Embed Size (px)
Citation preview

General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights.
Users may download and print one copy of any publication from the public portal for the purpose of private study or research.
You may not further distribute the material or use it for any profit-making activity or commercial gain
You may freely distribute the URL identifying the publication in the public portal If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim.
Downloaded from orbit.dtu.dk on: Dec 29, 2020
Unravelling a new catabolic pathway of C-19 steroids in Mycobacterium smegmatis
Fernández-Cabezón, Lorena; Galán, Beatriz; García, José L.
Published in:Environmental Microbiology
Link to article, DOI:10.1111/1462-2920.14114
Publication date:2018
Document VersionPeer reviewed version
Link back to DTU Orbit
Citation (APA):Fernández-Cabezón, L., Galán, B., & García, J. L. (2018). Unravelling a new catabolic pathway of C-19 steroidsin Mycobacterium smegmatis. Environmental Microbiology, 20(5), 1815-1827. https://doi.org/10.1111/1462-2920.14114

Unravelling a new catabolic pathway of C-19 steroids in
Mycobacterium smegmatis
Lorena Fernández-Cabezón1†
, Beatriz Galán1 and José L. García
1*
1Department of Environmental Biology. Centro de Investigaciones Biológicas. Consejo
Superior de Investigaciones Científicas. Ramiro de Maeztu 9, 28040 Madrid, Spain.
*For correspondence: E-mail [email protected]; Tel. +34 918373112; Fax +34 915360432
†Present address: The Novo Nordisk Foundation Center for Biosustainability, Technical
University of Denmark, Kemitorvet 220, 2800 Lyngby, Denmark
This article is protected by copyright. All rights reserved.
This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process which may lead to differences between this version and the Version of Record. Please cite this article as an ‘Accepted Article’, doi: 10.1111/1462-2920.14114

2
Summary
In this work, we have characterized the C-19+ gene cluster (MSMEG_2851 to
MSMEG_2901) of Mycobacterium smegmatis. By in silico analysis we have identified the
genes encoding enzymes involved in the modification of the A/B steroid rings during the
catabolism of C-19 steroids in certain M. smegmatis mutants mapped in the PadR-like
regulator (MSMEG_2868), that constitutively express the C-19+ gene cluster. By using
gene complementation assays, resting-cell biotransformations and deletion mutants, we
have characterized the most critical genes of the cluster, i.e, kstD2, kstD3, kshA2, kshB2,
hsaA2, hsaC2 and hsaD2. These results have allowed us to propose a new catabolic route
named C-19+ pathway for the mineralization of C-19 steroids in M. smegmatis. Our data
suggest that the deletion of the C-19+ gene cluster may be useful to engineer more
robust and efficient M. smegmatis strains to produce C-19 steroids from sterols.
Moreover, the new KshA2, KshB2, KstD2 and KstD3 isoenzymes may be useful to
design new microbial cell factories for the 9α-hydroxylation and/or Δ1-dehydrogenation
of 3-ketosteroids.
This article is protected by copyright. All rights reserved.

3
Introduction
Steroids are naturally occurring hydrophobic molecules that present a structure core formed
by four fused alicyclic rings named gonane. Steroid compounds are frequently found in the
biosphere and can be used as carbon and energy sources by different bacteria (e.g.
Mycobacterium, Pseudomonas, Sterolibacterium, Sphingomonas, Novosphingobium,
Comamonas) (Fujii et al., 2002, 2003; Tarlera and Denner, 2003; Horinouchi et al., 2003a,
2004a; Philipp et al., 2006; van der Geize et al., 2007; Roh and Chu, 2010; Leu et al., 2011;
Merino et al., 2013). Most bacteria capable of degrading aerobically sterols (e.g., cholesterol
and phytosterols), which are one of the most abundant steroid compounds in nature, belong to
the phylum Actinobacteria (e. g., Mycobacterium, Rhodococcus, Gordonia) (van der Geize et
al., 2007; Kendall et al., 2007, 2010; Drzyzga et al., 2009; Uhía et al., 2012; Fernández de las
Heras et al., 2013; Li et al., 2014; Bergstrand et al., 2016). The bacterial catabolism of
cholesterol has been investigated in detail because of its relevance in the pathogenicity of
certain bacteria such as Mycobacterium tuberculosis or Rhodococcus equi. These bacteria are
able to metabolize cholesterol located in the membranes of the host cells during the course of
infection, being the degradation of this sterol crucial for bacterial persistence (van der Geize
et al., 2007, 2011; Pandey and Sassetti, 2008). The investigation of cholesterol catabolism has
also attracted increasing interest because low-cost natural sterols (e.g., phytosterols) are
currently biotransformed at an industrial scale into valuable pharmaceutical steroidal
intermediates such as the C-19 steroids 4-androstene-3,17-dione (AD), 1,4-androstadiene-
3,17-dione (ADD) or 9α-hydroxy-4-androstene-3,17-dione (9OH-AD) (García et al., 2012;
Donova, 2017).
The aerobic 9,10-seco degradation for cholesterol pathway has been postulated based
on the compilation of multiple genetic and biochemical investigations carried out on different
Gram-positive bacteria, but this pathway is also partially shared by some Gram-negative
This article is protected by copyright. All rights reserved.

4
bacteria that degrade bile acids and testosterone (Fig. S1) (Barrientos et al., 2015; García et
al., 2012; Galán et al., 2017a). Briefly, in Actinobacteria, the cholesterol is transported by a
specific ATP-dependent transport system named Mce4 (Casali and Riley 2007; Pandey and
Sassetti, 2008; Mohn et al., 2008; Klepp et al., 2012; García-Fernández et al., 2017a) and is
subsequently transformed into cholest-4-en-3-one (cholestenone) by 3β-hydroxysteroid
dehydrogenase/isomerases enzymes (3β-HSD) and/or cholesterol oxidases (ChOx) (Kreit,
2017). Concomitantly with the formation of cholestenone, at least two P450 cytochromes,
named CYP125 and CYP142, initiate the side-chain degradation by performing sequential
oxidations of the methyl group at C-26 to generate a carboxylic acid (Capyk et al., 2009a;
McLean et al., 2009; Rosłoniec et al., 2009; Driscoll et al., 2010; Johnston et al., 2010;
Ouellet et al., 2010; García-Fernández et al., 2013). This modification enables the β-oxidation
of the side-chain that yields two molecules of propionyl-CoA, an acetyl-CoA and the 17-
ketosteroid AD (Fig. S1). Thereafter, the degradation of the A/B steroid rings of the
intermediate AD occurs via a central catabolic route called 9,10-seco pathway that results in
the generation of 2-hydroxyhexa-2,4.dienoic acid (HHD) and 9,17-dioxo-1,2,3,4,10,19-
hexanorandrostan-5-oic acid (HIP) (Fig. S1) (García et al., 2012; Galán et al., 2017a). The
degradation of HDD probably occurs through the action of enzymes similar to the TesE, TesF
and TesG proteins described in C. testosteroni (Horinouchi et al., 2005), while the
degradation of HIP (i.e., the mineralization of C/D steroid rings) takes place in a lower
catabolic pathway, recently studied in detail in M. tuberculosis and other Actinobacteria,
which leads to the formation of metabolites that enter the central metabolic pathways (Crowe
et al., 2017). The expression of the genes responsible for the catabolism of C/D rings is
controlled by the KstR2 repressor, a TetR-like transcriptional regulator (Kendall et al., 2010;
Casabon et al., 2013; Crowe et al., 2015; García-Fernández et al., 2015), while the expression
of the genes encoding the enzymes involved in the side-chain and the A/B rings degradation
This article is protected by copyright. All rights reserved.

5
as well as those encoding the sterol uptake system, is regulated by another TetR-like
transcriptional regulator, the KstR repressor (Kendall et al., 2007; 2010; Uhía et al., 2011).
Conventionally, it has been proposed that the C-19 steroids AD, ADD and 9OH-AD
are key intermediates of the (chole)sterol catabolic pathway due to multiple lines of evidence
(Fig. S1). Firstly, the C-19 compounds have been found in the culture medium of several
naturally sterols-degrading bacteria (Marsheck et al., 1972; Wei et al., 2010). Recently,
various mutant strains producing C-19 steroids from sterols have also been constructed
through metabolic engineering approaches by redirecting the metabolic flux of sterol
catabolism to the accumulation of C-19 compounds by gene deletions (Wilbrink et al., 2011;
Yeh et al., 2014; Yao et al., 2014; Galán et al., 2017b). On the other hand, several examples
of bacteria capable of using both cholesterol and C-19 steroids as the only carbon and energy
source have been described (e.g., Rhodococcus erythropolis strain SQ1, Rhodococcus ruber
strain Chol-4, Rhodococcus rhodochrous DSM 43269, Gordonia neofelifaecis NRRL B-
59395 (van der Geize et al., 2000; Petrusma et al., 2011; Fernández de las Heras et al., 2012;
Li et al., 2014). However, these bacterial strains have other sets of steroid-degrading
enzymes, different from those participating in the cholesterol catabolism, that are specifically
induced by different C-19 steroids. Therefore, in these strains, the different steroid catabolic
pathways could be partially overlapped creating a complex network of metabolic
intermediates that are degraded by a battery of redundant enzymes that have possibly led to
some misinterpretations of the pathways for sterol catabolism.
In a previous manuscript, we proposed that the C-19 steroids AD, ADD and 9OH-AD
are not the natural intermediates of the (chole)sterol catabolic pathway based on several lines
of evidence (Fernández-Cabezón et al., 2017). Firstly, different authors showed that certain
modifications introduced into the A/B rings during the catabolism of cholesterol (e.g., 9α-
hydroxylation and Δ1-dehydrogenation) might occur simultaneously with the side-chain
This article is protected by copyright. All rights reserved.

6
degradation. The enzymes involved in these catabolic steps appear to use as preferred
substrates the acyl-CoA derivatives resulting from the β-oxidation of the side-chain (i.e., most
probably compounds that still retain 3 or more carbons of the sterol side-chain) (Capyk et al.,
2011; Penfield et al., 2014). Secondly, by inhibiting the B-ring opening during the
degradation of cholesterol, it is possible to detect in the culture medium, in addition to the C-
19 steroids, other steroid intermediates that still retain part of the side-chain like 3-oxo-23,24-
bisnorchol-1,4-dien-22-oic acid (1,4-BNC), 3-oxo-23,24-bisnorchol-4-en-22-oic acid (4-
BNC), 22-hydroxy-23,24-bisnorchol-1,4-dien-3-one (1,4-HBC), or 22-hydroxy-23,24-
bisnorchol-4-en-3-one (4-HBC) (Szentirmai, 1990; Donova et al., 2005;Wilbrink et al., 2011;
Yeh et al., 2014; Galán et al., 2017b). Thirdly, we demonstrated that the C-19 steroids AD,
ADD and 9OH-AD cannot be efficiently mineralized through the cholesterol pathway in M.
smegmatis mc2155, since neither the wild-type strain nor its unregulated mutant ΔkstR are
able to degrade efficiently these compounds (Fernández-Cabezón et al., 2017). All these
results reinforce the hypothesis that certain modifications introduced into the A/B rings of
cholesterol occur simultaneously with the side-chain degradation and therefore, the C-19
steroids AD, ADD or 9OH-AD are actually side products of the cholesterol catabolic route
(Fig. S1).
On the other hand, we also reported the existence of a new silent and tightly-regulated
catabolic pathway in M. smegmatis, that is not induced by sterols (e.g., cholesterol,
phystosterols) nor by C-19 steroids (e.g., AD, ADD, 9OH-AD), but which, after its
desilencing/activation, confers to this bacterium the capacity to mineralize these and other C-
19 steroids (Fernández-Cabezón et al., 2017). The activation of this catabolic pathway is
found in some M. smegmatis spontaneous mutants mapped in the PadR-like regulator
(MSMEG_2868) that acquire the ability to efficiently metabolize C-19 steroids. The PadR
inactivation allows the expression of a set of genes named C-19+ cluster (MSMEG_2851 to
This article is protected by copyright. All rights reserved.

