Embed Size (px)
Citation preview

University of Groningen
Activation, apoptosis and clearance of neutrophils in Wegener's granulomatosisRossum, Aart Pieter van
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2005
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Rossum, A. P. V. (2005). Activation, apoptosis and clearance of neutrophils in Wegener's granulomatosis.s.n.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
The publication may also be distributed here under the terms of Article 25fa of the Dutch Copyright Act, indicated by the “Taverne” license.More information can be found on the University of Groningen website: https://www.rug.nl/library/open-access/self-archiving-pure/taverne-amendment.
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 09-11-2021

PART II
Clearance of apoptotic neutrophils in the presence of PTX3, a hypothesized candidate factor in the induction of
leukocytoclasia


Chapter 8
The prototypic tissue pentraxin PTX3, in contrast to the
short pentraxin SAP, inhibits phagocytosis of late
apoptotic neutrophils by macrophages
André P. van Rossum 1, Fausto Fazzini 2, Pieter C. Limburg 4, Angelo A. Manfredi 2,
Patrizia Rovere-Querini 2, Alberto Mantovani 3, Cees G.M. Kallenberg 1
1Department of Clinical Immunology, Internal Medicine, University Medical Center
Groningen, The Netherlands, 2Clinical Immunology Unit and Cancer Immunotherapy
& Gene Therapy Program, H. San Raffaele Scientific Institute, 20132 Milano, Italy, 3Instituto di Ricerche Farmacologiche “Mario Negri” and Università, Milan, Italy, and
4Department of Pathology and Laboratory Medicine and Department of
Rheumatology, University Medical Center Groningen, The Netherlands
Arthritis & Rheumatism 2004; 50 (8): 2267 - 2274

Chapter 8
100
ABSTRACT Objective. Phagocytosis of apoptotic cells can be facilitated by complement components and short pentraxins, like serum amyloid P (SAP). In contrast, the long pentraxin PTX3 was shown to inhibit phagocytosis of apoptotic Jurkat cells by dendritic cells and bind late apoptotic polymorphonuclear leukocytes (PMN). Recently, levels of the pentraxin PTX3 proved to parallel disease activity in small vessel vasculitis. Since small vessel vasculitis is often characterized by leukocytoclasia, a persistence of leukocyte remnants in the vessel wall, we hypothesized that PTX3 inhibits phagocytosis of late apoptotic PMN by macrophages, thereby leading to their accumulation in the vessel wall. Methods. Macrophages were allowed to phagocytose late apoptotic or secondary necrotic PMN that were incubated with or without PTX3 for 30 min prior to phagocytosis. Phagocytosis was allowed to occur in the presence of 30% normal human serum (NHS) with or without SAP and with or without depletion of complement. To discriminate between an inhibitory effect of PTX3 on binding or internalization of apoptotic PMN into macrophages, internalization was blocked by Cytochalasin B. Results. SAP and complement were both necessary for effective in vitro phagocytosis. In contrast, PTX3 inhibited phagocytosis dose-dependently, from 10% inhibition at 10 µg/ml up to almost complete inhibition at 100 µg/ml. Furthermore, PTX3 partly affected binding of apoptotic PMN to macrophages. Conclusion. PTX3, in contrast to SAP and complement, inhibits phagocytosis of late apoptotic PMN by monocyte-derived macrophages dose-dependently. Hence, PTX3 can play a role in the development of leukocytoclasia by affecting the clearance of apoptotic PMN, thereby inducing their accumulation in the vessel wall.

