Embed Size (px)
Citation preview

Vietnam J. Agri. Sci. 2019, Vol. 17, No. 3: 196-203 Tạp chí Khoa học Nông nghiệp Việt Nam 2019, 17(3): 196-203 www.vnua.edu.vn
196
TÍNH ĐA HÌNH ĐƠN NUCLEOTIDE
TRONG PHÂN ĐOẠN GEN MÃ HÓA ENZYME CATHEPSIN D ASPARTIC PROTEASE
CỦA LOÀI GIUN MÓC TRUYỀN LÂY Ancylostoma ceylanicum
Dương Đức Hiếu1*, Bùi Khánh Linh1, Nguyễn Thu Hương2, Lê Đức Vĩnh3, Nguyễn Thị Hoàng Yến1, Nguyễn Thị Hồng Chiên1, Nguyễn Văn Phương1
1Khoa Thú y, Học viện Nông Nghiệp Việt Nam 2Viện Sốt rét - Ký sinh trùng - Côn trùng TW
3Đại học Y Dược Phạm Ngọc Thạch
*Tác giả liên hệ: [email protected]
Ngày nhận bài: 02.04.2019 Ngày chấp nhận đăng: 01.06.2019
TÓM TẮT
Nghiên cứu được tiến hành nhằm xác định sự tồn tại của loài giun móc chó truyền lây Ancylostoma ceylanicum
trên người tại Việt Nam, đồng thời phân tích đặc điểm đa hình đơn nucleotide trong phân đoạn gen mã hóa enzyme
cathepsin D aspartic protease của A. ceylanicum, một trong những loại enzyme thủy phân đóng vai trò quan trọng
trong vòng đời phát triển của giun móc (Williamson et al., 2004). Bằng phương pháp KOD-PCR, sự tồn tại của loài
giun móc chó truyền lây Ancylostoma ceylanicum trên người và chó tại Việt Nam được khẳng định. Kết quả so sánh
các trình tự phân đoạn gen mã hóa enzyme cathepsin D aspartic protease của A. ceylanicum cho thấy xuất hiện
nhiều đột biến điểm tại vùng Exon 7, 8, 9. Đặc biệt, 3 đột biến điểm không đồng nghĩa (1 điểm đột biến trên vùng
exon 7 và 2 điểm đột biến trên vùng exon 9) được xác định gây ra sự thay đổi về trình tự axitamin của aspartyl
protease. Kết quả nghiên cứu về tính đa hình đơn nucleotide trên vùng gen mục tiêu này của chúng tôi là cơ sở khoa
học cho việc phát triển các loại vacxin phòng bệnh giun móc hiệu quả sử dụng kháng nguyên cathepsin D aspartic
protease tái tổ hợp.
Từ khóa: Ancylostoma ceylanicum, tính đa hình đơn nucleotide, cathepsin D aspartic protease.
Single Nucleotide Polymorphisms (SNPs) in the Gene Segment Coding for Cathepsin D Aspartic Protease of Zoonotic Hookworm Ancylostoma ceylanicum
ABSTRACT
This study was conducted to determine the presence of the canine zoonotic hookworm species, Ancylostoma
ceylanicum, in humans in Vietnam and to analyze the single nucleotide polymorphism in the gene segment encoding
A. ceylanicum cathepsin D aspartic protease, one of the hydrolytic enzymes with an important role in the life cycle of
hookworm. The existence of Ancylostoma ceylanicum in dogs and humans in Vietnam was confirmed by KOD-PCR.
Comparative analysis of tthe sequences encoding A. ceylanicum cathepsin D aspartic protease revealed, some
mutations in exon 7, 8, and 9. Three SNPs non-synonymous mutations (one from exon 7 and two from exon 9)
caused the changes in amino acid sequences of aspatyl protease. Our results can be considered for futher studies
on the development of effective vaccine against hookworm disease using recombinant cathepsin D aspartic protease
as an antigen.
Keywords: Ancylostoma ceylanicum, single nucleotide polymorphisms, cathepsin D aspartic protease.
