhiza co-dominated inland ward. Although the mean temperatures in Trat and Pang-nga
are quite comparable (27.3 - 27.6"C), the annual precipitation at Trat is relatively lower (2,663.7 mm)
than that in Pang-nga (3,014.8 mm) (Aksornkoae, 1998). The forest at Trat is affected by single day tide,
but at Pang-nga is double day tide (Royal Thai Navy, 2001).
Seven, 23, and 27 tree samples were selected for above-ground biomass studies in the open-forest
site, Pang-nga site, and Trat site, respectively (Table 1). In addition, data of 18 tree samples from
primary mangrove forests in Halmahera, Eastern Indonesia (Komiyama et ai., 1988b) and 26 tree
samples from Ranong, Southern Thailand (Tarnai et al., 1983) were used in this study (Table 1).
Table l. The size and above-ground weights of sampled trees in the five study sites.site Tree no. species DBH D RO.3 DB HB H W s W B W L
(em) (em) (em) (m) (m) (kg) (kg) (kg)
Open forest site 27 Sonneratia alba 4.3 6.1 0.45 3.46 2.25 2.55 1.17(n = 7) 24 S. alba 4.7 5.2 1.06 5.62 3.24 0.81 0.59
28 S. alba 5.2 7.0 0.60 4.49 3.67 4.09 2.2226 S. alba 10.5 10.5 1.41 7.98 20.06 15.38 3.7529 S. alba 12.7 15.0 0.96 10.54 38.23 35.67 5.4530 Xylocarpus granatum 6.7 6.0 1.42 4.17 5.31 3.03 1.1325 X granatum 7.9 7.0 1.69 5.30 8.95 6.41 2.94
Pang-nga 10 Bruguiera cylindrica 4.4 3.3 4.10 6.75 4.98 0.79 0.28(n = 23) 12 B. cylindrica 5.2 5.2 2.17 6.43 6.73 1.56 0.98
9 B. cylindrica 6.1 4.7 4.18 7.89 11.20 1.53 1.258 B. cylindrica 8.0 8 3.50 8.52 19.98 3.91 2.1011 B. cylindrica 10.3 10.7 1.41 8.15 32.60 16.82 5.157 B. cylindrica 11.9 12.7 1.43 10.71 49.10 25.21 5.9814 Rhizophora mucronata 4.7 5.1 4.8 2.37 6.97 5.75 1.08 0.8815 R. mucronata 5.6 6.3 5 3.26 8.28 9.96 1.25 1.4716 R. mucronata 7.7 7.6 7.3 2.34 8.81 17.80 4.99 2.9013 R. mucronata 8.7 8.4 7.2 3.91 9.78 24.23 4.37 2.8617 R. mucronata 9.5 10.9 11.8 2.80 10.39 33.89 23.73 10.3718 R. mucronata 10.0 8.4 7.4 4.14 8.90 22.10 6.31 3.8222 S. alba 4.2 3.6 2.60 5.77 2.72 0.29 0.1520 S. alba 5.5 3.8 4.89 8.00 6.30 1.07 0.6323 S. alba 7.3 5.1 5.96 10.74 14.78 1.70 0.7421 S. alba 9.1 7.8 2.94 9.90 18.04 3.91 1.1319 S. alba 10.6 9.0 4.66 13.44 33.88 6.30 1.393 X granatum 3.7 2.8 3.80 6.66 2.42 0.55 0.236 X granatum 4.4 4.6 1.71 7.07 4.18 2.05 0.342 X granatum 5.8 5.2 1.77 7.03 5.75 2.62 0.875 X granatum 6.6 5.9 4.00 8.05 9.46 3.34 0.91I X granatum 8.1 7.0 2.91 7.66 15.78 5.66 1.304 X granatum 12.7 14.1 1.12 8.07 29.46 32.28 5.06
Trat 666 Avicennia alba 4.8 4.8 1.51 5.94 3.34 1.50 0.68(n = 27) 665 A. alba 5.6 4.9 3.18 6.52 5.43 1.92 0.98
668 A. alba 6.2 6.7 1.16 6.17 5.82 4.48 1.68

