Embed Size (px)
Citation preview

www.elsevier.com/locate/cnsns
Communications in Nonlinear Science
and Numerical Simulation 12 (2007) 357–365
Transitions from bursting to spiking due to depolarizingcurrent in the Chay neuronal model
Zhuoqin Yang, Qishao Lu *
School of Science, Beihang University, Beijing 100083, China
Received 12 February 2004; received in revised form 20 December 2004; accepted 10 January 2005
Available online 25 May 2005
Abstract
Distinct transitions of firing activities from bursting to spiking induced by the depolarizing current I are
explored near the Hopf bifurcations in the Chay neuronal system. The period-1 ‘‘circle/homoclinic’’ burst-
ing at one rest state makes a transition slowly to repetitive spiking with the parameter I increasing. How-ever, the ‘‘Hopf/homoclinic’’ bursting via a ‘‘fold/homoclinic’’ hysteresis loop at another rest state may
transit to continuous spiking abruptly by increasing I.
� 2005 Elsevier B.V. All rights reserved.
PACS: 05.45.�a; 82.40.Bj
Keywords: Ursting; Spiking; Bifurcation; Fast/slow dynamic analysis
1. Introduction
Neuronal firing activities are of fundamental importance to reveal the mechanism of neuronalcoding. Many different dynamical models have been studied to explore their dynamical behavior,among which bursting is the most important one and has been investigated in many experiments
1007-5704/$ - see front matter � 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.cnsns.2005.01.014
* Corresponding author.
E-mail address: [email protected] (Q. Lu).

358 Z. Yang, Q. Lu / Communications in Nonlinear Science and Numerical Simulation 12 (2007) 357–365
[1–4] and theoretical studies [5–12]. Bursting means that the firing activity of neuron alternatesbetween a rest state and repetitive spiking (that is, a burst). Many mathematical models of neu-ronal bursters can be written in the following singular perturbation form [11]:
_x ¼ f ðx; uÞ; ð1Þ
_u ¼ lgðx; uÞ; ð2Þ
where l� 1 represents the ratio between the fast and slow time scales. x 2 Rm is the fast variableresponsible for repetitive spiking, and u 2 R is the slow variable to modulate spiking. When con-sidering the fast subsystem _x ¼ f ðx; uÞ with the one-dimensional slow variable u as a time-depen-dent bifurcation parameter, there are two important kinds of bifurcations of the fast subsystemassociated with bursting: the bifurcation of a rest state leading to repetitive spiking and that ofrepetitive spiking leading to a rest state. These bifurcations of the fast subsystem can reveal explic-itly how the spike-generating mechanism interacts with the slow dynamics to produce burstingbehavior and provide a complete classification scheme for bursting, see Table 4 in [11].
In contrast with bursting, periodic spiking only corresponds to the existence of a large stablelimit cycle and always lies near this limit cycle.
Many excitable cells at rest can exhibit interesting discharge activities in response to variousexternal stimuli. As a consequence, the firing patterns induced by depolarizing current, as wellas their dynamical behavior, are studied through the Chay system by means of numerical simu-lation and fast/slow dynamic analysis here. This paper is organized as follows. Section 2 presentsthe Chay model with depolarizing current. Transition phenomena from distinct types of burstingto spiking induced by the depolarizing current are discussed in Section 3. Finally, a conclusion isgiven in Section 4.
2. Chay model with depolarizing current
The following three equations form the Chay model with depolarizing current [6]:
dVdt¼ g�I m3
1h1ðV I � V Þ þ g�K;V ðV K � V Þn4 þ g�K;CC
1þ CðV K � V Þ þ g�LðV L � V Þ þ I; ð3Þ
dndt¼ n1 � n
sn; ð4Þ
dCdt¼ q½m3
1h1ðV C � V Þ � kCC�; ð5Þ
where three dynamical variables are V (the membrane potential), n (the probability of opening thevoltage-sensitive K+ channel) and C (the intracellular concentration of Ca2+ ions). VK, VC are thereversal potentials for K+ and Ca2+ channels, respectively, and VL, VI are that for other ionicchannels. The explicit expressions for m1, h1 and n1 can be written generally as y1 = ay/(ay + by), where y represents m, n or h, with

Z. Yang, Q. Lu / Communications in Nonlinear Science and Numerical Simulation 12 (2007) 357–365 359
am ¼ 0.1ð25þ V Þ=ð1� e�0.1�V�2.5Þ;bm ¼ 4e�ðVþ50Þ=18;
ah ¼ 0.07e�0.05V�2.5;
bh ¼ 1=ð1þ e�0.1V�2Þ;an ¼ 0.01ð20þ V Þ=ð1� e�0.1V�2Þ;bn ¼ 0.125e�ðVþ30Þ=80;
sn ¼ 1=knðan þ bnÞ.
