Embed Size (px)
Citation preview

Indian Journal of Experimental Biology Vol. 38, January 2000, pp. 6-17
Review Article
Transgenic strategies for genetic improvement of Basm~ i rice
Rajinder K. Jainl* & Sunita Jain2
IDepartment of Biotechnology and Molecular Biology, 2Department of Biochemistry, CCS Haryana Agricultural University, Hisar 125004, India
Transgenic approach offers an attractive alternative to conventional techniques for the genetic improvement of Basmati rice because they enable the introduction of one or more genes into a leading cultivar without affecting its genetic background. During the last ten years, a rapid progress has been made towards the development of transformation methods in rice. Several transformation methods including Agrobacterium, biolistic, and DNA uptake by protoplasts, have heen employed to produce transgenic rice. An array of useful genes is now available and many of these have already been transferred in rice to improve the resistance against biotic and abiotic stresses. In Basmati rice, a beginning has already been made regarding the development of tissue culture protocols, transformation methods and production of useful transgenic plants. The application and future prospects of transformation technology to engineer the resistance against insect pests (stem horer, leaf folder, brown plant hopper, gall midge), fungal diseases (blast, bakanaelfoot. rot), bacterial di seases (bacterial leaf blight , sheath blight), abiotic stresses (salinity and drought) and improved nutritional quality (accumulation of provitamin A and essential amino acids in endosperm) in Basmati rice, have been addressed.
Introgression of specific genes/ traits from various genetic resources into Basmati rice through conventional breeding based on sexual hybridization, genetic recombination and selection, has been cumbersome, time consuming and expensive. Transformation approaches offer an attractive alternative to the conventional techniques to introduce one or more novel useful genes to improve Basmati rice without disrupting their otherwise desirable genetic make-up. Improvement in the transformation ' technology for rice including indicas, has been remarkable in the past few years l
•2
• In addition to the direct gene-delivery methods (protoplast transformation, biolistic, electroporation), rice can also be transformed efficiently using the Agrobacterium method. Genes for several important traits are now available that can be transferred in to Basmati rice to improve its resistance/tolerance against insect pests, fungal diseases, drought and salinity and to improve its nutritional quality . This review article deals with the application of transgenic technology for the genetic improvement of Basmati rice.
Basmati rice breeding: present status, research priorities and problems
Among the various aromatic rice types available throughout the world, Basmati rice is the most pre-
*Fax: 91-1662-33884 E-mail: [email protected]
ferred3-5
. Basmati rice is characterized by superfine long slender grain, exquisite aroma, sweet taste, soft texture, delicate curvature and extra elongation with a least breadth-wise swelling on cooking. Cooked Basmati rice is non-sticky, and has longer shelf life and is easily ' digestible. Cooking and eating qualities of milled rice are mainly due to its specific starch properties; t 6-25% amylose content, intermediate to low starch gelatinization temperature, and medium to soft gel consistency. Basmati rice commends high premium in India as well as in International market. The value of export of Basmati rice from India has increased from t 53 million US $ in 1986-87 to 337 million $ in 1996-97. This accounts for a 5% of the total export of agricultural and allied products in t 996-97. The Basmati rice cultivars have long grains and a morphologically distinct phenotype that has been classified as part of the indica subspecies. However, isozyme data have shown that Basmati rice varieties (Group 5) are closer to japonica (Varietal Group 6) than indica (Group I ) rice varieties6
. Cultivation of Basmati rice is confined to the northwestern states of India (Haryana, Punjab, Himachal Pradesh , Western Uttar Pradesh and Jammu and Kasmir) and Pakistan.
Basmati rice varieties have a poor plant type (tall plant stature, light green and non-erect leaves) and are photoperiod sensitive and poor yielding. Basmati rice is host to a number of insect pests and diseases3.4.

JAIN & JAIN: TRANSGENIC STRATEGIES FOR GENETIC IMPROVEMENT OF RICE 7
Blast and bakanae/foot rot are important among the fungal diseases while stem borer and leaf folder are major insect pests. Though susceptible, incidence of bacterial leaf blight and sheath blight has been rare as these disease results from the use of nitrogenous fertilizers in higher doses, a practice, which is otherwise detrimental to Basmati crop because of lodging. No major outbreak of tungro, dwarf and stunt viral diseases have been reported in most of traditional Basmati rice varieties. However, not much data is available to show that Basmati rice varieties have true resistance against these viral diseases. Basmati varieties are moderately susceptible to sheath rot. Insect pests, such as yellow stem borer and leaf folder, brown plant hopper and gall midge invariably affect Basmati and cause huge yield losses. However, some of the Basmati rice varieties have been reported to be resistant to white backed plant hopper. Basmati rice collections are found susceptible to all major abiotic stresses (drought, salinity).
Basmati rice breeding has been difficult due to complicated nature of quality traits and poor combining ability of Basmati rice varieties. The inheritance of grain quality is very complicated due to epistasis, maternal and cytoplasmic effects, and triploid nature of endosperm? When Basmati rice varieties are crossed with dwarf, high yielding indica rice parents, the hybrids show a high degree of sterility thus preventing many desirable segregants to appear in the segregating population. The genetics of each of the Basmati grain quality components (aroma, intermediate amylose content, intermediate gel consistency and high grain elongation) may not be very complex but to find a desirable segregant possessing all these components require screening of segregants at a very large scale. The grain quality traits are also tightly linked to poor Basmati rice plant type. The breeding strategy adopted to combine Basmati rice grain quality and improved plant type and yield potential, has been quite cumbersome. It involved the crosses between the Basmati rice and dwarf, higher yielding non-Basmati parents and the selection of lines with improved plant type having one or two components of Basmati grain quality. These lines were then intercrossed to develop lines that have higher yield potential with all the components of Basmati grain quality. To give an example, this breeding approach has been used to develop Pusa Basmati I (a semi-dwarf, relatively photoperiod insensitive, high yielding Basmati variety) from the
Pusa 150 (indica) x Karnallocal (Basmati) crosses .
Application of transgenic technology in Basmati rice breeding
A rapid progress has been made towards the development of genetic transformation technology in rice. Initially, rice transformation was carried out by the direct gene-delivery methods such as DNA uptake into protoplasts by polyethylene glycol (PEG) treatment or electroporation and microprojectile bombardment. Hiei et af. 8 reported that rice could also be efficiently transformed using Agrohacteriummediated gene transfer method . Most success regarding rice transformation has been achieved in case of japonica rice cultivars where a number of potentially useful genes have been transferred and in some cases ttansgenic plants have been field tested for a number of years. Subsequently, a number of indica, javanica and Basmati rice varieties have also been transformed. Potentially useful genes has been isolated that may have a function in improving resistance against various biotic and abiotic stresses that severely affect the plant productivity lJ . Many of these genes have already been transferred in model plant species like tobacco, Arahidopsis, or rice and the transgenic plants have been field-tested for transgenic trait. Transformation technology can greatly strengthen Basmati rice breeding programs by transferring these novel genes to improve its resistance against insect/pests, fungal diseases, salinity, drought, etc. The other advantage of transformation is that a useful trait for example resistance to yellow stem borer, 'can be engineered in Basmati rice without altering its genetic background/quality traits which has been difficult to achieve through sexual hybridization involving crosses with the non-Basmati rice varieties .
