Embed Size (px)
Citation preview

HAL Id: hal-00899492https://hal.archives-ouvertes.fr/hal-00899492
Submitted on 1 Jan 1992
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Étude de l’action des ganglions cérébroïdes et des corpsdorsaux sur la synthèse d’ADN induite par la chaleur
dans l’ovotestis d’Helix aspersa en hibernationP Gomot, B Griffond, C Colard, L Gomot
To cite this version:P Gomot, B Griffond, C Colard, L Gomot. Étude de l’action des ganglions cérébroïdes et des corpsdorsaux sur la synthèse d’ADN induite par la chaleur dans l’ovotestis d’Helix aspersa en hibernation.Reproduction Nutrition Development, EDP Sciences, 1992, 32 (1), pp.55-66. �hal-00899492�

Article original
Étude de l’action des ganglions cérébroïdeset des corps dorsaux sur la synthèse d’ADN
induite par la chaleur dans l’ovotestis
d’Helix aspersa en hibernation
P Gomot, B Griffond C Colard L Gomot
SDI CNRS 6319, faculté des sciences, laboratoire de zoologie et embryologie,place Maréchal Leclerc, 25030 Besançon Cedex, France
(Reçu le 8 juillet 1991; accepté le 24 décembre 1991)
Résumé ― Chez des escargots en début d’hibernation, l’extirpation totale du cerveau (ganglionscérébroïdes : GC et corps dorsaux associés : CD) augmente la synthèse d’ADN des spermatogo-nies lorsque les animaux sont placés à 25 °C pendant un mois. Cet effet n’est pas observé si les ani-maux sont maintenus à 5 °C. La réimplantation immédiate de la totalité du cerveau à des escargotscérébrotomisés et placés à 25 °C ne corrige pas complètement les effets de l’ablation de leur cer-veau. L’implantation soit de CD soit de GC à des escargots cérébrotomisés montre que les CDramènent les synthèses d’ADN et les muliplications spermatogoniales au niveau des témoins, tandisque les GC les stimulent. Les GC et les CD exercent donc des effets antagonistes qui contrôlent lasynthèse d’ADN dans les spermatogonies de l’escargot durant l’hibernation.
Helix aspersa / spermatogenèse / cerveau / hibernation / température
Summary ― Effect of the cerebral ganglia and dorsal bodies on temperature-induced DNAsynthesis in the gonads of hibernating Helix aspersa. Brain extirpation of snails at the start oftheir natural hibernation increased the synthesis of DNA in spermatogonia when the animals weretransferred from 5 to 25 °C for a 4-week period. This effect did not occur if animals were maintainedat 5 °C. The reimplantation of brain (cerebral ganglia: CG + associated dorsal bodies: DB) in brain-ablated snails failed to correct the effects of brain extirpation. The implantation of either DB or CG incerebrotomized hosts showed that, compared to shams, DB restored the level of DNA synthesis andspermatogonial proliferations whereas CG stimulated it. The CG and associated DB were thereforefound to exert antagonistic effects which are responsible for the control of spermatogonial DNA syn-thesis in hibernating Helix aspersa.
Helix aspersa / spermatogenesis / brain / hibernation / temperature

INTRODUCTION
Chez les Mollusques pulmonés, la repro-duction et la gamétogenèse dépendentdes facteurs de l’environnement (Joosse,1984; Gomot et al, 1989) qui agissent parl’intermédiaire du système endocrine donton connaît plusieurs centres producteursd’hormones (Joosse, 1988).
Chez Helix aspersa en dormance, nousavons observé que la méiose mâle est in-hibée par les basses températures (6 à10 °C), tandis que les températures supé-rieures à 16 °C stimulent la multiplicationdes spermatogonies et la spermiogenèse(Gomot et al, 1986). Après extirpation ducerveau, nous avons ensuite montré, parmesure d’incorporation de thymidine tritiéeet observation autoradiographique, quecelui-ci exerce une action inhibitrice sur la
multiplication des spermatogonies et desspermatocytes induite par la chaleur audébut de l’hibernation (Gomot et Gomot,1989).
