Embed Size (px)
Citation preview

UNIVERSIDADE DO VALE DO ITAJAÍ
VANESSA DUARTE CLAUDINO
ESTUDO FITOQUÍMICO E AVALIAÇÃO DO POTENCIAL
ANTIPARASITÁRIO E ANTITUMORAL DO EXTRATO
CLOROFÓRMICO E DE MOLÉCULAS BIOATIVAS DE Drimys
brasiliensis Miers (Winteraceae)
Itajaí, 2011

UNIVERSIDADE DO VALE DO ITAJAÍ
PROGRAMA DE MESTRADO ACADÊMICO EM CIÊNCIAS
FARMACÊUTICAS
ÁREA DE CONCENTRAÇÃO EM PRODUTOS NATURAIS E
SUBSTÂNCIAS SINTÉTICAS BIOATIVAS
VANESSA DUARTE CLAUDINO
ESTUDO FITOQUÍMICO E AVALIAÇÃO DO POTENCIAL
ANTIPARASITÁRIO E ANTITUMORAL DO EXTRATO
CLOROFÓRMICO E DE MOLÉCULAS BIOATIVAS DE Drimys
brasiliensis Miers (Winteraceae)
Dissertação submetida à Universidade do Vale do Itajaí como parte dos requisitos para a obtenção do grau de Mestre em Ciências Farmacêuticas. Orientadora: Profª. Dra. Angela Malheiros Co-orientador: Prof. Dr. Valdir Cechinel Filho
Itajaí, Julho de 2011.

ESTUDO FITOQUÍMICO E AVALIAÇÃO DO POTENCIAL ANTIPARASITÁRIO E ANTITUMORAL DO EXTRATO
CLOROFÓRMICO E DE MOLÉCULAS BIOATIVAS DE Drimys brasiliensis Miers (Winteraceae)
VANESSA DUARTE CLAUDINO “Esta Dissertação foi julgada adequada para obtenção do Título de Mestre em Ciências Farmacêuticas, Área de Concentração Produtos Naturais e Substâncias Bioativas e aprovada em sua forma final pelo Programa de Mestrado em Ciências Farmacêuticas da Universidade do Vale do Itajaí.”
____________________________________ Angela Malheiros, Dra.
Orientadora
____________________________________
Valdir Cechinel Filho, Dr. Co-Orientador
__________________________________________________________ Tânia Mari Bellé Bresolin, Dra.
Coordenadora do Programa Mestrado em Ciências Farmacêuticas
Apresentado perante a Banca Examinadora composta pelos Professores:
______________________________________ Profa. Dra. Angela Malheiros (UNIVALI)
Presidente e Orientadora
______________________________________ Prof. Dr. Valdir Cechinel Filho (UNIVALI)
Co-orientador
______________________________________ Prof. Dr. Rivaldo Niero (UNIVALI)
Membro Interno
______________________________________ Prof. Dra. Rosi Zanoni da Silva (UEPG)
Membro Externo
Itajaí (SC), Julho de 2011

3
Dedico este trabalho aos meus pais que são o meu apoio e meu alicerce. À minha avó querida Malvina Filomena Duarte (in memorium) minha grande amiga e confidente.

4
AGRADECIMENTOS
Agradeço a Deus, por estar sempre presente e nas horas mais
difíceis ter me ajudado a escolher o caminho certo.
Agradeço aos meus familiares, em especial minha mãe Yvelise,
meu pai José por serem à base de meus princípios e de minha
formação. Um agradecimento em especial a minha avó Malvina (in
memorium) pelo apoio, dedicação, amizade, carinho, compreensão.
Aos orientadores, Profa. Dra. Angela Malheiros e Prof. Dr.
Valdir Cechinel Filho, agradeço pelo apoio, incentivo e pela
orientação científica, sem eles este trabalho não poderia ser
realizado.
Aos professores da pré-banca, Prof. Dr. Clóvis Antônio
Rodrigues e Prof. Dr. Rivaldo Niero, agradeço pelo apoio, incentivo e
pelas dicas para a melhoria desta pesquisa científica, tornando-a
melhor e mais elaborada.
Aos professores do PMCF da UNIVALI pelo conhecimento
passado até nós e pela amizade cultivada no decorrer do mestrado.
Ao Prof Dr. Franco Delle Monache do Centro Chimica
Recettori, Consiglio Nazionale delle Ricerche, Roma – Itália pelo
auxílio na identificação das substâncias isoladas.
Ao Dr. Alberto Giménez, Dr. Efraín Salamanca, Dra. Graice
Ruiz, do Instituto de Investigaciones Fármaco Bioquímicas –
IIFB/UMSA/La Paz – Bolívia pelos ensaios com Leishmannia e
Plasmodium.
Ao Gilberto Carlos Franchi Júnior e Alexandre Eduardo Nowill
do Centro Integrado de Pesquisas Oncohematológicas da Infância –
CIPOI/UNICAMP/SP pelos ensaios de atividade antitumoral.

5
Aos colaboradores que auxiliaram de forma direta ou
indiretamente neste trabalho, em especial os colegas do Laboratório
de Fitoquímica, dos Laboratórios de Farmacologia in vitro e in vivo,
aos Pedros por terem ajudado nas análises e aos amigos cultivados ao
longo dessa jornada: Joel, Maggie, Sheila, Ingrid, Bruna, Mariana,
Simone, Marcelo, Kelly, Jéssica, Sabrina, Paola, Mariel, Mirela,
Zhelmy, Marta, Lígia, Delair, e muitos outros.

6
“Não tenho um dom especial – Sou apenas apaixonadamente curioso.”
(Albert Einstein 1879 - 1955)

7
ESTUDO FITOQUÍMICO E AVALIAÇÃO DO POTENCIAL
ANTIPARASITÁRIO E ANTITUMORAL DO EXTRATO CLOROFÓRMICO E
DE SUBSTANCIAS ISOLADAS DE Drimys brasiliensis Miers
(Winteraceae)
VANESSA DUARTE CLAUDINO
Julho / 2011
Orientadora: Angela Malheiros, Dra. Co-orientador: Valdir Cechinel Filho, Dr. Área de Concentração: Produtos Naturais e Substâncias Sintéticas Bioativas. Número de Páginas: 132 Há uma crescente necessidade de estudos com plantas medicinais para o tratamento de diversas doenças. Estudos realizados com a Drimys brasiliensis descrevem-na como possuidora de diversas propriedades medicinais, entre elas no tratamento do câncer. Este trabalho avaliou as ações citotóxicas e antiparasitárias dos diferentes extratos das cascas, caules e folhas bem como as substâncias isoladas das cascas de D. brasiliensis. O extrato foi fracionado por meio de cromatografia em coluna, utilizando como fase estacionária sílica gel e fase móvel hexano, acetato de etila e metanol. Foram isolados os sesquiterpenos drimanos poligodial, 1-β-O-p-metoxi-E-cinamil-isodrimeninol, 1-β-O-p-metoxi-E-cinamil-5α-hidroxi-isodrimeninol, 1-β-(p-metoxicinamil)-poligodial, drimanial, 1-β-(p-cumaroiloxi)-poligodial, o aromadendrano espatulenol, a lignana (-) hinoquinina, e outra em fase de identificação, além de ácidos graxos insaturados e um glicerídeo. As substâncias foram identificadas por ressonância magnética nuclear de 1H e 13C e suas estruturas foram confirmadas por comparação com dados de literatura. Os extratos das cascas, caules e folhas bem como as substâncias isoladas foram avaliadas em células tumorogênicas de leucemia mielóide (K562), linfóide aguda B (Nalm6), adenocarcinoma mamário humano (MCF-7) e melanoma murino (B16F10) “in vitro” nas concentrações entre 0,0001 e 1000 µg/mL. O extrato clorofórmico das cascas foi o mais ativo para todos os modelos celulares testados, com CI50 entre 4 e 22 µg/mL. Nos mesmos modelos celulares foram testados quatro drimanos e um aromadendrano. Os drimanos foram seletivos para algumas células, com destaque para o 1-β-(p-metoxicinamil)-poligodial que foi o mais ativo para todas as linhagens testadas. Os extratos e as substâncias isoladas foram avaliados em duas formas promastigotas de Leishmania e uma forma trofozoíta de Plasmodium. O extrato também foi o mais ativo. Os drimanos apresentaram resultados bastante satisfatórios com CI50 entre 1 a 42 µM e dentre eles, o 1-β-(p-cumaroiloxi)-poligodial foi o mais ativo, com CI50 entre 1 a 6 µM. Os resultados indicam que os drimanos presentes nas cascas são os responsáveis pelas atividades atribuídas a D. brasiliensis, confirmando seu uso popular no tratamento de diversas patologias que acometem o homem e gerando perspectivas de futuros aprimoramentos como medicamento para o tratamento das patologias relacionadas. Palavras-chave: Drimys brasiliensis, sesquiterpenos drimanos, atividade citotóxica, atividade antiparasitária.

8
ABSTRACT
PHYTOCHEMICAL ANALISYS AND EVALUATION OF THE
ANTIPARASITIC AND ANTITUMOR POTENTIAL OF CHLOROFORM
EXTRACT AND COMPOUNDS ISOLATED FROM Drimys brasiliensis
MIERS (Winteraceae)
Vanessa Duarte Claudino (Tese)
July / 2011
Supervisor: Angela Malheiros, Dra. Co-supervisor: Valdir Cechinel Filho, Dr. Area of Concentration: Natural products and synthetic bioactive substances. Number of Pages: 132 There is a growing need for studies involving medicinal plants for the treatment of various diseases. Studies conducted with Drimys brasiliensis describe it as having various medicinal properties, making it potentially useful in the treatment of cancer. The study presented here evaluates the cytotoxicity and antiparasitic activities of different extracts of stem bark, branches and leaves, as well as compounds isolated from the stem bark of D. brasiliensis. The extract was fractionated by column chromatography using silica gel as stationary phase and hexane, ethyl acetate and methanol as mobile phase. The sesquiterpene drimanes polygodial, 1-β-O-p-methoxy-E-cinnamoyl-isodrimeninol, 1-β-O-p-methoxy-E-cinnamoyl-5α-hydroxy-isodrimeninol, 1-β-(p-methoxycinnamoyl)-polygodial, drimanial and 1-β-(p-coumaroiloxy)-polygodial, the aromadendrane spathulenol, the lignan (-) hinokinin, and another still being identified, were isolated, as well as unsaturated fatty acids and a glyceride. The substances were identified by nuclear magnetic resonance of 1H and 13C, and their structures were confirmed by comparison with the data in the literature. The extracts of the stem bark, branches and leaves, as well as the isolated compounds, were evaluated in vitro in tumorigenic cells of myeloid leukemia (K562), acute lymphoblastic B (Nalm6), mammary adenocarcinome (MCF-7) and murine melanome (B16F10) at concentrations of between 0.0001 and 1.000 µg/mL. The chloroform extract of the stem bark was the most active for all the cell models tested, with IC50 of between 4 and 22 µg/mL. In the same cell models, four drimanes and one aromadendrane were also tested. The drimanes were selective for some cells, especially 1-β-(p-methoxycinnamyl)-poligodial, which was the most active for all strains tested. The extracts and isolated compounds were evaluated in two promastigote forms of Leishmmania and a trophozoite form of Plasmodium. The drimanes showed satisfactory results, with IC50 of between 1 and 42 µM and of these, 1-β-(p-cumaroiloxi)-poligodial was the most active, with IC50 of between 1 and 6 µM. The results indicate that the drimanes present in the stem bark are responsible for the activities attributed to D. brasiliensis, confirming its popular use in the treatment of various diseases affecting humans, and generating prospects for future improvements as a medicine for the treatment of related pathologies. Key words: Drimys brasiliensis, sesquiterpenes drimanes, cytotoxicity activity, antiparasitary activity.

9
LISTA DE FIGURAS
Figura 1: Gênese das células sanguíneas e linfócitos. (Adaptado de: INCA, 2010). .......................................................................................................................... 28
Figura 2: Leishmania, (A) forma flagelada ou promastigota; (B) forma aflagelada ou amastigota (Adaptado de: BRASIL, 2007). ........................................................ 30
Figura 3: Ciclo biológico de Leishmania no vetor e no vertebrado. (Adaptado de: BRASIL, 2007)................................................................................................... 31
Figura 4: Ciclo de vida da malária no vetor e no vertebrado (Adaptado de: BRASIL, 2005). ................................................................................................................ 33
Figura 5: Drimys brasiliensis: aspecto geral da planta adulta. (Fonte: Autora). ....... 35 Figura 6: Fluxograma dos procedimentos cromatográficos do extrato clorofórmico
das cascas de D. brasiliensis. ........................................................................... 52 Figura 7: Metabolização do MTT em sais de formazan em células viáveis. ............ 53 Figura 8: Fluxograma dos procedimentos cromatográficos do extrato clorofórmico
de D. brasiliensis mostrando o isolamento do Espatulenol. .............................. 62 Figura 9: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBJ-70. ....................... 63 Figura 10: Ampliações do espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBJ-70:
a) de 7,5 a 3,5 ppm; b) de 2,5 a 1,5 ppm; c) de 1,4 a 0,4 ppm. ......................... 64 Figura 11: Espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBJ-70. ...................... 65 Figura 12: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBJ-70. .................... 65 Figura 13: Esquema para obtenção dos principais grupos de sesquiterpenos
gerados a partir do precursor farnesil pirofosfato (Adaptado de: DEWICK, 2009). ................................................................................................................ 67
Figura 14: Provável rota biossintética do sesquiterpeno Espatulenol. (Adaptado de CHAVES, et al., 2003). ...................................................................................... 68
Figura 15: Fluxograma dos procedimentos cromatográficos do extrato clorofórmico de D. brasiliensis mostrando o isolamento do Ácido Graxo Insaturado e do Glicerídeo. ......................................................................................................... 69
Figura 16: Ampliação do espectro de RMN de 1H do DBJ-100 (300 MHz, CDCl3/TMS) de 5,5 até 0,5 ppm. ....................................................................... 70
Figura 17: Espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBJ-100. .................... 71 Figura 18: a) Espectro de RMN 1H (300 MHz, CDCl3/TMS) do DBF-1; b) expansões
de 6,0 a 3,5 ppm; c) expansões de 2,4 a 0 ppm. ............................................. 72 Figura 19: a) Expansão do espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBF-1
de 120 a 14 ppm................................................................................................ 73 Figura 20: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBF-1. ..................... 73 Figura 21: Biossíntese dos lipídeos através do precursor acetil CoA (Adaptado de:
HORNSTRA, 2002; LENINGER; NELSON; COX, 2002). .................................. 75 Figura 22: Representação estrutural dos ácidos graxos ω-6 e ω-3 (Adaptado de:
HORNSTRA, 2002; LENINGER; NELSON; COX, 2002). .................................. 76 Figura 23: Fluxograma dos procedimentos cromatográficos do extrato clorofórmico
de D. brasiliensis mostrando o isolamento da (-) Hinoquinina e da lignana DBX-09. ..................................................................................................................... 78
Figura 24: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBK-29. ..................... 79 Figura 25: Espectro de RMN ¹3C (75 MHz, CDCl3/TMS) do DBK-29. ..................... 80 Figura 26: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBK-29. ................... 80 Figura 27: Biossíntese dos lignóides (Adaptado de: SIMÕES, et al., 2004). ........... 83 Figura 28: Rota biossintética provável da hinoquinina (Fonte: DEWICK, 2002). ..... 84 Figura 29: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBX-09. ..................... 85

10
Figura 30: Espectro de RMN ¹3C (75 MHz, CDCl3/TMS) do DBX-09. ..................... 86 Figura 31: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBX-09. ................... 86 Figura 32: Fluxograma dos procedimentos cromatográficos do extrato clorofórmico
de D. brasiliensis mostrando o isolamento dos sesquiterpenos drimanos Poligodial, 1-β-O-p-metoxi-E-cinamil-isodrimeninol, 1-β-O-p-metoxi-E-cinamil-5α-hidroxi-isodrimeninol, 1-β-(p-metoxicinamil)-poligodial, drimanial e 1-β-(p-cumaroiloxi)-poligodial, e o que está em fase de identificação o DBK-17. ........ 87
Figura 33: Espectro de RMN 1H (300 MHz, CDCl3/TMS) do DBI-11. ...................... 89 Figura 34: Espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBI-11. ....................... 89 Figura 35: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBI-11...................... 90 Figura 36: Rota biossintética provável do poligodial (Adaptado de: JANSEN e
GROOT, 2004). ................................................................................................. 92 Figura 37: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBK-17. ..................... 93 Figura 37: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBP-44. ..................... 94 Figura 38: Espectro de RMN ¹3C (75 MHz, CDCl3/TMS) do DBP-44. ..................... 94 Figura 39: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBP-44. ................... 95 Figura 40: Rota biossintética provável do 1-β-(p-metoxicinamil)-poligodial
(Adaptado de: JANSEN e GROOT, 2004). ........................................................ 96 Figura 41: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBP-11. ..................... 98 Figura 42: Espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBP-11. ..................... 98 Figura 43: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBP-11. ................... 99 Figura 43: Rota biossintética provável do 1-β-O-p-metoxi-E-cinamil-isodrimeninol.
(Adaptado de: JANSEN e GROOT, 2004). ........................................................ 99 Figura 44: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBV-25. ................... 101 Figura 45: Espectro de RMN ¹3C (75 MHz, CDCl3/TMS) do DBV-25. ................... 102 Figura 46: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBV-25. ................. 102 Figura 47: Rota biossintética provável do drimanial. (Adaptado de: JANSEN e
GROOT, 2004). ............................................................................................... 103 Figura 48: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBP-21. ................... 105 Figura 49: Espectro de RMN ¹3C (75 MHz, CDCl3/TMS) do DBP-21. ................... 106 Figura 50: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBP-21. ................. 106 Figura 51: Rota biossintética provável do 1-β-O-p-metoxi-E-cinamil-5α-hidroxi-
isodrimeninol. (Adaptado de: JANSEN e GROOT, 2004; ALOUCHE, et al., 2009). .............................................................................................................. 108
Figura 52: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBV-50. ................... 109 Figura 53: Espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBV-50. ................... 109 Figura 54: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBV-50. ................. 110 Figura 55: Rota biossintética provável do 1-β-(p-cumaroiloxi)-poligodial (Adaptado
de: JANSEN e GROOT, 2004). ....................................................................... 111 Figura 56: Biossíntese do pirofosfato de dimetilalila a partir do precursor ácido
mevalônico (Adaptado de: GEISSMAN; CROUT, 1969; MAGRI, 1999).......... 113 Figura 57: Biossíntese dos sesquiterpenos pela formação do núcleo germacreno
(Adaptado de: GEISSMAN; CROUT, 1969; MAGRI, 1999). ............................ 113 Figura 58: Rearranjo drimano do precursor germacreno (Adaptado de: JANSEN e
GROOT, 1991). ............................................................................................... 114

LISTA DE TABELAS
Tabela 1: Rendimentos das cascas, caules e folhas de D. brasiliensis Miers dos extratos CHCl3 e CH3OH em relação ao material vegetal utilizado. ............. 57
Tabela 2: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBJ-70 comparando com dados da literatura para o Espatulenol. ........................... 66
Tabela 3: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBK-29 comparando com os dados de literaturas para a Hinoquinina e para a (-) Hinoquinina. ................................................................................................. 81
Tabela 4: Valores de deslocamentos químicos () de RMN 1H e 13C para DBI-11 e dados da literatura para o Poligodial. ........................................................ 91
Tabela 5: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBP-44 comparando com dados da literatura para o 1-β-(p-metoxicinamil)-poligodial. ..................................................................................................................... 96
Tabela 6: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBP-11 comparando com dados da literatura para o 1-β-(p-metoxicinamil)-poligodial e o 1-β-O-p-metoxi-E-cinamil-6α-hidroxi-isodrimeninol. ............................. 100
Tabela 7: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBV-25 comparando com dados da literatura para o Drimanial. ............................. 103
Tabela 8: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBP-21 comparando com dados da literatura para o drimanial e o 1-β-O-p-metoxi-E-cinamil-6α-hidroxi-isodrimeninol. ................................................................ 107
Tabela 9: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBV-50 e dados da literatura para o 1-β-(p-cumaroiloxi)-poligodial. ....................... 111
Tabela 10: Avaliação da viabilidade celular de K562, Nalm6, MCF-7 e B16F10 em extratos brutos. ..................................................................................... 115
Tabela 11: Avaliação da viabilidade celular de K562, Nalm6, MCF-7 e B16F10 das substâncias isoladas. ........................................................................... 117
Tabela 12: Atividade leishmanicida dos extratos de Drimys brasiliensis in vitro nas formas promastigotas de Leishmania amazonensis e L. brasiliensis. . 119
Tabela 13: Atividade leishmanicida das substâncias isoladas das cascas de Drimys brasiliensis in vitro nas formas promastigotas de Leishmania amazonensis e L. brasiliensis. .................................................................... 120
Tabela 14: Atividade plasmodicida dos extratos, substâncias isoladas das cascas de Drimys brasiliensis in vitro nas formas trofozoítas de Plasmodium falciparum. .................................................................................................. 122

12
LISTA DE ABREVIATURAS E SIGLAS
AA – Ácido Araquidônico.
AcOEt – Acetato de Etila.
ACP – Proteína Carreadora de Acila.
AITC – Alil Isotiocianato.
ALA – Ácido Alfa-Linolênico.
AO – Ácido Oléico.
AS – Anisaldeído Sulfúrico.
ATP – Trifosfato de Adenosina.
ATPase – Enzima que hidrolisa o ATP.
α-TOH – α-Tocoferol.
BCRJ – Banco de Células da Universidade Federal do Rio de Janeiro.
BK – Bradicinina.
B16F10 – Melanoma Murino.
CC – Cromatografia em Coluna.
CCD – Cromatografia em Camada Delgada.
CCDP – Cromatografia em Camada Delgada Preparativa.
CF – Cromatografia Flash.
CG – Cromatografia Gasosa.
CGAR – Cromatografia Gasosa de Alta Resolução.
CG/MS – Cromatografia Gasosa Acoplada a Espectroscopia de Massas.
CHO – Célula Ovariana de Hamster.
CIM – Concentração Inibitória Mínima.
CIPOI – Centro Integrado de Pesquisa Oncológica da Infância.
CI50 – Concentração citotóxica ou inibitória da viabilidade em 50% das células.
CLAE – Cromatografia Líquida de Alta Eficiência.
CLC – Cromatografia Líquida Clássica.
δ – Deslocamento Químico.
DBF-1 – Drimys brasiliensis Coluna F (coluna do extrato bruto) fração 9-21)
identificada como Glicerídeo.
DBI-11 – Drimys brasiliensis Coluna I (junção da coluna 4-8 da coluna G (26-30)
fração 11-18) identificada como Poligodial.

13
DBJ-70 - Drimys brasiliensis Coluna J (fração 26-33 da coluna H (4-21 da coluna
do extrato bruto) fração 70-71) identificada como Espatulenol.
DBJ-100 - Drimys brasiliensis Coluna J (fração 26-33 da coluna H (4-21 da coluna
do extrato bruto) fração 100-105) identificada como Ácido Graxo Insaturado.
DBK-11 – Drimys brasiliensis Coluna K (fração 31-38 da coluna do extrato bruto)
fração 8-11) identificada como Poligodial.
DBK-17 - Drimys brasiliensis Coluna K (fração 31-38 da coluna do extrato bruto)
fração 17-21) substância não identificada.
DBK-29 - Drimys brasiliensis Coluna K (fração 31-38 da coluna do extrato bruto)
fração 29-43) identificada como (-) Hinoquinina.
DBP-11 - Drimys brasiliensis Coluna P (fração 48-55 da coluna do extrato bruto)
fração 11-20) identificada como 1-β-O-p-metoxi-E-cinamil-isodrimeninol.
DBP-21 - Drimys brasiliensis Coluna P (fração 48-55 da coluna do extrato bruto)
fração 21-30) identificada como 1-β-O-p-metoxi-E-cinamil-5α-hidroxi-isodrimeninol.
DBP-44 - Drimys brasiliensis Coluna P (fração 48-55 da coluna do extrato bruto)
fração 44-47) identificada como 1-β-(p-metoxicinamil)-poligodial.
DBV-25 - Drimys brasiliensis Coluna V (fração 56-59 da coluna do extrato bruto)
fração 25-46) identificada como Drimanial.
DBV-50 - Drimys brasiliensis Coluna V (fração 56-59 da coluna do extrato bruto)
fração 50-55) identificada como 1-β-(p-cumaroiloxi)-poligodial.
DBX-04 - Drimys brasiliensis Coluna X (fração 24-33 da coluna O (39-45 da coluna
do extrato bruto) fração 4-5) identificada como (-) Hinoquinina.
DBX-09 - Drimys brasiliensis Coluna X (fração 24-33 da coluna O (39-45 da coluna
do extrato bruto) fração 9) substância não identificada.
DCM – Diclorometano.
d – Dubleto.
dd – Duplo Dupleto.
ddd – Dupleto de Dupleto de Dupleto.
dddd – Dupleto de Dupleto de Dupleto de Dupleto.
DEPT – Distorção por transferência de polarização.
DMC – Distúrbios Mieloproliferativos Crônicos.
DMEM – Dulbecco’s Modified Eagle’s Minimum Essential Medium.
DMSO – Dimetilsulfóxido.
DNA – Ácido Desoxirribonucléico.

14
EC50 – Concentração efetiva que causa 50% de inibição em relação ao controle.
ED50 – Concentração efetiva que causa 50% de inibição em relação ao fármaco.
EtOH – Etanol.
GRI – Glóbulos Vermelhos Infectados.
GRL – Glóbulos Vermelhos não-Infectados.
HBR – Herbário Barbosa Rodrigues.
HEK293 – Células Humanas Embrionárias de Rim.
HELA – Adenocarcinoma Epitelial de Cérvix Humano.
Hex – Hexano.
EM – Espectrometria de Massas.
HPV – Vírus Papiloma Humano.
INCA – Instituto Nacional de Câncer.
IP – Inositol Fosfato.
J – Constante de Acoplamento.
K562 – Leucemia Mielóide Crônica Humana.
LA – Ácido Linoléico.
LMA – Leucemia Mielóide Aguda.
LTA – Leishmaniose Tegumentar Americana.
L929 – Fibroblasto Murino.
m – Multipleto.
mAChR – Receptores Muscarínicos Agonistas de Acetilcolina Ativados.
MCF-7 – Michigan Cancer Fondation – 7; Adenocarcinoma Mamário Humano.
MTT – Sal de brometo de 3-(4,5 dimetil-2-tiazolil)-2,5-difenil-2H-tetrazolium.
M2903 – Leishmania brasiliensis.
NA – Ácido Nervônico.
NADPH – Nicotinamida Adenina Dinucleotídeo Phosfato.
Nalm6 – Leucemia Linfóide Aguda B.
NIQFAR – Núcleo de Investigações Químico-Farmacêuticas da Universidade do
Vale do Itajaí.
NO – Óxido Nítrico.
OMS – Organização Mundial de Saúde.
PAF – Fator de Agregação Plaquetária.
PE – Fosfatidiletanolamina.
PH8 – Leishmania amazonensis.

15
PLC – Fosfolipase C.
PPI – Fosfatidil Inositídeo.
PS – Fosfatidilserina.
PTX – Toxina Pertussis.
q – Quarteto.
Rf – Fator de Retenção.
RMN – Ressonância Magnética Nuclear.
RMN de 1H – Ressonância Magnética Nuclear de Hidrogênio.
RMN de 13C – Ressonância Magnética Nuclear de Carbono Treze.
ROS – Espécies Reativas de Oxigênio.
RPMI – Instituto Roswell Park Memorial.
s – Simpleto.
sl – Simpleto Largo.
SFB – Soro Fetal Bovino.
SH-SY5Y – Células de Neuroblastoma Humano.
SM – Síndrome Mielodisplásica.
SP – Substância P.
t – Tripleto.
TMS – Tetrametilsilano.
TRPA1 – Receptores Potenciais Vanilóides Transitórios 1.
TRPV1 – Receptores Potenciais Vanilóides 1.
Фi – Diâmetro Interno.
%Inh. – Percentual de Inibição.
53 RP75 – Leishmania donovani.

16
SUMÁRIO
1 INTRODUÇÃO ..................................................................................................... 18
2 OBJETIVOS ......................................................................................................... 20
2.1 Objetivo geral: ........................................................................................ 20
2.2 Objetivos específicos: ........................................................................... 20
3 REVISÃO DA LITERATURA ............................................................................... 21
3.1 Aspectos gerais sobre plantas medicinais ......................................... 21
3.2 Aspectos gerais sobre neoplasias....................................................... 23
3.3 Aspectos sobre doenças parasitárias ................................................. 29
3.3.1 Leishmaniose................................................................................................ 29
3.3.2 Malária ........................................................................................................... 32
3.4 Aspectos botânicos do gênero Drimys ............................................... 34
3.5 Aspectos químicos e farmacológicos do gênero Drimys ................. 36
3.6 Técnicas de fracionamento, isolamento e identificação de produtos
naturais ......................................................................................................... 45
4 MATERIAL E MÉTODOS .................................................................................... 48
4.1 Coleta do material vegetal .................................................................... 48
4.2 Materiais e reagentes ............................................................................ 48
4.3 Equipamentos ........................................................................................ 49
4.4 Obtenção dos extratos de D. brasiliensis ........................................... 49
4.5 Fracionamento do extrato CHCl3 das cascas de D. brasiliensis ...... 50
4.6 Atividade biológica ................................................................................ 53
4.6.1 Avaliação da citotoxicidade ........................................................................ 53
4.6.2 Avaliação da atividade leishmanicida ........................................................ 54
4.6.3 Avaliação da atividade plasmodicida ......................................................... 55
4.7 Análise estatística ................................................................................. 56
5 RESULTADOS E DISCUSSÃO ........................................................................... 57
5.1 Extração .................................................................................................. 57
5.2 Fracionamento do extrato CHCl3 das cascas de Drimys brasiliensis58
5.3 Identificação das substâncias isoladas de D. brasiliensis ................ 59

17
5.3.1 Identificação do Espatulenol ....................................................................... 61
5.3.2 Identificação do Ácido Graxo Insaturado ................................................... 69
5.3.3 Identificação do Glicerídeo .......................................................................... 71
5.3.4 Identificação da (-) Hinoquinina .................................................................. 77
5.3.5 Identificação do DBX-09 .............................................................................. 84
5.3.6 Identificação dos Sesquiterpenos Drimanos ............................................. 87
5.3.6.1 Identificação do Poligodial ....................................................................... 88
5.3.6.2 Identificação do DBK-17 ........................................................................... 92
5.3.6.3 Identificação do 1-β-(p-metoxicinamil)-poligodial .................................. 93
5.3.6.4 Identificação do 1-β-O-p-metoxi-E-cinamil-isodrimeninol ..................... 97
5,3.6.5 Identificação do Drimanial ...................................................................... 100
5.3.6.6 Identificação do 1-β-O-p-metoxi-E-cinamil-5α-hidroxi-isodrimeninol. 104
5.3.6.7 Identificação do 1-β-(p-cumaroiloxi)-poligodial .................................... 108
5.4 Atividade biológica .............................................................................. 115
5.4.1 Avaliação da citotoxicidade ...................................................................... 115
5.4.2 Avaliação da atividade leishmanicida ...................................................... 119
5.4.3 Avaliação da atividade plasmodicida ....................................................... 121
6 CONCLUSÕES .................................................................................................. 123
REFERÊNCIAS ..................................................................................................... 124

18
1 INTRODUÇÃO
A utilização de plantas medicinais, para tratamento, cura e prevenção de
doenças, é uma das antigas formas de prática medicinal da humanidade. As
observações populares sobre o uso e a eficácia de plantas medicinais contribuem
de forma relevante para a divulgação das propriedades terapêuticas (BUTLER,
2008; KINGSTON, 2011).
Muitas pessoas utilizam plantas para o tratamento de várias doenças,
inclusive para o tratamento do câncer. Embora muitas das suposições quanto à
eficácia possam ser vistas com certo ceticismo, devido ao fato de o câncer, por ser
uma doença específica, muitos extratos ou substâncias isoladas de plantas vêm
sendo testados a fim de confirmar as suposições atribuídas às plantas como
possuidoras de atividades biológicas comprovando assim o seu uso (CRAGG;
NEWMAN, 2009). Para tanto se podemos citar os alcalóides Vincristina e
Vimblastina obtidos de Catharantus roseus, para linfomas e leucemia infantil, o
diterpeno taxol de Taxus brevifolia, para o câncer ovariano, mamário e pulmonar, a
camptotecina de Camptotheca acuminata, que atua no controle da proliferação
celular, entre outros que ainda se encontram em fase experimental (GURIB-FAKIM,
2006).
A busca por antiparasitários inovadores constitui outra área onde os
produtos naturais podem contribuir, pois existem poucos medicamentos no
mercado para o tratamento de diferentes doenças parasitárias (KYSER,
KIDERLEN, CROFT, 2003). Como exemplos de agentes antimaláricos pode-se
citar o sesquiterpeno lactona artemisinina de Artemisina annua e o alcalóide
quinina isolado de Chinchona succirubra. A anfotericina B e ivermectina são
antiparasitários importante isoladas de microorganismos como Streptomyces
(CRAGG; NEWMANN, 2007; WAGNER, 2009; BARREIRO; BOLZANI, 2009).
Considerando a necessidade presente em se descobrir novos e efetivos
agentes medicinais o gênero Drimys surge como uma fonte de princípios ativos.
Entre as indicações da medicina popular pode-se citar a sua utilização no
tratamento do câncer (FORSBY; WALUM, 1996; FORSBY, et al., 1998; SILVA,
2009). Esta planta vem sendo estudada há alguns anos por pesquisadores do
Núcleo de Investigações Químico Farmacêuticas da Universidade o Vale do Itajaí

19
(NIQFAR/UNIVALI) em colaboração com outros Grupos de Pesquisa. O potencial
terapêutico tem sido comprovado em diversos modelos experimentais e vários
artigos científicos foram publicados (MENDES, et al., 1998; 2000; CECHINEL-
FILHO, 1998; ANDRE, et al., 1999; MALHEIROS, 2001; CUNHA, et al., 2001;
SCHEIDT, et al., 2002; ANDRE, et al., 2004; MALHEIROS, et al., 2005; ANDRE, et
al., 2006; MUNÕZ-CONCHA, et al., 2007). Porém o que motivou a escolha da D.
brasiliensis foi á constatação em estudos preliminares da atividade em células
tumorogênicas Hela do extrato das cascas e dos sesquiterpenos drimanos,
drimanial e 1-β-(p-metoxiciamil)-poligodial (SILVA, 2009). Devido aos resultados
promissores, o nosso trabalho propõe dar continuidade para esclarecer qual ou
quais são os metabólitos responsáveis pelos efeitos apresentados, especialmente
em relação à possível atividade antileucêmica, bem como avaliar o potencial
antiparasitário desta espécie vegetal.

20
2 OBJETIVOS
2.1 Objetivo geral:
Estudar fitoquimicamente o extrato clorofórmico das cascas de D.
brasiliensis e as substâncias isoladas sobre linhagens de células tumorais K562,
Nalm6, MCF-7 e B16F10, bem como as suas atividades antiparasitárias sobre as
linhagens de Leishmania amazonensis, Leishmania brasiliensis e Plasmodium
falciparum.
2.2 Objetivos específicos:
Isolar e identificar as substâncias majoritárias presentes no extrato
clorofórmico das cascas;
Definir a concentração inibitória mínima 50 (CI50) dos extratos clorofórmico e
metanólico de cascas, caules e folhas e as substâncias isoladas em
modelos celulares de leucemias mielóide (K562) e linfóide aguda B (Nalm6),
adenocarcinoma mamário humano (MCF-7) e melanoma murino (B16F10);
Determinar a concentração inibitória mínima 50 (CI50) dos extratos e
substâncias sobre a atividade leishmanicida e antimalárica in vitro nas
formas promastigotas de L. amazonensis e L. brasiliensis e as formas
trofozoítas de P. falciparum.

21
3 REVISÃO DA LITERATURA
3.1 Aspectos gerais sobre plantas medicinais
O uso das plantas medicinais vem crescendo gradativamente ao longo dos
últimos anos. Apesar da grande evolução da medicina alopática a partir da
segunda metade do século XX, ainda existe obstáculos na utilização pelas
populações carentes, que vão desde o acesso aos centros de atendimento
hospitalar, à obtenção de exames e medicamentos. Estes motivos associados à
fácil obtenção e a grande tradição do uso de plantas medicinais contribuem para a
sua utilização pelas populações dos países em desenvolvimento (VERLI;
BARREIRO, 2005).
Para definir futuras pesquisas de plantas medicinais, deve-se enfatizar que,
das cerca de 2000 doenças agudas e crônicas registradas, somente 30 a 40% são
curáveis nos dias de hoje. O restante pode ser tratado somente sintomaticamente,
com maior ou menor efeito, e algumas doenças ainda não tem tratamento.
Portanto, existe uma grande necessidade de novos tratamentos, altamente
eficientes e orientados, para os quais se espera que a fitoterapia contribua
(WAGNER, 2009).
É expressivo o número de medicamentos disponíveis na terapêutica
moderna desenvolvida a partir de plantas medicinais. Os produtos naturais têm
exercido um importante papel ao longo dos anos para a descoberta de novos
fármacos, muitos deles são utilizados de forma pura ou associados a outras
substâncias naturais ou sintéticas, servindo com ponto de partida para novas
moléculas mais ativas e eficazes a partir de sua otimização estrutural (YUNES,
CECHINEL FILHO, 2007; 2009).
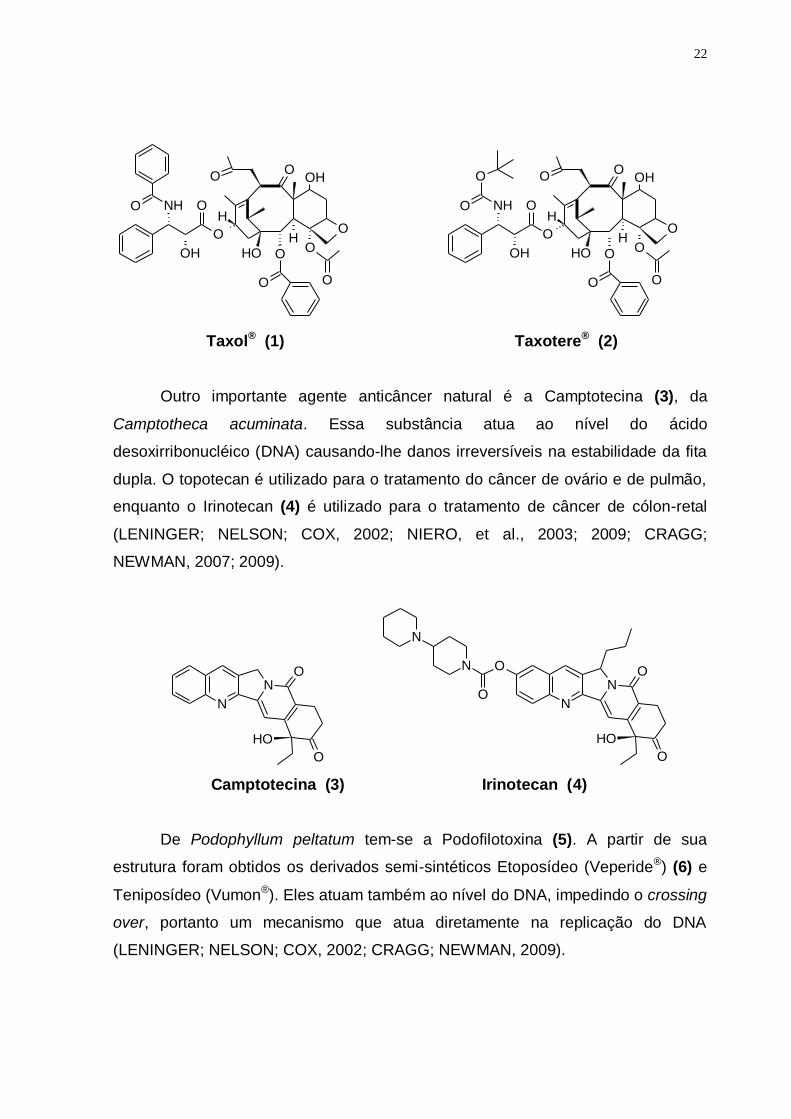
O diterpeno taxol ou Paclitaxel (Taxol®) (1) isolado da Taxus brevifolia vem
sendo utilizado para o tratamento de câncer ovariano, mamário, pulmonar, e
também tem demonstrado eficácia contra sarcoma de Kaposi. Além de ser utilizado
para o tratamento de esclerose múltipla, psoríase e artrite reumatóide. Um derivado
semissintético do taxol, denominado Docetaxol (Taxotere®) (2), é solúvel em água
e cerca de duas vezes mais potente que a substância natural (MILLER, et al.,
2008).
22
O
O
O
NHO
OH
O
OH
OH
OH O
O
HO
O
O
O
O
NHO
OH
O
O
OH
OH
OH O
O
HO
O
Taxol® (1) Taxotere® (2)
Outro importante agente anticâncer natural é a Camptotecina (3), da
Camptotheca acuminata. Essa substância atua ao nível do ácido
desoxirribonucléico (DNA) causando-lhe danos irreversíveis na estabilidade da fita
dupla. O topotecan é utilizado para o tratamento do câncer de ovário e de pulmão,
enquanto o Irinotecan (4) é utilizado para o tratamento de câncer de cólon-retal
(LENINGER; NELSON; COX, 2002; NIERO, et al., 2003; 2009; CRAGG;
NEWMAN, 2007; 2009).
N
N
O
OH
O
N
N
O
OH
OO
O
N
N
Camptotecina (3) Irinotecan (4)
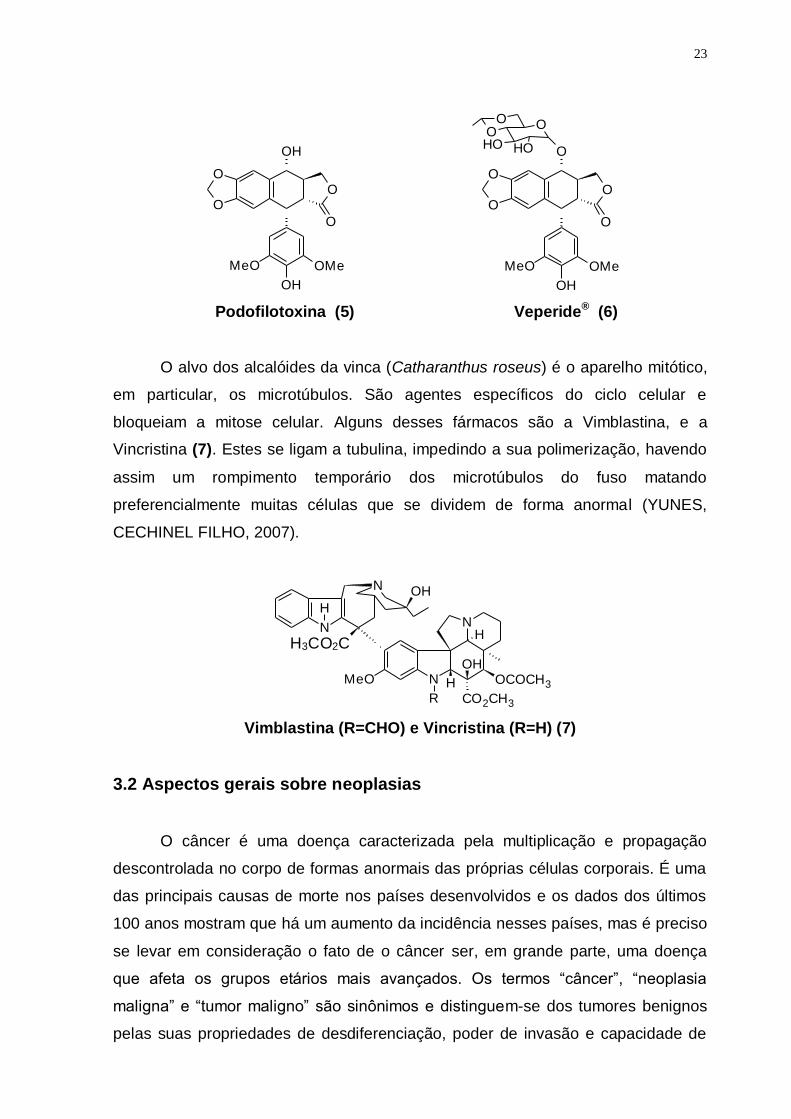
De Podophyllum peltatum tem-se a Podofilotoxina (5). A partir de sua
estrutura foram obtidos os derivados semi-sintéticos Etoposídeo (Veperide®) (6) e
Teniposídeo (Vumon®). Eles atuam também ao nível do DNA, impedindo o crossing
over, portanto um mecanismo que atua diretamente na replicação do DNA
(LENINGER; NELSON; COX, 2002; CRAGG; NEWMAN, 2009).
23
O
OO
O
OH
OH
OMeMeO
O
OO
O
O
OH
OMeMeO
OO
O
OHOH
Podofilotoxina (5) Veperide® (6)
O alvo dos alcalóides da vinca (Catharanthus roseus) é o aparelho mitótico,
em particular, os microtúbulos. São agentes específicos do ciclo celular e
bloqueiam a mitose celular. Alguns desses fármacos são a Vimblastina, e a
Vincristina (7). Estes se ligam a tubulina, impedindo a sua polimerização, havendo
assim um rompimento temporário dos microtúbulos do fuso matando
preferencialmente muitas células que se dividem de forma anormal (YUNES,
CECHINEL FILHO, 2007).
N
N OH
H3CO2C
MeO N
R
N
CO2CH3
OHOCOCH3
H
H
H
Vimblastina (R=CHO) e Vincristina (R=H) (7)
3.2 Aspectos gerais sobre neoplasias
O câncer é uma doença caracterizada pela multiplicação e propagação
descontrolada no corpo de formas anormais das próprias células corporais. É uma
das principais causas de morte nos países desenvolvidos e os dados dos últimos
100 anos mostram que há um aumento da incidência nesses países, mas é preciso
se levar em consideração o fato de o câncer ser, em grande parte, uma doença
que afeta os grupos etários mais avançados. Os termos “câncer”, “neoplasia
maligna” e “tumor maligno” são sinônimos e distinguem-se dos tumores benignos
pelas suas propriedades de desdiferenciação, poder de invasão e capacidade de

24
metastatizar-se (isto é, disseminar-se para outras partes do corpo) (ROBBINS, et
al., 2008).
As neoplasias são caracterizadas como um aumento descontrolado da
proliferação celular. As neoplasias em geral ocorrem paralelamente ao aumento do
crescimento e perda de diferenciação celular. Assim as células neoplásicas perdem
as suas características originais de diferenciação e se tornam atípicas. Células em
condições normais dependem de atuação coordenada de vários pontos
regulatórios determinados por genes que orientam a resposta aos estímulos
internos e externos. Já a célula neoplásica sofre alteração justamente nestes
pontos que regulam a sua função, e dessa forma, adquirem autonomia de
crescimento se tornando independente dos estímulos externos (BOGLIOLO;
BRASILEIRO FILHO, 2004).
Segundo o INCA (Instituto Nacional de Câncer) foram esperados para 2010
489.270 casos; em mulheres 192.590 (51,3%) e entre os homens 182.830 (48,7%).
O mais incidente deve ser o câncer de pele não melanoma, com 113.850 casos.
Em seguida estão os tumores de próstata com 52.350; mama feminina com 49.240;
cólon e reto com 28.110 e pulmão com 27.630 casos (INCA, 2010).
O Adenocarcinoma Epitelial de Cérvix Humano (Hela) mais conhecido como
câncer do colo do útero, é uma doença de crescimento lento e silencioso. A
detecção precoce do câncer do colo do útero ou de lesões precursoras é
plenamente justificável, pois a cura pode chegar a 100% e em grande número de
vezes, a resolução ocorrerá em nível ambulatorial. Alimentação saudável, sexo
seguro e diminuição da exposição ao tabaco contribuem para se evitar o
aparecimento da doença. A principal forma de detecção é através do exame
preventivo (Papanicolau), com o objetivo de identificar aquelas que possam
apresentar a doença em fase muito inicial, quando o tratamento pode ser mais
eficaz. A detecção precoce, associada ao tratamento em estágios iniciais, tem
resultado em uma diminuição de até 90% dos casos de câncer invasivo (INCA,
2010).
Em muitos casos uma infecção decorrente do vírus papiloma humana (HPV)
desencadeia o desenvolvimento do câncer do colo do útero. Existem 13 tipos de
HPV reconhecidos como oncogênicos pela Agência internacional de pesquisa
Sobre o Câncer. Destes os mais comuns são o HPV16 e o HPV18 (WHO, 2008;
INCA, 2010). Ele está presente em mais de 90% dos casos de câncer cervical.