A. Plant regeneration from cells, tissues and protoplasts
Before targeting a particular rice variety to ill vitro genetic transformation, there is a need to develop efficient procedure(s) for regeneration of green, fertile plants from explant tissues, cells or protoplasts with minimal somaclonal variation. Indeed, it is the totipotency of plant cells that underlies the efficiency of most plant transformation systems. Japonica rice varieties have shown to be more responsive to in vitro culture and plant regeneration than indica rice varieties. As a result, it has been easier to transform japonica rice varieties. However, a lot of research has

8 INDIAN J EXP BIOL, JANUARY 2000
been done to develop tissue culture protocols in indica rice varieties to achieve a workable efficiency of gene transfer, selection and regeneration of transformants. Most success has been achieved in this regard by using either the explants such as immature embryos with many regenerable cells or freshly initiated scutella-derived embryogenic calli . The choices were made based on the easy and round the year availability of the explant tissues with the option to keep tissue culture regime to the minimum. Detailed consideration of optimization of tissue culture systems useful for rice transformation is beyond the scope of this review, but the progress made and protocols developed for rice tissue culture, have been addressed in several reviews and technical manuals l,loo I2.
A number of factors such as the plant genotype and explant type, composition of the medium including carbohydrates and growth regulators, presence of nurse cells, partial desiccation, cryopreservation and water stress treatments, have been examined in attempts to break the recalcitrance of Basmati type indica rice varietiesl3018. In Basmati rice, immature embryos and embryogenic calli or cell suspensions initiated from mature/ immature seed scutella and microspores, have been used for in vitro genetic manipulation experiments. In general, addition of 2,4-0 to the medium causes callus induction and proliferation in rice explants, cells and tissues, whereas plant regeneration occurred in either hormone-free media or media containing a combination of auxin (NAA) and cytokinin (BAP or kinetin). Shoots are generally rooted in hormone-free media or an auxin (NAA) containing medium.
Jain et al. 13 reported an improved procedure for the protoplast culture of several rice varieties including Pusa Basmati I; the procedure involved plating of protoplasts isolated from embryogenic cell suspensions on the surface of filter membranes overlaying agarose-embedded feeder cells of OryZG ridleyi and Latium muitiflorum. Nurse cultures were essential for protoplast culture. L. muitiflorum feeder cells induced a 6-fold higher plating efficiency than feeder cells of O. ridieyi; the two types of feeder cells when used in combination showed an additive effect on protoplast plating efficiency. However, somaclonal variation and sterility problems marred the application of the protoplast system for transgenic plant production. Jain et al. 14 reported maltose to be preferential carbon source compared to sucrose, glucose, fructose, etc. for both somatic embryogenesis and plant regeneration
from cell suspension and protoplast-derived calli of Basmati rice varieties. Jain et ai. 16 reported severalfold increase in frequency of shoot regeneration fol~ lowing water stress treatment (partial desiccation for 24 hr, use of 1.0% agarose instead of 0.5% for medium solidification or O.4M mannitol-containing medium) of cell clumps in Pusa Basmati and Basmati 385 .. Best regeneration frequencies (54-98%) were obtained when 24 hr-desiccated calli were grown 011 regeneration with 1.0% (w/v) agarose. Jain et al.15
reported a simple freeze preservation procedure for the cryopreservation of embryogenic cells of Pusa Basmati I and Basmati 385; the cryopreservation treatment also led to the enrichment of embryogenic cells. Khanna and Raina l
!) reported that ill vitro green plant regeneration efficiency could be greatly enhanced through modification of nitrate-nitro~en and ammonium-nitrogen concentrations in the callusing medium. The highest frequency of plant regeneration (100%) and a maximum number green plants (7 per callus) were obtained in calli derived from the medium having 35 mM KNO~ and 5 mM (NH4h SO.j. Bishnoi et al. 20 reported efficient production of androgenic calli in several Basmati and non-Basmati indica rice varieties and their FI . hybrids/ F:~ plants using an improved anther culture medium (modified RZ21 medium). The calli regenerated green shoots with higher frequencies suggesting that androgenic calli can also be used as the starting material for transformation.
B. Gene transfer methods Protoplast-based direct gene transfer. biolistic
transformation and Agrobacterium-mediated gene transfer are the major techniques that are being routinely used for transgenic plant production in rice. Other methods such as tissue electroporationn .n , laser mediation24, pollen tube pathwa/' , DNA uptake by imbibition of dry embryos26, silicon carbide fibermediated transformation27
, etc . have been reported to produce the transgenic rice plants but have not found widespread use up to now. Several excellent re-
. I 2 d I b I P ox fO. vIews' an a oratory manua s -.- on trans ormatIOn methods have been published. There are still many problems that have to be solved in terms of reproducibility . Most of the above mentioned methods could deliver DNA into the cell, but the subsequent events in cells, nuclei and organelles are not controlled and the integration of foreign DNA is random. Gene silencing and interactions between multiple

JAIN & JAIN: TRANSGENIC STRATEGIES FOR GENETIC IMPROVEMENT OF RICE 9
copies of the same trans gene or different transgenes result in unexpected expression pattern of foreign genes29
-3J
_ Several independent transformants with a specific gene cassette are necessary, to find one transgenic plant with the desired transgene expression pattern and level.
Protoplast-mediated trallsjormatiofl-The first approach to transfer foreign DNA into rice cells was the direct gene transfer into protoplasts. Both polyethylene glycol (PEG) and electroporation methods have been used for rice transformation . Protoplasts are the ideal material for gene transfer. While the delivery of DNA into protoplasts has been quite easy, major problems relate to the establishment of embryogenic cell suspension cultures (source of protoplasts) and regeneration of green, fertile plants from protoplasts. Protoplast culture and regeneration procedure(s) are quite labor intensive and time consuming; for example it may take up to six months to establish embryogenic cell suspensions that are ideal for protoplast isolation. This method is strongly genotype dependent and has not been worked out for many elite indica rice varieties. Jain et at. J3 reported that protoplast culture in two indica rice varieties, Pusa Basmati I, Jaya, required the use of specific nurse cultures. In many cases, transgenic rice plants produced through protoplast transformation were reported to be sterile, carry extensive somaclonal variation, or multiple copies of the transgene. Inspite of these limitations, protoplasts have been successfully used for the production of transgenic plants containing agronomically useful genes in some responsive japonica and indica rice varieties34
.35 (Table I). This method of transfor
mation is no longer a method of choice for rice transformation. However, protoplasts may have an advantage to accomplish the transfer of large segments of DNA and organelles into a single cell.
Biolistic trallsjormation-Biolistic method employs high-velocity metal particles to deliver biologically active DNA into regenerable plant cells, tissues and organs. This method depends far less on sophisticated tissue culture procedures and provides a fairly genotype-independent transformation system bypassing Agrobacterium host specificity and protoplast-related regeneration difficulties. Chances of tissue culture raised somaclonal variation can also be minimized using the explants exhibiting direct shoot regeneration with a minimum of callusing phase. The transformation of several elite indica rice vanetles became possible due to the extended range of cell
tissues/ explants that can be targeted by microprojectiles. In rice, biolistic method has been successfully used for the transfer of many useful genes in over 40 japoniea, indica and Basmati rice varieties'6 (Table I). In most cases, embryogenic calli or suspension cells derived from immature and mature embryos, were used as 'the target tissue. Mostly, ~us gene has been used as the reporter gene and hygR and har genes have been used as selectable marker genes for bioi istic transformation. A number of factors including osmotic pre-conditioning of the target tissues, nature and size of metallic particles used for DNA delivery, humidity and temperature, DNA loading on microprojectiles, target cell distance, accelerating force and depth of penetration, have been reported to influence the biolistic transformation frequency 10,37. Biolistic has also been used for organelle transformation especially the plastids which is an attractive target for crop engineering due to the high level transgene expression,g,39. Besides plasmids, RNA, Y AC (yeast artificial chromosome) DNA clones, or even E. coil or Agrobacterillll1 cells have been successfully used for biolistic transformation40
. Biolistic method' has some drawbacks including the high cost of the equipment and consumables, integration of transgene in high copy number, transgene rearrangement, and gene silencing.