L’objet du présent travail est de cher-cher comment interviennent les 2 consti-tuants principaux du système neuro-
endocrine cérébral que sont les ganglionscérébroïdes (GC) et les corps dorsaux
(CD) de la gaine conjonctive périganglion-naire. Pour cela, nous avons procédéd’une part à l’extirpation du cerveau et
d’autre part à sa réimplantation ou à celledes GC ou des CD. Nous avons vérifiél’état de survie et le fonctionnement des
tissus transplantés par des études ultra-
structurales et cyto-immunologiques. Pources dernières, nous avons choisi de re-
chercher deux peptides, le FMRFamide etla méthionine-enképhaline, dont nous
connaissons la localisation dans le cer-
veau des escargots et qui jouent un rôleimportant dans le contrôle des corrélationsendocrines (Marchand et Dubois, 1986;Griffond et Mounzih, 1989; Marchand et al,1991). Nous avons ensuite comparé les
effets des différentes interventions sur la
multiplication des spermatogonies.
MATÉRIEL ET MÉTHODES
Les animaux
Les expériences ont été réalisées sur des escar-gots Helix aspersa nés au printemps, élevés enparcs extérieurs et récoltés à l’état adulte à leurentrée en hibernation naturelle au mois d’oc-tobre. Ils ont été ensuite conservés un mois enchambre froide à 5 °C à l’obscurité avant la réa-lisation des expériences.
Conditions expérimentales
Les escargots en hibernation sont réveillés pen-dant quelques heures afin de pratiquer les inter-ventions sur le cerveau selon la technique dé-crite précédemment (Gomot et al, 1990a).Aussitôt après extirpation (E) du cerveau, desescargots reçoivent l’implantation dans l’hémo-coele d’un cerveau entier (groupe E + 1, n = 12),tandis qu’à d’autres on implante seulement lesganglions cérébroïdes (groupe E + GC, n = 12)ou le conjonctif périganglionnaire cérébral conte-nant les corps dorsaux (groupe E + CD, n = 12).Des animaux témoins (T, n = 12) subissent uni-quement l’anesthésie et la section du tégument.Après l’opération, les escargots sont conservésà 5 °C pendant une semaine. Ensuite, 2 lotsd’animaux témoins et opérés sont constitués;l’un demeure à 5 °C (n = 24), tandis que l’autreest placé à 25 °C (n = 24).
Après 4 semaines à 5 °C ou 25 °C, 6 escar-gots de chaque groupe expérimental reçoiventdans l’hémocoele de la région viscérale une in-jection de thymidine-3H du CEA (Commissariatà l’Énergie Atomique - 91191 Gif-sur-Yvette)d’activité spécifique 1 Cilmmol, à raison de 5ycilg de poids frais dans 0,02 ml de liquide deRinger. Cinq heures plus tard, au moment dusacrifice, on prélève 2 morceaux des organesétudiés : un est fixé pour les examens cytologi-que et radioautographique, l’autre est pesé pourla détermination de la teneur en ADN et pour lamesure de la thymidine-3H incorporée.

Étude comparée de la synthèse d ADNdans la gonade et la glande à albumen(organe témoin) par mesurede l’incorporation de thymidine-3H
L’évaluation de la synthèse d’ADN est faite par lamesure de l’incorporation de thymidine-3H (Soko-love et al, 1983). Ces valeurs d’incorporation sontrapportées à la quantité d’ADN dosée par spec-trofluorimétrie (Brunk et al, 1979). Les comparai-sons des résultats, exprimés en dpm/pg d’ADN,sont faites par application du test non paramétri-que U de Mann et Withney (Siegel, 1956).
