Embed Size (px)
Citation preview

Biochimica et Biophysica Acta, 1154 (1993) 183-199 183 © 1993 Elsevier Science Publishers B.V. All rights reserved 0304-4157/93/$06.00
BBAREV 85425
Type I receptors for PACAP (a neuropeptide even more important than VIP?)
Jean Christophe *
Department of Biochemistry and Nutrition, Medical School, Universit~ Libre de Bruxelles, Building G/E, CP 611, 808 Route de Lennik, B-1070 Brussels (Belgium)
(Received 11 May 1993)
Contents
I. Discovery and structure of PACAP . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184
II. The PACAP precursor and the PACAP gene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184
III. Two classes of PACAP receptors and a few receptor subtypes . . . . . . . . . . . . . . . . . . . . . . . . 184
IV. Two effector systems for type I PACAP-preferring receptors . . . . . . . . . . . . . . . . . . . . . . . . . 186
V. The hypothalamo-pituitary complex and the pineal gland . . . . . . . . . . . . . . . . . . . . . . . . . . . . 187
VI. The brain and spinal cord . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 189
VII. Ocular distribution of type I receptors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 190
VIII. The digestive tract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 190
IX. The exocrine pancreas and pancreatic AR 4-2J cell line . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 191
X. The endocrine pancreas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 192
XI. The liver . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 192
XII. The adrenal medulla and the rat pheochromacytoma PCI2 and human neuroblastoma NB-OK cell lines . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 192
XIII. The respiratory tract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 193
XIV. The extra-pulmonary vascular system and heart . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 193
XV. Normal and neoplastic blood cells and fibroblasts . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 194
XVI. The reproductive function . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 194
XVII. Structural requirements for the occupancy of type I receptors coupled to adenylate cyclase activation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 194
* Corresponding author. Fax: + 32 2 5556230. Abbreviations: CGRP, calcitonin gene-related peptide; CRF, corticotropin releasing factor; GH, growth hormone; GIP, gastric inhibitory peptide; GRF, growth hormone releasing peptide; ir, immunoreactive; LH, luteinizing hormone; LHRH = GnRH, gonadotropin releasing hormone; PACAP, pituitary adenylate cyclase activating polypeptide; PHV, peptide histidine valinamide; PKC, protein kinase C; PLC, phospholipase C; PRL, prolactin; PRP, PACAP-related peptide; PTH, parathormone; VIP, vasoactive intestinal peptide.

184
XVIII . Ten ta t ive m o l e c u l a r s t r u c t u r e o f the type I P A C A P r e c e p t o r . . . . . . . . . . . . . . . . . . . . . . . . . 196
XIX. S u m m a r y a n d conc lu s ions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 196
A c k n o w l e d g e m e n t s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 197
R e f e r e n c e s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 197
I. Discovery and structure of PACAP
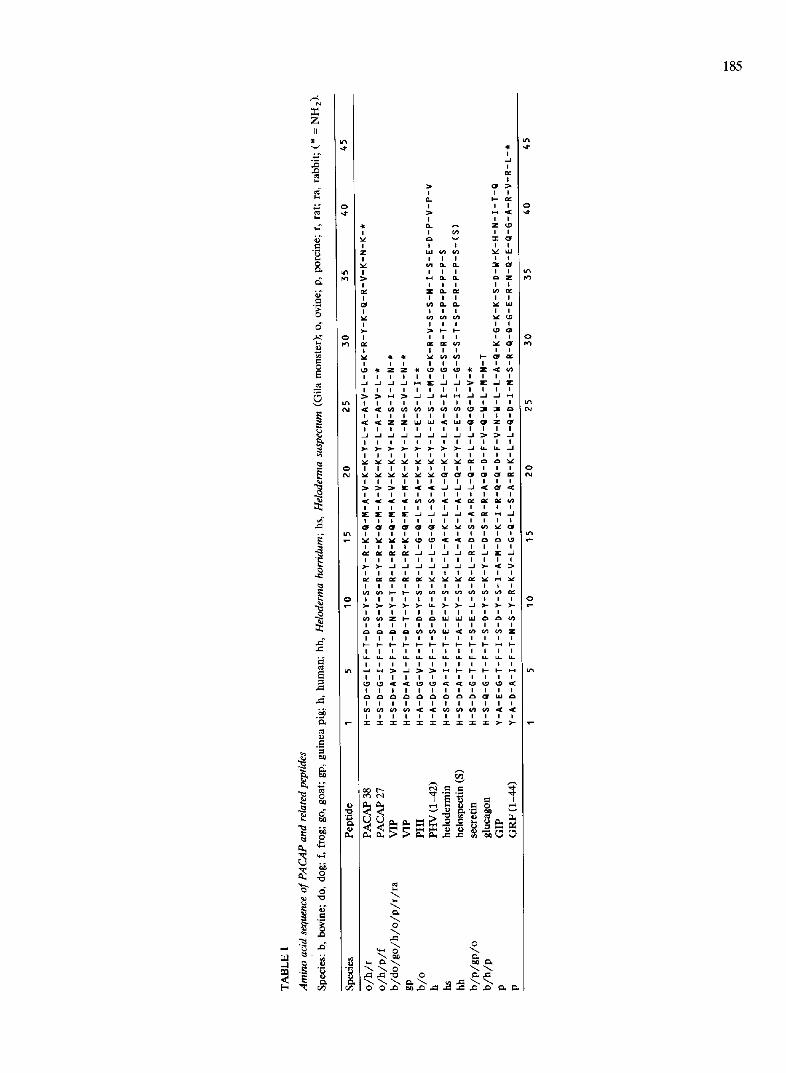
PACAP (the Pituitary Adenylate Cyclase Activating Polypeptide) is the latest discovered member of a su- perfamily that already includes several regulatory pep- tides, e.g., VIP, PHI, PHV, secretin, helodermin, helospectin I and II, glucagon, GIP and GRF (Table I). This biologically active neuropeptide exists in two ami- dated forms: PACAP(1-38)-NH 2 (in short PACAP-38) and PACAP(1-27)-NH 2 (in short PACAP-27). The deduced amino acid sequence of PACAP-38 in man [38] and rat [69] is identical to that of the isolated ovine PACAP-38. Ovine PACAP-38 was originally extracted from 5000 hypothalami, based on its ability to elevate cyclic AMP in cultured rat pituitary cells [55] and ovine PACAP-27 was isolated one year later from a side fraction [56]. It now appears that, in all tissues, PACAP-27 represents only a minor portion of total PACAP-ir. In PACAP-38 isolated from 1200 frog brains, there is one substitution of isoleucine for valine at position 35 [13] implying that the short PACAP-27 form is again identical. This extreme conservation of structure in vertebrates means that this neuropeptide is subjected to strong evolutionary pressure and may play a substantial role.
PACAP-27 exhibits 68% similarity to VIP, this being evident in the N-terminal (1-3) and (6-8) areas and in the central (14-23) region. In contrast, the (29-38) C-terminus of PACAP-38, with six cationic amino acid residues, shows no similarity to parent peptides of similar length (Table I).
II. The PACAP precursor and the PACAP gene
The molecular cloning of a single hybridizing cDNA for PACAP-38 and PACAP-27, from cDNA libraries of ovine hypothalamus, rat brain, and human testicle points at a highly homologous precursor sequence in the three animal species, and suggests that PACAP may be synthesized in several tissues. Ovine pro- PACAP-38 consists of 176 amino acid residues where PACAP-38 is located between positions 132 and 169 [38]. In the human precursor [32,38,70], some amino acid substitutions are observed in the 'pro' area. The human PACAP gene is assigned to the short arm of chromosome 18 in a region (locus 18pll) associated with hereditary holoprosencephaly, which may indicate that PACAP is involved in brain development. The last
exon encodes PACAP-38 and Y-untranslated se- quences, implying that PACAP-27 is not generated by alternative splicing. A PACAP-Related Peptide (PRP) of 29 amino acid residues is theoretically encoded by another exon arranged in tandem with the exon coding for PACAP. PRP shows low homology with PACAP itself. It is secreted into the cultured medium from CHO cells permanently transfected with the entire human PACAP precursor cDNA [72] but specific PRP receptors have not (yet) been described. The PACAP gene organization is compatible with the presence of two transcriptional regulatory response elements for cyclic AMP (CRE), one element for phorbol ester (TRE or AP-1 site), and two sequences homologous for pituitary-specific factor GHF-1 binding sites mediating the expression of the growth hormone gene [32]. The rat PACAP cDNA encodes a 175 amino acid precursor protein [69], due to the deletion of the amino acid present at position 29 from the N-terminus in ovine and human PACAP precursors and contains also a sequence for PRP.
Rat and human PACAP-38 are flanked, in their precursor, by basic processing sites and a carboxy- terminus amidation signal for the a-amidating mono- oxygenase system. Whether PACAP-27 is generated from mature PACAP-38 or both peptides are directly cleaved by alternative amidation and proteolysis from their common precursor remains to be established. At any rate PACAP-27 production requires the cleavage at the dibasic Lys-Arg site in position 29-30 of PACAP-38, and subsequent action on glycine in posi- tion 28 (Table I).
III. Two classes of PACAP receptors and a few receptor subtypes
At least two classes of PACAP receptors have been described in mammalian tissues and cell lines: type I PACAP-preferring receptors and type II receptors, which bind PACAP-27, PACAP-38 and VIP [11,88]. In addition, the first type is capable to display two sub- types, and the second type can be tentatively divided into three subtypes.
Type I is highly selective in that it recognizes PACAP-27 and PACAP-38 much more potently than VIP. PACAP-38 and PACAP-27 can thereafter acti- vate adenylate cyclase with the same high potency and efficacy. Typically, in human neuroblastoma NB-OK-1
185
e,i
z II
; .2
&
b ~
6 ° ~
0
e~
=
._=
e~
t
• Z Z •
~ Z Z ~
o .
ILl ( n
V ) O.
~-~ O.
Z O .
03 O.
( n ~n
~ , I - -
u~
(.o
u )
~C
> -
v
~C
._J
T ,_1
_J
>-
u~
Z
3
Z ~
~ J J
~ Z
T ~ T
._I
o .4-
uJ
z f ~
u4
I £
~-0
. - I
- - I
J
J
v
> -
~n
z
I -
u_
p-4
<c
~c
> -
0
0
0
0