PTX3 inhibits uptake neutrophils
101
INTRODUCTION Pentraxins can be divided into two structural classes: the classical short pentraxins, such as CRP and SAP, with a monomeric MW of ~25 kDa, and the recently described long pentraxins with a molecular weight between 40-50 kDa [reviewed in 1]. Recently, the long pentraxin PTX3 was identified as a novel acute-phase reactant in active vasculitis. Fazzini et al showed that PTX3 is produced at sites of inflammation and levels of PTX3 can be used as an independent laboratory indicator for disease activity in small vessel vasculitis 2. Serum levels of PTX3 only partially corresponded to levels of CRP. Furthermore, in the same study they showed that activated endothelial cells produce PTX3 at sites of active vasculitis, in contrast to hepatically produced CRP. Serum concentrations of CRP can increase up to 1000- fold within a few hours during the acute phase response 3. In mice, another short pentraxin, serum amyloid P (SAP), is responding as an acute phase reactant as well 3. The function of peripherally produced PTX3 is presently unclear. Recent studies with the pentraxins SAP and CRP have demonstrated an important facilitating role of these molecules in phagocytosis of apoptotic cells 4. PTX3 was shown to be involved in phagocytosis as well. Rovere et al described that PTX3 specifically binds to apoptotic Jurkat cells and subsequently inhibits their phagocytosis by dendritic cells 5. Additionally, they showed that late apoptotic polymorphonuclear leukocytes (PMN) were able to bind PTX3. In view of the persistence of late apoptotic, or secondary necrotic, PMN in active leukocytoclastic vasculitis 6 and the association between vasculitic disease activity and PTX3 serum levels 2, we hypothesized that PTX3, produced locally by activated endothelial cells, may play a role in the persistence of late apoptotic or secondary necrotic PMN in vasculitic lesions. The histopathologic substrate of small-vessel vasculitis is often designated as leukocytoclastic vasculitis 6. Leukocytoclasia, i.e. the accumulation of unscavenged dead neutrophils within the vessel wall, is normally an uncommon phenomenon. Leukocytoclastic lesions are particularly found in the skin, but other organs may be involved as well 7. In small vessel vasculitis, levels of pro-inflammatory cytokines, such as tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β), are elevated 8. TNF-α and IL-1β can activate endothelial cells resulting in adhesion and migration of neutrophils. Furthermore, it has been shown that pro-inflammatory signals induce the production of PTX3 in endothelial cells and PTX3 was shown to be expressed in endothelial cells and infiltrating leukocytes at sites of active vasculitis 2;9. In the present study, we investigated whether PTX3, in contrast to SAP and complement, inhibits the phagocytosis of late apoptotic PMN by macrophages, so explaining the phenomenon of leukocytoclasia in small-vessel vasculitis.

Chapter 8
102
MATERIALS AND METHODS Materials All chemicals used were from Sigma, Zwijndrecht, The Netherlands, unless otherwise indicated. Anti-coagulant tubes were from BD Vacutainer Systems, Plymouth, UK. Hanks Balanced Salt Solution (HBSS) and Gentamycin were purchased from Gibco / Life Technologies, Breda, The Netherlands. RPMI 1640 and FCS were from BioWhittaker Europe, Verviers, Belgium, and culture plates from Costar, Schiphol, The Netherlands. Human PTX3 was purified from Chinese hamster ovary (CHO) cells stably and constitutively expressing the protein, as described by Bottazzi et al 10. Isolation and culture of human neutrophils Neutrophils were isolated from blood of healthy human volunteers by Polymorphprep TM (Nycomed, Oslo, Norway) density gradient centrifugation. 