Embed Size (px)
Citation preview

Zbl. Vet. Med. A, 24, 789-798 (1977) @ 1977 Verlag Paul Parey, Berlin und Hamburg ISSN 0300-871 l/ASTM-Coden: ZVRAAX
Aus dem Institut fur Physiologie, Physiologische Chemie und Ernahrungsphysiologie
im Fachbereich Tiermedizin der Universitat Munchen
Untersuchungen zur Genese und Biochemie der Pansenacidose 4. Verteilung von D(-)Milchsaure nach Infusion bei Ziege, Schaf und Rind'
Von
M. STANGASSINGER und D. GIESECKE
Mit 4 Abbildungen und einer Tabelle
(Eingegangen am 12. Juli 1977)
D(-)Milchsaure ist nach den bisherigen Kenntnissen ein physiologisch nor- males Zwischenprodukt der Pansenfermentation, das von lactilytischen Bak- terien mit der gleichen - relativ niedrigen - Rate metabolisiert wird wie das L( +)Isomer (15). Bei Oberfiitterung mit Getreide kann Milchsaure im Pansen akkumulieren und zu schweren Schadigungen des Organs fuhren (8, 4, 6). Unter solchen Bedingungen nimmt der Isomerenanteil von D-Milchsaure mit sinkendem pH-Wert bis zu rund 50 O/o bei pH 4,5 zu (10). Die Resorption von Milchsaure in groi3eren Mengen fuhrt zur metabolischen Acidose mit charak- teristischen klinischen Erscheinungen, die insbesondere der D-Milchsaure zu- geschrieben werden (8,5,2,2O).
Hinsichtlich der acidotischen Wirkungsweise, die auf der chemischen Natur der Milchsaure als einer im Organismus weitgehend dissozierten or@- nischen Saure (pk = 3,85) beruht, kann zwischen D- und L-Milchsaure kein Unterschied bestehen. Maagebliche Diff erenzen zwischen beiden Isomeren mus- sen indessen in Hinblick auf ihre Beseitigung aus dem Organismus erwartet werden, weil nur L-Lactat als physiologische Form gilt, das D-Isomer hingegen nicht. Zwar ist bekannt, dai3 D-Milchsaurespiegel im Blut langer bestehen bleiben (81, aber uber die Wege der Elimination von D-MiIchsaure nach der Resorption liegen keine quantitativ verwertbaren Kenntnisse vor. In Fort- setzung fruherer Arbeiten wurden darum Infusionsversuche teils unter Ver- wendung von 1%-markierter D-Milchsaure durchgefuhrt, um zunachst die Verteilung (diese Arbeit) und spater die Eliminationskinetik, die renale Aus- scheidung und die Metabolisierung zu beschreiben. Fruheren Ergebnissen ent- sprechend, wurde in allen Infusionsversuchen DL-Milchsaure mit einem Isomerenanteil von 45 o/o D(-) verwendet*.
Mit Unterstutzung der Deutschen Forschungsgemeinschaft. * Fur die Oberlassung der Sonderanfertigung wird der Firma Boehringer und Sohne, Ingel- heim, gedankt
Zbl. Vet. Med., Reihe A, Bd. 24, Heft 10 54

790 STANGASSINGER und GIESECKE
Material und Methoden Versucbstiere und Futterung
Die Untersuchungen wurden an 3 erwachsenen Hammeln der Merino- landrasse (Q 91, T 76 und V 77 kg Korpergewicht), 4 erwachsenen ovarekto- mierten Deutschen Edelziegen (B und D je 30, A und C je 32 kg) und 1 Deut- schen Fleckviehkuh (570 kg) durchgefuhrt. Alle Tiere erhielten 2mal taglich Wiesenheu ad libitum; das Rind bekam zusatzlich 2mal taglich eine Kraft- futterration (1 kg DLG 111). Die Durchfuhrung der Versuche erfolgte etwa 15 Stunden nach der letzten Futterung in gewohnter Umgebung der Tiere.
Durch fuhrung der In fusionsversuche Die Versuche wurden annahernd zur gleichen Tageszeit, morgens zwischen
8 und 9 Uhr, begonnen. Die Infusionen erfolgten mit Hilfe einer Walzen- pumpe uber einen Polyathylendauerkatheter in die V. jugularis. Bei Dauer- infusionen wurde dem uber mehrere Stunden konstanten Dosisstrom eine Kurzzeitinfusion (,,priming dose") vorausgeschickt. Die Infusionslosungen waren in allen Versuchen auf p H 4,O teilneutralisierte (mit N a O H und KOH im Verhaltnis 20 : 1) racemische Milchsaurelosungen mit konstantem Isomeren- verhaltnis (55 O/o L( +)- und 45 " / o D( -)Milchsaure) aber unterschiedlichem Isomerengehalt.