186
cell membranes, the Kac t order is: PACAP-38 (0.2 nM)> PACAP-27 (0.4 nM) >> helodermin > VIP = PHV (1 /zM) [11]. Type I receptors may, however, be present as two coexisting subtypes (or states) based on radioligand binding. In rat brain membranes [12], the PACAP-A subtype exhibits hardly any preference for PACAP-38 over PACAP-27, whereas the PACAP-B subtype recognizes PACAP-38 with high affinity and PACAP-27 with low affinity. Accordingly: (a) Scatchard analysis of saturation curves for [~zSI]PACAP-38 as compared to [12s]PACAP-27 indicates a 2-fold higher number of receptors labelled by the first radioligand; and (b) the inhibition curve of [12SI]PACAP-38 binding by PACAP-38 is flatter (Hill coefficient < 1) than that obtained with PACAP-27, and the inhibition curve of [125I]PACAP-27 binding by PACAP-27 is flatter for PACAP-27 than that obtained with PACAP-38.
The type II receptor is the classical VIP receptor. Based on binding affinity and adenylate cyclase activa- tion, it now appears that the type II receptor is non- selective since it shows similar high affinity for PACAP-27, PACAP-38 and VIP (VIP > PACAP-27 = PACAP-38 >> helodermin > PHV). Type II receptors qualify, therefore, as VIP-PACAP receptors and are observed in large number in lung, liver, and vascular smooth muscle [77]. In rat liver membranes, a subclass of these type II receptors is specifically labelled with [125I]helodermin [79]. Another subclass of type II re- ceptors binds unlabelled helodermin with the highest affinity and is present in some neoplastic cell lines, including the human lymphoblastic SUP T1 cell line [27,76] and a human small cell lung carcinoma cell line [481.
We will focuse our attention to type I receptors and pass over the case for the already well documented VIP-PACAP (type II) receptors, that interact almost exclusively with adenylate cyclase.
IV. Two effector systems for type I PACAP-preferring receptors
The dissociation of [125I]PACAP-27 and [125I] PACAP-38 prebound to type I receptors in several membrane models (see, e.g., Refs. 11,12,46,71,85), in the presence of unlabelled PACAP, is moderately ac- celerated by GTP, suggesting a role for G protein(s) in stimulus-effector coupling. In many cases, it appears that type I receptors stimulate both adenylate cyclase and phospholipase C (PLC), this coupling to dual sig- naling cascades involving interactions with G proteins of the G s and Gq types.
1V-A. The adenylate cyclase system
When given intravenously to conscious mice, PACAP-27 and PACAP-38 are more effective than
VIP, helodermin, helospectin I and II, glucagon, PTH, calcitonin and CGRP in raising plasma cAMP [1]. In vitro, both PACAPs increase cAMP production through a GTP-dependent stimulation of adenylate cyclase in all membrane preparations endowed with type 1 recep- tors.
IV-B. The inositol phosphate cascade and cytosolic Ca 2 +
At variance with most type II receptors, type I receptors are often coupled to a second effector system elevating [Ca2+]i . Subsequent to PLC activation and through the inositol phosphate cascade, Ca 2 * mobiliza- tion is followed by a secondary Ca 2÷ entry. If we take human neuroblastoma NB-OK-1 cells as an example [17], PACAP-27 and PACAP-38 provoke a biphasic increase in [Ca2+]i: the transient elevation reflects the mobilization of intracellular Ca 2÷ (being unaffected by EGTA) while the secondary sustained Ca 2÷ entry is suppressed by EGTA. Concurrent IP 3 and IP 4 eleva- tions suggest that the intraceUular Ca 2 ÷ mobilization is due to IP 3 and the following Ca 2÷ entry to IP 4. The IP 3 and IP 4 productions, in response to PACAPs, are lower and short-lived as compared to those due to carbachol (acting through muscarinic M1 receptors) and this could be responsible for the lower sustained phase of [Ca2+]i elevation observed with PACAPs.
IV-C. Possible incidence of a parallel signalling network
Variations in intracellular pathways depend on a high number of parameters. They may reflect i.a. dif- ferences in receptor density, and the presence of iso- forms of adenylate cyclases or PKC. There is also evidence for regulatory links between PKC and adeny- late cyclase. In the example considered above, cAMP- mediated phosphorylations due to PACAP in NB-OK-1 cells could dampen the PACAP-stimulated PLC activ- ity, a phenomenon not engendered when muscarinic receptor occupancy provokes PLC activation only. This might account for PACAP receptors being less effi- ciently coupled to PLC than to adenylate cyclase in NB-OK-1 cells, and more rapidly desensitized at the PLC level, so that there is lesser Ca 2÷ handling than with muscarinic M1 receptors [17].
IV-D. Final biological responses
Fast responses typically include secretory and non secretory processes such as a relaxation of vascular smooth muscle. In the latter case [65] PACAP could conceivably trigger a cAMP-dependent phosphoryla- tion of myosin light chain kinase or the sequestration of Ca 2÷ into the sarcoplasmatic reticulum (the lower- ing of [Ca 2+ ]i should then reduce the formation of the

Ca2+-calmodulin complex hence reducing the actin- myosin interaction).
In vitro biphasic dose-response curves may reflect local desensitization or paracrine effects of one cell type on the other, and in vivo a parameter such as blood pressure may be influenced in opposite direc- tions by the release of catecholamines from the adrenal medulla and direct peripheral vasodilation (see below).
Even more importantly, cAMP and diglycerides may influence gene regulatory elements. Specific gene ex- pression can then favor cell growth and differentiation (e.g., neurite outgrowth in PC12 cells), survival or other basic cellular activities.
V. The hypothalamo-pituitary complex and the pineal gland
PACAP may exert several neuroendocrine functions in the regulation of the hypothalamo-pituitary complex. In vivo and in vitro data imply i.a. a role for PACAP in the regulation of reproduction, growth and food intake but further conclusive evidence is needed on these and other functions.
V-A. Presence of PACAP-ir neurons in the hypothalamus
PACAP-ir neurons are present in the hypothalamus of frog [114], sheep [42], rat [43,52], monkey and man [104]. In frog hypothalamus, numerous PACAP-ir neu- rons are observed in the preoptic nucleus (a region homologous to mammalian supraoptic and paraventric- ular nuclei) and in the dorsal and ventral infundibular nuclei that also contain TRH, somatostatin, NPY, ANF and a-MSH. In the caudal hypothalamus, a dense bundle of PACAP-ir fibers runs through the ventral infundibulum and terminates in the external vascular layer of the median eminence [114]. In ovine hypo- thalamus, PACAP-ir neurons are found in the supraoptic and paraventricular nuclei. There is a dense network of PACAP-ir fibers in both the external and internal zones of the median eminence and, in the external zone, they abut the hypophyseal portal capil- laries, pituitary stalk, septum and posterior pituitary [421.
Among seven nuclei of the rat hypothalamus, PACAP-ir perikarya are located in the supraoptic, paraventricular, suprachiasmatic, periventricular and arcuate nuclei [43]. PACAP-ir levels are moderate in the lateral hypothalamic area and in the mammillary body. In colchicine-treated rat, PACAP-ir fibers are organized in the stria terminalis pathway and an as- cending pathway from the suprachiasmatic nucleus to the paraventricular region [39,52]. Besides, VIP-ir in the suprachiasmatic nucleus is the highest among hy- pothalamic nuclei and the relationship between
187
PACAP and VIP in this nucleus should be investigated [43].
In the supraoptic and paraventricular nuclei of hu- man and spider monkey hypothalami, PACAP-ir neu- rons are again demonstrated [104]. Networks of PACAP-ir fibers are stained in both the external and internal zones of the tuber cinereum, close to the transition of the pituitary stalk (median eminence).
V-B. Hypothalamo-neurohypophyseal functions
PACAP may be involved in the regulation of poste- rior pituitary function. Indeed, PACAP-ir neurons are observed in the ovine posterior pituitary and PACAP-ir concentration is higher in the rat posterior lobe than in the anterior lobe [5,39]. Besides PACAP-ir magnocellu- lar cells in the supraoptic and paraventricular nuclei of the hypothalamus project to the internal zone of the median eminence and in the neurohypophysis (see above).
The paraventricular nucleus, rich not only in PACAP but also in CRF cell bodies, is involved in feeding behavior. In food-deprived mice, the intracerebroven- tricular (i.c.v.) injection of PACAP or CRF reduces food intake and increases grooming, it also antagonizes increased food intake resulting from administration of neuropeptide Y (NPY) [60]. Besides, the i.c.v, adminis- tration of PACAP raises plasma arginine vasopressin in conscious rat via type I receptors [64]. Arginine vaso- pressin originates in the supraoptic and paraventricular nuclei, and is transported to the posterior pituitary where it is stored and released. The posterior pituitary is outside the blood brain barrier and there is no circulation from hypothalamus to posterior pituitary. There results that i.c.v.-administered PACAP can not reach the posterior pituitary and probably acts on hypothalamic nuclei and hypothalamo-neurohypo- physeal fibers.
In cultured melanotropes from the rat neurointer- mediate gland, PACAP stimulates both cAMP forma- tion and the secretion of a-MSH. The effect of PACAP is additive with CRF on cAMP production but not on a-MSH release. On the other hand the activation of dopamine D2 receptors dampens both parameters [40].
V-C. Antehypophysiotropic functions of PACAP
PACAP encounters five criteria for being a hy- pophysiotropic neurohormone:
(1) As stated above, PACAP-ir neurons are present in the hypothalamus of primates, rat, sheep and frog. A dense network of PACAP-ir fibers terminates in the external zone of the ovine median eminence, in close contact with capillary loops, denoting a potential for PACAP release into portal vessels and transport to the anterior pituitary [42].

