Embed Size (px)
Citation preview

Aus d e m Institut fur Blutgruppenforschung der Tierzuchtforschung Munchen
Uber den erblichen Polymorphismus des roten Blutfarbstoffes bei Rindern der Hohenrassen
Von D. 0. SCHMID
Eingang des Ms. 5 . 6. 1963
Das Harnoglobin, der rote Blutfarbstoff des Menschen und der Tiere ist chemisch ein Chrornoproteid, eine Verbindung eines niedermolekularen Farb- stoffes, dem Ham als prosthetischer Gruppe, rnit einem Eiweigkorper, dern Globin. Das Globin ist ein histonahnliches basisches Protein, das in seinem hminosaureaufbau, Molekulargewicht und Verhalten im elektrischen Feld eine grofle Ahnlichkeit rnit dem Serumalbumin hat. Die chemische Struktur des Ham ist schon lange bekannt. Sie wurde von H. Fischer und Mitarbeitern aufgeklart und durch Totalsynthese bewiesen. Das Ham ist bei den Hamoglo- binen aller Wirbeltiere gleich. Das Hfirnoglobinmolekul besteht zu 96 O / o aus dem Protein Glohin und zu 4 O/O aus dern Farbstoff, dern Ham. Das Hamo- globin ist kein Strukturelernent der Erythrocytenrnembran, sondern wird nach seiner Bildung erst sekundar an diese Membran adsorbiert. Die biologische Be- deutung des Globins liegt darin, die Farbstoffgruppe in die Lage zu versetzten, eine leicht spaltbare Verbindung rnit Sauerstoff eingezugehen und daniit die Atmungsfunktion des Blutes zu ermoglichen.
Fur den Imrnungenetiker sind die genetisch kontrollierten Globin-Typen als Gen-Markierungssubstanzen von grofier Bedeutung.
Es darf angenommen werden, dafi die Globine und darnit auch die Hamo- globine beirn Menschen und den verschiedenen Tierarten nicht identisch sind. Irn Gegensatz zu den abnorrnen Harnoglobin-Typen des Menschen, deren Entstehung auf Mutation zuriickgefuhrt wird und bei dereii Auftreten gleich- zeitig Blutkrankheiten bestehen, ist bei vielen Tierarten eiii Polyrnorphismus des roten Blutfarbstoffes nachweisbar, der unabhangig von pathologischen Veranderungen auftritt.
Besonders eingehend wurde der Harnoglobin-Polyrnorphismus bei Maus, Schaf und Rind untersucht.
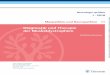
CABANNES und SERAIN (1955) sowie GRIMES und Mitarbeiter (1957, 1958) stellten rnit Hilfe der Papierelektrophorese irn Blut von Rindern neben dem foetalen Hamoglobin, das bereits fast 100 Jahre vorher von KORBER (1866) beschrieben wurde und heute ebenfalls elektrophoretisch von dem Hamo- globin erwachsenen Tiere abgrenzbar ist, zwei weitere Hamoglobine mit ver- schiedenen Wanderungsgeschwindigkeiten fest, die entweder einzeln im Falle einer homozygoten Merkmalsbildung oder aber in Kombination bei Hetero- zygotie nachweisbar sind (Abbildung).
