Embed Size (px)
Citation preview

Kidney International, Vol. 61 (2002), pp. 1365–1376
Troglitazone suppresses the secretion of type I collagen bymesangial cells in vitro
ROBERT EARL ROUTH, JOHN HARDWICK JOHNSON, and KEVIN JOHN MCCARTHY
School of Medicine, Louisiana State University Health Sciences Center, Shreveport, Louisiana, and Pfizer, Incorporated,Ann Arbor, Michigan, USA
Troglitazone suppresses the secretion of type I collagen by mes- The increased incidence/progression of diabetic ne-angial cells in vitro. phropathy, that is, the gradual pathophysiological changes
Background. Our laboratory has shown that troglitazone, a that occur within the nephron, has been linked to thethiazolidinedione and peroxisomal proliferator activated re-degree of glycemic control in individuals afflicted withceptor � (PPAR-�) agonist, prevents mesangial expansion andeither type I or type II diabetes mellitus [1]. In the earlyglomerulosclerosis in diabetic rats. We investigated and com-
pared the action of two PPAR agonists at the level of the mes- stages of type II diabetes mellitus, glycemic control inangial cell. most individuals can be maintained by a combination of
Methods. Rat mesangial cells were grown in medium contain- dietary restrictions, decreased caloric intake, and exer-ing 5 mmol/L glucose, 30 mmol/L glucose, or 5 mmol/L glucosecise. For individuals unresponsive to or unwilling to fol-plus 25 mmol/L mannitol. The cultures were either left untreated,
treated with 10 �mol/L troglitazone (PPAR-�), or 100 �mol/L low such regimens, oral hypoglycemic agents such asclofibrate (PPAR-�). The following parameters were used to tolbutamide and tolazamide were developed as an alter-assess mesangial cell responses: detection of PPAR-� and -� native to insulin injections [2]. The primary action ofmRNA, the degree of PPAR-� and -� activation, spread cell
these drugs, as well as second and third generation deriv-area, total protein production, and laminin and type I collagenatives, is to stimulate the release of insulin from existingproduction.
Results. Reverse transcription-polymerase chain reaction pancreatic beta cells. Although these agents have been(RT-PCR) showed the presence of PPAR-� and -� mRNA in successfully used to correct the hyperglycemia of type IIrat mesangial cells. PPAR-� and -� proteins are active in mesan- diabetes mellitus, often individuals become refractory togial cells and the extent of activation is affected by different
the actions of these drugs after prolonged usage due toglycemic conditions. Troglitazone and clofibrate treatment cor-beta cell failure [2].rected in part the increase in spread cell area seen in mesangial
cells in hyperglycemic conditions. However, neither agonist A new class of agents, the thiazolidinediones (troglita-corrected the increase in total protein production induced by zone, rosiglitazone, pioglitazone, and ciglitazone), hashyperglycemia. Treatment with troglitazone resulted in a sig-
provided an alternative therapeutic modality to the treat-nificant, specific decrease in type I collagen along with a slightment of the abnormal physiology of type II diabetes mel-decrease in laminin production in both medium conditions.
Clofibrate had no effect on laminin synthesis in either medium litus. Rather than act directly on the pancreas to increasecondition but did decrease type I collagen synthesis in cells the secretion of insulin by beta cells, the thiazolidinedi-grown in hyperglycemic conditions. ones exert their effects by sensitizing peripheral tissuesConclusion. PPAR-� and -� mRNA signaling pathways are
to the actions of insulin [3, 4]. Although several mecha-in place and active in mesangial cells. Both agonists affect thenisms of action for the insulin sensitizing effects of thephenotypic behavior of mesangial cells and ameliorate changes
resulting from hyperglycemia. The data indicate that the cor- thiazolidinediones have been proposed, it is currently be-rection of mesangial cell phenotype by troglitazone may influ- lieved that the thiazolidinediones exert their actions viaence production/deposition of a pathological fibrotic connec-
the activation one group of nuclear hormone receptorstive tissue matrix (that is, type I collagen) by these cells.known as peroxisomal proliferator activated receptor-�(PPAR-�) [5].
PPARs are members of the nuclear hormone receptorsuperfamily that include the retinoic acid and thyroidKey words: hyperglycemia, fibrosis, connective tissue matrix, glomeru-
losclerosis, diabetes, progressive renal disease. hormone receptors [reviewed in 3–5]. Besides PPAR-�,two other PPARs, -� and -�/�, have been identified. InReceived for publication September 28, 2000order to modulate gene transcription, PPARs must het-and in revised form November 15, 2001
Accepted for publication November 16, 2001 erodimerize with another member of the nuclear hor-mone receptor family, the retinoid X receptors (RXRs). 2002 by the International Society of Nephrology
1365

Routh et al: Troglitazone suppresses collagen secretion1366
PPAR/RXR heterodimers target DNA sequences known hyperglycemic conditions; (3) PPAR-� activation in mes-angial cells by clofibrate has different effects on matrixas PPAR response elements (PPREs) present in the pro-
moter regions of many genes, particularly ones involved production by mesangial cells in vitro; and (4) interven-tion with troglitazone may have a role in modulating thein the metabolic pathways associated with adipogenesis
[6–8]. Diversity in the pathway response is mediated by production/deposition of a pathological fibrotic connec-tive tissue matrix by mesangial cells.the ability of the RXR to heterodimerize with at least
three different PPAR family members. The PPAR par-ticipating in PPAR/RXR heterodimer formation and the
METHODSorganization of the PPRE confers specificity for modu-
Mesangial cell culturelating promoter activity of genes associated with thePPAR signaling pathway [9]. PPAR signaling can be Glomeruli were isolated from the kidneys of adult
Sprague-Dawley rats (Charles River, Raleigh, NC, USA)further modified by the actions of co-activators/repres-sors of PPAR/RXR activity [10, 11]. by sieving through a graded series of stainless steel sieves
according to published methods [21]. Isolated glomeruliPPAR ligands that move inactive PPARs to the activestate include natural activators such as fatty acids (oleic were grown in explant culture at 37�C, 10% CO2 in RPMI-
1640 media (BioWhitaker,Walkersville, MD, USA), 5and linoleic acids) and prostaglandins (15-deoxy-�12- and15-deoxy-�12,14 prostaglandin J2) [12, 13]. Pharmacologi- mmol/L d-glucose (Sigma, St. Louis, MO, USA), and
17% fetal bovine serum (FBS), 20 mL/liter penicillin-cal activators of these molecules are spread among sev-eral classes of drugs including the fibrates clofibrate and streptomycin (5000 IU/mL and 5000 �g/mL, respectively;
Mediatech, Herndon, VA, USA), 20 mL/L l-glutaminebenzofibrate (PPAR-� activators) [14, 15] and, as men-tioned above, the thiazolidinediones (PPAR-� activators). (200 mmol/L; Mediatech), insulin (5 �g/mL; Sigma),
transferrin (5�g/mL; Sigma), and sodium selenite (5 ng/At least one compound, the synthetic PPAR-� ligand,bisphenol A diglycidyl ether (or BADGE), has been mL), until outgrowth of mesangial cells was seen. The
mesangial cells were subsequently removed by trypsin-shown to antagonize PPAR-� activation by the thiazoli-dinediones in a dose-specific manner [16]. ization and reseeded into new flasks in the above medium
for further growth and expansion. The mesangial cellsSeveral recent studies, including one from our ownlaboratory, have shown that thiazolidinedione therapy were subsequently characterized by their stellate shape
and their ability to form hillocks in culture.can prevent the onset and progression of diabetic ne-phropathy in animal models of obesity and type II dia-
CMFDA staining for cell area measurementsbetes mellitus [17–19]. All show that a preventative regi-men (that is, therapeutic intervention at early onset of 5-Chloromethylfluorescein diacetate (CMFDA) stain-
ing (Molecular Probes, Eugene, OR, USA) was used todiabetes mellitus) with a thiazolidinedione was capableof preventing glomerulosclerosis, interstitial fibrosis, and label the cytoplasmic compartment of mesangial cells
grown in culture. CMFDA, a membrane permeable vitalproteinuria. These positive responses were coincidentwith the amelioration of the metabolic abnormalities of dye, is non-fluorescent until an ester group is enzymati-
cally cleaved by living cells, after which the dye becomeshyperglycemia, hypertriglyceridemia, and hypercholes-terolemia. The data from our study further showed that membrane impermeable and fluorescent at 494 nanome-
ter (a measure of wavelength). Using the extent ofthiazolidinedione intervention prevented the onset andprogression of mesangial expansion, that is, the abnor- CMFDA staining as a marker, spread cell area measure-
ments were made of mesangial cells grown in the pres-mal increase in the volume of the mesangium relativeto the volume of the glomerular tuft [20]. ence or absence of troglitazone (TRO-PPAR-� agonist;
Pfizer, Ann Arbor, MI, USA) or clofibrate (CLO-PPAR-�The results of our former study raise several questions.Was the inhibition of mesangial expansion in the thiazoli- agonist; C-6643; Sigma). Briefly, rat mesangial cells (pas-
sages 4 to 10) were seeded at a density of 1 � 103 cellsdinedione-treated animals due to the global correctionof metabolic defects, which, in turn, led to the correction per well into 24-well plates, with the wells coated prior
to cell seeding with type I collagen (Vitrogen; Cohesionof abnormal physiology in mesangial cells? Could thethiazolidinediones have direct effects in modulating mes- Technologies, Inc., Palo Alto, CA, USA) according to
the manufacturer’s instructions. Twenty-four hours afterangial cell behavior? If so, is the PPAR-� signaling path-way responsible in part for mediating these effects? Our seeding, the medium was exchanged and the wells orga-
nized into three medium conditions: 5 mmol/L glucose,current study shows that (1) rat mesangial cells are di-rectly capable of responding to PPAR-� and -� agonists 30 mmol/L glucose, or 5 mmol/L glucose plus 25 mmol/L
mannitol (osmotic control). Twenty-four hours after me-and possess the PPAR-� and -� signaling pathway thatallows mesangial cells to respond to PPAR-� and -� dium change, the cells grown in the three medium condi-
tions were further subdivided into triplicate treatmentactivation; (2) thiazolidinediones correct in part the ab-normal changes in mesangial cell behavior induced by groups of either 10 �mol/L TRO, or 100 �mol/L CLO.