In what follows, I denotes the depolarizing current and is considered as an adjustable controlparameter for the dynamic study of the firing activities of the Chay system (3)–(5) under sometypical values of VK. The values of other parameters used in numerical simulations here are iden-tical to that in [6].
The fast/slow dynamics analysis in the Chay system can be used on the basis of the fact that q isusually a small quantity, and then C varies much slower than the other variables. Therefore, thewhole Chay system can be divided into two subsystems: the fast subsystem (abbreviated as FS)given by Eqs. (3) and (4) and the slow subsystem (abbreviated as SS) given by Eq. (5). The dynam-ics of the fast subsystem is dependent on the slow variable C as a slowly-varying bifurcationparameter. Moreover, the nullcline of the slow variable C is given by
C ¼ ½m31ðV Þh1ðV ÞðV C � V Þ�=kC. ð6Þ
3. Transitions from distinct types of bursting to spiking due to depolarizing current
VK � �82.386 mV and �61.027 mV are two Hopf bifurcation points in the Chay model with-out depolarizing current. In order to investigate the effects of the depolarizing current I on firingactivities at the rest states of the Chay model, two values of VK near these Hopf bifurcation values,i.e. �83 mV and �61 mV, are chosen in the following study.
3.1. The slow transition from period-1 ‘‘circle/homoclinic’’ bursting to repetitive spiking
When VK = �83 mV, a period-1 firing pattern arises and its interspike intervals (ISI) get smallerwith I increasing, which can be seen from the bifurcation diagram of ISI vs. I as a bifurcationparameter in Fig. 1(a). The time histories of period-1 firings for I = 70 mA and I = 100 mA arepresented in Fig. 1(b) and (c), respectively.
The dynamical behavior of the period-1 firing patterns induced by the depolarizing current I forVK = �83 mV can be explored by the fast/slow dynamics analysis. No loss of generality, only theperiod-1 firings for I = 70 mA and 100 mA will be discussed here. For these two values of I, theequilibrium points of the fast subsystem (FS) with respect to the parameter C form the left andthe right Z-shaped curves in (C,V)-phase plane, respectively (see Fig. 2(a)). On the upper branchof the curves, the stable foci of FS become unstable via Hopf bifurcations (H1) and (H2), and thestable limit cycles, corresponding to period-1 firings, occur around the unstable focus are plotted
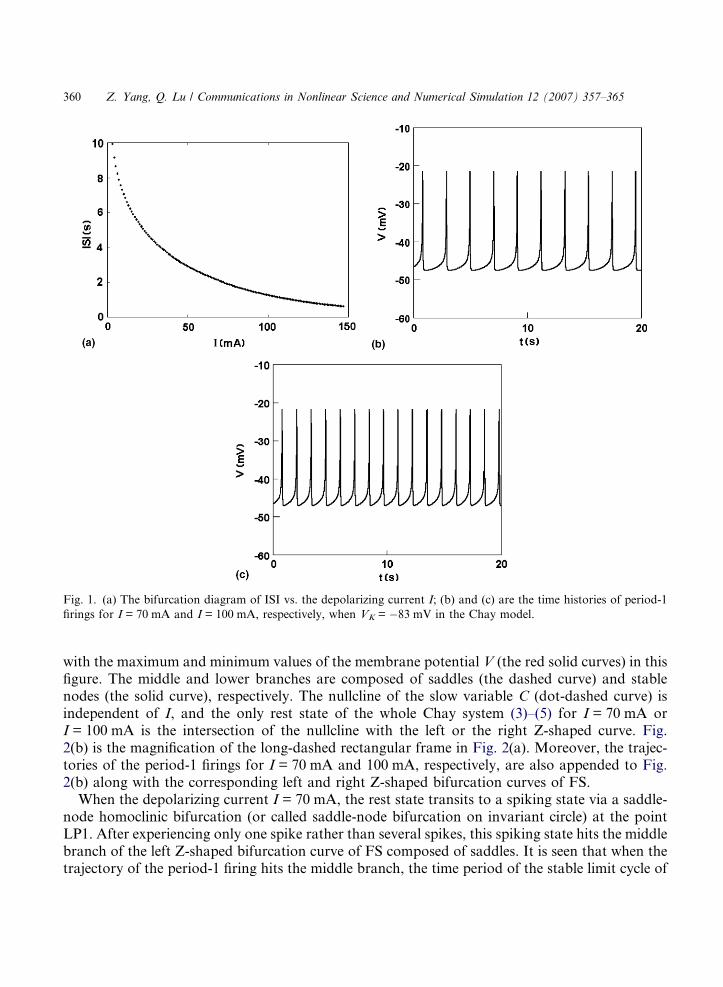
Fig. 1. (a) The bifurcation diagram of ISI vs. the depolarizing current I; (b) and (c) are the time histories of period-1
firings for I = 70 mA and I = 100 mA, respectively, when VK = �83 mV in the Chay model.
