Embed Size (px)
Citation preview

1
UNIVERSIDAD DE LA REPÚBLICA.
FACULTAD DE CIENCIAS.
CURSO CURRICULAR DE PALEONTOLOGÍA 2012
MATERIAL DE APOYO PARA PRÁCTICO DE ORIGEN DE LOS AMNIOTAS.
RESPONSABLE: Dr. Graciela Piñeiro
El origen de los amniotas se produce en algún momento del Carbonífero
(probablemente a mediados de este período), cuando algunos anfibios primitivos se
independizan totalmente del agua mediante el desarrollo de un huevo con membranas
extraembrionarias (huevo amniota). <Los restos más antiguos provienen de una
localidad llamada Joggins, del Carbonífero Tardío de Canadá.
Probables ancestros:
Los Diadectomorfos, un grupo de anfibios de mediano a gran tamaño presentes en el
Carbonífero Medio a Superior que incluyen formas herbívoras (ej. Diadectes) y
carnívoras (ej. Limnoscelis). Diadectes habría estado adaptado a una dieta sobre
vegetales duros y presentaba una marcada heterodoncia en su dentición marginal.
Existen problemas con este grupo referidos a su gran tamaño y robustez, comparado con
los diminutos y gráciles primeros amniotas. Por otra parte, algunos autores consideran
que los diadectomorfos podrían ser amniotas primitivos y probablemente comparten un
ancestro común con los pequeños amniotas hallados en Joggins.
Primitivos amniotas:
Los restos más antiguos de amniotas provienen del Carbonífero Superior de Norte
América, de territorios que ocupaban la franja cercana al Ecuador de la Pangea, y que
ostentaban climas bastante más benignos.

2
Joggins, una localidad del Carbonífero Tardío de Canadá.
En Gondwana, a finales del Carbonífero se iniciaba un período de glaciaciones y el
clima era hostil para el desarrollo de este grupo de vertebrados. No obstante, en el
comienzo del Pérmico, los mesosaurios representan a los amniotas más primitivos de
Gondwana y sus restos se encuentran representados en Uruguay, Brasil, el sur de
Paraguay y Africa del Sur.
Cómo reconocemos a estos primitivos amniotas a través de los fósiles?
De acuerdo a lo que indica el registro fósil, los representantes más primitivos eran de
pequeño tamaño y según algunos autores también sus ancestros más directos debieron
presentar esta característica. Sin embargo, los diadectomorfos, considerados el grupo
hermano de los amniotas eran formas robustas y en general de gran tamaño. Para la
adaptación a un modo de vida terrestre, el pequeño tamaño podría haber sido favorable
para el soporte del cuerpo en tierra sin modificaciones importantes del esqueleto.
La adquisición del huevo amniota fue un paso importante en el proceso de la
conquista del medio terrestre y pudo estar acompañado de la fertilización interna, la
reducción del tamaño del adulto, la reducción del número de huevos y el aumento de
tamaño de los mismos (fundamentalmente a partir de una mayor cantidad de yema
disponible para la alimentación de los embriones), y de estadios larvales cada vez más
cortos y con tendencia a la desaparición total. Más tarde (geológicamente hablando al
comienzo del Triásico) esa tendencia se revierte cuando aparece la posibilidad de
biomineralizar una cáscara de protección. Allí los períodos embrionarios se hacen más
largos nuevamente, y el número de huevos por camada puede aumentar de acuerdo a las
distintas especies. No obstante, un linaje de amniotas seguirá perfeccionándose hacia un
tipo de reproducción donde la retención del embrión en el útero materno será la
estrategia más común: el linaje de los synápsidos, incluyendo los mamíferos.
Los cambios osteológicos desde los anfibios a los primeros amniotas.
Los amniotas tempranos difieren de los anfibios que les dieron origen
básicamente en:

3
*la morfología de la columna vertebral (diferenciación de las vértebras
cervicales y la delimitación mayor del cuello),
*el mayor desarrollo de las cinturas y los apéndices
Estos cambios están ligados a la optimización de la locomoción en tierra.
Además, debemos tener en cuenta las modificaciones en la estructura del paladar
y la dentición, que están ligados a la adquisición de nuevos hábitos de alimentación.
Otros caracteres que nos ayudan a diferenciarlos de los anfibios son:
*ausencia de dientes laberintodontes (dientes cuyo corte transversal muestra pliegues de
dentina dentro de la cavidad pulpar).
*reducción de la escotadura ótica (estructura donde se inserta la membrana timpánica en
anfibios, principalmente en aquellos de hábitos acuáticos).
Estructura de la región temporal en la clasificación de los amniotas:
Se reconocen tres planes básicos en el cráneo de los amniotas, de acuerdo a la
presencia/ausencia de aberturas en la región temporal del cráneo (ventanas temporales)
Anápsidos (línea tortuga): sin ventanas temporales (el patrón ancestral)
Synápsidos (línea mamaliana): con una ventana a cada lado del cráneo de posición
inferior.
Diápsidos (línea reptiliana): con dos ventanas temporales a cada lado del cráneo (una
superior y otra inferior).

4
Hylonomus (a, b), el amniota más antiguo de Yoggins y
los patrones craneanos ya presentes en el Carbonífero Tardío: Anapsido (c), synapsido
(d) y diápsido (e).
Huesos que intervienen en la delimitación de las
ventanas temporales y los diferentes patrones y sus modificaciones (ver ejemplos en las
fotografías representadas a continuación).
Tortugas Dimetrodon (synapsido basal) Tuátara (diápsido generalizado)
Plesiosaurio (diápsido modificado a euriápsido= lepidosaurios)
En el Carbonífero Superior los tres patrones ya estaban presentes.
Los Mesosaurios
Fueron los primeros reptiles acuáticos conocidos, hábito que probablemente no fue
adquirido secundariamente, sino que es el que heredaron de sus ancestros hasta el
momento desconocidos. Probablemente los mesosaurios se diferencian como linaje

5
hacia finales del Carbonífero, cuando el clima se volvió más benigno luego de la
glaciación del Gondwana.
Procedencia geográfica y estratigráfica:
Sus restos se encuentran en depósitos del Pérmico Inferior de Uruguay (Formación
Mangrullo en Deptos. de Tacuarembó, Cerro Largo y Rivera), Brasil (Fm. Iratí) y
Africa del Sur (Fm. Whitehill). Esta distribución geográfica y la escasa probabilidad que
estos reptiles hayan podido cruzar el océano Atlántico tal como hoy lo conocemos,
sugiere que América del Sur y Africa estaban unidos durante el comienzo del Pérmico,
argumentación que apoya la hipótesis de la Deriva Continental y la existencia del
Gondwana. Si bien fueron organismos acuáticos, las adaptaciones anatómicas que
caracterizan a los vertebrados marinos estaban ausentes.
Biocrón: Pérmico Temprano (biocrón reducido, lo que indica que son buenos fósiles
guía de este corto período de tiempo).
Tafonomía:
Sus restos pueden encontrarse preservados de la siguiente manera:
*huesos o moldes de los distintos huesos desarticulados formando bone beds donde
todos los elementos del esqueleto pueden estar representados, pero costillas y vértebras
son los más frecuentes, ya que son los más numerosos en el esqueleto).
*esqueletos o moldes de esqueletos parcialmente articulados, representados
principalmente por fragmentos de la columna vertebral y costillas, que pueden estar
asociados a huesos de las cinturas y extremidades.
*esqueletos completos o moldes de esqueletos completos, con cráneo y cola
preservados en posición anatómica.
Características de los mesosaurios que los identifican entre otros amniotas primitivos:

6
*Cráneo corto en su región postorbital, pero con un largo hocico provisto de largos y
delgados dientes. Probablemente se alimentaran de crustáceos pigocefalomorfos, los
cuales se encuentran asociados a sus restos.
*Costillas dorsales paquiostóticas (ensanchadas en su parte media). Este carácter les
habría permitido sumergirse con menor gasto energético. Debido a su pequeño tamaño,
el aumento de peso en la región dorsal pudo ser adaptativa y les permitió una inmersión
más rápida en busca de su alimento. Sin embargo, podrían haber existido mesosaurios
que no desarrollaron paquiostosis, los cuales habrían tenido una vida más anfibia,
adaptada a las márgenes del cuerpo de agua y solo muy cortos períodos de tiempo
dentro del agua.
Naturaleza del cráneo de los mesosaurios:
Un enigma que generó mucha controversia entre los autores. Podrían haber existido
mesosaurios anápsidos (Modesto, 2006) y otros que habrían desarrollado ventanas
temporales como promulgan algunos autores? Los restos hallados en Uruguay sugieren
la presencia de una ventana temporal y no hemos hallado hasta el momento evidencias
de mesosaurios anápsidos.
LA LINEA DE LOS SYNAPSIDOS (línea mamaliana)
La presencia de una ventana temporal de posición inferior distingue a los sinápsidos del
resto de los amniotas, ya que todos sus integrantes sin excepciones presentan este
carácter. Por ello no existen dudas que este carácter es una sinapomorfía que diagnostica
al grupo de los sinápsidos.
Entre los integrantes de este grupo se encuentran los pelicosaurios, los terapsidos y los
mamíferos, conformando una línea evolutiva en este sentido:
Pelycosauria---------------------- Therapsida--------------------- Mammalia Carbonífero Sup y Pérmico---Pérmico Superior-Triásico Superior---Triásico Superior –Reciente.
En los pelicosaurios encontramos pocos caracteres mamalianos, pero en algunos de
ellos podemos observar una incipiente heterodoncia con el crecimiento de algunos
dientes en la región de los caninos. Algunos como Dimetrodon podrían haber regulado
de alguna manera su temperatura corporal mediante el desarrollo de una estructura
dorsal llamada vela que consistía en una membrana sobre el dorso sostenida por las
espinas de las vértebras dorsales, las cuales eran exageradamente altas.

7
En los terápsidos en cambio, muchos caracteres son compartidos con sus descendientes
los mamíferos. En efecto, los terápsidos más evolucionados en el Triásico, podrían
haber tenido pelo y glándulas mamarias.
LA LÍNEA DE LOS ANAPSIDA
Captorhinidae
Pareiasauridae
Procolophonoidea
Parareptilia,características:
Los parareptiles son los anápsidos dentro de los saurópsidos. No obstante, su posición
filogenética y su status dentro de Amniota no está muy clara.
Incluye:

8
Procolophonoidea:
Leptopleuron
Owenetta, procolophonoideo que tiene
como características primitivas,
dentición marginal homodonta
consistente de muchos dientes cónicos
similares, paladar con dentículos
dispersos, y dentición superior que
cubre a la inferior cuando las
mandíbulas están cerradas. Estos son
caracteres PRIMITIVOS,
PLESIOMÖRFICOS dentro de
Procolophonoidea.
Procolophon, procolophonoideo que presenta dentición marginal heterodonta,
consistente de pocos dientes anteriores incisiviformes y dientes maxilares molariformes
o ensanchados transversalmente. Además, los dientes palatinos están reducidos y
presentan un arreglo paracoanal (alrededor de la coana), como se aprecia en la figura
más a la derecha de la serie. Nótese que las piezas dentarias en las series marginales
superior e inferior se interdigitan cuando las mandíbulas están cerradas. Estos caracteres
son DERIVADOS para Procolophonoidea.
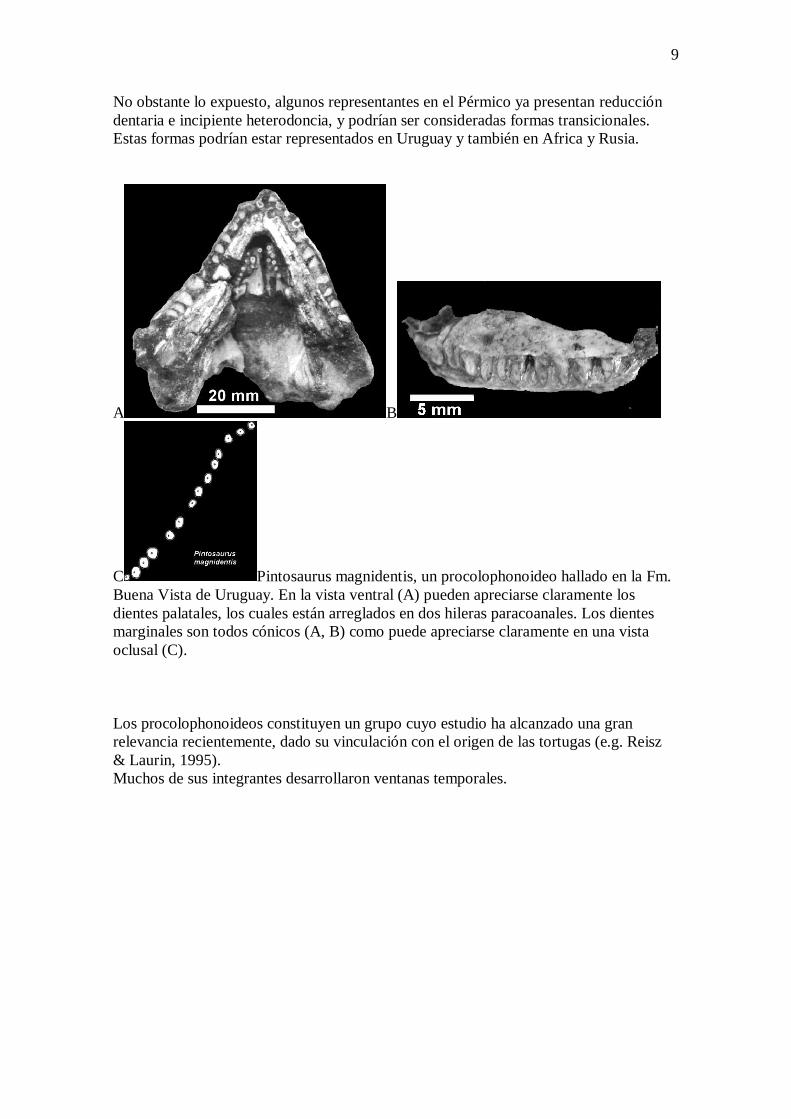
9
No obstante lo expuesto, algunos representantes en el Pérmico ya presentan reducción
dentaria e incipiente heterodoncia, y podrían ser consideradas formas transicionales.
Estas formas podrían estar representados en Uruguay y también en Africa y Rusia.
A
A B
C Pintosaurus magnidentis, un procolophonoideo hallado en la Fm.
Buena Vista de Uruguay. En la vista ventral (A) pueden apreciarse claramente los
dientes palatales, los cuales están arreglados en dos hileras paracoanales. Los dientes
marginales son todos cónicos (A, B) como puede apreciarse claramente en una vista
oclusal (C).
Los procolophonoideos constituyen un grupo cuyo estudio ha alcanzado una gran
relevancia recientemente, dado su vinculación con el origen de las tortugas (e.g. Reisz
& Laurin, 1995).
Muchos de sus integrantes desarrollaron ventanas temporales.

10
Cuadro que muestra un número de taxones representativo dentro de parareptilia,
resaltando la predominancia de formas con ventanas temporales (flechas grises gruesas)
sobre las que nunca las desarrollan (flechas negras finas).
Milleretidae
Los millerettidos son amniotas que vivieron durante el Pérmico fundamentalmente en
Africa. Fueron formas pequeñas e insectívoras y algunos taxones desarrollaron ventanas
temporales.
Los millerétidos presentan vértebras dorsales con morfología del arco neural
comprimido, en forma similar a la que prepondera en los synapsidos y los diápsidos.