Jain et al. 17•IS reported an improved, reproducible
biolistic procedure for the transformation of embryogenic suspension cells/calli of Pusa Basmati I. The procedure involved the osmotic pre-conditioning of cells for 24 hr on a medium supplemented with 0.25M mannitol prior to bombardment, use of gold particles for DNA delivery, and use of plant regeneration medium with 1.0% agarose. This procedure consistently produced over 600 transient transformants and at least five fertile plants showing integrative transformation per bombarded filter. Minhas et al. 41 reported the optimum parameters for the biolistic introduction of GlIS (uidA) reporter gene into embryogenic callus cultures of Basmati 370 rice variety . Khanna et al.42
reported the biolistic procedure for the transformation of a Basmati rice cultivar Kamal Local lIsing the plasmid carrying Gus and bar gene cassettes, five-day precu1tured mature embryos and glufosinatecontaining liquid medium for selection,
Agrobacterium -mediated trallsjormation-Severalmonocots including rice, are now among the plants that can be transformed routinely by A, tumejaciens.
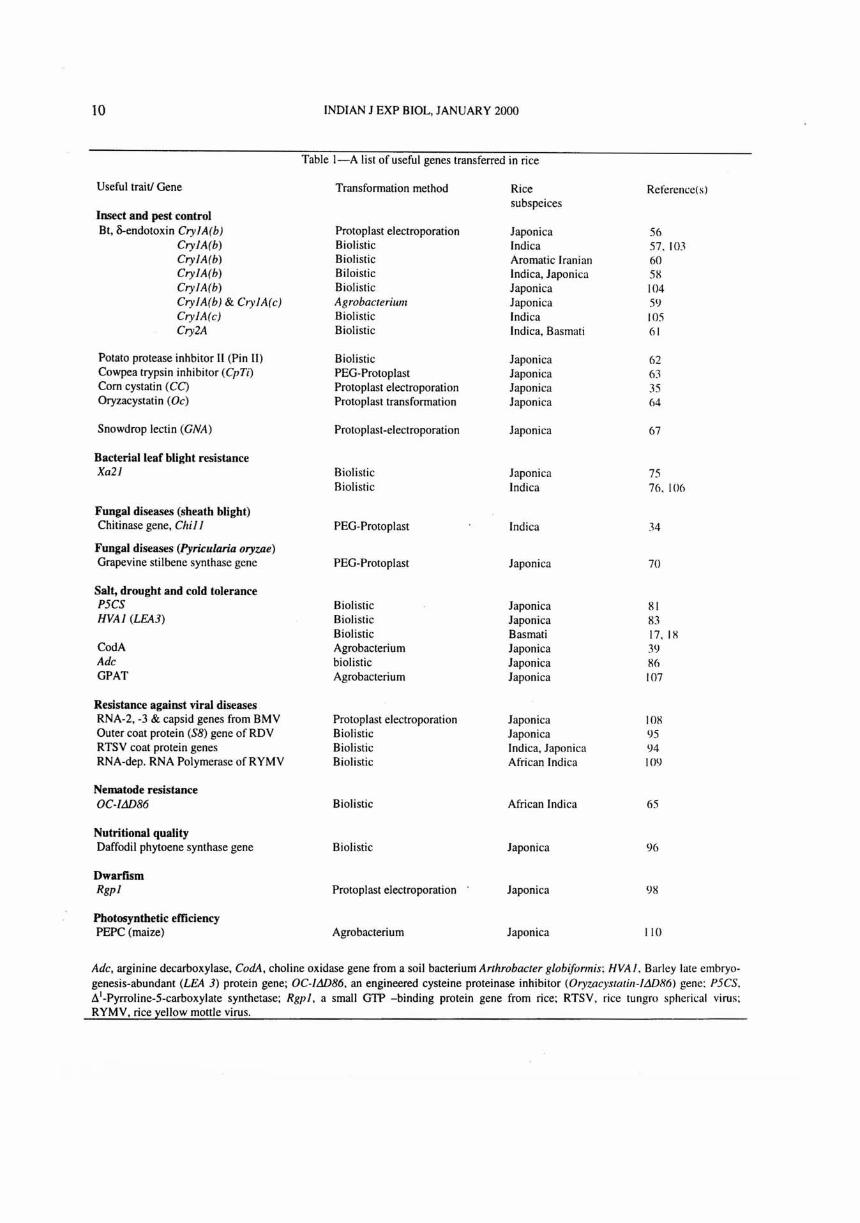
10 INDIAN J EXP BIOL, JANUARY 2000
Table I-A list of useful genes transferred in rice
Useful trait! Gene Transformation method Rice Reference( s) subspeices
Insect and pest control Bt, S-endotoxin Cry/Arb) Protoplast electroporation Japonica 56
Cry/Arb) Biolistic Indica 57.103 Cry/Arb) Biolistic Aromatic Iranian 60 Cry/Arb) Biloistic Indica, Japonica 5H Cry/Arb) Biolistic Japonica 104 Cry/Arb) & Cry/Arc) Agrobacterium Japonica 59 Cry/A (c) Biolistic Indica 105 Cry2A Biolistic Indica, Basmati 61
Potato protease inhbitor II (Pin II) Biolistic Japonica 62 Cowpea trypsin inhibitor (CpTi) PEG-Protoplast Japonica 63 Corn cystatin (CC) Protoplast electro po ration Japonica 35 Oryzacystatin (Oc) Protoplast transformation Japonica 64
Snowdrop lectin (GNA) Protoplast-electroporation Japonica 67
Bacterial leaf blight resistance Xa2/ Biolistic Japonica 75
Biolistic Indica 76. 106
Fungal diseases (sheath blight) Chitinase gene, Chil / PEG-Protoplast Indica 34
Fungal diseases (Pyricu/aria oryzae) Grapevine stilbene synthase gene PEG-Protoplast Japonica 70
Salt, drought and cold tolerance P5CS Biolistic Japonica 81 HVA/ (LEA3) Biolistic Japonica 83
Biolistic Basmati 17. IX CodA Agrobacterium Japonica 39 Adc biolistic Japonica 86 GPAT Agrobacterium Japonica 107
Resistance against viral diseases RNA-2, -3 & capsid genes from BMV Protoplast electroporation Japonica lOX Outer coat protein (S8) gene of RDV Biolistic Japonica 95 RTSV coat protein genes Biolistic Indica, Japonica 94 RNA-dep. RNA Polymerase ofRYMV Biolistic African Indica 109
Nematode resistance OC-ltID86 Biolistic African Indica 65
Nutritional quality Daffodil phytoene synthase gene Biolistic Japonica 96
Dwarfism Rgpl Protoplast electroporation Japonica 9H
Photosynthetic efficiency PEPC (maize) Agrobacterium Japonica 110
Adc, arginine decarboxylase, CodA, choline oxidase gene from a soil bacterium Arthrobacter globiformis; HVA /. Barley late embryogenesis-abundant (LEA 3) protein gene; OC-ltID86, an engineered cysteine proteinase inhibitor (Oryzacystatill-ltID86) gene: P5CS. t1 1-Pyrroline-5-carboxylate synthetase; Rgpl, a small GTP -binding protein gene from rice; RTSV. rice tungro spherical virus ; RYMV, rice yellow mottle virus.