7
MSMEG_2901) that encode putative steroid-degrading enzymes. The C-19+ cluster would
have evolved independently from the upper cholesterol kstR-regulon, since C-19+ cluster has
an independent regulation, but both C-19+ cluster and kstR-regulon would converge on the
lower cholesterol kstR2-regulon responsible for the metabolism of C/D steroid rings (Fig. S1).
However, this is only a working hypothesis that requires further confirmation. Other steroid-
degrading Actinobacteria have homologous C-19+ clusters (e.g., Mycobacterium neoaurum
ATCC 25795, Rhodococcus jostii RHA1, Gordonia neofelifaecis NRRL-B59395,
Nocardioides simplex VKM Ac-2033D), but it is interesting to note that it is absent in M.
tuberculosis and in certain industrial mycobacterial strains used to produce C-19 steroids
from sterols (e.g., Mycobacterium sp. NRRL B-3805) (Fernández-Cabezón et al., 2017).
In this work, we have characterized in more detail the C-19+ gene cluster in M.
smegmatis. Using different methodologies, we have investigated the biological function
and/or the essentiality of those genes of the C-19+ cluster that encode the enzymes involved
in the modification of A/B rings during the catabolism of C-19 steroids in this bacterium.
Results
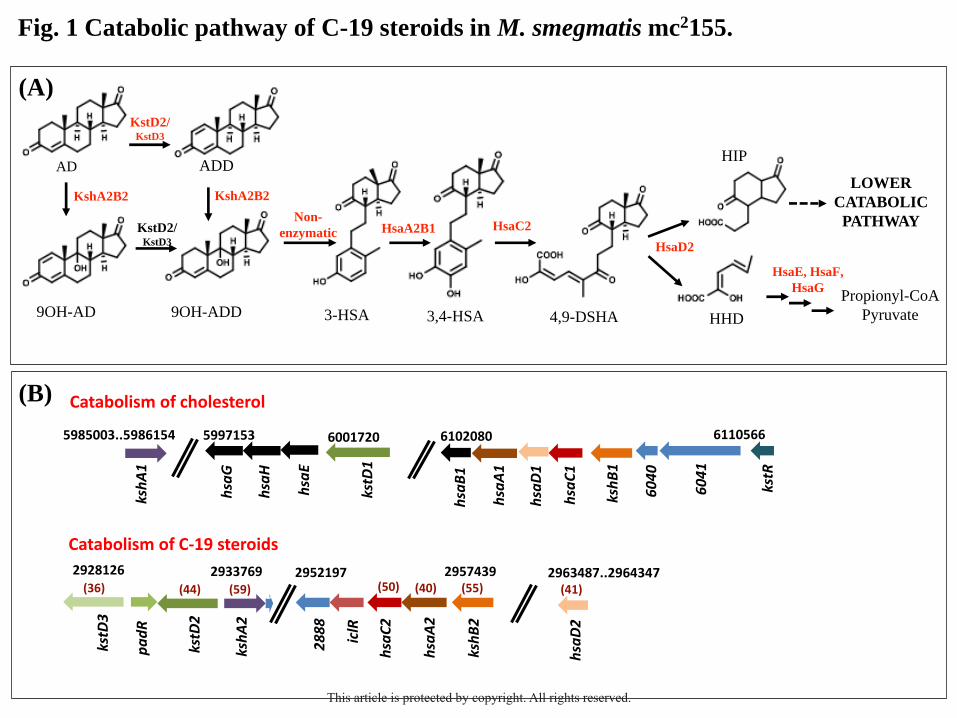
Identification of the key genes involved in the degradation of C-19 steroids in M. smegmatis
Based on the knowledge of the 9,10-seco pathway for the bacterial catabolism of steroids
(García et al., 2012; Galán et al., 2017a), we have proposed a catabolic route named C-19+
pathway for the catabolism of C-19 steroids in M. smegmatis (Fig. 1A) (Fernández-Cabezón
et al., 2017). By in silico analysis, we identified in the C-19+ gene cluster homologous genes
for almost all the postulated enzymatic steps involved in the A/B steroid ring modification
(Fig. 1B). The members of the kstR regulon involved in the cholesterol catabolism in M.
smegmatis have been used as reference genes (Kendall et al., 2007; Uhía et al., 2012). As it is
shown in the Fig. 1B, despite the existence of homologous genes in the two steroid catabolic
gene clusters found in M. smegmatis they do not share any synteny.
This article is protected by copyright. All rights reserved.

8
The genes MSMEG_2867 (kstD2) and MSMEG_2869 (kstD3) encode putative 3-
ketosteroid-Δ1-dehydrogenases sharing a 36% and 44% protein identity to KstD1
(MSMEG_5941), respectively. The MSMEG_2893 (kshB2) gene encodes a putative reductase
component of a 3-ketosteroid-9α-hydroxylase showing 55% protein identity to KshB1
(MSMEG_6039), whereas the MSMEG_2870 (kshA2) gene would encode the oxygenase
component of the same enzyme since it has a 59% protein identity to KshA1
(MSMEG_5925). The MSMEG_2892 (hsaA2) and MSMEG_2891 (hsaC2) genes may encode
a 3-HSA monooxygenase (oxygenase component) and a 3,4-HSA dioxygenase, that share
between 40-50% protein identity to HsaA1 (MSMEG_6038) and HsaC1 (MSMEG_6036),
respectively. Finally, the MSMEG_2900 (hsaD2) gene encodes a putative 4-9-DSHA
hydrolase with 41% protein identity to HsaD1 (MSMEG_6037). There are not homologous
genes to the hsaB1 (MSMEG_6035), hsaE (MSMEG_5940), hsaF (MSMEG_5939), and hsaG
(MSMEG_5937) genes within the C19+ cluster.
Determination of the biological function of the kstD2, kstD3, kshA2 and kshB2 genes
To investigate the role of the MSMEG_2893 (kshB2) gene, the mutant M. smegmatis
MS6039 (ΔMSMEG_6039, ΔkshB1) was complemented in trans with an original copy of the
kshB2 gene. Galán et al. (2017b) previously demonstrated that the MS6039 mutant is unable
to efficiently grow in sterols as the sole carbon and energy source, unlike the wild-type strain
of M. smegmatis, since the mutant lacks the KshB1 reductase activity necessary to mineralize
cholesterol. Analogously, to study the biological function of the MSMEG_2867 (kstD3) and
MSMEG_2869 (kstD2) genes, the mutant M. smegmatis MS5941 (ΔMSMEG_5941, ΔkstD1)
was constructed and complemented in trans with an original copy of the kstD2 or kstD3 genes
(Table S2). The KstD1 activity required to mineralize cholesterol is eliminated in the MS5941
mutant, and thus, this mutant is unable to efficiently grow in sterols as the sole carbon and
energy (Fig. 2A). However, when the MS5941 mutant is cultured in minimal media
This article is protected by copyright. All rights reserved.

9
containing glycerol as carbon and energy source and cholesterol (or phytosterols) as
feedstock, a large accumulation of 9OH-AD is observed (data not shown). According to our
hypothesis, the expression of a kshB gene (e.g., kshB1 or kshB2) or a kstD gene (e.g., kstD1,
kstD2 or kstD3) in the MS6039 or MS5941 mutants, respectively, will restore its capability to
grow in cholesterol. For this purpose, the MSMEG_2867 (kstD3), MSMEG_2869 (kstD2) and
MSMEG_2893 (kshB2) genes were cloned into the pMV261 plasmid generating the
corresponding recombinant plasmids pMV2867 (harbouring kstD3), pMV2869 (harbouring
kstD2) and pMV2893 (harbouring kshB2). The recombinant plasmids pMV5941 (harbouring
kstD1) and pMV6039 (harbouring kshB1) were also constructed and used as controls for the
complementation experiments. The plasmids harboring the kshB genes were transformed in
the MS6039 mutant (ΔkshB1), whereas the plasmids harbouring the kstD genes were
transformed into the MS5941 mutant (ΔkstD1).
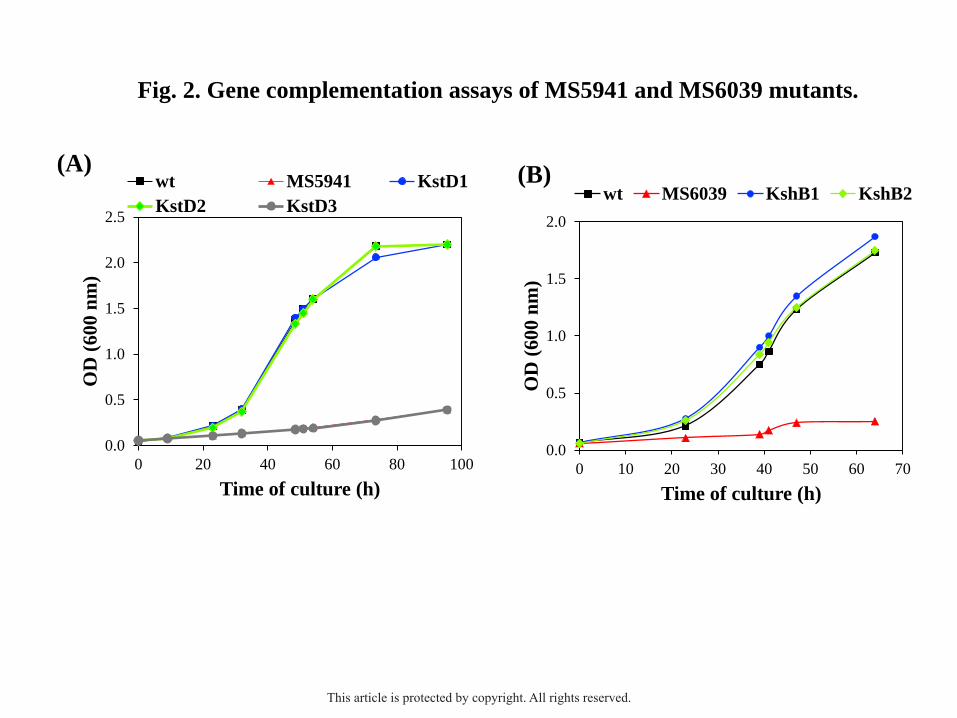
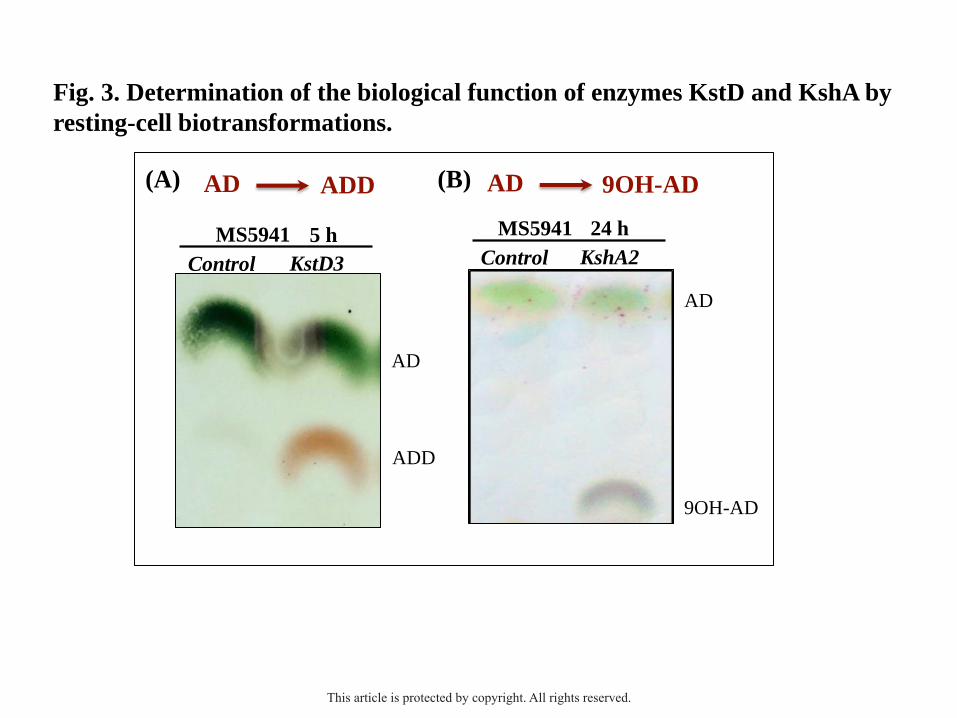
The overexpression of the kstD2 gene in the MS5941 strain restored its capability of
growing in cholesterol as the sole carbon and energy source, exhibiting a similar growth
pattern to the wild-type strain mc2155 and to the MS5941 (pMV5941) that is complemented
with the kstD1 gene (Fig. 2A). Otherwise, the overexpression of the kstD3 gene in the
MS5941 (pMV2867) recombinant did not restore the ability of MS5941 of growing in
cholesterol, suggesting that KstD3 present another enzymatic activity or a different substrate
specificity. To investigate this hypothesis, we monitored the bioconversion of several steroid
substrates by resting-cell biotransformation using the recombinant MS5941 (pMV2867). The
Figure 3A shows that this recombinant was able to transform AD into ADD, whereas the
control strain MS5941 (pMV261) harboring the empty plasmid was not able to, confirming
that KstD3 present 3-ketosteroid-Δ1-dehydrogenase activity. On the other hand, the
overexpression of the kshB2 gene in the MS6039 strain restored its capability of growing in
1.8 mM cholesterol as the sole carbon and energy source (Fig. 2B; Table S2)). The growth of
This article is protected by copyright. All rights reserved.

