Embed Size (px)
Citation preview

ORIGINAL ARTICLE Reproductive endocrinology
Ulipristal acetate does not impacthuman normal breast tissueLaudine Communal1, Myriam Vilasco1, Justine Hugon-Rodin1,2,†,Aurelie Courtin1,†, Najat Mourra3, Najiba Lahlou4, Sylvie Dumont3,Marc Chaouat5, Patricia Forgez1, and Anne Gompel1,2,*1INSERM-UPMC, UMRS 938, Hopital Saint-Antoine, 75012 Parris, France 2UF de Gynecologie-Endocrinienne, Universite Paris Descartes,AP-HP, Cochin, 75014 Parris, France 3Service d’Anatomie et Cytologie Pathologiques, AP-HP, Hopital Saint-Antoine, 75012 Parris, France4Service de Biologie Hormonale, Universite Paris Descartes, AP-HP, Hopital Cochin, 75005 Parris, France 5Service de Chirurgie Plastique,Universite Paris Diderot, AP-HP, Hopital Saint-Louis, 75011 Paris, France
*Correspondence address. UF de Gynecologie Endocrinienne, Cochin-Port Royal, 53 avenue de l’Observatoire, 75014 Paris, France.Fax: +33158413578; E-mail: [email protected]
Submitted on November 25, 2011; resubmitted on April 30, 2012; accepted on May 22, 2012
background: Antiprogestins are of growing interest for the development of new treatments in the gynecological field. Ulipristal acetate(UPA) is a progesterone receptor (PR) modulator considered for long-term administration in contraception and is currently being registeredfor the treatment of uterine fibroids. In light of the influences of hormonal dysfunction in breast pathologies, the secondary consequences ofchronic UPA therapy need to be established. The aim of this study was to determine UPA actions mediated by PR and glucocorticoid re-ceptor (GR) in normal and transformed breast.
methods: UPA, progesterone (P) and dexamethasone (DEX) effects were observed on PR and GR responsive genes and on proliferationand apoptosis of normal human breast epithelial (HBE) and breast cancer cells. Human normal breast tissue samples were xenografted inathymic mice and treated with estradiol (E2), or E2 + P, or E2 + P + UPA.
results: Analysis of PR and GR reporter gene transactivation and their respective endogenous target genes indicated that UPA exertedanti-progestational and anti-glucocorticoid activity in both types of cells with a more pronounced effect in cancer cells. When combined withP or DEX, UPA limits the proliferation of HBE cells but increases growth in breast cancer cell lines. UPA administration had no impact on themitotic index on xenografted human breast tissue exposed to gonadal hormones at similar concentrations to those present in normalwomen.
conclusions: Although further clinical trials are required to confirm that the results from our experimental models can be extrapo-lated to women treated with UPA, they suggest that such treatment would not be deleterious to normal breast tissue at least for a cycle (28days) of continuous administration.
Key words: Ki67 / cyclin A / BCL2 / FASN / IEX-1
IntroductionAntiprogestins have provided a significant therapeutic benefit towomen since their discovery and clinical exploitation. Antiprogestins,also known as selective progesterone receptor modulators (SPRMs),exert activities ranging from pure progesterone receptor (PR) antag-onism to mixed PR antagonist/agonist actions. SPRMs may alsodisplay glucocorticoid receptor (GR) antagonist properties. Themost well-known SPRM is mifepristone employed in medical termin-ation of pregnancy. Mifepristone was also shown to be effective in
myoma treatment (Eisinger et al., 2005; Esteve et al., 2012).However, because of its marked anti-glucocorticoid activity, long-termtherapies of mifepristone were controversial and led to the developedof other SPRMs (Cadepond et al., 1997). Ulipristal acetate (UPA), alsoknown as VA-2914 or CDB-2914, is an SPRM which efficiently bindsand inhibits PR in progesterone target tissues (Cook et al., 1994;Wagner et al., 1998; Wagner et al., 1996; Attardi et al., 2004). UPAis well characterized for its anti-fertility potency in various animalmodels (Reel et al., 1998; Hild et al., 2000; Gainer and Ulmann,2003; Brenner et al., 2010) and in women (Blithe et al., 2003;
† These two authors have contributed equally.
& The Author 2012. Published by Oxford University Press on behalf of the European Society of Human Reproduction and Embryology. All rights reserved.For Permissions, please email: [email protected]
Human Reproduction, Vol.27, No.9 pp. 2785–2798, 2012
Advanced Access publication on June 26, 2012 doi:10.1093/humrep/des221
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article-abstract/27/9/2785/627029 by guest on 04 April 2019

Chabbert-Buffet et al., 2007). In vitro studies have also determined itsbiological effects in normal myometrial and leiomyoma cells. UPA wasshown to exert an anti-proliferative activity in leiomyoma cells (Xuet al., 2005, 2006; Maruo et al., 2010; Yoshida et al., 2010).
UPA has largely been employed in the gynecological field, becauseof its reduced anti-glucocorticoid activity (Wagner et al., 1999; Attardiet al., 2004). In addition to being marketed as a new generation of emer-gency contraceptive pill (ellaOnew), UPA is currently being consideredfor regular contraception. Preclinical and phase II clinical trials investigat-ing the contraceptive activity of locally delivered UPA are in progress(http://www.popcouncil.org/pdfs/factsheets/RH_ContraceptiveDev.pdf). UPA was registered as treatment for uterine fibroids, and asso-ciated with decreasing bleeding (Graham and Clarke, 1997; Levenset al., 2008; Nieman et al., 2011). Some evidence suggest that UPAmight also exhibit benefits for endometriosis treatment, as well asovarian and breast cancer therapies (Spitz et al., 1996; Cadepondet al., 1997; Klijn et al., 2000; Chabbert-Buffet et al., 2005; Goyenecheet al., 2012).
Long-term UPA exposure may affect hormone-responsive tissues,and particularly breast tissue. Because of the controversial effectsof progesterone in the human normal breast tissue, the effectsof SPRMs need further characterization (Graham and Clarke, 1997).A study on normal breast tissue collected by fine needle aspirationin women treated with mifepristone suggested a potent anti-proliferative effect of this SPRM (Engman et al., 2008). However,UPA treatment has not been explored in normal human epithelialbreast cells and is limited to gene reporter transactivation studies inthe T-47D breast cancer cell line (Attardi et al., 2002, 2004). AsUPA has impending future clinical use, we investigated its PR- andGR-mediated activities on target gene expression, as well as on prolif-eration and apoptosis in primary cultures of normal human breast epi-thelial cells (HBE) and in breast cancer cell lines. We also evaluated therole of UPA in the context of ovarian hormones, using an in vivo ex-perimental model, with normal human breast tissues xenografted inthe nude mice. Our results show that treatment with UPA does notalter normal breast cell proliferation under these conditions.
Materials and Methods
SteroidsThe SPRM UPA and its mono-N-demethylated metabolite CDB-3877(CDB) were kindly provided by HRA-Pharma (Paris, France). 17b estradiol(E2), progesterone (P), dexamethasone (DEX) and mifepristone were pur-chased from Sigma (St Quentin Fallavier, France).
PatientsBreast tissue samples were obtained from 26 consecutive women (aged17–50 years) undergoing surgery for mammoplasty reduction between2008 and 2011. The patients had no history of breast disease and immu-nohistochemical studies including a hematoxylin–phloxine–saffron stainonly showed normal breast tissue. Some of the samples (n ¼ 6) wereused both for the in vitro experiments and xenografts.
Cell culture proceduresT-47D and MCF-7 cell lines were, respectively, maintained in RPMI 1640and in DMEM medium supplemented with 10% fetal calf serum (PAA La-boratory, Les Mureaux, France). T-47D cell line was derived from a human
ductal breast carcinoma and constitutively expressed high amounts of PRand estrogen receptor (ER). Normal HBE cell primary cultures wereobtained from 26 women breast tissues. The procedure used to cultureHBE cells is described in detail by Gompel et al. (1986). HBE cells weremaintained in HAM F10 medium (PAA Laboratory, Les Mureaux,France) containing hydrocortisone (5 ng/ml), Triiodo-L-thyronine(6.5 ng/ml), choleratoxin (10 ng/ml), transferrin (5 mg/ml), insulin(0.016 U/ml), epidermal growth factor (10 ng/ml) (Sigma, St Quentin Fal-lavier, France) and 5% human serum (Etablissement Francais du Sang). Wepreviously demonstrated that HBE primary cultures expressed epithelialmarkers as well as low levels of estradiol receptor (ER) and estradiolinduced PR (Malet et al., 1991; Courtin et al., 2012).
Steroid treatmentsAfter seeding, cells were cultured for 24 h in serum and phenol red freemedium. Then treatments were carried out in a phenol red free mediumcontaining 5% dextran–charcoal-stripped serum. Cells were treated withP or DEX (100 nM), alone or in combination with UPA, mifepristone(1 nM to 1 mM) or CDB (100 nM). Control cells were treated withethanol, at 1:1000 final ethanol concentration alone as vehicle or E2 (10 nM).
Reporter enzyme assaysCells were transfected with reporter gene plasmids containing glucocortic-oid and progesterone responsive elements (GRE/PRE): (i) the MMTV-Lucis a Mouse Mammary Tumor Virus Long Terminal Repeat promoter con-taining one GRE/PRE palindrome and three GRE/PRE hemi-palindromesupstream to firefly luciferase gene in pFC31 vector (ii) the GRE-Luc con-tains six copies of GRE/PRE palindrome upstream to firefly luciferase genein pBL vector. When indicated, HBE cells were transfected with human PRisoforms hPR-A and hPR-B expression plasmids constructed in POP3vectors. Rous Sarcoma Virus promoter upstream to beta galactosidasegene (pRSV-b-Gal) was transfected in each experiment as control. Trans-fections were performed according to the manufacturer instructions usingLipofectamine or Lipofectamine LTX Reagents (Invitrogen, Cergy-Pontoise, France) for breast cancer cell lines or HBE cells, respectively.After 24 h of transfection, breast cancer and HBE cells were treatedwith hormones for 24 or 48 h, respectively. At the end of the experiment,cells were lysed and luciferase activity was determined using the LuciferaseAssay System (Promega, Charbonnieres-les-bains, France). Beta galactosi-dase activity was assessed using the Galacto Star kit (Applied Biosystems,Courtaboeuf, France) to normalize luciferase activity data. Each independ-ent experiment was performed in triplicate.
