Embed Size (px)
Citation preview

Underwater walking
Joseph Ayers*
Department of Biology, Marine Science Center, Northeastern University, East Point, Nahant, MA 01908, USA
Received 8 March 2004; accepted 25 May 2004
Abstract
Lobsters are generalist decapods that evolved in a broad variety of niches in the Northwestern Atlantic. Due to their inherent buoyancy
they have acquired adaptations to reduced traction and surge. We have developed a biomimetic robot based on the lobster that features
artificial muscle actuators and sensors employing labeled-line codes. The central controller for this robot is based on the command neuron,
coordinating neuron central pattern generator model. A library of commands is released by sensor feedback to mediate adaptive sequences
and goal achieving behavior. Rheotaxic behaviors can mediate adaptations to achieve some of the advantages of the biological models.
q 2004 Elsevier Ltd. All rights reserved.
Keywords: Lobster; Robot; Biomimetic; Sensors; Artificial muscle; Locomotion
1. Introduction
The littoral region of the oceans presents a challenging
environment for any organism. Fundamental problems exist
in the shallow inshore. Heavy turbulence caused by surge,
wave action, tides and currents can cause severe stability
problems. These problems have been overcome by bottom
dwelling organisms such as crabs and lobsters that live with
impunity in these complex conditions (Sandeman and
Atwood, 1982; Cobb and Phillips, 1980). Lobsters have
navigated over the ocean bottom formillions of years and have
evolved robust systems for control of locomotion, sensing,
searching, and adaptation to surge (Factor, 1995). Lobsters
oftenmigrate into littoral regionswhere thebidirectional surge
due to wave action and persistent currents due to tidal
movements in channels require ongoing compensation. The
necessity of dealing with surge and currents and the inherent
traction problems are the primary factor differentiating
underwater walking from terrestrial walking. The sprawled
posture of thewalking legs helps prevent overturning in strong
lateral currents (Alexander, 1971). The ambulatory locomo-
tory movements, combined with hydrodynamic adaptability
of decapods are a proven solution to the stability problem
posed by both littoral and stream environments (Maude and
Williams, 1983; Martinez, 2001).
Decapods have evolved other mechanisms to address
surge. Lobsters and crayfish use their claws, abdomen and
swimmerets as hydrodynamic control surfaces and thrusters
(Fig. 1) affording considerable adaptation relative to
hydrodynamic perturbation during locomotion (Davis,
1968; Maude and Williams, 1983). A hydrodynamically
adaptable shape provides negative lift (Martinez, 2001), and
can hold a decapod to the bottom rather than allowing
current and surge to displace it. Maude and Williams (1983)
found that crayfish species with lower drag coefficients were
able to inhabit streams with higher flow velocities.
Although lobsters and crayfish are generally in mechan-
ical contact with the sea floor, their traction is reduced
relative to terrestrial organisms due to their buoyancy.
Hydrodynamic translational forces can often exceed
gravitational forces (Martinez, 2001). Having eight legs in
contact with the substrate therefore gives them more
traction, as does a wedge-like posture that generates a
hydrodynamic thrust vector into the substrate that further
enhances traction (Fig. 1). The additional legs also provide
for more mechanical stability in the pitch plane relative to
the alternating tripod gait of insects.
The mechanisms underlying behavior in decapods have
been formalized into a general model, the command neuron,
coordinating neuron, central pattern generator (CPG) mode
(Kennedy and Davis, 1977; Stein, 1978; Evoy and Ayers,
1982). The central component consists of segmental CPGs
that control the motor neurons and muscles of each limb
1467-8039/$ - see front matter q 2004 Elsevier Ltd. All rights reserved.
doi:10.1016/j.asd.2004.06.001
Arthropod Structure & Development 33 (2004) 347–360
www.elsevier.com/locate/asd
* Tel.: þ1-617-581-7370x309; fax: þ1-617-581-6076.
E-mail address: [email protected] (J. Ayers).

(Sillar and Skorupski, 1986), coordinating systems that
determine the phase relations or gaits between the CPGs of
different limbs and command systems that specify and
modulate the behavior generated by the CPGs. The
command systems represent the control locus at which the
decision to generate a particular behavior is achieved
(Edwards et al., 1999; Larimer, 2000). The peripheral
component consists of exteroceptive and proprioceptive
sensors (Atema and Voigt, 1995; Cattaert and Le Ray, 2001)
that generate adaptive reflexes at the command level
(exteroceptive or orientational reflexes), the CPG level
(phase modulating reflexes) and/or the motor neuron
level (amplitude modulating reflexes).
Arthropod systems have been subjected to considerable
reverse engineering over the past 25 years, adequate to
permit robust synthetic models of their underlying organ-
ization (Beer, 1991; Beer et al., 1998; Ayers et al., 2002). In
several cases the actual synaptic networks have been
established by electrophysiological stimulation and record-
ing (Burrows, 1996; Chrachri and Clarac, 1989; Cattaert and
Le Ray, 2001; Selverston, 1999). These biological models
can be readily adapted to robotic control (Beer et al., 1998;
Ayers et al., 2002). We submit that biologically based
reverse engineering is the most effective procedure both to
design autonomous underwater robots as well as to establish
detailed higher order control schemes for remote sensing
procedures. The control schemes by which a lobster
searches for and acquires prey provide excellent solutions
to the problem of how an underwater robot can successfully
search for and investigate underwater objects (Grasso,
2000).
2. Overview of the lobster walking system
The control of the decapod walking system has been
investigated at almost all levels, from the behavioral to the
neuromodulatory (Factor, 1995). Combined with compara-
tive data from other oscillatory systems in the crustacean
CNS, it is possible to construct reasonably complete control
structures based entirely on known components (Evoy and
Ayers, 1982; Ayers and Crisman, 1992; Ayers et al., 2002;
Cattaert and Le Ray, 2001; Beltz, 1995).