S. POUNGPARN, A. KOMIYAMA. P. PATANAPONPAIPOON, V. ]INTANA.
150 T. SANGTIEAN, P. TANAPERMPOOL, S. PIRIYAYOTA, C. MAKNUAL & S. KATO
660 A. alba 8.1 7.1 2.82 10.00 14.26 4.60 2.38651 A. alba 8.4 8.9 1.96 9.09 14.33 8.46 3.1829 A. alba 9.5 7.0 5.30 12.37 22.68 2.95 1.01
697 A. alba 11.6 9.2 3.26 10.91 25.49 13.29 3.4030 A. alba 15.1 15.1 2.61 14.61 72.48 31.35 7.7349 B. gymnorrhiza 4.8 5.1 4.1 2.34 9.25 5.29 1.92 1.1648 B. gymnorrhiza 6.5 6.9 6.2 5.30 9.68 13.08 2.92 2.2046 B. gymnorrhiza 7.0 7.9 6.0 4.13 9.75 15.58 3.61 1.6651 B. gymnorrhiza 7.4 8.2 6.5 3.63 10.70 19.38 4.99 2.4347 B. gymnorrhiza 9.2 10.6 8.1 5.17 9.54 25.63 11.88 3.5350 B. gymnorrhiza 11.0 11.4 9.2 4.02 10.88 36.99 16.25 4.85
924 R. apiculala 3.6 7.7 6.5 6.63 15.04 26.71 3.40 1.15822 R. apiculala 4.3 8.5 7.5 8.19 16.37 35.89 5.00 2.64911 R. apiculala 4.7 4.4 4.0 4.60 7.82 6.69 1.32 0.5244 R. apiculala 5.5 5.5 4.2 4.69 8.47 7.76 1.91 0.87708 R. apiculala 8.1 6.8 6.1 2.14 6.16 8.03 3.91 1.16806 R. apiculala 8.9 10.9 7.9 8.76 16.60 49.68 8.19 2.92780 R. apiculala 14.0 13.1 12.5 5.60 14.89 75.06 17.48 6.07821 R. apiculala 14.1 10.2 8.9 9.01 17.61 61.18 10.73 4.20838 R. mucronala 4.8 4.8 4.2 3.66 8.65 6.70 1.83 0.54837 R. mucronata 5.8 5.8 4.6 3.05 7.15 6.49 2.31 0.98772 R. mucronala 6.4 6.3 5.8 3.44 8.35 9.72 2.71 1.65849 R. mucronala 9.3 8.2 7.3 5.56 13.87 28.62 5.32 2.6938 R. mucronata 11.2 10.9 7.3 10.85 16.02 60.66 8.62 4.46
Halmahera 12 B. gymnorrhiza 9.7 11.0 9.5 1.80 11.10 33.30 14.60 6.89(n = 18) 11 B. gymnorrhiza 16.5 17.6 13.5 5.80 16.10 127.00 47.00 11.55
10 B. gymnorrhiza 19.2 20.6 17.5 7.60 20.30 229.10 70.80 11.0215 B. gymnorrhiza 48.9 38.6 28.3 10.20 30.60 1085.60 280.00 45.528 R. apiculala 3.6 3.7 3.2 2.20 5.50 2.70 1.60 0.486 R. apiculala 5.7 5.7 5.4 3.10 9.50 12.10 2.70 1.344 R. apiculala 8.1 8.2 7.5 4.20 14.00 29.30 5.20 2.682 R. apiculala 9.2 9.7 6.5 7.60 12.40 27.80 4.10 1.54
site Tree no. species DBH D RO.3 DB HB H Ws WB WL(em) (em) (em) (m) (m) (kg) (kg) (kg)
5 R. apiculala 12.9 11.9 11.4 4.30 17.50 77.20 15.80 5.681 R. apiculala 14.9 14.9 11.4 8.30 21.40 130.20 16.10 6.923 R. apiculala 19.8 24.4 20.8 12.60 26.30 506.50 78.90 8.037 R. apiculala 20.5 18.5 18.0 8.30 26.20 273.70 39.70 11.049 R. apiculala 25.4 47.0 31.3 19.90 34.30 2015.50 319.60 28.8619 R. apiculala 36.4 34.7 24.0 18.30 30.10 1067.30 152.00 1Q.4218 S. alba 6.7 7.2 1.30 7.30 9.20 3.30 0.6917 S. alba 21.7 19.6 3.80 22.60 219.70 27.10 9.2213 X moluccensis 11.8 11.5 1.80 13.50 36.20 9.40 1.8714 X granatum 18.6 17.2 1.80 13.40 95.40 56.30 10.50
Ranong 1 R. apiculala 4.4 5.7 4.0 4.15 7.85 8.08 3.18 1.79(n = 26) 4 R. apiculala 7.1 7.6 5.7 4.77 10.95 19.10 4.45 2.58