188
(2) The hypothalamus and anterior pituitary have type I PACAP-receptors and PACAP stimulates adenylate cyclase at both levels [5,25,50]. In rat hypo- thalamus a type IB subtype exhibits an eight-fold greater affinity for PACAP-38 compared with PACAP- 27 (PACAP-27 K d = 1452 pM; PACAP-38 K d = 175 pM) [25]. In general, type I receptors are in lower concentration in the anterior lobe of rat and man pituitary than in hypothalamus [25,45,90,91].
More precisely, in rat a high proportion of GH and PRL cells bind biotinyl-PACAP-38 as compared to a lesser number of ACTH cells and only a few LH, FSH and TSH cells [105]. These receptors are often coupled to the two effector systems but the biological hetero- geneity of responses among cell types or within a given category is obvious in the mammalian as well as am- phibian pituitary. In vitro in rat anterior pituitary cells, PACAP-38 is at least 1000-fold more potent than VIP in elevating cAMP [55]. It elevates cytosolic Ca 2+ in FSH cells (where it induces a 'Ca 2+ spike' or a Ca 2+- dependent 'Ca 2+ plateau' in 72% and 28% of the cases, respectively), GH cells (with a biphasic response or repetitive transients in 42-58% of the cases), more rarely in PRL and ACTH cells [9,113]. In freshly dis- persed rat anterior pituitary cells 96% of FSH and only 9% of PRL cells respond to PACAP by an increase in [Ca2+]i [105]. PACAP increases [Ca2+] i in cultured rat folliculo-stellate cells [98,113]. In cultured frog pitu- itary cells, PACAP stimulates adenylate cyclase activity [13]. It elevates [Ca2+]i in 45% of GH and PRL cells, 25% of ACTH and TSH cells, and only 16% of FSH cells through sustained stimulation, transient spike or oscillatory responses [28].
(3) PACAP variously affects hormone release from rat anterior pituitary depending on the cell type and experimental conditions. In static culture, dispersed cells fail to secrete pituitary hormones in response to PACAP [16,55]. However, PACAP weakly potentiates over 3 h the effects of L H R H on LH and FSH release in cell culture [16]. In a dynamic system with super- fused dispersed cells, PACAP increases both the num- ber of secreting GH cells and the amount of GH secreted per cell within 30 min [24]. Pulses of PACAP induce pulses of GH, ACTH, PRL and LH but the dose-response curve is bell-shaped for the first three hormones (due to desensitization at high dosis?) [55]. Others are unable to observe PACAP-induced GH or PRL secretion in this system [115]. Anterior pituitary folliculo-stellate (FS) cells endowed with type I recep- tors [98,105] produce interleukin-6 (IL-6) in response to PACAP, that stimulates (at relatively high concen- tration) the release of PRL, GH, FSH, and LH from cultured pituitary cells [98].
In frog, PACAP may stimulate the secretion of G H and PRL and, to a lesser extent, ACTH, TSH, and FSH [28].
(4) Delayed or atypical release of pituitary hor- mones in response to cAMP stimulation mediated by PACAP may reflect the interaction of a second mes- senger, induction of an additional transduction signal a n d / o r effects on transcription. For instance, PACAP inhibits PRL release as potently as dopamine from dispersed rat pituitary cells (implying that the stimula- tion of cAMP production in cultured pituitary cells is not due to an effect on PRL cells [35]). On dispersed rat cells in static culture, PACAP-38 effects (through PKC activation?) are significant after 4 h on the syn- thesis and release of LH and a-subunit while those on GH and ACTH levels (via IL-6 production?) require as much as 24 h for being meaningful [30]. Quantitative in situ hybridisation indicates that a prolonged (38 h) static incubation of a total pituitary cell suspension with PACAP-38 results in increased GH mRNA in GH cells and PRL mRNA in PRL cells (by paracrine action?) [115].
(5) In vivo an intraatrial injection of PACAP-38 evokes LH release much more than an intracere- broventricular injection in male rats (while VIP has almost no effect). This denotes that the LH releasing effect of PACAP-38 may be more potent via the ante- rior pituitary than via the hypothalamus [73]. PACAP stimulates hormone release in male rats [47]. An intra- venous PACAP infusion in hypothalamus-lesioned rats (with high serum PRL levels due to a lack of hypotha- lamic dopamine) rapidly stimulates PRL and GH re- lease. The apparently contradicting in vivo and in vitro observations on PRL suggest that PACAP stimulates the release of a paracrine factor (interleukin-6, fibro- blast growth factor, epidermal growth factor?) in the intact pituitary that outrides the direct inhibitory ac- tion of PACAP on PRL cells [35].
V-D. Cancerous pituitary cell lines
The PACAP receptors of the mouse pituitary clonal cell line AtT-20 and the rat pituitary clonal cell line GH3 are probably distinct [74]. In mouse AtT-20 corti- cotropes (proopiomelanocortin (POMC) cells), PACAP at low concentration stimulates cAMP production and the release of ACTH and 7B2 (a 185 amino acid protein found in secretory granules and in the plasma of patients with some endocrine tumors) over a 2-3 h incubation period. Addition of CRF-41 to PACAP produces an additive effect on ACTH release and a synergistic effect on 7B2 release while the co-addition of a somatostatin octapeptide agonist exerts opposite effects [40,74].
The cAMP content of rat G H 3 clonal pituitary cells is elevated by PACAP-27, PACAP-38, and VIP through type II VIP-PACAP receptors and there is no activa- tion in phosphatidyl inositol turnover. The three pep- tides show similar efficacy on the resulting hypersecre-

189
tion of GH, PRL and 7B2 over a 2-5 h incubation and are opposed by a somatostatin agonist [18,74].
V-E. PACAP receptors (of type I mostly) are present in human benign pituitary adenomas except in prolactino- mas
In membranes from 'null'-, gonadotropin-, ACTH- and GH-producing human pituitary adenomas, PACAP-27 and PACAP-38 stimulate adenylate cyclase activity potently (Kac t 3 nM) and equally well while VIP is 100-300-fold less potent. By contrast, both PACAPs are inactive in human prolactinomas [80]. In parallel, l~I-[AcHis-1]PACAP-27 binding is detected on membranes from all adenomas, except in prolacti- nomas. Taken together the data are compatible with the coexistence of a majority of type I receptors and a minority of type II receptors in gonadotropin- and ACTH-secreting adenomas.
V-F. The pineal gland
The density of type I receptors is 12-fold higher than that of type II receptors in the rat pineal gland [51]. This gland displays circadian synthesis and secre- tion of melatonin due to the nocturnal release of noradrenaline from sympathetic fibers. Melatonin is immediately released upon its synthesis. PACAP-27 proves to be a potent stimulator of melatonin synthesis from perifused and cultured dissociated pinealocytes. As expected, this PACAP stimulation is not inhibited by the VIP antagonist [4-CI-o-Phe-6,Leu-17]VIP whereas that of VIP is halved [89].
VI. The brain and spinal cord
VIA. Three PACAP forms in rat brain
PACAP is found by radioimmunoassay and immu- nocytochemistry in several parts of the brain. Total PACAP content is highest in hypothalamus (49.3 ng/g) and lower in hippocampus, frontal cortex, and cerebel- lum (6.1, 3.1, and 1.6 ng/g, respectively) [102]. Three PACAP forms can be separated by combining Fracto- gel chromatography and radioimmunoassays: PACAP- 38, PACAP-27 and a minor biologically inactive C- terminal fragment. PACAP-38 is the predominant form in all areas (from 91% of the total immunoreactivity in cerebellum to 73% in cortex) while the PACAP-27 fragment represents no more than 10% and may be PACAP(21-38)-NH 2, a degradation product resulting from cleavage of the 20-21 dibasic doublet (see also Ref. 72).
VI-B. Presence of PACAP in rat and frog central nervous system
In rat, apart from their presence in the hypothala- mus, most of the PACAP-ir neurons and fibers are
located in medial and ventral areas of the dien- cephalon, central thalamic nuclei, amygdala, bed nu-
cleus of stria terminalis, septum, hippocampus, and cingulate and entorhinal cortex [43]. The highest con- centrations of PACAP-ir are found in the substantia nigra and nucleus accumbens, followed by the septum, amygdala, globus pallidus and sacral spinal cord. This is accompanied by the presence of a PACAP mRNA of 2.7 kb [23]. By sandwich-enzyme immunoassay high amounts of PACAP-ir are observed in basal forebrain (lateral septal nucleus, intermediate part and diagonal band), central amygdaloid nucleus, brainstem (central gray, interpeduncular nucleus and dorsal raphe nuclei). Moderate amounts of PACAP-ir are found in basal forebrain in the dorsal part of the lateral septal nu- cleus and medial septal nucleus, in medial amygdaloid nucleus, thalamus (paraventricular, paratenial, central medial, ventromedial, reuniens and rhomboid nuclei), and brainstem (ventral tegmental area, interfascicular nucleus and locus coeruleus) [52].
In Rana ridibunda [114], PACAP-ir perikarya and fibers are also widely distributed. Perikarya are located in the medial and ventral diencephalon, i.e., the preop- tic nucleus, the ventral and dorsal infundibular nuclei, and very abundantly in the thalamus (nucleus postero- centralis thalami and ventral and ventrolateral areas). In the telencephalon, a few perikarya are found in the nucleus accumbens septi, the amygdala, the pallial commissure, and the bed nucleus of the pallial commis- sure. In the hindbrain, PACAP-ir perikarya are present in the torus semicircularis, the nucleus profundus and the nucleus anteroventralis tegmenti of the mesen- cephalon. PACAP-ir fibers extend from the central thalamus to the optic tectum, and rostrocaudally, from the nucleus accumbens septi to the nucleus entopedun- cularis. Other areas, such as the interpeduncular nu- cleus, the nucleus isthmi and the roots of cranial nerves V and III in the medulla oblongata, are densely inner- vated. PACAP-ir fibers extend through the dorsal tha- lamus, from the nucleus posterocentralis to the optic tectum. Concurrently, a dense bundle of PACAP-ir fibers is observed in the nucleus isthmi, which is at the origin of the tractus isthmotectalis, one of the afferent connections of the tectum [114].
VI-C. Type I receptors in the rat central nervous system
A high density of type I receptors is observed in brain membranes from several species (e.g., the human brain [90]) but we will consider more specifically the various rat brain areas [12]. In a crude approach, the cortex, olfactory bulb, hippocampus, hypothalamus, striatum, cerebellum, pons and cervico-dorsal spinal cord show a Bma x in the 1-2 pmol /mg protein range, i.e., 10-50-fold higher than that of type II VIP-PACAP receptors in the same areas, and in the same high

190
range as /3-adrenoceptors. Thus, 125I-[AcHis-1]- PACAP-27 that binds with similar affinity to both type I and type II receptors essentially identifies type I receptors.
More precisely, in rat forebrain sections [50,51] type I receptors are dense almost everywhere!
VI-D. Ontogenesis of PACAP and type I receptors in rat central nervous system
PACAP-ir levels in rat brain are high already at birth and do not change markedly during the first 3 weeks of post natal development, but the relative pro- portion of PACAP-27 increases somewhat during the first post natal week. If the density of type I receptors is stable over the same period, the coupling of these receptors to adenylate cyclase increases from birth to weaning, owing to increasing enzyme activity [103].
VI-E. Possible neurobiological roles of PACAP in ex- trahypothalamic areas of the central nervous system
These roles are not yet clearly established. Of course the abundance of PACAP-ir neurons in several brain areas and even more so the very broad distribution of type I receptors imply that PACAP-38 may exert neurotransmitter and /o r neuromodulatory activities. As expected, the PACAP-ir distribution in forebrain does not parallel the localization of type I receptors, e.g., low amounts of PACAP-ir are observed in the hippocampal formation which contains high densities of type I receptors. Additionally, PACAP is more widely distributed than VIP in brain [22] and PACAP and VIP neural circuits are at least partly independent, e.g., PACAP-ir levels are low in various regions of the cerebral cortex where VIP is highly concentrated [43]. The regional distribution pattern of type I receptors is also remarkably different from that of type II (VIP- PACAP) receptors [50,51].
In the rat mesencephalon, type I receptors are pre- sent in the superior colliculus and the medial genicu- late body, two structures mediating visual attention [50]. In frog hindbrain, ir-perikarya are located in tha- lamic and perithalamic (mesencephalic central gray) regions where PACAP may, there too, play a role in the integration of visual inputs [114].
On a neuropharmacological point of view, it would be of interest to study any contribution of PACAP as an amnestic [50] since: (a) neuropeptide Y (its antago- nist on food intake: see above) is a memory enhancer when injected intrahippocampal [60], and (b) the hip- pocampus is rich in type I receptors and receives afferents from septohippocampal cholinergic neurons playing an important role in memory. In the brainstem, the interpeduncular nucleus displays high PACAP-ir and ANF-ir, raising the question of PACAP influenc- ing heart rate and salt appetite centrally [114]. It would
also be relevant to investigate the effects of PACAP on meso-striatal and meso-limbic dopaminergic pathways and on serotonergic neurons in the dorsal raphe [51].
PACAP-38 increases cAMP levels in cultured rat neurons and cultured rat astrocytes [97] and [Ca2+]i in cultured rat hippocampal neurons from 18-day-old em- bryos (whereas VIP is inefficient) [99]. Alterations of cAMP and Ca 2+ homeostasis could support growth cone behavior during development, repair, and alter- ation in circuitry. PACAP may be particularly impor- tant for the development of rat brain: its early pres- ence and that of type I receptors in newborn rat brain is in line with this concept (see above). PACAP pro- tects mouse hippocampal neurons in culture from the lethal action of gp 120, the envelop protein of HIV virus (a preventive effect at least 100-fold greater than that of VIP) [4]. Whether this protection is due to the direct action of PACAP on neurons or mediated by neurotropic factors from astrocytes remains to be eluci- dated.
VII. Ocular distribution of type I receptors
Type I receptors constitute the major part of PACAP receptors in membranes from the retina (~ 90%) and choroid (~ 60%) of albino rabbit eye. Adenylate cy- clase is stimulated 5.2-fold in retinal membranes and 2.4-fold in choroid membranes by PACAP-38. VIP shows less potency and efficacy. It remains to be estab- lished whether type I receptors can influence the physi- ology of specific retinal cell populations, as well as vasodilation and aqueous humor flow in the choroid [68].
VIII. The digestive tract
VILLA. PACAP localization
In the esophagus of sheep, man and cat, PACAP-ir fibers are numerous and a few PACAP-ir neurons are seen in myenteric ganglia. In the esophagus of guinea- pig PACAP-ir fibers are scant [101]. In sheep, PACAP- ir fibers are present in all regions of the stomach and surround Brunner's glands in the submucosal layer of duodenum [6]. In ovine, porcine, feline, and murine gut wall, PACAP-ir fibers are mostly of intramural origin as inferred by extrinsic denervation experiments and predominate in myenteric ganglia and muscle. PAC- AP-ir neurons are numerous in submucous ganglia [93]. In human small and large intestine, PACAP-ir fibers and cell bodies are numerous in the mucosa as well as in both muscle layers [93]. In rat, PACAP-ir is higher in stomach, duodenum and jejunum than in other portions of the gut [5], PACAP-ir fibers are rarely observed in small intestine and PACAP-ir neurons are found in the myenteric plexus but rarely in the mucosal area [93].