Blutprobennahme und Analytik Die in regelmafiigen Abstanden aus der gegenuberliegenden V. jugularis
gewonnenen Blutproben wurden zur Enteiweiaung und Konservierung sofort mit gleichen Volumina 0,6 N HClO, versetzt, zentrifugiert, und die klaren Oberstande zur weiteren Aufarbeitung bei + 4 O C gelagert. Die Bestimmung von L( +)- und D( -)Milchsaure erfolgte enzymatisch, in Anlehnung an HOHORST, GAWEHN und BERGMEYER (II) , nach den Angaben der Fa. Boehrin- ger mit L( +)Lactatdehydrogenase (EC 1.1.1.27) aus Schweinemuskel und D( - )Lactatdehydrogenase (EC 1.1.1.28) aus Lactobacillus leichmanii. Be- stimmungen der ',C-Aktivitat und der spezifischen Aktivitat (SA) werden in der folgenden Arbeit (5. Mitteilung) dargelegt.
Begriffserlauterungen zur Verteilung Urn die Verteilung von D-Milchsaure zu beschreiben, wurden folgende
Grofien verwendet: Verteilungsraum (V) und Distributionskoeffizient (A) bezeichnen das-
jenige Volumen pro Tier bzw. pro kg Korpergewicht, das Milchsaure bei an- genommener Gleichverteilung im Organismus beanspruchen wurde:
(D = Dosis; G = Korpergewicht; Tf = Transfer = Dosisstrom; yx = Kon- zentrationsplateau im Blut bei Dauerinfusion; a = fiktive Anfangskonzen- tration im Blut nach Kurzzeitinfusion; k = Eliminationskonstante).
Kompartiment (Kp) ist eine Fraktion von V, die einen anatomisch um- grenzten Bezirk (z. B. Organ), bestimmte histologische Strukturen (2. B. Ge- webe) oder auch ein biokinetisch und funktionell einheitliches Ganzes dar- stellen kann:
Kp, + Kp, + . . . + Kpn = V (1)
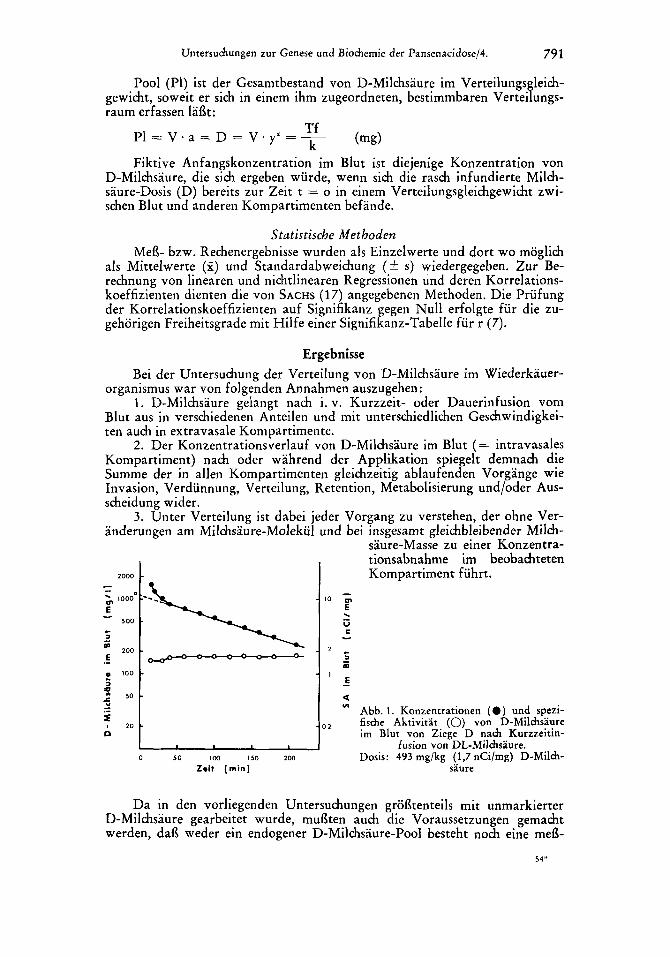
Untersuhungen zur Genese und Biohemie der Pansenacidose/4. 791
Pool (Pl) ist der Gesamtbestand von D-Milchsaure im Verteilungsgleich- gewicht, soweit er sich in einem ihm zugeordneten, bestimmbaren Verteilungs- raum erfassen lafit:
Tf k (mg) p1 = V - a = D = V . y" = -
Fiktive Anfangskonzentration im Blut ist diejenige Konzentration von D-Milchsaure, die sich ergeben wurde, wenn sich die rasch infundierte Milch- saure-Dosis (D) bereits zur Zeit t = o in einem Verteilungsgleichgewicht zwi- schen Blut und anderen Kompartimenten befande.
Statistische Methoden Me% bzw. Rechenergebnisse wurden als Einzelwerte und dort wo moglich
als Mittelwerte (Z) und Standardabweichung ( _+ s) wiedergegeben. Zur Be- rechnung von linearen und nichtlinearen Regressionen und deren Korrelations- koeffizienten dienten die von SACHS (1 7) angegebenen Methoden. Die Prufung der Korrelationskoeffizienten auf Signifikanz gegen Null erfolgte fur die zu- gehorigen Freiheitsgrade mit Hilfe einer Signifikanz-Tabelle fur r (7).
