Embed Size (px)
Citation preview

Development KM, 155-164(1988)Primed in Greal Britain © The Company of Biologists Limited 1988
155
Transcripts of a-cardiac and a-skeletal actins are early markers for
myogenesis in the mouse embryo
DAVID A. SASSOON, IAN GARNER and MARGARET BUCKINGHAM
Department of Molecular Biology, Pasteur Institute, 25 Rue de Dr Roux, Paris 75015, France
Summary
Among the first tissues to differentiate in the mam-malian embryo are cardiac and subsequently skeletalstriated muscle. We have developed specific cRNAprobes corresponding to the 5' noncoding regions of a-cardiac and a-skeletal actin mRNAs in order toinvestigate myogenesis in the mouse embryo. Tran-scripts coding for cardiac actin which is the majorisoform of the adult heart can first be detected between7-5 and 7-8 days p.c. in the developing heart and areobserved in all somites as they are formed. In ad-dition, a-skeletal actin transcripts are accumulated atmuch lower levels in cardiac tissue and newly formed
somites; both heart and skeletal muscle show co-expression of this actin gene pair at all stages ofdevelopment examined. The fact that cardiac actintranscripts can be observed in the myotomal portion ofthe somite prior to muscle fibre differentiation indi-cates that cardiac actin transcripts (and to a lesserextent skeletal actin transcripts) are markers not onlyof striated muscle tissue, but also of earlier stages ofthe myogenic programme in vivo.
Key words: sarcomeric actins, myotome, somites,cardiogenesis, mouse embryo.
Introduction
During vertebrate embryogenesis, striated muscle isone of the first tissues characteristic of the adult to beestablished (Rugh, 1968). In Xenopus, nuclease pro-tection assays and protein analyses have revealed thatmuscle type a--actin gene transcripts and their pro-teins are accumulated by the mid-neurula stage in themyotome region of the somites (Mohun et al. 1984;Cascio & Gurdon, 1987). These transcripts arealready detectable as early as the mid/late-gastrulastage, and thus a-actin gene activation is one of theearliest identifiable events leading to tissue specializ-ation. In higher vertebrates, actin gene expression atcomparably early stages has not been investigated. Incontrast to the frog, the establishment of the heartpreceeds somitogenesis and subsequent formation ofskeletal muscle takes place relatively more slowlythan in amphibians (Nieuwkoop & Faber, 1956;Rugh, 1968). In this context, it is therefore of interestto establish the pattern of expression of the a'-actingenes in a higher vertebrate.
In mice, as in most vertebrates, striated muscletissue contains almost exclusively o'-skeletal and a-cardiac actins. Skeletal actin is the major (>95 %)
isoform in adult skeletal muscle (Vandekerckhove etal. 1986). However, during fetal and neonatal devel-opment much higher levels of cardiac actin areobserved in this tissue. Indeed, it has been suggestedthat cardiac actin is the major actin isoform duringthe early stages of skeletal muscle formation. In latefetal (18-day) limb muscle of the mouse, cardiac actin(protein and mRNA) represents about 40% ofstriated muscle actin, and declines to 20% immedi-ately after birth (Minty et al. 1982; Vandekerckhoveet al. 1986). Earlier stages of mammalian myogenesishave not been investigated in vivo. For the chicken,the data are somewhat confusing, although in gen-eral, a'-cardiac actin and a-skeletal actin transcriptshave been detected as early as 5-5 days (stage 26/27)in the limb bud (Ordahl, 1986). During chick devel-opment in ovo, the relative levels of cr-cardiac and ct-skeletal actin mRNAs seem to differ depending onthe particular muscle examined (see Paterson &Eldridge, 1984; Hayward & Schwartz, 1986): post-hatching, as skeletal muscles mature, a-cardiac actinceases to be a major isoform.
In primary cultures derived from late fetal/new-born chick (Hayward & Schwartz, 1986) and human(Gunning et al. 1987) skeletal muscle, cr-cardiac

156 D. A. Sassoon, I. Garner and M. Buckingham
actin transcripts are already detectable prior to theformation of muscle fibres and subsequently cr-car-diac actin becomes the major actin gene to beexpressed as muscle fibres form; o--skeletal actintranscripts accumulate more slowly following myo-tube formation. Similar observations have been madewith mouse muscle cell lines (Minty et al. 1982, 1986;Baines et al. 1984). However, cardiac actin expressionis clearly not obligatory for muscle cell differentiationsince in the rat muscle cell line L6, which was derivedfrom neonatal skeletal muscle, skeletal but not car-diac actin mRNA accumulates as muscle fibres form(Buckingham et al. 1984).
In the case of cardiac muscle, coexpression ofskeletal and cardiac actin genes is also observed. Inlate fetal and new-born rat hearts, skeletal actin(mRNA and protein) accumulates although at thisstage cardiac actin is the predominant isoform (Mayeret al. 1984; Schwartz et al. 1986). In the chick heart,Ordahl (1986) has reported similar levels of a--cardiacand a--skeletal actin gene transcripts as early as2-5days in ovo. Again, it remains unclear when eachmember of this gene pair is activated during cardiacdevelopment.