2.1. Limb movements
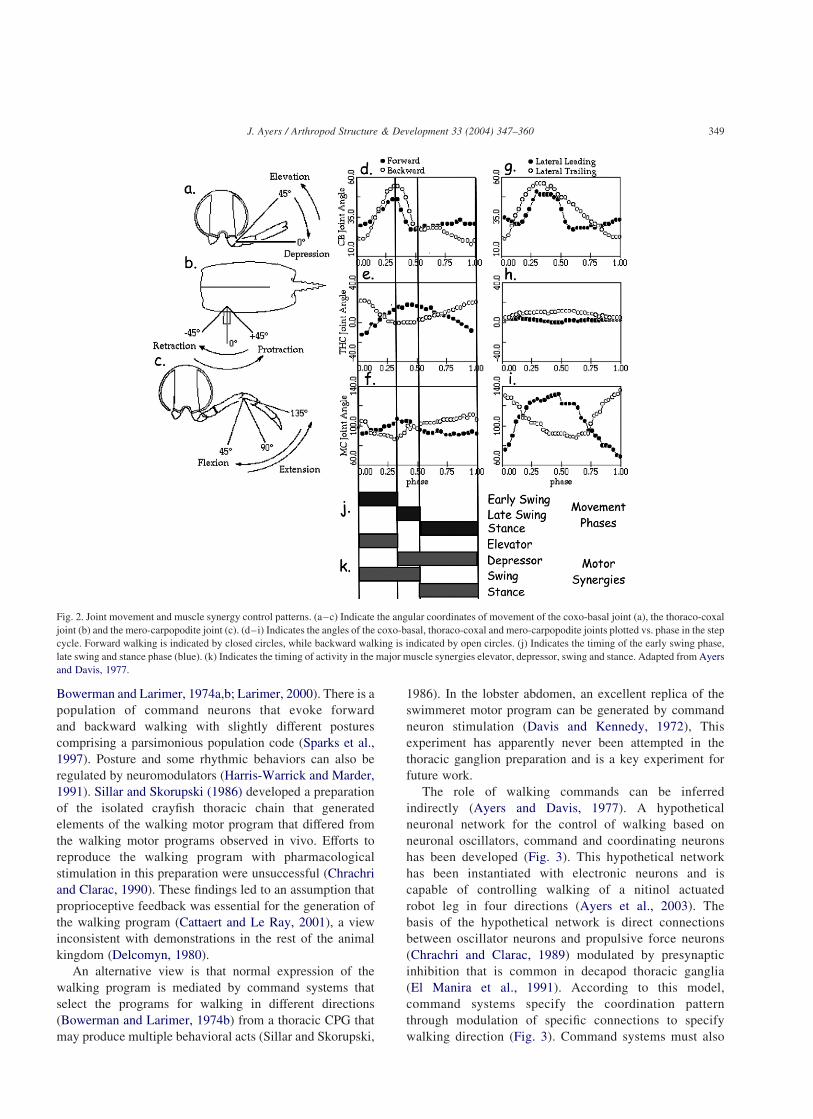
Lobster walking movements are performed around three
major limb joints (Fig. 2): the thoraco-coxal joint mediates
protraction and retraction movements (Fig. 2a), the coxo-
basal joint mediates elevation and depression movements
(Fig. 2b) while the mero-carpopodite joint mediates
extension and flexion movements (Fig. 2c; Ayers and
Davis, 1977; Ayers and Clarac, 1978). The movements of
the coxo-basal and mero-carpopodite joints occur in the
same plane while movements of the thoraco-coxal joint
occur in a plane at right angles to the others. Thus there is a
clear demarcation between opposing movements of the
basilar leg joints.
The lobster step cycle consists of three phases: an early
swing phase lifts the limb tip toward the initial position of
the stance, an early stance phase drops the leg to initiate the
stance and during the late stance phase the limb applies
propulsive force and compensates for gravity. During
walking, the swing phase is always constant in duration
and variation in the step period is mediated by variation in
the duration of the stance phase (Ayers and Davis, 1977).
Cyclic elevation and depression movements of the coxo-
basal joints underlay the swing and stance phase movements
respectively for walking in all four directions (Fig. 2d and
g). Movements of the thoraco-coxal and mero-carpopodite
joints generate propulsive forces (Fig. 2e and i). During
forward and backward walking propulsive forces are
generated by torques about the thoraco-coxal joint. During
lateral walking propulsive forces are generated by the
torques about the mero-carpopodite joint. Propulsive forces
that are synergistic with depression for walking in one
direction (retraction for forward walking, protraction for
backward walking) become antagonistic for walking in the
opposite direction (Fig. 2e and i).
2.2. Neuronal circuits
Many acts of the lobster and crayfish behavioral
repertoire can be evoked in situ by stimulation of single
descending command neurons (Davis and Kennedy, 1972;
Fig. 1. Macruran compensatory postures to low (a), high speed (b) and
backward (c) water currents (from Maude and Williams, 1983).
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360348
Bowerman and Larimer, 1974a,b; Larimer, 2000). There is a
population of command neurons that evoke forward
and backward walking with slightly different postures
comprising a parsimonious population code (Sparks et al.,
1997). Posture and some rhythmic behaviors can also be
regulated by neuromodulators (Harris-Warrick and Marder,
1991). Sillar and Skorupski (1986) developed a preparation
of the isolated crayfish thoracic chain that generated
elements of the walking motor program that differed from
the walking motor programs observed in vivo. Efforts to
reproduce the walking program with pharmacological
stimulation in this preparation were unsuccessful (Chrachri
and Clarac, 1990). These findings led to an assumption that
proprioceptive feedback was essential for the generation of
the walking program (Cattaert and Le Ray, 2001), a view
inconsistent with demonstrations in the rest of the animal
kingdom (Delcomyn, 1980).
An alternative view is that normal expression of the
walking program is mediated by command systems that
select the programs for walking in different directions
(Bowerman and Larimer, 1974b) from a thoracic CPG that
may produce multiple behavioral acts (Sillar and Skorupski,
1986). In the lobster abdomen, an excellent replica of the
swimmeret motor program can be generated by command
neuron stimulation (Davis and Kennedy, 1972), This
experiment has apparently never been attempted in the
thoracic ganglion preparation and is a key experiment for
future work.