Ergebnisse Bei der Untersuchung der Verteilung von D-Milchsaure im Wiederkauer-
organismus war von folgenden Annahmen auszugehen: 1. D-Milchsaure gelangt nach i. v. Kurzzeit- oder Dauerinfusion vom
Blut aus in verschiedenen Anteden und mit unterschiedlichen Geschwindigkei- ten auch in extravasale Kompartimente.
2. Der Konzentrationsverlauf von D-Milchsaure im Blut (= intravasales Kompartiment) nach oder wahrend der Applikation spiegelt demnach die Summe der in alien Kompartimenten gleichzeitig ablaufenden Vorgange wie Invasion, Verdunnung, Verteilung, Retention, Metabolisierung und/oder Aus- scheidung wider.
3. Unter Verteilung ist dabei jeder Vorgang zu verstehen, der ohne Ver- anderungen am Milchsaure-Molekiil und bei insgesamt gleichbleibender Milch-
saure-Masse zu einer Konzentra- tionsabnahme im beobahteten Kompartiment fuhrt.
0 5 0 100 150 2 0 0
Zeit (rnin)
Abb. 1 . Konzentrationen ( 0 ) und spezi- fische Aktivirat (0) yon D-Milmsaure irn Blut von Ziege D na& Kurzzeitin-
fusion von DL-Mildwaure. Dosis: 493 rng/kg (1,7 nCi/mg) D-Mikh-
saure
Da in den vorliegenden Untersuchungen groi3tenteils mit unmarkierter D-Milchsaure gearbeitet wurde, mufiten auch die Voraussetzungen gemacht werden, dai3 weder ein endogener D-Milchsaure-Pool besteht noch eine mefi-
54c
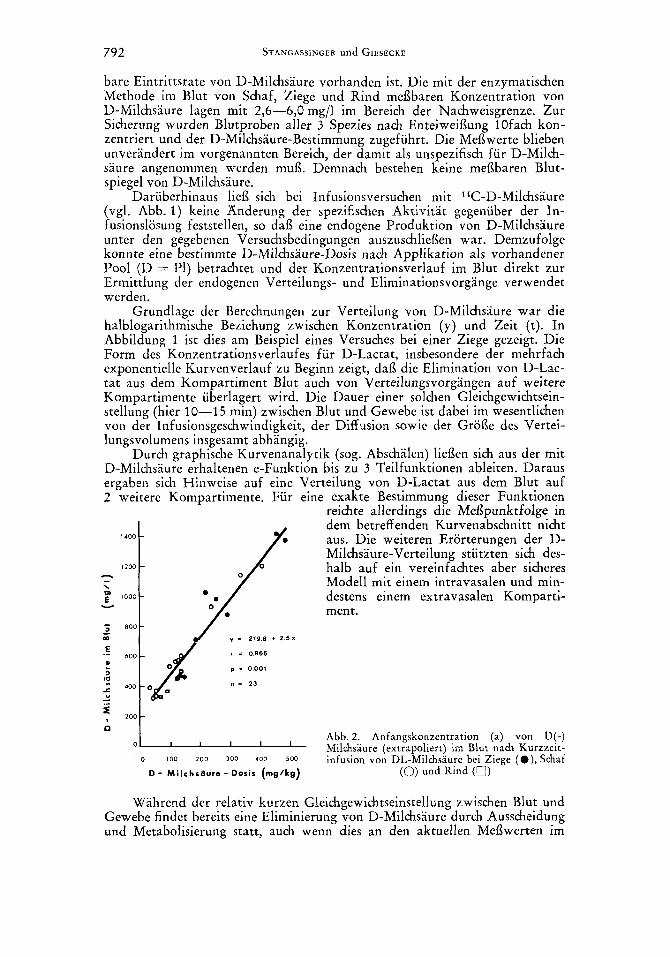
792 STANGASSINGER und GIESECKE
: 200
0
- Abb. 2. Anfangskonzentration (a) von D(-)
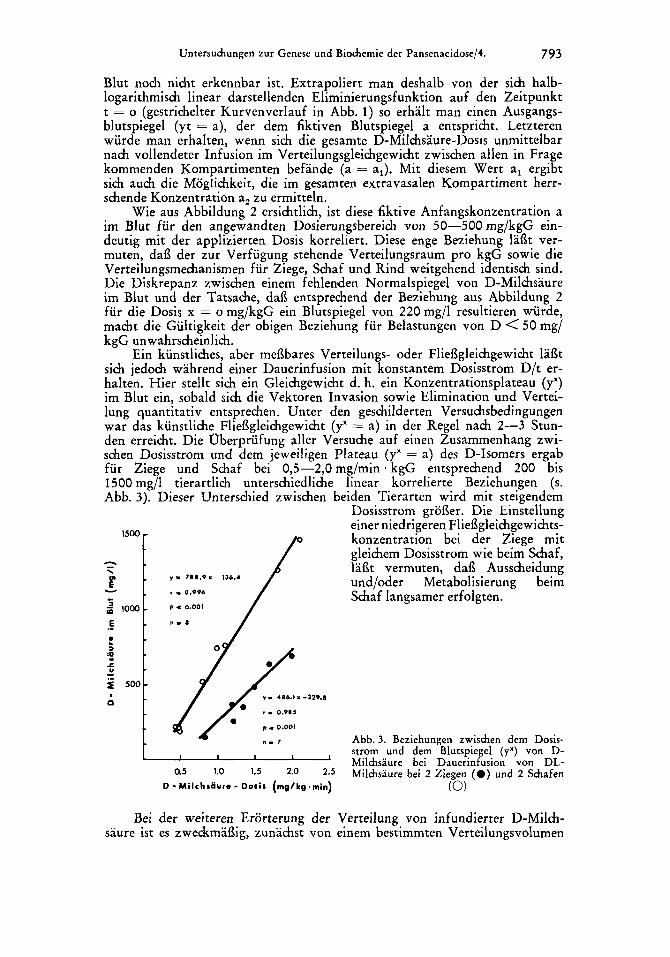
Untersuchungen zur Genese und Biochemie der Pansenacidose/4. 793
Blut noch nicht erkennbar ist. Extrapoliert man deshalb von der sich halb- logarithmisch linear darstellenden Eliminierungsfunktion auf den Zeitpunkt t = o (gestrichelter Kurvenverlauf in Abb. 1) so erhalt man einen Ausgangs- blutspiegel (yt = a), der dem fiktiven Blutspiegel a entspricht. Letzteren wurde man erhaIten, wenn sich die gesamte D-Milchsaure-Dosis unmittelbar nach vollendeter Infusion im Verteilungsgleichgewicht zwischen allen in Frage kommenden Kompartimenten befande (a = al). Mit diesem Wert a, ergibt sich auch die Moglichkeit, die im gesamten extravasalen Kompartiment herr- schende Konzentration a, zu ermitteln.