6 R. apiculala 12.4 13.2 9.7 1.42 6.15 73.38 16.14 7.077 R. apiculala 10.0 10.9 7.2 5.75 12.15 35.92 8.91 3.739 R. apiculala 12.3 11.7 10.1 6.20 17.10 77.34 14.26 5.8110 R. apiculala 10.1 11.5 8.1 6.98 15.30 56.56 8.83 4.6412 R. apiculala 8.4 8.6 7.0 5.25 14.42 30.41 7.24 4.1314 R. apiculala 7.5 8.0 5.4 6.95 12.95 22.57 3.30 1.4015 R. apiculala 17.2 17.9 14.0 7.30 20.38 196.59 49.41 11.8418 R. apiculala 14.4 15.5 11.0 7.20 17.80 109.30 19.22 5.5319 R. apiculala 37.9 39.9 31.1 15.30 28.50 1264.50 141.70 59.4220 R. apiculala 39.7 43.3 32.8 12.50 31.20 1467.40 502.75 33.9721 R. apiculala 24.0 24.8 22.6 5.30 24.10 413.32 182.50 20.8622 R. apiculala 20.3 20.0 18.5 7.50 22.60 285.66 90.71 16.2525 R. apiculala 30.0 30.7 22.4 13.50 28.60 780.78 106.08 9.483 B. cylindrica 5.3 5.1 2.20 7.80 8.87 3.53 2.062 B. cylindrica 5.5 4.8 3.10 9.85 11.61 2.84 1.268 B. cylindrica 5.7 4.8 4.20 9.85 11.56 2.27 1.4213 B. cylindrica 5.9 5.2 2.50 8.78 9.96 3.44 2.5811 B. cylindrica 8.0 7.4 2.34 11.10 21.18 8.43 4.865 B. cylindrica 9.8 9.2 4.35 12.75 35.88 10.73 6.33

Site-independent allometric relationshipsfor estimating above-ground weights of mangroves 151
16 B. cylindrica 10.7 8.6 4.80 15.05 49.00 12.39 5.8326 B. gymnorrhiza 18.0 15.9 7.45 17.95 164.16 48.71 9.0623 B. gymnorrhiza 22.5 20.8 6.55 20.27 314.54 109.47 18.5724 B. gymnorrhiza 27.2 25.0 10.00 22.30 568.81 109.32 11.4217 B. gymnorrhiza 33.4 27.1 8.65 24.95 790.99 124.05 28.55
Before weighing each tree, the stem diameters at ground level (Do), at 30 cm height (D03), at each
1-m interval (D L3 = DBH, D23, D3.3...), and at the height of lowest living branch (DB) were measured. For
Rhizophora species, stem diameter at 30 cm above the highest prop root (DRO.3) was also measured,
because we used DRO.3standing for DBH of Rhizophora species. The total tree height (H), and the height
of the lowest living branch (HB) were also recorded. Assuming a conical-shaped stem, the stem
diameters at each 1-m interval were used for calculating the stem volume (Vs) as described by
Komiyama et at. (2002).
Each sample tree was cut at ground level using handsaws and carried to the laboratory at the
Mangrove Operation Center. Each sample tree was manually separated into stem, branch, and leaf.
Those organs were weighed fresh using an electric balance with an accuracy of 0.1 g (Bonso Co. Ltd.,
model 339) and then stem weight, Ws; branch weight, WB; and leaf weight, WL; were converted to dry
weights. For the conversion, approximately samples of 500 g of each organ were oven-dried (llOt for
48 hours) to acquire the dry matter ratios.
The slopes of regression lines obtained from the relationships for respective sites were tested by
ANCOVA in order to judge the segregation of allometric relationships among sites. If the slopes were
not statistically different, then differences in intercepts were tested. The site independent allometric
relationship was to be established by pooling sample trees from all sites in the case of similar slope and
intercept. On the other hand, significantly different slopes or the same slopes but different intercepts
indicate the segregated allometric relationships (Crow, 1978). We dealt the site-independent allometric
relationships separately for the three species groups: Rhizophora species (R. apiculata and R.
mucronata, here after RH), Bruguiera species (B. cylindrica and B. gymnorrhiza, here after BG), and the
other species (5. alba, A. alba, X moluccensis here after OT). For the three species of OT, the specific
gravities of wood were similarly light (Walsh, 1977).
