Embed Size (px)
Citation preview

University of Groningen
Pathogenesis of nosocomial infections with Enterococcus faecalisWaar, Karola
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2004
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Waar, K. (2004). Pathogenesis of nosocomial infections with Enterococcus faecalis. s.n.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 14-05-2020

Pathogenesis of nosocomial infections withEnterococcus faecalis
Karola Waar


RIJKSUNIVERSITEIT GRONINGEN
Pathogenesis of nosocomial infectionswith Enterococcus faecalis
Proefschrift
ter verkrijging van het doctoraat in deMedische Wetenschappen
aan de Rijksuniversiteit Groningen op gezag van de
Rector Magnificus, Dr. F. Zwartsin het openbaar te verdedigen op
woensdag 17 november 2004om 14.45 uur
door
Karola Waar
geboren op 6 april 1971te Rotterdam

Promotores: Prof. Dr. J.E. DegenerProf. Dr. Ir. H.J. BusscherProf. Dr. H.C. van der Mei
Copromotor: Dr. H.J.M. Harmsen
Beoordelingscommissie: Prof. Dr. T.H. TheProf. Dr. M.J.H. SlooffProf. Dr. M.J.M. Bonten

Pathogenesis of nosocomial infections with Enterococcus faecalisKarola WaarPhD Thesis, University of Groningen – with references – with summary in Dutch
ISBN 90-367-2141-5
© K. Waar, Groningen, 2004
Printed by Ponsen & Looijen BV Wageningen
Cover design: DOKA design and Ponsen & Looijen BV
The cover shows the typical configuration ‘kameelzakligging’ of enterococci. The entero-coccus on the left is filled with a picture of a Gram stain of enterococci in the bile of a livertransplant patient, the enterococcus on the right is filled with a picture of a mixed culture ofE. faecalis and E. faecium hybridized with fluorescent DNA probes.
The studies described in this thesis were financially supported by a grant from the DutchOrganization for Scientific Research NWO (grant SGO-INF-007)
The publication of this thesis was financially supported by:AstraZeneca BVBayer BV Health CarebioMérieux Benelux BVGroningen University Institute for Drug Exploration GUIDE Faculteit Medische Wetenschappen RUGPfizer BVWyeth Pharmaceuticals BV

Paranimfen:Harriët HafkampMarion Waar

All the world’s a stage;And all the men and women merely players:
They have their exits and their entrancesAnd one man in his time plays many parts….
Shakespeare; As you like it.


Contents
Chapter 1 Pathogenesis of Enterococcus faecalis infections. 11Review of the literature.Scope of the thesis.
Part I Virulence of Enterococcus faecalis in the host 31
Chapter 2 Genogrouping and incidence of virulence factors of Enterococcusfaecalis in liver transplant patients differ from blood culture and fecalisolates.Journal of Infectious Diseases 2002; 185: 1121-1127 33
Chapter 3 Molecular epidemiology of Enterococcus faecalis in liver transplantpatients at the University Hospital Groningen.Journal of Hospital Infection 2003; 55:53-60 47
Chapter 4 Fluorescent in situ hybridization with specific DNA probes offersadequate detection of Enterococcus faecalis and Enterococcusfaecium in clinical samples.Submitted 59
Part II Enterococcus faecalis in biomaterial related infections 69
Chapter 5 Enterococcus faecalis surface proteins determine its adhesionmechanism to bile drain materials.Microbiology 2002; 148: 1863-1870 71
Chapter 6 Atomic force microscopy study on interaction forces betweenEnterococcus faecalis with and without aggregation substance.Submitted 83
Chapter 7 Adhesion to bile drain materials and physicochemicalb surfaceproperties of Enterococcus faecalis strains grown in the presence ofbile.Applied and Environmental Microbiology 2002; 68:3855-3858 93
9
CONTENTS

Chapter 8 Summarizing discussion 103
Chapter 9 Dutch summaryNederlandse samenvatting 113
References 121List of abbreviations 141Dankwoord 145Curriculum Vitae 149
10

CHAPTER
Pathogenesis of Enterococcus faecalis Infections
Review of the Literature 1


Introduction
Enterococci are Gram positive coccoid bacteria that belong to normal microbiota of thegastrointestinal tract of humans, most mammals, birds, and many other species. In the colon of nearly all humans enterococci can be found in numbers as high as 108
colony-forming units per gram of feces (164, 182). From the more than 13 enteroccocalspecies that are described, Enterococcus faecalis and Enterococcus faecium are isolatedmost frequently. In the normal healthy host enterococci seldom are causing infections, onlysome urinary tract infections are seen. However, surveillance data indicate that enterococci are becoming one of the leading causes of nosocomial infections (142, 159, 204).Nosocomial infections are infections that patients acquire in a health-care institution. Theseinfections can be caused by transmission of the bacterium from patient to patient or fromthe health care worker to the patient. Nosocomial infections with enterococci are frequentlyseen in critically ill patients at intensive care units, for example in liver transplant patients,which are often considered especially vulnerable to enterococcal infections (248, 250).Theproblem of nosocomial enterococcal infection is compounded by emerging antibioticresistance (113). Different studies describe a longer length of stay in hospital and increasedmortality due to vancomycin-resistant E. faecium compared to vancomycin-susceptible E.faecium (70, 140). However, resistance alone does not explain the increase of enterococciin nosocomial infections. Although resistance is relatively uncommon among E. faecalisisolates compared to resistance among E. faecium isolates (113), E. faecalis currentlyaccounts for the majority of clinical enterococcal isolates (up to 90 %), followed by E.faecium (142, 206). This disparity might be explained by the relative abundance of E.faecalis in the gastrointestinal tract (164, 182) or enhanced virulence of E. faecalis. Thisreport will focus on E. faecalis infections as these are more prominent among hospitalacquired infections. The pathogenesis of enterococcal infections is quite complex because under most circum-stances, enterococci stably coexist with the host and are often even considered beneficial to the host. However, situations can arise where this balanced commensalism is disrupted.This disruption may result from a breakdown in host mechanisms that hold commensalorganisms in control. For example, the natural barrier of the skin or mucosa might be disrupted in hospitalized patients that underwent large operations in the abdominalcavity or received indwelling biomaterial implants like drains or catheters. An increasingnumber of patients also receives immune suppressive therapy e.g. after solid organtransplantation, suppression of the immune response can also lead to infection withcommensal microorganisms. Disruption may further result from the organism acquiring new traits that enable the organism to circumvent the host defense or to adhere tobiomaterial implants and cause biomaterial associated infections. The goal of this introduction is to highlight different aspects of the pathogenesis and clinicalmanifestation of (nosocomial) E. faecalis infection, both from the side of the host as well theside of the microorganism.
13
CHAPTER
1

Virulence of E. faecalis
Enterococcal virulence factors can contribute to enterococcal disease in different ways; byenhancing colonization, adherence and invasion of host tissues, by modulation of the hostimmunity, and by inducing pathological changes in the host associated with increasedseverity of infection (123, 125, 167). Generally, the pathogenesis of enterococcal infectionsfollows a common sequence of events and different virulence factors of E. faecalis can playa role in each of these steps (95, 167). First, enterococci possessing various virulent traitsasymptomatically colonize the gastro-intestinal tract (233, 229). Subsequently, thispopulation is expanded and in some patients the enterococci can invade the host tissue andcause disease. Factors that enhance the virulence of enterococci through this way are foundmore frequently among isolates from hospitalized patients compared to isolates from thecommunity. Finally, enterococci can cause symptomatic disease at the level of tissuedestruction or toxicity (39, 117, 213, 223, 226). The virulence factors enhance patho-genicity through this way would not necessarily appear in increased numbers amongvarious clinical isolates but would be associated with more severe clinical presentation.Therefore, the contribution of certain enterococcal traits to virulence can be proven by ahigher incidence in nosocomial isolates and/or increased severity of disease in patients oranimals.The contribution of virulence factors to the pathogenesis of nosocomial infection issomewhat blurred due to the fact that patients with high immune suppression are alsosusceptible to commensal strains without virulence factors (9, 50, 93). Therefore, collectionsof infection-derived hospital isolates possibly contain a spectrum of types of strains, frompure commensals to strains harboring a combination of virulence factors. Strains that causehospital ward outbreaks among different groups of patients might represent the truevirulent enterococcal lineages; therefore the ability to cause outbreaks should be included inthe analysis of a possible virulence trait. Below the different stages of pathogenesis and therole of different virulence factors herein are discussed.
Colonization, adherence and invasion of host tissues
Clinical studies have shown that nosocomial enterococcal infections often arise throughtransfer of the enterococci between patients (7, 37, 114, 131, 172, 240, 247). Patientsoften get colonized with nosocomial enterococci previous to the development of anenterococcal infection (95, 275). One of the questions of nosocomial enterococcal infectionis why these nosocomial enterococci so easily colonize niches that are already occupied bymembers of the same species. These nosocomial enterococci might have extra capabilitiesto colonize, overgrow and invade host tissues. These capabilities might be supported by theuse of antibiotics without significant anti-enterococcal activity, thus outcompeting theindigenous flora and creating niches which can be readily colonized by the nosocomialenterococci (20, 273). This is supported by the fact that many nosocomial enterococci aremore resistant to antibiotics compared to commensal enterococci (113, 214). Amplifiedfragment length polymorphism (AFLP) analysis of E. faecium isolates from different clinicalsubgroups and particular hosts indicated that these isolates could be divided into
PATH
OG
ENES
IS O
F E.
FA
ECA
LIS
INFE
CTI
ON
S
14

genogroups with an AFLP similarity of > 65%. Interestingly, a clear dichotomy betweenisolates from hospitalized and non hospitalized persons was detected (261). This associationof a specific genogroup with a particular host or environment might hold the key to thequestion why nosocomial enterococci colonize the hospitalized patient and its environmentmore easily. Studies in mice showed that following colonization of the GI tract, bacteria may be able totranslocate over the epithelial lining of the lumen (225). The proposed mechanism for thetranslocation of intestinal bacteria is that epithelial cells and phagocytes uptake thegastrointestinal bacteria and the bacteria exit at the apical side or migrate in phagocytes tothe mesenteric lymph nodes, proliferate and spread to distant sites (14, 258). Evidence oftranslocation of E. faecalis across the intact intestinal epithelium was given by Wells et al.who cultured E. faecalis from liver, spleen and lymph nodes of mice with intestinalovergrowth of E. faecalis (255, 257). Furthermore, they showed that these translocatedenterococci caused systemic infections (225). In another study, Wells et al. suggested thatthe site of translocation in mice was the caecum or colon (254).In an in vitro study Guzman et al. provided evidence for the role of adherence in thepathogenesis of E. faecalis urinary tract infection and endocarditis (101, 102). They showedthat isolates from urinary tract infections or endocarditis adhered best to respectively urinarytract epithelial cells and heart cells and that these adhesive properties were induced bygrowth in serum (102).
Modulation of the host immunity
E. faecalis must overcome the clearance functions of the host system to successfully causeinfection. Polymorphonuclear leukocytes (PMNs) are a critical component of the humanhost response against bacterial infections. Invading bacteria may be coated by complementproteins or specific antibodies and subsequently phagocytosed and killed by PMNs. Thisprocess of coating of bacteria with complement proteins or antibodies to enhancephagocytosis is called opsonization. Studies involving the role of antibodies andcomplement in the phagocytic killing of enterococci revealed that PMN mediated killingdepended primarily on complement activation by either the classical or the alternativepathway (5, 106). Antibodies to E. faecalis enhanced the PMN mediated killing, howeverthey were not essential as different studies showed efficient killing also in the presence ofserum without gamma globulins (84, 106).Although antibodies to enterococci are found in humans with enterococcal infections (229),studies on the efficacy of antibodies to E. faecalis in the prevention of infections are quitecontradicting. Huebner et al. (110) found prophylactic and therapeutic efficacy of anti-bodies to a capsular polysaccharide in a mouse infective model. In addition, the role ofantibodies to the surface protein aggregation substance (Agg) in prevention of endocarditisis underscored by the absence of host antibodies specific for the Agg during the formationof endocardial vegetation. Thereby the bacteria are protected from the influence of theantibodies (155). However, another study to the efficacy of antibodies to Agg in theprevention of endocarditis in a rabbit model did not show any protection (154).E. faecalis has developed different strategies to overcome the immune response. Gentry-
15
CHAPTER
1

Weeks et al. (92) reported a prolonged intracellular survival of enterococci for up to 72 h inmouse peritoneal macrophages. This property might contribute to the pathogenesis ofinfections in the way that the enterococci migrate to distant sites in the body and beprotected from antimicrobial therapy within the macrophage. In line with these findings arethe results of other investigations reporting that Agg promotes direct, opsonin-independentbinding of E. faecalis to PMN and that through this opsonin-independent binding E. faecaliswas able to survive inside different phagocytes (201, 231, 246). Another study showed thatstrains expressing gelatinase, cytolysin, or Agg were not more resistant to neutrophil-mediated killing, but the in vitro assays were performed under circumstances that might notsupport expression of these traits or mimic the in vivo situation (5, 184).The structure of theenterococcal surface protein (Esp) with multiple repeat motifs in the encoding gene mightbe important in the immune evasion of infecting E. faecalis (217).
Pathological changes in the host
The last step in the pathogenesis of infections is the production of pathologic changes in thehost. Such changes can be induced by the host inflammatory response or by direct tissuedamage as a result of secreted toxins or proteases. Enterococcal lipoteichoic acid is mostfrequently described as one of the factors that modulates the host immune response andthereby causes tissue damage. Several groups found lipoteichoic acid to be as inflammatory aslipopolysaccharide of Gram-negative bacteria and a potent inducer of different cytokines (17,238). A study to the role of Agg and enterococcal binding substance to the pathogenesis ofendocarditis found that strains without Agg or enterococcal binding substance lacked theability to cause disease, strains with either Agg or enterococcal binding substance wereintermediate virulent and strains with both Agg and enterococcal binding substance on theirsurface exhibited the greatest ability to cause disease. Furthermore, none of the rabbitsreceiving Agg and enterococcal binding substance positive organisms showed gross pericardialinflammation. The lethality and lack of inflammation are consistent with the presence of asuperantigen (213). Secreted products of E. faecalis that can cause direct tissue damage arecytolysin and gelatinase (123). The effects of these toxins will be discussed below.
Virulence traits of E. faecalis
Several possible virulence traits are described in literature, including different enterococcaladhesins that are expressed on the surface of E. faecalis like Agg or Esp (95). These adhesinspossibly play a role in the adhesion to different biological surfaces like epithelial cells andsynthetic biomaterial surfaces like indwelling catheters. Furthermore, some virulence factorsare described that are excreted into the environment like cytolysin and gelatinase (95).These factors might play a role in the toxicity of the enterococcus and the survival of themicroorganism in different environments. In Table 1 the results of different studies reportingthe incidence of virulence factors in E. faecalis isolates from different sources aresummarized. The results of the different studies are quite variable, depending on thesources of the E. faecalis isolates and the methods of determination of the virulence factors.By adding up all results of the different studies for medical and commensal isolates, a
PATH
OG
ENES
IS O
F E.
FA
ECA
LIS
INFE
CTI
ON
S
16

significant difference between the two groups can be found for the cytolysin and thegelatinase trait and the presence of the esp gene. Comparing the different studies this waymight be controversial as different methods of detection were used. Recent studies showedan incongruence between the genotypic and phenotypic detection of cytolysin, gelatinaseand Agg with lower percentages in the phenotypic detection (46, 69). This may be due tothe presence of silent genes that are not expressed under the test conditions. Below thedifferent virulence factors and their possible contribution to the pathogenesis of infectionsare summarized. A schematic representation of most virulence factors and their workingmechanism is given in figure 1.
Aggregation substance and sex-pheromone plasmid systemThe Agg was first described as part of the sex-pheromone system, a very efficient systemthrough which E. faecalis can collect plasmids (63, 65). Figure 2 gives a schematic overview
17
CHAPTER
1
Figure 1. Schematic representation of the involvement of the different virulence factors in patho-
genesis.

of the transfer mechanism of this plasmid collecting system (41, 64, 265). The conjugationinvolves three steps: (i) the excretion of small linear peptides, the sex-pheromones, bystrains not possessing the corresponding sex-pheromone plasmid, (ii) the sex-pheromoneinduces a response in donor cells carrying the corresponding sex-pheromone plasmid, thesecells start to express a plasmid-encoded surface protein, the Agg, (iii) the Agg enables closecell-cell aggregation between donor and recipient and transfer of the sex-pheromoneplasmid. Many different sex-pheromone plasmids encoding different virulence traits andantibiotic resistances are described (265). DNA hybridization studies have shown that alldifferent sex-pheromone plasmids contain a homologous DNA region that encodes the Agg(e.g. Asa1 encoded on pAD1), except the Agg on plasmid pAM373 (Asa373), which doesnot fit the overall homology (50, 88, 175). Sequencing of the asa1 gene encoding Aggshowed that this adhesin contained two RGD (arg/gly/asp) motifs that are also present e.g.in fibronectin in which it mediates the binding to integrin receptors present on eukaryotic
PATH
OG
ENES
IS O
F E.
FA
ECA
LIS
INFE
CTI
ON
S
18
Figure 2. Schematic overview of the transfer mechanism of the sex-pheromone system. Theconjugation involves three steps: (1) excretion of sex-pheromones by strains not possessing thecorresponding sex-pheromone plasmid, (2) the sex-pheromone induces a response in donor cellscarrying the corresponding sex-pheromone plasmid, these cells start to express a plasmid-encodedsurface protein, the aggregation substance, (3) the aggregation substance enables close cell-cellcontact between donor and recipient and transfer of the plasmid. The production of pheromone in therecipient is shut down.

cells (87, 208). To date, the sex-pheromone system has been confined to E. faecalis andwas not observed in other enterococcal species although the transfer of plasmid throughthis system is an interesting object of study as this would explain transfer of antibioticresistance and virulence genes. The only evidence for interaction with other bacterialspecies is the fact that expression of Asa373 is also induced by a pheromone produced byStaphylococcus aureus (42, 176).The finding of the RGD motif in Asa1 led to the investigation of the in vitro adherence of E.faecalis expressing Agg to eukaryotic cells. Kreft et al. (132) found a sevenfold higheradhesion to renal tubular cells of E. faecalis constitutively expressing Agg compared to aplasmid free control strain. Interestingly, the binding capacity could be reduced by theaddition of RGD peptide to the epithelial cells prior to the binding assay thus blocking thereceptors on the epithelial cells. Further studies report the role of Agg in the invasion ofenterococci in intestinal epithelial cells or colonic mucosa and show that the presence ofAgg on the surfaces promotes the internalization of enterococci into these tissues to levelssimilar to that of Salmonella typhimurium (119, 185), however Agg did not increase theadhesion and internalization to cells derived from the ileum (212). Another argument forthe role of Agg in interaction with epithelial cells was the fact that Agg expression inLactococcus lactis also increased the internalization to cultured colonic enterocytes (259).Waters et al. (252) used mutations constructed in Asc10, the Agg encoded by the plasmidpCF10, to prove that the Asc10 functional domain and not the RGD motif was critical forefficient internalization by enterocytes. This is in contrary to earlier reports about the role orthe RGD motif in the interaction with epithelial cells. They also found that expression ofAsc10 in the non-aggregating E. faecalis strain INY3000 did not mediate the adhesionwhich indicated that apart from the Asc10 functional domain other enterococcal traitsmight play a role in the eukaryotic cell internalization. This might also explain the differencewith earlier reports on the role of the RGD motif in internalization.Table 1 shows that Asa1 is present in about 50 % of both medical and commensal isolates,providing evidence that although it might enhance host bacterium interaction, it is not aprerequisite for infection. A reason for the lack of enrichment for Asa1 among medicalisolates may be the variable expression of Asa1 in vivo. Agg is produced upon induction byits homologous pheromone. However, serum factors might also induce expression of Agg(108, 132). Only few studies investigated the presence of Asa373 in E. faecalis isolates,these studies showed that the general incidence is much lower than the incidence of Asa1.The role of Agg in pathogenesis was further studied in different animal models. However,results were not consistent and depended on the strains and models used. Two studies tothe role of Agg in endocarditis in rabbits found that Agg contributed to the development ofendocarditis. However, additional factors, cytolysin or binding substance, were necessary todevelop serious disease (39, 213). Another animal study with rats failed to find acontribution of Agg to the development of endocarditis (15), as well as an experimentalstudy to the development of endophthalmitis in rabbits (122) and a study to the virulenceof E. faecalis in the roundworm Caenorhabditis elegans (91). In conclusion, the Agg might contribute to the pathogenesis of E. faecalis infections atdifferent levels. First, it plays a role in the dissemination of plasmids encoding virulencefactors or antibiotic resistance. Furthermore, Agg plays a role in the immune evasion of E.
19
CHAPTER
1

PATH
OG
ENES
IS O
F E.
FA
ECA
LIS
INFE
CTI
ON
S
20
Tabl
e 1.
Prev
alen
ce o
f vi
rule
nce
fact
ors
in E
. fae
calis
isol
ates
fro
m v
ario
us s
ourc
es a
nd t
he e
xclu
sion
of
clon
al r
elat
ions
hip
as r
epor
ted
in t
he li
tera
ture
.
mat
eria
lno
. of
viru
lenc
e fa
ctor
isol
ates
aggr
egat
ion
subs
tanc
e
hem
olys
inge
latin
ase
Asa
1A
sa37
3Es
pef
aAcl
onal
ity
r
efer
ence
vario
us m
edic
al s
ampl
es97
58 (
60)
48 (
49)e
68 (
70)g
not
excl
uded
(118
)fe
ces
med
ical
stu
dent
s23
4 (1
7)a
bloo
d19
085
(45
)bno
t ex
clud
edi
(114
)
vario
us m
edic
al s
ampl
es11
824
(20
)cno
t ex
clud
ed(1
39)
endo
card
itis
354
(11)
14 (
40)
excl
uded
with
(112
)bl
ood
396
(16)
12 (
32)
PFG
Efe
ces
heal
thy
volu
ntee
rs14
0d9
(64)
d
endo
card
itis
447
(16)
24 (
54)
23 (
52)
excl
uded
with
(44)
vario
us m
edic
al s
ampl
es86
32 9
37)
50 (
58)
62 (
72)
PFG
Efe
ces
hosp
ital p
atie
nts
3210
(31
)20
(62
)18
(56
)fe
ces
heal
thy
volu
ntee
rs30
6 (2
0)b
8 (2
7)a
9 (3
0)d
endo
card
itis
153
(20)
1 (7
)hex
clud
ed w
ith(1
5)w
ell w
ater
112
(18)
hPF
GE
bloo
d89
14 (
16)c
49 (
55)e
14 (
16)d
not
excl
uded
(72)
endo
card
itis
3314
(42
)ex
clud
ed w
ith(2
17)
bloo
d10
029
(29
)PF
GE
fece
s he
alth
y vo
lunt
eers
341
93)d
star
ter
cultu
re2
0 (0
)1
(50)
2 (1
00)
0 (0
)2
(100
)no
t ex
clud
ed(6
9)fo
od9
4 (4
4)7
(78)
6 (6
7)3
933)
8 (8
9)va
rious
med
ical
sam
ples
94
(44)
d8
(89)
d7
(78)
d4
(44)
d9
(100
)d
vario
us m
edic
al s
ampl
es15
11 (
73)d
5 (3
3)d
9 (6
0)d
13 (
87)d
excl
uded
with
(68)
ribot
ypin
g
star
ter
cultu
re47
10 (
21)a
23 (
49)e
23 (
49)d
15 (
32)d
17 (
36)d
not
excl
uded
(83)
bloo
d21
9k23
911
)c14
1 (6
4)e
71 (
32)d
not
excl
uded
(247
)
vario
us m
edic
al s
ampl
es58
14 (
24)
46 (
79)
36 (
62)
8 (1
4)28
(48
)58
(10
0)ex
clud
ed w
ith(4
6)fe
ces/
thro
at h
ealth
y vo
lunt
eers
103
(30)
4 (4
0)9
(90)
1 (1
0)2
(20)
10 (
100)
PFG
Een
viro
nmen
t6
0d5
(83)
d2
(33)
d0d
3 (5
0)d
6 (1
00)
tota
l med
ical
isol
ates
1179
281/
1008
(28
)39
7/64
9 (6
1)19
4/42
2 (4
6)9/
73 (
12)
155/
434
(36)
80/8
2 (9
8)to
tal c
omm
ensa
l iso
late
s18
627
/141
(19
)*48
/104
(46
)*62
/129
(48
)18
/106
(15
)26
/108
(24
)*26
/27
(96)
Not
e: d
ata
are
no. (
%),
a de
term
ined
on
agar
pla
te w
ith h
uman
blo
od, b
dete
rmin
ed o
n ag
ar p
late
with
hum
an b
lood
and
with
a D
NA
pro
be, c
dete
rmin
ed o
n ag
ar p
late
with
hor
sebl
ood,
d de
term
ined
with
PC
R o
r D
NA
pro
be,e
dete
rmin
ed o
n ag
arpl
ate
with
gel
atin
, f de
term
ined
on
agar
plat
e w
ith g
elat
in a
nd a
n ag
arpl
ate
with
ski
m m
ilk,
g de
term
ined
with
clum
ping
ass
ay, h
dete
rmin
ed w
ith c
lum
ping
ass
ay, w
este
rnbl
ot a
nd D
NA
pro
bes,
i 12
H+
and
gent
amic
in R
isol
ates
wer
e id
entic
al o
n PF
GE,
khe
mol
ysin
det
erm
ined
on
211
isol
ates
,*
P ≤
0.05
(�
2te
st).

faecalis through prolonged intracellular survival in macrophages and finally, Agg plays a rolein adhesion to and internalization in epithelial cells a first step to infection.
Enterococcal surface proteinThe Esp was initially identified in E. faecalis MMH594, a clinical isolate that caused multipleinfections in a hospital ward outbreak. Esp is a large cell-surface-localized protein that hassimilar structure to other streptococcal surface proteins that have be shown to be virulencedeterminants and contribute to immune evasion through their different number of repeatmotifs (217). Esp was also found to be part of a pathogenicity island that codes for mostknown auxiliary traits that enhance virulence and a number of additional previouslyunknown genes that are rare in non-infection-derived isolates (215). In an animal model ofascending urinary tract infection, significantly higher numbers of E. faecalis expressing Espwere recovered from the bladder and urine but not from the kidney compared to an Espnegative strain. However, no difference in histopathological changes was seen between thetwo strains, suggesting that Esp only contributes to maintenance and colonization of E.faecalis in acute urinary tract infections and thus suggesting an adhesive role for this protein(216). Furthermore, analysis of the relationship between the presence of Esp and biofilmformation showed a high association with the biofilm formation capacity on polystyrenesurfaces (235). Table 1 shows an overall significant difference in the prevalence of Espamong medical and commensal E. faecalis isolates, confirming its role in pathogenesis.
Adhesin to collagen of E. faecalisAdhesion of bacteria to host cells or extracellular matrix proteins is the initial step ininfection. Many different studies to the enterococcal adherence to extracellular matrixcomponents, fibronectin, collagen(s) and laminin are performed. However, these studiesshowed conflicting results depending on the strains used, the experimental protocol or thegrowth conditions (136, 218, 220, 228, 266, 272). Xiao et al. (266) demonstrated anincrease in binding to immobilized laminin and collagen types I and IV when E. faecalis wasgrown at 46 ºC but not at 37 ºC. Searching the genome database of E. faecalis revealed aprotein that showed significant similarities with the collagen-binding protein, Cna, of S.aureus. This protein was given the name adhesin to collagen of E. faecalis (Ace) (203).Disruption mutation studies of the ace gene showed reduction in the 46 ºC growth-elicitedbinding to collagen type I and IV and laminin (179, 180). However, the role of this proteinin infection remains unclear as the ace gene was found in 62 of 62 E. faecalis isolatesregardless of their origin (62). Results from Nallapareddy et al. (180) indicate that Ace mightbe produced during infection because antibodies to Ace were found in 90 % of sera fromendocarditis patients.
E. faecalis antigen AThe E. faecalis antigen A (EfaA) was identified by two groups through screening of differentsurface proteins with sera from endocarditis patients (143, 267). In a mouse peritonitismodel, mice infected with an E. faecalis mutant that lacked the EfaA showed moreprolonged survival compared to mice infected with the parent strain (222). However, thecontribution of EfaA to pathogenesis could not be confirmed through a higher incidence in
21
CHAPTER
1

infection derived isolates as the gene was found in 42 of 42 E. faecalis isolates from bothinfection and healthy volunteers (222) and also in Table 1 EfaA was found in up to 90 % ofmedical and commensal E. faecalis isolates although total numbers are low.
Extracellular polysaccharidesDifferent research groups found several polysaccharides that might play a role in infections.Cloning a genomic library of E. faecalis in cosmid vectors and screening with rabbit immuneserum and serum from patients with enterococcal endocarditis revealed a non proteinantigen that was identified as a carbohydrate (267, 268). Further analysis of this cosmidclone identified a gene cluster, enterococcal polysaccharide antigen (epa), encodinghomologues of many genes involved in polysaccharide biosynthesis (267-269). Evidenceshowed that this epa gene is widespread among E. faecalis isolates and increases resistanceto neutrophilic killing of E. faecalis (232). Mutants with disrupted genes showed delayedkilling and a higher lethal dose in a mouse peritonitis model. Furthermore, the antigen wasdetected in two mucoid E. faecalis strains from chronic urinary tract infections (269).Another study by Huebner et al. (111) isolated and chemically characterized a capsularpolysaccharide antigen by immunization of rabbits with killed E. faecalis strains. Thiscapsular polysaccharide antigen was found on approximately one third of 15 E. faecalisstrains. Furthermore, they showed that antibodies against the capsular polysaccharideantigen were protective in a mouse model of infection (239). Another capsularpolysaccharide was detected by Hancock et al. (104). Evidence for the role of this capsularpolysaccharide in pathogenesis was given by the fact that insertional inactivated mutantswere enhanced susceptible to phagocytic killing and were significantly compromised in theirability to persist in lymphatic tissue in vivo (104).
CytolysinThe cytolysin/hemolysin of E. faecalis is a secreted, posttranslational modified protein thatis regulated by a two-component regulatory system via a quorum sensing mechanism(103). The cytolysin is distantly related to streptolysin S and also to members of a class ofbacteriocins known as the lanbiotics, it lyses human, rabbit and horse erythrocytes and isactive against many Gram-positive bacteria (10). Details of the discovery and charac-terization of the working mechanism are described in a recent review (43). The encodingoperon for cytolysin is either encoded on the sex-pheromone plasmids or integrated intothe chromosome within a pathogenicity island (115, 215). It has been hypothesized that thecytolysin may influence the pathogenesis of infection through lysis of erythrocytes or thedestruction of other host cells (43). Furthermore, the cytolysin may impact pathogenesis bylysing PMNs and macrophages as was shown by Miyazaki et al. (158).Isogenic mutants of the cytolysin operon were used to study the role of the cytolysin in thepathogenesis of infection in different animal models. Generally, these models showed anincreased virulence of cytolytic E. faecalis compared to non-cytolytic E. faecalis. First, Ike etal. (117) examined the contribution of cytolysin to pathogenicity in intraperitoneallyinfected mice. Strains exhibiting the normal cytolysin phenotype were significantly morevirulent than the non-cytolytic insertion mutants. A mutant plasmid with an increasedcytolytic phenotype rendered host strains more virulent than the wild-type cytolytic
PATH
OG
ENES
IS O
F E.
FA
ECA
LIS
INFE
CTI
ON
S
22

enterococci in mice. The role of cytolysin in the pathogenesis of peritonitis in mice wasconfirmed in different studies (67, 223). Dupont et al. (67) found the lowest 50 % lethaldose (LD50) with strains of E. faecalis expressing the cytolysin or a combination of thecytolysin with Agg. In addition, Singh et al. (223) reported a 35-fold lower LD50 with E.faecalis producing cytolysin in a mouse peritonitis model with added sterile rat fecalextracts. Cytolysin has also been shown to contribute to the mortality due to endocarditis ina rabbit endocarditis model (39). Mortality was significantly increased in animals given E.faecalis expressing Agg and cytolysin (55 %) compared with E. faecalis expressing only Agg(15 %) or only cytolysin (no mortality) (39). In an endophtalmitis model, E. fecalis positivefor cytolysin destroyed the neural tissue of the retina and its architecture whereas non-cytolytic strains produced few or no destructive changes (124). Furthermore, in a C. elegansmodel, the cytolysin was observed to enhance nematode killing by E. faecalis (91). Anindication of the virulence of cytolysin in human infections was the observation thatpatients with bacteremia with a cytolytic strain had a fivefold increased risk of mortality. Inthis study, the cytolytic strains were also gentamicin resistant (114). However, Caballero-Granado et al. (35) reported that the mortality associated with high-level gentamicinresistant E. faecalis bacteremia was not significantly different from mortality associated withgentamicin susceptible strains. The role of cytolysin in infection is confirmed by theepidemiological data in Table 1 which show a significant difference in the incidence ofcytolytic E. faecalis among commensal and medical isolates.
ProteasesTwo secreted proteases have been described for E. faecalis, namely gelatinase and serineprotease, and both are regulated by the fsr system that is described below. The gelatinasewas shown to hydrolyze gelatin, collagen, casein, lactoglobulin, porcine myofibrillarproteins, porcine sarcoplasmic proteins, and other small biologically active peptides (149).The gelatinase might therefore contribute to virulence by degradation of specific hostproteins and damaging host tissue. Experiments in animals confirmed the role of gelatinasein virulence. In a rabbit endocarditis model, proteolytic E. faecalis strains were associatedwith partial dissolution of the vegetation and a more severe clinical picture (100). In twomouse peritonitis models the gelatinase positive strains showed a lower LD50 than thegelatinase negative strain (67, 223), however this effect was not observed in a rat peritonitismodel (67). Finally, an E. faecalis deletion mutant defective in gelatinase production wasattenuated in its ability to kill C. elegans (221). Table 1 shows a significant differencebetween the incidences of gelatinase positive isolates from medical and commensal sourcesthus indicating a possible role in infection in humans.The serine protease was detected by sequencing the regions downstream of gelE, the genecoding for the gelatinase (199). Northernblot analysis revealed that gelE and the gene forthe serine protease (sprE) were cotranscribed (199). However, experiments in a mouseperitonitis model with a sprE mutant showed a prolonged survival of the mice compared tosurvival after infection with the parent strain, indicating that the serine protease isindependently important for infection in this model (199).Sequencing the regions upstream of the gelE gene revealed three open reading frames thatwere named the fsr (E. faecalis regulator) locus. The Fsr system is a quorum-sensing system
23
CHAPTER
1

that regulates the expression of gelatinase and serine protease (200). Disruption mutants ofall three open reading frames of the fsr locus showed a prolonged survival of the micecompared to the parent strains in a mouse peritonitis model (199, 221). In addition,mutants in one of the open reading frames of the fsr locus were attenuated in theirpathogenicity in the rabbit endophtalmitis model (178) and C. elegans model (91, 221). Therole of the Fsr system in virulence was confirmed by a retrospective epidemiological studythat found fsr in a significant higher percentage of endocarditis isolates versus of stoolisolates, however this difference was not found for other clinical isolates (195).
Nosocomial infections with E. faecalis
E. faecalis is a frequent cause of hospital acquired infections. In a recent study on thefrequency of isolation of different bacterial species from patients in intensive care units inEurope, E. faecalis was found in about 4 % of the isolates ranking number 8 in the list ofmost frequently isolated microorganisms (79). In many of the nosocomial enterococcalinfections local and/or systemic immunity is compromised for example by catheterization,surgery in areas that are normally colonized with enterococci or medical immunesuppression after transplantation (8, 40). A combination of these factors is present in manyof the critical ill patients at intensive care units, for example in liver transplant patients thatare often considered especially vulnerable to enterococcal infections (248, 250).
Nosocomial transmission and acquisition of enterococci
The intrinsic robustness of E. faecalis may allow members of this species to survive forextended periods of time, leading to its persistence and nosocomial spread. E. faecalis cangrow at 10 to 45 ºC, in 6.5 % NaCl, in the presence of 40 % bile salts and over a broadrange of pH (74). Originally, enterococcal infections were thought to arise from a patient’sown endogenous flora or to be introduced into the abdomen during transplant surgery orits complications (6, 73). However, molecular epidemiological studies provided evidence forepidemic spread of enterococci in a hospital setting and nosocomial acquisition ofenterococci (7, 131, 171). Zervos et al. (274, 275) were among the first to demonstratenosocomial acquisition and nosocomial spread of gentamicin-resistant E. faecalis, usingplasmid content as an epidemiologic marker. They suggested that the transmission mighthave been through the hands of personnel (275). Later, Livornese et al. (141) were the firstto document an inanimate object, in this case rectal thermometer probes, as the mode oftransmission of a vancomycin resistant E. faecium. Removal of the rectal thermometerprobes resulted in termination of the outbreak. These reports were followed by manyreports on nosocomial outbreaks and transmission of antibiotic resistant enterococci (131,172, 191). Risk factors for the nosocomial acquisition of enterococci that are described inthe majority of studies are: previous antimicrobial therapy, duration of hospitalization,severe underlying disease, or invasive procedures (20, 131). Nosocomial enterococcalacquisition and infection are often due to superinfection after the use of antibiotics withlittle or no anti-enterococcal activity like cephalosporins or quinolones (38, 273). Prevention
PATH
OG
ENES
IS O
F E.
FA
ECA
LIS
INFE
CTI
ON
S
24