VIII-B. Physiological roles
I/IlI-B.1. Motility The intramural PACAP innervation is probably re-
sponsible for muscle relaxation. Basically, it appears that PACAP ranks among the inhibitory non-adren- ergic, non-cholinergic neurotransmitters and can di- rectly relax intestinal smooth muscle by activating apamin-sensitive potassium channels [86]. Yet, some experimental data are difficult to interpret, e.g., in rat and chicken where PACAP and VIP coexist in neurons throughout the gut [92,93] or when cholinergic neurons play an interfering role.
PACAP relaxes smooth muscle in vitro in all por- tions of the rat gastrointestinal tract through type I receptors (e.g., its inhibitory effect in midcolon is 100- fold higher than that of VIP). It inhibits the contrac- tions induced by carbachol, cholecystokinin, substance P, and galanin. By contrast, VIP causes atropine-sensi- tive contraction of the longitudinal muscle layer of duodenum, implying the stimulating presence of type II receptors on cholinergic neurons. This VIP-induced contraction is inhibited by PACAP. Therefore, PACAP and VIP act in opposite directions in some areas of the intestine [63].
PACAP-27 and VIP directly relax the guinea-pig taenia caeci in the presence of scopolamine, guanethi- dine and tetrodotoxin. On the other hand, PACAP and VIP can indirectly contract the longitudinal muscle of guinea-pig ileum by releasing acetylcholine and a non- cholinergic neurotransmitter. Apamin, the blocker of calcium-activated potassium channels, prevents the re- laxation induced by PACAP-27 whereas the effect of VIP remains unaffected. This indicates that the effect of PACAP-27 on apamin-sensitive potassium channels is mediated by type I receptors [86].
In patients with Hirschprung's disease, the sustained contraction of a segment of the distal colon is due to a lack of ganglion cells in the submucous and myenteric plexuses. The scarcer PACAP innervation in this agan- glionic segment may contribute to the relaxation de- fect, in an area where fibers for VIP, PHI, substance P, GRP, and enkephalin are also reduced [87].
The contractile effects of intravenous PACAP-27 and PACAP-38 on the gallbladder in conscious dog are, on a molar basis, 0.11 and 0.04 as potent as CCK-8 in the digestive state, and 0.18 and 0.02 in the interdi- gestive state. By contrast, VIP inhibits gallbladder motility in the digestive state. PACAP effects are al- most abolished by atropine or hexamethonium and are mediated, therefore, by a preganglionic cholinergic mechanism [57].
VIII-B.2. Secretory processes In isolated chief cells from guinea pig stomach,
PACAP stimulates pepsinogen secretion through type
191
II receptors at low concentration and through secretin receptors at high concentration [20]. In conscious rat PACAP-27 inhibits pentagastrin-stimulated gastric acid secretion whereas VIP has no effect. This inhibitory effect of PACAP-27 is biphasic, since a large dose produces a lesser inhibitory effect. It is conceivable (but not proven) that PACAP might promote somato- statin release from D cells [62].
Very few type I receptors are found in rat intestinal membranes contrasting with numerous type II recep- tors in rat and human. For instance, a type II receptor (molecular mass 67 kDa) is identified in epithelial plasma membranes from human small intestine [83]. In practice, in vitro, PACAP-27, PACAP-38 and VIP act as anion secretory neuropeptides on mucosal sheets of rat jejunum. At low concentration, the rapid and tran- sient secretory effect of PACAP-27 is inhibited by tetrodotoxin, denoting a role for type I receptors on submucous neurons innervating the mucosal region. More prolonged and tetrodotoxin-insensitive secretory responses through basolateral epithelial receptors pre- dominate at higher PACAP-27 concentration [15]. In guinea-pig distal colon PACAP evokes chloride secre- tion by noncholinergic neuronal activation [44].
In the T84 colonocyte cell line, VIP, PACAP-38 and PACAP-27 activate electrogenic chloride secretion (relative potency: VIP > PACAP-27 > PACAP-38) and increase a cAMP-dependent phosphorylation of phos- phoproteins p18, p23 and p37. The PACAP response of the monolayer, observed in Ussing chamber, is po- tentiated by carbachol, inhibited by bumetanide and barium chloride, and not further stimulated by VIP addition. Thus, both PACAPs act via type II receptors through cAMP-dependent signaling [67].
IX. The exocrine pancreas and pancreatic AR 4-2J cell line
In the pancreas of rat and mouse, no PACAP neu- rons are seen but extrinsic PACAP-ir fibers innervate the pancreatic ganglia preferentially, with a minor in- nervation of the islets [21].
IX-A. Normal exocrine pancreas
During postnatal development, the bovine pancre- atic function may be regulated by type I receptors linked to cAMP generation. Indeed, the calf pancreas adapted to milk intake shows type I receptors (molecu- lar mass 55 kDa) with the following specificity: PACAP-38 > helodermin > VIP, PACAP-27 > PHM [46]. In vitro, in rat pancreatic acini, the inhibition of PACAP-38-induced cAMP production and amylase re- lease by two VIP antagonists ([Ac-Tyr-l,o-Phe- 2]GRF1-29NH z and [4-Cl-o-Phe-6,Leu-17]VIP) indi- cates that PACAP effects are mediated by type II

192
receptors [36,75]. Indeed, no type I receptors are de- tected in the rat pancreas. In vivo, in the anesthetized rat, a bolus PACAP-27 injection stimulates pancreatic amylase secretion more efficiently than VIP [61]. In conscious dog, PACAP-38 and PACAP-27, in contrast to VIP and secretin, stimulate protein secretion. This phenomenon is reduced by atropine, suggesting that PACAP provokes the secretion of hydrolases via a cholinergic mechanism (while having a VIP-secretin like action on pancreatic fluid and bicarbonate secre- tion) [66].
IX-B. The rat pancreatic carcinoma acinar cell line AR 4-2J
The AR 4-2J cell line derives from azaserine-in- duced hyperplastic nodules of the rat exocrine pan- creas and this cancerous state provokes the emergence of type I receptors normally absent in the exocrine pancreas but revealed by malignancy. Some time ago, in our hands, this cell line also exhibited a large number of type II receptors [94] but, after numerous transfers, this is no longer the case. In AR 4-2J cell membranes, the total number of type I receptors la- belled by [~25I]PACAP-38 is 2-fold higher than that labelled by [125I]PACAP-27. Inhibition curves of tracer binding with PACAP-27 are flatter than those obtained with PACAP-38. This suggests two subtypes (states) of type I receptors: one with high affinity (K d 0.3 nM) for PACAP-27 and PACAP-38 and a second one with high affinity for PACAP-38 (K d 0.3 nM) and low affinity for PACAP-27 (K~ 30 nM) [7,78,82]. In the present case, PACAP must be added to a list of mitogenic agents that already includes gastrin, insulin and epidermal growth factor: PACAP increases ornithine decarboxy- lase activity as an early event then induces, after 2 days, a 30% increase of cell proliferation. This mito- genic effect of PACAP depends on a pertussis-sensitive G protein and is suppressed by a somatostatin analog, perhaps through phosphorylation/dephosphorylation of tyrosine residues [8].
X. The endocrine pancreas
In the perfused rat pancreas, glucose-dependent insulin release is stimulated, with the following po- tency, by: truncated glucagon like peptide-1 > PACAP- 27 > VIP. Thus, a slight direct stimulatory action of PACAP on fl-cells is suggested on this preparation [37,58]. In vivo in mice, PACAP-38 augments markedly the basal and carbachol-stimulated glucagon secretion as well as plasma glucose, and this may be responsible for a modest elevation of plasma insulin. A preferential stimulation of glucagon secretion is also observed with VIP, PHI, helodermin, helospectin I and secretin [21].
XI. The liver
In rat liver, PACAP-27 binds to two receptors: (1) a minority (20%) of type I receptors with a K d for PACAP-38 of 0.5 nM; (2) a majority (80%) of type II receptors coexisting in two states: (a) the high affinity state specifically labelled by [ 125 I]helodermin that shows the following affinity: PACAP-27 > VIP > PACAP-38; and (b) the low affinity state tested by [~25I]VIP bind- ing, and for which PACAP-27 = PACAP-38 > VIP [79].
XII. The adrenal medulla and the rat pheochromacy- toma PC12 and human neuroblastoma NB-OK cell lines
XII-A. Adrenal medulla
The rat adrenal medulla is endowed with type I receptors and contains, among peripheral tissues, the highest concentration of PACAP next to the testis [5]. PACAP activates adenylate cyclase and elevates cy- tosolic Ca 2+ in cultured rat chromaffin cells [111]. Infusion of PACAP into the rat adrenal gland results in a long-term (over 120 min) hypersecretion of cate- cholamines [106]. Thus, PACAP is involved in acute synthesis and secretion of catecholamines from chro- maffin cells (by contrast acetylcholine, with Ca 2÷ as probable messenger, provokes only short term secre- tion). The rate-limiting enzyme tyrosine hydroxylase is phosphorylated by the cAMP-dependent protein ki- nase and, in addition, transcription of its gene is accel- erated by a cAMP responsive element in the promoter region. PACAP may also regulate sympathetic develop- ment since, like VIP, it potently enhances mitosis, stimulates neurite genesis, and prolongs survival of cultured sympathetic neuroblasts from the superior cervical ganglia of rat embryo [19].
XII-B. Rat pheochromocytoma PC12 cell line
In PC12 cells, derived from a transplantable rat adrenal pheochromocytoma, PACAP-38 and PACAP- 27 elevate cAMP levels. PACAP-38 additionally ele- vates inositol phosphates while PACAP-27 is 2-3 logs less potent. The minimal responsiveness to VIP sug- gests that type I receptors are involved [18,110]. The elevation of cAMP induces tyrosine hydroxylase. In addition, PACAP-38 is more efficient than nerve growth factor (NGF) in eliciting neurite outgrowth (longer but fewer neurites) [18,77]. By comparison, 8-bromo-cAMP results in only 23% cells bearing neu- rites after 3 days of treatment.
XII-C. Type I receptors in the human neuroblastoma cell line NB-OK
The human neuroblastoma NB-OK-1 cell line pos- sesses type I receptors and the structural requirements