JAIN & JAIN: TRANSGENIC STRATEGIES FOR GENETIC IMPROVEMENT OF RICE II
Agrobacterium-mediated gene transfer system is becoming the method of choice in rice due to its low costs, convenience, high transformation efficiency, high probability of single copy integration with minimal rearrangements, the transfer of relatively large segments of DNA and high fertility of transgenic plants. In 1994, Hiei et al. 8 reported an efficient Agrobacterium-mediated transformation procedure in japonica rice. Since then a rapid progress has been made towards the development of Agrobacteriummediated transformation protocols in various rice varieties including indicas and several useful genes have been transferred using this approach43 (Table I). In general, the procedure involved the cocultivation of actively dividing, embryogenic cells such as immature embryos, isolated shoot apices and calli induced from scutella in the presence of acetosyringone, which is a potent inducer of the virulence genes. Several Agrobacterium strains such as LBA4404 (pT0K233), EHAlOI (pIGI2IHm), and EHAlOl (pSMABuba), have been used for rice transformation. A series of improved binary vectors have been constructed that are suitable for monocot transformation. These binary vectors have either bar or hph resistance genes as selection markers. HphR gene .has been improved by the introduction of an intron into its coding so as to abolish its expression in A. tumefaciens, rendering the bacterium susceptible to the hygromycin B44. In addition, binary bacterial artificial chromosome (BIBAC) vector has been developed that is capable of transferring at least 150 kb of foreign DNA into plant nuclear genome45.46
. The BIBABC vector has the minimal origin of replication for both E. coli F and A. rhizogenes Ri plasmids and it replicates as a single copy in E. coli as well as A. tumefaciens. The large BIBAC T-DNAs in conjunction with the helper plasmid that carries additional copies of virulence genes, have been used for high frequency transformation. The ability to introduce high molecular weight DNA into plant chromosomes should accelerate the studies on gene identification, genome organization and genetic engineering of complex polygenic traits.
Rashid et al. 47 reported the production of genetically stable transgenic plants of three Basmati rice varieties (Basmati 370, Basmati 385 and Basmati ' 6129) essentially using the Hiei et aL 8 transformation procedure. They used acetosyringone at 50' ~ concentration (instead of I 00 ~ for cocultivation. Zhang et al.48 reported A. tumefaciens-mediated transformation of . several commercially important rice
cultivars including Pusa Basmati I using the matureseed scutella derived calli and Agrobacterium strain LBA4404 (pTOK233). Based on the total number of calli co-cultivated, the transformation frequency of independent transgenic Pusa Basmati rice plants was 13.5%. Data from Southern hybridization analysis proved that foreign genes on T-DNA were stably integrated into rice genome at low copy/ site numbers. Jain49 observed a positive correlation between embryogenic potential and regeneration capacities of Basmati rice calli and Agrobacterium (strain LBA4404 with pTOK233)-mediated transformation frequencies using the modified Hiei et al. x procedure. Pusa Basmati I calli in comparison to those of Taraori Basmati had higher regeneration potential and consequently also showed higher transformation frequency. Regeneration potential also varied with the age of callus cultures. In Pusa Basmati I, six-weekold micro-calli obtained after one subculture of mature seeds, had the maximum regeneration potential and were found . to be best for Agrobacteriummediated transformation50. We used a novel Agrobacterium-mediated transformation system for the introduction of useful insect-resistant and drought tolerant genes into Basmati rice. The procedure involved the mobilization of pCAMBIA vectors carrying a cloned useful gene cassette into Agrobacterium strain LBA4404 carrying pSB I plasmid (lacks TDNA but carries virulence genes as in case of pTOK233) by tri-parental mating. The resultant strains have been used successfully for Pusa Basmati 1 transformation5o.
c. Potentially useful genes that can improve the Basmati rice
Since the first report of fertile transgenic plant production in 1989, remarkable progress has been made towards the transfer of useful genes in japonica as well as indica rice varieties. A variety of useful genes conferring resistance against abiotic and biotic stresses have been transferred in rice and these have been summarized in Table I. The Table do not contain the exhaustive list of the reports on rice transformation, but it serves to illustrate the type of genes
. that can be 'used for the genetic improvement of Bas- , . .mati rice. The breeding objectives in Basmati rice that can be well addressed using the transgenic technology and the traits for which novel genes are available, are described as under:
Resistance against insectlpests-The 8-endotoxin

12 INDIAN J EXP BIOL. JANUARY 2000
crystal protein genes from Bacillus thuringiensis -and plant defensive genes, protease inhibitors and lectins, are among the insect resistance genes51
-55 that have
been transferred in japonica and indica (including Basmatij rice varieties (Table I). The transgenic plants have been reported to be genetically stable and display the improved resistance against a variety of stem borers, leaf-hoppers or leaf folder insects.
B.thuringiensis 8-endotoxin crystal (cry) proteins are part of a large and still growing family of homologous proteins. About 130 cry genes have been identified to date. Most Bt crystal proteins are synthesized in a protoxin form and are proteolytically converted into smaller toxic polypeptides in the insect midgut. These toxins kill insects by binding to midgut membranes causing lesions. Cry genes have been recently classified on the basis of amino acid homology into four major classes, cry /, cry2, cry3 and cry4, which are, respectively, specific for insects belonging to Lepidotera (caterpillars), Lepidotera and Diptera (flies and mosquitos), Coleoptera (beetles) and Diptera. In rice, cry genes belonging to first two classes have been used for transgenic plant production. Fujimoto et al. 56 produced the transgenic japonica rice plants containing truncated and codon-modified 8-endotoxin gene using the protoplast-electroporation method. The R2 generation transgenic plants were more resistant to striped stem borer (SSB) and leafhopper than the non-transgenic plants. Wiinn et al. 57
reported increased insecticidal effect on several lepidoteran insect pests (yellow stem borer, striped stem borer and leaf folder) in the Ro, RI and R2 generation transgenic IR58 plants expressing the synthetic cry 1 A(b) gene. Datta et al. 58 produced transgenic plants of several indica and japonica rice varieties containing cry/Arb) gene driven by either constitutive (CaMV35S, Actin I) or tissue-specific (PEP carboxylase, pith specific) promoters using the biolistic and protoplast transformation systems. The transgene, crylA, driven by different promoters showed a wide range of expression (low to high) but conferred enhanced resistance to yellow stem borer (YSB). Out of 800 Southern-positive plants that were bio-assayed, 81 transgenic plants showed 100% mortality of insect larvae of the YSB . Cheng et al. 59 reported the production of over 2600 transgenic j.aponica rice plants containing the cry/Arb) or cryJA(c) synthetic gene driven by maize ubiquitin, CaMV35S and Brassica Bp 10 gene promoters. The transgenic Ro plants accumulated the cry/Arb) and cry/Arc) protein at higher
levels (up to 3% of soluble protein). The RI generation transgenic plants were highly toxic to SSB and YSB with mortalities of 97-100% after 5 days of infestation. Ghareyazie et al. 60 reported the transfer of the cry/Arb) gene driven by the tissue-specific maize C4 PEP carboxylase gene promoter in an Iranian aromatic rice variety Tarom Molaii belonging to group V. The transgene was expressed in the leaf blades but was not expressed to a detectable level in dehulled mature grain . Transgenic plants showed enhanced resistance against to first-instar larvae of SSB and YSB. Maqbool et al. 61 reported the production of transgenic Basmati 370 and M7 plants expressing the cry2A gene via biolistic method. The gene product was expressed up to 5% of total leaf protein and the transgenic plants had higher insecticidal activity against YSB and rice leaf folder.
