Embed Size (px)
Citation preview

TARTU ÜLIKOOL
Bioloogia-geograafia teaduskond
Botaanika ja ökoloogia instituut
Merli Nagel
TURBASAMBLAD EESTIS: ÖKOLOOGIA,
LEVIK JA KAITSE
Bakalaureusetöö
Juhendaja: teadur Kai Vellak
Tartu 2006

2
Sisukord
1.TURBASAMBLAD JA NENDE ÖKOLOOGIA ...........................4
1.1 LIIGILINE MITMEKESISUS JA LEVIK...........................................................................4 1.2 TURBASAMMALDE UURIMISE AJALOOST EESTIS...............................................4 1.3 TURBASAMMALDE PRODUKTIIVSUS.........................................................................6 1.4 TURBASAMMALDE PALJUNEMINE .............................................................................7 1.5 TURBASAMMALDE SEOS VEEGA.................................................................................8 1.6 TURBASAMMALDE SEOS MINERAALAINETEGA ..................................................9 1.7 TURBASAMMALDE SEOSED SOONTAIMEDEGA ..................................................11 1.8 TURBASAMMALDE TÄHTSUS LOODUSES..............................................................13
2. KAITSEALUSTE TURBASAMMALDE LEVIK ....................14
2.1 LOIGU-TURBASAMMAL - SPHAGNUM INUNDATUM RUSS. .........................16 2.1.1 Levik ....................................................................................................................................16 2.1.2 Kasvukohad ja ökoloogia .................................................................................................19
2.2 LINDBERGI TURBASAMMAL – SPHAGNUM LINDBERGII SCHIMP. EX
LINDB.............................................................................................................................................19 2.2.1 Levik ....................................................................................................................................19 2.2.2 Kasvukohad ja ökoloogilised nõudlused ........................................................................21
2.3 VIIEREALINE TURBASAMMAL - SPHAGNUM QUINQUEFARIUM (LINDB. EX BRAITHW.) WARNST .......................................................................................................22
2.3.1 Levik ....................................................................................................................................22 2.3.2 Kasvukohad ja ökoloogilised nõudlused ........................................................................24
2.4 WULFI TURBASAMMAL – SPHAGNUM WULFIANUM GIRG. ........................24 2.4.1 Levik ....................................................................................................................................24 2.4.2 Kasvukohad ja ökoloogilised nõudlused ........................................................................27
KOKKUVÕTE .....................................................................................................................28
SUMM ARY .............................................................................................................................29
KASUTATUD KIRJANDUS..............................................................................................................30
KASUTATUD INTERNETIALLIKAD:.......................................................................................................33

3
Sisse juhatus
Eesti soodes ja märgades metsades kasvab üle poole Euroopas levinud
turbasammaldest. Siin on praeguseks teada 37 turbasamblalii ki, Euroopas on neid
kokku 54. Mõnd lii ki leidub meil peaaegu igas soos- või soostuvas koosluses, teised
seevastu on kasvukoha suhtes ülinõudlikud ning neid kohtab vaid väga harva (Vellak,
2004).
Turbasamblaid iseloomustab tohutu veeimamisvõime, mistõttu neid on
kasutatud nii rahvameditsiinis kui ka sõdade ajal sidumismaterjalina vati asemel
(Ingerpuu, 1994). Palju kasutatakse turbasamblaid aianduses ja isegi energeetikas, kus
turvast peetakse üheks alternatiiviks põlevkivienergeetikale. Turba kaevandamine
ületab Eestis turba loodusliku juurdekasvu 3,5 – 4,5 korda; kui võrrelda lubatud
kasutusmääraga, siis üle kuue korra (Ilomets, 2003). Seega on need pisikesd taimed
tähtsad nii ökoloogili selt kui ka majanduslikult ning on küll põhjust mõelda nende
esialgu veel suuremalt jaolt tavaliste sammalde kaitsele.
Eestis kuulub seaduse kaitse alla neli turbasambla lii ki. Need lii gid kuuluvad
kolmandasse kaitsekategooriasse, ning nende arvukust ohustab elupaikade ja
kasvukohtade hävimine.
Antud töö eesmärk on anda ülevaade turbasammaldest ja nende ökoloogiast,
selgitada nende suhteid soontaimedega ning iseloomustada Eesti looduskaitsealuste
turbasammalde seisundit ja levikut. Töö praktili seks osaks on koostada nende nelja
lii gi levikukaardid Eestis ning sisestada registreeritud andmestik kaitsealuste
samblalii kide andmebaasi. Levikukaartide aluseks on EMA Põllumajandus- ja
Keskkonnainstituudi herbaariumi (TAA) ja Tartu Ülikooli (TU) herbaariumi andmed.
Kaardid on koostatud programmiga MapInfo. Töös kasutatud Eesti esinevate lii kide
ladinakeelsete nimede aluseks on Eesti sammalde määrajas (Ingerpuu jt., 1998) ja
Eesti taimede määrajas (Leht jt., 1999) esitatud süsteemid.
Tahaksin tänada käesoleva töö valmimise eest oma juhendajat Kai Vellakut.

4
1.Turbasamblad ja nende ökoloog ia
1.1 Liigili ne mitmekesisus ja levik
Turbasamblad (Sphagnum L.), rahvapäraselt ka ka rabasamblad ja
valgesamblad (Laasimer et. al., 1954) kuuluvad lehtsammalde klassi, moodustades
turbasammalde alamklassi, millesse kuulub üks ainuperekonnaline sugukond
turbasamblalised (Sphagnaceae) (Ingerpuu & Vellak, 1998).
Turbasamblaid on maailmas umbes 300 lii ki (Clymo & Hayward, 1982),
Eestis on preaguseks teada 37 lii ki turbasamblaid (Vellak, 2004). Peamiselt kasvavad
turbasamblad parasvöötmes ja boreaalses kliimavöötmes (Clymo & Hayward, 1982).
Ka teistel laiuskraadidel kasvab turbasamblaid, kuid valdavalt vaid mäestike
pii rkondades, näiteks Austraalia lumistes mägedes, Andides ja Aafrika mägede
tippudes. Näiteks S. junghuhnianum ssp. junghuhnianum Dozy & Molk. kasvab
Malaisia mägedes, Hiinas ja Jaapanis (Clymo & Hayward, 1982). Kuigi üksikud lii gid
võivad kasvada väga ekstreemsetes kasvukohtades, valitsevad turbsamblad erinevat
tüüpi soodes ja soostuvates- või soometsades. Nii näiteks on alli kasoo turbasammalt
(Sphagnum teres) ja sulgjat turbasammalt (S. subnitens) leitud kasvamas Islandi
kuumaveealli kate juurest, kus vee pH alli kates varieerub väga happelisest kuni
aluseliseni ning veetemperatuur taimede vahel võib olla isegi kuni 40 °C (Clymo &
Hayward, 1982).
Looduses on turbasamblaid üsna raske üksteisest eristada, enamasti on vaja
luubi, mõnegi lii gi puhul isegi mikroskoobi abi. Ent teades, missugused lii gid milli ses
koosluses kasvavad, saame abi ka ümbrust silmitsedes: on lii ke, mis kasvavad
valdavalt märgades kohtades, mõned elavad ainult laugastes, osa jälle eelistab
madalsoid ja osa on rabade asukad ( Vellak, 2004).
1.2 Turbasammalde uurimise ajaloost Eestis
Kõige kuulsam Eestist pärit sfagnoloog on Edmund August Friedrich Russow
(8.03.1841 – 23.04.1897). 1865. aastal kaitses ta magistrikraadi turbasammalde
süstemaatikast. Magistritöös kirjeldab ta uut turbasamblalii ki, millele oma hea sõbra
auks paneb nimeks Sphagnum girgensohnii . Samas kirjeldab ta ka pudeva

5
turbasambla (Sphagnum cuspidatum) arvukaid varieteete, millede hulgast hilj em said
lii gi staatuse Sphagnum majus ja Sphagnum balticum (Ingerpuu, 2001).
Ta on seitsme põhjapoolkeral väga laialdaselt levinud turbasamblalii gi
ainukirjeldaja või kaaskirjeldaja. Lisaks üldtunnustatud taimelii kidele (kui jätta
kõrvale vaieldavad hunditubakalii gid ning saaremaa robirohi) ongi Eesti materjali
põhjal teadusele kõige enam uusi turbasamblalii ke kirjeldatud. Lisaks E. Russowi
kirjeldatud lii kidele on Eestist kirjeldanud uue turbasamblalii gi ka K. Girgensohn,
nimetades selle kohaliku mõisniku auks Wulfi turbasamblaks (Sphagnum wulfianum)
(Ingerpuu, 2001).
Ka E. Russowi enda auks on nimetatud kaks taimelii ki, mis mõlemad esinevad
Eestis, nendest üks on Eestis tavaline Russowi turbasammal – Sphagnum russowii
Warnst. ja teiseks, meil samuti tavaline orhidee Russowi sõrmkäpp (Dactylorhiza
russowii ) (Ingerpuu, 2001).
E. Russowi eelistatuimad uurimispii rkonnad asusid Tartu ümbruses,
(Tähtveres), Lõuna- Eestis (end. nimega Appelsee) ning põhjarannikul, Käsmu
poolsaarel, kus suvitas perega mitmetel aastatel ning mida ta nimetas “Eldoradoks”
sfagnoloogile. Käsmust kogus ta tohutu turbasammalde materjali , millest suur osa
paikneb ilmselt sõja ajal Peterburi sattunud ja alles 1988. aastal Eestisse tagasi
jõudnud kogus. Oma ekskursioonidel märkas E. Russow ka inimese poolt loodusele
tekitatud kahju. Oma 1894. aasta töös märgib ta turbasammalde kadumist Tähtvere
metsaaladel kraavituse ja raiete tagajärjel ning kirjutab: “Kultuur rikub loodust!”
(Ingerpuu, 2001).
Samuti uuris turbasamblaid kuulus rabauurija Viktor Masing (11.04.1925-
18.03.2001). Viktor Masing arvas, et soid saab uurida vaid nende pärisperemehi –
turbasamblaid hästi tundes ( Kalda, 2000).
Tänapäeval üks tuntum turbasammalde uurija on V. Masingu õpilane Mati
Ilomets. Turbasammalde kasvule ja produktiivsusele olid pühendatud nii tema
kursuse- kui ka diplomitöö ning geoloogia-mineraloogiateaduste kandidaadi
dissertatsioon "Turbasammalde ja sfagnumturba juurdekasvu uurimine turbalasundite
stratigraafili se jaotamise eesmärkidel" 1982. a. ja kümned teadusartiklid kogumikes ja
erialaajakirjades. Tema juhendamisel on turbasammaldest ja soode eri aspektidest
erinevate ülikoolide juures kaitstud mitukümmend kursuse- ja diplomitööd, 4
magistri- ning doktoritöö. Ta on olnud mitmete väitekirjade oponendiks (Truus,
Karofeld, 2000).