10
this recombinant strain MS6039 (pMV2893) was similar to the wild-type strain mc2155 and
to the MS6039 (pMV6039) that is complemented with the kshB1 gene. Finally, to investigate
the activity of MSMEG_2870 (kshA2) gene, we constructed the plasmid pMV2870
(harbouring kshA2) and followed the same resting cell approach. As expected, the
recombinant strain MS5941 (pMV2870) that produces constitutively the KshA2 protein, was
able to transform AD into 9OH-AD (Fig. 3B; Table S2).
These results confirmed that the kshA2 and kshB genes encode the oxygenase subunit
(KshA) and the ferredoxin reductase subunit (KshB) of a 3-ketosteroid-9α-hydroxylase
enzyme, while the kstD2 and kstD3 genes encode 3-ketoesteroid-Δ1-dehydrogenase proteins.
Determination of the biological function of the hsaA2, hsaC2 and hsaD2 genes
To investigate the biological role of the hsaA2 (MSMEG_2892), hsaC2 (MSMEG_2891) and
hsaD2 (MSMEG_2900) genes in the catabolism of C-19 steroids in M. smegmatis, we
constructed by double homologous recombination three mutant strains in M. smegmatis
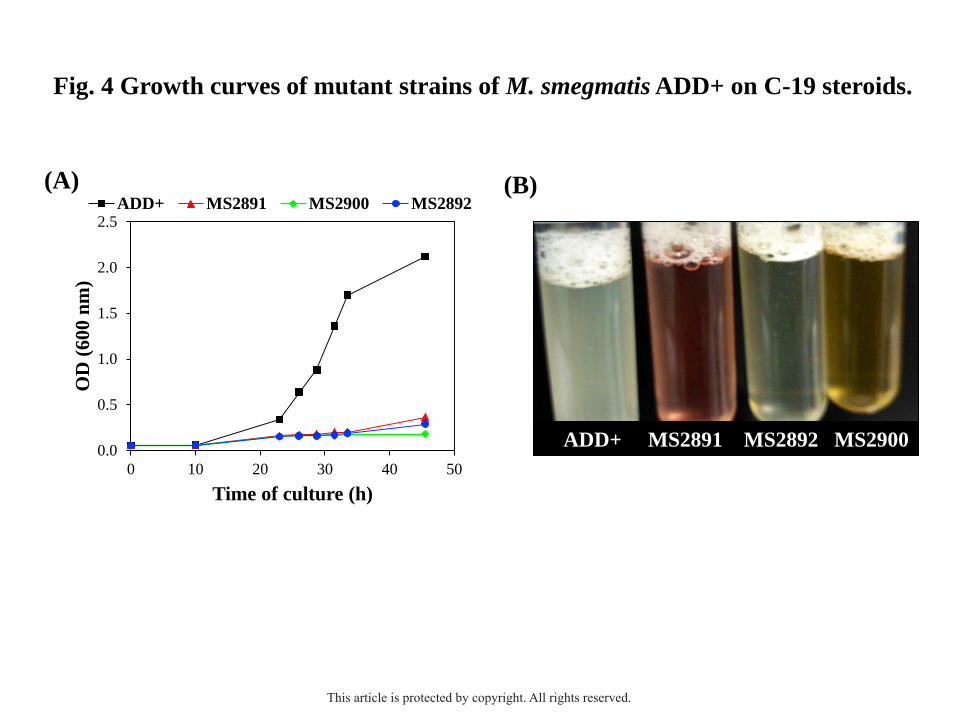
ADD+ named MS2891 (ΔhsaC2), MS2892 (ΔhsaA2) and MS2900 (ΔhsaD2) (Table 1). The
three engineered mutants were unable to efficiently grow in C-19 steroids (e.g., 1.8 mM AD)
as the sole carbon and energy source when compared with the parental strain ADD+ (Fig.
4A), although they perfectly grew using glycerol as a carbon source (data not shown).
Interestingly, a color alteration of the C-19 steroid-containing medium was observed during
the culture of the MS2891 and MS2900 mutants (Fig. 4B). A pink coloration of the medium
was developed in the culture of the MS2891 mutant (ΔhsaC2), possibly indicating the
accumulation of a catecholic derivate (e.g., 3,4-HSA) and its subsequent non-enzymatic
oxidation to quinone derivatives. This phenotype was previously observed in ΔhsaC mutants
of the cholesterol catabolism in R. jostii RHA1 and M. tuberculosis H37Rv (van der Geize et
al., 2007; Yam et al., 2009), as well as in the ΔtesB mutant of C. testosteroni TA441 in the
This article is protected by copyright. All rights reserved.

11
presence of ADD or cholic acid (Horinouchi et al., 2004b). On the other hand, a yellowish-
brown coloration in the medium was observed in the culture of the MS2900 mutant (ΔhsaD2)
likely due to the accumulation of 4,9-HSA (Gibson et al., 1966). This characteristic color was
also described in the mutant ΔtesD of C. testosteroni TA441 in the presence of testosterone
(Horinouchi et al., 2003b).
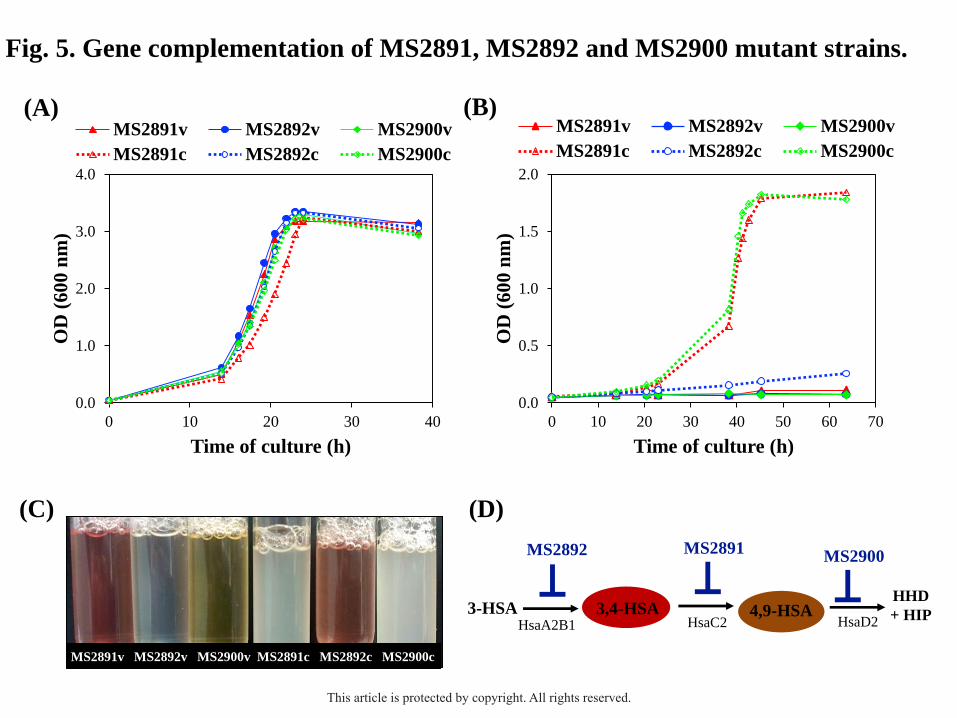
To confirm the absence of polar effects, the three mutant strains were complemented
in trans with an original copy of the corresponding deleted gene (MSMEG_2891,
MSMEG_2892 or MSMEG_2900). For that, each gene was cloned into the pMV261 plasmid
generating the recombinant plasmids pMV2891, pMV2892 and pMV2900 and then, they
were transformed into the mutant strains MS2891, MS2892 and MS2900, respectively (Table
1). Fig. 5B shows that the complemented MS2891 and MS2900 mutants (MS2891c and
MS2900c, respectively) recovered the capacity to grow in C-19 steroids as the sole carbon
and energy source (e.g., 1.8 mM AD) (Table S2). In contrast, the complemented MS2892
strain (MS2892c) was not able to grow in 1.8 mM AD as the sole carbon and energy source,
although it showed a normal growth in other substrates (e.g., 18 mM glycerol) (Fig. 5A).
However, the same pink coloration in the AD-containing medium described above with the
MS2891 mutant was also observed in the culture broths of MS2892c strain (Fig. 5C). This
result supports the hypothesis that the MSMEG_2892 gene encodes a 3-HSA monooxygenase,
which under overexpression conditions, could generate high amounts of 3,4-HSA to inhibit
bacterial growth. The high toxicity of these compounds to the cells has been previously
described (Yam et al., 2009).
Our results confirmed that the hsaA2, hsaC2 and hsaD2 genes play an essential role in
the catabolism of C-19 steroids in M. smegmatis. In addition, considering that the engineered
mutants were unable to efficiently grow in C-19 steroids, we can conclude that the HsaA2,
HsaC2 and HsaD2 activities cannot be replaced in these conditions by other enzymatic
This article is protected by copyright. All rights reserved.

12
activities. Notice that their corresponding homologous proteins involved in cholesterol
catabolism are not expressed in the tested conditions since the cholesterol pathway is not
induced by C-19 compounds.
Discussion
In this work, we have characterized the C-19+ gene cluster recently identified in M.
smegmatis (Fernández-Cabezón et al., 2017). First, we have identified by in silico analysis
several genes encoding enzymes putatively involved in the modification of the A/B steroid
rings for the catabolism of C-19 steroids in this bacterium. Then, we have used different
genetic and biochemical approaches to determine the biological function of these genes that
has allowed us to postulate the C-19+ pathway in M. smegmatis (Fig. 1A).
According to the classical 9,10-seco pathway, the transformation of AD into ADD is
the first enzymatic step necessary for the opening of A/B rings for the aerobic catabolism of
steroids. This reaction is catalyzed by 3-ketosteroid-Δ1-dehydrogenases named KstD in M.
smegmatis (Brzostek et al., 2005), R. erythropolis (van der Geize et al., 2000, 2001, 2002)
and M. tuberculosis (Brzostek et al., 2009), but named TesH in C. testosteroni (Horinouchi et
al., 2003a). The 3-ketosteroid-Δ1-dehydrogenase are FAD-dependent proteins that catalyze
the trans-axial removal of the hydrogen atoms C-1(α) and C-2(β) of 3-ketosteroids (EC
1.33.99.4) (Knol et al., 2008). We have demonstrated that the KstD2 protein is able to replace
the biological role of KstD1 in the catabolism of cholesterol in M. smegmatis MS5941
(ΔMSMEG_5941, ΔkstD1) (at least, under the tested conditions of kstD2 gene
overexpression), even though KstD2 is not involved in the catabolism of sterols in this
bacterium (Fig. 2A). This result agrees with the previous findings of Brzostek et al. (2005). In
contrast, the overexpresssion of the kstD3 gene (MSMEG_2867) in the MS5941 mutant is not
able to restore this capability (Fig. 2A), although the KstD3 protein has 3-ketosteroid-Δ1-
dehydrogenase activity in resting-cell biotransformations (Fig. 3A). This result suggests that
This article is protected by copyright. All rights reserved.