and control of transmission include the controlled use of antibiotics, active surveillancecultures to identify the reservoir for spread and stringent application of recommendedcontact precautions (171).A variant of the esp gene was detected in all epidemic vancomycin-resistant E. faecium inhospitals but not in non-epidemic animal isolates (260). This indicates that the surfaceprotein Esp is associated with enterococcal colonization and spread. Analysis of themechanism underlying the influence of this surface protein on enterococcal transmissionmight lead to new ways to prevent colonization and transmission.
Clinical manifestation of E. faecalis infections
Urinary tract infectionsThe most frequent hospital infections caused by enterococci are urinary tract infections.Enterococci are found in up to 15 % of urine isolates, ranking only second after Escherichiacoli (23, 24, 78, 142). Risk factors for enterococcal urinary tract infection have beenidentified as urinary tract instrumentation or catheterization, other genitourinary tractpathology and previous use of antibiotics, especially cephalosporin (165). The influence ofinstrumentation on infection and colonization was illustrated by a study by Gross et al. (98).In this study, 30 of 34 urinary tract infections associated with enterococci were preceded byurinary tract instrumentation and not related to antecedent antibiotic therapy. The clinicalmanifestations of enterococcal urinary tract infection are similar to those of otherorganisms. A reliable diagnosis of urinary tract infection can be difficult because enterococciare opportunistic pathogens that can also be colonizers or cause asymptomatic bacteriuria.Different studies were performed to investigate the role of surface proteins of E. faecalis inthe interaction with uroepithelial tissue (132, 216). Kreft et al. (132) showed a potential rolefor Agg in the adhesion of enterococci to renal epithelial cells. In addition, Esp was shownto contribute to colonization and persistence of E. faecalis at the urinary tract (216).
EndocarditisE. faecalis is associated with endocarditis in a significant percentage of cases. In a 2-yearstudy of all endocarditis cases in The Netherlands E. faecalis accounts for about 8 % of theisolates, thereby being the third most common species following Staphylococcus aureusand Streptococcus sanguis (241). The probable source of infection in cases of enterococcalendocarditis is the genitourinary tract (14 to 70 % of cases), followed by the gastrointestinaltract (3 to 27 % of cases) and dental tract (2 to 12 % of cases) (156). Manipulation, surgeryor disease in these areas is generally the immediate cause of the infection. Enterococci caninfect normal valves as well as those with pre-existing damage or pathology. Usually, nativevalve enterococcal endocarditis is a subacute infection with a more protracted course (138,148). In patients with valve replacements, E. faecalis is one of the predominant species inlate prosthetic valve endocarditis (241). Mortality due to enterococcal endocarditis has beenquoted as being between 10 and 40 % (99, 151, 156).Different enterococcal virulence mechanism might be involved in the pathogenesis ofendocarditis (153). As noted above, enterococci cultured in serum exhibit enhanced bindingto Girardi heart cells (101, 102). Furthermore, experiments with E. faecalis endocarditis in
25
CHAPTER
1

animal models suggest that both cytolysin and Agg are associated with increased mortalityand/or vegetation weight (39, 213) and proteases are associated with a more severe clinicalpicture (100).
BacteremiaAbout 5 to 10 % of bloodstream isolates is E. faecalis, thereby E. faecalis ranges number 6in the list of causative microorganisms of bacteremia (77, 142). However, up to 45 % ofenterococcal bacteremias are polymicrobial (90, 148, 219). Enterococcal bacteremia is muchmore common than enterococcal endocarditis. The proportion of significant episodes ofenterococcal bacteremia that was deemed to represent endocarditis ranged from 1 to 30 %,where the percentage generally depended on the definition of bacteremia in the underlyingstudy (156, 169). The sources of bacteremia without endocarditis are similar to the sourcesof endocarditis as mentioned above with soft tissue infections and burn wound infectionsadded to the list. However, for a significant number of bacteremias no source could beidentified (90) which stimulates the speculation that the translocation of enterococci acrossan intact intestinal epithelial barrier might lead to many of these bacteremias (255).The reported mortality among patients with enterococcal bacteremia has been high (up to45 %), most probably because of severe underlying complicating factors (138, 148, 150,219). Only few comparative studies on the attributable mortality and morbidity associatedwith enterococcal bacteremia are published and their results are contradictory partly due todifferences in design of the study. In a prospective, matched case-control study, Caballero-Granado et al. (34) found no increased mortality but an increased duration of stay amongpatients with enterococcal bacteremia. On the other hand, Landry et al. (137) reported anattributable mortality rate due to enterococcal bacteremia of 31 % in a retrospective cohortstudy. Factors predicting a fatal outcome of enterococcal bacteremia are: (i) infection witha resistant organism, (ii) inadequate antimicrobial therapy, (iii) prior use of antimicrobialtherapy, (iv) severe underlying disease, (v) use of intravascular catheter, (vi) renal dialysis,and (vii) prior need for mechanical ventilation (34, 137, 247). Risk factors for thedevelopment of enterococcal bacteremia are: (i) neutropenia, (ii) urinary catheter, and (iii)previous administration of cephalosporins and imipenem (33). These studies underline theimportance of appropriate use of antibiotics with no or little anti-enterococcal activity.One epidemiological study by Huycke et al. (114) showed a relation between the presenceof the virulence trait cytolysin and mortality due to enterococcal bacteremia. However, thisrelation could not be confirmed in a recent study investigating the association between thepresence of enterococcal virulence factors and mortality among patients with enterococcalbacteremia (247).
Intra-abdominal infectionsAlthough enterococci can be isolated in a significant number of intra-abdominal infections,usually as part of a polymicrobial infection, their role in these infections is controversial (55).Animal models of bacterial peritonitis showed that enterococci alone did not cause anyabscess formation (186), but a mixed inoculation of E. faecalis and other aerobe oranaerobe bacteria resulted in death and abscess formation suggesting a synergistic effect ofE. faecalis in the pathogenesis of bacterial peritonitis (152, 162, 163, 186). This finding is
PATH
OG
ENES
IS O
F E.
FA
ECA
LIS
INFE
CTI
ON
S
26

underscored by the fact that antibiotics that lack activity against enterococcus can often beemployed successfully in intra-abdominal infections, even when enterococci are present aspart of the polymicrobial flora (227, 262). However, others suggested that the role of E.faecalis in experimental peritonitis might depend on the presence of virulence factors (67,117). Furthermore, several reviews on bacteremia have revealed that the source ofenterococcal bacteremia is intra-abdominal infection in a number of cases (90, 148, 150,219) and some clinical report describe enterococci as the sole cause of intra-abdominalinfections (48, 273). Finally, Burnett et al. (29) reported enterococcus as a predictor oftreatment failure in complicated intra-abdominal infections. Predisposing factors for thepresence of enterococcus in intra-abdominal infections include age, severity of underlyingdisease, hospital length of stay, and the presence of foreign bodies e.g. drains and priorantibiotic use (29, 40, 273). A problem in evaluating the importance of E. faecalis in thepathogenesis of intra-abdominal infection in humans is that surgical drainage is usually partof the therapy, and drainage alone may be enough to achieve cure.
Skin and soft tissue infectionsE. faecalis accounts for up to 5 % of isolates from skin and soft tissue infections (78, 126).Enterococci generally cause infections only in previously damaged tissues and are notapparently responsible for primary cellulites. Especially in wound infections after abdominalsurgery enterococci are frequently cultured (8). However, since enterococci from skin andsoft tissue infections are frequently cultured in association with other pathogens, their rolein pathogenicity is unclear.
Biomaterial associated infectionsMany nosocomial enterococcal infections are associated with medical devices such asintravascular or urinary catheters, bile drains and prosthetic heart valves (54, 75, 97, 270).Biomaterial-centered infections are initiated by bacterial adhesion and biofilm formation onthe indwelling device. This biofilm can be a source of persistent infection, because thebiofilm mode of growth protects the adhering bacteria against the host defense and theaction of antimicrobial agents (45). These biomaterial related infections cause significantmorbidity and can be very difficult to treat, often removal of the device is necessary (210).Recently, Sandoe et al. (211) reported that E. faecalis isolated from intravascular catheterrelated bloodstream infections produced significant more biofilm compared to bloodstreamisolates from other sources. This suggests that the ability to form a biofilm might be amarker of virulence. By use of a polystyrene microtiter plate biofilm assay, Toledo-Arana etal. (235) confirmed the association of Esp and biofilm formation. However, also E. faecaliswithout Esp or Agg was shown to be able to produce biofilms (133). In this study, theproduction of gelatinase enhanced the biofilm formation of E. faecalis (133) but this wasnot confirmed in another study (127). Furthermore, different environmental conditions caninfluence the ability of E. faecalis to adhere and produce biofilms. Adherence of E. faecalisto different biomaterials increased by growth in the presence of 10 % serum or sub-inhibitory concentrations of ampicillin and vancomycin (85, 86). Biofilm formation alsoincreased in the presence of a nutrient poor medium but decreased with higher osmolarityof the medium (133). Possibly, the different virulence factors of E. faecalis work in concert
27
CHAPTER
1

in a biofilm; the surface proteins Esp and Agg function to mediate clumping or adherenceleading to biofilm formation and thereby increasing the local density of bacteria to the levelnecessary to induce the cell density dependent expression of cytolysin or gelatinaseregulated by the Fsr system (95).
Concluding remarks and future directions
E. faecalis is an emerging pathogen with an increasing incidence among hospitalizedpatients. Therefore E. faecalis attracts more and more attention from medical micro-biologists and infectious disease specialists. Many possible virulence traits of E. faecalis arestudied by use of animal models, in vitro models and isolates from different groups ofpatients. However, animal and in vitro models of infection are hampered by the fact thatthey often do not represent the human situation and different studies frequently showconflicting results. Furthermore, studies to the incidence of virulence factors amongdifferent patient groups are blurred due to the fact that patients with high immunesuppression are also susceptible to commensal strains without virulence factors. Despitethese numerous investigations, the mechanism of virulence of E. faecalis is still notcompletely understood. Results indicate a role for the known virulence traits of E. faecalis.However, other unknown factors might also play a role because enterococci without any ofthe known virulence factors can cause serious infections and some aspects of pathogenesiscan not be explained with the present theories. The recent publication of the entire genome of E. faecalis V583 (192) will lead to thedetection of new virulence traits. Analysis of these new and old virulence traits with the useof newly derived methods like DNA micro arrays might lead to new insights in thepathogenesis of infection and the discovery of ways to prevent or treat these infectionswithout the extensive use of antibiotics.
Scope of the thesis
At the University Hospital Groningen, about 55 liver transplants are performed every year.Liver transplant recipients are very vulnerable to infections and bacteria are the mostcommon cause, with a reported incidence ranging from 30 to 65 % (9, 93, 135, 146, 249,264). Most bacterial infections occur in the first 4 weeks after the transplantation (9, 146,193, 248). Gram positive cocci are the major organism isolated from liver transplant patientswith enterococci accounting for about 10 % of the bacterial episodes (189, 248, 249, 264).Risk factors contributing to bacterial infection after liver transplantation are prolongedsurgery, bleeding, use of choledochojejeunostomy (roux-en-Y) instead of a choledo-chocholedochostomy (duct-to-duct) for biliary anastomosis, repeated abdominal surgeryand CMV infection (189, 194). Furthermore, the isolation of predominantly Gram positivemicroorganisms might be associated with the prophylactic use of antibiotics with no anti-enterococcal activity (146, 189, 193, 276).The concern about E. faecalis as an emerging pathogen, its role in infections among liver
PATH
OG
ENES
IS O
F E.
FA
ECA
LIS
INFE
CTI
ON
S
28

transplant patients and the substantial morbidity and mortality associated with theseinfections led us to study the pathogenesis of E. faecalis infections in this group of patients.The general aim of this thesis was to gain insight in the pathogenesis of E. faecalis infectionswith special interest in liver transplant patients. The starting point for our investigation wasthe microorganism and to find virulence factors and mechanisms of E. faecalis that mightlead to infections or spread. To achieve the general aim of this thesis, the outline of ourstudies pertaining to the underlying questions will be addressed in the subsequent chapters. Part I focuses at the epidemiology and the role of virulence factors of E. faecalis in infectionsin liver transplant patients. In Chapter 2, the differences in prevalence of virulence factorsand genomes (genogrouping) among isolates from liver transplant patients and isolatesfrom other patient groups or healthy volunteers are analyzed. The central question in thischapter was whether E. faecalis from liver transplant patients is different from isolates fromother sources. In Chapter 3, the epidemiology of E. faecalis among liver transplant patientsis investigated. In Chapter 4, the development and evaluation of a rapid non-culturalmethod to detect enterococcal infections and to study their spatial distribution in the sampleis described.In Part II of the thesis the mechanisms of adhesion of E. faecalis to different biomaterials arestudied. Biofilms formed on biomaterials inserted into the human body are a frequentsource of infection. Adhesion is the first step in biofilm formation. In Chapter 5, the role ofthe surface proteins Agg and Esp of E. faecalis on its adhesion to different biomaterialsinvestigated and a proposal for the adhesion mechanism of E. faecalis is given. In Chapter6, the molecular interaction forces between E. faecalis expressing Agg is measured withatomic force microscopy and the influence of antibodies to Agg on these forces isinvestigated. In Chapter 7, the influence of the growth in the presence of (ox) bile of E.faecalis on the numbers of bacteria adhering to different biomaterials and the cell surfaceproperties involved in initial adhesion is studied. In Chapter 8, the results and conclusions ofthis thesis are summarized and discussed.
29
CHAPTER
1


PART IVirulence of Enterococcus faecalis
in the Host


2
CHAPTER
Genogrouping and Incidence of VirulenceFactors of Enterococcus faecalis in Liver
Transplant Patients Differ from BloodCulture and Fecal isolates
Karola Waar, Albrecht B. Muscholl-Silberhorn, Rob J.L. Willems, MaartenJ.H. Slooff, Hermie J.M. Harmsen, and John E. Degener
Journal of Infectious Diseases 2002; 185:1121-1127

Abstract
Enterococcus faecalis is a leading cause of infections in liver transplant patients. This studyreviewed the incidence of virulence factors such as hemolysin, gelatinase, aggregationsubstances (asa1 and asa373) or the enterococcal surface protein (Esp) in isolates from livertransplant patients. In total, 133 isolates from liver transplant patients were compared to 47isolates from feces of healthy volunteers and 66 isolates from blood cultures. Amplifiedfragment length polymorphism (AFLP) analysis indicates that the isolates from differentclinical subgroups can be divided into genogroups with an AFLP similarity of >80% anddifferent virulence factors. Hemolysin and asa1 might be associated with infection, as theyare more frequent in isolates from blood cultures and transplant patients. Esp might beassociated with colonization and spread, because it is more frequent in isolates from fecesof healthy volunteers and transplant patients. An epidemic esp gene-positive strain amongliver transplant patients supports this hypothesis.
E. F
AEC
ALI
SIN
LIV
ERTR
AN
SPLA
NT
PATI
ENTS
34

Introduction
In recent years enterococci have become significant nosocomial pathogens and are nowone of the leading causes of nosocomial infections (159). Enterococcus faecalis causes upto 28 % of the episodes of bacterial infection after liver transplantation (264). The reasonfor the prominent role of this species in liver transplant patients may be the selection ofmore virulent and/or resistant strains of E. faecalis during (anti-microbial) treatment.Different virulence factors of E. faecalis have been described elsewhere (reviewed in 123).Traits of E. faecalis that are often mentioned as possible virulence factors are hemolysin/bacteriocin (Hln) (124), gelatinase (Gel) (100), aggregation substance (Agg) (41) andenterococcal surface protein (Esp) (217). The Hln of E. faecalis lyses human, rabbit and horse erythrocytes and is active against manyGram-positive bacteria (10). Hln increases the virulence of E. faecalis in infection models ofdifferent animal species (117, 124). Gel is capable of hydrolyzing biologically activepeptides like gelatin or collagen (149). In an experimental endocarditis model, Gelproducing strains resulted in more severe clinical findings (100). Agg is a surface proteinencoded by sex-pheromone-responsive plasmids (265). All different sex-pheromoneplasmids contain a homologous DNA region that encodes the Agg (e.g. Asa1 encoded onpAD1), except for Agg on plasmid pAM373 (Asa373), which does not fit the overallhomology (88, 175). Agg increases the number of bacteria adhering to renal and intestinalepithelial cells, suggesting that Agg is important for colonization and translocation of hosttissues by E. faecalis (132, 185). Esp was discovered in an E. faecalis strain that causedmultiple infections within a hospital ward. Studies on the distribution of this surface proteinrevealed a significant enrichment in infection-derived E. faecalis isolates (217). A variant espgene was also found in vancomycin-resistant Enterococcus faecium spreading in hospitals(260).In this study, we used amplified fragment length polymorphism (AFLP) to investigate thegenetic relationship between different groups of epidemiologic nonrelated isolates and toexclude spread of one strain among isolates from one hospital ward. Pulsed field gelelectrophoresis (PFGE) is often considered the standard reference method for the study ofthe nosocomial spread of bacteria (134). However, by use of AFLP we were able toinvestigate the degree of genetic relatedness between non-related E. faecium strains thatwould show no similarity at all with PFGE (130). In addition, AFLP is also of value instudying hospital epidemics with vancomycin resistant E. faecium (VRE) (4).Earlier investigations determined the incidence of these virulence factors in differentpatients groups, but the results were contradictory (44, 114, 118). To our knowledge, untilnow no published research has described the distribution of virulence factors of E. faecalisin liver transplant patients. We determined the incidence of the virulence factors Hln, Gel,Agg and Esp in fecal isolates from healthy volunteers and isolates from blood cultures. Toinvestigate whether virulence factors of E. faecalis are more frequent in liver transplantpatients, we compared our results with the incidence of virulence factors in isolates fromdifferent clinical specimens of liver transplant patients.
35
CHAPTER
2

Materials and Methods
Study design
To investigate whether virulence factors are more prevalent in E. faecalis isolated from livertransplant patients, we determined the incidence of Hln, Gel, Agg and Esp in isolates fromdifferent clinical specimens of liver transplant patients. Weekly surveillance cultures were takenroutinely from stool and throat and other cultures were taken of different clinical sites whenthere were any signs of infection from consecutive patients who received a liver transplant atGroningen University Hospital (The Netherlands). E. faecalis isolates from these cultures wereincluded in the study. The specimens were collected during the first month after the firsttransplantation, patients with a retransplantation more than one month after the first trans-plantation and patients < 18 year were not included. As a control group of invasive clinicallyrelevant E. faecalis, we used blood isolates from nonliver transplant patients. As a controlgroup for nonclinical enterococci we used E. faecalis isolates from feces of healthy volunteers.To exclude multiple identical isolates, we performed AFLP analysis as described later.
Bacteria
From the specimens of the liver transplant group, 133 E. faecalis isolates were culturedbetween March 1998 and March 2000 by routine methodology. After isolation, enterococciwere subcultured on blood-agar plates for further use. The E. faecalis blood isolates (68)were from a collection obtained from patients with E. faecalis bacteremia in GroningenUniversity Hospital between January 1997 and January 2000. No additional clinical criteriawere considered in the group of blood cultures because in that hospital a blood culturepositive for E. faecalis is considered clinically significant. E. faecalis (38 isolates) from fecesof healthy volunteers were collected in 1999 (provided by N. Bruinsma, Academic HospitalMaastricht, The Netherlands) (27). The volunteers were part of a random sample of theopen population living around Groningen. Another 9 isolates were cultured from the fecesof University of Groningen students. The volunteers did not use any antibiotics and werenot hospitalized during the month before E. faecalis isolation.The isolates were identified using a commercially available kit (API 20 Strep; bioMérieux).We used the following E. faecalis strains as controls: OG1X containing pAD1 (asa1+, Hln+)(81), OG1S (Gel+) containing pCF10 (asa1+) (63), OG1X containing pAM373 (asa373+)(42), the plasmid-free strains OG1X (116) and JH2-2 (120) (Hln-, Gel-, Agg-) and the esp-positive strain MMH594 (217) (provided by V. Shankar, University of Oklahoma HealthSciences Center, Oklahoma City).
Hln production
Production of Hln was determined by plating the enterococci onto Todd-Hewitt agar(Oxoid) supplemented with 5 % human blood. Plates were incubated at 37 °C and obser-ved after 24 and 72 h. A clear zone of ß-hemolysis around the bacteria was consideredindicative of the production of Hln.
E. F
AEC
ALI
SIN
LIV
ERTR
AN
SPLA
NT
PATI
ENTS
36

Gel production
Production of Gel was determined by 2 different media. The first method used trypticasesoy agar (Oxoid) supplemented with 1.5 % skim milk. After inoculation, the plates wereincubated overnight at 37 °C. A clear halo around the colonies was considered positive forthe production of Gel. The second method used Todd-Hewitt agar containing gelatin (30 gL-1; Oxoid). After overnight incubation at 37 °C, the plates were cooled for 5 h at 4 °C.Hydrolysis of gelatin was determined by screening the plates for the appearance of a turbidhalo around the colonies.
Clumping assay
Aggregation (clumping) was detected as described elsewhere (65). The supernatant of an overnight culture of the pheromone producing strain JH2-2 (120) in Todd-Hewitt broth (Oxoid) was autoclaved and subsequently used as a source of pheromones. We mixed 2 ml of the supernatant with 2 ml of fresh Todd-Hewitt broth and added 50 µl of overnight cultured enterococci for testing. The mixtures were grown for 4 h at 37 °C in a rotating device at 65 rpm and examined for the formation of aggregates(clumping).
Isolation of DNA
Chromosomal DNA of E. faecalis was isolated according to Boom et al. (21) with someminor modifications. The bacteria were grown overnight in brain-heart infusion broth(Oxoid) at 37 °C on a rotating incubator at 250 rpm. The 1-ml culture pellet wasresuspended in 50 µl of H2O. The bacteria were lysed by incubation with mutanolysin (finalconcentration, 0.04 mg ml-1; ICN Biomedicals) for 30 minutes at 37 °C. After lysis, 950 µlof a mixture of diatom suspension (Acros Organics) and L6 buffer was added and mixed.After 10 min at room temperature, the tube was mixed again and centrifuged, and thesupernatant was discarded. The pellet was subsequently washed twice with ethanol 70 %and once with acetone. After disposal of the acetone, the tube was dried at 37 °C for 30min. The DNA was eluted by adding 200 µl of H2O, and the vessel was mixed briefly andincubated for 10 min at 56 °C, mixed again, and centrifuged for 5 min.
Hybridization with probes for asa1 and asa373
Chromosomal enterococcal DNA was restricted with PvuII prior to Southern blothybridization. The probe used for the detection of asa373 was a 2.1-kB BsrGI/HpaIIrestriction fragment of the gene asa373 cloned into vector pQE30 (Qiagen). Probes for thedetection of asa1-homologous genes were a 1.1- and 1.3-kB PstI restriction fragment ofpAD1 cloned, into vector pUC18. The probes cover great part of asa1 and asa373,respectively, except for the sequences corresponding to those encoding the signal peptideand membrane anchor and, in case of asa1, some adjacent nucleotides. Labeling of theprobes, hybridization and stringent washing was done using the ECL direct nucleic acid
37
CHAPTER
2

labeling and detection system (Amersham Pharmacia Biotech) as indicated by themanufacturer. The filters were hybridized for 5 h at 42 °C.
AFLP
We performed the AFLP as described by Willems et al. (261). In brief, the E. faecalis DNAwas simultaneously digested with EcoRI and CfoI and ligated to a single adapter thatresulted in circularized DNA molecules. The molecules were amplified using 2 primers basedon the adapter sequence, each with an extra selective base and one primer was labeled witha fluorescent dye. The amplification products were separated on a 96 capillary ABI PRISM3700 DNA analyzer. AFLP patterns were analyzed with BioNumerics software version 1.5(Applied Maths). We calculated the Pearson product moment correlation and used theunweighted pair group method with arithmetic averages for cluster analysis.
PCR for esp
The presence of the esp gene in DNA isolated from E. faecalis was determined by poly-merase chain reaction amplification by use of primers esp11 and esp12 as described byShankar et al. (217). According to Shankar et al. (217) detection of the Esp correlatedprecisely with the presence of the esp gene; therefore, isolates positive for the esp genewere considered also to express Esp on its surface.
Antibiotic sensitivity testing
Sensitivity of E. faecalis isolates was tested with agar disc diffusion. The inoculum wasprepared from isolated colonies on blood agar plates. The colonies were suspended inphysiologic saline to a turbidity of 0.5 McFarland U subsequently diluted 1:100 and appliedto iso-sensitest agar plates (Oxoid). After the plates were dried for �5 min at roomtemperature, we applied neosensitabs tablets with amoxicillin (30 µg), gentamicin (250 µg),vancomycin (5 µg), tetracycline (80 µg), erythromycin (78 µg) and ciprofloxacin (10 µg;Rosco). After 24 h at 37 °C the plates were checked for semi-confluent growth and theinhibition zones were determined, the zone of vancomycin was checked again after 48 h at37 °C. Minimal inhibitory concentration breakpoints and corresponding inhibition diameterswere used as determined for the University Hospital Groningen according to the Dutchnational guidelines. The sensitivity and resistance breakpoints were respectively ≤ 2 and > 16 µg ml-1 for amoxycillin, ≤ 1 and > 4 µg ml-1 for gentamicin, ≤ 4 and > 8 µg ml-1 forvancomycin, ≤ 1 and µ 4 µg ml-1 for tetracycline, ≤ 1 and > 2 µg ml-1 for erythromycin, and≤ 1 and > 2 µg ml-1 for ciprofloxacin.
Statistical analysis
Categorical data were analyzed by the �2 method with Yate’s correction. Significance wasdefined as P ≤ 0.05.
E. F
AEC
ALI
SIN
LIV
ERTR
AN
SPLA
NT
PATI
ENTS
38

Results
Bacteria and patients
During the study period 60 patients > 18 years old received liver transplants. We studied133 E. faecalis isolates from 43 of these patients. Of the 133 isolates, 98 were cultured fromroutine surveillance specimens from feces or throat of 40 different patients. Another 35isolates from 19 patients were cultured from clinical sites when an infection was suspected:sputum (5), ascites (8), wound (10), bile (10), blood (1), and intravascular catheter (1).From blood cultures of 66 patients, 68 E. faecalis isolates were collected. From feces of 46healthy volunteers, 47 E. faecalis isolates were cultured. Of the 43 liver transplant patientsin this study, 31 had > 1 isolate collected (mean, 3.9 isolates/patient). Ten (32 %) of thesepatients carried ≥ 2 nonidentical E. faecalis isolates.
Genogrouping of isolates by AFLP
To investigate the genetic relationship and to exclude multiple identical isolates, AFLPanalysis combined with analysis of the pattern of virulence factors was done. E. faecalisisolates with a similarity of ≥ 90 % in AFLP analysis and an identical pattern of virulencefactors were considered identical. The cutoff point of 90 % similarity was chosen accordingto Willems et al. (260) who considered VRE strains to be epidemic if the AFLP patternshowed ≥ 90 % similarity. The comparison was made by clinical subgroup; therefore,identical isolates could be from blood cultures, liver transplant patients or feces from healthyvounteers.Of the 248 investigated isolates 113 were considered nonidentical (34 from 21 livertransplant patients, 44 from blood cultures of 43 patients and 35 from 35 healthyvolunteers). Identical isolates were omitted from the study not using further criteria. Figure1 shows the genetic relationships, as determined by AFLP, of the 113 different E. faecalisisolates. Eleven genogroups were discerned in which the strains shared > 80 % of therestriction fragments. Table 1 shows the numbers of isolates per AFLP genogroup. Themajority of the isolates clustered in 3 groups: A, B and H. Most genogroup A isolates werefrom blood cultures, while the majority of the genogroup B isolates were from livertransplant patients. In genogroup H, most isolates were isolates from feces of healthyvolunteers.AFLP analysis also revealed a large group of 65 identical isolates from 23 liver transplantpatients with a high similarity (≥ 95 %) that were collected during a 1-year period (data notshown). Of these, 48 had an identical virulence pattern, namely Hln+, Gel-, asa1+, asa373-and esp+, 5 isolates were esp- and 12 were asa373+. Before or after this year no isolateswith the same AFLP pattern were collected during the study. These isolates clustered togenogroup B. This strain was not especially associated with infections in liver transplantpatients (data not shown).
39
CHAPTER
2

E. F
AEC
ALI
SIN
LIV
ERTR
AN
SPLA
NT
PATI
ENTS
40
Figure 1. Dendrogram of 113 nonidenticalE. faecalis isolates. Nos. on horizontal axisindicate percentage similarities (Pearsonproduct-moment correlation coefficient).A-K, different genogroups with >80%shared restriction fragments.

Presence of Hln, Gel, asa1, asa373 and esp among the E. faecalis isolates
Figure 2 shows the frequencies of virulence factors in the three largest genogroups: A, B andH. The predominant virulence factors by group were as follows: group A, Hln, Gel, andasa1; group B, Hln, asa1, and esp; and group H, Gel, asa1 and esp. The frequencies ofvirulence factors per genogroup were also divided into the different clinical subgroups(blood cultures, feces from health volunteers, liver transplant patients). However, thedistribution of the virulence factors in the genogroups per isolate group (data not shown)was very similar to the distribution of all the isolates (figure 2). Comparison of the virulencefactors of group A, B and H revealed that Hln+ and asa1+ isolates were significant morefrequent in group A (the blood culture isolates and group B (isolates from liver transplantpatients; �2, P ≤ 0.05). Whereas esp+ isolates are significant more frequent among group H,which primairly contained isolates from feces of healthy volunteers and group B, whichcomprised isolates from liver transplant patients (�2, P ≤ 0.05). Finally, Gel+ isolates weresignificantly more frequent among group H compared to group B (�2, P ≤ 0.05).Figure 3 shows the frequencies of the virulence factors per clinical subgroup. A significanthigher frequency of esp+ isolates was found in isolates from liver transplant patients (71 %)compared to isolates from feces of healthy volunteers (40 %) and blood cultures (45 %; �2,P ≤ 0.05). Also, in the isolates from liver transplant patients, the frequency of Gel+ isolateswas significant lower (32 %) than in the isolates from feces of healthy volunteers (66 %; �2,P ≤ 0.05). Asa373 was more frequent among isolates from liver transplant patients (15 %)than among isolates from blood cultures (5 %) or healthy volunteers (6 %).The isolates from liver transplant patient could be divided into 2 groups: those that werecultured from feces or throat during surveillance and isolates that were cultured fromspecimens that were taken if an infection was suspected. When we compared the incidenceof virulence factors in these two groups we did not find a significant difference (data not
41
CHAPTER
2
Table 1. Number of isolates per genogroup when the AFLP dendrogram is abridged at 80 % and allidentical isolates are omitted.
GroupNumber of E. faecalis isolates froma
Totalblood cultures Liver transplant patients feces of healthy volunteers (113 isolates)
(44 isolates, 43 patients) (34 isolates, 21 patients) (35 isolates, 35 volunteers)
A 11 (85) 2 (15) 13B 4 (24) 12 (70) 1 (6) 17C 2 (100) 2D 1 (33) 2 (67) 3E 1 (100) 1F 1 (50) 1 (50) 2G 1 (50) 1 (50) 2H 21 (34) 11 (18) 30 (48) 62I 1 (25) 3 (75) 4J 1 (100) 1K 1 (17) 5 (83) 6
a Numbers in parenthesis are percentage of total per genogroup.
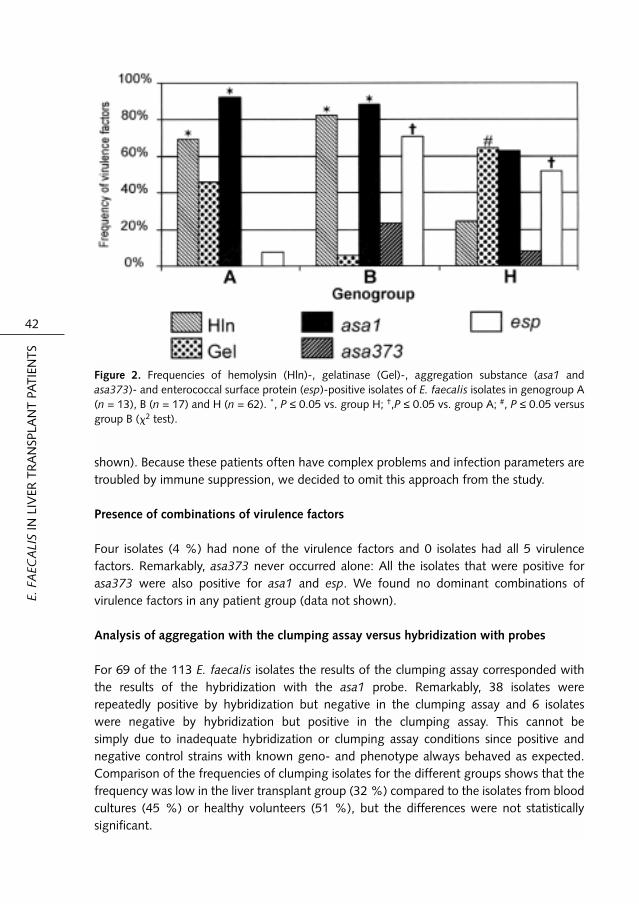
shown). Because these patients often have complex problems and infection parameters aretroubled by immune suppression, we decided to omit this approach from the study.
Presence of combinations of virulence factors
Four isolates (4 %) had none of the virulence factors and 0 isolates had all 5 virulencefactors. Remarkably, asa373 never occurred alone: All the isolates that were positive forasa373 were also positive for asa1 and esp. We found no dominant combinations ofvirulence factors in any patient group (data not shown).
Analysis of aggregation with the clumping assay versus hybridization with probes
For 69 of the 113 E. faecalis isolates the results of the clumping assay corresponded withthe results of the hybridization with the asa1 probe. Remarkably, 38 isolates wererepeatedly positive by hybridization but negative in the clumping assay and 6 isolates were negative by hybridization but positive in the clumping assay. This cannot be simply due to inadequate hybridization or clumping assay conditions since positive andnegative control strains with known geno- and phenotype always behaved as expected.Comparison of the frequencies of clumping isolates for the different groups shows that thefrequency was low in the liver transplant group (32 %) compared to the isolates from bloodcultures (45 %) or healthy volunteers (51 %), but the differences were not statisticallysignificant.
E. F
AEC
ALI
SIN
LIV
ERTR
AN
SPLA
NT
PATI
ENTS
42
Figure 2. Frequencies of hemolysin (Hln)-, gelatinase (Gel)-, aggregation substance (asa1 andasa373)- and enterococcal surface protein (esp)-positive isolates of E. faecalis isolates in genogroup A(n = 13), B (n = 17) and H (n = 62). *, P ≤ 0.05 vs. group H; †,P ≤ 0.05 vs. group A; #, P ≤ 0.05 versusgroup B (�2 test).