25
Esse contágio acontece pela falta de uso de preservativos, início precoce da
atividade sexual e múltiplos parceiros. Além disso, outros fatores também podem
desencadear o desenvolvimento deste câncer, como tabagismo e o uso prolongado
de pílulas anticoncepcionais. É o segundo tipo de neoplasia mais freqüente entre
mulheres, com aproximadamente 500 mil novos casos por ano no mundo, sendo
responsável pelo óbito de aproximadamente 230 mil mulheres por ano (ROBBINS,
et al., 2008).
Dados mostram que a incidência de câncer do colo do útero já é evidenciada
na faixa etária de 20 a 29 anos e o risco aumenta rapidamente até o seu pico
máximo que está entre 45 a 49 anos. Segundo o INCA (2010), o número de novos
casos de câncer do colo do útero esperado para o Brasil no ano de 2010 foi de
18.430, com um risco estimado de 18 casos a cada 100 mil mulheres.
Para o tratamento do câncer do colo do útero, os mais comuns são a cirurgia
e a radioterapia, salvo que, dependendo do estágio da doença, tamanho do tumor,
idade e desejo de ter filhos por parte da paciente, irá determinar qual o melhor
procedimento a ser realizado (ROBBINS, et al., 2008).
O Adenocarcinoma Mamário Humano (MCF-7), mais conhecido como
câncer de mama, é uma doença maligna onde há uma proliferação acelerada,
desordenada e sem controle das células dos tecidos da glândula mamária. O tumor
invade os tecidos vizinhos metastatizando para os órgãos distantes no corpo.
Como outros tumores malignos, o câncer de mama é conseqüência de alterações
na estrutura e função dos genes (ROBBINS, et al., 2008). Como principais
sintomas têm-se abaulamentos e retrações inclusive dos mamilos, aspecto
semelhante à casca de laranja, secreções no mamilo. O sintoma do câncer
palpável é o nódulo ou caroço no seio acompanhado ou não de dor mamária.
Também pode haver nódulos nas axilas (INCA, 2010).
Os cânceres de pele podem ser definidos como um crescimento anormal e
descontrolado das células que fazem parte da pele, podendo ser classificados em
câncer de pele não melanoma (carcinoma basocelular e epidermóide) e melanoma
(DE VITA; HELLMANN; ROSENBERG, 2005; FAURI, et al., 2010).
O câncer de pele não melanoma é o câncer mais frequente no Brasil
correspondendo a cerca de 25% de todos os tumores malignos registrados no país.
O número de casos novos estimados no Brasil no ano de 2010 para este tipo de
câncer foi de 53.410 entre homens e de 60.440 nas mulheres. De acordo com

26
estes dados o risco estimado foi de 56 novos casos a cada 100 mil homens e de 61
para cada 100 mil mulheres (INCA, 2010).
O câncer de pele conhecido como Melanoma Murino (B16F10) é a forma
mais agressiva de câncer de pele e que se expande mais rapidamente em termos
de incidência mundial. O melanoma é um tumor de origem neuroectodérmica
resultante da transformação dos melanócitos epidérmicos. Estes por sua vez, são
células encontradas ao longo da camada basal da epiderme cuja principal função é
a produção do pigmento melanina, importante para fotoproteção natural (DE VITA;
HELLMANN; ROSENBERG, 2005).
Apesar de o melanoma cutâneo representar apenas 4% de todos os tipos de
câncer de pele, este possui um grande potencial de metastatização, mesmo no
início de sua manifestação, razão pela qual se constitui numa das neoplasias mais
agressivas dentre as lesões cutâneas oriundas de processo tumoral, com elevado
índice de mortalidade, principalmente em casos de detecção tardia (INCA, 2010).
A Organização Mundial de Saúde (OMS) estima que anualmente ocorram
cerca de 132 mil novos casos deste câncer no mundo. Este aumento vem se
tornando expressivo nos últimos 40 anos, em principalmente, populações de pele
clara que apresentam uma maior susceptibilidade a este tipo de tumor. Quando
detectado no início é potencialmente curável, embora dados mostrem que quando
detectado tardiamente apresente letalidade elevada. A média mundial de sobrevida
é de 69% (WHO, 2008). Geralmente este tipo de tumor apresenta duas fases
distintas; na primeira a lesão se apresenta como um crescimento radial, plana e
pequena, e seu comportamento é bem menos agressivo. Na segunda fase, o
crescimento da lesão passa ser vertical, onde as células malignas se localizam
mais internamente na derme reticular podendo, inclusive invadir a camada de
tecido subcutâneo, apresentando um processo lesivo mais grave. Sua
agressividade é proporcional à sua capacidade de se infiltrar na derme na transição
de crescimento radial para crescimento vertical (ANGER, 2008).
A leucemia se caracteriza por uma proliferação neoplásica generalizada ou
acúmulo de células hematopoiéticas, com ou sem envolvimento periférico. Na
maioria dos casos, as células leucêmicas extravasam para o sangue, onde podem
ser vistas em grande número, podendo infiltrar-se no fígado, baço, linfonodos e
outros tecidos. A leucemia pode ser classificada de acordo com o grau de

27
maturidade das células afetadas; leucemia mielóide aguda e crônica e leucemia
linfóide aguda e crônica (HAMERSCHLAK, 2010).
A Leucemia Mielóide (K562), começa pela produção cancerosa de células
mielóides jovens na medula óssea e, a seguir, dissemina-se por todo o corpo, de
modo que os leucócitos são produzidos em muitos órgãos extra-medulares,
sobretudo nos linfonodos, no baço e no fígado (GUYTON; HALL, 2001).
O processo canceroso, algumas vezes, produz células parcialmente
diferenciadas, resultando em formas que poderiam ser denominadas leucemia
neutrofílica, eusinofílica, basofílica ou monocítica. Em geral, quanto mais
indiferenciada a célula mais aguda a leucemia, levando, freqüentemente, à morte
dentro de poucos meses se não for tratada. Com algumas das células mais
diferenciadas, o processo pode ser crônico, desenvolvendo-se algumas vezes,
lentamente no decorrer de 10 a 20 anos. As células leucêmicas, particularmente as
muito indiferenciadas, não costumam ser funcionais para proporcionar a proteção
habitual associada aos leucócitos contra infecção (RANG, et al., 2007).
Os tumores mielóides são tumores principalmente de adultos que se dividem
em três grupos: Leucemias Mielóides Agudas (LMA), Distúrbios Mieloproliferativos
Crônicos (DMC) e Síndrome Mielodisplásica (SM). Na LMA as células contém
diversas lesões genéticas que freqüentemente levam a expressão de fatores de
transcrição anormais que bloqueiam a diferenciação da célula mielóide.
Compreendem em células imaturas de linhagem mielóide (mieloblastos), que
substituem a medula óssea e suprimem a hematopoese normal (figura 1). No DMC
os tumores são indolentes nos quais a produção de células está inicialmente
aumentada, levando a contagens elevadas no sangue e hematopoese
extramedular. Na SM os grupos de tumores mielóides caracterizados por
hematopoese desordenada e ineficaz. A maior parte dos pacientes apresenta
pancitopenia, e muitos progridem para um estado da doença que é idêntico a LMA
(ROBBINS, et al., 2008).
As Leucemias Linfóides Agudas B (Nalm6) são causadas pela produção
cancerosa de células linfóides, que surgem, em geral, na medula ou nos blastos
linfóides chamados de linfonodos, ou outro tecido linfóide e, a seguir, disseminam-
se para outras áreas do corpo (GUYTON; HALL, 2001).
Já as neoplasias linfóides são classificadas com base na célula de origem e
no estágio de diferenciação. Os tipos mais comuns nas crianças são as leucemias
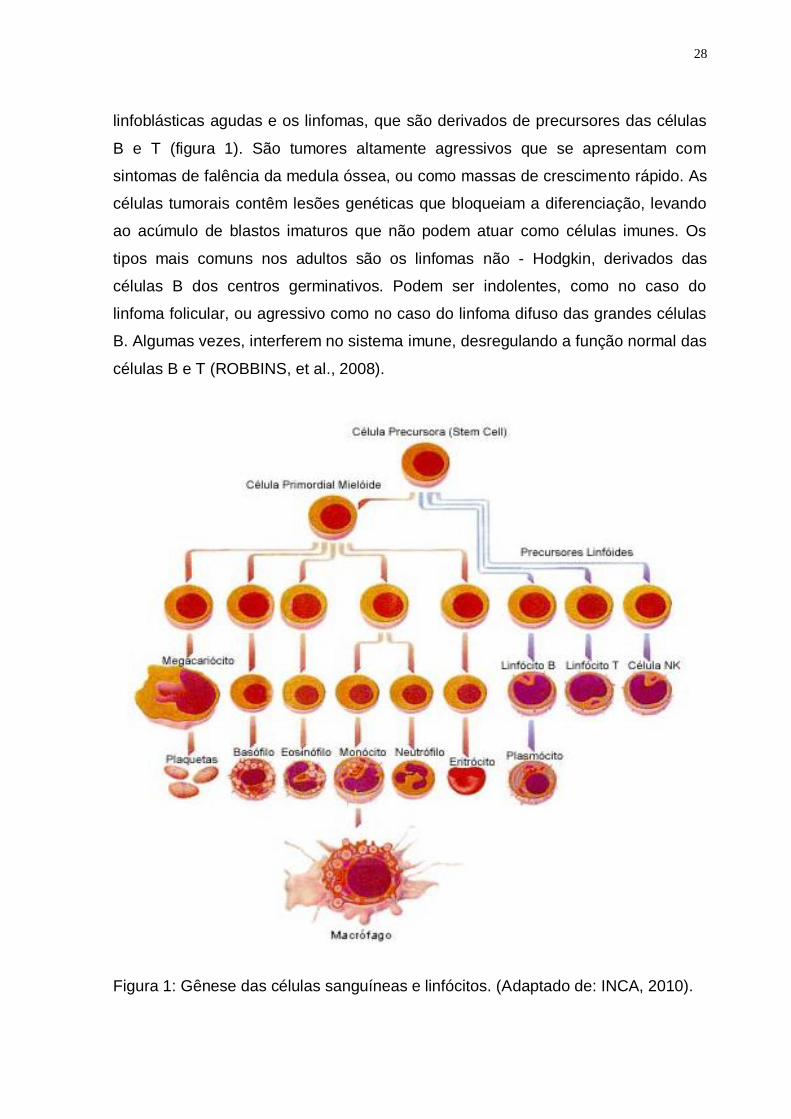
28
linfoblásticas agudas e os linfomas, que são derivados de precursores das células
B e T (figura 1). São tumores altamente agressivos que se apresentam com
sintomas de falência da medula óssea, ou como massas de crescimento rápido. As
células tumorais contêm lesões genéticas que bloqueiam a diferenciação, levando
ao acúmulo de blastos imaturos que não podem atuar como células imunes. Os
tipos mais comuns nos adultos são os linfomas não - Hodgkin, derivados das
células B dos centros germinativos. Podem ser indolentes, como no caso do
linfoma folicular, ou agressivo como no caso do linfoma difuso das grandes células
B. Algumas vezes, interferem no sistema imune, desregulando a função normal das
células B e T (ROBBINS, et al., 2008).
Figura 1: Gênese das células sanguíneas e linfócitos. (Adaptado de: INCA, 2010).

29
3.3 Aspectos sobre doenças parasitárias
As parasitoses continuam sendo responsáveis por elevadas taxas de
mortalidade no Brasil e em diversos países latino-americanos. Embora
consideráveis esforços destinados à pesquisa e desenvolvimento de vacinas
tenham ocorrido na última década, ainda não houve uma implantação de uma
imunoprofilaxia eficaz contra protozoários e parasitas. Além disso, programas de
erradicação de doenças vetoriais, baseados em combate ao inseto vetor,
freqüentemente são fadados ao fracasso devido ao elevado custo e resistência aos
inseticidas utilizados (BRESOLIN, CECHINEL FILHO, 2010).
As doenças parasitárias são consideradas como um problema de saúde
pública, pois disponíveis para o tratamento há um limitado número de fármacos, e
mesmo assim os que existem, apresentam significativos efeitos colaterais. Para
tanto, faz-se necessário uma busca de novos agentes antiparasitários a partir de
substâncias naturais que sejam menos nocivos ao organismo sadio e mais efetivo
no tratamento da parasitose. Embora muitas substâncias naturais apresentem
promissora atividade antiparasitária, o desenvolvimento de um novo medicamento
aprovado em todos os testes clínicos ainda é uma realidade distante. Um
levantamento realizado sobre os novos fármacos desenvolvidos entre 1975 e 1996
revelou que de 1.223 novos fármacos, somente 1% (11 novas substâncias e 2
formulações) foram destinados para o tratamento de doenças tropicais (BRASIL,
2007).
3.3.1 Leishmaniose
A Leishmaniose Tegumentar Americana (LTA) tem ampla distribuição
mundial e no continente Americano onde há registros de casos desde o extremo
Sul dos Estados Unidos até o Norte da Argentina, com exceção do Chile e do
Uruguai. Ela é um problema de saúde pública em 88 países distribuídos em quatro
continentes (Américas, Europa, África e Ásia), com registro anual de 1 a 1,5
milhões de casos. É considerada pela OMS, como uma das seis mais importantes
doenças infecciosas, pelo alto coeficiente de detecção e capacidade de produzir
deformidades. No Brasil a LTA é uma doença com uma diversidade de agentes, de
reservatórios e de vetores que apresenta diferentes padrões de transmissão e um
30
conhecimento ainda limitado sobre alguns aspectos, o que a torna de difícil controle
(BRASIL, 2007).
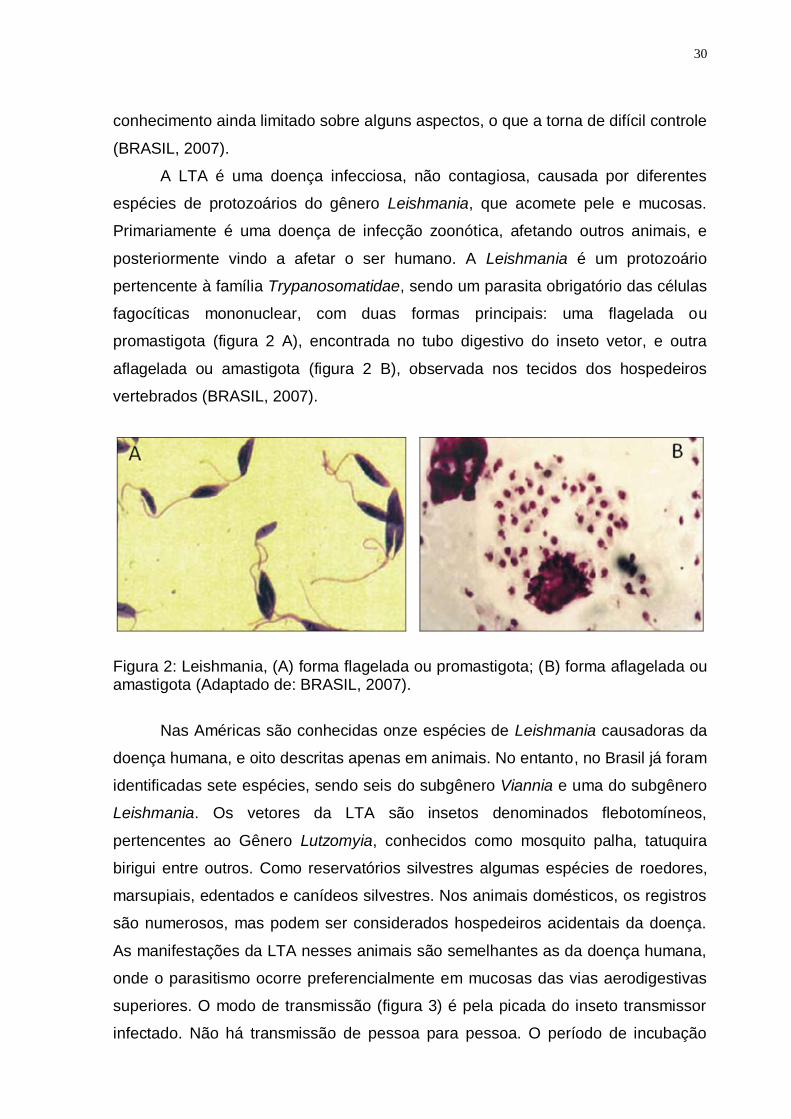
A LTA é uma doença infecciosa, não contagiosa, causada por diferentes
espécies de protozoários do gênero Leishmania, que acomete pele e mucosas.
Primariamente é uma doença de infecção zoonótica, afetando outros animais, e
posteriormente vindo a afetar o ser humano. A Leishmania é um protozoário
pertencente à família Trypanosomatidae, sendo um parasita obrigatório das células
fagocíticas mononuclear, com duas formas principais: uma flagelada ou
promastigota (figura 2 A), encontrada no tubo digestivo do inseto vetor, e outra
aflagelada ou amastigota (figura 2 B), observada nos tecidos dos hospedeiros
vertebrados (BRASIL, 2007).
Figura 2: Leishmania, (A) forma flagelada ou promastigota; (B) forma aflagelada ou amastigota (Adaptado de: BRASIL, 2007).
Nas Américas são conhecidas onze espécies de Leishmania causadoras da
doença humana, e oito descritas apenas em animais. No entanto, no Brasil já foram
identificadas sete espécies, sendo seis do subgênero Viannia e uma do subgênero
Leishmania. Os vetores da LTA são insetos denominados flebotomíneos,
pertencentes ao Gênero Lutzomyia, conhecidos como mosquito palha, tatuquira
birigui entre outros. Como reservatórios silvestres algumas espécies de roedores,
marsupiais, edentados e canídeos silvestres. Nos animais domésticos, os registros
são numerosos, mas podem ser considerados hospedeiros acidentais da doença.
As manifestações da LTA nesses animais são semelhantes as da doença humana,
onde o parasitismo ocorre preferencialmente em mucosas das vias aerodigestivas
superiores. O modo de transmissão (figura 3) é pela picada do inseto transmissor
infectado. Não há transmissão de pessoa para pessoa. O período de incubação
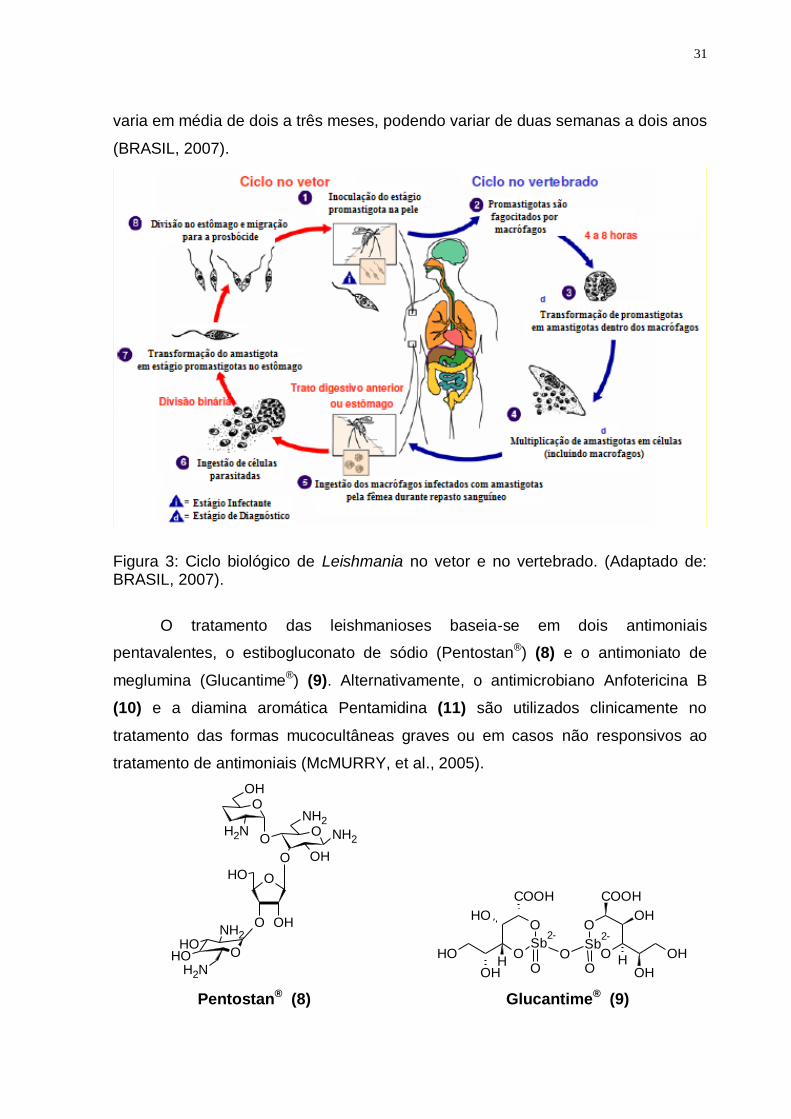
31
varia em média de dois a três meses, podendo variar de duas semanas a dois anos
(BRASIL, 2007).
Figura 3: Ciclo biológico de Leishmania no vetor e no vertebrado. (Adaptado de: BRASIL, 2007).
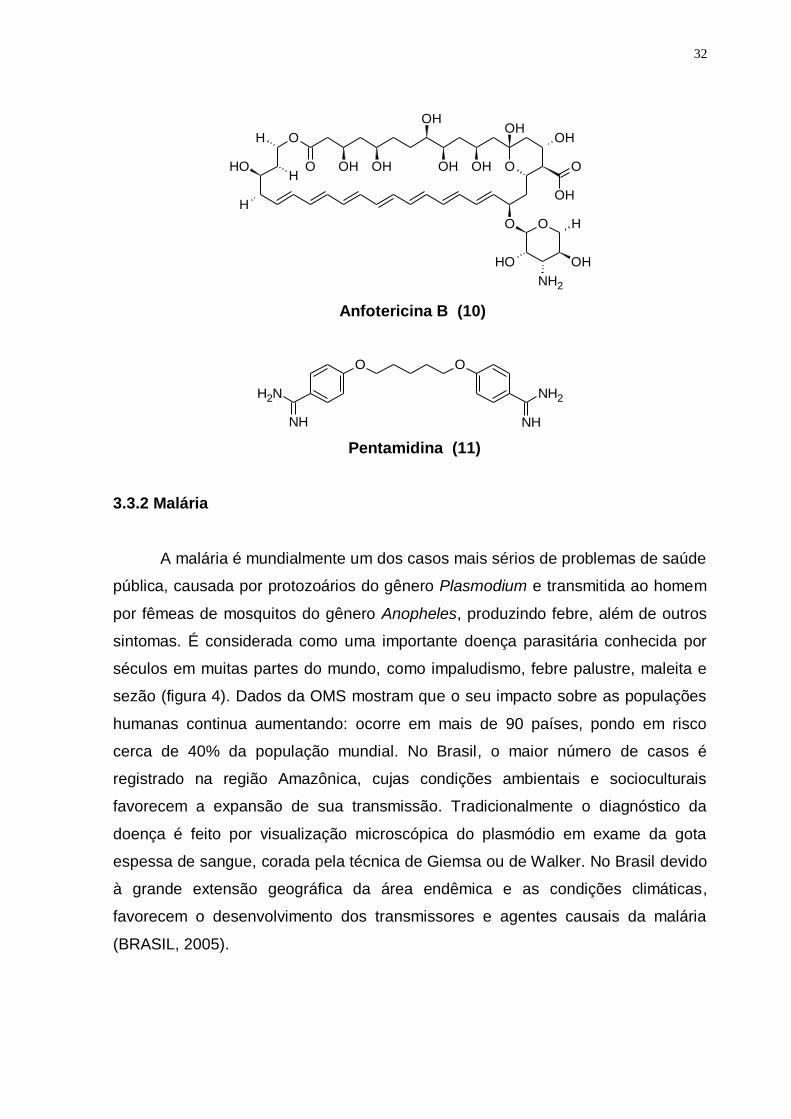
O tratamento das leishmanioses baseia-se em dois antimoniais
pentavalentes, o estibogluconato de sódio (Pentostan®) (8) e o antimoniato de
meglumina (Glucantime®) (9). Alternativamente, o antimicrobiano Anfotericina B
(10) e a diamina aromática Pentamidina (11) são utilizados clinicamente no
tratamento das formas mucocultâneas graves ou em casos não responsivos ao
tratamento de antimoniais (McMURRY, et al., 2005).
O
O
NH2O
OH
O
O
NH2
O
OH
O OH
NH2
NH2OH
OH
OH
NH2
OH
O
OSb
2-O
OSb
2-
O OH
OHH
O
OH
COOH
O
OH
COOH
OHH
Pentostan® (8) Glucantime® (9)
32
H
OH
O
HO OH OH OH OH O
OH
O
OHH
OO H
OH
NH2
OH
OHOH
Anfotericina B (10)
NH2
O O
NH
NH2
NH
Pentamidina (11)
3.3.2 Malária
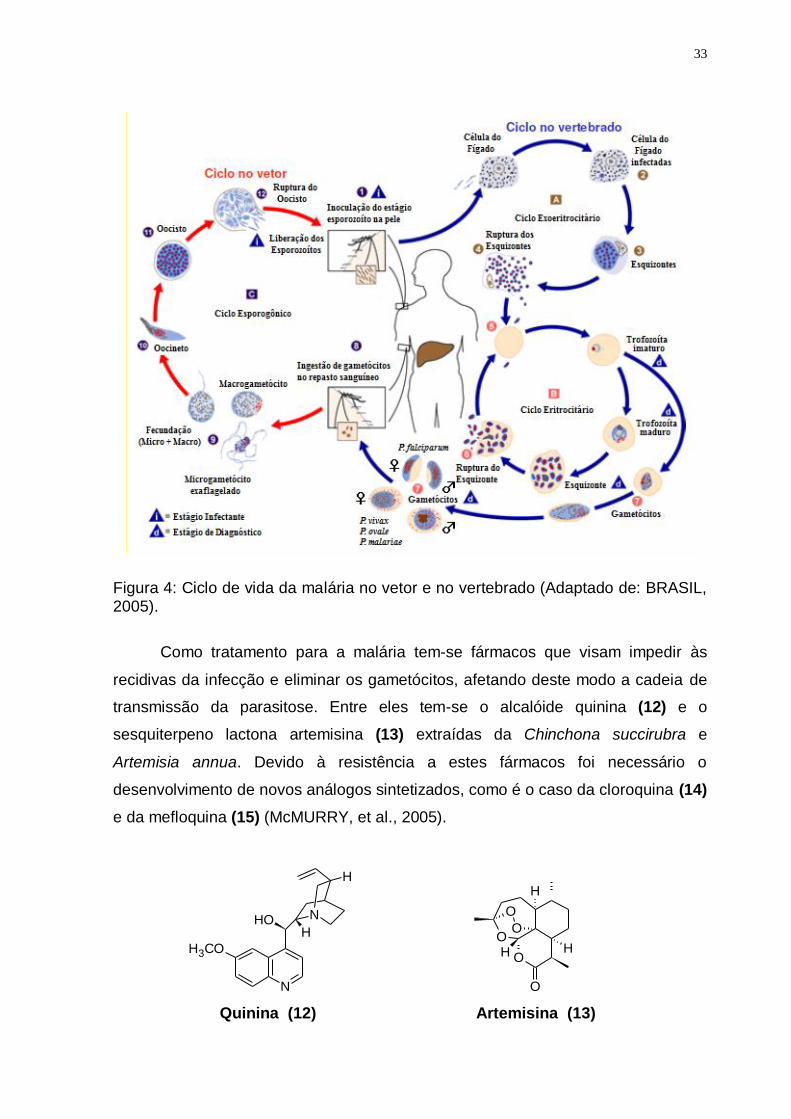
A malária é mundialmente um dos casos mais sérios de problemas de saúde
pública, causada por protozoários do gênero Plasmodium e transmitida ao homem
por fêmeas de mosquitos do gênero Anopheles, produzindo febre, além de outros
sintomas. É considerada como uma importante doença parasitária conhecida por
séculos em muitas partes do mundo, como impaludismo, febre palustre, maleita e
sezão (figura 4). Dados da OMS mostram que o seu impacto sobre as populações
humanas continua aumentando: ocorre em mais de 90 países, pondo em risco
cerca de 40% da população mundial. No Brasil, o maior número de casos é
registrado na região Amazônica, cujas condições ambientais e socioculturais
favorecem a expansão de sua transmissão. Tradicionalmente o diagnóstico da
doença é feito por visualização microscópica do plasmódio em exame da gota
espessa de sangue, corada pela técnica de Giemsa ou de Walker. No Brasil devido
à grande extensão geográfica da área endêmica e as condições climáticas,
favorecem o desenvolvimento dos transmissores e agentes causais da malária
(BRASIL, 2005).
33
Figura 4: Ciclo de vida da malária no vetor e no vertebrado (Adaptado de: BRASIL, 2005).
Como tratamento para a malária tem-se fármacos que visam impedir às
recidivas da infecção e eliminar os gametócitos, afetando deste modo a cadeia de
transmissão da parasitose. Entre eles tem-se o alcalóide quinina (12) e o
sesquiterpeno lactona artemisina (13) extraídas da Chinchona succirubra e
Artemisia annua. Devido à resistência a estes fármacos foi necessário o
desenvolvimento de novos análogos sintetizados, como é o caso da cloroquina (14)
e da mefloquina (15) (McMURRY, et al., 2005).
N
OHH
N
H
H3CO
O
O
O
O
O
H H
H
Quinina (12) Artemisina (13)

34
N
NHN
Cl
N
OHNH
CF3
CF3
Cloroquina (14) Mefloquina (15)
3.4 Aspectos botânicos do gênero Drimys
A família Winteraceae consiste de árvores e arbustos que são encontrados
em toda a América Central e do Sul. Ela contém aproximadamente cem espécies
com seis ou sete gêneros (WILLIANS; HARVEY, 1982).
A Drimys seção wintera é nativa do sul do Brasil e de alguns outros países
da América do Sul. Ela é conhecida como cataia, casca-d’-anta, caá-pororoca,
canela-amarga, capororoca-picante, carne-d’-anta, melambo, melambó, para-tudo,
pau-pra-tudo. A denominação winter é devido ao capitão Winter que utilizou as
cascas da Drimys para tratar a sua tripulação de escorbuto. A casca das espécies
de Drimys era comercializada como “Casca de Winter” ou “Cortex Winteranus”
(TRINTA; SANTOS, 1997). Pio Correa (1931) relata que o nome vernáculo casca-
da-anta provém do folclore dos aborígenes Araucanos onde estes relatam que
quando a anta (Tapirus americanus) se vê doente recorre a cascas desta espécie.
A Drimys brasiliensis também conhecida como casca-da-anta é muito
utilizada como substituto da pimenta do reino na culinária, além de ser utilizada na
medicina tradicional como carminativa e estomáquica. Ela também é altamente
recomendada para problemas gástricos e intestinais, incluindo dispepsia,
disenteria, náuseas, dores intestinais e cólicas bem como febre e anemia. Ainda
pode-se citar atividade antiespasmódica, aromática, contra hemorragia uterina e,
algumas vezes no tratamento do câncer (ANDRÉ, et al., 1999; 2003; 2004;
FORSBY; WALUM, 1996; FORSBY, et al., 1998; SILVA, 2009).
A planta apresenta-se como uma árvore com 4 a 8 metros de altura, dotada
de uma copa globosa e densa. O tronco é geralmente tortuoso, de 30 a 40 cm de
diâmetro, com casca rugosa de cor cinza. Folhas simples, glabras, de coloração
verde mais clara na face inferior, com 8 a 12 cm de largura. Inflorescências em
panículas terminais, com flores brancas. Fruto do tipo baga subglobosa, contendo 2

35
a 5 sementes pretas. A sua ocorrência é em todo o Brasil, em várias formações
florestais, porém principalmente em matas ciliares e na floresta semidecídua de
altitude. Planta perenifólia, heliófita, seletiva e higrófita. Dependendo do local de
ocorrência pode variar em porte avaliando-se de norte a sul do país, indo de 7 a 9
metros de altura (figura 5). Pode florescer até duas vezes por ano, porém com
maior intensidade em julho a agosto. E os frutos amadurecem em outubro a
novembro (LORENZI, 2002).
O gênero Drimys em Santa Catarina ocorre duas espécies, D. winteri e D.
brasiliensis, as quais são utilizadas na região do Planalto Catarinense como
condimento para carnes, transformando-se suas cascas em pó e utilizando-o como
substituto da pimenta-do-reino (SIMÕES, et al., 1986).
Figura 5: Drimys brasiliensis: aspecto geral da planta adulta. (Fonte: Autora).

36
A Drimys possui a seguinte classificação taxonômica (BARROSO, 1991):
Filo: Magnoliophyta (Angiosperma),
Classe I: Magnolitae (Dicotiledônea),
Subclasse: Magnoliidae,
Ordem: Magnoliales,
Família: Winteraceae,
Gênero: Drimys,
Espécie: Drimys brasiliensis.
3.5 Aspectos químicos e farmacológicos do gênero Drimys
O gênero Drimys destaca-se por apresentar metabólitos secundários
responsáveis por diversos efeitos farmacológicos comprovados cientificamente,
incluindo atividades antiinflamatória, antibacteriana, antifúngica, citotóxica,
moluscicida, piscicida, antihelmíntica e reguladora do crescimento de plantas. Entre
as principais classes de substâncias encontradas são os sesquiterpenos drimanos,
flavonóides e aromadendranos (MORTON, 1981; SIMÕES, et al., 1986; ANDRÉ, et
al., 1999; 2003; 2004; JANSEN e GROOT, 2004).
Mendes e colaboradores (1998), utilizando o extrato hidroalcoólico de
cascas de D. winteri e o sesquiterpeno isolado poligodial (16), investigaram as
propriedades antiiperalgésicas em modelo de contorções abdominais induzidas por
três agentes químicos irritantes: ácido acético, zimosan e kaolin e o modelo de
edema de pata causado por formalina e capsaicina. O extrato não exibiu efeito nos
modelos analisados, mas o poligodial (16) produziu uma inibição dose-dependente
das contorções abdominais provocadas pelos agentes químicos.
O extrato das cascas e o poligodial (16) também foram avaliados em
modelos experimentais afim de verificar as atividades antiinflamatórias e
antialérgicas: o modelo de edema de pata induzido por agentes químicos
(prostaglandina E2, bradicinina (BK), substância P (SP), dextrano, fator de ativação
plaquetária (PAF) e carragenina); o modelo de pleuresia (SP, BK, des-Arg9-BK,
histamina, carragenina e PAF); e o modelo de edema de orelha (ácido
araquidônico, capsaicina e óleo de cróton), em camundongos. Nos três modelos o
poligodial (16) exibiu resultados satisfatórios. Ele protegeu o tecido tratado com os
agentes flogísticos evitando a extensão da lesão tanto no edema de orelha quanto

37
de pata. também evitou o choque anafilático provocado pela ovoalbumina no
método de edema de pata. Para a pleuresia houve inibição significativa
comprovada pela diminuição da exudação e influxo celular para a área lesionada
do parênquima pulmonar e a resposta inflamatória foi reduzida, comprovando seu
efeito antiinflamatório e antialérgico (CUNHA, et al., 2001).
Cechinel Filho e colaboradores (1998) estudaram as frações diclorometano e
acetato de etila obtidas do extrato metanólico das cascas de D. winteri. Neste
estudo, verificou-se que o poligodial (16), o 1-β-(p-metoxicinamil)-poligodial (17), a
taxifolina (18) e a astilbina (19) estavam presentes nas frações obtidas. As
substâncias foram avaliadas em modelos de contorções induzidas por ácido
acético. O poligodial (16) e o 1-β-(p-metoxicinamil)-poligodial (17) foram
responsáveis pela ação antinociceptiva, sendo mais potentes que a aspirina e o
paracetamol, utilizados como fármacos de referência. Em trabalho similar, André e
colaboradores (2004), utilizando o poligodial (16) e o drimanial (20) em ratos
neonatos, observaram uma pronunciada inibição da nocicepção e da hiperalgesia
térmica quando adultos utilizando o modelo de indução por capsaicina.
CHO
CHO
H
O
O
MeO
CHO
CHO
Poligodial (16) 1-β-(p-metoxicinamil)-poligodial (17)
OOH
OH O
OH
OH
OH
OOH
OH O
ORamnose
OH
OH
Taxifolina (18) Astilbina (19)
Considerando os resultados promissores obtidos para o poligodial (16), foi
investigado o seu potencial de inibição de receptores muscarínicos agonistas de
acetilcolina ativados (mAChR) na transdução de sinal em células de neuroblastoma
humano SH-SY5Y. Este provocou uma inibição de carbacol e a produção de

38
inositol fosfato em 79 e 84%, respectivamente. Quando foi avaliada a sua ação
comparando com a toxina pertussis (PTX), utilizada como modelo padrão de
inibição, ele foi aproximadamente 50% mais potente que a toxina, em termos de
diminuição do carbacol induzido por afluxo de Ca2+. Dessa forma conclui-se que o
poligodial inibiu a transdução de sinal de mAChR em células SH-SY5Y e que o
local de ação inibitória não está localizado na mAChR, mas no nível da proteína-G
ou a jusante da via de transdução de sinal (FORSBY, et al., 1998).
A ação vasorrelaxante do poligodial (16) foi avaliada sobre a ação mediada
pela liberação de óxido nítrico (NO) ou por uma substância relacionada no
endotélio vascular. Ele produz também relaxamento nos anéis da artéria pulmonar
do coelho e na artéria pulmonar torácica da aorta e no endotélio da cobaia. Os
resultados sugerem que os canais de Ca2+ e K+ não estão envolvidos na ação
vasorelaxante na artéria pulmonar nem na artéria torácica da aorta. Mas, no
endotélio da cobaia os canais sensíveis ao trifosfato de adenosina (ATP) podem
ser ativados, e dessa forma, pode-se explicar em parte a sua ação vasorelaxante
(ANDRÉ, et al., 1999; 2004). Também foi testado a ação vasorrelaxante in vitro do
poligodial (16) sobre a veia porta de ratos. Constatou-se que houve uma
significativa diminuição das contrações sugerindo que houve um relaxamento
muscular provavelmente associada ao influxo de cálcio dos canais sensíveis a
voltagem e a interação dos mecanismos da proteína quinase-Ca dependente (EL
SAYAH, et al., 2000). Também foi observada a ação sobre a enzima que hidrolisa
o ATP (ATPase) mitocondrial do poligodial (16). Ele inibiu fortemente a síntese de
ATP acoplado a oxidação de partículas de succinato submitocondriais do coração.
Sendo assim, ele é um potente inibidor da síntese de ATP mitocondrial. Apesar
dele não aumentar a permeabilidade da membrana mitocondrial interna aos
prótons, o mais provável mecanismo de ação seria um desacoplamento na síntese
de ATP na cadeia transportadora de elétrons, onde pode ser verificado e explicado
pela mudança das propriedades elétricas da membrana, ou seja, há uma
despolarização quando na presença da substância e, consequentemente, o
colapso do potencial de membrana. Com isso há um desequilíbrio no pH interno
com o pH transmembrana causando assim, a ruptura da célula (CASTELLI, et al.,
2005).
Outro drimano que merece destaque é o drimanial (20) isolado das cascas
de D. winteri. Este apresentou ação antinociceptiva nos modelos de dor induzida

39
por ácido acético, formalina, capsaicina e glutamato. Comprovou-se sua ação em
ratos ao nível local, periférico, espinhal e supra-espinhal, além de haver uma
interação com os receptores de glutamato metabotrópicos, contribuindo assim para
os mecanismos de ação antinociceptiva. O drimanial (20) e poligodial (16) inibiram
a captação de glutamato pelos astrócitos, analisados em cortes histológicos do
córtex, hipocampo e corpo estriado. Foi observado um aumento da liberação de
glutamato por sinaptossomas. Estes efeitos simultâneos predispõem a um aumento
da concentração do glutamato extracelular, levando a possíveis efeitos
neurotóxicos (excitotoxicidade) destas substâncias. Por isso a necessidade de
alguma cautela na sua aplicação terapêutica (MALHEIROS, et al., 2001; SCHEIDT,
et al., 2002; MARTINI, et al., 2007).
O
O
MeO
CHO
CHO
OH
Drimanial (20)
Em preparações de membrana de córtex de camundongos o drimanial (20)
inibiu o “binding” de [3H]Glutamato. Nos experimentos de nocicepção induzida por
formalina, hiperalgesia térmica e edema de pata, hiperalgesia termal e
extravasamento plasmático induzido por capsaicina e tratamento neonatal também
foi verificado o efeito antiinflamatório e antinociceptivo desta substância (ANDRE,
et al., 2004).
Os sesquiterpenos drimanos, o drimanial (20) e poligodial (16) também
foram avaliados sobre os receptores potenciais vanilóides transitórios 1 (TRPV1)
em bexiga urinária isolada de ratos. Ambos produziram uma contração mediada
por taquicinina que foi inibida pela combinação dos receptores antagonistas. Estas
substâncias em conjunto com a capsaicina promoveram um aumento da captação
de Ca2+ em sinaptossomas de medula espinhal, e em níveis intracelulares em
culturas de neurônios trigêmeos de ratos. Em resumo, os resultados sugerem que
estes drimanos parecem ser parcialmente mediados pela ativação direta e/ou
indireta de TRPV1 (ANDRE, et al., 2006).

40
Witaicenis e colaboradores (2007) comprovaram a atividade antiulcerogênica
de extratos etanólicos de cascas e folhas de D. angustifolia. Os extratos das
cascas e folhas na dose de 250 mg/kg, apresentaram atividade antiulcerogênica no
modelo de etanol/HCl. Eles inibiram a lesão em 81,42% e 76,50%,
respectivamente, sendo maior que o fármaco de referência a carbenoxolona.
Possivelmente a atividade antiulcerogênica pode estar relacionada com a presença
dos sesquiterpenos drimanos. Em um estudo biodirecionado em plantas do gênero
Tasmannia utilizando o extrato metanólico das folhas de T. lanceolata, Matsuda e
colaboradores (2002), avaliaram o potencial inibitório contra úlcera gástrica
induzida por etanol em ratos. Foram isoladas quatro substâncias: o poligodial (16),
poligodial 12α-acetal (21), poligodial 12β-acetal (22) e metil isodrimeninol (23).
Entre eles, o poligodial (16) se mostrou muito potente sobre o efeito gastroprotetor
(ED50= 28 µg/kg, via oral). O trabalho sugere que a ação gastroprotetora do
poligodial (16) é particularmente mediada pelas interações com neurônios
sensoriais via receptores vanilóides.
H
O
OMe
MeO
H
O
OMe
MeO
H
OMeO
12α-acetal (21) Poligodial 12α-acetal (22) Metil isodrimeninol (23)
A atividade antifúngica também é relatada no gênero Drimys. Malheiros e
colaboradores (2005), realizaram um fracionamento biomonitorado do extrato
clorofórmico das cascas. Foram isolados os sesquiterpenos poligodial (16), 1-β-(p-
metoxicinamil)-poligodial (17), drimanial (20) e 1-β-(p-cumaroiloxi)-poligodial (24),
que foram seletivamente ativos contra os dermatófitos Epidermophyton floccosum e
Tricophyton rubrum. O poligodial (16) inibiu ativamente E. floccosum com valores
de concentração inibitória mínima (CIM) de 3 µg/mL. O 1-β-(p-metoxicinamil)-
poligodial (17), o drimanial (20) e o 1-β-(p-cumaroiloxi)-poligodial (24), inibiram
esses fungos em concentrações superiores ao poligodial (16), em torno de 25
µg/mL. Assim demonstrou-se que as três substâncias contribuem para a atividade
atifúngica observada nas cascas.