Real-time quantitative reverse transcriptionPCRTotal RNA was extracted using TriZOL Reagent (Invitrogen, Cergy-Pontoise, France). Two micrograms of total RNA were subjected toreverse transcription (RT) using random primers for 1 h at 378C. Twomicroliters of RT product were diluted (1:10) and subjected to a quanti-tative PCR using sequence-specific primers (375 nM) and Brillant SYBRGREEN QPCR master mix (Fermentas, Saint-Remy-les-Chevreuse,France) on an Mx3000P apparatus (Agilent Technologies, Massy,France). Sequence primers for target genes were: 36B4, upper,5′-gattggctacccaactgttg-3′; lower, 5′-caggggcagcagccacaaa-3′; FASN,upper, 5′-tacatcgactgcatcaggca-3′; lower, 5′-gatactttcccgtcgcatac-3′; ALPL,upper, 5′-tcactctccgagatggtggtggtgg-3′; lower, 5′-ttccttcatggtgcccgtgg-3′;cyclin A, upper, 5′-gcagacggcgctccaagagg-3′, lower 5′-ctgctgctgcgctagacccc-3′; BCL2, upper, 5′-ggacggggtgaactggggga-3′, lower 5′-agttccacaaaggcatccca-3′, IEX-1, upper, 5′-cggtcctgagatcttcacct-3′; lower, 5′-accctcttcagccatcagga-3′; GOS8, upper, 5′-ttcctctactcctgggaagc-3′, lower,
2786 Communal et al.
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article-abstract/27/9/2785/627029 by guest on 04 April 2019

5′-agtcttcacaggccagccag-3′. Thermocycling conditions were 1 cycle at958C for 10 min followed by 40 cycles at 958C for 30 s, 608C for 1 minand 728C for 30 s. Gene expression values were normalized to the house-keeping gene 36B4. Time of steroid treatment was chosen to get theoptimal stimulation for a given gene. ALPL and G0S8 mRNA were analyzedafter 6 h of treatment. IEX-1, FASN and BCL2 were analyzed after 24 h oftreatment. Cyclin A mRNA was analyzed after 24 h for GR responses or48 h for PR responses. Each independent experiment was performed induplicate.
Tritiated thymidine incorporationAfter 24 h of hormonal treatment, cells were incubated with [methyl-3H]thymidine (Perkin Elmer, Courtaboeuf, France) for 48 or 20 h for HBE orcancer cells, respectively, at 378C. After incubation cells were washedtwice with phosphate-buffered saline (PBS) 1X and once with 5% trichlor-oaetic acid (TCA). Cells were incubated in 5% TCA for 15 min at 48C andlysed in NaOH 0.1 N for 30 min at 378C. The total cell lysate was addedto 5 ml of Ecolite scintillation liquid (MP biomedical, Illkirch, France) andradioactivity was counted with a b-counter HIDEX 300SL (ScienceTec,Courtaboeuf, France). Each independent experiment was performed intriplicate.
Flow cytometry analysisAfter 96 or 48 h of hormonal treatment, respectively, for HBE or MCF-7and T-47D cells, cells were washed in PBS, matrix dissociated with accur-ate enzyme (PAA laboratory, Les Mureaux, France) and centrifuged 5 minat 300 g. Cells were fixed and frozen at 2208C in 70% ethanol. Beforeanalysis, cells were washed in PBS and stained with 10 mg/ml propidiumiodide in PBS (containing 0.835 U/ml RNase A; Sigma, St Quentin Falla-vier, France). For each sample at least 10 000 cells were counted in aBD LSR II flow cytometer (BD Biosciences, Le Pont de Claix, France).After gating out doublets and debris, cycle distribution was analyzedusing the ModFit LT software (Verity Software House, USA). Each inde-pendent experiment was performed in triplicate.
Human breast xenografts in mice andpellet treatmentsFour weeks old ovariectomized female NMRInu/nu athymic mice were pur-chased from Janvier laboratory (Le Genest Saint Isle, France). Breast tissuesamples were obtained from six consecutive premenopausal womenundergoing mammoplasty surgery and free of concomitant treatment.Their mean age was 36.5+5.16 years (range: 29–42). The day of themenstrual cycle at the time of the surgery was Days 3, 7, 13, 21, 25and 29 for patient 1–6, respectively. For each patient, human mammarygland structures were cut into 2 × 2 × 2 mm fragments, and four frag-ments were then subcutaneously xenografted onto the back of eachmouse of the experiment. Four treatment groups were performed per ex-periment: control, E2, E2 + P and E2 + P + UPA. Each group containedfour mice. Each patient was treated as a separate experiment. Therefore,for each patient, 64 fragments were grafted in 16 mice. Treatments wereadministered by subcutaneously grafting steroid pellets onto the neck ofeach mouse. The experimental conditions were initially determined byusing a dose range of hormones mixed with cholesterol into the pellets.Blood sample assays were performed after 2 and 4 weeks of treatmentto measure plasmatic hormonal concentrations. Finally, the dose of0.3 mg per pellet for E2 and 20 mg per pellet for P and UPA wereused, as it provides the appropriate blood concentrations. For thecontrol groups, pellets containing only cholesterol were used. To repro-duce menstrual cycle conditions, mice were grafted on the first day of ex-periment with cholesterol, E2 and/or UPA containing pellets and on the
fourteenth day with cholesterol or P containing pellets in control and E2groups or in E + P and E2 + P + UPA groups, respectively. Twenty-eightdays after the start of experiment, mice were sacrificed. Blood was col-lected for each mouse, and serum was frozen at 2208C until hormoneconcentrations analysis. Breast tissue fragments were collected and imme-diately fixed in paraformaldehyde solution for immunohistochemical ana-lysis. All study protocols and environmental conditions of the animalrooms were approved by the French Ethic committee for the care anduse of laboratory animals Charles Darwin.
Hormone concentration analysisEstradiol was measured by radioimmunoassay using Clinical AssaysEstradiol-2 (Sorin Biomedica Diagnostics SpA, Saluggia, Italia). Progester-one levels were evaluated by UPLC-MSMS using Acquity UPLC andQuattro Premier XE (Waters, Milford, MA, USA). UPA concentrationswere measured using LC-MS/MS technique by MPI Research (StateCollege, PA, USA).
Immunohistochemical analysisMitotic index was calculated using the Ki67 antibody, and was determinedfor each breast tissue grafted into mice. Immunohistochemical analyseswere performed using the BOND-MAX workstation (Leica, Nanterre,France). Paraffin sections of breast tissue xenografts were de-waxed andrehydrated before antigen retrieval using citrate retrieval solution (pH6.0) for Ki67 antibody or EDTA retrieval solution (pH 9.0) for PR andERalpha antibodies, for 30 min. Sections were then incubated with Ki67at 1:100 (Novocastra, NCL-L-Ki67-MM1), PR at 1:80 (Biogenex,MU-328-UC) or ER at 1:300 (Novocastra, NCL-L-ER-6F11) monoclonalantibodies. For signal detection, the Kit Bond Polymer Refine Detectionwas used. Reagents were purchased from Menarini-Diagnostic (Rungis,France). A negative control (omitting the first antibody) was included ineach set. For each marker, determination of the ratio of positive cellswas performed on a total of 1000 lobular and 1000 ductal luminal cellsin the four breast tissue fragments grafted into each mouse. For each ex-periment, the final percentages for each treatment were the mean of per-centages obtained in the four mice per group.
Ethical permissionsThe breast samples were obtained from women having signed an informedconsent according to the French law on clinical experimentation (L. 1243-3and L. 1243-4), as a part of a biomedical project including the collectionand conservation of cell cultures and xenografts of breast tissues obtainedfrom women undergoing plastic surgery. The authorization number filedfor this project is 11 826, from the French ethical committee ‘Comitede Protection des Personnes’.
Statistical analysisResults were expressed as mean+ SEM and n represents the number ofindependent experiments. To determine the statistical significance oftreatments, one-way analysis of variance and Tukey–Kramer multiplecomparisons tests were performed to compare the relative efficiency ofeach treatment with the Instat 3 software (GraphPad, USA). When onlytwo treatments were compared, an unpaired t-test was performed, withP , 0.05 considered as significant.
Ulipristal acetate in breast cells 2787D
ownloaded from
https://academic.oup.com
/humrep/article-abstract/27/9/2785/627029 by guest on 04 April 2019

Results
UPA and the PRUPA effects on PR gene transactivationIn order to analyze UPA antagonist properties on PR-induced genetransactivation, normal HBE cells and T-47D breast cancer cell linewere transfected with the MMTV-Luc reporter gene (Fig. 1). In HBEcells which expressed low and variable amounts of PR among patients(Malet et al., 1991), P induced a significant luciferase expression(1.38+0.11 control fold induction, P , 0.05; Fig. 1A). To betterstudy the dose effects of UPA on PR, HBE cells were co-transfectedwith hPR-A and hPR-B expression plasmids (Fig. 1B). UPA and mife-pristone inhibited P induced MMTV-Luc transactivation in a dose-dependent manner from 10 to 1 000 nM. HBE cells were then trans-fected with only hPR-A or hPR-B, to discriminate UPA actionsbetween PR isoforms. UPA was an effective antagonist no matterwhich PR isoform was increased (Fig. 1C and D). In the T-47D cellline, UPA and mifepristone displayed similar and potent PR antagonistactions from 10 to 1000 nM (Fig. 1E). Partial antagonist responseswere also detected at 1 nM (Fig. 1E). In both HBE and T-47D cells,UPA and mifepristone did not exhibit progestational agonist proper-ties on reporter gene transcription (Fig. 1B and E). These results indi-cated that UPA acted as a potent P antagonist in normal andcancerous breast cells.
UPA activity on mRNA expression of PR target genesA hundred nanomolar was the lowest concentration of UPA necessaryto exert a complete antagonist activity in HBE cells. This concentrationwas, therefore, chosen to further study the effects of UPA on specificP target genes in HBE and T-47D cells. Estradiol (E2) was added toHBE cells, in order to increase PR expression as reported in severalprevious studies (Clarke et al., 1997; Anderson, 2001; Courtinet al., 2012).