Étude cytologiqueet radioautographique
Les organes subissent une double fixation gluta-raldéhyde-acide osmique avant d’être inclus dansl’ERL 4206 et traités pour les examens cytologi-ques et radioautographiques selon les procédésdéjà décrits (Gomot et al, 1990b). La comparai-son du nombre et de la surface des organites descellules des corps dorsaux est réalisée par ana-
lyse d’images sur photos de microscopie électro-nique et application du test (t) de Student.
Observations cyto-immunologigues
Des cerveaux témoins et des greffons sont fixésdans du liquide de Bouin-Hollande/sublimé puisdébités en coupes de 7 jim d’épaisseur. Lescoupes sont traitées en immuno-enzymologieselon la procédure décrite par Marchand et ai(1982) pour le FMRFamide et Marchand et Du-bois (1986) pour la méthionine-enképhaline.
RÉSULTATS
État des animaux et des greffons
Survie
Les escargots cérébrotomisés ou implan-tés, maintenus en inactivité à 5 ou 25 °C,
survivent aussi bien que les témoins : au-cune mortalité n’est enregistrée pendant ladurée de l’expérience.
Aspect cytologiquedes transplants cérébraux
Les organes transplantés ne sont pas reje-tés, ils sont retrouvés dans l’hémocoele aumoment du sacrifice de l’animal.
L’étude comparée des CD et des GC aété conduite dans les transplants des es-cargots placés à 25 °C chez qui l’implanta-tion entraîne des modifications de la syn-thèse d’ADN de la lignée mâle.
Les cerveaux implantés entiers pren-nent une forme globuleuse et sont entou-rés par une épaisse capsule conjonctive. Àl’intérieur, les ganglions cérébroïdes limitéspar une lame basale conservent leur struc-ture et leurs relations avec les cellules des
corps dorsaux par l’intermédiaire d’axones.Certains neurones ont une réaction met-
enképhaline positive, tandis que d’autressont FMRFamide positifs. Les cellules descorps dorsaux ont le même aspect quedans les cerveaux témoins et présententune réaction positive à la met-enképhaline.Les organites cytoplasmiques (appareil deGolgi, mitochondries, inclusions lipidiques,grains de sécrétion) y ont le même déve-loppement que dans les cerveaux témoins(tableau 1).
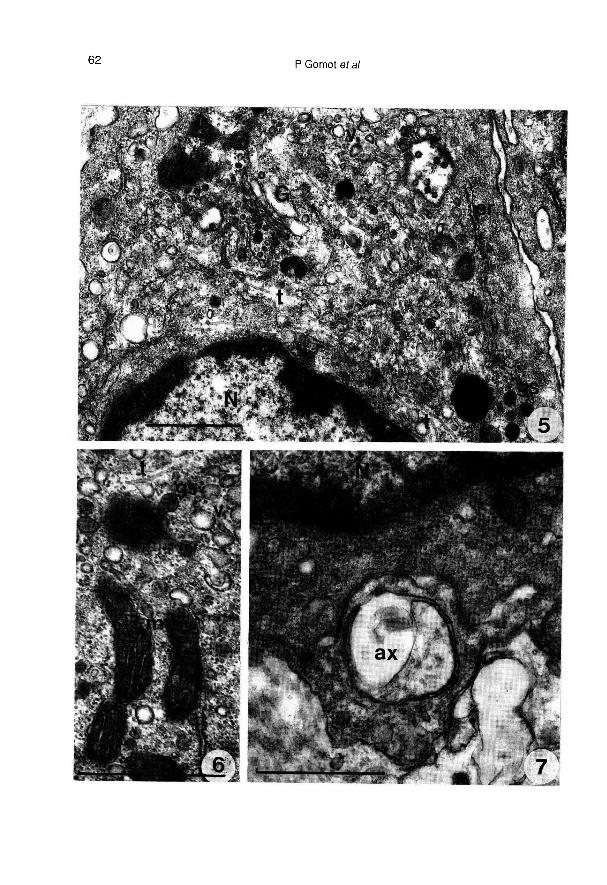
Lorsque les ganglions cérébroïdes iso-lés sont implantés, les neurones surviventbien (fig 1), mais leur disposition est forte-ment modifiée. L’activité des cellules neu-rosécrétrices est attestée par le dévelop-pement important de l’appareil de Golgi etles nombreux grains de sécrétion (fig 2)qui sont libérés par exocytose (fig 3) dansleur environnement immédiat.