RESULTS
Allometric relationship for stem weight
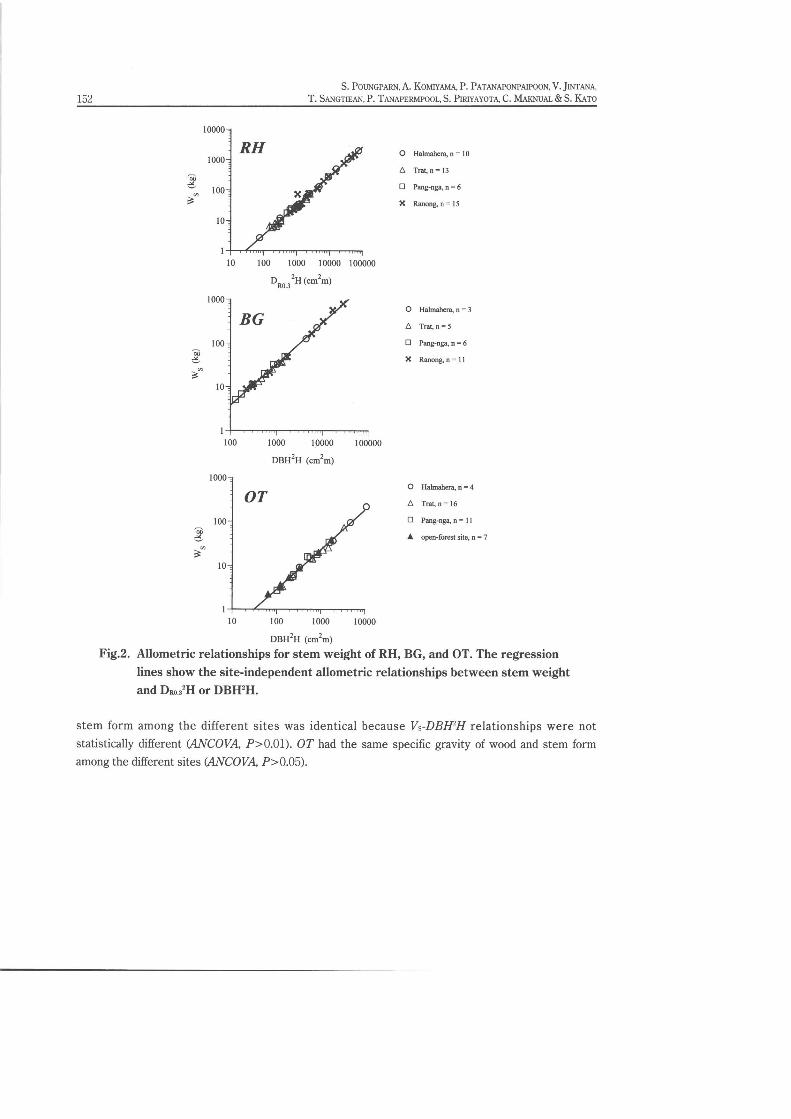
The allometric relationships for stem weight were established using the parameter of DRO}H for
RH, and DBHH for BG and OT. The allometric relationships for stem weight were site-independent for
RH, BG, and OT (Fig. 2). Neither slopes nor intercepts differed statistically (ANCOVA, P>0.05, Table
2) among the allometric relationships of respective sites. For obtaining the relationship for BG, we
excluded the two outlier samples (the smallest B. gymnorrhiza of the Trat site, No.49; and the biggest
B. gymnorrhiza of Halmahera site, No.15; Table 1) in the analysis.
In addition to the Ws - DRO}H and Ws - DBHH relationships, we checked the site-dependency of
the specific gravity of wood and that of stem form using Ws-Vs and Vs-DBHH (DRoiH for RH)
relationships, respectively (Table 2). RH and BG showed significant differences in specific gravity of
wood among different sites, indicated by the difference in intercept (ANCOVA, P<O.Ol). However,
152S. POUNGPARN. A. KOMIYAMA. P. PATANAPONPAIPOON, V. ]INTANA,
T. SANGTIEAN, P. TANAPERMPOOL, S. PIRIYAYOTA, C. MAKNuAL & S. KATO
o Halmahera, n = 3
)( Ranong, n = II
t; Tra~n=5
o Pang-ng., n = 6
t; Tra~ n = 13
o Halmahera,n= 10
o Pang-nga, n = 6
X Ranong, n = 15
10
100
10000
1000
OilC
100~'"
10
1
10 100 1000 10000 100000
DRO
.3 2H (cm2m)
1000
1100 1000 10000 100000
DBH2H (cm2m)
10000 Halmahera, n = 4
OT t; Trat, n= 16
100 0 Pang-nga, n = II
~ .6- open~forest site, n =7
~'"
10
110 100 1000 10000
DBH2H (cm2m)
Fig.2. Allometric relationships for stem weight of RH, BG, and OT. The regressionlines show the site-independent allometric relationships between stem weightand DRo.lH or DBH'H.
stem form among the different sites was identical because Vs-DBlPH relationships were not
statistically different (ANCOVA, P>O.Ol). OT had the same specific gravity of wood and stem form
among the different sites (ANCOVA, P>O.05).