Beirn Vergleich der Papierelektrophorese, dern bisher fast ausschliefllich benutzten Nachweisverfahren fur Hamoglobine, mit der Starkegel-Elektro-
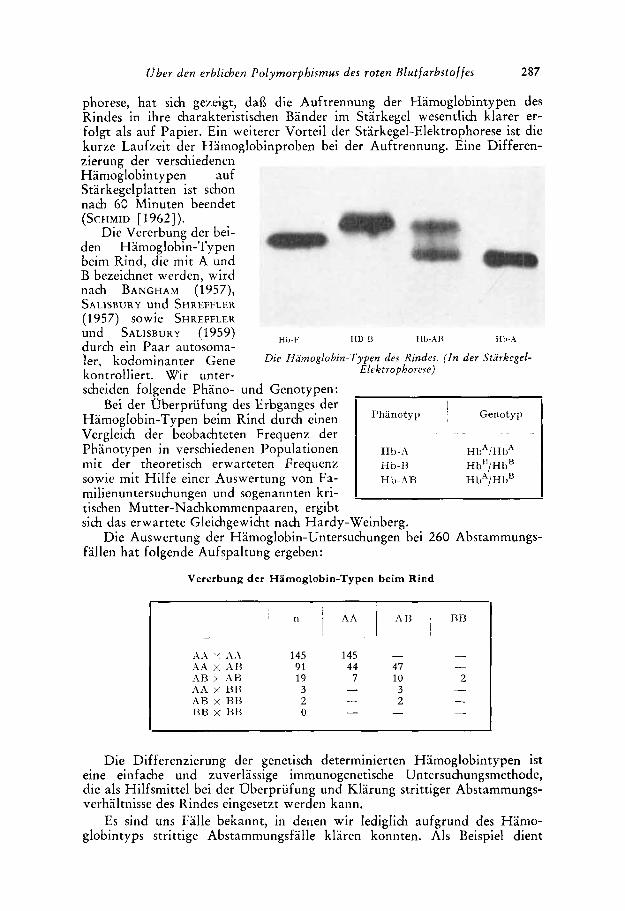
Bei der Oberpriifung des Erbganges der
Vergleich der beobachteten Frequenz der
mit der theoretisch erwarteten Frequenz
Hamoglobin-Typen beini Rind durch einen
Phanotypen in verschiedenen Populationen
sowie mit Hilfe einer Auswertung von Fa- milienuntersuhungen und sogenannten kri-
A.4 “ A.\ 145 145 A.4 x A H 91 44 A\B x A\B 19 7 Alh > nn 3 A B x RI3 2 BB x 131% 0
- - -
I ”hanotyi’ ~ Genoty~l
Hb-A HbA/HbA
Hb-AR HbA/HbU
~ ~~~ ~ .~~ ~~~
Hb-I3 Hb”/Hb”
Die Differenzierung der genetisch determinierten Hamoglobintypen ist eine einfache und zuverlassige immunogenetische Untersuchungsmethode, die als Hilfsmittel bei der Oberpriifung und Klarung strittiger Abstammungs- verhaltnisse des Rindes eingesetzt werden kann.
Es sind uns Falle bekannt, in denen wir lediglich aufgrund des Hamo- globintyps strittige Abstammungsfalle klsren konnten. Als Beispiel dient
288 D. 0. Schmid
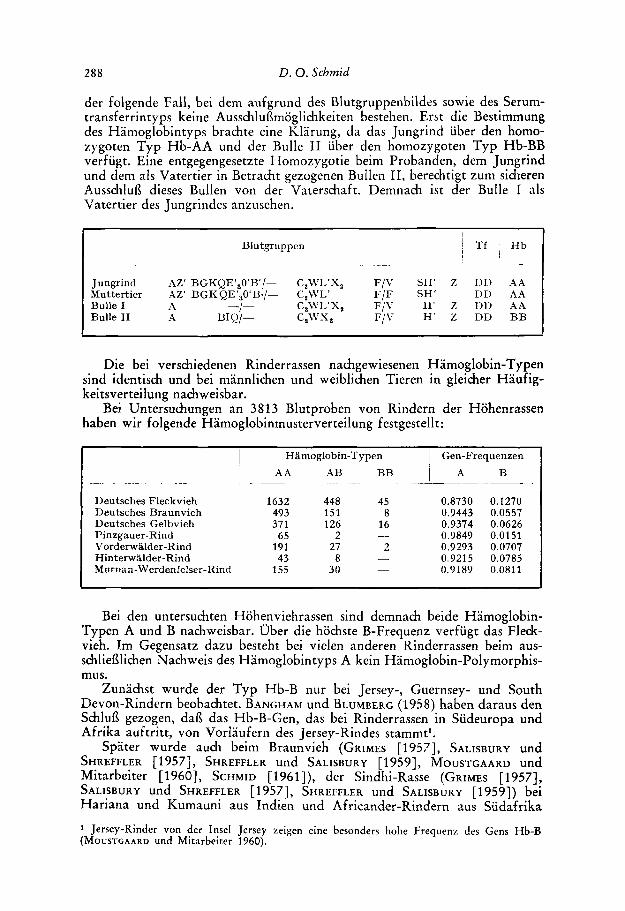
der folgende Fall, bei dem aufgrund des Blutgruppenbildes sowie des Serum- transferrintyps keine Ausschluflmoglichkeiten bestehen. Erst die Bestimmung des Hamoglobintyps brachte eine Klarung, da das Jungrind uber den homo- zygoten Typ Hb-AA und der Bulle I1 uber den homozygoten Typ Hb-BB verfugt. Eine entgegengesetzte Homozygotie beim Probanden, dem Jungrind und dem a h Vatertier in Betracht gezogenen Bullen 11, berechtigt zum sicheren Ausschlufl dieses Bullen von der Vaterschaft. Demnach ist der Bulle I nls Vatertier des Jungrindes anzusehen.