Routh et al: Troglitazone suppresses collagen secretion 1367
A third group, also run in triplicate, of untreated (UNT) medium condition/treatment groups as described in thewells served as the vehicle control (0.01% DMSO section above. Triplicate samples from each medium con-added). These growth conditions and treatment groups dition (5 mmol/L, 30 mmol/L, and osmotic control) andwere maintained for all experiments. Twenty-four hours corresponding treatment group (UNT, TRO, and CLO)after treatment groups were established, cells were stained were run simultaneously during one assay protocol towith CMFDA (2 �L/well of 10 mmol/L CMFDA) for control for intra-assay differences that might be intro-30 minutes at 37�C. Afterwards, the CMFDA solution duced during the blotting protocol. Briefly, 200 �L ofwas replaced with fresh media containing the appropriate sample from each well of the above treatment groupsmedium conditions and treatments as described above. was diluted with 100 �L of phosphate-buffered salineOne hundred cells for each medium condition/treatment (PBS). Afterwards, the samples were added individuallygroup were imaged/digitized using an Olympus IX-70 in-
to the wells of a slot blot apparatus (Bio-Rad) that hadverted microscope that was equipped for epifluorescencebeen fitted with a 0.45 �m, pore size neutral nylon mem-illumination and digital image acquisition (Sensys�;brane (Biodyne A; Gelman Laboratory, Pall Corp., AnnRoper Instruments, Tucson, AZ, USA). To measure cellArbor, MI, USA). The samples were allowed to interactarea the planimetry subroutine in the image analysisfor an hour via gravity filtration before vacuum was ap-software of IP Lab Spectrum (Scanalytics, Fairfax, VA,plied to clear the wells. The membrane was then incu-USA) was used to draw a line encompassing the entirebated in blocking buffer (1 � PBS, 0.2% I-Block�, 0.1%perimeter of each cell and the spread cell areas calcu-Tween-20 detergent; Calbiochem, La Jolla, CA, USA)lated. Statistical analysis of the data was done using thefor 45 minutes, according to the membrane manufactur-analysis of variance (ANOVA) subroutine in the Stat-er’s instructions. For the immunoassay either a lamininview (SAS, Cary, NC, USA) software package, and sig-
nificant differences between the groups were determined primary antiserum (polyclonal antiserum against lami-using the Bonferroni Dunn test. nin-1, a routine reagent from our lab), or a type I collagen
monoclonal antibody (C-2456; Sigma) was added at aEffects of PPAR activation on protein production 1:5000 concentration. Both antibodies were diluted with
Rat mesangial cells (passages 4 through 10, medium blocking buffer and allowed to interact with the mem-conditions as described in the section on mesangial cell brane for 45 minutes. Afterwards, the membrane wasculture) were seeded onto 24-well plates at a density of washed two times for five minutes in wash buffer (PBS,5 � 104 cells per well to give an approximately 80% with 0.1% Tween-20 detergent), and then incubated inconfluent cell monolayer. Twenty-four hours after seed-
a species-specific secondary antibody-alkaline phospha-ing, the wells were subdivided into three groups of ninetase conjugate (Jackson Immunoresearch, Malverne,wells and the medium in each group was replaced withPA, USA) diluted in blocking buffer for 45 minutesserum-free medium containing one the following glucoseat a 1:40,000 dilution. Afterwards, the membrane wasconditions: 5 mmol/L glucose, 30 mmol/L glucose, or 5washed three times for five minutes in wash buffer fol-mmol/L glucose with 25 mmol/L mannitol. The wellslowed by 2 � 2-minute rinses in 1 � assay buffer (200within each medium condition were further subdividedmmol/L Tris, pH 9.8, 10 mmol/L MgCl2). The blot wasinto triplicate wells of the treatment groups: untreatedair-dried and then incubated in CDP-Star (Tropix; PE(UNT, vehicle control), or treated with 10 �mol/L TROBiosystems, Bedford, MA, USA) for five minutes, andor 100 �mol/L CLO, the drugs replenished every 24
hours. After 72 hours under the above medium/treat- the chemiluminescence subsequently imaged on filmment conditions, the medium from each well was de- (Eastman Kodak, Rochester, NY, USA). Because ofcanted, clarified, and stored at 80�C until further use. slight differences in the exposure time of the chemilumi-A bicinchoninic acid (BCA) protein assay (Pierce, Rock- nescence signal to the film, quantitative interassay com-ford, IL, USA) was used to quantify total protein in the parisons were not possible. The data shown in this reportmedium according to developer’s instructions, and the represent a single slot blot immunoassay consisting ofresults of the assay were measured by absorbance at 562 triplicate samples for each medium condition and treat-nm using a microplate reader (BioRad, Hercules, CA, ment group. Each set of experiments in this study wasUSA). Statistical analysis of the data was done using the
repeated for a total of three times. Images of the slotANOVA subroutine in the Statview software package,blots were digitized from films using a gel documentationand statisically significant differences between the groupssystem, and the density of the bands calculated using thewere determined using the Bonferroni Dunn test.densitometry subroutine in IP Lab Spectrum. Statistical
Solid phase immunoassay of extracellular analysis of the data was done using the ANOVA subrou-matrix molecules tine in the Statview software package, and statistically
significant differences between the groups were deter-A solid phase immunoassay was used to measure levelsof laminin and type I collagen produced by the different mined using the Bonferroni Dunn test.