5 ml EDTA-anticoagulated blood was carefully layered over 5 ml solution, centrifuged at 500 x g for 30 minutes, and the middle band was harvested. Red blood cells were lysed twice by mixing the cells with 6 ml NH4Cl solution, incubation for 10 min on ice and centrifugation (1200 x g for 3 min.). Subsequently, neutrophils were washed in HBSS without calcium and magnesium and centrifuged at 1200 x g for 3 min. Finally, 1x106/ml neutrophils were reconstituted in RPMI 1640 supplemented with Gentamycin (60 µg/ml) and 5% FCS in 6- wells plates and aged for 72 hours at 37oC in 5% CO2 to yield late apoptotic, or secondary necrotic, PMN. Apoptosis and necrosis was measured by Annexin V and propidium iodide staining. For staining, 99 µl binding buffer (10mM HEPES [pH 7.4], 140 mM NaCl, 5 mM CaCl2), 10 µl propidium iodide (10 µg/ml; Molecular Probes, Leiden, The Netherlands), and 1 µl fluorescein isothiocyanate (FITC)-labeled annexin V (Nexins Research, Kattendijke, The Netherlands) diluted 1:10 were added to 1x106 PMN. Immediately after incubation for 10 minutes on ice in the dark, immunofluorescence analysis was performed on an Epics-Elite FACS equipped with a gated amplifier (Coulter Electronics, Mijdrecht, The Netherlands). Macrophage culture Peripheral blood mononuclear cells were isolated by Lymphoprep TM (Nycomed, Oslo, Norway) density gradient centrifugation from citrated blood. Healthy controls served as donors. Cells were suspended in medium containing RPMI, gentamycin, and 2% pooled normal human serum (NHS) at a concentration of 1x106/ml. Plastic coverslips (13 mm diameter; Nunc, Roskilde, Denmark) were placed in a 16 mm-diameter 24-wells plate (Costar, Schiphol, The Netherlands). In every well, 0.5 ml cell suspension was seeded, and, subsequently, monocytes were allowed to differentiate into macrophages during 7 days at 37oC in a 5% CO2 atmosphere. At day 2 and 5 the medium was supplemented with 0.5 ml fresh medium. Phagocytosis assay and scoring Coverslips with adherent monocyte-derived macrophages were washed with RPMI containing 1% NHS to remove non-adherent cells and were transferred into 24-wells plates. Late apoptotic PMN were incubated for 30 min. with various concentrations of PTX3 (0, 3.125, 6.25, 12.5, 25, 50, and 100 µg/ml) and were resuspended in RPMI

PTX3 inhibits uptake neutrophils
103
containing 30% normal human serum (NHS). Next, the PMN cell suspensions (0.3 ml/ well (5x105cells)) were added to the 24 wells- plates, containing monocyte-derived macrophages (5x104 cells/well), and cell interaction was allowed for 30 min. at 37oC in 5% CO2. Cell interaction was also allowed to occur in SAP-depleted and complement inactivated serum. SAP-depleted serum was made by bringing NHS on an agarose column enriched with high electroendosmosis agarose (lot.no. AG 0493; FMC Bioproducts, Rockland, ME) as described earlier 4. Complement-inactivated serum was made by heating at 56oC for 30 minutes. Subsequently, coverslips were washed with 0.4% HSA to remove non-phagocytosed cells. Scoring of phagocytosis was done as described 4;11. Briefly, macrophages were flattened by centrifugation at 25 x g for 10 min and air-dried. Cells were fixed in ethanol and stained for MPO as a marker for ingested PMN. To this end, samples were incubated for 30 min with anti-MPO –mAbs (266.6K2, IQProducts, Groningen, The Netherlands), washed three times with PBS and subsequently incubated with horseradish peroxidase (HRP)-conjugated goat anti-mouse antibody (Dako, Glostrup Denmark). Dilutions of antibodies were made according to the manufacturer’s protocol. Cells were washed and allowed to react with di-aminobenzidine (DAB) and H2O2. Finally, nuclear staining of monocyte-derived macrophages was performed with haematoxylin (Merck, Darmstadt, Germany). Preparations were scored at 400 x magnification by regular light microscopy and the number of ingested PMN per individual macrophage was determined by counting 100 monocyte-derived macrophages for each coverslip. Only PMN clearly within the perimeter of the macrophage were counted. Binding assessment Cytochalasin B (Sigma, Zwijndrecht, The Netherlands) was used to block internalization but not binding of apoptotic cells, and was added to the 24 wells plates with coverslip-adherent monocyte-derived macrophages (5x104 cells/well) for 30 minutes prior to interaction with late apoptotic neutrophils. Monocyte-derived macrophages were allowed to interact with apoptotic PMN for 30 min at 37oC in 5% CO2 in the presence of cytochalasin B (24 µg/ml). Scoring was done as described above with the exception that binding was scored by counting the number of bound PMN per individual macrophage per total of 100 monocyte-derived macrophages. STATISTICS Results are expressed as mean ± SEM of the number of independent experiments. Statistical analysis was performed using the unpaired Student’s t-test and GraphPad Prism (version 3.0; GraphPad software, San Diego, CA).