193
for their occupancy and for adenylate cyclase activation have been defined [26,81] (see below).
XIII. The respiratory tract
XIII-A. The respiratory wall
PACAP-ir fibers are found in the tracheo-bronchial wall of guinea-pig, rat, ferret, pig, sheep and squirrel monkey [100]. In particular, in the guinea-pig, a moder- ate supply of PACAP-ir fibers is seen around smooth muscle bundles and glands of the trachea and bronchi and in the nasal mucosa [10]. All PACAP-ir fibers store VIP but some VIP-ir fibers lack PACAP. A few PACAP-ir neurons are located in lungs and also in sphenopalatine and otic ganglia. Two classes of [125I]PACAP-27 receptors coexist in rat lung mem- branes with K d values of 1.0 nM and 5.2 nM, respec- tively. These two classes are less specific than those of the central nervous system and may reflect partial binding to VIP receptors [45].
PACAP-38 and PACAP-27 acting through cAMP are nonadrenergic, noncholinergic relaxants of guinea- pig tracheal smooth muscle. The first agonist has a longer-lasting action in vitro [3] and is slightly more dilatory than VIP on the trachea precontracted with histamine [10]. PACAP may, therefore, contribute to the balance between bronchodilators (/3-adrenergic, PACAP and VIP) and bronchoconstrictors.
Human small cell lung carcinoma cell lines possess VIP-helodermin preferring type II receptors [48] or PACAP-preferring type I receptors [59]. In the NCI- H345 cell line, PACAP increases cAMP and [Ca2+] i and stimulates clonal growth perhaps through c-los and bombesin/GRP gene expression [59].
XIII-B. The pulmonary artery
In the guinea-pig there is a rich supply of PACAP-ir fibers around blood vessels in the lung, and PACAP and VIP provoke the same dilation of the pulmonary artery contracted with histamine, noradrenaline or serotonine. This PACAP effect is endothelium-depen- dent (the removal of endothelium abolishes PACAP- induced vascular relaxation) whereas that to VIP is not, suggesting a different mechanism of action for PACAP and VIP [10]. In the anesthetized cat, injection of PACAP decreases the pulmonary lobar arterial pressure through vasodilation and type I receptors may contribute, since PACAP is 10-fold more potent than VIP in dilating the pulmonary vascular bed [53].
XIV. The extra-pulmonary vascular system and heart
XIV-A. Systemic arterial pressure
After intravenous PACAP administration to anes- thetized rat, the arterial pressure decreases transiently
[2,65]. By contrast, after i.c.v, administration of PACAP there is a transient moderate hypertensive peak at 5 min. In this case PACAP may influence the central cardiovascular control system by releasing arginine va- sopressin (AVP) [64]. That blood pressure reflects sev- eral parameters is illustrated in further studies on the anesthetized rat: PACAP causes a transient hypoten- sive effect at low dosis, the amplitude and duration of the response being comparable with VIP [65] and pos- sibly implemented through type II receptors and /o r the release of cAMP in blood [1]. At higher dosis, the biphasic change in blood pressure (decrease followed by increase) is accompanied by a biphasic change in systemic vascular resistance [2,54,108]. In the anes- thetized dog, intravenous PACAP-27 and PACAP-38 provoke hypotension at low dosis but hypertension and bradycardia after transient hypotension and tachycar- dia at high dosis [34]. In the anesthetized cat PACAP provokes an initial decrease in systemic arterial pres- sure that is followed by a secondary increase [53]. In addition PACAP increases right ventricular contractile force, left atrial pressure and heart rate. These in- creases are changed to decreases following the admin- istration of phentolamine or after adrenalectomy [54]. Thus, the pressor responses are due to the release of catecholamines from the adrenal gland (see above).
XIV-B. Vascular and microvascular beds
The species and the part of the vasculature tested may explain differences in response. In man, infusion of PACAP and VIP into the brachial artery increases the forearm blood flow [109]. In cat PACAP produces a biphasic change in perfusion pressure in the hindquarters vascular bed: an initial decrease is fol- lowed by a secondary increase and VIP is 3- to 10-fold more potent than PACAP in dilating the bed [53]. PACAP-38, PACAP-27 and VIP are equipotent in vitro in relaxing the phenylephrine-precontracted rat tail artery [2]. Accordingly, in rat, IC50 values for PACAP-27 and VIP binding are identical in ho- mogenates from aorta, and iliac and femoral vein and artery, suggesting that these peptides share type II receptors [65]. However, in isolated rabbit precon- tracted aortic rings, the relaxant action of both forms of PACAP is 100-fold more potent than VIP, surpris- ingly long-lasted, and endothelium-independent [107].
PACAP is a potent microvasodilator in human skin: intradermally injected PACAP and VIP cause a rapid 2-3 min flare which becomes erythematous after 5 min. The PACAP induced vasodilatation lasts 6 h, whereas the effect of VIP lasts less than 2 h [109]. In rabbit, based on 133Xe clearance from skin, 10-~2 mol PACAP-38 provokes the same in vivo microvasodila- tion as a 100-fold higher (10 -1° mol) dose of VIP per site. The vasodilation effect of PACAP-38 is still mea-

194
surable 2 h after the intradermal injection of 1 pmol. Thus, PACAP may contribute to both the hyperaemia and oedema constituents of inflammation [108].
XIV-C. Heart
In conscious sheep, PACAP and VIP provoke a tachycardia greater than expected from baroreceptor activation, that may not rely only on changes in blood pressure [84]. On isolated piglet heart, PACAP exerts a potent positive inotropic effect [54]. On isolated per- fused rat heart, a PACAP-38 bolus exerts a transient (maximum at 20 s) positive inotropic effect. On car- diomyocytes cultured from neonatal rat heart, PACAP- 38 causes a long-lasting increase in beating rate [112]. PACAP-38 and PACAP-27 stimulate adenylate cyclase in purified sarcolemmal membranes from porcine and canine ventricles with higher potency than VIP. By contrast, PACAP activation of adenylate cyclase in crude membranes and homogenates from porcine, ca- nine, and rat heart is comparable to that of VIP [112].
XV. Normal and neoplastic blood cells and fibroblasts
The proliferation of murine splenocytes induced by concanavalin A is inhibited when PACAP-27, PACAP- 38 or VIP bind to type II receptors [96]. Again PACAPs and VIP exhibit similar affinity in membranes from normal peripheral human monocytes, on which they double cAMP generation (VIP, PACAP-38 > PHV, PHM > PACAP-27, hpGRF and secretin) [14].
By contrast, it is through type I receptors that PACAP increases adenylate cyclase activity 5.3-fold in membranes from human THP-1 leukemic monocytes, with an affinity 50-fold higher than that of VIP. The potency order is PACAP-38 > helodermin, VIP > PHM, PHV, helospectin. The spontaneous or retinoic acid-induced differentiation of normal peripheral monocytes and leukemic THP-1 rnonocytes into macrophages reduces the functional expression of Gsa an the responsiveness to PACAP and VIP in terms of respiratory burst, lysosomal enzyme release, migration, phagocytosis and cytotoxicity [14].
In human Sup-T1 ceils (a childhood CD4 + T-cell lymphoma cell line), an original type II receptor sub- type recognizes helodermin with a higher potency than PACAP and VIP. The order of potency for binding and adenylate cyclase activation is: helodermin-35 > helodermin-27 > PACAP-38 > PACAP-27 > VIP [27].
PACAP-induced cAMP production is dampened in non-tumoral non-endocrine rat pituitary fibroblasts by the phorbol ester PMA but unchanged in Swiss 3T3 fibroblasts [41].
XVI. The reproductive function
A cDNA encoding PACAP-38 was cloned from a human testis Agtll cDNA library, suggesting that the
PACAP gene is expressed in this tissue [38]. In rat PACAP-ir is indeed as abundant in testis as in most brain areas while the ovary contains only a small con- centration of PACAP [5]. In the testis and epididymis, type I PACAP receptors are localized in Sertoli cells [31] and in germinal cells (especially in the spermato- zoa tail) [88].
PACAP-38, helodermin and helospectin suppress electrically evoked contractions in the rat vas deferens via receptors distinct from type II receptors [29]. LH and FSH secretion by the pituitary is potentiated by PACAP (see above). Together, PACAP may play an important role in spermatogenesis, sperm motility, and reproductive activity.
XVII. Structural requirements for the occupancy of type I receptors coupled to adenylate cyclase activation
XVII-A. Conformation of PACAP
Structural predictions suggest that PACAP-27 and PACAP-38 may have a first /3-bend at residues 2-4, a second /3-bend at residues 8-11 and an a-helix at residues 14-27. When determined by 1H-NMR and CD spectroscopies, the conformation of PACAP-27, in 25% methanol, includes extended or averaging confor- mation for residues 1-6, a type II /3 turn at residues 9-12, an a helix (residues 12-14), a looser helical conformation (residues 15-20), and an a helix (re- sidues 22-24). Two hydrophobic clusters contribute to energy stability: Tyr-10, Tyr-13 and Met-17 on the one hand, and Tyr-22, Leu-23 and Val-26 on the other hand [33].
XVII-B. Biological requirements for stimulus-effector coupling
Certain PACAP portions may be critically involved in in vitro receptor recognition and subsequent adeny- late cyclase activation. To tackle this problem, several PACAP analogs are compared for their ability (a) to compete with 125I-[AcHis-1]PACAP-27 (a tracer with low non-specific binding) for the occupancy of type I receptors and (b) to activate the enzyme. The constants evaluated include: (1) the IC50, median inhibitory con- centration of a given unlabelled ligand, that inhibits 50% of tracer binding (if the tracer concentration is low enough, the ICso is close to the concentration Ka required for half-maximal occupancy; (2) the Kact, con- centration exerting half-maximal stimulation of the en- zyme, and (3) the I.A., intrinsic activity (a fraction, taking the maximal effect of the reference peptide PACAP-27 as 1.0). When the peptide investigated is inactive or almost inactive (I.A. < 0.05), it is tested on the dose/effect curve of PACAP-27 on adenylate cy- clase, in order to evaluate the K~, concentration pro- yoking a 2-fold shift of the curve to the right.
We used three model membranes rich in type I