6
Peale M. Ilometsa on teinud oma lõputöö turbasammalde teemal Tartu
Ülikoolis veel neli õpilast. 1977. aastal uuris Vill em Laas turbasammalde
fütoproduktsiooni ja turbatekkekihis toimuvat turvastumist. 1982. aastal tegi Anne
Reitel oma lõputöö teemal “Turbasammalde kasv ja produktiivsus erinevais metsa- ja
sootüüpides (Leningradi oblasti kesk-taiga tingimusis)” ning 1996. aastal Katrin
Pajuste teemal “Sphagnum fuscum (Schimp.) Klinggr. ja S. rubellum Wils.
ökoloogili ne eristuvus Männikjärve raba mikroreljeefil ” . Aastal 1997 valmis Paljula
Raimol lõputöö pealkirjaga “Soomaa soode arengu ajalis-ruumiline dünaamika ja
seda mõjutavad tegurid“ .
1.3 Turbasammalde produ ktii vsus
Produktiivsus on defineeritud kui kuiv biomass, mis on toodetud aastas ühe
ruutkilomeetri kohta (Gunnarsson, 2005).
Enam biomassi produtseerivad turbasamblad kevadel ja sügisel, mil on
piisavalt niiskust nende intensiivseks kasvuks (Gunnarsoon, 2005). Turbasammalde
produktiivsus rabades on vahemikus 5- 550 g m-2 aastas (Malmer, 1993), Eestis on
mõõdetud produktiivsuseks 0,7 – 1,3 g dm-2 aastas (Ilomets, 1982).
Turbasamblad on kõige tähtsam turvast moodustav taimerühm ja neis sisaldub
rohkem süsinikku kui üheski teises taimeperekonnas (Clymo & Hayward, 1982).
Produktsiooni vähenedes arvatavasti väheneb ka turba produktsioon ning suuremal
skaalal võib see mõjutada globaalset süsiniku tasakaalu (Gunnarsson, 2000).
Erinevad turbasamblalii gid asustavad erinevaid raba kasvupaiku: neid võib
leida rabades mätaste tipuosas, älvestes ja laugastes. Erinevates mikroelupaikades on
HULQHY�WRLWDLQHWH�VHLVXQG�MD�HULQHY�YHHUHåLLP�MD�VHHSlUDVW�RQ�HULQHYDWHO�OLLNLGHO�erinevates elupaikades ka erinev biomassi produktiivsus (Gunnarsson, 2005).
Klimaatili sed tingimused koos geograafili se asetusega on tähtsad faktorid hindamaks
globaalselt turbasammalde produktiivsust. Tähtsamad faktorid on keskmine aastane
temperatuur ja sademed. Produktiivsus on madal kui need mõlemad faktorid on
madalad (Gunnarsson, 2005). Nii on kindlaks tehtud, et peamiselt sektsiooni
Cuspidata kuuluvatel turbasamblalii kidel, kes kasvavad märjemates ja
toitainerikkmates kasvukohtades (älveservad, peenrdevahed) on ka produktiivsus
kõrgem. Samas aga lii kidel sektsioonist Acutifolia, kes elavad rabades peamiselt

7
mätaste ülaosades ja toituvad valdavalt vihmaveest, jääb produktiivsus suhteliselt
madalaks (Gunnarsson, 2005).
Turbasambla võsu lineaarne juurdekasv on Eesti rabades 5 –65 mm, umbes
samas ulatuses muutub samblavarre alumine osa turbaks (Valk, 2005). Lineaarne
juurdekasv sõltub sademete ja auramise hulgast ning keskmisest õhutemperatuurist ja
soovetetasemest. Suhteliselt madalamal paiknevate lii kide, balti j a lill aka turbasambla
(S. balticum ja S. magellanicum) juurdekasv on kõige tugevamini seotud
sademetehulgaga. Tüüpili ste mättalii kide nagu pruun ja punane turbasammal (S.
fuscum ja S. rubellum) juurdekasv on seoses temperatuuri ja auramisega.
Keskkonnategurite mõju diferentseeritus turbasamblakooslustele tuleneb
turbasamblalii kide anatoomilis-morfoloogili stest erinevustest. Turbasamblavaiba
veetõstesüsteemi põhiosa – kapill aaride – läbimõõdu ja veetõstevõime vahel on otsene
negatiivne seos. Kompenseerimaks veetaseme langust suureneb okste tihedus
älveslii kidel, parandades nii vee kapill aarset neeldumist. Tulemusena langeb lineaarne
juurdekasv, kuid produktsioon ei tarvitse väheneda (Ilomets, 1980).
Mättalii kidel on kapill aaride läbimõõt piisavalt väikene tagamaks juurdekasvu
vähest seost veetaseme ja sademetega. Koos temperatuuriga suurenev auramine tõstab
taimedele kättesaadavate toitainete hulka. Toitainete ülejäägi kasvades pidurdub
taimede apikaalne kasv ja suurenb lateraalne, st. suureneb tihedus ja lineaarne
juurdekasv väheneb. Tiheduse tõus omakorda vähendab auramist. Selli ne
tagasisidemehhanismi olemasolu tagab turbasamblavaiba produktsiooni stabiil suse
muutuvates keskkonnatingimustes. Järelikult vaatamata lineaarse juurdekasvu
muutumisele (keskkonnategurite mõjul) jääb turbasamblakoosluste (olenemata
kasvukohast) produktsioon stabiil seks (Ilomets, 1980).
1.4 Turbasammalde paljunemine
Turbasammaldel erili si sigikehi ei teki, vegetatiivne paljunemine toimub neil
varre või oksa tükikestega, mis tekivad näiteks ladvapunga vigastamisel või võsude
murdumisel. Paljudel lii kidel, kel sporogoonid ehk eoskuprad esinevad harva, on
vegetatiivne paljunemine peaaegu ainsaks paljunemisviisiks (Laasimer et. al., 1954).

8
Vegetatiivne paljunemine võsude harunemise teel on arvatavasti kõige tähtsam
vahend paljunemiseks ja püsimiseks ühes kasvukohas. Vegetatiivset paljunemist
peetakse turbasammalde paljunemises eoselisest tähtsamaks seetõttu, et nende
püsimajäämine soodsas keskkonnas on taime elutsüklis odavam kui produtseerida
suurt hulka eoseid (Sundberg, 2000).
Enamus boreaalses regioonis kasvavatest turbasammaldest toodab regulaarselt
sporofüüte ning see seletab suurel määral turbasammalde laialdast levikut ka
toitainete vaeses kasvukohas (Sundberg, 2000). Katsetest turbasammaldega on
selgunud, et eosed püsivad suhteliselt kaua levisepangas ning võivad kaasa aidata
sambla taastekkele sobivate tingimuste saabudes. Mõne lii gi eosed ( S. balticum, S.
fuscum ) säilit asid oma idanevuse ka pärast 13 aastat külmutatult hoidmist (Sundberg,
2000). Ühes eoskupras olevate eoste arv on lii giti varieeruv, sõltudes nii kupra kui ka
eoste suurusest. Turbasammalde eoste suurus on vahemikus 20-50 µm (Sundberg,
2000). Rohkem eoseid on lii kidel, millel on suurem eoskupar ja väiksemad eosed.
Kupra suurus lii kidesiseselt on positii vselt korreleeritud aga kapiitulumi suurusega
(Sundberg & Rydin, 1998). Eosed valmivad ja vabanevad küll lii giti erinevatel
aegadel, kuid see jääb enamasti suve lõppu, olles eriti tundlik põua suhtes (Sundberg,
2002).
1.5 Turbasammalde seos veega
Turbasamblad vajavad elutgevuseks regulaarselt vett ja seepärast ongi nad
märgade elupaikade asukad (Gignac, 1993). Mõned lii gid võivad olla isegi täielikult
veesisesed taimed (Laasimer et. al., 1954). Turbasamblad puuduvad kõrbetes,
kaasaarvatud polaarsetes kõrbetes, kuivades metsades, kuivadel ja pool-kuivadel
rohumaadel, mis peegeldab nende pidevat veevajadust, mis selli stel aladel puudub
(Gignac, 1993).
Soode teke ja areng on alati seotud lii gniiskuse ja niiskuslembeste taimedega.
Selli sel metsamaal, kus vettpidav horisont (tavaliselt savikiht või nõrgkivi) on
maapinna lähedal ja tasane reljeef (pinnamood) ei võimalda lii gvee äravoolu ning
muld on muutunud lii gniiskeks, hakkavad levima sootaimed, eelkõige turbasamblad

9
(Valk, 2005). Turbasamblas olev vee mass on tavaliselt 10 – 14 korda suurem tema
kuivaine massist (Valk, 2005).
Kõige tundlikumad on tXUEDVDPEODG�YHHUHåLLPL�PXXWXVWH�VXKWHV��3LQQDVH�kuivendamine mõjub neile alati hävitavalt (Laasimer et. al., 1954). Paremaks vee
kogumiseks ja säilit amiseks on turbasammaldel väljakujunenud spetsii fili ne
anatoomiline ehitus. Lehtede suured surnud rakud (hüalotsüstid) on väliskeskkonnaga
ühenduses pooride kaudu. Liigniiskuse puhul täituvad need rakud veega, põua ajal
aga õhuga. Küllaldase niiskuse korral sisaldab enamik turbasamblaid vett üle 90%
oma esialgsest kaalust (Valk, 2005). Turbasammaldel on väga kõrge veesäilit amise
võime, mis tuleneb okste ja varte vahel kujunenud erili sest juhtesüsteemist. Nimelt
paiknevad turbasammalde oksad varrel kimpudena, millest osa on laiuvad ning
hangivad vett ümbtritsevast keskkonnast, osa oksi (nn. rippoksad) aga lii buvad piki
vart ning moodustavad koos vart ümbritseva epidermiga vee lii kumist tagava
kapill arsüsteemi (Laasimer et. al., 1954). Näiteks on leitud, et mätaste ülaosas kasvav
lii k teravlehine turbasammal (S. capillif olium) mahutab küllastumisel rohkem vett,
kaotab seda aeglasemalt ja seega püsib märjana kauem ka põuaperioodidel (Andrus,
1986).
1.6 Turbasammalde seos mineraalainetega
Seda, et turbasamblad eelistavad kasvada happelises keskkonnas ja on ise
selli se keskkonna loomisel abiks, on teatud juba pikka aega (Clymo & Hayward,
1982). Neil on võime muuta keskkonda happelisemaks, vabastades vesinikioone ja
saades vastu lahustuvaid katioone. Eelistatult absorbeeritakse kõrgema valentsiga
katioone (Ca2+, Mg2+, Fe3+), mida omastatakse kergemini kui madalama valentsiga
katioone (Na+, K+) (Andrus, 1986). On kindlaks tehtud, et mida kõrgemal veetasemest
taim kasvab, seda parem katioonide vahetusvõime tal on ja samuti efektiivne
katioonide vahetusvõime langeb kui pH langeb (Clymo & Hayward, 1982).
Nõudlikkuse järgi mineraaltoitainete suhtes võib turbasamblad jagada kolme
rühma: 1) oligotroofsed (kõige vähema nõudlikkusega, rabas kasvavad), 2)
mesotroofsed (keskmise nõudlikkusega, kasvavad sii rdesoodes ja rabastuvates