The role of walking commands can be inferred
indirectly (Ayers and Davis, 1977). A hypothetical
neuronal network for the control of walking based on
neuronal oscillators, command and coordinating neurons
has been developed (Fig. 3). This hypothetical network
has been instantiated with electronic neurons and is
capable of controlling walking of a nitinol actuated
robot leg in four directions (Ayers et al., 2003). The
basis of the hypothetical network is direct connections
between oscillator neurons and propulsive force neurons
(Chrachri and Clarac, 1989) modulated by presynaptic
inhibition that is common in decapod thoracic ganglia
(El Manira et al., 1991). According to this model,
command systems specify the coordination pattern
through modulation of specific connections to specify
walking direction (Fig. 3). Command systems must also
Fig. 2. Joint movement and muscle synergy control patterns. (a–c) Indicate the angular coordinates of movement of the coxo-basal joint (a), the thoraco-coxal
joint (b) and the mero-carpopodite joint (c). (d–i) Indicates the angles of the coxo-basal, thoraco-coxal and mero-carpopodite joints plotted vs. phase in the step
cycle. Forward walking is indicated by closed circles, while backward walking is indicated by open circles. (j) Indicates the timing of the early swing phase,
late swing and stance phase (blue). (k) Indicates the timing of activity in the major muscle synergies elevator, depressor, swing and stance. Adapted from Ayers
and Davis, 1977.
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360 349

specify the amplitude of propulsive movements through
recruitment from the motor unit pool (Davis and
Kennedy, 1972). This connectivity is capable of
explaining both the rapid changes in walking direction
seen in behaving specimens as well as diagonal walking
where both the thoraco-coxal and mero-carpopodite
joints participate in the generation of propulsive forces
(Ayers et al., 2002).
2.3. A biomimetic lobster robot
We have developed a biomimetic lobster-based robot
using the following process.
(1) Use of a biomimetic plant. We use the general body
form of the lobster. Our lobster robot has its legs
attached to a central hull that also supports claw and
tail-like hydrodynamic control surfaces.
(2) Use of a neuronal circuit-based controller. The brains
of our robot consist of a set of finite state machines
based on the command neuron, coordinating neuron,
CPG model of the organization of motor systems
(Stein, 1978).
(3) Use of myomorphic actuators. We employ artificial
muscle based linear actuators that can recruit force in
the same manner as lobster neuromuscular systems
(Atwood, 1976).
(4) Use of neuromorphic sensors. We employ sensors that
code environmental information in the same fashion as
the sensors of the model organism. They use a labeled
line code that quantizes the timing, amplitude and
orientation of the input stimulus (Bullock, 1978).
(5) Use of a behavioral library based on reverse
engineered sequences of behavior of the target
organisms (Ayers et al., 2002). Much of the behavior
of lobsters consists of sequences of stereotyped acts
that are triggered by environmental input. We are
developing an extensible library of command state
sequences that underlie different acts and are associ-
ated with sensor-based releasers.
The physical plant of this robot is composed of watertight
electronics and battery bays on which 8 three-degree of
freedom nitinol-actuated walking legs are mounted (Fig. 4).
The robot has anterior and posterior hydrodynamic control
surfaces to mimic the chelae and abdomen of the model
organism. Motor drives move these control surfaces as well
as a pair of flexible antennae that use strain gauges to
monitor their bending in response to contact and flow. A
sonar transponder allows supervisory control.
2.4. Neural circuit-based controller
Most previous biomimetic robots use behavioral
based controllers where finite state machines focus on
the implementation of different behavioral acts (Brooks,
1989). Other investigators have used genetic algorithms.
In contrast, we implemented the command neuron,
coordinating neuron central pattern generator (CCCPG)
model for omnidirectional walking as a finite state
machine (Ayers et al., 2002). Since the controller
communicates with the interface boards using a serial
bus the robot can be operated from an external lap top
computer or from an embedded controller (Persistor,
Fig. 3. Hypothetical neuronal network for the control of omnidirectional stepping (Adapted from Ayers and Davis, 1977; Ayers et al., 2002, 2003). Circles
indicate presynaptic or conventional inhibitory connections. Triangles indicate excitatory connections. The orange rectangle encloses an interneuron network
that is modulated to generate the rhythm.
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360350

Motorola 68020). The program treats the CCCPG model
components as objects which pass messages with regard
to status changes. Changes in walking direction, speed,
load etc., are effected by keystrokes on the laptop,
sensor input or sonar supervisory commands to the
embedded processor. The program generates strip chart
displays that are the basis of many of the figures of
experimental results in this manuscript.
At the single limb control level, the ambulation
controller relies on three major classes of components that
control the elevator, depressor, protractor, retractor, exten-
sor and flexor synergies (Fig. 5). The oscillator component
is a software clock that regulates the period of stepping as
well as the duration of the swing or elevator phase fraction
of the stepping cycle based on the dynamics of an
endogenous pacemaker neuron (Ayers and Selverston,
1979). The oscillator is embodied as the elevator synergy
and is the site of action of coordinating input from other
oscillators and phase modulating feedback.
The second major component of the ambulation
controller is the pattern generator. It determines the
pattern of discharge of bifunctional synergies based on
command input. The pattern generator determines
through a truth table what synergies should be active
and sets or clears the Boolean variables associated with
the on or off state of the different bifunctional
synergies. The truth table implements the presynaptic
inhibitory logic of the neuronal circuit model (Fig. 3)
and specifies the excitatory connections that would be
disabled by the directional command.
The third major component of the ambulation controller
is the recruiter that determines which of the elements of the
propulsive force synergies are active (Fig. 6a). The recruiter
responds to the desired period parameter and recruits motor
Fig. 4. The lobster-based robot.