41
O
O
OH
CHO
CHO
1-β-(p-cumaroiloxi)-poligodial (24)
Investigando a depleção de glutationa em Saccharomyces cerevisiae,
Machida e colaboradores (1999), utilizando a substância isolado poligodial (16),
verificaram houve uma forte atividade fermentativa, e detectaram a produção de
espécies reativas de oxigênio (ROS) em um nível mais significativo podendo ser
identificados com uma sonda fluorescente. A produção de ROS em células tratadas
com poligodial (16) foi confirmado pela sua eliminação e o acompanhado pelo
processo protetor contra os efeitos fermentativos na presença de antioxidantes tais
como ascorbato e α-tocoferol. Poligodial (16) poderia acelerar a produção de ROS
apenas em células da linhagem Grande do tipo selvagem, mas não nas mutantes
deficientes de respiração, indicando o seu papel interferente na cadeia mitocondrial
de transporte de elétrons na produção de ROS. Diferentemente do caso com uma
antimicina que acelera a produção de ROS por atacar diretamente o fluxo de
elétrons mitocondriais, o poligodial (16) causa a depleção de glutationa
citoplasmática e mitocondrial, que funciona em eliminar ROS inevitavelmente
gerados durante o crescimento aeróbio. O esgotamento do poligodial (16) mediada
pela glutationa intracelular pode, possivelmente, ser dependente de uma interação
direta entre a sua porção enal e o grupo sulfidrila da cisteína em glutationa.
Tentando descobrir como o poligodial (16) age como agente antifúngico,
Fujita e Kubo (2005), realizaram um ensaio de atividade antifúngica com
Saccharomyces cerevisiae. O poligodial (16) num primeiro momento atua como um
agente de superfície (surfactante) e, em seguida, torna-se envolvido em processos
bioquímicos. A capacidade de formar um derivado pirrol com um grupo de amina
primária de fosfatidiletanolamina (PE) e fosfatidilserina (PS) na monocamada
externa da membrana plasmática é provável que, em parte, seja um passo inicial
para a ação antifúngica do poligodial (16). A ação primária fungicida do poligodial
(16) vem de sua capacidade de agir como um surfactante que inespecíficamente
interrompe a interface de proteínas, lipídios, proteínas integrais, desnaturando a

42
sua conformação funcional. Depois o poligodial (16) entra no citoplasma,
destruindo a barreira de membrana, reagindo com L-cisteína contida no material
citoplasmático, com uma pequena molécula de glutationa, e com a enzima álcool
desidrogenase, para potencializar a ação antifúngica.
Em estudo mais recente realizado por Silveira (2011), foi estudada a
atividade antibacteriana, de diferentes extratos brutos, frações e substâncias de D.
brasiliensis ultilizando a metodologia de diluição de ágar. Neste mesmo estudo foi
avaliado o potencial citotóxico dos extratos brutos utilizando-se o microcrustáceo
Artemia salina. Nos ensaios de bioautografia das frações, verificou-se as frações
que continham as substâncias de interesses e que exerciam atividade biológica,
guiando assim a parte fitoquímica para o isolamento. O extrato clorofórmico de
cascas de D. brasiliensis e as substâncias isoladas espatulenol, poligodial e 1-β-(p-
metoxicinamil)-poligodial apresentaram promissora atividade inibitória contra as
bactérias Gram positivas com CIM entre 31,25 e 250 µg/mL, mas se mostraram
inativos contra as Gram-negativas. O poligodial apresentou melhor atividade contra
Helicobacter pylori, com CIM de 250 µg/mL. Tanto o extrato clorofórmico quanto as
frações apresentaram elevada toxicidade sobre a Artemia salina, evidenciando que
possam ter substâncias citotóxicas no extrato.
Existem alguns estudos avaliando o potencial antitumoral de sesquiterpenos
drimanos. A respeito da indução de inositol fosfato em linhagens de células de
neuroblatoma humano (SH-SY5Y), Forsby e Walum (1996), investigaram o efeito
do isoveleral (25), poligodial (16) e epipoligodial (26) sobre a formação de inositol
fosfato (IP) supondo que poderia haver uma correlação entre a liberação de Ca2+
intracelular e pungência das substâncias. Verificou-se que poligodial (16) induz a
mobilização de IP de uma forma dependente da concentração, enquanto isoveleral
(25) não teve efeito sobre a formação IP nas células SH-SY5Y. O volume de
fosfoinositídeo (PPI) foi ativada por epipoligodial (26), mas apenas em
concentrações 40 vezes superiores ao poligodial (16). Isto enfatiza a importância
da correta esteriometria na configuração dialdeído para a atividade biológica do
poligodial (16). A formação induzida do IP por poligodial (16) foi reduzida em 71%
sob condições extracelulares livres de cálcio, o que sugere interações entre a
formação IP e o aumento da [Ca2+]i para esclarecer uma ativação periódica da
fosfolipase C (PLC).

43
Num estudo mais recente realizado por Silva (2009), foi avaliada a ação
citotóxica dos extratos clorofórmicos das cascas, caules e folhas da D. brasiliensis
e os sesquiterpenos drimanos drimanial (20) e 1-β-(p-metoxicinamil)-poligodial (17),
isolados do extrato clorofórmico das cascas, em células tumorais Hela
(Adenocarcinoma Epitelial de Cérvix Humano) e não tumorogênica L929
(Fibroblasto Murino). Os resultados obtidos mostram que os extratos clorofórmicos
das folhas e do caule não apresentam elevado grau de citotoxicidade nos modelos
avaliados. O extrato clorofórmico das cascas apresentou um efeito tóxico em
células não tumorogênicas de fibroblasto L929 somente em altas concentrações
como a de 100,0 µg/mL com CI50 > 100,0 µg/mL sugerindo baixa citotoxicidade nos
tempos de 24 e 48 horas de incubação. Nas células tumorais Hela observou-se um
aumento na potência citotóxica com CI50 de 61,2 e 18,0 µg/mL para os tempos de
24 e 48 horas, respectivamente. O drimanial (20) apresentou grau de citotoxicidade
em células L929 com IC50 26,4 µg/mL e Hela 10,0 µg/mL para 24 horas de
incubação. Houve um aumento na potência citotóxica com o tempo de 48 horas de
incubação para células L929 e Hela com CI50 de 16,7 e 8,8 µg/mL,
respectivamente. O 1-β-(p-metoxicinamil)-poligodial (17) apresentou citotoxicidade
em células L929 e Hela após 48 horas de contato com CI50 de 10,1 e 7,7 µg/mL,
respectivamente, sugerindo um potencial citotóxico para ambos os drimanos
avaliados.
CHO
CHO
CHO
CHO
H
Isoveleral (25) Epipoligodial (26)
Investigando a possível atividade antialimentar, Zapata e colaboradores
(2009) investigaram os extratos da casca e do caule (hexano, acetona e
metanol:água [80:20]) e os drimanos isolados drimendiol (27), isodrimenol (28),
isotadeonal (29) e poligodial (16) isolados de D. winteri contra larvas e lagartas de
Spodoptera littoralis (Noctuidae). O extrato hexano (5000 ppm) e o poligodial (16)
(1000 ppm) apresentaram atividade antialimentar comprovada no crescimento e
desenvolvimento das larvas. Já para o teste de toxicidade as substâncias isoladas
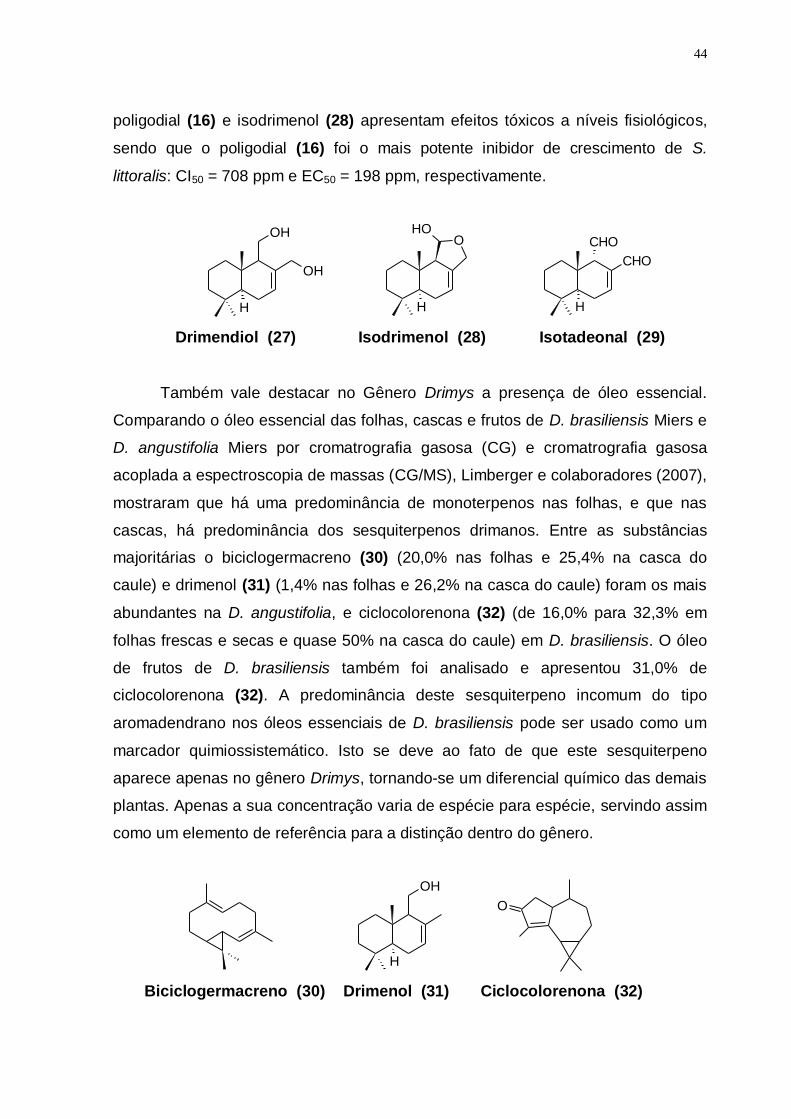
44
poligodial (16) e isodrimenol (28) apresentam efeitos tóxicos a níveis fisiológicos,
sendo que o poligodial (16) foi o mais potente inibidor de crescimento de S.
littoralis: CI50 = 708 ppm e EC50 = 198 ppm, respectivamente.
H
OH
OH
H
OOH
CHO
CHO
H
Drimendiol (27) Isodrimenol (28) Isotadeonal (29)
Também vale destacar no Gênero Drimys a presença de óleo essencial.
Comparando o óleo essencial das folhas, cascas e frutos de D. brasiliensis Miers e
D. angustifolia Miers por cromatrografia gasosa (CG) e cromatrografia gasosa
acoplada a espectroscopia de massas (CG/MS), Limberger e colaboradores (2007),
mostraram que há uma predominância de monoterpenos nas folhas, e que nas
cascas, há predominância dos sesquiterpenos drimanos. Entre as substâncias
majoritárias o biciclogermacreno (30) (20,0% nas folhas e 25,4% na casca do
caule) e drimenol (31) (1,4% nas folhas e 26,2% na casca do caule) foram os mais
abundantes na D. angustifolia, e ciclocolorenona (32) (de 16,0% para 32,3% em
folhas frescas e secas e quase 50% na casca do caule) em D. brasiliensis. O óleo
de frutos de D. brasiliensis também foi analisado e apresentou 31,0% de
ciclocolorenona (32). A predominância deste sesquiterpeno incomum do tipo
aromadendrano nos óleos essenciais de D. brasiliensis pode ser usado como um
marcador quimiossistemático. Isto se deve ao fato de que este sesquiterpeno
aparece apenas no gênero Drimys, tornando-se um diferencial químico das demais
plantas. Apenas a sua concentração varia de espécie para espécie, servindo assim
como um elemento de referência para a distinção dentro do gênero.
H
OH
O
Biciclogermacreno (30) Drimenol (31) Ciclocolorenona (32)

45
3.6 Técnicas de fracionamento, isolamento e identificação de produtos
naturais
Quando se destina a obtenção de substâncias bioativas numa planta, deve
se priorizar a preservação de sua integridade química e farmacológica, garantindo
a constância de sua ação biológica e a segurança de sua utilização, além de
valorizar o seu potencial terapêutico (PETROVICK, et al., 2007).
A preparação dos extratos brutos das plantas deve ser a etapa mais
importante e a que precisa ter mais cuidado para se realizar. A escolha do solvente
também é de grande importância para que a extração ocorra de forma a separar as
substâncias presentes no extrato. (FALKENBERG; SANTOS; SIMÕES, 2004).
Na investigação de constituintes presentes no extrato de uma planta, pode-
se recorrer a um fracionamento do extrato total a fim de se separar as diferentes
classes de substâncias presentes por meio de solventes que podem separar por
diferença de polaridade. A separação e a purificação dos constituintes químicos
são realizadas por técnicas cromatográficas, nas quais duas ou mais substâncias
em uma mistura podem ser separados fisicamente por diferença de polaridade e as
suas distribuições se dão entre duas fases: uma fase estacionária, que pode ser
um líquido ou um sólido ou um líquido suportado num sólido e uma fase móvel, um
gás ou um líquido, que flui continuamente pela fase estacionária (DI STASI, 1996).
Na cromatografia líquida clássica (CLC), a fase móvel é arrastada através da
coluna apenas por força da gravidade, enquanto que na cromatografia líquida de
alta eficiência (CLAE) e na cromatografia flash (CF) se utilizam fases estacionárias
de partículas menores, sendo necessário o uso de uma bomba de alta pressão
para eluição da fase móvel. As separações também podem ser realizados por
cromatografia gasosa simples (CG) e por cromatografia gasosa de alta resolução
(CGAR). A diferença entre as duas reside nos tipos de colunas utilizadas. Enquanto
na CGAR são utilizadas colunas capilares, nas quais a fase estacionária é um filme
depositado na coluna, a outra tem uma fase estacionária formada por sílica. Já o
cromatotron é uma cromatografia em camada delgada preparativa (CCDP) e
acelerada centrifugamente. É composto por um sistema com uma placa circular de
cromatografia em camada delgada inclinada, o que permite mais eficácia na coleta
das substâncias ou frações. A cromatografia em camada delgada (CCD) é uma

46
técnica de adsorção líquido/sólido. Nesse caso, a separação dos componentes da
mistura ocorre em função da migração diferencial sobre uma camada delgada de
adsorvente, fixo numa superfície plana, por meio de uma fase móvel (um líquido ou
misturas de líquidos) (AQUINO NETO, 2003).
Entre as técnicas de identificação, a espectroscopia de infravermelho, é
utilizada para identificação e comparação de substâncias orgânicas, verificação do
grau de pureza e acompanhamento de reações (SILVERSTEIN; BASSLER;
MORRIL, 1994; PAVIA, et al., 2010). É a técnica que abrange a faixa 4000 e 400
cm-1 no espectro eletromagnético, fornecendo informações sobre as características
da molécula como um todo, de seus grupamentos funcionais, originando bandas
que ocorrem na mesma frequência, independente da estrutura da molécula
(SILVERSTEIN; BASSLER; MORRIL, 1994; NIERO et al., 2003; PAVIA, et al.,
2010).
O método destina-se tanto à análise qualitativa quanto à quantitativa, de
sólidos, líquidos ou gases. A quantidade de analito usada é muito pequena, da
ordem de miligramas e a amostra não é destruída no ensaio podendo ser
recuperada após a medida do espectro. A interpretação do estado de absorção no
infravermelho é uma ferramenta essencial para caracterizar grupamentos
funcionais (COSTA NETO, 2004).
A Ressonância Magnética Nuclear (RMN) ocorre com átomos quando as
seguintes condições são atendidas: o átomo possuir um valor de spin nuclear maior
do que zero; o átomo, na molécula, estar sujeito a um campo magnético externo; o
sistema ser irradiado com radiação eletromagnética; ter relação entre valores do
spin nuclear, campo magnético externo e frequência de irradiação (SILVERSTEIN;
BASSLER; MORRIL, 1994; PAVIA, et al., 2010).
Na Ressonância Magnética Nuclear de Hidrogênio RMN de 1H, cada
hidrogênio eletronicamente distinto em uma molécula normalmente possui sua
própria absorção. Um dos usos da RMN de 1H é encontrar quantos tipos de
hidrogênios não equivalentes estão presentes (McMURRY, 2005; PAVIA, et al.,
2010). Três são as informações mais relevantes que o espectro de RMN de 1H
fornece para fins de caracterização de cadeias hidrocarbônicas: o deslocamento
químico, o desdobramento e a intensidade dos picos (COSTA NETO, 2004; PAVIA,
et al., 2010). Os dados obtidos com esse método espectroscópico são muito

47
importantes para a elucidação estrutural de praticamente todas as classes de
produtos naturais, incluindo metabólitos secundários vegetais.
Na Ressonância Magnética Nuclear de Carbono RMN de 13C é possível
observar o número de diferentes átomos de carbono em uma molécula de estrutura
desconhecida. O deslocamento químico de cada ressonância de carbono 13
depende do ambiente eletrônico do carbono na molécula (McMURRY, 2005).
Os espectros de RMN de hidrogênio e carbono 13 são os mais utilizados e
sua interpretação permite caracterizar o número e o tipo de átomos de hidrogênio e
carbono, em função da localização e do desdobramento dos sinais
correspondentes a absorção de energia eletromagnética (FALKENBERG;
SANTOS; SIMÕES, 2004; PAVIA, et al., 2010).
Outra técnica de identificação é a Espectrometria de Massas (EM). Ela
fornece a massa molecular de uma molécula. Além disso, frequentemente é
possível a informação estrutural a respeito de uma molécula, pela medida das
massas dos fragmentos produzidos quando as moléculas são fragmentadas
(McMURRY, 2005). A massa molecular permite estabelecer a fórmula molecular da
substância, enquanto o padrão de fragmentação pode ajudar a caracterizar a
presença, bem como a localização de certos grupos funcionais e de cadeias
laterais (FALKENBERG; SANTOS; SIMÕES, 2004). Ele pode ser acoplado ao
aparelho de cromatografia gasosa (CG/MS), que permite tanto a identificação como
a quantificação de componentes de baixo peso molecular, mesmo com misturas
complexas. É uma técnica muito utilizada para a análise de óleos voláteis
(FALKENBERG; SANTOS; SIMÕES, 2004; PAVIA, et al., 2010).
A espectroscopia de ressonância magnética nuclear de 1H e 13C
complementa a espectrometria de massas e a espectroscopia na região do
infravermelho, fornecendo um “mapa” do esqueleto carbono-hidrogênio de uma
molécula orgânica. Juntas, as três técnicas possibilitam a determinação das
estruturas de moléculas muito complexas (McMURRY, 2005).

48
4 MATERIAL E MÉTODOS
4.1 Coleta do material vegetal
Drimys brasiliensis foi coletada em Rancho Queimado, localizado no estado
de Santa Catarina em Maio de 2008. A identificação foi realizada anteriormente
pelos botânicos Jasper José Zanco (UNISUL) e Prof. Dr. Ademir Reis (UFSC) e
teve como resultado a espécie Drimys brasiliensis Miers subsp. sylvatica (Saint
Hilaire) Ehrendofer L. Gott sb. Uma exsicata está depositada no Herbário Barbosa
Rodrigues (HBR, Itajaí – SC) sob número HBR-2031.
4.2 Materiais e reagentes
O perfil cromatográfico por CCD dos extratos e substâncias puras obtidas foi
realizado utilizando-se placas de sílica gel 60 GF254, de 20 µm de espessura
preparada sobre folhas de alumínio da Merck. Diferentes sistemas de eluentes
foram utilizados dependendo da polaridade das amostras. Após a eluição as
cromatoplacas foram visualizadas sob luz ultravioleta antes da revelação destrutiva
da amostra. Como revelador na CCD foi utilizado o anisaldeído sulfúrico (AS)
(identificação de terpenos e esteróides). As cromatoplacas foram vaporizadas com
AS e, em seguida, aquecidas à temperatura de 105°C.
Nos procedimentos de cromatografia em coluna, foi utilizada como fase
estacionária, sílica gel 60 (Merck) de granulometria 70-230 mesh ( = 0,063 – 0,20
mm). O diâmetro e altura das colunas foram selecionados de acordo com a
quantidade do material a ser cromatografado. A eluição foi realizada com solventes
orgânicos em ordem crescente de polaridade. Os solventes utilizados foram
hexano (Hex), acetato de etila (AcOEt) e metanol (CH3OH) provenientes dos
Laboratórios Dinâmica, Quimex ou Vetec. As frações coletadas foram reunidas
conforme as semelhanças de fator de retenção (Rf) identificadas na CCD.
Para análise de RMN de 1H e RMN de 13C foram utilizados solventes
deuterados (acetona, clorofórmio e metanol), provenientes da Cambridge Isotope
Laboratories Inc.

49
Além dos reagentes citados acima, foram utilizados acetona, clorofórmio,
éter, metanol (CH3OH), diclorometano (DCM) e sulfato de sódio, provenientes
comercialmente dos Laboratórios Dinâmica, Quimex e Vetec.
4.3 Equipamentos
Para visualização da fluorescência das substâncias rastreadas por CCD, foi
utilizada radiação ultravioleta Minerallight ( = 254 e 366 nm) em câmara de UV-
DIST.
Os espectros de RMN de 1H e RMN de 13C foram realizados em
espectrômetro BRUKER AC-300F em 300 MHz; tendo como referência interna o
tetrametilsilano (TMS) ou o próprio solvente. Os deslocamentos químicos foram
registrados em valores adimensionais (ppm).
Os extratos foram concentrados em rotavapor TECNAL TE-2II com controle
de temperatura, e pesados em balança analítica SHIMADZU LIBROR-AEG-220 e
SHIMADZU LIBROR EB-33OD.
4.4 Obtenção dos extratos de D. brasiliensis
As cascas de D. brasiliensis (774,33 g) foram secas em estufa a 40ºC por
cinco dias, pulverizadas em um triturador, e posteriormente extraídas por
maceração com clorofórmio (CHCl3) e metanol (CH3OH) a temperatura ambiente
por 10 dias cada. Optou-se por este tipo de extração, pois o particionamento não
consegue separar as substâncias de interesse farmacológico presentes no material
vegetal. Já quando se utiliza como solventes primeiramente o CHCl3 e após ele o
CH3OH a separação é bem mais satisfatória. Os sesquiterpenos drimanos,
principal classe de compostos presentes na casca e objeto de estudo de nosso
trabalho, é extraído mais facilmente no extrato clorofórmico. Os extratos foram
concentrados em rotavapor com pressão reduzida para eliminação do solvente,
obtendo-se 31,2 e 62,6 g dos respectivos extratos.
Os caules (cerca de 700 g), foram processados da mesma forma obtendo-se
17,6 e 14,8 g de extrato de CHCl3 e CH3OH, respectivamente.

50
De 250 g de folhas foram obtidos similarmente rendendo 24,5 e 10,6 g de
extrato de CHCl3 e CH3OH, respectivamente.
4.5 Fracionamento do extrato CHCl3 das cascas de D. brasiliensis
Na figura 6 está apresentado o fluxograma dos procedimentos
cromatográficos realizados com o extrato clorofórmico das cascas de D. brasiliensis
Miers. Este extrato compreende a fração mais rica de substâncias que possuem
atividades biológicas comprovadas em estudos anteriores. O extrato e as frações
que foram submetidas ao processo cromatográfico foram eluídas com hexano
(Hex) 100%, posteriormente misturas de Hex e AcOEt aumentando gradativamente
a polaridade e finalizando com EtOH 100%. As frações de cada coluna foram
reunidas de acordo com a semelhança observada por CCD e reveladas por
anisaldeído sulfúrico (AS) sob aquecimento a 105°C.
O extrato CHCl3 das cascas (24,73 g) foi submetido à coluna cromatográfica
(CC) utilizando como fase estacionária sílica gel (238,62 g, int 5,4 cm). Esta coluna
foi denominada coluna 1. Foram coletadas 70 frações. O volume de cada fração
variou de 100 a 20 mL. Da fração 9-21 uma substância denominada de DBF-1
(100,80 mg) foi isolada.
Com o sobrenadante da fração 4-21 (1,47 g) da coluna 1 foi realizado nova
CC de sílica gel (86,61 g, int 3,5 cm), denominada coluna 2. Foram coletadas 105
frações com aproximadamente 15 mL cada. A coluna 3 foi realizada com a fração
26-35 (650,00 mg) proveniente da coluna 2. A coluna foi empacotada com sílica
gel (20,00 g, int 2,5 cm). Foram coletadas 111 frações com aproximadamente 15
mL cada. Da fração 70-71 foi obtida uma substância denominada DBJ-70 (360,00
mg). Da fração 100-105 foi obtida uma substância denominada DBJ-100 (15,40
mg).
A coluna 4 foi realizada com a fração 26-30 (1,29 g) da coluna 1, com sílica
gel (33,72 g, int 3,0 cm). Foram coletadas 61 frações com aproximadamente 15 mL
cada. A coluna 5 foi realizada com a fração 4-8 (690,30 mg) proveniente da
coluna 4, com sílica gel (25,90 g, int 2,5 cm). Foram coletadas 48 frações com
aproximadamente 15 mL cada. Da fração 11-18 foi obtida uma substância
denominada DBI-11 (52,75 mg).

51
A coluna 6 foi realizada com a fração 31-38 (1,59 g) da coluna 1. A coluna
foi empacotada com sílica gel (20,22 g, int 2,0 cm). Foram coletadas 135 frações
com aproximadamente 15 mL cada. Foi obtida da fração 8-11 uma substância
denominada DBK-11 (205,30 mg). Da fração 17-21 foi obtida uma substância
denominada DBK-17 (48,10 mg). Da fração 29-43 foi obtida uma substância
denominada DBK-29 (189,60 mg).
A coluna 7 foi realizada com a fração 39-45 (1,75 g) da coluna 1. Como
fase estacionária utilizou-se sílica gel (79,17 g, int 3,5 cm). Foram coletadas 70
frações com aproximadamente 15 mL cada. A coluna 8 foi realizada com a fração
24-33 (595,20 mg) da coluna 7, utilizando com fase estacionária sílica gel (54,19 g,
int 2,5 cm). Foram coletadas 30 frações com aproximadamente 15 mL cada. Foi
obtido da fração 04-05 uma substância denominada DBX-04 (39,30 mg) e da
fração 09 uma substância denominada DBX-09 (41,40 mg).
A coluna 9 foi realizada com a fração 48-55 (3,74 g) proveniente da coluna
1. Utilizou-se sílica gel (75,89 g, int 3,5 cm) para o empacotamento. Foram
coletadas 91 frações com aproximadamente 15 mL cada. Foi obtido da fração 11-
20 uma substância denominada DBP-11 (204,00 mg), da fração 21-30 uma
substância denominada DBP-21 (128,30 mg) e da fração 44-47 uma substância
denominada DBP-44 (1,61 g).
A coluna 10 foi realizada com a fração 56-59 (3,60 g) da coluna 1. Como
fase estacionária utilizou-se sílica gel (138,70 g, int 5,4 cm). Foram coletadas 60
frações com aproximadamente 15 mL cada. Foi obtida da fração 25-46 uma
substância denominada DBV-25 (2,00 g) e da fração 50-55 um substância
denominada DBV-50 (175,34 mg).
As substâncias isoladas nos diversos procedimentos foram analisadas por
RMN para elucidação de sua estrutura. Outras colunas foram realizadas na
tentativa do isolamento das substâncias de interesse farmacológico. No entanto,
foram omitidas no presente trabalho por não ser possível obter nenhuma
substância, com elevado grau de pureza.
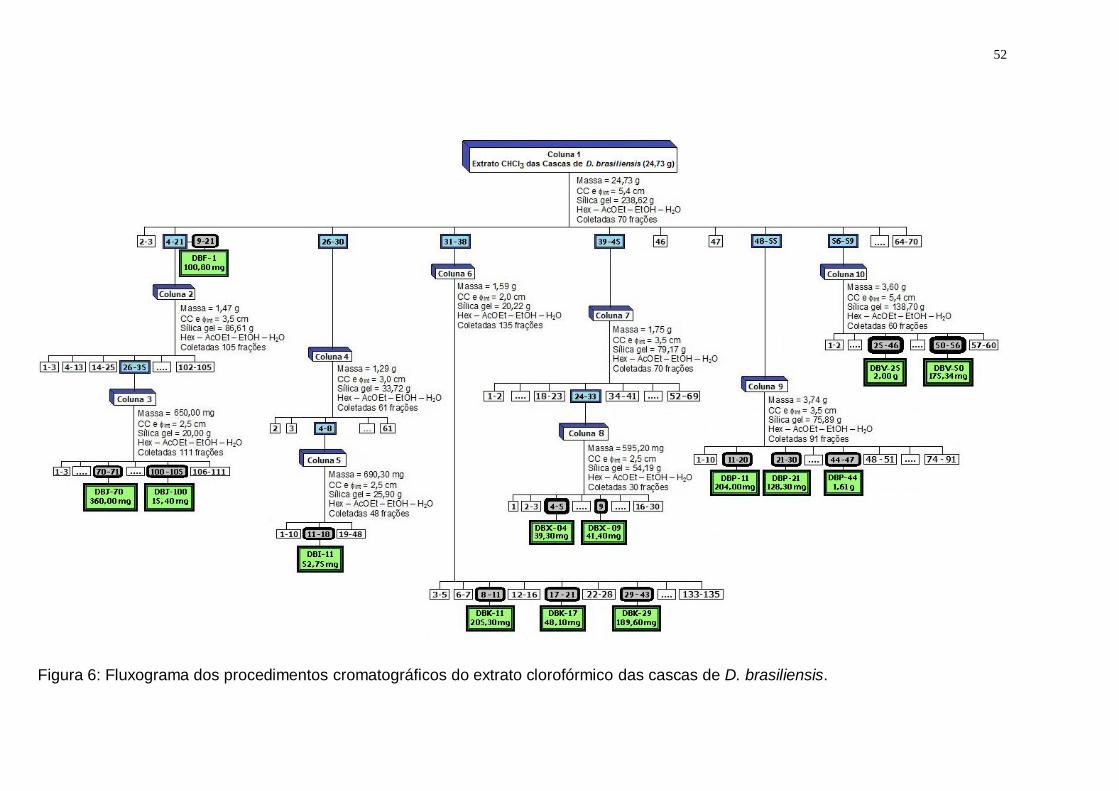
52
Figura 6: Fluxograma dos procedimentos cromatográficos do extrato clorofórmico das cascas de D. brasiliensis.

53
4.6 Atividade biológica
4.6.1 Avaliação da citotoxicidade
A citotoxicidade dos extratos e substâncias isoladas foram avaliadas nas
células K562 (Leucemia mielóide / eritroleucemia Ph+), Nalm6 (Leucemia Linfóide
Aguda B), MCF-7 (adenocarcinoma de mama humano (Homo sapiens) (BCRJ
CR119)) e B16F10 (melanoma murino (Mus musculus C57BL/6J)). Utilizou-se o
ensaio de redução do azul de tetrazólio ou 3-(4,5-dimetiltiazol-2-il)-2,5-
difeniltetrazólio brometo (MTT) (Sigma) que permite avaliar a viabilidade celular
daquelas que crescem aderidas à superfície das placas. Na figura 7 tem-se um
esquema de redução do MTT, que em meio biológico, ocorre pela enzima
mitocondrial succinato desidrogenase.
N+
N
N
N
N
S
N
NH
N
N
N
SBr-
Figura 7: Metabolização do MTT em sais de formazan em células viáveis.
Os testes de citotoxicidade das células K562, Nalm6, MCF-7 e B16F10 foram
realizados tanto no Centro Integrado de Pesquisa Oncológica da Infância (CIPOI),
UNICAMP sob a colaboração de Alexandre Nowill e Gilberto Carlos Franchi Júnior,
como também foram realizado no Laboratório de Cultivo Celular Animal da UNIVALI
sob a colaboração do Prof. Dr. Rilton Alves de Freitas.
Neste teste, o sal de tetrazólio (MTT), sofre uma redução pela enzima
desidrogenase nas mitocôndrias das células vivas, sendo convertido em sal de
formazan, onde há formação de cristais de cor azulada, que após diluição por
dimetilsulfóxido (DMSO), permite-se a quantificação da viabilidade celular em leitor
de microplacas. A atividade enzimática mitocondrial e a viabilidade celular são

54
proporcionais a intensidade da cor, quanto mais cor mais atividade é gerada
(MOSMANN, 1983).
As células foram plaqueadas, em placas de 96 poços, a 2 x 104 células/poço e
incubadas com 300 µL de meio Dulbecco’s Modified Eagle’s Minimum Essential
Médium (DMEM) suplementado com soro fetal bovino (SFB) a 10% (SFB), 1% de
aminoácidos não essenciais e 2 mM de L-glutamina, por 24h a 37ºC em 5% CO2. A
fim de manter todas as células em um mesmo estágio da divisão celular (Go), o meio
DMEM com SFB foi substituído por 180 μL de meio DMEM sem SFB e incubado por
1h a 37ºC em 5% CO2. Posteriormente, foram adicionadas 20 μL das substâncias
testes nas diferentes concentrações (0,01; 0,1; 1,00; 10, 100 e 1000 μg/mL), do
controle positivo DMSO e do controle negativo (meio DMEM sem SFB) e incubadas
por 48h a 37ºC em 5% CO2.
Após este tempo foi retirado o meio e adicionado 270 L de DMEM sem SFB
e 30 µl de MTT (5 mg/mL) e incubadas por 4h a 37ºC em 5% CO2. O meio
juntamente com o MTT foi retirado, e os cristais de formazan insolúveis foram
dissolvidos em 100 µL de DMSO. A determinação da absorbância foi realizada em
leitor de microplaca a 570 nm (MOSMANN, 1983). A viabilidade relativa das células
relacionadas ao controle sem as substâncias testes foi calculada conforme equação
1.
A utilização de fármacos antitumorais na avaliação da viabilidade celular foi
realizada para fins comparativos em relação aos extratos e substâncias isoladas. A
Vincristina e o Paclitaxel foram testados em apenas células K562, Nalm6 e B16F10.
O DMSO foi utilizado como controle interno positivo, enquanto o meio de cultura
DMEM sem SFB foi utilizado como controle negativo, com viabilidade celular igual a
100%.
Viabilidade relativa = Absorbância da amostra teste x 100 Equação 1
Absorbância do controle
4.6.2 Avaliação da atividade leishmanicida
Os testes de atividade leishmanicida em culturas in vitro das linhagens das
formas promastigotas de L. amazonensis (PH8), L. brasiliensis (M2903) e L.
donovani (PP75) foram realizados no Instituto de Investigaciones Fármaco

55
Bioquímicas (IIFB/UMSA), La Paz, Bolívia sob a colaboração da equipe do Dr.
Alberto Giménez, Dr. Efraín Salamanca e Dra. Graice Ruiz.
As culturas in vitro dos parasitas foram cultivadas em 26ºC em meio
Schineider suplementado com 5% de inativos (56ºC x 30 min) em SFB. Parasitas na
fase logarítmica de crescimento, na concentração de 1 x 106 parasitas/mL, foram
distribuídas em placas de 96 poços em diferentes concentrações das substâncias
(100, 50 e 25 μg/mL) foram adicionados. Pentamidina e a Glucantime (50-100
μg/mL) foram utilizadas como fármacos de referência durante as avaliações que
foram feitas em triplicata. A concentração inibitória CI50 das substâncias ativas foi
determinado pela interpolação linear da curva típica obtida plotando o logaritmo da
concentração versus o percentual de inibição, usando como programas de
informática para o Logit, o Probit e análise da regressão polinomial.
4.6.3 Avaliação da atividade plasmodicida
Os testes de atividade plasmodicida em culturas in vitro das linhagens das
formas trofozoítas de Plasmodium falciparum (F32-Tanzânia, linhagem sensível a
Cloroquina, gentilmente cedido pelo Dr. Fandeur T., do Instituto Pasteur, Kênia)
foram realizados no Instituto de Investigaciones Fármaco Bioquímicas (IIFB/UMSA),
La Paz, Bolívia sob a colaboração da equipe do Dr. Alberto Giménez, Dr. Efraín
Salamanca e Dra. Graice Ruiz.
Os parasitas foram cultivados em meio Instituto Roswell Park Memorial
(RPMI) 1640 suplementado com 10% de soro humano e 4% de hematócrito (sangue
do grupo O, RH+) a 37ºC em meio anaeróbico (TRAGER; JENSEN, 1976). Os
parasitas foram sincronizados até 1% de parasistemia e 2% de hematócrito e foram
distribuídos no volume de 100 μL em placas de 96 poços em duplicata. 100 μL de
cada diluição da substância (em DMSO, com no máximo 0,1% na concentração
final) foram adicionados nos poços. Em seguida os parasitas foram encubados a
37ºC por 48 horas. Um esfregaço foi fixado em CH3OH e corado com Giemsa. A
atividade antiplasmodial foi determinada pela observação no microscópio expressos
em percentual de inibição (% Inh.) de acordo com a equação 2:

56
% Inh. = (GRL - GRI) X 100 Equação 2
GRL
GRL = glóbulos vermelhos não-infectados;
GRI = células infectadas.
A concentração de inibição de 50% em maturação esquizonte (CI50) foi
realizada através do método gráfico usando o programa CRIKET GRAPH 1.3, sobre
os ativos que apresentavam um CI50 < 10 μg/mL.
4.7 Análise estatística
A CI50 foi determinada pelo método de linearização de PROBITOS das
viabilidades celulares em função da concentração ou aproximada através da
regressão sigmoidal utilizando o software Origin 5.0, para o experimento com
células, utilizando-se em função do log das concentrações aplicadas.
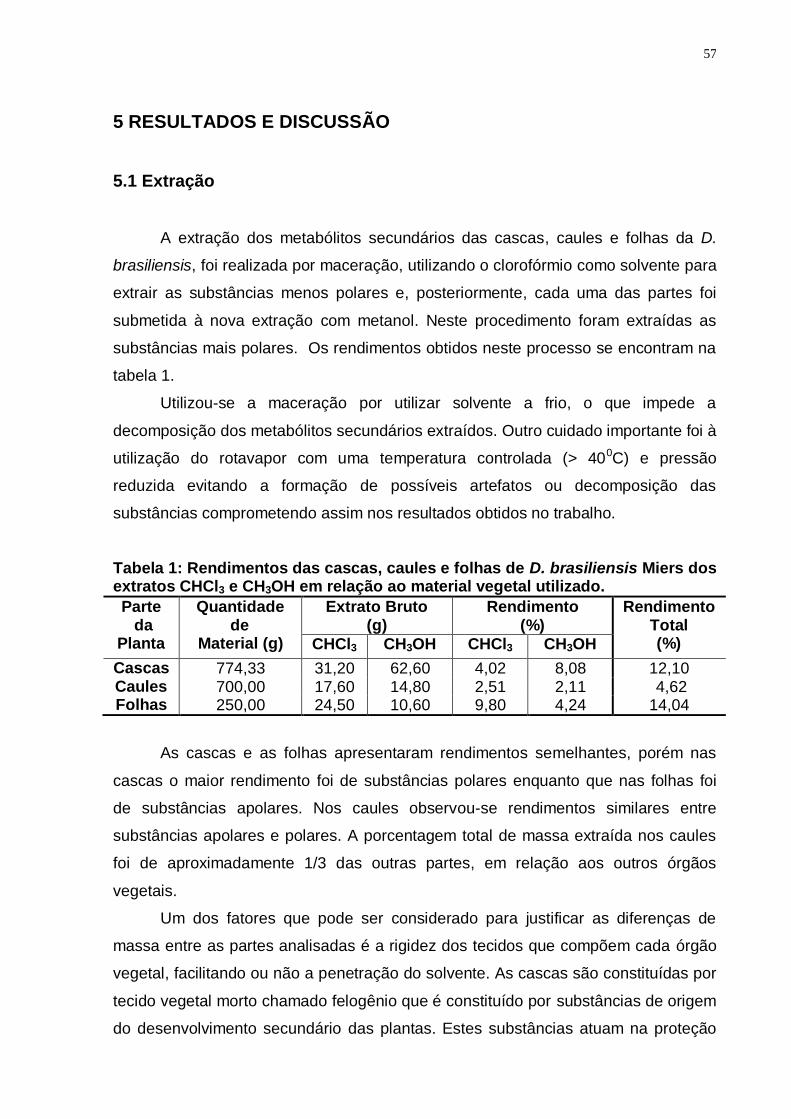
57
5 RESULTADOS E DISCUSSÃO
5.1 Extração
A extração dos metabólitos secundários das cascas, caules e folhas da D.
brasiliensis, foi realizada por maceração, utilizando o clorofórmio como solvente para
extrair as substâncias menos polares e, posteriormente, cada uma das partes foi
submetida à nova extração com metanol. Neste procedimento foram extraídas as
substâncias mais polares. Os rendimentos obtidos neste processo se encontram na
tabela 1.
Utilizou-se a maceração por utilizar solvente a frio, o que impede a
decomposição dos metabólitos secundários extraídos. Outro cuidado importante foi à
utilização do rotavapor com uma temperatura controlada (> 400C) e pressão
reduzida evitando a formação de possíveis artefatos ou decomposição das
substâncias comprometendo assim nos resultados obtidos no trabalho.
Tabela 1: Rendimentos das cascas, caules e folhas de D. brasiliensis Miers dos extratos CHCl3 e CH3OH em relação ao material vegetal utilizado.
Parte da
Planta
Quantidade de
Material (g)
Extrato Bruto (g)
Rendimento (%)
Rendimento Total (%) CHCl3 CH3OH CHCl3 CH3OH
Cascas 774,33 31,20 62,60 4,02 8,08 12,10 Caules 700,00 17,60 14,80 2,51 2,11 4,62 Folhas 250,00 24,50 10,60 9,80 4,24 14,04
As cascas e as folhas apresentaram rendimentos semelhantes, porém nas
cascas o maior rendimento foi de substâncias polares enquanto que nas folhas foi
de substâncias apolares. Nos caules observou-se rendimentos similares entre
substâncias apolares e polares. A porcentagem total de massa extraída nos caules
foi de aproximadamente 1/3 das outras partes, em relação aos outros órgãos
vegetais.
Um dos fatores que pode ser considerado para justificar as diferenças de
massa entre as partes analisadas é a rigidez dos tecidos que compõem cada órgão
vegetal, facilitando ou não a penetração do solvente. As cascas são constituídas por
tecido vegetal morto chamado felogênio que é constituído por substâncias de origem
do desenvolvimento secundário das plantas. Estes substâncias atuam na proteção