Here we report an investigation of actin geneexpression during embryogenesis in the mouse from6-5 days post coitum to 13-5 days. This is the periodencompassing late gastrulation (late primitivestreak), the appearance and morphological compart-mentalization of the heart into atria and ventricle aswell as somitogenesis and subsequent establishmentof primary skeletal muscle (e.g. Rugh, 1968). In orderto look at early stages, we have used in situ hybridiz-ation with specific RNA probes for the skeletal andcardiac actin mRNA sequences. By this approach wecan establish precisely the regions of the embryowhere these genes are first expressed. We demon-strate that cardiac actin is the major sarcomeric actintranscript in the mouse during early muscle forma-tion. It accumulates in the first myotome cells of theearly somites and is clearly detectable in the cardiactube by 7-8 days. Skeletal actin is always coexpressedwith cardiac actin, although at lower levels. Thus, thesarcomeric actin transcripts which are characteristicof terminally differentiated adult striated muscle, alsoserve as the early markers of these tissues and theiranlagen.
Materials and methods
Mouse embryosInbred lines of C3H or BALB/c mice were mated and themorning of vaginal plug was counted as post coital (p.c.)day 0-5. Pregnant females were anaesthetized by CO2 andsacrificed by cervical dislocation. Embryos or decidua wereremoved from surrounding tissue in cold phosphate-
buffered saline (PBS: 013M-NaCl, 7mM-Na2PO4, 3mM-NaH2PO4- 2H2O, pH7-4), somites were counted (in em-bryos from 8 days p.c.) under a dissection microscope andembryos carefully transferred to a solution of freshlyprepared 4% paraformaldehyde in PBS. After an over-night fixation, embryos were rinsed in the following sol-utions at the temperature and times indicated: PBS, 4°C(30min (30')), 0-85% saline solution 4°C (30'), 50%ETOH in 0-425 % saline (room temperature here and forthe following), (2x 15'), 70% ETOH (30'), 85% ETOH(30'), 95% ETOH (30'), 100% ETOH (2x 30'), xylene(2x 30'), and a 1:1 solution of xylene:melted paraplast60°C (45'). Tissue was transferred through 3x 20' paraplastat 60°C, then oriented in plastic moulds and cooled. Tissueblocks were stored at 4°C until cut.
In situ hybridizationEmbryos were cut at 5-7 ^m on a standard paraffinmicrotome and individual sections were placed on a drop ofwater to relax compression, on a 'subbed' slide (Gall &Pardue, 1971) at 50°C. Slides were blotted and drainedovernight at room temperature and stored at 4°C until use.The procedures used for section treatment, hybridizationand washings were based upon those used by Wilkinson etal. (1987) with modifications described here. Tissue sectionswere deparaffinized in xylene (2x10'), rehydrated througha graded series of ethanols (100 %-> 30 %), rinsed in 0-85 %saline solution (5'), 1 x PBS (5'), and postfixed in a freshlyprepared and filtered solution of 4 % paraformaldehyde inlx PBS buffer (30'). Slides carrying sections were thenrinsed in PBS (2x10') and treated with a fresh solution ofproteinase K (Merck, 20^gmr' , 8') in Tris-HCl, EDTA(50mM,5mM, pH7-2). Slides were then rinsed in PBS (5'),refixed in 4 % paraformaldehyde solution in PBS, dipped indistilled water and acetylated in a 0-09 M-solution of trietha-nolamine with acetic anhydride (l:400v/v) for lOmin.Slides were subsequently rinsed in lx PBS and 0-85%saline (5' each), and quickly dehydrated through a series ofethanols (30%-» 100%). Slides were allowed to dry atleast 2h prior to hybridization. Probe was applied directlyto tissue sections (10—13 ̂ zl; see 'Preparation of probe') at afinal adjusted concentration of 750O0disintsmin~ V ' inhybridization buffer (50% deionized formamide, 0-3 M-NaCl, 20mM-Tris-HCl (pH7-4), 5mM-EDTA, 10 raM-NaPO4 (pH8), 10% dextran sulphate, lx Denhart's,50 jig ml"1 total yeast RNA) and tissue and probe werecovered with a siliconized coverslip (22x22mm). Hybridiz-ation was carried out at 50°C for approximately 16 h in ahumid chamber. Coverslips were gently floated off in 5xSSC(lx SSC,045M-NaCl,0-015M-sodiumcitrate), lOmM-dithiothreitol(DDT) at 50°C and subsequently tissue wassubjected to a stringent washing at 60°C in 50% forma-mide, 2x SSC, O I M - D D T . Slides were then rinsed in awashing buffer (Wilkinson et al. 1987) and treated withRNase A (20^gml"') and Tl (2/igmP1) (Boehringer) inwashing buffer for 45min at 37°C. Following washes atroom temperature in 2x SSC (15') and 01 x SSC (15'),respectively, slides were rapidly dehydrated in a series ofethanols containing 0-3M-ammonium acetate. Slides wereprocessed for standard autoradiography using Kodak NTB-2 nuclear track emulsion and exposed for one week (unless

a-actin gene expression in mouse embryos 157
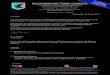
CCAAT TATA I II III IV V VI VII
+ 1
-15 +113
-46 + 127
Cardiac
Skeletal
1 10 20 30 40 SOGOAACAAAAGCrrGCATGCCTGCAGGTCCUCTCTAOAQO^TCCAAGAGCA
60 70 80 90 100CAAAA(X>CACnKnXjACAAGaTCCTGCACGOAG<>ACH3CrGGGACTCAC
UO 120 130 140 ISOCGTGfrrAGGr.GCGGfTGGATTrAGCTGGr.rTGGCGrTGGTGGCAGGCACT
160 170 180 190GGGTOTCAGTCOCCTGGGOATaxxxX3GTA(xGAGCTCGAATTC
1 10 20 30 40 SOGGAACAAAAGCTTCGCIXX:AGGTCGACGGATCCCCTACAAGCCCAAGGCT
60 70 80 90 100CAATAGCTTTCTTrGCTCXKnxrrcCAGGTCACCCTCCCCACCTCCACXCT
110 120 130 140 ISOACrTGrTGrTrTGACTrTGGrrCTGGGTGrCGAGGGTAGGAAGTGAGGfT
160 170 180 190 200If^OmXfilfilOACCGCCTGTCCCCTra^CAOOITrrTATATAGTCCC
210
Fig. 1. (A) Schematic representation ofthe shared intron-exon arrangement ofboth sarcomeric a--actin genes. For eachgene, a probe corresponding to the firstnoncoding exon was subcloned asdescribed in Materials and methods.(B) Sequence of insert used for thear-cardiac actin probe is between the twounderlined BamHl sites. Bluescribe andM13mp8 polylinker sequences are insubscript with the first nucleotide of T3promoter transcripts labelled nucleotide J.The DNA sequence complementary tothe first noncoding exon is underlined inbold type. (C) Sequence of insert used forthe o'-skeletal actin probe and is betweenthe underlined Hindlll and Sacl sites.Bluescribe and M13mp8 polylinkersequences are in subscript. The DNAcomplementary to the first noncodingexon of the skeletal actin gene isunderlined and in bold type.