Site-independent allometric relationshipsfor estimating above-ground weights of mangroves 153
Table 2. Allometric relationships for above-ground weight of RH, BG and OT. These
relationships were determined from data sets of all sites and had significant
P-values (P<O.Ol). The general form is W = aX", when W is plant weight, X =variable corresponding to stem diameter, a and b are constants presented as
Y-intercept and slope respectively. P value of a and b were calculated for
comparison of allometric relationships for respective sites by using ANCOVA.
Values in the last three columns (a, b, and R2) are for site-independent
models, which were calculated using the pooled data from all the sites.allometric Species Site P value P value a b R'
relationship inpendency ofa ofb (S.E.) (S.E.)
WS-DRO.,'H RH 0 0.881 0.995 0.0394 (0.0481) 0.966 (0.0146) 0.991
Ws-DBH'H BO 0 0.082 0.867 0.0472 (0.0390) 0.948 (0.0126) 0.996
Ws-DBH'H OT 0 0.055 0.745 0.0444 (0.041 I) 0.904 (0.0150) 0.992
Ws-V s RH X 0.0001 0.784 819.8 (0.0126) 1.021 (0.0091 I) 0.997
W s - Vs BO X 0.001 0.579 701.5 (0.0207) 0.992 (0.0144) 0.995Ws-V s OT 0 0.089 0.967 491.5 (0.0226) 0.995 (0.0134) 0.995
VS-DRO.,'H RH 0 0.341 0.947 0.00006 (0.0520) 0.944 (0.0158) 0.988
Vs-DBH'H BO 0 0.120 0.488 0.00006 (0.0522) 0.952 (0.0169) 0.993Vs-DBH'H OT 0 0.064 0.349 0.00009 (0.0355) 0.907 (0.0129) 0.994
W.-D RO.,' H RH X 0.017 0.0155 (0.127) 0.884 (0.0385) 0.926W.-DBH'H BO X 0.004 0.0117 (0.162) 0.968 (0.0523) 0.937W.-DBH'H OT ~ 0.034 0.694 0.0263 (0.291) 0.832 (0.106) 0.687
W.-D.' RH ~ 0.020 0.050 0.0416 (0.0840) 1.267 (0.0426) 0.955
W.-De' BO ~ 0.016 0.051 0.0411 (0. J08) 1.271 (0.0555) 0.958
W.-De' OT 0 0.298 0.685 0.0283 (0.133) 1.280 (0.0751) 0.912
WL-D RO./H RH X 0.003 0.036 0.0424 (0.132) 0.605 (0.040 I) 0.844WL-DBH'H BO X 0.008 0.0384 (0.179) 0.658 (0.0578) 0.849WL-DBH]H OT X <0.001 0.431 0.0276 (0.264) 0.643 (0.0961) 0.615
W L - De' RH ~ 0.021 0.022 0.0797 (0.0931) 0.878 (0.0472) 0.892
WL-De' BO ~ 0.023 0.072 0.0890 (0.133) 0.868 (0.0686) 0.874
WL-De' OT ~ 0.023 0.340 0.0269 (0.142) 1.009 (0.0798) 0.851
(0) indicates the site-independent relationship in which both the P value of a and b are more than 0.05.
(LI.) indicates the site-independent relationship in which the P value of a or b is 0.01 <P<0.05.
(X) indicates the site-dependent relationship in which the P value of a or b is less than 0.01.
Allometric relationship for branch weight
The allometric relationships for the branch weight (W.) of RH, BG, and OT were separately established
using two variables, DB/FH (DRO}H for RH) and DB2• Although the WB-DB/FH relationships were
significant (P<O.Ol) for RH and BG in each site (Fig. 3A), these relationships showed the segregation
by site for both genera, indicated by the significantly different slopes in the WB-DB/FH relationship
among the sites (ANCOVA, P<O.Ol, Table 2). The WB-DB/FH relationship for OT was also site
dependent, indicated by the different intercepts among the sites (ANCOVA, P<O.05, Table 2).