1 1 I Tf ' Hb
I Blu tgruppen
Jungrind AZ' BGKQE'30'B'/- C,WL'X, F / V SH' 2 D n A A Muttertier AZ' BGKQE',O'B./- C,WL' F/F SH' DD A A Bulle I A -I- C,WL'X, F/\- H' 2 DD A A Bulle I1 A BIQ/-- CZWX, F/V H' 2 DD BB
I
Die bei verschiedenen Rinderrassen nachgewiesenen Hamoglobin-Typen sind identisch und bei mannlichen und weiblichen Tieren in gleiher Haufig- keitsverteilung nachweisbar.
Bei Untersuchungen an 3513 Blutproben von Rindern der Hohenrassen haben wir folgende Hamoglobinmusterverteilung festgestellt:
Deutsches Fleckvieh Deutsches Braunvieh Deutsches Gelbvieh Pinzgauer-Rind Vorderwalder-Rind Hintemalder-Rind Murnan-Werdenfelser-Rind
Bei den untersuchten Hohenviehrassen sind demnach beide Hamoglobin- Typen A und B nachweisbar. Ober die hochste B-Frequenz verfugt das Fleck- vieh. Im Gegensatz dazu besteht bei vielen anderen Rinderrassen beim aus- schliefllichen Nachweis des Hamoglobintyps A kein Haniogiobin-Polymorphis- mus.
Zunachst wurde der Typ Hb-B nur bei Jersey-, Guernsey- und South Devon-Rindern beobachtet. BANGHAM und BLUMBERG (1958) haben daraus den Schlufl gezogen, dafl das Hb-B-Gen, das bei Rinderrassen in Sudeuropa und Afrika auftritt, von VorIaufern des Jersey-Rindes stammt'.
Spater wurde auch beim Braunvieh (GRIMES [1957], SALISBURY und SHREFFLER [ 19571, SHREFFLER und SALISBURY [ 19591, MOUSTGAARD und Mitarbeiter [1960], SCHMID [1961]), der Sindhi-Rasse (GRIMES [1957], SALISBURY und SHREFFLER [ 19571, SHREFFLER und SALISBURY [ 19591) bei Hariana und Kumauni aus Indien und Africander-Rindern aus Siidafrika
1 Jersey-Rinder von der Insel Jersey zeigen eine hesonders hohe Frequenz des Gens Hb-B (MOUSTGAARD und Mitarbeirer 1960).
Hamoglobin-T ypen
A A AB BB __ ~ .. ~-
1632 448 45 493 151 8 371 126 16
191 27 2
155 30 -
_- 65 2
43 8 -
1 Ge;Frequenzen
0.8730 0.1270 0.9443 0.0557 0.9374 0.0626 0.9849 0.0151 0.9293 0.0707 0.9215 0.0785 0.9189 0.0811
B ~ _ _

Uber den erblichen Polymorphisrntls des roten Bluifarbstojfes 289
(MOUSTGAARD und Mitarbeiter [ 1960]), Normanner-, Montbeliard- und Piebald-Rindern (NIKOLAJCZUK und Mitarbeiter [ 1961]), sowie beim Deut- schen Fleckvieh, Deutschen Gelbvieh, Pinzgauer-Rind, Vorderwalder-Rind und Hinterwalder-Rind (SCHMID [ 19611) der Hamoglobintyp B festgestellt.