10
metsades) ja 3) eutroofsed (suurima nõudlikkusega, kasvavad madalsoodes)
(Laasimer et. al., 1954).
Turbasamblad taluvad hästi mineraalainete vähesust, nad suudavad toituda
sademeveest ja õhus leiduvast mineraalsest tolmust. Kasvades kõige
toitesoolavaesemates tingimustes, moodustavad nad oligotroofset rabaturvast
mineraalainesisaldusega alla 5% (Valk, 2005). Teine toitainete alli kas samblakihile on
vee kapill aarne tõus. See protsess on tähtsam madalsoodes, kuna seal sisaldab soovesi
rohkem K+, Mg2+ ja Ca2+ ioone. Selle mehhanismi tähtsus kahaneb koos kasvava
veetasemega ja niiskusega. Soontaimede maapealne kõdu ja varis on suhteliselt
vähetähtis toitainete alli kas turbasammaldele (Malmer, 1993).
Turbasammaldel on soontaimedega võrreldes tunduvalt väiksem võime
reguleerida mineraalsete toitainete akumulatsiooni. See annab suurema elementide
variatsiooni sammaldes kui soontaimede maapealsetes osades (Malmer, 1993).
Makrotoitained nagu lämmastik, fosfor, kaalium ja väävel akumuleeruvad
turbasammaldes põhili selt läbi juurdevoolu klorofülli sisaldavatesse rakkudesse ning
magneesium ja kaltsium akumuleeruvad läbi adsorptsiooni (Malmer, 1993).
Kontsentratsioonid suvel ei erine kontsentratsioonidest sügisel ja kevadel.
Turbasammalde puhul lämmastiku, fosfori ja kaaliumi kontsentratsioonid on
suuremad apikaalsetes osades, magneesium ja kaltsium on küllaltki ühtlaselt jaotunud
ja Fe3+ ioonide kontsentratsioon on suurim basaalsetes osades. Fosfor ja ilmselt ka
lämmastik ja kaalium pidevalt paigutuvad ümber madalamatest, vananevatest osadest
aktiivselt kasvavatesse apikaalsetesse osadesse seesmise transpordi läbi. Mg2+ ja Ca2+
ioonide summa on umbes sama kõikides pii rkondades, näitates et vahetusvõime ei
muutu sambla vanusega ning et Mg2+ja Ca2+ kontsentratsioonid ei suurene vanusega
nagu mõnedes soontaimedes (Malmer, 1993).
Väga halvasti taluvad turbasamblad kaltsiumioonide suurt kontsentratsiooni.
See on ka üks põhjusi, miks nad Kirde – Eestis mõnedel lubjatolmuga saastunud
rabadel surevad (Ingerpuu, 1994).

11
1.7 Turbasammalde seosed soon taimedega
Turbasamblad domineerivad raba taimekooslustes – nad on edifikaatorid, mis
mõjustavad teiste taimede elu- ja keskkonnatingimusi; moodustades laiuvaid ja
omavahel liit uvaid padjandeid ning muutes keskkonnaolud oma kasvuks ja levikuks
järjest paremaks, teistele, nõudlikumatele taimedele, aga halvemaks, mistõttu need
taanduvad (Valk, 2005).
Tavalisemad soontaimed, mis kasvavad koos turbsammaldega on tupp-vill pea
(Eriophorum vaginatum), valge nokkhein (Rhynchospora alba), raba jänesvill
(Trichophorum cespitosum), kanarbik (Calluna vulgaris). Samuti mustikas
(Vaccinium myrtill us), sinikas (Vaccinium uliginosum), rabamurakas (Rubus
chamaemorus) ja pohl (Vaccinium vitis-idaea). Silmatorkavad põõsad on kukemari
(Empetrum nigrum) ja sookail (Ledum palustre) ja samuti putuktoidulised taimed
nagu ümaralehine huulhein (Valk, 2005). Laiade pehmete lehtedega taimed on
rabades haruldased, väljaarvatud märkimisväärne erand rabamurakas. Mõnedel nendel
lii kidel, nagu kanarbik, on vähe juuri või ei ole üldse juuri turbas. Teistel nagu tupp-
vill pea ja rabamurakas on vähemalt mõned juured suure aerenhüümiga, mis pakub
neile efektiivset kanalit hapniku transpordiks juurtest tippu. Enamus neist lii kidest on
risoomidega püsikud, üheaastased taimed on haruldased (Clymo & Hayward, 1982).
Rabaturba pideva pealekasvamise tõttu tõuseb järjekindlalt ka rabavee tase ja
selle tõttu jäävad rabas kasvavate taimede juured iga aastaga üha sügavamale turbasse
ja rabavette. Rabavesi tõuseb umbes niisama palju, nagu kasvab peale rabaturvast. Et
rabavesi on hapnikupuuduse ja toitesoolade vaesuse tõttu ebasoodsaks keskkonnaks
taimejuurtele, siis moodustavad paljud rabas kasvavad taimed (huulheinad, kanarbik,
tupp-vill pea jt.) pealekasvanud turbakihis uued juured (Valk, 2005).
Elus turbasamblakate on rabataimede seemnelisele uuendusele väga
ebasoodsaks substraadiks. Selle tõenäosteks põhjusteks on 1) trubasamblakatte
kohevus ja lagundunud taimeosade, seega ka mineraalsete toitesoolade vähesus, 2)
HEDVRRGXV�YHHUHåLLP�– pinnalt läbikuivamine, alumises osas lii gniiskus, 3) pidev
juurdekasv; võimalik, et pidurdavateks teguriteks on ka happene reaktsioon ja nõrk
nitrifikatsioon (Masing, 1958). Osa turbasammalde pinnale langenud seemneid idaneb
pinnal, kuid sureb tõusmena; suurem osa mattub aga turtbasammalde tiheda muru

12
sisemusse. Kohtades, kus turbasammaldest kate on nõrgenenud või hoopis hävinud,
leidub suuremal hulgal rabataimede seemneid. Seejuures rikkalikum seemnevaru
niiskemates kohtades, mis saavad ka kuivalembese mättataimede uuenduskolleteks
(Masing, 1958).
Sammalde elustrateegia esineb suuresti soontaimede omast. Turbasammalde
puhul toimub fotosüntees ja mineraaltoitainete omastamine kogu taime pinnaga, kuid
soontaimede puhul on need kaks tegevust eraldatud juurte ja varte vahel.
Turbasamblad on võimelised kasvama niikaua kui taime pind pole külmunud ja
kapiitulum on varustatud vajaliku niiskusega. Soontaimede puhul on juurdekasv
pii ratud paari kuuga nende varajasel vegetatsiooni perioodil . Selli ne erinevus kasvus
kergendab nende kooseksisteerimist (Malmer et.al., 2003). Siiski on trubasammaldel
pigem negatiivne mõju soontaimede elule sookooslustes.
Tihe turbasammalde vaip on kõige ohtlikum tõusmetele ja noortaimedele,
mille aastane juurdekasv on väike, samas kui turbasammalde juurdekasv võib aastas
küündida kuni 65 millimeetrini (Valk, 2005). Puurinde uuendust takistab otseselt
mitte turbasammalde juurdekasv, vaid selle toitainetevaesus ja kohevus (Masing,
1958). Rohu – puhmarinde taimedele pole niivõrd ohtlik turbasamblaiikide suur
lineaarne juurdekasv, kuivõrd juurdekasvava osa iseloom (selle tihedus jt. omadused).
Selletõttu pole kii rekasvulised älvestes kasvavad turbasamblad nii hukatuslikud, kui
tihedaid mättaid moodustavad lii gid, eriti pruun turbasammal (Sphagnum fuscum),
vähemal määral ka teravalehine (Sphagnum acutifolium), punane ja lill akas
turbasammal (S. rubellum ja S. magellanicum) (Masing, 1958).
Samas esineb ka soontaimede lii ke, millele on turbasamblavaip eelistatumaks
substraadiks, näit. jõhvika- ja huulheinalii gid; nende esinemine turbata aladel on kas
sekundaarne või ajutine nähe. Turbasammalde kate näib oma kohevuse tõttu olevat
soodsaks kasvukohaks ka niisugustele võsundili stele rabataimedele, nagu murakas,
rabakas ja ka mudatarn (Masing, 1958).
Kui soontaimed, omastades vajaliku koguse toitaineid mineralisatsioonist, on
võimelised säilit ama piisava produktiivsuse, mis pii raks turbasammalde kasvu, siis on
nad võimelised ära hoidma turba akumulatsiooni tõusu. Tihedad soontaimede varred
madalsoodes aitavad kaasa madala turba akumulatsioonitaseme säilit amisele ja
sellega vältida raba moodustumist (Malmer et.al., 2003).

13
Ka soontaimed võivad kujundada kasvukeskkonda turbasammalde jaoks
soodsamaks või ebasoodsamaks. Tiheda rohu-puhmarinde all on valgustus nõrgem.
Eriti tugevat varju annavad suured tupp-vill pea puhmikud ja tihe kanarbikurinne, mis
teevad võimatuks valguslembeste turbasammlade kasvu nende all . Mahalangenud
lehed, eriti muraka laiad lehelabad, varjutavad nende alla mattuvaid turbasamblaid,
kutsudes esile nende elutegevuse intensiivsuse muutusi, mis avaldub lehtede värvi
muutumises: turbasamblalii gid, mis valguse käes on punased, muutuvad varjus
rohelisteks. Tihe sookailu puhmastik soodustab metsalehtsamblalii kide kasvu ja
vähendab valguslembestele turbasammalde kasvu (Masing, 1958).
Puurinde mõju suurus on rabades väga varieeruv olenevalt puude mõõtmeist ja
arvust pinnaühikule. Juba üksikud puud lagedal rabal mõjutavad alumisi rindeid,
peamiselt varjamise ja aineringete kaudu (Masing, 1958). Väikesed tihedad
rabamännidki põhjustavad valguslembeste turbasammalde kadumist puu võra all j a
nende asendumist aja jooksul pärislehtsammaldega, peamisel harili ku palusambla
(Pleurozium schreberi) ja lainja kaksikhambaga (Dicranum polysetum) (Masing,
1958).
1.8 Turbasammalde tähtsus loodu ses
Nagu taimedki kasvavad enamus samblaid tipust, kuid mis teeb turbasamblad
omapäraseks on see, et surnud materjali kõdunemine on ebatavaliselt aeglane ja seega
surnud taimed akumuleeruvad turbana (Clymo & Hayward, 1982). Turbasamblad on
soodes peamised turbamoodustajad, rabas taimekoosluse dominanttaimed –
edifikaatorid, mis mõjustavad teiste taimede elu- ja keskkonnatingimusi (Valk, 2005).
Ükski sammalde perekond ei ole majanduslikult nii tähtis ja ökoloogili selt nii
domineeriv maailmas kui turbasamblad (Andrus, 1986). Turvast on intensiivselt
kaevandatud ja kasutatud aianduses, kütusena, biofilt ratsioonil , isoleerimiseks,
absorptsiooniks ja maitseainena Šoti viskis. Paljudes kohtades Euroopas ja Põhja –
Ameerikas suur enamus turbasambla rabasid on hävitatud turba eemaldamise tõttu ja
kuivendustööde tõttu.Võib väita, et turbasamblad on globaalselt tähtsad, nii
ökoloogili selt kui majanduslikult (Sundberg, 2000).