11
A B C Vértebras dorsales de procolophonoideos hallados en la Formación Buena Vista del Permo-Triásico de
Uruguay, poseyendo arcos neurales ensanchados (Ay B) y vértebra dorsal de un Dimetrodon con arco
neural comprimido ©. Las primeras son típicas de los parareptiles y los diadectomorfos y las segundas
caracterizan a la mayoría de los synapsidos y diápsidos.
Pareiasauridae
Pareiasaurus Elginia
Los pareiasauridos son un grupo de pareptiles de complexión generalmente robusta,
herbívoros y mayormente de gran tamaño. Nunca desarrollaron ventanas temporales.
Las vértebras dorsales tienen arcos neurales muy ensanchados lateralmente (proporción,
2 a 1, en vista dorsal). Sus restos se encuentran en rocas del Pérmico Medio alto y
Pérmico tardío de Europa (particularmente Rusia e Inglaterra), Africa y América del Sur
(Brasil). Son importantes, ya que algunos autores (e.g. Lee, 1996) los han considerado
ancestros de las tortugas.
Testudines
Proganochelys, tortuga primitiva del Triásico Tardío de
Alemania.

12
Las tortugas surgen en el Triásico Tardío, su origen ha sido relacionado con al menos
dos grupos de parareptiles, los pareisáuridos y los procolophonidos. Los primeros
fueron propuestos como ancestros por presentar osteodermos dorsales asociados al
esqueleto axial, que pueden ser homologados con las placas del caparazón dorsal en las
tortugas. No obstante no explican el origen del plastrón.
Otra hipótesis plantea un origen a partr de los procolophonoidea, asumiendo varios
caracteres osteomorfológicos similares entre los dos grupos y la reducción dentaria que
experimentan los procolophonidos en su evolución, que puede detectarse a través de sus
últimos representantes en el Triásico Tardío que presentan menos de cuatro dientes
marginales y el paladar está totalmente desprovisto de ellos. Es la hipótesis que ha sido
más aceptada en la última década, aunque existen dos hipótesis alternativas:
1- Las tortugas son diápsidos muy aberrantes que han perdido sus ventanas y
algunas de las características más sobresalientes del grupo. Esta hipótesis
incluye un origen a partir de ancestros marinos (considerando ese hábitat para
Odontochelys semitestacea, una forma recientemente hallada en el Noriano
(Triásico Tardío) de China.
Odontochelys, una forma relacionada a ambientes marinos fue hallada en el TriásicoTardío pero en
depósitos al menos 5 millones de años más antiguos que los que brindaron las tortugas más antiguas
conocidas (e.g. Proganochelys). Odontochelys es considerada muy cercana al origen de las tortugas, y
según los autores que la describieron presenta sólo plastrón y el caparazón dorsal está muy reducido o

13
ausente. Esta hipótesis sugiere un origen marino para las tortugas, a pesar que las formas más antiguas
son claramente terrestres. La presencia de plastrón en Odontochelys sugiere que esa estructura se formó
antes que el caparazón dorsal, pero algunas críticas ha tenido esta sugerencia, dado que algunos autores
creen que la ausencia de caparazón es en realidad una adaptación al medio marino.
2- Las tortugas derivan de ancestros terrestres y el caparazón surge, como una
estructura de adaptación a vivir en ese medio, a través de un contacto (al inicio
cartilaginoso) entre las costillas dorsales y el esternón y probablemente las
costillas ventrales. Esta hipótesis involucra a un probable parareptil,
Eunotosaurus africanus más relacionado con los millerétidos que con los
procolophonoideos.
Hipótesis que involucra a Eunotosaurus africanus con el origen de las tortugas.
Filogenia de Amniota mostrando la posición de los parareptiles y de las tortugas como posibles
parareptiles o como probables diápsidos aberrantes.