MOUSTGAARD (1 962) hat auch beim Siamesischen Wasserbuffel einen dem Rind vergleichbaren Hamoglobin-Polymorphismus mit den Typen Hb-A und Hb-B gefunden. Wahrend die Wanderungsgeschwindigkeit von Hb-A im elektrischen Feld bei Rind und Wasserbuffel iibereinstimmen, bleibt das fur den Hb-B-Typ charakteristische Band des Wasserbuffels im Vergleich zum B-Band des Rindes etwas zuruck.
Im Gegensatz zu den vorgenannten Rinderrassen ist der Typ Hb-B bisher bei den folgenden Rinderrassen nicht nachgewiesen: Ayrshire, Holstein-Friesian, Aberdeen-Angus, Hereford, Shorthorn, Rotes Danisches Milchvieh, Schwarzbuntes Danisches Milchvieh und Island-Rind (GRIMES [1957], SALISBURY und SHREFFLER [ 1957-19591, MOUSTGAARD und Mitarbeiter [ 19601, HOJGAAKD und Mitarbeiter [ 19601, BRAEND, RENDEL, GAHNE und ADALSTEINSSON [1962]).
BANGHAM und BLUMBERG (1 95 8) beobachteten eine Korrelation zwischen dem Fehlen des Hamoglobintyps B und einer relativ hohen Toleranz gegen- uber der Trypanosomiasis beim Rind. Das Gen Hb-A scheint damit eine ahnliche Rolle zu spieln wie das Gen Hb-S bei der Malaria des Menschen.
LEHMANN und ROLLINSON (1958) fanden bei der Untersuchung von Rin- dern der Ankole-Rasse und der kurzgehornten Zebus in Uganda beide Hamo- globintypen. Die relativ niedrige Frequenz beim Zebu wird von den Autoren auf den groi3eren asiatischen Einflufl auf diese Rinderrasse zuruckgefuhrt.
LEHMANN und ROLLINSON (1 958) diskutieren weiterhin den Zusammen- hang zwischen einer erhohten Resistenz gegenuber dem East Coast-Fever bei Kalbern mit in den ersten Lebensmonaten persistierendem foetalem Hb-F. Xormalerweise ist das Hb-F beim Kalb, das Hb-F-Band liegt zwischen dem Hb-A und Hb-B-Band (SCHMID [1962]), 20 Tage nach der Geburt in der Starkegel-Elektrophorese nicht mehr nachweisbar. Beim menschlichen Saug- ling ist wahrend der Dauer der physiologischen Hb-F-Persistenz eine damit vergleichbare Resistenz gegenuber Malaria festgestellt. Es ergeben sich sornit interessante Parallelen hinsichtlich der Hamoglobintypen und der Wider- standsfahigkeit gegenuber parasitaren Tropenkrankheiten.
Das Vorkommen des Hb-B-Gens beim Bos indicus veranlai3te LEHMANN (1 959) zu Hamoglobinuntersuchungen beim indischen Zebu-Rind. Beim Gir- Rind fand er die gleiche Genfrequenz fur Hb-A und Hb-13 mit einem hohen Uberschui3 an Heterozygoten. Er leitet daraus eine Bevorzugung der Hetero- zygoten im Zuge der naturlichen Selektion dieser Rinderrasse ab.
Zusammenfassung
Im Gegensatz zu vielen anderen Rinderrassen wurde bei den untersuchten Hohenrassen Deutsches Fleckvieh, Deutsches Braunvieh, Deutsches Gelbvieh, Pinzgauer-Rind, Vorderwalder-Rind, Hinterwalder-Rind und Murnau-Wer- denfelser-Rind ein genetisch determinierter Hamoglobin-Polymorphismus nachgewiesen.
Fur die einzelnen Hohenrassen wurden die Gen-Frequenzen fur die Ha- nioglobine A und B berechnet.