58
da planta contra doenças e pragas bem como em processos alelopáticos. Os caules
são os responsáveis pela passagem da seiva bruta e elaborada das raízes para as
folhas, além de sustentação da árvore e produção de folhas. Tem como principal
função o transporte dos nutrientes. Já as folhas são as responsáveis em captar a
energia solar e utilizá-la na fotossíntese para a produção das mais variadas
substâncias de que a planta precise. Desempenham importantes papéis
antialimentares e alelopáticos (GLORIA e GUERREIRO, 2003).
Quanto maior a fragmentação do material vegetal, maior será a superfície de
contato e consequentemente maior será o poder de penetração do solvente para a
dissolução dos princípios ativos vegetais, consequentemente maior será o
rendimento do processo extrativo.
5.2 Fracionamento do extrato CHCl3 das cascas de Drimys brasiliensis
Os extratos brutos das cascas, caules e folhas foram avaliados por CCD para
a verificação dos seus respectivos perfis cromatográficos. Foi observado que o perfil
cromatográfico dos extratos eram bem distintos entre si. Com base em estudos
anteriores utilizando o extrato das cascas, foi detectada a presença dos
sesquiterpenos drimanos, a principal classe de interesse, devido aos efeitos
farmacológicos promissores (MORTON, 1981; SIMÕES, et al., 1986; CECHINEL
FILHO, et al., 1998; MENDES, et al., 1998; CUNHA, et al., 2001; MALHEIROS,
2001; ANDRÉ, et al., 2004; JANSEN e GROOT, 2004; MALHEIROS, et al., 2005).
Neste aspecto, o extrato CHCl3 das cascas foi o selecionado para os processos
cromatográficos. Este então foi submetido a sucessivas colunas cromatográficas, e
as substâncias puras foram analisadas em RMN 1H e RMN 13C.
Para um melhor entendimento nos processos cromatográficos realizados,
optou-se apresentar um breve resumo das colunas, bem como as substâncias
isoladas, separando-as apenas por diferença de polaridade e classe estrutural.
Nas figuras 8, 15, 23 e 32 nas páginas 61, 68, 77 e 87, respectivamente,
estão demonstrados os procedimentos cromatográficos que foram realizados para o
isolamento das moléculas bioativas pertencentes a diferentes classes de metabólitos
secundários.
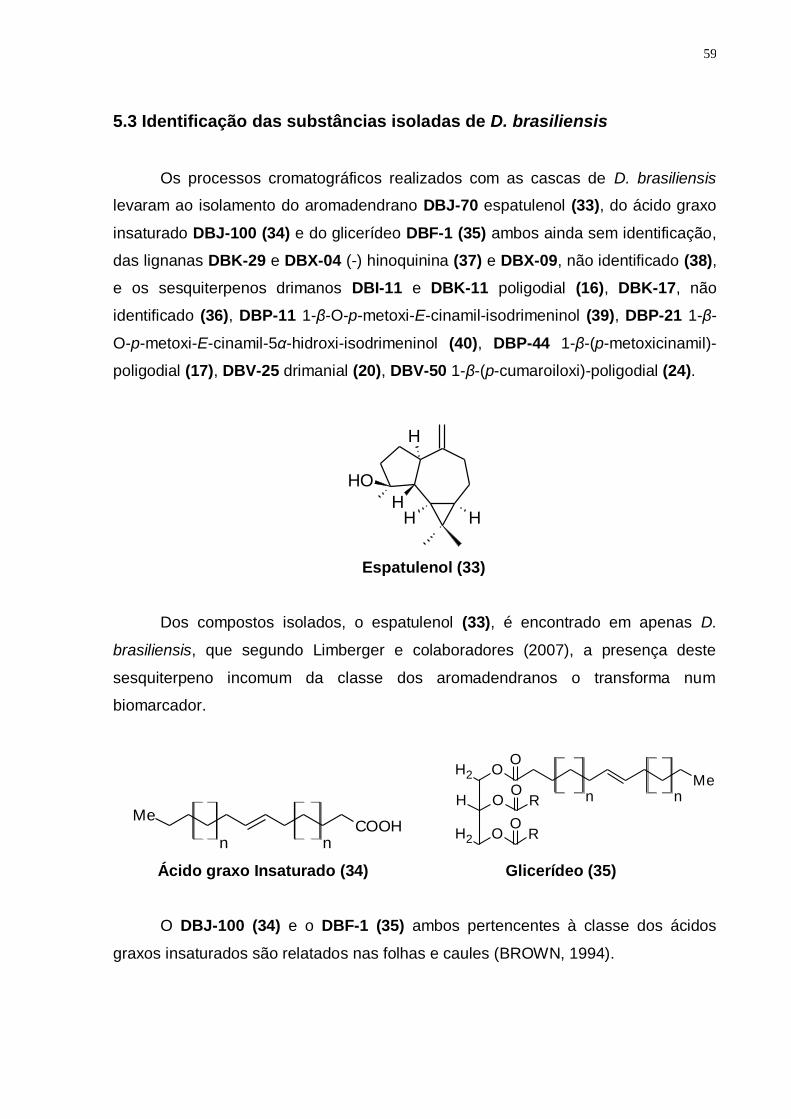
59
5.3 Identificação das substâncias isoladas de D. brasiliensis
Os processos cromatográficos realizados com as cascas de D. brasiliensis
levaram ao isolamento do aromadendrano DBJ-70 espatulenol (33), do ácido graxo
insaturado DBJ-100 (34) e do glicerídeo DBF-1 (35) ambos ainda sem identificação,
das lignanas DBK-29 e DBX-04 (-) hinoquinina (37) e DBX-09, não identificado (38),
e os sesquiterpenos drimanos DBI-11 e DBK-11 poligodial (16), DBK-17, não
identificado (36), DBP-11 1-β-O-p-metoxi-E-cinamil-isodrimeninol (39), DBP-21 1-β-
O-p-metoxi-E-cinamil-5α-hidroxi-isodrimeninol (40), DBP-44 1-β-(p-metoxicinamil)-
poligodial (17), DBV-25 drimanial (20), DBV-50 1-β-(p-cumaroiloxi)-poligodial (24).
H
HHH
OH
Espatulenol (33)
Dos compostos isolados, o espatulenol (33), é encontrado em apenas D.
brasiliensis, que segundo Limberger e colaboradores (2007), a presença deste
sesquiterpeno incomum da classe dos aromadendranos o transforma num
biomarcador.
n n
MeCOOH
n nMe
OO
H2
ROO
H
ROO
H2
Ácido graxo Insaturado (34) Glicerídeo (35)
O DBJ-100 (34) e o DBF-1 (35) ambos pertencentes à classe dos ácidos
graxos insaturados são relatados nas folhas e caules (BROWN, 1994).
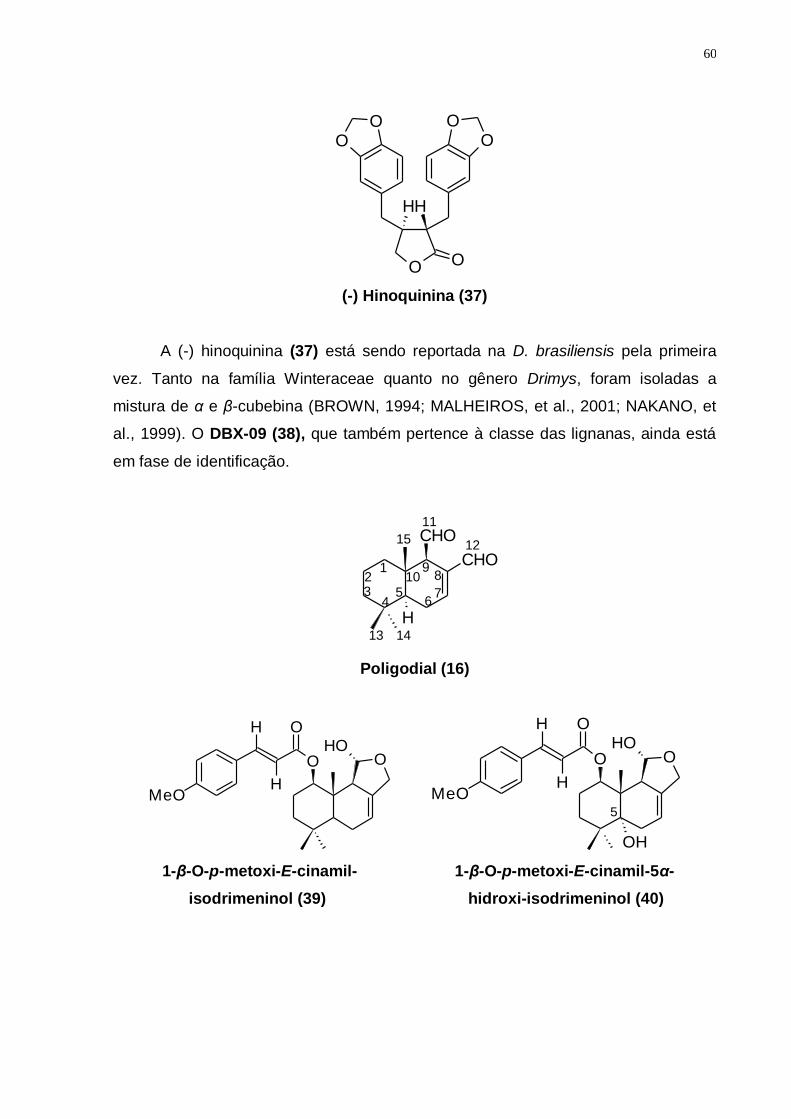
60
OOO
O
O O
HH
(-) Hinoquinina (37)
A (-) hinoquinina (37) está sendo reportada na D. brasiliensis pela primeira
vez. Tanto na família Winteraceae quanto no gênero Drimys, foram isoladas a
mistura de α e β-cubebina (BROWN, 1994; MALHEIROS, et al., 2001; NAKANO, et
al., 1999). O DBX-09 (38), que também pertence à classe das lignanas, ainda está
em fase de identificação.
CHO
CHO
H
123
45
67
89
11
12
10
13 14
15
Poligodial (16)
O
O
MeO
OOH
H
H
O
O
MeO
OH
OOH
H
H
5
1-β-O-p-metoxi-E-cinamil- 1-β-O-p-metoxi-E-cinamil-5α-
isodrimeninol (39) hidroxi-isodrimeninol (40)
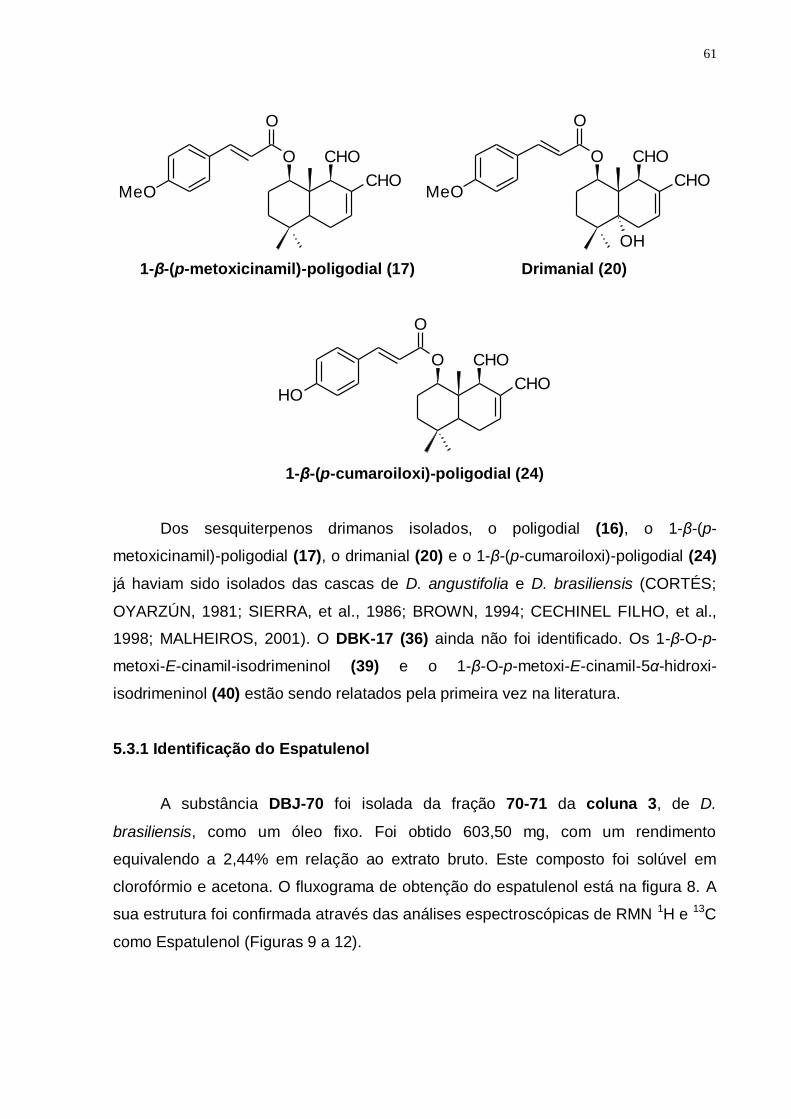
61
O
O
MeO
CHO
CHO
O
O
MeO
CHO
CHO
OH
1-β-(p-metoxicinamil)-poligodial (17) Drimanial (20)
O
O
OH
CHO
CHO
1-β-(p-cumaroiloxi)-poligodial (24)
Dos sesquiterpenos drimanos isolados, o poligodial (16), o 1-β-(p-
metoxicinamil)-poligodial (17), o drimanial (20) e o 1-β-(p-cumaroiloxi)-poligodial (24)
já haviam sido isolados das cascas de D. angustifolia e D. brasiliensis (CORTÉS;
OYARZÚN, 1981; SIERRA, et al., 1986; BROWN, 1994; CECHINEL FILHO, et al.,
1998; MALHEIROS, 2001). O DBK-17 (36) ainda não foi identificado. Os 1-β-O-p-
metoxi-E-cinamil-isodrimeninol (39) e o 1-β-O-p-metoxi-E-cinamil-5α-hidroxi-
isodrimeninol (40) estão sendo relatados pela primeira vez na literatura.
5.3.1 Identificação do Espatulenol
A substância DBJ-70 foi isolada da fração 70-71 da coluna 3, de D.
brasiliensis, como um óleo fixo. Foi obtido 603,50 mg, com um rendimento
equivalendo a 2,44% em relação ao extrato bruto. Este composto foi solúvel em
clorofórmio e acetona. O fluxograma de obtenção do espatulenol está na figura 8. A
sua estrutura foi confirmada através das análises espectroscópicas de RMN 1H e 13C
como Espatulenol (Figuras 9 a 12).

62
Figura 8: Fluxograma dos procedimentos cromatográficos do extrato clorofórmico de D. brasiliensis mostrando o isolamento do Espatulenol.
O espectro de RMN 1H (Figura 9 e 10) mostra sinais característicos de anel
ciclopropânico em δH 0,46 (dd, J = 11,3 e 9,6 Hz) referente ao H-6 e em δH 0,72 (m)
referente ao H-7. Estes sinais são indicativos de esqueleto aromadendrano. Os
sinais apresentam em regiões baixas devido ao efeito de blindagem exercido pelo
anel ciclopropânico. O sinal em δH 4,68 (d, J = 7,2 Hz) é referente ao hidrogênio da
dupla ligação ligado ao anel cicloheptânico H-14. Os hidrogênios ligados a uma
ligação dupla (hidrogênio vinila) são desblindados pela anisotropia da ligação dupla
adjacente. Os hidrogênios das metilas H-12, H-13 e H-15 estão em δH 1,25, 1,28 e
1,05, respectivamente. Os hidrogênios metilênicos encontram-se de δH 2,50 a 1,50.
Estes se encontram desblindados devido à proximidade com o oxigênio e a dupla
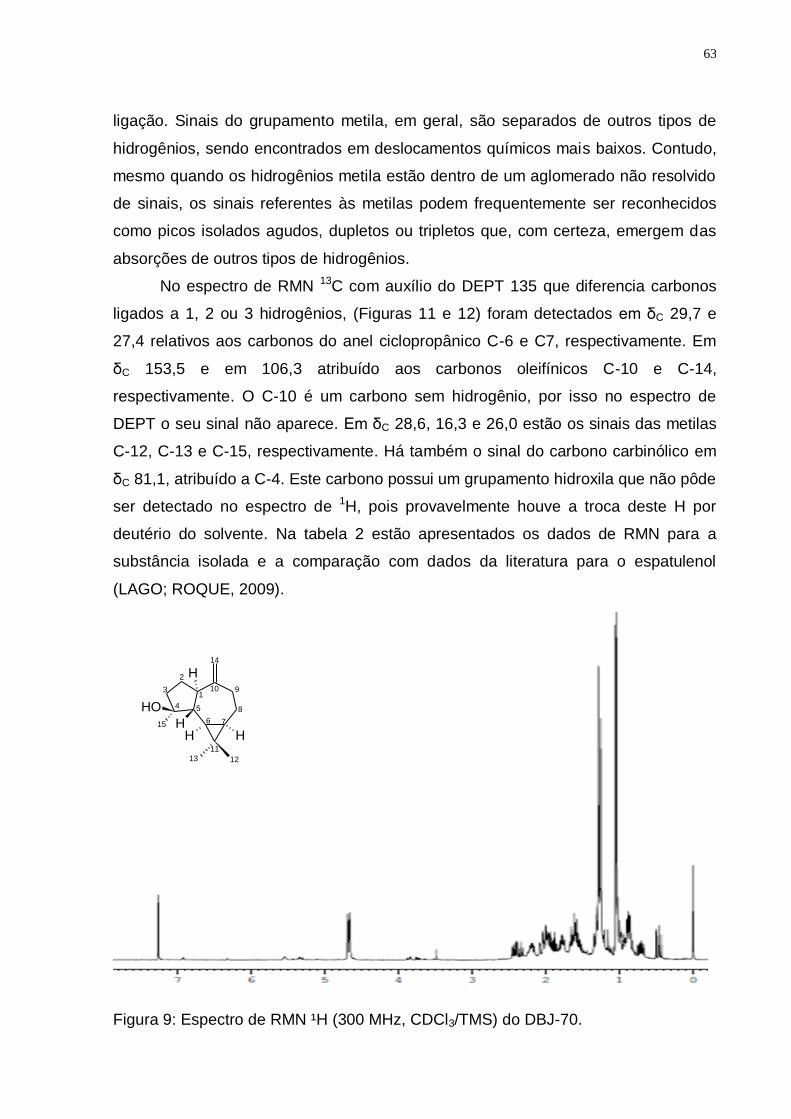
63
ligação. Sinais do grupamento metila, em geral, são separados de outros tipos de
hidrogênios, sendo encontrados em deslocamentos químicos mais baixos. Contudo,
mesmo quando os hidrogênios metila estão dentro de um aglomerado não resolvido
de sinais, os sinais referentes às metilas podem frequentemente ser reconhecidos
como picos isolados agudos, dupletos ou tripletos que, com certeza, emergem das
absorções de outros tipos de hidrogênios.
No espectro de RMN 13C com auxílio do DEPT 135 que diferencia carbonos
ligados a 1, 2 ou 3 hidrogênios, (Figuras 11 e 12) foram detectados em δC 29,7 e
27,4 relativos aos carbonos do anel ciclopropânico C-6 e C7, respectivamente. Em
δC 153,5 e em 106,3 atribuído aos carbonos oleifínicos C-10 e C-14,
respectivamente. O C-10 é um carbono sem hidrogênio, por isso no espectro de
DEPT o seu sinal não aparece. Em δC 28,6, 16,3 e 26,0 estão os sinais das metilas
C-12, C-13 e C-15, respectivamente. Há também o sinal do carbono carbinólico em
δC 81,1, atribuído a C-4. Este carbono possui um grupamento hidroxila que não pôde
ser detectado no espectro de 1H, pois provavelmente houve a troca deste H por
deutério do solvente. Na tabela 2 estão apresentados os dados de RMN para a
substância isolada e a comparação com dados da literatura para o espatulenol
(LAGO; ROQUE, 2009).
Figura 9: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBJ-70.
H
HHH
OH1
2
3
4 5
6 7
8
910
11
1213
14
15
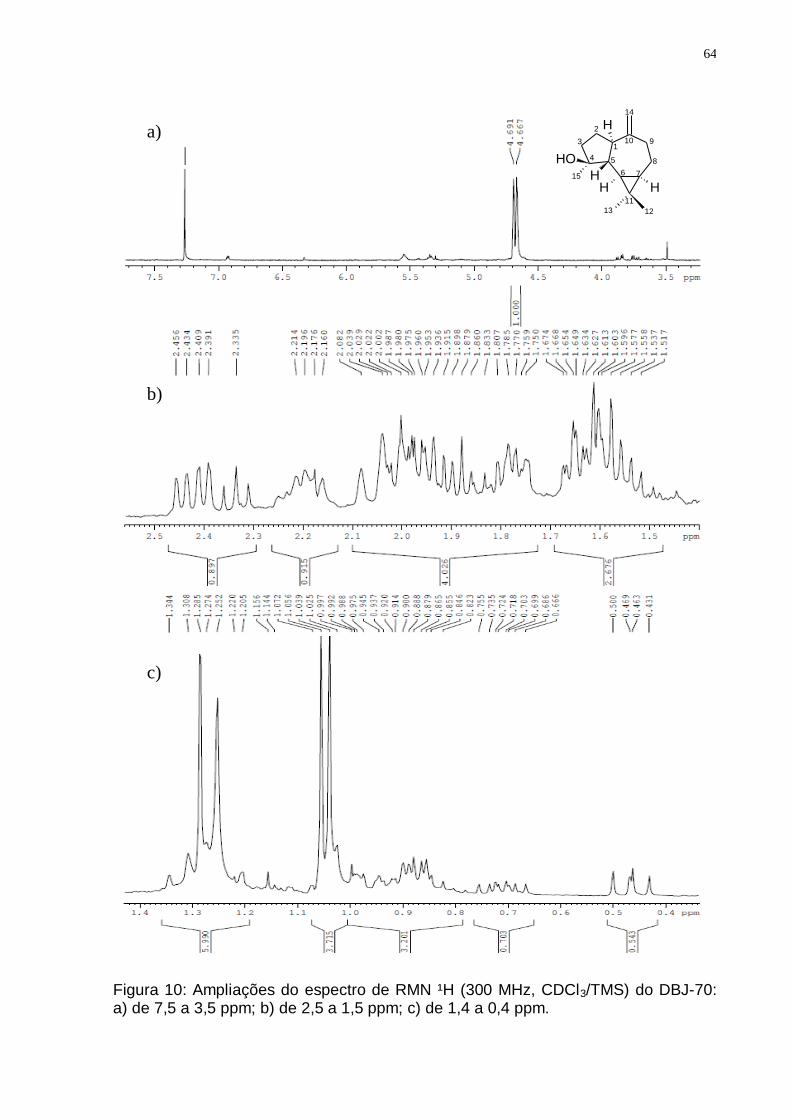
64
Figura 10: Ampliações do espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBJ-70: a) de 7,5 a 3,5 ppm; b) de 2,5 a 1,5 ppm; c) de 1,4 a 0,4 ppm.
a)
b)
c)
H
HHH
OH1
2
3
4 5
6 7
8
910
11
1213
14
15
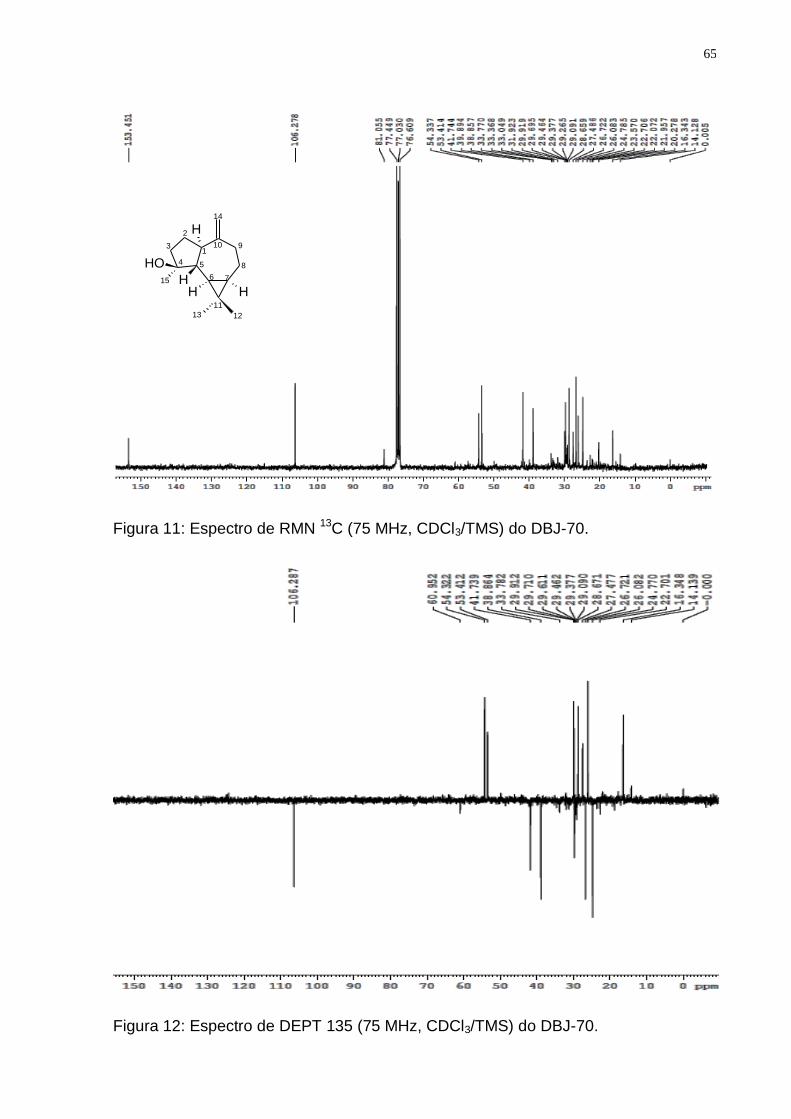
65
Figura 11: Espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBJ-70.
Figura 12: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBJ-70.
H
HHH
OH1
2
3
4 5
6 7
8
910
11
1213
14
15

66
Tabela 2: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBJ-70 comparando com dados da literatura para o Espatulenol.
H
HHH
OH1
2
3
4 5
6 7
8
910
11
1213
14
Posição
DBJ-70 1H (ppm) mult. J (Hz)
Espatulenol* 1H (ppm) mult. J (Hz)
DBJ-70 13
C / DEPT
Espatulenol* 13
C / DEPT
1 2,42, dd, (J= 14,1; 6,6) 2,39, dd, (J= 14,1; 6,3) 53,4 / CH 53,4 / CH
2 26,7 / CH2 26,7 / CH2
3 41,7 / CH2 41,8 / CH2
4 81,1 / C 81,0 / C
5 54,3 / CH 54,4 / CH
6 0,46, dd, (J = 11,3; 9,6) 0,46, dd, (J = 11,2; 9,5) 29,9 / CH 29,9 / CH
7 0,70, m 0,72, ddd, (J = 11,2; 9,5;
6,1)
27,5 / CH 27,5 / CH
8 24,8 / CH2 24,8 / CH2
9 38,9 / CH2 38,9 / CH2
10 153,5 / C 153,5 / C
11 20,3 / C 20,3 / C
12 1,25, s 1,25, s 28,7 / CH3 28,7 / CH3
13 1,28, s 1,28, s 16,3 / CH3 16,3 / CH3
14 4,69 / 4,68, d, (J = 7,2) 4,69 / 4,66, s 106,3 / CH2 106,3 / CH2
15 1,05, d, (J = 5,1) 1,06, s 26,1 / CH3 26,1 / CH3
Solvente CDCl3
300 MHz
CDCl3
200 MHz
CDCl3
75 MHz
CDCl3
50 MHz
*(LAGO; ROQUE, 2009).
Os aromadendranos derivam dos terpenóides. O precursor farnesil pirofosfato
pode formar tanto sesquiterpenos lineares quanto cíclicos devido ao aumento da sua
cadeia e das ligações duplas adicionais que realiza, permitindo assim um aumento
na flexibilidade da cadeia e a formação de um maior número de esqueletos. Estas
ciclizações podem dar origem a estruturas mono, di, ou tricíclicas. Também pode
haver mudança na estereoquímica da dupla ligação, podendo esta adotar a
configuração E (como no farnesil pirofosfato) ou Z por via da ionização como no
caso do geranil/neril pirofosfato (DEWICK, 2009).
Deste precursor diversos grupos são formados, de acordo com o tipo de
ciclização dos anéis (figura 13).
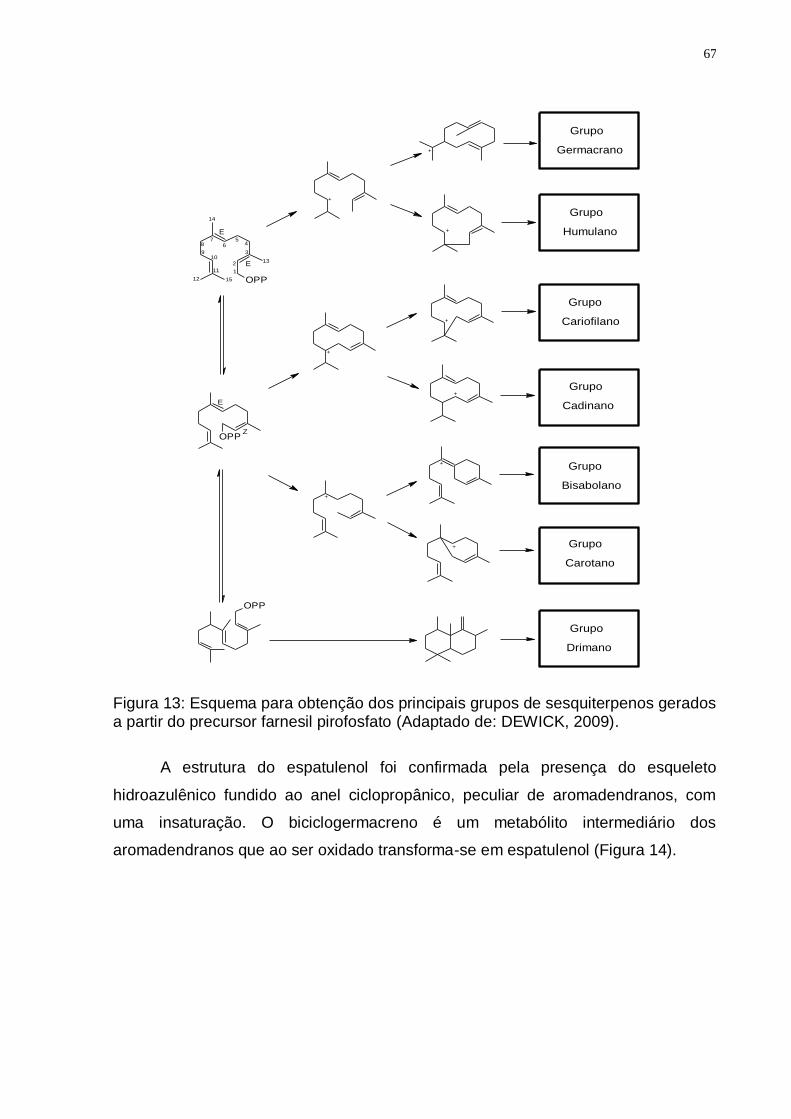
67
OPP
OPP
+
+
+
+
+
+
OPP
+
+
+
2
3
1
56
78
910
11
12
13
14
15
4
E
E
E
Z
Grupo
Germacrano
Grupo
Humulano
Grupo
Cariofilano
Grupo
Cadinano
Grupo
Bisabolano
Grupo
Carotano
Grupo
Drimano
Figura 13: Esquema para obtenção dos principais grupos de sesquiterpenos gerados a partir do precursor farnesil pirofosfato (Adaptado de: DEWICK, 2009).
A estrutura do espatulenol foi confirmada pela presença do esqueleto
hidroazulênico fundido ao anel ciclopropânico, peculiar de aromadendranos, com
uma insaturação. O biciclogermacreno é um metabólito intermediário dos
aromadendranos que ao ser oxidado transforma-se em espatulenol (Figura 14).

68
H
H
OH
H
HHH
OH
H
HOH
H
H
Germacreno-D Biciclogermacreno Macrocarp-11(15)-en-8-ol
Espatulenol
Figura 14: Provável rota biossintética do sesquiterpeno Espatulenol. (Adaptado de CHAVES, et al., 2003).
Os aromadendranos apresentam diversas atividades biológicas como:
antifúngica, antibacteriana e toxicidade (LIMA; BEATRIZ; RAMOS, 1997; MOREIRA,
et al., 2003), antitripanocida, antiviral, regulação no crescimento de plantas e
citotóxica (ATAWODI; ALAFIATAYO, 2007). Perez-Hernandez e colaboradores
(2008) avaliaram o efeito espasmódico em útero de ratas e demonstraram que o
espatulenol foi o terpeno mais potente. Ele possui um efeito relaxante direto sobre a
contração tônica induzida por soluções despolarizantes da musculatura lisa do útero
de forma dependente, impedindo a contração causada pelo acúmulo de Ca2+. O
efeito espasmolítico do espatulenol não depende de NO e/ou receptores
adrenérgicos, pois ele atua diretamente nos canais de cálcio sensíveis a voltagem-
dependente.
Tanto na família Winteraceae quanto no gênero Drimys são relatados
substâncias contendo o esqueleto aromadendrano. O biciclogermacreno e a
ciclocolorenona estão presentes no óleo essencial das folhas frescas e das cascas
de D. angustifolia (LIMBERGER et al., 2007), no óleo essencial de cascas de D.
winteri (CICCIÓ, 1997; BARRERO, et al., 2000) e no óleo das folhas e dos frutos de
D. granadensis (CICCIÓ, 1997). O óleo das folhas de D. brasiliensis mostrou-se rico
em ciclocolorenona, um sesquiterpeno incomum da classe dos aromadendranos
(SCHMIDT, et al., 1999). A presença do espatulenol apenas na D. brasiliensis o
transforma num candidato a biomarcador para o controle de matéria prima vegetal,

69
tendo em vista o alto rendimento em relação ao extrato bruto e sua excelente
atividade farmacológica.
5.3.2 Identificação do Ácido Graxo Insaturado
A substância DBJ-100 foi isolada da fração 100-105 da coluna 3, de D.
brasiliensis, como um óleo fixo. Foi obtido aproximadamente 15,40 mg, com um
rendimento equivalendo a 0,06% em relação ao extrato bruto. Este foi solúvel em
diclorometano e acetona. O fluxograma de obtenção do ácido graxo está na figura
15. A estrutura foi identificada como um derivado de ácido graxo insaturado (Figuras
16 e 17).
Figura 15: Fluxograma dos procedimentos cromatográficos do extrato clorofórmico de D. brasiliensis mostrando o isolamento do Ácido Graxo Insaturado e do Glicerídeo.
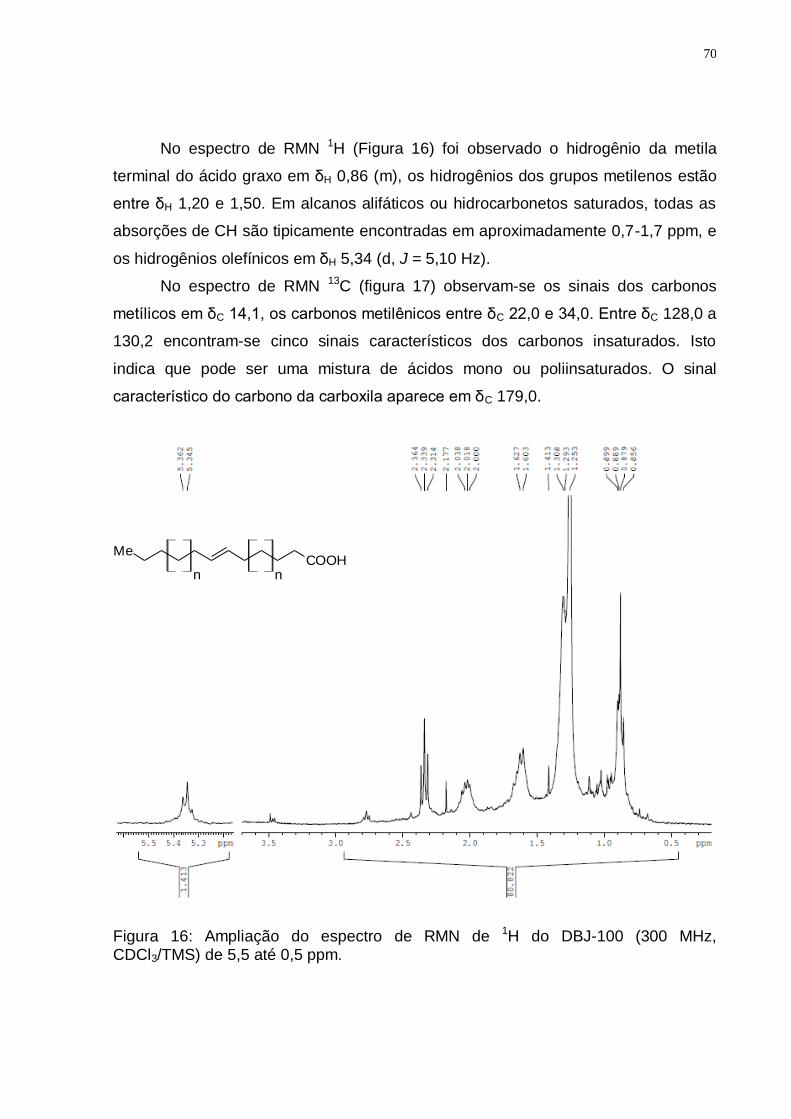
70
No espectro de RMN 1H (Figura 16) foi observado o hidrogênio da metila
terminal do ácido graxo em δH 0,86 (m), os hidrogênios dos grupos metilenos estão
entre δH 1,20 e 1,50. Em alcanos alifáticos ou hidrocarbonetos saturados, todas as
absorções de CH são tipicamente encontradas em aproximadamente 0,7-1,7 ppm, e
os hidrogênios olefínicos em δH 5,34 (d, J = 5,10 Hz).
No espectro de RMN 13C (figura 17) observam-se os sinais dos carbonos
metílicos em δC 14,1, os carbonos metilênicos entre δC 22,0 e 34,0. Entre δC 128,0 a
130,2 encontram-se cinco sinais característicos dos carbonos insaturados. Isto
indica que pode ser uma mistura de ácidos mono ou poliinsaturados. O sinal
característico do carbono da carboxila aparece em δC 179,0.
Figura 16: Ampliação do espectro de RMN de 1H do DBJ-100 (300 MHz, CDCl3/TMS) de 5,5 até 0,5 ppm.
n n
MeCOOH
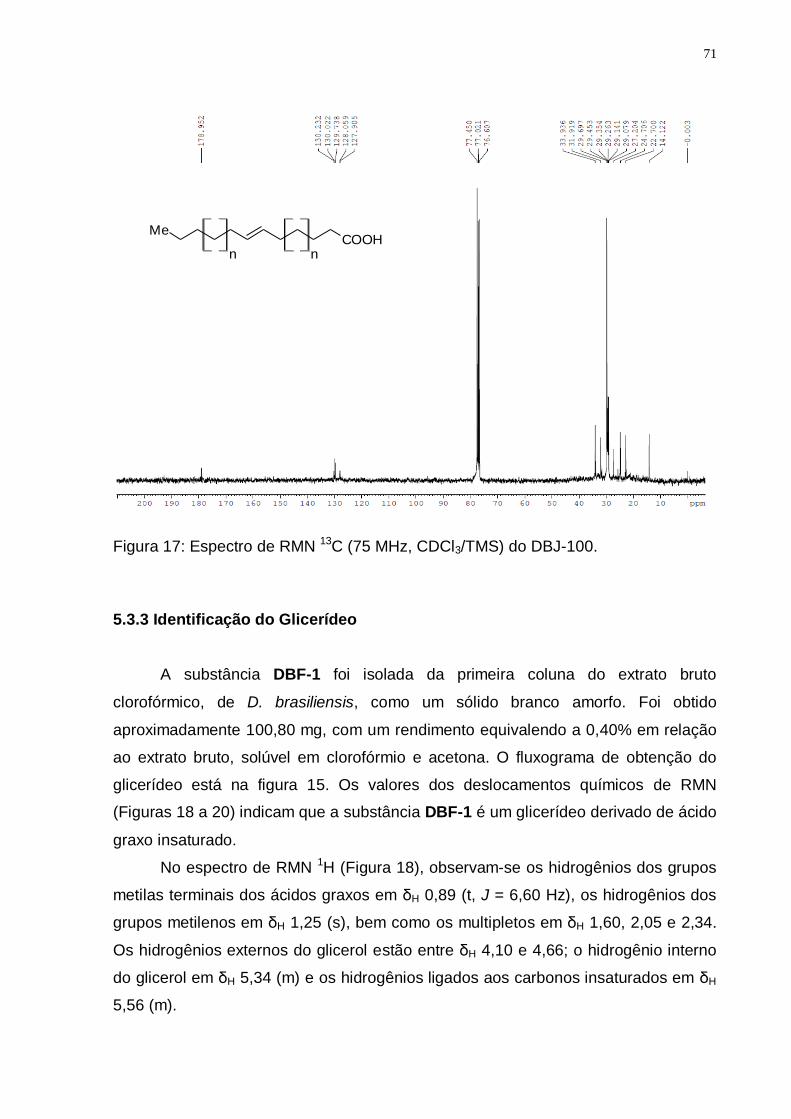
71
Figura 17: Espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBJ-100.
5.3.3 Identificação do Glicerídeo
A substância DBF-1 foi isolada da primeira coluna do extrato bruto
clorofórmico, de D. brasiliensis, como um sólido branco amorfo. Foi obtido
aproximadamente 100,80 mg, com um rendimento equivalendo a 0,40% em relação
ao extrato bruto, solúvel em clorofórmio e acetona. O fluxograma de obtenção do
glicerídeo está na figura 15. Os valores dos deslocamentos químicos de RMN
(Figuras 18 a 20) indicam que a substância DBF-1 é um glicerídeo derivado de ácido
graxo insaturado.
No espectro de RMN 1H (Figura 18), observam-se os hidrogênios dos grupos
metilas terminais dos ácidos graxos em δH 0,89 (t, J = 6,60 Hz), os hidrogênios dos
grupos metilenos em δH 1,25 (s), bem como os multipletos em δH 1,60, 2,05 e 2,34.
Os hidrogênios externos do glicerol estão entre δH 4,10 e 4,66; o hidrogênio interno
do glicerol em δH 5,34 (m) e os hidrogênios ligados aos carbonos insaturados em δH
5,56 (m).
n n
MeCOOH
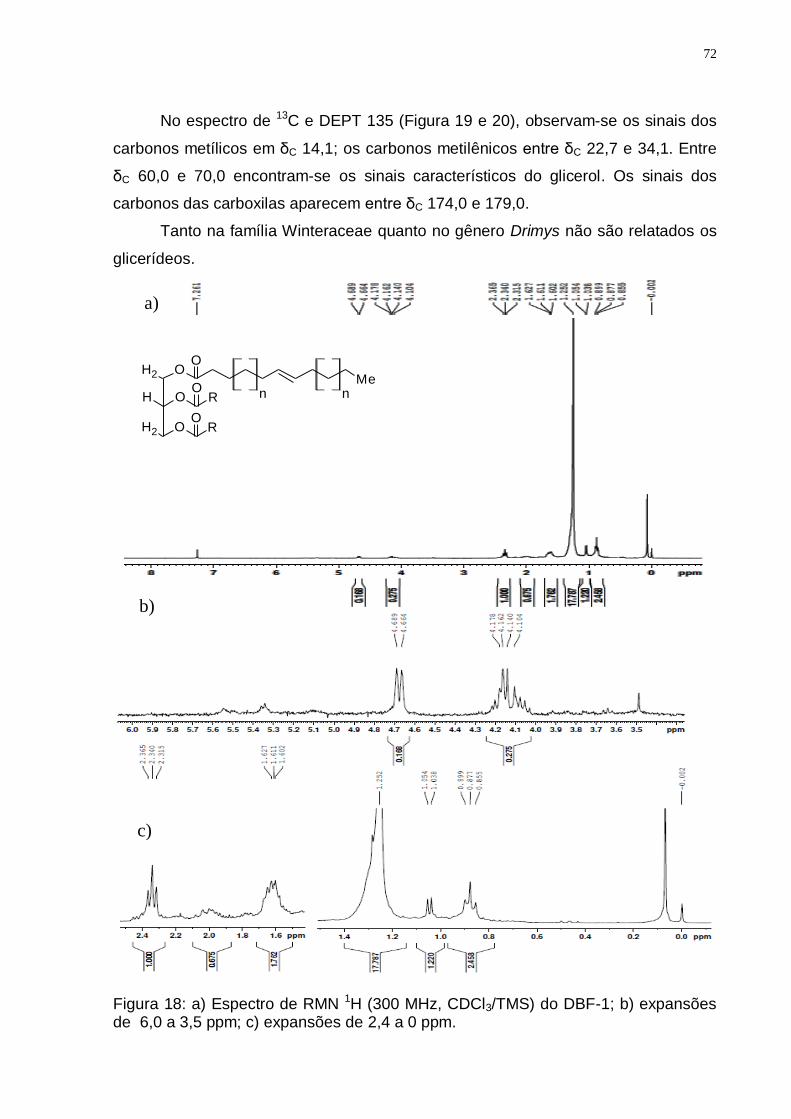
72
No espectro de 13C e DEPT 135 (Figura 19 e 20), observam-se os sinais dos
carbonos metílicos em δC 14,1; os carbonos metilênicos entre δC 22,7 e 34,1. Entre
δC 60,0 e 70,0 encontram-se os sinais característicos do glicerol. Os sinais dos
carbonos das carboxilas aparecem entre δC 174,0 e 179,0.
Tanto na família Winteraceae quanto no gênero Drimys não são relatados os
glicerídeos.
Figura 18: a) Espectro de RMN 1H (300 MHz, CDCl3/TMS) do DBF-1; b) expansões de 6,0 a 3,5 ppm; c) expansões de 2,4 a 0 ppm.
n nMe
OO
H2
ROO
H
ROO
H2
a)
b)
c)
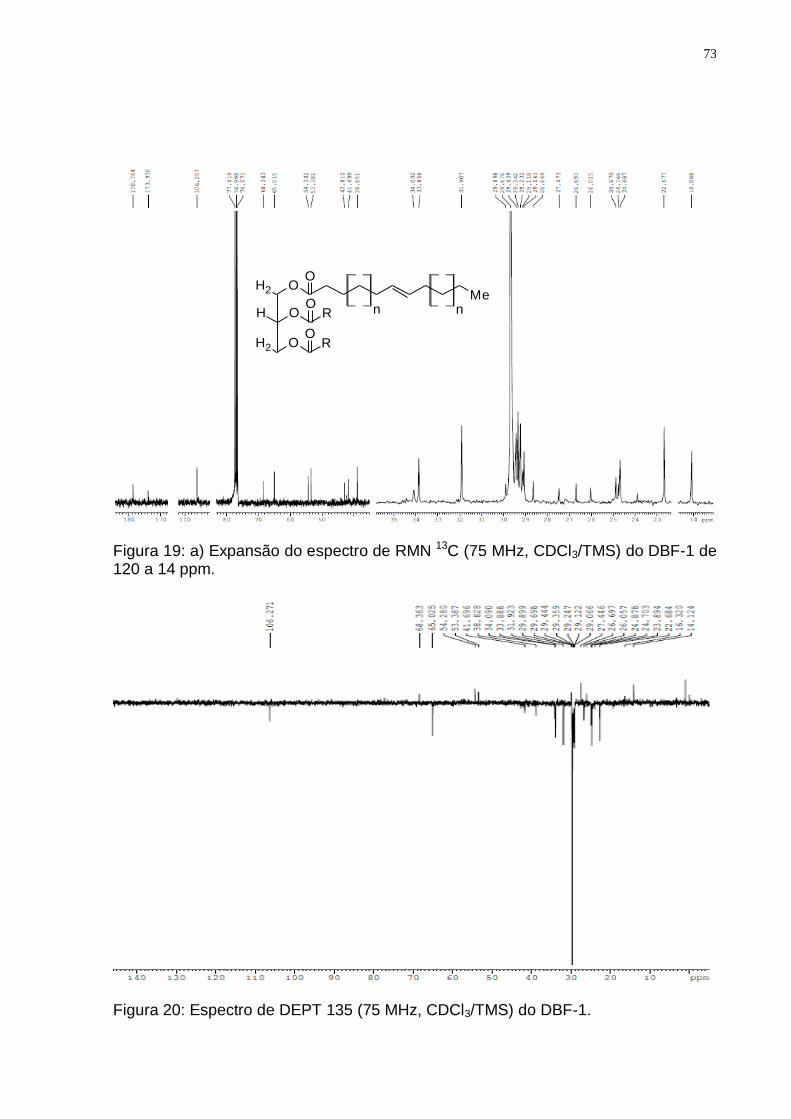
73
Figura 19: a) Expansão do espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBF-1 de 120 a 14 ppm.
Figura 20: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBF-1.
n nMe
OO
H2
ROO
H
ROO
H2

74
Os lipídeos (óleos fixos, gorduras e ceras) são ésteres de ácidos graxos e
alcoóis de cadeia longa, ou derivados afins. A principal diferença entre estas
substâncias é o tipo de álcool; nos óleos fixos e nas gorduras, o glicerol combina-se
com os ácidos graxos, já nas ceras o álcool tem um peso molecular diferente,
influenciando no comportamento da molécula (HORNSTRA, 2002).
Ambos são muito importantes, sendo utilizados principalmente para fins
farmacológicos, industriais e nutricionais. Diferem apenas no ponto de fusão; os que,
em temperatura ambiente são líquidos, recebem o nome de óleos fixos; os que são
sólidos ou semi-sólidos recebem o nome de gorduras. Os óleos e as gorduras
vegetais podem ocorrer em várias partes da planta, mas em vias de regras, as
sementes contêm quantidades maiores de gorduras e óleos que outras partes da
planta (LENINGER; NELSON; COX, 2002).
Os óleos fixos e as gorduras são compostos predominantemente por
triacilgliceróis, que tem ácidos graxos diferentes ou idênticos nas três posições
hidroxila da molécula de glicerol. O nome ácido graxo designa qualquer um dos
ácidos monocarboxílicos alifáticos que podem ser liberados por hidrólise de gorduras
e óleos naturais. Os principais ácidos graxos podem ser saturados ou insaturados,
com uma cadeia de carbono não ramificada de número par. Em geral, os
triacilgliceróis dos ácidos graxos insaturados são líquidos, já os saturados com
comprimento de cadeia suficiente são sólidos. A predominância de um tipo ou de
outro num óleo determina se a mistura é líquida ou sólida. Além dos triglicerídeos,
também existem em menores quantidades os monoacilgliceróis e os diacilgliceróis.
Estes são intermediários na síntese dos triacilgliceróis (VALENZUELA; NIETO, 2001;
2003; GONZÁLES, 2002; HORNSTRA, 2002; SILVA, et al., 2007).
Na biossíntese dos lipídeos (figura 21), as partes de ácido graxo são
formadas a partir de Acetil-CoA e de Malonil-CoA através de várias reações, onde
enzimas individuais catalisam a formação de ácidos graxos nos cloroplastos. Os
intermediários da síntese são ligados covalentemente aos grupos sulfidrila de uma
proteína carreadora de acila (ACP), que carreia todos os intermediários durante a
síntese. A cadeia de ácidos graxos em crescimento é alongada pela adição
consecutiva de duas unidades de carbono, derivadas da acetil-CoA, mas o doador
ativado dessas unidades é a malonil-ACP. A energia que ativa o processo de
alongamento está conjugada a carboxilação da acetil-CoA, dependente de ATP,
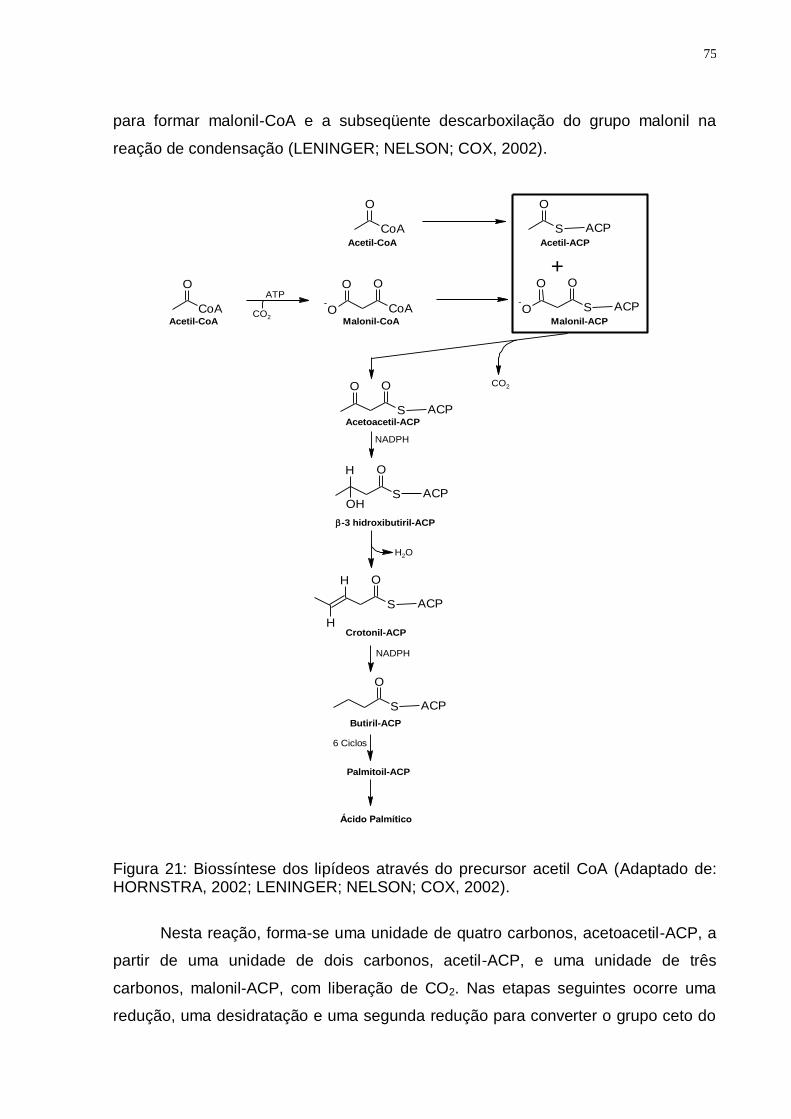
75
para formar malonil-CoA e a subseqüente descarboxilação do grupo malonil na
reação de condensação (LENINGER; NELSON; COX, 2002).
CoA
O O
-O
O
CoA
CoA
O
CO2
ATP
+
Acetil-CoA Acetil-ACP
Acetil-CoA Malonil-CoA Malonil-ACP
S
O
ACP
S
O
-O
O
ACP
S
OO
ACP Acetoacetil-ACP
CO2
NADPH
S
O
ACPOH
H
-3 hidroxibutiril-ACP
S
O
ACP
H
H
H2O
NADPH
S
O
ACP
Crotonil-ACP
Butiril-ACP
Palmitoil-ACP
6 Ciclos
Ácido Palmítico
Figura 21: Biossíntese dos lipídeos através do precursor acetil CoA (Adaptado de: HORNSTRA, 2002; LENINGER; NELSON; COX, 2002).
Nesta reação, forma-se uma unidade de quatro carbonos, acetoacetil-ACP, a
partir de uma unidade de dois carbonos, acetil-ACP, e uma unidade de três
carbonos, malonil-ACP, com liberação de CO2. Nas etapas seguintes ocorre uma
redução, uma desidratação e uma segunda redução para converter o grupo ceto do

76
C-3 do acetoacetil-ACP num grupo metileno que produz butiril-ACP, que completa o
primeiro ciclo de alongamento. Os ciclos de alongamento continuam até formar o
palmitoil-ACP. Este intermediário não é o substrato para a enzima condensadora,
por isso ele é hidrolisado para produzir o ácido palmítico e ACP. O ácido palmítico é
o precursor dos ácidos graxos saturados e insaturados de cadeia mais longa,
através de elongases e dessaturases. Esta ocorre pela inserção de sucessivas
moléculas de malonil-CoA com acil-CoA. As reações são seguidas por reduções
associadas ao Nicotinamida Adenina Dinucleotídeo Phosfato (NADPH) (VANZUELA;
NIETO, 2001; 2003; GONZÁLES, 2002).
Os ácidos graxos na sua grande maioria são os constituintes estruturais das
membranas celulares, cumprindo com as funções energéticas de reservas
metabólicas. Vários estudos apontam que sua utilização traz benefícios para a
saúde humana, prevenindo enfermidades cardiovasculares, câncer de cólon,
doenças imunológicas e favorecendo o desenvolvimento cerebral e da retina.
(VALENZUELA; NIETO, 2001; 2003; GONZÁLES, 2002; HORNSTRA, 2002; SILVA,
et al., 2007). O ω-6 e o ω-3 (Figura 22) são considerados precursores dos ácidos
graxos poliinsaturados de cadeia longa: ácido araquidônico, ácido
eicosapentaenóico, e ácido docosahexaenóico (HORNSTRA, 2002). O ácido
araquidônico (série ω-6) tem grande importância nos primeiros meses de vida,
sendo constituinte de estruturas celulares e precursores de mediadores inflamatórios
(GONZÁLES, 2002). Como agentes terapêuticos, utilizam-se essas substâncias que,
por estarem frequentemente deficientes na dieta, podem atuar em diversos
desequilíbrios metabólicos.
Figura 22: Representação estrutural dos ácidos graxos ω-6 e ω-3 (Adaptado de: HORNSTRA, 2002; LENINGER; NELSON; COX, 2002).