1. ĐẶT VẤN ĐỀ
Bệnh giun móc là một trong những căn
bệnh truyền lây được Tổ chức Y tế thế giới
WHO xếp vào danh mục những căn bệnh bị
lãng quên tại vùng nhiệt đới, ước tính trên 700
triệu người tại các quốc gia đang phát triển bị
nhiễm căn bệnh này (Hotez et al., 2004). Đặc
biệt, Ancylostoma ceylanicum, loài giun móc
chó duy nhất có khả năng lây nhiễm và phát

Dương Đức Hiếu, Bùi Khánh Linh, Nguyễn Thu Hương, Lê Đức Vĩnh, Nguyễn Thị Hoàng Yến, Nguyễn Thị Hồng Chiên, Nguyễn Văn Phương
197
triển thành dạng trưởng thành ở cơ thể người,
đã được ghi nhận với tỷ lệ nhiễm cao trên chó
và người tại khu vực Đông Nam Á (Traub et
al., 2008; Ngui et al., 2012a; Ngui et al., 2012b;
Conlan et al., 2012; Inpankaew et al., 2014),
trong đó có Việt Nam (Dinh et al., 2015; Duong
et al., 2018). Giun móc ký sinh trong ruột vật
chủ, lấy chất dinh dưỡng từ việc hút máu, ly
giải hồng cầu, phân hủy huyết sắc tố
Hemoglobin (Hb) và các loại protein khác bằng
con đường thủy phân protein (Williamson et
al., 2004). Mebendazole và albendazole là
những loại thuốc được sử dụng phổ biến trong
việc tẩy giun. Tuy nhiên, một số nghiên cứu đã
chỉ ra tính kháng thuốc trên nhóm giun truyền
qua đất, trong đó có loài giun móc Necator
americanus, do liên quan đến đột biến điểm di
truyền (Ambrose et al., 2018). Do vậy, việc
phát triển các loại thuốc tẩy giun mới, cũng
như vacxin được coi là cần thiết trong việc
phòng chống căn bệnh này. Cathepsin D
aspartic protease (aspartyl protease) là một
trong những loại enzyme thủy phân đóng vai
trò quan trọng trong việc phân hủy huyết sắc
tố trong quá trình tiêu hóa của giun móc
trưởng thành (Williamson et al., 2002;
Williamson et al., 2003a), cũng như phân hủy
mô của vật chủ trong quá trình xâm nhập và di
hành của ấu trùng (Brown et al., 1999;
Williamson et al., 2003b; Jolodar et al., 2004).
Do đóng vai trò quan trọng trong sự tồn tại và
phát triển của giun móc ở cơ thể vật chủ,
aspartyl protease được nhiều nghiên cứu đánh
giá cao về tiềm năng ứng dụng trong việc phát
triển vacxin phòng bệnh giun móc (Loukas et
al., 2005). Việc đánh giá tính đa hình đơn
nucleotide (SNPs) là cần thiết, đặc biệt là
những đột biến điểm không đồng nghĩa (non-
synonymous SNPs) dẫn đến sự sai khác trong
trình tự axit-amin có thể làm thay đổi cấu
trúc, chức năng của protein và ảnh hưởng tới
hiệu quả của vacxin phòng bệnh. Trong nghiên
cứu bước đầu này, phân đoạn gen (vùng Exon
7- Exon 8- Exon 9) mã hóa aspartyl protease
của loài giun móc truyền lây A. ceylanicum
được giải trình tự và phân tích tính đa hình
đơn nucleotide.
2. VẬT LIỆU VÀ PHƯƠNG PHÁP
2.1. Mẫu và nguồn gốc mẫu
Mẫu phân chó thu thập ngẫu nhiên tại các
hộ nuôi chó tại Hà Nội và Bắc Ninh được chuyển
về Phòng thí nghiệm Ký sinh trùng - Bộ môn Ký
sinh trùng, Khoa Thú y, Học viện Nông Nghiệp
Việt Nam để xét nghiệm. Mẫu ấu trùng giun móc
người thu từ các bệnh nhân dương tính với trứng
giun móc được cung cấp bởi Viện Sốt rét - Ký
sinh trùng - Côn trùng TW (NIMPE).
2.2. Phương pháp phù nổi Fullerborn
5 gram mẫu phân chó được cho vào cốc sạch
chứa khoảng 50-60 mL dung dịch nước muối
bão hòa, dùng đũa thủy tinh khuấy đều, lọc
dung dịch qua lưới lọc và chuyển dung dịch sang
ống fancol 15 cho đầy, đặt phiến kính lên. Sau
15-20 phút trứng giun sẽ nổi lên bám vào phiến
kính, lấy ra và soi dưới kính hiển vi quang học
(x10) xác định trứng giun móc theo khóa định
danh Bowman, 2009.
2.3. Phương pháp nuôi ấu trùng trên
thạch agar
Khoảng 5 gram mẫu phân chó dương tính
với trứng giun móc được chuyển lên bề mặt đĩa
thạch agar (2%). Trứng giun móc được nuôi
trong tủ ấm 27C và kiểm tra hàng ngày. Sau
khoảng 2-3 ngày nuôi cấy, ấu trùng giun móc
được thu và bảo quản trong dung dịch cồn 70.