13
KstD3 is not able to efficiently recognize the intermediates with side-chain that are possibly
the true substrates of KstD1 during the cholesterol catabolism (Fernández-Cabezón et al.,
2017).
Then, the 9α-hydroxylation step takes place by the action of the 3-ketosteroid-9α-
hydroxylases (Andor et al., 2006; van der Geize et al., 2007; Capyk et al., 2009b). These
enzymes are two-component monooxygenases consisting of a terminal oxygenase subunit
(KshA) and a ferredoxin reductase subunit (KshB) (E.C. 1.14.13.142) (Petrusma et al., 2014).
The KshA subunit specifically recognizes the steroid substrates, whereas the KshB
component transfers the reducing power required for the progression of the enzymatic
reaction. Petrusma et al. (2011) described the presence of 5 kshA genes involved in the
catabolism of different classes of steroids (e.g., cholesterol, cholate, AD and progesterone) in
R. rhodochrous DSM43269, each displaying unique steroid induction patterns and substrate
ranges. They demonstrated that the overexpression of only certain kshA genes in a null-kshA
mutant restores its growth on any of the different classes of steroids tested. The KshA/KshB
interaction does not seem to be very specific, since it is possible to find more kshA genes than
kshB genes in the genomes of actinobacterial species (e.g. R. jostii RHA1, N. simplex VKM
Ac-2033D or G. neofelifaecis NRRL B-59395) (Petrusma et al., 2011). This observation
suggests that a particular KshB protein can interact with several KshAs or even that, other
non-specific reductases present in the bacterial cell can act as donors of reducing power. In
agreement with this observation, we have demonstrated that the KshB2 of M. smegmatis is
able to replace KshB1 in the catabolism of cholesterol (Fig. 2B). In the same line, Yuan et al.
(2015) were able to reconstitute a functional KSH in Mycobacterium sp. NRRL B-3805 by
overexpressing the kshA gene of M. smegmatis and the kshB gene of G. neofelifaecis NRRL
B-59395. Moreover, we have observed that the co-expression of a kshB gene does not seem to
This article is protected by copyright. All rights reserved.

14
be always essential for the 9α-hydroxylations of 3-ketosteroids (Fig. 3B), as previously
demonstrated by other authors (Andor et al., 2005; Arnell et al., 2007).
The 3-HSA intermediate obtained after the action of the 3-ketosteroid-Δ1-
dehydrogenase and the 3-ketosteroid-9α-hydroxylase enzymes, is hydroxylated by a two-
component oxygenase (HsaA2B1), similar to TesA1A2 from C. testosteroni (Horinouchi et
al., 2004b) or to HsaAB from M. tuberculosis and R. jostii RHA1 (Dresen et al., 2010)
leading to the production of 3,4-DHSA. This catechol derivative is subsequently cleaved by a
meta-extradiol dioxygenase (HsaC2), similar to TesB in C. testosteroni (Horinouchi et al.,
2001) or to HsaC in R. jostii RHA1 (van der Geize et al., 2007) and M. tuberculosis H37Rv
(Yam et al., 2009), that yields 4,9-DSHA. This compound is subsequently hydrolyzed by a
meta-cleavage product (MCP) hydrolase (HsaD2) similar to TesD in C. testosteroni
(Horinouchi et al., 2003b) or to HsaD in M. tuberculosis and R. jostii RHA1 (van der Geize et
al., 2007; Lack et al., 2008, 2010) yielding HIP and HHD. To investigate the biological role
of the HsaA2, HsaC2 and HsaD2 proteins in the catabolism of C-19 steroids, in this work we
have constructed by double homologous recombination the mutants ADD+ MS2891
(ΔMSMEG_2891; ΔhsaC2), ADD+ MS2892 (ΔMSMEG_2892; ΔhsaA2) and ADD+ MS2900
(ΔMSMEG_2900; ΔhsaD2). The three mutants are unable to efficiently grow in C-19 steroids
as the sole carbon and energy source, confirming that the hsaA2, hsaC2 and hsaD2 genes are
essential for the catabolism of C-19 steroids in M. smegmatis (Fig. 4A). Moreover, the color
changes of culture media observed when the mutant strains are cultured in the presence of C-
19 compounds strongly support the C-19+ pathway postulated above.
Finally, the degradation of HDD should occur through the action of the enzymes
HsaE, HsaF and HsaG encoded by the unique hsaEFG operon of M. smegmatis (Fernández-
Cabezón et al., 2017). These genes are not always present in all steroid-degrading gene
clusters in Actinobacteria, because they encode enzymes that are common in many catabolic
This article is protected by copyright. All rights reserved.

15
pathways of aromatic compounds and thus, these enzymes can replace those (Bergstrand et
al., 2016).
Summarizing, the results presented above confirm that the C-19 steroids AD, ADD
and 9OH-AD are mineralized through the C-19+ catabolic pathway that is constitutively
expressed in the padR mutants of M. smegmatis. Although we have identified some of the key
enzymatic steps of the C-19+ pathway, additional studies are required to determine the roles
of other genes found in the C-19+ gene cluster of M. smegmatis (MSMEG_2851 to
MSMEG_2901), as well as to identify the chemical compound that induces this cluster in
vivo. A better understanding of the complexity of steroid catabolism in M. smegmatis and
other actinobacterial species can also be fundamental to optimize the current industrial
bioproduction of steroidal intermediates from sterols. The deletion of the C-19+ gene cluster
or at least of the most relevant genes located therein (e.g., kstD, kshA, kshB) will be useful to
increase the bioconversion yields in producer strains derived from M. smegmatis and other
cholesterol degrading bacteria. Yao et al. (2014) demonstrated that two residual enzymatic
activities KstD (KstD2 and KstD3) present in the mutant Mut(MN-KSTD1) (∆kstD1) of M.
neoaurum ATCC 25795, markedly decreases the conversion yield of 9OH-AD from sterols.
Interestingly, KstD2 and KstD3 are encoded by genes located in a homologous C-19+ cluster
that is not found in the close phylogenetic strains M. neoaurum VKM Ac-1815D and
Mycobacterium sp. NRRL B-3805, both used in industrial bioprocesses. On the other hand,
given the industrial relevance of the 9α-hydroxylation and Δ1-dehydrogenation reactions of 3-
ketosteroids, the new KshA2, KshB2, KstD2 and KstD3 enzymes identified could be used to
design new microbial cell factories for the production of steroids à la carte.
This article is protected by copyright. All rights reserved.

16
Experimental procedures
Chemicals
4-Androstene-3,17-dione (AD) and 1,4-androstadien-3,17-dione (ADD) were purchased from
TCI America. Cholesterol, Tween 80, tyloxapol, gentamicin and kanamycin were from Sigma
(Steinheim, Germany). Glycerol and glucose were purchased from Merck (Darmstardt,
Germany). 9α-Hydroxy-4-androstene-3,17-dione (9OH-AD) was kindly provided by Gadea
Biopharma (León, Spain).
Bacterial strains, plasmids and culture conditions
The plasmids and bacterial strains used in this work are listed in Table 1. M. smegmatis
strains were grown at 37°C in an orbital shaker at 200 r.p.m. Middlebrook 7H9 broth medium
(Difco) supplemented with 0.4% glycerol and 0.05% Tween 80 was used as rich medium.
7H9 broth without any supplement was used as minimal medium. 7H10 agar (Difco) plates
supplemented with 10% albumin-dextrose-catalase (Becton Dickinson) were used for solid
media. Kanamycin (20 µg ml-1
) or gentamicin (5 µg ml-1
), were used for strain selection when
appropriate. All the steroids were dissolved in 10% tyloxapol prior to its addition to the
minimal medium. Because of the low solubility of the steroid compounds, the stock solutions
were warmed at 80 ºC in agitation, sonicated in a bath for 1 h and then autoclaved.
Escherichia coli DH10B strain was used as a host for cloning. It was grown in LB
medium at 37°C in an orbital shaker at 200 r.p.m. LB agar plates were used for solid media.
Kanamycin (50 µg ml-1
) or Gentamicin (10 µg ml-1
), were used for plasmid selection and
maintenance.
This article is protected by copyright. All rights reserved.

17
DNA manipulations and sequencing
DNA manipulations and other molecular biology techniques were essentially as described by
Sambrook and Russell (2001). Oligonucleotides used at this work were purchased from
Sigma-Aldrich and are listed at Table S1. Genomic DNA purification from mycobacterial
strains was performed as previously described (Uhía et al., 2011). DNA amplification was
performed on a Mastercycler Gradient (Eppendorf) using Phusion High-Fidelity DNA
Polymerase from ThermoFisher Scientific. DNA fragments were purified with High Pure
PCR System Product Purification Kit (Roche). Restriction enzymes were obtained from
various suppliers and were used according to their specifications. Plasmid DNA was prepared
with a High Pure Plasmid Isolation Kit (Roche Applied Science). E. coli was transformed by
the rubidium chloride method (Wirth et al., 1989). M. smegmatis cells were transformed by
electroporation (Gene Pulser; Bio-Rad) (Parish and Stoker, 1998). All cloned inserts and
DNA fragments were confirmed by DNA sequencing through an ABI Prism 377 automated
DNA sequencer (Applied Biosystems Inc.) at Secugen S.L. (Madrid, Spain).
Gene deletion
The knock-out strains were constructed by double homologous recombination using the
plasmid pJQ200x, a derivative of the suicide vector pJQ200 that does not replicate in
Mycobacterium (Jackson et al., 2001). The followed approach consisted in amplifying by
PCR two fragments of ~700 bp containing the upstream region (UP fragment) and the
downstream region (DOWN fragment) of the target gene to be deleted (Table S1). The two
PCR fragments were purified, digested with the corresponding enzymes and ligated. Using
the ligation product as template, the UP+DOWN fragment was amplified by PCR using the
most external primers. The resulting amplicon was cloned into the plasmid pJQ200x using E.
coli DH10B competent cells. Then, the resulting pJQ200x-derivate plasmid was
This article is protected by copyright. All rights reserved.

18
electroporated into competent M. smegmatis ADD+ (Table 1). The single cross-overs were
selected on 7H10 agar plates containing gentamicin. The obtained single colonies were
contra-selected in 10% sucrose and in addition, the presence of the xylE gene marker was
confirmed by the appearance of yellow coloration after spreading catechol over each colony
replica. To select for double cross-overs, a single colony was grown in rich medium up to an
optical density of 0.8–0.9 and several dilutions of this culture were plated onto 7H10 agar
plates with 10% sucrose. Potential double cross-overs (i.e., sucrose-resistant colonies) were
subsequently screened for gentamicin sensitivity and the gene deletion was confirmed by
PCR.
Complementation of the knock-out mutants
To isolate the genes (e.g., MSMEG_2891) from M. smegmatis mc2155 for the
complementation experiments, its genomic DNA was extracted and amplified by PCR using
the corresponding primers forward and reverse (e.g., MSMEG_2891F and MSMEG_2891R)
(Table S1). The resulting amplicons were purified and digested with the corresponding
restriction enzymes to be cloned into the expression vector pMV261 (Stover et al., 1991), a
shuttle plasmid that replicates in E. coli and Mycobacterium. The resulting plasmids (e.g.,
pMV2891) were transformed into E. coli DH10B for cloning purposes and once their
sequences were verified, they were used to transform electrocompetent cells of the
corresponding M. smegmatis mutant strain (e.g., MS2891). The mutant strains harboring the
plasmid pMV261were used as controls.
Resting-cell biotransformations
The recombinant strains were grown in rich medium at 37 °C during 24 h. The cells were
harvested by centrifugation at 5000 x g for 20 min at 4 °C and washed once with 0.85% NaCl.
This article is protected by copyright. All rights reserved.

