Embed Size (px)
Citation preview

Indian Journal of Experimental Biology Vol. 41, April 2003, pp. 336-340
Toxicity of PCB 1232 on mitochondria of fish Arius cae latus (Valenciennes)
Dravyam Selvarani * & C Rajamanickam
Department of Biochemistry, School of Biological Sciences, Madurai Kamaraj University, Madurai 625 021, India
Received 19 February 2002, revised 24 January 2003
Mitochondrial proteins and phospholipids were estimated and SOH, Na+ - K+ - ATPase and Mg2+ - ATPase activities were analysed in the gill, liver and heart tissues of PCB 1232 (sublethal doses) treated fish A. cae/alUs. Protein and phospholipids were found to be decreased significantly and SOH, Na+ - K+ - ATPase, Mg2+ - ATPase and other enzyme systems displayed an inverse relationship with PCB dosage. Statistical analysis was carried out to indicate the relationship between sublethal doses of varying concentration and the activities of the enzyme systems involved in energy metabolism. The studies indicated impairment in mitochondrial functions .
Organochlorines have been found to uncouple mitochondrial oxidative phosphorylation and the respiratory chain. Ela et al. I presumed organochlorine like DDT to be an inhibitor of oxidative phosphorylation rather than an uncoupler. Uncouplers enhance ATPase acti vity by causing the break down of A TP2.
Na+-K+-ATPase and Mg2+-ATPase are both membrane bound3 and maintain ionic balance by active transport. They have been found to be affected by organochlorines4
,5 . DDT has also been reported to inhibit SOH activitl, which is an indication of impaired oxidative metabolic cycle. Cytochrome c oxidase, being the terminal enzyme of the electron transport system was found to be the site of organochlorine action7
.
Polychlorinated biphenyl (PCB)-an organochlorine is an universal priority toxic pollutantS, which is transported through the food chain and biomagnified at higher tropic levels9
• Hence in the present study the effects of PCB 1232 on the key enzymes of mitochondria of the gills, liver and heart of an estuarine fish Arius caelatus have been analysed.
Materials and Methods Fish Arius caelatus weighing 95-100 g were col
lected along the coast of Pazhayakayal near Tuticorin (Longitude East 79° of Greenwich and Latitude 8° North) and maintained in three separate netted enclosures in the estuary itself. The fishes were fed with beef. Five sets of experiments were performed. Each
* Address for correspondence: Department of Biology, Pope's College, Sawyerpuram 628 251, India. Phone: 0461 - 2327759
set consisted of 3 fishes . The first set which served as the control received ip injection of I ml corn oil. The fish in the second, third, fourth and fifth sets were given, 0 .6, 2, 4 and 6 mg PCB/kg respectively in I ml corn oil. PCB 1232 was purchased from NSI Environmental Solutions, North Carolina, USA. After 5 days of injection, the gills, liver and heart were removed and processed for isolation of mitochondria. All proceedings were carried out at 0°_5°C.
Isolation of mitochondria from gills-Mitochondria from the gills were separated 10. The gill bars were scraped off without any cartilage and after washing with 0.15 M KCl solution, were homogenised in 0.25 M sucrose containing 0.002 M Tris-HCI, pH 7.6 and 0.002 Methylene diamine tetra acetic acid (EDTA). The homogenate was centrifuged at 600 g for 10 min to remove nuclei and cell debris. The supernatant was again centrifuged at 10,000 g for 10 min to obtain mitochondrial pellet. The pellet was washed thrice and finally suspended in the same medium.
Isolation of mitochondria from heart and liverIsolation of mitochondria from heart and liver was carried out ll . The organs were blotted free of blood, weighed and washed several times in ice-cold 0.25 M sucrose solution containing ImM EDTA, 20mM trisHCI (PH 7.2) and 0.2% bovine serum albumen (BSA, pH 7.2-SET buffer). The tissues were sliced into small pieces. The whole liver was homogenised, while only the most metabolically active chambers of the heart, such as, the atrium and ventricle were used. The homogenisation was performed in 10 volume (w/v) ice-cold SET buffer. Nuclei and cell debris were removed and mitochondria were isolated from the homogenate by differential centrifugation.

SELVARANI & RAJAMANICKAM : TOXICITY OF PCB 1232 ON MITOCHONDRIA OF FISH ARIUS CAELATUS 337
Protein was estimated by the method of Lowry et al l2
. The phospholipids were extracted by. the method of Bligh and Dyerl3 and estimated by the method of Fiske and SUbbarow l4.