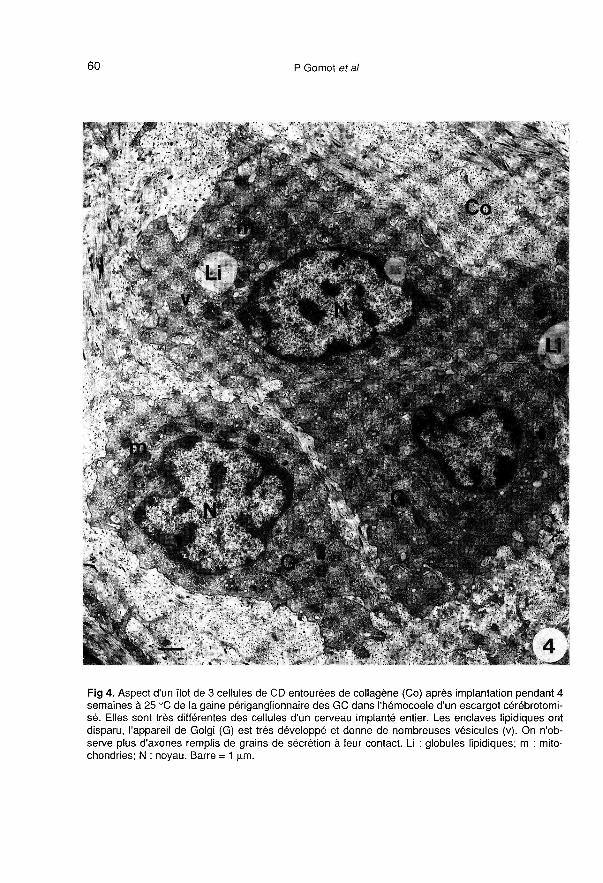
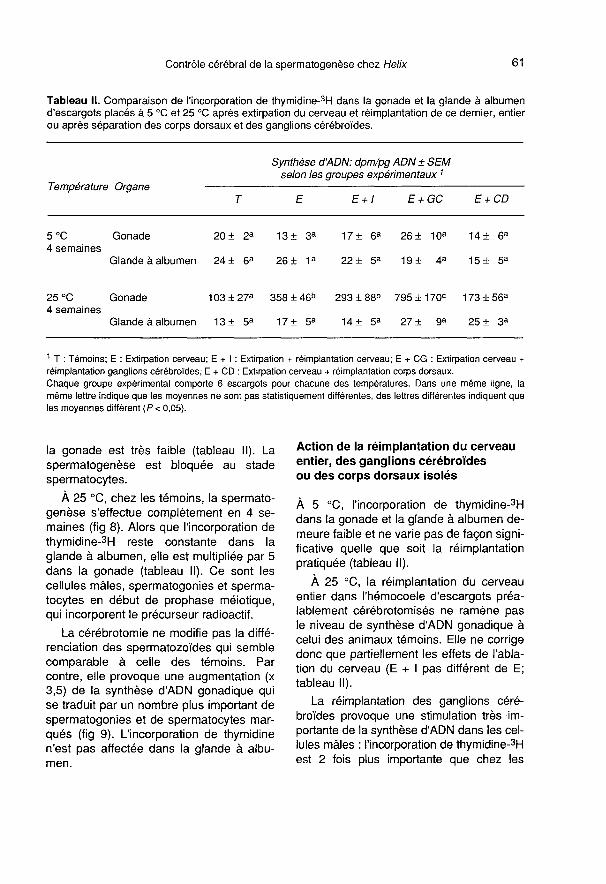
Les cellules des corps dorsaux (séparésdes ganglions cérébroïdes) implantés dansl’hémocoele (fig 4), prennent un aspect
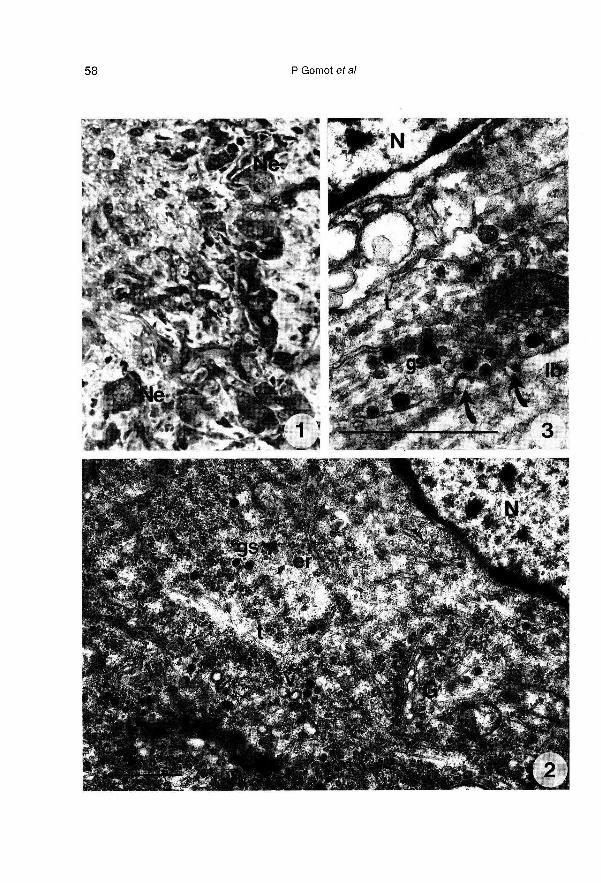

très différent de celui des cellules des CD
des cerveaux greffés entiers et des cer-veaux témoins. Les changements ultra-
structuraux les plus apparents, confirméspar l’étude morphométrique, concernent laforte diminution des enclaves lipidiques(tableau 1), le développement de l’appareilde Golgi (fig 5, tableau 1), l’apparition detrès nombreuses vésicules hérissées et devésicules de réticulum lisse (fig 5), la dimi-nution du nombre de grains de sécrétion(tableau 1). Les mitochondries moins nom-breuses ont une surface relative équiva-lente (tableau 1), leur taille augmente et
elles changent d’aspect; leur matrice de-vient dense aux électrons et leurs crêtess’orientent longitudinalement (fig 6). On re-
connaît encore quelques axones en
contact étroit avec les cellules des CD,mais ils ne contiennent plus de grains desécrétion (fig 7).