360 Z. Yang, Q. Lu / Communications in Nonlinear Science and Numerical Simulation 12 (2007) 357–365
with the maximum and minimum values of the membrane potential V (the red solid curves) in thisfigure. The middle and lower branches are composed of saddles (the dashed curve) and stablenodes (the solid curve), respectively. The nullcline of the slow variable C (dot-dashed curve) isindependent of I, and the only rest state of the whole Chay system (3)–(5) for I = 70 mA orI = 100 mA is the intersection of the nullcline with the left or the right Z-shaped curve. Fig.2(b) is the magnification of the long-dashed rectangular frame in Fig. 2(a). Moreover, the trajec-tories of the period-1 firings for I = 70 mA and 100 mA, respectively, are also appended to Fig.2(b) along with the corresponding left and right Z-shaped bifurcation curves of FS.
When the depolarizing current I = 70 mA, the rest state transits to a spiking state via a saddle-node homoclinic bifurcation (or called saddle-node bifurcation on invariant circle) at the pointLP1. After experiencing only one spike rather than several spikes, this spiking state hits the middlebranch of the left Z-shaped bifurcation curve of FS composed of saddles. It is seen that when thetrajectory of the period-1 firing hits the middle branch, the time period of the stable limit cycle of
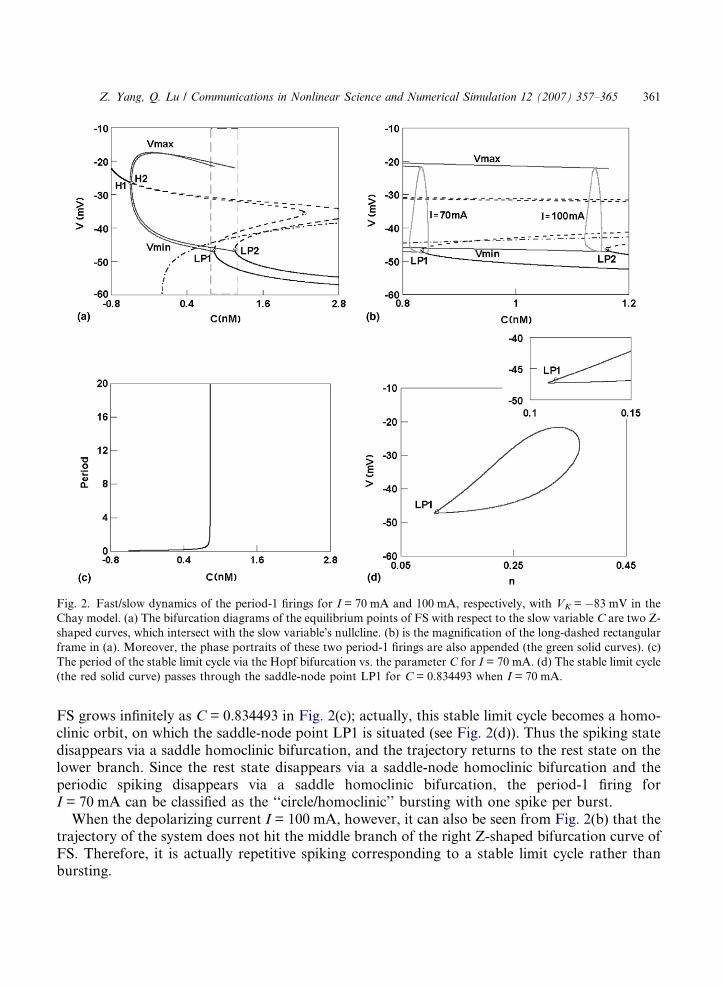
Fig. 2. Fast/slow dynamics of the period-1 firings for I = 70 mA and 100 mA, respectively, with VK = �83 mV in the
Chay model. (a) The bifurcation diagrams of the equilibrium points of FS with respect to the slow variable C are two Z-
shaped curves, which intersect with the slow variable�s nullcline. (b) is the magnification of the long-dashed rectangular
frame in (a). Moreover, the phase portraits of these two period-1 firings are also appended (the green solid curves). (c)
The period of the stable limit cycle via the Hopf bifurcation vs. the parameter C for I = 70 mA. (d) The stable limit cycle
(the red solid curve) passes through the saddle-node point LP1 for C = 0.834493 when I = 70 mA.