2.4. Phương pháp tách chiết DNA tổng số
Từng ấu trung giun móc riêng rẽ được rửa
sạch bằng dung dịch PBS-Tween20 (0,1%) và
chuyển vào ống Eppendorf chứa dung dịch đệm
lysis gồm Direct PCR Lysis Reagent (Tail)
(VIAGEN Biotech, Canada), 1 M Dithiothreitol
và Proteinase K (QIAGEN, Hà Lan). Ủ hỗn hợp
lysis trong 20 phút ở 60C, 10 phút ở 95C. DNA
tổng số của ấu trùng giun móc được bảo quản
trong tủ âm (-20C) đến khi dùng.
2.5. Phương pháp KOD-PCR nhân vùng gen
mt cox1 và phân đoạn gen aspartyl protease
Cặp mồi HPo1: 5’-TTACGTAGAAGGTCAA
TTTCTTTGG-3’ và HPo2: 5’- CTACTTAACCTA

Tính đa hình đơn nucleotide trong phân đoạn gen mã hóa enzyme cathepsin D aspartic protease của loài giun móc truyền lây Ancylostoma ceylanicum
198
TACTCCGGCCTTC-3’ được thiết kế dựa trên
trình tự gen ty thể của A. ceylanicum
AP017674.1 (NCBI) nhằm nhân phân đoạn gen
mt cox1 (983 bp). Phản ứng KOD- PCR được thực
hiện sử dụng KOD Fx Neo polymerase với chu kỳ
nhiệt như sau: 94C trong 2 phút, 35 chu kỳ lặp
lại (94C - 10 giây, 63C - 30 giây, 68C - 1 phút),
68C - 5 phút và giữ sản phẩm ở 4C. Tính đặc
hiệu của phản ứng KOD-PCR được kiểm tra sử
dụng mạch khuôn DNA của các loài giun móc
A. caninum, A. ceylanicum và N. americanus làm
đối chứng. Căn cứ vào trình tự genome của
A. ceylanicum JARK01001578.1 (NCBI), cặp mồi AceyAP3F: 5’-GGTGTTCGCTTTCTGGCTCA-3’
và AceyAP3R: 5’-GGTCAATACGTAATCTTCAC
CCTTG-3’ được thiết kế phủ vùng mã hóa Exon
7, 8, 9 của aspartyl protease gen. PCR master
mix trong 25 uL phản ứng gồm 12,5 uL dung
dịch đệm 2x KOD Fx PCR, 5 uL 2 mM dNTP, 0,5
uL KOD Fx Neo polymerase, 0,75 uL 10 uM mồi
xuôi và ngược, 1ul mạch khuôn DNA và nước cất
tinh sạch. Chu trình nhiệt cho phản ứng PCR
như sau: khởi đầu 94C trong 2 phút, 35 chu kỳ
lặp lại (biến tính 98C -10 giây, gắn mồi ở 63C -
30 giây, kéo dài ở 68C - 45 giây), kết thúc kéo
dài 68C - 5 phút và giữ sản phẩm ở 4C. Sản
phẩm PCR được kiểm tra bằng phương pháp điện
di trên thạch agarose 1,5% chứa GelRed (Wako,
Nhật Bản) trong 30 phút ở 100V trong dung dịch
đệm 1X TAE.
2.6. Phương pháp TA cloning
Sản phẩm PCR của phản ứng nhân phân
đoạn gen aspartyl protease được tinh sạch bằng
bộ kit QIAquick PCR Purification (QIAGEN, Hà
Lan) và được gắn với plasmid pCR 2.1-TOPO
bằng TOPO TA Cloning kits (Thermo Fisher
Scientific, Mỹ). Sau đó, vector tách dòng gen
được biến nạp vào tế bào khả biến E. coli DH5α
bằng phương pháp sốc nhiệt và được nuôi trong
môi trường SOC trong tủ ấm lắc 37C. Sau 1 giờ
nuôi cấy, tế bào E. coli được cấy chuyển trên bề
mặt thạch LB chứa Amplicilin (50 ug/mL) và
chất chỉ thị màu X-Gal, nuôi trong tủ ấm 37C
trong 12 giờ. Kiểm tra sự phát triển của khuẩn
lạc, chọn lọc các khuẩn dương tính (màu trắng)
và tiến hành phản ứng Colony PCR sử dụng cặp
mồi AceyAP3F và AceyAP3R.