Évolution des synthèses d ADNdans l’ovotestis
Action de la cérébrotomiesur la synthèse d’ADN gonadiqueen réponse à l’augmentationde la température
Chez les escargots témoins ou traitésmaintenus à 5 °C, l’incorporation de thymi-dine-3H dans la glande à albumen et dans
la gonade est très faible (tableau 11). Laspermatogenèse est bloquée au stade
spermatocytes.À 25 °C, chez les témoins, la spermato-
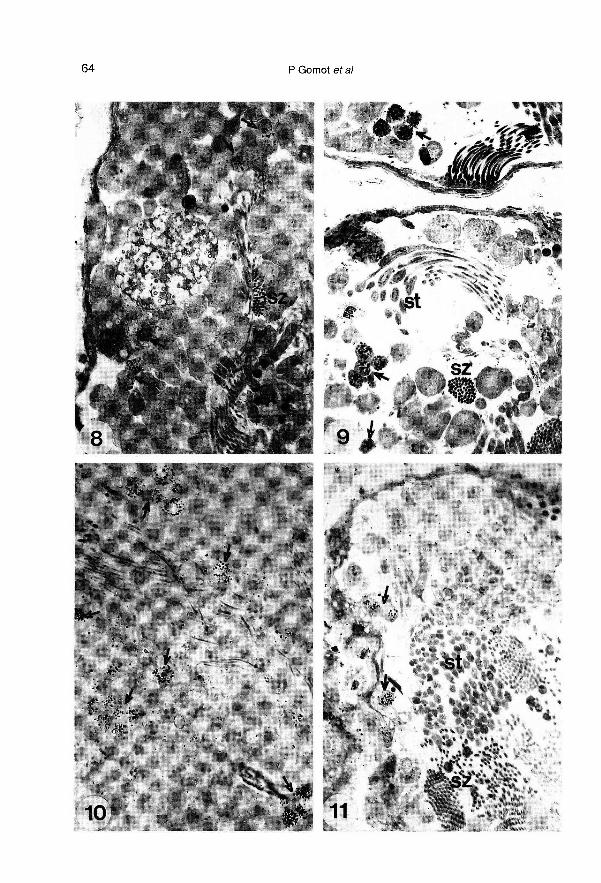
genèse s’effectue complètement en 4 se-maines (fig 8). Alors que l’incorporation dethymidine-3H reste constante dans la
glande à albumen, elle est multipliée par 5dans la gonade (tableau 11). Ce sont lescellules mâles, spermatogonies et sperma-tocytes en début de prophase méiotique,qui incorporent le précurseur radioactif.
La cérébrotomie ne modifie pas la diffé-renciation des spermatozoïdes qui semblecomparable à celle des témoins. Par
contre, elle provoque une augmentation (x3,5) de la synthèse d’ADN gonadique quise traduit par un nombre plus important despermatogonies et de spermatocytes mar-qués (fig 9). L’incorporation de thymidinen’est pas affectée dans la glande à albu-men.
Action de la réimplantation du cerveauentier, des ganglions cérébroïdesou des corps dorsaux isolés
À 5 °C, l’incorporation de thymidine-3Hdans la gonade et la glande à albumen de-meure faible et ne varie pas de façon signi-ficative quelle que soit la réimplantationpratiquée (tableau 11).À 25 °C, la réimplantation du cerveau
entier dans l’hémocoele d’escargots préa-lablement cérébrotomisés ne ramène pasle niveau de synthèse d’ADN gonadique àcelui des animaux témoins. Elle ne corrigedonc que partiellement les effets de l’abla-tion du cerveau (E + I pas différent de E;tableau 11).
La réimplantation des ganglions céré-
broïdes provoque une stimulation très im-
portante de la synthèse d’ADN dans les cel-lules mâles : l’incorporation de thymidine-3Hest 2 fois plus importante que chez les

escargots cérébrotomisés et 8 fois plus quechez les témoins (tableau II). Le nombre despermatogonies et de spermatocytes mar-qués est élevé (fig 10).
La réimplantation du conjonctif péricéré-bral contenant les CD diminue significati-vement la synthèse d’ADN gonadique desescargots cérébrotomisés et la ramène auniveau de celle des animaux témoins
(tableau 11). Ce même traitement diminuele nombre de cellules mâles marquées(fig 11 ).
DISCUSSION
Chez l’escargot (Gomot et al, 1990b),comme chez la limace (Sokolove et al,1983), lorsque les synthèses d’ADN gona-dique sont stimulées expérimentalement,ce sont essentiellement les cellules de la li-
gnée mâle (spermatogonies et spermato-cytes en début de prophase méiotique) quiincorporent la thymidine tritiée.