290 D. 0. Schmid
Die Bestimmung der erblichen Hamoglobin-Typen mit Hilfe der Star- kegel-Elektrophorese stellt eine einfache und zuverlassige immunogenetische Untersuchungsmethode zur Oberpriifung und Klarung strittiger Abstammungs- verhaltnisse dar.
Summary
In comparison to many other cattle breeds a haemoglobin-polymorphism genetically determined could be demonstrated in the investigated mountain breeds Deutsches Fleckvieh, Deutsches Braunvieh, Deutsches Gelbvieh, Pinz- gauer-Rind, Vorderwalder-Rind, Hinterwalder-Rind and Murnau-Werden- felser-Rind.
The gene frequency for haemoglobin A and B was calculated within the different mountain breed.
The determination of the heritable haemoglobin-types using starchgel- electrophoresis is a simple and reliable immunogenetical method for testing disputed pedigree certificates.
Literatur
1. BANGHAM, A. D., 1957: Distribution of electrophoretically different hemoglobins among cattle breeds of Great Britain. Nature 179, 467.
2. BANGHAM, A.-D., und BLUMBERG, B. S., 1958: Distribution of electrophoretically different hemoglobins among some cattle breeds of Europe and Africa. Nature 181, 1551.
3.
4.
9. 10. 1 1 .
12. 13.
14.
15.
16.
17.
~._..
BRAEND, M., RENDEL, J. GAfINE, B., und ADALSTEINSSO~~, S., 1962: Genetic studies on blood groups, transferrins and hemoglobins in Icelandic cattle. Hereditas 48, 264. CABANNES, R., und SERAIN, C., 1955: Heterogeneite de l’hemoglobine des bovides. Iden- tification electrophoretique de deux hemoglobines bovines. Conipt. rend. SOC. biol. 149, 7. GRIMES, R. M., und DUNCAN, C. W., 1959: Electrophoretic properties of the globins of bovine hemoglobins A and B. Arch. Biochem. Biophys. 84, 393. GRIMES, R. M., DUNCAN, c. w., und LASSISTER, c. A., 1957: Occurence of multiple hemoglobins in certain breeds of dairy cattle. J. Dairy Science 40, 1338. GRIMES, R. M., DUNCAN, C. W., und LASSISTER, C. A., 1958: Bovine fetal hemoglobin. Postnatal persistence and relation to adult hemoglobins. J. Dairy Science 41, 1527. HOJGAARD, N., MOUSTGAARD, J., und MOLLER, I., 1960: Report on Danish investi- gations concerning bovine serum beta-globulins. Rep. Sterility Res. Inst. Roy. Vet. Agric. Coll. 99. KORBER, E., 1866: Ifber Differenzen des Blutfarbstoffes. Inaugural-Dissertation. Dorpat. LEHMANN, H., 1959: The haemoglobins of 103 Gir cattle. Man 59, 66. LEHMANN, H., ROLLINSON, D. H. L., 1958: Themhaemoglobins of 211 cattle in Uganda Man 58, 62. MOUSTGAARD, J., 1962: Personliche Mitteilung. MOUSTGAARD, J., MOLLER, I., und HAVSKOV-SORENSEN, P., 1960: Report on Danish investigations concering polymorphism ob bovine hemoglobin, serum-beta-globulin and bata-lacto-globulin. Immunogenetica Edinburgensis, 122. NIKOLAJCZUK, M., ,COQUEI.ET, A., de TRAVERSE, P. M., 1961: Etude des hemoglobines des bovides a I’aide de l’electrophorese sur papier de la chromatographie et de la denaturation alcaline. Ann. Inst. Pasteur 103, 421. SALISBURY, G. W., SHREFFLER, D. C., 1957: Hemoglobin variants in dairy cattle. J. Dairy Science 40, 1198. SCHMID, D. O., 1962: Die genetische Bedeutung der Hihoglobin-Typen beim Tier. Zentralblatt fur Veterinarmedizin 9, 705. SHREFFLER, D. C., SALISBURY, G. W., 1959: Distribution and inheritance of hemoglobin variants in American cattle. J. Dairy Science 42, 1147.
Ansrhrift des Verfassers: Dr. D. 0. SCHMID, Munchen 22. Reitmorstr. 25