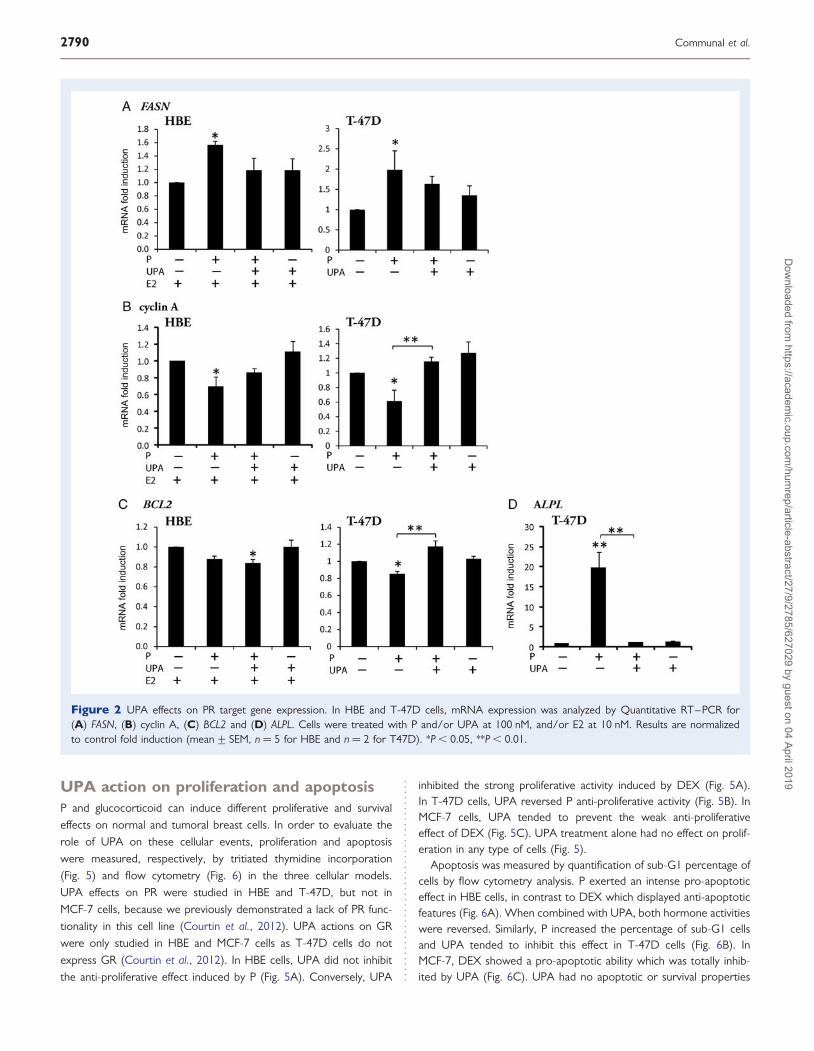
Fatty acid synthase (FASN) is implicated in normal breast cell differ-entiation as well as in mammary tumor progression (Joyeux et al.,1990; Kuhajda, 2006). An up-regulation of FASN mRNA by progestinsthrough PR was previously demonstrated in normal and tumoralbreast cells in vitro and in vivo (Joyeux et al., 1990; Chalbos et al.,1992; Courtin et al., 2012). As shown in Fig. 2A, UPA was able toprevent P induction of FASN mRNA expression in both HBE and inT-47D cells. We observed that P down-regulated cyclin A mRNA ex-pression in normal breast cells and in T-47-D cells (Fig. 2B), as previ-ously reported in MDA-MB-231 breast cancer cells transfected by PR(Lin et al., 2003). UPA reversed P-induced cyclin A mRNA down-regulation in HBE and T-47D cells (Fig. 2B). We previously reporteda decrease of the anti-apoptotic B-cell CLL/lymphoma 2 (BCL2)protein expression under progestin treatment in normal and tumoralbreast cells (Kandouz et al., 1996; Gompel et al., 2000). As shownin Fig. 2C, we also observed a decrease of BCL2 mRNA expressionby P in HBE and T-47D cells (Fig. 2C). Nevertheless, UPA did notreverse BCL2 mRNA down-regulation induced by P in HBE cells,whereas UPA antagonized the P inhibitory effect on this transcriptin T-47D cells (P , 0.01; Fig. 2C). Tissue non-specific alkaline phos-phatase (ALPL) is a P responsive gene implicated in metastasis ofbreast cancer (Di Lorenzo et al., 1993; Ritzke et al., 1998). InT-47D cells, UPA totally inhibited the strong P mediated induction
of ALPL transcript (Fig. 2D). In HBE cells, ALPL mRNA expressionwas not modified by P treatment (data not shown). UPA did notdisplay any PR agonist activity on these genes.
UPA and the GRDue to the similarities in the structure of nuclear receptors PR andGR, antiprogestins also display anti-glucocorticoid properties. Wetherefore evaluated GR antagonist/agonist activity of UPA in normaland tumoral breast cells.
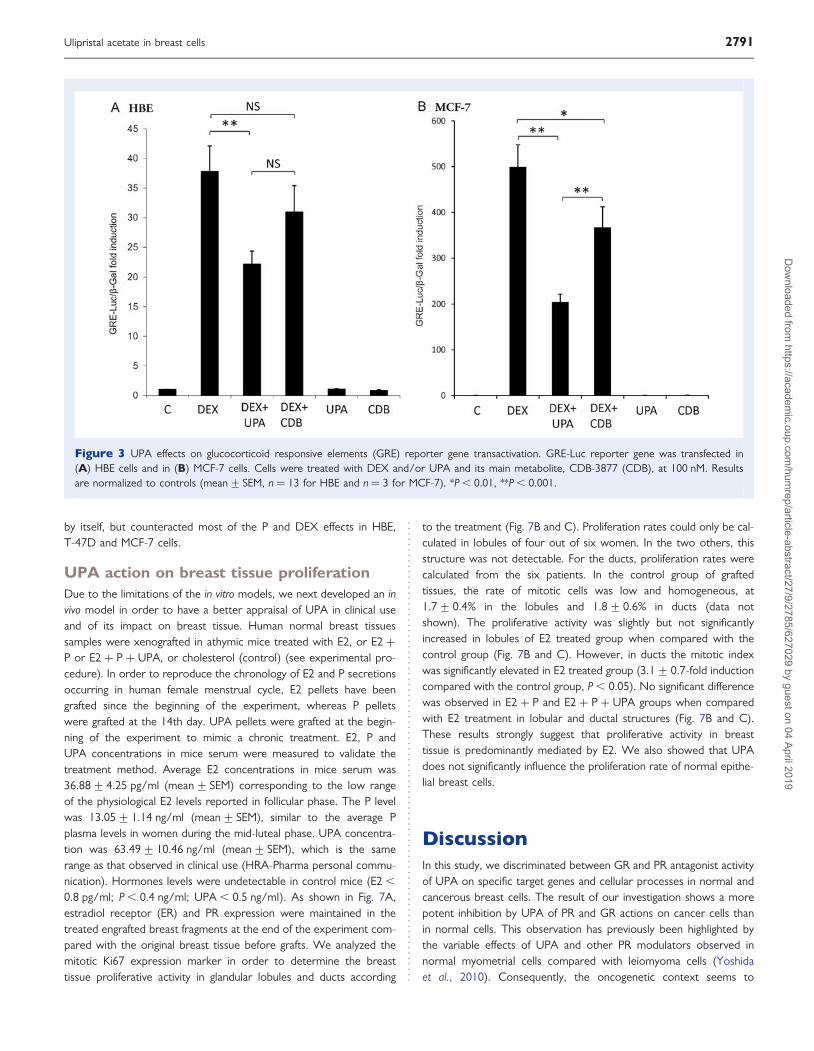
UPA effects on GR gene transactivationThe study was performed in HBE and MCF-7 cells, which expressedhigh amounts of GR as previously reported (Courtin et al., 2012).Cells were transfected with a GRE-Luc reporter gene and treatedwith DEX, UPA and its proximal mono-demethylated metabolite,CDB-3877 (CDB), known to exert less anti-glucocorticoid activitythan UPA (Attardi et al., 2004). We confirmed the reduced anti-glucocorticoid activity of this metabolite, since UPA inhibited by41.3+5.8% the DEX-induced luciferase transactivation, whereas itsmetabolite failed to inhibit DEX activity in HBE cells (18.1+11.8%;Fig. 3A). However, in MCF-7 cells, both UPA and CDB significantlyantagonized by 59+ 3.4 and 26.5+ 9.1%, respectively, the DEX-induced luciferase transactivation (Fig. 3B). These results suggest astronger anti-glucocorticoid potency of UPA in MCF-7 breast cancercells than in HBE.
UPA activity on mRNA expression of GR target genesIn order to better define UPA anti-glucocorticoid effects in HBE andMCF-7 cells, mRNA expressions of various glucocorticoid responsivegenes were analyzed after DEX and UPA treatment. Immediate earlyresponse 3 (IEX-1) and regulator of G-protein signaling 2 (G0S8) havebeen implicated in cell survival under stress conditions (Wu, 2003;Shen et al., 2006) and in G-protein signaling (Kehrl and Sinnarajah,2002), respectively. IEX-1 and G0S8 were characterized as gluco-corticoid responsive genes (Wan and Nordeen, 2002). As shown inFig. 4A, IEX-1 mRNA expression was down-regulated by DEX andUPA did not antagonize this response in HBE and MCF-7 cells. Onthe other hand, UPA exerted a slight and non-significant antagonisteffect on DEX-induced up-regulation of G0S8 mRNA in HBE cells,whereas its anti-glucocorticoid activity was strong in MCF-7 cells(P , 0.001; Fig. 4B). Cyclin A and BCL2 genes were previouslyshown to be regulated by DEX in vitro in osteoblasts and neuroblasto-mas, respectively (Sengupta et al., 2000; Gabet et al., 2011). DEX andUPA actions were therefore analyzed on cyclin A and BCL2 mRNA ex-pression in HBE and MCF-7 cells. Cyclin A was differentially regulatedby DEX in the two types of cells as mRNA expression was induced inHBE cells and repressed in MCF-7 cells (Fig. 4C; Courtin et al., 2012).In both cellular models, UPA partially inhibited DEX effect on cyclin AmRNA expression. BCL2 mRNA was down-regulated by DEX in HBEand in MCF-7 cells (Fig. 4D). However, UPA did not reverse the DEXdown-regulation of BCL2 mRNA in HBE cells, whereas it partiallyantagonized this effect in MCF-7 cells. UPA did not display any gluco-corticoid agonist activity on these genes (Fig. 4).
