Embed Size (px)
Citation preview

TRILOBITA Agnostids http://biosys-serv.biologie.uni ulm.de/sektion/dieter/noncrustacean/noncrustacean.html
Class TRILOBITA (the trilobites) Trilobites are the first successful arthropod experiment and are extensively used to zone the Paleozoic. As a matter of fact, the first appearance of the trilobite genus Ollenellus marks the classical beginning of the Paleozoic. They are characterized by a body that is divided in three parts, hence their name. Some were blind, others had well developed compound eyes. Many were mud or filter feeders, while still others may have been predators. Their ultimate demise may well have been due to being outcompeted by fishes.
Trilobites are one of the most successful Paleozoic groups. Because of their diversity and continued evolution they are one of the better groups for time zonation.
trilbt.jpg
Perenopsids are a group of small blind trilobites. This specimen is actually less than 1.5 cm long (about 1/2 in.)
Others were much larger. This trilobite is some 20cm (~8 in.) in length.
1

This is a detail of the trilobite above. Note that the trilobites had already evolved compound eyes, still used today by their relatives, the insects.
Others developed bizarre and ornate horns.
2

This is Phacops rana a common North American trilobite
Trilobites are hard-shelled, segmented creatures that existed over 300 million years ago in the Earth's ancient seas. They went extinct before dinosaurs even existed, and are one of the key signature creatures of the Paleozoic Era, the first era to exhibit a proliferation of the complex life-forms that established the foundation of life as it is today. Although dinosaurs are the most well-known fossil life forms, trilobites are also a favorite among those familiar with Paleontology (the study of the development of life on Earth).
ANCIENT ARTHROPODS Trilobites were among the first of the arthropods, a phylum of hard-shelled creatures with multiple body segments and jointed legs (although the legs, antennae and other finer structures of trilobites only rarely are preserved). They constitute an extinct class of arthropods, the Trilobita, made up of eight orders, over 150 families, about 5000 genera, and over 15,000 described species. New species of trilobites are unearthed and described every year. This makes trilobites the single most diverse group of extinct organisms, and within the generalized body plan of trilobites there was a great deal of diversity of size and form. The smallest known trilobite species is just under a millimeter long, while the largest include species from 30 to 70 cm in length (roughly a foot to two feet long!). With such a diversity of species and sizes, speculations on the ecological role of trilobites includes planktonic, swimming, and crawling forms, and we can presume they filled a varied set of trophic (feeding) niches, although perhaps mostly as detritivores, predators, or scavengers. Most trilobites are about an inch long, and part of their appeal is that you can hold and examine an entire fossil animal and turn it about in your hand. Try that with your average dinosaur!
3

THE TRILOBITE BODY PLAN
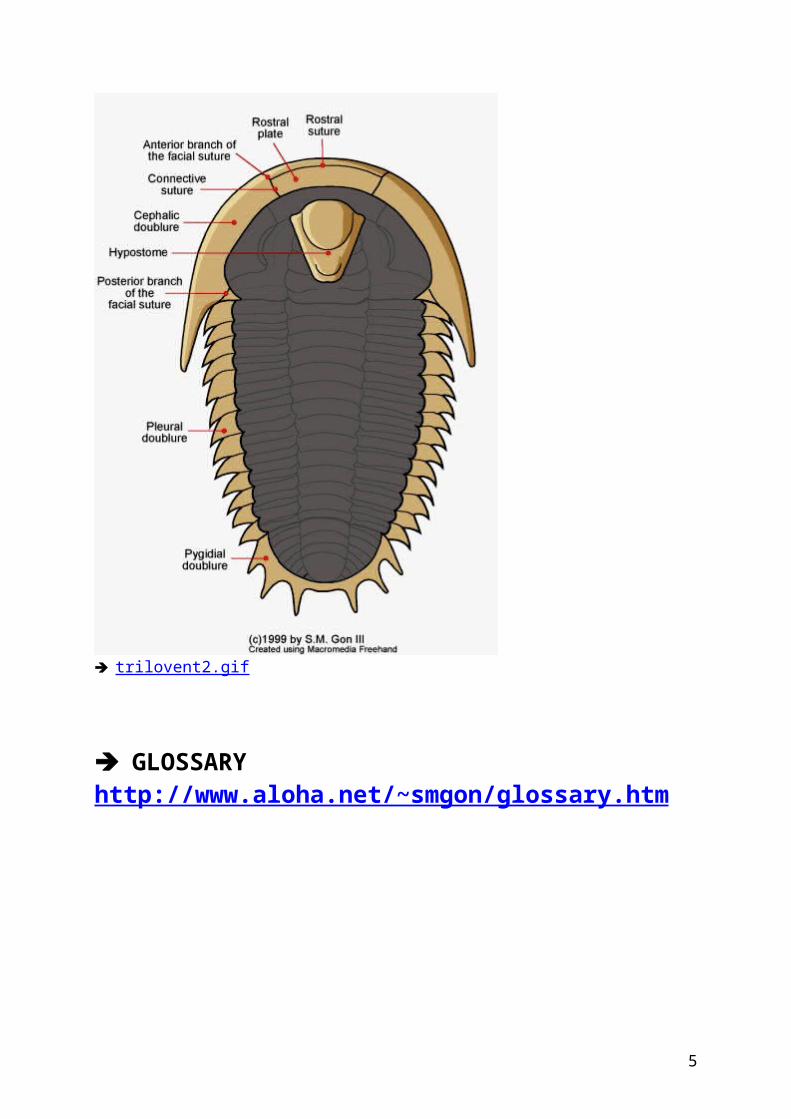
Whatever their size, all trilobite fossils have a similar body plan, being made up of three main body parts: a cephalon (head), a segmented thorax, and a pygidium (tail piece) as shown at left. However, the name "trilobite," which means "three lobed," is not in reference to those three body parts mentioned above, but to the fact that all trilobites bear a long central, or axial lobe, flanked on each side by right and left pleural lobes. These three lobes that run from the cephalon to the pygidium are what give trilobites their name, and are common to all trilobites despite their great diversity of form. You can examine the trilobite body plan in more detail using the links http://www.aloha.net/~smgon/trilomorph.htm ( TRILOBITE BODY )
trilovent2.gif
GLOSSARY http://www.aloha.net/~smgon/glossary.htm
4
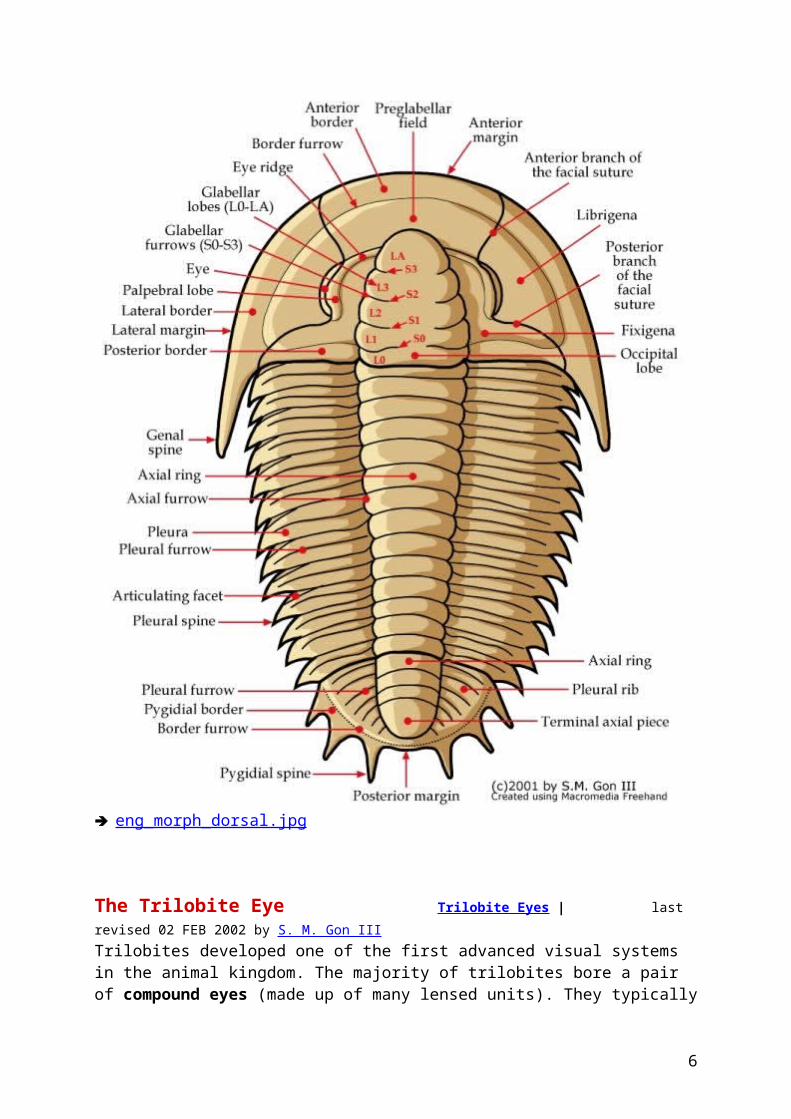
eng_morph_dorsal.jpg
The Trilobite Eye Trilobite Eyes | last revised 02 FEB 2002 by S. M. Gon III Trilobites developed one of the first advanced visual systems in the animal kingdom. The majority of trilobites bore a pair of compound eyes (made up of many lensed units). They typically occupied the outer edges of the fixigena (free cheeks) on either side of the glabella, adjacent to the facial sutures. At least one suborder of trilobites, the Agnostina, are thought to be primarily eyeless. None have ever been found with eyes. In contrast, a few secondarily eyeless species (in which a clear evolutionary trend toward reduced eye size with eventual
5
disappearence of eyes altogether) have developed within several groups, even those known for large, well-developed eyes (e.g., Phacopina).
Holochroal eyes of the Asaphoid trilobite Isotelus
The advantage of good eye design
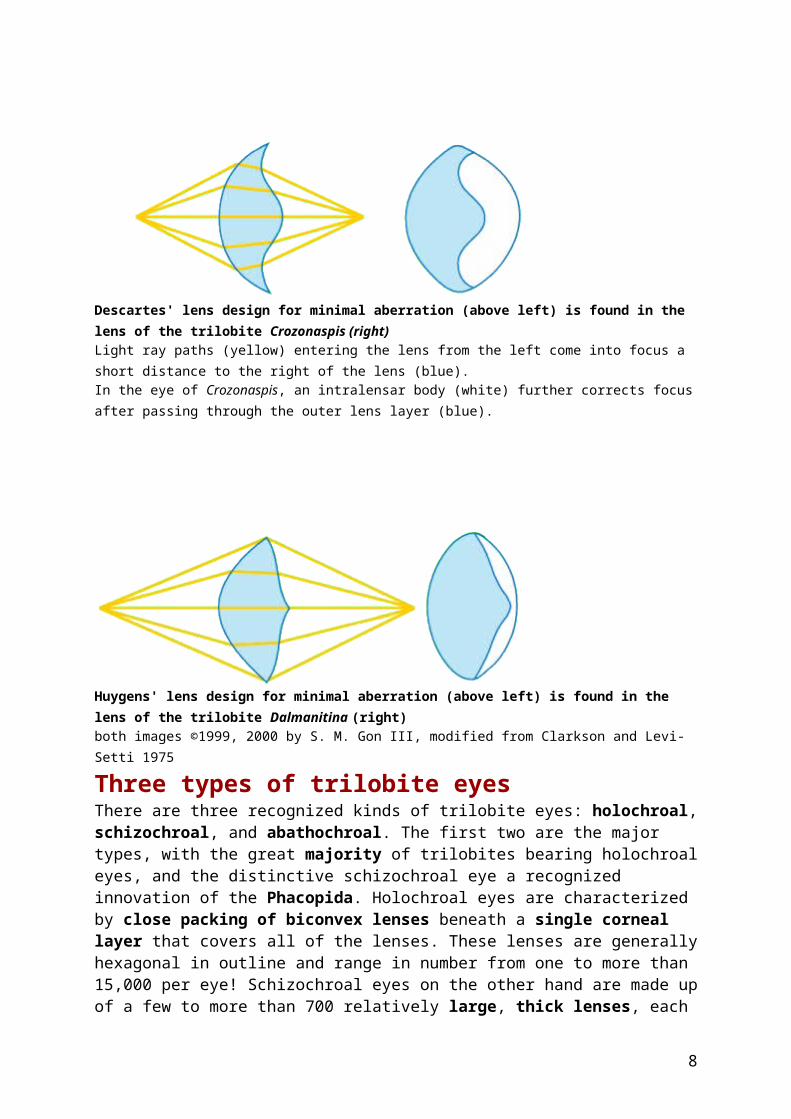
Compound eyes in living arthropods such as insects are very sensitive to motion, and it is likely that they were similarly important in predator detection in trilobites. It has also been suggested that stereoscopic vision was provided by closely spaced, but separate eyes. Vertebrate lenses (such as our own) can change shape (accomodate) to focus on objects at varying distances. Trilobite eyes, in contrast, had rigid, crystalline lenses, and therefore no accomodation. Instead, an internal doublet structure (two lens layers of different refractive indices acting in combination) corrected for focusing problems that result from rigid lenses. The shapes of some trilobite lenses, in fact, match those derived by optical scientists over 300 million years later to answer similar needs. Compare, for example, the optical designs of the 17th century physicists Descartes and Huygens shown below, with those of two trilobite species. The result is that, even without the benefit of accomodation, the rigid trilobite doublet lens had remarkable depth of field (that is, allowed for objects both near and far to remain in relatively good focus) and minimal spherical aberration (distortion of image).
Descartes' lens design for minimal aberration (above left) is found in the lens of the trilobite Crozonaspis (right) Light ray paths (yellow) entering the lens from the left come into focus a short distance to the right of the lens (blue). In the eye of Crozonaspis, an intralensar body (white) further corrects focus after passing through the outer lens layer (blue).
6