otherwise indicated) in light-tight boxes with desiccant at4°C. Photographic development was carried out in full-strength Kodak D-19 (3-5' at 16°C). Slides were fixed(Ilford rapid fix), and lightly stained in toluidine blue.Analysis was carried out using both light- and dark-fieldoptics on a Zeiss Axiophot microscope. All grain countingswere carried out on slides from parallel experiments and atleast 100 cells were counted per section. At least threeembryos of both C3H and B ALB/c mice were examined forcomparisons at all stages. Other than for cardiac tissuehybridized with skeletal actin, counts of grains were madeat regions of highest density, thus ratios presented reflectdifferences between highest levels of hybridization.
Preparation of plasmids and cRNA probesBoth sarcomeric actin genes possess very similar structuresconsisting of seven exons (Fig. 1 A). Regions containing thefirst noncoding exons of the ar-cardiac and ar-skeletal actingenes from nucleotides —15 to +113 (Garner et al. 1986)and -46 to +127 (Chien-Tsung Hu et al. 1986, and S.Alonso, unpublished data), respectively, were subclonedfrom ml3mp8 recombinants generated from sonicatedfragments as described in Garner et al. (1986). For the ar-cardiac actin gene, a BamHl fragment containing the aboveregion and eight nucleotides of mp8 polylinker was sub-cloned into Bluescribe plus (Stratagene). For the a'-skeletalactin gene, a Sacl-f//VidIII fragment containing the aboveregion and 28 nucleotides of mp8 polylinker was subclonedinto Bluescribe plus. These sequences have <0-l % hom-ology, are both unique in the mouse genome, and byNorthern analysis only recognize their respective RNAs
(data not presented). The sequence of each insert ispresented in Fig. 1B,C. High specific activity RNA probeswere synthesized using 35S-UTP (specific activity>1000 Ci mmole"', New England Nuclear) according to thesupplier's conditions. Probe length was reduced to a meansize of 50-60 nucleotides by alkali hydrolysis and unincor-porated nucleotides were removed by gel filtration on G-50sephadex. The final exon sequences complementary to thegene transcripts were 50 nucleotides for a'-cardiac(lOUTPs) and 60 nucleotides for ^-skeletal (12 UTPs), thusallowing for direct comparisons of both actin transcripts.
Results
Cardiac development
According to Rugh (1968), the first striated muscle toform during mouse development is the heart. Theheart begins to contract at approximately 8 days p.c.although the heart mesenchymal primordium can beidentified at about 7-0 days p.c. Using specific 35S-cRNA probes corresponding to the 5' noncodingregions of the ar-cardiac and ar-skeletal mRNAs, weobserve a weak hybridization which is discretelylocalized near the head fold at 7-5 days p.c, and by7-8days p.c, both actins are clearly detectablealthough the a--skeletal signal is much weaker(Fig. 2). By 8 days p.c, the primitive heart tubeshows strong hybridization with the cardiac actinprobe, and levels remain high at all subsequent stages

158 D. A. Sassoon, I. Garner and M. Buckingham
of embryonic development examined (see Fig. 6).For skeletal actin transcripts, we observe that levelsincrease from 8 days p.c., although they are always
fr'-' ' * '"
• • t
2A
Fig. 2. Coexpression of cardiac and skeletal a-actintranscripts in early heart (7-8 days p.c). (A) Phase-contrast photomicrograph. Arrows indicate amnion andyolk sac. (B) Dark-field photomicrograph of samesection, hybridized to the cardiac actin probe. (C) Dark-field photomicrograph of adjacent section to B hybridizedto the skeletal actin probe. As compared to cardiac actin(B), the degree of hybridization reveals fewer grainsfollowing an identical exposure time (1 week). Bar,50 j<m.
much lower than those of cardiac actin. We note thatalthough the hydridization with the skeletal actinprobe has overall lower levels in the heart (Fig. 3),there are often clusters of cells or patches that show agrain density which is comparable to that seen withthe cardiac actin probe following identical exposuretimes and hybridization conditions (Fig. 4). Suchskeletal actin hot spots or patches are not obviouslyassociated with particular structures in the heart.Other than regional levels of high skeletal actintranscripts, the average grain density ratio of a-cardiac/a'-skeletal is >25:1.