2788 Communal et al.
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article-abstract/27/9/2785/627029 by guest on 04 April 2019
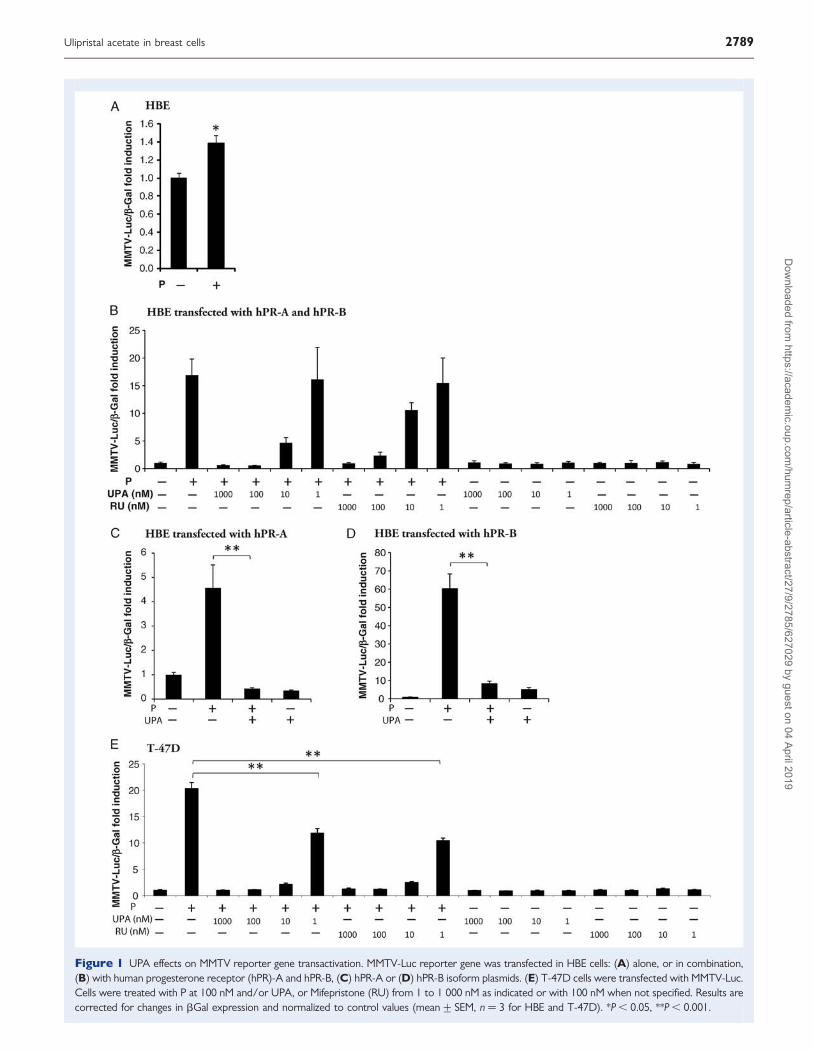
Figure 1 UPA effects on MMTV reporter gene transactivation. MMTV-Luc reporter gene was transfected in HBE cells: (A) alone, or in combination,(B) with human progesterone receptor (hPR)-A and hPR-B, (C) hPR-A or (D) hPR-B isoform plasmids. (E) T-47D cells were transfected with MMTV-Luc.Cells were treated with P at 100 nM and/or UPA, or Mifepristone (RU) from 1 to 1 000 nM as indicated or with 100 nM when not specified. Results arecorrected for changes in bGal expression and normalized to control values (mean+ SEM, n ¼ 3 for HBE and T-47D). *P , 0.05, **P , 0.001.
Ulipristal acetate in breast cells 2789D
ownloaded from
https://academic.oup.com
/humrep/article-abstract/27/9/2785/627029 by guest on 04 April 2019
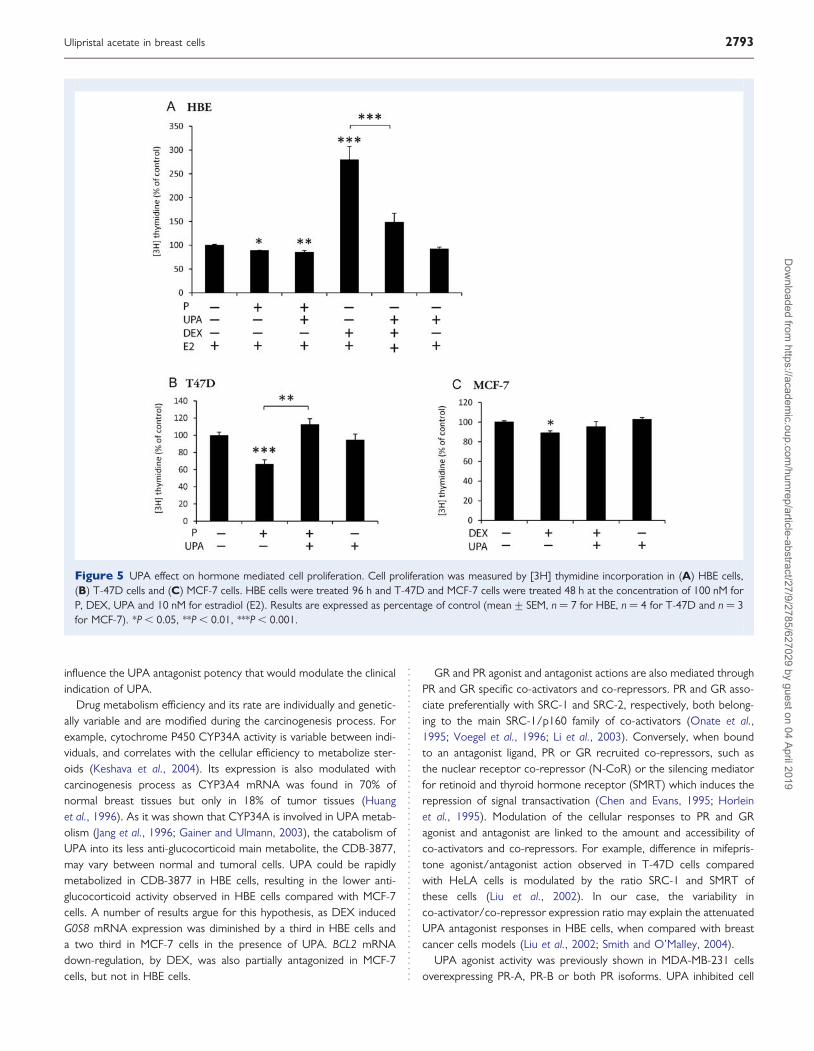
UPA action on proliferation and apoptosisP and glucocorticoid can induce different proliferative and survivaleffects on normal and tumoral breast cells. In order to evaluate therole of UPA on these cellular events, proliferation and apoptosiswere measured, respectively, by tritiated thymidine incorporation(Fig. 5) and flow cytometry (Fig. 6) in the three cellular models.UPA effects on PR were studied in HBE and T-47D, but not inMCF-7 cells, because we previously demonstrated a lack of PR func-tionality in this cell line (Courtin et al., 2012). UPA actions on GRwere only studied in HBE and MCF-7 cells as T-47D cells do notexpress GR (Courtin et al., 2012). In HBE cells, UPA did not inhibitthe anti-proliferative effect induced by P (Fig. 5A). Conversely, UPA
inhibited the strong proliferative activity induced by DEX (Fig. 5A).In T-47D cells, UPA reversed P anti-proliferative activity (Fig. 5B). InMCF-7 cells, UPA tended to prevent the weak anti-proliferativeeffect of DEX (Fig. 5C). UPA treatment alone had no effect on prolif-eration in any type of cells (Fig. 5).
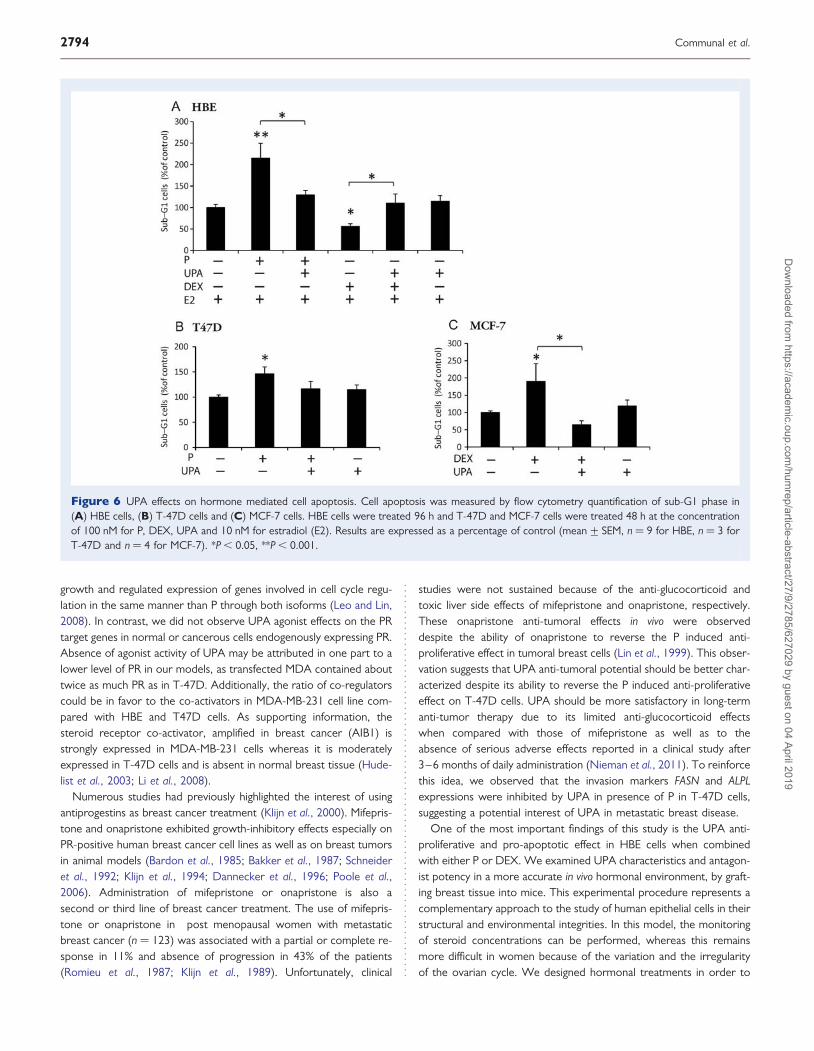
Apoptosis was measured by quantification of sub-G1 percentage ofcells by flow cytometry analysis. P exerted an intense pro-apoptoticeffect in HBE cells, in contrast to DEX which displayed anti-apoptoticfeatures (Fig. 6A). When combined with UPA, both hormone activitieswere reversed. Similarly, P increased the percentage of sub-G1 cellsand UPA tended to inhibit this effect in T-47D cells (Fig. 6B). InMCF-7, DEX showed a pro-apoptotic ability which was totally inhib-ited by UPA (Fig. 6C). UPA had no apoptotic or survival properties
Figure 2 UPA effects on PR target gene expression. In HBE and T-47D cells, mRNA expression was analyzed by Quantitative RT–PCR for(A) FASN, (B) cyclin A, (C) BCL2 and (D) ALPL. Cells were treated with P and/or UPA at 100 nM, and/or E2 at 10 nM. Results are normalizedto control fold induction (mean+ SEM, n ¼ 5 for HBE and n ¼ 2 for T47D). *P , 0.05, **P , 0.01.