14
LA LÍNEA DIAPSIDA
Los representantes más primitivos del linaje de vertebrados dominante hoy en día
A pesar que los diápsidos están representados desde el Carbonífero Tardío, no es sino
hasta el Triásico que se vuelven dominantes. Durante el Paleozoico, algunas pocas
formas son bien conocidas, tal es el caso de Petrolacosaurus y Araeoscelis del
Carbonífero Tardío y Pérmico Temprano de Norte América. Los diápsidos acuáticos,
asociados a ambientes marinos someros como los eosuchidos notosaurios y
pachypleurosaurios se vuelven dominantes hacia el final del Pérmico, así como los
Prolacertiformes y Younginiformes se diversifican en tierra.
Los diápsidos se caracterizan por presentar dos ventanas temporales a cada lado del
cráneo, pero algunas modificaciones surgen en la evolución del grupo que pueden
producir cráneos con una sola ventana (lagartos y serpientes), o sin ellas (aves).

15
Notosaurios y Plesiosaurios primitivos (Lepidosauromorpha) del Pérmico y el Triásico
Temprano.
Prolacertiformes, reptiles diápsidos del Pérmico y el Triásico Temprano. Vértebras de
estos reptiles han sido halladas en la Fm. Buena Vista de Uruguay de edad Permo-
Triásico. Dentro de estos grupos se encuentran los ancestros de los plesiosaurios,
reptiles que conquistan los mares a partir del Triásico y dominan durante todo el
Mesozoico.
Los Plesiosaurios estaban bien adaptados al medio

16
marino, y presentaban hiperfalangia e hiperdactilia para transformar manos y pies en
paletas natatorias. El cuerpo se hizo progresivamente hidrodinámico con cabezas
pequeñas y cuerpos alargados.
Otro grupo de diápsidos marinos dominantes en el Mesozoico son los Ictiosaurios, los
cuales fueron los reptiles más adaptados al medio marino.
Los ictiosaurios son los reptiles vivíparos más antiguos conocidos y sus fósiles
documentan muy bien esta estrategia reproductiva.
LOS ARCHOSAURIOS
Los archosaurios son diápsidos que entre otras cosas, se caracterizan por poseer una
fenestra anteorbitaria entre la órbita y la narina. Esta fenestra ha recibido la atención de
los especialistas, las cuales la han asociado con la presencia de una glándula de la sal,
con funciones neumáticas o asociadas con la ventilación y enfriamiento del aire que
entra a la cavidad nasal. Ellos derivan de un stock basal conoido como Tecodontos, los
cuales agrupan a aquellos reptiles que presentan dientes posicionados en un alvéolo.
Tecodontos del Triásico Superior

17
En el medio aéreo los arcosaurios fueron dominantes durante el Mesozoico a través de
los Pterosaurios. Estos reptiles presentaban una membrana alar que se sostenía mediante
un alargamiento del dedo IV de la mano.
Ranforincoideos: Pterosaurios primitivos con cola larga y cráneos menos especializados
a la función del vuelo.
Pterodactiloideos: Pterosaurios derivados con cola reducida y cráneos especializados
Algunos pterosaurios alcanzaron dimensiones gigantescas como se puede observar en la
figura en comparación con el tamaño de un hombre.

18
LA EVOLUCIÓN DE LOS COCODRILOS
Los primeros cocodrilos son conocidos desde el Triásico más Tardío y comienzos del
Jurásico. Las formas más primitivas estaban más adaptadas a la carrera, presentaban
miembros más largos y gráciles.
Triásico Jurásico Cretácico
En Uruguay, los cocodrilos están representados en la Fm. Tacuarembó del Jurásico-
Cretácico (Meridiosaurus) y en la Fm. Guichón (Uruguaysuchus) de edad Cretácico
Tardío.
Uruguaysuchus aznarezi
LOS DINOSAURIOS Y EL ORIGEN DE LAS AVES
En tierra, los diápsidos dominantes en el Mesozoico fueron los dinosaurios a partir del
Triásico Tardío y hasta el Cretácico Tardío en que se extinguen junto a los pterosaurios
y los reptiles dominantes en el ámbito marino.
También surgen del stock basal Tecodonte y Euparkeria y Lagosuchus son
considerados ancestros probables:
Podemos reconocer dos grandes grupos, Saurischia y Ornitischia:

19
Los Saurisquios incluyen formas cuadrúpedas y herbívoras (Sauropodomorfos) y
otras carnívoras, en su gran mayoría bípedas (Terópodos).
La pelvis presenta tres radios (ileon dorsal, isquion posterior y pubis anterior)
(pelvis trirradiada).
Pelvis saurisquia.
Los ornitisquios son todos herbívoros, incluyen formas bípedas y cuadrúpedas y su
pelvis presenta cuatro radios (ileon dorsal, isquion posterior y pubis con una rama
posterior y otra anterior que acompaña al pubis).
Pelvis tetrarradiada.
En Uruguay los dinosaurios están representados en las formaciones Tacuarembó del
Jurásico Tardío-Cretácico (dientes), Guichón (dientes, huesos y huevos) y Mercedes
(huesos fragmentarios y huevos), las dos últimas de edad Cretácico Tardío.

20
Dientes de ceratosaurios de la Fm. Tacuarembó (Jurásico-Cretácico de Uruguay).
Dientes de terópodos de la Fm. Tacuarembó (Jurásico-Cretácico de Uruguay).
Huevos de dinosaurios probablemente titanosaurios de la Fm. Mercedes (Cretácico Tardío) de Uruguay.

21
EL ORIGEN DE LAS AVES
Las aves surgen en el Jurásico Medio y derivan de dinosaurios saurisquios (por lo tanto
la pelvis de las aves es trirradiada).
El ave más antigua conocida es Archaeopteryx, un taxón solo conocido del Jurásico de
Alemania.
Archaeopteryx es conocida mediante el registro de 9 ejemplares hallados en una cantera
de explotación de calizas litográficas de la ciudad de Solenhofen, Alemania. Estos
depósitos son interpretados como lagos profundos de fondos anóxicos donde las aves
fueron preservadas. Esta característica permitió la preservación de estructuras tan
delicadas como las plumas.

22
Archaeopteryx litographica
Archaeopteryx presentaba más caracteres reptilianos que avianos. Se supone que las
plumas, derivadas de las escamas de los reptiles, fueron adquiridas originalmente como
órganos de regulación térmica. Posteriormente fueron utilizadas para el desarrollo del
vuelo.
Entre las diferencias más notorias entre
Archaeopteryx y las aves actuales se destaca la larga cola CON VÉRTEBRAS.

23
Los dinosaurios de la línea aviana eran formas gráciles, bípedas y ágiles corredoras.
Estas formas desarrollaron plumas y sus esqueletos fueron hallados en depósitos del
Cretácico de China.
Protarchaeopteryx

24
Sinosauropteryx
Así, se pudo reconstruir una línea evolutiva que deriva en las aves como sus
representantes más derivados y con representación en las faunas actuales.


![343o aos amniotas [Modo de Compatibilidade])colegiomichel.com.br/wp-content/uploads/2014/03/...aos-amniotas.pdf · Padrõesevolutivos • Surgimentodosamniotas Filo Chordata (Cordados)](https://img.pdfslide.net/doc/110x75/5bf4460009d3f2f85f8c80f2/343o-aos-amniotas-modo-de-compatibilidade-padroesevolutivos-surgimentodosamniotas.jpg)

![SP 10 Modelos [Modo de compatibilidad]cuencas.fcien.edu.uy/cursos/materiales/SP 10 Modelos.pdf · PetroMod Version 10 Service ... Basin modeling (also spelled basin modelling) is](https://img.pdfslide.net/doc/110x75/5b5d5ec77f8b9ad2198e3056/sp-10-modelos-modo-de-compatibilidad-10-modelospdf-petromod-version-10-service.jpg)
