Sensitivity to antibiotics
We compared the percentage of sensitivity among the nonidentical isolates by clinicalgroup. The results showed that the isolates from liver transplant patients and blood cultureswere, in general, more resistant than the fecal isolates from healthy volunteers. Thedifference was most clear for ciprofloxacin: All the isolates from feces of healthy volunteerswere sensitive compared with 56 % of isolates from liver transplant patients and 64 % ofblood culture isolates. Similar differences were seen for gentamicin (94 % vs. 52 and 39 %respectively), erythromycin (77 % vs. 36 and 33 %) and tetracycline (54 % vs. 20 and 28%). All isolates were sensitive to amoxicillin; only 1 isolate from blood cultures wasvancomycin resistant.
Discussion
The question addressed by this study was whether the incidence of some or all tested E.faecalis virulence factors were higher in liver transplant patients. Information about thefrequency of virulence factors in E. faecalis isolates from this group of patients might enableconclusions about their role in the high incidence of infections. We determined theincidence of the virulence factors Hln, Gel, Agg (asa1 and asa373 homologies) and esp inisolates from liver transplant patient specimens and compared the results with the incidenceof the virulence factors in blood culture isolates of nonliver transplant patients and fecalisolates from healthy volunteers. Analysis of the results per clinical subgroup showed thatesp was significantly higher and Gel significantly lower in isolates from liver transplant
43
CHAPTER
2
Figure 3. Presence of hemolysin (Hln), gelatinase (Gel), the aggregation substance (asa1 and asa373)and enterococcal surface protein (esp) in isolates from liver transplant patients (LT; n = 34), bloodculture (BC; n = 44), and feces of healthy volunteers (FHV; n =35). *,P ≤ 0.05 vs. FHV; †,P ≤ 0.05 vs.BC (�2 test).

patients. However, results from the genogrouping after AFLP analysis showed that theisolates from the different clinical subgroups clustered into different genogroups withdifferent predominant virulence factors.Comparison of these genogroups showed that Hln+ and asa1+ isolates might be associatedwith infections, as they are mainly found in the genogroups representing blood cultures andliver transplant patients. On the other hand, esp+ isolates might be associated withcolonization as they are mainly found in the genogroups represented by fecal isolates fromhealthy volunteers or liver transplant patients. Esp might also play a role in the spread of E.faecalis in liver transplant patients as the strain that spread over the liver transplantdepartment during one year was also esp+.The percentages of esp+ isolates reported in this study are much higher than in the study ofShankar et al. (217). They found 29 % of blood culture isolates and 3 % of isolates fromfeces of healthy volunteers were positive for esp, whereas we found much higherfrequencies (45 and 40 %, respectively). However, Eaton and Gasson (69) reported 44 %of medical E. faecalis isolates were positive, as were and 33 % of E. faecalis isolated fromfood.Clustering of enterococci in distinct genogroups by AFLP analysis was described by Willemset al. (261). By use of AFLP analysis, they showed that E. faecium genogroups wereassociated with particular hosts and environments (so called ecovars). They reported thatisolates from healthy volunteers or hospitalized patients from different countries withdifferent genotypes belonged to the same genogroup; however, there was a cleardichotomy between isolates from hospitalized and nonhospitalized persons. These resultsare in line with our results for E. faecalis.Although the E. faecalis conjugative plasmid pAM373 was originally found in a clinicalisolate (42), to our knowledge this is the first study on the occurrence of this plasmid. Arelatively low frequency of asa373 encoding the Agg of this plasmid was found among ourisolates from blood cultures (5 %) and feces of healthy volunteers (6 %) but a somewhathigher frequency was found in isolates from liver transplant patients (15 %). Surprisingly, allisolates positive for asa373 were also positive for asa1 and esp. Thus, it is possible that thesetraits are linked. If so, it will be difficult to study the role of asa373 in infections alone. The discrepancy between the results of the clumping assay and the hybridization withprobes for asa1 might indicate a difference between phenotype (expression of aggregationsubstance) and genotype (presence of the gene encoding for the aggregation substance).The clumping reaction is sometimes difficult to interpret and this assay is possibly much lesssensitive than the hybridization. The clumping phenotype of strains obviously lacking Aggmay, of course, indicate the existence of unrelated, still unknown factors also mediating cellclumping of E. faecalis. The in vivo expression of Agg is very difficult to test. Both thehybridization and the clumping assay do not analyze the in vivo expression of Agg. Asfactors other than pheromone (e.g. serum) are described that might induce expression ofAgg (108) we think that the incidence of the gene encoding Agg is still the best measure tostudy the possible presence of Agg in vivo.AFLP analysis revealed a large group of E. faecalis isolates with a ≥ 95 % similar AFLPpattern among liver transplant patients, which is highly suggestive of spread of bacteriabetween liver transplant patients. Using AFLP it is very difficult to obtain patterns that are
E. F
AEC
ALI
SIN
LIV
ERTR
AN
SPLA
NT
PATI
ENTS
44

100 % identical; Willems et al. (261) reported 95-99 % similarity between quadruplicates.Thus, we might conclude one strain that spread throughout the department. The epidemicstrain in our study was mostly isolated directly after the transplant, suggesting that transferoccurred very quickly on the intensive care unit or during admissions for pretransplantmonitoring. In this study, E. faecalis isolates with a similarity ≥ 90 % in AFLP analysis wereconsidered identical. In a hospital setting, epidemic spread of one clone was considered ifthe AFLP pattern showed ≥ 90 % similarity. The lower similarity of a clone in an epidemicalhospital setting might be due to the transfer of small genetic elements (260). Because ourstudy was carried out in one hospital and the surrounding geographical area, we thought itessential to exclude possible clones that might obscure the analyses. However, this wouldalso mean that nonidentical isolates might be omitted from the study. Thus, to identify asmany nonidentical isolates as possible, we included the virulence pattern in the analysis.A factor that might play a role in colonization of liver transplant patients is antibioticresistance and treatment with antibiotics. As prophylaxis during the liver transplant, thepatients receive amoxicillin with clavulanic acid and ciprofloxacin. For selective boweldecontamination, colistin and tobramycin are administered. We tested the sensitivity of theE. faecalis isolates for different antibiotics and found a higher percentage of resistant strainsamong the liver transplant patients. Some of the sex-pheromone plasmid encoded virulencefactors are associated with antibiotic resistance (265). Treatment with antibiotics might leadto selection of more virulent strains.In summary, we show that E. faecalis isolates from different groups of patients and healthyvolunteers can be divided into different genogroups with > 80 % shared restrictionfragments and each genogroup with specific predominant virulence factors. By combininggenogroups and virulence factors, we show that hemolysin and asa1 might be associatedwith infection and esp with colonization in liver transplant patients. The finding of anepidemic esp+ strain among liver transplant patients supports the last hypothesis. Theseresults might be important in understanding the high frequency of infections with E. faecalisin liver transplant patients.
We thank the staff of the diagnostic department of Medical Microbiology, for collecting theE. faecalis isolates of liver transplant patients, Henk Jans, Kor Kooi, Carmen Lehner,Margriet Postma and Marga van Santen-Verheuvel, for excellent technical assistance, andRudi Tonk, for reproduction of the figures in this article.
45
CHAPTER
2


Molecular Epidemiology ofEnterococcus faecalis in Liver
Transplant Patients at the UniversityHospital Groningen
Karola Waar, Rob J. L. Willems, Maarten J. H. Slooff, Hermie J. M. Harmsen, and John E. Degener
Journal of Hospital Infection 2003; 55:53-60
3
CHAPTER

Abstract
We report the molecular epidemiology of Enterococcus faecalis in liver transplant patientstransplanted at the University Hospital Groningen (The Netherlands) as determined byamplified fragment length polymorphism (AFLP) typing. A total of 133 E. faecalis isolateswere cultured from the faeces and throat (95 isolates) or clinical sites (35 isolates) of 43 livertransplant patients. Among these 133 isolates, 15 different AFLP types could be identifiedwith 90 % AFLP similarity. Of these 15 groups, nine contained isolates from more than onepatient, which may indicate transmission of E. faecalis isolates between patients. In five ofthese groups transmission could be explained by the fact that patients carrying identicalstrains were staying in the same ward at the same time. One of these epidemic isolates(AFLP type K) distinguished itself by colonizing 23 liver transplant patients during 15months. Antimicrobial susceptibility testing did not reveal any multi-resistant isolates. Thisstudy showed that transmission of susceptible E. faecalis isolates occurs frequently on theliver transplant wards. Detection of this transmission and understanding of the mechanismis important, as it might also be an indicator of possible transmission of enterococci resistantto antibiotics.
MO
LEC
ULA
R E
PID
EMIO
LOG
Y O
F E.
FA
ECA
LIS
IN L
IVER
TR
AN
SPLA
NTA
TIO
N
48

Introduction
In recent years enterococci have become one of the leading causes of nosocomial infectionsand their incidence is increasing (131, 159). The seriousness of infections with enterococcihas been underscored by different reports, which have shown that enterococcal bacteremiais associated with an increased mortality and prolonged duration of hospital stay (34, 137).Enterococcal infections are becoming more difficult to treat due to emerging antibioticresistance (71). Enterococcus faecalis is an important cause of episodes of bacterial infectionin patients receiving a liver transplant (LT) (93, 249, 264). The reason for the prominent roleof this species in LT patients is not clearly understood. Selection and spread of more virulentand/or resistant strains during anti-microbial treatment may play a role as enterococci arenot included in the spectrum of most prophylactic antibiotic regimens (6, 189).Originally, enterococcal infections were thought to arise from a patient’s own endogenousflora or to be introduced into the abdomen during transplant surgery or its complications (6,37). However, molecular epidemiological studies provided evidence for epidemic spread ofenterococci in a hospital setting and nosocomial acquisition of enterococci (7, 131).Information about the nosocomial spread of enterococci is important to detect and preventtransmission of strains that might be virulent or resistant and cause infections in time. In anearlier study of the incidence of virulence factors in E. faecalis isolates from LT patients wereported that some E. faecalis surface proteins, like the enterococcal surface protein, mightbe associated with spread between LT patients (chapter 2).Different molecular techniques are used to study the molecular epidemiology of bacteria.Pulsed field gel electrophoresis (PFGE) is often considered the standard reference method inthe study of the nosocomial spread of strains in hospitals (134, 236). However, amplifiedfragment length polymorphism (AFLP) has similar power and has been more robust instudying hospital epidemics with vancomycin resistant Enterococcus faecium (VRE) (260,261) and it has been suggested that AFLP could serve as “gold standard” for molecularepidemiology of VRE (4). Furthermore, AFLP typing has been used to study the molecularepidemiology of a wide range of bacterial species (52, 244).In the present study, we report the screening of LT patients for E. faecalis combined withDNA typing using AFLP to investigate the molecular epidemiology of E. faecalis amongthese patients. Aim of this study was to gain insight in the epidemiology and the (high)infection rate with E. faecalis in LT patients.
Materials and Methods
Study design
Weekly routine surveillance cultures from stool and throat and other cultures from clinicalsites with signs of infection were taken from consecutive patients who received a LT atUniversity Hospital Groningen (The Netherlands) between March 1998 and March 2000.The liver transplant unit consisted of two wards (ward 1 and 2) and one intensive care unit(ICU). Ward 1 consisted of 31 beds for LT patients and patients with other gastro-intestinal
49
CHAPTER
3

diseases. LT patients generally stayed on this ward during pre- and post-transplantevaluation. Ward 2 consisted of 30 beds for LT patients and patients undergoing hepato-biliary or vascular surgery. LT patients generally stayed on this ward for direct post-transplant treatment. The ICU contained 12 beds and patients demanding intensive careafter surgery stayed on this unit. Nurses were generally dedicated to one ward, whereasmedical doctors worked on the three different wards.From every culture, one E. faecalis isolate was included in the study, presuming thatenterococci from one culture with similar colony morphology and identification wereidentical isolates. The specimens were collected during the first month after the first LT.Patients with a re-transplant more than one month after the first transplantation andpatients younger than 18 year were not included. To investigate the clonal relationship of E.faecalis isolates AFLP analysis was performed.
Patients and bacteria
The throat and stool specimens were cultured on blood agar plates CM55 (Oxoid)containing 5 % sheep blood and blood agar plates with 20 µg ml-1 aztreonam added. Everyputative enterococcus colony was subcultured on blood agar for further identification. Theisolates were identified using a commercially available kit (API 20 Strep; bioMérieux).
AFLP
Chromosomal DNA of E. faecalis was isolated according to Boom et al. (21) with someminor modifications. The bacteria were grown overnight in brain-heart infusion broth(Oxoid) at 37 °C on a rotating incubator at 250 rpm. After centrifugation, the 1-ml culturepellet was resuspended in 50 µL water. The bacteria were lysed by incubation withmutanolysin (final concentration: 0.04 mg ml-1; ICN Biomedicals) for 30 minutes at 37 °C.After lysis, 950 µl of a mixture of diatom suspension (Acros Organics) and L6 buffer (21)was added and mixed. After 10 min at room temperature, the tube was mixed again andcentrifuged, and the supernatant was discarded. The pellet was subsequently washed twicewith ethanol 70 % and once with acetone. After disposal of the acetone, the tube was driedat 37 °C for 30 min. The DNA was eluted by adding 200 µL water, and the vessel was mixedbriefly and incubated for 10 min at 56 °C, mixed again, and centrifuged for 5 min.The AFLP analysis was performed as described by Willems et al. (261). In brief, the E.faecalis DNA was simultaneously digested with EcoRI and CfoI and ligated to a singleadapter that resulted in circularised DNA molecules. The molecules were amplified usingtwo primers based on the adapter sequence, each with an extra selective base, and oneprimer was labelled with a fluorescent dye. The amplification products were separated on a96 capillary ABI PRISM 3700 DNA analyser. AFLP patterns were analysed with BioNumericssoftware version 1.5 (Applied Maths). The Pearson product moment correlation wascalculated and the unweighted pair group method with arithmetic averages was used forcluster analysis. In a previous study the similarity percentage of AFLP banding patterns ofepidemiological related strains in a hospital outbreak was > 90 % (260, chapter 2).Therefore, in this study, strains were considered to be identical when AFLP banding patterns
MO
LEC
ULA
R E
PID
EMIO
LOG
Y O
F E.
FA
ECA
LIS
IN L
IVER
TR
AN
SPLA
NTA
TIO
N
50

were more than 90 % similar. Isolates were considered epidemic when the same AFLP typewas found in two or more patients, who could have been in contact, having stayed at thesame ward during overlapping time periods.
Antibiotic susceptibility testing
From each patient, one E. faecalis isolate per AFLP group (total 56 isolates) was tested withagar disk diffusion for susceptibility to gentamicin (250 µg), erythromycin (78 µg),tetracycline (80 µg), ciprofloxacin (10 µg), amoxicillin (30 µg), and vancomycin (5 µg). Theappropriate inoculum was derived from one or more colonies on blood-agar platessuspended in physiologic saline to a turbidity of 0.5 McFarland units, subsequently diluted1:100 and applied to iso-sensitest agar plates (Oxoid). After the plates were dried for about5 minutes at room temperature, the neosensitabs tablets (Rosco) were applied. After 24 hat 37 °C the plates were checked for semiconfluent growth and the inhibition zones weredetermined, the zone of vancomycin was checked again after 48 h at 37 °C. Inhibition zonediameters corresponding to minimal inhibitory concentration breakpoints andcorresponding were read as determined for the University Hospital Groningen according tothe Dutch national guidelines (51). The sensitivity and resistance breakpoints wererespectively ≤ 1 and > 4 µg ml-1 for gentamicin, ≤ 1 and > 2 µg ml-1 for erythromycin, ≤ 1and > 4 µg ml-1 for tetracycline, ≤ 1 and > 2 µg ml-1 for ciprofloxacin, ≤ 2 and > 16 µg ml-1
for amoxicillin, and ≤ 4 and > 8 µg ml-1 for vancomycin. Data from the antimicrobialsusceptibility testing were analysed by the �2 method. Significance was defined as P ≤ 0.05.
Results
Bacteria and patients
During the study period 60 patients older than 18 years received LTs. We studied 133 E.faecalis isolates from 43 of these patients (chapter 2). Of these isolates, 35 were culturedfrom clinical sites: wound (10), bile (10), sputum (5), ascites (8), blood (1) and intravascularcatheter (1). These clinical sites were cultured when there were signs of infection, however,it was not proven that the E. faecalis isolates from these sites caused infection. From 31 ofthe 43 LT patients in this study more than one isolate was collected (mean, 3.9isolates/patient) and nine of these patients (patient number 2, 6, 7, 10-12, 18, 19, 22)carried isolates of more than one AFLP type. In 14 of these 31 patients (patient number 1,2, 6, 12, 18, 22, 23, 25, 29, 30, 34, 40-42), E. faecalis with identical AFLP types (similarity> 90 %) were isolated from both routine cultures and clinical sites. This indicates that inthese patients the faeces or throat might provide reservoirs for enterococci that might causeinfections. However, a common source (e.g., environment or hands) could also have led tocolonisation of both faeces and/or throat and the clinical site.
51
CHAPTER
3

Epidemiology of E. faecalis
Figure 1 shows the genetic relationship of the 133 E. faecalis isolates recovered from 43 LTpatients. Among these isolates 15 different AFLP types (similarity > 90 %) were identifiedof which nine AFLP types (A, B, G, H, I, J, K, M and O) contained isolates from more thanone patient, which is compatible with spread of E. faecalis isolates between patients.Transmission from patient to patient could be explained by geographic location andtemporal relation between patients carrying isolates with AFLP type I, J, K, M and O. Of thefive patients carrying AFLP type H isolates, transmission within two clusters of respectivelytwo (patient 11 and 12) and three (patient 7, 9 and 10) patients could be explained bygeographic location and temporal relation. However, transmission between the two clusterscould not be explained. Transmission of the AFLP type G isolates between the two patientscould be explained by survival of the E. faecalis on the hands of personnel or surfaces forone day, whereas transmission of AFLP type A and B isolates between the two patientscould only be explained by survival of E. faecalis for long periods of two and one monthrespectively. Transmission from patient to patient of AFLP type A, B, I, J, G and H could haveoccurred on LT ward 1, transmission of AFLP type K and M on LT ward 1 or 2, whereastransmission of AFLP type O could only have occurred on the ICU.Remarkable was the transmission of a genetically highly homologous strain (AFLP type K,similarity generally > 95 %) among 23 patients between September 1998 and December1999. Nine other patients that also received a LT during this period and stayed at the samewards did not acquire this epidemic strain. Of these nine patients, six were transplantedduring the first or last two months of the period that AFLP type K strain was predominanton the LT ward. Figure 2 shows the stay of the LT patients carrying AFLP type K E. faecalison the different LT wards during the study period. All patients carrying the epidemic strainstayed on the same ward during the same period before or after the LT, which illustrates thepossibility of transmission on the LT wards. Although cultures were only taken after the LT,transmission likely occurred during a previous stay in the hospital, e.g. for pretransplantmonitoring, as in most cases the epidemic strain K was isolated immediately after theoperation. The E. faecalis strain AFLP type K was not isolated during the study period beforeSeptember 1998 or after December 1999 from LT patients nor from the faeces of healthyvolunteers from the same geographical area or blood culture isolates from patients at thesame hospital during the same period (data not shown) (chapter 2). These datademonstrate epidemic spread of one E. faecalis clone (AFLP type K) among LT patients.The duration of hospital stay during one year before LT varied between one and 135 days,but the average duration did not differ significantly between the patients with and withoutthe epidemic strain K. Three patients were not colonized with isolates from any of theepidemic groups, their average duration of stay before transplantation also did not differsignificantly from the other patients.
Characteristics of epidemic isolates
The characteristics of the epidemic isolates (AFLP types A, B, G, H, I, J, K, M, and O; total126 isolates), with respect to isolation site and antibiotic resistance, were compared with
MO
LEC
ULA
R E
PID
EMIO
LOG
Y O
F E.
FA
ECA
LIS
IN L
IVER
TR
AN
SPLA
NTA
TIO
N
52

53
CHAPTER
3
Figure 1. Dendrogram of 133 E. faecalisisolates recovered from 43 liver transplantpatients. Percentage on horizontal axisindicates percentage similarities (Pearsonproduct-moment correlation coefficient). A-O,different AFLP-types with a similarity of > 90%. Patients are numbered 1 to 43; numbersbehind the branches in the dendrogramindicate the patient from which the E. faecaliswas isolated and the source (routine or clinicalisolate).aRoutine surveillance isolates from faeces orthroat, bincludes wound (10), bile (10), ascites(8), sputum (5) blood (1) and intravascularcatheter (1), cfive patients had two differentisolates and four patients had three differentisolates

non-epidemic isolates (AFLP type C, D, E, F, L, and N; total 7 isolates). Twenty-five percentof the epidemic isolates originated from clinical sites, in contrast to 43 % of the non-epidemic isolates. In total, 26 % of the isolates came from clinical sites. These data suggestthat the epidemic isolates may spread well but mainly colonize the patients. Table 1 showsthe susceptibility of 56 E. faecalis isolates to gentamicin (high level resistance),erythromycin, tetracycline, and ciprofloxacin as determined by agar disk diffusion. Allisolates were susceptible to amoxicillin and vancomycin. The results of the �2 test showed asignificant difference in antibiotic resistance between epidemic and non-epidemic isolatesfor gentamicin, erythromycin, and ciprofloxacin; the epidemic strains were more resistant.
Discussion
We report the molecular epidemiology of E. faecalis in LT patients, a group prone toinfections with this microorganism. We found that E. faecalis isolates susceptible toantibiotics, such as vancomycin or amoxicillin, had spread among LT patients. The findingof one particular clone that colonised 23 LT patients over a 15 months period was especiallystriking. During the remaining nine months of the study period this strain was not culturedagain from a LT patient. This indicates that the strain might have disappeared from thetransplantation wards. However, the end of transmission did not coincide with any changein antibiotic regimen, selective bowel decontamination or infection prevention on the LTwards. For five of the nine E. faecalis strains that were isolated from multiple patients, theepidemiology could be fully explained by the fact that patients carrying identical strainsstayed in the same wards during overlapping periods of time. These findings suggest thatthere was transmission of E. faecalis strains on a LT ward, and this might have been causedby carriage on the hands of personnel, direct contact between the patients or adhesion andsurvival on abiotic surfaces e.g. medical equipment. Only one of the epidemic isolatesdescribed in this study was possibly transmitted on the ICU. This shows that both on ICUsand on other wards hygienic precautions should be carefully followed to preventtransmission of bacteria between patients.
MO
LEC
ULA
R E
PID
EMIO
LOG
Y O
F E.
FA
ECA
LIS
IN L
IVER
TR
AN
SPLA
NTA
TIO
N
54
Table 1. Antimicrobial susceptibility of the E. faecalis isolates as determined by agar disk diffusion.
Epidemic isolates N (%) Non-epidemic isolates N (%)Antibiotic tested
Resistent Intermediate Sensitive Resistant Intermediate Sensitive
Gentamicina 43 (86) 7 (14) 1 (17) 5 (83)Erythromycin 43 (86) 7 (14) 1 (17) 2 (33) 3 (50)Tetracycline 39 (78) 7 (14) 4 (8) 3 (50) 3 (50)Ciprofloxacin 41 (82) 1 (2) 8 (16) 1 (17) 5 (83)Vancomycin 50 (100) 50 (100)Amoxicillin 50 (100) 50 (100)
The epidemic isolates are AFLP types A, B, G, H, I, J, K, M, and O (50 isolates), the non-epidemicisolates are AFLP type C, D, E, F, L, and N (six isolates).a high level resistance was tested

The genetic relatedness of E. faecalis strains was studied using AFLP. AFLP is a highthroughput typing method, providing a high level of discrimination between isolates and isapplicable without previous knowledge of genome sequences. AFLP has already proven itsvalue in molecular epidemiological studies of various pathogens including E. faecium (28,260, 261). In a previous study we applied AFLP to study the genetic relatedness of E.faecalis isolates recovered from faeces and clinical sites of liver transplant patients, otherpatients and volunteers. By AFLP we were able to discern different E. faecalis genogroups,where the strain within each group shared > 80 % of their restriction fragments. In thisstudy AFLP was used to identify epidemic clones. The cut-off point of 90 % similarity todefine identical isolates was chosen according to Willems et al. (260) who considered VREstrains to be epidemic if the AFLP pattern showed > 90 % similarity.Reports of transmission of E. faecalis susceptible to clinical important antibiotics likeamoxicillin and vancomycin are scarce as most reports focus on resistant enterococci.Sabria-Leal et al. (209) reported one E. faecalis strain, sensitive to clinically important
55
CHAPTER
3
Figure 2. Admission of the LT patients carrying AFLP type K E. faecalis to the different LT wards duringthe study period. Horizontal bars indicate the time of stay. Patients are numbered according to thedendrogram in Figure 1.

antibiotics, that was found in 11 out of 45 patient on an ICU as determined by PFGE.Information about the molecular epidemiology of E. faecalis still susceptible to clinicallyimportant antibiotics like amoxicillin and vancomycin, and not causing infections, isimportant to detect and prevent ‘silent’ transmission of isolates. As enterococci are able toacquire resistant determinants relatively easily, unrecognised transmission of susceptibleisolates may finally result in outbreaks of multi-resistant strains. An example illustrating thiswas reported recently in which a VRE outbreak in a hospital in Poland originated from avancomycin-susceptible strain that was endemic in the hospital, and that acquired the vanBtransposon through horizontal transmission (129).The incidence of colonization with E. faecalis in our study was 72 % (43 of 60 patients),which is relatively high compared to the incidence of 2.4 % found among all patientsadmitted to a University Hospital in eastern France (166). This difference might beexplained by the fact that most of the risk factors for nosocomial acquisition of multi-resistant enterococci are present in the LT patients in our study. All patients have peri-operative antibiotic prophylaxis, previous surgery and most patients have longerhospitalisation and the need for advanced nursing care, factors frequently described as riskfactors for acquisition of multi-resistant enterococci (131). Although the isolates we foundwere not multi-resistant, these risk factors might also apply to their acquisition. This issupported by the fact that the epidemic strain K was resistant to ciprofloxacin, one of thedrugs used as prophylaxis during the LT. Conversely, data regarding colonization are oftendifficult to compare because culturing methods are very different. In this study we focusedon the epidemiology of E. faecalis, therefore we cultured only weekly for four weeks.However to study the exact dynamics of colonization with E. faecalis, longer and morefrequent culturing is necessary.Epidemic isolates were cultured less frequently from clinical sites compared with non-epidemic isolates, which indicates that although these epidemic isolates might be capable ofspreading, they are not more pathogenic. Previously, we reported that isolates of E. faecalisfrom LT patients, blood cultures and the faeces of healthy volunteers were geneticallydifferent and could be subdivided into different AFLP genogroups with > 80 % sharedrestriction fragments (chapter 2). The disclosure of different genogroups for isolates causinginfections (e.g. blood culture isolates) and isolates from LT patients suggested the existenceof genetically distinct pathogenic subpopulations. Our present finding that epidemicisolates from LT patients are not frequently isolated from clinical sites is in agreement withthis previous study.Adhesion of bacteria to biomaterial surfaces plays an important role in the initiation ofinfections and spread of bacteria (22, 45). Better adhesion to abiotic surfaces might explainthe spread of E. faecalis, such as epidemic strain K in this study. We reported previously thatE. faecalis strains expressing enterococcal surface protein or aggregation substance, surfaceproteins that are associated with infection and spread, adhere better to biomaterialscompared to E. faecalis strains not expressing these surface proteins (chapter 5). Usingpolymerase chain reaction (PCR) we demonstrated the presence of the esp gene in theepidemic strain K. Furthermore, we determined that this strain was able to adhere to siliconerubber in comparable high numbers to other esp positive E. faecalis strains (data notshown). These data suggest that the observed high adherence to biomaterial surfaces
MO
LEC
ULA
R E
PID
EMIO
LOG
Y O
F E.
FA
ECA
LIS
IN L
IVER
TR
AN
SPLA
NTA
TIO
N
56

facilitated the epidemic spread of this strain. We have also reported that growth in bilesignificantly increased adhesion to biomaterial surfaces of esp positive strains (chapter 7).This underscores the possible role of esp of strain K in transmission as many LT patients havea bile drain for prolonged time after LT.In summary, this study showed the epidemic spread of E. faecalis isolates susceptible toclinically important antibiotics at the LT wards of a university hospital. The spread of onestrain among 23 patients during 15 months was especially interesting. The epidemic isolateswere not more frequently isolated from clinical sites compared with the non-epidemicisolates. However, detection and prevention of epidemic spread of susceptible enterococcalstrains is important, as it could be an indicator of and may precede transmission of, multi-resistant enterococcal strains.
We thank Kor Kooi for performing the antibiotic susceptibility testing and Jan Arends forhelpful discussion and reproduction of Figure 2 of this article. 57
CHAPTER
3


4
CHAPTER
Fluorescent in Situ Hybridization with Specific DNA Probes offers
Adequate Detection of Enterococcus faecalis andEnterococcus faecium in
Clinical Samples
Karola Waar, John E. Degener, Marja J. van Luyn, and Hermie J.M. Harmsen
Submitted

Abstract
Enterococcus faecalis and Enterococcus faecium are among the leading causes of hospitalacquired infections. Reliable and quick identification of E. faecalis and E. faecium isimportant for accurate treatment and understanding their role in the pathogenesis ofinfections. Fluorescent in situ hybridization (FISH) of whole bacterial cells with oligo-nucleotides targeted at the 16S rRNA molecule leads to a reduction of time to identification.Furthermore, the abundance, spatial distribution and bacterial cell morphology can beobserved in situ. We describe the design of two fluorescent labelled oligonucleotides that respectively detect the 16S rRNA of E. faecalis and the combination of E. faecium,Enterococcus hirae, Enterococcus mundtii and Enterococcus saccharolyticus. Differentprotocols for the application of these oligonucleotides with FISH in clinical samples likefaeces or blood cultures are given. We were also able to visualize enterococci in a biofilmattached to a biomaterial. Embedding of the biomaterial optimal preserved the morphologyand the architecture of the biofilm could be observed. The studies that use FISH fordetection of enterococci are generally hampered by the fact that they are only focused onone material and one protocol to detect the enterococci. However, the results of this studyshow that the probes can be used both in the routine laboratory to detect and determinethe enterococcal species in different clinical samples and in a research setting to enumerateand detect the enterococci in their physical environment.
DET
ECTI
ON
OF
E. F
AEC
ALI
SA
ND
E. F
AEC
IUM
WIT
H F
ISH
60

Introduction
Enterococcus faecalis and Enterococcus faecium, which are considered part of the normalintestinal flora, are among the leading causes of nosocomial infection. E. faecalis accountsfor 80-90 % and E. faecium for 5-10 % of clinical enterococcal isolates in human infection(159). Treatment of these infections can be difficult due to emerging antimicrobialresistance (113, 170).Reliable identification of enterococci, especially E. faecalis and E. faecium, is important foraccurate diagnosis and treatment. Current methods of identification are based on pheno-typic characteristics which may show variation on testing, are difficult to perform and oftentake up to several days for a final result (73). In recent years, authors have describedmolecular methods for the detection and identification of Enterococcus spp. by the use oflabelled oligonucleotide probes based on both 16S and 23S rRNA genes (11, 12, 16, 161,198). However, most of these studies use direct DNA hybridization techniques for theidentification of enterococci. Only few studies describe whole cell in situ hybridization foridentification and enumeration of enterococci (12, 105). Fluorescent in situ hybridization(FISH) of whole bacterial cells with oligonucleotides that detect the 16S rRNA molecule hasthe advantage that bacteria do not need to be cultured before detection and this would leadto a reduction of time to identification of the infecting organism (121). Also, differentstudies showed that only part of the bacteria in a specimen can be cultured because thebacteria have entered a nonculturable state, the culture conditions are not suitable or thepatient has already been treated with antibiotics (2).The studies that use FISH for detection of enterococci that are published until now arehampered by the fact that the specificity of the probes is not tested in different mixedbacterial populations or that they are only focused on one specific type of material (12, 105,121). In this study, we describe the design of two fluorescent labelled oligonucleotides that specifically detect the 16S rRNA of E. faecalis and the combination of E. faecium,Enterococcus hirae, Enterococcus. mundtii and Enterococcus saccharolyticus. Furthermore,we describe different protocols for the application of these oligonucleotides in FISH ondifferent clinical samples.
Materials and Methods
Culture strains and media
All reference strains used in this study are listed in Table 1. The strains were obtained fromdifferent sources as indicated in the table: DSM(Z) is Deutsche Sammlung von Mikro-organismen und Zellkulturen (Braunsweig, Germany), ATCC is the American Type CultureCollection (Rockville, Md.), and MMB is the Laboratory for Medical Microbiology(Groningen, The Netherlands). Clinical isolates of E. faecalis and E. faecium were obtainedfrom various clinical samples of different patients staying at the University HospitalGroningen, The Netherlands. DSM or ATCC strains were cultivated on the media asdescribed in the respective catalogues. All anaerobic strains were cultivated in anoxic
61
CHAPTER
4

peptone-yeast extract-glucose (PYG) medium (109) under anaerobic conditions at 37 °C.Facultative anaerobes were cultured on 5 % sheep blood agar (Oxoid) and one colony wasinoculated in brain heart infusion medium (Oxoid) at 37 °C. All MMB strains are clinical orhuman faecal isolates from local and regional public health laboratories and have beenidentified by routine procedures.
DET
ECTI
ON
OF
E. F
AEC
ALI
SA
ND
E. F
AEC
IUM
WIT
H F
ISH
62
Table 1. Reference organisms, sources, and FISH results for the probes Enfl84 and Enfm93.
Species SourceReaction with probe
Enfl84 Enfm93
Enterococcus faecalis 15 clinical isolates + –Enterococcus faecium 15 clinical isolates – +
Enterococcus faecalis DSMa 20478 + –Enterococcus faecium DSM 20477 – +Enterococcus hirae DSM 20160 – +Enterococcus mundtii DSM 4838 – +Enterococcus saccharolyticus DSM 20726 – +Enterococcus avium DSM 20679 – –Enterococcus casseliflavus DSM 20680 – –Enterococcus dispar DSM 6630 – –Enterococcus durans DSM 20633 – –Enterococcus gallinarum DSM 20628 – –Enterococcus raffinosus DSM 5633 – –
Bacteroides thetaiotaomicron MMBb – –Bifidobacterium pseudolongum MMB – –Clostridium perfringens MMB – –Eubacterium contortum DSM 3982 – –Eubacterium cylindroids MMB – –Lactobacillus acidophilus DSM 2007 – –Lactobacillus reuteri DSM 20016 – –Lactococcus lactis sub. cremoris DSM 20069 – –Peptostreptococcus asaccharolyticus MMB – –
Streptococcus agalactiae MMB – –Streptococcus bovis MMB – –Streptococcus intermedius DSM 20573 – –Streptococcus mitis MMB – –Streptococcus mutans DSM 20523 – –Streptococcus oralis DSM 20627 – –Streptococcus salivarius MMB – –Streptococcus sanguis DSM 20567 – –
Escherichia coli ATCCc 25922 – –aDSM, Deutsche Sammlung von Mikroorganismen und Zellkulturen.bMMB, Laboratory for Medical Microbiology Groningen.cATCC, American Type Culture Collection.

Design and testing of DNA probes
Oligonucleotide probes were designed with the ARB software package (145) and rRNAsequences were obtained in an aligned form from the Ribosomal Database Project (RDP)(147) supplemented with newly deposited rRNA sequences from GenBank. Fluoresceinlabelled oligonucleotides against selected specific target sequences of E. faecalis and E.faecium were synthesised commercially (Eurogentec) and tested for specificity against a setof reference organisms listed in Table 1. All strains were cultured in the appropriate fluidmedium until log-phase; this medium was diluted 1:50 in PBS (8 g of NaCl, 0.2 g of KCl,1.44 g of Na2HPO4, and 0.24 g of KH2PO4 each per liter). From this cell suspension, 10-20µl was smeared on a glass slide and air-dried at room temperature (RT). The cells on theglass slides were fixed for 5 minutes in ethanol (96 %) and again air-dried. To allowpermeabilization of their cell membrane, the Gram-positive strains were treated withenzymes prior to hybridization. The other strains were hybridized immediately after fixation.The permeabilization buffer consisted of a fresh mixture of 100 mM Tris-HCl pH 7.5 and 50 mM EDTA. For the permeabilization of Streptococcus spp., Enterococcus spp., Bifido-bacterium spp., Eubacterium spp. and Peptostreptococcus spp. permeabilization bufferwith 1 mg ml-1 lysozyme (Boehringer) was applied for 30 min at 37 ºC. For the perme-abilization of Clostridium spp., Lactobacillus spp. and Lactococcus spp. permeabilizationbuffer with 5 mg lysozyme and 0.1 mg ml-1 pancreatic lipase (Sigma) was applied for 30 min at 37 ºC. To the cell smears, 20 µl of the enzyme mixture was added and incubatedfor 30 min at 37 ºC in a buffer-saturated chamber. After treatment the slides were shortlyrinsed with milli-Q water and air-dried. Subsequently, 10 µl of hybridization buffer (0.9 MNaCl, 20 mM Tris-Cl [pH 7.5] and 0.1 % [w/v] SDS) containing 10 ng µl-1 of FITC- or Cy3 labelled probe was added to the smears. The formamide concentration was optimizedby varying the percentage of it in the hybridization buffer. The smears were incubatedunder a coverslip at 50 ºC for 2 h in a buffer-saturated chamber. Finally, the slides werewashed in 50 ml wash solution (0.9 M NaCl, 20 mM Tris-Cl [pH 7.5]), air-dried andmounted in Vectashield (Vector Labs). In all experiments, the Eub338 probe was used as apositive control (1). The slides were evaluated with an Olympus BH2 epifluorescencemicroscope.
Detection of E. faecalis and E. faecium in clinical samples of faeces and blood cultures
The application of the probes on different faecal samples was tested by enumerating theenterococci in diluted fecal samples that yielded enterococci on routine culture. The faecesof healthy adult volunteers and 9 premature babies between 3 and 21 days old (average of10 days) and a gestational time between 32 weeks 1 day and 35 weeks (average 33weeks4 days) was tested. Portions (0.5 g) of each stool were fixed with paraformaldehyde (PFA)as described previously (82). The fixed solution was mixed 1:1 with 5 % (w/w) Tween 80,10 µl was smeared on a glass slide and air-dried at RT. Further treatment was performed asdescribed above with modifications as listed in Table 2. Positive blood cultures with Gram-positive cocci in chains on the gram stain were selectedfor the detection of E. faecalis and E. faecium. A total of 15 µl from a positive blood culture
63
CHAPTER
4

was smeared onto a glass slide and air-dried at RT. Further treatment was performed asdescribed above with modifications as listed in Table 2.
Detection of E. faecalis and E. faecium in biofilms on bile drains
The drains were cut into pieces of 0.5 cm and fixed in 4 % (w/v) PFA for 48 h at 4 ºC andsubsequently stored in 50 % ethanol/PBS (v/v) at 4 ºC. The drains were rinsed in distilledwater, dehydrated in graded ethanol (50, 70, 96 and 100 %) and embedded in Technovit7100 hydroxyethylmethacrylate (Kulzer Histo-Technik, Klinipath). Cross sections of 10 µmwere cut and hybridized as describe above with modifications as listed in Table 2. Results ofthe hybridization were confirmed by routine culture.
Results
Probe design and specificity
To design oligonucleotide probes that specifically detect E. faecalis and E. faecium,alignments of 16S rRNA sequences retrieved from the RDP were analyzed and target sideswere identified that are present in E. faecalis or in E. faecium. In Table 3 the sequences ofthe Enfl84 and Enfm93 probe are given. In Table 1 the results of the FISH of probes Enfl84 and Enfm93 with different clinicalenterococcal isolates and reference organisms are shown. As reference organisms we chosestreptococci because they look very similar to enterococci on Gram-stain and other relevantorganisms that are frequently found in the faeces to confirm that the enterococci could bedistinguished in this complex environment. All microorganisms hybridized with the Eub338probe. The Enfl84 probe only hybridized with E. faecalis species. The Enfm93 probe alsohybridized with E. hirae, E. mundtii and E. saccharolyticus. Table 3 shows the selectedoligonucleotide sequences of Enfl84 and Enfm93, their target sites and alignments of thecorresponding sites of the enterococci that were used as reference organism. Enfl84 has aminimum of 8 mismatches with the enterococci used as reference; this explains the high
DET
ECTI
ON
OF
E. F
AEC
ALI
SA
ND
E. F
AEC
IUM
WIT
H F
ISH
64
Table 2. Hybridization conditions on different samples for the detection of E. faecalis and E. faecium
Bacterial culture Faeces Blood cultures Biofilms
Fixation 96 % ethanol 4 % PFA 96 % ethanol 4 % PFA(5’, RT) (overnight, 4 ºC) (5’, RT) (48 h, 4 °C)
96 % ethanol 50 % ethanol(10’, RT) (4 °C)
Preparation glass slide glass slide glass side Technovit 7100air dried air dried air dried cross sections
Permeabilization lysozyme no lysozyme lysozyme(30’, 37 °C) (15’, 37 °C) (15’, 37 °C)
Formamide 10 % 10 % 10 % 10 %
Hybridization 50 °C, 2 h 50 °C, overnight 50 °C, 30’ 50 °C overnight

specificity of the probe. Enfm93 has a full match with E. hirae so it is also specific for thisstrain. The probe has only one weak T-G mismatch with E. mundtii and two G-T mismatchesat the extreme 5’ site of the target with E. saccharolyticus. The hybridization conditionsmight not have been stringent enough to prevent hybridization with these targets.
Optimization of hybridization protocols of the clinical samples
The faeces and blood culture samples were prepared and fixed for hybridization in differentways; therefore material specific optimized protocols were established. A summary of thespecific hybridization conditions is shown in Table 2.Hybridization of the faecal samples was optimal without prior permeabilization. Probably,the longer fixation in PFA already permeabilized the bacteria. The blood cultures werepermeabilized and hybridized shorter than the pure bacterial cultures, this was necessary forthe use in the routine laboratory where quick detection had the priority. The protocolresulted in less clear signals; however detection and identification was still possible.
Enumeration of enterococci in the faeces of healthy volunteers
In the faeces of the adult volunteers, only occasionally enterococci were detected by FISHwhich indicated that the enterococci were below the detection limit for enumeration.However, in the faecal samples of seven of the nine babies high numbers of enterococciwere found. From the total number of DAPI stained cells up to 41 % (average 8.4 %) was E. faecalis and up to 23 % (average 6.4 %) was E. faecium with a total number of up to 2.6 x 1010 cells g-1 wet weight (average 3.46 x109) for E. faecalis and up to 1.1 x 1010 cellsg-1 wet weight (average 1.71 x109) for E. faecium. Figure 1 shows the fluorescence of E. faecium hybridised with the Enfm83 probe labelled with FITC in a faecal suspension of apremature baby.
65
CHAPTER
4
Table 3. Oligonucleotide sequences of probes Enfl84 specific for E. faecalis and Enfm93 that hybridizes withE. faecium, E. hirae, E. mundtii and E. saccharolyticus, their target sites and alignments of the correspondingsites of the enterococci used as reference strains.
Enterococcal Accession Enfl84 Enfm93species number
E. faecalis AB012212E. faecium Y18294E. hirae AF061011E. mundtii AF061013E. saccharolyticus AF061004E. dispar AF061007E. durans Y18359E. avium Y18274E. casseliflavus Y18161E. gallinarum AF039898E. raffinosus Y18296

DET
ECTI
ON
OF
E. F
AEC
ALI
SA
ND
E. F
AEC
IUM
WIT
H F
ISH
66 Figure 1. Fluorescence (a) and phase contrast (b) micrograph of a fecal suspension hybridized with theprobe Enfm93 labelled with FITC. Comparison of the phase contrast and the fluorescence micrographsshows that not all bacteria hybridized with the probe which indicates that other bacteria besides E.faecium are present. Bar is 5 µm.
Figure 2. Fluorescence micrograph of a 5-10 µm section of a bile drain hybridized with probes Enfl84(Cy3 labelled) and Eub338 (FITC labelled). The micrograph shows that the biofilm contained a layer ofother bacteria that hybridised with the Eub338 probe and a microcolony of E. faecalis that hybridizedwith the Enfl84 probe. Bar is 5 µm.