When we use the square of the diameter at the height of the lowest living branch (J)i) as the
independent variable (Fig. 3B), the site-dependence in allometric relationships for branch weight
became smaller than that of when we used the variable of DB/FH for all species groups, which was
indicated by inflation ofR2 (Fig. 3, Table 2). However, site-independence was still significant (ANCOVA,
P<O.05) except for OT (Table 2).

154S. POUNGPARN. A. KOMIYAMA. P. PATANAPONPAIPOON. V. ]INTANA.
T. SANGTlEAN. P. TANAPERMPOOL. S. PIRIYAYOTA. C. MAKNuAL & S. KATO
(A) (B)
RH
1000 10000 100000 100 1000 10000
DRO
.3
2H (cm2m) DB
2 (cm2)
1000 1000
100 100
'Oilc-10 10
Ol::: BG
0.10.1100 1000 10000 100000 10 100 1000 10000
DBH 2H (cm2m) DB
2 (cm2)
100 100
10 10'OilC-
Ol:::
0.1 0.1 +...,..-'\'~mr~~mr~~""10 100 1000 10000 10 100 1000
DBH 2H (cm2m) DB
2 (cm2)
Fig.3. Allometric relationships for branch weight of RH, BG, and OT. Symbols and
number of tree samples were the same as shown in Fig. 2. (A) The regression
lines show the allometric relationships between branch weight and the
parameter of DRoiH or DBH2H for respective sites. The allometric relationships
showed complete segregation by site. (B) The regression lines show the site
independent allometric relationship between branch weight and DB2
•
Allometric relationship for leaf weight
The WL-DBlFH relationship was segregated by site (Fig. 4A), like in the case of the allometric
relationship for branch weight. However, when the D.2 was used for the variable in the allometric
relationship (Fig. 4B), the site-dependence was reduced, though still significant for the three groups of
mangroves (ANCOVA, P<0.05, Table 2). The slope of the WL- D B2 relationship was not statistically
different from 1.0 atP<0.05 for BG and OT, atP<O.Ol for RH (t-test).

Site-independent allometric relationshipsfor estimating above-ground weights of mangroves 155
(A)100
10
(B)
RH
1000010001000.1 +-~~'"'..,.....,~mr~...,...,.""
1010000 10000010000.1 +..,.....,~mr..,.....,~mr...,....,""T"n""
100
10000
10
0.1 -+-"'-....,..,..,.,.",.--"...,...,.."...,..,........,.....,.-rTT"'!
10
100
1000 10000 100000
DBH2H (cm2m)
10
0.1+-.......,...,.,.",.,..............,..,.,."",......,...,..,..,."",
100
100
1000100101000010001000.1 +..,.....,"\<mmr-..,.....,~mr...,....,""T"n""
10
10
100
FigA. Allometric relationships for leaf weight of RH, BG, and QT. Symbols and number
of tree samples were the same as shown in Fig. 2. (A) The regression lines show
the allometric relationships between leaf weight and the parameter of DRoiH or
DBH'H for respective sites. The allometric relationships show complete
segregation by site. (B) The regression lines show the site-independent allometric
relationship between leaf weight and Do'.
DISCUSSION
The site-dependence of allometric relationships for above-ground weights of mangroves is summarized
in Table 2. The allometric relationship for stem weight, the Ws-DBlPH relationship, was site
independent (ANCOVA, P>O.05) for RH, OT, and BG. Therefore, we can establish a common equation
for each species group. Stem weight is a function of stem volume and specific gravity, whereas the

156S. POUNGPARN, A. KOMIYAMA. P. PATANAPONPAIPOON, V. ]INTANA,
T. SANGTIEAN. P. TANAPERMPOOL. S. PIRIYAYOTA, C. MAKNUAL & S. KATO
value of DBHH is proportional to the volume of a cylinder with diameter (DBll) and height (ll). If the
wood specific gravity and the stem form are identical among the different sites, a common equation can
be obtained (Crow, 1978). The specific gravity of wood differed among the different sites for RH and
BG. The site-independency of the Ws-DBHH relationship proved that this difference in specific gravity
was ignorably small for attaining a site-dependence in allometric relationship for stem weight. Recently,
the allometric relationships using the specific gravity of stem as a part of the variable have been
proposed by some authors for establishing species- and site-specific allometric relationships (Brown etai., 1989; Ketterings et at., 2001; Komiyama et at., 2002).