14
Eesti rabades leiduv vähelagunenud samblaturvas on tähtis ekspordiartikkel. Hea
veeimavuse tõttu kasutatakse seda kasvusubstraadina kurkide, tomatite ja lill ede
kasvatamisel (Kink et. al., 1998). Vähelagunenud turvast toodetakse nii frees- kui
plokkturbana, millest just viimane on hinnatud välisriikides. Aastatoodang on Eesti
Turbalii du 1997.a. andmeil u. 0,65 milj . tonni (Kink et. al., 1998).
Boreaalsed sood sisaldavad umbes 300 – 455 Gt süsinikku, mis on võrdne 20 –
30 % globaalse süsiniku sisaldusega. Enamus sellest süsinikust sisaldub sfagnumi-
turbas ja on arvatud, et turbasammaldes sisaldub rohkem süsinikku kui üheski teises
taimeperekonnas (Clymo ja Hayward, 1982).
Põhjapoolkera turbaalad sekvesteerivad umbes 12% inimese tekitatud süsiniku
emissioonidest aastas. Kuna turbasamblad kõdunevad aeglasemalt kui soontaimed,
siis süsiniku tasakaal turbaaladel sõltub sfagnumiturba olemasolust ja seega on tundlik
muutustele turbasammalde ja soontaimede vahel (Limpens et. al., 2004).
Inimese tegevus on mõjutanud soode arengut läbi aegade , kasvanud on
lämmastiku ja väävli depositsioon ja temperatuur (Gunnarsson et.al., 2004), samuti on
soid kuivendatud ja turvast kaevandatud. 19. sajandi keskel hakati Eestis soid
kuivendama nii põllumajanduse tarbeks, metsakasvutingimuste parandamiseks kui
turba kaevandamiseks. Esimesed teadaolevad sookraavitused tehti Eestimaal juba 17.
sajandil . Kuivenduse tulemusel intensiivistub puude kasv, väheneb maapinnale
langeva valguse hulk ning rabas alaneb veetase. Kõige selle tagajärjel hävivad
turbasamblad ja katkeb rabanõlvadel turba akumulatsioon (Ilomets, 2005). Nii on
kuivendamise tõttu hävinud ka Eesti ainus teadaolev Ångstroemi turbasambla
(Sphagnum aongstroemii ) leiukoht Käsmu poolsaarel (Lill eleht, 1998).
2. Kaitsealuste turbasammalde levik
Sammalde kaitsmise ajalugu ei ole nii pikk nagu pilkupüüdvamate ja seetõttu
sageli ka ohustatumate soontaimede puhul. Esimene ohustatud sammaltaimede
nimestiku käsikiri koostati alles 1987. aastal. Riikliku kaitse alla võeti samblad
esmakordselt 1994. aastal kinnitatud looduskaitseseadusega (Vellak, 2005).

15
Eesti rabades on turbasamblad kõige suurema katteväärtusega ja kõige
sagedamini esinevad taimed. Turbasamblad kujundavad ka ise ümbritsevaid
keskkonnatingimusi, tugevdavad ja kii rendavad soostumisprotsesse, uute soode,
soostike ja rabamassiivide teket (Valk, 2005). Samas esineb turbasammalde hulgas ka
lii ke, mida leidub vaid vähestes kasvukohtades ja on väga nõudlikud oma
kasvutingimuste suhtes. Kolmekümne seitsmest praegu Eestist teadaolevast
turbasamblalii gist võib tavaliseks pidada vaid 15 (Laasimer et. al., 1954). Üheksal
lii gil on teada vähem kui 12 leiukohta ning need kuuluvad Eestis haruldaste lii kide
hulka (Vellak, 2004).
Eesti punasesse raamatusse on kantud seitse turbasammalt (Lill eleht, 1998).
1994. aastal vastu võetud kaitstavate loodusobjektide seadusega võeti kaitse alla II
kaitsekategooria lii gina ka üks turbasamblalii k – Lindbergi turbasammal (Sphagnum
lindbergii ) (Ingerpuu & Vellak, 1995). Alates 2004. aastast kuulub looduskaitse alla
neli lii ki turbasamblaid, li saks eelpoolnimetatule veel – loigu turbasammal
(Sphagnum inundatum), viierealine turbasammal (Sphagnum quinquefarium) ja Wulfi
turbasammal (Sphagnum wulfianum). Praegu kuuluvad kõik need lii gid kaitsealuste
lii kide III kategooriasse (Riigi Teataja, 2004).
Looduskaitseseaduse kohaselt III kaitsekategooriasse arvatakse:
1) lii gid, mille arvukust ohustab elupaikade ja kasvukohtade hävimine või rikkumine
ja mille arvukus on vähenenud sedavõrd, et ohutegurite toime jätkumisel võivad nad
sattuda ohustatud lii kide hulka;
2) lii gid, mis kuulusid I või II kaitsekategooriasse, kuid on vajalike kaitseabinõude
rakendamise tõttu väljaspool hävimisohtu (1).
Lindbergi turbasambla kaitsekategooria muutmine ongi tulenenud sellest, et
lii gile on seadustevahelisel perioodil li sandunud uusi leiukohti, nagu nähtub
registreeritud herbaarandmetest.
Kümme aastat tagasi lähtuti lii kide kaitsekategooria määramisel peamiselt
nende esinemissagedusest ja ohustatusest Eestis ja lähiümbruses. Nüüdse nimestiku
koostamisel pöörati enam tähelepanu sammaldele, kes on ohustatud ja mida peetakse
kaitset väärivaks Euroopas laiemalt: kuuluvad direktiividega kaitstavate lii kide eri
nimistutesse ja/või Euroopa sammalde punasesse raamatusse. Samas lülitati
kaitsenimistusse lii ke, mille levik Eestis on aja jooksul tugevasti vähenenud või mis

16
on meil oma levila pii ril . Oluliseks peeti ka seda, kas vastava lii gi kaitse on tegelikult
võimalik (Vellak, 2005).
Samblaid saab kaitsta eeskätt nende kasvukohtade kaitsmise kaudu. Nii on
kõik turbasamblad lülitatud Euroopa Loodusdirektiivi V lisassse, mis reguleerib
lii kide elupaikade säästlikku kasutamist (EU Habitats Directive, 1992). Paljud
samblalii gid võivad võrdsel määral kasvada väga erinevates biotoopides, kuid on
lii ke, mis on väga nõudlikud ja seetõttu kergesti kaduvad juba väikeste muutuste läbi
kasvukoha ökoloogili stes tingimustes. Nii võib Eesti floorast kadunuks pidada
Ångstroemi (S. aongstroemii ) ja kõrv-turbasammalt (S. auriculatum), sest nende
teadaolnud ainsad kasvukohad Käsmu poolsaarel on tänaseks hävinud (Ingerpuu &
Vellak, 1995).
Allj ärgnevalt iseloomustatakse nelja Eestis kaitse all oleva turbasambla
levikut, lii gi ökoloogili si nõudlusi ning kaitse alla võtmise põhjendatust.
2.1 Loigu-turbasammal - Sphagnu m inund atum Russ.
2.1.1 Levik
Loigu turbasammal on üks seitmsest E. Russowi poolt kirjeldatud lii gist. Liiki
kirjeldab ta esmakordselt kahe turbasammalde keeruka sektsiooni taksonoomiat
käsitlevas töös (Russow, 1894). Läbi aja on lii k leidnud käsitlemist nii i seseisva
lii gina kui ka koos kõrv-turbasamblaga (S. auriculatum Schimp.) nimetuse all S.
denticulatum Brid., ka Eesti nimestikes on ta olnud erinevate nimetuste all (Laasimer
jt., 1954; Ingerpuu jt., 1994). Norra sfagnoloogi K. Flatbergi taksonoomiliste tööde
põhjal (Flatberg, 1985) eelistatakse siiski E. Russowi käsitlust antud lii gi kohta.
Liik kasvab merelise kliimaga pii rkondades (Ulvinen, et. al., 2002). Liigi
üldareaal hõlmab põhjapoolkera (Hiina, Jaapan, Euroopa ja Põhja-Ameerika),
lehtmetsavööndi põhjaosa ja okasmetsavööndi (Ulvinen, et. al., 2002). Skandinaavias
esineb regiooni lõuna- ja keskosas (Joonis 1). Soomes esineb põhili selt lõunaosas
(Ahvenamaal, Varsinais- Suomi maakonnas, Lõuna- Karjalas). Loigu-turbasammal on
intensiivse sookuivenduse tagajärjel muutunud mitmes pii rkonnas haruldaseks ning
võetud kaitse alla. Soome punases raamatus (Rassi et al., 2001) kuulub lii k peaaegu