Detection of enterococci in biofilms on indwelling devices
After a liver transplantation the bile is often diverted outside the body of the patient forprolonged times by a bile drain inserted into the bile duct. These drains often get colonizedwith enterococci; therefore we selected these materials for the detection of enterococci in biofilms attached to biomaterials. Figure 2 shows a micrograph of a bile drain coveredwith a thick layer of bacteria that hybridize with the FITC labelled Eub338 probe with a microcolony of E. faecalis on top of it that also hybridizes with the Cy3 labelled Enfl84 probe. Routine culture of this drain contained E. faecalis, Klebsiella pneumoniae,Klebsiella oxytoca and Escherichia coli. The presence of high numbers of Entero-bacteriaceae in this biofilm was confirmed by FISH with a DNA probe that hybridizes withEnterobacteriaceae (197).
Discussion
In this study we have described the application of two DNA probes one for E. faecalis andone for E. faecium E. hirae, E. mundtii and E. saccharolyticus in various clinical materials.The results show that the probes can be used both in the routine laboratory to easily andquickly detect and determine the enterococcal species and in a research setting toenumerate and detect the enterococci in their physical environment and study the spatialdistribution in situ.The Enfl84 probe is specific only for E. faecalis and the Enfm93 probe hybridized with E.faecium, E. hirae, E. mundtii and E. saccharolyticus. Hybridization with these enterococcicould be explained by the presence of few or no mismatches (Table 3). E. hirae and E.mundtii have been isolated from human sources but only rarely and mostly theseenterococcal species only colonize the human and are not isolated from blood cultures orother clinically important material (73, 94, 128, 206). E. saccharolyticus has not beenisolated from human sources (74). When using FISH with the Enfm93 probe on mixedbacterial populations, the most probable organism to give a positive signal is E. faecium butthe possibility of one of the other three enterococcal species should be accounted for.Different materials require varying FISH protocols, however in this study we show that onlyminor adjustments are necessary to make the protocol applicable for new materials. Theenterococci could be detected in faeces, one of the most complex bacterial populations ofthe human body; this implies that the probes also can be used on other clinical materialscontaining mixed bacterial populations like sputum or pus from abdominal abscesses.Positive blood cultures often contain a single bacterial species that can be identified easily,however this can take up to 24 or 48 hours and antibiotic treatment is necessaryimmediately. A study by Munson et al. showed that especially the notification of the Gram-stain of a positive blood culture influenced the antibiotic therapy; fewer interventionsappeared after the release of antimicrobial susceptibility data (168). As E. faecium isgenerally more resistant to antibiotics compared to E. faecalis, early discrimination betweenthese species can be important for the empirical therapy. The application of FISH on positiveblood cultures was previously reported and the enterococcal DNA probe used in this study
67
CHAPTER
4

only identified E. faecalis (121). However, analysis with the ARB software package of theoligonucleotide sequence revealed that the probe used in that study detected many morebacterial species that are not commonly found in blood cultures but may be a problemwhen the probe is applied to a mixed bacterial population. Another study by Beimfohr et al.described two probes specific for the detection of E. faecalis and E. faecium in milk (12).These probes were based on the 23S rRNA. The 23S rRNA database is not as extensive asthe 16S rRNA database. Therefore it might be difficult to use these probes in clinical samplescontaining a complex mixture of bacteria as e.g. faeces because the specificity of the probescan not be compared in sillico to many other bacteria.Enumeration of the enterococci in faeces revealed that the adult volunteers carried numbersunder the detection limit for enumeration, but in a group of premature baby’s high numberswere found. This showed that FISH is a suitable tool to study the enterococcal content ofthe faeces and the development of early bacterial gastro-intestinal colonization in this agegroup. Furthermore, many hospitalized patients use antibiotics with little or no anti-enterococcal activity, this might increase the number of enterococci in their faeces andtherefore this method will also be useful for samples from this group of patients.Interestingly, we were able to detect enterococci with the FISH in a biofilm attached to anexplanted biomaterial. Embedding the material in Technovit 7100 preserved the morpho-logy very well and we were able to observe the architecture of the biofilm. These biofilmsare a frequent source of persistent infections and many studies are conducted to prevent theformation of these biofilms (45, 188). Recently it was reported that bacterial biofilms play arole in orthopaedic implant failure even if the routine culture did not reveal a pathogenicbacterium and it was suggested that microscopic analysis of the surface of the explantedprosthesis could improve the detection of implant infections (181). With this protocol weprovide a strong tool to detect bacteria in the biofilm and to observe its in vivo composition.Direct observation of the distribution of enterococci in biofilms might lead to better insightin their development and the discovery of ways to prevent their formation on indwellingbiomaterials.In other studies the 16S rRNA based probe technique has been used to identify differententerococcal species (11, 161, 190, 198). Only few however combined the advantage ofdirect visualization of the enterococci by FISH and the identification through the 16S or 23SrRNA (12, 105, 121). The studies that use the FISH for detection of enterococci arehampered by the fact that they are only focused on one material and therefore one protocolto detect the enterococci. In this study, we describe two DNA probes that can be used forthe clinically important enterococci E. faecalis and E. faecium and the hybridizationprotocols on three different materials. Therefore it can be concluded that these probes areideal for the detection of enterococci in various clinical materials in a microbial laboratorysetting where research and routine diagnostics are combined.
We would like to thank Marga Wester, Nelleke van Tilburg, Margriet Postma and LindaBrouwer for excellent technical assistance. We would like to thank Aad van der Berg and ElsHaagsma for collecting the bile drains.
DET
ECTI
ON
OF
E. F
AEC
ALI
SA
ND
E. F
AEC
IUM
WIT
H F
ISH
68

PART IIEnterococcuss faecalis in Biomaterial
related Infections


5
CHAPTER
Enterococcus faecalis Surface Proteinsdetermine its Adhesion Mechanism to
Bile Drain Materials
Karola Waar, Henny C. van der Mei, Hermie J. M. Harmsen, John E. Degener andHenk J. Busscher
Microbiology 2002; 148; 1863-1870

Abstract
An important step in infections associated with biliary drains is adhesion of microorganismsto the surface. In this study the role of three surface proteins of Enterococcus faecalis(enterococcal surface protein, aggregation substances 1 and 373) in the adhesion to siliconerubber, fluoro-ethylene-propylene and polyethylene was examined. Four isogenic E.faecalis strains with and without aggregation substances and one strain expressingenterococcal surface protein were used. The kinetics of enterococcal adhesion to thematerials was measured in situ in a parallel plate flow chamber. Initial deposition rates weresimilar for all strains, whereas the presence of surface proteins increased the total numberof adhering bacteria. Nearest neighbour analysis demonstrated that enterococci expressingthe whole sex-pheromone plasmid encoding aggregation substances 1 or 373 adhered inhigher numbers through mechanisms of positive cooperativity, which means that adhesionof bacteria enhances the probability of adhesion of other bacteria near these bacteria.Enterococci with the enterococcal surface protein did not adhere through this mechanism.These findings indicate that the surface proteins of E. faecalis play a key role in the adhesionto bile drains and bile drain associated infections.
SUR
FAC
E PR
OTE
INTS
OF
E. F
AEC
ALI
SA
ND
AD
HES
ION
72

Introduction
After an operation involving the common bile duct (CBD), for instance a livertransplantation, the bile is often diverted for prolonged times using a bile drain (T-tube).Infections are a frequent complication of T-tubing of the CBD (270). Normally, the biliarytract is sterile but bacterial colonization occurs in up to 75 % of patients within severalweeks after insertion of an in-dwelling foreign body (270). Especially in immune-compromised liver transplant patients the bile may be drained with a T-tube for up to threemonths and infections pose a major problem (13). In animal experiments, T-tubing of theCBD resulted invariably in contamination of bile by intestinal bacteria within 6 weeks (130).After insertion of the bile drain in the CBD, bile-colonizing bacteria may form a biofilm thatcan be a source of persistent infection, because the biofilm mode of growth protects theadhering bacteria against the host defence and the action of antimicrobial agents (45). Theinitial step in the formation of a microbial biofilm is the transport and adhesion ofmicroorganisms to the surface of the in-dwelling foreign body. This adhesion is oftendetermined by the presence of a host-derived coating of the foreign body and differentbacterial cell surface structures, like fimbriae, fibrillae or specific surface proteins (3, 22, 30).In a physico-chemical approach, these specific structural and chemical cell surfaceproperties are included in overall cell surface properties, as hydrophobicity and zetapotential (3, 22, 30). Insight into the involvement of these bacterial surface properties in theadhesion to biomaterials, as an important initial step in biofilm formation may lead tostrategies to reduce or prevent biomaterial-centred infections.Enterococcus faecalis is one of the Gram-positive microorganisms commonly found ininfections associated with foreign bodies in the bile duct (13, 160, 271). Due to theemerging antibiotic resistance of this microorganism, these infections are often difficult toprevent or treat (169). Some surface proteins of E. faecalis are found more frequently inenterococci causing infections and are therefore reported to be associated with enterococcaladhesion and infection (15, 44, 123, 217). One of these proteins, aggregation substance(Agg), is encoded by sex-pheromone-responsive plasmids. Agg is expressed in response topeptide pheromone induction as a sticky ‘hair-like’ structure on the cell wall (89) resultingin formation of aggregates of donor and recipient bacteria and transfer of plasmids (41). Allsex-pheromone plasmids, except pAM373, contain a homologous DNA region thatencodes the Agg (e.g. Asa1 encoded on pAD1). The Agg encoded on plasmid pAM373(Asa373) does not fit the overall homology as no similarities could be detected by Southernor Western blots using Asa1-specific DNA probes or antibodies (88, 175). Finally, the DNAsequence of asa373 that was determined proved to be totally different from the DNAsequence of the asa1 gene (50). The deduced amino-acid sequence of Agg contains twotripeptide arginine-glycine-aspartic acid (RGD) sequences (87). Since RGD is the cellattachment site of a large number of adhesive extracellular matrix, blood, and cell surfaceproteins (207), it has been suggested that Agg might play a role in the adherence to, andthe colonization of, host tissues by E. faecalis. This has been confirmed by different groups,showing that Agg is correlated with the binding of E. faecalis to renal tubular cells andintestinal epithelial cells (132, 185, 212).The second surface protein, enterococcal surface protein (Esp), was discovered in an E.
73
CHAPTER
5

faecalis strain that caused multiple infections within a hospital ward (217). Recently, it wasshown that the presence of the esp gene was highly associated with the capacity of E.faecalis to form a biofilm on a polystyrene surface (235). Also, the presence of Espcontributed to colonization and persistence of E. faecalis in the urinary tract the in an animalmodel of urinary tract infection (216). A variant esp gene was found in vancomycin-resistant Enterococcus faecium spreading in hospitals (260). The fact that surface proteinsAgg and Esp are associated with E. faecalis causing infections or epidemics suggests thatthese proteins may be associated with the ability of this microorganism to adhere to andsubsequently infect host tissue, or to spread within the hospital.The purpose of this study was to investigate whether surface proteins Agg (Asa1 andAsa373) encoded on sex-pheromone-responsive plasmids and Esp of E. faecalis influenceits adhesion to different bile drain materials. To this end, we compared the adhesion of fiveE. faecalis strains with different surface proteins to fluoro-ethylene-propylene (FEP),polyethylene (PE) and silicone rubber (SR) under flow conditions similar to those in a biledrain.
Materials and Methods
Strains and growth conditions
Four isogenic E. faecalis strains were used in this study: the plasmid-free strain OG1X (116);OG1X containing the sex-pheromone-responsive plasmid pAD1 encoding the Agg Asa1,with a positive regulator gene inserted that induces constitutive expression of this plasmid(denoted as OG1XE:pAD1, E indicates the positive regulator gene) (81, 173); OG1Xcontaining the plasmid pW-e-Asa1, a derivative of the Escherichia coli – Enterococcusfaecalis shuttle vector pWM401 which contains the asa1 gene and constitutively expressesAsa1 but no other proteins encoded on pAD1 nor any proteins that might be encoded onthe vector (personal communication with Albrecht B. Muscholl-Silberhorn, Thetis-IBN,Hamburg, Germany) (denoted as OG1X:pWeAsa1) (173) and OG1X containing theplasmid pAM373 which expresses Asa373 after induction with pheromones (denoted asOG1X:pAM373) (42). The four isogenic OG1X derived strains did not contain the esp geneas was confirmed by PCR. Expression of the Agg was checked by immunofluorescence withpolyclonal antibodies against Asa1 or Asa373. A similar level of Agg expression on all threeAgg positive strains was detected (data not shown). A. B. Muscholl-Silberhorn, kindlyprovided OG1X strains and antibodies. V. Shankar (University of Oklahoma Health SciencesCentre, Oklahoma City, USA) provided MMH594, the Esp expressing strain (217).MMH594 did not express Agg as was confirmed by immunofluorescence.The strains were streaked and grown overnight at 37 °C from a frozen stock on blood agarplates. The plate was then kept at 4 °C, but never longer than 4 weeks. Several colonieswere used to inoculate 3 ml of Todd-Hewitt broth (THB; Oxoid) that was incubated at 37°C in ambient air for 24 h. From this preculture, 2 ml was used to inoculate a second cultureof 200 ml THB that was grown for 18 h. E. faecalis OG1X:pWeAsa1 was grown in THB with20 µg erythromycin ml-1 and 20 µg chloramphenicol ml-1, and MMH594 was grown in THB
SUR
FAC
E PR
OTE
INTS
OF
E. F
AEC
ALI
SA
ND
AD
HES
ION
74

with 500 µg gentamicin ml-1. Bacteria from the second culture were harvested bycentrifugation at 10 000 g for 5 min at 10 °C and washed twice with demineralized water.Subsequently, bacteria were sonicated on ice for 2 x 10 s to separate cell clusters andcounted in a Bürker-Türk counting chamber. The cells were resuspended in phosphatebuffered saline (PBS) (10 mM potassiumphosphate and 0.15 M NaCl at pH 7) at aconcentration of 3 x 108 cells ml-1.E. faecalis JH2-2 excretes all known sex-pheromones of E. faecalis into the growth mediumand was used to collect pheromone (120). After 24 h growth at 37 °C in THB, the culturewas centrifuged at 10 000 g for 10 min at 10 °C and the supernatant containing thepheromones was autoclaved. To induce expression of the Agg in strain OG1X:pAM373,cells must be cultivated in the presence of pheromone, therefore the second culture of strainOG1X:pAM373 consisted of 100 ml fresh THB and 100 ml pheromone containing THBsupernatant.
Biomaterials
Implant-grade SR was obtained from Medin. Poly(tetrafluoroethylene-co-hexafluoro-propylene) (fluoro-ethylene-propylene, FEP) was supplied by Fluorplast and low density PEfrom Goodfellow. To ensure biomaterial surfaces were clean, they were sonicated for 3 minin a surfactant solution (2 % RBS 35 in water, Omniclean), rinsed thoroughly with waterand then washed with methanol and demineralized water before use.
Parallel plate flow chamber, image analysis and adhesion
The flow chamber (internal dimensions: l x w x h, 76 x 38 x 0.6 mm) and image analysissystem have been described in detail previously (31). Images were taken from the bottomplate (58 x 38 mm) of the parallel plate flow chamber. The Perspex bottom plate wascompletely covered with the material under study for FEP and PE. For SR a thin square (15x 15 mm) of the material was affixed centrally into the groove (15 x 15 mm) of a thicker(2.0 mm) Perspex plate. The depth of the groove was adapted to the thickness of thebiomaterial in such way that the silicone rubber surface was at the same height as thesurface of the Perspex plate. The top plate of the chamber was always made of glass. Theflow chamber was cleaned with Extran (Merck) and thoroughly rinsed with water anddemineralized water. Prior to each experiment, all tubes and the flow chamber were filledwith PBS, taking care to remove all air bubbles from the system. Once the system was filled,a bacterial suspension of 3 x 108 cells ml-1 in PBS was allowed to flow through the systemat a flow rate of 1.44 ml min-1, corresponding with a shear rate of 10.6 s-1. This shear rateis similar to the shear rate in a bile drain of 2 mm diameter at a bile production of 30 ml h-1. Deposition was observed with a CCD-MXRi camera (High Technology) mounted on aphase-contrast microscope (Olympus BH-2) equipped with a 40 x ultra-long-workingdistance lens (Olympus ULWD-CD Plan 40 PL). The camera was coupled to an imageanalyser (TEA, Difa). The bacterial suspension was perfused through the system for 4 h withre-circulation at room temperature, and images were taken at different time intervals andanalysed. All adhesion experiments were performed in triplicate with separately cultured
75
CHAPTER
5

bacteria. To exclude the influence of the growth in presence of the antibiotics erythromycinand chloramphenicol on the adhesion of strain OG1X:pWeAsa1, we tested the adhesion ofthis strain after the growth in medium without antibiotics and the results fell within therange found for adhesion after growth in the presence of antibiotics (data not shown).However, as the vector used is very unstable if the bacteria are grown without antibiotics,strain OG1X:pWeAsa1 was grown in the presence of antibiotics for better reproducibility.
Statistical analysis
Data were analysed with one-way analysis of variance followed by the Bonferonni t-test forpairwise multiple comparisons. The Kruskal-Wallis test followed by the Mann-Whitney testwere used for non-parametric data. Significance was defined as P ≤ 0.05.
Results
To measure the adhesion of E. faecalis strains to different bile drain materials, a bacterialsuspension was perfused through a flow chamber system for 4 h during which images weretaken from the material with adhering bacteria. The number of adhering bacteria in theimages were transformed to bacteria per unit area and least-square-fitted to an exponentialcurve. The number of bacteria adhering at stationary end point, n at t∞, could be estimatedfrom this exponential curve (SigmaPlot for Windows, version 5.00, SPSS), as well as acharacteristic time constant, �, describing the approach of the adhesion process to astationary state (49). The initial increase in the number of adhering bacteria over time wasexpressed in so-called initial deposition rate, j0, the increase in the number of adheringbacteria per unit area and time.
Initial deposition rates
In Table 1 the results of the flow chamber adhesion experiments are shown, including initialdeposition rates (j0), characteristic adhesion times (�) and number of bacteria at stationaryend point (n at t∞). The initial deposition rates (j0) did not differ for the E. faecalis strainswith or without surface proteins on FEP and PE. On SR, however, the initial deposition rateof strain MMH594 was significantly higher than of strain OG1X, while in addition the mean initial deposition rates, averaged for all strains, were significantly higher on FEP than on SR (P ≤ 0.05). The experimentally observed initial deposition rates are all in excessof the theoretical initial deposition rate (j0*) for deposition in the parallel plate flow chamberunder the current experimental conditions according to Von Smoluchowski-Levich, i.e. 692cm-2 s-1 (25). Consequently, the mean deposition efficiency (j0/j0*) of this collection ofenterococci amounts to 2.1 ± 0.4, which indicates that these strains have a high affinity forthe hydrophobic substrata used.
SUR
FAC
E PR
OTE
INTS
OF
E. F
AEC
ALI
SA
ND
AD
HES
ION
76

Number of bacteria adhering at the stationary end point
The number of bacteria at the stationary end point (n at t∞) in the flow chamber wassignificantly higher for E. faecalis with surface proteins compared to OG1X without surfaceproteins (Table 1). The mean total number of bacteria for all strains on FEP was significantlyhigher than on SR (P ≤ 0.05).
Characteristic adhesion time constant
The characteristic adhesion time constant � (= n∞ j0-1) describes the way in which the
adhesion process approaches a stationary state. For E. faecalis strains with surface proteins,� is higher than for OG1X without surface proteins and for most strains this difference issignificant, indicating that it takes longer for these bacteria to attain stationary stateadhesion. No significant difference could be detected between the different substrataaveraged over all strains.
Positive cooperativity
Positive cooperativity in microbial adhesion to surfaces is defined as the ability of oneadhering organism to stimulate the adhesion of other organisms in its immediate vicinity.Originally, positive cooperativity was indirectly inferred from Scatchard plots of bound overunbound organisms. However, the Scatchard analysis involves several assumptions that arenot always met for microbial adhesion (242). Sjollema & Busscher (1990) have pointed outthat positive cooperativity is directly reflected in the spatial arrangement of adheringorganisms over a substratum surface, provided the spatial arrangement is fully preserved asexisting during adhesion. Positive cooperativity is then concluded from high local relativedensities around a given adhering organism. The spatial arrangement of adheringenterococci over a substratum surface was analysed by radial pair distribution functions.Each adhering bacterium was taken once as a centre point and local densities of adheringenterococci were determined in circular shells at a distance r from this centre point. Theselocal densities were normalized with respect to the mean density of adhering bacteria,resulting in the radial pair distribution function g(r). When enterococci are randomly
77
CHAPTER
5
Table 1. Adhesion of E. faecalis strains to 3 different bile drain materials
Strain j0 (102 cm-2 s-1) � (103 s-1) n at t∞ (106 cm-2)
FEP PE SR FEP PE SR FEP PE SR
OG1X (Agg-, Esp-) 16 16 8 3.7 2.9 3.0 6.1 4.9 2.6OG1XE:pAD1 (Agg+) 17 16 10 8.3* 5.0 11.1* 14.0* 8.0* 11.2*OG1X:pWeAsa1 (Agg+) 17 16 12 8.3* 8.3* 6.7* 14.9* 13.3* 7.8*OG1X:pAM373 (Agg+) 16 12 13 7.1* 7.1* 6.3* 11.5* 8.3* 7.9*MMH594 (Esp+) 17 16 18* 5.9 7.7* 3.8 10.3* 12.6* 7.1*
Numbers show the initial deposition rate (j0), characteristic adhesion time (�) and no. of bacteria at thestationary end point (n at t∞). Experiments performed in triplicate with separate bacterial culturesyielded a SD < 30 %. *P ≤ 0.05 versus OG1X.

distributed over the entire substratum surface, g(r) = 1. However, if there is preferentialadhesion at a given separation distance r between adhering bacteria, then g(r) > 1. Themaximum value of g(rp) indicates the preference for bacterial deposition near an alreadyadhering bacterium and is a direct measure for positive cooperativity. The distance at whichthis maximum occurs (rp) indicates the preferential adhesion distance of the bacteria. Themaximum value of g(rp) was calculated for 3 different fields of view after 4 h of flow.Figure 1 compares the spatial arrangements of strains OG1X, OG1XE:pAD1 and MMH594on FEP after 4 h of flow and in Table 2 the accessory values for g(rp) and rp are shown.Previously, for inert polystyrene particles g(rp) values ranging from 1.2 to 1.4 were reported,which provides a criterion for the absence of positive cooperativity (224). The strain used inthis study can be divided into three groups with regard to their strength of cooperativity. (1)E. faecalis strains OG1X and OG1X:pWeAsa1 expressing weak, possibly non-specificcooperativity, as the spatial arrangement of strain OG1X shows mainly singly adheringorganisms with some small aggregates (Figure 1a). (2) E. faecalis strains OG1XE:pAD1 andOG1X:pAM373 expressing strong cooperativity, i.e. on the substratum surface distinctaggregates are formed (Figure 1b), in line with an elevated value of g(rp) (see Table 2). (3)Strain MMH594 adhering exclusively as single cells (Figure 1c), i.e. g(rp) equals unity,indicating the absence of positive cooperativity (see Table 2).
Discussion
This study addressed the question whether different surface proteins on E. faecalis influenceits adhesion to bile drain materials. Surface proteins do not influence the initial deposition ofE. faecalis on different hydrophobic materials. However, the total number of bacteriaadhering at stationary end point was significantly higher for E. faecalis expressing surfaceproteins. The surface proteins enhance the total number of adhering bacteria via a differentmechanism. The expression of Agg (Asa1 and Asa373) and other gene products encoded
SUR
FAC
E PR
OTE
INTS
OF
E. F
AEC
ALI
SA
ND
AD
HES
ION
78
Figure 1. Spatial arrangements of E. faecalis after 4 h of flow on FEP for: OG1X (a), OG1XE:pAD1 (b),MMH594 (c). The images show the presence of small aggregates of E. faecalis on the surface forOG1X (a), the formation of distinct aggregates if the Agg is expressed (b) and the absence ofaggregates if Esp is expressed (c). The distribution of OG1X:pWeAsa1 was very similar to OG1X, whilethe distribution of OG1X:pAM373 was very similar to OG1XE:pAD1; therefore, these images are notshown. Bar, 15 µm.

on the sex-pheromone plasmids, but not solely Agg (Asa1), invokes positive cooperativity.This means that adhering individual enterococci stimulate adhesion of more enterococci inthe more advanced stages of the adhesion process, whereas the expression of Esp or solelyAgg without the sex-pheromone plasmid enhances the number of adhering enterococcithrough direct interactions with the material and not via positive cooperativity. The initial deposition rates reflect the direct interaction between the microorganism and thesubstratum surface, without influences of already adhering bacteria (22). The initial contactbetween the bacterium and the substratum is determined by the presence of hydrophobicor electrostatic groups. When surface proteins are only scarcely present on the bacterial cellsurface, these physico-chemical interaction forces will be influenced little by the presence ofthe surface proteins. Whereas the initial deposition rates vary little among the differentstrains, deposition rates of enterococci on FEP were higher than on PE and SR. Thisdifference is likely to be a result of a combination of the high surface hydrophobicity of FEPand a different chemical composition. FEP is a very inert material with only fluorine groupson its surface in contrast to e.g. SR with methyl groups on its surface (3).In contrast to the initial deposition, the total number of bacteria at the stationary end pointwas increased by the presence of surface proteins on E. faecalis. The number of bacteria atthe a stationary end point is influenced not only by fundamental interaction forcesoriginating from the substratum surface, but also by more specific interactions between thebacteria. Positive cooperativity is one of the mechanisms that might play a role in theincrease in total number of bacteria if surface proteins are present. Adhesion of bacteriagives rise to new adhesion sites or diminishes the influence of antagonistic sites, and newlydepositing bacteria will adhere preferably in the neighbourhood of already adhering ones.As a corollary of positive cooperativity, organisms adhere close to each other and appear notrandomly distributed. Positive cooperativity on saliva-coated substrata has been explainedpredominantly by the capacity of adhering cells to induce conformational changes in thepellicle or on the surfaces of approaching cells (58). However, positive cooperative pheno-mena have also been described for oral streptococci adhering to inert substrata (242).Proposed explanations are based on the hypothesis that the magnitude of the interactionforces between adhering cells is higher than the interaction forces between adhering cellsand their substrata or between cells in suspension (56, 242).
79
CHAPTER
5
Table 2. Degree of positive cooperativity involved in the adhesion of E. faecalis strains to FEP, PE andSR, as derived from radial distribution functions.
g(rp) rp (µm)Strain FEP PE SR FEP PE SR
OG1X (Agg-, Esp-) 1.4 1.8 2.3 1.2 1.2 1.3OG1XE:pAD1 (Agg+) 3.2* 2.7* 2.8 1.8* 1.7 1.8OG1X:pWeAsa1 (Agg+) 1.7 1.5 2.2 1.8* 1.9* 1.5OG1X:pAM373 (Agg+) 2.0 2.7* 2.6 1.5* 1.3 1.5MMH594 (Esp+) 1.0 1.0* 1.1* 2.8* 2.6* 2.3*
Positive cooperativity is concluded from g(rp) values in excess of 1.4 and rp indicates the preferentialadhesion distance. Experiments performed in triplicate with separate bacterial cultures yielded a SD < 30 %. *P ≤ 0.05 versus OG1X.

A model for the adhesion of the different E. faecalis strains to an inert substratum surface,accounting for the role of surface proteins, is proposed in Figure 2. The model shows threedifferent mechanisms of adhesion to the substratum and interactions between bacteria, asfound for the different strains in this study. Strain OG1X, without any surface proteins,shows a weak cooperativity between adhering enterococci that might be due to non-specific interactions (Figure 2a). The E. faecalis strains expressing the sex-pheromoneplasmids (OG1XE:pAD1 and OG1X:pAM373) adhere in high numbers to the substratumdue to specific interaction between the bacteria (positive cooperativity) (Figure 2b). Thishigh cooperativity between sex-pheromone-plasmid-expressing bacteria might beexplained by the fact that the plasmid plays a role in contact between bacteria for its owntransfer (265). The Esp-positive E. faecalis strain (MMH594) adheres in high numbers to thesubstratum, but does not show any interaction between the bacteria (Figure 2c).
SUR
FAC
E PR
OTE
INTS
OF
E. F
AEC
ALI
SA
ND
AD
HES
ION
80
Figure 2. Proposed mechanism of the adhesion of E. faecalis with different surface proteins tohydrophobic substrata. (a) Strain OG1X without surface proteins shows weak cooperativity, possiblydue to non-specific interactions and adheres in low numbers, (b) E. faecalis strains expressing the sex-pheromone plasmids (OG1XE:pAD1 and OG1X:pAM373) adhere in high numbers to thesubstratum due to specific interaction between the bacteria (positive cooperativity), (c) Esp positive E.faecalis strain MMH594 does not show any interaction between the bacteria, but yet adheres in highnumbers to the substratum as a result of strong interaction forces with the substratum due to thepresence of Esp on the surface.

Consequently, the interaction force with the substratum of this strain must be strong,possibly due to the presence of Esp on the surface. This result is confirmed by a recent study(235) in which it was shown that Esp increases the number of bacteria adhering to abioticsurfaces by comparing the adhesion with Esp-deficient mutants. The strain OG1X:pWeAsa1expressing Asa1 but not the sex-pheromone plasmid, does not fit very well into this model.This strain adheres in high numbers, although its cooperativity is similar to that of strainOG1X. Both strain OG1X:pWeAsa1 as well as the strains that express the whole sex-phero-mone plasmid showed high numbers of adhering bacteria at stationary end point; this mightindicate that not the whole sex-pheromone plasmid, but only the Agg is necessary to reachhigh numbers of adhering bacteria. On the other hand, the lower cooperativity of strainOG1X:pWeAsa1 compared to the strains with the whole sex-pheromone plasmid indicatesthat other factors than Agg encoded on the sex-pheromone plasmid play a role in positivecooperativity. The difference in cooperativity could also be explained by differences in theexpression level of Agg between OG1X:pWeAsa1 and OG1XE:pAD1 since the promotorconstructed in OG1X:pWeAsa1 is weaker than the promotor constructed in OG1XE:pAD1(Albrecht B. Muscholl-Silberhorn, personal communication). However, a similar level ofexpression was seen in the immunofluorescence assay, which indicates that the differencein expression might not be very high. The sex-pheromone plasmid transfer system is verycomplicated with many factors involved in its control. The functions of some genes on theplasmids are still not completely understood (50, 66, 80, 265). The question of which ofthese factors is involved in positive cooperativity on the surface is intriguing and still needsto be answered more fully.The surface proteins described in this paper are possible virulence factors associated withinfections in humans. However, the exact role of these proteins in the pathogenesis ofinfections is still not known. Many of the infections in hospitalised patient are associatedwith in-dwelling medical devices, especially bile drains. One way of initiating thesebiomaterial-centered infections is by adhesion to the device. In this study we found that thesurface proteins of E. faecalis are possible virulence factors because they enhance thenumber of bacteria adhering to bile drain materials. Higher total numbers of bacteriaadhering to in-dwelling medical devices may lead to more colonization and biomaterial-centered infections. In addition, the question might arise whether the surface proteinsstudied under in vitro conditions will be expressed in patients. Esp is always expressed onthe surface if the bacterium contains the esp gene, therefore it is likely to be expressed invivo (217). The in vivo expression of Agg is more difficult to describe. To our knowledgeuntil now no research has been published on the expression of Agg in bile or the humangut. However, it has been shown that factors other than pheromone (e.g. serum) caninduce expression of Agg in vivo (108, 132). Therefore, we think that it is reasonable toassume that Agg is expressed in vivo.In summary, enterococci expressing Esp or Agg adhere in higher numbers to hydrophobicbile drain materials. However, E. faecalis with different surface proteins adheres to thesubstratum through different mechanisms. Enterococci expressing the sex-pheromoneplasmid encoding the Asa1 or Asa373 adhere in high numbers through positivecooperativity between adhering bacteria. Enterococci positive for Esp also adhere in highnumbers, but do not utilise positive cooperative mechanisms in their adhesion to these
81
CHAPTER
5

materials. These findings indicate that interfering with the positive cooperativity betweenadhering enterococci, as resulting from the expression of sex-pheromone plasmids, orinterfering with the interaction between surface proteins and materials, might yield a wayto prevent bile drains associated infections.
We thank Albrecht Muscholl-Silberhorn for helpful discussion. We also thank Rudi Tonk andJoop de Vries for reproduction of the figures in this article.
SUR
FAC
E PR
OTE
INTS
OF
E. F
AEC
ALI
SA
ND
AD
HES
ION
82

6
CHAPTER
Atomic Force Microscopy study on Interaction Forces between
Enterococcus faecalis with and withoutAggregation Substance
Karola Waar, Henny C. van der Mei, Hermie J. M. Harmsen, Joop de Vries, Jelly Atema-Smit, John E. Degener and Henk J. Busscher
Submitted

Abstract
Enterococcus faecalis is becoming one of the leading causes of hospital acquired infectionsand especially indwelling medical devices are prone to enterococcal infection. E. faecalisexpressing Aggregation substance (Agg) adheres to biomaterials surfaces by means of so-called positive cooperativity, i.e. the ability of one adhering organism to stimulate theadhesion of other organisms in its immediate vicinity. In this study, Atomic Force Microscopy(AFM) was used to measure the interaction forces between E. faecalis with and withoutAgg. Bacteria were attached to both a substratum surface and a tip-less cantilever. Two E.faecalis strains expressing different forms of Agg showed nearly two-fold higher interactionforces between bacterial cells than a strain lacking Agg (adhesive force Fadh = -1.3 nN). Thestrong interaction forces between the strains with Agg could be reduced by adsorption ofantibodies against Agg from -2.6 nN and -2.3 nN to -1.2 nN and -1.3 nN, respectively.Comparison of the results of the AFM interaction forces with the positive cooperativity afteradhesion to a biomaterial in a parallel plate flow chamber showed that in the absence ofstrong interaction forces between the cells, either due to lack of Agg or due to adsorptionof antibodies against Agg, positive cooperativity was absent too. In conclusion, this is thefirst time that the influence of specific antibodies on interaction forces between E. faecalishas been demonstrated by AFM and that the interaction forces measured have a directimpact on the way these bacteria colonize a biomaterials surface.
AFM
STU
DY
ON
TH
E IN
TER
AC
TIO
N F
OR
CES
OF
E. F
AEC
ALI
S
84

Introduction
Enterococci are becoming one of the leading causes of hospital acquired infections (204),with Enterococcus faecalis accounting for up to 90 % of clinical enterococcal isolates (206).Indwelling medical devices are a frequent source of these enterococcal infections (53, 270).A bacterial biofilm attached to the indwelling device can be a source of persistent infection.Microbial adhesion and aggregation are crucial steps in biofilm formation and therefore inbiomaterial centered infections (45).Aggregation substance (Agg) is a plasmid encoded surface protein of E. faecalis associatedwith infection, aggregation and biofilm formation (123, chapter 2 and 5). Two differentforms of Agg are described, Asa1 and Asa373 (50, 87). Previously, we reported that E.faecalis expressing Agg adheres in significantly higher numbers to biomaterials compared toisogenic strains without Agg by studying their adhesion in a parallel plate flow chamber.This increase in adhesion was through specific interaction between the bacteria on thesurface mediated by the Agg (chapter 5). This specific interaction was expressed as thedegree of positive cooperativity, i.e. the ability of one adhering organism to stimulate theadhesion of other organisms in its immediate vicinity. Positive cooperativity is directlyreflected in the spatial arrangement of adhering organisms over a substratum surface and isconcluded from high local relative densities around a given adhering organism (224).Atomic force microscopy (AFM) is a surface imaging technique, which operates by sensingthe force between a very sharp probe attached to a flexible cantilever and the samplesurface (18). Recently, AFM has emerged as a powerful tool to measure molecular inter-action forces (61). AFM force measurements were further applied to microbial systemsmeasuring the interaction between bacteria and a substratum surface (26, 144, 202). Oneapproach to study these microbial interactions is to attach the bacteria directly onto theAFM probe or cantilever and study the interaction with the substratum. However, insight inthe process of microbial aggregation in biofilm formation would require the investigation ofthe direct interaction between two bacteria. If the bacteria are attached to both thecantilever and a substratum surface it is possible to study the interaction forces between thebacteria by AFM. Here, we report the use of AFM force measurements to study the interaction between E.faecalis strains expressing the Agg Asa1 or Asa373 by attaching the bacteria to both asubstratum surface and a tip-less cantilever. The results were compared with the positivecooperativity after adhesion in a parallel plate flow chamber to fluoro-ethylene-propylene(FEP), a frequently used biomaterial. The role of Agg in the interaction and biofilmformation was confirmed by incubating the bacteria with antibodies specific for Asa1 orAsa373 prior to both the AFM force measurements and adhesion in the parallel plate flowchamber. Insight into the mechanism of direct interaction between enterococci, the role ofAgg in this interaction and the influence of specific antibodies on this interaction might leadto ways to prevent biofilm formation of enterococci on indwelling medical devices.
85
CHAPTER
6