An additional structure, the dorsal aorta, which isconnected to the heart and spans the rostral-caudalaxis of the embryo during embryonic life is alsoobserved to hybridize weakly with <r-cardiac actin(Fig. 5). We do not observe labelling above back-ground levels with skeletal actin although this mayreflect that, as for the heart, levels of this transcriptare lower than cardiac actin, and thus below the levelof detection.
Skeletal muscle developmentThe majority of skeletal muscle is derived fromsomites, which arise from a segmentation of paraxialmesoderm (e.g. Tarn, 1981; Tarn & Beddington,1987). The first somites can be observed at about 8days p.c. and continue to be generated in a rostral-caudal direction with the last somites (65 in total)generated at 15 days p.c. in the tail (Rugh, 1968; Tarn,1981). Thus, by using serially collected parasagitalsections, we observe a gradient of somite differen-tiation, the most mature somites located rostrally andthe newly established somites and presegmentedmesoderm (somitomeres) at the most caudal location(Fig. 6). At 9 days p.c, embryos possess approxi-mately 14-16 somites (Rugh, 1968; and our obser-vations). Due to body torsions characteristic ofmouse embryos, no one section contains all of thesomites, however, analysis of serially collected tissuesections reveals that all somites show moderate tohigh levels of hybridization with the <r-cardiac actinprobe (Fig. 8) and very weak hybridization with thea-skeletal actin probe (data not shown). The averagegrain density ratio of cr-cardiac/o'-skeletal at themyotomal portion at this stage is >40:l. Presomiticparaxial mesoderm shows no significant labelling,however, we note that the last somite present im-mediately adjacent to presomitic mesoderm showsweak labelling of cells along the inner lining of thesomitocoel prior to its transition from an epithelialstructure to a somite containing an interior compart-ment of cells. Upon initial formation, somites aredivided into at least three compartments: derma-tome, sclerotome and a central packet of cells calledthe myotome (Rugh, 1968). It is the latter group of

a-actin gene expression in mouse embryos 159
Fig. 3. Coexpression of cardiac and skeletal a-actintranscripts in embryonic heart (9 days p.c). (A) Mid-parasagital section hybridized to the cardiac actin probeshowing the early heart-forming presumptive atrial (a)and ventricular (v) compartments. Arrows denote pointsat which cells that are hybridized with the cardiac actinprobe border with cells that do not (see Fig. 5B).(B) Dark-field micrograph of same section showing stronghybridization of cardiac actin probe. (C) Dark-fieldmicrograph of an adjacent section hybridized to theskeletal actin probe. As compared to cardiac actin (B),the degree of hybridization reveals fewer grains followingan identical exposure time (1 week). Bar, 100nm.
cells that are suggested to be the progenitor orfounding cells for skeletal muscle. Labelling in thesomites appears to be restricted to the myotomalportion, thus consistent with the proposed fate of thissomite compartment.
By 10 days p.c. (approximately 30 somites stage)the most anterior somites begin to establishmyotomal muscle, whereas the posterior somites aremorphologically similar to somites observed at 9 daysp.c. At this stage, we observe different stainingpatterns between these two actins at the level of thesomites. Caudal somites at 10 days p.c, whichpossess a similar morphology to the somites at 9 daysp.c, show hybridization of cardiac and to a lesserextent skeletal transcripts in the round cells in thecentral myotomal compartment (Fig. 8) which aresimilar to the cells in this region at 9 days p.c. In someregions of the dermatome, labelling with the cardiacactin probe appears to exceed background levels seenelsewhere in the embryo following longer exposuretimes (see Fig. 8B). It is possible that this reflects lowlevels of transcripts in cells in the dermatome com-partment. Grain-density ratios of Q'-cardiac/a'-skel-etal in the myotome of caudal somites is approxi-mately 5:1. In contrast, somites located rostrallycontain at least two 'myotomal' cell types which canbe distinguished morphologically: the first type areround cells which show high levels of accumulatedcardiac actin transcripts, but show only very lowlevels of hybridization with the skeletal actin probe(ratio ae/ask > 50:1); the second population of cellswhich are located more ventrally and are elongated inmorphology hybridize strongly to the o--cardiac actinprobe, but also show strong hybridization with the a-skeletal actin probe (ratio ac/ask > 4:1) (Fig. 9). Wesuggest that the appearance of the elongated cellscorresponds to the initial establishment of themyotomal muscle whose most differentiated progenyare located in the ventral portion of the somite.Under standard light field optic and phase contrast,the cells appear mononucleate and occasional crossstriations are observed. Muscle fibres are detectedshortly afterwards and at all subsequent stages exam-ined, cr-cardiac and ar-skeletal actin-positive myo-tome muscles increase progressively in the caudalportion of the embryo. By 13-5 daysp.c, somites 1 to50 are replaced by segmental blocks of dermatomeand sclerotome (see Fig. 6), which show conden-sation into vertebrae and ribs as well as segmentedblocks of myotomal muscles whose fibres run pre-dominantly in a dorsal-to-ventral direction. At 11days, ar-cardiac and ar-skeletal actin-positive cells canbe seen at the level of both front and hind limb budssuggesting a very rapid migration of myotomal cellsfrom the somites and subsequent establishment ofmyofibres. The appearance of sarcomeric actin-posi-

160 D. A. Sfissnnn, J Gnrnpr nnd M Buckingham
Fig. 4. Coexpression of cardiac and skeletal a--actin transcripts in embryonic heart (10 days p.c). Adjacent parasagittalsections hybridized to the cardiac actin probe (A,C) and the skeletal actin probe (B,D) reveal strong labelling ofpresumptive right atria (A) and ventricle (C) with the cardiac actin probe whereas labelling with the skeletal actin probeis weak in an adjacent section (B,D). In addition, several regions (arrow) strongly hybridize with the skeletal actinprobe. (E) Phase-contrast image at lower magnification showing entire heart. The arrow denotes the regions presentedin A and B that show highly local hybridization with the skeletal actin probe. (A-D) Bar, 0-05 mm; (E) Bar, 0-15 m.
