Embed Size (px)
Citation preview

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2013 669
TOKSISITAS CENDAWAN Beauveria bassiana Vuill. (Balsamo) TERHADAP TELUR DAN LARVA PENGGEREK UBIJALAR
Cylas formicarius (Coleoptera: Curculionidae)
Yusmani Prayogo Balai Penelitian Tanaman Aneka Kacang dan Umbi
Jl. Raya Kendalpayak KM 08, Kotak Pos 66 Malang, 65101 E-mail: [email protected]
ABSTRAK Cylas formicarius merupakan hama utama pada ubijalar yang dapat menyebabkan
kehilangan hasil mencapai 100%. Usaha pengendalian yang dilakukan petani yaitu aplikasi insektisida kimia, namun belum diperoleh hasil yang memuaskan. Penelitian ini bertujuan untuk mempelajari toksisitas cendawan Beauveria bassiana terhadap telur dan larva C. formicarius. Penelitian dilakukan di laboratorium Entomologi, Balai Penelitian Tanaman Kacang-kacangan dan Umbi-umbian (Balitkabi). Percobaan disusun menggunakan rancangan acak lengkap, setiap perlakuan diulang lima kali. Perlakuan adalah berbagai umur telur dan berbagai stadia larva C. formicarius. Toksisitas cendawan dihitung berdasarkan persentase telur yang tidak menetas dan jumlah larva yang mati terinfeksi cendawan B. bassiana. Hasil penelitian menunjukkan bahwa toksisitas cendawan B. bassiana ditentukan oleh umur telur dan umur stadia larva. Cendawan B. bassiana sangat toksik sehingga mampu menggagalkan penetasan telur C. formicarius mencapai 100%, terutama telur yang baru diletakkan imago (0-4 hari). Aplikasi cendawan B. bassiana pada telur C. formicarius yang berumur lima sampai dengan tujuh hari juga toksik karena masih mampu membunuh telur berkisar dari 43-74%. Selain itu, telur C. formicarius yang terinfeksi cendawan B. bassiana mengalami terlambat menetas hingga mencapai enam hari setelah aplikasi. Cendawan B. bassiana juga toksik terhadap seluruh stadia larva C. formicarius, yaitu mulai stadia larva instar I sampai dengan stadia larva instar V. Hasil penelitian ini dapat disimpulkan bahwa cendawan B. bassiana sangat toksik terhadap stadia telur maupun larva C. formicarius. Dengan demikian, cendawan tersebut dapat digunakan sebagai salah satu alternatif untuk mengurangi pestisida kimia dalam mengendalikan penggerek ubijalar C. formicarius.
Kata kunci: toksisitas, B. bassiana, telur, larva, C. formicarius.
ABSTRACT Toxicity of the Beauveria bassiana against eggs and larva of Cylas formicarius.
Cylas formicarius is a major pest on sweet potato which cause yield losses of up to 100%. Control technology were used by farmers, namely the application of chemical insecticides, but has not been recorded optimal results. The research to study the toxicity of the Beauveria bassiana against eggs and larva of C. formicarius. The research was conducted in the laboratory of Entomology, Indonesian Legumes and Tuber Crops Research Institute (Iletri). The expe-riments were used complete randomized design, each treatment was repeated five times. Treatment was various ages eggs and various stage C. formicarius. Toxicity of fungi was calcu-lated based on the number of eggs that did not hatching and the mortality of larvae infected by fungus. The results showed that the toxicity of B. bassiana is determined by the age of the eggs C. formicarius. The fungus was highly toxic in suppressing hatching egg of C. formicarius reaches 100%, especially the newly laid (0-4 days). Application of B. bassiana was highly toxic to eggs C. formicarius of five to seven days age caused kill eggs between 43-74%. In addition, eggs C. formicarius were infected by fungus indicate later hatching up to six days after

Prayogo: Toksisitas Beauveria bassiana terhadap telur Cylas formicarius 670
application. The fungus also toxic to all larvae stages of C. formicarius included of I to V larvae stage. The results of this study that B. bassiana is highly toxic to eggs and larvae of C. formicarius. Therefore, the fungus can be used as an alternative to substitute chemical pesticides to control of sweet potato borer C. formicarius.
Keywords: toxicity, B. bassiana, eggs, larvae, C. formicarius.
PENDAHULUAN Penggerek ubijalar atau disebut dengan hama boleng (Cylas formicarius) merupakan
salah satu hama utama pada ubijalar di berbagai negara termasuk di Indonesia (Kandori et al. 2006; Sharah et al. 2012; Reddy et al. 2012). Kerugian hasil yang disebabkan oleh hama ini hingga mencapai 100%, sementara itu umbi yang sudah tergerek tidak layak untuk dikonsumsi manusia karena dapat meracuni atau menyebabkan penyakit kanker. Teknologi pengendalian hama penggerek umbi menggunakan insektisida kimia yang biasa digunakan petani hingga saat ini belum mampu menunjukkan hasil yang signifikan (Chira-njeevi et al. 2002; Smith & Hammond 2006; Kumano et al. 2007; Leng & Reddy 2012). Hal ini disebabkan stadia serangga yang merusak adalah larva, dan stadia tersebut berkembang di dalam umbi sehingga bahan aktif insektisida sintesis yang diaplikasikan tidak mengenai serangga sasaran. Berbagai upaya untuk menanggulangi hama boleng terus dikembangkan, salah satunya adalah pengendalian biologis menggunakan cendawan entomopatogen (Yasuda et al. 2000; Ondiaka et al. 2008; Rosfiansyah 2009; Ratissa 2011; Komi 2011; Ahdiaty 2013).
Beauveria bassiana merupakan salah satu jenis cendawan entomopatogen yang mampu menginfeksi hampir semua ordo dan berbagai stadia serangga sehingga cukup prospektif digunakan sebagai alternatif untuk mengurangi insektisida kimia karena berbagai kelebihannya (Kirkland et al. 2004; Batta 2007; Meyling & Eilenberg 2007; Kafie et al. 2011). Hasil eksplorasi terhadap berbagai isolat cendawan B. bassiana telah dilakukan oleh Prayogo (2010) di berbagai lokasi di pulau Jawa telah diperoleh tiga isolat yang cukup efektif karena mampu membunuh imago, larva maupun menggagalkan pene-tasan telur hingga mencapai 100%. Penelitian lebih lanjut mengindikasikan bahwa tiga isolat B. bassiana yang diperoleh dari Bogor, Probolinggo dan Tumpang (Malang) memi-liki keampuhan yang lebih tinggi dalam membunuh larva maupun imago dibandingkan dengan isolat yang diperoleh dari lokasi lain (Prayogo 2010). Hal ini mengindikasikan bahwa perbedaan asal isolat memiliki keragaman virulensi atau keampuhan yang berbeda (Bidochka et al. 2002; Thakur et al. 2005; Cruz et al. 2006; Takatsuka 2007; Sevim et al. 2012).