17
ohustatud lii kide kategooraisse, kuna lii gi levik on viimastel aastalel vähenud ning
peamisteks ohuteguriteks peetakse just keskkonna saatatuse tõusu, Tšehhis on ta
WlKHOHSDQXYllULYDWH�OLLNLGH�QLPLVWXV��.XþHUD��9i�D��������/ätis on lii k haruldane
(2), Leedus on lii gi levik ebaselge (Jukoniene, 2003).
Eestis lii gil on teada vaid kuus leiukohta, neist kolm on suhteliselt vanad leiud
(Joonis 2), seega on lii k Eestis haruldane (Tabel 1, Ingerpuu jt., 1998). Väheste leiude
arvu tõttu ja kasvukohatingimuste suhtes nõudliku lii gina on loigu-turbasammal
arvatud Eestis looduskaitsealuste lii kide hulka.
Tabel 1. Loigu-turbasamba Eesti leiukohtade andmed.
Leiukoht Leidja Aasta Alli kas
Pärnumaa, Sindi S. Talts 1949 TAA
Pärnumaa, Kesu raba E. Varep 1954 TAA
Põlvamaa, Rainova ja Võõpsu vahel H. Krall 1957 TAA
Läänemaa, Vensi soo L. Laasimer 1981 TAA
Põlvamaa, Nohipalu, Valgjärv A. Mäemets 1994 TAA
Saaremaa, Kihelkonna vald, Viki ja
Taavi vahel
N. Ingerpuu 1997 TAA (2 eks)
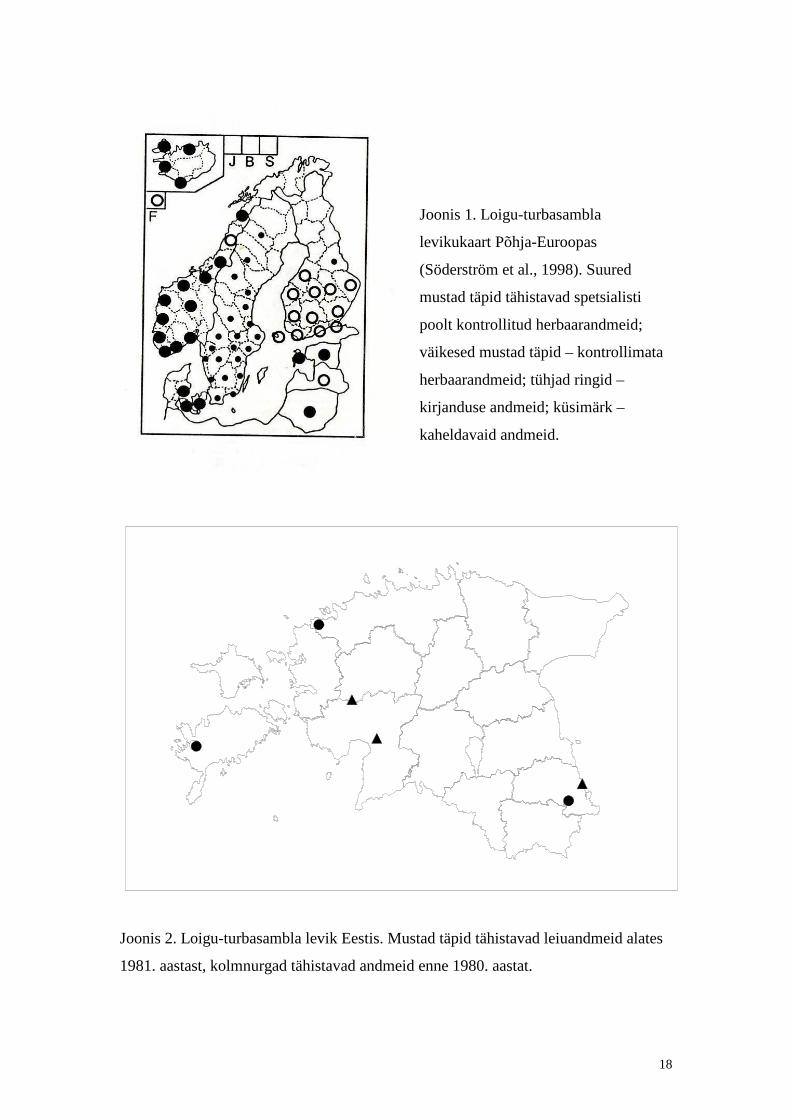
18
Joonis 1. Loigu-turbasambla
levikukaart Põhja-Euroopas
(Söderström et al., 1998). Suured
mustad täpid tähistavad spetsialisti
poolt kontrollit ud herbaarandmeid;
väikesed mustad täpid – kontrollimata
herbaarandmeid; tühjad ringid –
kirjanduse andmeid; küsimärk –
kaheldavaid andmeid.
Joonis 2. Loigu-turbasambla levik Eestis. Mustad täpid tähistavad leiuandmeid alates
1981. aastast, kolmnurgad tähistavad andmeid enne 1980. aastat.