The allometric relationship of branch and leaf weights was site-dependent for all the species
groups when we used DBlFH as the independent variable. The assemblage of both living and dead
pipes in the stem at breast height may cause this site-dependency. Nevertheless, when the parameter
of DB2 was used, the allometric relationship for branch and leaf weights was less site-dependent for each
species group. Since the thickness of stem at the lowest living branch gathers living pipes carrying the
total amount of leaves and branches, the fairly site-independent allometric relationship was obtained
usingDl.
The score of 1.0 for the slope of the WL - Dl relationship of the three species groups implies that
the WL - DB2 relationship is proportional. A similar result was shown by Shinozaki et at. (1964b) for the
leaf-allometric relationships of a Japanese coniferous tree growing in different stands. It is noteworthy
that establishing a site-independent allometric relationship for branch and leaf weights of mangroves
could be accomplished by the application of the Pipe Model. The Pipe Model was also successfully used
for establishing a common allometric relationship for leaf weight of some broad-leaved trees (Hoffmann
et at., 2002).
This study shows that a site-independent allometric relationship can be obtained by selecting
suitable variables, such as DBlFH and DB2, to be used in the equation. Some of the former allometric
equations on mangroves which used only the variable of DBlFH (Ong et at., 1982; Suzuki et at., 1983;
Kusmana et at., 1992), or D (Clough et at., 1989; Slim et at., 1996; Clough et at., 1997) should be site
dependent, especially for branch and leaf weights. Although the measurement of DBH and H is
relatively easy in the field compared to the DB measurement, Kanto (1999) and Matsushita et at. (2000)
recently introduced devices to measure DB easily.
As the site-independent allometric relationships established in this study are applicable to the
mangroves both in primary and secondary forests, these allometric relationships may be applied to
various sites with wide geographical difference at least in Southeast Asia.
ACKNOWLEDGEMENTS We are grateful to the National Research Council of Thailand and Royal
Forest Department of Thailand for allowing research in the study sites. We thank Mangrove Operation
Center No.1 and 2 for providing accommodation during our field studies. Prof. K. Ogino and Prof. S.
Tarnai are appreciated for favoring with biomass data of the mangrove forest collected in Halmahera,
Eastern Indonesia, and Ranong, Southern Thailand. We also thank Dr. A. Sumida for comments and
suggestions on the manuscript.

Site-independent allometric relationshipsfor estimating above-ground weights of mangroves
REFERENCES
157
Aksornkoae, S. 1998. Mangroves ecology and management (in Thai). 2nd edition. Kasetsart University
Press, Bangkok, pp.277.
Brown, S., Gillespie, A.J.R. & Lugo, A.E. 1989. Biomass estimation methods for tropical forests with
applications to forest inventory data. Forest Science 35: 881-902.
Clough, B.F. & Scott K 1989. Allometric relationships for estimating above-ground biomass in six
mangrove species. Forest Ecology and Management 27: 117-127.
Clough, B.F., Dixon, P. & Dalhaus, O. 1997. Allometric relationships for estimating biomass in multi
stemmed mangrove trees. Australian Journal ofBotany 45: 1023-1031.
Crow, T.R. 1978. Common regressions to estimate tree biomass in tropical stands. Forest Science 7: 110
-114.
Eamus, D., McGuinness, K & Burrows, W. 2000. Review ofallometric relationships for estimating woody
biomass for Queensland, the northern territory and western Australia. In the national carbon
accounting system, technical report no. Ea. Australian Greenhouse Office, pp. 56.
Fromard, F., Puig, H., Mougin, E., Marty, G., Betoulle, ].1. & Cadamuro, 1. 1998. Structure above
ground biomass and dynamics of mangrove ecosystems: New data from French Guiana. Oecologia
115: 39-53.
Haase, R. & Haase, P. 1995. Above-ground biomass estimates for invasive trees and shrubs in the
Pantanal of Mato Grosso, Brazil. Forest ecology and Management 73: 29-35.
Hoffmann, C.W. & Usoltsev, V.A. 2002. Tree-crown biomass estimation in forest species of the Ural
and of Kazakhstan. Forest ecology and Management 158: 59-69.
Kanto, T., 1999. Inventions of the new instruments for pointing and measuring of any heights of trunks
and for remote-measuring the upper diameters by using the red-laser-pointer. J. Jpn. For. Soc. 81:85-88.