2790 Communal et al.
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article-abstract/27/9/2785/627029 by guest on 04 April 2019
by itself, but counteracted most of the P and DEX effects in HBE,T-47D and MCF-7 cells.
UPA action on breast tissue proliferationDue to the limitations of the in vitro models, we next developed an invivo model in order to have a better appraisal of UPA in clinical useand of its impact on breast tissue. Human normal breast tissuessamples were xenografted in athymic mice treated with E2, or E2 +P or E2 + P + UPA, or cholesterol (control) (see experimental pro-cedure). In order to reproduce the chronology of E2 and P secretionsoccurring in human female menstrual cycle, E2 pellets have beengrafted since the beginning of the experiment, whereas P pelletswere grafted at the 14th day. UPA pellets were grafted at the begin-ning of the experiment to mimic a chronic treatment. E2, P andUPA concentrations in mice serum were measured to validate thetreatment method. Average E2 concentrations in mice serum was36.88+4.25 pg/ml (mean+ SEM) corresponding to the low rangeof the physiological E2 levels reported in follicular phase. The P levelwas 13.05+ 1.14 ng/ml (mean+ SEM), similar to the average Pplasma levels in women during the mid-luteal phase. UPA concentra-tion was 63.49+10.46 ng/ml (mean+ SEM), which is the samerange as that observed in clinical use (HRA-Pharma personal commu-nication). Hormones levels were undetectable in control mice (E2 ,
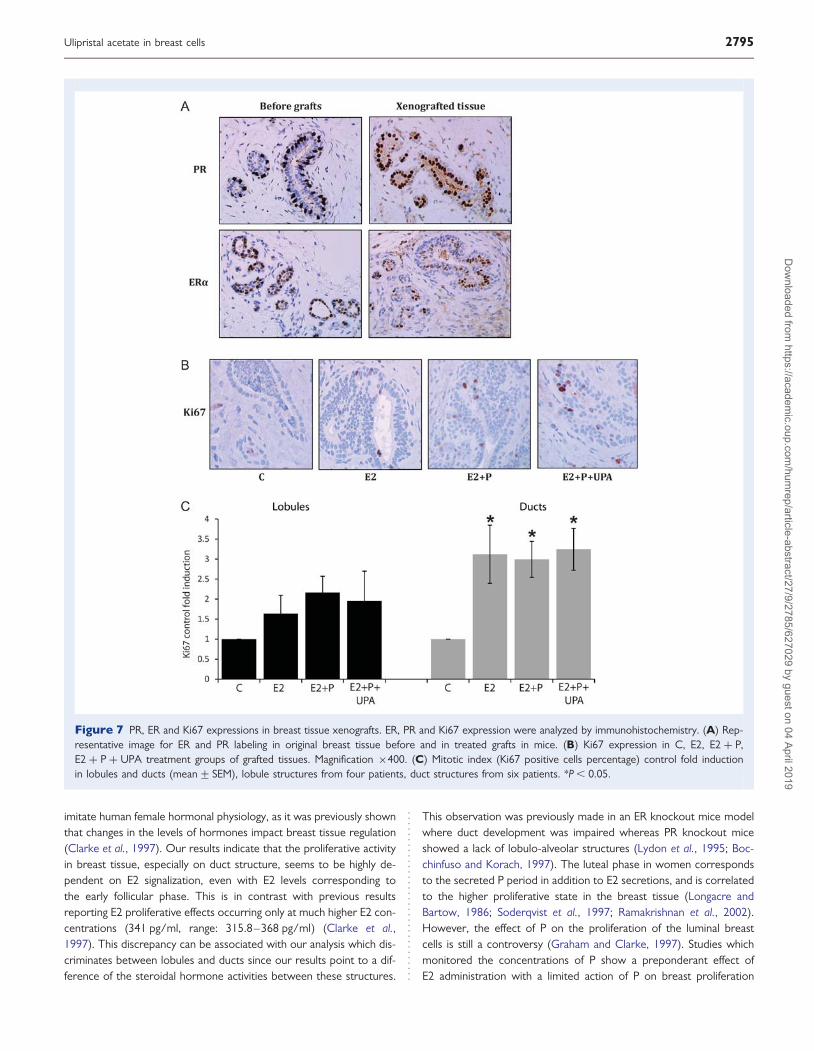
0.8 pg/ml; P , 0.4 ng/ml; UPA , 0.5 ng/ml). As shown in Fig. 7A,estradiol receptor (ER) and PR expression were maintained in thetreated engrafted breast fragments at the end of the experiment com-pared with the original breast tissue before grafts. We analyzed themitotic Ki67 expression marker in order to determine the breasttissue proliferative activity in glandular lobules and ducts according
to the treatment (Fig. 7B and C). Proliferation rates could only be cal-culated in lobules of four out of six women. In the two others, thisstructure was not detectable. For the ducts, proliferation rates werecalculated from the six patients. In the control group of graftedtissues, the rate of mitotic cells was low and homogeneous, at1.7+ 0.4% in the lobules and 1.8+ 0.6% in ducts (data notshown). The proliferative activity was slightly but not significantlyincreased in lobules of E2 treated group when compared with thecontrol group (Fig. 7B and C). However, in ducts the mitotic indexwas significantly elevated in E2 treated group (3.1+ 0.7-fold inductioncompared with the control group, P , 0.05). No significant differencewas observed in E2 + P and E2 + P + UPA groups when comparedwith E2 treatment in lobular and ductal structures (Fig. 7B and C).These results strongly suggest that proliferative activity in breasttissue is predominantly mediated by E2. We also showed that UPAdoes not significantly influence the proliferation rate of normal epithe-lial breast cells.
DiscussionIn this study, we discriminated between GR and PR antagonist activityof UPA on specific target genes and cellular processes in normal andcancerous breast cells. The result of our investigation shows a morepotent inhibition by UPA of PR and GR actions on cancer cells thanin normal cells. This observation has previously been highlighted bythe variable effects of UPA and other PR modulators observed innormal myometrial cells compared with leiomyoma cells (Yoshidaet al., 2010). Consequently, the oncogenetic context seems to
Figure 3 UPA effects on glucocorticoid responsive elements (GRE) reporter gene transactivation. GRE-Luc reporter gene was transfected in(A) HBE cells and in (B) MCF-7 cells. Cells were treated with DEX and/or UPA and its main metabolite, CDB-3877 (CDB), at 100 nM. Resultsare normalized to controls (mean+ SEM, n ¼ 13 for HBE and n ¼ 3 for MCF-7). *P , 0.01, **P , 0.001.
Ulipristal acetate in breast cells 2791D
ownloaded from
https://academic.oup.com
/humrep/article-abstract/27/9/2785/627029 by guest on 04 April 2019
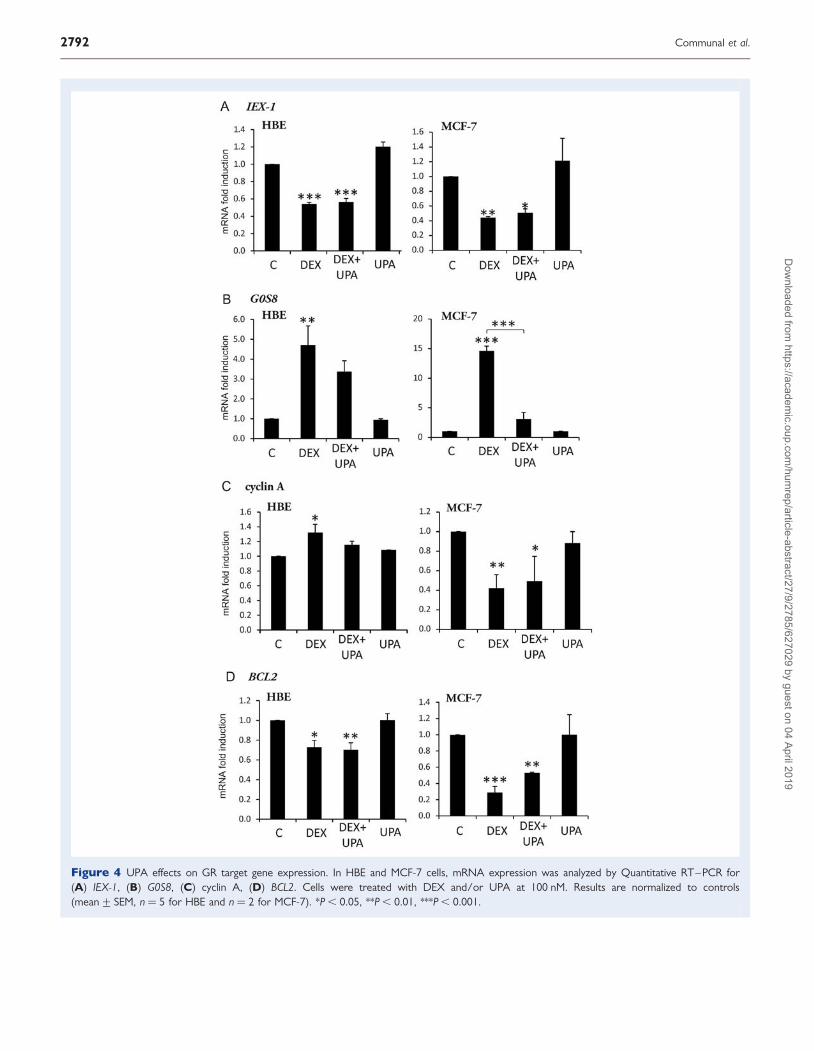
Figure 4 UPA effects on GR target gene expression. In HBE and MCF-7 cells, mRNA expression was analyzed by Quantitative RT–PCR for(A) IEX-1, (B) G0S8, (C) cyclin A, (D) BCL2. Cells were treated with DEX and/or UPA at 100 nM. Results are normalized to controls(mean+ SEM, n ¼ 5 for HBE and n ¼ 2 for MCF-7). *P , 0.05, **P , 0.01, ***P , 0.001.
2792 Communal et al.
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article-abstract/27/9/2785/627029 by guest on 04 April 2019
influence the UPA antagonist potency that would modulate the clinicalindication of UPA.