Z. Yang, Q. Lu / Communications in Nonlinear Science and Numerical Simulation 12 (2007) 357–365 361
FS grows infinitely as C = 0.834493 in Fig. 2(c); actually, this stable limit cycle becomes a homo-clinic orbit, on which the saddle-node point LP1 is situated (see Fig. 2(d)). Thus the spiking statedisappears via a saddle homoclinic bifurcation, and the trajectory returns to the rest state on thelower branch. Since the rest state disappears via a saddle-node homoclinic bifurcation and theperiodic spiking disappears via a saddle homoclinic bifurcation, the period-1 firing forI = 70 mA can be classified as the ‘‘circle/homoclinic’’ bursting with one spike per burst.
When the depolarizing current I = 100 mA, however, it can also be seen from Fig. 2(b) that thetrajectory of the system does not hit the middle branch of the right Z-shaped bifurcation curve ofFS. Therefore, it is actually repetitive spiking corresponding to a stable limit cycle rather thanbursting.
362 Z. Yang, Q. Lu / Communications in Nonlinear Science and Numerical Simulation 12 (2007) 357–365
More detailed fast/slow bifurcation analyses confirm the following fact. It seems that whenVK = �83 mV all the firing patterns are period-1 by appearance for different values of I, however,there is a slow transition from period-1 ‘‘circle/homoclinic’’ bursting to repetitive spiking atI � 89.5 mA in fact.
3.2. The abrupt transition from ‘‘Hopf/homoclinic’’ bursting to continuous spiking
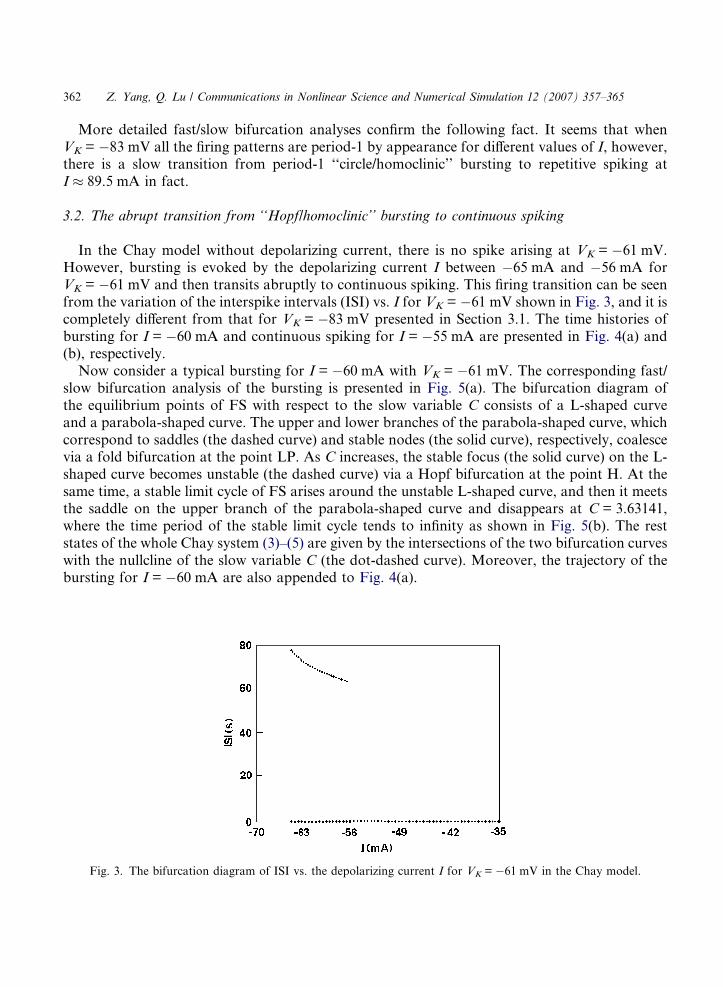
In the Chay model without depolarizing current, there is no spike arising at VK = �61 mV.However, bursting is evoked by the depolarizing current I between �65 mA and �56 mA forVK = �61 mV and then transits abruptly to continuous spiking. This firing transition can be seenfrom the variation of the interspike intervals (ISI) vs. I for VK = �61 mV shown in Fig. 3, and it iscompletely different from that for VK = �83 mV presented in Section 3.1. The time histories ofbursting for I = �60 mA and continuous spiking for I = �55 mA are presented in Fig. 4(a) and(b), respectively.