Les expériences d’extirpation du cer-
veau suivies, soit de la réimplantation ducerveau entier (GC + CD), soit de la réim-plantation des GC ou des CD, mettent enévidence les fonctions respectives des GCet des CD dans la régulation de la sperma-togenèse des escargots. De plus, elles
confirment les résultats antérieurs : rôle dela température dans la réalisation de la
spermiogenèse (Gomot et al, 1990b), exis-tence d’une activité cérébrale inhibitrice dela multiplication des spermatogonies et
des spermatocytes (Gomot et Gomot,1989).
L’implantation des GC et des CD isolésapporte des éléments nouveaux qui témoi-gnent de l’existence d’interrelations fonc-tionnelles étroites entre ces 2 organes.
Lorsqu’ils sont séparés, les GC et les CDexercent individuellement une action oppo-sée sur la synthèse d’ADN des spermato-gonies et des spermatocytes. Les CD im-plantés seuls inhibent la synthèse d’ADN.Leurs cellules ont un aspect fortement acti-vé qui peut s’expliquer par la suppressiondu contrôle inhibiteur de type nerveux
exercé par les GC (Vincent et al, 1984b).L’action des GC sur les CD pourrait s’effec-tuer par l’intermédiaire des neurones
FMRFamide positifs et de leurs axones quisont en contact avec les cellules des CD
(Griffond et Mounzih, 1990) car le FMRFamide synthétique inhibe in vitro la syn-thèse des protéines des CD (Griffond et
Mounzih, 1989).Inversement, l’implantation des GC iso-
lés augmente très nettement la synthèsed’ADN des spermatogonies. Ceci peut ré-sulter de l’absence d’un effet inhibiteur di-rect des CD sur les cellules mâles et/ou
d’un effet stimulateur des GC sur ces
mêmes cellules; la seconde hypothèsesuggère une influence inhibitrice des CDsur les GC, comme cela avait été supposépar Bohmig (1931) chez Planorbarius.
Cette action inhibitrice des CD sur la sper-
matogenèse opposée à leur action stimula-trice sur la vitellogenèse (Vincent et al,

1984a) apporte un élément nouveau decompréhension de la régulation de l’her-
maphrodisme chez les escargots.Les résultats des associations in vitro
de CD isolés et de gonades (Vincent et al,1984a) ou des implantations de GC et deCD séparés décrites dans ce travail met-tent en évidence la coexistence dans le
cerveau d’Helix aspersa de facteurs stimu-lateurs et de facteurs inhibiteurs. Aussi,peut-on penser, comme Mathieu (1987)chez Mytilus, que le contrôle de la multipli-cation goniale fait intervenir 2 facteurs an-tagonistes dont les taux respectifs détermi-nent les phases de la gamétogenèse enfonction des conditions d’environnement.
En conclusion, nos expériences appor-tent la preuve que le cerveau exerce une
influence globale inhibitrice sur la multipli-cation des spermatogonies et des sperma-tocytes au début de l’hibernation. Cette ac-tion semble résulter de 2 influences
antagonistes des CD et des GC qui peu-vent produire respectivement un facteur in-hibiteur (CD) et un facteur mitogène (GC).Cette action inhibitrice du cerveau contrôlela multiplication des spermatogonies et
des spermatocytes pendant la période d’hi-bernation.
REMERCIEMENTS
Les auteurs remercient vivement Brigitte Joli-bois pour la préparation du manuscrit.