Another classes of genes that have been used for improving insect/pest resistance are the protease inhibitor or lectin genes of plant origin. whose expression have shown anti-metabolic effects against certain insects. Expression of protease inhibitor genes have been useful for the control of insects which feed by chewing plant tissues, such as insects belonging to Lepidotera and Coleoptera, while that of lectin genes have been toxic to the sap-sucking insects belonging to the order Homoptera (brown plant hopper, green plant hopper). Duan et al. 62 and Xu et al. (,:1 produced transgenic japonica rice plants containing potato proteinase inhibitor II (Pinll) and cowpea trypsin inhibitor (CpTi) genes, respectively. The transgenes were driven by wound-inducible Pinll gene promoter with rice actin 1 intron and rice actin I promoter, respectively. Transgenic plants had high level accumulation of the PinIII CpTi proteins and their progenies showed increased resistance against pink stem borer and/or SSB. Hosoyama et al. 64 and Irie et al. 1
) produced 'transgenic rice plants containing oryzacystatin (Dc) and corncystatin (Cc) genes, respectively, which showed potent inhibitory activity against the cysteine proteinases that occur in the gut of insect pests. Vain et al. 65 reported increased nematode resistance in transgenic plants containing an engineered cysteine proteinase inhibitor (Oryzacystatin -/ t1D86) gene. These reports clearly indicate that proteinase inhibitor genes can be used to engineer the resistance against insect pests and nematodes in rice.
Among the various plant lectins, snowdrop lectin (Galanthus nivalis agglutinin: GNA) has been reported to be most toxic to the brown plant hopper

JAIN & JAIN: TRANSGENIC STRATEGIES FOR GENETIC IMPROVEMENT OF RICE 13
(BPH) and non-toxic to mammals66• GNA is a tet
rameric protein consisting of identical sub-units of 12 kD. Rao et al. 67 reported the production of transgenic rice plants containing snowdrop lectin gene driven by either phloem-specific rice sucrose synthase-l gene promoter (RSsl; Shi et a1. 68
) or the constitutive maize ubiquitin-l promoter (ubi). Phloem-specific promoters have been used to specifically express this gene in phloem tissue of transgenic rice plants conferring resistance against sap-sucking insects. Some of the transgenic plants had GNA at levels of up to 2.0% of total protein. Insect bioassays and feeding studies showed that GNA expressed in the transgenic plants decreased the insect survival rate, retarded insect development, and had a deterrent effect on BPH feeding.
Resistance against fungal diseases-Rice blast (Pyricularia grisea, oryzae) and Bakanae/foot rot (Fusarium moniliforme) are the two major fungal diseases that cause severe yield losses in Basmati rice4
.
In response to fungal attack, plants synthesize an assortment of new proteins commonly known as pathogenesis-related (PR) proteins including chitinases and ~-1 ,3-glucanases34
,69. Chitinase preparations alone and in combination with ~-1,3-glucanases, have been reported to inhibit fungal growth in vitro. Transgenic approach based on constitutive expression of a chitinase gene in transgenic tobacco have been shown to result in an increased ability to survive in soil infested with fungal pathogens and delayed development of disease symptoms69
. Lin et al. 34 developed transgenic indica rice plants that expressed chitinase gene constitutively. The plants with high levels of chitinase exhibited increased resistance to infection by the sheath blight pathogen, R. solani.
Stark-Lorenzen et al.70 reported active transcription of grapevine stilbene-synthase gene in transgenic rice plants after incubation with the fungus of the rice blast P. oryzae. Preliminary results indicated an enhanced resistance of the transgenic rice to P. oryzae. Stilbene synthase in some plant species synthesize a phytoalexin trans-resveratrol that seems to have a role in the early protection of plants against fungal pathogens. The effectiveness of stilbene synthase genes in enhancing resistance to another fungal pathogen, Phytophthora infestans, has been demonstrated in transgenic tomatoes71
• Tada et al.72 reported that transgenic rice plants expressing bar-gene showed decreased symptoms of rice blast disease caused by Magnporthe grisea following bialaphos treatment.
Resistance against bacterial leaf blight-Basmati rice is also susceptible to bacterial leaf blight (BLB) although incidence of bacterial leaf blight has been low probably due to the non-use of nitrogenous fertilizers for Basmati rice cultivation. A significant progress has been made towards the genetic engineering for BLB resistance in rice7.l. A dominant gene for resistance to BLB was transferred from a wild species, O. longistaminata, to the cultivated variety "IR24" (Khush et al. 74). This gene designated as Xa21, confers resistance to all known races of Xanthomonas oryzae pv. oryzae (Xoo) . Wang et aC5 reported that transgenic rice plants expressing Xa21 , gene conferred multi-isolate resistance to 29 diverse isolates from eight countries indicating that a single cloned gene is sufficient to confer multi-i solate resistance. Tu et aC6 reported higher resistance against two prevalent races (4 and 6) of Xoo in transgenic IR72 plants containing Xa21 gene. Resistance against race 4 was higher due to the pyramiding of transgene Xa21 and Xa4 (already present in IR72) . The Xa21 encodes a receptor kinase-like protein and it may have a role in cellular signaling for plant disease resistance.
Salinity and drought tolerance-A number of transgenic strategies are being used to increase tolerance to salinity and drought in plants77
-x1
. These include the late embryogenesis abundant proteins, overproduction of enzymes responsible for biosynthesis of osmolytes, and detoxification enzymes. This approach has met with preliminary success and there are now several reports on transgenic plant production containing stress-tolerant genes. Since expression of these genes individually confers marginal tolerance to abiotic stresses, it may be necessary to pyramid these different genes for higher level of stress tolerance. It may also be desirable to use stress-inducible promoters instead of constitutive promoters for the
. f I gry expreSSIon 0 stress to erant genes -.
Late-embryogenesis abundant protein (LEA) genes are normally expressed in the seed during maturation stage that involves desiccation and also in vegetative tissues during water deficit. LEA genes are induced by ABA and by osmotic stress resulting from drought, salinity stress, or cold temperatures. LEA proteins ha~e long been suggested as important in water retention or protection, ion sequestration, and as molecular chaperones, although their precise functioning is not clear. Xu et al. 83 produced the transgen ic japonica rice plants containing barley LEA3 (HVAJ)

14 INDIAN J EXP BIOL, JANUARY 2000
gene driven by a constitutive rice actin I promoter. The transgenic plants showed the constitutive accumulation of the HV A I protein both in leaves and roots. The second generation transgenic plants showed increased tolerance to both water and salt stresses. Artus et al. 84 reported that constitutive expression of a LEA-related gene Cori5a, in transgenic Arabidopsis plants enhanced the freezing tolerance of both chloroplasts and protoplasts. Cor J 5a expression has been suggested to affect the cryostability of the plasma membrane possibly through the interaction of corISa polypeptide with lipid bilayers.
During ()smotic stress, plant cells accumulate low molecular weight osmolytes to prevent water loss and maintain turgor. Transgenic approach has been used to engineer biosynthesis pathways of some of these osmolytes. Transgenic plants containing key genes encoding enzymes involved in the production of mannitol, proline, fructans, trehalose, glycine-betaine, D-ononitol and polyamines have been produced in model plant species like Arabidopsis, tobacco and . . . 777980 Th . Japomca nce . '. e transgemc plants have been reported to accumulate the corresponding osmolyte at higher levels and consequently showed a marginal to significant increase in the dehydration, salinity and/or cold tolerance. To give a few examples in rice; Zhu et at. 85 reported that overexpression of ABAIstressinducible promoter driven moth bean pyrroline-5-carboxylate synthetase (P5CS) gene resulted in up to 2.S-fold higher proline accumulation under stress conditions. The second generation transgenic plants compared to the non-transgenic plants had higher shoot and root biomass under salt- and water-stress conditions. Sakamoto et al. 39 reported that transgenic rice plants over-expressing the choline oxidase (codA; isolated from a soil bacterium Arthrobacter globiformis) gene accumulated glycine betaine at higher levels and were more tolerant to salt and low temperature stresses. The study also showed that such a gene if targeted to chloroplast was more effective in improving the stress tolerance. Capell et al. 86 produced the transgenic rice plants over-expressing the oat arginine decarboxylase (ADC) gene. The transgenic plants showed improved drought tolerance in terms of chlorophyll loss, however, constitutive expression of this gene severely affected the development patterns in vitro.