Materials and Methods
Bacterial strains, culture conditions and harvesting
Three isogenic E. faecalis strains were used in this study: the plasmid-free strain OG1X(116); OG1X containing the sex-pheromone-responsive plasmid pAD1 encoding the AggAsa1, with a positive regulator gene inserted which induces constitutive expression of thisplasmid (depicted as OG1XE:pAD1, E indicates the positive regulator gene) (173); andOG1X containing the plasmid pAM373 which expresses Asa373 after induction withpheromones (depicted as OG1X:pAM373) (42). Expression of the Agg was checked byimmune fluorescence with polyclonal antibodies against Asa1 or Asa373. A similar level ofAgg expression on both Agg positive strains was detected (data not shown). A. B.Muscholl-Silberhorn, Thetis-IBN, Hamburg, Germany, kindly provided OG1X strains andpolyclonal antiserum. The strains were streaked and grown overnight at 37 °C from a frozen stock on blood agarplates. Several colonies were used to inoculate 3 ml of Todd-Hewitt broth (Oxoid) that wasincubated at 37 °C in ambient air for 24 h. From this preculture, 2 ml was used to inoculatea second culture of 200 ml THB that was grown for 18 h. If necessary, bacteria from thesecond culture were incubated with polyclonal antiserum (1:600) for 30 min at 37 °C. In apilot study, different dilutions of serum were tested in the parallel plate chamber and a dose-effect relation was observed, from these experiments the dilution of 1:600 was chosen tobe used in this study. Bacteria were harvested by centrifugation at 10 000 x g for 5 min at10 °C and washed twice with demineralized water. Subsequently, bacteria were suspendedin PBS (10 mM potassiumphosphate and 0.15 M NaCl at pH 7) and sonicated on ice for 2x 10 s to separate cell clusters. For the parallel plate flow chamber experiments, bacteriawere counted in a Bürker-Türk counting chamber and diluted to a concentration of 3 x 108
cells ml-1.E. faecalis JH2-2 excretes all known sex-pheromones of E. faecalis into the growth mediumand was used to collect pheromone (120). After 24 h growth at 37 °C in THB, the culturewas centrifuged at 10 000 x g for 10 min at 10 °C and the supernatant containing thepheromones was autoclaved. To induce the expression of Agg in strain OG1X:pAM373,growth in the presence of pheromone is necessary, therefore the second culture of strainOG1X:pAM373 consisted of 100 ml fresh THB and 100 ml of pheromone containing THBsupernatant.
Polyclonal antibodies
Purified Asa1 or Asa373 was used for the production of polyclonal antiserum. The genesencoding Asa1 or Asa373 were constructed in vector pQE30-32 (Qiagen), expressed inEscherichia coli cloning strain JM109 and purified as described by Muscholl-Silberhorn(174, 175). Eurogentec (Belgium) carried out immunization according to a standardizedprocedure (injections on day 0, 14, 28, 56 and bleeds on day 0, 38, 64). Prior toimmunization serum was tested for absence of cross-reactivity with aggregations substanceor unrelated proteins from E. faecalis and E. coli. The polyclonal antiserum was tested for
AFM
STU
DY
ON
TH
E IN
TER
AC
TIO
N F
OR
CES
OF
E. F
AEC
ALI
S
86

specificity for Asa1 or Asa373 using western blots with purified Agg and crude proteinpreparation of enterococci expressing different forms of Agg.
Atomic force microscopy
Bacteria were attached through electrostatic interactions (physical adsorption) to both aglass slide and a tip-less cantilever, made positively charged through adsorption of poly-L-lysine (19). In order to coat the glass surface with poly-L-lysine, the glass slide was cleanedby dipping in methanol and rinsing with demineralized water, after which a drop of 0.01 %(w/v) poly-L-lysine (Sigma) solution was added and spread over the surface. After air-drying of the glass slide, a few drops of the undiluted bacterial suspension in PBS wereadded. After 15 min, the bacteria-coated slide was rinsed with PBS to remove looselyattached bacteria and transferred to the AFM. The tip-less AFM cantilever (NP-0, Veeco)was dipped into a drop of 0.01 % (w/v) poly-L-lysine solution and allowed to dry;afterwards the cantilever was dipped into a drop of bacterial suspension and dried again.AFM force-distance measurements were made at room temperature under PBS using aDimension 3100 system (Nanoscope III Digital Instrument). “V”-shaped tip-less siliconnitride cantilevers from Veeco with a spring constant of 0.06 N m-1 were used. Individualforce curves were collected between the bacteria-coated AFM tip and the top of physicallyadsorbed bacteria on randomly selected locations with z-displacement of 1000-2000 nm atz-scan rates ≤ 1 Hz. The slope of the retraction force curves in the region where probe andsample are in contact were used to convert the voltage into cantilever deflection. Theconversion of deflection into force was carried out as has been previously described byothers (59). Approach curves were fitted to an exponential function. Retraction curvesgenerally showed multiple adhesion peaks and the magnitude of the peaks was recordedand averaged. Results represent the average of 2 separate runs with a total of 30 force-distant curves taken on 6 different bacteria (5 curves per bacterium). The results werefurther normalised with respect to the mean of both runs.
Parallel plate flow chamber, image analysis and adhesion
The flow chamber (internal dimensions: l x w x h = 76 x 38 x 0.6 mm) and image analysissystem have been described in detail previously (31). Poly(tetrafluoroethylene-co-hexafluoropropylene) (fluoro-ethylene-propylene, FEP) was obtained from Fluorplast.Images were taken from the Perspex bottom plate (58 x 38 mm) of the parallel plate flowchamber that was completely covered with FEP. Surfaces were sonicated for 3 min in asurfactant solution (2 % RBS 35 in water, Omniclean), rinsed thoroughly with water, andthen washed with methanol and demineralized water before use. The flow chamber wascleaned with Extran (Merck) and thoroughly rinsed with water and demineralized water.Prior to each experiment, all tubes and the flow chamber were filled with PBS, taking careto remove all air bubbles from the system. Once the system was filled, a bacterialsuspension of 3 x 108 cells ml-1 in PBS was allowed to flow through the system at a flow rateof 1.44 ml min-1 corresponding with a shear rate of 10.6 s-1. Deposition was observed witha CCD-MXRi camera (High Technology) mounted on a phase-contrast microscope
87
CHAPTER
6

(Olympus BH-2) equipped with a 40 x ultra-long-working distance lens (Olympus ULWD-CD Plan 40 PL). The camera was coupled to an image analyzer (TEA, Difa). The bacterialsuspension was perfused through the system for 4 h with re-circulation at roomtemperature and images were taken at different time intervals and analyzed. Adhesionexperiments were performed in triplicate with separate bacterial cultures, except OG1Xwith sera containing polyclonal antibodies that were performed in duplicate.
Results
The interaction forces measured between the E. faecalis cells are plotted as a function of theseparation distance in Figure 1. E. faecalis strain OG1X showed similar force-distance curveswith and without incubation with antibodies to Asa1 or Asa373, and therefore only thecurve without incubation is shown (Figure 1a). The force-distance curves of the E. faecalisstrains expressing Agg (Figure 1b and d) show high adhesion forces upon retraction over along distance. Comparing the force-distance curves of the E. faecalis strains expressing Agg(Figure 1b and d) with the force-distance curves after incubation with specific antibodiesreveals a remarkable decrease in adhesion upon retraction (Figure 1c and e). Approachcurves did not show significant differences between the E. faecalis strains and all showed arepulsive force upon contact of about 5.5 nN with a characteristic decay length of about 64nm.After adhesion to FEP in the parallel plate flow chamber, bacteria per unit area were least-square fitted to an exponential curve and the number of bacteria adhering at stationaryend-point (n at t∞) could be estimated from this exponential curve. Furthermore, the degreeof positive cooperativity between adhering bacteria was assessed by comparing the localdensities of bacterial adhesion around all individual bacteria with the average bacterialdensity over the substratum surface, yielding the radial pair distribution function g(r)(chapter 5). When enterococci are randomly distributed over the entire substratum surface,g(r) = 1. However, if there is preferential adhesion at a given separation distance r betweenadhering bacteria through positive cooperative mechanisms, then g(r) > 1.Table 1 summarizes the interaction forces between the E. faecalis cells (Fadh), their degree ofpositive cooperativity [g(rp)] and the average bacterial density on FEP at stationary end-point (n at t∞). The average interaction forces upon retraction (Fadh) are stronger for the E.faecalis strains expressing Agg (OG1XE:pAD1 and OG1X:pAM373) compared to the strainwithout Agg (OG1X). This is in line with the higher positive cooperativity measured for thestrains expressing Agg. The Fadh, and g(rp) values decreased remarkably for the E. faecalisstrains expressing Agg after incubation with the specific antiserum which indicates specificinterference of the antibodies with the interaction between the strains. Interestingly, thenumber of bacteria at stationary end point also decreased for the E. faecalis strainsexpressing Agg after incubation with polyclonal antibodies against the Agg. This decreasewas not seen for the OG1X strain after incubation with the antibodies which indicates thatthe decrease was due to specific interaction with the Agg.
AFM
STU
DY
ON
TH
E IN
TER
AC
TIO
N F
OR
CES
OF
E. F
AEC
ALI
S
88

89
CHAPTER
6
Figure 1. Representative examples of AFMforce-distance curves between E. faecalis strainson a silicone nitride tip-less cantilever and apositively-charged glass surface.(a) strain OG1X without Agg; (b) strainOG1XE:pAD1 expressing aggregation substanceAsa1; (c) strain OG1XE:pAD1 incubated withantibodies to Asa1; (d) strain OG1X:pAM373expressing aggregation substance Asa373 and(e) strain OG1X:pAM373 incubated withantibodies to Asa373. The arrows indicate thedirection of movement of the tips.

Discussion
Microbial aggregation and adhesion are crucial events in the formation of biofilms inmedicine and nature. Understanding the individual interaction force between aggregatingbacteria can give more insight in molecular basis of these phenomena. In this paper, weshow that the interaction forces between bacteria can be measured by use of the AFM forcemeasurements and attaching the bacteria to both a substratum surface and a tip-lesscantilever. The interaction forces between different strains of E. faecalis with and withoutsurface Agg and after incubation with antibodies to Agg were measured and comparedwith the positive cooperativity observed after their adhesion to a biomaterials surface. Aggis a surface protein of E. faecalis that enables close cell-cell aggregation between bacteriaand transfer of plasmids (65). Previously, we showed that Agg enhances the adhesion tobiomaterials through positive cooperativity, which is interaction between the bacteria on thebiomaterial surface. Comparing the adhesion force upon retraction measured with AFMand the positive cooperativity after adhesion in the parallel plate flow chamber showed agood correlation. Therefore it can be concluded that the interaction forces measured withthe AFM are relevant for the macroscopic colonization of biomaterials surfaces by E. faecaliscells and that antibodies to Agg can block this interaction.In recent years, AFM force measurements have made it possible to explore the physicalproperties of microbial cells (reviewed in 60). These studies generally used the approach ofcoating the bacteria either to the tip of the cantilever or the substratum and measure theinteraction with different materials. Alternatively, AFM probes were functionalized withbiomolecules in order to measure discrete receptor-ligand interactions at cell surfaces (107,237). However, in this study we describe the unique approach of coating both a tip-lesscantilever tip and a substratum surface with bacteria. This enables us to analyse the directinteraction between bacteria as during in vivo colonization of a surface.To confirm the role of Agg in the interaction between the E. faecalis strains, the bacteria
AFM
STU
DY
ON
TH
E IN
TER
AC
TIO
N F
OR
CES
OF
E. F
AEC
ALI
S
90
Table 1. Comparison of AFM interaction forces and adhesion characteristics of E. faecalis
Strain Pre-incubation Fadha g(rp)
b n at t∞c
OG1X no -1.3±0.5 1.4±0.1 6.1±0.5OG1X anti Asa1d -1.2±0.5 1.2±0.1 7.1± 0.2OG1X anti Asa373e -1.5±0.4 1.2±0.1 6.9±1.1
OG1XE:pAD1 no -2.6±0.5 3.2±0.8 14.0±1.1OG1XE:pAD1 anti Asa1 -1.2±0.4 1.9±0.1 10.5±0.8
OG1X:pAM373 no -2.3±0.6 2.0±0.4 11.5± 0.5OG1X:pAM373 anti Asa373 -1.3±0.4 1.2±0.1 10.4±0.7a Fadh, average adhesion force upon retraction (nN); b g(rp), degree of positive cooperativity; c n at t∞,number of bacteria at stationary end point (in 106 cm-2); d Asa1, aggregation substance 1; e Asa373,aggregation substance 373.Adhesion experiments were performed in triplicate with separate bacterial cultures, except OG1X withsera containing polyclonal antibodies that were performed in duplicate. AFM data are results of 30measurements on 6 different bacteria in 2 separate runs.

were incubated with antibodies specific for Asa1 or Asa373 prior to AFM forcemeasurements and adhesion experiments. The results showed a clear decrease in both theadhesion force upon retraction and the positive cooperativity, indicating that the interactionwas through Agg and could be inhibited with antibodies to the Agg. Non specific influencesof the antibodies were excluded by incubating the strain without Agg (OG1X) withantibodies, but this did not yield a change in interaction forces or positive cooperativity. After incubation with polyclonal antiserum the presence of antibodies on the bacteria insuspension was checked with immune fluorescence with FITC labeled mouse anti-rabbitantibodies (data not shown). The immune fluorescence assay showed only antibody coatingfor E. faecalis expressing Agg when incubated with the matching polyclonal antiserumwhich showed that it were the specific antibodies and not other components of the serumthat interacted with the forces between the E. faecalis strains.Remarkably, not only the positive cooperativity but also the number of adhering bacteria atstationary end point decreased after incubation with antibodies to Agg. Possible, a role inthe prevention of biomaterial related enterococcal infections might be granted to theseantibodies through their interference with positive cooperative mechanisms of adhesion.Previous studies performed on the influence of antibodies on bacterial adhesion are in linewith our current results and showed a decrease in the number of bacteria adhering atstationary end-point (245) or initial deposition rate (196). Agg is associated withenterococci causing infections or colonizing hospitalized patients. Here it is shown thatantibodies to Agg, obtained passively or actively via immunization could play a role in theprevention of infections with enterococci in hospitalized patients.
In conclusion, this is the first time that the influence of specific antibodies oninteraction forces between E. faecalis cells has been demonstrated by AFM. Theseinteraction forces are affected by adsorption of antibodies specific to Agg and have a directimpact on the way these bacteria colonize a biomaterials surface.
91
CHAPTER
6


7
CHAPTER
Adhesion to Bile Drain Materials andPhysicochemical Surface Properties ofEnterococcus faecalis Strains Grown
in the Presence of Bile
Karola Waar, Henny C. van der Mei, Hermie J. M. Harmsen, John E. Degener and Henk J. Busscher
Applied and Environmental Microbiology 2002; 68:3855-3858

Abstract
The aim of this study is to determine whether growth in the presence of bile influences thesurface properties and adhesion to hydrophobic bile drain materials of Enterococcus faecalisstrains expressing aggregation substance (Agg) or enterococcal surface protein (Esp), twosurface proteins that are associated with infections. After growth in the presence of bile, thestrains were generally more hydrophobic by water contact angles and zeta potentials weremore negative than when the strains were grown in the absence of bile. Nitrogen wasfound in lower surface concentrations upon growth in the presence of bile, whereas highersurface concentrations of oxygen were measured by X-ray photoelectron spectroscopy.Moreover, an up to twofold-higher number of bacteria adhered after growth in bile for E.faecalis not expressing Agg or Esp and E. faecalis with Esp on its surface. E. faecalisexpressing Agg did not adhere in higher numbers after growth in bile, possible because theymainly adhere through positive cooperativity and less through direct interactions with asubstratum surface. Since adhesion of bacteria is the first step in biomaterial-centeredinfection, it can be concluded that growth in bile increases the virulence of E. faecalis.
GR
OW
TH O
F E.
FA
ECA
LIS
IN B
ILE
INC
RES
ES A
DH
ESIO
N
94

Introduction
Biliary drains play an important role in biliary surgery and drainage, but infections associatedwith biliary drains constitute a major complication (270). One of the predominant microbialspecies isolated from biliary drain devices is Enterococcus faecalis (270).Biomaterial-centered infections are initiated by bacterial adhesion and biofilm formation onthe indwelling device (e.g. bile drain) (45). Initial microbial adhesion is generally believed todepend on the physicochemical properties of the microbial and biomaterial surfaces, likehydrophobicity or electrostatic charge (22). These physicochemical properties can be linkedto the chemical composition of the bacterial cell surface as measured by X-ray photo-electron spectroscopy (XPS) (243). Some reports indicate that bile plays a role in theadhesion of bacteria. For example, the presence of bile in the gut might be beneficial, as itinhibits bacterial invasion of enterocytes and bacterial translocation through the gut wall(256). E. faecalis is resistant to the bactericidal effects of bile and this resistance is inducedby expression of a large number of stress proteins (76).Two surface proteins of E. faecalis, the aggregation substance (Agg) and enterococcalsurface proteins (Esp) are reported to be associated with infections, suggesting that theseproteins may increase the ability of this microorganism to adhere (123, 217). Recently, itwas also shown that the presence of the esp gene was associated with the capacity of E.faecalis to form a biofilm on a polystyrene surface (235).The aim of the study is to determine whether growth in the presence of (ox) bile of E.faecalis strains expressing the surface proteins Agg (Asa1 and Asa373) and Esp influencesthe numbers of bacteria adhering to different bile drain materials. Also, the influence ofgrowth in the presence of bile on cell surface properties involved in initial adhesion(hydrophobicity, electrostatic charge and chemical composition) was determined.
Materials and Methods
Strains and growth conditions
Three isogenic E. faecalis strains were used in this study: the plasmid-free strain OG1X(116), OG1X carrying the sex-pheromone-responsive plasmid pAD1 (OG1XE:pAD1)encoding the Agg Asa1, with a positive regulator gene inserted which induces consti-tutive expression of this plasmid (173); and OG1X containing the plasmid pAM373(OG1X:pAM373) which expresses Asa373 (175) The three isogenic OG1X derived strainsdid not contain the esp gene, as confirmed by PCR. Expression of Agg was checked using afluorescent-antibody technique with polyclonal antibodies against Asa1 or Asa373, and thepresence of the gene encoding Asa1 or Asa373 was checked with Southernblothybridization. E. faecalis MMH594 expresses Esp but not Agg as was confirmed byimmunological fluorescence microscopy. The presence or absence of these characteristicswas checked at regular time intervals.The strains from a frozen stock were streaked on blood agar plates (blood agar base CM55supplemented with 5 % sterile sheep blood; Oxoid) and grown overnight at 37 °C. Several
95
CHAPTER
7

colonies were used to inoculate 3 ml of Todd-Hewitt broth (THB; Oxoid) that was incubatedat 37 °C for 24 h. From this preculture, 2 ml was used to inoculate 200 ml THB with orwithout 50 mg of ox bile ml-1 (BactoTM oxgall, Difco). This second culture was incubated for20 h at 37 °C without shaking. Ox bile was chosen instead of human bile, because it waseasier to obtain and its composition more consistent. The powdered bile was dissolved indemineralized water and filter sterilized immediately before use. The final concentration ofox bile was similar to the physiological concentration of ox bile. E. faecalis MMH594 wasgrown in THB with 500 µg gentamycin ml-1. E. faecalis OG1X:pAM373 was grown in thepresence of pheromone as described previously (120). Bacteria from the second culturewere harvested by centrifugation at 10 000 x g for 5 min at 10 °C and washed twice withdemineralized water. Subsequently, bacteria were sonicated on ice twice for 10 s each toseparate cell clusters, and a fraction was counted in a Bürker-Türk counting chamber. Thebacteria were diluted in phosphate-buffered saline (PBS) (10 mM potassiumphosphate and0.15 M NaCl at pH 7) to a concentration of 3 x 108 cells ml-1.To check the growth phase during adhesion experiments, growth curves with and withoutox bile were compared. Bacteria were suspended in THB with or without 50 mg ox bile ml-1 at a concentration of 106 cells ml-1 and grown for 24 h at 37 °C. The optical densitywas measured at regular time intervals at 540 nm.
Microbial cell surface characterization
The surface properties of the E. faecalis strains were characterized after growth in THB withor without ox bile added.For zeta potential measurements, each bacterial strain was resuspended in 50 ml PBS to adensity of approximately 107 cells ml-1. The electrophoretic mobility of each sample wasmeasured at 150 V using a Lazer Zee Meter 501 (PenKem). The mobility of the bacteriaunder the applied voltage was converted to an apparent zeta potential using the Helmholtz-Smoluchowski equation (183). The zeta potentials were measured in triplicate withseparately cultured bacteria.Water contact angles were determined by the sessile drop technique as previously described(32). Briefly, bacteria were resuspended in demineralized water and deposited onto a 0.45-µm-pore-size filter (Millipore) using negative pressure. A lawn of approximately 50 stackedlayers of bacteria was produced on the filter. The filters were left to air dry until so-calledplateau water contact angles could be measured (approximately 30 min) using anautomated contour monitor. For each strain, contact angles were measured in triplicate withseparately cultured bacteria.The chemical composition of the bacterial surfaces was determined by XPS. The washedpellets were transferred to stainless steel troughs, frozen in liquid nitrogen, and sub-sequently freeze-dried in a Leybold Heraus Combitron CM30 freeze drier. The samples werepressed into small stainless steel cups, put into the XPS chamber (S-Probe; Surface ScienceInstruments) and analyzed as described by Rouxhet and Genet (205). X-Ray production (10kV, 22 mA) at a spot size of 250 x 1000 µm occurred using an aluminum anode. Scans weremade of the overall spectrum in the binding energy range of 0 to1100 eV at low resolution(pass energy 150 eV). The area under each peak, after Shirley background subtraction, was
GR
OW
TH O
F E.
FA
ECA
LIS
IN B
ILE
INC
RES
ES A
DH
ESIO
N
96

used to calculate peak intensities, yielding elemental surface concentration ratios fornitrogen (N), oxygen (O) and phosphorus (P) to carbon (C). Two samples from separatecultures of each strain were examined.
Substratum surfaces
Implant-grade silicone rubber (SR) was obtained from Medin and poly(tetrafluoroethylene-co-hexafluoropropylene) (fluoro-ethylene-propylene, FEP) was supplied by Fluorplast. Forcleaning, substrata were sonicated for 3 min in a surfactant solution (2 % RBS 35 in water;Omniclean), rinsed thoroughly with water, and then washed with methanol anddemineralized water before use, yielding two hydrophobic surfaces with water contactangles of 115 and 108 º for SR and FEP, respectively.
Parallel plate flow chamber, image analysis and adhesion
The flow chamber (internal dimensions: l x w x h, 76 x 38 x 0.6 mm) and image analysis systemhave been described in detail previously (31). Images were taken from the bottom plate (58 x 38mm) of the parallel plate flow chamber with the material under study attached. The flowchamber was cleaned with the detergent Extran (Merck) and thoroughly rinsed with water anddemineralized water. A bacterial suspension of 3 x 108 cells ml-1 in PBS was allowed to flowthrough the system at a flow rate of 1.44 ml min-1 for 4 h with re-circulation at roomtemperature, and images were taken at different time intervals and analyzed. The shear rate was10.6 s-1 which corresponds to the shear rate in a bile drain of 2 mm diameter at a bile productionof 30 ml h-1. The adhesion of the different E. faecalis strains after growth in the absence andpresence of ox bile was compared. All adhesion experiments were performed in triplicate withseparately cultured bacteria. E. faecalis MMH594 is highly resistant to gentamicin and wasgrown in the presence of gentamicin to keep the selective pressure. Growth in the presence ofgentamycin did not influence the adhesion because the ranges of values for adhesion of strainMMH594 after growth in medium with and without antibiotics were the same (data not shown).
Statistical analysis
Data with and without growth in ox bile were compared with the Students’ t-test assumingnormal distribution of the data. Significance was defined at P ≤ 0.05.
Results
Growth in ox bile
Growth curves showed that the E. faecalis strains reach the logarithmic growth phase afterabout the same time period in medium with and without bile and were harvested in theearly log phase. However, total numbers of bacteria were lower when bacteria were grownin the presence of ox bile.
97
CHAPTER
7

Microbial cell surface characterization
Table 1 shows that all E. faecalis strains became significantly more hydrophobic whengrown in the presence of ox bile, except for OG1XE:pAD1 (difference not statisticallysignificant). The zeta potentials of all E. faecalis strains in PBS are negative and becomeeven more negative for most strains after growth in the presence of ox bile (Table 1). Thechemical cell surface compositions determined by XPS are also shown in Table 1. The N/Cand P/C elemental surface concentration ratios decreased when the strains were grown inthe presence of ox bile, while the O/C surface concentration ratios increased, except forOG1X:pAM373, which had a lower O/C surface concentration in the presence of ox bile.
Initial deposition rates and numbers of bacteria adhering in a stationary end point
The numbers of bacteria in the images taken were transformed to numbers of bacteriaadhering per unit area and fitted to an exponential curve by the least-squares method. Thenumber of bacteria adhering at stationary end point of the adhesion process could beestimated from this exponential curve (SigmaPlot for Windows, version 5.00, SPSS Inc.).The initial increase in the number of adhering bacteria over time was expressed in an initialdeposition rate. Table 2 shows that upon growth in the presence of bile, initial depositionrates of OG1X and OG1XE:pAD1 on SR became significantly higher, while on FEP, theincreases for these strains were not statistically significant. After growth in ox bile, numbersof bacteria adhering in a stationary end point of adhesion were significantly higher for E.faecalis OG1X and MMH594, both on FEP and SR.
GR
OW
TH O
F E.
FA
ECA
LIS
IN B
ILE
INC
RES
ES A
DH
ESIO
N
98
Table 1. Water contact angles, zeta potentials, and elemental surface composition ratios as determinedby XPS for E. faecalis strains expressing Agg or Esp grown in the absence or presence of ox bile.
Elemental surface compositiond
Strain Ox bilea Phenotype �wb �c N/C O/C P/C
OG1X – Agg-, Esp- 35 -17 0.11 0.46 0.03+ 40* -17 0.10 0.56 0.03
OG1XE:pAD1 – Agg+, Esp- 47 -12 0.12 0.47 0.02+ 48 -19* 0.12 0.49 0.01
OG1X:pAM373 – Agg+, Esp- 33 -14 0.13 0.58 0.03+ 41* -18* 0.09 0.44 0.01
MMH594 – Agg-, Esp+ 33 -20 0.10 0.50 0.02+ 37* -28* 0.10 0.53 0.02
a E. faecalis strains were grown in the absence (-) or presence (+) of ox bile; b �w, water contact angles(in degrees). The water contact angles and zeta potentials were measured on three separate culturesyielding a standard deviation of less than 15%; c �, zeta potentials (in mV). The zeta potentials weremeasured in PBS on three separate cultures yielding a standard deviation of less than 20 %; d The elemental surface compositions of carbon, oxygen, nitrogen, and phosphorus were measured ontwo separate cultures yielding a standard deviation of less than 30 %.* Significantly difference from the corresponding strain grown in the absence of ox bile (P ≤ 0.05).

Positive cooperativity
Positive cooperativity in microbial adhesion to surfaces is defined as the ability of oneadhering organism to stimulate the adhesion of other organisms in its immediate vicinityand reflected by the spatial arrangement of adhering organisms as analyzed by radial pairdistribution functions g(r) (chapter 5). When enterococci are randomly distributed over theentire substratum surface, g(r) = 1. However, if there is preferential adhesion at a givenseparation distance r between adhering bacteria, then g(r) > 1. Table 2 summarizes thedegree of positive cooperativity. OG1XE:pAD1 adhered most through positive cooperativemechanisms, and its degree of positive cooperativity became significantly lower aftergrowth in ox bile. The weak positive cooperativity of strain OG1X also became significantlylower after growth in ox bile.
Discussion
The main finding of this study is that growth in the presence of ox bile stimulates theadhesion of some E. faecalis strains, as up to twofold-more bacteria adhered at stationaryend point of adhesion for strain OG1X not expressing Agg or Esp and strain MMH594expressing Esp on its surface. These results indicate that growth in bile changes theadhesion properties of E. faecalis and thereby possibly increases its capability to adhere andinfect. Analysis of the cell surface properties revealed that the strains were generally morehydrophobic and zeta potentials more negative after growth in the presence of ox bile,concurrent with decreases in N/C and P/C elemental surface concentration ratios and anincrease in O/C surface concentration ratio (except for OG1X:pAM373).Hydrophobic and charge interactions contribute to the initial adhesion to surfaces of most
99
CHAPTER
7
Table 2. Initial depostion rate, number of bacteria at stationary end point of adhesion, and degree ofpositive cooperativity as derived from radial distribution functions involved in the adhesion of E.faecalis strains to FEP and SRa
j0b n at t∞
c g(rp)d
Strain Oxbilea Phenotype FEPd SRe FEP SR FEP SR
OG1X – Agg-, Esp- 1646 839 6.1 2.6 1.4 2.3+ 1819 1669* 8.2* 5.0* 1.2* 1.6*
OG1XE:pAD1 – Agg+, Esp- 1703 990 14.0 11.2 3.2 2.8+ 1775 1637* 14.0 11.8 2.0* 1.8*
OG1X:pAM373 – Agg+, Esp- 1619 1260 11.5 7.9 2.0 2.6+ 1639 1285 9.9 6.9 2.3* 2.5
MMH594 – Agg-, Esp+ 1719 1806 10.3 7.1 1.0 1.1+ 1843 1818 22.3* 10.6* 1.0 1.1
a E. faecalis strains were grown in the absence (-) or presence (+) of ox bile; b j0, initial depostion rate(in cm-2s-1); c n at t∞,number of bacteria at stationary end point of adhesion (in 106 cm-2); d g(rp),degree of positive cooperativity. Experiments were performed in triplicate with separate bacterialcultures and yielded a standard deviation of less than 30 %.* Significantly different from the corresponding strain grown on the absence of ox bile (P ≤ 0.05)

pathogens and a relationship between hydrophobicity and infection has been described fordifferent microorganisms (57). Different growth conditions can alter the hydrophobicity andcharge of a microorganism (157). In this study, we found an increase in hydrophobicitywhen bacteria were grown in the presence of ox bile and a more negative zeta potential.These changes may be the effect of the expression of different surface proteins in responseto bile salts present in the growth medium (76). A negative surface charge is oftenassociated with the presence of oxygen-rich groups, whereas a more positive charge isassociated with the presence of nitrogen-rich groups (243). In this respect, it is interestingthat all enterococcal strains possessing a higher number of oxygen-rich groups (and asimilar or slightly lower amount of nitrogen-rich groups) after growth in the presence of oxbile have a more negative zeta potential. Alternatively, strain OG1X:pAM373 has a lowernumber of oxygen-rich groups upon growth in the presence of ox bile, but this strain gainsnegative charge through a relatively large loss of positively charged, nitrogen-rich groups.In oral streptococci, lower N/C elemental surface concentration ratios and higher O/C ratioshave been associated with decreased expression of proteinaceous, fibrillar structures on thesurface and a loss of cell surface hydrophobicity (243). Despite the loss of nitrogen-richsurface groups upon growth in the presence of ox bile, strains did not become morehydrophilic but showed minor increases in cell surface hydrophobicity, except for E. faecalisOG1XE:pAD1, which was most hydrophobic when grown in the absence of ox bile. In this study, we found an increase in deposition rates and/or numbers of bacteria adheringat stationary end point of the E. faecalis strains after growth in ox bile. This is in line withthe generally higher cell surface hydrophobicity after growth in bile, as hydrophobic strainsadhere preferentially to hydrophobic substrata. Evidently, the stronger electrostaticrepulsion between the strains and the negatively charged substrata caused by their morenegative zeta potentials is overruled by the increased cell surface hydrophobicity. In a previous study, we reported that enterococci expressing the sex-pheromone plasmidencoding Asa1 or Asa373 adhere in high numbers to FEP and SR through positivecooperative mechanisms via specific interactions between the bacteria mediated by Agg. E.faecalis strains MMH594 and OG1X did not adhere to the substratum through strongspecific positive cooperativity (chapter 5). These differences in adhesion mechanism alsoexplain the difference in reaction to the growth in ox bile as reported in this study. Thepositive cooperativity as described for the strains expressing Agg probably diminishes theinfluence of the growth in the presence of ox bile on the adhesion. However, the positivecooperativity exhibited by OG1XE:pAD1 is partly inhibited by growth in ox bile, possiblydue to interference of components of the ox bile with receptors involved in the specificinteraction between the bacteria (Table 2). The positive cooperativity exhibited byOG1X:pAM373 is not inhibited by growth in ox bile. The difference in loss of positivecooperativity between the two strains expressing Agg can be explained by the difference inchange of elemental surface composition and water contact angels after growth in ox bile(Table 1). Finally, the weak positive cooperativity exhibited by OG1X was also inhibited bybile. This inhibition might be due to changes in physico-chemical surface properties after thegrowing E. faecalis OG1X in ox bile (Table 1).Several reports described a possible association between the presence of Agg and infection.This might not be in line with our findings that growth in bile only increases the adhesion of
GR
OW
TH O
F E.
FA
ECA
LIS
IN B
ILE
INC
RES
ES A
DH
ESIO
N
100

the strains not expressing Agg. However, in a previous study to the association betweenAgg of E. faecalis and infection in liver transplant patients, we found an associationbetween Agg and infection only if the E. faecalis isolates were divided into differentgenogroups specific for liver transplant patients (chapter 2). Therefore, we therefore thinkthat not only the presence of surface proteins but also the environment, growth conditions,and selection of specific genogroups may contribute to virulence of E. faecalis.In conclusion, growth in the presence of (ox) bile changes the surface properties of the E.faecalis strains and increases adhesion of the strain expressing Esp (MMH594) and thestrain not expressing Esp or Agg (OG1X), but not of the strains expressing Agg (Asa1 andAsa373). The findings of this study will stimulate further research to the effect of bile on E.faecalis. Questions that need to be answered are whether and which surface proteins areregulated or modified in reaction to the presence of bile, since bile components may bind tothe surface of E. faecalis.
We thank A. B. Muscholl-Silberhorn for providing the OG1X strains and antibodies and V.Shankar for providing E. faecalis MMH594. We also thank Joop de Vries for performingXPS.
101
CHAPTER
7