Wie aus Abbildung 2 ersichtlich, ist diese fiktive Anfangskonzentration a im Blut fur den angewandten Dosierungsbereich von 50-500 mg/kgG ein- deutig mit der applizierten Dosis korreliert. Diese enge Beziehung 12Bt ver- muten, dai3 der zur Verfugung stehende Verteilungsraum pro kgG sowie die Verteilungsmehanismen fur Ziege, Schaf und Rind weitgehend identisch sind. Die Diskrepanz zwischen einem fehlenden Normalspiegel von D-Milchsaure im Blut und der Tatsache, dafi entsprechend der Beziehung aus Abbildung 2 fur die Dosis x = o mg/kgG ein Blutspiegel von 220 mg/l resultieren wurde, macht die Giiltigkeit der obigen Beziehung fur Belastungen von D < 50 mg/ kgG unwahrscheinlich.
Ein kunstliches, aber mei3bares Verteilungs- oder Fliei3gleichgewicht laat sich jedoch wahrend einer Dauerinfusion mit konstantem Dosisstrom D/t er- halten. Hier stellt sich ein Gleichgewicht d. h. ein Konzentrationsplateau (y") im Blut ein, sobald sich die Vektoren Invasion sowie Elimination und Vertei- lung quantitativ entsprechen. Unter den geschilderten Versuchsbedingungen war das kunstliche Fliefigleichgewicht (y" = a) in der Regel nach 2-3 Stun- den erreicht. Die Oberprufung aller Versuche auf einen Zusammenhang zwi- schen Dosisstrom und dem jeweiligen Plateau (y" = a) des D-Isomers ergab fur Ziege und Schaf bei 0,5-2,0 mg/min kgG entsprechend 200 bis 1500 mg/l tierartlich unterschiedliche linear korrelierte Beziehungen (s. Abb. 3). Dieser Unterschied zwischen beiden Tierarten wird mit steigendem
y 9 718.9 I 136.4
r I 0.996
P - 0.001
y I 486.1 i -229.8
a5 1.0 1.5 2.0 2.5
D -MilchsGure- Doris (mg/kg.min)
Dosisstrom groi3er. Die Einitellung einer niedrigeren Fliefigleichgewihts- konzentration bei der Ziege mit gleichem Dosisstrom wie beim Schaf, lai3t vermuten, dai3 Ausscheidung und/oder Metabolisierung beim Schaf langsamer erfolgten.