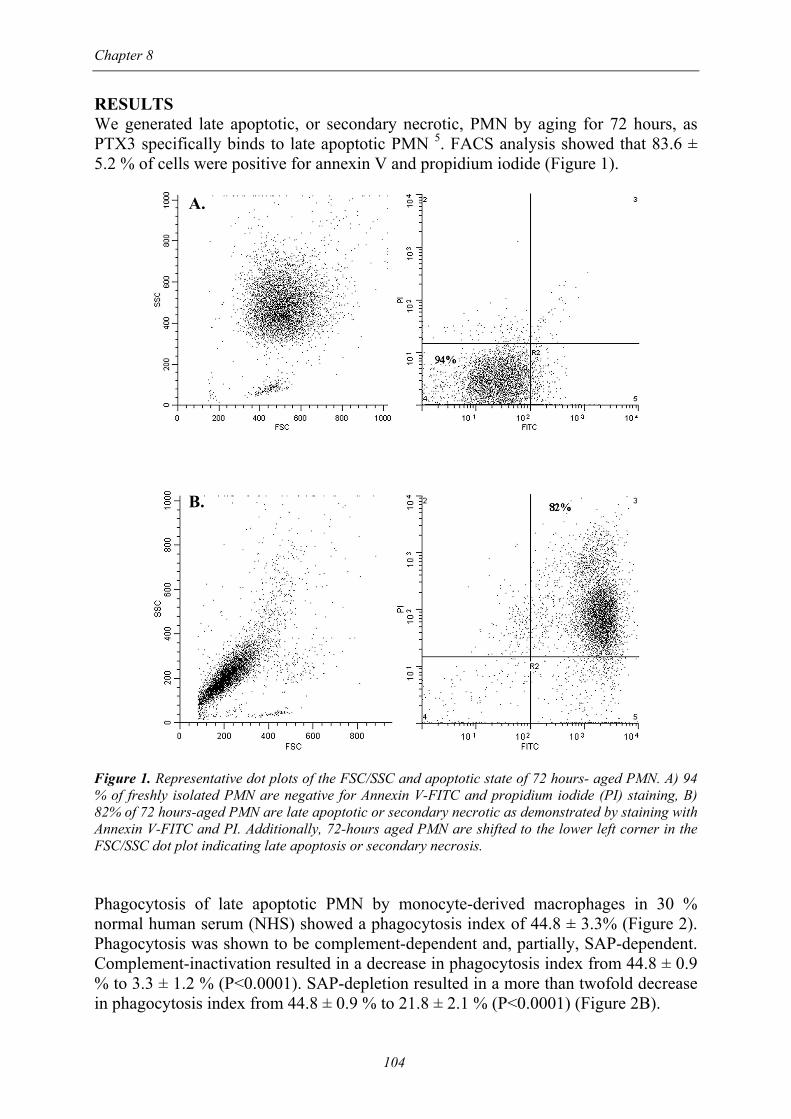
Chapter 8
104
RESULTS We generated late apoptotic, or secondary necrotic, PMN by aging for 72 hours, as PTX3 specifically binds to late apoptotic PMN 5. FACS analysis showed that 83.6 ± 5.2 % of cells were positive for annexin V and propidium iodide (Figure 1). A. B. Figure 1. Representative dot plots of the FSC/SSC and apoptotic state of 72 hours- aged PMN. A) 94 % of freshly isolated PMN are negative for Annexin V-FITC and propidium iodide (PI) staining, B) 82% of 72 hours-aged PMN are late apoptotic or secondary necrotic as demonstrated by staining with Annexin V-FITC and PI. Additionally, 72-hours aged PMN are shifted to the lower left corner in the FSC/SSC dot plot indicating late apoptosis or secondary necrosis. Phagocytosis of late apoptotic PMN by monocyte-derived macrophages in 30 % normal human serum (NHS) showed a phagocytosis index of 44.8 ± 3.3% (Figure 2). Phagocytosis was shown to be complement-dependent and, partially, SAP-dependent. Complement-inactivation resulted in a decrease in phagocytosis index from 44.8 ± 0.9 % to 3.3 ± 1.2 % (P<0.0001). SAP-depletion resulted in a more than twofold decrease in phagocytosis index from 44.8 ± 0.9 % to 21.8 ± 2.1 % (P<0.0001) (Figure 2B).