receptors and (almost) free of type II receptors: those from the human neuroblastoma cell line NB-OK-1, the rat hippocampus, and the rat pancreatic acinar cell line AR 4-2J [26,78,81,82]. The general conclusions are the same for the three membrane systems and, unless specifically quoted, data on NB-OK-1 membranes are mentioned below. We consider that slight quantitative (but not qualitative) differences among the three sys- tems, such as a general tendency towards higher ICs0 and lesser I.A. values with a given series of analogs, merely reflect lower coupling of type I receptors but not the existence of a distinct receptor variant.
XVII-C. Mono- or bi-substituted analogs
Starting with position 1, the acetylation of His-1 in PACAP-27 decreases moderately (2-fold) the ICs0 i.e., it increases the affinity: a free ionizable a-amino group is obviously unimportant for optimal Kac r More pre- cisely, in AR 4-2J membranes, deprotonated His-1 is a major determinant for PACAP-27 recognition: the K d of PACAP-27 decreases 6-fold between pH 6.0 (3.0 nM) and 8.0 (0.5 nM) and the pH effect on PACAP-27 binding is amplified at 15°C as compared to 37°C. However, the Ka of PACAP-38 (0.3 nM) is pH-inde- pendent. The imidazole ring itself is important for optimal coupling, since the replacement of His-1 by Phe-1 reduces the I.A. by 40% but its presence is not mandatory, considering the I.A. of 0.9 achieved with PACAP(2-27) and PACAP(2-38).
Altering the /3-turn potential in the 1-3 region can provoke major changes. The chemistry and size of the amino acid in position 2 is determining for coupling: isosteric substitution of Ser-2 is well tolerated (L-Ala-2 and D-Ala-2 derivatives are full agonists) but L-Arg-2 or D-Arg-2 and L-Phe-2 or D-Phe-2 are partial agonists only and the substitution of p-C1-D-Phe-2 in PACAP-38 gives a potent antagonist (ICs0 2 nM) with very little I.A. (0.1). The presence of the acidic Asp-3 residue in PACAP-27 is critical as its substitution by Asn reduces 5000-fold the peptide affinity. This suggests an essen- tial ionic bound between the ligand and its receptor counterpart and any hindrance in this respect is poorly supported since the I.A. of [Glu-3]PACAP-27 is re- duced to 0.4. When Gly-4 in PACAP-38 is replaced by an Aib residue, in order to constrain the /3-turn, the biological parameters remain similar to those of the natural ligand.
Further down the peptide, Met-17 can be easily replaced by Nle-17. Finally, the hydrophilic residues Lys-20,Lys-21 are important for binding: the combined a-helix-breaking Gly-20,Gly-21 substitution reduces the binding affinity 500-fold.
XVII-D. Discovery of N-shortened PACAP antagonists
Progressive N-terminal shortening in the PACAP-27 and PACAP-38 series reduces markedly the binding
195
potency. PACAP(2-27) is already 50-fold less potent than PACAP-27 for binding. The evolution of IC50 values is somewhat discontinuous: for instance the IC50 of PACAP(6-27) and PACAP(6-38) are lower, i.e., the affinity is higher than for the longer fragments PACAP(3-27) and PACAP(3-38), respectively.
PACAP(2-27) retains 90% of its I.A. on adenylate cyclase but the deletion of His-l-Ser-2, as well as further deletion suppress the capability to stimulate the enzyme (I.A. = 0). Fittingly shortened fragments are, therefore, candidate competitive antagonists of PACAP effects: they shift indeed the adenylate cyclase activation curve of PACAP-27 to the right. The most potent full inhibitors in each series are PACAP(6-27) and PACAP(6-38) with a K i of 100 nM and 1.5 nM, respectively.
PACAP(6-38) is a potent antagonist of PACAP effects but does not allow to discriminate type I from type II receptors in terms of binding and adenylate cyclase activation as PACAP itself binds to both recep- tors. Perhaps PACAP(6-38) inhibition of [Ca2÷]i ele- vation could be more discriminative in some in vitro systems since type I receptors are much better coupled to the PLC cascade than type II receptors. At any rate, when both receptors coexist, their relative contribution could be better ascertained by using a potent and selective VIP antagonist, whenever available.
XVII-E. Importance of the C-terminal extension in PACAP-38
The (28-38) C-terminal extension of PACAP-38 contains six hydrophilic amino acids. When His-1 is normally present, PACAP-27 and PACAP-38 show similar potency and I.A., indicating that the hy- drophilic C-terminal extension in PACAP-38 is then not important for enzyme activation. This is no longer true after N-terminal deletion or after several mono- substitutions. The resulting analogs show higher affin- ity when derived from PACAP-38 rather than from PACAP-27, except in the case of [Ala-2]PACAP-27. The smallest difference (a 10-fold factor) is between PACAP(2-27) and PACAP(2-38) and the highest dif- ference (75-fold) is between PACAP(9-27) and PACAP(9-38). This observation can be extended to gac t or g i values that are disproportionately high for low affinity PACAP-27 analogs, compared to their longer PACAP-38 counterparts.
XVII-F. Conclusions
Three anchorage domains of PACAP-38 contribute to drive stimulus-effector coupling in PACAP type I receptors: the N-terminal triplet His-Ser-Asp, the (11- 27) area and the C-terminal (28-38) extension. His- 1,Ser-2,Asp-3 and the (11-27) area are key elements

196
for binding (see IC50 values) and activation (see Kac t value), while the (3-5) bridge opposes binding if not preceded by His-l,Ser-2. The reinforcing interaction of the large (29-38) highly cationic C-terminal extension of PACAP-38 accounts for the higher potency and efficacy (i.e., better coupling) of N-terminally short- ened or modified PACAP-38 analogs as compared to PACAP-27 equivalents, and may imply a conforma- tional modification of the NH 2-terminus in PACAP-38, whose properties are then less pH-dependent than in PACAP-27 (Ref. 82 and see above).
XVIII. Tentative molecular structure of the type I PACAP receptor
In membranes from rat brain [12,57], rat cultured astrocytes [97], human neuroblastoma NB-OK-1 cells [11], and rat pancreatic acinar cell line AR 4-2J [7], the cross-linking of radioiodinated PACAP reveals a pro- tein species of 57-62 kDa. Type I receptors solubilized from rat-brain plasma membranes with Triton X-100 form a 400 kDa complex in association with G protein(s) [85]. Those from bovine brain membranes (molecular mass 57 kDa) when solubilized with CHAPS retain high affinity (Ka 200 pM) and specificity for PACAP [49,71].
The full permanent expression of cloned type I PACAP receptor cDNA is required to establish the binding characteristics and coupling properties of the receptor with specific G proteins. To approach this goal, two eDNA fragments have been PCR amplified at low-stringency, from the rat eDNA of AR 4-2J acinar cells differentiated with dexamethasone by using two degenerated primers based on conserved se- quences in the 2nd and 6th transmembrane segments of secretin receptors [95]. Apart from a canonic se- cretin receptor fragment, a longer band contains a related yet original sequence in which an added 84 bp insert corresponds to 28 more amino acids in the putative third intracellular loop (with four hemi-cystines and several serine, threonine and basic residues). This subcloned fragment, when utilized as probe for North- ern blot analysis, allows to observe a high molecular mass transcript (about 7 kb) in AR 4-2J cells and also in rat brain. After screening a lambda-ZAP bacterio- phage library of AR 4-2J eDNA, one clone with an ORF of 1269 bp, when translated to a 423 amino acid peptide, shows 201 identities with VIP receptors and 185 identities with secretin receptors. About 200 bp are still missing at the 5' end. This almost complete se- quence may correspond to PACAP receptors.
XIX. Summary and conclusions
Among vertebrates, there is an extreme conserva- tion in amino acid sequence for the neuropeptide
PACAP-38 and its C-terminal shortened derivative PACAP-27. The PACAP gene is assigned to chromo- some 18 in man and its organization has been charac- terized. PACAP-38 and its minor derivative PACAP-27 are widely distributed in the central nervous system. PACAP-38 is particularly abundant in hypothalamus. The mapping of the afferentation and efferentation of PACAP systems are progressively delineated, including a search for the colocalization with other neurotrans- mitters. In several peripheral organs positive neuronal perikarya and fibers are also seen.
PACAP acts through two types of receptors: (1) the highly selective type 1 that displays a 500 to 2000 selectivity for PACAP-38 and PACAP-27 as compared to VIP; (2) type II is the so-called VIP receptor show- ing similar high affinity for PACAP-38, PACAP-27 and VIP. It is less selective, therefore, than previously thought. This is why this second receptor, qualifying as an unspecific VIP-PACAP receptor, is hardly consid- ered here.
Type I receptors can stimulate two enzymes: the adenylate cyclase and phospholipase C (whose activa- tion leads to the inositol phosphate-cytosolic Ca 2+ cascade). This dual coupling may have several distal consequences including on gene expression, cell growth and differentiation.
Although a relatively comprehensive spectrum of pharmacological activities has already been established we still need to limit the physiological roles of PACAP as neurotransmitter and /o r neuromodulator.
Concerning the hypothalamo-pituitary axis, PACAP reduces food intake in mice and raises plasma arginine vasopressin in rat, probably through PACAP-ir neu- rons in paraventricular and supraoptic nuclei project- ing to the neurohypophysis. PACAP originating in the hypothalamus may also be transported to the anterior pituitary through portal vessels. Data on the antehy- pophysis suggest a role on i.a. reproduction and growth. PACAP stimulates adenylate cyclase and increases [Ca 2+] in gonadotropes, somatotropes, and folliculo- stellate cells. It elevates the secretion of a-MSH from melanotropes, and that of interleukin-6 from pituitary folliculo-stellate ceils. PACAP potentiates the effects of LHRH on LH and FSH secretion. More clearly perhaps, PACAP increases the synthesis of LH, GH, PRL and ACTH after 1-2 days. In human pathology, PACAP-27 and PACAP-38 stimulate adenylate cyclase activity in membranes from 'null' -, gonadotropin-, GH-, and ACTH-producing pituitary adenomas but are inactive in prolactinomas.
In brain, and outside the hypothalamus, PACAP stimulates the synthesis of melatonin from pinealocytes and may play a role in the integration of visual inputs and in memory. The widespread distribution of specific type I PACAP receptors contrasts with the more fo- cused localization of type II VIP-PACAP receptors.