19
The biotransformation was carried out with an optical density (OD600) of 5 in a 100 mL shake
flask containing 40 mL of reaction mixture: 0.1 M phosphate buffer (pH 8.0), 2 mM AD
(substrate) and 0.05% Tween 80. An additional carbon source (1% glucose) was added in the
9alpha-hydroxylation biotransformations. The substrate AD was incorporated into the
medium as a solution with randomly methylated β-cyclodextrin (1:10.3, molar ratio) (Klein et
al., 1995) or alternatively, was dissolved in 10% tyloxapol as mentioned above. The steroid
bioconversion was monitored by thin layer chromatography (TLC) (García-Fernández et al.,
2017b).
Bioinformatic analysis
Sequence alignments were carried out using Clustal W (Thompson et al., 1994) and different
BLAST algorithms from the National Centre of Biotechnology Information Server (NCBI)
were also used.
Acknowledgements
The technical work of A. Valencia is greatly appreciated. This work was supported by grants
from the Ministry of Science and Innovation (BFU2006-15214-C03-01, BFU2009-11545-
C03-03) and Ministry of Economy and Competitiveness (BIO2012-39695-C02-01). LFC was
supported by an FPU fellowship from the Spanish Ministry of Education, Culture and Sports.
Conflict of interest
None declare
This article is protected by copyright. All rights reserved.

20
References
Andor, A., Jekkel, A., Hopwood, D.A, Jeanplong, F., Ilkoy, E., Kónya, A., Kurucz, I., and
Ambrus, G. (2006) Generation of useful insertionally blocked sterol degradation pathway
mutants of fast-growing mycobacteria and cloning, characterization, and expression of
the terminal oxygenase of the 3-ketosteroid 9alpha-hydroxylase in Mycobacterium
smegmatis mc(2)155. Appl Environ Microbiol 72: 6554-6559.
Arnell, R., Johannisson, R., Lindholm, J., Fornstedt, T., Ersson, B., Ballagi, A., et al. (2007).
Biotechnological approach to the synthesis of 9alpha-hydroxylated steroids. Prep.
Biochem Biotechnol 3: 309-321.
Barrientos, A., Merino, E., Casabon, I., Rodríguez, J., Crowe, A.M., Holert, J., et al. (2015)
Functional analyses of three acyl-CoA synthetases involved in bile acid degradation in
Pseudomonas putida DOC21. Environ Microbiol 17: 47-63.
Bergstrand, L.H., Cardenas, E., Holert, J., Van Hamme, J.D., and Mohn, W.W. (2016)
Delineation of Steroid-Degrading Microorganisms through Comparative Genomic
Analysis. MBio 7: e00166.
Brzostek, A., Sliwiński, T., Rumijowska-Galewicz, A., Korycka-Machała, M., and Dziadek,
J. (2005) Identification and targeted disruption of the gene encoding the main 3-
ketosteroid dehydrogenase in Mycobacterium smegmatis. Microbiology 151: 2393-2402.
Brzostek, A., Pawelczyk, J., Rumijowska-Galewicz, A., Dziadek, B., and Dziadek, J. (2009)
Mycobacterium tuberculosis is able to accumulate and utilize cholesterol. J Bacteriol 191:
6584-6591.
Capyk, J.K., Kalscheuer, R., Stewart, G.R., Liu, J., Kwon, H., Zhao, R., et al. (2009a)
Mycobacterial cytochrome P450 125 (Cyp125) catalyzes the terminal hydroxylation of
C27-steroids. J Biol Chem 284: 35534–35542.
This article is protected by copyright. All rights reserved.

21
Capyk, J.K., D'Angelo, I., Strynadka, N.C., and Eltis, L.D. (2009b) Characterization of 3-
ketosteroid 9α-hydroxylase, a Rieske oxygenase in the cholesterol degradation pathway
of Mycobacterium tuberculosis. J Biol Chem 284: 9937-9946.
Capyk, J.K., Casabon I., Gruninger R., Strynadka N.C., and Eltis L.D. (2011) Activity of 3-
Ketosteroid 9α-hydroxylase (KshAB) indicates cholesterol side chain and ring
degradation occur simultaneously in Mycobacterium tuberculosis. J Biol Chem 286:
40717–40724.
Casabon, I., Zhu, S.H., Otani, H., Liu, J., Mohn, W.W., and Eltis, L.D. (2013) Regulation of
the KstR2 regulon of Mycobacterium tuberculosis by a cholesterol catabolite. Mol
Microbiol 89: 1201-1212.
Casali, N., and Riley, L.W. (2007) A phylogenomic analysis of the Actinomycetales mce
operons. BMC Genomics 8: 60.
Crowe, A.M., Stogios, P.J., Casabon, I., Evdokimova, E., Savchenko, A., and Eltis, L.D.
(2015) Structural and functional characterization of a ketosteroid transcriptional regulator
of Mycobacterium tuberculosis. J Biol Chem 290: 872-882.
Crowe, A.M., Casabon, I., Brown, K.L., Liu, J., Lian, J., Rogalski, J.C., et al. (2017)
Catabolism of the Last Two Steroid Rings in Mycobacterium tuberculosis and Other
Bacteria. MBio 8 pii: e00321-17. doi: 10.1128/mBio.00321-17.
Donova, M.V. (2017). Steroid Bioconversions. Methods Mol Biol. 1645: 1-13.
Donova, M.V., Gulevskaya, S.A., Dovbnya, D.V., and Puntus,I.F. (2005) Mycobacterium sp .
mutant strain producing 9alpha-hydroxyandrostenedione from sitosterol. Appl Microbiol
Biotechnol 67: 671–678.
This article is protected by copyright. All rights reserved.

22
Dresen, C., Lin, L.Y., D'Angelo, I., Tocheva, E.I., Strynadka, N., and Eltis, L.D. (2010) A
flavin dependent monooxygenase from Mycobacterium tuberculosis involved in
cholesterol catabolism. J Biol Chem 285: 22264-22275.
Driscoll, M.D., McLean, K.J., Levy, C., Mast, N., Pikuleva, I.A., Lafite, P., et al. (2010)
Structural and biochemical characterization of Mycobacterium tuberculosis CYP142:
evidence for multiple cholesterol 27-hydroxylase activities in a human pathogen. J Biol
Chem 285: 38270-38282.
Drzyzga, O., Navarro Llorens, J.M., Fernández de Las Heras, L., García Fernández, E., and
Perera, J. (2009) Gordonia cholesterolivorans sp. nov., a cholesterol-degrading
actinomycete isolated from sewage sludge. Int J Syst Evol Microbiol 59: 1011-1015.
Fernández de las Heras, L., van der Geize, R., Drzyzga, O., Perera, J., and Navarro-Llorens,
M.J. (2012) Molecular characterization of three 3-ketosteroid-Δ(1)-dehydrogenase
isoenzymes of Rhodococcus ruber strain Chol-4. J Steroid Biochem Mol Biol 132: 271-
281.
Fernández de Las Heras, L., Alonso, S., de la Vega de León, A., Xavier, D., Perera, J., and
Navarro Llorens, J.M. (2013) Draft Genome Sequence of the Steroid Degrader
Rhodococcus ruber Strain Chol-4. Genome Announc 16. pii: e00215-13.
Fernández-Cabezón, L., García-Fernández, E., Galán, B., and García, J.L. (2017). Molecular
characterization of a new gene cluster for steroid degradation in Mycobacterium
smegmatis. Environ Microbiol 19: 2546-2563.
Fujii, K., Kikuchi, S., Satomi, M., Ushio-Sata, N., and Morita, N. (2002) Degradation of
17beta-estradiol by a gram-negative bacterium isolated from activated sludge in a sewage
treatment plant in Tokyo, Japan. Appl Environ Microbiol 68: 2057-2060.
This article is protected by copyright. All rights reserved.

23
Fujii, K., Satomi, M., Morita, N., Motomura, T., Tanaka, T., and Kikuchi, S. (2003)
Novosphingobium tardaugens sp. nov., an oestradiol-degrading bacterium isolated from
activated sludge of a sewage treatment plant in Tokyo. Int J Syst Evol Microbiol 53: 47-
52.
Galán, B., García-Fernández, J., Felpeto-Santero, C., Fernández-Cabezón, L., and García, J.L.
(2017a). Bacterial metabolism of Steroids. In Handbook of Hydrocarbon and Lipid
Microbiology Series. Aerobic Utilization of Hydrocarbons, Oils and Lipids. Fernando
Rojo (ed). doi: 10.1007/978-3-319-39782-5.
Galán, B., Uhía, I., García-Fernández, E., Martínez, I., Bahíllo, E., de la Fuente, J.L. et al.
(2017b). Mycobacterium smegmatis is a suitable cell factory for the production of
steroidic synthons. Microb Biotechnol 10: 138-150.
García, J.L., Uhía, I., and Galán, B. (2012) Catabolism and biotechnological applications of
cholesterol degrading bacteria. Microb Biotechnol 5: 679-699.
García-Fernández, E., Frank, D.J., Galán, B., Kells, P.M., Podust, L.M., García, J.L., and
Ortiz de Montellano, P.R. (2013) A highly conserved mycobacterial cholesterol catabolic
pathway. Environ Microbiol 15: 2342-2359.
García-Fernández, J., Galán, B., Medrano, F.J., and García, J.L. (2015) Characterization of
the KstR2 regulator responsible of the lower cholesterol degradative pathway in
Mycobacterium smegmatis. Environ Microbiol Rep 7: 155-163.
García-Fernández, J., Papavinasasundaram, K., Galán, B., Sassetti, C.M., and García, JL.
(2017a). Molecular and functional analysis of the mce4 operon in Mycobacterium
smegmatis. Environ Microbiol 19: 3689-3699.
This article is protected by copyright. All rights reserved.

24
García-Fernández, J., Martínez, I., Fernández-Cabezón, L., Felpeto-Santero, C., García, J.L.,
and Galán, B. (2017b). Bioconversion of Phytosterols into Androstadienedione by
Mycobacterium smegmatis CECT 8331. Methods Mol Biol 1645: 211-225.
Gibson, D.T.K.C., Wang, C.J., Sih, C.J., and Whitlock, H. Jr. (1966). Mechanisms of steroid
oxidation by microorganisms. IX. On the mechanism of ring A cleavage in the
degradation of 9,10-seco steroids by microorganisms. J Biol Chem 241: 551–559.
Horinouchi, M., Yamamoto, T., Taguchi, K., Arai, H., and Kudo, T. (2001) Metacleavage
enzyme gene tesB is necessary for testosterone degradation in Comamonas testosteroni
TA441. Microbiology 147: 3367-3375.
Horinouchi, M., Hayashi, T., Yamamoto, T., and T. Kudo. (2003a). A new bacterial steroid
degradation gene cluster in Comamonas testosteroni TA441 which consists of aromatic-
compound degradation genes for seco-steroids and 3-ketosteroid dehydrogenase genes.
Appl Environ Microbiol 69: 4421–4430.
Horinouchi, M., Hayashi, T., Koshino, H., Yamamoto, T., and Kudo, T. (2003b) Gene
encoding the hydrolase for the product of the meta-cleavage reaction in testosterone
degradation by Comamonas testosteroni. Appl Environ Microbiol 69: 2139-2152.
Horinouchi, M., Kurita, T., Yamamoto, T., Hatori, E., Hayashi, T., and Kudo, T. (2004a)
Steroid degradation gene cluster of Comamonas testosteroni consisting of 18 putative
genes from meta-cleavage enzyme gene tesB to regulator gene tesR. Biochem Biophys
Res Commun 324: 597-604.
Horinouchi, M., Hayashi, T., and Kudo, T. (2004b) The genes encoding the hydroxylase of 3-
hydroxy-9,10-secoandrosta-1,3,5(10)-triene-9,17-dione in steroid degradation in
Comamonas testosteroni TA441. J Steroid Biochem Mol Biol 92: 143-154.
This article is protected by copyright. All rights reserved.