Routh et al: Troglitazone suppresses collagen secretion1368
PPAR-� inhibitory assay PPAR-� and PPAR-� activation assayA dual-luciferase assay system (Promega, Madison, WI,To determine if the actions of troglitazone on mesan-
USA) was used to measure the changes in activation ofgial cell collagen secretion were mediated in part throughPPAR-� and -� in the presence of TRO, and CLO, andthe PPAR-� signaling pathway, BADGE, a syntheticif hyperglycemia affected PPRE activation in mesangialantagonist of PPAR-� activation [16], was used to blockcells. Briefly, rat mesangial cells grown in 5 mmol/L glu-its activation in mesangial cells. Dose response studiescose (passages 4 through 10) were seeded onto 24 wellindicated that treating mesangial cells with 50 �mol/Lplates at a density of 5 � 104 cells per well (�80% con-BADGE decreased the response of a PPRE-luciferasefluence). Twenty-four hours after seeding, the mediumreporter construct (see below) approximately 75% forconditions in triplicate groups of wells were switched tocells treated with troglitazone (inset, Fig. 5).either 5 mmol/L glucose, 30 mmol/L glucose, or the os-Rat mesangial cells were seeded in 24-well plates atmotic control, 5 mmol/L glucose with 25 mmol/L manni-a density of 8.0 � 104 cells/well. Twenty-four hours aftertol. Twenty-four hours later, triplicate wells within each
seeding the medium was changed to the following condi- medium condition were left untreated (UNT, vehicletions: 5 mmol/L glucose, 30 mmol/L glucose, or 5 mmol/L control as above) or treated with 10 �mol/L TRO orglucose with 25 mmol/L mannitol (total 18 wells/medium 100 �mol/L CLO. Twenty-four hours after treatment thegroup) and allowed to grow for an additional 24 hours. cells were used in transfection assays. All plasmids usedAfterwards one set of cell cultures (that is, 9 wells/ in this study were isolated with the Endofree Plasmidmedium group) was additionally treated with 50 �mol/L Maxi Purification� kit (Qiagen, Valencia, CA, USA).of BADGE while the other set served as a vehicle con- Plasmid tranfections were done using Effectene� (Qia-trol. Triplicate wells from each medium treatment group gen) according to the manufacturer’s instructions. Towere further treated with TRO, CLO, or UNT groups measure the extent of PPAR-� or -� activation in mesan-as described above. Twenty-four hours after BADGE gial cells, the cells were transfected with a firefly lucifer-
ase reporter plasmid bearing a PPRE capable of re-treatment the medium in the wells was exchanged forsponding to PPAR-� and -� activation (a gift from Toddserum-free medium and the respective treatment condi-Leff, Ph.D.; Pfizer/Parke-Davis Research). The PPREtions (glucose, BADGE, and drug) maintained as ini-reporter construct consisted of a TKpGL3 plasmid (Pro-tially described. BADGE and drug treatments weremega) with three PPRE sequence (5-TGACCTtTGTadded on a daily basis for a total exposure time of 72CCT-3) inserts upstream of the TK promoter. This PPREhours. Afterwards, the media were collected and colla-sequence corresponds to that found in the acyl CoAgen production was analyzed by slot blot immunoassayoxidase (ACO) promoter, and both PPAR-� and -� areas described above.able to bind to this PPRE, activating the firefly luciferasereporter. The cells were simultaneously co-transfectedReverse transcription-polymerase chain reactionwith a renilla luciferase-thymidine kinase plasmid (pRL-
Isolation of total mRNA from rat mesangial cells in cul- TK; Promega), which constitutively expresses the renillature was done using Trizol (Life Technologies, Gaithers- luciferase protein. Afterwards, dual luciferase assay wasburg, MD, USA) according to the manufacturer’s instruc- performed according to the developer’s instructions (Pro-tions. Samples were stored at 4�C prior to use. Reverse mega) to measure PPAR activation. Luciferase activitytranscription-polymerase chain reaction (RT-PCR) was was measured using a TD-20/20 luminometer (Turnerperformed using a commercially available kit (RETRO- Designs, Sunnyvale, CA, USA). The firefly luciferase data
were normalized to the luminescence data obtained fromscript kit; Ambion, Austin, TX, USA). The sequence ofthe co-transfected renilla luciferase to control for intra-primers used for PCR follows: PPAR-� (forward) 5-CTwell assay variability due to possible differences in cellTCGGAATCAGCTCTGTGGAC-3, (backward) 5-GCnumber, viability, or transfection efficiency (Promega).ATCCTTCACAAGCA TGGACTC-3, or (backward)The normalized data were expressed as the ratio of the5-ACCCTTGC ATCCTTCACAAGC-3, and PPAR-�firefly luciferase luminescence to the renilla luciferase lu-(forward) 5-TTTTGTGGGCTGGAGGGTTC-3, (back-minescence. Statistical analysis of the data was done usingward) 5-TCTCCGAGGGACTGAGAAATCTC-3, orthe ANOVA subroutine in the Statview software package(backward) 5-TCCTCTCCGAGGGACTGAGAAATC-3.and statistically significant differences between the groupsCycling profiles were as follows: 95�C for 5 minutes, 94�C were determined using the Bonferroni Dunn test.
for 20 seconds, 55�C for 30 seconds, 72�C for 40 seconds,and repeated 29 times. Appropriate controls consisted
RESULTSof PCR reactions with no RT product and PCR reactionsExpression of PPAR-� and -� in mesangial cellswith the RNA isolation product having no RT reactionusing RT-PCRperformed. Samples were electrophoresed on a 1% aga-
rose gel, at 50 volts for two hours, and stained with ethid- Reverse transcription-polymerase chain reaction wasused to determine whether rat mesangial cells containium bromide, and imaged on a gel documentation system.
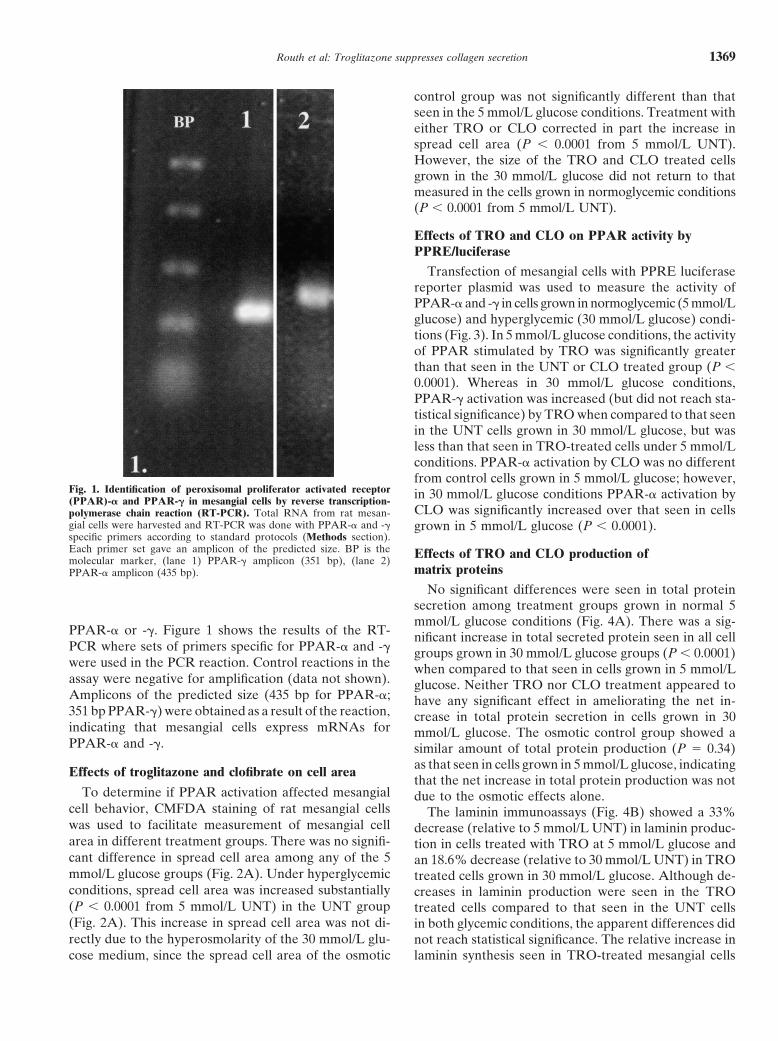
Routh et al: Troglitazone suppresses collagen secretion 1369
control group was not significantly different than thatseen in the 5 mmol/L glucose conditions. Treatment witheither TRO or CLO corrected in part the increase inspread cell area (P � 0.0001 from 5 mmol/L UNT).However, the size of the TRO and CLO treated cellsgrown in the 30 mmol/L glucose did not return to thatmeasured in the cells grown in normoglycemic conditions(P � 0.0001 from 5 mmol/L UNT).
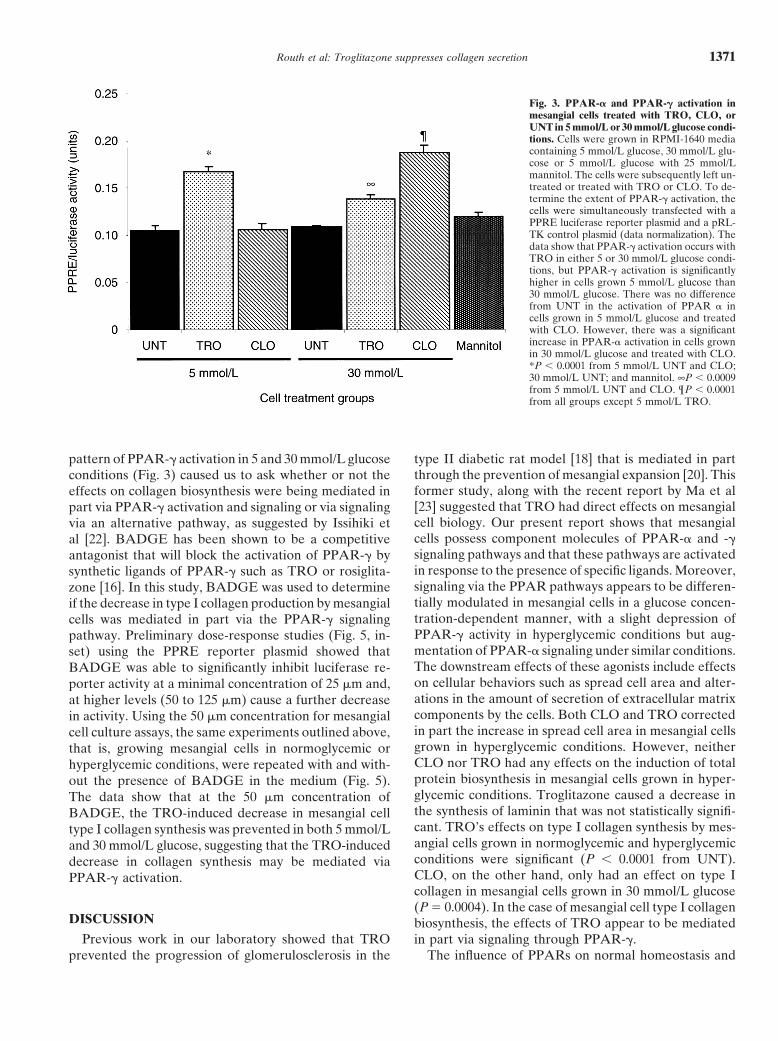
Effects of TRO and CLO on PPAR activity byPPRE/luciferase
Transfection of mesangial cells with PPRE luciferasereporter plasmid was used to measure the activity ofPPAR-� and -� in cells grown in normoglycemic (5 mmol/Lglucose) and hyperglycemic (30 mmol/L glucose) condi-tions (Fig. 3). In 5 mmol/L glucose conditions, the activityof PPAR stimulated by TRO was significantly greaterthan that seen in the UNT or CLO treated group (P �0.0001). Whereas in 30 mmol/L glucose conditions,PPAR-� activation was increased (but did not reach sta-tistical significance) by TRO when compared to that seenin the UNT cells grown in 30 mmol/L glucose, but wasless than that seen in TRO-treated cells under 5 mmol/Lconditions. PPAR-� activation by CLO was no differentfrom control cells grown in 5 mmol/L glucose; however,
Fig. 1. Identification of peroxisomal proliferator activated receptor in 30 mmol/L glucose conditions PPAR-� activation by(PPAR)-� and PPAR-� in mesangial cells by reverse transcription-CLO was significantly increased over that seen in cellspolymerase chain reaction (RT-PCR). Total RNA from rat mesan-
gial cells were harvested and RT-PCR was done with PPAR-� and -� grown in 5 mmol/L glucose (P � 0.0001).specific primers according to standard protocols (Methods section).Each primer set gave an amplicon of the predicted size. BP is the Effects of TRO and CLO production ofmolecular marker, (lane 1) PPAR-� amplicon (351 bp), (lane 2)
matrix proteinsPPAR-� amplicon (435 bp).