Fig. 5. Expression of cardiac actin transcripts in the dorsal aorta of 10 days p.c. embryo. (A) Parasagittal sectionhybridized with the cardiac actin probe, v, ventricle; a, atria; nf, neural fold; sc, spinal canal; da, dorsal aorta. Dottedlines ( ) are used to indicate location of somites shown in bright field. (B) Same section with dark-field opticsshowing dense accumulation of grains over somites and heart and a weaker accumulation of grains over the dorsalaorta. Exposure time was 2 weeks. Bar, 0-4 mm.

a-actin gene expression in mouse embryos 161
Fig. 6. Representative mid-parasagittal sections ofembryos hybridized with the cardiac actin probe. Dark-field optics reveal intense labelling of the heart at allstages and increasing intensity of labelling as somitesundergo the transition to myotomal muscle. (A) 9 daysp.c. h, heart; 5, somites. Note that in this section, not allsomites present at this stage are included in this plane ofsection. (B) 10 days p.c. m, myotomal muscle. Also noteweak hybridization of dorsal aorta (unmarked, seeFig. 5). (C) 11 days p.c. Heart has now developed clearlydefined ventricle (v) and atrial (a) compartments. Notethat in this section, anterior myotomes are not included.Bar, 0-5 mm.
tive cells coincides with an observed condensation ofprecartilage mesoderm in the limb bud which mayimply that the processes of chondrogenesis and myo-genesis are coordinately regulated during develop-ment.
Discussion
Sarcomeric actins as early markers for myogenesisWe demonstrate here that cardiac (and to a lesserextent skeletal) actin transcripts accumulate in mono-nucleated cells, in the myotome region of the somites.It is generally considered that muscle actins andmyosins are markers of terminal differentiation ofmuscle tissue (Buckingham, 1985). It has been pro-posed that mesodermal cells undergo a 'commitment'step to become myoblasts (Dienstman & Holtzer,1977; Konigsberg etal. 1978) and then withdraw fromthe cell cycle, undergo fusion and subsequent myo-fibre maturation. The formation of myotubes by cellfusion correlates with a rapid accumulation of musclemRNAs and proteins in the developing fibre (seeBuckingham, 1985). In the cell culture situation, it isclear that cell fusion is not an obligatory step for theexpression of muscle genes. This will take place inmononucleated cells or myocytes when fusion isinhibited, and in muscle cell cultures, where fusionoccurs normally, mononucleated cells expressingmuscle markers are also observed especially just priorto fibre formation (e.g. Emerson & Beckner, 1975).
In studies on developing chicken embryos employ-ing fluorescent antibodies to sarcomeric myosin,Holtzer etal. (1957) first detected staining at the levelof the rostral somites in the elongated myotomalcells. They showed that these cells contracted in thepresence of ATP and that some of the cells containedcross-striations. No staining was detected at earlierstages in these somites, and the caudal somites whichwere presumably less advanced in their developmentwere negative. Jockusch et al. (1984) performedsimilar experiments with antibodies to muscle myo-sin, a-actinin and tropomyosin on mouse embryosand observed that only heart tissue was positive forantibody staining during initial somite formation. Atday 10p.c., they observed weak but positive stainingat the level of the somites although no indication isgiven as to whether all somites or only the mostanterior were positive. In this study, we show thatfrom 9 days p.c. for the more rostral somites of themouse, elongated cells present in the ventral regionof the somite have accumulated (^-skeletal and a-cardiac gene transcripts. They have also accumulatedmuscle myosin heavy chain mRNA (results notshown). We propose that these cells are equivalent tothose described in the somites of the chicken by

162 D. A. Sassoon, I. Garner and M. Buckingham
Holtzer et al. (1957), and that they correspond todifferentiated mononucleated cells or myocyteswhich contribute to the formation of the myotomalmuscles. At this very early stage of muscle formationit is clear that cardiac and skeletal actin gene tran-scripts coaccumulate.
A striking finding from the in situ analysis pre-sented here is that a-cardiac actin is already a majortranscript in the very earliest myotomal cells of themouse somite, at least a day before the first appear-ance of elongated muscle cells. Unlike the latter,these precursor cells show very low accumulation ofcr-skeletal actin transcripts. However, the signaldetected is above background and would suggest thathere also the two striated muscle actin genes arecoactivated. Cardiac actin is therefore a molecularmarker for these early myotomal cells. We do notdetect any hybridization in presomitic paraxial meso-derm. However, this does not exclude the possibilitythat individual cells may be positive. At this level ofresolution, it is difficult to distinguish a weak positivesignal above background by in situ hybridization;expression of a marker gene such as ^-galactosidaseunder the regulation of the cardiac actin promoterwould be easier to follow by conventional stainingprocedures.