Hasil aplikasi cendawan entomopatogen B. bassiana pada umbi mampu melindungi kerusakan umbi dari serangan larva hama penggerek umbi karena larva yang terinfeksi cendawan B. bassiana tidak mampu merusak umbi dan telur yang terinfeksi cendawan tersebut tidak mampu menetas. Hal ini disebabkan cendawan B. baasiana mampu meng-infeksi berbagai jenis serangga maupun berbagai stadia hama mulai stadia telur, larva maupun imago (El-Sinary 2002; Gayathri et al. 2010). Oleh karena itu, pengendalian hama boleng menggunakan cendawan entomopatogen B. bassiana mempunyai prospek yang baik dan dapat dikembangkan karena sangat toksik terhadap hama sasaran dan aman terhadap serangga berguna serta tidak mencemari lingkungan (Broza et al. 2001; Thungrabeab & Tongma 2007; Zimmermann 2007; Meikle et al. 2008; Costa et al. 2010).

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2013 671
Pengendalian hama penggerek umbi menggunakan agens hayati cendawan entomo-patogen B. bassiana di Indonesia belum berkembang pesat. Hal ini disebabkan petani masih lemah dalam mengidentifikasi jenis dan bioekologi hama. Sementara itu, jenis dan bioekologi hama sangat berkaitan dengan keberhasilan teknologi pengendalian yang dilakukan. Setiap jenis cendawan memiliki kisaran inang yang berbeda, selain itu setiap stadia serangga memiliki kerentanan yang berbeda pula terhadap toksisitas cendawan. Penelitian ini bertujuan untuk mempelajari toksisitas cendawan B. bassiana terhadap stadia telur dan larva hama penggerek ubijalar.
BAHAN DAN METODE Penelitian dilakukan di laboratorium hama dan penyakit, Balai Penelitian Tanaman
Kacang-kacangan dan Umbi-umbian (Balitkabi) mulai bulan Januari sampai dengan Juli 2012. Penelitian disusun menggunakan rancangan acak lengkap, ulangan lima kali. Perlakuan umur telur C. formicarius setelah diletakkan imago yaitu; (1) 0 hari (baru diletakkan imago), (2) satu hari setelah diletakkan imago (SDI), (3) dua hari SDI, (4) tiga hari SDI, (5) empat hari SDI, (6) lima hari SDI, (7) enam hari SDI, (8) tujuh hari SDI, dan (9) kontrol (tanpa aplikasi cendawan B. bassiana). Perlakuan larva C. formicarius adalah sebagai berikut; (1) larva instar I, (2) larva instar II, (3) larva instar III, (4) larva instar IV, dan (5) larva instar V.
Pembiakan Serangga C. formicarius untuk Mendapatkan Telur dan Larva
Imago penggerek ubijalar diperoleh dari eksplorasi beberapa umbi ubijalar yang rusak di Pasar Gadang (Malang) pada bulan Desember 2011. Umbi-umbi yang rusak dima-sukkan ke dalam kotak plastik yang berukuran tinggi 30 cm dan berdiameter 20 cm. Setiap kotak plastik diisi dua buah umbi ubijalar yang sudah rusak tergerek kemudian ditambah dua buah umbi yang masih segar dengan tujuan sebagai calon makanan bagi larva mau-pun imago. Kotak plastik ditutup menggunakan kain kasa dengan tujuan agar imago yang dipelihara tidak keluar. Pemeliharaan serangga dipertahankan terus menerus hingga mem-peroleh jumlah serangga yang cukup berlimpah. Pemeliharaan serangga ini ditujukan untuk memperoleh kelompok telur yang berbagai umur dengan jumlah yang memadai sebagai bahan perlakuan.
Kelompok telur yang sudah diletakkan oleh imago dipisahkan berdasarkan umur sesuai perlakuan kemudian diletakkan di permukaan umbi dan dimasukkan ke dalam cawan petri berdiameter 16 cm yang telah dilapisi kertas tissue berwarna untuk memudahkan pengamatan dalam membedakan warna telur.
Perbanyakan Cendawan B. bassiana
Cendawan B. bassiana yang digunakan dalam penelitian ini adalah isolat dari Probo-linggo hasil eksplorasi pada tahun 2010. Berdasarkan hasil uji virulensi bahwa isolat tersebut memiliki kriteria virulensi sangat tinggi dalam membunuh imago hama penggerek ubijalar. Isolat cendawan ditumbuhkan pada media beras jagung manis yang sudah di-sterilisasi di dalam autoclave selama 45 menit pada temperatur 141 oC. Tujuan penggu-naan beras jagung manis sebagai media tumbuh adalah untuk mendapatkan konidia dalam jumlah yang banyak karena media tumbuh tersebut kaya karbohidrat dan protein.
Beras jagung dibilas dengan air hingga bersih kemudian dimasak/dikukus setengah matang. Setelah itu, dipaparkan hingga kering kemudian dimasukkan ke dalam kantong

Prayogo: Toksisitas Beauveria bassiana terhadap telur Cylas formicarius 672
plastik untuk disterilisasi di dalam autoclave pada suhu 121 oC selama 15 menit. Inokulum suspensi konidia B. Bassiana sebagai starter dengan kerapatan konidia 106/ml diinokulasi-kan pada media tumbuh di dalam kantong plastik menggunakan jarum steril. Biakan cendawan selanjutnya diinkubasi di dalam suhu ruangan dengan temperatur kurang lebih 27 oC. Pada umur 21 hari setelah inokulasi (HSI), biakan cendawan dimasukkan ke dalam erlenmeyer dan ditambah air kurang lebih 100 ml kemudian dikocok menggunakan shaker selama 30 menit untuk merontokkan konidia yang terbentuk. Suspensi konidia yang sudah terkumpul disaring menggunakan kain kasa halus kemudian ditambah Tween 80 sebanyak 2 ml per liter dengan tujuan konidia bercampur homogen karena bersifat hidropobik. Selanjutnya, suspensi konidia dihitung menggunakan haemocytometer yang diamati menggunakan mikroskop untuk memperoleh kerapatan konidia mencapai 107/ml.