197
Type I receptors in the rabbit eye may contribute to the physiology of the retina and choroid. In general, the neuronal increases in cAMP and Ca 2÷ could con- tribute to early brain development, repair, and modifi- cation of circuitry, with the help of neurotropic factors from astrocytes that are also stimulated by PACAP.
Type I receptors represent a minority (20%) as compared to type II receptors in rat liver. PACAP directly relaxes the digestive tract motility via activa- tion of apamin-sensitive calcium-activated K ÷ channels and is an anion secretory neuropeptide on rat jejunal mucosa, possibly through submucous neurons. PACAP contracts the gallbladder in conscious dog by a pregan- glionic mechanism. In vivo it increases amylase secre- tion via a cholinergic mechanism in dog and glucagon release in mice.
PACAP provokes the hypersecretion and synthesis of catecholamines from the rat adrenal and facilitates mitosis, neurite genesis, and survival of cultured rat sympathetic neuroblasts. A bolus i.v. injection of PACAP induces a biphasic change in blood pressure (decrease followed by increase) that is accompanied by a biphasic change in systemic vascular resistance. The pressor response is due to the release of catechol- amines from the adrenal gland. The relaxant action of PACAP in isolated rabbit precontracted aortic rings is 100-fold more potent than VIP, surprisingly durable, and endothelium-independent. PACAP is also a potent microvasodilator in human skin and here its action is long-lasting again. PACAP-38 causes an increase in beating rate in neonatal rat cardiomyocytes and stimu- lates adenylate cyclase in purified sarcolemmal mem- branes from porcine and canine ventricles (with higher potency than VIP). PACAP relaxes the airway and vascular smooth muscle in guinea-pig, rat and cat lung. PACAP-ir is abundant in the rat testis and type I receptors in spermatozoa may perhaps play a role in sperm motility.
In addition to this already imposing 'hochepot' of actions on a variety of target organs, PACAP may intervene in ontogenetic processes in the nervous sys- tem and modulate mitogenesis and differentiation (e.g., neurite outgrowth) in several cell lines where malig- nancy is sometimes at the origin of type I receptor emergence. The case for mouse AtT-20 corticotropes, the human neuroblastoma cell line NB-OK, rat pheochromocytoma PC12 cells, the rat pancreatic aci- nar cell line AR 4-2J and human THP-1 leukemic monocytes is briefly considered.
Finally, an in vitro molecular pharmacology ap- proach with several analogs makes it possible to demonstrate that PACAP-38 derivatives are in general more potent than the corresponding PACAP-27 analogs, and that PACAP(6-38) is a potent N-termi- nally shortened competitive antagonist. The require- ments for PACAP binding and stimulus-effector cou-
piing with G s and Gq will be better understood once a permanent expression of cloned type I PACAP recep- tors has been achieved. At present a large part of a candidate receptor is already sequenced.
Acknowledgements
Experiments from our laboratory referred to in this work were supported by Grants 3.4525.91 from the Fund for Medical Scientific Research (Belgium), grant SCI-CT91-0632 from the European Economic Com- munity, and the 'Concerted Research Action' 87/92408 from the Ministry of Scientific Policy (Belgium). I am indebted to my colleagues A. Cauvin, C. Delporte, P. Robberecht, M.-C. Vandermeers-Piret, A. Vander- meers, P. Vertongen, J. Winand and M.-C. Woussen- Colle for sharing a common enthusiasm for PACAP and to our visiting guests L. Buscail, X. Hou, S. Nilsson and L. Will-Shabab for their distinguished participa- tion. I wish also to thank the skillful contribution of M. Sti6venart and N. Peuchot in preparing this manuscript.
References
1 Absood, A., Chen, D. and H?~kanson, R. (1992) Regul. Pept. 40, 311-322.
2 Absood, A., Chen, D., Wang, Z.-Y. and H~tkanson, R. (1992) Regul. Pept. 40, 323-329.
3 Araki, N. and Takagi, K. (1992) Eur. J. Pharmacol. 216, 113-117. 4 Arimiira, A., Brenneman, D.E. and Kitada, C. (1990) The En-
docrine Society, 72nd Annual Meeting, Atlanta. 5 Arinura, A., Somogyv~ri-Vigh, A., Miyata, A., Mizuno, K., Coy,
D,H. and Kitada, C. (1991) Endocrinology 129, 2787-2789. 6 Arimura, A. (1992) Regul. Pept. 37, 287-303. 7 Buscall, L., Gourlet, P., Cauvin, A., De Neef, P., Gossen, D.,
Arimura, A., Miyata, A., Coy, D.H., Robberecht, P. and Christophe, J. (1990) FEBS Lett. 262, 77-81.
8 Buscail, L., Cambillau, C., Seva, C., Scemama, J.L., De Neef, P., Robberecht, P., Christophe, J., Susini, C. and Vaysse, N. (1992) Gastroenterology 103, 1002-1008.
9 Canny, B.J., Rawlings, S.R. and Leong, D.A. (1992) Endocrinol- ogy 130, 211-215.
10 GardeIl, L.O., Uddman, R., Luts, A. and Sundler, F. (1991) Regul. Pept. 36, 379-390.
11 Cauvin, A., Buscail, L., Gourlet, P., De Neef, P., Gossen, D., Arimura, A., Miyata, A., Coy, D.H., Robberecht, P. and Christophe, J. (1990) Peptides 11,773-777.
12 Cauvin, A., Robberecht, P., De Neef, P., Gourlet, P., Vander- meers, A., Vandermeers-Piret, M.-C. and Christophe, J. (1991) Regul. Pept. 35, 161-173.
13 Chartrel, N., Tonon, M.C., Vaudry, H. and Conlon, J.M. (1991) Endocrinology 129, 3367-3371.
14 Chedeville, A., Mirossay, L., Chastre, E., Hurbain-Kosmath, I., Lopez, M. and Gespach, C. (1993) FEBS Lett. 319, 171-176.
15 Cox, H.M. (1992) Br. J. Pharmacol. 106, 498-502. 16 Culler, M,D. and Paschall, C.S. (1991) Endocrinology 129,
2260-2262, 17 Delporte, C., Van Praet, A., Herchuelz, A., Winand, J. and
Christophe, J., (1993) Peptides, in press. 18 Deutsch, P.J. and Sun, Y. (1992) J. Biol. Chem. 267, 5108-5113. 19 DiCicco-Bloom, E. and Deutsch, P.J. (1992) Regul. Pept. 37,
319.

198
20 Felley, C.P., Qian, J.-N., Mantey, S., Pradhan, T. and Jensen, R.T. (1992) Am. J. Physiol. 263, G901-G907.
21 Fridolf, T., Sundler, F. and Ahr~n, B. (1992) Cell Tissue Res. 269, 275-279.
22 Chatei, M.A., Takahashi, K., Suzuki, H., Gardiner, J.V., Jones, P.M. and Bloom, S.R. (1992) Biomed. Res. 13, 337-342.
23 Ghatei, M.A., Takahashi, K., Suzuki, Y., Gardiner, J., Jones, P.M. and Bloom, S.R. (1993) J. Endocrinol. 136, 159-166.
24 Goth, M.I., Lyons, C.E., Canny, B.J. and Thorner, M.O. (1992) Endocrinology 130, 939-944.
25 GottschalI, P.E., Tatsuno, I. and Arimura, A. (1991) FASEB J. 5, 194-199.
26 Gourlet, P., Woussen-Colle, M.C., Robberecht, P., De Neef, P., Cauvin, A., Vandermeers-Piret, M.C., Vandermeers, A. and Christophe, J. (1991) Eur. J. Biochem. 195, 535-541.
27 Gourlet, P., De Neef, P., Woussen-Colle, M.C., Vandermeers, A., Vandermeers-Piret, M.C., Robberecht, P. and Christophe, J. (1991) Biochim. Biophys. Acta 1066, 245-251.
28 Gracia-Navarro, F., Lamacz, M., Tonon, M.C. and Vaudry, H. (1992) Endocrinology 131, 1069-1074.
29 Grundemar, L. and H~kanson, R. (1992) Regul. Pept. 40, 331- 337.
30 Hart, G.R., Gowing, H. and Burrin, J.M. (1992) J. Endocrinol. 134, 33-41.
31 Heindel, J.J., Powell, C.J,, Paschall, C.S., Arimura, A. and Culler, M.D. (1992) Biol. Reprod. 47, 800-806.
32 Hosoya, M., Kimura, C., Ogi, K., Ohkubo, S., Miyamoto, Y., Kugoh, H., Shimizu, M., Onda, H., Oshimura, M., Arimura, A. and Fujino, M. (1992) Biochim. Biophys. Acta 1129, 199-206.
33 lnooka, H., Endo, S., Kitada, C., Mizuta, E. and Fujino, M. (1992) Int. J. Pept. Prot. Res. 40, 456-464.
34 Ishizuka, Y., Kashimoto, K., Mochizuki, T., Sato, K., Ohshima, K. and Yanaihara, N. (1992) Regul. Pept. 40, 29-39.
35 Jarry, H., Leonhardt, S., Schmidt, W.E., CreutzfeIdt, W. and Wuttke, W. (1992) Life Sci. 51,823-830.
36 Kashimura, J., Shimosegawa, T., Kikuchi, Y., Koizumi, M. and Toyota, T. (1991) Tohoku J, Exp. Med. 164, 309-318.
37 Kawai, K., Ohse, C., Watanabe, Y., Suzuki, S., Yamashita, K. and Ohashi, S. (1992) Life Sci. 50, 257-261.
38 Kimura, C., Ohkubo, S., Ogi, K., Hosoya, N., Itoh, Y., Onda, H., Miyata, A., Jiang, L., Dahl, R.R., Stibbs, H.H., Arimura, A. and Fujino, M. (1990) Biochem. Biophys. Res. Commun. 166, 81-89.
39 Kivipelto, L., Absood, A., Arimura, A., Sundler, F., H~kanson, R. and Panula, P. (1992) J. Chem. Neuroanat. 5, 85-94.
40 Koch, B. and Lutz-Bucher, B. (1992) Regul. Pept. 38, 45-53. 41 Koch, B. and Lutz-Bucher, B. (1992) Mol. Cell. Endocrinol. 87,
79-86. 42 K6ves, K., Arimura, A., Somogyvfiri-Vigh, A., Vigh, S. and
Miller, J. (1990) Endocrinology 127, 264-271. 43 K6ves, K., Arimura, A., G6rcs, T.G. and Somogyvfiri-Vigh, A.
(1991) Neuroendocrinology 54, 159-169. 44 Kuwahara, A., Kuwahara, Y., Suzuki, T., Sakai, Y., Mochizuki,
T. and Yanaihara, N. (1992) Biomed. Res. 13, 343-346. 45 Lam, H.-C., Takahashi, K., Ghatei, M.A., Kanse, S.M, Polak,
J.M. and Bloom, S.R. (1990) Eur. J. Biochem. 193, 725-729. 46 Le Meuth, V., Farjaudon, N., Bawab, W., Chastre, E., Rosselin,
G., Guilloteau, P. and Gespach, C. (1991) Am. J. Physiol. 260, G265-G274.
47 Leonhardt, S., Jarry, H., Kreipe, A., Werstler, K. and Wuttke, W. (1992) Neuroendocrinol. Lett. 14, 319-328.
48 Luis, J. and Said, S.I. (1990) Peptides 11, 1239-1244. 49 Masuda, Y., Ohtaki, T., Kitada, C., Tsuda, M., Arimura, A. and
Fujino, M. (1990) Biochem. Biophys. Res. Commun. 172, 709- 714.
50 Masuo, Y., Ohtaki, T., Masuda, Y., Nagai, Y., Suno, M., Tsuda, M. and Fujino, M. (1991) Neurosci. Lett. 126, 103-106.
51 Masuo, Y., Ohtaki, T., Masuda, Y., Tsuda, M. and Fujino, M. (1992) Brain Res. 575, 113-123.
52 Masuo, Y., Suzuki, N., Matsumoto, H., Tokito, F., Matsumoto, Y., Tsuda, M. and Fujino, M. (1993) Brain Res. 602, 57-63.
53 Minkes, R.K., McMahon, T.J., Hood, J.S., Murphy, W.A., Coy, D.H., McNamara, D.B. and Kadowitz, P.J. (1992) J. Appl. Physiol. 72, 1212-1217.
54 Minkes, R.K., McMahon, T.J., Higuera, T.R., Murphy, W.A., Coy, D.H. and Kadowitz, P.J. (1992) Am. J. Physiol. 263, H1659-H1669.
55 Miyata, A., Arimura, A., Dahl, R.R., Minamino, N., Uehara, A.. Jiang, L., Culler, M.D. and Coy, D.H. (1989) Biochem. Biophys. Res. Commun. 164, 567-574.
56 Miyata, A., Jiang, L., Dahl, R.D., Kitada, C., Kubo, K., Fujino, M., Minamino, N. and Arimura, A. (1990) Biochem. Biophys. Res. Commun. 170, 643-648.
57 Mizumoto, A., Fujimura, M., Ohtawa, M., Ueki, S., Hayashi, N., itoh, Z., Fujino, M. and Arimura, A. (1992) Regul. Pept. 42, 39-50.
58 Mochizuki, T., Sato, K., Ohshima, K., Takahata, H., Kakuyama, H. and Yanaihara, N. (1992) Biomed. Res. 13, 389-392.
59 Moody, T.W., Zia, F. and Makheja, A. (1993) Peptides 14, 241-246.
60 Morley, J.E., Horowitz, M., Morley, P.M.K. and Flood, J.F. (1992) Peptides 13, 1133-1135.
61 Mungan, Z., Ertan, A., Hammer, R.A. and Arimura, A. (1991) Peptides 12, 559-562.
62 Mungan, Z., Ozmen, V., Ertan, A. and Arimura, A. (1992) Regul. Pept. 38, 199-206.
63 Mungan, Z., Arimura, A., Ertan, A., Rossowski, W.J. and Coy, D.H. (1992) Scand. J. Gastroenterol. 27, 375-380.
64 Murase, T., Kondo, K., Otake, K. and Oiso, Y. (1993) Neuroen- docrinology, in press.
65 Nandha, K.A., Benito-Orfila, M.A., Smith, D.M., Ghatei, M.A. and Bloom, S.R. (1991) J. Endocrinol. 129, 69-73.
66 Naruse, S., Suzuki, T. and Ozaki, T. (1992) Pancreas 7, 543-547. 67 Nguyen, T.D., Heintz, G.G. and Cohn, J.A. (1992) Gastroen-
terology 103, 539-544. 68 Nilsson, S.F.E., De Neef, P., Robberecht, P. and Christophe, J.
(1993) Invest. Ophthalmol. Vis. Sci. 34, 825. 69 Ogi, K., Kimura, C., Onda, H., Arimura, A. and Fujino, N.
(1990) Biochem. Biophys. Res. Commnn. 173, 1271-1279. 70 Ohkubo, S., Kimura, C., Ogi, K., Okazaki, K., Hosoya, M.,
Onda, H., Miyata, A., Arimura, A. and Fujino, M. (1992) DNA Cell Biol. 11, 21-30.
71 Ohtaki, T., Watanabe, T., lshibashi, Y., Kitada, C., Tsuda, M., Gottschall, P.E., Arimura, A. and Fujino, M. (1990) Biochem. Biophys. Res. Commun. 171, 838-844.
72 Okazaki, K., Kimura, C., Kosaka, T., Watanabe, T., Ohkubo, S., Ogi, K., Kitada, C., Onda, H. and Fujino, N. (1992) FEBS Lett. 298, 49-56.
73 Osuga, Y., Mitsuhashi, N. and Mizuno, M. (1992) Endocrinol. Jpn. 39, 153-156.
74 Propato-Mussafiri, R., Kanse, S.M., Ghatei, M.A. and Bloom, S.R. (1992) J. Endocrinol. 132, 107-113.
75 Raufman, J.P., Malhotra, R. and Singh, L. (1991) Regul. Pept. 36, 121-129.
76 Robberecht, P., De Neef, P., Gourlet, P., Cauvin, A., Coy, D.H. and Christophe, J. (1989) Regul. Pept. 26, 117-126.
77 Robberecht, P., Cauvin, A., Gourlet, P. and Christophe, J. (1990) In Second Workshop on Nonadrenergic Noncholinergic Mechanisms (Pauwels, R., Lefebvre, R.A. and Herman, A.G., eds.), Arch. Int. Pharmacodyn. Ther., Vol. 303, pp. 51-66.
78 Robberecht, P., Woussen-Colle, M.C., De Neef, P., Gourlet, P., Buscail, L., Vandermeers, A., Vandermeers-Piret, M.C. and Christophe, J. (1991) FEBS Lett. 286, 133-136.