Abb. 3. Beziehungen zwischen dem Dosis- strom und dem Blutspiegel (y") von D- Milchsaure bei Dauerinfusion von DL- Milchsaure bei 2 Ziegen ( 0 ) und 2 Schafen
(0)
Bei der weiteren Erorterung der Verteilung von infundierter D-Milch- saure ist es zweckmaflig, zunachst von einem bestimmten Verteilungsvolumen
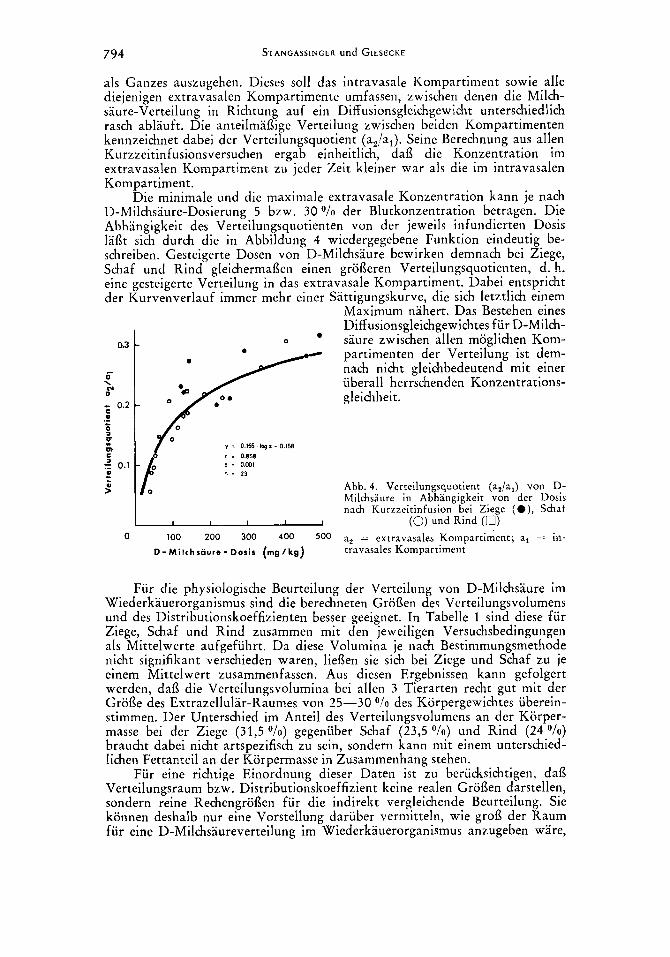
794 STANGASSINGER und GIESECKE
als Ganzes auszugehen. Dieses sol1 das intravasale Kompartiment sowie alle diejenigen extravasalen Kompartimente umfassen, zwischen denen die Milch- saure-Verteilung in Richtung auf ein Diff usionsgleichgewicht unterschiedlich rasch ablauft. Die anteilrnaflige Verteilung zwischen beiden Kompartimenten kennzeichnet dabei der Verteilungsquotient (a2/al). Seine Berechnung aus allen Kurzzeitinfusionsversuchen ergab einheitlich, dafl die Konzentration im extravasalen Kompartiment zu jeder Zeit kleiner war als die irn intravasalen Kornpartiment.
Die minimale und die maximale extravasale Konzentration kann je nach D-Milchsaure-Dosierung 5 bzw. 30 o/o der Blutkonzentration betragen. Die Abhangigkeit des Verteilungsquotienten von der jeweils infundierten Dosis Iai3t sich durch die in Abbildung 4 wiedergegebene Funktion eindeutig be- schreiben. Gesteigerte Dosen von D-Milchsaure bewirken demnach bei Ziege, Schaf und Rind gleichermaflen einen grofleren Verteilungsquotienten, d. h. eine gesteigerte Verteilung in das extravasale Kompartiment. Dabei entspricht der Kurvenverlauf immer mehr einer Sattigungskurve, die sich letztlich einem
Maximum nahert. Das Bestehen eines Diff usionsgleichgewichtes fur D-Milch-
partimenten der Verteilung ist dem- nach nicht gleichbedeutend mit einer uberall herrschenden Konzentrations- gleichheit.
0.3 - 0 saure zwischen allen moglichen Kom-
y I 0 165. I q x - 0.158 I = 0.858 p i O.M1 n = 23
Abb. 4. Verteilungsquotient (an/al) von D- Milchsaure in Abhangigkeit von der Dosis nach Kurzzeitinfusion bei Ziege ( O ) , Schat
0 100 200 300 400 500 a, = extravasales Kompartiment; a, = in- I I I I I (0) und Rind (0)
D - M i l c h s a u r e - Doris (mg/kg) travasales Kompartiment
Fur die physiologische Beurteilung der Verteilung von D-Milchsaure im Wiederkauerorganismus sind die berechneten Groflen des Verteilungsvolumens und des Distributionskoeffizienten besser geeignet. In Tabelle 1 sind diese fur Ziege, Schaf und Rind zusammen mit den jeweiligen Versuchsbedingungen als Mittelwerte aufgefuhrt. Da diese Volumina je nach Bestimmungsmethode nicht signifikant verschieden waren, lieflen sie sich bei Ziege und Schaf zu je einem Mittelwert zusammenfassen. Aus diesen Ergebnissen kann gefolgert werden, dafl die Verteilungsvolumina bei allen 3 Tierarten recht gut mit der Grofle des Extrazellular-Raumes von 25-30 O/o des Korpergewichtes uberein- stimmen. Der Unterschied irn Anteil des Verteilungsvolumens an der Korper- masse bei der Ziege (31,5 o/o) gegenuber Schaf (23,5 O/o) und Rind (24 "n) braucht dabei nicht artspezifisch zu sein, sondern kann rnit einem unterschied- lichen Fettanteil an der Korpermasse in Zusammenhang stehen.