Ghi chú: M: thang chuẩn DNA; giếng 1,2: mẫu ấu trùng giun móc từ chó; giếng 3,4: mẫu ấu trùng giun móc từ người
Hình 1. (A) Tính đặc hiệu của phản ứng KOD-PCR sử dụng cặp mồi HPo1
và HPo2 nhân phân đoạn gen ty thể mt cox1 nhằm sàng lọc loài giun móc A. ceylanicum;
(B) Ảnh điện di sản phẩm PCR nhân phân đoạn gen ty thể mt cox1 của A. ceylanicum
trên gel Agarose 1,5%

Dương Đức Hiếu, Bùi Khánh Linh, Nguyễn Thu Hương, Lê Đức Vĩnh, Nguyễn Thị Hoàng Yến, Nguyễn Thị Hồng Chiên, Nguyễn Văn Phương
199
2.7. Phương pháp Sequencing PCR và giải
trình tự gen
Sản phẩm Colony PCR sau khi tinh sạch
được sử dụng là mạch khuôn cho phản ứng
Sequencing PCR bằng bộ kit BigDye
Terminator v3.1 Cycle Sequencing (Thermo
Fisher Scientific, Mỹ). Chu trình nhiệt của phản
ứng Sequencing PCR như sau: ủ 96C trong 1
phút, 25 chu kỳ lặp lại (biến tính 96C - 10 giây,
gắn mồi ở 50C - 5 giây, kéo dài ở 60C - 4 phút)
và giữ sản phẩm ở 4C. Sản phẩm Sequencing
PCR được tinh sạch bằng hạt từ tính - Magnetic
Bead Agencourt AMPure XP (Beckman Coulter,
Mỹ) và chuyển vào đĩa PCR 96 giếng. Trình tự
gen được giải mã sử dụng phương pháp Sanger
Sequencing bằng hệ thống máy giải trình tự gen
ABI Prism 3100 genetic analyzer (Applied
Biosystems, Mỹ).
2.8. Xử lý số liệu
Toàn bộ kết quả trình tự gen được xử lý,
phân tích và so sánh bằng phần mềm 4Peak và
MEGA7, đồng thời sử dụng hệ thống dữ liệu về
trình tự gen, trình tự protein từ NCBI để
tham khảo.
3. KẾT QUẢ VÀ THẢO LUẬN
3.1. Tính đặc hiệu của phản ứng KOD-PCR
và kết quả sàng lọc loài giun móc truyền lây
Ancylostoma ceylanicum thu tại Việt Nam
Tính đặc hiệu của phản ứng KOD-PCR
phân biệt loài giun móc truyền lây
A. ceylanicum được kiểm tra bằng các mẫu DNA
đối chứng được tách chiết từ giun móc trưởng
thành của các loài A. caninum, A. ceylanicum và
N. americanus đã được định loài bằng hình thái
học. Kết quả thể hiện ở hình 1A cho thấy với
mẫu A. ceylanicum chỉ cho 1 dải băng duy nhất
có kích thước như dự đoán ở vị trí 983 bp, với
mẫu A. caninum thu được nhiều dải băng không
đặc hiệu và không có kết quả dương tính nào với
mẫu N. americanus. Như vậy, phản ứng KOD-
PCR sử dụng cặp mồi HPo1 và HPo2 đảm bảo
tính đặc hiệu khi phân loại loài giun
móc A. ceylanicum.
Kết quả phân lập ấu trùng giun móc truyền
lây A. ceylanicum từ chó và người bằng KOD-
PCR sử dụng cặp mồi đặc hiệu nhân phân đoạn
gen ty thể mt cox1 (983bp) được thể hiện ở hình
1B. Kết quả điện di cho thấy một dải băng sáng
rõ nét duy nhất ở mỗi làn chạy có kích thước
983 bp. Như vậy, trong các mẫu ấu trùng giun
móc thu được từ chó và người, chúng tôi đều
phát hiện sự tồn tại và phân lập thành công loài
giun móc chó truyền lây A. ceylanicum trên cả 2
vật chủ (người và chó).
3.2. Tách dòng gen aspartyl protease của
Ancylostoma ceylanicum bằng vector tách
dòng TA cloning
Với các mẫu DNA tổng số của ấu trùng giun
móc A. ceylanicum sau khi định danh bằng
phản ứng KOD-PCR, chúng tôi tiếp tục sử dụng
làm mạch khuôn nhằm nhân phân đoạn gen
aspartic protease mục tiêu có kích thước khoảng
810 bp (Hình 2).