RÉFÉRENCES
Bohmig L (1931) Zur Kenntnis der sogenanntenDorsallapen des Gehirns von Lymnaea sta-gnalis (L) Lam und Planorbis corneus (L)Pfeiff. Sber Akad Wiss Wien Math Nat KI
140, 319-335
Brunk CF, Jones KC, James TW (1979) Assayfor nano-gram quantities of DNA in cellularhomogenates. Anal Biochem 92, 497-500
Gomot P, Gomot L (1989) Inhibition of tempera-ture-induced spermatogenic proliferation by abrain factor in hibernating Helix aspersa (Mol-lusca). Experientia 45, 349-351
Gomot P, Griffond B, Gomot L (1986) Effets dela température sur la spermatogenèse d’Es-cargots Helix aspersa maintenus en repos ar-tificiel. CR Séances Acad Sci Paris 302, 27-32
Gomot P, Gomot L, Griffond B (1989) Evidencefor a light compensation of the inhibition ofreproduction by low temperatures in the snailHelix aspersa. Ovotestis and albumen glandresponsiveness to different conditions of pho-toperiods and temperatures. Biol Reprod 40,1237-1245

Gomot P, Gomot L, Marchand CR, Colard C(1990a) Extirpation and transplantation ofthe brain of the snail Helix aspersa: a studyof the survival of the animal and the implant.Can J Zoo168, 1505-1512 2
Gomot P, Griffond B, Gomot L (1990b) Actionde la température sur la multiplication descellules mâles et la spermatogenèse d’Helixaspersa en hibernation. J Thermal Biol 15,267-280
Griffond B, Mounzih K (1989) In vitro inhibitionof methionine incorporation in the dorsal bo-dies of Helix aspersa by synthetic FMRFamide. Comp Biochem Physiol 92C, 45-49
Griffond B, Mounzih K (1990) Innervation of thedorsal body cells of Helix aspersa: immuno-cytochemical evidence for the presence ofFMRFamide-like substances in nerves and
synapses. Tissue & Cell22, 741-748
Joosse J (1984) Photoperiodicity, rhythmicityand endocrinology of reproduction in the
snail Lymnaea stagnatis. ln: PhotoperiodicRegulation of Insect and Molluscan Hor-
mones. Ciba Foundation Symposium, vol
104, Pitman, Londres, 204-220
Joosse J (1988) The hormones of molluscs. ln:Endocrinology of selected invertebrate types(H Laufer, GH Downer, eds), Alan R Liss Inc,89-140
Marchand CR, Dubois MP (1986) Mise en évi-dence par immunocytochimie de substancesapparentées à la méthionine-enképhalinedans divers tissus de l’escargot Helix asper-sa Müller. Haliotis 15, 241-247
Marchand CR, Wijdenes J, Schot LPC (1982)Localisation par la technique cyto-immuno-
enzymologique d’un neuropeptide cardio-excitateur (le FMRF-amide) dans le colliernerveux périoesophagien d’Helix aspersaMüller (Gastéropode, Pulmoné, Stylommato-phore). CR Séances Acad Sci Paris Ser fil294, 39-44
Marchand CR, Griffond B, Mounzih K, Colard C
(1991) Distribution of methionine-enkephalin-like and FMRFamide-like immunoreactivitiesin the central nervous system (including dor-sal bodies) of the snail Helix aspersa Müller.Zool Sci 8, 909-913 3
Mathieu M (1987) Étude expérimentale descontrôles exercés par les ganglions nerveuxsur la gamétogenèse et les processus méta-boliques associés chez la moule Mytilus edu-lis. Thèse Doct Etat, Université de Caen
Siegel S (1956) The case of two independentsamples in non parametric statistics. Interna-tional Student Ed, McGraw Hill InternationalBook Co, Auckland, p 116 6
Sokolove PG, Melrose GR, Gordon TM, O’NeillMC (1983) Stimulation of spermatogonialDNA synthesis in slug gonad by a factor re-leased from cerebral ganglia under the in-fluence of long days. Gen Comp Endocrinol50, 95-104
Vincent C, Griffond B, Gomot L, Bride J (1984a)Étude in vitro de l’influence des corps dor-saux sur l’ovogenèse d’Helix aspersa Müll.Gen Comp Endocrinol54, 230-235
Vincent C, Griffond B, Wijdenes J, Gomot L(1984b) Contrôle d’une glande endocrine :les corps dorsaux par le système nerveuxcentral chez Helix aspersa. CR SéancesAcad Sci Paris Sér /// 299, 421-426





