No significant differences were seen in total proteinsecretion among treatment groups grown in normal 5mmol/L glucose conditions (Fig. 4A). There was a sig-
PPAR-� or -�. Figure 1 shows the results of the RT- nificant increase in total secreted protein seen in all cellPCR where sets of primers specific for PPAR-� and -� groups grown in 30 mmol/L glucose groups (P � 0.0001)were used in the PCR reaction. Control reactions in the when compared to that seen in cells grown in 5 mmol/Lassay were negative for amplification (data not shown). glucose. Neither TRO nor CLO treatment appeared toAmplicons of the predicted size (435 bp for PPAR-�; have any significant effect in ameliorating the net in-351 bp PPAR-�) were obtained as a result of the reaction, crease in total protein secretion in cells grown in 30indicating that mesangial cells express mRNAs for mmol/L glucose. The osmotic control group showed aPPAR-� and -�. similar amount of total protein production (P � 0.34)
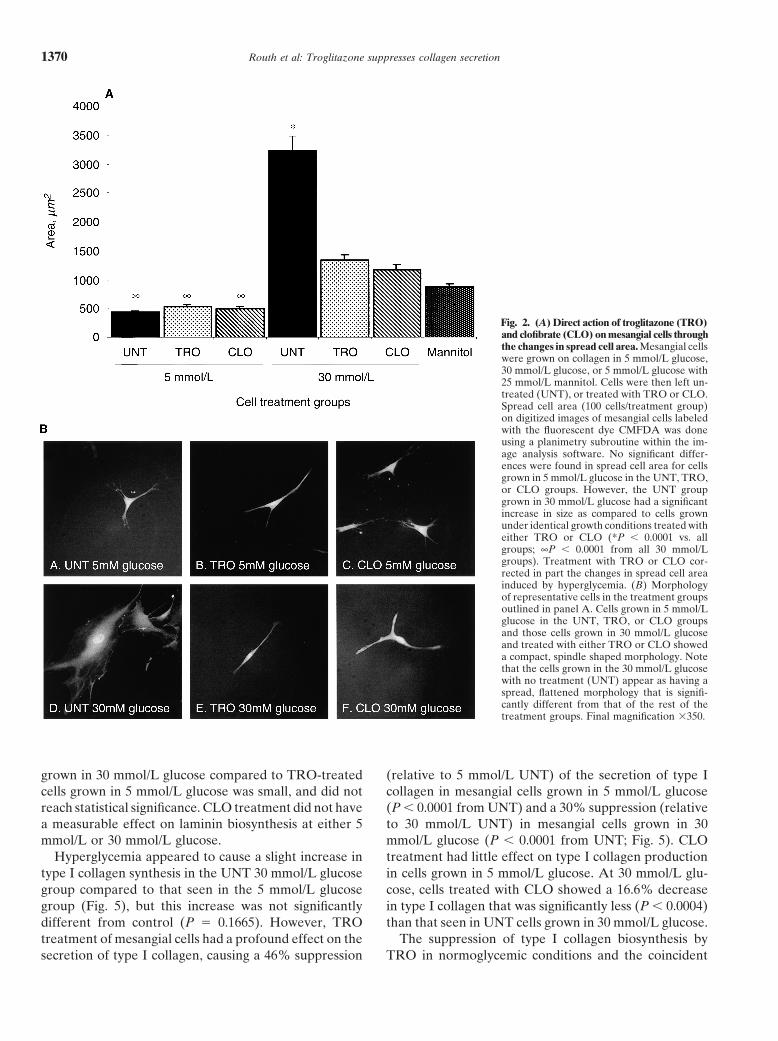
as that seen in cells grown in 5 mmol/L glucose, indicatingEffects of troglitazone and clofibrate on cell areathat the net increase in total protein production was not
To determine if PPAR activation affected mesangial due to the osmotic effects alone.cell behavior, CMFDA staining of rat mesangial cells The laminin immunoassays (Fig. 4B) showed a 33%was used to facilitate measurement of mesangial cell decrease (relative to 5 mmol/L UNT) in laminin produc-area in different treatment groups. There was no signifi- tion in cells treated with TRO at 5 mmol/L glucose andcant difference in spread cell area among any of the 5 an 18.6% decrease (relative to 30 mmol/L UNT) in TROmmol/L glucose groups (Fig. 2A). Under hyperglycemic treated cells grown in 30 mmol/L glucose. Although de-conditions, spread cell area was increased substantially creases in laminin production were seen in the TRO(P � 0.0001 from 5 mmol/L UNT) in the UNT group treated cells compared to that seen in the UNT cells(Fig. 2A). This increase in spread cell area was not di- in both glycemic conditions, the apparent differences didrectly due to the hyperosmolarity of the 30 mmol/L glu- not reach statistical significance. The relative increase in
laminin synthesis seen in TRO-treated mesangial cellscose medium, since the spread cell area of the osmotic
Routh et al: Troglitazone suppresses collagen secretion1370
Fig. 2. (A) Direct action of troglitazone (TRO)and clofibrate (CLO) on mesangial cells throughthe changes in spread cell area. Mesangial cellswere grown on collagen in 5 mmol/L glucose,30 mmol/L glucose, or 5 mmol/L glucose with25 mmol/L mannitol. Cells were then left un-treated (UNT), or treated with TRO or CLO.Spread cell area (100 cells/treatment group)on digitized images of mesangial cells labeledwith the fluorescent dye CMFDA was doneusing a planimetry subroutine within the im-age analysis software. No significant differ-ences were found in spread cell area for cellsgrown in 5 mmol/L glucose in the UNT, TRO,or CLO groups. However, the UNT groupgrown in 30 mmol/L glucose had a significantincrease in size as compared to cells grownunder identical growth conditions treated witheither TRO or CLO (*P � 0.0001 vs. allgroups; ∞P � 0.0001 from all 30 mmol/Lgroups). Treatment with TRO or CLO cor-rected in part the changes in spread cell areainduced by hyperglycemia. (B) Morphologyof representative cells in the treatment groupsoutlined in panel A. Cells grown in 5 mmol/Lglucose in the UNT, TRO, or CLO groupsand those cells grown in 30 mmol/L glucoseand treated with either TRO or CLO showeda compact, spindle shaped morphology. Notethat the cells grown in the 30 mmol/L glucosewith no treatment (UNT) appear as having aspread, flattened morphology that is signifi-cantly different from that of the rest of thetreatment groups. Final magnification �350.