Much of the injury to plants caused by abiotic stresses, is associated with oxidative damage at the cellular level87
. The production of transgenic plants
with increased capacity for detoxification and scavenging reactive oxygen intermediates (ROIs) could be an another important strategy to engineer stress tolerance. Several groups have developed transgenic alfalfa and tobacco plants expressing a oxidative stress-
I t d h . d d' xx X<) I re a e gene suc as superoxi e Ismutase " ,g uta-thione-S-transferase90
, and iron-binding protein ferritin91
• The transgenic plants were reported to display greater tolerance to oxidative damage, salt stress, water deficit, and/or freezing.
Rec~ntly, Jaglo-Ottosen et ul.'!2 reported that overexpression of a regulator gene, CBF I, whose product is a transcriptional activator, induced COR gene expression and increased the freezing tolerance in transgenic Arabidopsis plants. Kasuga et ai.')1 transformed Arabidopsis with a gene encoding DREBIA, a homologue of CBFI, driveQ by a stress-inducible rd29A promoter. The overexpression"" of this gene in transgenic plants activated the expression of many stress-tolerance genes, rd 17 and rd29A (LEA like proteins), Kin J, Cor 6.6, Cor 15a, and P5CS; the transgenic plants showed greater tolerance to drought, salt and freezing stresses. The transformation of plants using such regulatory genes could be more rewarding to develop tolerance against complex traits like drought and salt tolerance.
Viral resistance-Research is also in progress to transfer genes for resistance against viruses (RTSV, rice tungro spherical virus; RTBV, rice tungro bacilliform virus; RDV, rice dwarf virus) in rice. Sivamani et al. 94 produced transgenic indica and japonica rice plants .containing RTSV coat protein genes. Most of transgenic plants, as well as their Rio R2 and/or Rl progeny that contained the target gene, showed the moderate levels of protection to RTSV infection and a significant delay of virus replication. Zheng et al.')S
developed the transgenic plants expressing the rice dwarf virus coat protein gene (S8), which is an important step towards studying the function of RDV genes and obtaining RDV -resistant rice plants.
Improved nutritional quality-Milled rice lacks provitamin A and is also deficient in essential amino acids (cysteine, methionine, lysine). Insufficient dietary provitamin A leads to severe clinical symptoms (eye disease xeropthalmia leading to blindness, increased susceptibility to other diseases such as diarrhoea, respiratory problems, measles, etc) . Burkhardt et al. 96 reported accumulation of phytoene. which is a key intermediate of provitamin A biosynthetic pathway, in endosperm tissue of the transgenic rice plants

JAIN & JAIN: TRANSGENIC STRATEGIES FOR GENETIC IMPROVEMENT OF RICE 15
expressing the daffodil phytoene synthase gene driven by the endospenn specific promoter. Further work is in progress to engineer the metabolic pathways for the synthesis of provitamin A and essential art}ino acids so as produce them in sufficient quantities in rice endosperm97.
Dwarfism-One of the major breeding objective in basmati rice is to develop short-statured plant type. Recently, Kisaka et al. 98 reported the production of transgenic japonica rice plants expressing the transgene, rgpJ (encodes for a GTP binding protein). These transgenic plants had distinct characteristics; namely, dwarfism, early flowering and high grain yield, and these characteristics were stable and heritable. The exact physiological function of GTPbinding proteins remains to be worked out. However, rgpJ gene has been shown to result in the reduction of apical dominance and high-level synthesis of cytokinins in transgenic tobacc099. It will be worthwhile to study the impact of such GTP-binding protein genes in Basmati rice.
Conclusions and future prospects Basmati rice breeding has been difficult due to
complicated nature of quality traits and poor combining ability of Basmati rice varieties . Major breeding objectives in Basmati rice includes the development of short-statured, photoperiod-insensitive and high yielding genotypes with increased resistance against insect pests (stem borer, leaf folder, brown plant hopper, gall midge), fungal diseases (blast, bakanae/foot rot) , bacterial diseases (bacterial leaf blight, sheath blight), and abiotic stresses (salinity and drought) . Transgenic approach holds great promise to achieve several of these objectives in Basmati rice without affecting its desirable genetic background and traits (aroma, grain quality, cooking quality). Rice is already a model monocot plant species for genetic transfonnation studies and is being routinely transformed by both direct gene transfer (biolistic, DNA uptake by protoplasts) and A. tumefaciens methods. Valuable genes for improving several important traits are now available and many of these have already been transferred in rice to improve the resistance against insect pests, fungal and viral diseases, salinity and drought stresses. Efforts are underway to engineer biosynthetic pathways for provitamin A and certain essential amino acids to improve the nutritional quality of rice.
Transgenic research to engineer complex agronomic traits (e.g. yield, drought tolerance, grain quality) and metabolic or regulatory pathways involving many genes or gene complexes, is likely to gain momentum in future 1oo
. This is pretty evident from the progress made towards the genome mapping and gene tagging research in rice and other plant species, development of tissue specific or stress-inducible promoters, and development of the ARrohacterium vectors for the transfer of fairly large DNA fragments. The problem of gene silencing can be tackled at least partially, by preferably using the ARrohacterium method of transfonnation4
\ selecting the transgenic plants with only one copy of the transgene.1 l, or by using the MAR (Matrix Attachment Regions) sequences which has been shown to reduce the position effect)OI.lo2. Issues such as safety and environmental concerns, public acceptance and the fees to be paid to the legal patent owner for the use of a particular technology or product (promoter, transgene, etc .), have to be taken into account before the application of transgenic technology.
Further research may be necessary to transform elite, commercially important indica rice varieties including Basmati rice varieties. A good beginning has already been made towards the development of tissue culture and transformation protocols in several commercially cultivated Basmati rice varieties including Pusa Basmati I and Basmati 370. Transgenic Basmati 370 plants containing Bt (cry2A) gene have been shown to be resistant to yellow stem borer and leaf folder61
• With the recent advances in rice molecular and transformation technologies, it should be feasible to design various transgenic strategies for the genetic improvement of Basmati rice and to increase its productivity.
Acknowledgement Thanks are due to Drs Ray Wu and M Maheswaran
for critically reading of the manuscript and di scussion. We would also like to thank the Rockefeller Foundation, New York, USA, for providing research grants for rice biotechnology research at CCS Haryana Agricultural University, Hisar (India) and Cornell, Ithaca, NY (USA).
References I Ayers N M & Park W D. Critical Rev Plallt Sci. 13 (1994)
219. 2 Tyagi A K. Mohanty A. Bajaj S. Chaudhury A & Mahesh
wari S C, CriticaL Rev BiotecllllOl. 19 ( 19(9) 41 .

16 INDIAN J EXP BIOL, JANUARY 2000
3 Ahuja S C, Panwar D V S, Ahuja U & Gupta K R, Basmati rice - The scented pearl (CCS Haryana Agricultural University, Hisar 125004, India) 1995.