ATPase was assayed following the procedure of Tzagoloffl5 with minor modifications. The reaction mixture contained a total volume of 1ml, 50mM Tris HCI (PH 8.5), 4 mM ATP, 5 mM MgS04 and the mitochondrial protein. Ouabain (2 mM was added when estimating Mg2+-ATPase activity. The reaction was initiated by adding ATP and stopped by the addition of 1ml of 10% TCA and kept at 10°C for 15 min. The supernatant was centrifuged at 10000 rpm for 10 min and assayed for inorganic phosphate by the method of Fiske and SUbbarow '4 . Na+-K+-ATPase activity was calculated as the difference between inorganic phosphate liberated in presence or absence of 2 mM ouabain. Quabain insensitive ATPase was considered Mg2+-ATPase. SDH I6 and cytochrome c oxidase '7
were assayed. The statistical analyses followed in the present
study are (l) standard deviation, (2) students t test, (3) simple linear regression and correlation, (4) one-way analysis of variation, (5) two-way analysis of variation, and (6) Student Newman-Keul's test.
Results and Discussion Irrespective of the tissues, injection of the lowest
PCB dose stimulated protein synthesis (Table 1). This may be an adjustment to detoxify the toxicant. Further increase in PCB dose gradually suppressed protein synthesis especially in gill and liver mitochondria. As a result the protein content of the fish that received the highest PCB dose was less than that of the control. According to Syverson lS the protein synthesis is inhibited by toxicants. The present study reveals that a part of the protein content may be utilized to counteract the pesticide stress as evidenced by the decline of
protein content and another part may be transformed into nitrogenous wastes like ammonia as evidenced by the elevation of ammonia excretion l9. This may also indicate rapid utilization of protein or that dietary protein consumed by the fish is not stored. The decline of protein is suggestive of intensive proteolysis supporting the hypothesis of Kabeer20. Moreover PCB treated fish would have obtained the extra energy from body tissue protein.
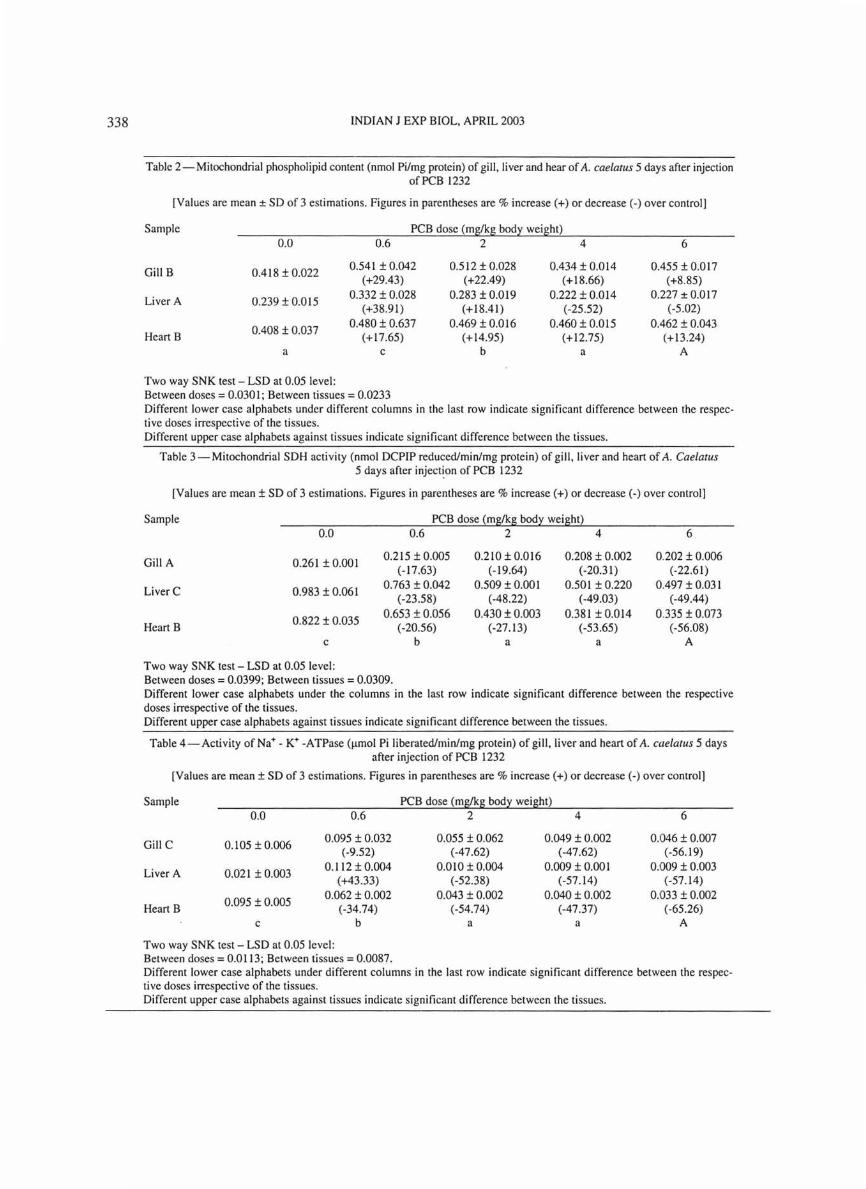
Injection of 0.6 mg PCB/kg body weight dose increased the phospholipid content which decreased significantly with further increase to 2, 4 and 6 mg/kg body weight (Table 2). At the highest dose the mitochondrial phospholipid content decreased from the level of sham treated fish .