79 Robberecht, P., Gourlet, P., Cauvin, A., Buscail, L., De Neef, P., Arimura, A. and Christophe, J. (1991) Am. J. Physiol. 260, G97-G102.
80 Robberecht, P., Vertongen, P., Velkeniers, B., De Neef, P., Vergani, P., Raftopoulos, C., Brotchi, J., Hooghe-Peters, E. and Christophe, J.J., Clin. Endocrinol. Metab., in press.
81 Robberecht, P., Gourlet, P., De Neef, P., Woussen-Colle, M.C., Vandermeers-Piret, M.C., Vandermeers, A. and Christophe, J. (1992) Eur. J. Biochem. 207, 239-246.
82 Robberecht, P., Gourlet, P., De Neef, P., Woussen-Coile, M.C., Vandermeers-Piret, M.C., Vandermeers, A. and Christophe, J. (1992) Mol. Pharmacol. 42, 347-355.
83 Salomon, R., Couvineau, A., Rouyer-Fessard, C., Voisin, T., Lavall6e, D., Blais, A., Darmoul, D. and Laburthe, M. (1993) Am. J. Physiol. 264, E294-E300.
84 Sawangjaroen, K., Dallemagne, C.R., Cross, R.B. and Curlewis, J.D. (1992) Peptides 13, 1029-1032.
85 Sch~ifer, H., Schwarzhoff, R., Creutzfeldt, W. and Schmidt, W.E. (1991) Eur. J. Biochem. 202, 951-958.
86 Schw6rer, H., Katsoulis, S., Creutzfeldt, W. and Schmidt, W.E. (1992) Naunyn-Schmiedeberg's Arch. Pharmacoi. 346, 511-514.
87 Shen, Z., Larsson, L.T., Malmfors, G., Absood, A., Hgkanson, R. and Sundler, F. (1992) Cell Tissue Res. 269, 369-374.
88 Shivers, B.D., G6rcs, T.J., Gottschall, P.E. and Arimura, A. (1991) Endocrinology 128, 3055-3065.
89 Simonneaux, V., Ouichou, A. and P6vet, P. (1993) Brain Res. 603, 148-152.
90 Suda, K., Smith, D.M., Ghatei, M.A., Murphy, J.K. and Bloom, S.R. (1991) J. Clin. Endocrinol. Metab. 72, 958-964.
91 Suda, K., Smith, D.M., Ghatei, M,A. and Bloom, S.R. (1992) Neurosci. Lett. 137, 19-23.
92 Sundler, F., Ekblad, E. and H~kanson, R. (1991) Acta Oncol. 30, 419-427.
93 Sundler, P., Ekblad, E., Absood, A., H~kanson, R., K6ves, K. and Arimura, A. (1992) Neuroscience 46, 439-454.
94 Svoboda, M., De Neef, P., Tastenoy, M. and Christophe, J. (1988) Eur. J. Biochem. 176, 707-713.
95 Svoboda, M., Tastenoy, M., Ciccarelli, E. and Christophe, J. (1993) Summer Neuropeptide Conference (Martha's Vinegard, USA, June 20-24, 1993), abstract.
96 Tatsuno, I., Gottschall, P.E. and Arimura, A. (1991) Endocrinol- ogy 128, 728-734.
97 Tatsuno, I., Gottschall, P.E. and Arimura, A. (1991) Peptides 12, 617-621.
98 Tatsuno, I., Somogyv~ri-Vigh, A., Mizuno, K., Gottschall, P.E.,
199
Hidaka, H. and Arimura, A. (1991) Endocrinology 129, 1797- 1804.
99 Tatsuno, I., Yada, T., Vigh, S., Hidaka, H. and Arimura, A. (1992) Endocrinology 131, 73-81.
100 Uddman, R., Luts, A., Arimura, A. and Sundler, F. (1991) Cell Tissue Res. 265, 197-201.
101 Uddman, R., Luts, A., Absood, A., Arimura, A., Ekelund, M., Desai, H., H~kanson, R., Hambreaus, G. and Sundler, F. (1991) Regul. Pept. 36, 415-422.
102 Vertongen, P., Woussen-Colle, M.-C., Cauvin, A., Robberecht, P. and Christophe, J. (1992) Biomed. Res. 13, 377-382.
103 Vertongen, P., Hou, X., Robberecht, P., R6sibois-Gr6goire, A. and Christophe, J. (1992) Regul. Pept. 40, 271.
104 Vigh, S., Arimura, A., K6ves, K., Somogyv~ri-Vigh, A., Sitton, J. and Fermin, C.D. (1991) Peptides 12, 313-318.
105 Vigh, S., Arimura, A., Gottschall, P.E., Kitada, C., Somogyv~ri- Vigh, A. and Childs, G.V. (1993) Peptides 14, 59-65.
106 Wakade, A.R., Guo, X., Strong, R., Arimura, A. and Haycock, J. (1992) Regul. Pept. 37, 331.
107 Warren, J.B., Donnelly, L.E., Cullen, S., Robertson, B.E., Ghatei, M.A., Bloom, S.R. and MacDermot, J. (1991) Eur. J. Pharmacol. 197, 131-134.
108 Warren, J.B., Larkin, S.W., Coughlan, M., Kajekar, R. and Williams, T.J. (1992) Br. J. Pharmacol. 106, 331-334.
109 Warren, J.B., Cockcroft, J.R., Larkin, S.W., Kajekar, R., Macrae, A., Ghatei, M.A. and Bloom, S.R. (1992) J. Cardiovasc. Pharma- col. 20, 83-87.
110 Watanabe, T., Ohtaki, T., Kitada, C., Tsuda, M. and Fujino, M. (1990) Biochem. Biophys. Res. Commun. 173, 252-258.
111 Watanabe, T., Masuo, Y., Matsumoto, H., Suzuki, N., Ohtaki, T., Masuda, Y., Kitada, C., Tsuda, M. and Fujino, M. (1992) Biochem. Biophys. Res. Commun. 182, 403-411.
112 Will-Shabab, L., Kiittner, I., Wallukat, G., Bartel, S., Rob- berecht, P. and Christophe, J. (1993) 1st International Sympo- sium on VIP, PACAP and related regulatory peptides (Bischen- berg, France, September 19-23, 1993), abstract.
113 Yada, T., Vigh, S. and Arimura, A. (1993) Peptides 14, 235-239. 114 Yon, L., Feuilloley, M., Chartrel, N., Arimura, A., Conlon, J.M.,
Fournier, A. and Vaudry, H. (1992) J. Comp. Neurol. 324, 485 -499.
115 Zheng, L., Velkeniers, B., Kazemzadeh, M., Robberecht, P., Christophe, J. and Hooghe-Peeters, E.L. (1993) 6th Meeting European Neuroendocrine Association (Lisboa, Portugal, Octo- ber 6-9, 1993), abstract.










![Neuropeptide Receptors in Pain Circuitries: Useful Targets ... · tion of receptors is sensitive to a multitude of endogenous chemical stimuli. Alterations in ionic environment [15]](https://img.pdfslide.net/doc/110x75/60136b714552cf5a802a7715/neuropeptide-receptors-in-pain-circuitries-useful-targets-tion-of-receptors.jpg)

![EDITORIAL Where now for obesity research? · Neuropeptide Y is a widely distributed peptide that acts on one of five receptors in the brain [5,6]. Activation of two of these NPY receptors,](https://img.pdfslide.net/doc/110x75/5f0ef7d87e708231d441d437/editorial-where-now-for-obesity-research-neuropeptide-y-is-a-widely-distributed.jpg)
