The results obtained in tissue culture certainlysuggest that cardiac actin is an early marker ofdifferentiated muscle cells, although there is noevidence that it is transcribed in actively dividingmyoblasts rather than myocytes. Indeed, this posesthe question as to whether myoblasts present in thedeveloping myotome are different from those presentin muscle masses at later stages of development,although almost all myoblasts originate from thesomite. It is possible that myoblasts (or theirprogeny) that migrate out from the myotome are nolonger capable of accumulating high levels of cardiacactin transcripts. We suggest that the myoblastspresent in the somite may represent a distinct class ofmuscle precursor cells which differ from the myo-blasts of myogenic cell lines. There is already evi-dence that the mononucleated precursor cells of fetaland adult muscle differ in regard to their expressionof certain muscle markers (e.g. Cossu et al. 1988;Mouly et al. 1987; Chevallier et al. 1986; Miller &Stockdale, 1986; Vivarelli & Cossu, 1986).
We do not know whether the cardiac actin proteinaccumulates in myotomal cells, although given theclose correlation between actin mRNA and protein inall other situations examined (Vandekerckhove et al.1986), this seems probable. Should this be the case,the role of cardiac actin in the apparent absence ofother muscle isoforms in such cells, is unclear.Alternatively, if cardiac actin does not have a func-tional role, it is possible that regulatory events
leading to the establishment of early myogenic cellswill result in the accumulation of cardiac actin tran-scripts whereas additional steps may be required forthe accumulation of other muscle-specific transcripts.The observation that cardiac rather than skeletalactin is the major early transcript from this gene pairmay reflect the evolutionary history of these twogenes. It has been suggested that cardiac actin is theancestral sarcomeric actin type on the basis of itspresence in the Pleurodeles, in the apparent absenceof the skeletal actin isoform (see Alonso, 1987). InXenopus laevis, both skeletal- and cardiac-like actinsequences are present, but these (genes al and a3)are coexpressed in the adult cardiac and skeletaltissues as well as during development, indicating thatthe phenomenon of coexpression probably precededthat of tissue specificity for this actin gene pair (Stutz& Spohr, 1986).
Sarcomeric actin expression in the heart temporallyprecedes that in skeletal muscle duringembryogenesisWe show here that cardiac actin transcripts areaccumulated already in the cardiac tube of mouseembryos. At this and subsequent stages, skeletal actintranscripts are also present in relatively low amounts,again suggesting that these genes are probably coacti-vated during development. The expression of thesarcomeric actins in the heart is of interest since this isone of the first adult tissues to be established in thedeveloping mouse embryo. The heart plays a criticalrole in feeding and removing wastes from the rapidlygrowing embryo, which may explain our observationsthat cr-cardiac/skeletal actin as well as )3MHC (datanot shown) can be detected as early as 7-8 days p.c. inthe heart. Using immunofluorescent techniques withmyosin antibodies, the presence of cardiac myosinhas also been detected in the cardiac tube of thechicken as early as stage 10 (li days) in ovo (Sweeneyet al. 1987). Unlike the somites, which present agradient of differentiated states in the post-8-dayp.c.embryo, we cannot determine for certain whether thetranscripts for the sarcomeric actins precede actualmyocyte differentiation. Previous studies (Snow,1981) using isolated in vitro preparations of earlymouse embryos (<7 days p.c.) have determined thateither cells or an inductive wave responsible for theestablishment of the heart primordium originate atthe posterior end of the embryo during primitivestreak formation. In complete series of 6-5-7-25 daysp.c. embryos, we do not observe any localized hybrid-ization although for reasons not yet understood, ourcRNA probe to cardiac actin produces markedlyhigher background in 7-0-7-5 days p.c. embryos thanat later stages (data not shown).
In addition to hybridization in the cardiac tube and

Fig. 7. Expression of cardiac actin transcripts in somites of a 9 days p.c. embryo. (A) Bright-field view of caudalsomites hybridized to the cardiac actin probe. Nuclei are stained light blue, d, dermatome, m, myotome, s, sclerotome.(B) Same section viewed with dark-field optics revealing dense accumulation of grains over myotomal (m) region.Bar, 50 /im.
d
m
§A
Fig. 8. Coexpression of cardiac and skeletal actintranscripts in caudal somites of a 10 days p.c. embryo.(A) Bright-field photomicrograph of somites hybridizedwith the cardiac actin probe. Nuclei are stained lightblue, cl, dermatome; HI, myotome; s, sclerotome.(B) Same section viewed with dark-field optics revealingdense accumulation of grains over myotomal (m) region.(C) Adjacent section hybridized with the skeletal actinprobe, again showing accumulation of grains overmyotomal region. Although these somites are reminiscentin morphology of somites at 9 days p.c, expression of theskeletal actin transcripts is much higher by comparison.Exposure times for both probes were extended to twoweeks, resulting in higher background in nonmyotomalregions of somites (see Fig. 7 for comparison). Bar,30 Jim.

M# *JT f4h. f*> n. « •
Fig. 9. Coexpression of cardiac and skeletal actin transcripts in rostral somites undergoing the transition to myotomalmuscle (same 10 days p.c. embryo shown in Fig. 8). (A) Bright-field view of somites hybridized with the cardiac theactin probe. Nuclei are stained light blue, d, dermatome; m, myotome. The arrow indicates level below whichmyotomal cell nuclei and cells (using phase contrast, data not shown) are elongated. (B) Same section viewed withdark-field optics revealing dense accumulation of grains over myotomal (in) region above and below arrow.(C) Immediately adjacent section hybridized with the skeletal actin probe viewed with bright-field optics. Arrowindicates level below which myotomal cell nuclei (stained here) and cells (using phase contrast, data not shown) areelongated. (D) Dark-field image of same section again showing accumulation of grains over myotomal region, howeverwe note that skeletal actin transcripts are less abundant or absent in cells above arrow. Exposure times for both probeswere extended to two weeks resulting in higher background in non-myotomal regions of somites. Serial sections throughmyotomal muscle consistently reveal a more dorsal extent of cardiac actin transcripts within the presumptive myotomeas compared to skeletal actin transcripts. Bar, 30/tm.