Huygens' lens design for minimal aberration (above left) is found in the lens of the trilobite Dalmanitina (right) both images ©1999, 2000 by S. M. Gon III, modified from Clarkson and Levi-Setti 1975
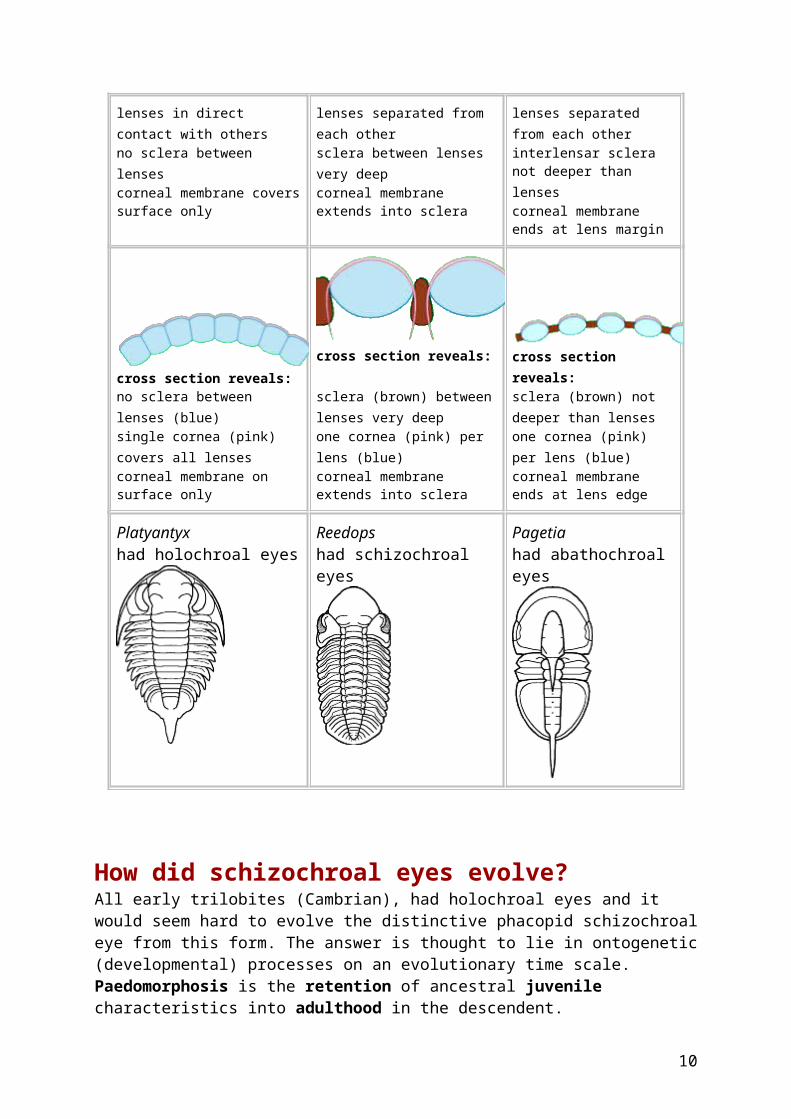
Three types of trilobite eyes There are three recognized kinds of trilobite eyes: holochroal, schizochroal, and abathochroal. The first two are the major types, with the great majority of trilobites bearing holochroal eyes, and the distinctive schizochroal eye a recognized innovation of the Phacopida. Holochroal eyes are characterized by close packing of biconvex lenses beneath a single corneal layer that covers all of the lenses. These lenses are generally hexagonal in outline and range in number from one to more than 15,000 per eye! Schizochroal eyes on the other hand are made up of a few to more than 700 relatively large, thick lenses, each covered by a separate cornea. Each lens is positioned in a conical or cylindrical mounting and is separated from its neighbors by sclera (cuticular exoskeleton material) that extends deeply, providing an anchor for the corneal membrane, which extends downward into the sclera, where it is called intrascleral membrane.The abathochroal eye is seen in only a few Cambrian trilobites and is somehat similar to the schizochroal eye, but differs in some important respects: the sclera is notthick,and the corneal membrane does not extend downward, but ends at the edge of the lens.
The table below illustrates and contrasts the characters of the three eye types.
Holochroal eye
from Clarkson 1975
Schizochroal eye
from Levi-Setti 1993
Abathochroal eye
from Zhang & Clarkson 1990
found in nearly all Orders found in some Phacopida only found in Cambrian Eodiscina only
7
few to very many lenses (to >15,000!)
lenses typically small, numerous one corneal layer covers all lenses lenses in direct contact with others no sclera between lenses corneal membrane covers surface only
typically fewer lenses (to ca 700) lenses much larger, fewer each lens bears an individual cornea lenses separated from each other sclera between lenses very deep corneal membrane extends into sclera
few lenses (to ca 70) lens size small, not numerous each lens bears an individual cornea lenses separated from each other
interlensar sclera not deeper than lenses corneal membrane ends at lens margin
cross section reveals: no sclera between lenses (blue) single cornea (pink) covers all lenses corneal membrane on surface only
cross section reveals: sclera (brown) between lenses very deep one cornea (pink) per lens (blue) corneal membrane extends into sclera
cross section reveals: sclera (brown) not deeper than lenses one cornea (pink) per lens (blue) corneal membrane ends at lens edge
Platyantyx had holochroal eyes
Reedops had schizochroal eyes
Pagetia had abathochroal eyes
How did schizochroal eyes evolve? All early trilobites (Cambrian), had holochroal eyes and it would seem hard to evolve the distinctive phacopid schizochroal eye from this form. The answer is thought to lie in ontogenetic (developmental) processes on an evolutionary time scale. Paedomorphosis is the retention of ancestral juvenile characteristics into adulthood in the descendent. Paedomorphosis can occur three ways: Progenesis (early sexual maturation in an otherwise juvenile body), Neoteny (reduced rate of morphological development), and Post-displacement (delayed growth of certain structures relative to others). The development of schizochroal eyes in phacopid trilobites is a good example of post-displacement paedomorphosis. The eyes of immature holochroal Cambrian trilobites were basically
8