77
Alguns dos ácidos graxos essenciais com importância fisiológica são:
Série ω-3: Ácido Alfa-Linolênico, Ácido Eicosapentaenóico e Ácido
Docosahexaenóico.
Série ω-6: Ácido Linoléico, Ácido Gama - Linolênico e Ácido Araquidônico.
Série ω-9: Ácido Oléico e Ácido Nervônico.
Os glicerídeos são definidos como produtos da reação de uma molécula de
glicerol com até três moléculas de ácido graxo. Através da reação de esterificação,
os ácidos graxos são incorporados à molécula de glicerol. A reação inversa é a
reação de hidrólise, que provoca o desmembramento das moléculas de ácido graxo
do glicerol. Os monoglicerídeos apresentam uma molécula de ácido graxo
incorporado à molécula de glicerol. Di e triglicerídeos apresentam duas e três
moléculas de ácido graxo incorporados ao glicerol, respectivamente Os glicerídeos
insaturados convertem-se em maior ou menor grau em glicerídeos saturados, que
são estáveis e se solidificam em temperatura ambiente (LENINGER; NELSON; COX,
2002).
5.3.4 Identificação da (-) Hinoquinina
A (-) hinoquinina foi isolada da fração 29-43 (DBK-29) proveniente da coluna
6 e da fração 24-33 (DBX-04) da coluna 8, de D. brasiliensis, como um sólido
amorfo. Foi obtido 228,90 mg, com um rendimento equivalendo a 0,92% em relação
ao extrato bruto. Esta substância foi solúvel em acetona, clorofórmio, e metanol. O
fluxograma de obtenção das (-) hinoquininas está na figura 23. A estrutura foi
confirmada através das análises espectroscópicas de RMN 1H e 13C como a lignana
(-) hinoquinina (3,4-bis-(1,3-benzodioxol-5-ilmetill)-oxolan-2-ona) (Figuras 24 a 26).
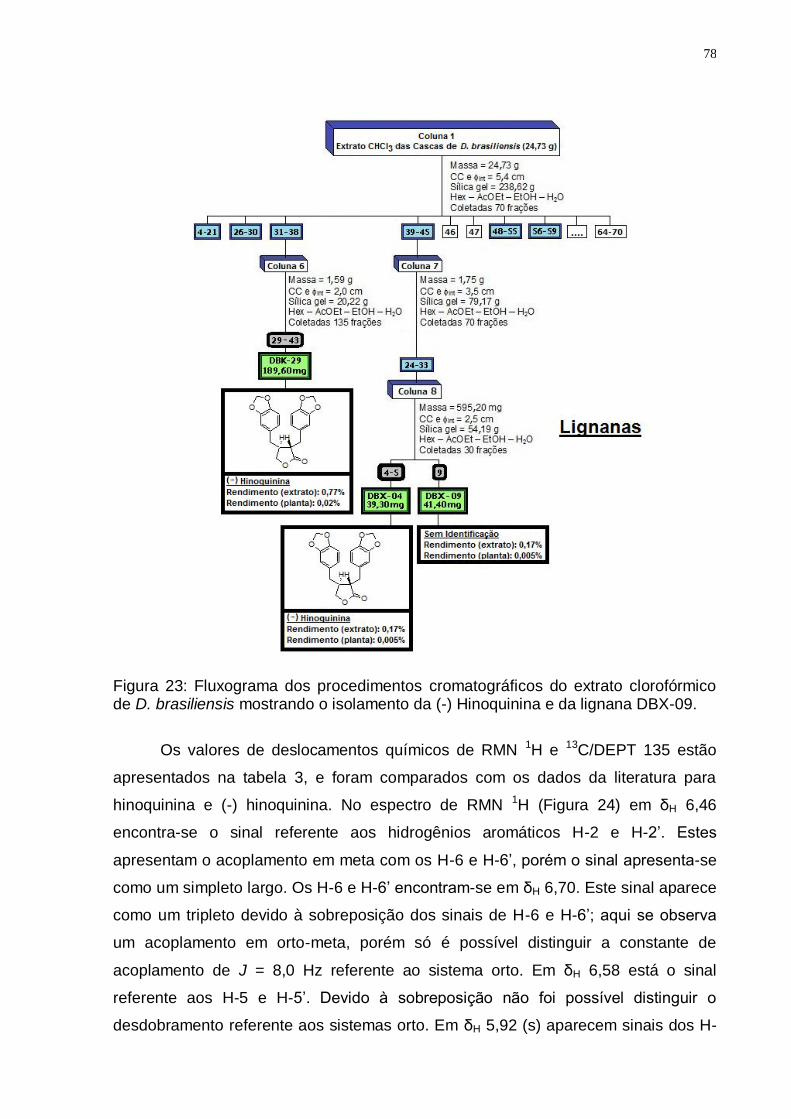
78
Figura 23: Fluxograma dos procedimentos cromatográficos do extrato clorofórmico de D. brasiliensis mostrando o isolamento da (-) Hinoquinina e da lignana DBX-09.
Os valores de deslocamentos químicos de RMN 1H e 13C/DEPT 135 estão
apresentados na tabela 3, e foram comparados com os dados da literatura para
hinoquinina e (-) hinoquinina. No espectro de RMN 1H (Figura 24) em δH 6,46
encontra-se o sinal referente aos hidrogênios aromáticos H-2 e H-2’. Estes
apresentam o acoplamento em meta com os H-6 e H-6’, porém o sinal apresenta-se
como um simpleto largo. Os H-6 e H-6’ encontram-se em δH 6,70. Este sinal aparece
como um tripleto devido à sobreposição dos sinais de H-6 e H-6’; aqui se observa
um acoplamento em orto-meta, porém só é possível distinguir a constante de
acoplamento de J = 8,0 Hz referente ao sistema orto. Em δH 6,58 está o sinal
referente aos H-5 e H-5’. Devido à sobreposição não foi possível distinguir o
desdobramento referente aos sistemas orto. Em δH 5,92 (s) aparecem sinais dos H-
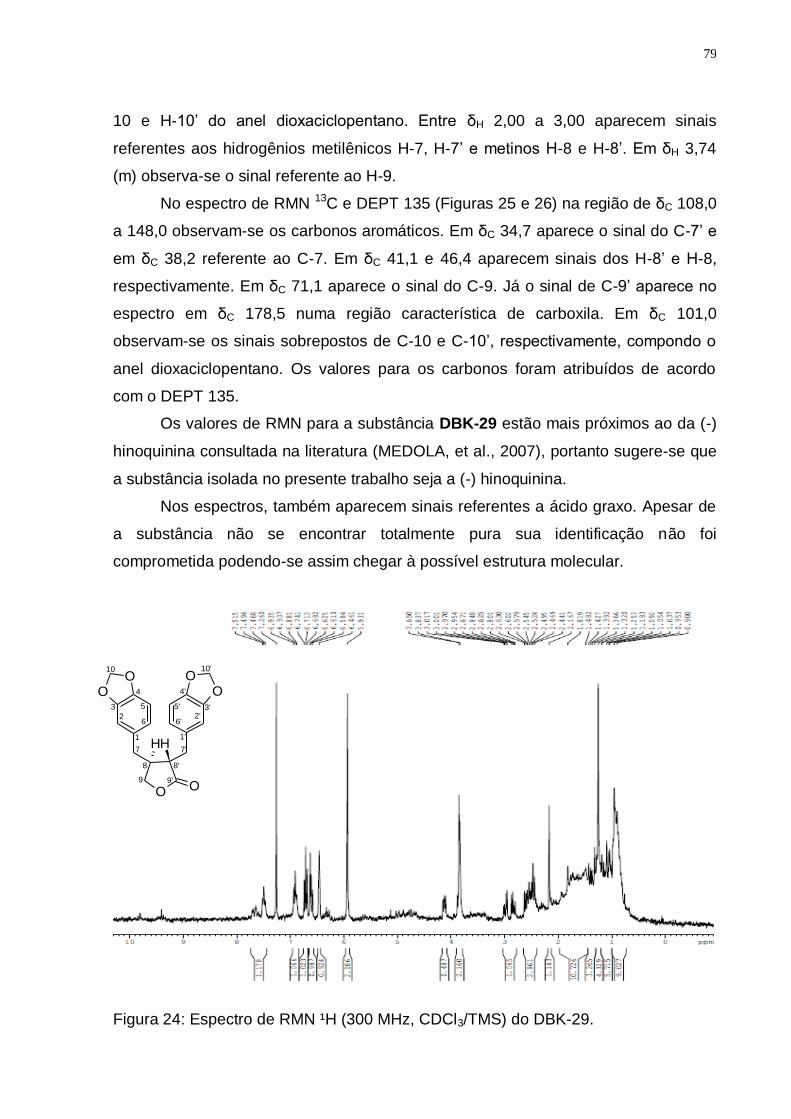
79
10 e H-10’ do anel dioxaciclopentano. Entre δH 2,00 a 3,00 aparecem sinais
referentes aos hidrogênios metilênicos H-7, H-7’ e metinos H-8 e H-8’. Em δH 3,74
(m) observa-se o sinal referente ao H-9.
No espectro de RMN 13C e DEPT 135 (Figuras 25 e 26) na região de δC 108,0
a 148,0 observam-se os carbonos aromáticos. Em δC 34,7 aparece o sinal do C-7’ e
em δC 38,2 referente ao C-7. Em δC 41,1 e 46,4 aparecem sinais dos H-8’ e H-8,
respectivamente. Em δC 71,1 aparece o sinal do C-9. Já o sinal de C-9’ aparece no
espectro em δC 178,5 numa região característica de carboxila. Em δC 101,0
observam-se os sinais sobrepostos de C-10 e C-10’, respectivamente, compondo o
anel dioxaciclopentano. Os valores para os carbonos foram atribuídos de acordo
com o DEPT 135.
Os valores de RMN para a substância DBK-29 estão mais próximos ao da (-)
hinoquinina consultada na literatura (MEDOLA, et al., 2007), portanto sugere-se que
a substância isolada no presente trabalho seja a (-) hinoquinina.
Nos espectros, também aparecem sinais referentes a ácido graxo. Apesar de
a substância não se encontrar totalmente pura sua identificação não foi
comprometida podendo-se assim chegar à possível estrutura molecular.
Figura 24: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBK-29.
OOO
O
O O
HH1 1'
2 2'3 3'
4 4'
5 5'
6 6'
7 7'
8 8'
9 9'
10 10'
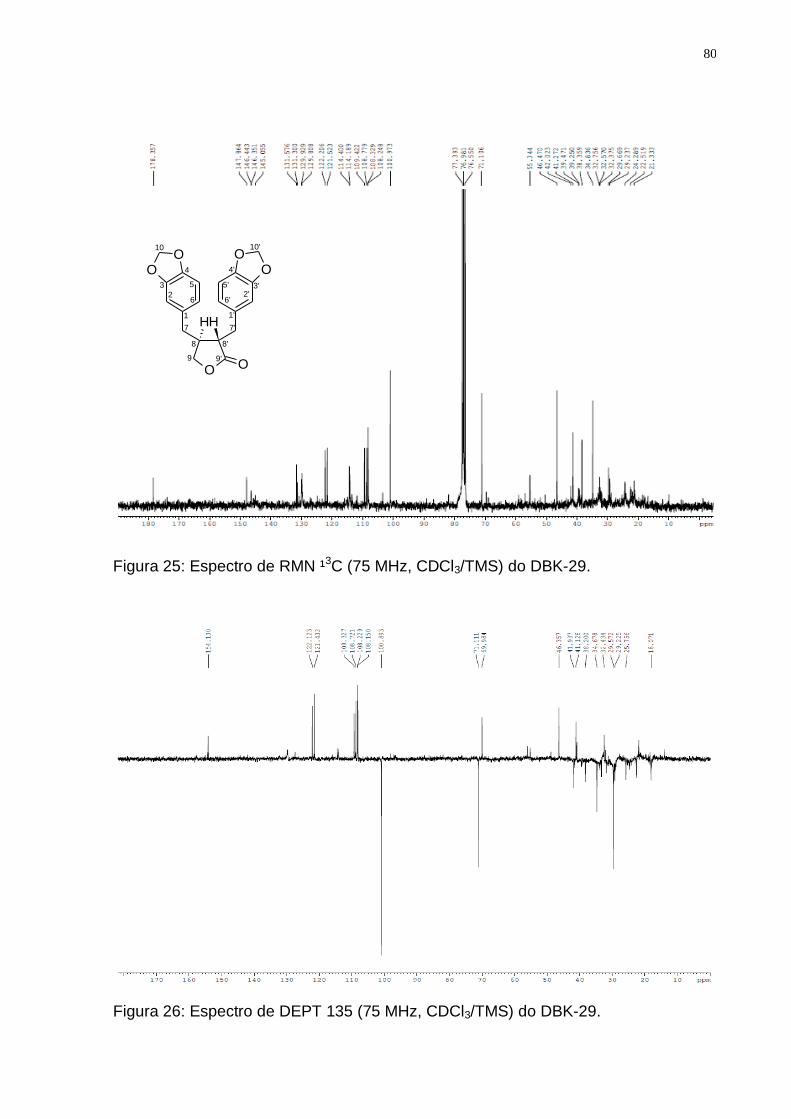
80
Figura 25: Espectro de RMN ¹3C (75 MHz, CDCl3/TMS) do DBK-29.
Figura 26: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBK-29.
OOO
O
O O
HH1 1'
2 2'3 3'
4 4'
5 5'
6 6'
7 7'
8 8'
9 9'
10 10'
81
Em trabalhos de Medola e colaboradores (2007) a hinoquinina é relatada
como tendo promissoras atividades tripanocidas, cogitando-se assim um potencial
fármaco antichagásico. Tanto no gênero Drimys quanto na família Winteraceae entre
as lignanas isoladas estão a α e β-cubebina (MALHEIROS, et al., 2001, NAKANO, et
al., 1999). Por outro lado, a (-) hinoquinina é a primeira vez que está sendo
reportada nesta espécie.
Tabela 3: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBK-29 comparando com os dados de literaturas para a Hinoquinina e para a (-) Hinoquinina.
OOO
O
O O
HH1 1'
2 2'3 3'
4 4'
5 5'
6 6'
7 7'
8 8'
9 9'
10 10'
Posição
DBK-29
(ppm) mult. J (Hz)
Hinoquinina*
(ppm) mult. J (Hz)
(-) Hinoquinina**
(ppm) mult. J (Hz)
DBK-29 13
C / DEPT
Hinoquinina*
13C / mult.
(-) Hinoquinina**
13C / mult.
1 131,2 / C 132,2 / C 131,4 / C 2 6,46, sl 6,24, d, (J = 1,9) 109,3 / CH 109,2 / CH 108,7 / CH 3 146,3 / C 146,9 / C 146,4 / C 4 147,7 / C 148,3 / C 147,9 / C 5 6,62, d, (J =
7,2) 6,54, d, (J = 7,9) 108,1 / CH 108,4 / CH 109,4 / CH
6 6,70, d, (J = 8,1)
6,08, dd, (J = 7,9; 1,9)
121,4 / CH 121,7 / CH 122,1 / CH
7 2,53, m; 1,76, m
2,05, dd, (J = 13,4; 5,8); 1,74, dd, (J = 13,4; 8,2)
2,45, m 38,2 / CH2 38,1 / CH2 38,4 / CH2
8 2,10, m 1,92, ddddd, (J= 14,2; 8,2; 8,0; 7,5 e 5,8)
3,00, dd, (J = 14,2; 5,1)
41,1 / CH 41,3 / CH 41,3 / CH
9 3,74, m 3,53, dd, (J = 9,0; 7,5)
3,85, dd, (J = 9,1; 7,1)
71,1 / CH2 70,5 / CH2 71,0 / CH2
10 5,93, sl 5,30, d, (J = 1,2) 5,90, sl 100,9 / CH2 100,9 / CH2 100,9 / CH2 1’ 131,5 / C 132,3 / C 131,5 / C 2’ 6,46, sl 6,56, d, (J = 1,9) 108,7 / CH 108,4 / CH 108,7 / CH 3’ 146,2 / C 146,7 / C 146,4 / C 4’ 147,7 / C 148,3 / C 147,9 / C 5’ 6, 62, d, (J =
7,2) 6,57, d, (J = 7,9) 108,2 / CH 109,9 / CH 109,4 / CH
6’ 6,70, d, (J = 8,1)
6,39, dd, (J = 7,9; 1,9)
121,1 / CH 122,6 / CH 122,5 / CH
7’ 2,96, dd, (J = 14,1; 4,8); 2,84, dd, (J = 13,8; 6,9)
2,76, dd, (J = 13,9; 5,0); 2,56, dd, (J = 13,9; 7,1)
2,85, dd, (J = 14,2; 7,3); 2,55, m
34,7 / CH2 34,9 / CH2 34,9 / CH2
8’ 2,44, m 1,96, ddd, (J = 14,2; 7,1; 5,0)
2,40, m 46,3 / CH 46,4 /CH 46,5 / CH
9’ 178,5 / C 177,5 / C 178,8 / C 10’ 5,93,sl 5,34, d, (J = 1,4) 100,9 / CH2 100,9 / CH2 100,9 / CH2
Solvente CDCl3 300 MHz
CDCl3 200 MHz
CDCl3 400 MHz
CDCl3 75 MHz
CDCl3 50 MHz
CDCl3 100 MHz
*HELENO, et al., 2006. **MEDOLA, et al., 2007.

82
As lignanas e as neolignanas costumam ser encontradas como derivados
fenilpropanoídicos, formados por duas unidades destes ligados através do carbono
β. São provenientes da via do ácido chiquímico, passando pela fenilalanina que
sofre desaminação enzimática pela fenilalanina-amonialiase ao ácido cinâmico
correspondente. O ácido p-hidroxicinâmico formado é o principal precursor de todos
os fenilpropanóides, inclusive as lignanas, ligninas, neolignanas e fenilpropenos. As
lignanas e as neolignanas são formadas pela dimerização oxidativa das unidades de
álcool coniferílico. Duas dessas unidades são unidas através de uma variedade de
possíveis ligações covalentes dos átomos de carbono e de oxigênio presentes na
cadeia lateral C-3. Reações subseqüentes levam a substituição dos anéis
aromáticos e a produção de compostos policíclicos (HELENO, et al., 2006).
Uma condição estrutural necessária para o acoplamento oxidativo resulta nos
diferentes tipos de lignóides é a presença de uma hidroxila livre na posição para das
unidades monoméricas. Outras oxigenações adicionais ocorrem freqüentemente nas
posições C-3 e C-5 dos anéis aromáticos, e os substituintes em geral são do tipo
hidroxi, metoxi e metilenodioxílico. A oxigenação do anel C ocorre após o
acoplamento das unidades monoméricas (Figura 27). Devido às variações de níveis
de oxidação, seu grau de substituições do anel, e sua complexidade estrutural a
transforma numa classe de substâncias de origem natural com uma grande
variedade estrutural (SIMÕES, et al., 2004).
Formam uma classe de produtos naturais que incluem um grande número de
estruturas conhecidas, mostrando uma grande variedade de atividades biológicas.
São relatadas atividades tripanocida, antitumoral, anti-HIV, anticonvulsionante,
antiespasmódica, sobre o fator de agregação plaquetária, antialérgica e
antiinflamatória (HELENO, et al., 2006, SIMÕES, et al., 2004).

83
Figura 27: Biossíntese dos lignóides (Adaptado de: SIMÕES, et al., 2004).
Gottlieb (1982) mostrou que um dos indicativos evolutivos em angiospermas
são os lignóides desempenhando num importante papel na adaptação ecológica.
Segundo ele, esta classe de substâncias pode estar relacionada com interações das
plantas com fungos, insetos e com outras plantas. Como fonte de medicamentos
tem-se a lignana podofilotoxina, extraída de Podophyllum.
De acordo com a rota biossintética dos lignóides, pode-se sugerir a provável
modificação para dar origem à hinoquinina (figura 28). Os lignóides são originados
da via do ácido chiquímico e seu principal precursor é o ácido p-hidroxicinâmico.
Este sofre uma oxidação no carbono 4 e se transforma no ácido caféico. O ácido
caféico por sua vez é metilado na hidroxila do carbono 4, dando origem ao ácido
ferúlico. Este por sua vez após uma série de modificações se transforma no ácido
coniferílico. Na porção oxigenada ligada ao anel aromático ocorre a ciclização
formando um anel metilenodioxifenil. Pela dimerização destas unidades formadas
podemos propor que esta seja a rota de formação da hinoquinina.
84
OH
O
OH
OH
O
OH
OH
OH
O
OH
OMe
S
O
OH
OMe
CoAOH
O
OHOH
OO
OH
OO
OH
OO
+
OOO
O
O O
HH
OH- Me
SCoA-
OSCoA-H+
Ácido p-hidroxicinâmico Ácido Caféico Ácido Ferúlico
Feriloil-CoAÁcido Coniferílico
Hinoquinina
OO O
O
OHOH
OO O
O
OHOH
-
H+
Figura 28: Rota biossintética provável da hinoquinina (Fonte: DEWICK, 2002).
5.3.5 Identificação do DBX-09
A substância DBX-09 foi isolada da fração 24-33 da coluna 8, de D.
brasiliensis, como um sólido amorfo. Foi obtido 41,40 mg, com um rendimento
equivalendo a 0,16% em relação ao extrato bruto. Esta substância foi solúvel em
acetona, clorofórmio e metanol. A substância está em fase de purificação.
No espectro de RMN 1H (figura 29) na região de δH 6,40 a 7,50 é possível
observar os sinais dos hidrogênios aromáticos. Em δH 5,91 o simpleto é
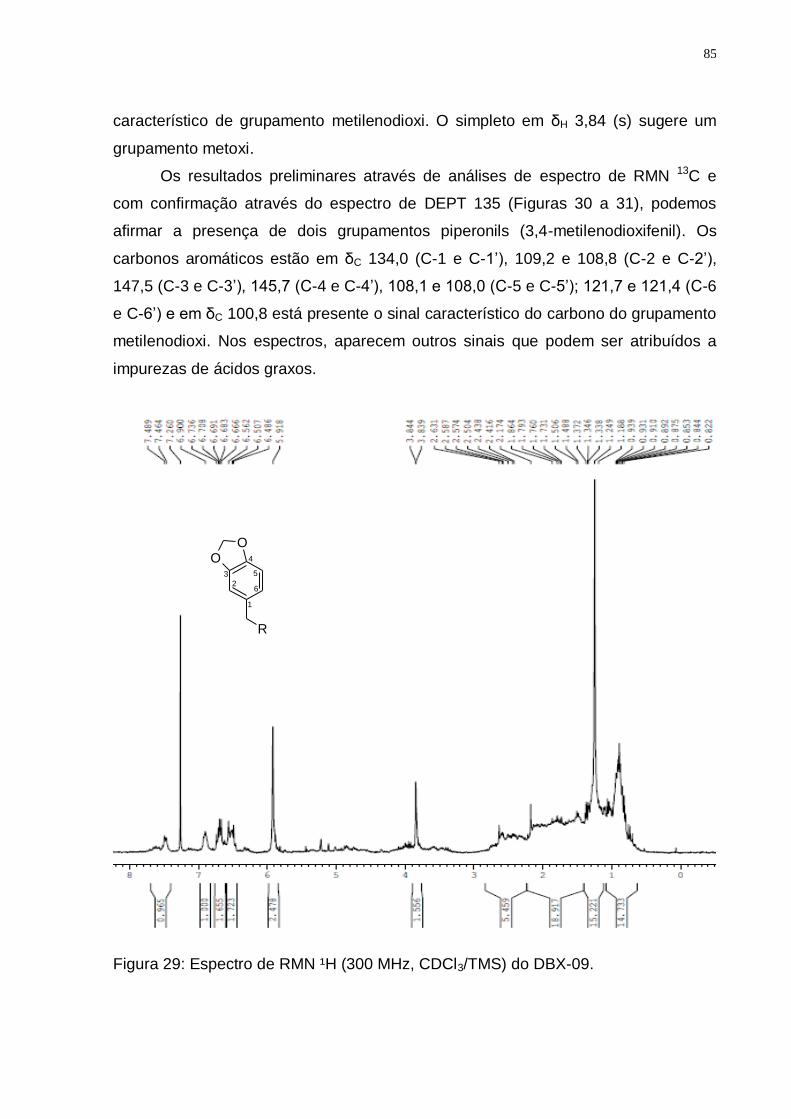
85
característico de grupamento metilenodioxi. O simpleto em δH 3,84 (s) sugere um
grupamento metoxi.
Os resultados preliminares através de análises de espectro de RMN 13C e
com confirmação através do espectro de DEPT 135 (Figuras 30 a 31), podemos
afirmar a presença de dois grupamentos piperonils (3,4-metilenodioxifenil). Os
carbonos aromáticos estão em δC 134,0 (C-1 e C-1’), 109,2 e 108,8 (C-2 e C-2’),
147,5 (C-3 e C-3’), 145,7 (C-4 e C-4’), 108,1 e 108,0 (C-5 e C-5’); 121,7 e 121,4 (C-6
e C-6’) e em δC 100,8 está presente o sinal característico do carbono do grupamento
metilenodioxi. Nos espectros, aparecem outros sinais que podem ser atribuídos a
impurezas de ácidos graxos.
Figura 29: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBX-09.
R
OO
1
2
3
4
5
6
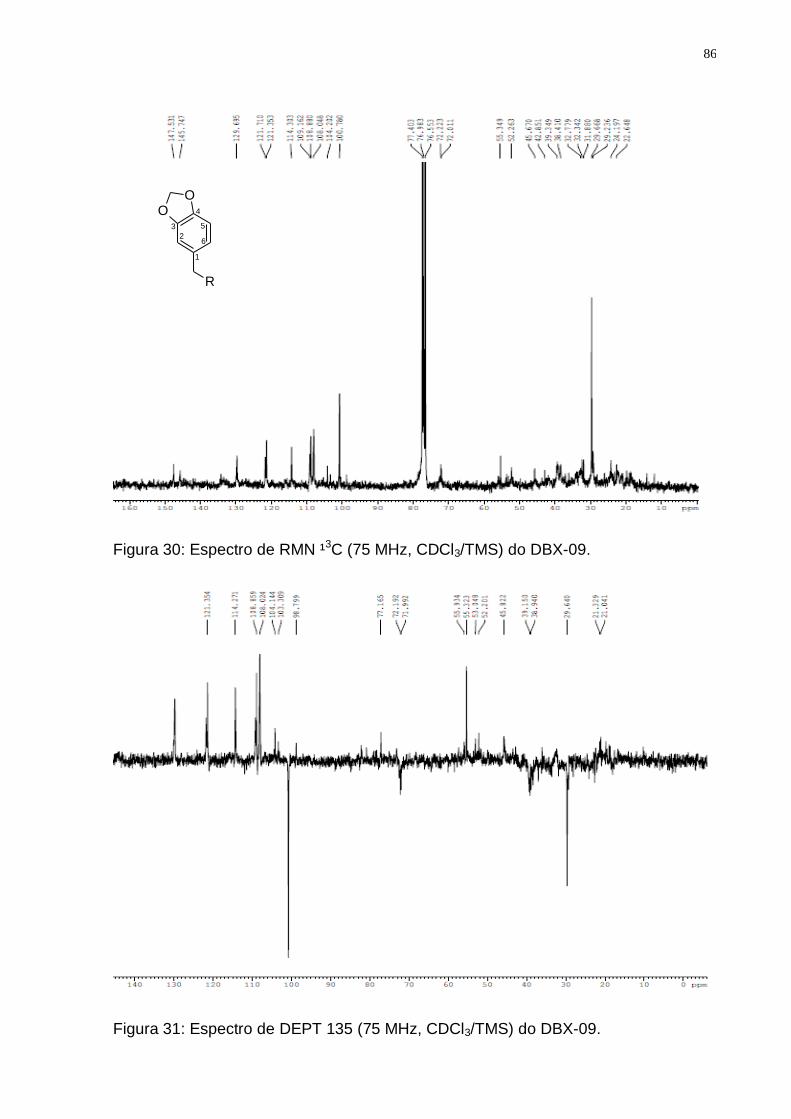
86
Figura 30: Espectro de RMN ¹3C (75 MHz, CDCl3/TMS) do DBX-09.
Figura 31: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBX-09.
R
OO
1
2
3
4
5
6
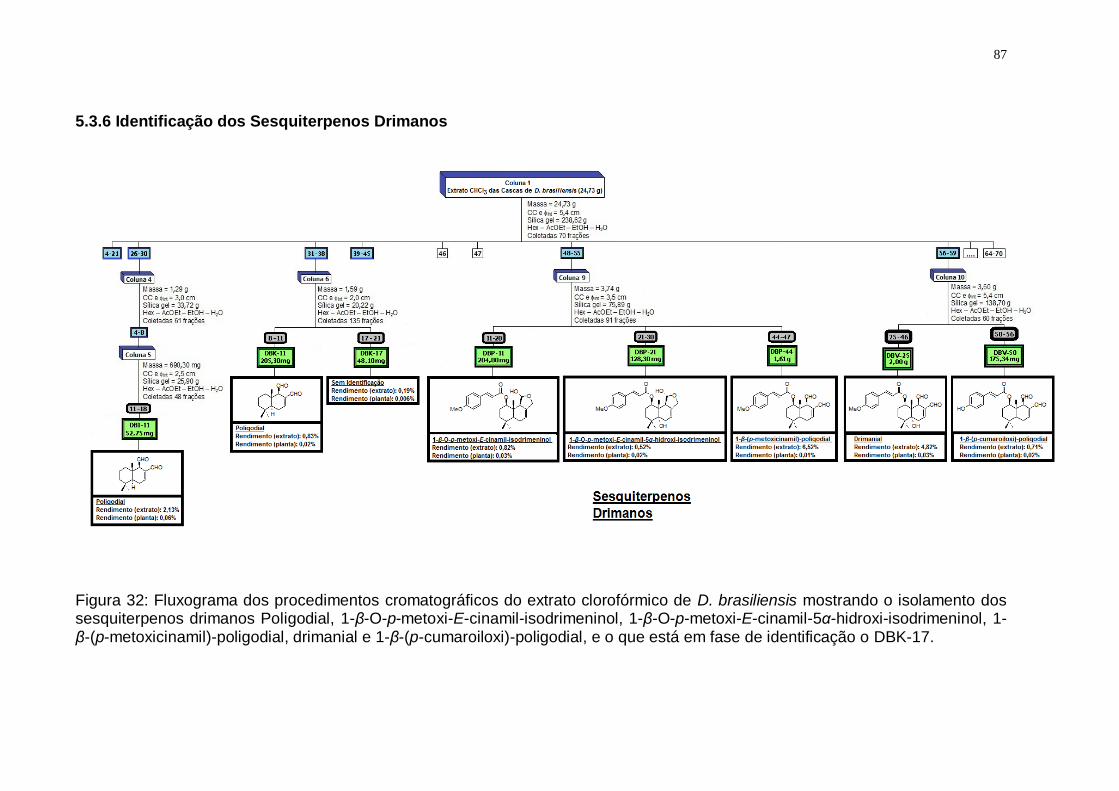
87
5.3.6 Identificação dos Sesquiterpenos Drimanos
Figura 32: Fluxograma dos procedimentos cromatográficos do extrato clorofórmico de D. brasiliensis mostrando o isolamento dos sesquiterpenos drimanos Poligodial, 1-β-O-p-metoxi-E-cinamil-isodrimeninol, 1-β-O-p-metoxi-E-cinamil-5α-hidroxi-isodrimeninol, 1-β-(p-metoxicinamil)-poligodial, drimanial e 1-β-(p-cumaroiloxi)-poligodial, e o que está em fase de identificação o DBK-17.

88
5.3.6.1 Identificação do Poligodial
O poligodial (16) (9--drim-7-en-11,12-dial) foi isolado das frações 11-18 da
coluna 5 e da fração 8-11 da coluna 6. Eles foram identificados com os códigos DBI-
11 e o DBK-11, respectivamente. Foi obtido 73,28 mg, com um rendimento
equivalendo a 2,96% em relação ao extrato bruto. Este foi solúvel em acetona,
clorofórmio, diclorometano e etanol. A sua estrutura foi confirmada através das
análises espectroscópicas de RMN 1H e 13C/DEPT 135 (Figuras 33 a 35). Esta
substância foi isolada anteriormente por Malheiros (2001) da D. brasiliensis e D.
angustifolia. A tabela 4 mostra os valores de deslocamentos químicos de RMN 1H e
13C para o poligodial, comparados com os valores obtidos para o DBI-11.
No espectro de RMN 1H (figura 33), observa-se os hidrogênios referentes aos
grupamentos aldeídicos. Em δH 9,54 está um dupleto (J = 4,5 Hz) referente ao H-12.
Próximo a ele, está um simpleto em δH 9,46, referente ao segundo grupamento
aldeídico do poligodial o H-11.
Em δH 7,13 há um multipleto, referente ao hidrogênio oleofínico H-7. O
hidrogênio no carbono adjacente ao grupo aldeído δH 2,83 é desblindado por causa
do grupo carbonila, mas por estar mais distante o efeito sobre ele é menor. Já entre
δH 2,48 e 2,36 aparece um multipleto referente ao H-6. Os sinais referentes aos
carbonos das três metilas estão em δH 0,92, 0,95 e 0,96.
No espectro de 13C/DEPT 135 (Figuras 34 e 35), os carbonos referentes aos
grupamentos aldeídicos estão em δC 202,0 (CH) para o C-11 e δC 193,2 (CH) para o
C-12. Em δC 154,3 está o C-7 (CH) e em δC 138,3 está o C-8 (C) referentes aos
carbonos olefínicos. O carbono C-9 adjacente ao grupo aldeídico está em δC 60,3
(CH). Entre 15 e 50 ppm estão às metilas, metilenos e metinos, com destaque
especial para as três metilas estão em δc 33,2 (C-13), 22,0 (C-14) e 15,2 (C-15).

89
Figura 33: Espectro de RMN 1H (300 MHz, CDCl3/TMS) do DBI-11.
Figura 34: Espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBI-11.
CHO
CHO
H
12
34
56
7
89
10
11
12
1314
15
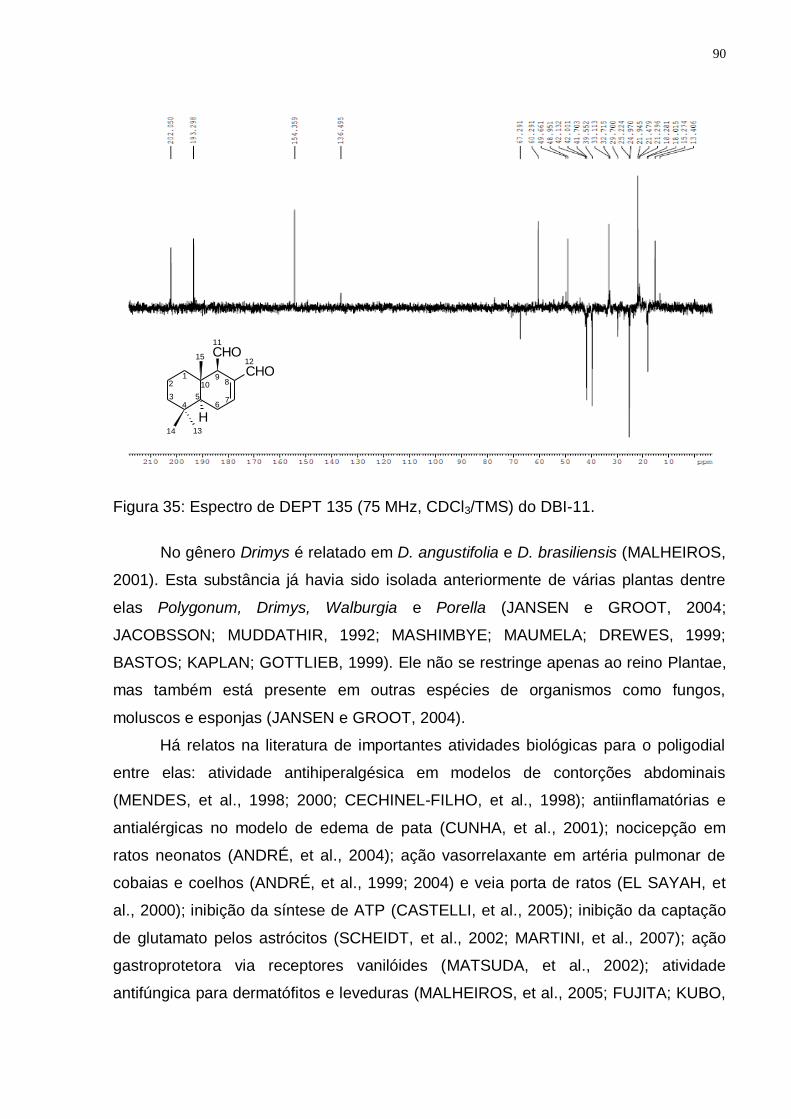
90
Figura 35: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBI-11.
No gênero Drimys é relatado em D. angustifolia e D. brasiliensis (MALHEIROS,
2001). Esta substância já havia sido isolada anteriormente de várias plantas dentre
elas Polygonum, Drimys, Walburgia e Porella (JANSEN e GROOT, 2004;
JACOBSSON; MUDDATHIR, 1992; MASHIMBYE; MAUMELA; DREWES, 1999;
BASTOS; KAPLAN; GOTTLIEB, 1999). Ele não se restringe apenas ao reino Plantae,
mas também está presente em outras espécies de organismos como fungos,
moluscos e esponjas (JANSEN e GROOT, 2004).
Há relatos na literatura de importantes atividades biológicas para o poligodial
entre elas: atividade antihiperalgésica em modelos de contorções abdominais
(MENDES, et al., 1998; 2000; CECHINEL-FILHO, et al., 1998); antiinflamatórias e
antialérgicas no modelo de edema de pata (CUNHA, et al., 2001); nocicepção em
ratos neonatos (ANDRÉ, et al., 2004); ação vasorrelaxante em artéria pulmonar de
cobaias e coelhos (ANDRÉ, et al., 1999; 2004) e veia porta de ratos (EL SAYAH, et
al., 2000); inibição da síntese de ATP (CASTELLI, et al., 2005); inibição da captação
de glutamato pelos astrócitos (SCHEIDT, et al., 2002; MARTINI, et al., 2007); ação
gastroprotetora via receptores vanilóides (MATSUDA, et al., 2002); atividade
antifúngica para dermatófitos e leveduras (MALHEIROS, et al., 2005; FUJITA; KUBO,
CHO
CHO
H
12
34
56
7
89
10
11
12
1314
15

91
2005); antitumoral em linhagens SH-SY5Y, Hela e L929 (FORSBY, et al., 1998;
FORSBY; WALUM, 1996; SILVA, 2009); antialimentar de lagartas (ZAPATA, 2009).
Tabela 4: Valores de deslocamentos químicos () de RMN 1H e 13C para DBI-11 e dados da literatura para o Poligodial.
CHO
CHO
H
12
34
56
7
89
10
11
12
1314
15
Posição
DBI-11
(ppm) mult. J (Hz)
Poligodial*
(ppm) mult. J (Hz)
DBI-11 13
C / DEPT
Poligodial*
13C / mult.
1 39,5 / CH2 39,8 / t
2 18,0 / CH2 18,0 / t
3 41,7 / CH2 42,0 / t
4 33,1 / C 33,0 / s
5 1,49, m 48,9 / CH 49,0 / d
6 2,48; 2.36, m 25,2 / CH2 25,3 / t
7 7,13, m 7,15, m 154,3 / CH2 152,2 / d
8 138,3 / C 138,0 / d
9 2,83, sl 2,85, m 60,3 / CH 60,0 / d
10 36,8 / C 37,0 / s
11 9,54, d, (J= 4,5) 9,55, d, (J= 4,4) 202,0 / CH 202,0 / d
12 9,46, s 9,45, s 193,2 / CH 194,0 / d
13 0,92, s 0,8, s 33,1 / CH3 33,0 / q
14 0,94, s 0,9, s 21,9 / CH3 22,0 / q
15 0,96, s 1,0, s 15,2 / CH3 15,0 / q
Solvente CDCl3
300 MHz
CDCl3
200 MHz
CDCl3
75 MHz
CDCl3
50 MHz
*(MALHEIROS, 2001).
De acordo com a rota biossintética dos drimanos, pode-se sugerir a provável
modificação estrutural do esqueleto drimano para dar origem ao poligodial (figura 36).
A metila do carbono 3 é retirada e posteriormente a dupla ligação do carbono 4 é
rompida para estabilização do anel. Após isso, a estrutura sofre uma oxidação na
metila do carbono 11 transformando-se em aldeído. Novamente ocorre uma segunda
oxidação na metila do carbono 12 transformando-se em aldeído.