Aplikasi Suspensi Konidia B. bassiana pada Telur dan Larva
Telur penggerek ubijalar sebagai perlakuan ditempelkan pada permukaan kulit umbi ubijalar menggunakan kuas halus. Masing-masing umbi ubijalar berisi telur 25 butir selan-jutnya dimasukkan ke dalam cawan Petri yang berdiameter 16 cm. Inokulasi dilakukan dengan cara menyemprotkan suspensi konidia B. bassiana sebanyak 10 ml/25 butir telur pada seluruh permukaan ubijalar yang sudah diinfestasi telur. Sementara itu, larva peng-gerek ubijalar sebagai perlakuan dimasukkan ke dalam toples plastik yang sudah berisi umbi segar kemudian disemprot menggunakan suspensi konidia B. bassiana sebanyak 10 ml/25 ekor larva hingga mengenai seluruh permukaan tubuh larva. Pengamatan
Peubah yang diamati adalah; (1) jumlah telur yang tidak menetas hingga 14 hari setelah aplikasi, (2) waktu penetasan telur setelah aplikasi B. bassiana, (3) jumlah larva yang mati terinfeksi cendawan B. bassiana, dan (4) proses kolonisasi miselium cendawan pada permukaan masing-masing larva yang mati terinfeksi cendawan, serta (5) ketebalan kolonisasi miselium cendawan pada permukaan inang (telur & larva). Analisis Data
Data yang diperoleh dianalisa menggunakan program Minitab 14, selanjutnya apabila terdapat perbedaan diantara perlakuan yang diuji maka dilanjutkan uji jarak berganda (Duncan multiple range test) α=(0,05).
HASIL DAN PEMBAHASAN
Pengaruh Aplikasi Cendawan B. bassiana terhadap Penetasan Telur
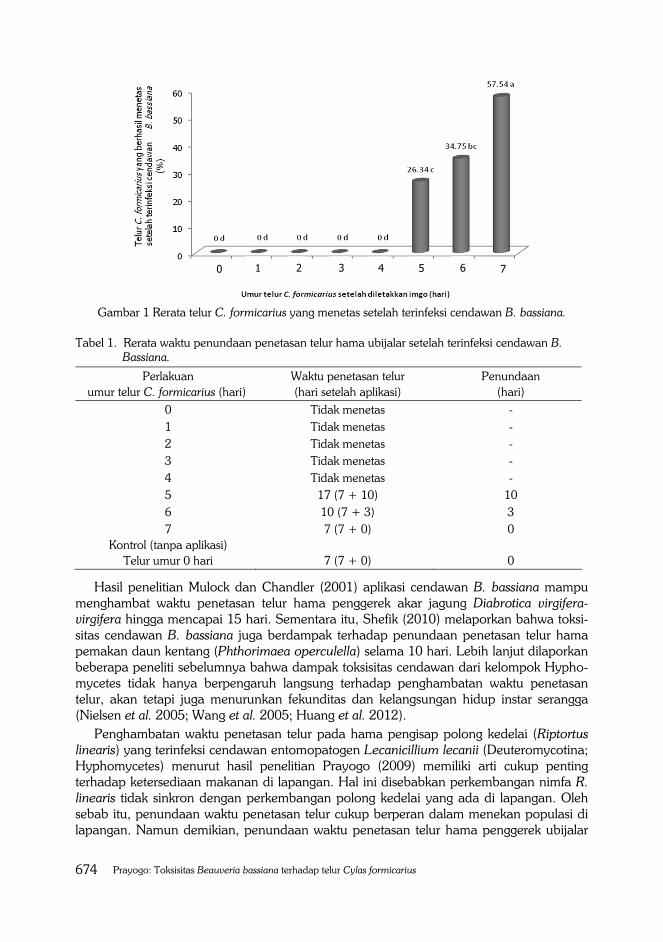
Toksisitas cendawan terhadap telur penggerek ubijalar dinilai dari jumlah telur yang tidak menetas setelah terinfeksi cendawan. Hasil penelitian menunjukkan bahwa semakin lama umur telur penggerek ubijalar diletakkan oleh imago, semakin tahan terhadap infeksi cendawan. Dengan kata lain bahwa cendawan B. bassiana sangat toksik pada telur hama penggerek ubijalar yang baru diletakkan imago. Hasil aplikasi suspensi cendawan pada telur yang baru diletakkan imago (nol hari) mampu membunuh telur hingga mencapai 100% (Gambar 1).
Cendawan entomopatogen B. bassiana juga toksik terhadap telur penggerek ubijalar yang berumur satu sampai dengan empat hari karena telur tersebut juga tidak menetas hingga mencapai 100% dan secara statistik tidak berbeda nyata dengan perlakuan 0 hari.

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2013 673
Oleh karena itu, cendawan B. bassiana sangat toksik terhadap telur penggerek ubijalar yang berumur 0 sampai dengan empat hari sehingga cendawan tersebut memiliki keampuhan yang besar dalam membunuh stadia telur C. formicarius terutama yang baru diletakkan imago. Hasil penelitian ini mengindikasikan bahwa cendawan B. bassiana bersifat ovisidal atau membunuh stadia telur sehingga sangat efektif dalam menekan perkembangan serangga (Shi et al. 2004; Gindin et al. 2006; Shi et al. 2008; Santos et al. 2009; Li et al. 2012).
Aplikasi suspensi cendawan B. bassiana dinilai masih cukup toksik terhadap telur hama penggerek ubijalar yang berumur lima hari karena mampu menekan penetasan telur hingga mencapai 73,66%. Sementara itu, toksisitas cendawan pada telur yang berumur enam dan tujuh hari setelah diletakkan imago masing-masing 34,75% dan 57,54%. Hasil penelitian ini mengindikasikan bahwa cendawan B. bassiana dapat mematikan telur terutama yang baru diletakkan. Tingginya kematian telur yang baru diletakkan disebabkan cendawan ini menghasilkan senyawa enzim protease, lipase dan khitinase yang mampu mendegradasi dinding sel telur terluar (chorion) yang mengandung karbohidrat, khitin dan protein. Sementara itu, Tikhonov et al. (2002) menyatakan bahwa kolagenase merupakan salah satu enzim yang lebih penting selain protease dan kitinase. Enzim tersebut biasanya berperan dalam mendegradasi komposisi struktur kulit telur maupun integumen inang (Shinya et al. 2008). Menurut hasil penelitian Carneiro et al. (1999) dan Regaieg et al. (2010) cendawan B. bassiana menghasilkan berbagai jenis senyawa enzim maupun toksin yang dapat mendegradasi dinding sel telur serangga Anocentor nitens (Neumann) (Acari: Ixodidae) sehingga telur serangga tersebut tidak mempunyai peluang untuk menetas.