19
2.1.2 Kasvukohad ja ökoloog ia
Loigu-turbasammal on kasvukohtade suhtes nõudlik, eelistades kasvada
märgadel üleujutatavail rabaniitudel, lodudes, turbaaukudes, rabalaugastes (Laasimer
et. al., 1954), sageli ka vees (Ingerpuu jt., 1998). Loigu-turbasammal on väga
heterogeenne lii k; olenevalt kasvukoha niiskusemäärast võivad taimed olla varieeruva
suurusega, oksalehtede asetusega ning avade hulgaga neis. Nende ökoloogili ste
erisuste alusel on eristatud mitmeid vorme (Laasimer et. al., 1954).
2.2 Lindb ergi turbasammal – Sphagnu m lindb ergii Schimp. ex
Lindb .
2.2.1 Levik
Lindbergi turbasammal on tsirkumpolaarse areaaliga. Skandinaavias ja
Venemaal on ta levinud peamiselt boreaalses ja arktili ses vööndis (Joonis 3). Lõuna
pool põhilevilast on vaid üksikuid leiukohti Läänemereäärsetel tasandikel ja Šotimaa
ning Kesk – Euroopa mägedes (Stamer, 1998). Soomes on Lindbergi turbasammal
ohustatud (Rassi et al., 2001). See turbasambla lii k kuulub ka Leedu sammalde
punasesse raamatusse ohustatud lii kide kategooriasse, kuna teadaolevaid leiukohti on
Leedus vaid üks (Jukoniene, 1996) ning samas kategoorias ka Läti punases raamatus
��EROLQD���������7ãHKKL�SXQDVHVVH�UDDPDWXVVH�RQ�DUYDWXV�/LQGEHUJL�WXUEDVDPPDO�NXL�WlKHOHSDQX�YllULY��NXLG�VHQL�YHHO�PLWWH�RKXVWDWXG�VRROLLN��.XþHUD��9i�D�������
Eestis on lii gil teada vaid kümme leiukohta, neist kuus on suhteliselt vanad
leiud (Joonis 4), seega on lii k meilgi suhteliselt haruldane (Tabel 2). Lindbergi
turbasammalt ohustab soode kuivendamine ja turbavõtmine ja seetõttu on ta li satud ka
Eesti punasesse raamatusse (Lill eleht, 1998), kui haruldane lii k. Samas on teada aga,
et see põhjapoolsematel aladel tavaline lii k levib lõunapoole, asustades just vanu
turbavõtuauke (Lönell et. al., 1998). Kuna lii k kasvab raskesti lii gipääsetavatel
märgaladel, on lii gi levikut Eestis raske kontrolli da. Uued lisandunud leiukohad ei
tähenda veel lii gi leviku laienemist Eestis, vaid nendel aladel pole varasematel
20
aegadel li htsalt käidud. Samas ei pruugi vanadest leiukohatdest lii ki enam üles leida
(Vellak jt., 1996).
Tabel 2. L indbergi turbasamba Eesti leiukohtade andmed.
Leiukoht Leidja Aasta Alli kas
Läänemaa, Laiküla J.Mikutowicz 1904 TU
Pärnumaa, Ruunasoo R. Stamer 1998 Stamer, 1998
Ida-Virumaa, Tärivere E. Parmasto 1950 TAA (2 eks.)
Ida-Virumaa, Raja soo E. Parmasto 1950 TAA
Hiiumaa, Partsi soo S. Talts 1951 TAA
Sutlepa M. Kask 1952 TAA (2 eks.)
Jõgeva, Endla LKA, Männikjärve raba R. Stamer 1996 TAA (4 eks.)
Pärnumaa, Kahvena raba N. Ingepuu 2000 TU
Pärnumaa, Tolkuse raba K. Vellak 2004 TU
Raplamaa, Atla raba T. Ploompuu 2006 e-mail
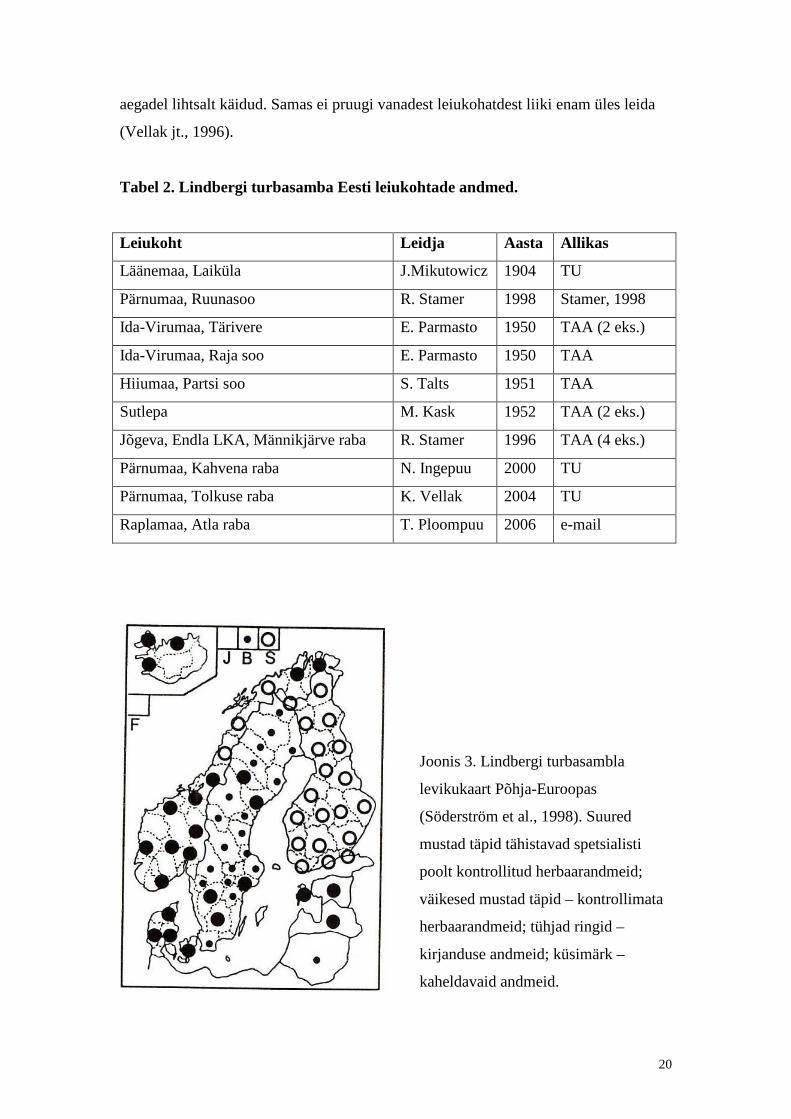
Joonis 3. Lindbergi turbasambla
levikukaart Põhja-Euroopas
(Söderström et al., 1998). Suured
mustad täpid tähistavad spetsialisti
poolt kontrollit ud herbaarandmeid;
väikesed mustad täpid – kontrollimata
herbaarandmeid; tühjad ringid –
kirjanduse andmeid; küsimärk –
kaheldavaid andmeid.
21
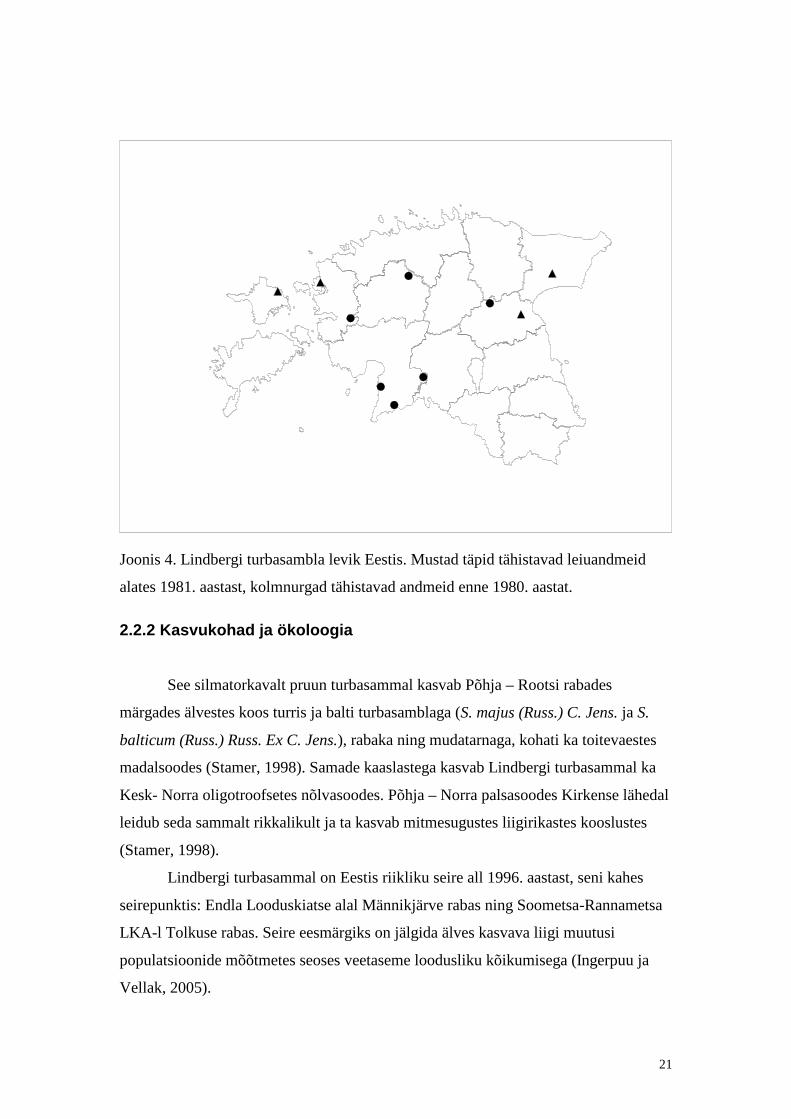
Joonis 4. Lindbergi turbasambla levik Eestis. Mustad täpid tähistavad leiuandmeid
alates 1981. aastast, kolmnurgad tähistavad andmeid enne 1980. aastat.
2.2.2 Kasvukohad ja ökoloog ia
See silmatorkavalt pruun turbasammal kasvab Põhja – Rootsi rabades
märgades älvestes koos turris ja balti turbasamblaga (S. majus (Russ.) C. Jens. ja S.
balticum (Russ.) Russ. Ex C. Jens.), rabaka ning mudatarnaga, kohati ka toitevaestes
madalsoodes (Stamer, 1998). Samade kaaslastega kasvab Lindbergi turbasammal ka
Kesk- Norra oligotroofsetes nõlvasoodes. Põhja – Norra palsasoodes Kirkense lähedal
leidub seda sammalt rikkalikult ja ta kasvab mitmesugustes lii girikastes kooslustes
(Stamer, 1998).
Lindbergi turbasammal on Eestis rii kliku seire all 1996. aastast, seni kahes
seirepunktis: Endla Looduskiatse alal Männikjärve rabas ning Soometsa-Rannametsa
LKA-l Tolkuse rabas. Seire eesmärgiks on jälgida älves kasvava lii gi muutusi
populatsioonide mõõtmetes seoses veetaseme loodusliku kõikumisega (Ingerpuu ja
Vellak, 2005).
22
2.3 Viierealine turbasammal - Sphagnu m quinqu efarium
(Lindb . ex Braithw.) Warnst
2.3.1 Levik
Viierealine turbasammal on üldlevikuliselt mereline (Ulvinen, et. al., 2002),
kuid harvaesinev kogu areaali ulatuses (Savitš-Ljubitskaja & Smirnova, 1968). See
lii k on levinud Kaukaasias, Siberis kuni Sahalini saareni. Esineb eelistatult Põhja-,
Kesk – ja atlantili ses Euroopas, Jaapanis ja Põhja – Ameerikas (Laasimer et. al.,
1954).
Skandinaavias levib viierealine turbasammal põhili selt lõunaosades ja puudub
põhjaosadest (Joonis 5, Ulvinen, et. al., 2002). Soomes levib paigutiselt põhili selt
lõunaosas ja ka Rootsis esineb paiguti (Ulvinen, et. al., 2002). Lätis leidub viierealist
turbasammalt üsna harva (8-12 leiukohta) (2). See lii k kuulub Leedu punasesse
raamatusse ebapiisavate andmetega lii kide kategooriasse, kuna lii gil on teada vaid
kaks leiukohta (Jukoniene, 1996).
Eesti punase raamatu ilmumise ajal oli lii gil teada vaid üks vana leiukoht ning
lii k arvati punase raamatu määratlemata staatusega lii kide kategooriasse (Lill eleht,
1998). Käesolevaks ajaks on herbaarandmete põhjal selgunud, et lii k on Eestis siiski
olemas kuid, väga haruldane, esinedes vaid kolmes leiukohas (Tabel 3, Joonis 6).
Tabel 3. Viierealise turbasamba Eesti leiukohtade andmed.
Leiukoht Leidja Aasta Alli kas
Saaremaa, Vedruka raba H. Tuvikene 1948 TAA
Lääne–Virumaa, Lahemaa RP, Oandu N. Cronberg 2003 TAA (1 eks.), TU
(2 eks.)
Hiiumaa, Kõpu poolsaar, Mägipe K. Vellak 2004 TU
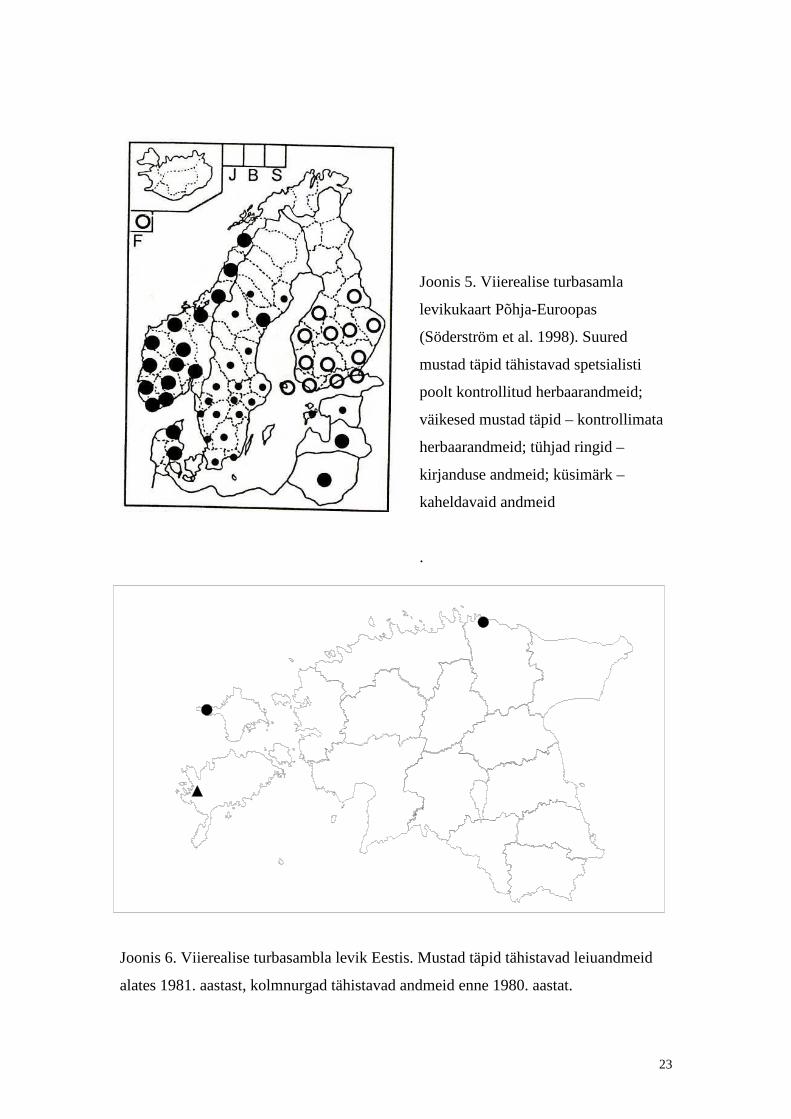
23
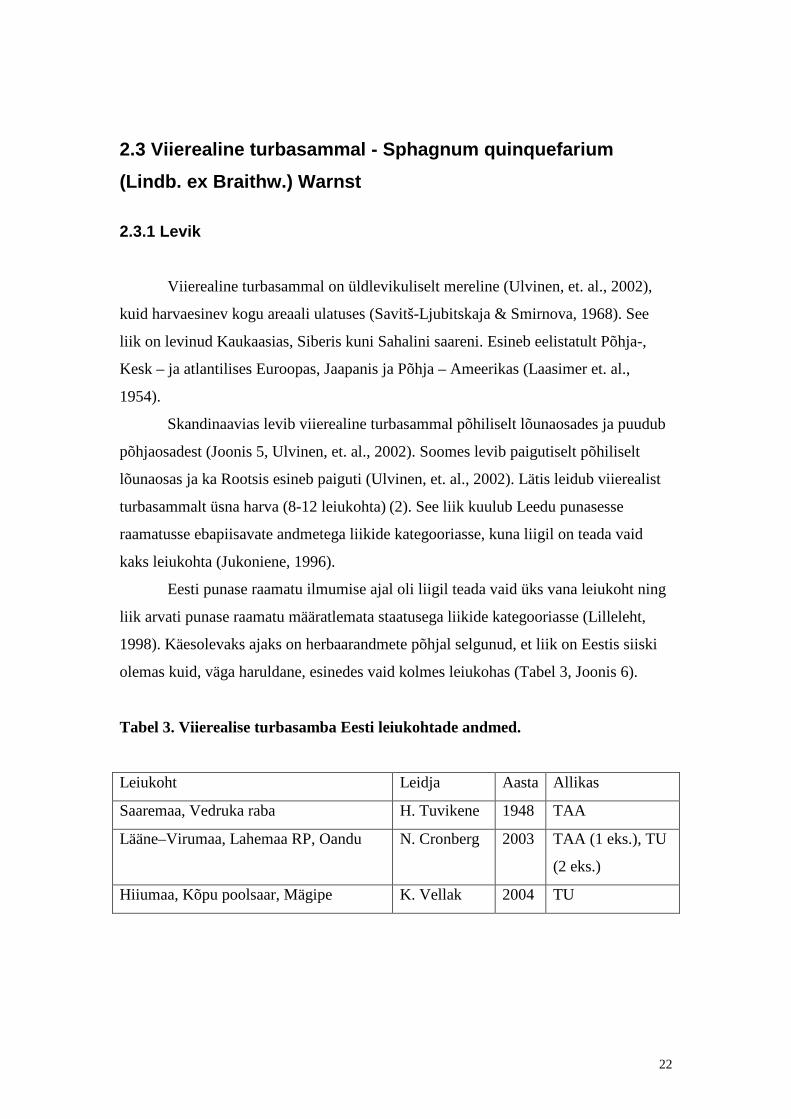
Joonis 5. Viierealise turbasamla
levikukaart Põhja-Euroopas
(Söderström et al. 1998). Suured
mustad täpid tähistavad spetsialisti
poolt kontrollit ud herbaarandmeid;
väikesed mustad täpid – kontrollimata
herbaarandmeid; tühjad ringid –
kirjanduse andmeid; küsimärk –
kaheldavaid andmeid
.
Joonis 6. Viierealise turbasambla levik Eestis. Mustad täpid tähistavad leiuandmeid
alates 1981. aastast, kolmnurgad tähistavad andmeid enne 1980. aastat.