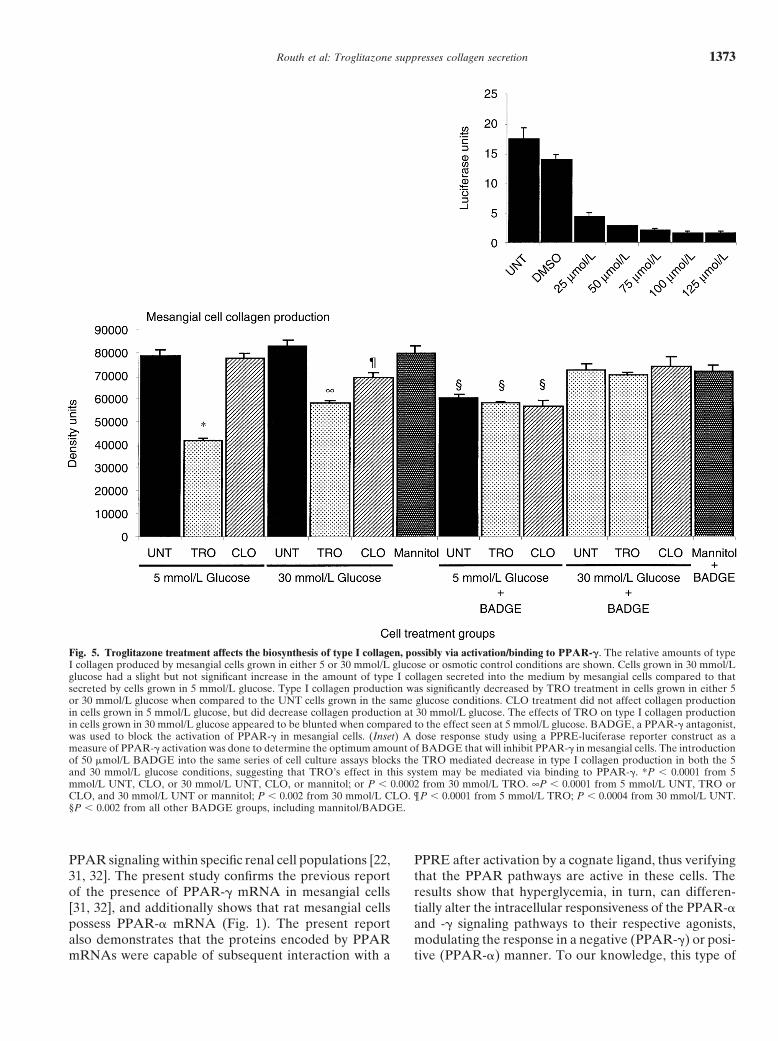
grown in 30 mmol/L glucose compared to TRO-treated (relative to 5 mmol/L UNT) of the secretion of type Icollagen in mesangial cells grown in 5 mmol/L glucosecells grown in 5 mmol/L glucose was small, and did not
reach statistical significance. CLO treatment did not have (P � 0.0001 from UNT) and a 30% suppression (relativeto 30 mmol/L UNT) in mesangial cells grown in 30a measurable effect on laminin biosynthesis at either 5
mmol/L or 30 mmol/L glucose. mmol/L glucose (P � 0.0001 from UNT; Fig. 5). CLOtreatment had little effect on type I collagen productionHyperglycemia appeared to cause a slight increase in
type I collagen synthesis in the UNT 30 mmol/L glucose in cells grown in 5 mmol/L glucose. At 30 mmol/L glu-cose, cells treated with CLO showed a 16.6% decreasegroup compared to that seen in the 5 mmol/L glucose
group (Fig. 5), but this increase was not significantly in type I collagen that was significantly less (P � 0.0004)than that seen in UNT cells grown in 30 mmol/L glucose.different from control (P � 0.1665). However, TRO
treatment of mesangial cells had a profound effect on the The suppression of type I collagen biosynthesis byTRO in normoglycemic conditions and the coincidentsecretion of type I collagen, causing a 46% suppression
Routh et al: Troglitazone suppresses collagen secretion 1371
Fig. 3. PPAR-� and PPAR-� activation inmesangial cells treated with TRO, CLO, orUNT in 5 mmol/L or 30 mmol/L glucose condi-tions. Cells were grown in RPMI-1640 mediacontaining 5 mmol/L glucose, 30 mmol/L glu-cose or 5 mmol/L glucose with 25 mmol/Lmannitol. The cells were subsequently left un-treated or treated with TRO or CLO. To de-termine the extent of PPAR-� activation, thecells were simultaneously transfected with aPPRE luciferase reporter plasmid and a pRL-TK control plasmid (data normalization). Thedata show that PPAR-� activation occurs withTRO in either 5 or 30 mmol/L glucose condi-tions, but PPAR-� activation is significantlyhigher in cells grown 5 mmol/L glucose than30 mmol/L glucose. There was no differencefrom UNT in the activation of PPAR � incells grown in 5 mmol/L glucose and treatedwith CLO. However, there was a significantincrease in PPAR-� activation in cells grownin 30 mmol/L glucose and treated with CLO.*P � 0.0001 from 5 mmol/L UNT and CLO;30 mmol/L UNT; and mannitol. ∞P � 0.0009from 5 mmol/L UNT and CLO. ¶P � 0.0001from all groups except 5 mmol/L TRO.
pattern of PPAR-� activation in 5 and 30 mmol/L glucose type II diabetic rat model [18] that is mediated in partthrough the prevention of mesangial expansion [20]. Thisconditions (Fig. 3) caused us to ask whether or not theformer study, along with the recent report by Ma et aleffects on collagen biosynthesis were being mediated in[23] suggested that TRO had direct effects on mesangialpart via PPAR-� activation and signaling or via signalingcell biology. Our present report shows that mesangialvia an alternative pathway, as suggested by Issihiki etcells possess component molecules of PPAR-� and -�al [22]. BADGE has been shown to be a competitivesignaling pathways and that these pathways are activatedantagonist that will block the activation of PPAR-� byin response to the presence of specific ligands. Moreover,synthetic ligands of PPAR-� such as TRO or rosiglita-signaling via the PPAR pathways appears to be differen-zone [16]. In this study, BADGE was used to determinetially modulated in mesangial cells in a glucose concen-if the decrease in type I collagen production by mesangialtration-dependent manner, with a slight depression ofcells was mediated in part via the PPAR-� signalingPPAR-� activity in hyperglycemic conditions but aug-pathway. Preliminary dose-response studies (Fig. 5, in-mentation of PPAR-� signaling under similar conditions.set) using the PPRE reporter plasmid showed thatThe downstream effects of these agonists include effectsBADGE was able to significantly inhibit luciferase re-on cellular behaviors such as spread cell area and alter-porter activity at a minimal concentration of 25 �m and,ations in the amount of secretion of extracellular matrixat higher levels (50 to 125 �m) cause a further decreasecomponents by the cells. Both CLO and TRO correctedin activity. Using the 50 �m concentration for mesangialin part the increase in spread cell area in mesangial cellscell culture assays, the same experiments outlined above,grown in hyperglycemic conditions. However, neitherthat is, growing mesangial cells in normoglycemic orCLO nor TRO had any effects on the induction of totalhyperglycemic conditions, were repeated with and with-protein biosynthesis in mesangial cells grown in hyper-out the presence of BADGE in the medium (Fig. 5).glycemic conditions. Troglitazone caused a decrease inThe data show that at the 50 �m concentration ofthe synthesis of laminin that was not statistically signifi-BADGE, the TRO-induced decrease in mesangial cellcant. TRO’s effects on type I collagen synthesis by mes-type I collagen synthesis was prevented in both 5 mmol/Langial cells grown in normoglycemic and hyperglycemicand 30 mmol/L glucose, suggesting that the TRO-inducedconditions were significant (P � 0.0001 from UNT).decrease in collagen synthesis may be mediated viaCLO, on the other hand, only had an effect on type IPPAR-� activation.collagen in mesangial cells grown in 30 mmol/L glucose(P � 0.0004). In the case of mesangial cell type I collagen
DISCUSSION biosynthesis, the effects of TRO appear to be mediatedPrevious work in our laboratory showed that TRO in part via signaling through PPAR-�.