Now consider a typical bursting for I = �60 mA with VK = �61 mV. The corresponding fast/slow bifurcation analysis of the bursting is presented in Fig. 5(a). The bifurcation diagram ofthe equilibrium points of FS with respect to the slow variable C consists of a L-shaped curveand a parabola-shaped curve. The upper and lower branches of the parabola-shaped curve, whichcorrespond to saddles (the dashed curve) and stable nodes (the solid curve), respectively, coalescevia a fold bifurcation at the point LP. As C increases, the stable focus (the solid curve) on the L-shaped curve becomes unstable (the dashed curve) via a Hopf bifurcation at the point H. At thesame time, a stable limit cycle of FS arises around the unstable L-shaped curve, and then it meetsthe saddle on the upper branch of the parabola-shaped curve and disappears at C = 3.63141,where the time period of the stable limit cycle tends to infinity as shown in Fig. 5(b). The reststates of the whole Chay system (3)–(5) are given by the intersections of the two bifurcation curveswith the nullcline of the slow variable C (the dot-dashed curve). Moreover, the trajectory of thebursting for I = �60 mA are also appended to Fig. 4(a).
Fig. 3. The bifurcation diagram of ISI vs. the depolarizing current I for VK = �61 mV in the Chay model.
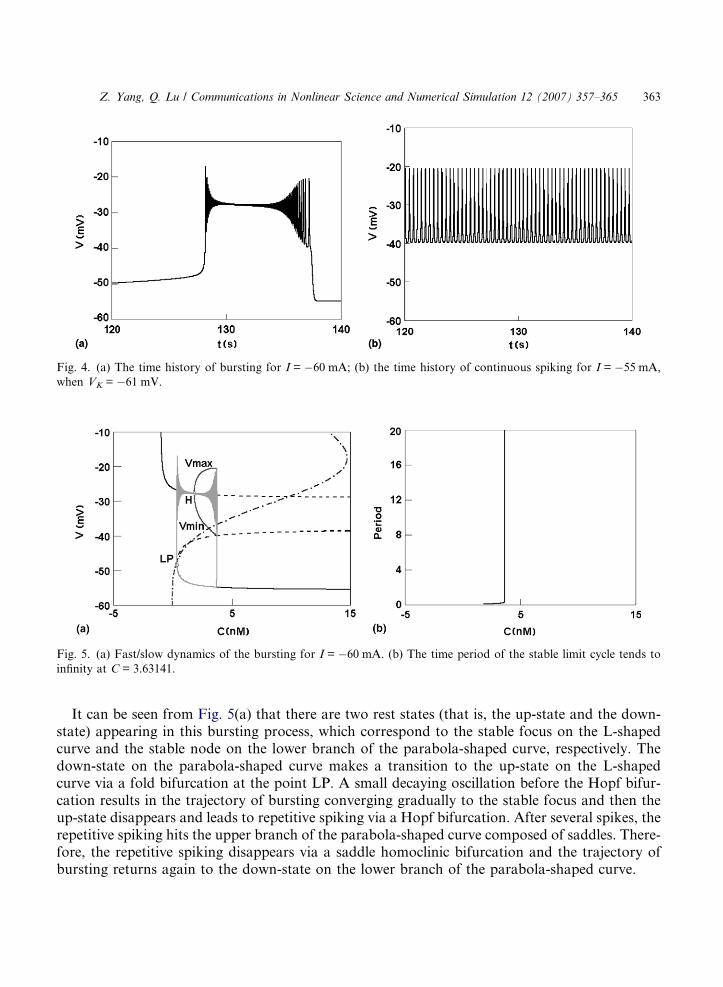
Fig. 4. (a) The time history of bursting for I = �60 mA; (b) the time history of continuous spiking for I = �55 mA,
when VK = �61 mV.
Fig. 5. (a) Fast/slow dynamics of the bursting for I = �60 mA. (b) The time period of the stable limit cycle tends to
infinity at C = 3.63141.