Data on SDH activity (Table 3) point out that the lower two PCB doses significantly decreased the activity and with the higher doses there was no significant decline of the enzyme activity. Similar results were obtained when Channa striatus was subjected to DDT6 . Sakthivel et al. 2I observed decreased SOH activity in Cyprinus carpio in response to textile dye stuff effluents and stated that it was a indication of depression of cellular metabolism and that the metabolic pathway may have turned anaerobic to meet the increased energy demands during stress caused by the pollutant. Joyce et al. 22 in their study using malathion on Clarias batrachus remarked that decreased SOH activity indicated decreased funnelling of intermediates into the citric acid cycle.
Like SDH, Na+-K+-ATPase (Table 4) and Mg2+_ ATPase (Table 5) activities decreased with increasing PCB dose. Influence of ATPase in fish by PCB is varied. Koch et.al. 5 have reported Na+-K+ -ATPase inhibition in kidney, liver and brain of Pimephales promelas while Folmar et.al. 23 did not observe any change in Oncorhynchus kistuch. Similarly organochlorine DDT decreased Na+-K+-ATPase activity of intestinal
Table 1 - Mitochondrial protein content (mg/g tissue) of gill, liver and heart of A. caelatus 5 days after injection of PCB 1232
[Values are mean ± SD of 3 estimations. Figures in parentheses are % increase (+) or decrease (-) over controll
Sample PCB dose (mg/kg bod~ weight) LSD value 0.0 0.6 2 4 6
Gill 4.633 ± O.073c 5.247 ± 0.085d 4 .938 ± O. I03cd 3.870 ± 0.773b 3.220 ± 0.257" 0.3328
(+13 .26) (+6.58) ( -16.47) ( -30.5)
Liver 11.286 ± 0.178b 12.925 ± 0.192c 11.335 ± 0.173b 9.318 ± 0.400' 9.400 ± 0.176' 0 .5352
(+ 14.52) (+0.43) (-17 .44) (-16.71)
Heart 4 .870 ± 0.271" 6.695 ± O.346b 6.273 ± 0. 149b 6.160 ± 0.244b 5.267 ± 0.261"
0.5661 (-37.47) (+28.81) (+ 26.49) (+8.15)
Note: Values in a row superscribed by different alphabets are significantly different at P < 0.05 (SNK Test)
338 INDIAN J EXP BIOL, APRIL 2003
Table 2 - Mitochondrial phospholipid content (nmol Pi/mg protein) of gill, liver and hear of A. cae/atus 5 days after injection of PCB 1232
[Values are mean ± SD of 3 estimations. Figures in parentheses are % increase (+) or decrease (-) over control]
Sample PCB dose (mg/kg body weight) 0.0 0.6 2 4 6
Gill B 0.418 ± 0.022 0.541 ± 0.042 0.512 ± 0.028 0.434 ± 0.014 0.455 ± 0.017
(+29.43) (+22.49) (+18.66) (+8 .85)
Liver A 0.239 ± 0.015 0.332 ± 0.Q28 0.283 ± 0.019 0.222 ± 0.014 0.227 ± 0.017
(+38.91) (+18.41) (-25 .52) (-5 .02)
0.408 ± 0.037 0.480 ± 0.637 0.469 ± 0.016 0.460 ± 0.Dl5 0.462 ± 0.043
Heart B (+17.65) (+14.95) (+12.75) (+13.24) a c b a A
Two way SNK test - LSD at 0.05 level: Between doses = 0.0301 ; Between tissues = 0.0233 Different lower case alphabets under different columns in the last row indicate significant difference between the respective doses irrespective of the tissues. Different upper case alphabets against tissues indicate significant difference between the tissues.