Konnstantopoulou dan Mazomenos (2004), Wang et al. (2005), Prasad dan Veerwal (2010) melaporkan juga bahwa cendawan B. bassiana menghasilkan senyawa toksin yang bersifat toksik terhadap syaraf serangga sehingga menyebabkan sistem syaraf terganggu yang diakhiri dengan kematian serangga. Laporan dari beberapa peneliti lain juga menun-jukkan bahwa cendawan entomopatogen yang termasuk dalam kelompok Hyphomycetes mampu menghasilkan senyawa toksin bassianolid, bassiacridin, oosperin dan cyclosporin yang dapat membunuh stadia telur maupun larva berbagai jenis serangga (Tikhonov et al. 2002; Mukhtar & Pervaz 2003; Vega et al. 2008). Ditinjau dari berbagai keunggulan cendawan tersebut maka B. bassiana berpeluang besar dapat digunakan sebagai salah satu alternatif untuk agens pengendalian hayati terhadap C. formicarius dan sebagai alternatif pengganti insektisida kimia.
Hasil penelitian ini menunjukkan bahwa waktu penetasan telur penggerek ubijalar yang terinfeksi cendawan B. bassiana mengalami penundaan berkisar tiga sampai dengan 10 hari jika dibandingkan dengan kontrol (Tabel 1). Hasil pengamatan mengindikasikan bahwa telur yang berumur lima dan enam hari yang tertunda waktu penetasannya yaitu masing-masing tiga dan 10 hari. Sementara itu, telur yang berumur tujuh hari tidak mengalami penundaan waktu penetasan karena telur tersebut memang sudah waktunya menetas seperti pada perlakuan kontrol (tanpa aplikasi cendawan B. bassiana). Telur yang baru diletakkan (no lhari) hingga berumur empat hari tidak menetas 100% sehingga tidak ada yang dapat diamati waktu penetasan telurnya. Dengan demikian, cendawan B. bassiana selain toksik juga mampu menghambat waktu penetasan telur hama penggerek ubijalar.
Prayogo: Toksisitas Beauveria bassiana terhadap telur Cylas formicarius 674
Gambar 1 Rerata telur C. formicarius yang menetas setelah terinfeksi cendawan B. bassiana.
Tabel 1. Rerata waktu penundaan penetasan telur hama ubijalar setelah terinfeksi cendawan B. Bassiana.
Perlakuan umur telur C. formicarius (hari)
Waktu penetasan telur (hari setelah aplikasi)
Penundaan (hari)
0 Tidak menetas - 1 Tidak menetas - 2 Tidak menetas - 3 Tidak menetas - 4 Tidak menetas - 5 17 (7 + 10) 10 6 10 (7 + 3) 3 7
Kontrol (tanpa aplikasi) Telur umur 0 hari
7 (7 + 0)
7 (7 + 0)
0 0
Hasil penelitian Mulock dan Chandler (2001) aplikasi cendawan B. bassiana mampu menghambat waktu penetasan telur hama penggerek akar jagung Diabrotica virgifera-virgifera hingga mencapai 15 hari. Sementara itu, Shefik (2010) melaporkan bahwa toksi-sitas cendawan B. bassiana juga berdampak terhadap penundaan penetasan telur hama pemakan daun kentang (Phthorimaea operculella) selama 10 hari. Lebih lanjut dilaporkan beberapa peneliti sebelumnya bahwa dampak toksisitas cendawan dari kelompok Hypho-mycetes tidak hanya berpengaruh langsung terhadap penghambatan waktu penetasan telur, akan tetapi juga menurunkan fekunditas dan kelangsungan hidup instar serangga (Nielsen et al. 2005; Wang et al. 2005; Huang et al. 2012).
Penghambatan waktu penetasan telur pada hama pengisap polong kedelai (Riptortus linearis) yang terinfeksi cendawan entomopatogen Lecanicillium lecanii (Deuteromycotina; Hyphomycetes) menurut hasil penelitian Prayogo (2009) memiliki arti cukup penting terhadap ketersediaan makanan di lapangan. Hal ini disebabkan perkembangan nimfa R. linearis tidak sinkron dengan perkembangan polong kedelai yang ada di lapangan. Oleh sebab itu, penundaan waktu penetasan telur cukup berperan dalam menekan populasi di lapangan. Namun demikian, penundaan waktu penetasan telur hama penggerek ubijalar
0 1 2 3 4 5 6 7
Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2013 675
yang terinfeksi cendawan B. bassiana perlu diteliti lebih detail dampak positifnya terhadap perkembangan hama di lapangan. Hal ini disebabkan belum banyak diketahui secara pasti penelitian tersebut karena penelitian pengendalian hama penggerek ubijalar menggunakan cendawan entomopatogen belum banyak dilaporkan.
Mortalitas Larva Hama Penggerek Ubijalar
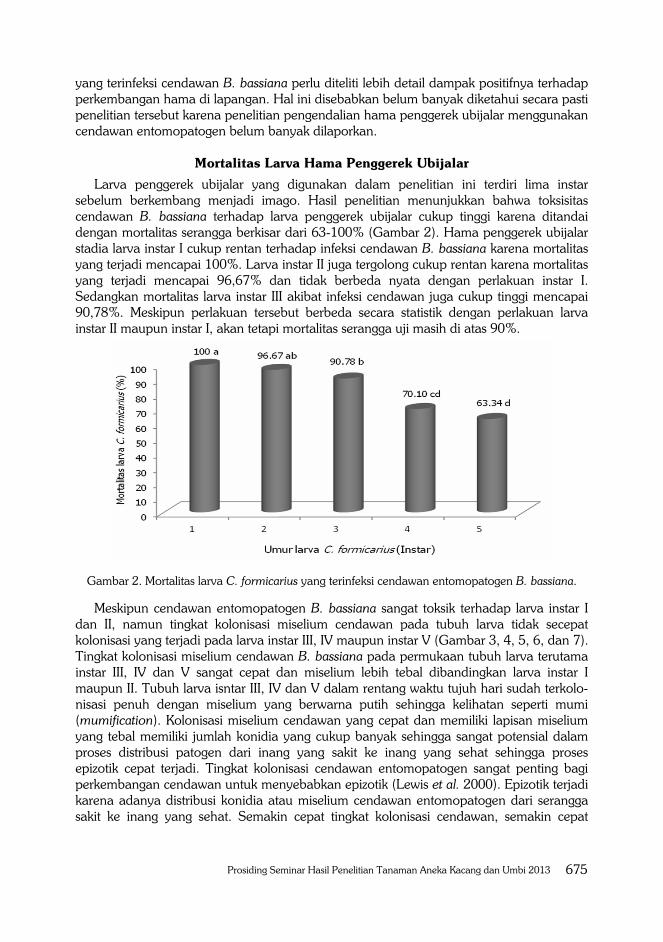
Larva penggerek ubijalar yang digunakan dalam penelitian ini terdiri lima instar sebelum berkembang menjadi imago. Hasil penelitian menunjukkan bahwa toksisitas cendawan B. bassiana terhadap larva penggerek ubijalar cukup tinggi karena ditandai dengan mortalitas serangga berkisar dari 63-100% (Gambar 2). Hama penggerek ubijalar stadia larva instar I cukup rentan terhadap infeksi cendawan B. bassiana karena mortalitas yang terjadi mencapai 100%. Larva instar II juga tergolong cukup rentan karena mortalitas yang terjadi mencapai 96,67% dan tidak berbeda nyata dengan perlakuan instar I. Sedangkan mortalitas larva instar III akibat infeksi cendawan juga cukup tinggi mencapai 90,78%. Meskipun perlakuan tersebut berbeda secara statistik dengan perlakuan larva instar II maupun instar I, akan tetapi mortalitas serangga uji masih di atas 90%.