24
2.3.2 Kasvukohad ja ökoloog ia
Erinevalt eelmistest kaitsealustest turbasammaldest on viierealine
turbasammal märgade metsade asukas, eelistades kasvada rabastuvates ja soostuvates
okasmetsades (Ingerpuu jt., 1998). Kasvukoha tingimuste suhtes nõudlik, kasvab
toitaineterikkamas, pika metsajärjepidevusega okasmetsades ning on tundlik nii
kuivenduse kui metsaraie suhtes (Ulvinen, et. al., 2002).
2.4 Wulfi turbasammal – Sphagnu m wulfianum Girg.
2.4.1 Levik
Wulfi turbasammal on lii k, mille tüüpmaterjal pärineb Eestist. Selle kirjeldas
Tartu lähedalt Tähtvere rabast kogutud materjali põhjal hobi-brüoloog K. Girgensohn,
nimetades selle maaomaniku nime järgi Wulfi turbasamblaks (Girgensohn, 1860).
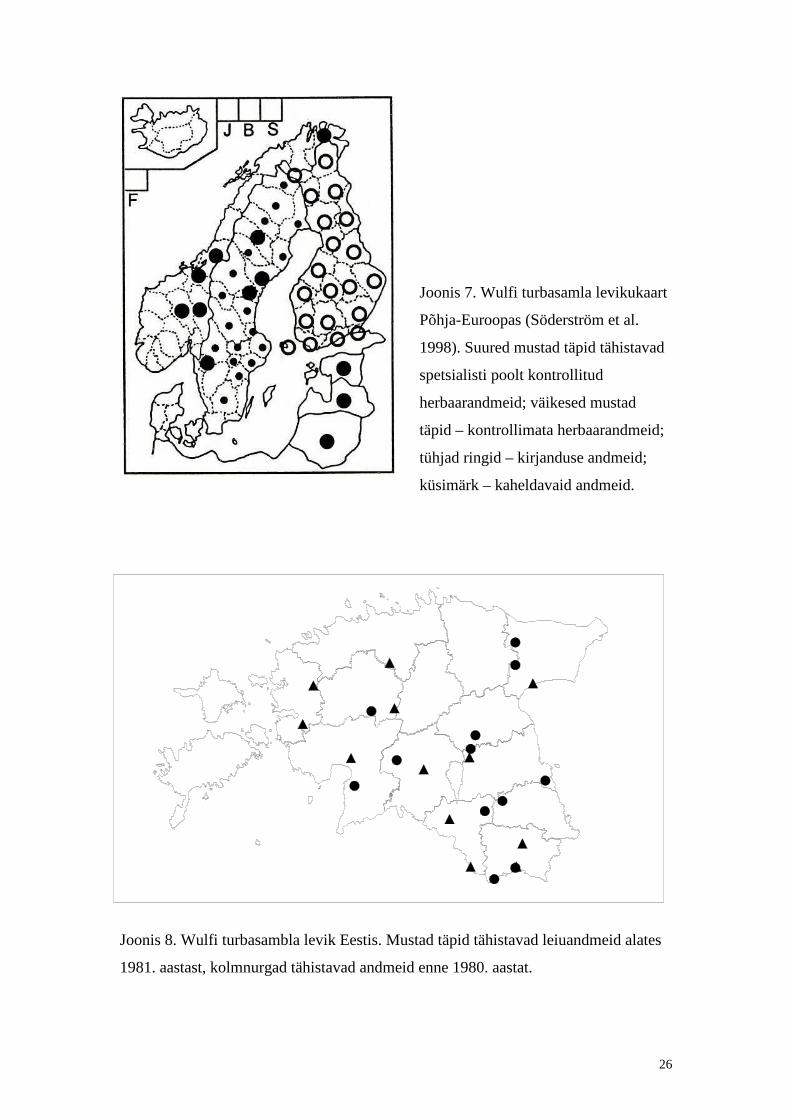
Liigi üldlevikuala on peamiselt mandrili ne. See lii k levib Euroopa põhja- ja
idaosas, peale selle Põhja – Ameerikas ja Aasias (Laasimer et. al., 1954).
Skandinaavias levib põhili selt põhjaosas (Joonis 7). Soomes levib see lii k kogu üle
maa ja on küllaltki sagedane (Ulvinen, et. al., 2002). Samas puudub see lii k Taanis
(Ulvinen, et. al., 2002) ning Norras on ta väheneva areaaliga ning arvatud tähelepanu
väärivate lii kide hulka nende punases raamatus (3).
Wulfi turbasammal esineb Eestis pill atult (Ingerpuu jt., 1998). Eestis on teada
selle lii gi leiukohti kakskümmend neli , millest kolmteist on suhteli selt vanad leiud
(Tabel 4, Joonis 8). Wulfi turbasammal kuulub Leedu punasesse raamatusse
kolmandasse kategooriasse, ning teadaolevalt on Leedus kaks leiukohta (Jukoniene,
1996). Lätis on Wulfi turbasammal oma levikult üsna sage (2).
Tabel 4. Wulfi turbasamba Eesti leiukohtade andmed.
Leiukoht Leidja Aasta Alli kas
Valgamaa, Taheva vald, Laanemets S. Talts 1949 TAA (3 eks.)
Tartumaa, Laeva maakond T. Aimla 1949 TAA
25
Pärnumaa, Rääma raba S. Talts 1949 TAA
Võrumaa, Reedu – Kubja vahel S. Talts 1950 TAA
Läänemaa, Marimetsa raba A. Kalda 1952 TAA
Raplamaa, Alli pa raba E. Varep 1952 TAA
Järvamaa, Lõõra soo S. Talts 1952 TAA
Võrumaa, Luhasoo L. Vilj asoo 1954 TAA
Valgamaa, Tõrva 1954 TAA
Vilj andimaa, Vilj andi Umbjärv V. Masing 1969 TU (2 eks.)
Harjumaa, Ämmusoo V. Masing 1969 TU (5 eks.)
Ida – Virumaa, Lohusuu E. Varep TAA
Tartumaa, Järvselja ürgmets K. Vellak 1983 TAA
Tartumaa, Kursi maakond K. Vellak 1994 TAA
Ida – Virumaa, Oonurme K. Vellak 1996 TAA
Tartumaa, Alam –Pedja LKA N. Ingerpuu 1996 TAA
Vilj andimaa, Soomaa RP K. Vellak 1998 TAA
Ida –Virumaa, Maidla maakond N. Ingerpuu 1998 TAA
Võrumaa, Krabi, Kellamäe N. Ingerpuu 2000 TAA
Põlvamaa, Krüüdneri N. Ingerpuu 2001 TU
Võrumaa, Vastse – Roosa, Peetri jõe
ääres
K. Vellak 2002 TAA
Pärnumaa, Surju maakond K. Vellak 2003 TAA
Raplamaa, Nõlvasoo raba N. Ingerpuu 2005 TU
Valgamaa, Otepää Looduspark M. Leis 2005 TU
26
Joonis 7. Wulfi turbasamla levikukaart
Põhja-Euroopas (Söderström et al.
1998). Suured mustad täpid tähistavad
spetsialisti poolt kontrollit ud
herbaarandmeid; väikesed mustad
täpid – kontrollimata herbaarandmeid;
tühjad ringid – kirjanduse andmeid;
küsimärk – kaheldavaid andmeid.
Joonis 8. Wulfi turbasambla levik Eestis. Mustad täpid tähistavad leiuandmeid alates
1981. aastast, kolmnurgad tähistavad andmeid enne 1980. aastat.

27
2.4.2 Kasvukohad ja ökoloog ia
Wulfi turbasammal kasvab varjulistes, küllalt kuivades kohtades rabastunud
sega- ja okasmetsades, harvem ka soostunud metsades (Ingerpuu jt., 1998).
Inimpelgliku lii gina on ta tundlik nii kuivenduse kui ka metsaraie suhtes (Vellak,
2004).
Eestis on just rohkete vanade leiukohatde arv ning inimtegevuse suhtes
tundliku märgade metsade asukana võib lii k kasvukohtade tingimuste muutuste tõttu
kergesti oma leiukohtadest kaduda. Selleks, et kindlaks teha, mis seda lii ki kõige
enam ohustab, alustati 2003. aastal lii gi seirega. 2005. aastal li sandus esimesele alale
veel üks (Ingerpuu, Vellak, 2005).

28
Kokkuvõte
Antud töö eesmärgiks oli anda ülevaade turbasammaldest ja nende
ökoloogiast, selgitada nende suhteid soontaimedega ning iseloomustada Eesti
looduskaitsealuste turbasammalde seisundit ja levikut. Töö praktili seks osaks oli
koostada Eesti nelja kaitsealuse turbasamblalii gi levikukaardid herbaarandmete põhjal
ning sisestada levikuandmed andmebaasi.
Herbaarandmete põhjal on loigu-turbasamblal kuus leiukohta, Lindbergi
turbasamblal kümme leiukohta ning Wulfi turbasamblal isegi koguni kakskümmend
neli l eiukohta. Kahjuks on paljud nende lii kide varasemad leiukohad ebaselged või
hävinud. Kuigi viierealisel turbasamblal on teada vaid kolm leiukohta, on neist kaks
leitud viimastel aastatel ja see kinnitab lii gi püsimist Eesti floras.
Lindbergi turbasammal ja Wulfi turbasammal on võetud seire alla kaitsealuste
taimelii kide rii kliku seire programmi raames vastavalt aastast 1996 ja 2003.
Turbasammaldele on kõige suuremaks ohuks soode kuivendamine ja
turbavõtmine ja seetõttu on Lindbergi turbasammal li satud ka Eesti punasesse
raamatusse ja samuti ka Läti ja Leedu punasesse raamatusse.
Turvas on Eesti üks väärtuslikemaid maavarasid: maailmas on vähe riike, kus
teda leiduks sama palju kui meil . Meiegi turbarabasid ähvardab kuivendamine ja
ülekaevandamine ning seegasäilit ades soid ja soiseid metsakooslusi, tagatakse ka
kaitsealuste turbasammalde püsimine Eesti flooras.