Table 3 - Mitochondrial SDH activity (nmol DCPIP reduced/min/mg protein) of gill. liver and heart of A. Cae/atus 5 days after inject~on of PCB 1232
[Values are mean ± SD of 3 estimations. Figures in parentheses are % increase (+) or decrease (-) over control]
Sample PCB dose (m~kg bod:t weight) 0.0 0.6 2 4 6
Gill A 0.261 ± 0.001 0.215 ± 0.005 0.210 ± 0.016 0.208 ± 0.002 0.202 ± 0.006
(-17.63) (-19.64) (-20.31) (-22.61)
Liver C 0.983 ± 0.061 0.763 ± 0.042 0.509 ± 0.001 0.501 ± 0.220 0.497 ± 0.031
( -23.58) (-48.22) (-49.03) (-49.44)
0.822 ± 0.035 0.653 ± 0.056 0.430 ± 0.003 0.381 ± 0.014 0.335 ± 0.073
Heart B (-20.56) (-27.13) (-53.65) (-56.08) c b a a A
Two way SNK test - LSD at 0.05 level: Between doses = 0.0399; Between tissues = 0.0309. Different lower case alphabets under the columns in the last row indicate significant difference between the respective doses irrespective of the tissues. Different upper case alphabets against tissues indicate significant difference between the tissues.
Table 4-Activity of Na+ - K+ -ATPase (Ilmol Pi liberated/min/mg protein) of gill, liver and heart of A. cae/alus 5 days after injection of PCB 1232
[Values are mean ± SD of 3 estimations. Figures in parentheses are % increase (+) or decrease (-) over control]
Sample PCB dose (mg/kg bod:t weight) 0.0 0.6 2 4 6
Gill C 0.105 ± 0.006 0.095 ± 0.032 0.055 ± 0.062 0.049 ± 0.002 0.046 ± 0.007
( -9.52) (-47 .62) (-47.62) (-56.19)
Liver A 0.021 ± 0.003 0.112 ± 0.004 0.010 ± 0.004 0.009 ± 0.00 1 0.009 ± 0.003
(+43.33) (-52.38) (-57.14) (-57 .14)
0.095 ± 0.005 0.062 ± 0.002 0.043 ± 0.002 0.040 ± 0.002 0.033 ± 0.002
Heart B (-34.74) (-54.74) (-47 .37) (-65.26) c b a a A
Two way SNK test - LSD at 0.05 level: Between doses = 0.0113; Between tissues = 0.0087. Different lower case alphabets under different columns in the last row indicate significant difference between the respecti ve doses irrespecti ve of the tissues. Different upper case alphabets against tissues indicate significant difference between the tissues .

SELVARANI & RAJAMANICKAM: TOXICITY OF PCB 1232 ON MITOCHONDRIA OF FISH ARlUS CAELATUS 339
Table 5 - Mitochondrial Mg2+-ATPase (flmol Pi liberated / min / mg protein) of gill. liver and heart of A. cae/alliS 5 days after injection of PCB 1232.
[Values are mean ± SD of 3 estimations. Figures in parentheses are % increase (+) or decrease (-) over control]
Sample PCB dose (mg/kg body weight) 0.0 0.6 2 4 6
Gill B 0.161 ±0.010 0.079 ± 0.001 0.077 ± 0.001 0.176 ± 0.003 0.076 ± 0.005 (-50.93) (-52.17) ( +9.32) ( -52.8)
Liver A 0.043 ± 0.002 0.029 ± 0.003 0.021 ± 0.002 0.020 ± 0.002 0.018 ± 0.002 (-32.56) (-51.16) ( -53.49) (-58.14)
0. 128 ± 0.007 0.109 ± 0.007 0.094 ± 0.001 0.088 ± 0.003 0.085 ± 0.003
Heart C (-14.84) ( -26.56) (-31.25) ( -33.59) c b a a A
Two way SNK test - LSD at 0.05 level: Between doses = 0.0053; Between ti ssues = 0.0041 . Different lower case alphabets under different columns in the last row indicate significant difference between the respective doses irrespective of the tissues. Different upper case alphabets against tissues indicate significant difference between the tissues.