Gambar 2. Mortalitas larva C. formicarius yang terinfeksi cendawan entomopatogen B. bassiana.
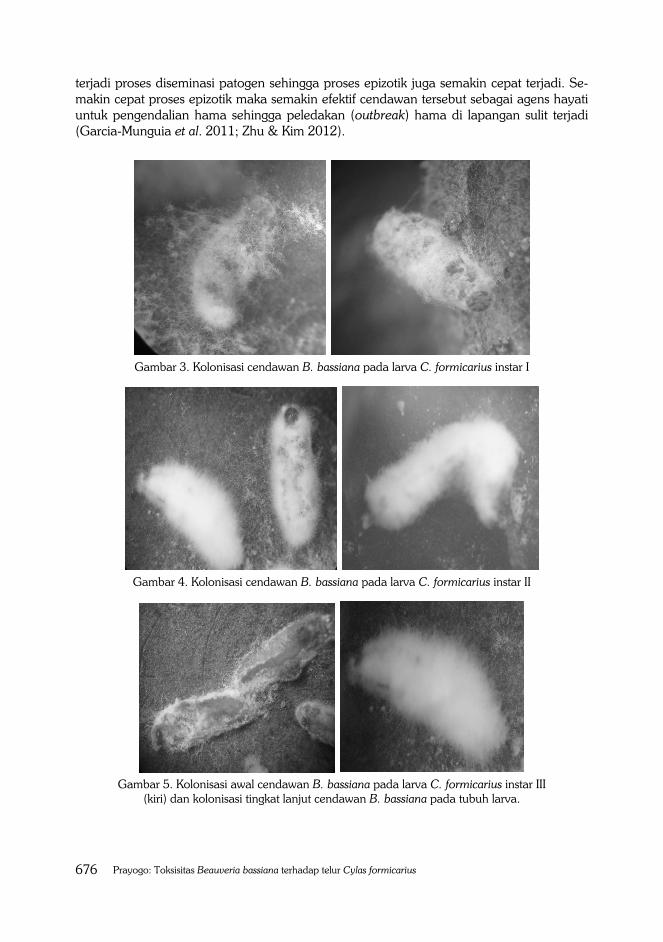
Meskipun cendawan entomopatogen B. bassiana sangat toksik terhadap larva instar I dan II, namun tingkat kolonisasi miselium cendawan pada tubuh larva tidak secepat kolonisasi yang terjadi pada larva instar III, IV maupun instar V (Gambar 3, 4, 5, 6, dan 7). Tingkat kolonisasi miselium cendawan B. bassiana pada permukaan tubuh larva terutama instar III, IV dan V sangat cepat dan miselium lebih tebal dibandingkan larva instar I maupun II. Tubuh larva isntar III, IV dan V dalam rentang waktu tujuh hari sudah terkolo-nisasi penuh dengan miselium yang berwarna putih sehingga kelihatan seperti mumi (mumification). Kolonisasi miselium cendawan yang cepat dan memiliki lapisan miselium yang tebal memiliki jumlah konidia yang cukup banyak sehingga sangat potensial dalam proses distribusi patogen dari inang yang sakit ke inang yang sehat sehingga proses epizotik cepat terjadi. Tingkat kolonisasi cendawan entomopatogen sangat penting bagi perkembangan cendawan untuk menyebabkan epizotik (Lewis et al. 2000). Epizotik terjadi karena adanya distribusi konidia atau miselium cendawan entomopatogen dari serangga sakit ke inang yang sehat. Semakin cepat tingkat kolonisasi cendawan, semakin cepat
Prayogo: Toksisitas Beauveria bassiana terhadap telur Cylas formicarius 676
terjadi proses diseminasi patogen sehingga proses epizotik juga semakin cepat terjadi. Se-makin cepat proses epizotik maka semakin efektif cendawan tersebut sebagai agens hayati untuk pengendalian hama sehingga peledakan (outbreak) hama di lapangan sulit terjadi (Garcia-Munguia et al. 2011; Zhu & Kim 2012).
Gambar 3. Kolonisasi cendawan B. bassiana pada larva C. formicarius instar I
Gambar 4. Kolonisasi cendawan B. bassiana pada larva C. formicarius instar II
Gambar 5. Kolonisasi awal cendawan B. bassiana pada larva C. formicarius instar III
(kiri) dan kolonisasi tingkat lanjut cendawan B. bassiana pada tubuh larva.
Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2013 677
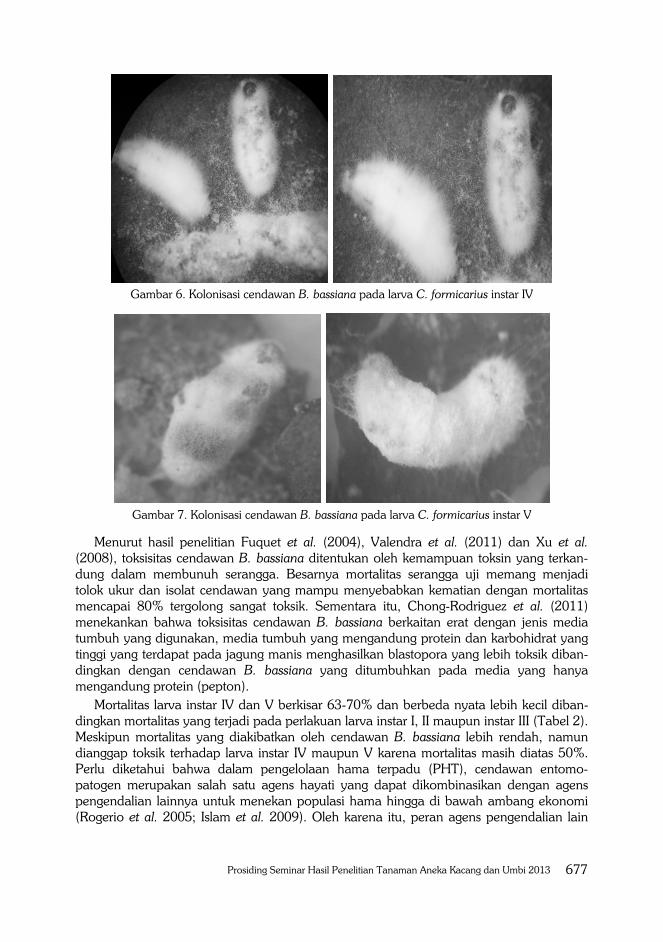
Gambar 6. Kolonisasi cendawan B. bassiana pada larva C. formicarius instar IV
Gambar 7. Kolonisasi cendawan B. bassiana pada larva C. formicarius instar V
Menurut hasil penelitian Fuquet et al. (2004), Valendra et al. (2011) dan Xu et al. (2008), toksisitas cendawan B. bassiana ditentukan oleh kemampuan toksin yang terkan-dung dalam membunuh serangga. Besarnya mortalitas serangga uji memang menjadi tolok ukur dan isolat cendawan yang mampu menyebabkan kematian dengan mortalitas mencapai 80% tergolong sangat toksik. Sementara itu, Chong-Rodriguez et al. (2011) menekankan bahwa toksisitas cendawan B. bassiana berkaitan erat dengan jenis media tumbuh yang digunakan, media tumbuh yang mengandung protein dan karbohidrat yang tinggi yang terdapat pada jagung manis menghasilkan blastopora yang lebih toksik diban-dingkan dengan cendawan B. bassiana yang ditumbuhkan pada media yang hanya mengandung protein (pepton).
Mortalitas larva instar IV dan V berkisar 63-70% dan berbeda nyata lebih kecil diban-dingkan mortalitas yang terjadi pada perlakuan larva instar I, II maupun instar III (Tabel 2). Meskipun mortalitas yang diakibatkan oleh cendawan B. bassiana lebih rendah, namun dianggap toksik terhadap larva instar IV maupun V karena mortalitas masih diatas 50%. Perlu diketahui bahwa dalam pengelolaan hama terpadu (PHT), cendawan entomo-patogen merupakan salah satu agens hayati yang dapat dikombinasikan dengan agens pengendalian lainnya untuk menekan populasi hama hingga di bawah ambang ekonomi (Rogerio et al. 2005; Islam et al. 2009). Oleh karena itu, peran agens pengendalian lain

Prayogo: Toksisitas Beauveria bassiana terhadap telur Cylas formicarius 678
untuk dikombinasikan dengan agens ini masih sangat diperlukan agar mencapai hasil yang optimal. Dengan demikian, mortalitas yang terjadi pada stadia larva instar IV dan V masih memberi kontribusi yang cukup besar jika dikombinasikan dengan agens pengendalian lain atau teknologi pengendalian lainnya.
KESIMPULAN DAN SARAN 1. Cendawan entomopatogen B. bassiana bersifat ovisidal dan larvisidal sehingga toksik
terhadap stadia telur dan larva C. formicarius. 2. Cendawan B. bassiana sangat toksik terhadap telur C. formicarius terutama telur yang
baru diletakkan imago sampai dengan umur empat hari. Cendawan B. bassiana juga toksik terhadap seluruh stadia larva C. formicarius.
3. Penelitian metode dan waktu aplikasi cendawan B. bassiana untuk mengendalikan C. formicarius di lapangan perlu dilakukan.
DAFTAR PUSTAKA Ahdiaty I. 2013. Pengaruh umur cendawan Beauveria bassiana (Balsamo) Vuillemen terhadap
infektivitasnya pada Cylas formicarius (Fabricius) (Coleoptera: Brentidae). Institut Pertanian Bogor, Fakultas Pertanian, Departemen Proteksi. [skripsi]. 96 hlm.
Batta YA. 2007. Biocontrol of almond bark beetle (Scolytus amygdale Geurin-Meneville) (Coleoptera: Scolytidae) using Beauveria bassiana (Bals.) Vuill (Deuteromycotidae: Hyphomycetes). J Appl Microbiol 103(5):1406-1414.
Bidochka MJ, Menzies FV, Kamp AM. 2002. Genetic groups of the insect pathogenic fungus Beauveria bassiana are associated with habitat and thermal growth preferences. Arch Microbiol 178:531-537.
Broza M, Pereira PM, Stimac JL. 2001. The non susceptibility of soil Collembola to insect pathogens and their potential as Scavengers of microbial pesticides. Pedobiol 45:523-534.
Carneiro ME, Monteiro SGM, Daemon E, Bettencourt VREP. 1999. Effect of isolate 986 of Beauveria bassiana (Bals) Vuill on eggs of the Ttick Anocentor nitens (Acari: Ixodidae). Rev Bras Parasitol Vet 8(1):59-62.
Chiranjeevi Ch, Reddy BDR, Gour TB, Reddy YN, Abida S. 2002. Chemical control sweet potato weevil Cylas formicarius (Fabricius) in sweet potato. Indian J of Plant Protect 30(1):59-62.
Chong-Rodriguez MJ, Maldonado-Blanco MG, Hernandez-Escareno JJ, Galan-Wong LJ, Sandoval-Coronado CF. 2011. Study of Beauveria bassiana growth, blastospore yield, desiccation-tolerance, viability and toxic activity using different liquid media. African J of Biotechnol 10(30):5736-5742.
Costa J, Alves LFA, Daros AA. 2010. Safety of Beauveria bassiana (Bals.) Vuillemen to Galus domesticus L. Braz Arch of Biol and Technol 53(2):465-471.
Cruz LP, Gaitan AL, Gongora CE. 2006. Exploiting the genetic diversity of Beauveria bassiana for improving the biological control of the coffee berry borer through the use of strain mixtures. Appl Microbiol Biotechnol 71:918-926.
El-Sinary GH. 2002. Influence of the entomopathogenic fungus Beauveria bassiana (Balsamo) on the mature larvae of the potato tuber moth Phthorimaea operculella (Zeller) under different degrees of temperature and relative humidity. J Agric Sci 27:4150-4161.