The influence of PPARs on normal homeostasis andprevented the progression of glomerulosclerosis in the
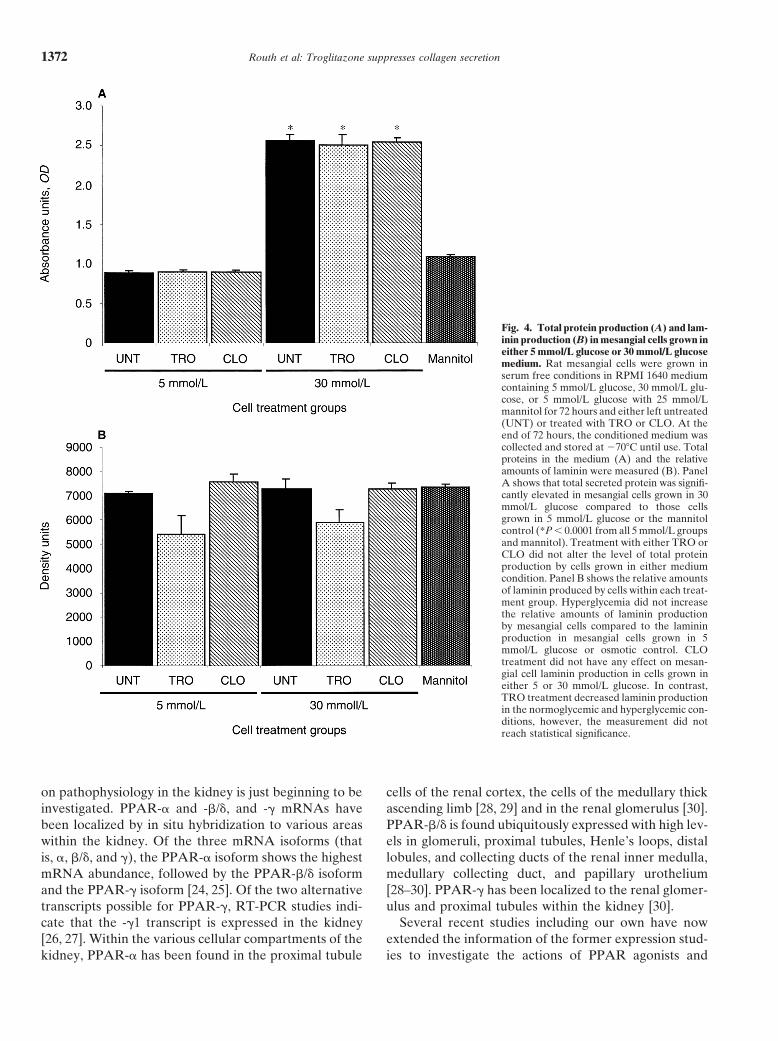
Routh et al: Troglitazone suppresses collagen secretion1372
Fig. 4. Total protein production (A) and lam-inin production (B) in mesangial cells grown ineither 5 mmol/L glucose or 30 mmol/L glucosemedium. Rat mesangial cells were grown inserum free conditions in RPMI 1640 mediumcontaining 5 mmol/L glucose, 30 mmol/L glu-cose, or 5 mmol/L glucose with 25 mmol/Lmannitol for 72 hours and either left untreated(UNT) or treated with TRO or CLO. At theend of 72 hours, the conditioned medium wascollected and stored at 70�C until use. Totalproteins in the medium (A) and the relativeamounts of laminin were measured (B). PanelA shows that total secreted protein was signifi-cantly elevated in mesangial cells grown in 30mmol/L glucose compared to those cellsgrown in 5 mmol/L glucose or the mannitolcontrol (*P � 0.0001 from all 5 mmol/L groupsand mannitol). Treatment with either TRO orCLO did not alter the level of total proteinproduction by cells grown in either mediumcondition. Panel B shows the relative amountsof laminin produced by cells within each treat-ment group. Hyperglycemia did not increasethe relative amounts of laminin productionby mesangial cells compared to the lamininproduction in mesangial cells grown in 5mmol/L glucose or osmotic control. CLOtreatment did not have any effect on mesan-gial cell laminin production in cells grown ineither 5 or 30 mmol/L glucose. In contrast,TRO treatment decreased laminin productionin the normoglycemic and hyperglycemic con-ditions, however, the measurement did notreach statistical significance.
on pathophysiology in the kidney is just beginning to be cells of the renal cortex, the cells of the medullary thickascending limb [28, 29] and in the renal glomerulus [30].investigated. PPAR-� and -�/�, and -� mRNAs have
been localized by in situ hybridization to various areas PPAR-�/� is found ubiquitously expressed with high lev-els in glomeruli, proximal tubules, Henle’s loops, distalwithin the kidney. Of the three mRNA isoforms (that
is, �, �/�, and �), the PPAR-� isoform shows the highest lobules, and collecting ducts of the renal inner medulla,medullary collecting duct, and papillary urotheliummRNA abundance, followed by the PPAR-�/� isoform
and the PPAR-� isoform [24, 25]. Of the two alternative [28–30]. PPAR-� has been localized to the renal glomer-ulus and proximal tubules within the kidney [30].transcripts possible for PPAR-�, RT-PCR studies indi-
cate that the -�1 transcript is expressed in the kidney Several recent studies including our own have nowextended the information of the former expression stud-[26, 27]. Within the various cellular compartments of the
kidney, PPAR-� has been found in the proximal tubule ies to investigate the actions of PPAR agonists and
Routh et al: Troglitazone suppresses collagen secretion 1373
Fig. 5. Troglitazone treatment affects the biosynthesis of type I collagen, possibly via activation/binding to PPAR-�. The relative amounts of typeI collagen produced by mesangial cells grown in either 5 or 30 mmol/L glucose or osmotic control conditions are shown. Cells grown in 30 mmol/Lglucose had a slight but not significant increase in the amount of type I collagen secreted into the medium by mesangial cells compared to thatsecreted by cells grown in 5 mmol/L glucose. Type I collagen production was significantly decreased by TRO treatment in cells grown in either 5or 30 mmol/L glucose when compared to the UNT cells grown in the same glucose conditions. CLO treatment did not affect collagen productionin cells grown in 5 mmol/L glucose, but did decrease collagen production at 30 mmol/L glucose. The effects of TRO on type I collagen productionin cells grown in 30 mmol/L glucose appeared to be blunted when compared to the effect seen at 5 mmol/L glucose. BADGE, a PPAR-� antagonist,was used to block the activation of PPAR-� in mesangial cells. (Inset) A dose response study using a PPRE-luciferase reporter construct as ameasure of PPAR-� activation was done to determine the optimum amount of BADGE that will inhibit PPAR-� in mesangial cells. The introductionof 50 �mol/L BADGE into the same series of cell culture assays blocks the TRO mediated decrease in type I collagen production in both the 5and 30 mmol/L glucose conditions, suggesting that TRO’s effect in this system may be mediated via binding to PPAR-�. *P � 0.0001 from 5mmol/L UNT, CLO, or 30 mmol/L UNT, CLO, or mannitol; or P � 0.0002 from 30 mmol/L TRO. ∞P � 0.0001 from 5 mmol/L UNT, TRO orCLO, and 30 mmol/L UNT or mannitol; P � 0.002 from 30 mmol/L CLO. ¶P � 0.0001 from 5 mmol/L TRO; P � 0.0004 from 30 mmol/L UNT.§P � 0.002 from all other BADGE groups, including mannitol/BADGE.
PPAR signaling within specific renal cell populations [22, PPRE after activation by a cognate ligand, thus verifyingthat the PPAR pathways are active in these cells. The31, 32]. The present study confirms the previous report
of the presence of PPAR-� mRNA in mesangial cells results show that hyperglycemia, in turn, can differen-tially alter the intracellular responsiveness of the PPAR-�[31, 32], and additionally shows that rat mesangial cells
possess PPAR-� mRNA (Fig. 1). The present report and -� signaling pathways to their respective agonists,modulating the response in a negative (PPAR-�) or posi-also demonstrates that the proteins encoded by PPAR
mRNAs were capable of subsequent interaction with a tive (PPAR-�) manner. To our knowledge, this type of

Routh et al: Troglitazone suppresses collagen secretion1374
modulation of PPAR signaling has not been previously by mesangial cells in culture than the PPAR-� agonistCLO. Clofibrate had no effect on laminin synthesis byreported in mesangial cells. The activation of PPAR-�
by TRO in 30 mmol/L glucose was decreased compared mesangial cells and only a modest effect on type I colla-gen biosynthesis in cells grown in 30 mmol/L glucose.to the 5 mmol/L glucose group, but was still higher than
the extent of activation seen in either the 5 mmol/L UNT The results of our TRO assays on laminin synthesis docorrelate in part with the findings of Isshiki et al, who(P � 0.0007, significant) or 30 mmol/L UNT (P � 0.0018,
did not reach significance) groups. In contrast, PPAR-� showed that glomeruli isolated from diabetic animalsthat had been treated with TRO had a significant de-activation by CLO was significantly increased (P �
0.0001) in 30 mmol/L glucose compared to 5 mmol/L crease in fibronectin and type IV collagen, both at themRNA and protein levels [22]. However, they did notglucose. The negative modulation of PPAR-� by hyper-
glycemia may be of significance because it implies that report a significant suppression of either fibronectin ortype IV collagen secretion in glomeruli isolated frombeneficial effects of the thiazolidinediones may be
blunted in individuals whose hyperglycemia in not en- TRO-treated control rats. The lack of effect of PPAR-�agonists on extracellular matrix (ECM) synthesis by cellstirely controlled. The mechanism by which these particu-
lar effects are differentially modulated by hyperglycemia has been observed in other cell systems [53]. However,the decrease in type I collagen synthesis by CLO inis currently unknown and an area of active investigation.