PTX3 inhibits uptake neutrophils
105
In order to evaluate the effects of PTX3 on phagocytosis, late apoptotic PMN were incubated for 30 minutes at room temperature with various concentrations of PTX3 in the presence of 30% NHS. Subsequently, phagocytosis was assayed of PTX3- incubated PMN. Incubation of late apoptotic PMN with 100 µg/ml PTX3 resulted in a significant decrease in phagocytosis index from 44.8 ± 0.9 % to 3.6 ± 0.8 % (Figure 2). The inhibitory effect of PTX3 on phagocytosis of late apoptotic PMN was dose-dependent (Figure 2C).
A. B. C. Figure 2. Inhibitory effect of PTX3 on the phagocytosis of late apoptotic PMN by monocyte-derived macrophages. A) Phagocytosis of late apoptotic PMN by monocyte-derived macrophages. PMN were stained by immunocytochemistry using peroxidase-conjugated antibodies to myeloperoxidase, a neutrophil- specific marker. Nuclear staining of monocyte-derived macrophages was performed by haematoxylin. B) Effect of SAP, PTX3 and complement on the phagocytosis of late apoptotic PMN by monocyte-derived macrophages. PTX3 (100 µg/ml) was added to PMN for 30 min prior to phagocytosis. Subsequently, PMN were allowed to interact with monocyte-derived macrophages for 30 minutes in 30% normal human serum. Phagocytosis was scored by counting the number of ingested PMN per individual macrophage per total of 100 macrophages (phagocytosis index). SAP depletion of normal human serum resulted in a decrease in phagocytosis index (P<0.0001). Complement inactivated- serum also resulted in decreased phagocytosis (P<0.0001). PTX3 (100 µg/ml) almost completely inhibited phagocytosis (P<0.0001). C) Dose-dependent inhibitory effect of PTX3 on the phagocytosis of late apoptotic PMN by monocyte-derived macrophages. Various concentrations of PTX3 were added to PMN 30 min prior to phagocytosis. Results are expressed as mean ± SEM of at least three independent experiments. Next, we investigated if the inhibitory effect of PTX3 was due to a defect in binding of late apoptotic PMN. Therefore, we incubated macrophages with cytochalasin B. Cytochalasin B- treated cells are incapable of ingesting particles but are still able to bind particles to membrane receptors 12. Cytochalasin B (24 µg/ml) almost completely disrupted internalization, whereas binding was still visible (Figure 3). When PMN

Chapter 8
106
incubated with PTX3 (50 µg/ml) were allowed to interact with cytochalasin B- treated monocyte-derived macrophages, binding was significantly decreased from 76.5 ± 1.5 % to 37 ± 2.0 % (P<0.005) (Figure 3C). A. B. C. Figure 3. PTX3 decreases binding of late apoptotic PMN. A) Immunohistochemical staining of binding of late apoptotic PMN to monocyte-derived macrophages. Cytochalasin B (24 µg/ml) was added for 30 min prior to interaction with PMN. PMN were allowed to bind for 30 min in 30% normal human serum to cytochalasin B-incubated macrophages. Arrows indicate bound PMN to monocyte-derived macrophages. PTX3 (50µg/ml) was added to late apoptotic PMN for 30 min prior to interaction. Cytochalasin B (24 µg/ml) was added to macrophages for 30 min prior to interaction. Cells were allowed to interact for 30 min at 37oC in 5% CO2. Phagocytosis (B) or binding (C) was scored by counting the number of ingested or bound PMN, respectively, per individual macrophage of a total of 100 macrophages. Results are expressed as mean ± SEM of at least three independent experiments. DISCUSSION Apoptotic cells are specifically recognized and rapidly engulfed by phagocytic cells such as macrophages and dendritic cells (DCs). The mechanisms of recognition and removal are complex and incompletely understood. Apoptosis results in a variety of surface changes, such as exposure of phosphatidylserine on the outer membrane of apoptotic cells. In addition, carbohydrates like fucose and N-acetylglucosamine are increasingly expressed during apoptosis 13;14. Subsequently, collectins and collectin-like molecules can bind to these newly expressed molecules 15-18. The collectin-like