miniature schizochroal eyes. In Phacopida, these were retained, via delayed growth of these immature structures (post-displacement), into the adult form.
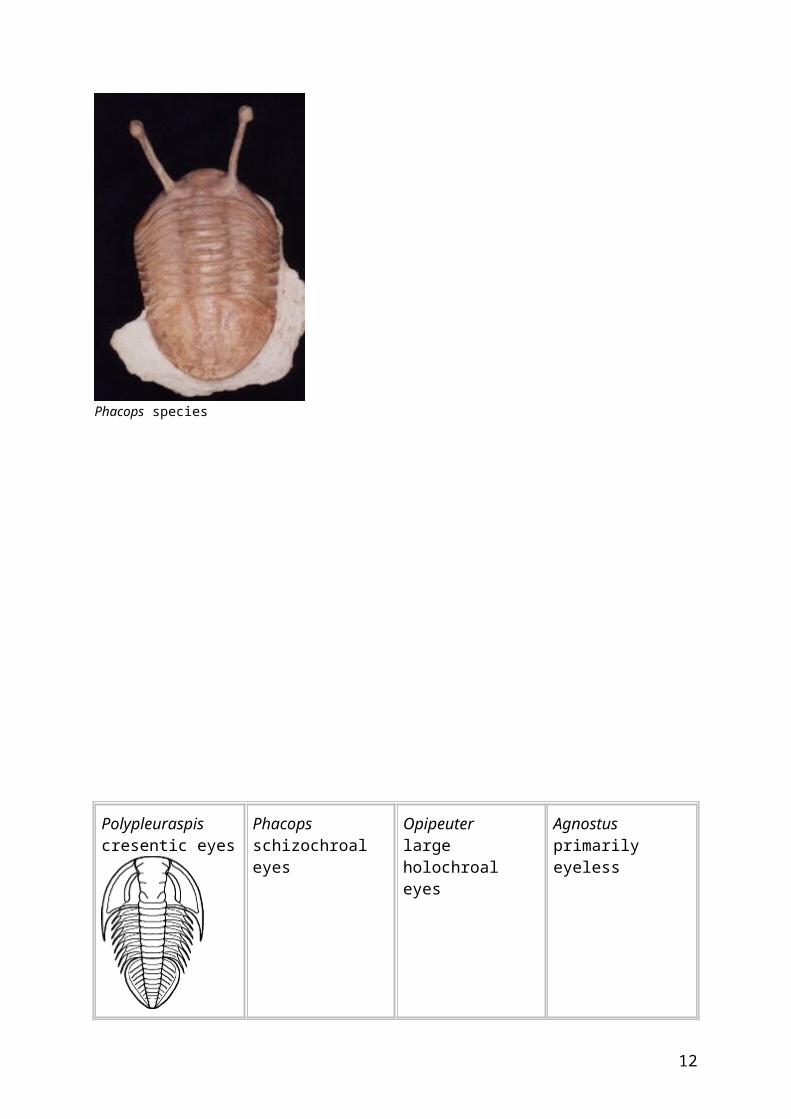
Variation in trilobite eyes As with other aspects of the trilobite body, there was a huge variation of size and form among trilobite eyes, which in many cases seems related to the ecological life style of different species. The figures below show some of these variations. Many of the earliest trilobite eyes were cresentic, such as those of the Corynexochid Polypleuraspis. A conical section of schizochroal eyes gave species such as Phacops an excellent field of vision. In some trilobites, such as the free-swimming pelagic trilobite Opipeuter, the eyes were so large that they dominated the cephalon, providing a 360 degree visual field. Planktonic forms, such as Agnostus, seem to have been entirely blind. Others, such as the Trinucleoid Cryptolithus were bottom feeders with a large, pitted sensory fringe, and eyes were reduced or lost. In species moving through a benthic layer of loose debris or algal growth, eyes raised above the body on stalks could peer about for danger, such as in the strange Russian Asaphoid Neoasaphus (left). Species living on the bottom in deeper waters would have little or no need for eyes at all, and species with reduced eyes, such as Trimerus and secondarily lost eyes, such as Conocoryphe are the result.
Phacops species
9
Polypleuraspis cresentic eyes
Phacops schizochroal eyes
Opipeuter large holochroal eyes
Agnostus primarily eyeless
Cryptolithus secondarily eyeless
Neoasaphus stalked eyes
Trimerus reduced eyes
Conocryphe secondarily eyeless
10