Sau khi tinh sạch, 17 sản phẩm PCR nhân
phân đoạn gen mã hóa enzyme aspartyl
protease của A. ceylanicum phân lập từ 11 mẫu
ấu trùng giun móc từ chó và 6 mẫu từ người
được lựa chọn gắn với vector tách dòng TOPO-
TA cloning và biến nạp vào tế bào khả biến
E. coli DH5α. Ít nhất 12 khuẩn lạc dương tính
(màu trắng) của từng mẫu được lựa chọn ngẫu
nhiên (Hình 3A), kiểm tra bằng Colony PCR
(Hình 3B) và tiến hành giải trình tự gen.
3.3. Tính đa hình đơn nucleotide trong
phân đoạn gen mã hóa enzyme aspartyl
protease của Ancylostoma ceylanicum
Sau khi phân tích kết quả giải trình tự gen
và BLAST trên NCBI, chúng tôi xác định được 22
điểm SNPs trên phân đoạn gen mã hóa enzyme
aspartyl protease của loài giun móc chó truyền
lây A. ceylanicum phân lập tại Việt Nam.
Kết quả ở bảng 1 cho thấy số lượng SNPs ở
vùng Exon 8 là nhiều nhất, tuy nhiên các đột
biến điểm này đều là đột biến điểm đồng nghĩa
và không hề ảnh hưởng tới sự sai khác về trình
tự axit-amin của aspartic protease. Trong khi
đó, 3 đột biến điểm (1 SNP tại vùng Exon 7 và 2

Tính đa hình đơn nucleotide trong phân đoạn gen mã hóa enzyme cathepsin D aspartic protease của loài giun móc truyền lây Ancylostoma ceylanicum
200
SNPs tại vùng Exon 9) đều dẫn đến sự thay đổi
về axit amin. Cụ thể, codon ACA mã hóa
Threonine tại vùng Exon 7 chuyển thành bộ ba
ATA mã hóa Isoleucine (Hình 4). Tại vùng Exon
9, CTC mã hóa Leucine thành CCC mã hóa
Proline và CCT mã hóa Proline chuyển thành
TCT mã hóa Serine (Hình 5). Tính đa hình đơn
SNPs trên một số vùng gen chức năng khác của
loài giun móc cũng đã được nghiên cứu như
beta-tubulin gen mã hóa protein beta-tubulin,
một loại protein quan trọng tham gia vào quá
trình phân chia tế bào. Cơ chế của các loại thuốc
tẩy giun phổ biến như albendazole hay
mebendazole là kìm hãm quá trình hình thành
thoi vô sắc từ beta-tubulin trong thời kỳ phân
bào, do vậy tiêu diệt được giun móc (Kwa et al.,
1993; Kwa et al., 1994). Tuy nhiên, các đột biến
điểm ở vị trí F167Y hoặc F200Y trên vùng gen
mã hóa beta-tubulin trên loài giun móc Necator
americanus được xác định là marker chỉ thị cho
sự kháng thuốc tẩy giun. Sự tồn tại những đột
biến điểm này trong kiểu gen của giun móc đã
được ghi nhận ở Ghana (Ambrose et al., 2018),
Bzazil (Furtado et al., 2014) và Kenya (Diawara
et al., 2013). Một nghiên cứu của Barry et al.
(2006) về phân tích tính đa dạng trong kiểu gen
mã hóa enzyme aspartyl protease của ký sinh
trùng sốt rét Plasmodium falciparum cũng đã
chỉ ra nhiều điểm SNPs trên bề mặt cấu trúc
của protein này và có thể liên quan đến sự thay
đổi vai trò đặc tính của enzyme. Thí nghiệm
trên chuột cho thấy kháng thể kháng enzyme
aspartyl protease có thể làm giảm 16-26% số
lượng ấu trùng L3 xâm nhập và di hành qua da
chuột (Williamson et al., 2003b). Theo Loukas et
al. (2005), vacxin chế tạo từ protein aspatyl
protease tái tổ hợp sử dụng thử nghiệm đã làm
giảm số lượng giun móc trong ruột chó và giảm
số lượng trứng bài thải ra môi trường so với
nhóm đối chứng, đồng thời làm giảm mất máu
và các biến đổi bệnh tích ở ruột của chó. Tuy
nhiên, những nghiên cứu trong và ngoài nước về
tính đa hình trong kiểu gen cathepsin D
aspartic protease của giun móc, về đột biến
điểm liên quan đến sự thay đổi cấu trúc và chức
năng của loại enzyme đó, về ảnh hưởng của sự
thay đổi này đến hiệu quả của vacxin phòng
bệnh còn rất hạn chế.