Fur eine richtige Einordnung dieser Daten ist zu berucksichtigen, dafl Verteilungsraum bzw. Distributionskoeffizient keine realen Groflen darstellen, sondern reine Rechengroflen fur die indirekt vergleichende Beurteilung. Sie konnen deshalb nur eine Vorstellung daruber vermitteln, wie grofl der Raum fur eine D-Milchsaureverteilung im Wiederkauerorganismus anzugeben ware,
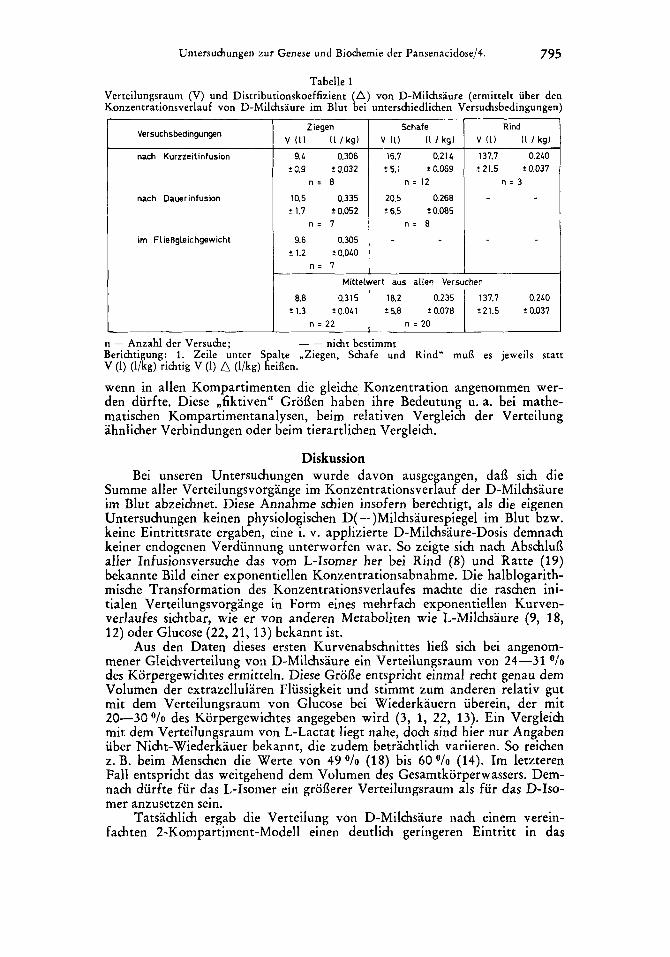
Untersuchungen zur Genese und Biochemie der Pansenacidose/4. 795
8.8 0.315 18.2 0.235 51.3 tO.041 t5.8 20.078
n = 22 n = 20
Tabelle 1 Verteilungsraum (V) und Distributionskoeffizient (A) yon D-Milchsaure (ermittelt uber den Konzen'trationsverlauf von D-Milchsaure im Blut bei unterschiedlichen Versuchsbedingungen)
137.7 0.240 221.5 tO.037
Versuchsbedingungen
nach Kurzzeitinfusion
nach Dawrinfusion
im FLieOgleichgewicht
11 = Anzahl der Versuche:
Ziegen V (11 ( I l k g )
9.4 0.306 r0.9 iO.032
n = 8 10.5 0.335 t 1.7 r0.052
n = 7
9.6 0.305 *- 1.2 *O.OLO
n : 7
Schafe V (1) (1 / kg)
16.7 0.214 t 5.1 r 0.069
n = 12 20.5 0.268 t6.5 20.085
n = 8
Rind V ( 1 ) (1 / kg)
137.7 0.240 t21.5 20.037
n.3
Berichtigung: 1 . Zeile unter Spalte ,Ziegen, Schafe und Rind" mui3 es jeweils statt V (1) (l/kg) richtig V (1)
wenn in allen Kompartimenten die gleiche Konzentration angenommen wer- den durfte. Diese ,fiktiven" Grof3en haben ihre Bedeutung u. a. bei mathe- matischen Kompartimentanalysen, beim relativen Vergleich der Verteilung ahnlicher Verbindungen oder beim tierartlichen Vergleich.
(l/kg) hei8en.