8
CHAPTER
Summarizing Discussion


In the normal healthy host the bacterium Enterococcus faecalis colonizes the gastro-intestinal tract and seldom causes infections. However, in hospitalized patients thismicroorganism is at the increase as a cause of infections and liver transplant patients areespecially vulnerable to these infections. Why these patients are more frequently infectedwith E. faecalis compared to other patients is not clear. In the pathogenesis of infectiongenerally two factors play a role. On the one hand, the immunity that normally protects thepatients can be diminished by immune suppressive treatment and medical interventions e.g.surgery or implantation of an indwelling biomaterial that will break the natural defense ofthe skin and mucous membranes. On the other hand, bacteria that are normally notpathogenic in the healthy host can acquire special traits that enhance virulence or increaseadhesion to biomaterials and therefore cause infections in the hospital environment. Theaim of this thesis was to gain insight in the pathogenesis of infections with the bacterium E.faecalis with special interest in liver transplant patients. The focus of our investigation wasthe microorganism and to find traits and mechanisms of E. faecalis that might lead to moresevere infection, spread or biomaterial associated infections. In Part I of this thesis theinvestigations to the role and epidemiology of E. faecalis in infections in the host aredescribed, Part II focuses on biomaterial centered infections with E. faecalis and the role ofsome surface proteins of E. faecalis. In the current chapter, the different investigations thatwere conducted to answer these questions are summarized, the results are discussed, theshortcomings are described and future directions are given.Many different traits of E. faecalis that possibly play a role in infections are described inliterature (Chapter 1). To study the possible role of these virulence factors in infections inliver transplant patients, we compared the prevalence of these traits among E. faecalisisolates from liver transplant patients to the prevalence among isolates from other patientsand healthy volunteers (Chapter 2). The prevalence of the virulence factors cytolysin(hemolysin), gelatinase, enterococcal surface protein (Esp) and the genes asa1 and asa373encoding two forms of aggregation substance (Agg) was determined among isolates fromliver transplant patients (133 isolates), blood culture isolates from non-liver transplantpatients (66 isolates) and a control group of non-clinical enterococci from feces of healthyvolunteers (47 isolates). To exclude multiple identical isolates, amplified fragment lengthanalysis (AFLP) was performed. In addition, the AFLP analysis furthermore discerned 11genogroups in which the strains shared > 80 % of the restriction fragments. The majority ofthe isolates clustered in three of these genogroups that respectively consisted of bloodcultures, isolates from liver transplant patients or isolates from feces of healthy volunteers.Combination of the genogrouping and the presence of virulence factors showed thatcytolysin (hemolysin) and asa1 might be associated with infections, because they weremainly found among isolates from the genogroups representing blood cultures and livertransplant patients. Furthermore, Esp might be associated with colonization because thegene encoding the protein is mainly found in isolates from the genogroups represented byfecal isolates from healthy volunteers or liver transplant patients.Studying the role of virulence factors based on their incidence among isolates from differentgroups of patients can be difficult because almost certainly the collection of isolates will bea mixture of virulent and non-virulent isolates, especially among liver transplant patientsthat are immune compromised and might be vulnerable to both virulent and non-virulent
105
CHAPTER
8

strains. Future studies possibly should also involve other groups of patients to rule out thepossibility of inclusion of non-virulent strains. Very interesting in this study is the finding ofdifferent genogroups among the E. faecalis isolates related to the different hosts that werestudied. This indicates that some enterococci might have special traits that allow them tosurvive in a special environment, e.g. the hospital. The enterococci isolated from thisenvironment are therefore genetically more related compared to enterococci from otherenvironments. This finding opens the opportunity to a new approach in the study of thepathogenesis of E. faecalis infections. Comparing the genomes of the different genogroupsmight lead to the identification of certain traits that the enterococci need to survive in acertain environment. Interference with these traits or colonization of these environmentswith less virulent traits might lead to ways to prevent infections with E. faecalis. The AFLP analysis of the E. faecalis isolates from liver transplant patients described inChapter 2 revealed one particular epidemic clone that colonized 23 of the 43 liver transplantpatients over a 15 months period. Chapter 3 describes the epidemiology and characteristicsof this and other epidemic isolates on the liver transplant ward. Among the 133 E. faecalisisolates from liver transplant patients that are described in Chapter 2, 15 different AFLPtypes could be identified with 90 % similarity which was considered the cut-off point foridentical isolates. Nine of these groups contained isolates from more than one patient whichmight indicate transmission. In five of these groups transmission could be explained by thefact that patients carrying identical strains were staying in the same ward at the same time.Analysis of the characteristics of the epidemic isolates in this study showed that theseisolates did not cause more infections compared to the non-epidemic isolates. Furthermore,the epidemic isolates were more resistant to the antibiotics gentamicin, erythromycin andciprofloxacin but none of the isolates was resistant to the clinically important antibioticsvancomycin or amoxicillin. Reports about the transmission of enterococci susceptible tomost antibiotics are scarce; therefore, the most important message of Chapter 3 is thedescription of the transmission of a susceptible E. faecalis strain. Knowledge about thetransmission of these strains is important because enterococci are able to acquire resistantdeterminants relatively easily and unrecognized transmission of susceptible isolates mayfinally result in outbreaks of multi-resistant strains. Another important finding was that theepidemic clone was positive for the surface protein Esp. This confirms the role of Esp intransmission and spread and emphasizes the possibility of interference with transmissionthrough interaction with Esp.In Chapter 4 the design and testing of two DNA oligonucleotides specific for the 16S rRNAgenes of respectively E. faecalis and Enterococcus faecium are described. When theseoligonucleotides are labeled with a fluorescent dye they can be used for fluorescent in situhybridization (FISH). FISH is a method to directly visualize and identify bacteria in a clinicalsample without any culturing. Testing of the oligonucleotides on other enterococci showedthat the E. faecalis probe only reacted with E. faecalis species. The E. faecium probe alsohybridized with Enterococcus hirae, Enterococcus mundtii and Enterococcus saccharo-lyticus as could be expected from the sequence information. We further describe FISHprotocols for the detection of E. faecalis and E. faecium in fecal samples, blood cultures andbiofilms on indwelling devices. The studies that use the FISH for detection of enterococci aregenerally hampered by the fact that they are only focused on one material. By describing
SUM
MA
RIZ
ING
DIS
CU
SSIO
N
106

the use of FISH on different clinical samples we show that the oligonucleotides can bebroadly applied to different materials. The most interesting fact of this report is that wewere able to show that the enterococci indeed are present on a biofilm attached to a biledrain. This opens the possibility to study the distribution of enterococci in biofilms that areformed in vivo.In Part II of this thesis the focus shifts towards biomaterial related infections with E. faecalis.When a biomaterial is introduced into the body it frequently gets colonized with bacteria.These bacteria form a biofilm on the biomaterial. This biofilm can be a source of persistentinfections because bacteria in a biofilm are generally more resistant to antibiotics. The firststep in biofilm formation is adhesion of the bacteria to the material. Insight in themechanisms of adhesion can lead to the development of methods to prevent this adhesionand therefore biofilm formation. Our investigations were focused on liver transplantpatients and in these patients the bile drain that is used to divert the bile outside the bodyis a frequent source of infection. Therefore we chose to study the adhesion of E. faecalis todifferent bile drain materials that are used at the University Hospital Groningen.First, the role of three surface proteins of E. faecalis (Esp and two forms of Agg; Asa1 andAsa373) in the adhesion to silicone rubber, fluoro-ethylene-propylene and polyethylenewas examined (Chapter 5). Four isogenic E. faecalis strains with and without Agg and onestrain expressing Esp were used. With the use of a parallel plate flow chamber the kineticsof initial adhesion of the strains to the materials were studied in detail. The results indicatethat the surface proteins of E. faecalis play a key role in the adhesion and increased the totalnumber of adhering bacteria. However, the different surface proteins increased theadhesion of E. faecalis through different mechanisms. Analysis of the distribution of theenterococci after the initial adhesion showed that the enterococci expressing Agg adheredin higher numbers through mechanisms of positive cooperativity, which means thatadhesion of bacteria enhances the probability of adhesion of other bacteria near thesebacteria. Enterococci positive for Esp also adhered in high numbers, but did not utilizepositive cooperative mechanisms in their adhesion to the biomaterials. Possibly, Espincreased the strength of the direct interaction between E. faecalis and the material. Thesefindings indicate some possibilities to design drains that prevent biomaterial relatedinfections. This might be through interference with the positive cooperativity betweenadhering enterococci, as resulting from the expression of Agg e.g. by adding antibody toAgg or through interference with the interaction between surface proteins and materialse.g. by changing the surface properties of the biomaterial.In the search for ways to prevent biomaterial related infections, we investigated whetherantibodies to Agg can interfere with the positive cooperativity between E. faecalisexpressing Agg (Chapter 6). The interaction forces between E. faecalis strains with andwithout Agg (Asa1 and Asa373) were measured at a molecular level by use of atomic forcemicroscopy (AFM). E. faecalis expressing Agg showed nearly two-fold higher interactionforces between bacterial cells than a strain lacking Agg. Interestingly, these stronginteraction forces between the strains with Agg could be reduced by adsorption withantibodies against Agg. Comparison of the results of the AFM interaction forces with thepositive cooperativity after adhesion to a biomaterial in a parallel plate flow chambershowed that in the absence of strong interaction forces between the bacteria, positive
107
CHAPTER
8

cooperativity was absent too. The absence of positive cooperativity further resulted in alower total number of adhering bacteria. The results of this study show that specificantibodies influence the interaction forces between E. faecalis and that these interactionforces have a direct impact on the way these bacteria colonize a biomaterial surface.Furthermore, a role in the prevention of biomaterial related enterococcal infections might begranted to these antibodies.Evidence from literature indicates that E. faecalis is resistant to the bactericidal effects of bileand this resistance is induced by expression of a large number of stress proteins (76).Furthermore, some reports indicate that bile plays a role in the adhesion of bacteria (230,256). Therefore, we investigated the influence of growth in the presence of bile on theadhesion of E. faecalis to bile drain materials and measured the physicochemical propertiesof the bacterial cell surface (Chapter 7). Because the previous study showed that the surfaceproteins Esp and Agg played a role in adhesion, we used strains expressing these proteins.After growth in the presence of bile, the strains were generally more hydrophobic and zetapotentials were more negative than when the strains were grown in the absence of bile.Moreover, E. faecalis not expressing Agg or Esp and E. faecalis with Esp on its surfaceadhered in an up to twofold-higher number after growth in bile. E. faecalis expressing Aggdid not adhere in higher numbers after growth in bile, possible because they mainly adherethrough positive cooperativity and less through direct interactions with a substratumsurface. The results of this study show that growth in bile can increase the virulence of E.faecalis as adhesion of bacteria is the first step to biomaterial related infection. Analysis ofthe effect of bile on the expression of surface proteins might lead to new ways to preventbile drain related infections.The parallel plate flow chamber model that was used in the studies to biomaterial relatedinfections is a very adequate model to study the adhesion mechanisms in detail. Thedisadvantage of this in vitro model is that is it very difficult to imitate the in vivo situation. Invivo many more factors influence the adhesion and biofilm formation than the variablesincluded in our study. In the study to the influence of bile on the adhesion, not only growth inbile but also the presence of bile during the adhesion process and the coating of bile drain withbile components prior to the adhesion might influence the adhesion. These variables could bethe subject of future studies. Furthermore to the study of the use of antibodies to preventadhesion to biomaterials it should be noticed that coating the bacteria with antibodies beforeadhesion is not a measure that can be performed in vivo. However, the study of isolatedvariables that influence adhesion is the only way to gain more insight in the exact mechanismsof adhesion. The results of these experiments will have to be confirmed in an in vivo model.
General conclusions
● E. faecalis isolates from different groups of patients and healthy volunteers can bedivided into different genogroups with > 80 % shared restriction fragments and eachgenogroup with specific predominant virulence factors.
● Virulence factors cytolysin (hemolysin) and Asa1 might be associated with infection andEsp with colonization and spread between (liver transplant) patients.
SUM
MA
RIZ
ING
DIS
CU
SSIO
N
108

● E. faecalis with different surface proteins adheres to the substratum through differentmechanisms. Enterococci expressing Agg adhere in high numbers through positivecooperativity between adhering bacteria but enterococci positive for Esp do not adherethrough this mechanism.
● Antibodies to Agg interfere with the interaction forces between E. faecalis expressingAgg as measured with AFM and these forces have a direct impact on the way thesebacteria colonize a biomaterial surface.
● Growth in the presence of (ox) bile changes the surface properties of E. faecalis andincreases adhesion of some particular strains.
Concluding remarks and future directions
In this thesis, we analyzed the role of the different virulence factors of E. faecalis known inliterature in the pathogenesis of infections. We investigated the prevalence of virulencefactors in E. faecalis isolates from different hosts as well as their role in biomaterial relatedinfections. The results showed that the virulence factors cytolysin (hemolysin) and the Asa1form of the surface protein Agg are involved in infection and the surface protein Esp isassociated with colonization. In addition, the surface proteins Agg and Esp play a role in theadhesion to biomaterials. However, another important conclusion from this thesis is thatalso other factors are involved in the pathogenesis of infection; E. faecalis without any ofthe virulence factors which are known thus far also caused infections and strains withoutany of the studied surface proteins adhered to biomaterials.Beside the bacterium also the host plays a role in the pathogenesis of bacterial infections. Inmodern medicine severely immune compromised patients are no exception, this makes theanalysis of pathogenesis of opportunistic pathogens more complicated as it seems difficultto apply the postulates of Koch on these patients. When the immune defense of the host iscompromised also non-virulent bacteria can cause infections and collections of isolates thatcause infections in these patients might be a mixture of virulent and non-virulent isolates.Furthermore, evidence suggests that after admission to a hospital the gastrointestinal tractof patients quickly gets colonized with enterococci of nosocomial origin (234, 275). A factorthat predisposes to nosocomial colonization with enterococci is the use of antibiotics withlittle or no anti-enterococcal activity. The question raised is why the indigenous enterococcido not take advantage of their presence and immediately fill the niches that are left by thebacteria killed by the antibiotic regime. The nosocomial enterococci that initially areoutnumbered by the indigenous enterococci finally colonize the gastrointestinal tract andcause infections, this suggests that these nosocomial enterococci poses traits that give themadvantages in this environment (96). The trait that nosocomial enterococci poses is notnecessarily antibiotic resistance because in this thesis we showed that also enterococcisusceptible to the clinically relevant antibiotics can spread between hospitalized patients.This implicates that in future the pathogenesis of a “commensal pathogen” like E. faecalismay not only be studied by direct search for straight virulence factors but also by analysis offactors that enable the bacterium to survive in an environment from where it can easilyinfect the host as soon as the immunity decreases. The environment that nosocomial
109
CHAPTER
8

enterococci colonize before causing an infection might not necessarily be the gastro-intestinal tract of the patient but also the colonization of other niches e.g. the skin orrespiratory tract of patients and the hands of personnel has to be taken into account.Colonization of this kind could be defined as ‘nosocomial colonization’, a term that wouldinclude the colonization of all sites from where the patient can be infected and that wouldalso include competition with the indigenous flora that can be present on this site. The needfor a redefinition of virulence that is implicated by the former statement was recognized byseveral infectious disease specialists (36, 251). They suggested a classification of microbialpathogenicity that takes into account the host immune response, allocating one class formicroorganisms that only cause damage in situations of weak immune response (36). Inaddition, a classification for virulence genes was proposed that divides the genes into truevirulence genes, virulence associated genes and genes essential for a pathogenic life-style(251). Genes that increase the ability of a microorganism to colonize the gastro-intestinaltract as described above could be classified as pathogenic life-style genes. However, inanalogue to the different steps in pathogenesis, virulence factors could also be classified intoclasses that represent the different stages of pathogenesis. This would solve the problem ofdefinition of virulence among opportunistic pathogens as these can have factors that areinvolved in the first steps of pathogenesis. Table 1 gives a suggestion of the classification ofvirulence factors and a suggested arrangement of the known virulence factors of E. faecalisover the different classes as far as the function of the virulence factor is known.The adjustment of the definition of virulence as described above implies also a change in theinfection models that are used to prove the role of virulence factors in pathogenesis. Up tonow, many infection models include the determination of the lethal dose in animals or atleast the development of infection in healthy animals. New infection models should be moredirected towards earlier steps in pathogenesis e.g. colonization or infections in immunecompromised hosts. The publication of the whole genome sequence of the vancomycin resistant E. faecalisstrain V583 (for genome data see http://www.tigr.org/) (192) enables us to study thegenomics of the pathogenesis of E. faecalis infections in more detail. Construction of a DNAmicroarray containing al known genes of E. faecalis will make it possible to analyze theexpression of genes in different environments (47, 187, 253). In analogue with theabundant presence of E. faecalis in the gastrointestinal tract and the rapid colonization ofthe gastrointestinal tract with nosocomial strains, it will be interesting to study the
SUM
MA
RIZ
ING
DIS
CU
SSIO
N
110
Table 1. Suggested classification of the known virulence factors related to the stages of pathogenesis
Virulence factors involved in Virulence factor of E. faecalisa
nosocomial colonization Esp, Aggadhesion to cellular or abiotic surfaces Esp, Agg, Aceinvasion or internalization of human tissue Agg, cyttissue destruction cyt, gelimmune evasion escape from cellular immunity Agg, epa, cyt
escape from humoral immunity Espa Esp, enterococcal surface protein; Agg, aggregation substance; Ace, Adhesin to collagen; epa,enterococcal polysaccharide antigen; cyt, cytolysin; gel, gelatinase

expression of genes upon contact with intestinal epithelial cell lines in vitro. Furthermore, itmight be interesting to study the influence of fecal components on the expression ofdifferent genes as the bacteria in the intestine possibly have more contact with feces thanwith the epithelial lining of the gut. These experiments might be expanded to the analysisof interaction with gelatinase and serine protease other host cells and human secretions thatare frequently involved in enterococcal infections e.g. uroepithelial cells, urine or bile.Furthermore, the host response to enterococcal infections can be analyzed by DNAmicroarray analysis of the host cell lines. Comparison of the response of the host cell linesto isogenic strains of E. faecalis lacking particular virulence factors might also gain furtherinsight in the host-pathogen interaction. These experiments might result in the definition ofnew putative virulence factors.The next step in the analysis of new virulence factors will be to prove that the gene is trulyassociated with virulence and/or colonization. This can be done in different ways: (i) theconstruction of mutants lacking the gene under study and using this mutant in an animalinfection model, (ii) prove of an immunological response to the microorganism and thatspecific immunity against the microorganism can protect against the infection and (iii) bycomparative genetics, showing that the gene encoding the virulence factor is similar toknown virulence factors in other species (251). However, all three methods might bedifficult for virulence factors that are not immediately correlated with pathogenicity butwith colonization and infection in immune compromised hosts by “commensal pathogens”.(i) The problem with the generation of mutants is that the mutant often also will beattenuated in its virulence if the gene is necessary for house keeping of the bacterium.Furthermore, if the gene under study is involved in the colonization of e.g. the humangastrointestinal tract this will be difficult to prove in an animal infection model. (ii) Factorsthat enhance the colonization of the host most likely will not elicit an immune response, and(iii) comparative genetics for genes that are associated with colonization can be difficult asnot many genes are defined yet as colonization genes.Other approaches to verify that a gene is necessary for survival and growth under certainconditions are genetic footprinting or signature-tagged mutagenesis. Both methods involvescanning of the genome by exhaustive transposon mutagenesis and subsequently screeningthe mutants en masse for functional properties (253). Comparison of pools of mutantsgrown under different conditions e.g. in broth culture and in the presence of fecal extractwill lead to the identification of genes that are essential for the growth in certainenvironments because these mutants are not present in the pool grown under theseconditions. This method can also be used to identify new virulence genes.One drawback of the use of microarrays to study pathogenesis is the fact that microarrayscan not measure expression of genes that are absent from the reference strain used toconstruct the microarray. Therefore, microarray analysis always needs to be completed withthe search for new unknown genes. The identification of different genotypes associatedwith specific hosts or environments (Chapter 2) and one clone that spread between manypatients (Chapter 3) can support this analysis of new virulence traits. In addition to this, newgenogroups of E. faecalis can be identified by AFLP or multilocus sequence typing (MLST)of isolates from different patient groups and healthy volunteers from different geographicalregions. Furthermore, new epidemic clones of enterococci that spread between patients can
111
CHAPTER
8

be recognized by analyzing enterococci from different clinical samples with molecularepidemiological methods. Representatives from the new and known genogroups andepidemic clones can be analyzed in the search for new virulence factors.The analysis and comparison of the genomes of selected E. faecalis isolates in the search fornew virulence factors can be done with different methods: (i) the different AFLP patternscan be compared and then DNA sequence of the differing bands can be analyzed. (ii)Another possibility is random sequencing of the genomes and searching for sequencesencoding e.g. a surface protein. (iii) A sophisticated method of comparing genomes andidentifying genes that are present in one genome but absent from another is substractivehybridization (263). (iv) The ultimate method is sequencing of the different isolates anddirect comparison of the genomes. The role in virulence or colonization of the genesidentified with these methods should be analyzed taking into account that these genesmight not be directly virulent in an animal model.At the moment, infections with E. faecalis seem to be mainly restricted to immunecompromised and severely ill patients. However, the emerging resistance of E. faecalis andthe increasing number of infections in hospitalized patients might lead to a point where notonly immune compromised patients get infected with this microorganisms but also personswith a healthy immune system. The dominant presence of E. faecalis might give it theopportunity to acquire virulence factors from other pathogens and combine them with itsown virulence factors into one strain. If a strain subsequently also acquires resistancedeterminants against many antibiotics this will finally result in a “superbug” that is able toinfect the normal healthy host and in addition is difficult to treat. The fact that this scenariois very well possible, is given by reports about the transfer of vancomycin resistancebetween strains (129) and the presence of components of the sex-pheromone plasmidtransfer system in Staphylococcus aureus.Therefore, it is important to keep on searching for the mechanisms of virulence and theAchilles heel of E. faecalis that can be used to treat and prevent infections with thismicroorganism.
SUM
MA
RIZ
ING
DIS
CU
SSIO
N
112

9
CHAPTER
Dutch SummaryNederlandse Samenvatting


Enterococcus faecalis is een ronde bacterie die vooral in de darm van de mens voorkomt. Inde gezonde gastheer veroorzaakt E. faecalis nauwelijks infecties, echter bij patiënten in hetziekenhuis worden regelmatig infecties met E. faecalis gezien. De laatste jaren worden dezeinfecties zelfs steeds vaker gevonden. Verschillende verklaringen voor deze stijging zijnmogelijk; enterokokken zijn steeds minder gevoelig voor de gangbare antibiotica en door demoderne geneeskunde komen er steeds meer patiënten die extra gevoelig zijn voorinfecties. Patiënten die een levertransplantatie hebben ondergaan zijn echter gevoeligervoor infecties met E. faecalis dan vergelijkbare patiënten die een andere ingreep hebbenondergaan, de oorzaak voor dit verschil in gevoeligheid is niet duidelijk.Voor het ontstaan van infecties zijn in het algemeen twee factoren van belang. Allereerst deafweer van de gastheer die normaal bescherming biedt tegen de infectie. Door debehandeling in het ziekenhuis kan deze afweer worden doorbroken. Dit kan gebeuren dooreen operatie of het inbrengen van een biomateriaal zoals een drain of katheter waardoor denatuurlijke barrière van de huid en de slijmvliezen wordt verbroken. Ook het geven vanmedicijnen die de immunologisch afweer tegen micro organismen onderdrukken kan denormale bescherming doorbreken. De tweede factor die een rol kan spelen bij het ontstaanvan infecties is de bacterie zelf. Een bacterie die normaal niet in staat is infecties teveroorzaken kan onder bepaalde omstandigheden eigenschappen verwerven die hem instaat stellen wel infecties te veroorzaken. Deze eigenschappen worden virulentiefactorengenoemd.Het doel van dit proefschrift was om inzicht te krijgen in de manier waarop E. faecalisinfecties bij levertransplantatie patiënten in het ziekenhuis veroorzaakt, hierbij hebben wevooral gekeken naar de eigenschappen van de bacterie. In Deel I van het onderzoek hebbenwe gekeken naar de interactie van E. faecalis met de gastheer en in Deel II naar de interactievan E. faecalis met biomaterialen. Biomaterialen zijn hulpmiddelen van kunststof die voorde behandeling bij de patiënt worden ingebracht, bij voorbeeld infusen of drains waardoorlichaamsvloeistoffen kunnen aflopen. Deze biomaterialen zijn vaak een bron van infecties,daarom hebben wij geprobeerd om de manier waarop E. faecalis deze infecties veroorzaaktte analyseren.De verschillende virulentiefactoren van E. faecalis die beschreven zijn in de literatuur en hunmogelijke rol in infecties zijn samengevat in Hoofdstuk 1. In Hoofdstuk 2 is gekeken naarde rol van deze virulentiefactoren van E. faecalis in het ontstaan van infecties bij lever-transplantatie patiënten. De aanwezigheid van de verschillende virulentiefactoren werdvergeleken tussen E. faecalis isolaten uit klinische materialen van levertransplantatiepatiënten (totaal 133), isolaten uit het bloed van andere patiënten (totaal 66) en isolaten uitde feces van gezonde vrijwilligers (totaal 47). De isolaten uit het bloed werden gekozenomdat zij E. faecalis vertegenwoordigden die bewezen hadden een infectie te kunnenveroorzaken en de isolaten uit de feces werden gekozen omdat zij juist geen infectie haddenveroorzaakt. Om uit te sluiten dat gelijke isolaten werden geanalyseerd die zich tussen depatiënten hadden verspreid is het genetische materiaal (DNA) van de enterokokkengeanalyseerd met amplified fragment length analysis (AFLP). Met deze methode kan opbasis van het DNA dat in stukken wordt geknipt en zichtbaar wordt gemaakt een uitspraakworden gedaan over de mate van genetische verwantschap tussen de verschillendebacteriële isolaten. De resultaten van deze AFLP analyse gaven aan dat de E. faecalis
115
CHAPTER
9

isolaten in drie grote groepen konden worden ingedeeld met minimaal 80 % genetischeovereenkomst. Omdat deze groepen op basis van de genetische verwantschap warengedefinieerd werden ze genogroepen genoemd. De eerste genogroep bestond vooral uitisolaten uit het bloed van niet levertransplantatie patiënten, de tweede genogroep bevattevooral isolaten van levertransplantatie patiënten en de derde genogroep bestond vooral uitisolaten uit de feces van gezonde vrijwilligers. Combinatie van de resultaten van dezegenogroepering en de analyse van het vóórkomen van de verschillende virulentiefactorenliet zien dat het oppervlakte eiwit “aggregatie substantie” (Agg) en het enzym “cytolysine”van E. faecalis geassocieerd zijn met infecties. Deze virulentiefactoren komen namelijkvooral voor in de genogroep van de levertransplantatie patiënten en de bloed isolaten en ditzijn genogroepen die zijn geassocieerd met infecties. Het oppervlakte eiwit “enterococcalsurface protein” (Esp), dat werd ontdekt in een E. faecalis stam die een epidemieveroorzaakte onder ziekenhuispatiënten in de Verenigde Staten bleek in ons onderzoekgeassocieerd met kolonisatie. Het Esp eiwit werd namelijk vooral gevonden bij isolaten uitde derde genogroep die geen infectie hadden veroorzaakt. De belangrijkste conclusie uit ditonderzoek is dat de enterokokken kunnen worden verdeeld in genogroepen die zijngeassocieerd met een specifieke gastheer of omgeving (bloed, feces of levertransplantatiepatiënten) en met bepaalde virulentiefactoren.De AFLP analyse van de 133 E. faecalis isolaten van de levertransplantatie patiënten uitHoofdstuk 2 onthulde verder nog dat verschillende patiënten precies dezelfde bacterie bijzich droegen. Dit wijst er op dat de bacterie is overgedragen van de ene op de anderepatiënt. In Hoofdstuk 3 worden deze overdracht en de eigenschappen van de over-gedragen bacteriën geanalyseerd. De meest opvallende bevinding was één E. faecalis stamdie zich gedurende 15 maanden over 23 levertransplantatie patiënten verspreidde. Verderveroorzaakten de overgedragen E. faecalis stammen niet meer infecties dan anderestammen, ze waren wel vaker resistent tegen de antibiotica gentamicine, erytromycine andciprofloxacine maar niet tegen de belangrijke antibiotica vancomycine en amoxicilline. Deoverdracht van E. faecalis stammen die gevoelig zijn voor deze laatste antibiotica is nogzelden beschreven. Het is wel belangrijk om naar de overdracht van deze gevoeligeenterokokken te kijken omdat enterokokken zeer goed in staat zijn om resistent te wordentegen allerlei antibiotica. Op deze manier kan de overdracht van gevoelige enterokokkenresulteren in een uitbraak van enterokokken die resistent zijn tegen veel antibiotica. Eenander belangrijk gegeven uit Hoofdstuk 3 is het feit dat de stam die zich over de 23levertransplantatie patiënten verspreidde positief was voor het Esp eiwit. Dit bevestigt deconclusie uit Hoofdstuk 2 dat Esp is geassocieerd met kolonisatie en verspreiding en opentmogelijkheden om deze verspreiding te onderbreken door middelen die een interactieaangaan met het Esp eiwit.In Hoofdstuk 4 wordt het ontwerpen en testen van twee DNA sondes beschreven diespecifiek kunnen binden aan het 16S rRNA gen van respectievelijk E. faecalis enEnterococcus faecium. Deze enterokokken veroorzaken samen meer dan 95 % van deinfecties met enterokokken. Als deze DNA sondes worden voorzien van een fluorescerendelabel kunnen ze worden gebruikt voor fluorescerende in situ hybridisatie (FISH). FISH is eenmethode waarmee bacteriën door binding met de fluorescerende DNA sonde zichtbaarkunnen worden gemaakt, een voordeel hierbij is dat de bacterie niet eerst gekweekt hoeft
NED
ERLA
ND
SE S
AM
ENV
ATTI
NG
116

te worden en direct zichtbaar is in het materiaal. In Hoofdstuk 4 stellen wij verschillendeprotocollen op waarmee E. faecalis en E. faecium met de FISH methode kunnen wordengedetecteerd in verschillende klinische materialen zoals bloed of feces. Veel onderzoekenwaarin DNA sondes voor de FISH methode wordt gebruikt beschrijven het gebruik in slechtséén materiaal. Door het gebruik van deze twee sondes met de FISH methode inverschillende materialen te combineren laten we zien dat dit een breed inzetbare methodeis om enterokokken in klinisch materiaal te detecteren.In Deel II van dit proefschrift verschuift de aandacht naar biomateriaal gerelateerde infectiesmet E. faecalis. Als een lichaamsvreemd materiaal, b.v. een biomateriaal in het lichaamwordt ingebracht raakt het meestal snel gekoloniseerd met bacteriën. Deze bacteriënhechten zich aan het oppervlak van het materiaal en vormen daar een biofilm. Een biofilmis een laag bacteriën die meestal zijn ingebed in een laag slijm en soms ook eiwittenafkomstig van de gastheer. Een biofilm op een biomateriaal kan een aanhoudende bron van infecties zijn omdat de bacteriën kunnen loslaten uit de biofilm en elders in het lichaam infecties kunnen veroorzaken. Bovendien is een dergelijke biofilm moeilijk teverwijderen omdat de bacteriën in de biofilm moeilijk te bereiken zijn door antibiotica. De eerste stap in biofilm vorming is de hechting van de bacteriën aan het oppervlak. Het is daarom van belang om dit proces van hechting te bestuderen. Inzicht in de mechanismenachter dit proces kan leiden tot het ontwikkelen van methoden om de hechting en daar-mee de biofilm vorming te voorkómen. Dit onderzoek is gericht op levertransplantatiepatiënten, die na de transplantatie meestal een galdrain in de buik krijgen waardoor de galgedurende enige weken kan aflopen. Deze drains zijn vaak een bron van infecties metonder andere E. faecalis. Daarom hebben we ervoor gekozen om de hechting teonderzoeken aan materiaal waar de galdrains in het Academisch Ziekenhuis Groningen vangemaakt zijn.Allereerst hebben we de invloed van drie verschillende oppervlakte eiwitten (Esp en tweevormen van Agg) op de binding van E. faecalis aan de galdrain materialen bestudeerd(Hoofdstuk 5). Hiervoor werd gebruik gemaakt van E. faecalis stammen die alleenverschilden in de expressie van de betreffende eiwitten. De kinetica van de hechting van de bacteriën werd bestudeerd in een parallelle plaat stroom kamer. De resultaten toondenaan dat meer bacteriën met de oppervlakte eiwitten aan het materiaal hechtten vergelekenmet bacteriën zonder de oppervlakte eiwitten. De verschillende oppervlakte eiwittenbeïnvloedden de hechting echter op verschillende manieren. Analyse van de verdeling vande bacteriën op het oppervlak liet zien dat de enterokokken met Agg clusters vormden ophet oppervlak. Deze vorming van clusters wordt positieve coöperativiteit genoemd en geeftaan dat de binding van een bacterie op het oppervlak de binding van andere bacteriën in debuurt stimuleert. Enterokokken met Esp op het oppervlak hechten niet via positievecoöperativiteit maar waarschijnlijk versterkt Esp de directe interactie met het materiaal.Deze resultaten geven een aantal indicaties voor manieren om biomateriaal gerelateerdeinfecties te voorkómen. Enerzijds kan worden gedacht aan onderbreking van de positievecoöperativiteit, bij voorbeeld met antilichamen gericht tegen Agg, anderzijds is het mogelijkeigenschappen van het biomateriaal te veranderen om de interactie met Esp teverminderen.In de zoektocht naar mogelijkheden tot preventie van biomateriaal gerelateerde infecties
117
CHAPTER
9

hebben we met behulp van atomaire krachten microscopie de invloed van antilichamentegen Agg op de positieve coöperativiteit tussen E. faecalis bacteriën onderzocht(Hoofdstuk 6). Met de atomaire krachten microscopie kan de interactiekracht tussen enkelebacteriën zeer nauwkeurig gemeten worden. De metingen toonden aan dat enterokokkenmet Agg op het oppervlak tot twee keer hogere interactiekrachten hadden danenterokokken zonder Agg op het oppervlak. Bovendien namen deze interactiekrachtenduidelijk af wanneer de bacteriën met Agg werden geïncubeerd met antilichamen tegenAgg. Vergelijking van de gemeten interactiekrachten met de positieve coöperativiteit nabinding aan een biomateriaal liet een duidelijke correlatie zien tussen de gemeten krachtenen de positieve coöperativiteit. Bij kleinere krachten was er minder coöperativiteit enuiteindelijk werden ook minder bacteriën gebonden aan het oppervlak. Dit wijst er op dater mogelijk een rol is weg gelegd voor antilichamen tegen Agg in het voorkomen vanbiomateriaal gerelateerde infecties met enterokokken.Uit de literatuur is bekend dat enterokokken in de aanwezigheid van gal verschillendeeiwitten aanmaken en dat gal daardoor mogelijk de hechting van bacteriën kanbeïnvloeden. Daarom hebben we de invloed van het opgroeien in gal op de hechting van E.faecalis aan galdrain materiaal bestudeerd (Hoofdstuk 7). Omdat in de vorige studie wasaangetoond dat Esp en Agg de binding kunnen beïnvloeden, is gekozen voor E. faecalisstammen die deze eiwitten tot expressie brengen. De resultaten lieten zien dat na groei ingal twee keer zo veel E. faecalis met Esp en E. faecalis zonder Esp of Agg aan het oppervlakhechtten. Groei in gal had geen invloed op de hechting van E. faecalis met Agg op het oppervlak, waarschijnlijk omdat deze stam vooral via positieve coöperativiteit hecht ende gal hier geen invloed op heeft. Hechting aan een materiaal is de eerste stap naarbiomateriaal gerelateerde infecties, daarom kan worden gesteld dat groei in gal hetinfecterende vermogen van enterokokken vergroot.In Hoofdstuk 8 van dit proefschrift worden de bevindingen van het onderzoek samengevat,de algemene conclusies worden gegeven en een aantal mogelijkheden voor verderonderzoek worden aangevoerd. Een belangrijke conclusie uit dit onderzoek is dat het ergmoeilijk is om de virulentiefactoren te onderzoeken van een bacterie zoals E. faecalis dieniet altijd infecties veroorzaakt maar ook gewoon bij de gastheer aanwezig kan zijn zonderdat deze er last van heeft. Bij het ontstaan van infecties spelen daarom meer factoren eenrol waarmee ook rekening gehouden zou moeten worden bij de analyse. Zo is het ookmogelijk dat sommige enterokokken eigenschappen bezitten waardoor ze vooral beterkunnen overleven in de omgeving van het ziekenhuis en de darm van patiënten in hetziekenhuis. Zodra de afweer van de patiënten dan vermindert, kunnen deze enterokokkendan infecties veroorzaken. Toekomstig onderzoek zal meer gericht moeten zijn op hetopsporen van deze eigenschappen van E. faecalis, daarbij zal ook moeten worden gekekennaar de verschillende genogroepen zoals beschreven in dit proefschrift.
NED
ERLA
ND
SE S
AM
ENV
ATTI
NG
118

De belangrijkste conclusies van dit onderzoek zijn:
● Enterokokken kunnen worden verdeeld in genogroepen die zijn geassocieerd met eenspecifieke gastheer of omgeving en bepaalde virulentiefactoren.
● Het oppervlakte eiwit Agg en het enzym “cytolysine” van E. faecalis zijn geassocieerdmet infecties en het oppervlakte eiwit Esp met kolonisatie en verspreiding.
● Oppervlakte eiwitten Esp en Agg van E. faecalis spelen een rol bij de binding aanbiomaterialen en daarmee bij het ontstaan van infecties.
● Antilichamen tegen Agg hebben invloed op de interactie tussen E. faecalis bacteriën metAgg en verminderen daarmee de binding aan biomaterialen.
● Het opgroeien in de aanwezigheid van gal verhoogt de binding van bepaaldeenterokokken aan de onderzochte biomaterialen.
Slotwoord
De infecties met E. faecalis beperken zich tot op heden vooral tot patiënten met eenverminderde afweer of een ernstige onderliggende ziekte. Dit is echter geen reden om hetgevaar van deze infecties te onderschatten. E. faecalis wordt steeds vaker gevonden bijpatiënten in het ziekenhuis. Deze toenemende aanwezigheid in het ziekenhuis zou er toekunnen leiden dat E. faecalis virulentiefactoren van andere soorten bacteriën gaatovernemen en deze gaat combineren met de eigen virulentiefactoren. Een dergelijke stamzou meer virulent kunnen zijn dat de E. faecalis stammen die we tot nu toe kennen. Als dezestam dan ook nog resistent wordt tegen de meest gebruikte antibiotica, is een“superbacterie” ontstaan die mogelijk ook in staat is infecties te veroorzaken in gezondepersonen, die vervolgens moeilijk te behandelen zijn. Dat dit scenario mogelijk is, blijkt uitrecente publicaties over de overdracht van genetisch materiaal tussen E. faecalis en anderesoorten bacteriën en over het ontstaan van resistentie in voorheen gevoelige E. faecalisstammen. Daarom is het van belang om het onderzoek naar de virulentie van E. faecalisvoort te zetten met als doel nieuwe manieren te vinden om deze infecties te voorkómen enhet ontstaan van een dergelijke “superbacterie” voor te zijn.
119
CHAPTER
9


References


1. Amann, R. I., B. J. Binder, R. J. Olson, S. W. Chisholm, R. Devereux, and D. A. Stahl. 1990.Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzingmixed microbial populations. Appl. Environ. Microbiol. 56:1919-1925.
2. Amann, R. I., W. Ludwig, and K. H. Schleifer. 1995. Phylogenetic identification and in situdetection of individual microbial cells without cultivation. Microbiol. Rev. 59:143-169.
3. An, Y. H., and R. J. Friedman. 1998. Concise review of mechanisms of bacterial adhesion tobiomaterial surfaces. J. Biomed. Mater. Res. 43:338-348.
4. Antonishyn, N. A., R. R. McDonald, E. L. Chan, G. Horsman, C. E. Woodmansee, P. S. Falk, andC. G. Mayhall. 2000. Evaluation of fluorescence-based amplified fragment length polymorphismanalysis for molecular typing in hospital epidemiology: comparison with pulsed-field gelelectrophoresis for typing strains of vancomycin-resistant Enterococcus faecium. J. Clin.Microbiol. 38:4058-4065.
5. Arduino, R. C., B. E. Murray, and R. M. Rakita. 1994. Roles of antibodies and complement inphagocytic killing of enterococci. Infect. Immun. 62:987-993.
6. Arnow, P. M., K. C. Zachary, J. R. Thistlethwaite, K. D. Thompson, J. L. Bova, and K. A. Newell.1998. Pathogenesis of early operative site infections after orthotopic liver transplantation.Transplantation. 65:1500-1503.
7. Bakir, M., J. L. Bova, K. A. Newell, J. M. Millis, J. F. Buell, and P. M. Arnow. 2001. Epidemiologyand clinical consequences of vancomycin-resistant enterococci in liver transplant patients.Transplantation. 72:1032-1037.
8. Barie, P. S., N. V. Christou, E. P. Dellinger, W. R. Rout, H. H. Stone, and J. P. Waymack. 1990.Pathogenicity of the enterococcus in surgical infections. Ann. Surg. 212:155-159.
9. Barkholt, L., B. G. Ericzon, J. Tollemar, A. S. Malmborg, A. Ehrnst, H. Wilczek, and J. Andersson.1993. Infections in human liver recipients: different patterns early and late after transplantation.Transpl. Int. 6:77-84.
10. Basinger, S. F., and R. W. Jackson. 1968. Bacteriocin (hemolysin) of Streptococcus zymogenes. J.Bacteriol. 96:1895-1902.
11. Behr, T., C. Koob, M. Schedl, A. Mehlen, H. Meier, D. Knopp, E. Frahm, U. Obst, K. Schleifer, R.Niessner, and W. Ludwig. 2000. A nested array of rRNA targeted probes for the detection andidentification of enterococci by reverse hybridization. Syst. Appl. Microbiol. 23:563-572.
12. Beimfohr, C., A. Krause, R. I. Amann, W. Ludwig, and K. H. Schleifer. 1993. In situ identificationof lactococci, enterococci and streptococci. Syst. Appl. Microbiol. 16:450-456.
13. Ben-Ari, Z., L. Neville, B. Davidson, K. Rolles, and A. K. Burroughs. 1998. Infection rates withand without T-tube splintage of common bile duct anastomosis in liver transplantation. Transpl.Int. 11:123-126.
14. Berg, R. D. 1995. Bacterial translocation from the gastrointestinal tract. Trends. Microbiol. 3:149-154.
15. Berti, M., G. Candiani, A. Kaufhold, A. Muscholl, and R. Wirth. 1998. Does aggregation substanceof Enterococcus faecalis contribute to development of endocarditis? Infection. 26:48-53.
16. Betzl, D., W. Ludwig, and K. H. Schleifer. 1990. Identification of lactococci and enterococci bycolony hybridization with 23S rRNA-targeted oligonucleotide probes. Appl. Environ. Microbiol.56:2927-2929.
17. Bhakdi, S., T. Klonisch, P. Nuber, and W. Fischer. 1991. Stimulation of monokine production bylipoteichoic acids. Infect. Immun. 59:4614-4620.
123
REFERENCES