Fig. 5. Organization of finite state machine controller. A neuronal oscillator
organizes the four-phase pattern. The neuronal oscillator receives phase
modulating inputs from coordinating neurons and proprioceptors as well as
parametric modulation from command neurons. The pattern generator
selects patterns for sub-behaviors in response to command logic. The
compensator receives inputs from pitch and roll sensors to modulate
posture. The propulsive recruiter responds to parametric modulation and
load sensors to control the synergies producing propulsive force. The
compensator end propulsive recruiter activate motor synergies controlling
different leg joints.
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360 351

units of increasing size (Davis, 1971). Fig. 6b and c is a
graphical realization of the output of the ambulation
controller demonstrating adaptation to speed during forward
and backward walking. In propulsive force synergies the
width of the trace indicates the degree of recruitment within
the pool. Notice that during increase in speed the period of
stepping decreases while propulsive force synergies are
selectively recruited.
The outputs of the finite state machine are control signals
that specify the timing and amplitude of actuator action.
These signals are used to gate power transistors at different
duty cycles to activate contractions of the artificial muscle
just as motor neuron action potentials activate muscle. Fig.
6d–g indicates the signals controlling the thoraco-coxal,
coxo-basal and mero-carpopodite joints during walking in
all four directions. Note that movement synergies that are
synergistic for walking in one direction become antagonistic
for walking in the opposite direction. Antagonist muscles of
joints that serve a postural function in a particular walking
direction are co-activated at low amplitude (Ayers and
Clarac, 1978).
2.5. Coordinating systems
Coordinating systems operate by providing timing
information from a governing oscillator to a governed
oscillator. The governed oscillator responds by resetting it’s
timing to match the period of the governing oscillator
(Stein, 1976; Ayers and Selverston, 1979; Pinsker and
Ayers, 1983). The emergent behavior of the known
intersegmental coordinating interneurons is the alternating
tetrapod gait (Wilson, 1967). In the walking system, there
are coordinating systems between contralateral pattern
generators of the same segment as well as coordinating
systems between adjacent segments. Although the network
basis is not established, contralateral coordination appears
Fig. 6. Motor programs for omnidirectional walking generated by the finite state machine. (a) Recruitment circuit that mediates size-ordered recruitment of
actuators. (b and c) Adaptation to speed indicating size ordered recruitment of propulsive synergies indicated by the width of the trace. (d–g) Omnidirectional
patterns indicating co-activation of antagonists for joints that serve a postural function. Ele, elevator synergy; Dep, depressor synergy; Pro, protractor synergy;
Ret, retractor synergy; Ext, extensor synergy; Flx, flexor synergy. The time marks occur once/second.
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360352

to be mediated by reciprocal inhibition (Sillar et al., 1987).
Intersegmental connections consist of a central synchronous
component that is probably most significant in the context of
leg waving behavior (Pasztor and Clarac, 1983), as well as
an effective indirect inhibitory component mediated by the
load sensitive funnel canal organs in the dactyl of each limb
which is appropriate to the gaits seen during walking
(Libersat et al., 1987).
In the controller, the limb coordinating systems are
implemented as the elevator synergies that pass messages to
the elevators of adjacent limbs (Fig. 7a). Fig. 7b and c
indicates stable alternating tetrapod coordination generated
by the ambulation controller with the empirically estab-
lished contralateral phase lag of 0.5 and an ipsilateral phase
lag of 0.4 (Clarac and Barnes, 1985).
2.6. Myomorphic actuators
We have had excellent results with artificial muscle
fabricated from the shape memory alloy nitinol (Fig. 8a).
Shape memory metals undergo a state transformation
(Fig. 8b) from a deformable state (martensite) to a
remembered state (austenite). When formed into wires,
nitinol may change its length by as much as 5% during
contraction (Duering et al., 1990). Nitinol has a maximum
and minimum range of action specified by its stretched and
remembered lengths. Within this range its control is
proportional, depending upon the level of current passed
or the linear extent of the wire through which current is
passed (Witting and Safak, 2000).
Nitinol wires can be activated to contract through the
heat generated by the pulses of electrical current. We use
pulse width duty cycle modulation to grade the proportion
of martensite that transforms to austenite. Discrete duty
cycles are used to mimic small medium and large motor
units (Fig. 8c). Increases in duty cycle increase the speed
and amplitude of contraction (Fig. 8d). These properties
map directly on the architecture of the ambulation
controller.
2.7. Neuromorphic sensors
We have developed several biomimetic sensors necess-
ary to mediate reactive tactile navigation on the ocean
bottom. All sensors code information with a labeled line
code (Bullock, 1978). Each sensor is represented by a byte,
Fig. 7. (a) Messages passed by the coordinating systems. The bars indicate the time of activity in the elevator synergy of legs 1–4 on the left and right sides. (b)
Coordination during increasing speed of walking. (c) Interlimb coordination during a turn to the left indicated by the horizontal bar.
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360 353

each bit of which corresponds to a labeled line. The labeled
line represents three characteristics of the stimulus (1) The
sensory modality, (2) the receptive field or orientation
relative to the body and (3) the amplitude of the stimulus.
All sensors are polled by the state machine and return a byte
representing their status. This byte is compared to bit masks
representing different releasers.
2.7.1. Antennae
To realize antennal sensors (Fig. 4), we rely on the use of
strain gauges to monitor bending. The strain gauge is
mounted in the middle of the antennae and interfaced to the
mothor board using a Wheatstone bridge circuit. A PIC
microcontroller digitizes the analog signal converts it to
seven discrete states represented by a seven bit labeled line
code. Each of the bits represents one of seven bend states:
straight, three magnitudes of bending to lateral or medial
and an eighth bit that indicates when the antenna is buckled
by a head on collision.
These antennae can be projected at four different angles
by the motor drive depending on context. They are capable
of distinguishing water currents from collisions with solid
objects such as rocks. The byte masks associated with the
two antennae must be interpreted in the context of the
position of the antennae relative to the hull. For example,
the buckling bit can be set by a head-on collision if the
antennae are projected forward or by a collision to either
side if they are projected laterally. Thus identification of
releasers requires that the returned data are interpreted
through a decision tree based on the position or movement
of the antennae. Responses must also be interpreted in the
context of movements between positions that can bend the
antennae.