Diskussion Bei unseren Untersuchungen wurde davon ausgegangen, dai3 sich die
Summe aller Verteilungsvorgange im Konzentrationsverlauf der D-Milchsaure im Blut abzeichnet. Diese Annahme schien insofern berechtigt, als die eigenen Untersuchungen keinen physiologischen D( -)Milchsaurespiegel im Blut bzw. keine Eintrittsrate ergaben, eine i. v. applizierte D-Milchsaure-Dosis demnach keiner endogenen Verdunnung unterworfen war. So zeigte sich nach AbschluQ aller Infusionsversuche das vom L-Isomer her bei Rind (8) und Ratte (19) bekannte Bild einer exponentiellen Konzentrationsabnahme. Die halblogarith- mische Transformation des Konzentrationsverlaufes machte die raschen ini- tialen Verteilungsvorgange in Form eines mehrfach exponentiellen Kurven- verlaufes sichtbar, wie er von anderen Metaboliten wie L-Milchsaure (9, 18, 12) oder Glucose (22,21, 13) bekannt ist.
Aus den Daten dieses ersten Kurvenabschnittes liei3 sich bei angenom- mener Gleichverteilung von D-Milchsaure ein Verteilungsraum von 24-31 O / o
des Korpergewihtes ermitteln. Diese Grof3e entspricht einmal recht genau dem Volumen der extrazellularen Flussigkeit und stimmt zum anderen relativ gut mit dem Verteilungsraum von Glucose bei Wiederkauern uberein, der mit 20-30 O / o des Korpergewichtes angegeben wird (3, 1 , 22, 13). Ein Vergleich mit dem Verteilungsraum von L-Lactat liegt nahe, doch sind hier nur Angaben uber Nicht-Wiederkauer bekannt, die zudem betrachtlich variieren. So reichen z. B. beim Menschen die Werte von 49 O / o (18) bis 60 O / o (14). Im letzteren Fall entspricht das weitgehend dem Volumen des Gesamtkorperwassers. Dem- nach durfte fur das L-Isomer ein grof3erer Verteilungsraum als fur das D-ISO- mer anzusetzen sein.
Tatsachlich ergab die Verteilung von D-Milchsaure nach einem verein- fachten 2-Kompartiment-Model1 einen deutlih geringeren Eintritt in das

796 STANGASSINGER u n d GIESECKE
extravasale Kompartiment. Auch bei hochsten Belastungen wurden hier, einer Sattigungskurve folgend (vgl. Abb. 7), nur etwa 30 o/o der Blutkonzentration erreicht. Als Bestatigung fur diese ungleiche Verteilung sind Untersuchungen am isolierten Rattenzwerchfell (16) zu sehen. Sie ergaben zwischen Muskel- fasern und umgebendem Medium fur D-Milchsaure (2,3-118 mM) einen Ver- teilungsquotienten im steady state von innen/auGen = 0,35, und bei Konzen- trationen unter 2,3 mM Werte von = 0,l . Fur das L-Isomer betrug der Quo- tient unter identischen Bedingungen 0,4. Als Ursache werden fur D- und L- Milchsaure verschiedenartige ,,Mechanismen" des Eintritts in die Gewebe diskutiert. Fur die Eliminierung von D-Milchsaure aus dem Organismus des Wiederkauers kommt den erorterten Verteilungsprozessen und dem Eintritt in bestimmte Kompartimente sicherlich eine besondere Bedeutung zu.
Zusammenfassung Um die Verteilung von D(-)Milchsaure im Organismus von Wieder-
kauern zu charakterisieren, wurden an 4 Ziegen, 3 Schafen und 1 Rind intra- venose Infusionen von DL-Milchsaure mit 45 "/n D-Isomer und Berechnungen anhand des Konzentrationsverlaufes im Blut durchgefuhrt.
Fur D-Lactat bestand kein endogener Pool. Der initiale mehrfach expo- nentielle Kurvenverlauf nach Kurzzeitinfusion deutete auf eine Verteilung von D-Milchsaure aus dem Blut auf 2 weitere Kompartimente hin.
Unter Zugrundelegung eines Zwei-Kompartimentmodells ergab sich der Gesamtverteilungsraum bei Ziege, Schaf und Rind zu 9,8, 18,2 und 137,7 1. Diese Werte entsprachen mit 24-3 1 O / o des Korpergewichtes weitgehend dem Extrazehlarraum.
Steigende Dosen von D-Milchsaure bewirkten bei den 3 Tierarten eine Zunahme des Verteilungsquotienten, gemai3 einer Sattigungskurve. Bei maxi- maler intravasaler Konzentration von 1370 mg/l wurde ein Quotient von 0,32 und damit eine extravasale Konzentration von 438 mg/l erreicht.
Die Moglichkeiten einer isomerenspezifischen Verteilung von Milchsaure werden diskutiert.
.
Summary Studies on the genesis and biochemistry of rumen acidosis
4. Distribution of D(-)lactic acid after infusion in goats, sheep and cattle
In order to characterize the distribution of D(-)lactic acid in ruminants intravenous infusions of DL-lactic acid containing 45 "/o of the D-isomer were given to 4 goats, 3 sheep and one bovine and calculations were made from the changes in blood concentration.
For D-lactate there is no endogenous pool. The initially multiple ex- ponential curve following short-time infusion indicates partition of the D- lactic acid from the blood into two further compartments.