18. Binnig, G., C. F. Quate, and C. Gerber. 1986. Atomic force microscope. Physical review letters.56:930-933.
19. Bolshakova, A. V., O. I. Kiselyova, A. S. Filonov, O. Y. Frolova, Y. L. Lyubchenko, and I. V.Yaminsky. 2001. Comparative studies of bacteria with an atomic force microscopy operating indifferent modes. Ultramicroscopy. 86:121-128.
20. Bonten, M. J., C. A. Gaillard, F. H. van Tiel, S. van der Geest, and E. E. Stobberingh. 1995.Colonization and infection with Enterococcus faecalis in intensive care units: the role ofantimicrobial agents. Antimicrob. Agents Chemother. 39:2783-2786.
21. Boom, R., C. J. Sol, M. M. Salimans, C. L. Jansen, P. M. Wertheim-van Dillen, and J. van derNoordaa. 1990. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol.28:495-503.
22. Bos, R., H. C. van der Mei, and H. J. Busscher. 1999. Physico-chemistry of initial microbialadhesive interactions-its mechanisms and methods for study. FEMS Microbiol. Rev. 23:179-230.
23. Bouza, E., R. San Juan, P. Munoz, A. Voss, and J. Kluytmans. 2001. A European perspective onnosocomial urinary tract infections I. Report on the microbiology workload, etiology andantimicrobial susceptibility (ESGNI-003 study). European Study Group on Nosocomial Infections.Clin. Microbiol. Infect. 7:523-531.
24. Bouza, E., R. San Juan, P. Munoz, A. Voss, and J. Kluytmans. 2001. A European perspective onnosocomial urinary tract infections II. Report on incidence, clinical characteristics and outcome(ESGNI-004 study). European Study Group on Nosocomial Infection. Clin. Microbiol. Infect.7:532-542.
25. Bowen, B. D., S. Levine, and N. Epstein. 1976. Fine particles depostion in laminar flow throughparallel plate and cilindrical channels. J. Colloid Interface Sci. 54:375-390.
26. Bowen, W. R., A. S. Fenton, R. W. Lovitt, and C. J. Wright. 2002. The measurement of Bacillusmycoides spore adhesion using atomic force microscopy, simple counting methods, and aspinning disk technique. Biotechnol. Bioeng. 79:170-179.
27. Bruinsma, N., J. M. Hutchinson, H. Giamarellou, A. Boogaard van den, J. Degener, P. Smet de,and E. Stobberingh. 2000. Importance of Population Density on the Prevalence of AntibioticResistance in the Fecal Flora of Healthy Volunteers [Abstract 101]. American Society forMicrobiology. 40th Interscience Conference on Antimicrobial Agents and Chemotherapy.Toronto, Ontario, Canada.
28. Bruinsma, N., R. J. Willems, A. E. van den Bogaard, M. Santen-Verheuvel, N. London, C.Driessen, and E. E. Stobberingh. 2002. Different Levels of Genetic Homogeneity in Vancomycin-Resistant and -Susceptible Enterococcus faecium Isolates from Different Human and AnimalSources Analyzed by Amplified-Fragment Length Polymorphism. Antimicrob. Agents Chemother.46:2779-2783.
29. Burnett, R. J., D. C. Haverstock, E. P. Dellinger, H. H. Reinhart, J. M. Bohnen, O. D. Rotstein, S.B. Vogel, and J. S. Solomkin. 1995. Definition of the role of enterococcus in intraabdominalinfection: analysis of a prospective randomized trial. Surgery. 118:716-723.
30. Busscher, H. J., R. Bos, H. C. van der Mei, and P. S. Handley. 2000. Physicochemistry of MicrobialAdhesion From an Overall Approach to the Limits., p. 431-458. In A. Baszkin and W. Norde (ed.),Physical Chemistry of Biological Interfaces. Marcel Dekker, New York, Basel.
31. Busscher, H. J., and H. C. van der Mei. 1995. Use of flow chamber devices and image analysismethods to study microbial adhesion. Methods Enzymol. 253:455-477.
32. Busscher, H. J., A. H. Weerkamp, H. C. van der Mei, A. W. van Pelt, H. P. de Jong, and J. Arends.
124

1984. Measurement of the surface free energy of bacterial cell surfaces and its relevance foradhesion. Appl. Environ. Microbiol. 48:980-983.
33. Caballero-Granado, F. J., B. Becerril, J. M. Cisneros, L. Cuberos, I. Moreno, and J. Pachon. 2001.Case-control study of risk factors for the development of enterococcal bacteremia. Eur. J. Clin.Microbiol. Infect. Dis. 20:83-90.
34. Caballero-Granado, F. J., B. Becerril, L. Cuberos, M. Bernabeu, J. M. Cisneros, and J. Pachon.2001. Attributable mortality rate and duration of hospital stay associated with enterococcalbacteremia. Clin. Infect. Dis. 32:587-594.
35. Caballero-Granado, F. J., J. M. Cisneros, R. Luque, M. Torres-Tortosa, F. Gamboa, F. Diez, J. L.Villanueva, R. Perez-Cano, J. Pasquau, D. Merino, A. Menchero, D. Mora, M. A. Lopez-Ruz, andA. Vergara. 1998. Comparative study of bacteremias caused by Enterococcus spp. with andwithout high-level resistance to gentamicin. J. Clin. Microbiol. 36:520-525.
36. Casadevall, A., and L. A. Pirofski. 1999. Host-pathogen interactions: redefining the basicconcepts of virulence and pathogenicity. Infect. Immun. 67:3703-3713.
37. Chenoweth, C., and D. Schaberg. 1990. The epidemiology of enterococci. Eur. J. Clin. Microbiol.Infect. Dis. 9:80-89.
38. Chow, J. W., A. Davidson, E. Sanford, III, and M. J. Zervos. 1997. Superinfection withEnterococcus faecalis during quinupristin/dalfopristin therapy. Clin. Infect. Dis. 24:91-92.
39. Chow, J. W., L. A. Thal, M. B. Perri, J. A. Vazquez, S. M. Donabedian, D. B. Clewell, and M. J.Zervos. 1993. Plasmid-associated hemolysin and aggregation substance production contribute tovirulence in experimental enterococcal endocarditis. Antimicrob. Agents Chemother. 37:2474-2477.
40. Chylak, J., W. Michalska, M. Drews, R. Marciniak, and M. Lange. 2000. Comparison of bacterialflora found in the peritoneal cavity and drains after intraabdominal surgery. Med. Sci. Monit.6:285-290.
41. Clewell, D. B. 1993. Bacterial sex pheromone-induced plasmid transfer. Cell. 73:9-12.
42. Clewell, D. B., F. Y. An, B. A. White, and C. Gawron-Burke. 1985. Streptococcus faecalis sexpheromone (cAM373) also produced by Staphylococcus aureus and identification of aconjugative transposon (Tn918). J. Bacteriol. 162:1212-1220.
43. Coburn, P. S., and M. S. Gilmore. 2003. The Enterococcus faecalis cytolysin: a novel toxin activeagainst eukaryotic and prokaryotic cells. Cell Microbiol. 5:661-669.
44. Coque, T. M., J. E. Patterson, J. M. Steckelberg, and B. E. Murray. 1995. Incidence of hemolysin,gelatinase, and aggregation substance among enterococci isolated from patients with endocarditisand other infections and from feces of hospitalized and community-based persons. J. Infect. Dis.171:1223-1229.
45. Costerton, J. W., P. S. Stewart, and E. P. Greenberg. 1999. Bacterial biofilms: a common cause ofpersistent infections. Science. 284:1318-1322.
46. Creti, R., M. Imperi, L. Bertuccini, F. Fabretti, G. Orefici, R. Di Rosa, and L. Baldassarri. 2004.Survey for virulence determinants among Enterococcus faecalis isolated from different sources. J.Med. Microbiol. 53:13-20.
47. Cummings, C. A., and D. A. Relman. 2000. Using DNA microarrays to study host-microbeinteractions. Emerg. Infect. Dis. 6:513-525.
48. Curry, N., R. W. McCallum, and P. H. Guth. 1974. Spontaneous peritonitis in cirrhotic ascites. Adecade of experience. Am. J. Dig. Dis. 19:685-692.
125
REFERENCES

49. Dabros, T., and T. G. M. van de Ven. 1982. Kinetics of coating by colloid particles. J. ColloidInterface Sci. 89:232-244.
50. De Boever, E. H., D. B. Clewell, and C. M. Fraser. 2000. Enterococcus faecalis conjugativeplasmid pAM373: complete nucleotide sequence and genetic analyses of sex pheromoneresponse. Mol. Microbiol. 37:1327-1341.
51. Degener, J. E., and I. Phillips. 2001. Comparison of antimicrobial susceptibility test breakpoints ofnational societies. Clin. Microbiol. Infect. 7:51-54.
52. Desai, M., A. Tanna, R. Wall, A. Efstratiou, R. George, and J. Stanley. 1998. Fluorescentamplified-fragment length polymorphism analysis of an outbreak of group A streptococcalinvasive disease. J. Clin. Microbiol. 36:3133-3137.
53. Dickinson, G. M., and A. L. Bisno. 1989. Infections associated with indwelling devices: infectionsrelated to extravascular devices. Antimicrob. Agents Chemother. 33:602-607.
54. Donlan, R. M., and J. W. Costerton. 2002. Biofilms: survival mechanisms of clinically relevantmicroorganisms. Clin. Microbiol. Rev. 15:167-193.
55. Dougherty, S. H. 1984. Role of enterococcus in intraabdominal sepsis. Am. J. Surg. 148:308-312.
56. Doyle, R. J. 1991. Strategies in experimental microbial adhesion research, p. 291-316. In N.Mozes, P. S. Handley, H. J. Busscher, and P. G. Rouxhet (ed.), Microbial Cell Surface Analysis -Structural and Physicochemical Methods. VCH Publishers Inc., New York.
57. Doyle, R. J. 2000. Contribution of the hydrophobic effect to microbial infection. Microbes. Infect.2:391-400.
58. Doyle, R. J., W. E. Nesbitt, and K. G. Taylor. 1982. On the mechanism of adhesion ofStreptococcus sanguis to hydroxylapatite. FEMS Microbiol. Lett. 15:1-5.
59. Dufrene, Y. F. 2000. Direct characterization of the physicochemical properties of fungal sporesusing functionalized AFM probes. Biophys. J. 78:3286-3291.
60. Dufrene, Y. F. 2002. Atomic force microscopy, a powerful tool in microbiology. J. Bacteriol.184:5205-5213.
61. Dufrene, Y. F. 2003. Recent progress in the application of atomic force microscopy imaging andforce spectroscopy to microbiology. Curr. Opin. Microbiol. 6:317-323.
62. Duh, R. W., K. V. Singh, K. Malathum, and B. E. Murray. 2001. In vitro activity of 19 anti-microbial agents against enterococci from healthy subjects and hospitalized patients and use of anace gene probe from Enterococcus faecalis for species identification. Microb. Drug Resist. 7:39-46.
63. Dunny, G., C. Funk, and J. Adsit. 1981. Direct stimulation of the transfer of antibiotic resistanceby sex pheromones in Streptococcus faecalis. Plasmid. 6:270-278.
64. Dunny, G. M. 1990. Genetic functions and cell-cell interactions in the pheromone- inducibleplasmid transfer system of Enterococcus faecalis. Mol. Microbiol. 4:689-696.
65. Dunny, G. M., B. L. Brown, and D. B. Clewell. 1978. Induced cell aggregation and mating inStreptococcus faecalis: evidence for a bacterial sex pheromone. Proc. Natl. Acad. Sci. U. S. A.75:3479-3483.
66. Dunny, G. M., B. A. Leonard, and P. J. Hedberg. 1995. Pheromone-inducible conjugation inEnterococcus faecalis: interbacterial and host-parasite chemical communication. J. Bacteriol.177:871-876.
67. Dupont, H., P. Montravers, J. Mohler, and C. Carbon. 1998. Disparate findings on the role of
126

virulence factors of Enterococcus faecalis in mouse and rat models of peritonitis. Infect. Immun.66:2570-2575.
68. Dupre, I., S. Zanetti, A. M. Schito, G. Fadda, and L. A. Sechi. 2003. Incidence of virulencedeterminants in clinical Enterococcus faecium and Enterococcus faecalis isolates collected inSardinia (Italy). J. Med. Microbiol. 52:491-498.
69. Eaton, T. J., and M. J. Gasson. 2001. Molecular screening of Enterococcus virulence determinantsand potential for genetic exchange between food and medical isolates. Appl. Environ. Microbiol.67:1628-1635.
70. Edmond, M. B., J. F. Ober, J. D. Dawson, D. L. Weinbaum, and R. P. Wenzel. 1996. Vancomycin-resistant enterococcal bacteremia: natural history and attributable mortality. Clin. Infect. Dis.23:1234-1239.
71. Eliopoulos, G. M. 1993. Increasing problems in the therapy of enterococcal infections. Eur. J. Clin.Microbiol. Infect. Dis. 12:409-412.
72. Elsner, H. A., I. Sobottka, D. Mack, M. Claussen, R. Laufs, and R. Wirth. 2000. Virulence factorsof Enterococcus faecalis and Enterococcus faecium blood culture isolates. Eur. J. Clin. Microbiol.Infect. Dis. 19:39-42.
73. Facklam, R. R., and M. D. Collins. 1989. Identification of Enterococcus species isolated fromhuman infections by a conventional test scheme. J. Clin. Microbiol. 27:731-734.
74. Facklam, R. R., Sahm, D. F., and Teixeira, L. M. 1999. Enterococcus, p. 297-305. In P. R. Murray,E. J. Baron, M. A. Pfaller, F. C. Tenover, and R. H. Yolken (ed.), Manual of Clinical Microbiology.ASM Press, Washington D.C.
75. Felmingham, D., A. P. Wilson, A. I. Quintana, and R. N. Gruneberg. 1992. Enterococcus speciesin urinary tract infection. Clin. Infect. Dis. 15:295-301.
76. Flahaut, S., J. Frere, P. Boutibonnes, and Y. Auffray. 1996. Comparison of the bile salts andsodium dodecyl sulfate stress responses in Enterococcus faecalis. Appl. Environ. Microbiol.62:2416-2420.
77. Fluit, A. C., M. E. Jones, F. J. Schmitz, J. Acar, R. Gupta, and J. Verhoef. 2000. Antimicrobialsusceptibility and frequency of occurrence of clinical blood isolates in Europe from the SENTRYantimicrobial surveillance program, 1997 and 1998. Clin. Infect. Dis. 30:454-460.
78. Fluit, A. C., F. J. Schmitz, and J. Verhoef. 2001. Frequency of isolation of pathogens frombloodstream, nosocomial pneumonia, skin and soft tissue, and urinary tract infections occurring inEuropean patients. Eur. J. Clin. Microbiol. Infect. Dis. 20:188-191.
79. Fluit, A. C., J. Verhoef, and F. J. Schmitz. 2001. Frequency of isolation and antimicrobial resistanceof gram-negative and gram-positive bacteria from patients in intensive care units of 25 Europeanuniversity hospitals participating in the European arm of the SENTRY Antimicrobial SurveillanceProgram 1997-1998. Eur. J. Clin. Microbiol. Infect. Dis. 20:617-625.
80. Francia, M. V., W. Haas, R. Wirth, E. Samberger, A. Muscholl-Silberhorn, M. S. Gilmore, Y. Ike,K. E. Weaver, F. Y. An, and D. B. Clewell. 2001. Completion of the nucleotide sequence of theEnterococcus faecalis conjugative virulence plasmid pAD1 and identification of a second transferorigin. Plasmid. 46:117-127.
81. Franke, A. E., and D. B. Clewell. 1981. Evidence for a chromosome-borne resistance transposon(Tn916) in Streptococcus faecalis that is capable of “conjugal” transfer in the absence of aconjugative plasmid. J. Bacteriol. 145:494-502.
82. Franks, A. H., H. J. Harmsen, G. C. Raangs, G. J. Jansen, F. Schut, and G. W. Welling. 1998.Variations of bacterial populations in human feces measured by fluorescent in situ hybridization
127
REFERENCES

with group-specific 16S rRNA-targeted oligonucleotide probes. Appl. Environ. Microbiol.64:3336-3345.
83. Franz, C. M., A. B. Muscholl-Silberhorn, N. M. Yousif, M. Vancanneyt, J. Swings, and W. H.Holzapfel. 2001. Incidence of virulence factors and antibiotic resistance among enterococciisolated from food. Appl. Environ. Microbiol. 67:4385-4389.
84. Gaglani, M. J., C. J. Baker, and M. S. Edwards. 1997. Contribution of antibody to neutrophil-mediated killing of Enterococcus faecalis. J. Clin. Immunol. 17:478-484.
85. Gallardo-Moreno, A. M., M. L. Gonzalez-Martin, C. Perez-Giraldo, J. M. Bruque, and A. C.Gomez-Garcia. 2002. Serum as a factor influencing adhesion of Enterococcus faecalis to glass andsilicone. Appl. Environ. Microbiol. 68:5784-5787.
86. Gallardo-Moreno, A. M., H. C. van der Mei, H. J. Busscher, M. L. Gonzalez-Martin, J. M. Bruque,and C. Perez-Giraldo. 2001. Adhesion of Enterococcus faecalis 1131 grown under subinhibitoryconcentrations of ampicillin and vancomycin to a hydrophilic and a hydrophobic substratum.FEMS Microbiol. Lett. 203:75-79.
87. Galli, D., F. Lottspeich, and R. Wirth. 1990. Sequence analysis of Enterococcus faecalisaggregation substance encoded by the sex pheromone plasmid pAD1. Mol. Microbiol. 4:895-904.
88. Galli, D., and R. Wirth. 1991. Comparative analysis of Enterococcus faecalis sex pheromoneplasmids identifies a single homologous DNA region which codes for aggregation substance. J.Bacteriol. 173:3029-3033.
89. Galli, D., R. Wirth, and G. Wanner. 1989. Identification of aggregation substances ofEnterococcus faecalis cells after induction by sex pheromones. An immunological andultrastructural investigation. Arch. Microbiol. 151:486-490.
90. Garrison, R. N., D. E. Fry, S. Berberich, and H. C. Polk, Jr. 1982. Enterococcal bacteremia: clinicalimplications and determinants of death. Ann. Surg. 196:43-47.
91. Garsin, D. A., C. D. Sifri, E. Mylonakis, X. Qin, K. V. Singh, B. E. Murray, S. B. Calderwood, andF. M. Ausubel. 2001. A simple model host for identifying Gram-positive virulence factors. Proc.Natl. Acad. Sci. U. S. A. 98:10892-10897.
92. Gentry-Weeks, C. R., R. Karkhoff-Schweizer, A. Pikis, M. Estay, and J. M. Keith. 1999. Survivalof Enterococcus faecalis in mouse peritoneal macrophages. Infect. Immun. 67:2160-2165.
93. George, D. L., P. M. Arnow, A. S. Fox, A. L. Baker, J. R. Thistlethwaite, J. C. Emond, P. F.Whitington, and C. E. Broelsch. 1991. Bacterial infection as a complication of livertransplantation: epidemiology and risk factors. Rev. Infect. Dis. 13:387-396.
94. Gilad, J., A. Borer, K. Riesenberg, N. Peled, A. Shnaider, and F. Schlaeffer. 1998. Enterococcushirae septicemia in a patient with end-stage renal disease undergoing hemodialysis. Eur. J. Clin.Microbiol. Infect. Dis. 17:576-577.
95. Gilmore, M. S., Coburn, P. S., Nallapareddy, S. R., and Murray, B. E. 2002. Enterococcal virulence,p. 301-354. In M. S. Gilmore, D. B. Clewell, P. Courvalin, G. M. Dunny, B. E. Murray, and L. B. Rice(ed.), The Enterococci. Pathogenesis, Molecular Biology, and Antibiotic Resistance. ASM Press,Washington D.C.
96. Gilmore, M. S., and J. J. Ferretti. 2003. Microbiology. The thin line between gut commensal andpathogen. Science. 299:1999-2002.
97. Gray, J., P. J. Marsh, D. Stewart, and S. J. Pedler. 1994. Enterococcal bacteraemia: a prospectivestudy of 125 episodes. J. Hosp. Infect. 27:179-186.
128

98. Gross, P. A., L. M. Harkavy, G. E. Barden, and M. F. Flower. 1976. The epidemiology ofnosocomial enterococcal urinary tract infection. Am. J. Med. Sci. 272:75-81.
99. Gullberg, R. M., S. R. Homann, and J. P. Phair. 1989. Enterococcal bacteremia: analysis of 75episodes. Rev. Infect. Dis. 11:74-85.
100. Gutschik, E., S. Moller, and N. Christensen. 1979. Experimental endocarditis in rabbits. 3.Significance of the proteolytic capacity of the infecting strains of Streptococcus faecalis. ActaPathol. Microbiol. Scand. [B. ]. 87:353-362.
101. Guzman, C. A., C. Pruzzo, G. LiPira, and L. Calegari. 1989. Role of adherence in pathogenesisof Enterococcus faecalis urinary tract infection and endocarditis. Infect. Immun. 57:1834-1838.
102. Guzman, C. A., C. Pruzzo, M. Plate, M. C. Guardati, and L. Calegari. 1991. Serum dependentexpression of Enterococcus faecalis adhesins involved in the colonization of heart cells. Microb.Pathog. 11:399-409.
103. Haas, W., B. D. Shepard, and M. S. Gilmore. 2002. Two-component regulator of Enterococcusfaecalis cytolysin responds to quorum-sensing autoinduction. Nature. 415:84-87.
104. Hancock, L. E., and M. S. Gilmore. 2002. The capsular polysaccharide of Enterococcus faecalisand its relationship to other polysaccharides in the cell wall. Proc. Natl. Acad. Sci. U. S. A.99:1574-1579.
105. Harmsen, H. J. M., P. Elfferich, F. Schut, and G. W. Welling. 1999. A 16S rRNA-targeted probefor detection of lactobacilli and enterococci in faecal samples by fluorescent in situ hybridization.Microb. Ecol. Health Dis. 11:3-12.
106. Harvey, B. S., C. J. Baker, and M. S. Edwards. 1992. Contributions of complement andimmunoglobulin to neutrophil-mediated killing of enterococci. Infect. Immun. 60:3635-3640.
107. Hinterdorfer, P., W. Baumgartner, H. J. Gruber, K. Schilcher, and H. Schindler. 1996. Detectionand localization of individual antibody-antigen recognition events by atomic force microscopy.Proc. Natl. Acad. Sci. U. S. A. 93:3477-3481.
108. Hirt, H., P. M. Schlievert, and G. M. Dunny. 2002. In vivo Induction of virulence and antibioticresistance transfer in Enterococcus faecalis mediated by the sex pheromone-sensing system ofpCF10. Infect. Immun. 70:716-723.
109. Holdeman, L. V., E. P. Cato, and W. E. C Moore. 1977. L. V. Holdeman, E. P. Cato, and W. E. C.Moore (ed.), Anaerobe laboratory manual. Virginia Polytechnic Institute and State University,Blacksburg, Va.
110. Huebner, J., A. Quaas, W. A. Krueger, D. A. Goldmann, and G. B. Pier. 2000. Prophylactic andtherapeutic efficacy of antibodies to a capsular polysaccharide shared among vancomycin-sensitive and -resistant enterococci. Infect. Immun. 68:4631-4636.
111. Huebner, J., Y. Wang, W. A. Krueger, L. C. Madoff, G. Martirosian, S. Boisot, D. A. Goldmann,D. L. Kasper, A. O. Tzianabos, and G. B. Pier. 1999. Isolation and chemical characterization of acapsular polysaccharide antigen shared by clinical isolates of Enterococcus faecalis andvancomycin-resistant Enterococcus faecium. Infect. Immun. 67:1213-1219.
112. Huycke, M. M., and M. S. Gilmore. 1995. Frequency of aggregation substance and cytolysingenes among enterococcal endocarditis isolates. Plasmid. 34:152-156.
113. Huycke, M. M., D. F. Sahm, and M. S. Gilmore. 1998. Multiple-drug resistant enterococci: thenature of the problem and an agenda for the future. Emerg. Infect. Dis. 4:239-249.
114. Huycke, M. M., C. A. Spiegel, and M. S. Gilmore. 1991. Bacteremia caused by hemolytic, high-level gentamicin-resistant Enterococcus faecalis. Antimicrob. Agents Chemother. 35:1626-1634.
129
REFERENCES

115. Ike, Y., and D. B. Clewell. 1992. Evidence that the hemolysin/bacteriocin phenotype ofEnterococcus faecalis subsp. zymogenes can be determined by plasmids in differentincompatibility groups as well as by the chromosome. J. Bacteriol. 174:8172-8177.
116. Ike, Y., R. A. Craig, B. A. White, Y. Yagi, and D. B. Clewell. 1983. Modification of Streptococcusfaecalis sex pheromones after acquisition of plasmid DNA. Proc. Natl. Acad. Sci. U. S. A.80:5369-5373.
117. Ike, Y., H. Hashimoto, and D. B. Clewell. 1984. Hemolysin of Streptococcus faecalis subspecieszymogenes contributes to virulence in mice. Infect. Immun. 45:528-530.
118. Ike, Y., H. Hashimoto, and D. B. Clewell. 1987. High incidence of hemolysin production byEnterococcus (Streptococcus) faecalis strains associated with human parenteral infections. J.Clin. Microbiol. 25:1524-1528.
119. Isenmann, R., M. Schwarz, E. Rozdzinski, R. Marre, and H. G. Beger. 2000. Aggregationsubstance promotes colonic mucosal invasion of Enterococcus faecalis in an ex vivo model. J.Surg. Res. 89:132-138.
120. Jacob, A. E., and S. J. Hobbs. 1974. Conjugal transfer of plasmid-borne multiple antibioticresistance in Streptococcus faecalis var. zymogenes. J. Bacteriol. 117:360-372.
121. Jansen, G. J., M. Mooibroek, J. Idema, H. J. Harmsen, G. W. Welling, and J. E. Degener. 2000.Rapid identification of bacteria in blood cultures by using fluorescently labeled oligonucleotideprobes. J. Clin. Microbiol. 38:814-817.
122. Jett, B. D., R. V. Atkuri, and M. S. Gilmore. 1998. Enterococcus faecalis localization inexperimental endophthalmitis: role of plasmid-encoded aggregation substance. Infect. Immun.66:843-848.
123. Jett, B. D., M. M. Huycke, and M. S. Gilmore. 1994. Virulence of enterococci. Clin. Microbiol.Rev. 7:462-478.
124. Jett, B. D., H. G. Jensen, R. E. Nordquist, and M. S. Gilmore. 1992. Contribution of the pAD1-encoded cytolysin to the severity of experimental Enterococcus faecalis endophthalmitis. Infect.Immun. 60:2445-2452.
125. Johnson, A. P. 1994. The pathogenicity of enterococci. J. Antimicrob. Chemother. 33:1083-1089.
126. Jones, M. E., F. J. Schmitz, A. C. Fluit, J. Acar, R. Gupta, and J. Verhoef. 1999. Frequency ofoccurrence and antimicrobial susceptibility of bacterial pathogens associated with skin and softtissue infections during 1997 from an International Surveillance Programme. SENTRYParticipants Group. Eur. J. Clin. Microbiol. Infect. Dis. 18:403-408.
127. Joyanes, P., A. Pascual, L. Martinez-Martinez, A. Hevia, and E. J. Perea. 1999. In vitroadherence of Enterococcus faecalis and Enterococcus faecium to plastic biomaterials. Clin.Microbiol. Infect. 5:382-386.
128. Kaufhold, A., and P. Ferrieri. 1991. Isolation of Enterococcus mundtii from normally sterile bodysites in two patients. J. Clin. Microbiol. 29:1075-1077.
129. Kawalec, M., M. Gniadkowski, M. Zaleska, T. Ozorowski, L. Konopka, and W. Hryniewicz.2001. Outbreak of vancomycin-resistant Enterococcus faecium of the phenotype VanB in ahospital in Warsaw, Poland: probable transmission of the resistance determinants into anendemic vancomycin-susceptible strain. J. Clin. Microbiol. 39:1781-1787.
130. Koivusalo, A., H. Makisalo, M. Talja, A. Siitonen, J. Vuopio-Varkila, M. Ruutu, and K.Hockerstedt. 1996. Bacterial adherence and biofilm formation on latex and silicone T-tubes inrelation to bacterial contamination of bile. Scand. J. Gastroenterol. 31:398-403.
130

131. Korten, V., and B. E. Murray. 1993. The nosocomial transmission of enterococci. Curr. Opin.Infect. Dis. 6:498-505.
132. Kreft, B., R. Marre, U. Schramm, and R. Wirth. 1992. Aggregation substance of Enterococcusfaecalis mediates adhesion to cultured renal tubular cells. Infect. Immun. 60:25-30.
133. Kristich, C. J., Y. H. Li, D. G. Cvitkovitch, and G. M. Dunny. 2004. Esp-independent biofilmformation by Enterococcus faecalis. J. Bacteriol. 186:154-163.
134. Kuhn, I., L. G. Burman, S. Haeggman, K. Tullus, and B. E. Murray. 1995. Biochemicalfingerprinting compared with ribotyping and pulsed-field gel electrophoresis of DNA forepidemiological typing of enterococci. J. Clin. Microbiol. 33:2812-2817.
135. Kusne, S., J. S. Dummer, N. Singh, S. Iwatsuki, L. Makowka, C. Esquivel, A. G. Tzakis, T. E.Starzl, and M. Ho. 1988. Infections after liver transplantation. An analysis of 101 consecutivecases. Medicine (Baltimore). 67:132-143.
136. Lambert, P. A., P. J. Shorrock, E. J. Aitchison, P. A. Domingue, M. E. Power, and J. W. Costerton.1990. Effect of in vivo growth conditions upon expression of surface protein antigens inEnterococcus faecalis. FEMS Microbiol. Immunol. 2:51-54.
137. Landry, S. L., D. L. Kaiser, and R. P. Wenzel. 1989. Hospital stay and mortality attributed tonosocomial enterococcal bacteremia: a controlled study. Am. J. Infect. Control. 17:323-329.
138. Lewis, C. M., and M. J. Zervos. 1990. Clinical manifestations of enterococcal infection. Eur. J.Clin. Microbiol. Infect. Dis. 9:111-117.
139. Libertin, C. R., R. Dumitru, and D. S. Stein. 1992. The hemolysin/bacteriocin produced byenterococci is a marker of pathogenicity. Diagn. Microbiol. Infect. Dis. 15:115-120.
140. Linden, P. K., A. W. Pasculle, R. Manez, D. J. Kramer, J. J. Fung, A. D. Pinna, and S. Kusne.1996. Differences in outcomes for patients with bacteremia due to vancomycin- resistantEnterococcus faecium or vancomycin- susceptible E. faecium. Clin. Infect. Dis. 22:663-670.
141. Livornese, L. L., Jr., S. Dias, C. Samel, B. Romanowski, S. Taylor, P. May, P. Pitsakis, G. Woods,D. Kaye, M. E. Levison, and . 1992. Hospital-acquired infection with vancomycin-resistantEnterococcus faecium transmitted by electronic thermometers. Ann. Intern. Med. 117:112-116.
142. Low, D. E., N. Keller, A. Barth, and R. N. Jones. 2001. Clinical prevalence, antimicrobialsusceptibility, and geographic resistance patterns of enterococci: results from the sentryantimicrobial surveillance program, 1997-1999. Clin. Infect. Dis. 32 Suppl 2:S133-S145.
143. Lowe, A. M., P. A. Lambert, and A. W. Smith. 1995. Cloning of an Enterococcus faecalisendocarditis antigen: homology with adhesins from some oral streptococci. Infect. Immun.63:703-706.
144. Lower, S. K., M. F. Hochella, Jr., and T. J. Beveridge. 2001. Bacterial recognition of mineralsurfaces: nanoscale interactions between Shewanella and alpha-FeOOH. Science. 292:1360-1363.
145. Ludwig, W., O. Strunk, S. Klugbauer, N. Klugbauer, M. Weizenegger, J. Neumaier, M.Bachleitner, and K. H. Schleifer. 1998. Bacterial phylogeny based on comparative sequenceanalysis. Electrophoresis. 19:554-568.
146. Lumbreras, C., M. Lizasoain, E. Moreno, J. M. Aguado, R. Gomez, I. Garcia, I. Gonzalez, C.Loinaz, C. Cisneros, and A. R. Noriega. 1992. Major bacterial infections following livertransplantation: a prospective study. Hepatogastroenterology. 39:362-365.
147. Maidak, B. L., G. J. Olsen, N. Larsen, R. Overbeek, M. J. McCaughey, and C. R. Woese. 1997.The RDP (Ribosomal Database Project). Nucleic Acids Res. 25:109-111.
131
REFERENCES

148. Maki, D. G., and W. A. Agger. 1988. Enterococcal bacteremia: clinical features, the risk ofendocarditis, and management. Medicine (Baltimore). 67:248-269.
149. Makinen, P. L., D. B. Clewell, F. An, and K. K. Makinen. 1989. Purification and substratespecificity of a strongly hydrophobic extracellular metalloendopeptidase (“gelatinase”) fromStreptococcus faecalis (strain 0G1-10). J. Biol. Chem. 264:3325-3334.
150. Malone, D. A., R. A. Wagner, J. P. Myers, and C. Watanakunakorn. 1986. Enterococcalbacteremia in two large community teaching hospitals. Am. J. Med. 81:601-606.
151. Mandell, G. L., D. Kaye, M. E. Levison, and E. W. Hook. 1970. Enterococcal endocarditis. Ananalysis of 38 patients observed at the New York Hospital-Cornell Medical Center. Arch. Intern.Med. 125:258-264.
152. Matlow, A. G., J. M. Bohnen, C. Nohr, N. Christou, and J. Meakins. 1989. Pathogenicity ofenterococci in a rat model of fecal peritonitis. J. Infect. Dis. 160:142-145.
153. McCormick, J. K., H. Hirt, G. M. Dunny, and P. M. Schlievert. 2000. Pathogenic mechanisms ofenterococcal endocarditis. Curr. Infect. Dis. Rep. 2:315-321.
154. McCormick, J. K., H. Hirt, C. M. Waters, T. J. Tripp, G. M. Dunny, and P. M. Schlievert. 2001.Antibodies to a surface-exposed, N-terminal domain of aggregation substance are not protectivein the rabbit model of Enterococcus faecalis infective endocarditis. Infect. Immun. 69:3305-3314.
155. McCormick, J. K., T. J. Tripp, G. M. Dunny, and P. M. Schlievert. 2002. Formation of vegetationsduring infective endocarditis excludes binding of bacterial-specific host antibodies toEnterococcus faecalis. J. Infect. Dis. 185:994-997.
156. Megran, D. W. 1992. Enterococcal endocarditis. Clin. Infect. Dis. 15:63-71.
157. Millsap, K. W., R. Bos, H. J. Busscher, and H. C. van der Mei. 1999. Surface aggregation ofCandida albicans on glass in the absence and presence of adhering Streptococcus gordonii in aparallel-plate flow chamber: A surface thermodynamical analysis based on acid-baseinteractions. J. Colloid Interface Sci. 212:495-502.
158. Miyazaki, S., A. Ohno, I. Kobayashi, T. Uji, K. Yamaguchi, and S. Goto. 1993. Cytotoxic effectof hemolytic culture supernatant from Enterococcus faecalis on mouse polymorphonuclearneutrophils and macrophages. Microbiol. Immunol. 37:265-270.
159. Moellering, R. C., Jr. 1992. Emergence of Enterococcus as a significant pathogen. Clin. Infect.Dis. 14:1173-1176.
160. Molinari, G., V. Pugliese, G. C. Schito, and C. A. Guzman. 1996. Bacteria involved in theblockage of biliary stents and their susceptibility to antibacterial agents. Eur. J. Clin. Microbiol.Infect. Dis. 15:88-92.
161. Monstein, H. J., M. Quednau, A. Samuelsson, S. Ahrne, B. Isaksson, and J. Jonasson. 1998.Division of the genus Enterococcus into species groups using PCR- based molecular typingmethods. Microbiology. 144:1171-1179.
162. Montravers, P., A. Andremont, L. Massias, and C. Carbon. 1994. Investigation of the potentialrole of Enterococcus faecalis in the pathophysiology of experimental peritonitis. J. Infect. Dis.169:821-830.
163. Montravers, P., J. Mohler, J. L. Saint, and C. Carbon. 1997. Evidence of the proinflammatory roleof Enterococcus faecalis in polymicrobial peritonitis in rats. Infect. Immun. 65:144-149.
164. Moore, W. E. C., and L. V. Holdeman. 1974. Human fecal flora:The normal flora of 20 Japanese-Hawaiians. Applied Microbiology. 27:961-979.
132