Drug metabolism efficiency and its rate are individually and genetic-ally variable and are modified during the carcinogenesis process. Forexample, cytochrome P450 CYP34A activity is variable between indi-viduals, and correlates with the cellular efficiency to metabolize ster-oids (Keshava et al., 2004). Its expression is also modulated withcarcinogenesis process as CYP3A4 mRNA was found in 70% ofnormal breast tissues but only in 18% of tumor tissues (Huanget al., 1996). As it was shown that CYP34A is involved in UPA metab-olism (Jang et al., 1996; Gainer and Ulmann, 2003), the catabolism ofUPA into its less anti-glucocorticoid main metabolite, the CDB-3877,may vary between normal and tumoral cells. UPA could be rapidlymetabolized in CDB-3877 in HBE cells, resulting in the lower anti-glucocorticoid activity observed in HBE cells compared with MCF-7cells. A number of results argue for this hypothesis, as DEX inducedG0S8 mRNA expression was diminished by a third in HBE cells anda two third in MCF-7 cells in the presence of UPA. BCL2 mRNAdown-regulation, by DEX, was also partially antagonized in MCF-7cells, but not in HBE cells.
GR and PR agonist and antagonist actions are also mediated throughPR and GR specific co-activators and co-repressors. PR and GR asso-ciate preferentially with SRC-1 and SRC-2, respectively, both belong-ing to the main SRC-1/p160 family of co-activators (Onate et al.,1995; Voegel et al., 1996; Li et al., 2003). Conversely, when boundto an antagonist ligand, PR or GR recruited co-repressors, such asthe nuclear receptor co-repressor (N-CoR) or the silencing mediatorfor retinoid and thyroid hormone receptor (SMRT) which induces therepression of signal transactivation (Chen and Evans, 1995; Horleinet al., 1995). Modulation of the cellular responses to PR and GRagonist and antagonist are linked to the amount and accessibility ofco-activators and co-repressors. For example, difference in mifepris-tone agonist/antagonist action observed in T-47D cells comparedwith HeLA cells is modulated by the ratio SRC-1 and SMRT ofthese cells (Liu et al., 2002). In our case, the variability inco-activator/co-repressor expression ratio may explain the attenuatedUPA antagonist responses in HBE cells, when compared with breastcancer cells models (Liu et al., 2002; Smith and O’Malley, 2004).
UPA agonist activity was previously shown in MDA-MB-231 cellsoverexpressing PR-A, PR-B or both PR isoforms. UPA inhibited cell
Figure 5 UPA effect on hormone mediated cell proliferation. Cell proliferation was measured by [3H] thymidine incorporation in (A) HBE cells,(B) T-47D cells and (C) MCF-7 cells. HBE cells were treated 96 h and T-47D and MCF-7 cells were treated 48 h at the concentration of 100 nM forP, DEX, UPA and 10 nM for estradiol (E2). Results are expressed as percentage of control (mean+ SEM, n ¼ 7 for HBE, n ¼ 4 for T-47D and n ¼ 3for MCF-7). *P , 0.05, **P , 0.01, ***P , 0.001.
Ulipristal acetate in breast cells 2793D
ownloaded from
https://academic.oup.com
/humrep/article-abstract/27/9/2785/627029 by guest on 04 April 2019
growth and regulated expression of genes involved in cell cycle regu-lation in the same manner than P through both isoforms (Leo and Lin,2008). In contrast, we did not observe UPA agonist effects on the PRtarget genes in normal or cancerous cells endogenously expressing PR.Absence of agonist activity of UPA may be attributed in one part to alower level of PR in our models, as transfected MDA contained abouttwice as much PR as in T-47D. Additionally, the ratio of co-regulatorscould be in favor to the co-activators in MDA-MB-231 cell line com-pared with HBE and T47D cells. As supporting information, thesteroid receptor co-activator, amplified in breast cancer (AIB1) isstrongly expressed in MDA-MB-231 cells whereas it is moderatelyexpressed in T-47D cells and is absent in normal breast tissue (Hude-list et al., 2003; Li et al., 2008).
Numerous studies had previously highlighted the interest of usingantiprogestins as breast cancer treatment (Klijn et al., 2000). Mifepris-tone and onapristone exhibited growth-inhibitory effects especially onPR-positive human breast cancer cell lines as well as on breast tumorsin animal models (Bardon et al., 1985; Bakker et al., 1987; Schneideret al., 1992; Klijn et al., 1994; Dannecker et al., 1996; Poole et al.,2006). Administration of mifepristone or onapristone is also asecond or third line of breast cancer treatment. The use of mifepris-tone or onapristone in post menopausal women with metastaticbreast cancer (n ¼ 123) was associated with a partial or complete re-sponse in 11% and absence of progression in 43% of the patients(Romieu et al., 1987; Klijn et al., 1989). Unfortunately, clinical
studies were not sustained because of the anti-glucocorticoid andtoxic liver side effects of mifepristone and onapristone, respectively.These onapristone anti-tumoral effects in vivo were observeddespite the ability of onapristone to reverse the P induced anti-proliferative effect in tumoral breast cells (Lin et al., 1999). This obser-vation suggests that UPA anti-tumoral potential should be better char-acterized despite its ability to reverse the P induced anti-proliferativeeffect on T-47D cells. UPA should be more satisfactory in long-termanti-tumor therapy due to its limited anti-glucocorticoid effectswhen compared with those of mifepristone as well as to theabsence of serious adverse effects reported in a clinical study after3–6 months of daily administration (Nieman et al., 2011). To reinforcethis idea, we observed that the invasion markers FASN and ALPLexpressions were inhibited by UPA in presence of P in T-47D cells,suggesting a potential interest of UPA in metastatic breast disease.
One of the most important findings of this study is the UPA anti-proliferative and pro-apoptotic effect in HBE cells when combinedwith either P or DEX. We examined UPA characteristics and antagon-ist potency in a more accurate in vivo hormonal environment, by graft-ing breast tissue into mice. This experimental procedure represents acomplementary approach to the study of human epithelial cells in theirstructural and environmental integrities. In this model, the monitoringof steroid concentrations can be performed, whereas this remainsmore difficult in women because of the variation and the irregularityof the ovarian cycle. We designed hormonal treatments in order to
Figure 6 UPA effects on hormone mediated cell apoptosis. Cell apoptosis was measured by flow cytometry quantification of sub-G1 phase in(A) HBE cells, (B) T-47D cells and (C) MCF-7 cells. HBE cells were treated 96 h and T-47D and MCF-7 cells were treated 48 h at the concentrationof 100 nM for P, DEX, UPA and 10 nM for estradiol (E2). Results are expressed as a percentage of control (mean+ SEM, n ¼ 9 for HBE, n ¼ 3 forT-47D and n ¼ 4 for MCF-7). *P , 0.05, **P , 0.001.
2794 Communal et al.
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article-abstract/27/9/2785/627029 by guest on 04 April 2019
imitate human female hormonal physiology, as it was previously shownthat changes in the levels of hormones impact breast tissue regulation(Clarke et al., 1997). Our results indicate that the proliferative activityin breast tissue, especially on duct structure, seems to be highly de-pendent on E2 signalization, even with E2 levels corresponding tothe early follicular phase. This is in contrast with previous resultsreporting E2 proliferative effects occurring only at much higher E2 con-centrations (341 pg/ml, range: 315.8–368 pg/ml) (Clarke et al.,1997). This discrepancy can be associated with our analysis which dis-criminates between lobules and ducts since our results point to a dif-ference of the steroidal hormone activities between these structures.
This observation was previously made in an ER knockout mice modelwhere duct development was impaired whereas PR knockout miceshowed a lack of lobulo-alveolar structures (Lydon et al., 1995; Boc-chinfuso and Korach, 1997). The luteal phase in women correspondsto the secreted P period in addition to E2 secretions, and is correlatedto the higher proliferative state in the breast tissue (Longacre andBartow, 1986; Soderqvist et al., 1997; Ramakrishnan et al., 2002).However, the effect of P on the proliferation of the luminal breastcells is still a controversy (Graham and Clarke, 1997). Studies whichmonitored the concentrations of P show a preponderant effect ofE2 administration with a limited action of P on breast proliferation
Figure 7 PR, ER and Ki67 expressions in breast tissue xenografts. ER, PR and Ki67 expression were analyzed by immunohistochemistry. (A) Rep-resentative image for ER and PR labeling in original breast tissue before and in treated grafts in mice. (B) Ki67 expression in C, E2, E2 + P,E2 + P + UPA treatment groups of grafted tissues. Magnification ×400. (C) Mitotic index (Ki67 positive cells percentage) control fold inductionin lobules and ducts (mean+ SEM), lobule structures from four patients, duct structures from six patients. *P , 0.05.
Ulipristal acetate in breast cells 2795D
ownloaded from
https://academic.oup.com
/humrep/article-abstract/27/9/2785/627029 by guest on 04 April 2019

during the luteal phase (Clarke et al., 1997; Foidart et al., 1998). In arecent study, E2 + P treatment was administered to post-menopausalwomen and did not increase epithelial cell proliferation whereas con-jugated estrogens associated with the progestin medroxyprogesteroneacetate (MPA) treatment was proliferative (Murkes et al., 2011). In thesame vein, in mammary tissues collected from ovariectomized cyno-molgus macaque female adults, E2 + MPA treatment was mitogenicand activated growth factor signalization. This effect was not seenwith E2 + P treatment (Wood et al., 2007, 2009). Our results corrob-orate those results as we observed that addition of P or UPA, to E2treatment did not markedly alter cell proliferation when comparedwith E2 alone. Nevertheless, our in vivo model offers some limitations,with the continuous deliverance of steroids through a pellet whichwould have a different release kinetic when compared with steroid se-cretion from the human ovary. In addition, to detect small amplitudeof P on cell proliferation, a higher number of experiments should beperformed with breast samples from women at the same period ofthe menstrual cycle.
The impact of SPRMs on normal mammary tissue especially inhuman is difficult to evaluate. Using fine needle aspiration, Engmanet al. (2008) showed that mifepristone was a potent anti-proliferativecompound on normal breast cells. The mechanism underlying theseactions is still not clarified. A reasonable hypothesis is that the mife-pristone anti-proliferative effect could be mediated through its anti-gonadotropic action (Marions et al., 2002; Leminen et al., 2005)rather than by or in addition to a specific anti-progesterone effecton breast cell proliferation. It is also known that mifepristone andMPA interact with GR. The influence of SPRMs on glucocorticoid ac-tivity should therefore be taken into account for the final mitogenic re-sponse as we showed the important impact of glucocorticoids onnormal luminal cells proliferation (Courtin et al., 2012).
The experiment performed in this study used a rate of UPA similarto the steady-state concentration of UPA after clinical chronic admin-istration in women (HRA pharma personal data). This level ensured amaximum potential to inhibit progesterone actions. Our resultssuggest that UPA does not exert growth stimulation on normalbreast tissue and could be used as an antiprogestin molecule targetinghormone-responsive organs, such as uterus, for long-term therapy.Further clinical studies in women are nevertheless necessary toconfirm its non-detrimental effect in normal breast tissue.