2.7.2. Active touch function
The antennae are controlled by a motor drive that allows
them to sweep over arbitrary ranges between four basic
positions. If the antennae collide with an object in a sweep
range it will indicate bending to high magnitudes and a
relative position is indicated by the time of the bending
within the sweep (Zeil et al., 1985).
2.7.3. Collision detection function
One of the characteristics of the underwater operation of
these sensors is that they buckle when they make a head on
collision with an object. This is associated with a rapid left–
right bend of the antennae. The PIC microcontroller is
programmed to recognize such events and sets the eighth bit
for each antennae for 150 ms following such collisions.
2.7.4. Water motion function
The antennae have been calibrated in a laminar flow
system and also indicate water flow. For example if the
antennae are statically projected forward and both are bent
to the right this would indicate flow from the left. If the
antennae are projected statically laterally to the body axis,
they become a flow sensor for axial currents and can
Fig. 8. (a) A nitinol myomorphic actuator. (b) The shape memory effect (from Duering et al., 1990). (c) Pulse width duty cycle modulation. (d) Graded
contractions produced by a nitinol muscle element. The upper trace in each panel represents the shortening of a muscle produced by the pulse trains in the lower
trace. Upward deflections indicate shortening. The duty cycle is indicated to the right of each panel.
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360354

indicate when the vehicle is oriented into flow or surge on
the basis of symmetric deflections on both sides.
2.7.5. Inclinometers
Lobsters normally sense the orientation of their bodies
using vestibular organs called the statocysts (Cohen, 1955).
Here individual labeled lines code for orientation relative to
gravity in the pitch and roll planes. For biomimetic
implementation we use a MEMs accelerometer and convert
the analog signal into eight discrete sectors of resolution for
an 808 range of motion relative to vertical. The labeled lines
are represented by eight bits for each of the eight sectors.
2.7.6. Bump sensors
Observations of movements of blinded lobsters using a
‘lobster cam’ indicate that they use a ‘bump’ sense mediated
by their chelipeds to navigate around low objects like rocks.
We have implemented bump sensors for the claws based on
a solid-state accelerometer. These sensors take the analog
signal coming form the accelerometer, full wave rectify it,
low pass filter it and threshold the low pass signal to
generate a bump signal.
2.7.7. Compass
It has recently been demonstrated that spiny lobsters can
sense the earths magnetic field and orient their migrations
relative to it (Boles and Lohmann, 2003). To confer the
robot with a similar ‘sense of direction’ we have integrated a
flux gate compass with interface that provides three bits of
resolution (eight heading sectors).
2.8. Behavioral controller
We are currently in the process of implementing the
behavioral controller of the robot. All of the behavior of
the robot can currently be controlled by teleoperation,
pre-programmed behavioral sequences can be played
and all of the sensors are functional. We are currently
in the process of implementing the sensory-motor
reflexes by optimizing the connections between sensor
objects and command objects. In the following sections
I present a progress report of the present organization of the
software and our plans for implementation of reactive
autonomous behavior.
The higher order controller of the ambulatory vehicle is
based on command objects. The operational state of the
appendages is specified by a set of 11 internal state variables
that configure the appendage state machines on the fly. The
basic structure of the controller is a real time loop that polls
sensors, manages a command stack, selects behavioral acts
and updates the leg state machines. An attention module
polls each of the sensors at a different rate depending on
behavioral context. The attention module keeps a list of the
clock time at which subsequent poll of a sensor is to occur
in temporal order and updates this list every time a sensor
is read.
Fig. 9. Behavioral quantization. Upper left panel: Screen shot of the program ColorImage in the process of decomposing a behavioral sequence into underlying
commands. The output of ColorImage is a command sequence table (bottom panel), where each row is a frame of the movie and each column represents the
changes underlying rheotaxic behavior. The upper right panel is a cartoon of a rheotaxic sequence that can be displayed on an animator.
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360 355

2.9. Behavioral libraries
The control architecture includes a command sequencing
capability that can store pre-programmed action patterns,
reverse engineered from observations of the adaptations of
the animal model that form the basis of a behavioral library
(Ayers et al., 2002). The fundamental assumption of this
behavioral control model is that the posture and action of the
different body parts is specified by a set of command
systems (Bowerman and Larimer, 1974a,b) that command
the task group to generate different states. To derive the
underlying commands states, we determine the posture or
action of the different appendages in each of the frames of a
movie. We categorize these actions or movements into one
of a set of states (see below). The movies are selected to
capture the model organism performing the act that one
wants the vehicle to mimic (Fig. 9). This analysis therefore
represents an inference of the states of the underlying
command neuron activity. We have found the following set
of command states adequate to define the ongoing behavior
of both the lobster and the robotic vehicle.
Thorax Pitch: rostrum up, level, rostrum down.
Thorax Roll: left up, level, right up
Thorax Yaw: hard left, easy left, straight, easy right, hard
right
Thorax Height: high, normal, low
Walking direction: standing, forward, backward, lateral
leading lateral trailing
Walking speed: slow, medium, fast, stop
Claw Pitch: up, normal, down
Claw Yaw: (left and right): extended, normal, meral
spread, lateral spread
Antennae Yaw: protracted, normal, lateral, retracted
Uropod Posture: flared, normal, adducted
Abdominal Pitch: extended, elevated, normal,
depressed, flexed
To derive robotic control sequences from video data, we
perform finite state analysis of the task groups that mediate
locomotion and searching individually to determine which
synergistic sets are operant during different behavioral acts.
A panel of buttons that represent different states of the task
groups (e.g. elevation vs. depression of the chelipeds, etc.)
are available to the investigator to specify which groups are
active (Fig. 9). By clicking on the appropriate buttons for
each frame, it is possible to efficiently quantify the activity
of all task groups at high temporal resolution from
videotapes of specimens behaving in a variety of situations.
The result of this analysis is a command sequence; a table of
numbers where each row represents a frame of video and
each column represents the state of a particular task group.