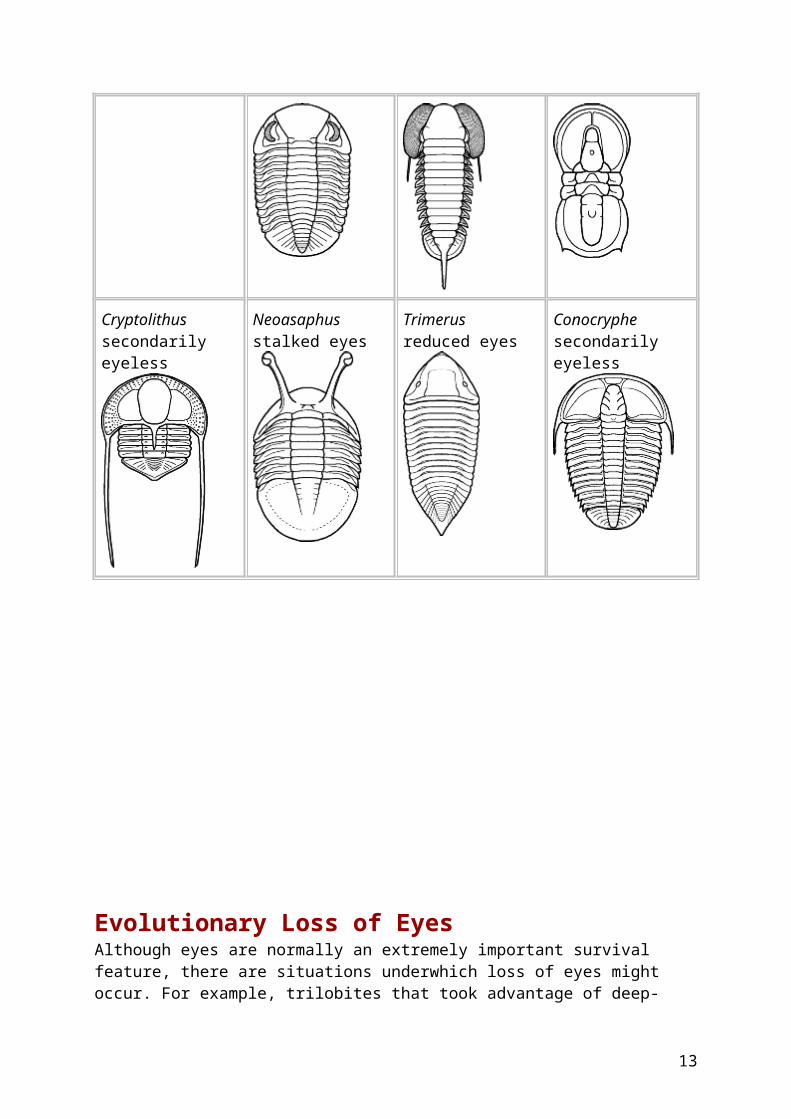
Evolutionary Loss of Eyes Although eyes are normally an extremely important survival feature, there are situations underwhich loss of eyes might occur. For example, trilobites that took advantage of deep-water benthic (bottom-feeding) habitats where light was dim or lacking might have gradually lost their eyes without suffering an adaptive disadvantage. Such eyeless trilobite assemblages are called atheloptic. Such evolutionary trends are repeatedly seen in a variety of trilobite orders, and two examples are shown below. In both cases, these are Devonian trilobites that started with ancestors bearing large, functional eyes. In one sequence, eyes of a Phacopid clade were lost, and facial sutures associated with eyes were also reduced and marginalized. In the other example, involving a Proetid clade, eyes were also reduced and lost, but the basic facial suture pattern was retained. In the figures below (after Fortey & Owens 1999), the eyes are show blue and facial sutures in red
The ancestral Phacops species had large eyes and typical phacopid proparian facial sutures
The proetid Pterocoryphe had large eyes associated with opisthoparian sutures.
Eyes large and typical
Reduction of eyes and a migration forward on the cephalon is seen in the descendant Cryphops.
Greatly reduced eye size marked the genus Pteroparia, descendant of Pterocoryphe.
Eyes reduced in size
Eventually the eyes were lost althogether and the sutures were left along the anterior margin of the cephalon in the genus Trimerocephalus.
Although the eyes are entirely lost in this Pteroparia species, the facial suture patterns are largely unchanged.
11
Eyes lost entirely
all line drawings this page ©1999, 2000, 2001 by S. M. Gon III SEM images of the trilobite schizochroal eye courtesy of Dr. Riccardo Levi-Setti from his superb book "Trilobites" CITATIONS
Clarkson, E. N. K. 1975. The evolution of the eye in trilobites. Fossils and Strata 4:7-31. Clarkson, E. N. K. & R. Levi-Setti. 1975. Trilobite eyes and the optics of Des Cartes and Huygens. Nature 254 (1975): 663-667. Fortey, R. A. & R. M. Owens. 1999. The Trilobite Exoskeleton. in: Functional Morphology of the Invertebrate Skeleton. John Wiley & Sons. Levi-Setti, R. 1993. Trilobites (2nd Ed.). University of Chicago Press. Moore, R. & Kaesler 1997. Treatise on Invertebrate Paleontology, Part O, Arthropoda 1, Trilobita, Revised. Zhang, Xiguang & E. N. K. Clarkson. 1990. The eyes of lower Cambrian eodiscid trilobites. Palaeontology 33:911-932.
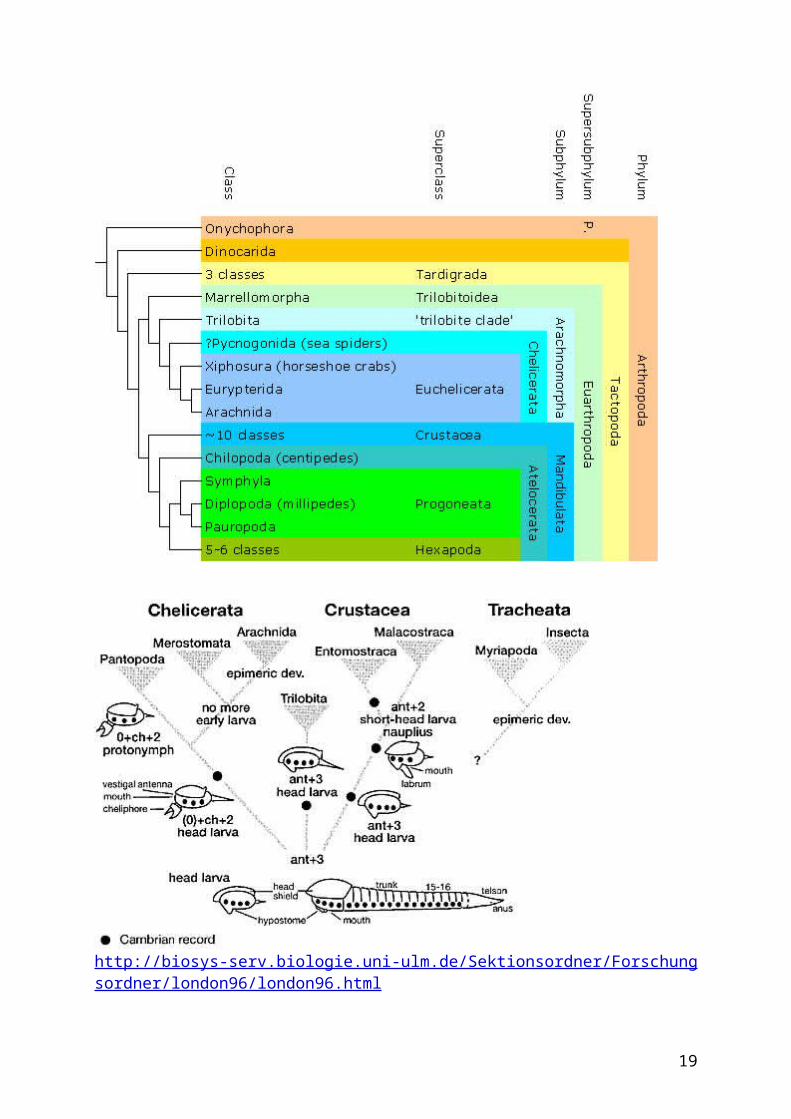
Systrematics & phylogeny trilobitomorpha.gif
As traditionally defined, the true Trilobita are divided into two main groups, the Agnostida and the Polymerida. Agnostids have sometimes been considered non-trilobites -- even crustaceans, by some -- but recent analyses (e.g. Fortey and Theron 1994) support the inclusion of the Agnostida in the Trilobita.
12
A third group, the Nektaspida, was not identified until the 1980s; its members include soft-bodied arthropods from the Burgess Shale such as Naraoia. Some, such as Gould (1991), have classified the Nektaspida as true trilobites; others (e.g. Simonetta and Delle Cave, 1991) have disagreed. On this cladogram, which is based on the work of Fortey and Theron (1994) and Wills et al. (1993), the Nektaspida are the sister taxon to the polymerid and agnostid trilobites; whether the Nektaspida should be considered "true" trilobites or not is largely a matter of definition.
Many other Cambrian arthropods, especially those from the world-famous Burgess Shale, were once considered probable trilobite relatives, or "trilobitomorphs." While many of these problematic arthropods are no longer thought to be related to the true trilobites, recent studies have shown that a number of them are closer to the trilobites than to any other arthropods. The term "Trilobitomorpha" is used here in this more restricted sense. Exactly how these arthropods are related to the trilobites is not yet perfectly clear, but the non-trilobite "trilobitomorphs" probably compose several clades. One of the clades that probably fits in here is called the Emeraldellida.
Yet another group of arthropods, the aglaspids, have traditionally been grouped with the chelicerates, but may be closer to trilobites. Some authors also group the emeraldellids with the cheliceramorphs as well (e.g. Simonetta and Delle Cave, 1991). Whether or not these groups actually belong to the cheliceramorphs, it seems likely that the cheliceramorphs are the closest living group to the trilobites and their relatives.
Sources:
Simonetta, A.M. and Delle Cave, L. 1991. Early Paleozoic arthropods and problems of arthropod phylogeny, with some notes on taxa of doubtful affinities. In: Simonetta, A.M. and Conway Morris, S. (eds.) The Early Evolution of Metazoa and the Significance of Problematic Taxa. Cambridge University Press, Cambridge.
Fortey, R.A. and Theron, J.N. A new Ordovician arthropod, Soomaspis, and the agnostid problem. Palaeontology 37(4): 841-861.
Gould, S. J. 1989. Wonderful Life. W. W. Norton, New York.
Wills, M. A., D. E. G. Briggs, and R. A. Fortey. 1994. Disparity as an evolutionary index: a comparison of Cambrian and Recent arthropods. Paleobiology 20(2): 93-130.
13

TRILOBIETEN http://www.bloggen.be/evodisku/archief.php?ID=57
Een van de overvloedig voorkomende fossielen vanaf het vroege cambrium ( Cambrian ) , behoren tot de uitgebreide en de succesvolle substam (biologie) der Trilobieten (Trilobites trilobites ) .... ( zie ook http://www.fossiel.net/informatie/glossarium.php?term=trilobieten http://www.trilobites.info/ )
De thans uitgestorven trilobieten waren de meest voorkomende wezens in de zeeën tijdens het cambrium, ongeveer 500 miljoen jaar geleden. Uit de vroege soorten ontwikkelden zich vele andere soorten en ze bereikten hun hoogtepunt in de ordovicium-periode. Ze waren nog steeds van belang tijdens het silurium, maar daarna bleven er slechts enkele soorten meer over. De laatste trilobieten kwamen nog voor gedurende perm-periode, zo'n 220 miljoen jaar geleden, waarna ze voorgoed verdwenen ( de grote uitstervinsgolf op het einde van het perm) .
ARTHROPODA CLADE 30 Jul 2006 http://www.peripatus.gen.nz/Taxa/Arthropoda/Index.html
14

http://biosys-serv.biologie.uni-ulm.de/Sektionsordner/Forschungsordner/london96/london96.html
15