AcknowledgementsWe thank Drs Michele Resche-Rigon, Andre Ulmann and Erin Gainer(HRA Pharma, Paris) for helpful discussions, for providing the antipro-gestins UPA and CDB-3377 and for their contribution on mice serumUPA concentrations analysis. We also thank Steve K. Nordeen (Univer-sity of Colorado, Denver) for sequences of IEX-1 primers. We are verygrateful to Aurelie Scriva for its technical assistance in immunohisto-chemistry. We thank Dr Neil Insdorf for his kind help in editing themanuscript, and Dr William Rostene for providing the pellet procedure.
Authors’ rolesA.G.: conception of the study, data interpretation, manuscript writingand final approval of manuscript. L.C.: collection and assembly of data,
data analysis and interpretation and manuscript writing. M.V.: collec-tion of data, data analysis and interpretation and manuscript writing.J.H. and A.C.: collection of data, data analysis and interpretation andreviewing of the manuscript. N.M.: data analysis and interpretationand reviewing of the manuscript. N.L.: performing estradiol and pro-gesterone dosages in the mice samples and reviewing of the manu-script. S.D.: collection and assembly of data. M.C.: access andcollection of materials. P.F.: data interpretation, reviewing andediting of the manuscript.
FundingThis research was supported by grants from INSERM-UPMC, HRAPharma and the Institut National du Cancer (INCa, France). LaudineCommunal was the recipient of a CIFRE grant from HRA Pharmaand the government. Myriam Vilasco was the recipient of a post-docfellowship (INCa). Aurelie Courtin was a recipient of a grant fromthe Association pour la Recherche sur le Cancer (ARC).
Conflict of interestL.C. was the recipient of a CIFRE grant from HRA Pharma and thegovernment part of the study was supported by a grant from HRA-Pharma. None were declared by the other authors.
ReferencesAnderson E. Ovarian steroids and control of proliferation in the normal
human breast. Breast 2001;10:273–278.Attardi BJ, Burgenson J, Hild SA, Reel JR, Blye RP. CDB-4124 and its
putative monodemethylated metabolite, CDB-4453, are potentantiprogestins with reduced antiglucocorticoid activity: in vitrocomparison to mifepristone and CDB-2914. Mol Cell Endocrinol 2002;188:111–123.
Attardi BJ, Burgenson J, Hild SA, Reel JR. In vitro antiprogestational/antiglucocorticoid activity and progestin and glucocorticoid receptorbinding of the putative metabolites and synthetic derivatives ofCDB-2914, CDB-4124, and mifepristone. J Steroid Biochem Mol Biol2004;88:277–288.
Bakker GH, Setyono-Han B, Henkelman MS, de Jong FH, Lamberts SW,van der Schoot P, Klijn JG. Comparison of the actions of theantiprogestin mifepristone (RU486), the progestin megestrol acetate,the LHRH analog buserelin, and ovariectomy in treatment of ratmammary tumors. Cancer Treatment Reports 1987;71:1021–1027.
Bardon S, Vignon F, Chalbos D, Rochefort H. RU486, a progestin andglucocorticoid antagonist, inhibits the growth of breast cancer cells viathe progesterone receptor. J Clin Endocrinol Metab 1985;60:692–697.
Blithe DL, Nieman LK, Blye RP, Stratton P, Passaro M. Development of theselective progesterone receptor modulator CDB-2914 for clinicalindications. Steroids 2003;68:1013–1017.
Bocchinfuso WP, Korach KS. Mammary gland development andtumorigenesis in estrogen receptor knockout mice. J Mammary GlandBiol Neoplasia 1997;2:323–334.
Brenner RM, Slayden OD, Nath A, Tsong YY, Sitruk-Ware R. Intrauterineadministration of CDB-2914 (Ulipristal) suppresses the endometrium ofrhesus macaques. Contraception 2010;81:336–342.
Cadepond F, Ulmann A, Baulieu EE. RU486 (mifepristone): mechanisms ofaction and clinical uses. Annu Rev Med 1997;48:129–156.
2796 Communal et al.
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article-abstract/27/9/2785/627029 by guest on 04 April 2019

Chabbert-Buffet N, Meduri G, Bouchard P, Spitz IM. Selectiveprogesterone receptor modulators and progesterone antagonists:mechanisms of action and clinical applications. Hum Reprod Update2005;11:293–307.
Chabbert-Buffet N, Pintiaux-Kairis A, Bouchard P. Effects of theprogesterone receptor modulator VA2914 in a continuous low doseon the hypothalamic-pituitary-ovarian axis and endometrium innormal women: a prospective, randomized, placebo-controlled trial.J Clin Endocrinol Metab 2007;92:3582–3589.
Chalbos D, Joyeux C, Galtier F, Rochefort H. Progestin-induced fatty acidsynthetase in human mammary tumors: from molecular to clinicalstudies. J Steroid Biochem Mol Biol 1992;43:223–228.
Chen JD, Evans RM. A transcriptional co-repressor that interacts withnuclear hormone receptors. Nature 1995;377:454–457.
Clarke RB, Howell A, Anderson E. Estrogen sensitivity of normal humanbreast tissue in vivo and implanted into athymic nude mice: analysis of therelationship between estrogen-induced proliferation and progesteronereceptor expression. Breast Cancer Res Treat 1997;45:121–133.
Cook CE, Lee YW, Wani MC, Fail PA, Petrow V. Effects of D-ring sub-stituents on antiprogestational (antagonist) and progestational (agonist)activity of 11 beta-aryl steroids. Hum Reprod 1994;9 Suppl 1:32–39.
Courtin A, Communal L, Vilasco M, Cimino D, Mourra N, de Bortoli M,Taverna D, Faussat AM, Chaouat M, Forgez P et al. Glucocorticoidreceptor activity discriminates between progesterone andmedroxyprogesterone acetate effects in breast cells. Breast Cancer ResTreat 2012;131:49–63.
Dannecker C, Possinger K, Classen S. Induction of TGF-beta by anantiprogestin in the human breast cancer cell line T-47D. Ann Oncol1996;7:391–395.
Di Lorenzo D, Gianni M, Savoldi GF, Ferrari F, Albertini A, Garattini E.Progesterone induced expression of alkaline phosphatase is associatedwith a secretory phenotype in T47D breast cancer cells. BiochemBiophys Res Commun 1993;192:1066–1072.
Eisinger SH, Bonfiglio T, Fiscella K, Meldrum S, Guzick DS. Twelve-monthsafety and efficacy of low-dose mifepristone for uterine myomas. J MinimInvasive Gynecol 2005;12:227–233.
Engman M, Skoog L, Soderqvist G, Gemzell-Danielsson K. The effect ofmifepristone on breast cell proliferation in premenopausal womenevaluated through fine needle aspiration cytology. Hum Reprod 2008;23:2072–2079.
Esteve JL, Acosta R, Perez Y, Campos R, Hernandez AV, Texido CS.Treatment of uterine myoma with 5 or 10 mg mifepristone daily during 6months, post-treatment evolution over 12 months: double-blind ran-domised clinical trial. Eur J Obstet Gynecol Reprod Biol 2012;161:202–208..
Foidart JM, Colin C, Denoo X, Desreux J, Beliard A, Fournier S, deLignieres B. Estradiol and progesterone regulate the proliferation ofhuman breast epithelial cells. Fertil Steril 1998;69:963–969.
Gabet Y, Noh T, Lee C, Frenkel B. Developmentally regulated inhibition ofcell cycle progression by glucocorticoids through repression of cyclin Atranscription in primary osteoblast cultures. J Cell Physiol 2011;226:991–998.
Gainer EE, Ulmann A. Pharmacologic properties of CDB(VA)-2914.Steroids 2003;68:1005–1011.
Gompel A, Malet C, Spritzer P, Lalardrie JP, Kuttenn F, Mauvais-Jarvis P.Progestin effect on cell proliferation and 17 beta-hydroxysteroiddehydrogenase activity in normal human breast cells in culture. J ClinEndocrinol Metab 1986;63:1174–1180.
Gompel A, Somai S, Chaouat M, Kazem A, Kloosterboer HJ, Beusman I,Forgez P, Mimoun M, Rostene W. Hormonal regulation of apoptosis inbreast cells and tissues. Steroids 2000;65:593–598.
Goyeneche AA, Seidel EE, Telleria CM. Growth inhibition induced byantiprogestins RU-38486, ORG-31710, and CDB-2914 in ovarian cancer
cells involves inhibition of cyclin dependent kinase 2. Invest New Drugs2012;30:967–980.
Graham JD, Clarke CL. Physiological action of progesterone in targettissues. Endocr Rev 1997;18:502–519.
Hild SA, Reel JR, Hoffman LH, Blye RP. CDB-2914: anti-progestational/anti-glucocorticoid profile and post-coital anti-fertility activity in ratsand rabbits. Hum Reprod 2000;15:822–829.
Horlein AJ, Naar AM, Heinzel T, Torchia J, Gloss B, Kurokawa R, Ryan A,Kamei Y, Soderstrom M, Glass CK et al. Ligand-independent repressionby the thyroid hormone receptor mediated by a nuclear receptorco-repressor. Nature 1995;377:397–404.
Huang Z, Fasco MJ, Figge HL, Keyomarsi K, Kaminsky LS. Expression ofcytochromes P450 in human breast tissue and tumors. Drug MetabDispos 1996;24:899–905.
Hudelist G, Czerwenka K, Kubista E, Marton E, Pischinger K, Singer CF.Expression of sex steroid receptors and their co-factors in normaland malignant breast tissue: AIB1 is a carcinoma-specific co-activator.Breast Cancer Res Treat 2003;78:193–204.
Jang GR, Wrighton SA, Benet LZ. Identification of CYP3A4 as the principalenzyme catalyzing mifepristone (RU 486) oxidation in human livermicrosomes. Biochem Pharmacol 1996;52:753–761.
Joyeux C, Chalbos D, Rochefort H. Effects of progestins and menstrualcycle on fatty acid synthetase and progesterone receptor in humanmammary glands. J Clin Endocrinol Metab 1990;70:1438–1444.
Kandouz M, Siromachkova M, Jacob D, Chretien Marquet B, Therwath A,Gompel A. Antagonism between estradiol and progestin on Bcl-2expression in breast-cancer cells. Int J Cancer 1996;68:120–125.
Kehrl JH, Sinnarajah S. RGS2: a multifunctional regulator of G-proteinsignaling. Int J Biochem Cell Biol 2002;34:432–438.
Keshava C, McCanlies EC, Weston A. CYP3A4 polymorphisms–potentialrisk factors for breast and prostate cancer: a HuGE review. Am JEpidemiol 2004;160:825–841.