Fuquet R, Therraud M, Vey A. 2004. Production in vitro of toxic macromolecules by strains of Beauveria bassiana and purification of a chitosanase like protein secreted by a melanizing

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2013 679
isolate. Comp Biochem Physiol Toxicol Pharmacol 138(2):149-161. Garcia-Munguia AM, Garza-Hernandez JA, Rebollar-Tellez MA, Rodriguez-Perez MA, Reyes-
Villanueva F. 2011. Transmission of Beauveria bassiana from male to female Aedes aegypti mosquitoes. Parasite & Vectors 4:24-29.
Gayathri G, Balasubramanian C, Moorthi PV, Kubendran T. 2010. Larvicidal potential of Beauveria bassiana (Balsamo) Vuillemin and Paecilomyces fumosoroseus (Wize.) Brown and Smith on Culex quinquefasciatus (Say.). J of Biopes 3(1):147-151.
Gindin G, Levski S, Glazen I, Soroker V. 2006. Evaluation of the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana against the Red Palm weevil Rhynchophorus ferrugineus. Phytopar 34(4):370-379.
Huang Z, Ali S, Ren S, Wu J, Zhang Y. 2012. Influence of the entomopathogenic fungus Beauveria bassiana on Prynocaria congener (Billberg) (Coleoptera: Coccinellidae) under laboratory conditions. Pakistan J Zool 4(1):209-2016.
Islam MT, Castle SJ, Ren SX. 2009. Compatibility of the insect pathogenic fungus Beauveria bassiana with neem against sweet potato whitefly Bemisia tabaci on eggplant. Entomol Experiment et Appl 134:28-34.
Kafie L, Wu WJ, Kao SS, Shin CJ. 2011. Efficacy of Beauveria bassiana against the red imported fire ant Solenopsis invicta (Hymenoptera: Formicidae) in Taiwan. Pest Manag Sci 67(11):14321-1438.
Kandori I, Kimura T, Tsumuki H, Sugimoto T. 2006. Cold tolerance of the sweet potato weevil Cylas formicarius (F.) (Coleoptera: Curculionidae) from the South Western Islands of Japan. Appl Entomol & Zool 41:217-222.
Kanstantopoulou MA, Mazomenos BE. 2004. Evaluation of Beauveria bassiana and B. brongniartii strains and four wild-type fungal species against adults a Bactrocera oleae and Ceratitis capitata. Biocont 50:293-305.
Kirkland BH, Westwood GS, Keyhani NO. 2004. Pathogenicity of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae to Ixodidae tick species Dermacentor variabilis, Rhipicephalus sanguineus and Ixodes scapularis. J Med Entomol 41(4):76-711.
Komi K. 2011. Eradication of sweet potato weevil Cylas formicarius Fabricius from Muroto city, Kochi, Japan. http://www.Fftc.imita.org/htmlarea/library/ 201107120759/eb493c.pdf [1 April 2013].
Kumano N, Kohama T, Ohno S. 2007. Effects of irradiation on dispersal ability of male sweetpotato weevils (Coleoptera: Brentidae) in the field. J Econ Entomol 100(3):730-736.
Leng PH, Reddy GVP. 2012. Bioactivity of selected ecofriendly pesticides against Cylas formicarius (Coleoptera: Brentidae). Florida Entomol Soc 95(4):1040-1047.
Lewis LC, Bruck DJ, Gunnarson RD, Bidne KG. 2000. Colonization of entomopathogenic fungus Beauveria bassiana (Deuteromycotina: Hyphomycetes) on Bt transgenic corn (Zea maize Still) and their genetic isolines and assesement of possible plant pathogenicity. Crop Sci 39:191-200.
Li MV, Lin HF, Li SG, Xu AM, Feng MF. 2012. Efficiency of entomopathogenic fungi in the control of eggs of the brown planthopper Nilaparvata lugens Stal. (Homoptera: Delphacidae)/ African J of Microbiol Res 6(44):7162-7167.
Luz C, Tai MHH Tai, Santos AH, Rocha LFN, Albemax CDAS, Silva HHG. 2007. Ovicidal activity of entomopathogenic hyphomycetes on Aedes aegypti (Diptera: Culicidae) under laboratory conditions. J of Medical Entomol 144(5)799-804.
Meikle NG, Mercadier G, Holst N, Girod V. 2008. Impact of two treatments of a formulation of Beauveria bassiana (Deuteromycotina: Hyphomycetes) conidia on Varroa mites (Acari: Verroidae) and on honeybee (Hynemoptera: Aphididae) colony health. Exp Appl Acarol 46:105-117.
Meyling NV & Eilenberg J. 2007. Ecology of the entomopathogenic fungi Beauveria bassiana

Prayogo: Toksisitas Beauveria bassiana terhadap telur Cylas formicarius 680
and Metarhizium anisopliae in temperate agroecosystems: Potential for conservation biological control. J Microbiol 104(2):1401-1411.
Mulock BS, Chandler LD. 2001. Effect of Beauveria bassiana on the fecundity of western corn rootworm (Diabrotica virgifera virgifera) (Coleoptera: Chrysomelidae). Biol Contr 22:16-21.
Mukhtar T, Pervaz I. 2003. In vitro evaluation of ovicidal and larvicidal effects of culture filtrate of Verticillium chlamydosporium against Meloidogyne javanica. Int J Agric Biol 4:576-579.
Nielsen C, Skovgard H, Steenberg T. 2005. Effects of Metarhizium anisopliae (Deuteromycotina: Hyphomycetes) on survival and reproduction of the filth fly parasitoid Spalangia cameroni (Hymenoptera: Pteromalidae). Environ Entomol 34:133-139.
Ondiaka S, Maniania NK, Nyamasyo GHN, Nderitu JH. 2008. Virulence of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae to sweetpotato weevil Cylas puncticollis and effects on fecundity and egg viability. Ann of Appl Biol 153(1):41-48.
Prassad A, Verwal B. 2010. Biotoxicity of entomopathogenic fungus Beauveria bassiana (Balsamo) Vuillemen against early larval instars of Anopheline mosquitoes. J of Herbal Medicine and Toxicol 4(2):181-188.
Prayogo Y. 2009. Kajian cendawan entomopatogen Lecanicillium lecanii (Zimm.) Zare & Gams untuk Menekan perkembangan telur hama pengisap polong kedelai Riptortus linearis (F.) (Hemiptera: Alydidae). Institut Pertanian Bogor, Fakultas Pertanian, Departemen Proteksi. [disertasi]. 129 hlm.
. 2010. Virulensi beberapa isolate cendawan entomopatogen Beauveria bassiana (Balsamo) untuk mengendalikan penggerek ubijalar Cylas formicarius. Makalah disampaikan pada Seminar Nasional Hasil Penelitian BALITKABI Tahun 2012. [belum terbit].