Deze oude arthropoda worden door zowel YEC als OEC creationisten ook veelal aangevoerd als een "overduidelijk icoon//bewijs" van de creationistische verklaring voor ( =de plotse creatie van de soorten door een ID -er )tijdens de "cambrische explosie " ...(1)
Eigenlijk beweren creationisten dat ook binnen de soortenexplosie van de trilobieten (ruim 17.000 soorten bekend en er worden er regelmatig nieuwe gevonden ), geen overgangsvormen(=transitionnals ) kunnen worden waargenomen ....Overgangsvormen bestaan immers niet volgens deze creato's en -dat derhalve ook de trilobieten niet zijn verder geevolueerd - noch verder opgesplits (= speciatie en radiatie ) in verschillende soorten waarvan de vele fossiele soorten die ondertussen zijn bekend ____ zoals dit wordt verklaard ; Conform de huidige consensus in de paleontologie , zijnde de "beste "verklaring van de waarnemingen en het fossielen archief ....
The Evolution of Trilobite Body PatterningFirst published online as a Review in Advance on January 29, 2007 The Evolution of Trilobite Body PatterningNigel C. Hughes
Department of Earth Sciences, University of California, Riverside, California 92521; email: [email protected]
Abstract
The good fossil record of trilobite exoskeletal anatomy and ontogeny, coupled with information on their nonbiomineralized tissues, permits analysis of how the trilobite body was organized and developed, and the various evolutionary modifications of such patterning within the group. In several respects trilobite development and form appears comparable with that which may have characterized the ancestor of most or all euarthropods, giving studies of trilobite body organization special relevance in the light of recent advances in the understanding of arthropod evolution and development. The Cambrian diversification of trilobites displayed modifications in the patterning of the trunk region comparable with those seen among the closest relatives of Trilobita. In contrast, the Ordovician diversification of trilobites, although contributing greatly to the overall diversity within the clade, did so within a narrower range of trunk conditions. Trilobite evolution is consistent with an increased premium on effective enrollment and protective strategies, and with an evolutionary trade-off between the flexibility to vary the number of trunk segments and the ability to regionalize portions of the trunk.
bron ;http://arjournals.annualreviews.org/doi/abs/10.1146/annurev.earth.35.031306.140258?journalCode=earth
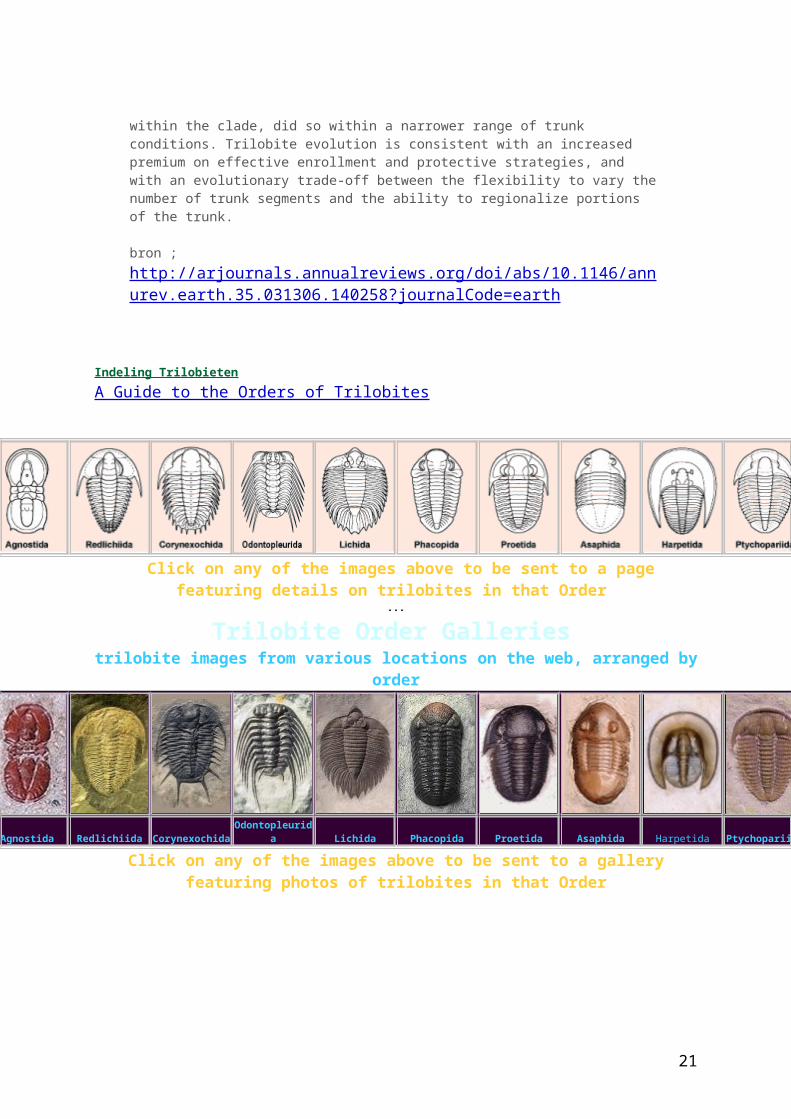
Indeling Trilobieten A Guide to the Orders of Trilobites
16

Click on any of the images above to be sent to a page featuring details on trilobites in that Order
...
Trilobite Order Galleries trilobite images from various locations on the web, arranged by order
Agnostida Redlichiida Corynexochida Odontopleurida Lichida Phacopida Proetida Asaphida Harpetida Ptychopariida
Click on any of the images above to be sent to a gallery featuring photos of trilobites in that Order
De evolutie van de trilobieten is namelijk zo uitgebreid ,divers en relatief( geologisch ) zo snel (2) gebeurt dat deze vondsten uitstekende gidfossielen (3) blijken te zijn
Trilobieten komen over de gehele wereld als fossiel voor (http://www.fossiel.net/vindplaatsen/vindplaatsensoort.php?soort=trilobieten) :
Een vroeg voorbeeld waarbij de oudste en meest "primitieve )trilobieten ( voornamelijk de orde Redlichiida (in het bijzonder de Suborde Olenellina) in evolutionaire en geologische( stratigrafische ) series(4) voorkomen ( samen met andere getuigen van de toenmalige fauna's ) zijn te vinden in california ( http://www.ucmp.berkeley.edu/cambrian/marblemts.html )
Trilobieten series in de Marble & Providence Mountains ca
Bekijk de afbeelding op ware grootte.
17

The Latham Shale, Marble Mountains, San Bernadino County, California.
www.ucmp.berkeley.edu/cambrian/marblemts.html
http://members.cox.net/jdmount/cambmojave1.html
18
Bristolia insolens, trilobietkop-pantser uit de bovenste/jongste laag van de formatie .
Bristolia Bristolia insolens (Resser, 1928). Cephalon. Onder- Cambrium, Latham shale, dicht bij de top van de formatie /Cadiz, San Bernadino County, CA.
Text-fig. A
19

Bristolia bristolensis trilobietkop-pantser uit de midden laag van de serie .
Latham shale,( vroeg cambrium) San Bernadino County, CA.
Olenellus mohavensis, trilobietkop-pantser uit de midden- laag van de formatie.
20
Olenellus fremonti, trilobietkop-pantser uit de onderste/oudste laag van de serie .
Olenellus fremonti (Walcott, 1910). >laag Cambrium , Latham shale, about 10m boven de basis van de formatie /Cadiz, San Bernadino County, CA
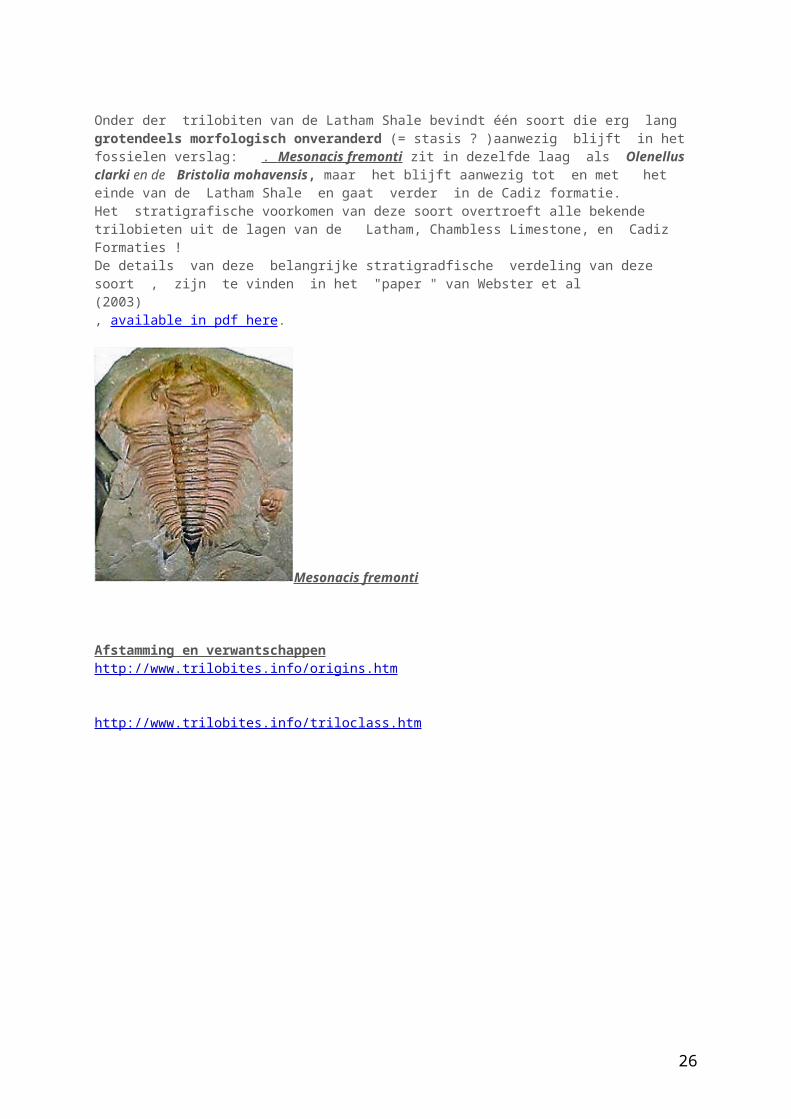
Onder der trilobiten van de Latham Shale bevindt één soort die erg lang grotendeels morfologisch onveranderd (= stasis ? )aanwezig blijft in het fossielen verslag: . Mesonacis fremonti zit in dezelfde laag als Olenellus clarki en de Bristolia mohavensis, maar het blijft aanwezig tot en met het einde van de Latham Shale en gaat verder in de Cadiz formatie. Het stratigrafische voorkomen van deze soort overtroeft alle bekende trilobieten uit de lagen van de Latham, Chambless Limestone, en Cadiz Formaties ! De details van deze belangrijke stratigradfische verdeling van deze soort , zijn te vinden in het "paper " van Webster et al (2003), available in pdf here.
21