92
CHO
+H2O +H2O
CHO
CHO
H
Poligodial
Figura 36: Rota biossintética provável do poligodial (Adaptado de: JANSEN e GROOT, 2004).
5.3.6.2 Identificação do DBK-17
A substância DBK-17 foi isolada da fração 17-21 da coluna 6, de D.
brasiliensis, como um sólido cristalino branco. Foi obtido 48,10 mg, com um
rendimento equivalendo a 0,19% em relação ao extrato bruto, solúvel em clorofórmio
e acetona. A substância está em fase de purificação.
No espectro de RMN 1H (Figura 37) são encontrados dois simpletos em δH 9,72
e 9,41, referentes aos hidrogênios aldeídicos, demonstrando assim a presença de
outro grupamento em C-9. Em 6,84 observa-se um sinal referente a hidrogênio
oleifínico e o simpleto em δH 5,95 indica outra função oxigenada na molécula. Em δH
3,84 um simpleto é visível relativo a mais um grupamento oxigenado, talvez uma
metoxila. Abaixo de 3,00 observam-se sinais referentes a metilas, metilenos e
metinos. Esses sugerem que a substância DBK-17 pertence à classe dos drimanos.
Porém fazem-se necessárias outras análises para a correta identificação, assim como
a sua purificação.
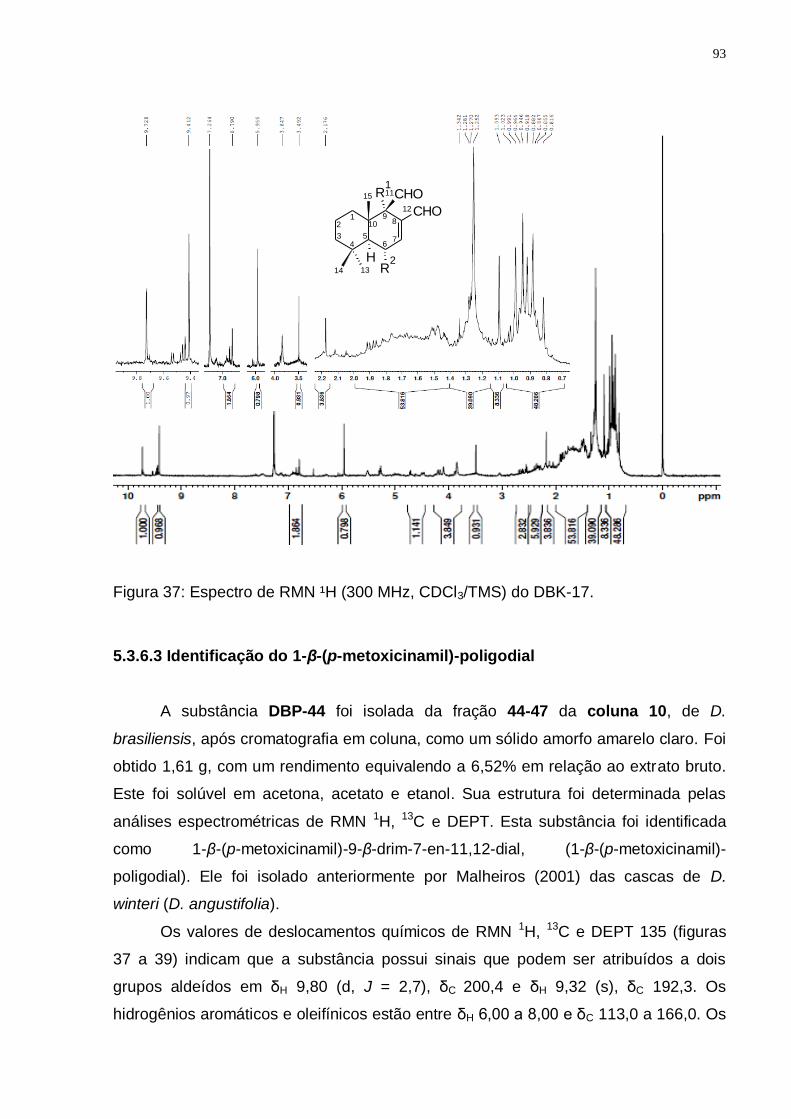
93
Figura 37: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBK-17.
5.3.6.3 Identificação do 1-β-(p-metoxicinamil)-poligodial
A substância DBP-44 foi isolada da fração 44-47 da coluna 10, de D.
brasiliensis, após cromatografia em coluna, como um sólido amorfo amarelo claro. Foi
obtido 1,61 g, com um rendimento equivalendo a 6,52% em relação ao extrato bruto.
Este foi solúvel em acetona, acetato e etanol. Sua estrutura foi determinada pelas
análises espectrométricas de RMN 1H, 13C e DEPT. Esta substância foi identificada
como 1-β-(p-metoxicinamil)-9-β-drim-7-en-11,12-dial, (1-β-(p-metoxicinamil)-
poligodial). Ele foi isolado anteriormente por Malheiros (2001) das cascas de D.
winteri (D. angustifolia).
Os valores de deslocamentos químicos de RMN 1H, 13C e DEPT 135 (figuras
37 a 39) indicam que a substância possui sinais que podem ser atribuídos a dois
grupos aldeídos em δH 9,80 (d, J = 2,7), δC 200,4 e δH 9,32 (s), δC 192,3. Os
hidrogênios aromáticos e oleifínicos estão entre δH 6,00 a 8,00 e δC 113,0 a 166,0. Os
CHO
CHO
H
R1
R2
12
34
56
7
89
10
11
12
1314
15
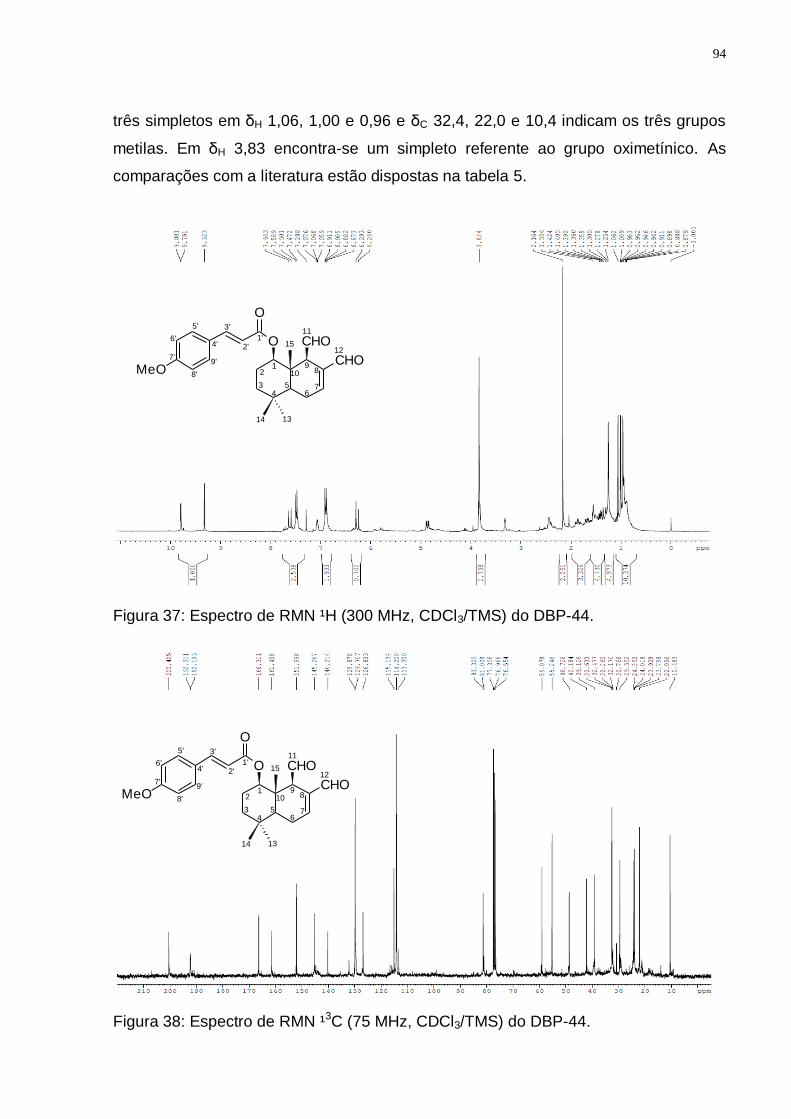
94
três simpletos em δH 1,06, 1,00 e 0,96 e δC 32,4, 22,0 e 10,4 indicam os três grupos
metilas. Em δH 3,83 encontra-se um simpleto referente ao grupo oximetínico. As
comparações com a literatura estão dispostas na tabela 5.
Figura 37: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBP-44.
Figura 38: Espectro de RMN ¹3C (75 MHz, CDCl3/TMS) do DBP-44.
O
O
MeO
CHO
CHO1
2
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
O
O
MeO
CHO
CHO1
2
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
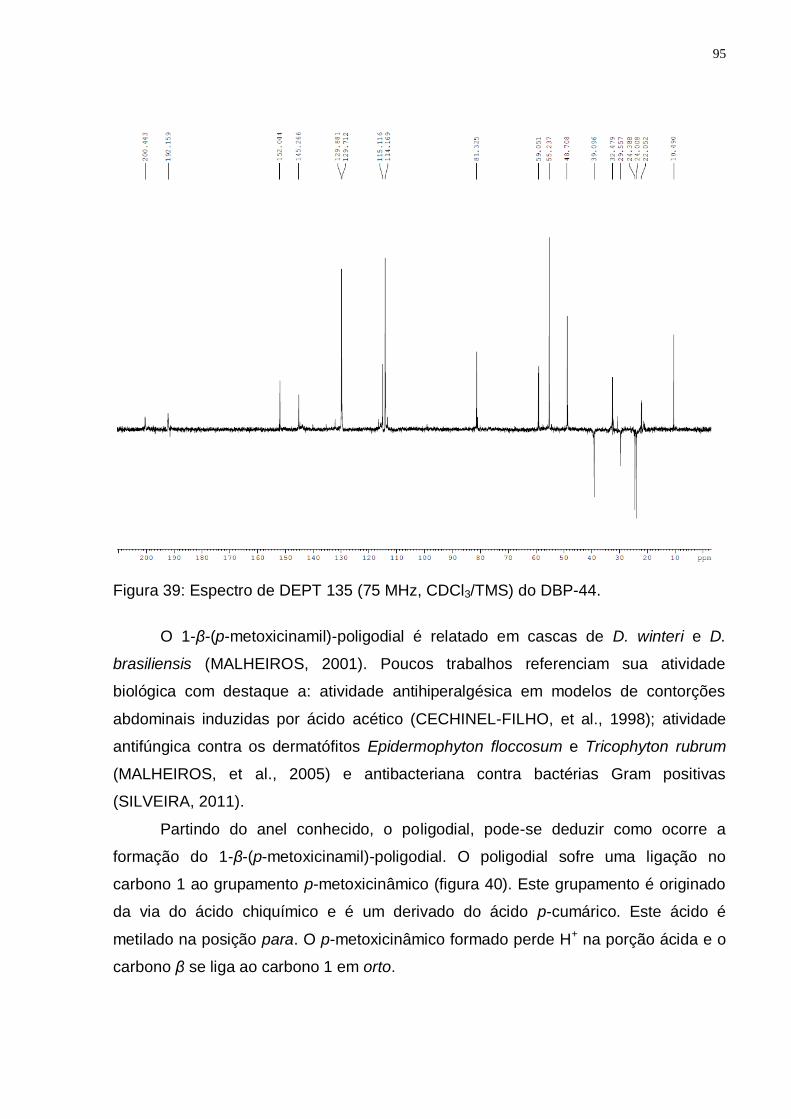
95
Figura 39: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBP-44.
O 1-β-(p-metoxicinamil)-poligodial é relatado em cascas de D. winteri e D.
brasiliensis (MALHEIROS, 2001). Poucos trabalhos referenciam sua atividade
biológica com destaque a: atividade antihiperalgésica em modelos de contorções
abdominais induzidas por ácido acético (CECHINEL-FILHO, et al., 1998); atividade
antifúngica contra os dermatófitos Epidermophyton floccosum e Tricophyton rubrum
(MALHEIROS, et al., 2005) e antibacteriana contra bactérias Gram positivas
(SILVEIRA, 2011).
Partindo do anel conhecido, o poligodial, pode-se deduzir como ocorre a
formação do 1-β-(p-metoxicinamil)-poligodial. O poligodial sofre uma ligação no
carbono 1 ao grupamento p-metoxicinâmico (figura 40). Este grupamento é originado
da via do ácido chiquímico e é um derivado do ácido p-cumárico. Este ácido é
metilado na posição para. O p-metoxicinâmico formado perde H+ na porção ácida e o
carbono β se liga ao carbono 1 em orto.
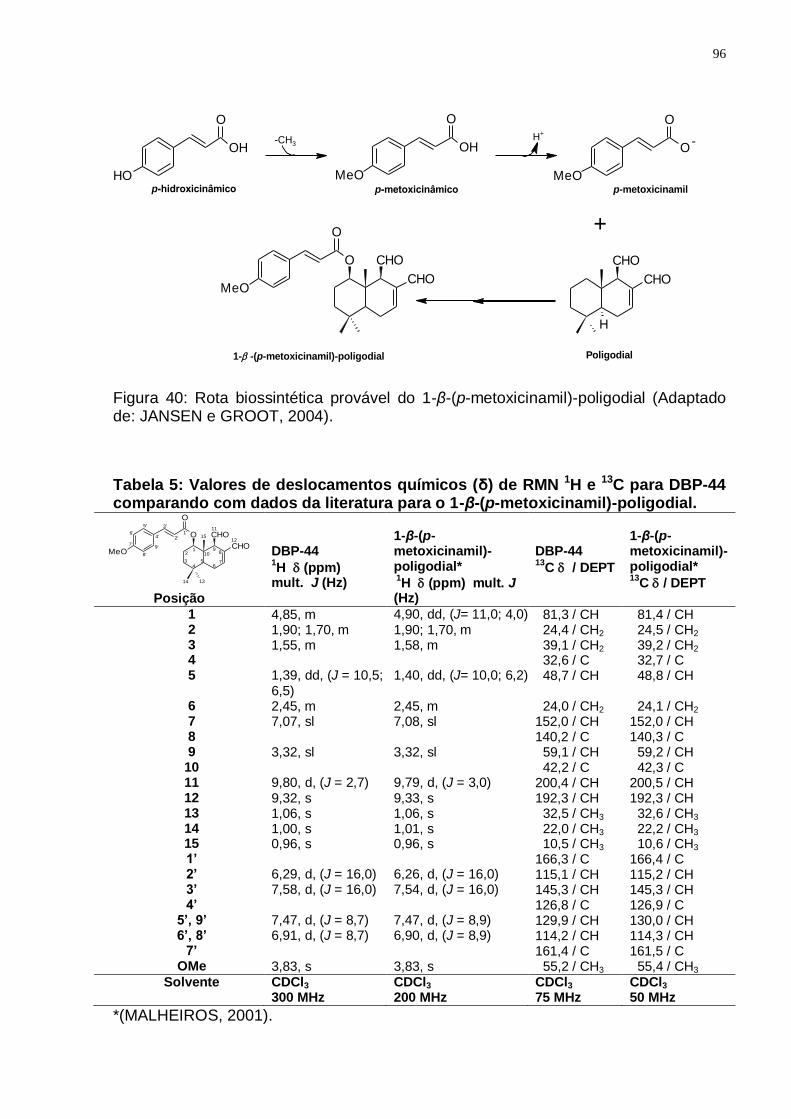
96
CHO
CHO
H
OH
O
OH
OH
O
MeO
-CH3
O
MeO
OH-
+
O
O
MeO
CHO
CHO
p-hidroxicinâmico p-metoxicinâmico
H+
p-metoxicinamil
Poligodial1--(p-metoxicinamil)-poligodial
Figura 40: Rota biossintética provável do 1-β-(p-metoxicinamil)-poligodial (Adaptado de: JANSEN e GROOT, 2004).
Tabela 5: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBP-44 comparando com dados da literatura para o 1-β-(p-metoxicinamil)-poligodial.
O
O
MeO
CHO
CHO1
2
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
Posição
DBP-44 1H (ppm)
mult. J (Hz)
1-β-(p-metoxicinamil)-poligodial* 1H (ppm) mult. J
(Hz)
DBP-44 13
C / DEPT
1-β-(p-metoxicinamil)-poligodial* 13
C / DEPT
1 4,85, m 4,90, dd, (J= 11,0; 4,0) 81,3 / CH 81,4 / CH 2 1,90; 1,70, m 1,90; 1,70, m 24,4 / CH2 24,5 / CH2
3 1,55, m 1,58, m 39,1 / CH2 39,2 / CH2 4 32,6 / C 32,7 / C 5 1,39, dd, (J = 10,5;
6,5) 1,40, dd, (J= 10,0; 6,2) 48,7 / CH 48,8 / CH
6 2,45, m 2,45, m 24,0 / CH2 24,1 / CH2 7 7,07, sl 7,08, sl 152,0 / CH 152,0 / CH 8 140,2 / C 140,3 / C 9 3,32, sl 3,32, sl 59,1 / CH 59,2 / CH
10 42,2 / C 42,3 / C 11 9,80, d, (J = 2,7) 9,79, d, (J = 3,0) 200,4 / CH 200,5 / CH 12 9,32, s 9,33, s 192,3 / CH 192,3 / CH 13 1,06, s 1,06, s 32,5 / CH3 32,6 / CH3 14 1,00, s 1,01, s 22,0 / CH3 22,2 / CH3 15 0,96, s 0,96, s 10,5 / CH3 10,6 / CH3 1’ 166,3 / C 166,4 / C 2’ 6,29, d, (J = 16,0) 6,26, d, (J = 16,0) 115,1 / CH 115,2 / CH 3’ 7,58, d, (J = 16,0) 7,54, d, (J = 16,0) 145,3 / CH 145,3 / CH 4’ 126,8 / C 126,9 / C
5’, 9’ 7,47, d, (J = 8,7) 7,47, d, (J = 8,9) 129,9 / CH 130,0 / CH 6’, 8’ 6,91, d, (J = 8,7) 6,90, d, (J = 8,9) 114,2 / CH 114,3 / CH
7’ 161,4 / C 161,5 / C OMe 3,83, s 3,83, s 55,2 / CH3 55,4 / CH3
Solvente CDCl3 300 MHz
CDCl3 200 MHz
CDCl3 75 MHz
CDCl3 50 MHz
*(MALHEIROS, 2001).

97
5.3.6.4 Identificação do 1-β-O-p-metoxi-E-cinamil-isodrimeninol
A substância DBP-11 foi isolada da fração 11-20 da coluna 9, de D.
brasiliensis, como um sólido amorfo. Foi obtido 204,00 mg, com um rendimento
equivalendo a 0,82% em relação ao extrato bruto. Este foi solúvel em acetona,
clorofórmio, acetato de etila e metanol. Pode-se sugerir que seja da classe dos
sesquiterpenos drimanos sem a presença dos grupamentos aldeídicos, pois não
foram detectados sinais entre 9,00 a 10,00 ppm no espectro de RMN 1H e entre 190,0
a 203,0 ppm no espectro de RMN 13C. A estrutura foi comparada através de análises
espectroscópicas de RMN 1H e 13C com o 1-β-(p-metoxicinamil)-poligodial, 1-β-O-E-
cinamil-6α-hidroxi-isodrimeninol e 1-β-O-p-metoxi-E-cinamil-6α-hidroxi-isodrimeninol
(MALHEIROS, 2001; ALOUCHE, et al., 2009).
Os valores de deslocamentos químicos de RMN 1H e 13C/DEPT 135 (Figuras
41 a 43), indicam que o 1-β-O-p-metoxi-E-cinamil-isodrimeninol possui similaridade
dos deslocamentos dos hidrogênios e carbonos do esqueleto drimano e o grupamento
p-metoxicinamil com o 1-β-(p-metoxicinamil)-poligodial, onde observam-se os
hidrogênios e carbonos aromáticos e olefínicos entre δH 6,00 a 8,00 e δC 114,0 a
168,0, respectivamente. Os três simpletos em δH 1,24, 1,17 e 0,98 e δC 32,8, 24,5 e
9,7, indicam os três grupos metilas. Em δH 3,81 encontra-se um simpleto referente ao
grupo oximetínico.
A ausência dos aldeídos nos carbonos 11 e 12 são as principais diferenças
encontradas. Também destaca-se a mudança do sinal do carbono 1 para 81,3 ppm
indicando a presença de outro substituinte no carbono 9. Utilizou-se uma série de
substâncias relatadas na literatura que apresentavam ausência dos grupamentos
aldeídos nos carbonos 11 e 12 e formação de um anel furano. Desta série de
compostos analisados dois se destacaram o 1-β-O-E-cinamil-6α-hidroxi-isodrimeninol
e o 1-β-O-p-metoxi-E-cinamil-6α-hidroxi-isodrimeninol. O anel furano pode ser
confirmado no DBP-11 pelos sinais dos hidrogênios 11 e 12 em δH 6,10, m e 4,54, s e
carbonos δC 99,6 e 68,1, respectivamente.
Na tabela 6 encontram-se os valores de deslocamentos químicos comparando
o DBP-11 com o 1-β-(p-metoxicinamil)-poligodial, e 1-β-O-p-metoxi-E-cinamil-6α-
hidroxi-isodrimeninol.
Alguns sinais de impurezas de ácidos graxos também podem ser observados.
Neste aspecto, será realizada a sua purificação e posteriormente a obtenção do
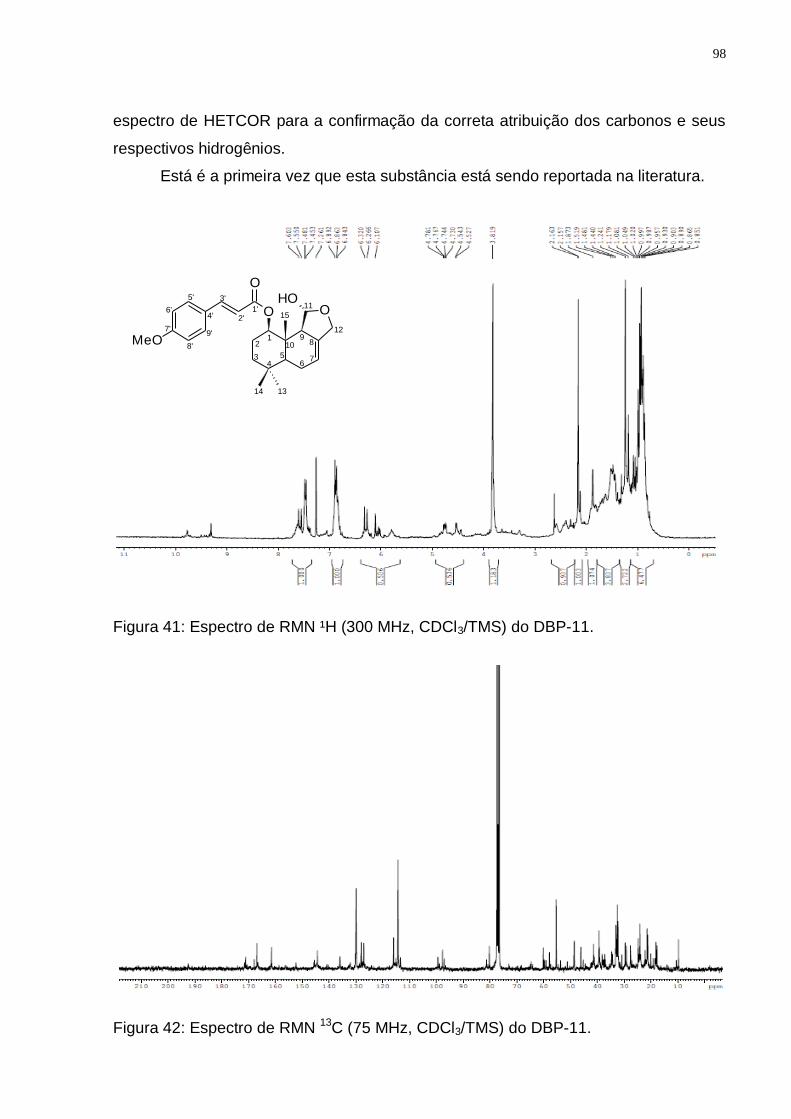
98
espectro de HETCOR para a confirmação da correta atribuição dos carbonos e seus
respectivos hidrogênios.
Está é a primeira vez que esta substância está sendo reportada na literatura.
Figura 41: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBP-11.
Figura 42: Espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBP-11.
O
O
MeO
OOH
12
34
56
7
89
10
11
12
13
151'
2'
3'
4'
5'
6'
7'
8'
9'
14
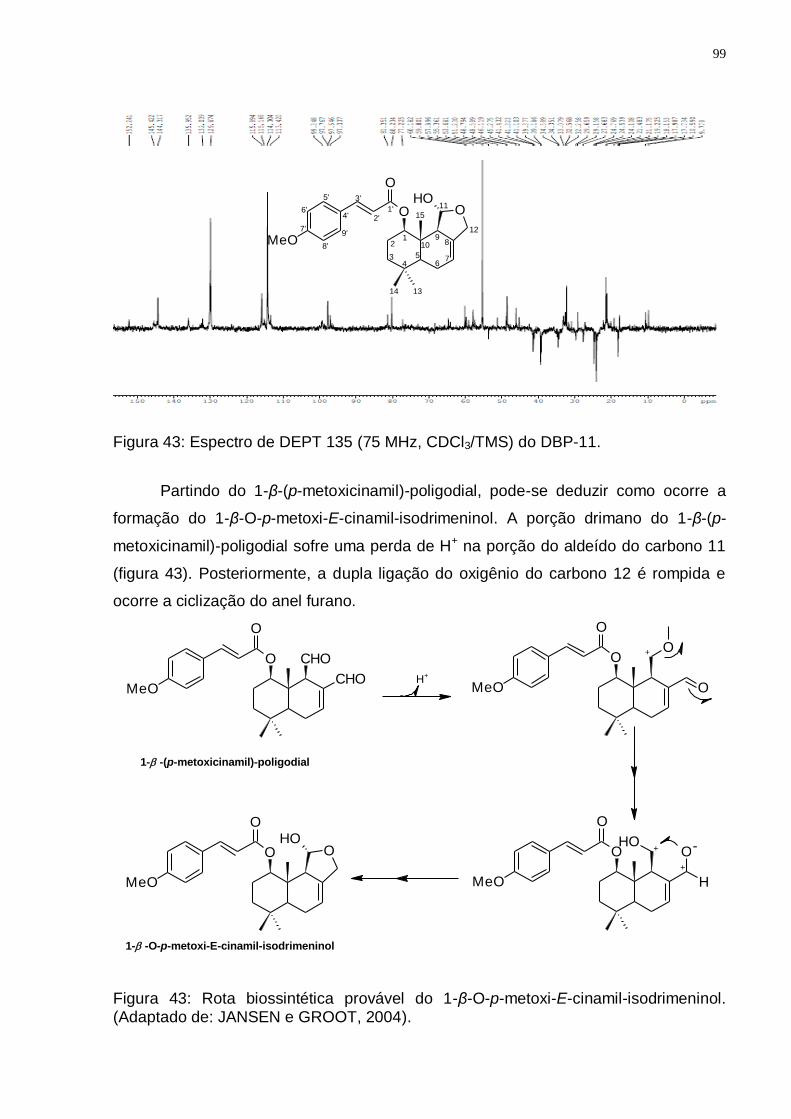
99
Figura 43: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBP-11.
Partindo do 1-β-(p-metoxicinamil)-poligodial, pode-se deduzir como ocorre a
formação do 1-β-O-p-metoxi-E-cinamil-isodrimeninol. A porção drimano do 1-β-(p-
metoxicinamil)-poligodial sofre uma perda de H+ na porção do aldeído do carbono 11
(figura 43). Posteriormente, a dupla ligação do oxigênio do carbono 12 é rompida e
ocorre a ciclização do anel furano.
O
O
MeO
CHO
CHO
1--(p-metoxicinamil)-poligodial
H+
O
O
MeO
O
O
+
O
O
MeO H
OHOH
+ -O
O
MeO
OOH
1--O-p-metoxi-E-cinamil-isodrimeninol
+
Figura 43: Rota biossintética provável do 1-β-O-p-metoxi-E-cinamil-isodrimeninol. (Adaptado de: JANSEN e GROOT, 2004).
O
O
MeO
OOH
12
34
56
7
89
10
11
12
13
151'
2'
3'
4'
5'
6'
7'
8'
9'
14
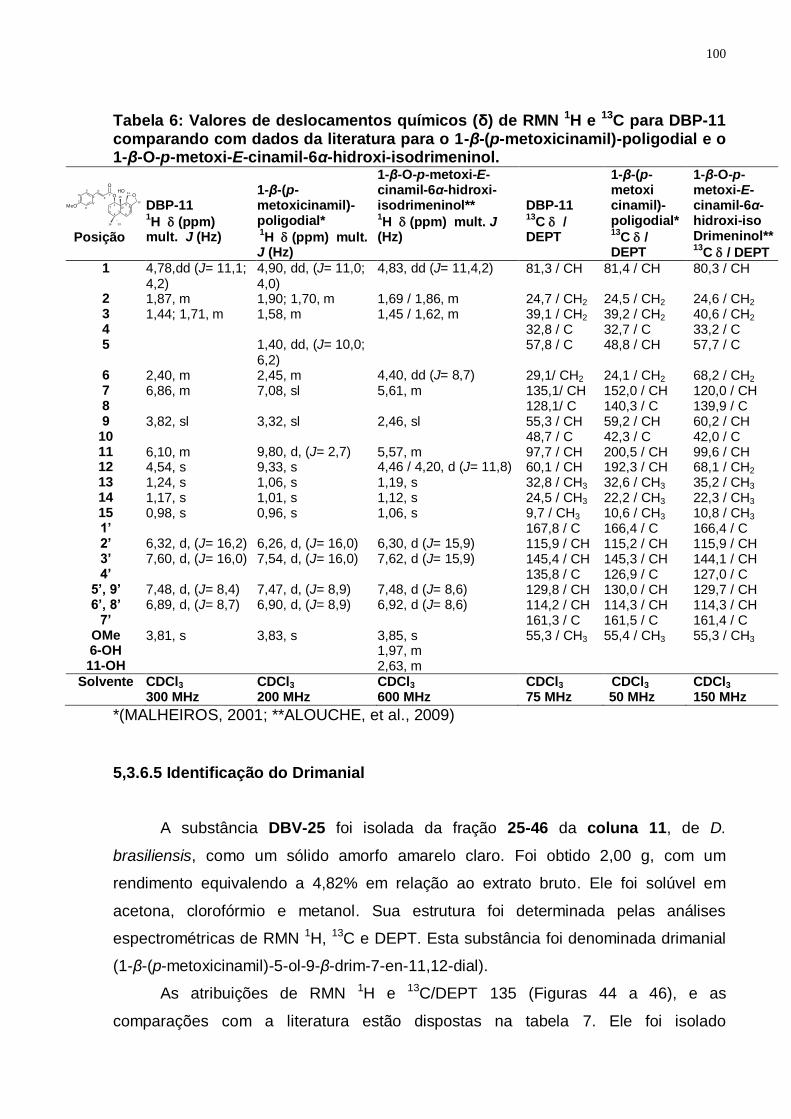
100
Tabela 6: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBP-11 comparando com dados da literatura para o 1-β-(p-metoxicinamil)-poligodial e o 1-β-O-p-metoxi-E-cinamil-6α-hidroxi-isodrimeninol.
O
O
MeO
OOH
12
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
Posição
DBP-11 1H (ppm)
mult. J (Hz)
1-β-(p-metoxicinamil)-poligodial* 1H (ppm) mult.
J (Hz)
1-β-O-p-metoxi-E-cinamil-6α-hidroxi-isodrimeninol** 1H (ppm) mult. J
(Hz)
DBP-11 13
C / DEPT
1-β-(p-metoxi cinamil)-poligodial* 13
C / DEPT
1-β-O-p-metoxi-E-cinamil-6α-hidroxi-iso Drimeninol** 13
C / DEPT 1 4,78,dd (J= 11,1;
4,2) 4,90, dd, (J= 11,0; 4,0)
4,83, dd (J= 11,4,2) 81,3 / CH 81,4 / CH 80,3 / CH
2 1,87, m 1,90; 1,70, m 1,69 / 1,86, m 24,7 / CH2 24,5 / CH2 24,6 / CH2 3 1,44; 1,71, m 1,58, m 1,45 / 1,62, m 39,1 / CH2 39,2 / CH2 40,6 / CH2 4 32,8 / C 32,7 / C 33,2 / C 5 1,40, dd, (J= 10,0;
6,2) 57,8 / C 48,8 / CH 57,7 / C
6 2,40, m 2,45, m 4,40, dd (J= 8,7) 29,1/ CH2 24,1 / CH2 68,2 / CH2 7 6,86, m 7,08, sl 5,61, m 135,1/ CH 152,0 / CH 120,0 / CH 8 128,1/ C 140,3 / C 139,9 / C 9 3,82, sl 3,32, sl 2,46, sl 55,3 / CH 59,2 / CH 60,2 / CH
10 48,7 / C 42,3 / C 42,0 / C 11 6,10, m 9,80, d, (J= 2,7) 5,57, m 97,7 / CH 200,5 / CH 99,6 / CH 12 4,54, s 9,33, s 4,46 / 4,20, d (J= 11,8) 60,1 / CH 192,3 / CH 68,1 / CH2 13 1,24, s 1,06, s 1,19, s 32,8 / CH3 32,6 / CH3 35,2 / CH3 14 1,17, s 1,01, s 1,12, s 24,5 / CH3 22,2 / CH3 22,3 / CH3 15 0,98, s 0,96, s 1,06, s 9,7 / CH3 10,6 / CH3 10,8 / CH3 1’ 167,8 / C 166,4 / C 166,4 / C 2’ 6,32, d, (J= 16,2) 6,26, d, (J= 16,0) 6,30, d (J= 15,9) 115,9 / CH 115,2 / CH 115,9 / CH 3’ 7,60, d, (J= 16,0) 7,54, d, (J= 16,0) 7,62, d (J= 15,9) 145,4 / CH 145,3 / CH 144,1 / CH 4’ 135,8 / C 126,9 / C 127,0 / C
5’, 9’ 7,48, d, (J= 8,4) 7,47, d, (J= 8,9) 7,48, d (J= 8,6) 129,8 / CH 130,0 / CH 129,7 / CH 6’, 8’ 6,89, d, (J= 8,7) 6,90, d, (J= 8,9) 6,92, d (J= 8,6) 114,2 / CH 114,3 / CH 114,3 / CH
7’ 161,3 / C 161,5 / C 161,4 / C OMe 3,81, s 3,83, s 3,85, s 55,3 / CH3 55,4 / CH3 55,3 / CH3 6-OH 1,97, m 11-OH 2,63, m
Solvente CDCl3 300 MHz
CDCl3 200 MHz
CDCl3 600 MHz
CDCl3 75 MHz
CDCl3
50 MHz CDCl3 150 MHz
*(MALHEIROS, 2001; **ALOUCHE, et al., 2009)
5,3.6.5 Identificação do Drimanial
A substância DBV-25 foi isolada da fração 25-46 da coluna 11, de D.
brasiliensis, como um sólido amorfo amarelo claro. Foi obtido 2,00 g, com um
rendimento equivalendo a 4,82% em relação ao extrato bruto. Ele foi solúvel em
acetona, clorofórmio e metanol. Sua estrutura foi determinada pelas análises
espectrométricas de RMN 1H, 13C e DEPT. Esta substância foi denominada drimanial
(1-β-(p-metoxicinamil)-5-ol-9-β-drim-7-en-11,12-dial).
As atribuições de RMN 1H e 13C/DEPT 135 (Figuras 44 a 46), e as
comparações com a literatura estão dispostas na tabela 7. Ele foi isolado
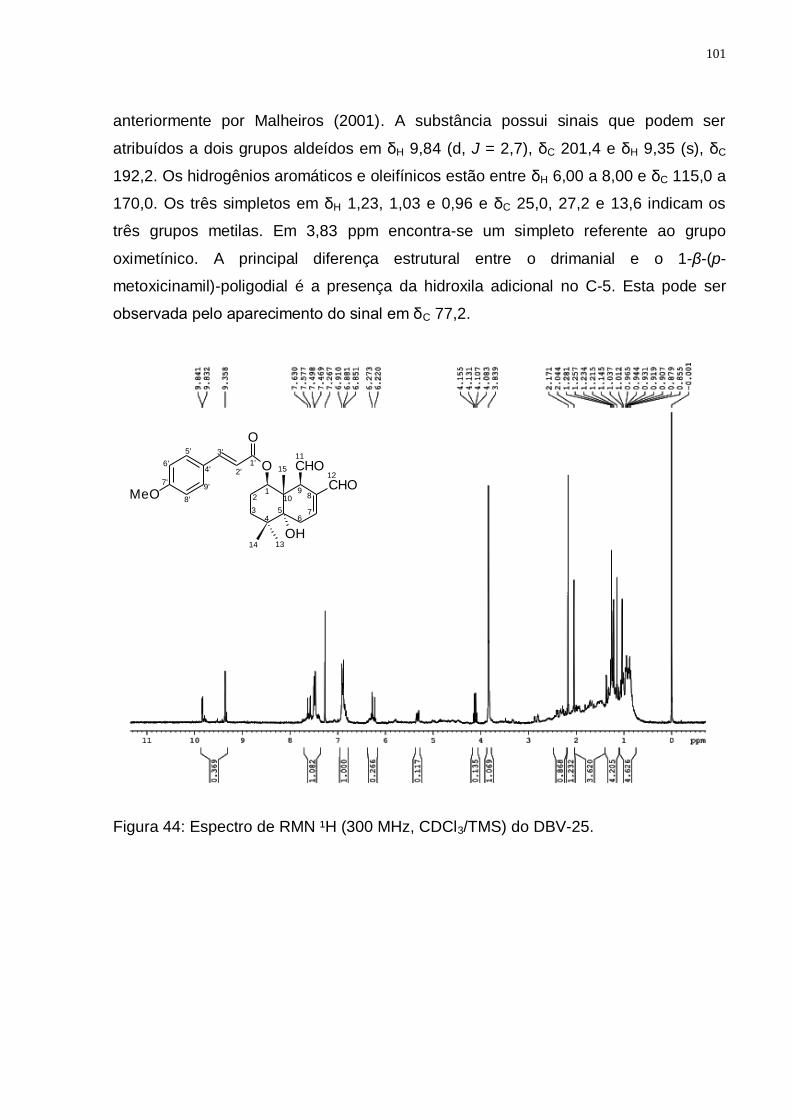
101
anteriormente por Malheiros (2001). A substância possui sinais que podem ser
atribuídos a dois grupos aldeídos em δH 9,84 (d, J = 2,7), δC 201,4 e δH 9,35 (s), δC
192,2. Os hidrogênios aromáticos e oleifínicos estão entre δH 6,00 a 8,00 e δC 115,0 a
170,0. Os três simpletos em δH 1,23, 1,03 e 0,96 e δC 25,0, 27,2 e 13,6 indicam os
três grupos metilas. Em 3,83 ppm encontra-se um simpleto referente ao grupo
oximetínico. A principal diferença estrutural entre o drimanial e o 1-β-(p-
metoxicinamil)-poligodial é a presença da hidroxila adicional no C-5. Esta pode ser
observada pelo aparecimento do sinal em δC 77,2.
Figura 44: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBV-25.
O
O
MeO
CHO
CHO
OH
12
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
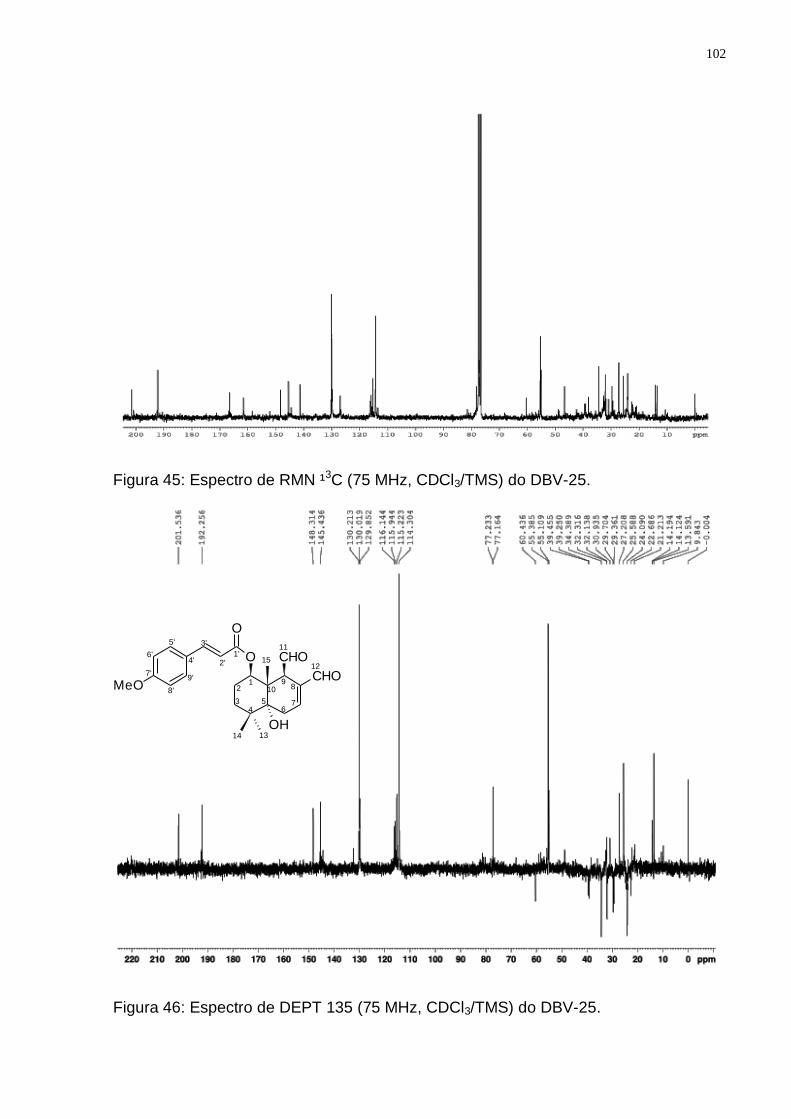
102
Figura 45: Espectro de RMN ¹3C (75 MHz, CDCl3/TMS) do DBV-25.
Figura 46: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBV-25.
O
O
MeO
CHO
CHO
OH
12
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
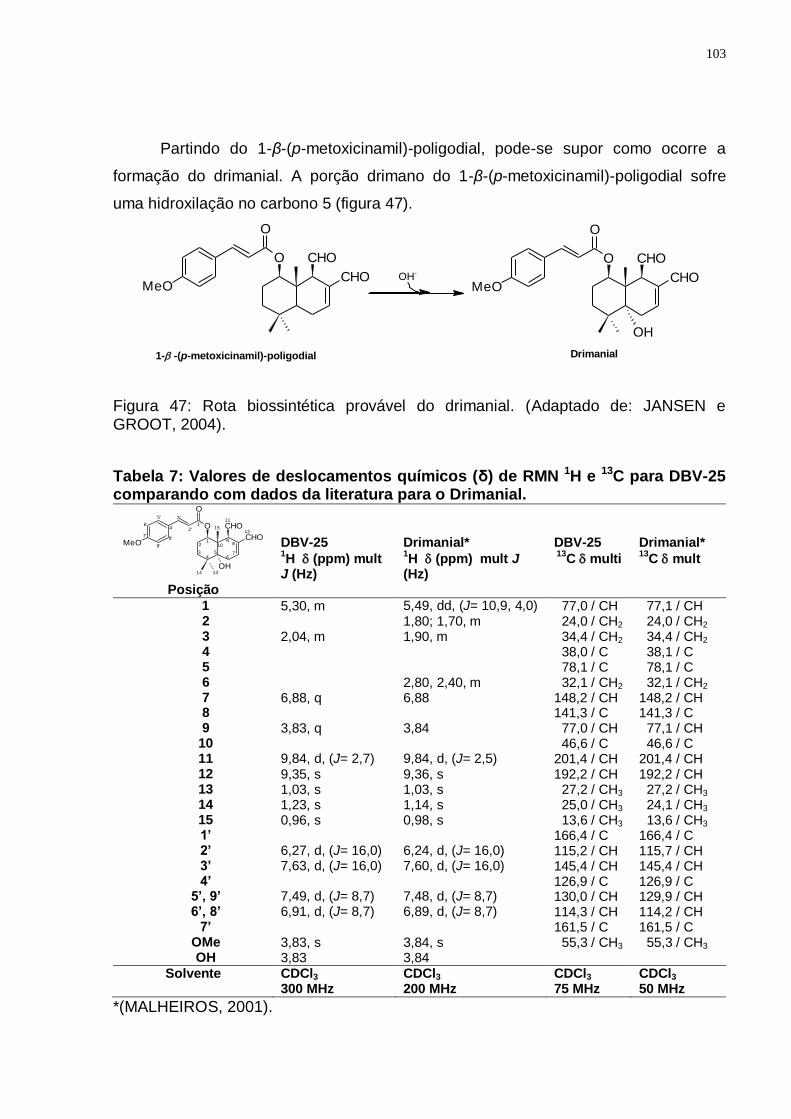
103
Partindo do 1-β-(p-metoxicinamil)-poligodial, pode-se supor como ocorre a
formação do drimanial. A porção drimano do 1-β-(p-metoxicinamil)-poligodial sofre
uma hidroxilação no carbono 5 (figura 47).
O
O
MeO
CHO
CHO
1--(p-metoxicinamil)-poligodial
OH-
O
O
MeO
CHO
CHO
OH
Drimanial
Figura 47: Rota biossintética provável do drimanial. (Adaptado de: JANSEN e GROOT, 2004).
Tabela 7: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBV-25 comparando com dados da literatura para o Drimanial.
O
O
MeO
CHO
CHO
OH
12
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
Posição
DBV-25 1H (ppm) mult
J (Hz)
Drimanial* 1H (ppm) mult J
(Hz)
DBV-25 13
C multi
Drimanial* 13
C mult
1 5,30, m 5,49, dd, (J= 10,9, 4,0) 77,0 / CH 77,1 / CH 2 1,80; 1,70, m 24,0 / CH2 24,0 / CH2 3 2,04, m 1,90, m 34,4 / CH2 34,4 / CH2 4 38,0 / C 38,1 / C 5 78,1 / C 78,1 / C 6 2,80, 2,40, m 32,1 / CH2 32,1 / CH2 7 6,88, q 6,88 148,2 / CH 148,2 / CH 8 141,3 / C 141,3 / C 9 3,83, q 3,84 77,0 / CH 77,1 / CH
10 46,6 / C 46,6 / C 11 9,84, d, (J= 2,7) 9,84, d, (J= 2,5) 201,4 / CH 201,4 / CH 12 9,35, s 9,36, s 192,2 / CH 192,2 / CH 13 1,03, s 1,03, s 27,2 / CH3 27,2 / CH3 14 1,23, s 1,14, s 25,0 / CH3 24,1 / CH3 15 0,96, s 0,98, s 13,6 / CH3 13,6 / CH3 1’ 166,4 / C 166,4 / C 2’ 6,27, d, (J= 16,0) 6,24, d, (J= 16,0) 115,2 / CH 115,7 / CH 3’ 7,63, d, (J= 16,0) 7,60, d, (J= 16,0) 145,4 / CH 145,4 / CH 4’ 126,9 / C 126,9 / C
5’, 9’ 7,49, d, (J= 8,7) 7,48, d, (J= 8,7) 130,0 / CH 129,9 / CH 6’, 8’ 6,91, d, (J= 8,7) 6,89, d, (J= 8,7) 114,3 / CH 114,2 / CH
7’ 161,5 / C 161,5 / C OMe 3,83, s 3,84, s 55,3 / CH3 55,3 / CH3 OH 3,83 3,84
Solvente CDCl3 300 MHz
CDCl3 200 MHz
CDCl3 75 MHz
CDCl3 50 MHz
*(MALHEIROS, 2001).