These tables are abstracted by the sequencer to the start and
stop times of particular commands for sequencing. This
expanding set of command sequences constitutes the
behavioral library of the vehicles. These libraries are
evoked by sensor input and played out by a sequencer that
is composed of the following components.
2.9.1. Releasers
When a sensor is polled, the result is compared to a list of
byte masks defining particular releasers. For example if an
antennal poll detected that the left antenna is bending
medially and the right antenna is bending laterally this
would constitute the releaser for a rheotaxic rotational turn
to the left. When the attention module identifies a behavioral
releaser, a look up table is used to identify the behavioral
sequence table associated with that particular releaser.
2.9.2. Sequencer
Whenever a releaser is identified the sequencer sets up
the sequence of command state changes that mediate the
evoked behavior. The sequencer actually performs two
tasks. First the releaser may specify a suppressor or table of
incompatible action components. For example, if the
evoked behavior involves forward walking, any instances
of backward walking command action components must be
cleared from the command stack. This suppressor action is
the locus of implementation of behavioral choice between
incompatible commands. Secondly, the releaser sets up the
behavior by pushing the behavioral state changes on the
stack in temporal order.
Animals normally can evoke behavioral acts at different
levels of intensity and this is typically mediated by
neuromodulators. The behavioral sequencer allows the
command state transitions to be pushed on the stack at
different temporal compressions to vary intensity. In
addition, the pulse width duty cycles associated with the
low, medium and high levels of recruitment of the nitinol
actuators can be varied. Thus at high intensity the sequence
of command state changes are compressed on the stack and
the recruitment state pulse width duty cycle mapped to a
high level of intensity. In contrast, at low intensity the
sequence of command state changes are expanded on the
stack and the recruitment state pulse width duty cycle
mapped to a low level of intensity.
During operation, releasers are detected by sensors and
evoke the sequencer to push the sequence of command state
changes on the command stack. As the robot continues
operation the command state changes are played out in
temporal order to modulate the operation of the appendage
state machines. As behaviors may superimpose or switch in
response to sensor input these operations are mediated by
the sequencer.
2.10. Sensory-motor reflexes
The robot features an expanding set of exteroceptive
reflexes in response to deviations from a desired heading or
posture, collision and surge. Exteroceptive reflexes act on
command systems and or intersegmental modulatory
interneurons by inducing lateral biases (Davis and Ayers,
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360356

1972; Kennedy and Davis, 1977). The way we have
implemented these reflexes can be best illustrated by control
of responses to surge by the antennae.
Depending on the habitat, the robot may be exposed to
low flow (Fig. 10a), unidirectional flow from tides (Fig. 10b)
or bidirectional surging flow from waves (Fig. 10c). In
surging flow, the releaser is flow detected by the antennae
that are held laterally. Antennal labeled lines responding to
lateral bending of the antennae project to the contralateral
walking command while labeled lines responding to medial
bending project to the ipsilateral forward walking command
(Fig. 10d). Due to this logic, if the surge is approaching from
off the midline, it will bias the output of the walking
commands such that the vehicle will yaw into the flow if the
flow is coming from the front but yaw away from the flow if
it is coming from the rear. This combination of reflexes
facilitates locomotion in surging water. When surge is
coming from the front, the vehicle will load compensate by
increasing the output of the walking commands. When
surge is coming from the rear, the vehicle will run with it.
Examples of control signals to the left and right leg when
yawing are shown in Fig. 10d. The robot has been modeled
dynamically to account for both hydrodynamic and
frictional forces (Safak, 2001). In still water the simulation
indicates the robot can locomote at speeds of up to,6 cm/s
which is above the observed speed of the present vehicle
(,3 cm/s). In the absence of hydrodynamic compensation,
the simulation indicates that the vehicle will stall when
walking into flow velocities of between 10 and 12 cm/s
depending on the coefficient of friction, and a maximum
speed of ,20 cm/s when walking with a 30 cm/s assisting
flow (Safak, 2001). Above opposing flow velocities of
12 cm/s the vehicle will slip.
In addition to yaw plane compensation, lobsters and
crayfish use their chelae, abdomen and uropods as
hydrodynamic control surfaces (Fig. 1) and in real lobsters
these surfaces are large relative to the thorax (Fig. 11a).
Compensatory reflex responses to water currents involve
simultaneous compensation of the thorax in the pitch plane
as well as postural responses of the abdomen and chelipeds.
Fig. 11 b and c indicates the compensatory response of the
anterior and posterior control surfaces in the lobster robot.
The ambulation controller supports antigravity recruiters for
each leg that act on the depressor synergy and mediates
pitch and roll compensation. Pitch control ‘interneurons’
bias the depressor recruiters along the body axis and
mediate pitch compensation (Fig. 11d–f). Pitching the hull
forward will provide additional hydrodynamic compen-
sation (Martinez, 2001). These compensatory responses to
water currents and surge can simultaneously integrate both
yaw and pitch plane components independent of the
direction and speed of walking.
2.11. Navigation
The yaw components of navigation are mediated in
biological systems by taxes and kineses (Loeb, 1918;
Braitenberg, 1984). Positive yaw taxes or attraction occur
Fig. 10. (a–c) Hydrodynamic environments that the robot might be expected to operate in (from Martinez, 2001). (a) Still water. (b) Unidirectional flow. (c)
Bidirectional surge. (d) Reflex circuitry to mediate rheotaxic exteroceptive reflexes resulting from water currents from off center. See text for description of
operation. (d) Yaw plane modulating motor patterns during forward walking. The traces indicate the elevator, depressor and retractor synergies of the left and
right sides.
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360 357

when sensor bias directs locomotion toward source and are
generally mediated by contralateral causality between
sensors and effectors. Negative yaw taxes or avoidance
occur when sensor bias directs locomotion away from
source and are generally mediated by ipsilateral causality
between sensors and effectors. In the ambulation controller,
such inputs send messages to both the parametric command
as well as the recruiting component. In attractive reflexes
the messages are sent to the contralateral command objects
while during avoidance reflexes (negative taxis) the
messages are sent to the ipsilateral command objects. As a
result, the controller both biases the period on the two sides
(walking faster on the side turned away from) as well as the
recruiters on the two sides to generate more propulsive force
on the faster side (Fig. 9e). Control schemes like that of
Fig. 9d can be easily adapted to a variety of beacon tracking,
avoidance, docking and search behavior through acoustic,
magnetic, optical, tactile, gravitational, flow and chemical
sensors.