Table 6- Mitochondrial cytochrome c oxidase activity (flmol cytochrome c reduced/min protein) of gill. liver and heart of A. cae/alliS 5 days after injection of PCB 1232.
[Values are mean ± SD of 3 estimations. Figures in parentheses are % increase (+) or decrease (-) over control]
Sample PCB dose (mg/kg body weight) 0.0 0.6 2 4 6
Gill A 0.159 ± 0.060 0. 139 ± 0.027 0.102 ± 0.002 0.099 ± 0.005 0.083 ± 0.002
(-12.58) (-35 .9) (-37.74) (-47.8)
Liver B 0.313 ± 0.027 0.26 1 ± 0.027 0.226 ± 0.02 0.217 ± 0.014 0.213 ± 0.020
(-16.61) (-27.8) (-30.67) (-31.95)
0.666 ± 0.020 0.617 ± 0.040 0.435 ± 0.004 0.404 ± 0.013 0.363 ± 0.021
Heart C (-7.36) (-34.69) (-39.34) ( -45.5) d c b ab A
Two way SNK test - LSD at 0.05 level: Between doses = 0.0211; Between ti ssues = 0.0164. Different lower case alphabets under different columns in the last row indicate significant difference between the respective doses irrespective of the tissues. Different upper case alphabets against tissues indicate significant difference between the tissues.
epithelium of Anguilla rostrata24 and brain Mg2+_ ATPase activity of Channa striatui5
. In Channa gachua 0.0022 mg/l endosulfan enhanced Na+-K+ATPase activity in the liver while 0.0056 mg/l enhanced Mg2+-ATPase activit/6
. Therefore it can be concluded that organochlorines seriously interfere with SOH, Na+-K+-ATPase and Mg2+-ATPase and enzymes of oxidative phosphorylation.
As SOH and A TPases, cytochrome C oxidase of sham treated A. caelatus displayed highest activity (Table 6). Injection of 0.6 and 2.0 mg/kg dose resulted in marked decrease in the enzyme activity. Cytochrome C oxidase like phospholipid is embedded in the inner mitochondrial membrane27. Hence the inhibition by the highest PCB dose can be related to the depletion of phospholipid. Surprisingly the phospholipid content of A. caelatus subjected to 0.6 mg PCB/kg increased while the cytochrome C oxidase activity reduced. Exposure of channel catfish28 to
stressor like hydrogen sulphide has been reported to cause similar reduction in the activity of cytochrome C oxidase. Hence cytochrome C oxidase inhibition is a response to stress.
The present study reveals that eventhough PCB 1232 has only 32% chlorine it is also hazardous as other PCBs with more chlorine content.
Acknowledgement One of the authors (OS) acknowledges the finan
cial assistance from UGC, New Delhi through a Teacher Fellowship.
References lEla R, Chefurka W & Robinson J B, In vivo glucose metabo
lism in the normal and poisoned cockroach Periplaneta americana, J Insect Physio/, 16 (1970) 2137.
2 Fukami 1. Insecticides as inhibitors of respiration in Insecticide biochemistry and physiology, edited by C F Wilkinson (Plenum Press, New York) 1976, 353.

340 INDIAN J EXP BIOL. APRIL 2003
3 Jowett R E. Rhead M M & Mayne B L. In vivo changes in the activity of gill ATPases and haemolymph ions of CarCillllS means exposed to p, p'-DDT and reduced salinities, COlllP Biochelll Physiol, 69C (1981) 399.
4 Leadam T P, Campell R C & Johnson 0 W, Osmoregulatory responses to DDT and varying salinities in Salmo goirdneri -I Gill Na+-K+-ATPase, Comp Biochem Physiol , 49A (1974) 197.
5 Koch R B, Desaiah 0, Yap H H & Cutkomp K L, Polychlorinated piphenyls: Effect of long term exposure on ATPase activity in fish Pimephales promelas. Bull Ellviroll ContOIll Toxicol, 7 (1972) 87.
6 Bhaskaran R, Effect of DDT and methyl parathion on the mitochondrial respiration SOH and ATPase activity of an airbreathing fish Chollllo striatlls, Ellvir & Ecol. 6 (1988) 198.