4 Ahuja S C, Thakur D P & Srivastava M P, Rice diseases and disorders (Directorate of Publication, CCS Haryana Agricultural University, Hisar 125004, India) 1997.
5 Nene Y L, Asian Agri-History, 2 (3) (1998) 175. 6 Glaszmann J C, Theor Appl Genet, 74 (1987) 21 . 7 He P, Li S G, Qian Q, Ma Y Q, Li J Z, Wang W M, Chen Y
& Zhu L H, Theor Appl Genet, 98 (1999) 502. 8 Hiei Y, Ohta S, Komari T & Kumashiro T, Plant J, 6 (1994)
271. 9 Toenniessen G H, in Rice biotechnology, edited by G S
Khush, G H Toenniessen (CAB International, IRRI) 1997, 253.
10 Christou P, Rice biotechnology and genetic engineering (Technomic Pub. Co. Inc., Lancaster) 1994.
II Jain R K, Indian J Exp Bioi, 35 (1997) 232. 12 Potrykus I & Spangenberg G, Gene transfer to plants
(Springer-Verlag, Berlin) 1995. 13 Jain R K, Khehra G S, Lee S-H, Blackhall N W, Marchant R,
Davey M R, Power J B, Cocking E C & Gosal S S, Plant Cell Rep, 14 (1995) 515.
14 Jain R K, Davey M R, Cocking E C & Wu R, J Exp Bot, 48 (1997) 751.
15 Jain S, Jain R K & Wu R, Plant Cell Rep, 15 (1996) 712. 16 Jain R K, Jain S & Wu R, Plant Cell Rep, 15 (1996) 449-
454. 17 Jain R K, Jain S, Wang B & Wu R, Plant Cell Rep, 15
(1996) 963. 18 Jain R K, Jain S, Wang B Y & Wu R, International Rice
Research Notes, 22 (1997) 10. 19 Khanna H & Raina S K, J Plant Biochem Biotech, 6 (1997)
85. 20 Bishnoi U, Jain R K, Rohilla J S, Chowdhury V K, Gupta K
R & Chowdhury J B, Euphytica (in press) (1999). 21 Raina S K & Zapata F J, Plant Breed, 116 (1997) 305. 22 Chaudhury A, Maheshwari S C & Tyagi A K, Plant Cell
Rep, 14 (1995) 215. 23 Arencibia A, Gentinetta E, Cuzzoni E, Castiglione S, Kohli
A, Vain P, Leech M, Christou P & Sala A, Mole Breed, 4 (1998) 99.
24 Guo Y, Liang H & Berns M W, Physiol Plant, 93 (1995) 19. 25 Luo Z & Wu R, Plant Mol Bioi Rep, 7 (1989) 69. 26 Yoo J & Jung G, Physiol Plant, 94 (1995) 453. 27 Matsushita J, Otani M, Wakita Y, Tanaka 0 & Shimada T,
Breed Sci, 49-1 (1999) 21 . 28 Gamborg 0 L & Philips G C, PLant cell. tissue and organ
culture: Fundamental methods (Springer-Verlag, Berlin) 1995.
29 Chareonpornwattana S, Thara K V, Wang L, Datta S K, Panbangred W & Muthukrishnan S, Theor Appl Genet, 98 (1999) 371.
30 Depicker A & Van Montagu M, Curr Opin Cell Bioi. 9 (1997) 373.
31 Kumpatla S P, Chandrasekharan M B,lyer L M, Li G & Hall T C, Trends Plant Sci, 3 (1998) 97.
32 Matzke M A & Matzke A J M, Plant Physiol, 107 (1995) 679.
33 Morino K, Olsen O-A & Shimamoto K, Plant J, 17 (1999) 275.
34 Lin W, Anuratha C S, Datta K. Potrykus L Muthukrisnan S & Datta S K, Bio/teelmology, 13 (1995) 6X6.
35 lrie K, Hosoyama H, Takeuchi T, Iwabuchi K, Watanabe H, Abe M, Abe K & Arai S, Plant Mole Bioi. 30 (1996) 149.
36 Datta S K, in Molecular imjlrovemenl ()( cereal crops. Vol 5. edited by I K Vasil (Kluwer Academic Publishers. The Netherlands) 1998, (In press).
37 Christou P, Plalll Mole Bioi, 35 (1997) 197. 38 McBride K E, Svab Z. Schaaf D J. Hogan P S. Slalker D M
& Maliga P, Bio/techllology, 13 (1995) 362. 39 Sakamoto A, Alia & Murata N. Plant Mole BioI. 3X (1998)
1011. 40 Siemens J & Schieder 0 , Plant Tissue ClIlllIre Bioteell/wl
(IAPTC), 2 (1996) 66. 41 Minhas D, Bajaj S, Grover A & Rajam M V. Cllrr Sci. 12
(1996) 1005 42 Khanna H K, Raina S K, Srinivaslu & Kumar K. Rice Bio
technol Quart, 28 (1996) 15. 43 Hiei Y, Komari T & Kubo T, Plallt Moll' BioI. 35 (1997)
205. 44 Wang M B, Li Z Y, Upadhyaya N M. Brettell R I S & Wa
terhouse P M, J Genet Plallt Breed. 51 (1997) 325. 45 Hamilton C M, Frary A. Lewis C & Tanksley S D. Proc Natl
Acad Sci (USA), 93 ( 1996) 9975. 46 Hamilton C M, (Jelle. 200 (1997) 107. 47 Rashid H, Yokoi S, Toriyama K & Hinata K. Planl Cell Rep.
15 (1996) 727. 48 Zhang J, Xu R J, Eliott M C & Chen D F, Mole Biolechnol.
8 (1997) 223. 49 Jain R K, Rice BioteellllOl Quell'l. 36 (199X) 12. 50 Jain R K, Rohilla J S, Wu R, upublished dala. 51 Estruch J J, Carozzi N B, Desai N. Duck N B. Warren G W
& Koziel M G, Nature Bioleell/IOI. 15 (1997)137. 52 Katiyar S K, Chandel G & Pratibha R. Indiall .I E~JI Bioi. 37
(1999) 211. 53 De Maagd R A, Bosch D & Stiekema W. Trellds Plan I Sci. 4
(1999) 9. 54 Ryan C A, Annu Rev Phytopal/lOl, 28 (1990) 25. 55 Schuler T H, Poppy G M. Kerry B R & Denholm L T1B
TECH, 16 (1998) 168. 56 Fujimoto H, Itoh K, Yamamoto M. Kyozuka J & Shimamoto
K, Bio/technology, II (1993) 1151. 57 Wiinn J, Kltiti A, Burkhardt P K. Ghosh Biswas G C. Launis
K, Iglesias V A & Potrykus L Bio/lechnolog,l'. 14 (1996) 171.
58 Datta K, Vasquez A, Tu J. Torrizo L. Alam M F. Oliva N. Abrigo E, Khush G S & Datta S K. TheaI' Apjl/ Genel. 97 (1998) 20.
59 Cheng X, Sardana R. Kaplan H & Altosaar I. Proc Nail Acad Sci (USA), 95 (1998) 2767.
60 Ghareyazie B, Ainia F, Menguito C A. Rubia L G. De Palma J M, Liwanag E A. Cohen M B. Khush G S & Bennett J, Mole Breed, 3 (1997) 40 I .