25
Horinouchi, M., Hayashi, T., Koshino, H., Kurita, T., and Kudo, T. (2005) Identification of
9,17-dioxo-1,2,3,4,10,19-hexanorandrostan-5-oic acid, 4-hydroxy-2-oxohexanoic acid,
and 2-hydroxyhexa-2,4-dienoic acid and related enzymes involved in testosterone
degradation in Comamonas testosteroni TA441. Appl Environ Microbiol 71: 5275-5281.
Jackson, M., Reinaldo Camacho, L., Gicquel, B., and Guilhot, C. (2001) Gene replacement
and transposon delivery using the negative selection marker sacB. Methods Mol Med 54:
59-75.
Johnston, J.B., Ouellet, H., and Ortiz de Montellano, P.R. (2010) Functional redundancy of
steroid C26-monooxygenase activity in Mycobacterium tuberculosis revealed by
biochemical and genetic analyses. J Biol Chem 285: 36352-36360.
Kendall, S.L., Withers, M., Soffair, C.N., Moreland, N.J., Gurcha, S., Sidders, B., et al.
(2007) A highly conserved transcriptional repressor controls a large regulon involved in
lipid degradation in Mycobacterium smegmatis and Mycobacterium tuberculosis. Mol
Microbiol 65: 684–699.
Kendall, S.L., Burgess, P., Balhana, R., Withers, M., Ten Bokum, A., Lott, J.S., et al. (2010)
Cholesterol utilization in mycobacteria is controlled by two TetR-type transcriptional
regulators: kstR and kstR2. Microbiology 156: 1362–1371.
Klein, U., Gimpl, G., and Fahrenholz, F. (1995) Alteration of the myometrial plasma
membrane cholesterol content with b-cyclodextrin modulates the binding affinity of the
oxytocin receptor. Biochemistry 34: 13784–13793.
Klepp, L.I., Forrellad, M.A., Osella, A.V., Blanco, F.C., Stella, E.J., Bianco, M.V., et al.
(2012) Impact of the deletion of the six mce operons in Mycobacterium smegmatis.
Microbes Infect 14: 590-599.
This article is protected by copyright. All rights reserved.

26
Knol, J., Bodewits, K., Hessels, G.I., Dijkhuizen, L., and van der Geize, R. (2008) 3-Keto-
5alpha-steroid Delta(1)-dehydrogenase from Rhodococcus erythropolis SQ1 and its
orthologue in Mycobacterium tuberculosis H37Rv are highly specific enzymes that
function in cholesterol catabolism. Biochem J 410: 339-346.
Kreit, J. (2017) Microbial catabolism of sterols: focus on the enzymes that transform the
sterol 3β-hydroxy-5-en into 3-keto-4-en. FEMS Microbiol Lett 364. doi:
10.1093/femsle/fnx007.
Lack, N., Lowe, E.D., Liu, J., Eltis, L.D., Noble, M.E., Sim, E., and Westwood, I.M. (2008)
Structure of HsaD, a steroid-degrading hydrolase, from Mycobacterium tuberculosis.
Acta Crystallogr Sect F Struct Biol Cryst Commun 64: 2-7.
Lack, N.A., Yam, K.C., Lowe, E.D., Horsman, G.P., Owen, R.L., Sim, E., and Eltis, L.D.
(2010) Characterization of a carbon-carbon hydrolase from Mycobacterium tuberculosis
involved in cholesterol metabolism. J Biol Chem 285: 434-443.
Leu, Y.L., Wang, P.H., Shiao, M.S., Ismail, W., and Chiang, Y.R. (2011) A novel
testosterone catabolic pathway in bacteria. J Bacteriol 193: 4447-4455.
Li, W., Ge, F., Zhang, Q., Ren, Y., Yuan, J., He, J., et al. (2014). Identification of gene
expression profiles in the actinomycete Gordonia neofelifaecis grown with different
steroids. Genome 57: 345-353.
Marsheck, W.J., Kraychy, S., and Muir, R.D. (1972) Microbial degradation of sterols. Appl
Microbiol 23: 72-77.
McLean, K.J., Lafite, P., Levy, C., Cheesman, M.R., Mast, N., Pikuleva IA, et al. (2009) The
Structure of Mycobacterium tuberculosis CYP125: molecular basis for cholesterol
binding in a P450 needed for host infection. J Biol Chem 284: 35524-35533.
This article is protected by copyright. All rights reserved.

27
Merino, E., Barrientos, A., Rodríguez, J., Naharro, G., Luengo, J.M., and Olivera, E.R. (2013)
Isolation of cholesterol- and deoxycholate-degrading bacteria from soil samples:
evidence of a common pathway. Appl Microbiol Biotechnol 97: 891-904.
Mohn, W.W., van der Geize, R., Stewart, G.R., Okamoto, S., Liu, J., Dijkhuizen, L., and
Eltis, L.D. (2008) The actinobacterial mce4 locus encodes a steroid transporter. J Biol
Chem 283: 35368–35374.
Ouellet, H., Guan, S., Johnston, J.B., Chow, E.D., Kells, P.M., Burlingame, A.L., et al. (2010)
Mycobacterium tuberculosis CYP125A1, a steroid C27 monooxygenase that detoxifies
intracellularly generated cholest-4-en-3-one. Mol Microbiol 77: 730–742.
Pandey, A.K., and Sassetti, C.M. (2008) Mycobacterial persistence requires the utilization of
host cholesterol. Proc Natl Acad Sci USA 105: 4376–4380.
Parish, T., and Stoker, N.G. (1998) Electroporation of mycobacteria. Methods Mol Biol 101:
129-144.
Penfield, J.S., Worrall, L.J., Strynadka, N.C., and Eltis, L.D. (2014) Substrate specificities
and conformational flexibility of 3-ketosteroid 9α-hydroxylases. J Biol Chem 289:
25523-25536.
Petrusma, M., Hessels, G., Dijkhuizen, L., and van der Geize, R. (2011) Multiplicity of 3-
Ketosteroid-9α-Hydroxylase enzymes in Rhodococcus rhodochrous DSM 43269 for
specific degradation of different classes of steroids. J Bacteriol 193: 3931-3940.
Petrusma, M., van der Geize, R., and Dijkhuizen, L. (2014) 3-Ketosteroid 9α-hydroxylase
enzymes: Rieske non-heme monooxygenases essential for bacterial steroid degradation.
Antonie Van Leeuwenhoek 106: 157-172.
Philipp, B., Erdbrink, H., Suter, M.J., and Schink, B. (2006) Degradation of and sensitivity to
cholate in Pseudomonas sp. strain Chol1. Arch Microbiol 185: 192-201.
This article is protected by copyright. All rights reserved.

28
Roh, H., and Chu, K. H. (2010) A 17beta-estradiol-utilizing bacterium, Sphingomonas strain
KC8: part I- characterization and abundance in wastewater treatment plants. Environ Sci
Technol 44: 4943-4950.
Rosłoniec, K.Z., Wilbrink, M.H., Capyk, J.K., Mohn, W.W., Ostendorf, M., van der Geize,
R., et al. (2009) Cytochrome P450 125 (CYP125) catalyzes C26-hydroxylation to initiate
sterol side chain degradation in Rhodococcus jostii RHA1. Mol Microbiol 74: 1031–
1043.
Sambrook, J., and Russell, D.W. (2001) Molecular Cloning: A Laboratory Manual, 3rd edn.
New York, NY, USA: Cold Spring Harbor Laboratory Press.
Snapper, S.B., Melton, R.E., Mustafa, S., Klesser, T., and Jacobs, W.R. Jr. (1990) Isolation
and characterizaton of efficient plasmid transformation mutants of Mycobacterium
smegmatis. Mol Microbiol 4: 1911-1919.
Stover, C.K., de la Cruz, V.F., Fuerst, T.R., Burlein, J.E., Benson, L.A., Bennett, L.T., et al.
(1991) New use of BCG for recombinant vaccines. Nature 351: 456-460.
Szentirmai, A. (1990) Microbial physiology of sidechain degradation of sterols. J Ind
Microbiol 6: 101-115.
Tarlera, S., and Denner, E.B. (2003) Sterolibacterium denitrificans gen. nov., sp. nov., a
novel cholesterol oxidizing, denitrifying member of the beta-Proteobacteria. Int J Syst
Evol Microbiol 53: 1085-1091.
Thompson, J.D., Higgins, D.G., and Gibson, T.J. (1994) CLUSTAL W: improving the
sensitivity of progressive multiple sequence alignment through sequence weighting,
position-specific gap penalties and weight matrix choice. Nucl Acids Res 22: 4673–4680.
Uhía, I., Galán, B., Morales, V., and García, J.L. (2011) Initial step in the catabolism of
cholesterol by Mycobacterium smegmatis mc2155. Environ Microbiol 13: 943–59.
This article is protected by copyright. All rights reserved.

29
Uhía, I., Galán, B., Kendall, S.L., Stoker, N.G., and García, J.L. (2012) Cholesterol
metabolism in Mycobacterium smegmatis. Environ Microbiol Rep 4: 168-182.
van der Geize, R., Hessels, G.I., van Gerwen, R., Vrijbloed, J.W., van Der Meijden, P., and
Dijkhuizen, L. (2000) Targeted disruption of the kstD gene encoding a 3-kestosteroid
delta(1)-dehydrogenase isoenzyme of Rhodococcus erythropolis strain SQ1. Appl
Environ Microbiol 66: 2029-2036.
van der Geize, R., Hessels, G.I., van Gerwen, R., van der Meijden, P., and Dijkhuizen, L.
(2001) Unmarked gene deletion mutagenesis of kstD, encoding 3-ketosteroid Delta1-
dehydrogenase, in Rhodococcus erythropolis SQ1 using sacB as counter-selectable
marker. FEMS Microbiol Lett 205: 197-202.
van der Geize, R., Hessels, G.I., and Dijkhuizen L. (2002) Molecular and functional
characterization of the kstD2 gene of Rhodococcus erythropolis SQ1 encoding a second
3-ketosteroid Δ1- dehydrogenase isoenzyme. Microbiology 148: 3285-3592.
van der Geize, R., Yam, K., Heuser, T., Wilbrink, M.H., Hara, H., Anderton, M.C., et al.
(2007) A gene cluster encoding cholesterol catabolism in a soil actinomycete provides
insight into Mycobacterium tuberculosis survival in macrophages. Proc Natl Acad Sci
USA 104: 1947-1952.
van der Geize, R., Grommen, A.W., Hessels, G.I., Jacobs, A.A., and Dijkhuizen, L. (2011)
The steroid catabolic pathway of the intracellular pathogen Rhodococcus equi is
important for pathogenesis and a target for vaccine development. PLoS Pathog 7:
e1002181.
Wei, W., Fan, S., Wang, F., and Wei, D. (2010) A new steroid-transforming strain of
Mycobacterium neoaurum and cloning of 3-ketosteroid 9alpha-hydroxylase in NwIB-01.
Appl Biochem Biotechnol 162: 1446-1456.
This article is protected by copyright. All rights reserved.