Mesonacis fremonti
Afstamming en verwantschappen http://www.trilobites.info/origins.htm
http://www.trilobites.info/triloclass.htm
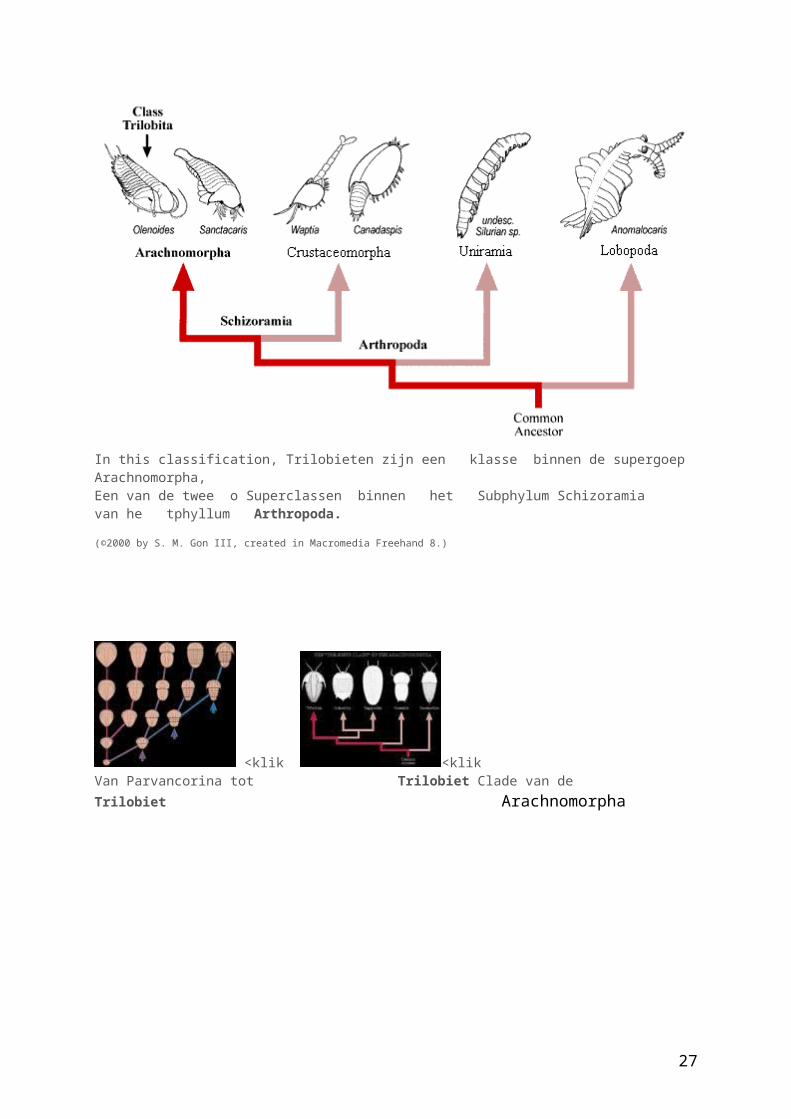
In this classification, Trilobieten zijn een klasse binnen de supergoep Arachnomorpha, Een van de twee o Superclassen binnen het Subphylum Schizoramia van he tphyllum Arthropoda. (©2000 by S. M. Gon III, created in Macromedia Freehand 8.)
22

<klik <klikVan Parvancorina tot Trilobiet Clade van de Trilobiet Arachnomorpha
Parvancorina minchami – een van de oudste arthropoden( Laat precambrium/begin cambrium) , een vroege voorloper- trilobiet(?) (Fortey et al. 1996).http://www.peripatus.gen.nz/Taxa/Arthropoda/Trilobita/TriOri.htmlhttp://gsa.confex.com/gsa/2003AM/finalprogram/abstract_59267.htm
Systematische verwantschap en verdeling over de geologische tijd van de verschillende trilobieten orden
23

In deze figuur is Redlichiida , voorgesteld als de basale ( primitieve )trilobieten - orde ____ te vinden in de afzettingen striktbepaerkt tot het cambrium http://www.trilobites.info/ordredlichiida.htm
( ©2007 by S. M. Gon III, created using Macromedia Freehand and PaintShop Pro Thanks to Nigel Hughes for stimulating discussions leading to revisions of this figure.As of June 2007, a version of this figure was published in Hughes 2007.http://www.trilobites.info/triloclass.htm#orders )
NOTEN
(1) bijvoorbeeld
De Yec/ID creationist Peter Borger ; "....Het waren gewoon multipurpose genomes waar de trilobieten over beschikten. Vol met genetiese redundanties en Variatie Inducerende Genetiese Elementen (VIGEs) die chromosomen transloceren en aldus de bouwplannen wijzigen. ".....Het gaat hier echter wel over de verwantschappen en soortvorming van 15.000 t/m 17000 bekende species gespreid vanaf het vroegste cambrium t/m het einde van het Perm....zie ook (2b)
( Uit de onnoemelijke YEC site " Evobeliever " /( vertalingen van artikels uit "creation science" ) " ....Neem de trilobieten als voorbeeld. Deze fossielen zijn zo gewoon dat u er een kan kopen onder de 15 €, maar er werden nooit fossielen van een voorouder gevonden!XML:NAMESPACE PREFIX = O />
..."
24