104
O drimanial foi isolado anteriormente por Malheiros e colaboradores (2001), e
várias atividades biológicas foram atribuídas a sua estrutura como: atividade
antihiperalgésica em modelos de contorções abdominais (CECHINEL-FILHO, et al.,
1998); antiinflamatórias e antialérgicas (CUNHA, et al., 2001); nocicepção em ratos
neonatos (ANDRÉ, et al., 2004); inibição da captação de glutamato pelos astrócitos
(SCHEIDT, et, al., 2002; MARTINI, et al., 2007); ação gastroprotetora via receptores
vanilóides (MATSUDA, et al., 2002); mediador de receptores TRPV1 (ANDRÉ, et al.,
2006); atividade antifúngica (MALHEIROS, et al., 2005) e antitumoral em linhagens de
Hela e L929 (SILVA, et al., 2009).
5.3.6.6 Identificação do 1-β-O-p-metoxi-E-cinamil-5α-hidroxi-isodrimeninol
A substância DBP-21 foi isolada da fração 21-30 da coluna 9, de D.
brasiliensis, como um sólido amorfo. Foi obtido 128,30 mg, com um rendimento
equivalendo a 0,52% em relação ao extrato bruto. Este foi solúvel em acetona,
clorofórmio, acetato de etila e metanol. Pode-se sugerir que seja da classe dos
sesquiterpenos drimanos sem a presença dos grupamentos aldeídicos, pois não
foram detectados sinais entre 9,00 a 10,00 ppm no espectro de RMN 1H e entre 192,2
a 201,4 ppm no espectro de RMN 13C. A estrutura foi comparada através de análises
espectroscópicas de RMN 1H e 13C com o 1-β-(p-metoxicinamil)-poligodial, 1-β-O-E-
cinamil-6α-hidroxi-isodrimeninol e 1-β-O-p-metoxi-E-cinamil-6α-hidroxi-isodrimeninol
(MALHEIROS, 2001; ALOUCHE, et al., 2009).
Os valores de deslocamentos químicos de RMN 1H e 13C/DEPT 135 (Figuras
48 a 50), indicam que o 1-β-O-p-metoxi-E-cinamil-isodrimeninol possui similaridade
dos deslocamentos dos hidrogênios e carbonos do esqueleto drimano e o grupamento
p-metoxicinamil com o 1-β-(p-metoxicinamil)-poligodial, onde observam-se os
hidrogênios e carbonos aromáticos e olefínicos entre δH 6,00 a 8,00 e δC 114,0 a
166,5, respectivamente. Os três simpletos em δH 1,21, 1,15 e 0,98 e δC 32,2, 24,4 e
9,4, indicam os três grupos metilas. Em δH 3,83 encontra-se um simpleto referente ao
grupo oximetínico.
A ausência dos aldeídos nos carbonos 11 e 12 são as principais diferenças
encontradas. Também destaca-se a mudança do sinal do carbono 1 para 81,0 ppm
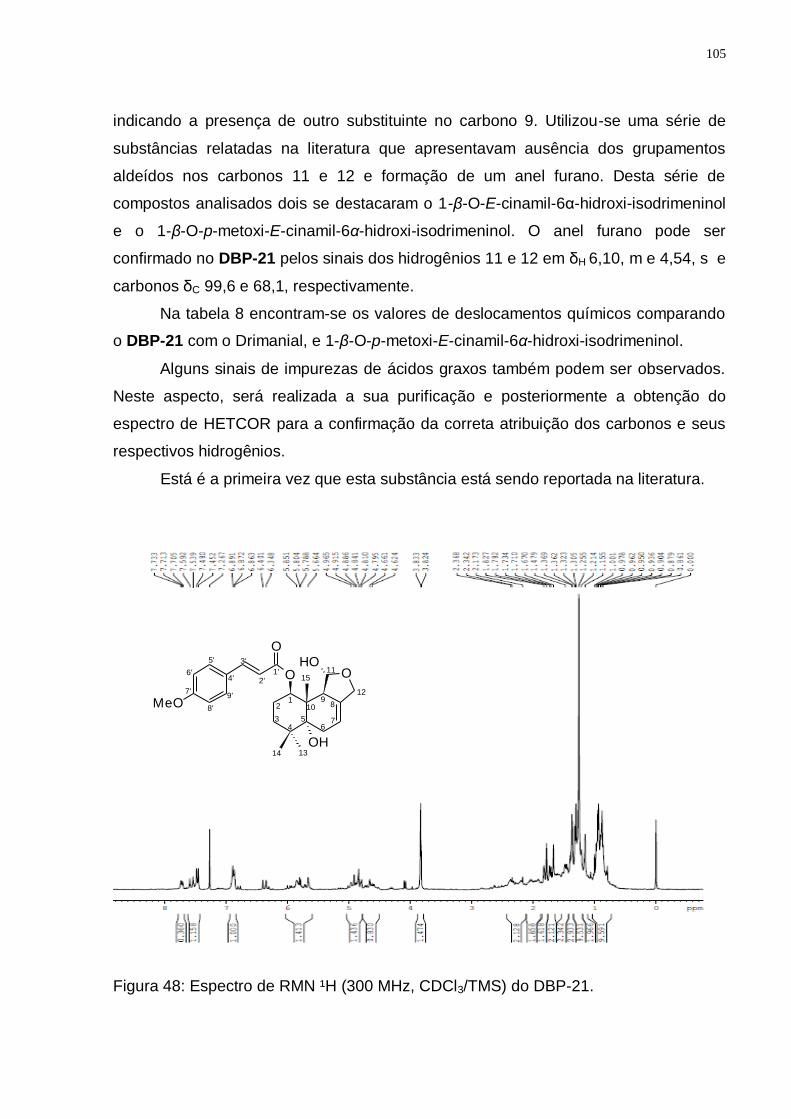
105
indicando a presença de outro substituinte no carbono 9. Utilizou-se uma série de
substâncias relatadas na literatura que apresentavam ausência dos grupamentos
aldeídos nos carbonos 11 e 12 e formação de um anel furano. Desta série de
compostos analisados dois se destacaram o 1-β-O-E-cinamil-6α-hidroxi-isodrimeninol
e o 1-β-O-p-metoxi-E-cinamil-6α-hidroxi-isodrimeninol. O anel furano pode ser
confirmado no DBP-21 pelos sinais dos hidrogênios 11 e 12 em δH 6,10, m e 4,54, s e
carbonos δC 99,6 e 68,1, respectivamente.
Na tabela 8 encontram-se os valores de deslocamentos químicos comparando
o DBP-21 com o Drimanial, e 1-β-O-p-metoxi-E-cinamil-6α-hidroxi-isodrimeninol.
Alguns sinais de impurezas de ácidos graxos também podem ser observados.
Neste aspecto, será realizada a sua purificação e posteriormente a obtenção do
espectro de HETCOR para a confirmação da correta atribuição dos carbonos e seus
respectivos hidrogênios.
Está é a primeira vez que esta substância está sendo reportada na literatura.
Figura 48: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBP-21.
O
O
MeO
OH
OOH
12
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
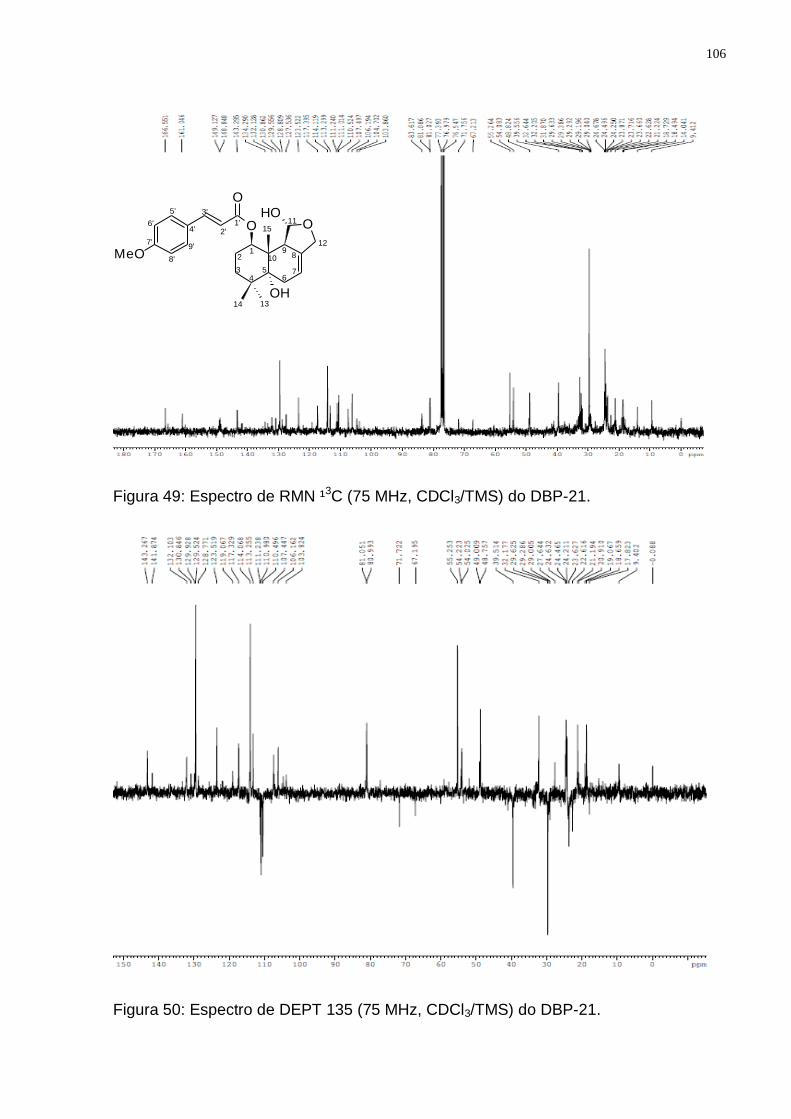
106
Figura 49: Espectro de RMN ¹3C (75 MHz, CDCl3/TMS) do DBP-21.
Figura 50: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBP-21.
O
O
MeO
OH
OOH
12
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
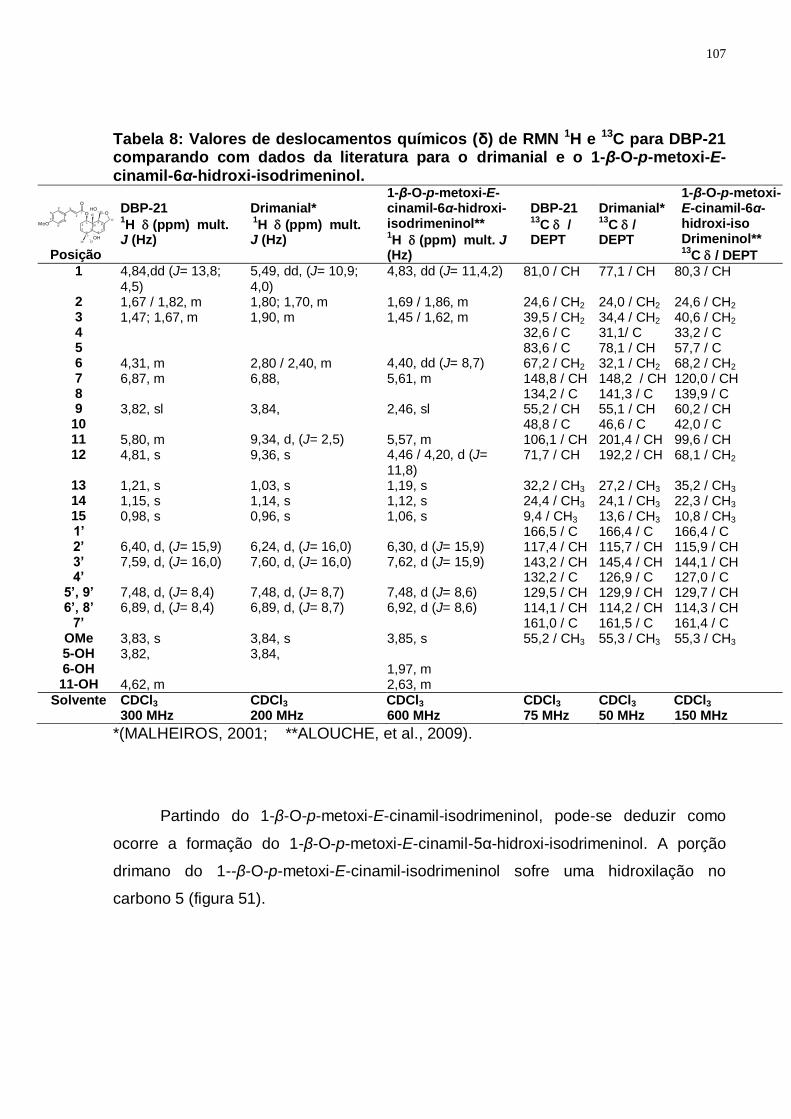
107
Tabela 8: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBP-21 comparando com dados da literatura para o drimanial e o 1-β-O-p-metoxi-E-cinamil-6α-hidroxi-isodrimeninol.
O
O
MeO
OH
OOH
12
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
Posição
DBP-21 1H (ppm) mult.
J (Hz)
Drimanial* 1H (ppm) mult.
J (Hz)
1-β-O-p-metoxi-E-cinamil-6α-hidroxi-isodrimeninol** 1H (ppm) mult. J
(Hz)
DBP-21 13
C / DEPT
Drimanial* 13
C / DEPT
1-β-O-p-metoxi-E-cinamil-6α-hidroxi-iso Drimeninol** 13
C / DEPT 1 4,84,dd (J= 13,8;
4,5) 5,49, dd, (J= 10,9; 4,0)
4,83, dd (J= 11,4,2) 81,0 / CH 77,1 / CH 80,3 / CH
2 1,67 / 1,82, m 1,80; 1,70, m 1,69 / 1,86, m 24,6 / CH2 24,0 / CH2 24,6 / CH2 3 1,47; 1,67, m 1,90, m 1,45 / 1,62, m 39,5 / CH2 34,4 / CH2 40,6 / CH2 4 32,6 / C 31,1/ C 33,2 / C 5 83,6 / C 78,1 / CH 57,7 / C 6 4,31, m 2,80 / 2,40, m 4,40, dd (J= 8,7) 67,2 / CH2 32,1 / CH2 68,2 / CH2 7 6,87, m 6,88, 5,61, m 148,8 / CH 148,2 / CH 120,0 / CH 8 134,2 / C 141,3 / C 139,9 / C 9 3,82, sl 3,84, 2,46, sl 55,2 / CH 55,1 / CH 60,2 / CH
10 48,8 / C 46,6 / C 42,0 / C 11 5,80, m 9,34, d, (J= 2,5) 5,57, m 106,1 / CH 201,4 / CH 99,6 / CH 12 4,81, s 9,36, s 4,46 / 4,20, d (J=
11,8) 71,7 / CH 192,2 / CH 68,1 / CH2
13 1,21, s 1,03, s 1,19, s 32,2 / CH3 27,2 / CH3 35,2 / CH3 14 1,15, s 1,14, s 1,12, s 24,4 / CH3 24,1 / CH3 22,3 / CH3 15 0,98, s 0,96, s 1,06, s 9,4 / CH3 13,6 / CH3 10,8 / CH3 1’ 166,5 / C 166,4 / C 166,4 / C 2’ 6,40, d, (J= 15,9) 6,24, d, (J= 16,0) 6,30, d (J= 15,9) 117,4 / CH 115,7 / CH 115,9 / CH 3’ 7,59, d, (J= 16,0) 7,60, d, (J= 16,0) 7,62, d (J= 15,9) 143,2 / CH 145,4 / CH 144,1 / CH 4’ 132,2 / C 126,9 / C 127,0 / C
5’, 9’ 7,48, d, (J= 8,4) 7,48, d, (J= 8,7) 7,48, d (J= 8,6) 129,5 / CH 129,9 / CH 129,7 / CH 6’, 8’ 6,89, d, (J= 8,4) 6,89, d, (J= 8,7) 6,92, d (J= 8,6) 114,1 / CH 114,2 / CH 114,3 / CH
7’ 161,0 / C 161,5 / C 161,4 / C OMe 3,83, s 3,84, s 3,85, s 55,2 / CH3 55,3 / CH3 55,3 / CH3 5-OH 3,82, 3,84, 6-OH 1,97, m 11-OH 4,62, m 2,63, m
Solvente CDCl3 300 MHz
CDCl3 200 MHz
CDCl3 600 MHz
CDCl3 75 MHz
CDCl3
50 MHz CDCl3 150 MHz
*(MALHEIROS, 2001; **ALOUCHE, et al., 2009).
Partindo do 1-β-O-p-metoxi-E-cinamil-isodrimeninol, pode-se deduzir como
ocorre a formação do 1-β-O-p-metoxi-E-cinamil-5α-hidroxi-isodrimeninol. A porção
drimano do 1--β-O-p-metoxi-E-cinamil-isodrimeninol sofre uma hidroxilação no
carbono 5 (figura 51).

108
O
O
MeO
OOH
1--O-p-metoxi-E-cinamil-isodrimeninol
O
O
MeO
OOH
OH1--O-p-metoxi-E-cinamil-5-hidroxi-isodrimeninol
OH-
Figura 51: Rota biossintética provável do 1-β-O-p-metoxi-E-cinamil-5α-hidroxi-isodrimeninol. (Adaptado de: JANSEN e GROOT, 2004; ALOUCHE, et al., 2009).
5.3.6.7 Identificação do 1-β-(p-cumaroiloxi)-poligodial
A substância DBV-50 foi isolada da fração 50-56 da coluna 11, de D.
brasiliensis. Foi obtido aproximadamente, 175,34 mg, com um rendimento
equivalendo a 0,71% em relação ao extrato bruto. Este foi solúvel em clorofórmio,
acetato de etila, metanol e acetona. Sua estrutura foi confirmada como sendo 1-β-(p-
cumaroiloxi)-poligodial (1-β-(p-cumaroiloxi)-9-β-drim-7-en-11,12-dial). Ele havia sido
isolado anteriormente das cascas de Drimys brasiliensis, e D. angustifolia
(MALHEIROS, 2001).
Os valores dos deslocamentos químicos de ¹H e ¹³C e DEPT 135 (Figuras 52 a
54) indicam que a substância DBV-50 é um sesquiterpeno drimano semelhante ao 1-
β-(p-metoxicinamil)-poligodial. Observam-se sinais para os dois grupos aldeídicos em
δ 9,75 (d, J = 3,0) e 9,32 (s). O sinal para o metino adjacente ao aldeído é verificado
em δ 3,29 (sl) e o próton oleofínico em δ 7,16 (sl). Porém não é observado no
espectro de RMN ¹H o singleto em δ 3,83, o que caracteriza o grupamento OH em C-
7’. A tabela 9 mostra os valores e deslocamentos químicos de RMN de ¹H e ¹³C
suportados pelo DEPT 135 para o DBV-50 e dados da literatura para 1-β-(p-
cumaroiloxi)-poligodial.
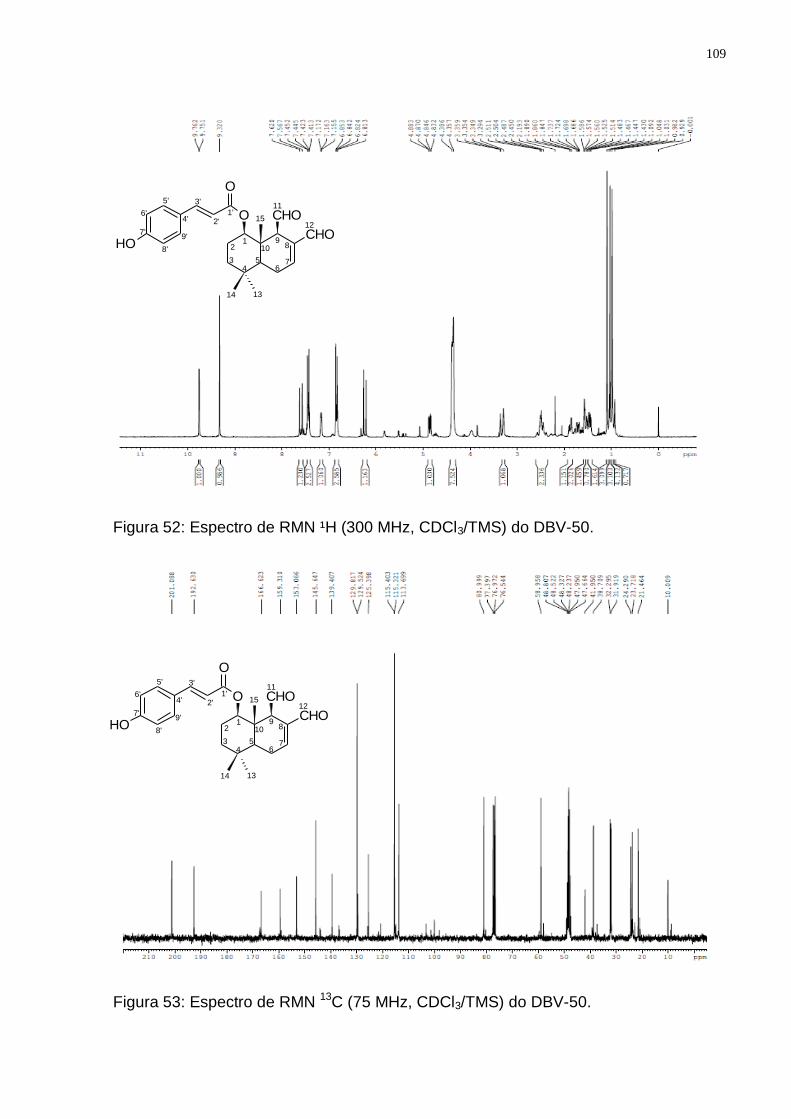
109
Figura 52: Espectro de RMN ¹H (300 MHz, CDCl3/TMS) do DBV-50.
Figura 53: Espectro de RMN 13C (75 MHz, CDCl3/TMS) do DBV-50.
12
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
O
O
OH
CHO
CHO
12
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
O
O
OH
CHO
CHO
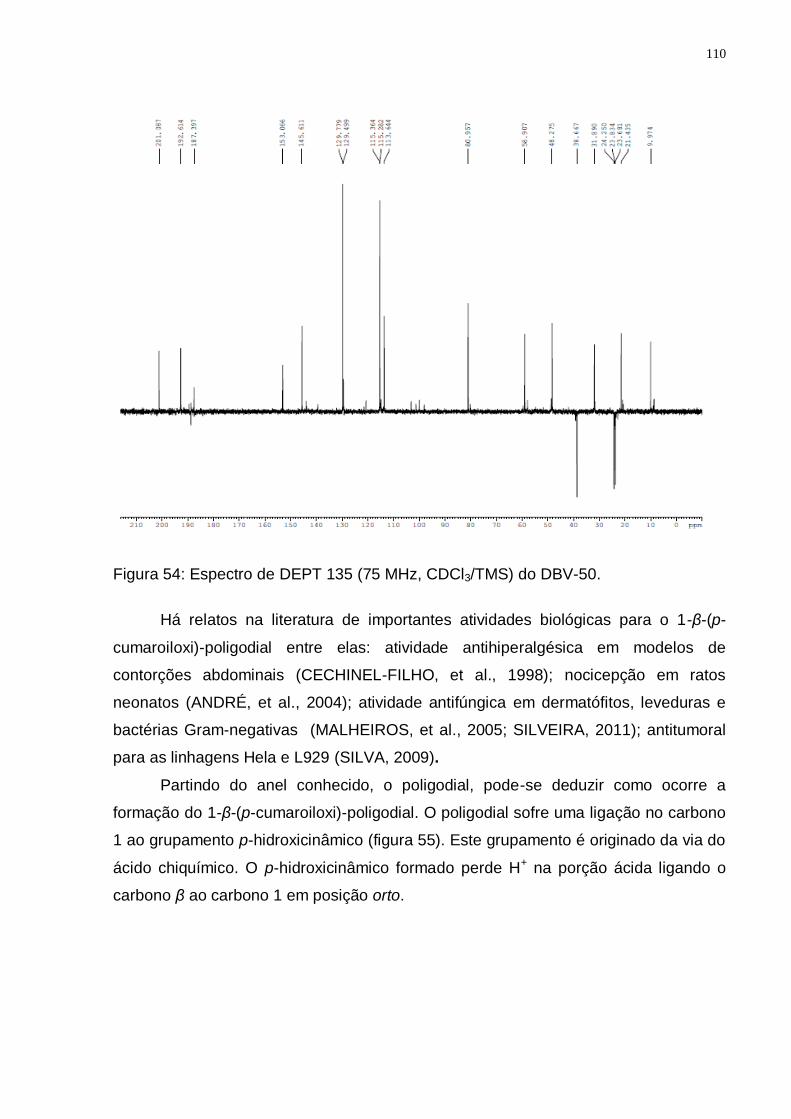
110
Figura 54: Espectro de DEPT 135 (75 MHz, CDCl3/TMS) do DBV-50.
Há relatos na literatura de importantes atividades biológicas para o 1-β-(p-
cumaroiloxi)-poligodial entre elas: atividade antihiperalgésica em modelos de
contorções abdominais (CECHINEL-FILHO, et al., 1998); nocicepção em ratos
neonatos (ANDRÉ, et al., 2004); atividade antifúngica em dermatófitos, leveduras e
bactérias Gram-negativas (MALHEIROS, et al., 2005; SILVEIRA, 2011); antitumoral
para as linhagens Hela e L929 (SILVA, 2009).
Partindo do anel conhecido, o poligodial, pode-se deduzir como ocorre a
formação do 1-β-(p-cumaroiloxi)-poligodial. O poligodial sofre uma ligação no carbono
1 ao grupamento p-hidroxicinâmico (figura 55). Este grupamento é originado da via do
ácido chiquímico. O p-hidroxicinâmico formado perde H+ na porção ácida ligando o
carbono β ao carbono 1 em posição orto.
111
CHO
CHO
H
OH
O
OH
O
OH
OH-
+
O
O
OH
CHO
CHO
p-hidroxicinâmico
H+
p-hidroxicinamil
Poligodial1--(p-cumaroiloxi)-poligodial
Figura 55: Rota biossintética provável do 1-β-(p-cumaroiloxi)-poligodial (Adaptado de: JANSEN e GROOT, 2004).
Tabela 9: Valores de deslocamentos químicos (δ) de RMN 1H e 13C para DBV-50 e dados da literatura para o 1-β-(p-cumaroiloxi)-poligodial.
12
34
56
7
89
10
11
12
1314
151'
2'
3'
4'
5'
6'
7'
8'
9'
O
O
OH
CHO
CHO
Posição
DBV-50 1H δ (ppm) mult. J
(Hz)
1-β-(p-cumaroiloxi)-poligodial)* 1H δ (ppm) mult. J
(Hz)
DBV-50 13
C δ (ppm)
1-β-(p-cumaroiloxi)-poligodial)* 13
C δ (ppm)
1 4,88, dd, (J= 9,0; 3,0) 4,89, dd, (J= 10,5, 4,6)
81,5 / CH 81,9 / CH
2 1,86; 1,85, m 1,82; 1,71, m 23,3 / CH2 24,6 / CH2 3 1,57, m 1,58, m 39,9 / CH2 39,6 / CH2 4 33,4 / C 33,2 / C 5 1,43, dd, (J= 12,0, 6,0) 1,44, dd, (J= 11,0,
6,0) 48,8 / CH 49,2 / CH
6 2,49, m 2,48, m 24,9 / CH2 25,2 / CH2 7 7,16, sl 7,09, sl 153,3 / CH 154,0 / CH 8 140,9 / C 140,3 / C 9 3,29, sl 3,34, sl 58,9 / CH 59,8 / CH
10 42,3 / C 42,8 / C 11 9,75, d, (J= 3,0) 9,82, d, (J= 2,8) 200,1 / CH 201,9 / CH 12 9,32, s 9,35, s 193,2 / CH 193,5 / CH 13 1,09, s 1,09, s 32,7 / CH3 32,8 / CH3 14 1,03, s 1,03, s 22,2 / CH3 22,3 / CH3 15 0,98, s 0,98, s 10,9 / CH3 10,9 / CH3 1’ 166,6 / C 167,5 / C 2’ 6,26, d, (J= 16,0) 6,23, d, (J= 16,0) 115,9 / CH 114,6 / CH 3’ 7,62, d, (J= 16,0) 7,69, d, (J= 16,0) 145,6 / CH 148,5 / CH 4’ 127,0 / C 126,3 / C
5’, 9’ 6,85, d, (J= 8,7) 6,87, d, (J= 8,8) 131,0 / CH 130,7 / CH 6’, 8’ 7,42, d, (J= 8,7) 7,39, d, (J= 8,8) 116,6 / CH 116,3 / CH
7’ 160,6 / C 160,2 / C Solvente
CDCl3/ MeOD 75 MHz
CDCl3 200 MHz
CDCl3/ MeOD 300 MHz
CDCl3 50 MHz
*(MALHEIROS, 2001).

112
Os sesquiterpenos drimanos derivam dos terpenóides. Esta classe de
substâncias é conhecida por suas importantes funções biológicas e fisiológicas, fato
este que justifica o seu uso na área farmacêutica. Acreditava-se que os terpenóides
em plantas superiores eram sintetizados a partir da via do acetato-mevalonato a partir
da conjugação de unidades do isopreno ligando-se orientados no sentido inverso
(cabeça-cauda). Mais recentemente, com a descoberta da via do não mevalonato, o
conceito de biossíntese passou por algumas modificações; a via principal, via do
acetato-mevalonato, age ao nível citoplasmático, mais precisamente nas mitocôndrias
sintetizando preferencialmente esteróides e sesquiterpenos. Já a via secundária, via
do não mevalonato age nos plastídios sintetizando todas as outras classes de
terpenos, incluindo os carotenóides e as clorofilas (DEWICK, 2009; DE LAS HERAS,
et al., 2009).
Até o momento são conhecidos cerca de 30.000 terpenos, que são
classificados de acordo com o número de unidades isoprênicas: hemiterpenóides (C5),
monoterpenóides (C10), sesquiterpenóides (C15), diterpenóides (C20), triterpenóides
(C30) e tetraterpenóides (C40) (DUBEY, et al., 2009).
Nos vegetais eles apresentam diversas funções: os monoterpenos são os
principais constituintes dos óleos voláteis, atuando na atração dos polinizadores; os
sesquiterpenos atuam nos sistemas de defesa das plantas contra fungos, bactérias e
insetos, com características deterrentes; e os diterpenóides originam os hormônios
vegetais importantes para as mais variadas funções dentro da planta. Os triterpenos e
os derivados esteroidais também atuam na proteção contra herbivoria, podendo
alguns serem antimitóticos, influenciando na germinação de sementes e na inibição
do crescimento da raíz, ou seja, possuem características alelopáticas (VICKERY;
VICKERY, 1981; HARBONE; BAXTER, 1995; VERPOORTE, 2009; MAHMOUD;
CROTEAU, 2009; GERSHENZON; DUDAREVA, 2009).
A unidade isoprênica é produzida naturalmente, a partir do ácido mevalônico.
Este é duplamente fosforilado por ATP levando ao ácido 5-pirofosfato-mevalônico.
Sua desidratação e descarboxilação produzem o pirofosfato de isopentenila, que sofre
isomerização ao pirofosfato de dimetilalila (figura 56) (GEISSMAN; CROUT, 1969;
MAGRI, 1999).
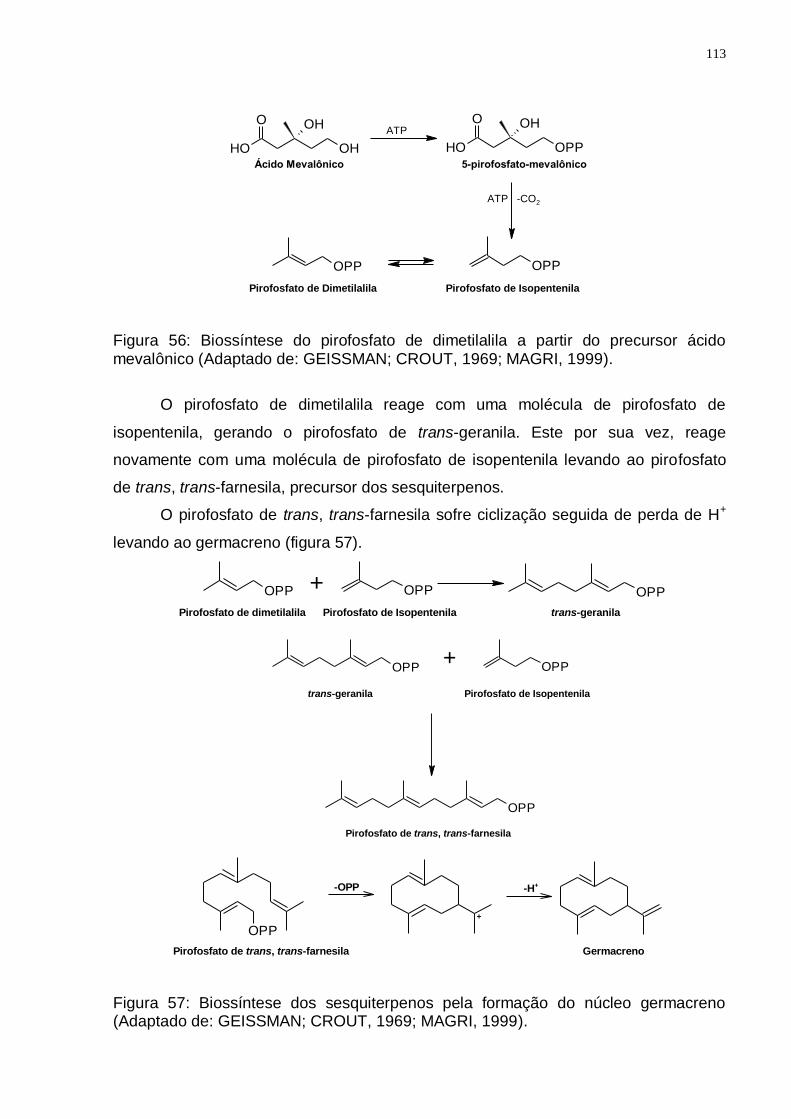
113
OH OH
O OHATP
OH OPP
O OH
OPPOPP
ATP -CO2
Ácido Mevalônico 5-pirofosfato-mevalônico
Pirofosfato de Dimetilalila Pirofosfato de Isopentenila
Figura 56: Biossíntese do pirofosfato de dimetilalila a partir do precursor ácido mevalônico (Adaptado de: GEISSMAN; CROUT, 1969; MAGRI, 1999).
O pirofosfato de dimetilalila reage com uma molécula de pirofosfato de
isopentenila, gerando o pirofosfato de trans-geranila. Este por sua vez, reage
novamente com uma molécula de pirofosfato de isopentenila levando ao pirofosfato
de trans, trans-farnesila, precursor dos sesquiterpenos.
O pirofosfato de trans, trans-farnesila sofre ciclização seguida de perda de H+
levando ao germacreno (figura 57).
OPP OPP+ OPP
Pirofosfato de dimetilalila Pirofosfato de Isopentenila trans-geranila
OPPOPP
OPP
+
Pirofosfato de trans, trans-farnesila
trans-geranila Pirofosfato de Isopentenila
OPP
+
Pirofosfato de trans, trans-farnesila Germacreno
-OPP -H+
Figura 57: Biossíntese dos sesquiterpenos pela formação do núcleo germacreno (Adaptado de: GEISSMAN; CROUT, 1969; MAGRI, 1999).

114
O germacreno pode levar a formação de todos os outros esqueletos
sesquiterpênicos por transformações diversas, com ciclização, rompimento do anel,
rearranjo e migração do grupo metila (GEISSMAN; CROUT, 1969; MAGRI, 1999).
O esqueleto drimano (figura 58) é formado pelo fechamento do anel do
precursor germacreno. Logo após ocorre um rearranjo estrutural com a migração do
grupamento metila do carbono C-4 para o C-3 e a ligação da dupla exocíclica
permanece no C-4 (JANSEN e GROOT, 1991).
Germacreno
Rearranjo Drimano
Figura 58: Rearranjo drimano do precursor germacreno (Adaptado de: JANSEN e GROOT, 1991).
Os drimanos estão presentes em diversas famílias, dentre elas a Winteraceae,
principalmente no gênero Drimys. Mas a ocorrência dos drimanos não está limitada
apenas as plantas, pois, já foram encontradas também em outros organismos como
os fungos, moluscos e esponjas (JANSEN e GROOT, 2004).
Esta classe de substâncias vem sendo extensivamente estudadas para o
tratamento de diversas doenças, inclusive o câncer (FORSBY; WALUM, 1996;
FORSBY, et al., 1998; SILVA, 2009). Na literatura outras atividades biológicas
comprovadas cientificamente são relatadas, incluindo atividades antiinflamatórias
(MENDES, et al., 1998; CECHINEL FILHO, et al., 1998; EL SAYAH, et al., 2000;
CUNHA, et al., 2001; MALHEIROS, et al., 2001; MATSUDA, et al., 2002; SCHEIDT, et
al., 2002; ANDRÉ, et al., 1999; 2004; CASTELLI, et al., 2005; MARTINI, et al., 2007;
WAITAINCENIS, et al., 2007), antibacterianas (MACHIDA, et al., 1999; FUJITA e
KUBO, 2005; MALHEIROS, et al., 2005; SILVEIRA, 2011), molusculicida, piscicida,
antihelmíntico e reguladora do crescimento (MORTON, 1981; SIMÕES, et al., 1986;
JANSEN e GROOT, 2004).

115
5.4 Atividade biológica
5.4.1 Avaliação da citotoxicidade
A ação citotóxica do extrato clorofórmico das cascas, caules e folhas da D.
brasiliensis, bem como algumas substâncias isoladas, foram avaliados nas linhagens
tumorais de células de leucemia mielóide (K562), de linfóide aguda B (Nalm6), de
adenocarcinoma mamário humano (MCF-7) e de melanoma murino (B16F10). Na
tabela 10 e 11 estão apresentados os resultados obtidos.
Tabela 10: Avaliação da viabilidade celular de K562, Nalm6, MCF-7 e B16F10 em extratos brutos.
Amostra Drimys brasiliensis
K562 Nalm6 MCF-7 B16F10
CI50 (µg/mL) CI50 (µg/mL) CI50 (µg/mL) CI50 (µg/mL)
DBCHCl3 Cascas 21,36 ± 0,81 4,23 ± 0,20 23,60 ± 1,50 9,35 ± 4,95 DBCHCl3 Caules 41,03 ± 1,56 16,04 ± 1,10 --------- -------- DBCHCl3 Folhas 67,66 ± 2,30 19,11 ± 0,95 --------- --------
Vincristina 2,97 ± 0,66 0,123 ± 0,001 --------- --------
Paclitaxel --------- --------- 0,98 ± 0,4 0,31 +0,032
Todos os extratos avaliados apresentaram resultados promissores para a
linhagens de células leucêmicas Nalm6 com CI50 entre 4 e 20 µg/mL e para K562
entre 20 e 70 µg/mL. Entre eles, o extrato clorofórmico das cascas apresentou os
melhores resultados com CI50 de 4,23 µg/mL para Nalm6 e 21,36 µg/mL para K562.
Os extratos foram menos ativos que o controle positivo, porém tratando-se de
extratos, os resultados podem ser considerados bons a moderados.
Segundo Cañedo (2005) a linhagem celular K562 é utilizada em experimentos
que envolvem o mecanismo de diferenciação do câncer, ou seja, uma forma de
avaliar o tratamento de células neoplásicas através de agentes indutores da
diferenciação celular. Esta linhagem celular em especial apresenta defeitos nas vias
de ativação das caspases, havendo um aumento nos níveis dos inibidores de
caspase-8 (FLIP) e uma baixa nos níveis da APAF-1, quando comparadas com
outras linhagens celulares, como, por exemplo, a Jurkat. Também é observado que
esta linhagem também é sensível aos inibidores da COX-2, com ativação de células
G0/G1 e da caspase-8 (PENG, et al., 2008).
Devido os resultados obtidos para o extrato clorofórmico das cascas, estes
foram avaliados para as linhagens de adenocarcinoma mamário humano (MCF-7) e

116
de melanoma murino (B16F10). Este também apresentou resultados significativos
para estas células com valores de CI50 de 23,60 e 9,35 µg/mL, respectivamente. O
câncer de pele melanoma cutâneo é resultante da modificação dos melanócitos que
compõem a pele, cuja principal função é a produção de melanina, um pigmento
natural muito importante para a fotoproteção da pele (DE VITA; HELLMANN;
ROSENBERG, 2005). No geral é o tipo de câncer que apresenta grande potencial
metastático, mesmo em fases iniciais de sua origem, fato este que constitui numa
das patologias mais agressivas dentre todas as lesões de pele, incluindo um alto
índice de mortalidade, quando diagnosticado tardiamente (INCA, 2010).
A prevenção segundo o INCA (2010) inclui ações primárias, como proteção
contra a luz solar, que são efetivas e de baixo custo. O auto-exame também é
essencial para o diagnóstico precoce da patologia. No caso do portador o tratamento
consiste em excisão cirúrgica, quimioterapia, imunoterapia, e uso de medicamentos,
como por exemplo, vincristina, vimblastina, paclitaxel e docetaxol, que no geral são
bastante agressivos e com efeitos adversos. Seu uso em longo prazo pode acarretar
em danos ao sistema fisiológico (GOLDMANN; AUSIELLO, 2005).
As substâncias isoladas, exceto as lignanas, foram testadas nas mesmas
linhagens tumorais e sob as mesmas condições dos extratos. Todos apresentaram
bons resultados frente às células testadas. Dentre as substâncias testadas o 1-β-(p-
metoxicinamil)-poligodial apresentou os melhores resultados com CI50 de 2,80; 0,22;
10,10 e 7,70 µM para as linhagens K562, Nalm6, MCF-7 e B16F10,
respectivamente.
Dentre os drimanos avaliados, quando a atividade é comparada entre o 1-β-
(p-metoxicinamil)-poligodial e o drimanial, verificou-se que a introdução do
grupamento OH no carbono C-5 de drimanial, a atividade diminuiu ligeiramente. Já o
1-β-(p-cumaroiloxi)-poligodial possui um grupamento OH no carbono C-7’ e com a
introdução deste grupamento tornou-se menos efetivo que os demais. É possível
que o grupamento OH tanto na posição C-7’ quanto na posição C-5 afete na ligação
da molécula ao receptor da célula e que o grupamento metoxi na posição C-7’
favoreça a ligação no receptor, e com isso, o efeito citotóxico se apresenta.
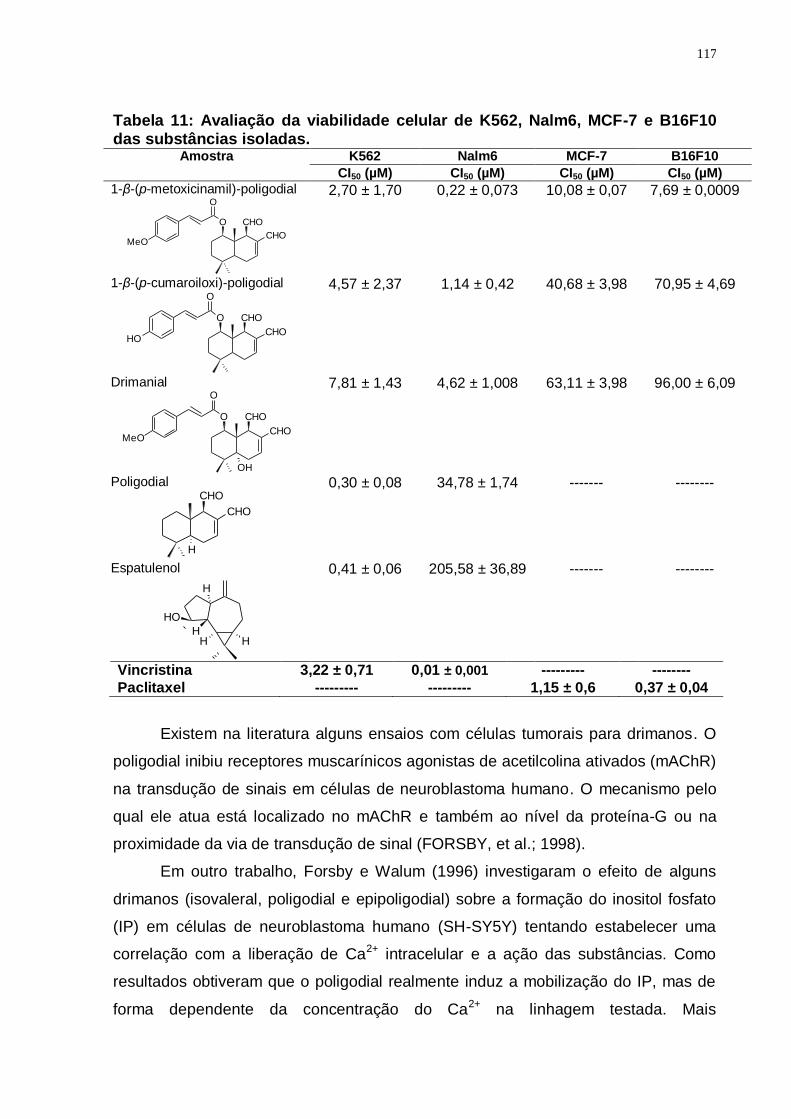
117
Tabela 11: Avaliação da viabilidade celular de K562, Nalm6, MCF-7 e B16F10 das substâncias isoladas.
Amostra
K562 Nalm6 MCF-7 B16F10
CI50 (µM) CI50 (µM) CI50 (µM) CI50 (µM)
1-β-(p-metoxicinamil)-poligodial
O
O
MeO
CHO
CHO
2,70 ± 1,70
0,22 ± 0,073
10,08 ± 0,07
7,69 ± 0,0009
1-β-(p-cumaroiloxi)-poligodial
O
O
OH
CHO
CHO
4,57 ± 2,37
1,14 ± 0,42
40,68 ± 3,98
70,95 ± 4,69
Drimanial
O
O
MeO
CHO
CHO
OH
7,81 ± 1,43
4,62 ± 1,008
63,11 ± 3,98
96,00 ± 6,09
Poligodial CHO
CHO
H
0,30 ± 0,08
34,78 ± 1,74
------- --------
Espatulenol
H
HHH
OH
0,41 ± 0,06
205,58 ± 36,89
------- --------
Vincristina 3,22 ± 0,71 0,01 ± 0,001 --------- --------
Paclitaxel --------- --------- 1,15 ± 0,6 0,37 ± 0,04
Existem na literatura alguns ensaios com células tumorais para drimanos. O
poligodial inibiu receptores muscarínicos agonistas de acetilcolina ativados (mAChR)
na transdução de sinais em células de neuroblastoma humano. O mecanismo pelo
qual ele atua está localizado no mAChR e também ao nível da proteína-G ou na
proximidade da via de transdução de sinal (FORSBY, et al.; 1998).
Em outro trabalho, Forsby e Walum (1996) investigaram o efeito de alguns
drimanos (isovaleral, poligodial e epipoligodial) sobre a formação do inositol fosfato
(IP) em células de neuroblastoma humano (SH-SY5Y) tentando estabelecer uma
correlação com a liberação de Ca2+ intracelular e a ação das substâncias. Como
resultados obtiveram que o poligodial realmente induz a mobilização do IP, mas de
forma dependente da concentração do Ca2+ na linhagem testada. Mais