a-actin gene expression in mouse embryos 163
maturing heart, we also detect lower but significanthybridization with the cardiac actin probe, in thedorsal aorta. To our knowledge, the dorsal aorta hasnot been reported to play an active role in contractionin the mouse embryo. It is tempting to speculate thatduring early embryogenesis in the mouse, the heart,which functions as a simple tube (see VanMierop,1979) similar to that found in primitive vertebrates(Romer, 1955), is aided by contraction in the dorsalaorta. An alternative explanation may be that thecommon embryonic origin of the heart and dorsalaorta (see VanMierop, 1979) may result in a transientexpression of the myogenic programme in an ectopicpopulation of cells present in the dorsal aorta.
The fact that cardiac and skeletal actin transcriptsaccumulate in early cardiac structures prior to theirexpression in the myotome, and that transcripts arenot detected in the presomitic mesoderm wouldsuggest that the embryonic or in utero environmentdoes not 'globally' induce sarcomeric actin expressionin presumptive muscle tissue, but that cardiac andskeletal muscle follow independent developmentalprogrammes. The developmental programmes of car-diac and skeletal muscle precursor cells are thereforenot temporally synchronized. The observationsreported here that cr-cardiac actin (and to a lesserextent a--skeletal actin) transcripts are accumulated inskeletal muscle precursor cells of the somite indicatesthat the accumulation of these transcripts precedesthe terminal differentiation of myoblasts in vivo andtheir presence thus serves as a marker not only forearly striated muscle but also for their anlagen. Ourresults indicate that the early expression of bothsarcomeric actin genes is a characteristic part of theearly myogenic programme in the mouse embryo.
We are grateful to our colleagues for advice and encour-agement during the course of these experiments andespecially to Dr Serge Alonso for the skeletal actin frag-ment and Philippe Billen for assistance with tissue prep-aration. We thank also Drs Stefano Schiaffino, AndrewMcMahon, David Wilkinson, Juliet Bailes and Janet Cham-pion for their advice on in situ hybridization. D.S. is arecipient of a postdoctoral fellowship grant from theAmerican Cancer Society. I.G. is a recipient of a researchfellowship from the Association Franchise contre les Myo-pathies. This work was supported by the French Ministry ofResearch, the Centre National de la Recherche Scientifiqueand the Institut National de la Sant6 et de la RechercheM6dicale and the Muscular Dystrophy Association ofAmerica.
References
ALONSO, S. (1987). Coexpression and evolution of the
two sarcomeric actin genes in vertebrates. Biochimie69, 1119-1125.
BAINES, W., PONTE, P., BLAU, H. & KEDES, L. (1984).
Cardiac actin is the major actin gene product inskeletal muscle cell differentiation in vitro. Molec. Cell.Biol. 4, 1449-1451.
BUCKINGHAM, M. E., ALONSO, S., BARTON, P., BUGAISKY,
G., COHEN, A., DAUBAS, P., GARNER, I., MINTY, A.,
ROBERT, B. & WEYDERT, A. (1984). Actin and myosingenes, and their expression during myogenesis in themouse. In Molecular Biology of Development, vol. 19(ed. E. H. Davidson & R. A. Firtel), pp. 275-292.New York: Alan R. Liss, Inc.
BUCKINGHAM, M. E. (1985). Actin and myosin multigenefamilies: their expression during the formation ofskeletal muscle. In Essays in Biochemistry 20, 77-109.
CASCIO, A. & GURDON, J. B. (1987). The initiation ofnew gene transcription during Xenopus gastrulationrequires immediately preceding protein synthesis.Development 100, 297-305.
CHEVALLIER, A., PAUTOU, M. P., HARRIS, A. J. & KIENY,
M. (1986). On the non-equivalence of skeletal musclesatellite cells and embryonic myoblasts. In Archivesd'Anatomie microscopique 75, pp. 161-166.
CHIEN-TSUNG HU, M., SHARP, S. B. & DAVIDSON, N.
(1986). The complete sequence of the mouse skeletalor-actin gene reveals several conserved and invertedrepeat sequences outside of the protein-coding region.Molec. Cell. Biol. 6, 15-25.
Cossu, G., RANALDI, G., SENNI, M. I., MOLINARO, M. &
VIVARELLI, E. (1988). "Early" mammalian myoblastsare resistant to phorbol ester-induced block ofdifferentiation. Development 102, 65-69.
DIENSTMAN, S. R. & HOLTZER, H. (1977). Skeletalmyogenesis: Control of proliferation in a normal celllineage. Expl Cell Res. 107, 355-364.
EMERSON, C. P. & BECKNER, S. K. (1975). Activation ofmyosin synthesis in fusing and mononucleatedmyoblasts. J. Mol. Biol. 93, 431-447.
GALL, J. G. & PARDUE, M. L. (1971). Nucleic acidhybridization in cytological preparations. MethodsEnzymol. 21,470-480.
GARNER, I., MINTY, A. J., ALONSO, A., BARTON, P. J. &
BUCKINGHAM, M. E. (1986). A 5' duplication of the a'-cardiac actin gene'in BALB/c mice is associated withabnormal levels of a--cardiac and a-skeletal actinmRNAs is adult cardiac tissue. EMBO J. 5, 2559-2567.
GUNNING, P., HARDEMAN, E., WADE, R., PONTE, P.,
BAINS, W., BLAU, H. M. & KEDES, L. (1987).