PTX3 inhibits uptake neutrophils
107
component of complement C1q was shown to be involved in apoptotic cell recognition 17;18. C1q can opsonize apoptotic cells and interact with complement receptors like CR3 and CR4 19. C1q is thought to play an important role in phagocytosis. Macrophages from C1q deficient mice showed a reduced capacity to phagocytose apoptotic thymocytes 19;20. Binding of complement is a rather late event during apoptotic cell death and an immediate early feature of necrotic cells. Therefore, complement might serve as an opsonin for late apoptotic or secondary necrotic cells which have escaped normal clearing mechanisms 21. Binding to apoptotic cells has also been demonstrated for the pentraxins 5;15;22. The short pentraxins SAP and CRP can bind to apoptotic and necrotic cells, and, possibly by interaction with Fcγ receptors, facilitate phagocytosis 23. Recently, Bijl et al demonstrated that SAP binds to late apoptotic Jurkat cells and facilitates their phagocytosis by monocyte-derived macrophages 3;4. In the present study we demonstrate that SAP also facilitates phagocytosis of late apoptotic PMN by monocyte-derived macrophages, since SAP-depletion resulted in more than 50% decrease in phagocytosis. Phagocytosis of late apoptotic PMN was shown to be complement-dependent as well. The pentraxin PTX3 also appeared to be involved in phagocytosis 5,24. Rovere et al 5 demonstrated that PTX3 binds specifically to apoptotic cells and inhibits phagocytosis of apoptotic Jurkat cells by dendritic cells. Binding of PTX3 to apoptotic Jurkat cells was dose-dependent and saturable. Furthermore, they showed that only late apoptotic neutrophils bind PTX3. For that reason, we generated late apoptotic or secondary necrotic neutrophils by aging PMN for 72 hours. 72 hrs aged PMN stained positive with AnnexinV and propidium iodide. PTX3 proved to inhibit phagocytosis of late apoptotic PMN by monocyte-derived macrophages. This inhibition was dose-dependent. To investigate if PTX3 interfered with the opsonizing effects of SAP, SAP-depleted serum was used. PTX3 still inhibited phagocytosis of late apoptotic PMN by macrophages in the presence of SAP-depleted serum. To assess whether this was due to a disturbance in binding of apoptotic cells, we pre-incubated macrophages with cytochalasin B which interferes with membrane dynamics thereby hampering internalization 12. Cytochalasin B-treated macrophages did bind apoptotic PMN whereas internalization was almost completely blocked. When PTX3-coated PMN were added to cytochalasin B-treated macrophages, binding was significantly affected. Binding of PTX3- incubated PMN was reduced twofold compared to non-incubated PMN, which suggests that inhibition of phagocytosis by PTX3 is partly due to reduced binding whereas the residual inhibitory effect is, probably, due to effects on internalization. Rovere et al. 5 have suggested that PTX3 did not influence binding of apoptotic cells to immature dendritic cells but only influenced their internalization. Physico-chemical differences in the plasma membranes, the different expression of membrane receptors between macrophages and immature dendritic cells or the lack of exogenous serum cofactors in the latter study may all be relevant to explain these apparent discrepancies. Opsonization of apoptotic cells by classical pentraxins has been suggested to lead to direct or indirect recognition by phagocytic cells. Indirect recognition occurs by activating complement, thereby enabling complement receptor –dependent uptake via CR3 and CR4, whereas direct recognition takes place via Fcγ receptors (FcγR) 19. CRP binds to FcγRI and FcγRII, whereas SAP can additionally interact with FcγRIII 25-27.