Ketterings, Q.M., Coe, R., Van Noordwijk, M., Ambagau, Y. & Plam, C.A. 2001. Reducing uncertainty in
the use of allometric biomass equations for predicting above-ground tree biomass in mixed
secondary forests. Forest Ecology and Management 146: 199-209.
Komiyama, A., Havanond, S., Srirawatt, W., Mochida, Y., Fujimoto, K, Ohnishi, T., Ishihara, S. &
Miyagi, T. 2000. Top/root biomass ratio of a secondary mangrove (Ceriops tagal (Perr.) C.B. Rob.)
forest. Forest Ecology and Management 139: 127-134.
Komiyama, A., Jintana, V., Sangtiean, T. & Kato, S. 2002. A common allometric equation for predicting
stem weight of mangroves. Ecological Research 17: 415-418.
Komiyama, A., Moriya, H., Prawiroatmodjo, S., Toma, T. & Ogino, K 1988a. Floristic composition and
stand structure. In: Ogino, K & M. Chihara (Eds.) Biological system ofmangroves, Ehime University,pp.85-96.
Komiyama, A., Moriya, H., Prawiroatmodjo, S., Toma, T. & Ogino, K 1988b. Primary productivity of
mangrove forest. In: Ogino, K. & M.Chihara (Eds.) Biological system of mangroves, Ehime
University, pp. 97-117.
Kusmana, C., Sabiham, S., Abe, K & Watanabe, H. 1992. An estimation of above ground tree biomass of
a mangrove forest in east Sumatra, Indonesia. Tropics 1: 243-257.
Matsushita, T., Kato, S. & Komiyama, A. 2000. Accuracy of an equipment for remote measuring of tree-

158S. POUNGPARN. A. KOMIYAMA. P. PATANAPONPAIPOON. V. ]INTANA.
T. SANGTIEAN. P. TANAPERMPOOL. S. PlRIYAYOTA. C. MAKNuAL & S. KATO
trunk diameters. Presentation in The Chubu Branch of the Japanese Forestry Society Mie, October
14,2000. Mie University.
Ogawa, H. & Kira, T. 1977. Methods of estimating forest biomass. In: Shidei, T. & Kira, T., (Eds.),
Primary productivity ofJapanese forests: Productivity of terrestrial communities, University of Tokyo,
JIBP Synthesis 16: 45-52.
Ong, J.E., Gong, W.K & Wong, C.H. 1982. Studies on nutrient levels in standing biomass, litter and
slash in a mangrove forest. BIOTROP, Bogor, pp. 44.
Royal Thai Navy. 2001. Tide tables of Thailand. Bangkok, pp. 313. (In Thai)
Shinozaki, K, Yoda. K, Hozumi, K & Kira, T. 1964a. A quantitative analysis of plant form - The pipe
model theory. I. Basic analysis.]ap.]. Ecol. 14: 97-105.
Shinozaki, K, Yoda. K, Hozumi, K & Kira, T. 1964b. A quantitative analysis of plant form - The pipe
model theory. II. Further evidence of the theory and its application in forest ecology. Jap. ]. Ecol.14: 133-139.
Slim, F.j., Gwada, P.M., Kodjo, M. & Hemminga, M.A. 1996. Biomass and litterfall of Ceriops tagal and
Rhizophora mucronata in the mangrove forest of Gazi bay, Kenya. Mar. Freshwater Res. 47: 999
1007.
Suzuki, E. & Tagawa, E. 1983. Biomass of a mangrove forest and a sedge marsh on Ishigaki island,
south Japan.]ap.]. EcoZ. 33: 231-234.
Tarnai, S., Nakasuga, T., Tabuchi, R. & Ogino, K 1983. Ecological studies of mangrove forests in
Southern Thailand: Standing structure and biomass. Mangrove ecology in Thailand, Thai-Japanese
cooperative research project on mangrove productivity and development, pp. 1-15.
Tarnai, S., Nakasuga, T., Tabuchi, R. & Ogino, K 1986. Standing biomass of mangrove forests in
southern Thailand.]. Jpn. For. Soc. 68: 384-388.
Ter-Mikaelian, M.T. & Korzukhin, M.D. 1997. Biomass equations for sixty-five north American tree
species. Forest Ecology and Management 97: 1-24.
Walsh, G.E. 1977. Exploitation of mangai. In: Chapman v.]. (Ed.), Wet coastal ecosystems: Ecosystem ofthe world, Amsterdam, pp. 347-362.
Received June 4, 2002
AcceptedJan.9,2003





