30
Wilbrink, M.H., Petrusma, M., Dijkhuizen, L., and van derGeize, R. (2011) FadD19 of
Rhodococcus rhodochrous DSM43269, a steroid-coenzyme A ligase essential for deg-
radation of C-24 branched sterol side chains. Appl EnvironMicrobiol 77: 4455–4464.
Wirth, R., Friesenegger, A., and Fieldler, S. (1989) Transformation of various species of
gram-negative bacteria belonging to 11 different genera by electroporation. Mol Gen
Genet 216: 175–177.
Yam, K.C, D'Angelo, I., Kalscheuer, R., Zhu, H., Wang, J.X., Snieckus, V., et al. (2009)
Studies of a ring-cleaving dioxygenase illuminate the role of cholesterol metabolism in the
pathogenesis of Mycobacterium tuberculosis. PLoS Pathog. 5: e1000344.
Yao, K., Xu, L.Q., Wang, F.Q., and Wei, D.Z. (2014) Characterization and engineering of 3-
ketosteroid-△1-dehydrogenase and 3-ketosteroid-9α-hydroxylase in Mycobacterium
neoaurum ATCC 25795 to produce 9α-hydroxy-4-androstene-3,17-dione through the
catabolism of sterols. Metab Eng 24: 181-191.
Yeh, C.H., Kuo, Y.S., Chang, C.M., Liu, W.H., Sheu, M.L., and Meng, M. (2014) Deletion of
the gene encoding the reductase component of 3-ketosteroid 9α-hydroxylase in
Rhodococcus equi USA-18 disrupts sterol catabolism, leading to the accumulation of 3-
oxo-23,24-bisnorchola-1,4-dien-22-oic acid and 1,4-androstadiene-3,17-dione. Microb
Cell Fact 13: 130.
Yuan, J., Chen, G., Cheng, S., Ge, F., Qiong, W., Li, W., and Li, J. (2015) Accumulation of
9α-hydroxy-4-androstene-3,17-dione by co-expressing kshA and kshB encoding
component of 3-ketosteroid-9α-hydroxylase in Mycobacterium sp. NRRL B-3805. Sheng
Wu Gong Cheng Xue Bao 31: 523-533 (article in Chinese).
This article is protected by copyright. All rights reserved.
31
TABLES
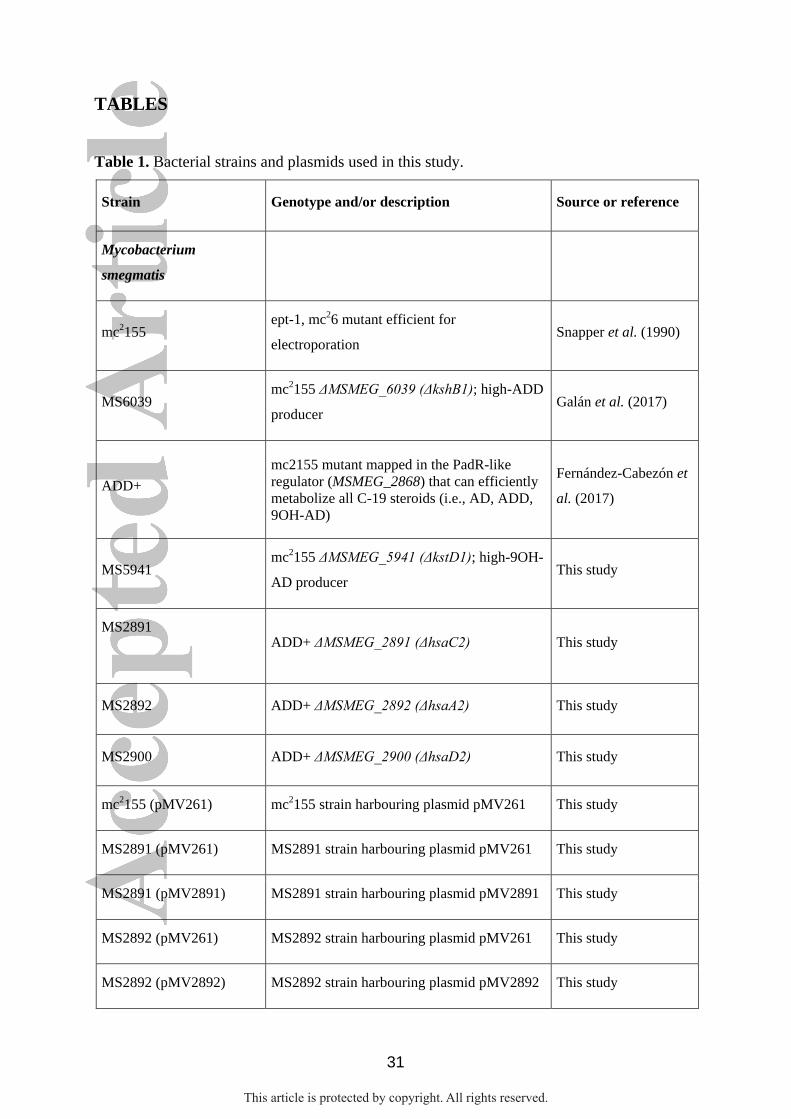
Table 1. Bacterial strains and plasmids used in this study.
Strain Genotype and/or description Source or reference
Mycobacterium
smegmatis
mc2155 ept-1, mc26 mutant efficient for
electroporation Snapper et al. (1990)
MS6039 mc2155 ΔMSMEG_6039 (ΔkshB1); high-ADD
producer Galán et al. (2017)
ADD+
mc2155 mutant mapped in the PadR-like
regulator (MSMEG_2868) that can efficiently
metabolize all C-19 steroids (i.e., AD, ADD,
9OH-AD)
Fernández-Cabezón et
al. (2017)
MS5941 mc2155 ΔMSMEG_5941 (ΔkstD1); high-9OH-
AD producer This study
MS2891
ADD+ ΔMSMEG_2891 (ΔhsaC2) This study
MS2892 ADD+ ΔMSMEG_2892 (ΔhsaA2) This study
MS2900 ADD+ ΔMSMEG_2900 (ΔhsaD2) This study
mc2155 (pMV261) mc2155 strain harbouring plasmid pMV261 This study
MS2891 (pMV261) MS2891 strain harbouring plasmid pMV261 This study
MS2891 (pMV2891) MS2891 strain harbouring plasmid pMV2891 This study
MS2892 (pMV261) MS2892 strain harbouring plasmid pMV261 This study
MS2892 (pMV2892) MS2892 strain harbouring plasmid pMV2892 This study
This article is protected by copyright. All rights reserved.
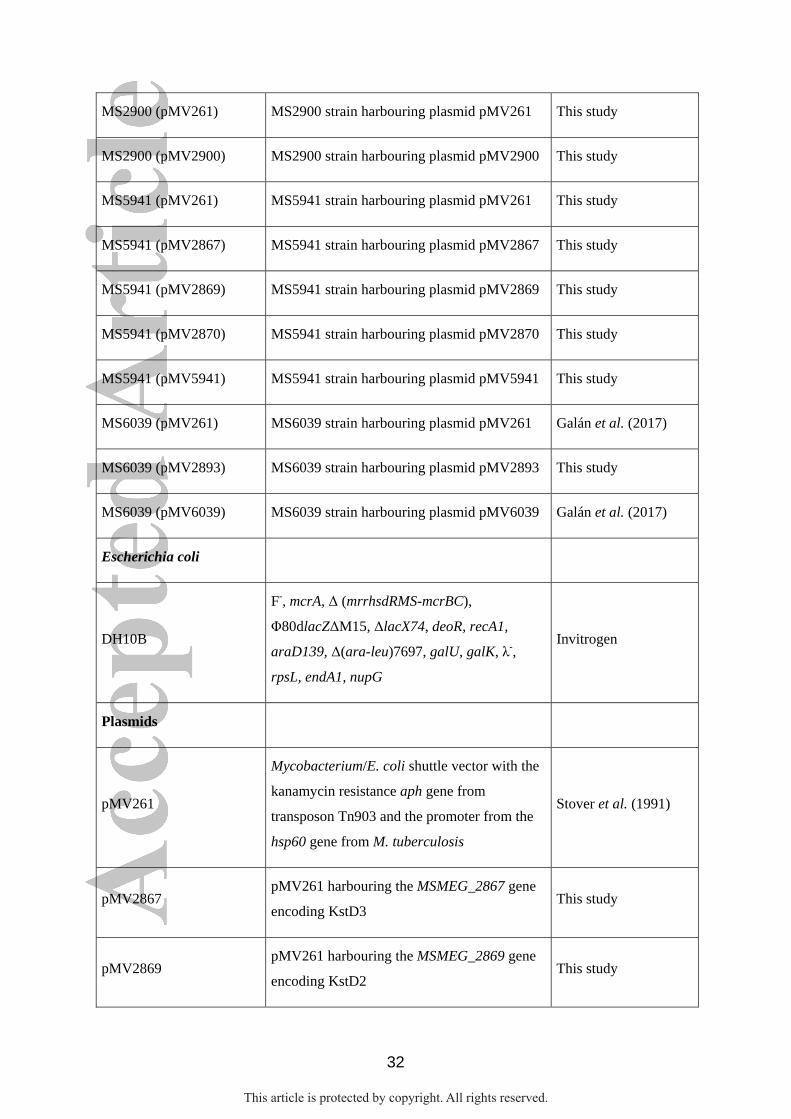
32
MS2900 (pMV261) MS2900 strain harbouring plasmid pMV261 This study
MS2900 (pMV2900) MS2900 strain harbouring plasmid pMV2900 This study
MS5941 (pMV261) MS5941 strain harbouring plasmid pMV261 This study
MS5941 (pMV2867) MS5941 strain harbouring plasmid pMV2867 This study
MS5941 (pMV2869) MS5941 strain harbouring plasmid pMV2869 This study
MS5941 (pMV2870) MS5941 strain harbouring plasmid pMV2870 This study
MS5941 (pMV5941) MS5941 strain harbouring plasmid pMV5941 This study
MS6039 (pMV261) MS6039 strain harbouring plasmid pMV261 Galán et al. (2017)
MS6039 (pMV2893) MS6039 strain harbouring plasmid pMV2893 This study
MS6039 (pMV6039) MS6039 strain harbouring plasmid pMV6039 Galán et al. (2017)
Escherichia coli
DH10B
F-, mcrA, Δ (mrrhsdRMS-mcrBC),
Φ80dlacZΔM15, ΔlacX74, deoR, recA1,
araD139, Δ(ara-leu)7697, galU, galK, λ-,
rpsL, endA1, nupG
Invitrogen
Plasmids
pMV261
Mycobacterium/E. coli shuttle vector with the
kanamycin resistance aph gene from
transposon Tn903 and the promoter from the
hsp60 gene from M. tuberculosis
Stover et al. (1991)
pMV2867 pMV261 harbouring the MSMEG_2867 gene
encoding KstD3 This study
pMV2869 pMV261 harbouring the MSMEG_2869 gene
encoding KstD2 This study
This article is protected by copyright. All rights reserved.

33
pMV2870 pMV261 harbouring the MSMEG_2870 gene
encoding KshA2 This study
pMV2891 pMV261 harbouring the MSMEG_2891 gene
encoding HsaC2 This study
pMV2892 pMV261 harbouring the MSMEG_2892 gene
encoding HsaA2 This study
pMV2893 pMV261 harbouring the MSMEG_2893 gene
encoding KshB2 This study
pMV2900 pMV261 harbouring the MSMEG_2900 gene
encoding HsaD2 This study
pMV5941 pMV261 harbouring the MSMEG_5941 gene
encoding KstD1 This study
pMV6039 pMV261 harbouring the MSMEG_6039 gene
encoding KshB1 Galán et al. (2017)
pJQ200x
Suicide vector used to perform allelic
exchange mutagenesis in mycobacteria, P15A
ori, sacB, GmR, xylE
Jackson et al. (2001)
pJQ2891
pJQ200x containing one fragment upstream
and another fragment downstream of
MSMEG_2891 gene
This study
pJQ2892
pJQ200x containing one fragment upstream
and another fragment downstream of
MSMEG_2892 gene
This study
pJQ2900
pJQ200x containing one fragment upstream
and another fragment downstream of
MSMEG_2900 gene
This study
This article is protected by copyright. All rights reserved.