Chapter 8
108
However, other studies have suggested that CRP does not bind to Fcγ-receptors 28. Van der Winkel et al 29 indicated that usage of IgG1 anti-CRP monoclonal antibodies to demonstrate binding of CRP to FcγR raises technical problems since these antibodies can bind through their Fc part to FcγRII. Usage of biotinylated anti-CRP or anti-CRP F(ab’)2 for this purpose did not show binding of CRP to FcγRII 29. It therefore remains a matter of debate whether CRP really binds to Fcγ- receptors. The receptor capable of binding PTX3 has as yet not been identified and the mechanisms whereby PTX3 inhibits uptake of apoptotic cells inhibits are therefore unclear. In vitro, PTX3 inhibits clearance of apoptotic cells substantially at a concentration of 50 µg/ml whereas PTX3 in serum of active untreated vasculitis patients reach levels of only 6.17 ± 4.77 ng/ml 2, so questioning the relevance of the in vitro findings. However, as PTX3 is locally produced in inflammatory tissue, serum levels of PTX3 do not reflect what is locally present at the site of inflammation. The biological meaning of the inhibitory capacity of PTX3 is still speculative. It has been proposed that pentraxin- mediated clearance of apoptotic cells, as complement, represents a back-up mechanism for the clearance of late apoptotic cells in situations where apoptotic cell load is high and clearance capacity low 30. Rovere et al 5 suggested that PTX3 acts as a local regulator inhibiting local inflammatory uptake of late apoptotic cells by immature dendritic cells, thereby preventing antigen presentation by APCs, which can be relevant for preventing induction of autoimmunity. The inhibitory effect of PTX3 on phagocytosis of apoptotic PMN may be important in view of leukocytoclasia. Small vessel vasculitides are histologically characterized by leukocytoclasia, i.e. the accumulation of unscavenged apoptotic or necrotic PMN in or around the vessel wall. The defective clearance of cell debris can in turn be involved in the maintenance of peripheral inflammation. PTX3, a long extra-hepatically produced pentraxin, is released by a variety of cells in vitro such as fibroblasts, endothelial cells and cells of the monocytic lineage 31 As PTX3 can be produced by endothelial cells in active skin lesions of patients 2 with vasculitis, we hypothesize that inhibition by peripherally produced PTX3 of the phagocytosis of apoptotic neutrophils is responsible for the phenomenon of leukocytoclasia in small-vessel vasculitis. It, therefore, seems relevant to demonstrate PTX3 in lesional tissue from patients with leukocytoclastic vasculitis. Such studies are underway in our laboratory. REFERENCES 1 Goodman AR, Cardozo T, Abagyan R, Altmeyer A, Wisniewski HG, Vilcek J. Long pentraxins:
an emerging group of proteins with diverse functions. Cytokine Growth Factor Rev. 1996; 7: 191-202.
2 Fazzini F, Peri G, Doni A et al. PTX3 in small-vessel vasculitides: an independent indicator of disease activity produced at sites of inflammation. Arthritis Rheum. 2001; 44: 2841-50.
3 Pepys MB, Baltz ML. Acute phase proteins with special reference to C-reactive protein and related proteins (pentaxins) and serum amyloid A protein. Adv.Immunol. 1983; 34: 141-212.
4 Bijl M, Horst G, Bijzet J, Bootsma H, Limburg PC, Kallenberg CG. Serum amyloid P component binds to late apoptotic cells and mediates their uptake by monocyte-derived macrophages. Arthritis Rheum. 2003; 48: 248-54.

PTX3 inhibits uptake neutrophils
109
5 Rovere P, Peri G, Fazzini F et al. The long pentraxin PTX3 binds to apoptotic cells and regulates their clearance by antigen-presenting dendritic cells. Blood 2000; 96: 4300-6.
6 Koutkia P, Mylonakis E, Rounds S, Erickson A. Leucocytoclastic vasculitis: an update for the clinician. Scand.J.Rheumatol. 2001; 30: 315-22.
7 Jessop SJ. Cutaneous leucocytoclastic vasculitis: a clinical and aetiological study. Br.J.Rheumatol. 1995; 34: 942-5.
8 Grau GE, Roux-Lombard P, Gysler C et al. Serum cytokine changes in systemic vasculitis. Immunology 1989; 68: 196-8.
9 Breviario F, d'Aniello EM, Golay J et al. Interleukin-1-inducible genes in endothelial cells. Cloning of a new gene related to C-reactive protein and serum amyloid P component. J.Biol.Chem. 1992; 267: 22190-7.
10 Bottazzi B, Vouret-Craviari V, Bastone A et al. Multimer formation and ligand recognition by the long pentraxin PTX3. Similarities and differences with the short pentraxins C-reactive protein and serum amyloid P component. J.Biol.Chem. 1997; 272: 32817-23.
11 Licht R, Jacobs CW, Tax WJ, Berden JH. An assay for the quantitative measurement of in vitro phagocytosis of early apoptotic thymocytes by murine resident peritoneal macrophages. J.Immunol.Methods 1999; 223: 237-48.