Acknowledgements
Supported by the Defense Advanced Research Projects
Agency and the Office of Naval Research.
References
Alexander, R., 1971. Size and Shape, Edward Arnold, London, p. 59.
Atema, J., Voigt, R., 1995. Behavior and sensory biology. In: Factor, J.R.,
(Ed.), The Biology of the Lobster, Homarus americanus, Academic
Press, San Diego, pp. 313–344.
Atwood, H.L., 1976. Organization and synaptic physiology of crustacean
neuromuscular systems. Progress in Neurobiology 7, 291–391.
Fig. 11. Hydrodynamic compensation. (a) Dorsal view of Homarus indicating the relatively large hydrodynamic control surfaces formed by the chelae,
abdomen and uropods (adapted from Huxley, 1973). (b) The lobster robot with abdomen and ‘chelae’ in the horizontal position. (c) The lobster robot with
abdomen and chelae in the wedge configuration (see Fig. 1). (d–f) Network combinatorics between modulatory interneurons and depressor motor synergies to
mediate rostrum (row) down (d), normal (e) and rostrum up (f) postures.
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360358

Ayers, J., Clarac, F., 1978. Neuromuscular strategies underlying different
behavioral acts in a multifunctional crustacean leg joint. Journal of
Comparative Physiology A: Sensory, Neural and Behavioral Physi-
ology 128, 81–94.
Ayers, J., Crisman, J., 1992. The lobster as a model for an omnidirectional
robotic ambulation control architecture. In: Beer, R., Ritzmann, R.,
McKenna, T. (Eds.), Biological Neural Networks in Invertebrate
Neuroethology and Robots, pp. 287–316.
Ayers, J., Davis, W.J., 1977. Neuronal control of locomotion in the lobster,
Homarus americanus. I. Motor programs for forward and backward
walking. Journal of Comparative Physiology A: Sensory, Neural and
Behavioral Physiology 115, 1–27.
Ayers, J., Selverston, A.I., 1979. Monosynaptic entrainment of an
endogenous pacemaker network: a cellular mechanism for von Holst’s
magnet effect. Journal of Comparative Physiology A: Sensory, Neural
and Behavioral Physiology 129, 5–17.
Ayers, J., Davis, J., Rudolph, A., 2002. Neurotechnology for Biomimetic
Robots, MIT Press, Cambridge.
Ayers, J., Volkovski, A., Rulkov, N., Selverston, A., Abarbanel, H.,
Rabinovich, M., 2003. Building a CNS for the lobster robot with INLS
electronic neurons. Society for Neuroscience Abstracts 27: 278. 14.
Beer, R.D., 1991. Intelligence as Adaptive Behavior: An Experiment in
Computational Neuroethology, Academic Press, New York.
Beer, R.D., Chiel, H.J., Quinn, R.D., Ritzmann, R.E., 1998. Biorobotic
approaches to the study of motor systems. Current Opinion in
Neurobiology 8, 777–782.
Beltz, B., 1995. Neurobiology and neuroendocrinology. In: Factor, J.R.,
(Ed.), The Biology of the Lobster, Homarus americanus, Academic
Press, San Diego, pp. 267–285.
Boles, L.C., Lohmann, K.J., 2003. True navigation and magnetic maps in
spiny lobsters. Nature 421, 60–63.
Bowerman, R.F., Larimer, J.L., 1974a. Command fibers in the circumoe-
sophageal connectives of crayfish. I. Tonic fibers. Journal of
Experimental Biology 60, 95–117.
Bowerman, R.F., Larimer, J.L., 1974b. Command fibers in the circumoe-
sophageal connectives of crayfish. II. Phasic fibers. Journal of
Experimental Biology 60, 119–134.
Braitenberg, V., 1984. Vehicles: Experiments in Synthetic Psychology,
MIT Press, Cambridge.
Brooks, R.A., 1989. A robot that walks; emergent behaviors from a
carefully evolved network. Neural Computation 1, 253–262.
Bullock, T.H., 1978. An Introduction to Neuroscience, Freeman, San
Francisco.
Burrows, M., 1996. The Neurobiology of an Insect Brain, Oxford
University Press, Oxford, 696 pages.
Cattaert, D., Le Ray, D., 2001. Adaptive motor control in crayfish. Progress
in Neurobiology 63, 199–240.
Chrachri, A., Clarac, F., 1989. Synaptic connections between motor
neurons and interneurons in the fourth thoracic ganglion of the crayfish,
Procambarus clarkii. Journal of Neurophysiology 62, 1237–1250.
Chrachri, A., Clarac, F., 1990. Fictive locomotion in the fourth thoracic
ganglion of the crayfish, Procambarus clarkii. The Journal of
Neuroscience 10, 707–719.
Clarac, F., Barnes, W.J.P., 1985. Peripheral influences on the coordination
of the legs during walking in decapod crustaceans. In: Bush, B.M.H.,
Clarac, F. (Eds.), Coordination of Motor Behavior, Cambridge
University Press, Cambridge, pp. 249–269.
Cobb, J., Phillips, B. (Eds.), 1980. The Biology and management of
lobsters, Physiology and Behavior, Vol. I. Academic Press, New York.
Cohen, M.J., 1955. The function of receptors in the statocyst of the lobster,
(Homarus americanus). Journal of Physiology 130, 9–33.
Davis, W.J., 1968. Lobster righting responses and their neural control.
Proceedings of the Royal Society B 70, 435–456.
Davis, W.J., 1971. Functional significance of motor neuron size and soma
position in the swimmeret system of the lobster. Journal of
Neurophysiology 34, 274–288.
Davis, W.J., Ayers, J., 1972. Locomotion: control by positive feedback
optokinetic responses. Science 177, 183–185.