165. Morrison, A. J., Jr., and R. P. Wenzel. 1986. Nosocomial urinary tract infections due toEnterococcus. Ten years’ experience at a university hospital. Arch. Intern. Med. 146:1549-1551.
166. Mulin, B., P. Bailly, M. Thouverez, V. Cailleaux, C. Cornette, M. J. Dupont, and D. Talon. 1999.Clinical and molecular epidemiology of hospital Enterococcus faecalis isolates in eastern France.Clin Microbiol Infect. 5:149-157.
167. Mundy, L. M., D. F. Sahm, and M. Gilmore. 2000. Relationships between enterococcal virulenceand antimicrobial resistance. Clin. Microbiol. Rev. 13:513-522.
168. Munson, E. L., D. J. Diekema, S. E. Beekmann, K. C. Chapin, and G. V. Doern. 2003. Detectionand treatment of bloodstream infection: laboratory reporting and antimicrobial management. J.Clin. Microbiol. 41:495-497.
169. Murray, B. E. 1990. The life and times of the Enterococcus. Clin. Microbiol. Rev. 3:46-65.
170. Murray, B. E. 1997. Vancomycin-resistant enterococci. Am. J. Med. 102:284-293.
171. Murray, B. E., K. V. Singh, J. D. Heath, B. R. Sharma, and G. M. Weinstock. 1990. Comparisonof genomic DNAs of different enterococcal isolates using restriction endonucleases withinfrequent recognition sites. J. Clin. Microbiol. 28:2059-2063.
172. Murray, B. E., K. V. Singh, S. M. Markowitz, H. A. Lopardo, J. E. Patterson, M. J. Zervos, E.Rubeglio, G. M. Eliopoulos, L. B. Rice, and F. W. Goldstein. 1991. Evidence for clonal spread ofa single strain of beta-lactamase- producing Enterococcus (Streptococcus) faecalis to sixhospitals in five states. J. Infect. Dis. 163:780-785.
173. Muscholl, A., D. Galli, G. Wanner, and R. Wirth. 1993. Sex pheromone plasmid pAD1-encodedaggregation substance of Enterococcus faecalis is positively regulated in trans by traE1. Eur. J.Biochem. 214:333-338.
174. Muscholl-Silberhorn, A. 1998. Analysis of the clumping-mediating domain(s) of sex pheromoneplasmid pAD1-encoded aggregation substance. Eur. J. Biochem. 258:515-520.
175. Muscholl-Silberhorn, A. 1999. Cloning and functional analysis of Asa373, a novel adhesinunrelated to the other sex pheromone plasmid-encoded aggregation substances of Enterococcusfaecalis. Mol. Microbiol. 34:620-630.
176. Muscholl-Silberhorn, A., E. Samberger, and R. Wirth. 1997. Why does Staphylococcus aureussecrete an Enterococcus faecalis- specific pheromone? FEMS Microbiol. Lett. 157:261-266.
177. Muto, C. A., J. A. Jernigan, B. E. Ostrowsky, H. M. Richet, W. R. Jarvis, J. M. Boyce, and B. M.Farr. 2003. SHEA guideline for preventing nosocomial transmission of multidrug-resistant strainsof Staphylococcus aureus and Enterococcus. Infect. Control Hosp. Epidemiol. 24:362-386.
178. Mylonakis, E., M. Engelbert, X. Qin, C. D. Sifri, B. E. Murray, F. M. Ausubel, M. S. Gilmore, andS. B. Calderwood. 2002. The Enterococcus faecalis fsrB gene, a key component of the fsrquorum-sensing system, is associated with virulence in the rabbit endophthalmitis model. Infect.Immun. 70:4678-4681.
179. Nallapareddy, S. R., X. Qin, G. M. Weinstock, M. Hook, and B. E. Murray. 2000. Enterococcusfaecalis adhesin, Ace, mediates attachment to extracellular matrix proteins collagen type IV andlaminin as well as collagen type I. Infect. Immun. 68:5218-5224.
180. Nallapareddy, S. R., K. V. Singh, R. W. Duh, G. M. Weinstock, and B. E. Murray. 2000. Diversityof ace, a gene encoding a microbial surface component recognizing adhesive matrix molecules,from different strains of Enterococcus faecalis and evidence for production of Ace during humaninfections. Infect. Immun. 68:5210-5217.
181. Neut, D., J. R. van Horn, T. G. van Kooten, H. C. van der Mei, and H. J. Busscher. 2003.
133
REFERENCES

Detection of biomaterial-associated infections in orthopaedic joint implants. Clin. Orthop. 261-268.
182. Noble, C. J. 1978. Carriage of group D streptococci in the human bowel. J. Clin. Pathol.31:1182-1186.
183. Noordmans, J., J. Kempen, and H. J. Busscher. 1993. Automated image analysis to determinezeta potential distributions in particulate microelectrophoresis. J. Colloid Interface Sci. 156:394-399.
184. Novak, R. M., T. J. Holzer, and C. R. Libertin. 1993. Human neutrophil oxidative response andphagocytic killing of clinical and laboratory strains of Enterococcus faecalis. Diagn. Microbiol.Infect. Dis. 17:1-6.
185. Olmsted, S. B., G. M. Dunny, S. L. Erlandsen, and C. L. Wells. 1994. A plasmid-encoded surfaceprotein on Enterococcus faecalis augments its internalization by cultured intestinal epithelialcells. J. Infect. Dis. 170:1549-1556.
186. Onderdonk, A. B., J. G. Bartlett, T. Louie, N. Sullivan-Seigler, and S. L. Gorbach. 1976. Microbialsynergy in experimental intra-abdominal abscess. Infect. Immun. 13:22-26.
187. Paine, K., and D. R. Flower. 2002. Bacterial bioinformatics: pathogenesis and the genome. J.Mol. Microbiol. Biotechnol. 4:357-365.
188. Pascual, A. 2002. Pathogenesis of catheter-related infections: lessons for new designs. Clin.Microbiol. Infect. 8:256-264.
189. Patel, R., A. D. Badley, J. Larson-Keller, W. S. Harmsen, D. M. Ilstrup, R. H. Wiesner, J. L. Steers,R. A. Krom, D. Portela, F. R. Cockerill, and C. V. Paya. 1996. Relevance and risk factors ofenterococcal bacteremia following liver transplantation. Transplantation. 61:1192-1197.
190. Patel, R., K. E. Piper, M. S. Rouse, J. M. Steckelberg, J. R. Uhl, P. Kohner, M. K. Hopkins, F. R.3. Cockerill, and B. C. Kline. 1998. Determination of 16S rRNA sequences of enterococci andapplication to species identification of nonmotile Enterococcus gallinarum isolates. J. Clin.Microbiol. 36:3399-3407.
191. Patterson, J. E., A. Wanger, K. K. Zscheck, M. J. Zervos, and B. E. Murray. 1990. Molecularepidemiology of beta-lactamase-producing enterococci. Antimicrob. Agents Chemother.34:302-305.
192. Paulsen, I. T., L. Banerjei, G. S. Myers, K. E. Nelson, R. Seshadri, T. D. Read, D. E. Fouts, J. A.Eisen, S. R. Gill, J. F. Heidelberg, H. Tettelin, R. J. Dodson, L. Umayam, L. Brinkac, M. Beanan,S. Daugherty, R. T. DeBoy, S. Durkin, J. Kolonay, R. Madupu, W. Nelson, J. Vamathevan, B.Tran, J. Upton, T. Hansen, J. Shetty, H. Khouri, T. Utterback, D. Radune, K. A. Ketchum, B. A.Dougherty, and C. M. Fraser. 2003. Role of mobile DNA in the evolution of vancomycin-resistant Enterococcus faecalis. Science. 299:2071-2074.
193. Paya, C. V., P. E. Hermans, J. A. 2. Washington, T. F. Smith, J. P. Anhalt, R. H. Wiesner, and R.A. Krom. 1989. Incidence, distribution, and outcome of episodes of infection in 100 orthotopicliver transplantations. Mayo Clin. Proc. 64:555-564.
194. Paya, C. V., R. H. Wiesner, P. E. Hermans, J. J. Larson-Keller, D. M. Ilstrup, R. A. Krom, S. Rettke,and T. F. Smith. 1993. Risk factors for cytomegalovirus and severe bacterial infections followingliver transplantation: a prospective multivariate time- dependent analysis. J. Hepatol. 18:185-195.
195. Pillai, S. K., G. Sakoulas, H. S. Gold, C. Wennersten, G. M. Eliopoulos, R. C. Moellering, Jr., andR. T. Inouye. 2002. Prevalence of the fsr locus in Enterococcus faecalis infections. J. Clin.Microbiol. 40:2651-2652.
134

196. Poelstra, K. A., H. C. van der Mei, B. Gottenbos, D. W. Grainger, J. R. van Horn, and H. J.Busscher. 2000. Pooled human immunoglobulins reduce adhesion of Pseudomonas aeruginosain a parallel plate flow chamber. J. Biomed. Mater. Res. 51:224-232.
197. Poulsen, L. K., T. R. Licht, C. Rang, K. A. Krogfelt, and S. Molin. 1995. Physiological state ofEscherichia coli BJ4 growing in the large intestines of streptomycin-treated mice. J. Bacteriol.177:5840-5845.
198. Pryce, T. M., R. D. Wilson, and J. K. Kulski. 1999. Identification of enterococci by ribotyping with horseradish-peroxidase- labelled 16S rDNA probes. J. Microbiol. Methods.36:147-155.
199. Qin, X., K. V. Singh, G. M. Weinstock, and B. E. Murray. 2000. Effects of Enterococcus faecalisfsr genes on production of gelatinase and a serine protease and virulence. Infect. Immun.68:2579-2586.
200. Qin, X., K. V. Singh, G. M. Weinstock, and B. E. Murray. 2001. Characterization of fsr, aregulator controlling expression of gelatinase and serine protease in Enterococcus faecalisOG1RF. J. Bacteriol. 183:3372-3382.
201. Rakita, R. M., N. N. Vanek, K. Jacques-Palaz, M. Mee, M. M. Mariscalco, G. M. Dunny, M.Snuggs, W. B. van Winkle, and S. I. Simon. 1999. Enterococcus faecalis bearing aggregationsubstance is resistant to killing by human neutrophils despite phagocytosis and neutrophilactivation. Infect. Immun. 67:6067-6075.
202. Razatos, A., Y. L. Ong, M. M. Sharma, and G. Georgiou. 1998. Molecular determinants ofbacterial adhesion monitored by atomic force microscopy. Proc. Natl. Acad. Sci. U. S. A.95:11059-11064.
203. Rich, R. L., B. Kreikemeyer, R. T. Owens, S. LaBrenz, S. V. Narayana, G. M. Weinstock, B. E.Murray, and M. Hook. 1999. Ace is a collagen-binding MSCRAMM from Enterococcus faecalis.J. Biol. Chem. 274:26939-26945.
204. Richards, M. J., J. R. Edwards, D. H. Culver, and R. P. Gaynes. 2000. Nosocomial infections incombined medical-surgical intensive care units in the United States. Infect. Control Hosp.Epidemiol. 21:510-515.
205. Rouxhet, P. G. , and Genet, M. J. 1991. Chemical composition of the microbial cell surface by x-ray photoelectron spectroscopy, p. 173-220. In N. Mozes, P. S. Handley, H. J. Busscher, and P. G.Rouxhet (ed.), Microbial cell surface analysis: structural and physicochemical methods. VCHPublishers, New York.
206. Ruoff, K. L., L. de la Maza, M. J. Murtagh, J. D. Spargo, and M. J. Ferraro. 1990. Speciesidentities of enterococci isolated from clinical specimens. J. Clin. Microbiol. 28:435-437.
207. Ruoslahti, E. 1996. RGD and other recognition sequences for integrins. Annu. Rev. Cell Dev.Biol. 12:697-715.
208. Ruoslahti, E., and M. D. Pierschbacher. 1987. New perspectives in cell adhesion: RGD andintegrins. Science. 238:491-497.
209. Sabria-Leal, M., M. A. Pfaller, V. H. Morthland, S. A. Young, R. J. Hollis, L. Werkmeister, R. L.Kleiman-Wexler, and K. S. Ephgrave. 1994. Molecular epidemiology of gastric colonization byEnterococcus faecalis in a surgical intensive care unit. Diagn. Microbiol. Infect. Dis. 19:197-202.
210. Sandoe, J. A., I. R. Witherden, H. K. Au-Yeung, P. Kite, K. G. Kerr, and M. H. Wilcox. 2002.Enterococcal intravascular catheter-related bloodstream infection: management and outcome of61 consecutive cases. J. Antimicrob. Chemother. 50:577-582.
211. Sandoe, J. A., I. R. Witherden, J. H. Cove, J. Heritage, and M. H. Wilcox. 2003. Correlation
135
REFERENCES

between enterococcal biofilm formation in vitro and medical-device-related infection potential invivo. J. Med. Microbiol. 52:547-550.
212. Sartingen, S., E. Rozdzinski, A. Muscholl-Silberhorn, and R. Marre. 2000. Aggregationsubstance increases adherence and internalization, but not translocation, of Enterococcusfaecalis through different intestinal epithelial cells In vitro. Infect. Immun. 68:6044-6047.
213. Schlievert, P. M., P. J. Gahr, A. P. Assimacopoulos, M. M. Dinges, J. A. Stoehr, J. W. Harmala,H. Hirt, and G. M. Dunny. 1998. Aggregation and binding substances enhance pathogenicity inrabbit models of Enterococcus faecalis endocarditis. Infect. Immun. 66:218-223.
214. Schmitz, F. J., J. Verhoef, and A. C. Fluit. 1999. Prevalence of aminoglycoside resistance in 20European university hospitals participating in the European SENTRY Antimicrobial SurveillanceProgramme. Eur. J. Clin. Microbiol. Infect. Dis. 18:414-421.
215. Shankar, N., A. S. Baghdayan, and M. S. Gilmore. 2002. Modulation of virulence within apathogenicity island in vancomycin-resistant Enterococcus faecalis. Nature. 417:746-750.
216. Shankar, N., C. V. Lockatell, A. S. Baghdayan, C. Drachenberg, M. S. Gilmore, and D. E.Johnson. 2001. Role of Enterococcus faecalis surface protein Esp in the pathogenesis ofascending urinary tract infection. Infect. Immun. 69:4366-4372.
217. Shankar, V., A. S. Baghdayan, M. M. Huycke, G. Lindahl, and M. S. Gilmore. 1999. Infection-derived Enterococcus faecalis strains are enriched in esp, a gene encoding a novel surfaceprotein. Infect. Immun. 67:193-200.
218. Shiono, A., and Y. Ike. 1999. Isolation of Enterococcus faecalis clinical isolates that efficientlyadhere to human bladder carcinoma T24 cells and inhibition of adhesion by fibronectin andtrypsin treatment. Infect. Immun. 67:1585-1592.
219. Shlaes, D. M., J. Levy, and E. Wolinsky. 1981. Enterococcal bacteremia without endocarditis.Arch. Intern. Med. 141:578-581.
220. Shorrock, P. J., and P. A. Lambert. 1989. Binding of fibronectin and albumin to Enterococcus(Streptococcus) faecalis. Microb. Pathog. 6:61-67.
221. Sifri, C. D., E. Mylonakis, K. V. Singh, X. Qin, D. A. Garsin, B. E. Murray, F. M. Ausubel, and S.B. Calderwood. 2002. Virulence effect of Enterococcus faecalis protease genes and the quorum-sensing locus fsr in Caenorhabditis elegans and mice. Infect. Immun. 70:5647-5650.
222. Singh, K. V., T. M. Coque, G. M. Weinstock, and B. E. Murray. 1998. In vivo testing of anEnterococcus faecalis efaA mutant and use of efaA homologs for species identification. FEMSImmunol. Med. Microbiol. 21:323-331.
223. Singh, K. V., X. Qin, G. M. Weinstock, and B. E. Murray. 1998. Generation and testing ofmutants of Enterococcus faecalis in a mouse peritonitis model. J. Infect. Dis. 178:1416-1420.
224. Sjollema, J., and H. J. Busscher. 1990. Deposition of polystyrene particles in a parallel plate flowcell. 2. Pair distribution functions between deposited particles. Colloids and Surfaces. 47:337-352.
225. Steffen, E. K., and R. D. Berg. 1983. Relationship between cecal population levels of indigenousbacteria and translocation to the mesenteric lymph nodes. Infect. Immun. 39:1252-1259.
226. Stevens, S. X., H. G. Jensen, B. D. Jett, and M. S. Gilmore. 1992. A hemolysin-encoding plasmidcontributes to bacterial virulence in experimental Enterococcus faecalis endophthalmitis. Invest.Ophthalmol. Vis. Sci. 33:1650-1656.
227. Stone, H. H., E. S. Morris, C. E. Geheber, L. D. Kolb, and W. E. Dunlop. 1982. Clinicalcomparison of cefotaxime with gentamicin plus clindamycin in the treatment of peritonitis andother soft-tissue infections. Rev. Infect. 4 Suppl.: S439-S443.
136

228. Styriak, I., A. Laukova, C. Fallgren, and T. Wadstrom. 1999. Binding of selected extracellularmatrix proteins to enterococci and Streptococcus bovis of animal origin. Curr. Microbiol.39:327-0335.
229. Sulaiman, A., R. M. Rakita, R. C. Arduino, J. E. Patterson, J. M. Steckelberg, K. V. Singh, and B.E. Murray. 1996. Serological investigation of enterococcal infections using western blot. Eur. J.Clin. Microbiol. Infect. Dis. 15:826-829.
230. Sung, J. Y., E. A. Shaffer, K. Lam, I. Rususka, and J. W. Costerton. 1994. Hydrophobic bile saltinhibits bacterial adhesion on biliary stent material. Dig. Dis. Sci. 39:999-1006.
231. Sussmuth, S. D., A. Muscholl-Silberhorn, R. Wirth, M. Susa, R. Marre, and E. Rozdzinski. 2000.Aggregation substance promotes adherence, phagocytosis, and intracellular survival ofEnterococcus faecalis within human macrophages and suppresses respiratory burst. Infect.Immun. 68:4900-4906.
232. Teng, F., K. D. Jacques-Palaz, G. M. Weinstock, and B. E. Murray. 2002. Evidence that theenterococcal polysaccharide antigen gene (epa) cluster is widespread in Enterococcus faecalisand influences resistance to phagocytic killing of E. faecalis. Infect. Immun. 70:2010-2015.
233. Tokars, J. I., T. Gehr, W. R. Jarvis, J. Anderson, N. Armistead, E. R. Miller, J. Parrish, S. Qaiyumi,M. Arduino, S. C. Holt, F. C. Tenover, G. Westbrook, and P. Light. 2001. Vancomycin-resistantenterococci colonization in patients at seven hemodialysis centers. Kidney Int. 60:1511-1516.
234. Tokars, J. I., S. Satake, D. Rimland, L. Carson, E. R. Miller, E. Killum, R. L. Sinkowitz-Cochran,M. J. Arduino, F. C. Tenover, B. Marston, and W. R. Jarvis. 1999. The prevalence of colonizationwith vancomycin-resistant Enterococcus at a Veterans’ Affairs institution. Infect. Control Hosp.Epidemiol. 20:171-175.
235. Toledo-Arana, A., J. Valle, C. Solano, M. J. Arrizubieta, C. Cucarella, M. Lamata, B. Amorena,J. Leiva, J. R. Penades, and I. Lasa. 2001. The enterococcal surface protein, Esp, is involved inEnterococcus faecalis biofilm formation. Appl. Environ. Microbiol. 67:4538-4545.
236. Tomayko, J. F., and B. E. Murray. 1995. Analysis of Enterococcus faecalis isolates fromintercontinental sources by multilocus enzyme electrophoresis and pulsed-field gelelectrophoresis. J. Clin. Microbiol. 33:2903-2907.
237. Touhami, A., B. Hoffmann, A. Vasella, F. A. Denis, and Y. F. Dufrene. 2003. Aggregation of yeastcells: direct measurement of discrete lectin-carbohydrate interactions. Microbiology. 149:2873-2878.
238. Tsutsui, O., S. Kokeguchi, T. Matsumura, and K. Kato. 1991. Relationship of the chemicalstructure and immunobiological activities of lipoteichoic acid from Streptococcus faecalis(Enterococcus hirae) ATCC 9790. FEMS Microbiol. Immunol. 3:211-218.
239. Van den Braak, N., A. Ott, A. van Belkum, J. A. Kluytmans, J. G. Koeleman, L. Spanjaard, A.Voss, A. J. Weersink, C. M. Vandenbroucke-Grauls, A. G. Buiting, H. A. Verbrugh, and H. P.Endtz. 2000. Prevalence and determinants of fecal colonization with vancomycin-resistantEnterococcus in hospitalized patients in The Netherlands. Infect. Control. Hosp. Epidemiol.21:520-524.
240. Van den Braak, N., A. van Belkum, D. Kreft, R. te Witt, H. A. Verbrugh, and H. P. Endtz. 1999.The prevalence and clonal expansion of high-level gentamicin-resistant enterococci isolated fromblood cultures in a Dutch university hospital. J. Antimicrob. Chemother. 44:795-798.
241. Van der Meer, J. T., W. van Vianen, E. Hu, W. B. van Leeuwen, H. A. Valkenburg, J. Thompson,and M. F. Michel. 1991. Distribution, antibiotic susceptibility and tolerance of bacterial isolatesin culture-positive cases of endocarditis in The Netherlands. Eur. J. Clin. Microbiol. Infect. Dis.10:728-734.
137
REFERENCES

242. Van der Mei, H. C., S. D. Cox, G.I., R. J. Doyle, and H. J. Busscher. 1993. A critical appraisal ofpositive cooperativity in oral streptococcal adhesion: Scatchard analyses of adhesion data versusanalyses of the spatial arrangement of adhering bacteria. J. Gen. Microbiol. 139:937-948.
243. Van der Mei, H. C., J. de Vries, and H. J. Busscher. 2000. X-ray photoelectron spectroscopy forthe study of microbial cell surfaces. Surface Science Reports. 39:1-24.
244. Van der Zwet, W. C., G. A. Parlevliet, P. H. Savelkoul, J. Stoof, A. M. Kaiser, A. M. van Furth,and C. M. Vandenbroucke-Grauls. 2000. Outbreak of Bacillus cereus infections in a neonatalintensive care unit traced to balloons used in manual ventilation. J. Clin. Microbiol. 38:4131-4136.
245. Van Raamsdonk, M., H. C. van der Mei, G. I. Geertsema-Doornbusch, J. J. De Soet, H. J.Busscher, and J. de Graaf. 1995. Physicochemical aspects of microbial adhesion — influence ofantibody adsorption on the deposition of Streptococcus sobrinus in a parallel-plate flowchamber. Colloids and Surfaces B: Biointerfaces. 4:401-410.
246. Vanek, N. N., S. I. Simon, K. Jacques-Palaz, M. M. Mariscalco, G. M. Dunny, and R. M. Rakita.1999. Enterococcus faecalis aggregation substance promotes opsonin- independent binding tohuman neutrophils via a complement receptor type 3-mediated mechanism. FEMS Immunol.Med. Microbiol. 26:49-60.
247. Vergis, E. N., N. Shankar, J. W. Chow, M. K. Hayden, D. R. Snydman, M. J. Zervos, P. K. Linden,M. M. Wagener, and R. R. Muder. 2002. Association between the presence of enterococcalvirulence factors gelatinase, hemolysin, and Enterococcal surface protein and mortality amongpatients with bacteremia due to Enterococcus faecalis. Clin. Infect. Dis. 35:570-575.
248. Wade, J., N. Rolando, J. Philpott-Howard, and J. Wendon. 2003. Timing and aetiology ofbacterial infections in a liver intensive care unit. J. Hosp. Infect. 53:144-146.
249. Wade, J. J., N. Rolando, K. Hayllar, J. Philpott-Howard, M. W. Casewell, and R. Williams.1995. Bacterial and fungal infections after liver transplantation: an analysis of 284 patients.Hepatology. 21:1328-1336.
250. Warren, R. E. 1988. Difficult streptococci. J. Hosp. Infect. 11 Suppl A:352-7.:352-357.
251. Wassenaar, T. M., and W. Gaastra. 2001. Bacterial virulence: can we draw the line? FEMSMicrobiol. Lett. 201:1-7.
252. Waters, C. M., C. L. Wells, and G. M. Dunny. 2003. The aggregation domain of aggregationsubstance, not the RGD motifs, is critical for efficient internalization by HT-29 enterocytes.Infect. Immun. 71:5682-5689.
253. Weinstock, G. M. 2000. Genomics and bacterial pathogenesis. Emerg. Infect. Dis. 6:496-504.
254. Wells, C. L., and S. L. Erlandsen. 1991. Localization of translocating Escherichia coli, Proteusmirabilis, and Enterococcus faecalis within cecal and colonic tissues of monoassociated mice.Infect. Immun. 59:4693-4697.
255. Wells, C. L., R. P. Jechorek, and S. L. Erlandsen. 1990. Evidence for the translocation ofEnterococcus faecalis across the mouse intestinal tract. J. Infect. Dis. 162:82-90.
256. Wells, C. L., R. P. Jechorek, and S. L. Erlandsen. 1995. Inhibitory effect of bile on bacterialinvasion of enterocytes: possible mechanism for increased translocation associated withobstructive jaundice. Crit. Care Med. 23:301-307.
257. Wells, C. L., R. P. Jechorek, M. A. Maddaus, and R. L. Simmons. 1988. Effects of clindamycinand metronidazole on the intestinal colonization and translocation of enterococci in mice.Antimicrob. Agents Chemother. 32:1769-1775.
138

258. Wells, C. L., M. A. Maddaus, and R. L. Simmons. 1988. Proposed mechanisms for thetranslocation of intestinal bacteria. Rev. Infect. Dis. 10:958-979.
259. Wells, C. L., E. A. Moore, J. A. Hoag, H. Hirt, G. M. Dunny, and S. L. Erlandsen. 2000. Inducibleexpression of Enterococcus faecalis aggregation substance surface protein facilitates bacterialinternalization by cultured enterocytes. Infect. Immun. 68:7190-7194.
260. Willems, R. J., W. Homan, J. Top, M. van Santen-Verheuvel, D. Tribe, X. Manzioros, C. Gaillard,C. M. Vandenbroucke-Grauls, E. M. Mascini, E. van Kregten, J. D. van Embden, and M. J.Bonten. 2001. Variant esp gene as a marker of a distinct genetic lineage of vancomycin-resistantEnterococcus faecium spreading in hospitals. Lancet. 357:853-855.
261. Willems, R. J., J. Top, N. van den Braak, A. van Belkum, H. Endtz, D. Mevius, E. Stobberingh,A. van Den Bogaard, and J. D. van Embden. 2000. Host specificity of vancomycin-resistantEnterococcus faecium. J. Infect. Dis. 182:816-823.
262. Willey, S. H., R. G. Hindes, G. M. Eliopoulos, and R. C. Moellering, Jr. 1989. Effects ofclindamycin and gentamicin and other antimicrobial combinations against enterococci in anexperimental model of intra- abdominal abscess. Surg. Gynecol. Obstet. 169:199-202.
263. Winstanley, C. 2002. Spot the difference: applications of subtractive hybridisation to the studyof bacterial pathogens. J. Med. Microbiol. 51:459-467.
264. Winston, D. J., C. Emmanouilides, and R. W. Busuttil. 1995. Infections in liver transplantrecipients. Clin. Infect. Dis. 21:1077-1089.
265. Wirth, R. 1994. The sex pheromone system of Enterococcus faecalis. More than just a plasmid-collection mechanism? Eur. J. Biochem. 222:235-246.
266. Xiao, J., M. Hook, G. M. Weinstock, and B. E. Murray. 1998. Conditional adherence ofEnterococcus faecalis to extracellular matrix proteins. FEMS Immunol. Med. Microbiol. 21:287-295.
267. Xu, Y., L. Jiang, B. E. Murray, and G. M. Weinstock. 1997. Enterococcus faecalis antigens inhuman infections. Infect. Immun. 65:4207-4215.
268. Xu, Y., B. E. Murray, and G. M. Weinstock. 1998. A cluster of genes involved in polysaccharidebiosynthesis from Enterococcus faecalis OG1RF. Infect. Immun. 66:4313-4323.
269. Xu, Y., K. V. Singh, X. Qin, B. E. Murray, and G. M. Weinstock. 2000. Analysis of a gene cluster of Enterococcus faecalis involved in polysaccharide biosynthesis. Infect. Immun. 68:815-823.
270. Yu, J. L., R. Andersson, and A. Ljungh. 1996. Infections associated with biliary drains. Scand. J.Gastroenterol. 31:625-630.
271. Yu, J. L., R. Andersson, H. Parsson, E. Hallberg, A. Ljungh, and S. Bengmark. 1996. Abacteriologic and scanning electron microscope study after implantation of foreign bodies in thebiliary tract in rats. Scand. J. Gastroenterol. 31:175-181.
272. Zareba, T. W., C. Pascu, W. Hryniewicz, and T. Wadstrom. 1997. Binding of extracellular matrixproteins by enterococci. Curr. Microbiol. 34:6-11.
273. Zervos, M. J., A. E. 3. Bacon, J. E. Patterson, D. R. Schaberg, and C. A. Kauffman. 1988.Enterococcal superinfection in patients treated with ciprofloxacin. J. Antimicrob. Chemother.21:113-115.
274. Zervos, M. J., S. Dembinski, T. Mikesell, and D. R. Schaberg. 1986. High-level resistance togentamicin in Streptococcus faecalis: risk factors and evidence for exogenous acquisition ofinfection. J. Infect. Dis. 153:1075-1083.
139
CAPTER
3

275. Zervos, M. J., C. A. Kauffman, P. M. Therasse, A. G. Bergman, T. S. Mikesell, and D. R.Schaberg. 1987. Nosocomial infection by gentamicin-resistant Streptococcus faecalis. Anepidemiologic study. Ann. Intern. Med. 106:687-691.
276. Zwaveling, J. H., J. K. Maring, I. J. Klompmaker, E. B. Haagsma, J. T. Bottema, M. Laseur, H. L.Winter, P. J. van Enckevort, E. M. TenVergert, H. J. Metselaar, H. A. Bruining, and M. J. Slooff.2002. Selective decontamination of the digestive tract to prevent postoperative infection: arandomized placebo-controlled trial in liver transplant patients. Crit Care Med. 30:1204-1209.
140

List of Abbreviations


Ace adhesin to collagen of E. faecalisAgg aggregation substanceAFLP amplified fragment length polymorphismAFM atomic force microscopyCBD common bile ductEfaA E. faecalis antigen AEpa enterococcal polysaccharide antigenEsp enterococcal surface proteinFEP fluoro-ethylene-propyleneFISH fluorescent in situ hybridizationGel gelatinaseHln cytolysin (hemolysin)ICU intensive care unitLT liver transplantPE polyethylenePFGE pulsed field gel electrophoresePMN polymorphonuclear leukocyteSR silicone rubberTHB Todd-Hewitt brothVRE vancomycin resistant E. faeciumXPS X-ray photoelectron spectroscopy
143
ABBREVIATI
ONS


Dankwoord


Dankwoord
Het dankwoord is bijna het moeilijkste deel om te schrijven omdat je zo veel wilt zeggen enzo veel mensen wilt bedanken. Hoe breng ik mijn waardering onder woorden en hoe kijk ikterug op deze tijd?Met de inkt op mijn bul nog nat kwam ik 7 jaar geleden vanuit Leiden naar het voor mij nogonbekende Hoge Noorden. Als ik nu terug kijk is het een heel goede keus geweest dezespannende stap te nemen. Ik heb het altijd erg naar mijn zin gehad bij de MedischeMicrobiologie. Dankzij velen heb ik mijn onderzoek goed kunnen afronden. Graag wil ikeen aantal mensen specifiek noemen.
Prof. Dr. J.E. Degener, Beste John, 7 jaar geleden begonnen we bijna te gelijk bij de MMB,jij werd zowel mijn promotor als mijn opleider. Dit werd het begin van een prettige enintensieve samenwerking want zowel het onderzoek als de opleiding in een AGIKO trajectstonden nog in de kinderschoenen. Bedankt dat je me zelf mijn weg hebt laten vinden enop de juiste momenten hebt bijgestuurd.
Prof. Dr. H.J. Busscher en Prof. Dr. H.C. van der Mei, Beste Henk en Henny, de hartelijkheidwaarmee ik ben ontvangen en de vanzelfsprekendheid waarmee ik werd opgenomen injullie onderzoeksgroep heb ik enorm gewaardeerd. Ik heb erg veel geleerd van onzebesprekingen en jullie benadering van het onderzoek.
Beste Hermie, hartelijk bedankt dat je regelmatig als denktank wilde functioneren als ikeven niet meer wist hoe het verder moest, jouw enthousiasme voor het onderzoek werkterg aanstekelijk.
Prof. Dr. M.J.M. Bonten, Prof. Dr. T.H. The, Prof. Dr. M.J.H. Slooff, hartelijk dank voor desnelle beoordeling van het manuscript.
Beste Rob Willems, op een congres in Canada waren wij de enige twee Nederlanders, datwas het begin van een erg prettige en leerzame samenwerking, hartelijk dank voor jebijdrage aan hoofdstuk 2 en 3 van dit proefschrift en de inspirerende gesprekken.
Lieber Albrecht Muscholl-Silberhorn, vielen Dank für den herzlichen Empfang damals inRegensburg und die Einweihung in die Geheime der Enterokokken molekular Forschung.
Beste Astrid, Florence, Gerda, Arnout, Dewi, Hans en alle andere medewerkers en AIO’s vande BME, ik vond het erg leuk om een tijd bij jullie te gast te zijn en ik voelde me erg welkom.
Margriet, Henk Jans, Rudi, Kor en anderen, bedankt voor de grotere of kleiner steentjes diejullie hebben bijgedragen.
Gjalt en Sytske, jullie betrokkenheid bij het wel en wee van de collega’s levert een grotebijdrage aan de gezelligheid op de MMB, Ank, Bernadette, Linda, Matty, Marieke, Albert-
147
DANKWOORD

Jan, Danny, Erwin, en andere medewerkers van de disciplinegroep, het was altijd gezelligom met een kopje koffie over het onderzoek of andere zaken te filosoferen.
Door de AGIKO constructie waarin ik mijn promotie heb gedaan zijn voor mij onderzoek ende opleiding tot microbioloog met elkaar verweven daarom zou ik ook de mensen die iktijdens mijn opleiding heb ontmoet willen bedanken. Allereerst natuurlijk de arts-assistentenEveline, Greetje, Niek, Annelies, Jutte, Glen, Astrid, ik vond het altijd erg gezellig omkamers, computers en lief en leed te delen. De microbiologen, Hilly, Willem, Nico, Rik, Janen René die het geduld hebben opgebracht mij de opleiding in etappes te laten voltooien.De analisten en andere medewerkers van het AZG en SLGD bedankt voor de gezellige tijden alles wat ik van jullie geleerd heb.
Lieve Marion, ik heb veel bewondering voor de manier waarop jij altijd voor familie envrienden klaar staat, het is ontzettend fijn om zo’n meuke te hebben. Lieve Harriët, na veelgezamenlijke avonturen tijdens de studie vertrokken we op dezelfde dag uit Leiden vooropleidingen in Maastricht en Groningen, de afstand werd groter de vriendschap bleef.Bedankt dat jullie mijn paranimfen willen zijn.
Toneelvrienden van het eerste uur, Inge, Karin, Marleen, Jan, en Mark, een mislukte proefna een dag hard werken was altijd snel vergeten tijdens een gezellige repetitie avond of eenavondje uit, hopelijk volgen er nog vele.
Lieve oma Wieke, aan mijn schier eindeloze “studie” is nu eindelijk een einde gekomen, ikga niet trouwen maar er is wel een feestje! Lieve opa Jo, fijn dat u altijd zo veel interesseheeft in het wel en wee van de (achter)kleinkinderen.
Lieve pa en ma, ver weg maar toch dichtbij, te veel om te noemen, gewoon bedankt!
Heit en mem, bedankt voor de altijd hartelijke ontvangst op jullie mooie plekje aan het water,onder de perenboom achter jullie huis komen soms de beste ideeën voor het onderzoek.
Lieve Hidde, voor jouw dagelijkse ontdekkingen is geen laboratorium nodig, blijfnieuwsgierig en blijf je verbazen!
Lieve Douwe, aan wat voor ons het begin was is nu ook voor mij een einde gekomen. Opnaar nieuwe uitdagingen en avonturen, samen met jou, ik heb er zin in!
En tot slotte, dank aan de talloze koolmeesjes, roodborstjes, merels en vinken die zich tegoed hebben gedaan aan de vetbollen voor mijn raam en met hun gezellige aanwezigheidde laatste loodjes van dit proefschrift enigszins hebben verlicht.
148

Curriculum vitae


Curriculum Vitae
De schrijfster van dit proefschrift werd geboren op 6 april 1971 te Rotterdam. Zij verhuisdeop de leeftijd van 8 jaar naar München, Duitsland, waar zij in 1989 het baccalaureaatsdiploma aan de Europese School behaalde. Hetzelfde jaar begon zij aan de studie Bio-medische Wetenschappen aan de Rijksuniversiteit Leiden. In 1991 startte zij tevens met destudie Geneeskunde. De opleiding werd afgerond met onderzoek naar de seroprevalentievan Strongyloides stercoralis aan de Anton de Kom Universiteit van Parimaribo onderleiding van Prof. Dr. A.M. Polderman en Prof. Dr. B.F.J.Oeseburg. In 1997 werden beidestudies afgerond en begon zij aan het huidige AGIKO traject aan de RijksuniversiteitGroningen dat resulteerde in het onderliggende proefschrift. Volgend jaar hoopt zij haaropleiding tot arts-microbioloog af te ronden. Zij deelt het huis met Douwe en haar zoontjeHidde.
151
CURRICULUM
VITAE






