It is well known that hyperglycemia associated with mesangial cells grown in 30 mmol/L glucose is novel,and suggests that the fibrate compounds may have somediabetes mellitus has profound effects on the biosynthe-
sis of extracellular matrix components within the glomer- effect in ameliorating the progression of glomeruloscle-rosis. Further studies must be done to discern the mecha-ulus [33–38]. Common to many studies is the observation
of a net increase in extracellular matrix mRNA expres- nism by which this effect is mediated and its overalleffect on the progression of glomerulosclerosis.sion and protein biosynthesis by mesangial cells in hyper-
glycemic conditions, those observations providing a par- Troglitazone significantly suppresses the biosynthesis/secretion of type I collagen by mesangial cells grown intial explanation for the accumulation of extracellular
matrix within the mesangial matrix of individuals af- 5 mmol/L and 30 mmol/L glucose, albeit an increase seenin type I collagen production in TRO-treated cells grownflicted with diabetes. However, the effects of hyperglyce-
mia on other aspects matrix protein biosynthesis/modifi- in 30 mmol/L glucose (P � 0.0002) was seen comparedto the TRO cells grown in 5 mmol/L glucose. Down-cation patterns, such as changes in the extent of sulfation
of proteoglycans [39–43], changes in the pattern of ex- regulation of collagen production was seen in the pancre-atic islets of TRO-treated ZDF/Gmi�fa rats in our previ-pression of laminin isoforms [44], or the abnormal ex-
pression of interstitial matrix components (see below) ous report [18]. How the thiazolidinediones affect colla-gen biosynthesis remains unclear. The report by Isshiki etmay eventually be proven as mediating functionally pro-
found deficits on the homeostasis of the glomerulus. Both al suggests that some of the effects of TRO on glomerularECM production might be mediated via a negative regu-this present report and the work of Isshiki et al [22]
indicate that TRO may act by decreasing mRNA or latory effect on the DAG-PKC-ERK pathway [22]. How-ever, our studies indicate that PPAR-� activation mayprotein synthesis of specific basement membrane compo-
nents by the renal glomerulus proper or, more specifi- play a role in modulating the secretion of type I collagenby mesangial cells. The extent of the PPAR-� activationcally, mesangial cells.
In vivo the predominant extracellular matrix compo- in mesangial cells in our luciferase studies is correlativewith the degree of suppression of collagen synthesis seennents secreted by mesangial cells consist of type IV colla-
gen, laminin, fibronectin, and proteoglycans [45, 46]. in mesangial cells grown in the same medium conditions.The complete loss of the type I collagen suppressionHowever, under pathophysiological conditions the mes-
angial cells are stimulated to abnormally initiate de novo induced by TRO using the PPAR-� antagonist, BADGE(Fig. 5) [16], add further support to the fact PPAR-�synthesis of types I and III collagens, [45, 47, 48]. These
abnormally secreted fibrillar collagens are routinely activation as mediating the type I collagen suppression.In addition, similar results of thiazolidinedione-inducedfound in sclerotic but not normal glomeruli [49]. When
mesangial cells are isolated and grown in culture, they suppression of type I collagen synthesis, and the antago-nism of that suppression by an entirely different PPARnot only secrete the former compliment of basement
membrane components in a constitutive manner, but also antagonist, has been recently reported in a study focusingon the effects of PPARs in hepatic stellate cells [53].initiate and maintain the production of type I collagen in
culture [50, 51]. In turn, hyperglycemia has been shown Taken together, this information suggests that TRO maybe exerting its effects on type I collagen biosynthesis viato cause an up-regulation of type I collagen synthesis in
mesangial cells grown in culture [52]. the activation of PPAR-�. However, to the best of ourknowledge the promoter region for type I collagen hasOur results show that TRO has a greater effect on the
biosynthesis of both laminin and type I collagen secretion not been reported to have a DR-1 region that would

Routh et al: Troglitazone suppresses collagen secretion 1375
permit PPAR-�/RXR heterodimer binding. This sug- REFERENCESgests that it is likely that another mechanism of action 1. Molitch ME, Steffes MW, Cleary PA, Nathan DM: Baseline
analysis of renal function in the Diabetes Control and Complica-or alternative gene activation pathway for activatedtions Trial. The Diabetes Control and Complications Trial Re-PPAR-� must be in place to mediate these significantsearch Group (corrected). Kidney Int 43:668–674, 1993
changes in collagen production. One possibility may be 2. Zimmerman BR: Sulfonylureas. Endocrinol Metab Clin North Am26:511–522, 1997an effect of TRO/PPAR-� activation on the global mod-
3. Willson TM, Lambert MH, Kliewer SA: Peroxisome prolifera-ulation/differentiation of these cells, moving them fromtor-receptor � and metabolic disease. Ann Rev Biochem 70:341–367,
an activated phenotype (matrix over-secreting) to a qui- 20014. Bar-Tana J: Peroxisome proliferator-activated receptor gammaescent phenotype as suggested by Asano et al [31]. Inves-
(PPARgamma) activation and its consequences in humans. Toxicoltigations focusing upon the effect of PPAR-� agonistsLett 120:9–19, 2001
on vascular smooth muscle migration and proliferation 5. Bishop-Bailey D: Peroxisome proliferator-activated receptors inthe cardiovascular system. Br J Pharmacol 129:823–834, 2000lend support to Asano et al’s premise [54–57].
6. Dreyer C, Krey G, Keller H, et al: Control of the peroxisomalIn summary, TRO has a direct effect on mesangialoxidation pathway by a novel family of nuclear hormone receptors.
cell behaviors under normal and pathological conditions. Cell 68:879–887, 19927. Kliewer SA, Umesono K, Noonan DJ, et al: Convergence ofTRO inhibits an abnormal increase in mesangial spread
9-cis retinoic acid and peroxisome proliferator signalling pathwayscell area under hyperglycemic conditions. Further workthrough heterodimer formation of their receptors. Nature 358:771–
is required to precisely elucidate the mechanism by which 774, 19928. Tugwood JD, Issemann I, Anderson RG, et al: The mouse peroxi-TRO has specific effects on the biosynthesis of one ma-
some proliferator activated receptor recognizes a response elementtrix component verses another. It is likely that corepres-in the 5 flanking sequence of the rat acyl CoA oxidase gene.
sors/co-activators of PPARs may play a role in causing EMBO J 11:433–439, 19929. Schoonjans K, Staels B, Auwerx J: Role of the peroxisome pro-some of these differential effects, possibly by preventing
liferator-activated receptor (PPAR) in mediating the effects of fi-the differentiation of the mesangial cell into an activatedbrates and fatty acids on gene expression. J Lipid Res 37:907–925,
phenotype. Research in this direction may lead to a 199610. McInerney EM, Rose DW, Flynn SE, et al: Determinants ofgreater understanding and final prevention of glomerulo-
coactivator LXXLL motif specificity in nuclear receptor transcrip-sclerosis in the diabetic patient, and other disease states tional activation. Genes Dev 12:3357–3368, 1998that result in glomerulosclerosis and end-stage renal dis- 11. DiRenzo J, Soderstrom M, Kurokawa R, et al: Peroxisome prolif-
erator-activated receptors and retinoic acid receptors differentiallyease.control the interactions of retinoid X receptor heterodimers withligands, coactivators, and corepressors. Mol Cell Biol 17:2166–2176, 1997NOTE ADDED IN PROOF 12. Kliewer SA, Lenhard JM, Wilson TM, et al: A prostaglandin J2metabolite binds peroxisome proliferator-activated receptor gammaDuring the review of this manuscript, a third reportand promotes adipocyte differentiation. Cell 83:813–819, 1995(Nicholas SB, Kawano Y, Wakino S, et al, Hypertension 13. Kliewer SA, Sundseth SS, Jones SA: Fatty acids and eicosanoids
37:722–727, 2001) described the presence of PPAR-� in regulate gene expression through direct interactions with peroxi-some proliferator-activated receptors alpha and gamma. Proc Natlmesangial cells. This report also corroborates the obser-Acad Sci USA 94:4318–4323, 1997vances of Fujii et al (Metabolism 46:981–983) showing 14. Ogawa S, Takeuchi K, Sugimura K: Bezafibrate reduces blood
that administration of thiazolidinediones in the STZ rat glucose in type 2 diabetes mellitus. Metabolism 49:331–334, 200015. Forman BM, Chen J, Evans RM: Hypolipidemic drugs, polyunsat-model of type I diabetes ameliorates the albuminuria
urated fatty acids, and eicosanoids are ligands for peroxisome pro-that develops in that model. The study also reports the liferator-activated receptors alpha and delta. Proc Natl Acad Sciinhibition of platelet-derived growth factor (PDGF)- USA 94:4312–4317, 1997
16. Wright HM, Clish CB, Mikami T, et al: A synthetic antagonistinduced mesangial cell proliferation by PPAR-� agonistsfor the peroxisome proliferator-activated receptor gamma inhibitsand the reduction of Ang II-induced expression of PA-I. adipocyte differentiation. J Biol Chem 275:1873–1877, 2000
In addition, treatment of STZ-diabetic animals with 17. Smith SA, Lister CA, Toseland CD, Buckingham RE: Rosiglita-zone prevents the onset of hyperglycaemia and proteinuria in thePPAR agonists decreased the expression of PAI-I toZucker diabetic fatty rat. Diabetes Obes Metab 2:363–372, 2000normal levels in diabetic animals. 18. McCarthy KJ, Routh RE, Shaw W, et al: Troglitazone halts dia-betic glomerulosclerosis by blockade of mesangial expansion. Kid-ney Int 58:2341–2350, 2000ACKNOWLEDGMENTS
19. Buckingham RE, Al-Barazanji KA, Toseland CD, et al: Peroxi-Work on this project was supported by a Research Grant from the some proliferator-activated receptor-gamma agonist, rosiglitazone,
American Diabetes Association (KJM), grant #1-RO1-DK48055 from protects against nephropathy and pancreatic islet abnormalities inNIH/NIDDK (KJM), and a Research Fellowship from the Louisiana Zucker fatty rats. Diabetes 47:1326–1334, 1998Affiliate of the American Heart Association (RER). These data were 20. Steffes MW, Osterby R, Chavers B, Mauer SM: Mesangialsubmitted in partial fulfillment of a Ph.D. degree (RER) at LSU Health expansion as a central mechanism for loss of kidney function inSciences Center, Shreveport, LA, USA. diabetic patients. Diabetes 38:1077–1081, 1989
21. Mistra RP: Isolation of glomeruli from mammalian kidneys bygraded sieving. Am J Clin Pathol 58:135–139, 1972Reprint requests to Kevin J. McCarthy, Ph.D., Department of Pathol-
ogy, Louisiana State University Health Sciences Center, 1501 King’s 22. Isshiki K, Haneda M, Koya D, et al: Thiazolidinedione compoundsameliorate glomerular dysfunction independent of their insulin-Highway, Shreveport, Louisiana 71130-3932, USA.