Z. Yang, Q. Lu / Communications in Nonlinear Science and Numerical Simulation 12 (2007) 357–365 363
It can be seen from Fig. 5(a) that there are two rest states (that is, the up-state and the down-state) appearing in this bursting process, which correspond to the stable focus on the L-shapedcurve and the stable node on the lower branch of the parabola-shaped curve, respectively. Thedown-state on the parabola-shaped curve makes a transition to the up-state on the L-shapedcurve via a fold bifurcation at the point LP. A small decaying oscillation before the Hopf bifur-cation results in the trajectory of bursting converging gradually to the stable focus and then theup-state disappears and leads to repetitive spiking via a Hopf bifurcation. After several spikes, therepetitive spiking hits the upper branch of the parabola-shaped curve composed of saddles. There-fore, the repetitive spiking disappears via a saddle homoclinic bifurcation and the trajectory ofbursting returns again to the down-state on the lower branch of the parabola-shaped curve.

364 Z. Yang, Q. Lu / Communications in Nonlinear Science and Numerical Simulation 12 (2007) 357–365
The down-state transits to the up-state via a fold bifurcation, and the up-state transits to thestate of repetitive spiking via Hopf bifurcation, and repetitive spiking terminates and returns tothe down-state via a saddle homoclinic bifurcation. Accordingly, the bursting for I = �60 mAcan be classified as the ‘‘Hopf/homoclinic’’ bursting via ‘‘fold/homoclinic’’ hysteresis loop.
4. Conclusion
Neurons at rest states can exhibit diverse firing activities in response to the external stimuli. Thefiring transitions from bursting to spiking induced by the depolarizing current I under differentvalues of VK were explored by numerical simulation and fast/slow dynamic analysis in the Chayneuronal system. When VK = �83 mV, the period-1 firing activity underwent a slow transitionfrom period-1 ‘‘circle/homoclinic’’ bursting to repetitive spiking with the increasing of the depo-larizing current I. However, when VK = �61 mV the firing pattern experienced an abrupt transi-tion from ‘‘Hopf/homoclinic’’ bursting via ‘‘fold/homoclinic’’ hysteresis loop to continuousspiking, evoked by the depolarizing current I.
In conclusion, the induced firing patterns of the Chay model at the rest states may be differentunder the actions of the depolarizing current. Similar to the Chay model, the depolarizing currentare essential for the occurrence of various firing patterns in many neuronal models, so the presentresults are also instructive to the complexity of neuronal firing activities in response to abundantexternal stimuli.
Acknowledgement
This work was supported by the National Natural Science Foundation of China (No.10432010). The bifurcation diagrams in this paper were obtained by means of the packageCONTENT.
References
[1] Deschenes M, Roy JP, Steriade M. Thalamic bursting mechanism: an invariant slow current revealed by membrane
hyperpolarization. Brain Res 1982;239:289–93.
[2] Harris-Warrick RM, Flamm RE. Multiple mechanisms of bursting in a conditional bursting neuron. J Neurosci
1987;7:2113–28.
[3] Ashcroft F, Rorsman P. Electrophysiology of the pancreatic b-cell. Prog Biophys Molec Biol 1989;54:87–143.
[4] Johnson SW, Seutin V, North RA. Burst firing in dopamine neurons induced by N-methyl-D-aspartate: role of
electrogenic sodium pump. Science 1992;258:665–7.
[5] Bertram R, Butte MJ, Kiemel T, Sherman A. Topological and phenomenological classification of bursting
oscillations. Bull Math Biol 1995;57:413–39.
[6] Chay TR. Chaos in a three-variable model of an excitable cell. Physica D 1985;16:233–42.
[7] Kepecs A, Wang XJ. Analysis of complex bursting in cortical pyramidal neuron models. Neurocomputing
2000;32–33:81–187.
[8] Rinzel J, Lee YS. Dissection of a model for neuronal parabolic bursting. J Math Biol 1987;25:653–75.
[9] Rush ME, Rinzel J. Analysis of bursting in thalamic neuron model. Biol Cybern 1994;71:281–91.

Z. Yang, Q. Lu / Communications in Nonlinear Science and Numerical Simulation 12 (2007) 357–365 365
[10] Soto-Trevino C, Kopell N, Watson D. Parabolic bursting revisited. J Math Biol 1996;35:114–28.
[11] Izhikevich EM. Neural excitability, spiking and bursting. Int J Bif Chaos 2000;10:1171–266.
[12] Chay TR, Fan YS, Lee YS. Bursting, spiking, chaos, fractals, and university in biological rhythms. Int J Bif Chaos
1995;5:595–635.





