Ghi chú: M: thang chuẩn DNA
Hình 2. Ảnh điện di sản phẩm PCR nhân phân đoạn gen mã hóa enzyme aspartyl protease
của A. ceylanicum trên gel Agarose 1,5%
Bảng 1. Số lượng đột biến điểm (SNPs), đột biến điểm đồng nghĩa
và đột biến điểm không đồng nghĩa phát hiện trên vùng Exon 7, Exon 8 và Exon 9
của phân đoạn gen mã hóa enzyme aspartyl protease của A. ceylanicum
Exon 7 Exon 8 Exon 9
SNPs: 7 11 4
Đột biến điểm đồng nghĩa (synonymous SNPs) 6 11 2
Đột biến điểm không đồng nghĩa (non-synonymous SNPs) 1 2
Dương Đức Hiếu, Bùi Khánh Linh, Nguyễn Thu Hương, Lê Đức Vĩnh, Nguyễn Thị Hoàng Yến, Nguyễn Thị Hồng Chiên, Nguyễn Văn Phương
201
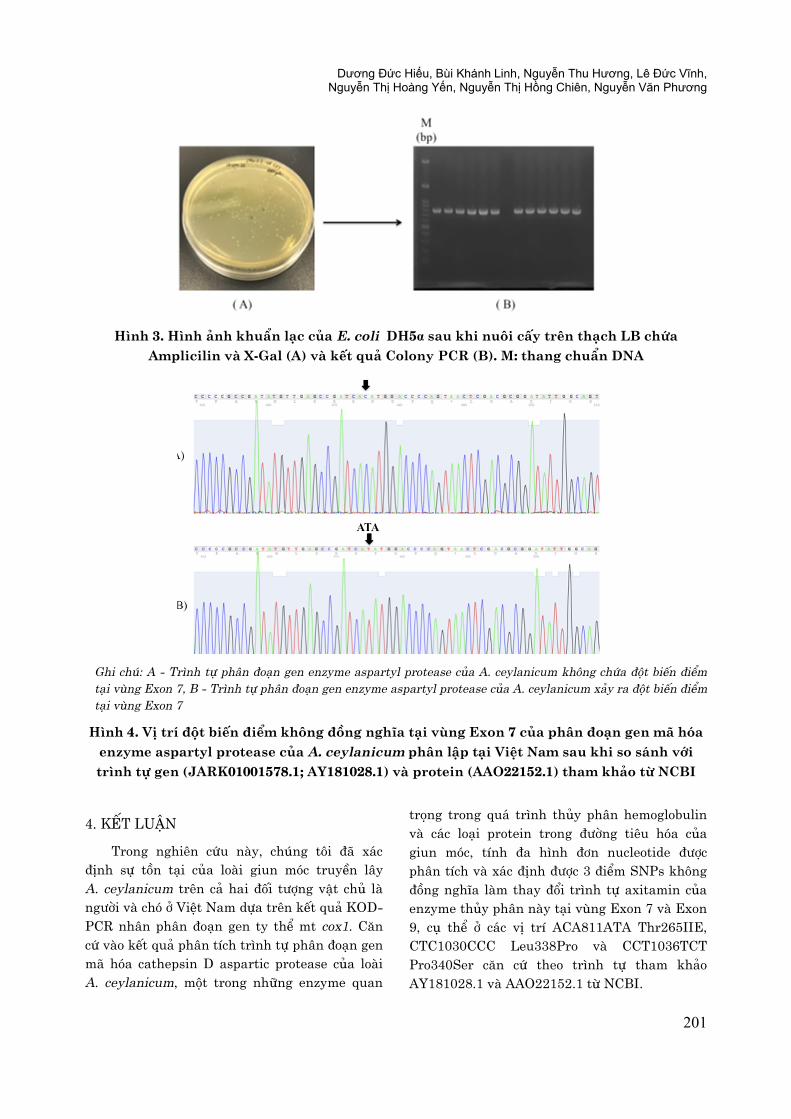
Hình 3. Hình ảnh khuẩn lạc của E. coli DH5α sau khi nuôi cấy trên thạch LB chứa
Amplicilin và X-Gal (A) và kết quả Colony PCR (B). M: thang chuẩn DNA
Ghi chú: A - Trình tự phân đoạn gen enzyme aspartyl protease của A. ceylanicum không chứa đột biến điểm
tại vùng Exon 7, B - Trình tự phân đoạn gen enzyme aspartyl protease của A. ceylanicum xảy ra đột biến điểm
tại vùng Exon 7
Hình 4. Vị trí đột biến điểm không đồng nghĩa tại vùng Exon 7 của phân đoạn gen mã hóa
enzyme aspartyl protease của A. ceylanicum phân lập tại Việt Nam sau khi so sánh với
trình tự gen (JARK01001578.1; AY181028.1) và protein (AAO22152.1) tham khảo từ NCBI
4. KẾT LUẬN
Trong nghiên cứu này, chúng tôi đã xác
định sự tồn tại của loài giun móc truyền lây
A. ceylanicum trên cả hai đối tượng vật chủ là
người và chó ở Việt Nam dựa trên kết quả KOD-
PCR nhân phân đoạn gen ty thể mt cox1. Căn
cứ vào kết quả phân tích trình tự phân đoạn gen
mã hóa cathepsin D aspartic protease của loài
A. ceylanicum, một trong những enzyme quan
trọng trong quá trình thủy phân hemoglobulin
và các loại protein trong đường tiêu hóa của
giun móc, tính đa hình đơn nucleotide được
phân tích và xác định được 3 điểm SNPs không
đồng nghĩa làm thay đổi trình tự axitamin của
enzyme thủy phân này tại vùng Exon 7 và Exon
9, cụ thể ở các vị trí ACA811ATA Thr265IIE,
CTC1030CCC Leu338Pro và CCT1036TCT
Pro340Ser căn cứ theo trình tự tham khảo
AY181028.1 và AAO22152.1 từ NCBI.