61 Maqbool S B, Husnain T, Riazuddin. Masson L & Christou P, Mole Breed, 4 (1998) 501.
62 Duan X, Li X, Xue Q, Abo-EI-Saad M. Xu D & Wu R. Nature Biotechnol, 14 (1996) 494.
63 Xu D, Xue Q, McElroy D. Mawal Y. Hilder V A & Wu R. Mole Breed, 2 (1996) 167.
64 Hosoyama H, lrie K, Abe K & Ami S, Plalll Cdl Rep, 15 (1995) 174.

JAIN & JAIN: TRANSGENIC STRATEGIES FOR GENETIC IMPROVEMENT OF RICE 17
65 Vain P, Worland B, Clarke M C, Richard G, Beavis M, Liu H, Kohli A, Leech M, Snape J, Christou P & Atkinson H, Theor Appl Genet, 96 (1998) 266.
66 Boulter D, Edwards G A, Gatehouse A M R, Gatehouse J A & Hilder V A, Crop Protection , 9 (1990) 351 .
67 Rao K V, Rathore K S, Hodges T K, Fu X, Stoger E, Sudhakar D, Williams S, Christou P, Bharathi M, Bown D P, Powell K S, Spence J, Gatehouse A M R & Gatehouse J A, Plant J, 15 (1998) 469.
68 Shi Y, Wang M-B, Powell K S, Van Damme E, Hilder V A, Gatehouse A M R, Boulter D & Gatehouse J A, J Exp Bot, 45 (1994) 623.
69 Broglie K, Chet I, Holliday M, Cressman R, Biddle P, Knowlton S, Mauvais J & Broglie R M, Science, 254 (1991) 1194.
70 Stark-Lorenzen P, Nelke B, HanGler G, MOhlbach H P & Thomzik J E, Plant Cell Rep, 16 (1997) 668.
71 Thomzik J E, Stenzel K, Stoecker R, Schreier PH, Hain R & Stahl D J, Physiol Mole Plant Pathol, 51 (1997) 265.
72 Tada T, Kanazaki H, Norita E, Uchimiya H & Nakamura I, Mol Plant-Microbe Interact , 9 (1996) 758.
73 Fauquet C M, Huet H, Ong C A, Sivamani E , Chen L, Viegas P, Marmey V P, Wang P, Daud M, de Kochko A & Beachy R N, in Abstracts, General Meeting of the International Programme on Rice Biotechnology (Malacca, Malyasia) 1997,59.
74 Khush G S, Bacalangco E & Ogawa T , Rice Genetics Newslett, 7 (1990) 121.
75 Wang G-L, Song W-L, Ruan D-L, Sideris S & Ronald P C, Mol Plant-Microbe Interact, 9 (1996) 850.
76 Tu J, Ona I, Zhang Q, Mew T W, Khush G S & Datta S K, Theor Appl Genet, 97 (1998) 31.
77 Bartels D & Nelson D, Plallt Cell Environ, 17 (1994) 659. 78 Bohnert H J & Jensen R G, Aust J Plant Physiol 23 (1996a)
661. 79 Bohnert H J & Jensen R G, Trends Biotech, 14 (1996b) 89. 80 Holmberg N & BOlow L, Trends Plant Sci, 3 (1998) 61. 81 Zhu J-K, Hasegawa P M & Bressan R A, Crit Rev Plant Sci,
16 (1997) 253. 82 Su J, Shen Q, Ho T-H D & Wu R, Plant Physiol, 117 (1998)
913 . 83 Xu D, Duan X, Wang B, Hong B, Ho T-H D & Wu R, Plant
Physiol, 110 (1996) 249. 84 Artus N N, Uemura M, Steponkus P L, Gilmour S J, Lin C &
Thomashow M F, Proc Nat Acad Sci (USA) 93 (1996) 13404.
85 Zhu B, Su J , Chang M C, Verma D P S, Fan Y L & Wu R, Plant Sci, 139 (1998) 41.
86 Capell T, Escobar C , Liu H, Burtin D, Lepri 0 & Christou P, Theor App/ Genet, 97 (1998) 246.
87 Allen R D, PLant PhysioL, 107 (1995) 1049.
88 Gupta A S, Heinen J L, Holaday A S, Burke J J & Allen R D. Proc Natl Acad Sci (USA), 90 (1993) 1629.
89 Mckersie B D, Bowley S R & Jones K S, Plant Ph-"siol. 119 (1999) 839.
90 Roxas V P, Smith R K Jr, Allen E R & Allen R D. Naltlre Biotechnol, 15 (1997) 988.
91 Deak M, Horvath G V, Davletova S. Torok K. Sass L. Vass I, Barna B, Kiraly Z & Dudits D. Nalllre Biotedlllol. 17 (1999)192.
92 Jaglo-Ottosen K R, Gilmour S J, Zarka D G. Schabenberger 0& Thomashow M F, Science. 280 (1998) 104.
93 Kasuga M, Liu Q, Miura S. Yamaguchi-Shinozaki K & Shinozaki K, Nature BiotecllllOl, 17 (1999) 287 .
94 Sivamani E & Huet H with : Shen p , Ong C A. de Kocko A, Fauquet C & Beachy R N, Mole Breed. 5 ( 1999) 177.
95 Zheng H H, Li Y, Yu Z H, Li W. Chen M Y. Ming X T. Casper R & Chen Z L, Theor AJlJlI Genet. 94 (1997) 522.
96 Burkhardt P K, Beyer P, WOnn J, Kltiti A. Armstrong G A. Schledz M, von Lintig J & Potrykus I. Plant J. II (1997) 1071.
97 Potrykus I, Burkhardt P K, Datta S K. FOtterer GhoshBiswas G C, Kltiti A, Spangenberg G & WOnn J. EIIJlhyticlI . 85 (1995) 441.
98 Kisaka H, Sano H & Kameya T, Them' AIIJlI Gellet, 97 (I 998} 810.
99 Sano H, Seo S, Orudgev E, Yosselian S. Ishizuka K & Ohashi V, Proc NatL Acad Sci (USA). 91 (1994) 10556.
100 Chen L, Marmey P, Taylor N J. Brizard J-P . Espinoza C , D'Cruz P, Huet H, Zhang S, de Kochko A. Beachy R N & Fauquet C M, Nature Biotechnol, 16 (1998) 1060.
10 I Spiker S & Thompson W F. PIli/It Physiol. I 10 (19%) 15.
102 Holmes-Davis R & Comai L. Trends Plant Sci. 3 (1998) 91.
103 Alam M F, Firoz M, Datta K. Abrigo E. Vasquez A, Sen ad-hira D & Datta S K, Plant Sci, 135 (1998) 25.
104 Wu C, Fan y , Zhang C, Oliva N & Dalla S K. Plan! Cell Rep, 17 (1997) 129.
105 Nayak P, Basu D, Das S. Basu A, Ghosh 1'.1 & Sen S K. Pmc Na!L Acad Sci (USA), 94 (1997) 2111 .
106 Zhang S, Song W-Y, Chen L. Ruan D, Taylor N. Ronald P. Beachy R & Fauquet C , Mole Breed. 4 (1998) 551 .
107 Yokoi S, Higashi S-I, Kishitani S. Murata N & Toriyama K, MoLe Breed, 4 (I 998} 269.
108 Huntley C C & Hall T C, Mol Plant-Microhe Interact, 9 (1996) 164.
109 Pinto Y M, Kok R P & Baulcombe D C. Natllre Biotedlllol. 17 (1999) 702.
110 Ku SBM, Agarie S, Nomura M. Fukayama H, Tsuchida H, Kazuko 0 , Hirose S. Toki S, Miyao M and Matsuoka M. Nature BiotechnoL, 17 (1999) 76.





