7 Anderson A 0, March R B & Metcalf R L. Inhibition of succinooxidase system of susceptible and resistant housenies by DDT and related compounds, AIIII Emomol Soc Alii. 47 (1954) 595.
8 Nowak B J, Biological significance of xenobiotic residues, TEN,4(1997) 149.
9 Wit C, Jansson de B, Bergek S, Hjelt M, Rappe C, Olsson M & Andersson 0, Polychlorinated dibenzo-p-dioxin and dibenzofuran levels and patterns in fish and fish eating wildlife in the Baltic sea, Chemosphere, 25 (1992) 185.
10 Meenakshi S, Rajamanickam C & Jeyaraman J, J Adaptation to salinity by fish, Biosci. 1(1979) 427.
II Irving 0 ° & Watson K, Mitochondrial enzymes of tropical fish. COlllP Biochem Physiol. 54B (1976) 81.
12 Lowry ° H. Rosenbrough N J. Farr A L & Randall R J, Protein measurement with the Folin phenol reagent, J Bioi Chelll. 20 (1951) 265.
13 Bligh E G & Dyer W J, A rapid method of total lipid extraction and purification, Call J Biochem Physiol, 37 (1959) 911.
14 Fiske C H & Subbarow Y, The calorimetric detcrmination of phosphatases, J Bioi Chem, 66 (1925) 375.
15 Tzagoloff A, The adenosine triphosphatase complex of mitochondria. In: The enzymes of biological membranes, editcd by Martinosi, A S, Vol4 (Plenum Press, New York) 1976, 103.
16 Green 0 E, Mil S & Kohout P M, SlIIdies on the termillal electroll trallsport systelll 1 in Succillic dehydrogellase, 217 (1955) 551.
17 Wharton 0 C & Tzagoloff A, in Methods ill enzymology, edited by R W Estabrook & M E Pullman. Vol X. (Aademic Press, New York) 1967, 245.
18 Syversen T L M, Effects of repeated dosing of methylmercury of in vivo protein synthesis of isolated neurons, Acta Pharmocol Toxicol. 50 (1982) 391.
19 Selvarani 0, SOllie aspeCTS of aqualic ecotoxic%gy - ToxiciTy of PCB 1232 all Arius caelallls, Ph 0 Thesis, Madurai Kamaraj University, Madurai, 1999.
20 Kabeer A S J, SlIIdies on sOllie aspects ofproteil1metabolislII alld associaled ellZYllle sysTellls ill the freshwaTer Te/eosT Tilapia /llossambicus to lIIalathion exposure, Ph 0 Thesis, Sri Venkateswara University, Tirupati, 1979.
21 Sakthivel M, Sampath K & Pandian T J, Sublethal effects of dye stuff on enzymes and tissue respiration of Cyprillllls carpio (Linn), Illdiall J Exp Bioi, 29 (1991) 979.
22 Joyce V, Venkateshwarla P & Janaiah C, Impact of sublethal concentrations of malathion on certain aspects of metabolism in freshwater fish, Clarias baTractus (Linn). COIllP Physiol Ecol, 615 (1990) 13.
23 Folmar L 0, Dickhoff W W, Zaugg W S & Hodgkins H 0, The effects of Aroclor 1254 and No 2 fuel oil on smoltilication and sca water adaptation of Coho salmon (OncorhYIIchlls kistIlCh) , Aquat Toxicol, 2 (1982) 291 .
24 Desaiah 0, Cutkomp L K, Koch R B & Jarvinen A, DDT: Effect of continuous exposure on ATPase activity in fish, Pilllephaies prolllelos. Arch Ellviron COI1TOIII Toxicol. 3 (1975) 132.
25 Bhaskaran R, Biological stlldies 011 chosell thermo cOllforlller (Chollllo striallls), Ph 0 Thesis submitted to Madurai Kamaraj University, Madurai 1980.
26 Sharma R M, Effect of endosulfan on adenosine triphosphatase (ATPase) activity in liver, kidney and muscles of Chollllo goclllla, BI/II Ell viroll COlltalll Toxicol, 41 (1988) 317.
27 AI-Tai W F, Jones M G & Wilson T, Adaptive changes in respiratory movements of an air-breathing fish , Challllo sTriallls (Blecker) exposed to carbamate pesticide, COIllP Biochelll Physiol, 77B (1984) 609.
28 Torrans E L & Clemens H P, Physiological and biochemical effects of acute exposure of fish to hydrogen sulphide, Comp Biochem Physiol, 171C (1982) 183.