Tính đa hình đơn nucleotide trong phân đoạn gen mã hóa enzyme cathepsin D aspartic protease của loài giun móc truyền lây Ancylostoma ceylanicum
202
Ghi chú: A - Trình tự phân đoạn gen enzyme aspartyl protease của A. ceylanicum không chứa đột biến điểm
tại vùng Exon 9, B - Trình tự phân đoạn gen enzyme aspartyl protease của A. ceylanicum xảy ra đột biến điểm
tại vùng Exon 9
Hình 5. Các vị trí đột biến điểm không đồng nghĩa tại vùng Exon 9 của phân đoạn gen mã
hóa enzyme aspartyl protease của A. ceylanicum phân lập tại Việt Nam sau khi so sánh với
trình tự gen (JARK01001578.1; AY181028.1) và protein (AAO22152.1) tham khảo từ NCBI
TÀI LIỆU THAM KHẢO
Ambrose R.O., Josephine E.Q., Peter S., Santosh G., Lisa M.H., Fabio P.D., Benjamin E., Adalgisa C., Debbie H., Michael D.W. & Michael C. (2018). Genetic markers of benzimidazole resistance among human hookworms (Necator americanus) in Kintampo North Municipality, Ghana. The American Journal of Tropical Medicine and Hygiene. https://doi.org/10.4269/ajtmh.18-0727.
Barry A.E., Leliwa-Sytek A., Man K., Kasper J.M., Hartl D.L. & Day K.P. (2006). Variable SNP density in aspartyl-protease genes of the malaria parasite Plasmodium falciparum. Gene. 376(2): 163-173.
Brown A., Girod N., Billett E.E. & Pritchard D.I. (1999). Necator americanus (human hookworm) aspartyl proteinases and digestion of skin macromolecules during skin penetration. Am J Trop Med Hyg. 60: 840-847.
Bowman D.D., Georgi J.R. & Saunders. (2009). Georgis’ Parasitology for Veterinarians. 10th ed., Elsevier.
Conlan J.V., Khamlome B., Vongxay K., Elliot A., Pallant L., Sripa B., Blacksell S.D., Fenwick S. & Thompson R.C. (2012). Soil-transmitted helminthiasis in Laos: a community-wide cross-sectional study of humans and dogs in a mass drug administration environment. The American Journal of Tropical Medicine and Hygiene. 86: 624-34.

Dương Đức Hiếu, Bùi Khánh Linh, Nguyễn Thu Hương, Lê Đức Vĩnh, Nguyễn Thị Hoàng Yến, Nguyễn Thị Hồng Chiên, Nguyễn Văn Phương
203
Diawara A., Halpenny C.M., Churcher T.S., Mwandawiro C., Kihara J., Kaplan R.M., Streit T.G., Idaghdour Y., Scott M.E., Basáñez M.G. & Prichard R.K. (2013). Association between response to albendazole treatment and β-tubulin genotype frequencies in soil-transmitted helminthes. PLOS Neglected Tropical Diseases. 7(5): 2247.
Dinh N.N., Sze F.H., Van-Anh T.N., Trong V.N., Dien V.N. & Rebecca J.T. (2015). Re-evaluation of the species of hookworms infecting dogs in Central Vietnam. Parasites & Vectors. 8: 401.