E-mail: [email protected] sensitizing action in diabetic rats. Diabetes 49:1022–1032, 2000

Routh et al: Troglitazone suppresses collagen secretion1376
23. Ma LJ, Marcantoni C, Linton MF, et al: Peroxisome proliferator- of [35S]-labeled glomerular basement membrane. Diabetes 33:8–12, 1984activated receptor-gamma agonist troglitazone protects against non-
41. Cohen MP, Klepser H, Van-Yu W: Undersulfation of glomerulardiabetic glomerulosclerosis in rats. Kidney Int 59:1899–1910, 2001basement membrane heparan sulfate in experimental diabetes and24. Cullingford TE, Bakhoo K, Peuchen S, et al: Distribution oflack of correction with aldose reductase inhibition. DiabetesmRNAs encoding the peroxisome proliferator-activated receptor37:1324–1327, 1988alpha, beta, and gamma and the retinoid X receptor alpha, beta,
42. Kanwar YS, Rosenweig CJ, Linker A, Jakubowski ML: De-and gamma in rat central nervous system. J Neurochem 70:1366–creased de novo synthesis of glomerular proteoglycans in diabetes:1375, 1998Biochemical and autoradiographic evidence. Proc Natl Acad Sci25. Kliewer SA, Forman BM, Blumberg B, et al: Differential expres-USA 80:2272–2275, 1983sion and activation of a family of murine peroxisome proliferator-
43. van den Born, van Kraats AA, Bakker MAH, et al: Reductionactivated receptors. Proc Natl Acad Sci USA 91:7355–7359, 1994of heparan sulphate-associated anionic sites in the glomerular base-26. Auboeuf D, Rieussert J, Fajas L, et al: Tissue distribution andment membrane of rats with streptozotocin-induced diabetic ne-quantification of the expression of mRNAs of peroxisome prolifer-phropathy. Diabetologia 38:1169–1175, 1995ator-activated receptors and liver X receptor-alpha in humans: no
44. Ha TS, Barnes JL, Stewart JL, et al: Regulation of renal lamininalteration in adipose tissue of obese and NIDDM patients. Diabetesin mice with type II diabetes. J Am Soc Nephrol 10:1931–1939, 199946:1319–1327, 1997
45. Rupprecht HD, Schocklmann HO, Sterzel RB: Cell-matrix27. Fajas L, Auboeuf D, Raspe E, et al: The organization, promoterinteractions in the glomerular mesangium. Kidney Int 49:1575–analysis, and expression of the human PPARgamma gene. J Biol1582, 1996Chem 272:18779–18789, 1997
46. McCarthy KJ: Development of the glomerular basement mem-28. Guan Y, Zhang Y, Davis L, Breyer MD: Expression of peroxi-brane and mesangial matrix: Asynchronous maturation of glomeru-some proliferator-activated receptors in urinary tract of rabbitslar extracellular matrices. Microsc Res Tech 39:231–253, 1997and humans. Am J Physiol 273:F1013–F1022, 1997 47. Savill J, Johnson RJ: Glomerular remodelling after inflammatory29. Yang T, Michele DE, Park I, et al: Expression of peroxisomal injury. (editorial) Exp Nephrol 3:149–158, 1995proliferator-activated receptors and retinoid X receptors in the 48. Johnson RJ: The glomerular response to injury: Progression or
kidney. Am J Physiol 277(6 Pt 2):F966–F973, 1999 resolution? (review) Kidney Int 45:1769–1782, 199430. Braissant O, Foufelle F, Scotto C, et al: Differential expression 49. Glick AD, Jacobson HR, Haralson MA: Mesangial deposition
of peroxisome proliferator-activated receptors (PPARs): Tissue of type I collagen in human glomerulosclerosis. Hum Pathol 23:distribution of PPAR-alpha, -beta, and -gamma in the adult rat. 1373–1379, 1992Endocrinology 137:354–366, 1996 50. Haralson MA, Jacobson HR, Hoover RL: Collagen polymor-
31. Asano T, Wakisaka M, Yoshinari M, et al: Peroxisome prolifera- phism in cultured rat kidney mesangial cells. Lab Invest 57:513–tor-activated receptor gamma1 (PPARgamma1) expresses in rat 523, 1987mesangial cells and PPARgamma agonists modulate its differentia- 51. Ohyama K, Seyer JM, Raghow R, Kang AH: Extracellular matrixtion. Biochem Biophys Acta 1497:148–154, 2000 phenotype of rat mesangial cells in culture. Biosynthesis of collagen
32. Iwashima Y, Eto M, Horiuchi S, Sano H: Advanced glycation types I, III, IV, and V and a low molecular weight collagenouscomponent and their regulation by dexamethasone. J Lab Clinend product-induced peroxisome proliferator-activated receptorMed 116:219–227, 1990gamma gene expression in the cultured mesangial cells. Biochem
52. Ziyadeh FN, Sharma K, Ericksen M, Wolf G: Stimulation ofBiophys Res Commun 264:441–448, 1999collagen gene expression and protein synthesis in murine mesangial33. Østerby R: Lessons from kidney biopsies. Diabetes/Metabolismcells by high glucose is mediated by autocrine activation of trans-Reviews 12:151–174, 1996forming growth factor-�. J Clin Invest 93:536–542, 199434. Adler S: Structure-function relationships in diabetic nephropathy:
53. Miyahara T, Schrum L, Rippe R, et al: Peroxisome proliferator-lessons and limitations. Kidney Int 52(Suppl 60):S42–S45, 1997activated receptors and hepatic stellate cell activation. J Biol Chem35. Sharma K, Ziyadeh FN: Hyperglycemia and diabetic kidney dis-275:35715–35722, 2000ease. The case for transforming growth factor-beta as a key media-
54. Benson S, Padmanabhan S, Kurtz TW, Pershadsingh HA: Li-tor. Diabetes 44:1139–1146, 1995gands for the peroxisome proliferator-activated receptor-gamma36. Ziyadeh FN: Mediators of hyperglycemia and the pathogenesis ofand the retinoid X receptor-alpha exert synergistic antiproliferativematrix accumulation in diabetic renal disease. Miner Electrolyte effects on human coronary artery smooth muscle cells. Mol Cell
Metab 21:292–302, 1995 Biol Res Commun 3:159–164, 200037. Adler S: Structure-function relationships associated with extracel- 55. Law RE, Goetz S, Xi XP, et al: Expression and function of PPAR-
lular matrix alterations in diabetic glomerulopathy. J Am Soc Ne- gamma in rat and human vascular smooth muscle cells. Circulationphrol 5:1165–1172, 1994 101:1311–1318, 2000
38. Striker GE, He CJ, Liu ZH, et al: Pathogenesis of non-immune 56. Law RE, Meehan WP, Xi XP, et al: Troglitazone inhibits vascularglomerulosclerosis: Studies in animals and potential applications smooth muscle cell growth and intimal hyperplasia. J Clin Investto humans. Lab Invest 73:596–605, 1995 98:1897–1905, 1996
39. Vernier RL, Steffes MW, Sisson-Ross S, Mauer SM: Heparan 57. Marx N, Schonbeck U, Lazar MA, et al: Peroxisome proliferator-sulfate proteoglycan in the glomerular basement membrane in type activated receptor gamma activators inhibit gene expression andI diabetes mellitus. Kidney Int 41:1070–1080, 1992 migration in human vascular smooth muscle cells. Circ Res 83:1097–
1103, 199840. Cohen MP, Surma ML: Effect of diabetes on in vivo metabolism





