Klijn JG, de Jong FH, Bakker GH, Lamberts SW, Rodenburg CJ,Alexieva-Figusch J. Antiprogestins, a new form of endocrine therapyfor human breast cancer. Cancer Res 1989;49:2851–2856.
Klijn JG, Setyono-Han B, Sander HJ, Lamberts SW, de Jong FH,Deckers GH, Foekens JA. Pre-clinical and clinical treatment ofbreast cancer with antiprogestins. Hum Reprod 1994;9 Suppl1:181–189.
Klijn JG, Setyono-Han B, Foekens JA. Progesterone antagonists andprogesterone receptor modulators in the treatment of breast cancer.Steroids 2000;65:825–830.
Kuhajda FP. Fatty acid synthase and cancer: new application of an oldpathway. Cancer Res 2006;66:5977–5980.
Leminen R, Raivio T, Ranta S, Oehler J, von Hertzen H, Janne OA,Heikinheimo O. Late follicular phase administration of mifepristonesuppresses circulating leptin and FSH - mechanism(s) of action inemergency contraception? Eur J Endocrinol 2005;152:411–418.
Leo JC, Lin VC. The activities of progesterone receptor isoform A and Bare differentially modulated by their ligands in a gene-selective manner.Int J Cancer 2008;122:230–243.
Levens ED, Potlog-Nahari C, Armstrong AY, Wesley R, Premkumar A,Blithe DL, Blocker W, Nieman LK. CDB-2914 for uterineleiomyomata treatment: a randomized controlled trial. Obstet Gynecol2008;111:1129–1136.
Li X, Wong J, Tsai SY, Tsai MJ, O’Malley BW. Progesterone andglucocorticoid receptors recruit distinct coactivator complexes andpromote distinct patterns of local chromatin modification. Mol CellBiol 2003;23:3763–3773.
Li LB, Louie MC, Chen HW, Zou JX. Proto-oncogene ACTR/AIB1promotes cancer cell invasion by up-regulating specific matrixmetalloproteinase expression. Cancer Lett 2008;261:64–73.
Ulipristal acetate in breast cells 2797D
ownloaded from
https://academic.oup.com
/humrep/article-abstract/27/9/2785/627029 by guest on 04 April 2019

Lin VC, Ng EH, Aw SE, Tan MG, Chan VS, Ho GH. Progestins inhibit thegrowth of MDA-MB-231 cells transfected with progesterone receptorcomplementary DNA. Clin Cancer Res 1999;5:395–403.
Lin VC, Woon CT, Aw SE, Guo C. Distinct molecular pathways mediateprogesterone-induced growth inhibition and focal adhesion.Endocrinology 2003;144:5650–5657.
Liu Z, Auboeuf D, Wong J, Chen JD, Tsai SY, Tsai MJ, O’Malley BW.Coactivator/corepressor ratios modulate PR-mediated transcriptionby the selective receptor modulator RU486. Proc Natl Acad Sci USA2002;99:7940–7944.
Longacre TA, Bartow SA. A correlative morphologic study of humanbreast and endometrium in the menstrual cycle. Am J Surg Pathol1986;10:382–393.
Lydon JP, DeMayo FJ, Funk CR, Mani SK, Hughes AR, Montgomery CA Jr,Shyamala G, Conneely OM, O’Malley BW. Mice lacking progesteronereceptor exhibit pleiotropic reproductive abnormalities. Genes Dev1995;9:2266–2278.
Malet C, Gompel A, Yaneva H, Cren H, Fidji N, Mowszowicz I, Kuttenn F,Mauvais-Jarvis P. Estradiol and progesterone receptors in cultured normalhuman breast epithelial cells and fibroblasts: immunocytochemical studies.J Clin Endocrinol Metab 1991;73:8–17.
Marions L, Hultenby K, Lindell I, Sun X, Stabi B, Gemzell Danielsson K.Emergency contraception with mifepristone and levonorgestrel:mechanism of action. Obstet Gynecol 2002;100:65–71.
Maruo T, Ohara N, Yoshida S, Nakabayashi K, Sasaki H, Xu Q, Chen W,Yamada H. Translational research with progesterone receptor modulatormotivated by the use of levonorgestrel-releasing intrauterine system.Contraception 2010;82:435–441.
Murkes D, Conner P, Leifland K, Tani E, Beliard A, Lundstrom E,Soderqvist G. Effects of percutaneous estradiol-oral progesteroneversus oral conjugated equine estrogens-medroxyprogesteroneacetate on breast cell proliferation and bcl-2 protein in healthywomen. Fertil Steril 2011;95:1188–1191.
Nieman LK, Blocker W, Nansel T, Mahoney S, Reynolds J, Blithe D, Wesley R,Armstrong A. Efficacy and tolerability of CDB-2914 treatment forsymptomatic uterine fibroids: a randomized, double-blind, placebo-controlled, phase IIb study. Fertil Steril 2011;95:767–772 e761–762.
Onate SA, Tsai SY, Tsai MJ, O’Malley BW. Sequence and characterizationof a coactivator for the steroid hormone receptor superfamily. Science1995;270:1354–1357.
Poole AJ, Li Y, Kim Y, Lin SC, Lee WH, Lee EY. Prevention ofBrca1-mediated mammary tumorigenesis in mice by a progesteroneantagonist. Science 2006;314:1467–1470.
Ramakrishnan R, Khan SA, Badve S. Morphological changes in breast tissuewith menstrual cycle. Mod Pathol 2002;15:1348–1356.
Reel JR, Hild-Petito S, Blye RP. Antiovulatory and postcoital antifertilityactivity of the antiprogestin CDB-2914 when administered as single,multiple, or continuous doses to rats. Contraception 1998;58:129–136.
Ritzke C, Stieber P, Untch M, Nagel D, Eiermann W, Fateh-Moghadam A.Alkaline phosphatase isoenzymes in detection and follow up of breastcancer metastases. Anticancer Res 1998;18:1243–1249.
Romieu G, Maudelonde T, Ulmann A, Pujol H, Grenier J, Cavalie G,Khalaf S, Rochefort H. The antiprogestin RU486 in advanced breastcancer: preliminary clinical trial. Bull Cancer 1987;74:455–461.
Schneider MR, Michna H, Habenicht UF, Nishino Y, Grill HJ, Pollow K. Thetumour-inhibiting potential of the progesterone antagonist Onapristonein the human mammary carcinoma T61 in nude mice. J Cancer Res ClinOncol 1992;118:187–189.
Sengupta S, Vonesch JL, Waltzinger C, Zheng H, Wasylyk B. Negativecross-talk between p53 and the glucocorticoid receptor and its role inneuroblastoma cells. EMBO J 2000;19:6051–6064.
Shen L, Guo J, Santos-Berrios C, Wu MX. Distinct domains for anti- andpro-apoptotic activities of IEX-1. J Biol Chem 2006;281:15304–15311.
Smith CL, O’Malley BW. Coregulator function: a key to understanding tissuespecificity of selective receptor modulators. Endocr Rev 2004;25:45–71.
Soderqvist G, Isaksson E, von Schoultz B, Carlstrom K, Tani E, Skoog L.Proliferation of breast epithelial cells in healthy women during themenstrual cycle. Am J Obstet Gynecol 1997;176:123–128.
Spitz IM, Croxatto HB, Robbins A. Antiprogestins: mechanism of actionand contraceptive potential. Annu Rev Pharmacol Toxicol 1996;36:47–81.
Voegel JJ, Heine MJ, Zechel C, Chambon P, Gronemeyer H. TIF2, a160 kDa transcriptional mediator for the ligand-dependent activationfunction AF-2 of nuclear receptors. EMBO J 1996;15:3667–3675.
Wagner BL, Pollio G, Leonhardt S, Wani MC, Lee DY, Imhof MO,Edwards DP, Cook CE, McDonnell DP. 16 alpha-substituted analogsof the antiprogestin RU486 induce a unique conformation in thehuman progesterone receptor resulting in mixed agonist activity. ProcNatl Acad Sci USA 1996;93:8739–8744.
Wagner BL, Norris JD, Knotts TA, Weigel NL, McDonnell DP. Thenuclear corepressors NCoR and SMRT are key regulators of bothligand- and 8-bromo-cyclic AMP-dependent transcriptional activity ofthe human progesterone receptor. Mol Cell Biol 1998;18:1369–1378.
Wagner BL, Pollio G, Giangrande P, Webster JC, Breslin M, Mais DE,Cook CE, Vedeckis WV, Cidlowski JA, McDonnell DP. The novelprogesterone receptor antagonists RTI 3021–012 and RTI 3021–022exhibit complex glucocorticoid receptor antagonist activities:implications for the development of dissociated antiprogestins.Endocrinology 1999;140:1449–1458.
Wan Y, Nordeen SK. Overlapping but distinct gene regulation profiles byglucocorticoids and progestins in human breast cancer cells. MolEndocrinol 2002;16:1204–1214.
Wood CE, Register TC, Lees CJ, Chen H, Kimrey S, Cline JM. Effects ofestradiol with micronized progesterone or medroxyprogesteroneacetate on risk markers for breast cancer in postmenopausalmonkeys. Breast Cancer Res Treat 2007;101:125–134.
Wood CE, Register TC, Cline JM. Transcriptional profiles of progestogeneffects in the postmenopausal breast. Breast Cancer Res Treat 2009;114:233–242.
Wu MX. Roles of the stress-induced gene IEX-1 in regulation of cell deathand oncogenesis. Apoptosis 2003;8:11–18.
Xu Q, Takekida S, Ohara N, Chen W, Sitruk-Ware R, Johansson ED,Maruo T. Progesterone receptor modulator CDB-2914down-regulates proliferative cell nuclear antigen and Bcl-2 proteinexpression and up-regulates caspase-3 and poly(adenosine5′-diphosphate-ribose) polymerase expression in cultured humanuterine leiomyoma cells. J Clin Endocrinol Metab 2005;90:953–961.
Xu Q, Ohara N, Chen W, Liu J, Sasaki H, Morikawa A, Sitruk-Ware R,Johansson ED, Maruo T. Progesterone receptor modulator CDB-2914down-regulates vascular endothelial growth factor, adrenomedullinand their receptors and modulates progesterone receptor content incultured human uterine leiomyoma cells. Hum Reprod 2006;21:2408–2416.
Yoshida S, Ohara N, Xu Q, Chen W, Wang J, Nakabayashi K, Sasaki H,Morikawa A, Maruo T. Cell-type specific actions of progesteronereceptor modulators in the regulation of uterine leiomyoma growth.Semin Reprod Med 2010;28:260–273.
2798 Communal et al.
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article-abstract/27/9/2785/627029 by guest on 04 April 2019





