Davis, W.J., Kennedy, D., 1972. Command interneurons controlling
swimmeret movements in the lobster 1. Types of effects on
motoneurons. Journal of Neurophysiology 35, 1–12.
Delcomyn, F., 1980. Neural basis of rhythmic behavior in animals. Science
210, 492–498.
Duering, T.M., Stockel, K.N., Wayman, D.C.M., 1990. Engineering
Aspects of Shape Memory Alloys, Butterworth Heinemann, Ltd,
London.
Edwards, D.H., Heitler, W.J., Krasne, F.B., 1999. Fifty years of the
command neuron: the neurobiology of escape behavior in the crayfish.
Trends in Neuroscience 22, 153–161.
El Manira, A., DiCaprio, R.A., Cattaert, D., Clarac, F., 1991. Monosynaptic
interjoint reflexes and their central modulation during fictive
locomotion in crayfish. European Journal of Neuroscience 3,
1219–1231.
Evoy, W., Ayers, J., 1982. Locomotion and control of limb movements. In:
Atwood, D.C., Sandeman, H. (Eds.), The Biology of Crustacea, Neural
Integration and Behavior, Vol. 4. Academic Press, New York,
pp. 62–106.
Factor, J.R., 1995. Biology of the Lobster, Homarus americanus, Academic
Press, New York.
Grasso, F.W., 2000. Flow and chemo-sense for robot and lobster guidance
in tracking chemical sources of turbulence. In: Ayers, J., Davis, J.,
Rudolph, A. (Eds.), Neurotechnology for Biomimetic Robots, MIT
Press, Cambridge, pp. 553–571.
Harris-Warrick, R.M., Marder, E., 1991. Modulation of neural networks for
behavior. Annual Review of Neuroscience 14, 39–57.
Huxley, T.H., 1973. The crayfish: an introduction to the study of zoology,
MIT Press, Cambridge.
Kennedy, D., Davis, W.J., 1977. Organization of invertebrate motor
systems. In: Geiger, R., Kandel, E., Brookhart, J.M. (Eds.), Handbook
of Physiology, American Physiology Society, Bethesda, MD, pp.
1023–1087.
Larimer, J.L., 2000. The interneurons of the abdominal positioning system
of the crayfish. Brain, Behavior and Evolution 55, 241–247.
Libersat, F., Clarac, F., Zill, S., 1987. Force sensitive mechanoreceptors of
the dactyl of the crab: single unit responses during walking and
evaluation of function. Journal of Neurophysiology 57, 1601–1607.
Loeb, J., 1918. Forced Movements, Tropisms and Animal Conduct,
Lippincott, Philadelphia.
Martinez, M.M., 2001. Running in the surf: hydrodynamics of the shore
crab Grapsus tenuicrustatus. Journal of Experimental Biology 204,
3097–3112.
Maude, S.H., Williams, D.D., 1983. Behavior of crayfish in water currents:
hydrodynamics of eight species with reference to their distribution
patterns in Southern Ontario. Canadian Journal of Fisheries and Aquatic
Sciences 40, 68–77.
Pasztor, V.M., Clarac, F., 1983. An analysis of waving behavior: an
alternative motor program for the thoracic appendages of decapod
crustacea. Journal of Experimental Biology 102, 59–77.
Pinsker, H.M., Ayers, J., 1983. Neuronal Oscillators. In: Willis, W.D.,
(Ed.), The Clinical Neurosciences. Section Five. Neurobiology,
Chapter, Churchill Livingstone Publishers, London, pp. 203–266,
Chapter 9.
Safak, K., 2001. Modeling and Analysis of a Biomimetic Underwater
Robot, PhD Dissertation Northeastern University.
Sandeman, D.C., Atwood, H.L., 1982. The Biology of crustacea, Neural
Integration and Behavior, Vol. 4. Academic Press, New York.
Selverston, A., 1999. General principles of rhythmic motor pattern
generation derived from invertebrate CPGs. Progress in Brain Research
123, 247–257.
Sillar, K.T., Skorupski, P., 1986. Central input to primary afferent
neurons in crayfish, Pacifastacus lenuisculus, is correlated with
rhythmic motor output of thoracic ganglia. Journal of Neurophysiology
55, 678–688.
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360 359

Sillar, K.T., Clarac, F., Bush, B.M.H., 1987. Intersegmental coordination of
central neural oscillators for rhythmic movements of the walking legs in
crayfish, Pacifastacus lenuisculus. Journal of Experimental Biology
131, 245–264.
Sparks, D.L., Kristan, W.B., Shaw, B.K., 1997. The role of population
coding in the control of movement. In: Stein, P.S.G., Grillner, S.,
Selverston, A., Stuart, D. (Eds.), Neurons Networks and Motor
Behavior, MIT Press, Cambridge, MA, pp. 21–32.
Stein, P.S.G., 1976. Mechanisms for interlimb phase control. In: Herman,
R.M., Grillner, S., Stein, P.S.G., Stuart, D.G. (Eds.), Neural Control of
Locomotion, Plenun Publishing, New York, pp. 465–487.
Stein, P.S.G., 1978. Motor systems, with specific reference to the control of
locomotion. Annual Review of Neuroscience 1, 61–81.
Wilson, D.M., 1967. Stepping patterns in tarantula spiders. Journal of
Experimental Biology 47, 133–151.
Witting, J., Safak, K., 2000. Shapememory alloy actuators applied to biomimetic
underwater robots. In: Ayers, J., Davis, J., Rudolph, A. (Eds.), Neurotechnol-
ogy for Biomimetic Robots, MIT Press, Cambridge, pp. 117–135.
Zeil, J., Sandeman, R., Sandeman, D., 1985. Tactile localization: the
function of active antennal movements in the crayfish Cherax
destructor. Journal of Comparative Physiology A: Sensory, Neural
and Behavioral Physiology 157, 607–617.
J. Ayers / Arthropod Structure & Development 33 (2004) 347–360360





