118
recentemente, Silva (2009), avaliou a ação citotóxica dos extratos clorofórmicos das
cascas, caules e folhas, além dos sesquiterpenos drimanos drimanial, 1-β-(p-
metoxicinamil)-poligodial, isolados do extrato clorofórmico das cascas, em células
tumorais Hela (Adenocarcinoma Epitelial de Cérvix Humano) e não tumorogênica
L929 (fibroblasto murino). O extrato clorofórmico das cascas e os compostos
isolados mostraram-se citotóxicos para a linhagem tumoral.
Além dos drimanos, neste trabalho, também foram isoladas do extrato
clorofórmico das cascas aromadendranos e lignanas. Estes têm apresentado
potencial citotóxico (ATAWODI; ALAFIATAYO, 2007; HELENO, et al., 2006). Para
estas classes, destacam-se ainda atividades, antifúngica, antibacteriana (LIMA;
BEATRIZ; RAMOS, 1997; MOREIRA, et al., 2003), antitripanocida, antiviral,
antialérgica e antiinflamatória (ATAWODI; ALAFIATAYO, 2007; HELENO, et al.,
2006; SIMÕES, et al., 2004).
A partir do exposto, pode-se sugerir que os drimanos contribuam com os
efeitos citotóxicos apresentados pelo extrato clorofórmico das cascas. Porém pode
haver um sinergismo, pois de maneira geral o extrato foi mais efetivo do que os
compostos isolados. As substâncias isoladas apresentam por vezes apenas um alvo
molecular e são na grande maioria mais eficazes quando estão combinados, como
por exemplo, no extrato bruto. Elas podem atuar em vários alvos simultaneamente, e
dessa forma aumentam o potencial terapêutico. Fármacos monoalvos segundo Keith
e Zimmermann (2004) não são capazes de combater de forma eficiente condições
patológicas como o câncer e doenças infecciosas. Estratégias multialvo podem ser
aplicadas no tratamento do câncer, por ele ser multigênico. O sinergismo com alvos
múltiplos pode potencializar o efeito terapêutico.
As substâncias testadas no presente estudo, isolados de D. brasiliensis, ainda
não foram plenamente investigados para a atividade biológica antitumoral em outras
linhagens de câncer, desta forma necessitando de maiores investigações a respeito
disso. Alguns testes estão sendo realizados para sanar esta lacuna, com outras
linhagens tumorais. Visto que os alvos celulares podem ser diferentes dependendo
das células avaliadas, alguns resultados que podem ser representativos para
algumas linhagens podem não ser tão significativos para outras e o inverso também
é verdadeiro. Dessa forma faz-se necessário a continuidade destes estudos com
diversas linhagens celulares para a verificação da especificidade e do grau de ação.
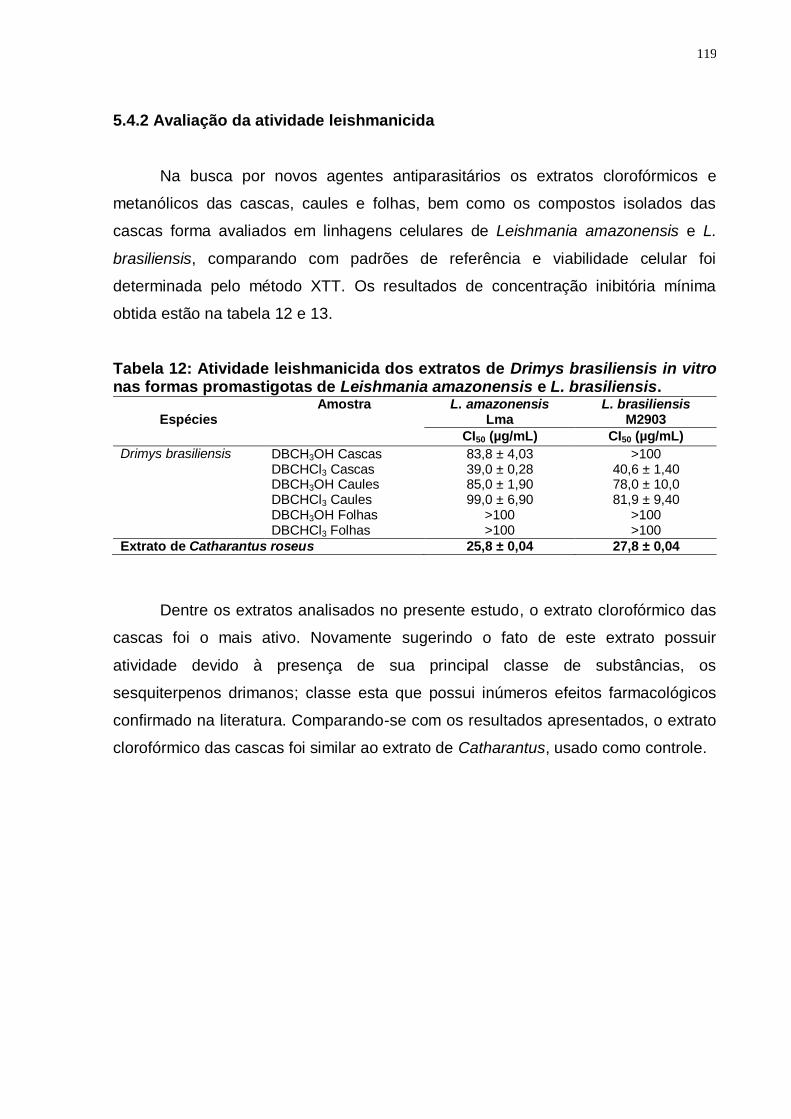
119
5.4.2 Avaliação da atividade leishmanicida
Na busca por novos agentes antiparasitários os extratos clorofórmicos e
metanólicos das cascas, caules e folhas, bem como os compostos isolados das
cascas forma avaliados em linhagens celulares de Leishmania amazonensis e L.
brasiliensis, comparando com padrões de referência e viabilidade celular foi
determinada pelo método XTT. Os resultados de concentração inibitória mínima
obtida estão na tabela 12 e 13.
Tabela 12: Atividade leishmanicida dos extratos de Drimys brasiliensis in vitro nas formas promastigotas de Leishmania amazonensis e L. brasiliensis.
Espécies
Amostra L. amazonensis Lma
L. brasiliensis M2903
CI50 (µg/mL) CI50 (µg/mL)
Drimys brasiliensis DBCH3OH Cascas 83,8 ± 4,03 >100 DBCHCl3 Cascas 39,0 ± 0,28 40,6 ± 1,40 DBCH3OH Caules 85,0 ± 1,90 78,0 ± 10,0 DBCHCl3 Caules 99,0 ± 6,90 81,9 ± 9,40 DBCH3OH Folhas >100 >100 DBCHCl3 Folhas >100 >100
Extrato de Catharantus roseus 25,8 ± 0,04 27,8 ± 0,04
Dentre os extratos analisados no presente estudo, o extrato clorofórmico das
cascas foi o mais ativo. Novamente sugerindo o fato de este extrato possuir
atividade devido à presença de sua principal classe de substâncias, os
sesquiterpenos drimanos; classe esta que possui inúmeros efeitos farmacológicos
confirmado na literatura. Comparando-se com os resultados apresentados, o extrato
clorofórmico das cascas foi similar ao extrato de Catharantus, usado como controle.

120
Tabela 13: Atividade leishmanicida das substâncias isoladas das cascas de Drimys brasiliensis in vitro nas formas promastigotas de Leishmania amazonensis e L. brasiliensis.
L. amazonensis Lma
L. brasiliensis M2903
CI50 (µM) CI50 (µM)
Substâncias 1-β-(p-metoxicinamil)-poligodial
O
O
MeO
CHO
CHO
15,85 ± 0,41 17,80 ± 0,68
1-β-(p-cumaroiloxi)-poligodial
O
O
OH
CHO
CHO
5,55 ± 0,98 2,52 ± 0,78
Drimanial
O
O
MeO
CHO
CHO
OH
20,42 ± 1,31 12,67 ± 1,15
Poligodial CHO
CHO
H
41,45 ± 1,79 25,64 ± 1,12
Espatulenol
H
HHH
OH
81,82 ± 0,63 89,10 ± 0,90
Anfotericina-B 0,21 ± 0,04 1,20 ± 0,04
Todos os drimanos apresentaram uma alta seletividade contra as formas
promastigotas, porém todos apresentaram CI50 bem superiores ao fármaco de
referência anfotericina B. Entre os compostos, o 1-β-(p-cumaroiloxi)-poligodial
apresentou a melhor atividade com CI50 = 5,55 e 2,52 µM, respectivamente, contra
as formas de Leishmania amazonensis e L. brasiliensis. Comparando a atividade de
ésteres cinamílicos 1-β-(p-metoxicinamil)-poligodial, 1-β-(p-cumaroiloxi)-poligodial e
o drimanial, nota-se que a mudança de um grupo metoxila em 1-β-(p-metoxicinamil)-
poligodial e drimanial por um grupo hidroxila no anel aromático em 1-β-(p-
cumaroiloxi)-poligodial, provocou um aumento da atividade leishmanicida contra as
duas formas de Leishmania analisadas. O poligodial é um drimano sem o
grupamento cinamílico e os resultados são semelhantes aos do drimanial. Esses

121
resultados indicam a importância do grupo p-hidroxicinamil à atividade leishmanicida
dos drimanos analisados.
5.4.3 Avaliação da atividade plasmodicida
Os extratos e os compostos 1-β-(p-metoxicinamil)-poligodial e o 1-β-(p-
cumaroiloxi)-poligodial também foram avaliados quanto à sua atividade antimalárica.
Para a atividade plasmodicida foram realizados por incubação de uma suspensão de
parasitas nas formas da cultura de trofozoítas de cepa de Plasmodium falciparum
F32-Tanzânia (cepa cloroquina-sensível). A triagem inicial dos extratos mostrou
atividade plasmodicida apenas para o extrato clorofórmico das cascas, com CI50 =
3,0 µg/mL e inibição de 97% (Tabela 14).
Os compostos revelaram promissora atividade plasmodicida, com 100% de
inibição e CI50 de 1,01 e 4,88 µM para o 1-β-(p-cumaroiloxi)-poligodial e 1-β-(p-
metoxicinamil)-poligodial, respectivamente. A diferença estrutural entre os dois
consiste na mudança do grupamento presente em C-7’. Enquanto em 1-β-(p-
cumaroiloxi)-poligodial o grupamento ligado é OH, no 1-β-(p-metoxicinamil)-
poligodial o grupamento metoxi está ligado. A presença do grupamento OH pode
estar influenciando na alteração das atividades físico-químicas da molécula como
hidrofilicidade, entre outras causando uma mudança na reatividade química.
Em Drimys angustifolia Miers predominam os drimanos poligodial, 1-β-(p-
metoxicinamil)-poligodial e o drimanial. Estes compostos apresentam potencial
antifúngico (CASTELLI, et al., 2005; MALHEIROS, et al., 2005), antinociceptivo (EL
SAYAH, et al., 1998; 2000), antiinflamatório (CECHINEL-FILHO, et al., 1998;
MALHEIROS, et al., 2001; CUNHA, et al., 2001) e antihiperalgésico (MENDES, et
al., 1998; MENDES, et al., 2000; MARTINI, et al., 2007). Estudos recentes indicaram
que o extrato bruto hexânico de Drimys brasiliensis e o poligodial mostraram
atividade contra Leishmania spp. na faixa entre 22 e 62 µg/mL (265 µM). O poligodial
também apresentou uma excelente atividade alta seletividade parasitária para
tripomastigotas de Trypanosoma cruzi (2 µg/mL; 8,5 µM) (CORREA, et al., 2011).

122
Tabela 14: Atividade plasmodicida dos extratos, substâncias isoladas das cascas de Drimys brasiliensis in vitro nas formas trofozoítas de Plasmodium falciparum.
Amostra % Inibição
CI50*
P. falciparum
FcR3
Drimys brasiliensis DBCHCl3 Cascas 97 ± 3,0 3,00 ± 0,4 ATIVO DBCH3OH Cascas 0 > 10 INATIVO DBCHCl3 Caules 0 > 10 INATIVO DBCH3OH Caules 0 > 10 INATIVO DBCHCl3 Folhas 40±6,0 > 10 INATIVO DBCH3OH Folhas 0 > 10 INATIVO
Substâncias
1-β-(p-metoxicinamil)-poligodial
O
O
MeO
CHO
CHO
100 4,88 ± 0,97 ATIVO
1-β-(p-cumaroiloxi)-poligodial
O
O
OH
CHO
CHO
100 1,01 ± 0,10 ATIVO
Cloroquina 100 0,1 ± 0,01 ATIVO * Os valores dos extratos estão em extratos estão em µg/mL. Já os valores das substâncias estão em µM.
Nossos resultados, juntamente com os da literatura, sugerem que os
drimanos podem ser agentes promissores contra as doenças causadas por
protozoários. No entanto, outros estudos (in vitro e in vivo) devem ser realizados a
fim de compreender os mecanismos de ação, bem como avaliar a toxicidade,
visando seu uso como fármaco.

123
6 CONCLUSÕES
Os resultados obtidos no presente trabalho mostram que o extrato
clorofórmico das cascas de D. brasiliensis é uma importante fonte de sesquiterpenos
drimanos e lignanas. Deste extrato foi possível isolar os drimanos poligodial, 1-β-(p-
metoxicinamil)-poligodial, drimanial e 1-β-(p-cumaroiloxi)-poligodial. Outro drimano
está em fase de identificação. Destes o 1-β-O-p-metoxi-E-cinamil-isodrimeninol e o
1-β-O-p-metoxi-E-cinamil-5α-hidroxi-isodrimeninol estão sendo relatados pela
primeira vez na literatura. Da classe das lignanas, foi isolada a (-) hinoquinina, que
está sendo relatada pela primeira vez para o gênero Drimys. Outra lignana está em
fase de identificação. Além destes, foi isolado um aromadendrano denominado
espatulenol e também foram isolados um ácido graxo e um glicerídeo. Todas as
substâncias foram identificadas por comparações de dados de ressonância
magnética nuclear com valores da literatura.
A ação citotóxica dos extratos clorofórmicos de cascas, caules e folhas e
substâncias isoladas foi verificada em modelos celulares de leucemia mielóide
(K562) e linfóide aguda B (Nalm6), adenocarcinoma mamário (MCF-7) e melanoma
murino (B16F10). O extrato clorofórmico das cascas foi o mais ativo para todos os
modelos celulares testados, e dentre os drimanos o 1-β-(p-metoxicinamil)-poligodial
que foi o mais ativo para todas as linhagens testadas.
Os extratos e as substâncias isoladas também foram avaliados em duas
formas promastigotas de Leishmania e uma forma trofozoíta de Plasmodium. O
extrato clorofórmico das cascas também foi o mais ativo. Os drimanos apresentaram
resultados bastante satisfatórios dentre eles, o 1-β-(p-cumaroiloxi)-poligodial foi o
mais ativo.
Os resultados do presente trabalho mostram a importância química e
medicinal da Drimys brasiliensis devido à presença dos sesquiterpenos drimanos
nas cascas que contribuem para sua atividade, confirmando seu uso popular no
tratamento de diversas patologias que acometem o homem.

124
REFERÊNCIAS
ALOUCHE, N.; APEL, C.; MARTIN, M. T.; DUMONTET, V.; GUÉRITTE, F.; LITAUDON, M. Cytotoxic sesquiterpenoids from Winteraceae of Caledonian rainforest. Phytochemistry, v. 70, p. 546-553, 2009. ANDRÉ, E.; MALHEIROS, A.; CECHINEL FILHO, V.; CALIXTO, J. B.; YUNES, R. A. Mechanisms underlying the relaxation caused by the sesquiterpene polygodial in vessels from rabbit and guinea-pig. European Journal of Pharmacology, v. 386, p. 47-53, 1999. ANDRÉ, E.; MALHEIROS, A.; CECHINEL FILHO, V.; CALIXTO, J. B.; YUNES, R. A. Role of nitric oxide and K+ channels in the relaxation induced by polygodial in the rabbit corpus cavernosum in vitro. Journal Cardiovascular Pharmacology, v. 41, p. 300-306, 2003.
ANDRÉ, E.; FERREIRA, J.; MALHEIROS, A.; YUNES, R. A.; CALIXTO, J. B. Evidence for the involvement of vanilloid receptor in the antinociception produced by the dialdeydes unsaturated sesquiterpenes polygodial and drimanial in rats. Neuropharmacology, n. 46, p. 590-597, 2004. ANDRÉ, E; CAMPI, B.; TREVISANI, M.; FERREIRA, J.; MALHEIROS, A.; YUNES, R. A.; CALIXTO, J. B.; GEPPETTI, P. Pharmacological characterization of the plant sesquiterpenes polygodial and drimanial as vanilloid receptor agonists. Biochemical Pharmacology, n. 71, p. 1248-1254, 2006. ANGER, M. Expressão de proteínas da apoptose em melanoma cutâneo primário.
2008. 97f. Tese de Doutorado. Universidade de São Paulo, São Paulo, 2008. AQUINO NETO, F. R. de. Cromatografia: princípios básicos e técnicas afins. Rio de
Janeiro: Interciência, 2003. ATAWODI, S. E.; ALAFIATAYO, A. A. Assessment of the phytochemical and antitrypanosomal properties of some extracts of leaves, stem and root bark of Landolphia sp., P. Beauv. Journal of Ethnopharmacology, v. 114, p. 207-211, 2007.
BARRERO, A. F., HERRADOR, M. M., ARTEAGA, P., LARA, A., CORTES, M. Chemical composition of the essential oil from Drimys winteri Forst. Wood. Journal of Essential Oil
Research, n. 12, p. 685-688, 2000. BARREIRO, E. J.; BOLZANI, V. da S. Biodiversidade: fonte potencial para a descoberta de fármacos. Química Nova, v. 32, n. 3, p. 679-688, 2009. BARROSO, M. G. Sistemática de angiospermas do Brasil. v. 2, Viçosa: UFV/Imprensa
Universitária, 1991. BASTOS, J. K.; KAPLAN, M. A. C.; GOTTLIEB, O. R. Drimane-type sesquiterpenoids as chemosystematics markers of Canellaceae. Journal of the Brazilian Chemical Society, v.
10, n. 2, p. 136-139, 1999. BOGLIOLO, L.; BRASILEIRO FILHO, G. Bogliolo: patologia geral. 3º ed. Rio de Janeiro:
Guanabara Koogan, 2004. BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Manual de diagnóstico laboratorial da malária. Brasília: Editora do Ministério da Saúde, 2005. 112 p. – (Série A.
Normas e Manuais Técnicos).

125
BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Manual de Vigilância da Leishmaniose Tegumentar Americana. 2. ed. Brasília: Editora do Ministério da Saúde,
2007. 180 p. - (Série A. Normas e Manuais Técnicos). BRESOLIN, T. M. B.; CECHINEL FILHO, V. Fármacos e medicamentos: uma abordagem
multidisciplinar. 1. ed. São Paulo: Santos, 2010. BROWN, G. D. Drimendiol, a sesquiterpene from Drimys winteri. Phytochemistry, v. 35, n.
4, p. 975-977, 1994. BUTLER, M. S. Natural products to drugs: Natural product – derived compounds in clinical trials. Natural Product Reports, n. 25, p. 475-516, 2008. CANHEDO, A. D. Indução da síntese de hemoglobina em células K562 por doxorrubicina e aclarrubicina: em busca de um mecanismo em comum. Tese de
doutorado. Universidade Federal do Rio Grande do Sul, 2005. CASTELLI, V. M.; LODEYRO, F. A.; MALHEIROS, A.; ZACCHINO, S. A. S.; ROVERI, A. O. Inhibition of the mitochondrial ATP synthesis by polygodial, a naturally occurring dialdehyde unsaturated sesquiterpene. Biochemical Pharmacology, v. 70, 2005, p. 82-89. CECHINEL FILHO, V.; YUNES, R. A. Estratégias para obtenção de compostos farmacologicamente ativos a partir de plantas medicinais. Conceitos sobre modificação estrutural para otimização da atividade. Química Nova, v. 21, n. 1, 1998, p. 99-105. CHAVES, M. H.; LAGO, J. H. G.; ROQUE, N. F. Macrocarpane from the leaves of Porcelia macrocarpa. Journal of the Brazilian Chemical, v. 14, n. 1, p. 16-19, 2003.
CICCIÓ, J. F. Aceites esenciales de las hojas y de los frutos verdes de Drimys granadensis
(Winteraceae). Revista de Biología Tropical, n. 44, p. 29-33, 1997. CORTÉS, M. & OYARZÚN, M. L. Tadeonal and Isotadeonal from Drimys winteri. Fitoterapia, v. 52, n.1, p. 33-35, 1981. CORRÊA, D. S.; TEMPONE, A. G.; REIMÃO, J. Q.; TANIWAKI, N. N.; ROMOFF, P.; FÁVERO, O. A.; SARTORELLI, P.; MECCHI, M. C., LAGO, J. H. Anti-leishmanial and anti-trypanosomal potential of polygodial isolated from stem barks of Drimys brasiliensis Miers
(Winteraceae). Parasitology Research, 2011. COSTA NETO, C. Análise Orgânica: métodos e procedimentos para caracterização de organoquímicos. Rio de Janeiro: UFRJ, p. 547-728, 2004. CUNHA, F. M.; FRÖDE, T. S.; MENDES, G. L.; MALHEIROS, A.; CECHINEL FILHO, V.; YUNES, R. A.; CALIXTO, J. B. Additional evidence for the anti-inflammatory and anti-allergic properties of the sesquiterpene polygodial. Life Sciences, n. 70, p. 159-169, 2001.
CRAGG, G.; NEWMAN, D. J. Natural Products as Source of New Drugs over the Last 25 Years. Journal of Natural Products, v. 70, p. 461-477, 2007.
CRAGG, G. M.; NEWMAN, D. J. Biodiversidade um componente essencial na descoberta de fármacos. In: YUNES, R. A., CECHINEL FILHO, V. Química de produtos naturais, novos fármacos e a moderna farmacognosia. 2. ed. Itajaí: UNIVALI, p. 53-81, 2009.

126
DEWICK, P. M. The mevalonate and deoxyxylulose phosphate pathways: Terpenoids and Esteroids. In: Química de produtos naturais, novos fármacos e a moderna farmacognosia, YUNES, R. A.; CECHINEL FILHO, v. 2. ed, UNIVALI, Itajaí, 2009. DE LAS HERAS, B.; RODRÍGUEZ, B.; BOSCÁ, L.; VILLAR, A. M. In: Química de produtos naturais, novos fármacos e a moderna farmacognosia, YUNES, R. A.; CECHINEL
FILHO, v. 2. ed, UNIVALI, Itajaí, 2009. DE VITA, V. T. Jr.; HELLMANN, S.; ROSEMBERG, S. A. Cancer: principles e practice of
oncology. 7 ed. Philadelphia: Lippincott Williams’s e Wilkins, 2005. DI STASI, L. C. Plantas medicinais: arte e ciência. Um guia de estudo interdisciplinar. São
Paulo: Universidade Estadual Paulista, 1996. DUBEY, V. S.; BHALLA, R.; LUTHRA, R. In: Química de produtos naturais, novos fármacos e a moderna farmacognosia, YUNES, R. A.; CECHINEL FILHO, v. 2. ed,
UNIVALI, Itajaí, 2009. El SAYAH, M.; MALHEIROS, A.; CECHINEL FILHO, V.; YUNES, R. A.; CALIXTO, J. B. Action of polygodial on agonist induced contraction of the rat portal vein in vitro. Journal of Cardiovascular Pharmacology, v. 35, n. 4, 2000, p. 670-675. FALKENBERG, M. B.; SANTOS, R. I.; SIMÕES, C. M. O. Introdução a análise fitoquímica. In: SIMÕES, C. M. O.; SCHENKEL, E. P.; GOSMANN, G.; MELLO, J. C. P.; MENTZ, L. A.; PETROVICK, P. R. Farmacognosia: da planta ao medicamento. 5. ed. Porto
Alegre/Florianópolis: UFRGS/UFSC, 2004. FAURI, J. A. C.; DIEHL, E. S.; CARTELL, A.; BAKOS, L.; EDELLWEISS, M. I. A. A proteína p16 e o melanoma cutâneo. Revista da Associação Médica do Rio Grande do Sul, Porto Alegre, v. 5, n. 1, p. 81-90, 2010. FORSBY, A.; NATHANSON, M.; HASANVAN, H.; WALUM, E. Polygodial inhibits carbachol-induced muscarinic acethylcholine receptor signal transduction. Toxicology in Vitro, n. 8, p. 585, 1998. FORSBY, A.; WALUM, E. Polygodial induces inositol phosphate turnover in human Neuroblastoma SH-SY5Y cells. Neuroscience Letters, n. 217, p. 50-54, 1996. FUJITA, K.; KUBO, I. Multifunctional action of antifungal polygodial against Saccharomyces cerevisiae: involvement of pyrrole formation on cell surface in antifungal action. Bioorganic
& Medicinal Chemistry, n. 13, p. 6742-6747, 2005. GEISSMAN, T. A.; CROUT, D. H. G. Organic Chemistry of Secondary Plant Metabolism.
Freeman, São Francisco, 1969. GERSHENZON, J.; DUDAREVA, N. In: Química de produtos naturais, novos fármacos e a moderna farmacognosia, YUNES, R. A.; CECHINEL FILHO, v. 2. ed, UNIVALI, Itajaí, 2009. GOLDMANN, L.; AUSIELLO, D. Cecil: Tratado de medicina Interna., 22 ed. Rio de Janeiro:
Editora Elsevier, 2005. GONZÁLES, M. I. Ácidos grasos omega 3: benefícios y fuentes. Interciencia, n. 27, p.128-136, 2002.

127
GOTTLIEB, O. R. Neolignans. In: Micromolecular evolution, systematic and ecology, an essay into a novel botanical discipline. Heidelberg: Springer, 1982.
GURIB-FAKIN, A. Review Medicinal plants: traditions of yesterday and drugs of tomorrow. Molecular Aspects of Medicine, n. 17, p. 1-93, 2006. GUYTON, A. C. M. D.; HALL, J. E. Tratado de Fisiologia Médica. 10. ed. Rio de Janeiro: Guanabara Koogan, 2001. HARBONE, J. B.; BAXTER, H. Phytochemical dictionary: a handbook of bioactive compounds from plants. Taylor & Francis, London, 1995. HAMMERSCHLAK, N. Manual de Hematologia: Programa Integrado de Hematologia e
Transplante de Medula Óssea. São Paulo: Manole, 2010. HELENO, V. C. G.; da SILVA, R.; PEDERSOLI, S.; ALBUQUERQUE, S. de; BASTOS, J. K.; SILVA, M. L. A. e; DONATE, P. M.; da SILVA, G. V. J.; LOPES, J. L. C. Detailed 1H and 13C NMR structural assignment of three biologically active lignan lactones. Spectrochimica Acta Part A, n. 63, p. 234-239, 2006. HORNSTRA, G. Omega-3 long-chain polyunsaturated fatty acids and health benefits. Neuilly-sur-seine: Nutriscience Roche Vitamins, 2002. INCA. Instituto Nacional de Câncer. Estimativa 2010 – Incidência de Câncer no Brasil. Disponível em: http://www1.inca.gov.br/estimativa/2010/index.asp?link=conteudo_view.asp&ID=1. Acessado em 07 Jan. 2010. INVITTOX; ERGATT; FRAME, MTT citotoxicity test, 1990.
JANSEN, B. J. M.; GROOT, A. the occurrence and biological activity of drimane sesquiterpenoids. Natural Product Reports, v. 8, n. 3, p. 309-318, 2004. JACOBSSON, U.; MUDDATHIR, A. K. Four biologically active sesquiterpenes of the drimane type isolated from Polygonum glabrum. Phytochemistry, v. 31, n. 12, p. 4207-4211, 1992.
KINGSTON, D. G. Modern natural products drug discovery and its relevance to biodiversity conservation. Journal of Natural Products, v. 74, p. 456-511, 2011. KEITH, C. T.; ZIMMERMANN, G. R. 2004. In: BRESOLIN, T. M. B.; CECHINEL FILHO, V. Fármacos e medicamentos: uma abordagem multidisciplinar. 1. ed. São Paulo: Santos, 2010. KYSER, O.; KIDERLEN, A. F.; CROFT, S. L. Natural products as antiparasitic drugs. Parasitology Reseach, v. 90, p. 55–62, 2003. LAGO, J. H. G.; ROQUE, N. F. Estudo fitoquímico da madeira de Guarea macrophylla (Meliaceae). Química Nova, v. 32, n. 9, p. 2351-2354, 2009. LENINGER, A.; NELSON, D. L.; COX, M. M. Princípios de Bioquímica. 3. ed. São Paulo:
Sarvier, 2002. LIMA, D. P. de.; BEATRIZ, A.; RAMOS, A. A. Transformações químicas do (+)-10-β, 14-diol-allo-aromadendrano, isolado de Duguetia glabriuscular. E. Fries (R. E. Fries) (Annonaceae)

128
e avaliações biológicas de alguns derivados obtidos. Química Nova, v. 20, n. 6, p. 616-620, 1997. LIMBERGER, R. P.; SIMÕES-PIRES, C. A.; SOBRAL, M.; MENUT, C.; BESSIERE, J-M.; HENRIQUES, A. T. Essential oils from Calyptranthes concinna, C. lucida and C. rubella
(Myrtaceae). Brazilian Journal of Pharmaceutical Sciences, v. 38, n. 3, p. 355-360, 2002. LIMBERGER, R. P.; SCOPEL, M.; SOBRAL, M.; HENRIQUES, A. T. Comparative analysis of volatiles from Drimys brasiliensis Miers and D. angustifolia Miers (Winteraceae) from
Southern Brazil. Biochemical Systems and Ecology, n. 35, p. 130-137, 2007. LORENZI, H. Árvores Brasileiras: Manual de identificação e Cultivo de Plantas Arbóreas
do Brasil. 3. ed. São Paulo: Nova Odessa, Instituto Plantarum de Estudos da Flora, 2002. MACHIDA, K.; TANAKA, T.; TANIGUCHI, M. Depletion of glutathione as a cause of the promotive effects of polygodial, a sesquiterpene on the production of reactive oxygen species in Saccharomyces cerevisiae. Journal of Bioscience and Bioengineering, v. 88, n. 5, p. 526-530, 1999. MAGRI, F. M. M. Determinação Estrutural e Simulação de Espectros de 13C de sesquiterpenos lactonizados utilizando métodos computacionais. Tese (Doutorado em
Ciências, Área Química). Instituto de Química da Universidade de São Paulo. Universidade de São Paulo, São Paulo, 1999. MAHMOUD, S. S.; CROTEAU, R. In: Química de produtos naturais, novos fármacos e a moderna farmacognosia, YUNES, R. A.; CECHINEL FILHO, v. 2. ed, UNIVALI, Itajaí,
2009. MALHEIROS, A.; CECHINEL FILHO, V.; SCHMITT, C. B.; YUNES, R. A.; ESCALANTE, A.; SVETAZ, L.; ZACCHINO, S.; DELLE MONACHE, F. Antifungal activity of drimane sesquiterpenes from Drimys brasiliensis using bioassay-guided fractionation. Núcleo de
Investigações Químico-Farmacêuticas, Universidade do Vale do Itajaí, Itajaí, Brasil. Journal of Pharmacy & Pharmaceutical Sciences, v. 8, n. 2, p. 335-339, 2005. MALHEIROS, A. Estudos químicos farmacológicos e alelopáticos das espécies Drimys angustifolia e Drimys brasiliensis (Winteraceae). Tese (Doutorado em Química). Centro
de Ciências Físicas e Matemáticas. Universidade Federal de Santa Catarina. Florianópolis, 2001. MALHEIROS, A.; CECHINEL FILHO, V.; SCHMITT, C. B.; SANTOS, A. R. S.; SCHEIDT, C; CALIXTO, J. B.; MONACHE, F. D.; YUNES, R. A. A sesquiterpene drimane with antinociceptive activity from Drimys winteri bark. Phytochemistry, v. 57, Maio, p. 103-107,
2001. MARTINI, L. H; JUNG, F.; SOARES, F.A.; ROTTA, L.N.; VENDITE, D. A.; FRIZZO, M. E. S.; YUNES, R. A.; CALIXTO, J. B; WOFCHUCK, S.; SOUZA, D. O. Naturally occurring compounds affect glutamatergic neurotransmission in rat brain. Neurochemical Research, n.11, p. 1950-1956, 2007. MASHIMBYE, M. J.; MAUMELA, M. C.; DREWES, S. E. A drimane sesquiterpenoid lactone from Walburgia salutaris. Phytochemistry, v. 51, p. 435-438, 1999. MATSUDA, H.; PONGPIRIYADACHA, Y.; MORIKAWA, T.; KASHIMA, Y.; NAKANO, K.; YOSHIKAWA, M. Protective Effects of Polygodial and Related Compounds on Ethanol-

129
Induced Gastric Mucosal Lesions in Rats: Structural Requirements and Mode of Action. Bioorganic & Medicinal Chemistry Letters, v. 12, p. 477–482, 2002. McMURRY, J. Química Orgânica. 6. ed. São Paulo: Pioneira Thomson Learning, 2005.
MEDOLA, J. F.; CINTRA, V. P.; SILVA, E. P. P. e; ROYO, V. de A.; da SILVA, R.; SARAIVA, J.; ALBUQUERQUE, S.; BASTOS, J. K.; SILVA, M. L. A. e; TAVARES, D. C. (-) Hinokinin causes antinogenotoxicity but not genotoxicity in peripheral blood of wistar rats. Food and Chemical Toxicology, n. 45, p. 638-642, 2007. MENDES, G. L.; SANTOS, A. R. S.; CAMPOS, M. M.; TRATSK, K. S.; YUNES, R. A.; CECHINEL FILHO, V.; CALIXTO, J. B. Anti-hiperalgesic properties of the extract and the main sesquiterpene polygodial isolated from the barks Drimys winteri. Life Sciences, v. 63,
n. 5, p. 369-381, 1998. MENDES, G. L.; MALHEIROS, A.; SANTOS, A. R. S.; CECHINEL FILHO, V.; YUNES, R. A.; CALIXTO, J. B. Assessment of Mechanisms Involved in Antinociception Caused by sesquiterpene Polygodial. The Journal of Pharmacology and Experimental Therapeutics, v. 292, n. 1, p. 164-172, 2000. MILLER, K.; NEILAN, B.; SZE, D. M. Recent patents anticancer. Drug Discovery, v.3, n.1, 2008. MOREIRA, I. C.; LAGO, J. H. G.; YOUNG, M. C. M.; ROQUE, N. F. Antifungal Aromadendrane Sesquiterpenoids from the Leaves of Xylopia brasiliensis. Journal of the
Brazilian Chemical Society, v. 14, n. 5, p. 828-831, 2003. MORTON, J. F. Winteraceae, Drymis family. In: Thomas CC Atlas of medical plants in middle America-Bahamas to Yucatan. Ilinóis: Springfield Thomas, p. 219, 1981.
MOSMANN, T. Rapid colorimetric assay for cellular growth and survival: application to ploriferation and cytotoxicity assay. Journal of Immunological Methods, v. 65, p. 55-63, 1983. MUNHOZ-CONCHA, D.; VOGEL, H.; YUNES, R.; RAZMILIC, I.; BRESCIANI, L.; MALHEIROS, A. Presence of polygodial and drimenol in Drimys populations from Chile.
Biochemical Systems and Ecology, n. 35, p. 434-438, 2007. NAKANO, T.; VILLAMIZAR, J. E.; MAILLO, M. A. Highly Efficient Síntesis of Optically Active Drimanic Sesquiterpenes, (+) Fuegim, (+) Epifutronolide (7-β-Hidroxyisodrimenin) and (+)-7-
Ketoisodrimenin. Tetrahedron, v. 55, p. 1561-1568, 1999. NIERO, R.; MALHEIROS, A. Principais aspectos químicos e biológicos de terpenos. In: CECHINEL, V. F. Química de produtos naturais, novos fármacos e a moderna farmacognosia. 2. ed. Itajaí: UNIVALI, p. 239-257, 2009.
NIERO, R.; MALHEIROS, A.; BITTENCOURT, C. M. S.; BIAVATTI, M. W.; LEITE, S. N.; FILHO, V. C. Aspectos químicos e biológicos de plantas medicinais e considerações sobre fitoterápicos. In: CECHINEL, V. F., BRESOLIN, T. M. B. Ciências Farmacêuticas:
contribuição ao desenvolvimento de novos fármacos e medicamentos. Itajaí: UNIVALI, p. 11-56, 2003. PAVIA, D. L.; LAMPMAN, G. M.; KRIZ, G. S.; VYVYAN, J. R. Introdução a espectroscopia.
São Paulo: Cengage Learning, 2010.

130
PENG, H.; ZHANG, G.; LIU, J., F.; LI, R. Dup-697, a specific COX-2 inhibitor, suppresses growth and induces apoptosis on K562 leukemia cells by cell-cycle arrest and caspase-8 activation. Annals of Hematology, v. 87, n. 2, p. 121-129, 2008. PEREZ-HERNANDEZ, N.; PONCE-MONTER, H.; MEDINA, J. A.; JOSEPH-NATHAN, P. Spasmolytic effect of constituents from Lepechinia caulescens on rat uterus. Journal of
Ethnopharmacology, v. 115, n. 1, p. 30-35, 2008. PETROVICK, P. R.; BASSANI, V. L.; GONZÁLEZ ORTEGA, G.; SONAGLIO, D. Desenvolvimento tecnológico e produção de fitoterápicos. In: SIMÕES, C. M. O.; SCHENKEL, E. P.; GOSMANN, G.; MELLO, J. C. P.; MENTZ, L. A.; PETROVICK, P. R. (org.) Farmacognosia: da planta ao medicamento. 6. ed., Porto Alegre: UFRGS,
Florianópolis: UFSC, cap. 13, p. 289-326, 2007. PIO CORRÊA, M. Dicionário das plantas úteis do Brasil e das tóxicas cultivadas. v. 6.
Rio de Janeiro: Imprensa Nacional, p. 1926-1978, 1931. RANG, H. P.; DALE, M. M.; RITTER, J. M.; FLOWER, R. J. Rang & Dale Farmacologia. 6.
ed. Rio de Janeiro: Elsevier, 2007. ROBBINS, S. L.; COTRAN, R. S.; KUMAR, V.; TARANTO, G. Patologia estrutural e funcional. 4. ed. Rio de Janeiro: Guanabara Koogan, p. 817-826, 2008.
SCHEIDT, C.; SANTOS, A. R.; FERREIRA, J.; MALHEIROS, A.; CECHINEL FILHO, V.; YUNES, R. A.; CALIXTO, J. B. Evidence for the involvement of glutamatergic receptors in the antinociception caused in mice by the sesquiterpene drimanial. Neuropharmacology, n. 43, p. 340-347, 2002. SCHMIDT, C. O.; BOUWMEESTER, H. J.; BÜLOW, N.; KÖNIG, W. A. Isolation, characterization, and mechanistic studies of (-)-alpha-gurjunene synthase from Solidago canadensis. Archives of Biochemistry and Biophysics, n. 364, p. 167-177, 1999.
SCHMEITS, B. L.; COOK, J. A.; VANDERJAGT, D. J.; MAGNUSSEN, M. A.; BHATT, S. K.; BOBIK, E. G.; HUANG, Y. S.; GLEW, R. H. Fatty acid composition of the milk lipids of women in Nepal. Nutrition Research, n. 19, p. 1339-1348, 1999. SIERRA, J. R., LÓPEZ, J. T. & CORTÉS, M. J. (-)-3-β-acetoxydrimenin from the leaves of Drimys winteri. Phytochemistry, v. 25, n. 1, p. 253-54, 1986.
SILVA, D. R. B. da; MIRANDA JÚNIOR; P. F.; SOARES, E. de A. A importância dos ácidos graxos poliinsaturados de cadeia longa na gestação e lactação. Revista Brasileira de Saúde Materna Infantil, v. 7, n.2, p.123-133, 2007. SILVA, K. C. da. Estudos Adicionais Sobre a Composição Química do Extrato Clorofórmico da Drimys brasiliensis e Avaliação Citotóxica de Células HELA (Tumorogênica) e L929 (Linhagem Não Tumorogênica). Monografia (Graduação em
Farmácia). Centro de Ciências da Saúde, Universidade do vale do Itajaí. Itajaí, 2009. SILVEIRA, A. C. de O. Análise da Atividade Antibacteriana e Toxicidade de Drimys brasiliensis. Dissertação (Mestrado em Ciências Farmacêuticas). Centro de Ciências da
Saúde, Universidade do Vale do Itajaí. Itajaí, 2011. SILVERSTEIN, R. M.; BASSLER, G. C.; MORRIL, T. C. Identificação Espectrométrica de Compostos Orgânicos. 5. ed. Rio de Janeiro: Guanabara Koogan, S. A., p. 387, 1994.

131
SIMÕES, C.M.O.; SCHENKEL, E. P.; GOSMANN, G.; MELLO, J. C. P.; MENTZ, L. A.; PETROVICK, P. R. Farmacognosia: da planta ao medicamento. 5. ed. Porto
Alegre/Florianópolis: UFRGS/UFSC, 2004. SIMÕES, C. M. O.; MENTZ, L. A.; SCHENKEL, E. P.; IRGANG, B. E.; STHEHMANN, J. R. Plantas Medicinais do Rio Grande do Sul. Porto Alegre: UFRGS, 1986.
TOMIO, T. A. Atividade Citotóxica e Antitumoral dos Extratos e Substâncias Isolados de Rapanea ferruginea e Piper aduncum. Dissertação (Mestrado em Ciências
Farmacêuticas). Universidade do Vale do Itajaí. Itajaí, 2011. TRAGER, W.; JENSEN, J.B. Human malaria in continuous culture. Science, v. 193, p. 673-675, 1976. TRINTA, E. F.; SANTOS, E. Winteráceas. Flora ilustrada Catarinense. Itajaí: Herbário
Barbosa Rodrigues, 1997. VALENZUELA, A. B.; NIETO, M. S. Acido docosahexaenoico (DHA) en el desarrolo fetal y en la nutrición materno-infantil. Revista Médica de Chile, n. 129, p. 1203-1211, 2001. VALENZUELA, A. B.; NIETO, S. K. Ácidos grasos omega-6 y omega-3 en la nutrición perinatal: su importância em el desarrolo del sistema nervioso y visual. Revista Chilena de Pediatría, n. 74, p.149-157, 2003.
VERLI, H.; BARREIRO, E. Um paradigma da química medicinal: a flexibilidade dos ligantes e receptores. Química Nova, v. 28, n. 1, p. 95-102, 2005. VERPOORTE, R. In: Química de produtos naturais, novos fármacos e a moderna farmacognosia, YUNES, R. A.; CECHINEL FILHO, v. 2. ed, UNIVALI, Itajaí, 2009.
VICKERY, M. L.; VICKERY, B. Secondary Plant Metabolism. The Macmillan Press Ltd,
Hong Kong, 1981. WAITAINCENIS, A.; ROLDÃO, E. F.; SEITO, L. N.; ROCHA, N. P.; DI STASI, L. C. Pharmacological and toxicological studies of Drimys angustifolia Miers (Winteraceae). Journal of Ethnopharmacology, n. 111, p. 541-546, 2007.
WAGNER, H. Pesquisa Fitomédica no Novo Milênio: Tendências e mudanças. In: YUNES, R. A., CECHINEL, V. F. Química de produtos naturais, novos farmacos e a moderna farmacognosia. 2. ed. Itajaí: UNIVALI, p. 53-81, 2009.
WILLIAMS, C. A.; HARVEY, W. J. Leaf. Leaf flavonoid patterns in the Winteraceae. Phytochemistry, v. 21, n. 2, p. 329-337, 1982. WHO. World Health Organization. World Cancer Report, 2008. YUNES, R. A.; CECHINEL FILHO, V. Química de produtos naturais, novos fármacos e a moderna farmacognosia. Itajaí, UNIVALI, 2007.
YUNES, R. A., CECHINEL FILHO, V. Química de produtos naturais, novos fármacos e a moderna farmacognosia. 2. ed. Itajaí: UNIVALI, 2009. ZAPATA, N; BUDIA, F.; VIÑUELA, E.; MEDINA, P. Antifeedant and growth inhibitory effects of extracts and drimanes of Drimys winteri stem bar against Spodoptera littoralis (Lep. Noctuidae). Industrial Crops and Products, n. 30, 2009, p. 119-125, 2009.





