Ratissa DA. 2011. Keefektifan cendawan entomopatogen Beauveria bassiana (Balsamo) Vuillemen terhadap Cylas formicarius (Fabricius) (Coleoptera: Brentidae) dan pengaruhnya pada keperidian. Institut Pertanian Bogor, Fakultas Pertanian, Departemen Proteksi. [skripsi]. 67 hlm.
Regaieg H, Ciancio A, Raovani NH, Grasco G, Rosso L. 2010. Effects of culture filtrates from the nematophagous fungus Verticillium leptobactrium on viability of the root-knot nematode Meloidogyne incognita. World J Microbiol Biotechnol 31(3):1-5.
Rogerio A, Depieri S, Martinez S, Menezes AO. 2005. Compatibility of the fungus Beauveria bassiana (Bals.) Vuill (Deuteromycetes) with extracts of neem seeds and leaves and the emulsible oil. Neotrop Entomol 34(4):601-606.
Rosfiansyah. 2009. Pengaruh aplikasi Beauveria bassiana (Balsamo) Vuillemen dan Heterorhabditis sp. Terhadap serangan hama ubijalar Cylas formicarius (Fabricius) (Coleoptera: Brentidae). Institut Pertanian Bogor, Fakultas Pertanian Departemen Proteksi. [tesis]. 74 hlm.
Santos AH, Tai MHH, Rocha LFN, Silva HHG, Luz C. 2009. Dependence of Metarhizium anisopliae on high humidity for ovicidal activity on Aedes aegypti. Biol Contr 50:37-42.
Sevim A, Hofte M, Demirbag Z. 2012. Genetic variability of Beauveria bassiana and Metarhizium anisopliae var. anisopliae isolates obtained from the Eastern Black Sea Region of Turkey. Turk J Biol 36:255-265.
Sharah HA, Sodangi BI, Abdurrahman M. 2012. The economic implication of Cylas spp. and rot organisms infestation on the matketability of sweet potato (Ipomoea batatas L.) in three markets in Maiduguri Metropolis Barno State Nigeria. International J of Econ Develop Res & Investment 3(3):32-38.
Shefik MAA. 2010. Biological effect of the entomopathogenic fungus Beauveria bassiana (Balsamo) Vuillemen on the potato tuber moth (Phthorimaea operculella Seller). Iraq J of

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2013 681
Sci 51(2):243-248. Shi WB, Feng MG. 2004. Lethal effect of Beauveria bassiana, Metarhizium anisopliae,
Paecilomyces fumosoroseus on the eggs of the Tetranychus cinnabarius (Acari: Tetranycidae) with a description of a mite eggs bioassay system. Biol Contr 30:165-173.
Shi WB, Feng MG, Liu SS. 2008. Sprays of emulsifiable Beauveria bassiana formulation are ovicidal towards Tetranychus urticae (Acari: Tetranychidae) at various regimes of temperature and humidity. J Exp & Appl Acarol 46(1-6):247-257.
Shinya R, Aiuchi D, Kushida A, Tani M, Kuramochi K, Koike M. 2008. Effect of fungal culture filtrate of Verticillium lecanii (Lecanicillium spp.) hybrid strains on Heterodera glycines eggs and juveniles. J Invertebr Pathol 97:291-297.
Smith TP, Hammond AM. 2006. Comparative susceptibility of sweetpotato weevil (Coleoptera: Brentidae) to selected insecticides. J Econ Entomol 99(6):2024-2029.
Takatsuka J. 2007. Characterization of Beauveria bassiana isolates from Japan using inter-simple-sequence-repeat-anchored polymerase chain reaction (ISSR-PCR) amplification. Appl Entomol Zool 42:563-571.
Thakur R, Rajak RC, Sandhu SS. 2005. Biochemical and molecular characteristics of indigenous strains of the entomopathogenic fungus Beauveria bassiana of central India. Biocont Sci Technol 15:733-744.
Thungrabeab M, Tongma S. 2007. Effect of entomopathogenic fungi Beauveria bassiana (Balsamo) and Metarhizium anisoliae (Metsch.) on non target insects. Kmitl Sci Tech J 7:8-12.
Tikhonov VE, Lopez-Llorca LV, Salinas J, Jansson HB. 2002. Purification and characterization of chitinase from the nematophagous fungi Verticillium chlamydosporium and V. suchlasporium. Fungi Gen Biol 35:67-78.
Valendra JW, Gaitan BAL, Jimenez AV, Grossi de-Sa MF. 2011. Cytotoxic activity of fungal metabolites from the pathogenic fungus Beauveria bassiana: an intraspecific evaluation of Beauvericin production. Curr Microbiol 63(3):306-312.
Vega FE, Posada F, Aime MC, Pova-Ripoll M, Infante F, Rehner SA. 2008. Entomopathogenic fungal endophytes. Biol Contr 46:72-82.
Wang L, Huang J, You M, Liu B. 2005. Effects of toxin from two strains of Verticillium lecanii (Fungi: Hyphomycetes) on bioatributes of a predatory ladybeetle Delphastus cata linae (Coleoptera: Coccinellidae). J Appl Entomol 129:32-38.
Xu Y, Orozco R, Wijeratne FM, Gunatilaka AA, Stack SP, Moinar I. 2008. Biosynthesis of the cyclooligomer depsipeptide, beauvericin, a virulence factor of the entomopathogenic fungus Beauveria bassiana. Chem Biol 15(9):898-907.
Yasuda K, Toyosato T, Takaesu K. 2000. Enhanced infectivity of oil formulations of Beauveria bassiana to Cylas formicarius (Fabricius) (Coleoptera: Curculionidae). Japanese J of Appl Entomol and Zool 44(4):241-243.
Zimmermann G. 2007. Review on safety of the entomopathogenic fungi Beauveria bassiana and B. brongniarti. Biocont Sci Technol 17:553-596.
Zhu H, Kim JJ. 2012. Target oriented dissemination of Beauveria bassiana conidia by the predator Harmonia oxyridis (Coleoptera: Coccinellidae) and Chrysoperla carnea (Neuroptera: Chrysopidae) for biocontrol of Myzus persicae. Biocont Sci & Technol 22(4):393-406.
















![Virulence of Beauveria bassiana (Bals.) Vuill. (Deuteromycotina: … · 2020-01-03 · Lepidoptera, such as Castnia licus Drury [9], Ostrinia nu- bilalis Hübner [10], Plutella xylostella](https://img.pdfslide.net/doc/110x75/5eb0d83c425ff45ef61877a5/virulence-of-beauveria-bassiana-bals-vuill-deuteromycotina-2020-01-03-lepidoptera.jpg)