Duong D. H., Bui K. L., Nguyen T. H., Tran T. D., Eiji N., Haruhiko M., Ayako Y. & Nariaki N. (2018). Phylogenetic relationship between Ancylostoma ceylanicum populations found in dogs and humans in Vietnam. Vietnam Journal of Infectious Diseases. 6: 53.
Furtado L. F., Bello A.C., Dos Santos H.A., Carvalho M.R., Rabelo E.M. (2014). First identification of the F200Y SNP in the β-tubulin gene linked to benzimidazole resistance in Ancylostoma caninum? Veterinary Parasitology. 206 (3-4): 313-316.
Hotez P.J., Brooker S., Bethony J.M., Bottazzi M.E., Loukas A. & Xiao S. (2004). Hookworm infection, The New England Journal of Medicine. 351: 799-807.
Inpankaew T., Schar F., Dalsgaard A., Khieu V., Chimnoi W., Chhoun C., Sok D., Marti H., Muth S., Odermatt P. & Traub R.J. (2014). High prevalence of Ancylostoma ceylanicum hookworm infections in humans, Cambodia. Emerging Infectious Diseases. 20: 976-82.
Jolodar A., Fischer P., Buttner D.W., Miller D.J., Schmetz C. & Brattig N.W. (2004). Onchocerca volvulus: expression and immunolocalization of a nematode cathepsin D-like lysosomal aspartic protease. Experimental Parasitology. 107: 145-156.
Kwa M.S., Kooyman F.N., Boersema J.H. & Roos M.H. (1993). Effect of selection for benzimidazole resistance in Haemonchus contortus on beta-tubulin isotype 1 and isotype 2 genes Biochemical and Biophysical Research Communications. 191: 413-419.
Kwa M.S., Veenstra J.G. & Roos M.H. (1994). Benzimidazole resistance in Haemonchus contortus is correlated with a conserved mutation at amino acid 200 in beta-tubulin isotype 1. Molecular and Biochemical Parasitology. 63: 299-303.
Loukas A., Bethony J.M., Mendez S., Fujiwara R.T., Goud G.N., Ranjit N., Zhan B., Jones K., Bottazzi M.E. & Hotez P.J. (2005). Vaccination with recombinant aspartic hemoglobinase reduces parasite load and blood loss after hookworm infection in dogs. PLOS Medicine. 2: 295.
Ngui R., Lim Y.A., Traub R., Mahmud R. & Mistam M.S. (2012a). Epidemiological and genetic data supporting the transmission of Ancylostoma ceylanicum among human and domestic animals. PLOS Neglected Tropical Diseases. 6: 1522.
Ngui R., Ching L.S., Kai T.T., Roslan M.A., Lim Y.A. (2012b). Molecular identification of human hookworm infections in economically disadvantaged communities in Peninsular Malaysia. The American Journal of Tropical Medicine and Hygiene. 86: 837-42.
Traub R.J., Inpankaew T., Sutthikornchai C., Sukthana Y. & Thompson R.C. (2008). PCR-based coprodiagnostic tools reveal dogs as reservoirs of zoonotic ancylostomiasis caused by Ancylostoma ceylanicum in temple communities in Bangkok. Veterinary Parasitology. 155: 67-73.
Williamson A.L., Brindley P.J., Abbenante G., Prociv P., Berry C., Girdwood K., Pritchard D.I., Fairlie D.P., Hotez P.J., Dalton J.P. & Loukas A. (2002). Cleavage of hemoglobin by hookworm cathepsin D aspartic proteases and its potential contribution to host specificity. The FASEB Journal. 16: 1458-1460.
Williamson A.L., Brindley P.J., Abbenante G., Datu B.J., Prociv P., Berry C., Girdwood K., Pritchard D.I, Fairlie D.P., Hotez P.J., Zhan B. & Loukas A. (2003a). Hookworm aspartic protease, Na-APR-2, cleaves human hemoglobin and serum proteins in a host-specific fashionThe Journal of Infectious Diseases. 187: 484-494.
Williamson A.L., Brindley P.J. & Loukas A. (2003b). Hookworm cathepsin D aspartic proteases: contributing roles in the host-specific degradation of serum proteins and skin macromolecules. Parasitology. 126: 179-185.
Williamson A. L., Paolo Lecchi, Bejamin E.T, Youngchol C., Peter J.H., James H.K, Lewwis C.K., Mohammed S., Charles S.C. & Alex L. (2004). A multi-enzyme cascade of hemoglobin proteolysis in the intestine of blood-feeding hookworms. Journal of Biological Chemistry. 279: 35950-35957.




