12 Roos D, Goldstein IM, Kaplan HB, Weissmann G. Dissociation of phagocytosis, metabolic stimulation and lysosomal enzyme release in human leukocytes. Agents Actions 1976; 6: 256-9.
13 Duvall E, Wyllie AH, Morris RG. Macrophage recognition of cells undergoing programmed cell death (apoptosis). Immunology 1985; 56: 351-8.
14 Russell L, Waring P, Beaver JP. Increased cell surface exposure of fucose residues is a late event in apoptosis. Biochem.Biophys.Res.Commun. 1998; 250: 449-53.
15 Gershov D, Kim S, Brot N, Elkon KB. C-Reactive protein binds to apoptotic cells, protects the cells from assembly of the terminal complement components, and sustains an antiinflammatory innate immune response: implications for systemic autoimmunity. J.Exp.Med. 2000; 192: 1353-64.
16 Holmskov U, Malhotra R, Sim RB, Jensenius JC. Collectins: collagenous C-type lectins of the innate immune defense system. Immunol.Today 1994; 15: 67-74.
17 Korb LC, Ahearn JM. C1q binds directly and specifically to surface blebs of apoptotic human keratinocytes: complement deficiency and systemic lupus erythematosus revisited. J.Immunol. 1997; 158: 4525-8.
18 Nauta AJ, Trouw LA, Daha MR et al. Direct binding of C1q to apoptotic cells and cell blebs induces complement activation. Eur.J.Immunol. 2002; 32: 1726-36.
19 Mevorach D, Mascarenhas JO, Gershov D, Elkon KB. Complement-dependent clearance of apoptotic cells by human macrophages. J.Exp.Med. 1998; 188: 2313-20.
20 Taylor PR, Carugati A, Fadok VA et al. A hierarchical role for classical pathway complement proteins in the clearance of apoptotic cells in vivo. J.Exp.Med. 2000; 192: 359-66.
21 Gaipl US, Kuenkele S, Voll RE et al. Complement binding is an early feature of necrotic and a rather late event during apoptotic cell death. Cell Death.Differ. 2001; 8: 327-34.
22 Familian A, Zwart B, Huisman HG et al. Chromatin-independent binding of serum amyloid P component to apoptotic cells. J.Immunol. 2001; 167: 647-54.
23 Mold C, Baca R, Du Clos TW. Serum Amyloid P Component and C-Reactive Protein Opsonize Apoptotic Cells for Phagocytosis through Fcgamma Receptors. J.Autoimmun. 2002; 19: 147-54.
24 Garlanda C, Hirsch E, Bozza S et al. Non-redundant role of the long pentraxin PTX3 in anti-fungal innate immune response. Nature 2002; 420: 182-6.
25 Bharadwaj D, Mold C, Markham E, Du Clos TW. Serum amyloid P component binds to Fc gamma receptors and opsonizes particles for phagocytosis. J.Immunol. 2001; 166: 6735-41.
26 Bharadwaj D, Stein MP, Volzer M, Mold C, Du Clos TW. The major receptor for C-reactive protein on leukocytes is fcgamma receptor II. J.Exp.Med. 1999; 190: 585-90.
27 Stein MP, Edberg JC, Kimberly RP et al. C-reactive protein binding to FcgammaRIIa on human monocytes and neutrophils is allele-specific. J.Clin.Invest 2000; 105: 369-76.
28 Hundt M, Zielinska-Skowronek M, Schmidt RE. Lack of specific receptors for C-reactive protein on white blood cells. Eur.J.Immunol. 2001; 31: 3475-83.

Chapter 8
110
29 Saeland E, van Royen A, Hendriksen K et al. Human C-reactive protein does not bind to FcgammaRIIa on phagocytic cells. J.Clin.Invest 2001; 107: 641-3.
30 Nauta AJ, Daha MR, Kooten C, Roos A. Recognition and clearance of apoptotic cells: a role for complement and pentraxins. Trends Immunol. 2003; 24: 148-54.
31 Alles VV, Bottazzi B, Peri G, Golay J, Introna M, Mantovani A. Inducible expression of PTX3, a new member of the pentraxin family, in human mononuclear phagocytes. Blood 1994; 84: 3483-93.





