29
Summary
The aim of the present study was to give a survey about peat mosses and their
ecology, to explain interferences between Sphagnum species and vascular plants and
to characterize Estonian peat mosses that are included in the third category of the
protected plant species of Estonia. This study’s practical part was to compile the
distribution maps for these species according to herbarium data and supplement
database of Estonian protected bryophyte species.
There are six localiti es for Sphagnum inundatum, ten localiti es for Sphagnum
lindbergii and twenty-four localiti es for Sphagnum wulfianum, but only less than half
of them are discovered recently. Although there are only three localiti es for
Sphagnum quinquefarium, two of them are found lately and that proves the species
persistence in Estonian flora.
The biggest threat on peat mosses is drainage mires and wet forests, and
drainage is one of the reasons why Sphagnum lindbergii is included to the red list of
Estonian bryophytes and also to Lithuania and Latvia’s red lists.
Sphagnum lindbergii and S. wulfianum are under the state monitoring program
of protected species since 1996 and 2003 respectively.
Peat is one of the most valuable natural resource of Estonia; there are few
countries where are so many peat resources as in Estonia. But amelioration and peat
cutting is the biggest threat to our peatlands. Strict rules for peatlands protection help
to protect species inhabit there as well .

30
Kasutatud kirjandu s
�EROLQD��$��������/DWYLMDV�UHW�V�XQ�DL]VDUJ�MDP�V�VÌQDV��9LGHV�DL]VDUG]¯ED�/DWYLM�����5¯JD�
Andrus, R. E. 1986. Some aspects of Sphagnum ecology. Can. J. Bot. 64: 416 - 426.
Clymo, R. S., Hayward, P. M. 1982. The ecology of Sphagnum. Smith, A. J. E.
(ed.), Bryophyte ecology. Chapman and Hall , 229 – 289.
EU Habitats Directive, 1992. EU Habitats Directive (92/43/EEC). Consolidated
Text. Off ice for Off iacial Publication of The European Union. CONSLEG:
1992LOO43-01.05-2004.
Flatberg, K.I . 1985. Taxonomy of crispate morphotypes in Sphagnum Sect.
Seubsecunda. – Lindbergia 11: 99-113.
Gignac, L . D. 1993. Distribution of Sphagnum Species, Communities, and habitats in
Relation to Climate. Advances in Bryology 5: 187 – 222.
Girgensohn, G.K. 1860. Naturgeschichte der Laub- und Lebermoose Liv-, Ehst- und
Kurlands. – Arch. Naturk. Liv-, Ehst- und Kurlands 2(2): 1 - 488.
Gunnarsson, U. 2000. Vegetation changes on Swedish mires. Effects of raised
temperature and increased nitrogen and sulphur influx. Acta Universitatis
Upsaliensis. Comprehensive Summaries of Upppsala Dissertations from the
Faculty of Science and Technology 581. 37 p.
Gunnarsson, U., Rydin, H. 2000. Nitrogen fertili zation reduces Sphagnum
production in bog communities. New Phytologist 147: 527- 537.
Gunnarsson, U., Granberg, G., Nilsson, M. 2004. Growth, production and
interspecific competition in Sphagnum: effects of temperature, nitrogen and
sulphur treatments on boreal mire. New Phytologist 163: 349 – 359.
Gunnarson, U. 2005. Global patterns of Sphagnum productivity. Journal of Bryology
2005, 27: 269-279.

31
I lomets, M. 1980. Turbasamblakoosluste juurdekasvu mõjustavaist tegureist
Männikjärve rabas. - T. Paaver (toim.) Bioloogili ne produktiivsus ja seda
määravad tegurid, Tartu. 49 - 53.
I lomets, M. 1982. The productivity of Sphagnum communities and the rate of peat
accumulation in Estonian bogs. – In: V. Masing (ed.) Estonian Peatlands. Est.
Contr. IBP, IX, Talli nn, Valgus. 102-116.
I lomets, M. 2003. Mill e arvel kaevandame turvast? Eesti Loodus 2/3: 20-24
I lomets, M. 2005. Eesti soode taastamine – vajadused, printsiibid, hetkeseis. Eesti
Looduseuurijate Aastaraamat 83: 72-95.
Ingerpuu, N. 1994. Igal samblal oma nägu 9. Soosamblad (järg). Eesti Loodus 8:
242.
Ingerpuu, N., Kalda, A., Kannukene, L., Krall , H., Leis, M. ja Vellak, K. 1994. Eesti
sammalde nimestik. List of the Estonian bryophytes. – Abiks Loodusevaatlejale
94: 1-175.
Ingerpuu, N., Vellak, K. 1995. Kaitsealused taimelii gid. Samblad. Eesti Loodus 7:
182-183.
Ingerpuu, N., Vellak, K. 2005. Haruldaste samblalii kide seire. 2005. aasta aruanne.
Käsikiri. Tartu. 41 lk.
Ingerpuu, N. (koost.), Kalda, A., Kannukene, L., Krall , H., Leis, M. ja Vellak, K.
(koost.) 1998. Eesti sammalde määraja. Eesti Loodusfoto, Tartu, 239 lk.
Ingerpuu, N. 2001. Edmund August Fridrich Russow 160. Samblasõber 4: 17 – 19.
Jukoniene, I .,�������/LHWXYRV�NLPLQDL�LU�åDOLRVLRV�VDPDQRV��%RWDQLNRV�LQVWLWXWR�leidykla, Vilnius.
Jukoniene, I . 1996. Rare and threatened bryophyte species in Lithuania. Botanica
Lithuania. 2 (4): 327-341.
Kalda, A. 2000. Viktor Masing - samblal sammuja - 75. Samblasõber 3: 10-11.
Kink, H., Andresmaa, M., Orr u, M. 1998. Eesti soode hüdrogeoökoloogia. Talli nna
Tehnikaülikooli Geoloogia Instituut. Teaduste Akadeemia Kirjastus. Talli nn.
Krall , H., Kukk, T., Kull , T., Kuusk, V., Leht, M., Oja, T., Reier, Ü., Sepp, S.,
Zingel, H., Tuulik, T. 1999. Eesti taimede määraja. EPMÜ ZBI. Eesti
Loodusfoto. Tartu.

32
.XþHUD��-���9i�D��-� 2003. Check- and Red list of bryophytes of the Szech Republic.
– Preslia 75: 193-222.
Laasimer, L ., Talts, S., Varep, E. 1954. Eesti NSV turbasamblad, Eesti rii klik
kirjastus, Talli nn.
Lill eleht, V. (toim.) 1998. Eesti punane raamat. – Tartu
Limpens, J., Berendse, F., Klees, H. 2004. How Phosphorus Availabilit y Affects the
Impact of Nitrogen Deposition on Sphagnum and Vascular Plants in Bogs.
Ecosystems 7: 793 – 804.
Lönell , N., Sundberg, S., Gunnarsson, U. 1998. Björnvitmossa Sphagnum
lindbergii I Södermanland och Uppland. Myrinia 8: 12-14.
Malmer, N. 1993. Mineral Nutrients in Vegatation and Surface Layers of Sphagnum-
Dominanted Peat-Forming Systems. Advances in Bryology 5: 223 – 248.
Malmer, N., Albinsson, C., Svensson B. M., Wallén B. 2003. Interferences
between Sphagnum and vascular plants: effects on plant community structure
and peat formation. Oikos 100: 469 – 482.
Masing, V. 1958. Ida-Eesti rabade taimekooslused ja nende dünaamika.
Kandidaadidisertatsioon. Käsikiri Tartu Ülikooli Teaduslikus Raamatukogus.
Rassi, P., Alanen, A., Kanerva, T. & Mannerkoski, I . (eds.). 2001. The Red List of
Finnish Species. – Ministry of the Environment and Finnish Environment
Institute, Helsinki. 432 pp.
Riigi Teataja 2004. III kaitsekategooria lii kide kaitse alla võtmine. – KKM, RTL
27.05.2004, 69, 31134.
Russow, E. 1894. Zur Kenntnis der Subsecundum- und Cymbifoliumgruppe
europäischer Torfmoose. – Arch. Naturk. Liv-, Ehst- und Kurlands 2(4): 1-167.
Savitš-L jubitskaja, L .I ., Smirnova, Z.N. 1968. Opredelitel’ sfagnovõkh mkhov
SSSR. – Nauka, Leningrad. 109 s.
Stamer, R. 1998. Lindbergi turbasambla uued leiud Eestis. Eesti Looduseuurijate
Seltsi Aastaraamat 78: 49-55.

33
Sundberg, S., Rydin, H. 1998. Spore number in Sphagnum and its dependence on
spore and capsule size. Journal of Bryology 20: 1-16.
Sundberg, S. 2000. The ecological significanceof sexual reproduction in peat mosses
(Sphagnum). Acta Universitatis Upsaliensis. Comprehensive Summaries of
Uppsala Dissertations from the Faculty of Science and Technology 581.
Sundberg, S. 2002. Sporophyte production and spore dispersal phenology in
Sphagnum: the importance of summer moisture and patch characteristics. - Can.
J. Bot. 80: 543-556.
Söderström, L. (ed.), Abolina, A., Blom, H. H., Damsholt, K., Fagersten, R.,
Flatberg, K. I ., Fr isvoll , A. A., Haapasaari, M., Hedenäs, L., Heegaard, E.,
Ingerpuu, N., Johannson, B., Jukoniene, I ., Kannukene, L., Koponen, T.,
Leis, M., Lewinsky-Haapasaari, J., Presto, T., Thingsgaard, K., Ulvinen, T.,
Vellak, K., Vir tanen, R., Weibull , H. 1998. Preliminary Distribution Maps of
Bryophytes in Northwestern Europe.Vol. 3 Musci (J-Z). – Trondheim, p. 1-69.
Truus, L ., Karofeld, E. 2000. Mati Ilomets 50. Samblasõber 3: 12-13.
Ulvinen, T., Syr jänen, K., Antt ila, S. 2002. Suomen sammalet – levinneisyys,
ekologia, uhanalaisuus. Suomen ympäristokeskus. Helsinki.
Valk, U. 2005. Eesti rabad. Ökoloogili s-metsanduslik uurimus, Tartu.
Vellak, K. 2004. Turbasamblad ja valvik. Eesti Loodus 3: 142-143.
Vellak, K. 2005. Samblaid on nüüd kaitse all poole rohkem. Eesti Loodus 8: 46-48.
Vellak, K., Ingerpuu, N., Leis, M., Trass, H., Roosam, A. 1996. Nigula
Looduskaitseala soontaimede, sammalde ja suursamblike lii gili ne mitmekesisus.
26 lk. Käsikiri.
Kasutatud internetialli kad:
1. Elektroonili ne Riigi Teataja. Looduskaitseseadus.
[https://www.riigiteataja.ee/ert/act.jsp?replstring=33&dyn=1&id=893104#pr1
0lg4] 02.05.2006

34
2. Mosses of Latvia.
[http://latvijas.daba.lv/scripts/db/saraksti/saraksti.cgi?d=suunas&l=en]
09.05.2006
3. Red List of threatened mosses in Norway
[http://www.toyen.uio.no/botanisk/mose/red_grup.htm]
11.05.2006





