de "oude aarde" ( islamisme ) creationist Harun Yayah :
" levende wezens die in de laag gevonden worden die tot het Cambrium behoorde, verschenen opeens in het fossielenarchief - er zijn geen voorouders die eerder bestonden. De fossielen die in de rotsen van het Cambrium gevonden zijn, zijn van slakken, trilobieten, sponzen, aardwormen, kwallen, zee-egels en andere ingewikkelde ongewervelden. Dit omvangrijke mozaïek van levende wezens vormde zo'n groot aantal ingewikkelde levensvormen, die zo plotseling verschenen, dat deze wonderlijke gebeurtenis in de geologische literatuur wordt aangeduid met de 'Explosie van het Cambrium'.
Trilobieten-fossielen met hun skelet, complexe ogen en ledematen, 'verschijnen' verbazingwekkend in de lagen van de "ongeëvenaarde " cambrische explosie van leven op aardeLevensvormen ( ook de trilobieten )blijken complex te zijn, zelfs die welke werden aangetroffen in de ''oudste' lagen van het fossiele verleden. Zo blijken verschillende soorten trilobieten een zeer geavanceerd gezichtsvermogen te hebben. Toch beweren evolutionisten dat deze wezentjes zich waarschijnlijk geleidelijk ontwikkeld hebben in de tijd dat de eerste meervoudige levensvormen zich beginnen te ontwikkelen, zo'n veronderstelde 620 miljoen jaren geleden 1.- Veel primitievere (= de oudste ) trilobieten bezitten GEEN ogen 2.- Hoe meer variatie een soort( en uberhaupt later gedivergeerde verschillende soorten ) kent, hoe meer ruw materiaal natuurlijke selectie heeft om mee te werken.( voozichtige creationisten claimen enkel ;)In 'oudere' aardlagen zijn geen voorlopers van trilobieten gevonden.Ze verschenen dus volledig gevormd, met al hun organen en structuren. Maar de trilobieten stierven ook volledig uit /in hoeverre is dit verzoenbaar met creatie ? Wat is het nut geweest van deze dieren, als het einddoel de mens en de natuur om hem heen is? Waartoe hebben de dinosauriërs geleefd, en de trilobieten?Voor de richtingloze natuurlijke selectie is de wereld op elk moment af.Waarom de trilobieten 350 miljoen jaar geleden bleken verdwenen te zijn is niet meer te achterhalen maar we hebben de zekerheid dat dit ons ook te wachten staat en alle logica laat vermoeden dat daarmee ook het zelfbewustzijn zal verdwenen zijn van deze aarde.
(2)a) Het tempo van de veranderingen( en aanpassingen ) werden vooral veroorzaakt door de vele tektonische , klimatologische e.a. geologische en fysico-chemische fenomenen die aan het begin en tijden het cambrium schering en inslag waren op deze planeet...
http://findarticles.com/p/articles/mi_qa3790/is_199903/ai_n8829266/pg_1
Testing the Darwinian legacy of the Cambrian radiation using Trilobite phylogeny and biogeography
25
Journal of Paleontology, Mar 1999 by Lieberman, Bruce S
ABSTRACT-Since the publication of Darwin (1859), the biological meaning of the Cambrian radiation * has been debated. Most commentators agree, however, that the Cambrian radiation is fundamentally a time of major metazoan cladogenesis. In and of itself this does not necessarily mean that unique evolutionary processes operated during the Cambrian radiation. Phylogenetic analysis has been used to study the tempo of speciation during the radiation, and thus far there is no need to invoke special rules relating to the tempo of evolution. Instead, what seems unique about the Cambrian radiation is its place as an important episode in the history of life-that is, as the first major radiation of the Metazoa. Although the tempo of evolution during the Cambrian radiation may not have been uniquely high, there were largely unique tectonic events that transpired during the late Neoproterozoic and Early Cambrian, such as extensive cratonic fragmentation. Biogeographic analysis of Early Cambrian olenelloid trilobites reveals that these tectonic events powerfully influenced evolutionary and distributional patterns in this diverse and abundant trilobite group.
This emphasizes the importance of physical earth history in generating evolutionary patterns. In the general study of macroevolutionary patterns and processes, earth history phenomena emerge as powerful forces influencing the history of life and provide insights into evolution that can best be inferred by paleontological data.
* Cambrian radiation ( =cambrische "vertakkingen " van de levensboom in phyla ) = Creationisten spreken natuurlijk liever van "cambrian explosion "
b) Snelle evolutie ( althans = SPECIATIE , ( specifieering=soorten-onstaan door bijvoorbeeld ecomorfen ) en het geologisch erg vlug onstaan van soortenzwermen ) is ook geopperd bij de CICHLIDEN , ANOLIS HAGEDISSEN en (vermoedelijk ) de oude fossiele Coelacanthen ....
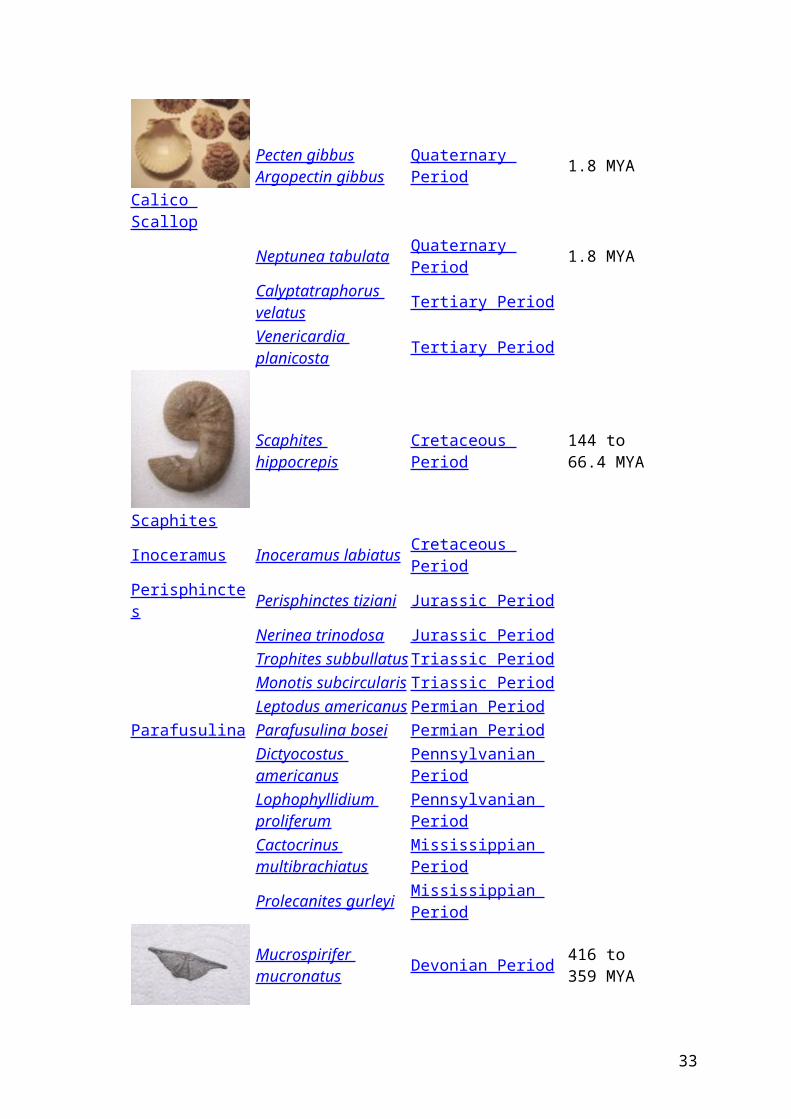
(3)voorbeeld van enkele Gidsfossielen ( =Index fossil )
26
List of Common Index Fossils
Fossil Scientific Name Time Period Million Years Ago
Calico Scallop
Pecten gibbusArgopectin gibbus Quaternary Period 1.8 MYA
Neptunea tabulata Quaternary Period 1.8 MYACalyptatraphorus velatus Tertiary Period
Venericardia planicosta Tertiary Period
27

Scaphites
Scaphites hippocrepis Cretaceous Period 144 to 66.4 MYA
Inoceramus Inoceramus labiatus Cretaceous PeriodPerisphinctes Perisphinctes tiziani Jurassic Period
Nerinea trinodosa Jurassic PeriodTrophites subbullatus Triassic PeriodMonotis subcircularis Triassic PeriodLeptodus americanus Permian Period
Parafusulina Parafusulina bosei Permian PeriodDictyocostus americanus Pennsylvanian Period
Lophophyllidium proliferum Pennsylvanian Period
Cactocrinus multibrachiatus Mississippian Period
Prolecanites gurleyi Mississippian Period
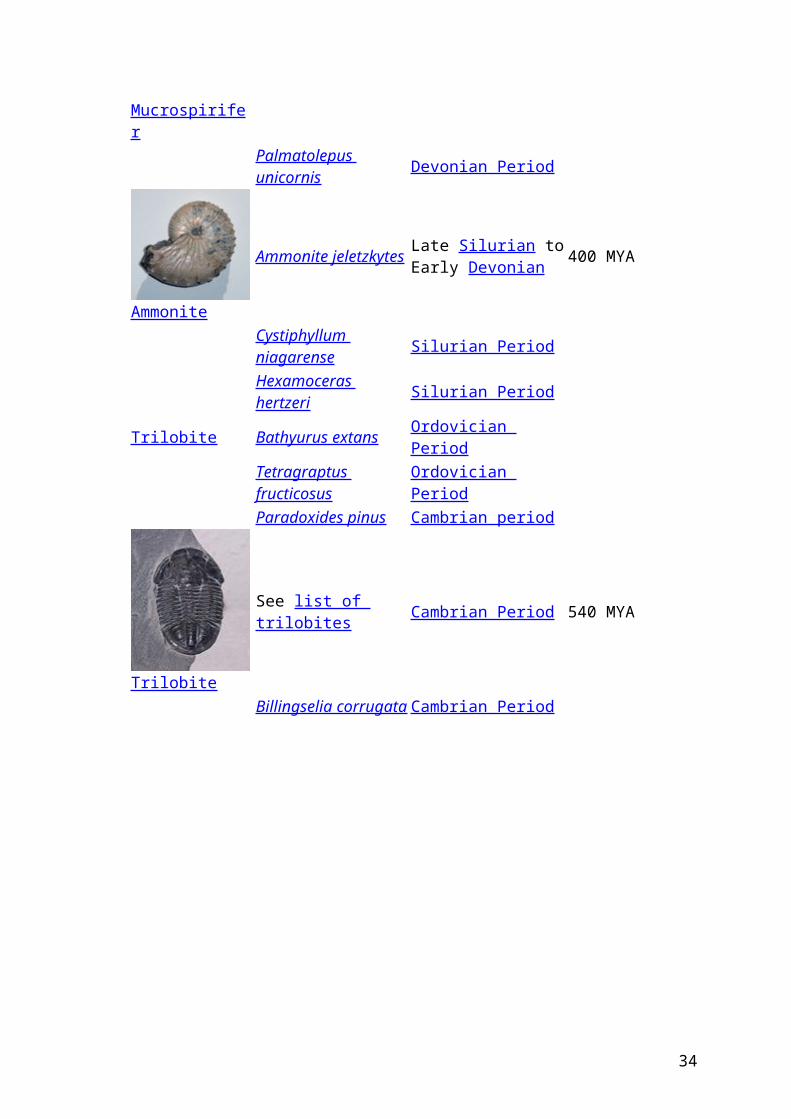
Mucrospirifer
Mucrospirifer mucronatus Devonian Period 416 to 359
MYA
Palmatolepus unicornis Devonian Period
Ammonite
Ammonite jeletzkytes Late Silurian to Early Devonian 400 MYA
Cystiphyllum niagarense Silurian Period
Hexamoceras hertzeri Silurian PeriodTrilobite Bathyurus extans Ordovician Period
Tetragraptus fructicosus Ordovician Period
Paradoxides pinus Cambrian period
28

Trilobite
See list of trilobites Cambrian Period 540 MYA
Billingselia corrugata Cambrian Period
Enkele cambrische trilobieten van groot biogeografisch /biostratigrafisch belang
29