34
pJQ5941
pJQ200x containing one fragment upstream
and another fragment downstream of
MSMEG_5941 gene
Galán et al. (2017)
This article is protected by copyright. All rights reserved.

35
FIGURE LEGENDS
Fig. 1. Catabolic pathway of C-19 steroids in M. smegmatis mc2155. A) Scheme of the C-19+
route proposed for the degradation of C-19 steroids (i.e., AD, ADD and 9OH-AD) in this
bacterium. The upper catabolic pathway consisting of the degradation of A/B rings of C-19
steroids is carried out by enzymes encoded by the C-19+ gene cluster. The resulting metabolic
intermediates (HHD and HIP) converge with the cholesterol catabolic pathway where the
degradation of C/D steroid rings occurs. The upper catabolic pathway is transcriptionally
regulated by the PadR repressor (padR regulon), while the lower catabolic pathway is
controlled by the KstR2 regulator (kstR2 regulon). B) Comparative genomic analysis of genes
encoding A/B steroid ring modification enzymes in the catabolism of cholesterol or C-19
steroids in M. smegmatis. Black numbers indicate the relative position of these gene clusters
in the genome of M. smegmatis (NC_008596.1). Homologous genes are shown with the same
color. Gene annotation: MSMEG_(number). Protein identities are indicated in parentheses.
Abbreviations of enzymes: 3-ketosteroid-Δ1-dehydrogenase (KstD); 3-ketosteroid-9α-
hydroxylase (KshAB); 3-HSA monooxygenase (HsaAB); 3,4-HSA dioxygenase (HsaC); (4,9-
DSHA) hydroxylase (HsaD); 2-hydroxy-2,4-hexadienoic acid hydratase (HsaE); 4-hydroxy-2-
hydroxy-2-ketovalerate aldolase (HsaF); acetaldehyde dehydrogenase (HsaG). Abbreviations
of metabolites: 4-androstene-3,17-dione (AD); 1,4-androstadiene-3,17-dione (ADD); 9α-
hydroxy-4-androstene-3,17-dione (9OH-AD); 9α-hydroxy-1,4-androstadiene-3,17-dione
(9OH-ADD); 3-hydroxy-9,10-secoandrosta-1,3,5(10)-trien-9,17-dione (3-HSA); 3,4-
dihydroxy-9,10-secoandrosta-1,3,5(10)-trien-9,17-dione (3,4-HSA); 4,5,9,10-diseco-3-
hydroxy-5,9,7-trioxoandrosta-1(10),2-diene-4-oic acid (4,9-DSHA); 2-hydroxy-2,4-
hexadienoic acid (HHD); 3aα-H-4α(3´-propionic acid)-7aβ-methylhexahydro-1,5-indanedione
(HIP).
This article is protected by copyright. All rights reserved.

36
Fig. 2. Gene complementation assays of MS5941 and MS6039 mutant strains in shake flasks
with minimal medium containing 1.8 mM cholesterol as the only carbon and energy source.
(A) Gene complementation of MS5941 (ΔMSMEG_5941, ΔkstD1) mutant: mc2155
(pMV261) [wt, black]; MS5941 (pMV261) [MS5941, red]; MS5941 (pMV5941) [KstD1,
blue]; MS5941 (pMV2869) [KstD2, green]; and MS5941 (pMV2867) [KstD3, grey]. (B)
Gene complementation of the MS6039 (ΔMSMEG_6039, ΔkshB1) mutant: mc2155 (pMV261)
[wt, black]; MS6039 (pMV261) [MS6039, red]; MS6039 (pMV6039) [KshB1, blue]; and
MS6039 (pMV2893) [KshB2, green].
Fig. 3. Determination of the biological function of enzymes KstD and KshA by resting-cell
biotransformations. The bioconversion of 1.8 mM AD into ADD (A) or 9-OHAD (B) by
recombinant strains of M. smegmatis was monitored by thin layer chromatography (TLC).
Nomenclature of M. smegmatis strains used: MS5941 (pMV261) [Control]; MS5941
(pMV2867) [KstD3]; and MS5941 (pMV2870) [KshA2].
Fig. 4. Growth curves of mutant strains of M. smegmatis ADD+ on C-19 steroids. (A) The
strains ADD+ [black], ADD+ MS2891 (ΔMSMEG_2891, ΔhsaC2) [red], ADD+ MS2892
(ΔMSMEG_2892, ΔhsaA2) [blue] and ADD+ MS2900 (ΔMSMEG_2900, ΔhsaD2) [green]
were cultivated in shake flasks with minimal medium containing 1.8 mM AD as the sole
carbon and energy source. (B) Photograph taken of a final sample of the bacterial cultures in
the presence of AD.
Fig. 5. Gene complementation of MS2891, MS2892 and MS2900 mutant strains. The knock-
out mutants were complemented in trans with an original copy of the corresponding deleted
This article is protected by copyright. All rights reserved.

37
gene [MS2891c, MS2892c and MS2900c strains; dotted lines] or transformed with the empty
plasmid as control [MS2891v, MS2892v and MS2900v strains; continuous lines]. The
recombinant strains were cultured in shake flasks with minimal medium containing 18 mM
glycerol (A) or 1.8 mM AD (B) as the sole carbon and energy sources. (C) Photograph taken
of a final sample of the bacterial cultures in the presence of AD. (D) Scheme of several key
enzymatic steps postulated in the catabolism of C-19 steroids in M. smegmatis.
This article is protected by copyright. All rights reserved.

38
Supplementary information
Table S1. Oligonucleotides used in this study.
Fig. S1. Scheme of the cholesterol catabolic pathway conventionally proposed in
Actinobateria. The cholesterol pathway has been traditionally considered to take place
through three consecutive stages. Firstly, the side-chain degradation and 3β-dehydrogenation
of the cholesterol molecules occurs in the upper catabolic pathway apparently yielding two
molecules of propionyl-CoA, an acetyl-CoA and the 17-ketosteroid AD. Then, the
modification of A/B rings of AD in the central catabolic pathway leads to the generation of
the HHD and HIP acids. This last compound is mineralized through the lower catabolic
pathway. The expression of the genes encoding the enzymes involved in the side-chain and
the A/B rings degradation as well as those encoding the sterol uptake system, is regulated by
the KstR repressor (kstR regulon), while the expression of the genes responsible for the
catabolism of C/D rings is controlled by the KstR2 repressor (kstR2 regulon). Abbreviations
of enzymes: 3-ketosteroid-Δ1-dehydrogenase (KstD); 3-ketosteroid-9α-hydroxylase
(KshAB); 3-HSA monooxygenase (HsaAB); 3,4-HSA dioxygenase (HsaC); (4,9-DSHA)
hydroxylase (HsaD). Abbreviations of metabolites: 4-androstene-3,17-dione (AD); 1,4-
androstadiene-3,17-dione (ADD); 9α-hydroxy-4-androstene-3,17-dione (9OH-AD); 9α-
hydroxy-1,4-androstadiene-3,17-dione (9OH-ADD); 3-hydroxy-9,10-secoandrosta-1,3,5(10)-
trien-9,17-dione (3-HSA); 3,4-dihydroxy-9,10-secoandrosta-1,3,5(10)-trien-9,17-dione (3,4-
HSA); 4,5,9,10-diseco-3-hydroxy-5,9,7-trioxoandrosta-1(10),2-diene-4-oic acid (4,9-DSHA);
2-hydroxyhexa-2,4.dienoic acid (HHD); 3aα-H-4α(3´-propionic acid)-7aβ-methylhexahydro-
1,5-indanedione (HIP).
Fig. S2. Chemical structures of relevant steroidal intermediates detected in the culture
medium of several naturally sterols-degrading bacteria. Abbreviations: AD (4-androstene-
This article is protected by copyright. All rights reserved.

39
3,17-dione); ADD (1,4-androstadiene-3,17-dione); 9OH-AD (9α-hydroxy-4-androstene-3,17-
dione); 4-HBC (22-hydroxy-23,24-bisnorchol-4-en-3-one); 1,4-HBC (22-hydroxy-23,24-
bisnorchol-1,4-dien-3-one); 4-BNC (3-oxo-23,24-bisnorchol-4-en-3-oic acid); 1,4-BNC (3-
oxo-23,24-bisnorchol-1,4-dien-3-oic acid).
This article is protected by copyright. All rights reserved.
KstD2/ KstD3
HsaA2B1
ADD AD
9OH-ADD 9OH-AD 3-HSA
KshA2B2
HsaC2
3,4-HSA HHD
KshA2B2
KstD2/ KstD3 HsaD2
HIP
LOWER
CATABOLIC
PATHWAY
Propionyl-CoA
Pyruvate
HsaE, HsaF,
HsaG
4,9-DSHA
Non-
enzymatic
Fig. 1 Catabolic pathway of C-19 steroids in M. smegmatis mc2155.
(A)
(B) Catabolism of cholesterol
6110566 6102080
ksh
B1
hsa
D1
hsa
B1
60
40
kstR
hsa
C1
hsa
A1
60
41
5997153 6001720
kstD
1
hsa
E
hsa
H
hsa
G
ksh
A1
5985003..5986154
(41)
ksh
A2
kstD
2
cv
c2
pa
dR
kstD
3
iclR
hsa
C2
hsa
A2
ksh
B2
hsa
D2
28
88
2928126
Catabolism of C-19 steroids
2963487..2964347 2957439 2952197 2933769 (55) (40) (50) (44) (59) (36)
This article is protected by copyright. All rights reserved.
(A) (B)
0.0
0.5
1.0
1.5
2.0
2.5
0 20 40 60 80 100
OD
(600 n
m)
Time of culture (h)
wt MS5941 KstD1
KstD2 KstD3
0.0
0.5
1.0
1.5
2.0
0 10 20 30 40 50 60 70
OD
(600 n
m)
Time of culture (h)
wt MS6039 KshB1 KshB2
Fig. 2. Gene complementation assays of MS5941 and MS6039 mutants.
This article is protected by copyright. All rights reserved.
AD
ADD
Control KstD3
MS5941 Control KshA2
MS5941
9OH-AD
AD
5 h 24 h
(A) (B) AD ADD AD 9OH-AD
Fig. 3. Determination of the biological function of enzymes KstD and KshA by
resting-cell biotransformations.
This article is protected by copyright. All rights reserved.
(A) (B)
0.0
0.5
1.0
1.5
2.0
2.5
0 10 20 30 40 50
OD
(600 n
m)
Time of culture (h)
ADD+ MS2891 MS2900 MS2892
ADD+ MS2891 MS2892 MS2900
Fig. 4 Growth curves of mutant strains of M. smegmatis ADD+ on C-19 steroids.
This article is protected by copyright. All rights reserved.
(A) (B)
0.0
1.0
2.0
3.0
4.0
0 10 20 30 40
OD
(600 n
m)
Time of culture (h)
MS2891v MS2892v MS2900v
MS2891c MS2892c MS2900c
0.0
0.5
1.0
1.5
2.0
0 10 20 30 40 50 60 70
OD
(600 n
m)
Time of culture (h)
MS2891v MS2892v MS2900v
MS2891c MS2892c MS2900c
Fig. 5. Gene complementation of MS2891, MS2892 and MS2900 mutant strains.
(C)
MS2891v MS2892v MS2900v MS2891c MS2892c MS2900c
HsaD2 3-HSA 3,4-HSA 4,9-HSA
HsaA2B1 HsaC2
MS2892 MS2891
HHD
+ HIP
MS2900
(D)
This article is protected by copyright. All rights reserved.





