Differential patterns of transcript accumulation duringhuman myogenesis. Molec. Cell. Biol. 7, 4100—4114.
HAYWARD, L. J. & SCHWARTZ, R. J. (1986). Sequentialexpression of chicken actin genes during myogenesis. J.Cell Biol. 102, 1485-1493.
HOLTZER, H., MARSHALL, J. M. JR & FINCK, H. (1957).
An analysis of myogenesis by the use of fluorescentantimyosin. J. Biophys. Biochem. Cytol. 3, 705-723.
JOCKUSCH, H., MOLLER, U. & JOSKUSCH, B. M. (1984).
Accumulation and spatial distribution of structuralproteins in developing mammalian muscle. Expl Biol.med. 9, 121-125.

164 D. A. Sassoon, I. Garner and M. Buckingham
KONIGSBERG, I. R . , SOLLMAN, P . A . & MlXTER, L.
O. (1978). The duration of the terminal Gl offusing myoblasts. Devi Biol. 63, 11-26.
MAYER, Y., CZOSNECK, H., ZEELON, P. E., YAFFE,
D. & NUDEL, U. (1984). Expression of the genescoding for the skeletal muscle and cardiac actinsin the heart. Nucl. Acids Res. 12, 1087-1100.
MILLER, J. B. & STOCKDALE, F. E. (1986).
Developmental origin of skeletal muscle fibers:clonal analysis of myogenic cell lineages based onfast and slow heavy chain expression. Proc. natn.Acad. Sci. U.S.A. 83, 3860-3864.
MINTY, A. J., ALONSO, S., CARAVATTI, M. &
BUCKINGHAM, M. (1982). A fetal skeletal muscleactin mRNA in the mouse, and its identity withcardiac actin mRNA. Ce//30, 185-192.
MINTY, A., BLAU, H. & KEDES, L. (1986). Two-
level regulation of cardiac actin genetranscription: Muscle-specific modulating factorscan accumulate before gene activation. Molec.Cell. Biol. 6, 2137-2148.
MOHUN, T. J., BRENNAN, S., DATHAN, N.,
FAIRMAN, S. & GURDON, J. B. (1984). Cell type-specific activation of actin genes in the earlyamphibian embryo. Nature, Lond. 311, 716-721.
MOULY, V., TOUTANT, M. & FlSZMAN, M. Y.(1987). Chick and quail limb bud myoblasts,isolated at different times during muscledevelopment, express stage specific phenotypeswhen differentiated in culture. Cell Diff. 20,17-25.
NIEUWKOOP, P. D. & FABER, J. (1956). Normal tableo/Xenopus laevis (Daudin). Amsterdam: NorthHolland.
ORDAHL, C. P. (1986). The skeletal and cardiac o--actin genes are coexpressed in early embryonicstriated muscle. Devi Biol. 117, 448-492.
PATERSON, B. M. & ELDRIDGE, J. D. (1984). a-
cardiac actin is the major sarcomeric isoformexpressed in embryonic avian skeletal muscle.Science 224, 1436-1438.
ROMER, A. S. (1955). The Vertebrate Body 2ndedition. Philadelphia: W. B. Saunders Co.
RUGH, R. (1968). The Mouse: Its Reproduction and
Development. Minneapolis, Minesota: Burgess.SCHWARTZ, K., DE LA BASTIE, D., BOUVERET, P.,
OLIVI^RO, P., ALONSO, S. & BUCKINGHAM, M.
(1986). ^-skeletal muscle actin mRNA'saccumulate in hypertrophied adult rat hearts.Circulat. Res. 59, 551-555.
SNOW, M. H. L. (1981). Autonomous developmentof parts isolated from primitive-streak-stagemouse embryos. Is development clonal? J.Embryol. exp. Morph. 65, 269-287.
STUTZ, F. & SPOHR, G. (1986). Isolation and
characterization of sarcomeric actin genesexpressed in Xenopus laevis embryos. J. molec.Biol. 187, 349-361.
SWEENEY, L. J., ZAK, R. & MANASEK, F. J. (1987).
Transitions in cardiac isomyosin expression duringdifferentiation of the embryonic chick heart.Circulat. Res. 61, 287-295.
TAM, P. P. L. (1981). The control of somitogenesisin mouse embryos. J. Embryol. exp. Morph. 65,103-128.
TAM, P. P. L. & BEDDINGTON, R. S. P. (1987). Theformation of mesodermal tissues in the mouse •embryo during gastrulation and earlyorganogenesis. Development 99, 109-126.
VANDEKERCKHOVE, J., BUGAISKY, G. &
BUCKINGHAM, M. (1986). Simultaneous expressionof skeletal muscle and heart actin proteins invarious striated muscle tissues and cells. J. biol.Chem. 261, 1838-1843.
VAN MIEROP, L. H. S. (1979). Morphologicaldevelopment of the heart. In Handbook ofPhysiology: The Cardiovascular System (ed. R.M. Berne, N. Sperelakis & S. R. Geiger), pp.1-28. Bethesda, USA: American PhysiologicalSociety.
VIVARELLI, E. & Cossu, G. (1986). Neural controlof early myogenic differentiation in cultures ofmouse somites. Devi Biol. 117, 319-325.
WILKINSON, D. G., BAILES, J. A., CHAMPION, J. E.
& MCMAHON, A. P. (1987). A molecular analysisof mouse development from 8 to 10 days postcoitum detects changes only in embryonic globinexpression. Development 99, 493-500.
(Accepted 31 May 1988)