Biostratigrafie
< <
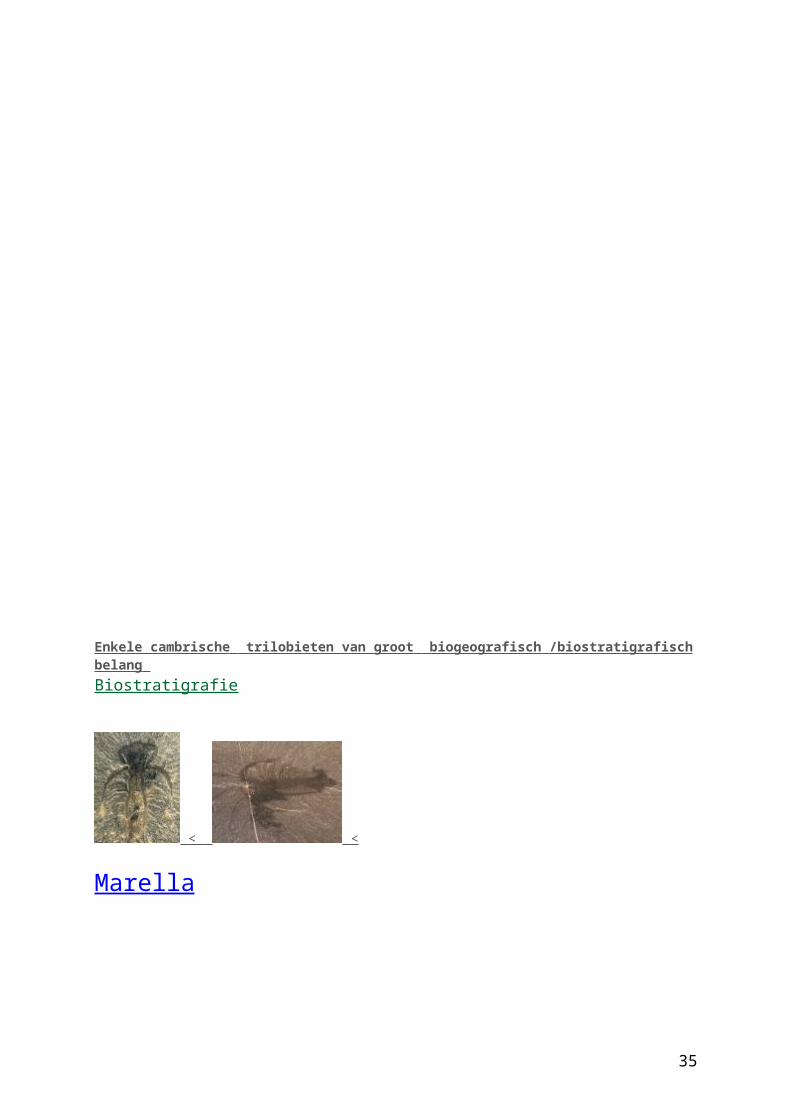
Marella
This fascinating reconstruction of Marrella, prepared using the ink wash technique, was found in the drawers containing archival collections belonging to Charles Doolittle Walcott (1850-1927), fourth secretary of the Smithsonian Institution and discoverer and collector of the famous fossils of the Burgess Shale. Marrella is one of the most common fossils found at the Cambrian Burgess Shale locality in British Columbia, Canada.
Publication:Walcott, Charles Doolittle. Addenda to Descriptions of Burgess Shale Fossils. Smithsonian Miscellaneous Collections, Volume 85, Number 3. Figure 9. Published by the Smithsonian Institution, City of Washington, June 29, 1931.
30

http://www.volkskrantblog.nl/bericht.php?id=40809 http://www.volkskrantblog.nl/bericht.php? id=18516
Marrella-achtigen
Wetenschappelijke naam: Marrellomorpha (klasse of onderklasse binnen de Geleedpotigen)
Marrella-achtigen zijn uitgestorven geleedpotigen (Arthropoda) die leefden van het Cambrium tot en met het Devoon (ca. 540-360 miljoen jaar geleden). Aan hun lichaam hadden zij tweetakkige aanhangsels met kieuwen. De kop droeg stekels en twee paar aanhangsels. Marrella-achtigen waren vrijzwemmende zeedieren.
http://www.museumkennis.nl/nnm.dossiers/museumkennis/i000440.html
31
oudste trilobieten (e.g., Fallotaspis longa)FAD Series 2, Stage 3
Oryctocephalus indicusFAD Series 3, Stage 5
Lejopyge laevigatusFAD Series 3, Stage 7
Agnostotes orientalisFAD Furongian Series, Stage 9
Olenellus gilberti (e.a. olenellids)FAD
Series 2, Stage 4
Ptychagnostus atavusFAD
Series 3, Drumian Stage (6)
Glyptagnostus reticulatus FAD
Furongian Series, Paibian Stage (8)
Lotagnostus americanusFAD
Furongian Series, Stage 10
Geokolom en trilobieten.docThe Late PaleozoicOrdovician
Ohio State Fossil Ohio Ordovician Isotelus
[PDF] GeoFacts No. 5 [PDF]
GeoFacts No. 6 , Isotelus : Ohio's state fossil
32
Marokko --> Onnia superba Onnia superba Moroccan Trilobites Death Assemblage
Onnia superba
Trilobites Order Asaphida, Family Trinucleidae
Geological Time: Ordovician
Size: (25.4mm=1 inch): see descritption
Fossil Site: Blekos, Morocco
This is SUPERB death assemblage of a member of the Order Asaphida, Family Trinucleidae from the Ordovician deposits of Blekos, Morocco known as Onnia superba. Cointained within this 6.7 inch by 5.7 inch matrix plaque are TEN examples of the type 0.5 inch - 0.8 inch wide at the genal spines. This is a NATURAL occurence, NOT a composite. There are many fine details present, such as the tiny occipital spine and the fine pitted cephalic ring. The trinucleids made their living by kicking up detritus from the seafloor, filtering it through the cephalic ring.
What catastrophe resulted in this mass death?
click to enlarge
33
about Russian Trilobites Illaenus Russian Trilobite Image Gallery
Name: Illaenus incisus Russian trilobites
Geologic Time: Middle Ordovician
Size (25.4 mm = 1 inch): 79 mm long by 39 mm wide on a 90 by 30 mm matrix
Location: Asery Horizon deposits of the Wolchow River region near Saint Petersburg, Russia
Description: This is an example of the rarely offered trilobite, Illaenus incisus. The genus derives its specific name from the cut (incised) appearance to its thoracic segments, This is one of the 50-odd members of the genus, and known for the high profile of the cephalon which is thought to have aided the trilobite in plowing through the substrate in search of a meal. This one is presented wrapped atop its matrix that has a flattened base for display from any aspect.
Click picture to enlarge
34
Wisconsin State Fossil Wisconsin Ordovician – Silurian
Calymene celebra
..The Devonian Pennsylvania State Fossil Pennsylvania Devonian Phacops rana
Marokkaanse trilobieten uit het devoon Coltraenia oufatenensisDicranurus monstrosusWalliserops trifurcatus..The Carboniferous: Coal Swamps and Glaciers
..The Permian http://noorderlicht.vpro.nl/artikelen/22046992/..The Great Permian Extinction
http://noorderlicht.vpro.nl/artikelen/18544626/
(4)
35

Een andere uitgebreide "serie " die duidelijk de verwantschappen en afstammingslijnen aantoont ( inclusief transitionnals tussen verschillende species ) is te vinden in het fossielen archief van het genus Phacops ( Phacops rana Pennsylvania Geological Survey: The State Fossil ) Phacopida
(; Phacops rana ;Eldredge, Niles, 1972.1974 Systematics and evolution of Phacops rana (Green, 1832) Phacops iowensis Delo, 1935 (Trilobita) from the Middle Devonian of North America. ;Strapple 1978).
Er wordt aangevoerd dat de onderzochte / waargenomen gevallen van ( allopatrische ) speciatie te maken hebben met de ramificatie van de ring- soort ( met mondiale verspreiding) Phacops rana , in verschillende subspecies ( ondersoorten of rassen ) (= creationisten spreken in dit verband van microevolutie )
De over gang tussen soorten is vaak gradueel terwijl de naamgeving een strikte scheiding doet vermoeden. Zo heb je bijvoorbeeld soort A die kan kruisen met B en vruchtbare nakomelingen kan krijgen B kan kruisen met A en C maar A en C kunnen niet met elkaar kruisen, dit heet ring soort.
Een bekend voorbeeld is een zogenaamde "ringsoort" bij salamanders( http://www.santarosa.edu/lifesciences2/enreview.htm )maar er zijn nog veel meer voorbeelden bekend.... ringsoortvorming komt vooral voor bij soorten die over een groot uitgestrekt gebied leven maar zelf niet zo mobiel zijn.( = veel kosmopoliete planten )
* Echter ook bij gespecialiseerde diersoorten ( = sommigen zitten vast tijdens hun volwassen leven (= sessiel ) , maar verspreiden zich als "larven " die rondzwemmen in het plankton ) ; * Ook " honden " zijn eigenlijk een ringsoort ; een chihuahua kan nooit paren met een deense dog , om evidente anatomische redenen .
36


























![Kuliah 3 arthropoda-trilobita celicherata [compatibility mode]](https://img.pdfslide.net/doc/110x75/5873102d1a28ab99088b7a4f/kuliah-3-arthropoda-trilobita-celicherata-compatibility-mode.jpg)