On the basis of this two-compartment model, the total partition space in the goat, sheep and bovine was 9.8, 18.2 and 137.7 litres, respectively. These values represented 24-31 O/O of the total extracellular space.
Increasing doses of D-lactic acid in all three species brought about an increase in the partition quotients, conforming to a saturation curve. With a maximal i. v. concentration of 1370 mg./ml. a quotient of 0.32 and thereby an extravascular concentration of 438 mg./l. were reached.
The possibilities of an isomer-specific partition of lactic acid are discussed.

Untersuchungen zur Genese und Biochemie der Pansenacidose/4.
Resume Recherches sur la genhse et la biochimie de l’acidose de la panse
4. RCpartition de D(-)acide lactique aprts infusion chez la chtvre, le mouton et le bovin
On a prockdk B des infusions intraveineuses de DL-acide lactique avec 45 O/o D-isomke chez 4 chkvres, 3 moutons et 1 bovin pour caractkriser la pool endogkne pour D-lactate: Une pool endogkne pour D-lactate n’existe pas. Le cours re kt& initial de la courbe exponentielle a p r b une infusion de courte
partir du sang. Sur la base d’un modkle A deux compartiments, l’ktendue de la rkpartition
zotale fut 9,8 chez la chkvre, 18,2 chez le mouton et 137,71 chez le bovin. Ces valeurs correspondent largement l’espace extracellulaire avec 24-3 1 O/o du poids corporel.
Des doses augmentkes de D-acide lactique occasionnkrent chez les 3 espkces une augmentation des quotients de rkpartition, conformkment ?i une courbe de saturation. Un quotient de 0,32 fut atteint avec une concentration maximale intravasale de 1379 mg/l et il s’en suivit une concentration extra- vasale de 438 mg/l.
Les possibilitks d’une rkpartition isomkrique spkcifique de l’acide lactique sont discutkes.
797
d u d e signi P ia une rkpartition de D-acide lactique sur deux compartiments A
Resumen Estudios sobre la genesis y bioquimica de la acidosis ruminal
4. Distribucidn de icido D(-)ldctico tras infusi6n en cabra, oveja y vaca Con el fin de caracterizar la distribucih de icido D(-)lictico en el
organism0 de rumiantes se llevaron a cab0 infusiones intravenosas de icido DLlictico con 45 O/o del isbmero D en 4 cabras, 3 ovejas y una vaca, y se efectuaron cAlculos a la vista del curso de concentraciones en sangre.
Para lactato D no existia ninguna rebalsa endbgena. El curso exponencial mliltiple inicial de la curva que seguia a una infusiitn de duraci6n corta seiiala hacia la distribucidn de icido D(-)lictico de la sangre a otros dos compartimientos.
Sobre la base de un modelo de dos compartimientos, el espacio de repar- timiento total en cabra, oveja y vaca fuC de 9.8, 18.2 y 137.7 litros, respectiva- mente. Estos valores correspondian con el 24-31 O/O del peso corporal, a grandes rasgos, con el espacio extracelular total.
Dosis crecientes de k i d 0 D(-)lictico ocasionaron en las 3 especies ani- males un aumento del cociente de distribucibn, conforme a una curva de saturacidn. Con concentracih intravasal mixima de 1370 mg/l, se logr6 un cociente de 0.32 y, con ello una concentraci6n extravasal de 438 mg/l.
Se discuten las posibilidades de una distribuci6n isomkrica especifica del icido lictico.
Literaturverzeichnis 1. BERGMANN, E. N., and D. E. HOWE, 1967: Glucose turnover and oxidation rates in
lactating sheep. Am. J. Physiol. 213, 1378-1384. 2. BRAIDE, B. G., and R. H. DUNLOP, 1969: Pharmacologic differences between sodium
D-lactate and sodium L-lactate in sheep. Am. J. Vet. Res. 30, 1281-1288 3. DAVIS, C. L., and R. E. BROWN, 1962: Availability and metabolism of various substrates
in ruminants. IV. Glucose metabolism in the young calf and growing steer. J. Dairy. Sci. 45, 513-516.
4. DIRKSEN, G., 1965: Uber die Pansenacidose des Rindes. Vet. Med. Na&r. 2, 79-108.

798 STANGASSINGER und GIESECKE
5. DIRKSEN, G., 1967: Beitrag zur Biochemie der Pansenacidose des Rindes. Schweiz. Arch.
6. DIRKSEN, G., 1970: Acidosis. In: A. T. PiiiLLirsoN: Physiology of digestion and meta-
7. Documenta Geigy, 1968: Wissenschaftliche Tabellen, 7. Auflage, Redaktion: K. DIEM und
8. DUNLOP. R. H.. and P. B. HAMMOND. 1965: D-lactic acidosis of ruminants. An. N.Y.
Tierheilk. 109, 28-34.
bolism in the ruminant S. 612-625, Oriel Press, Newcastle upon Tyne.
C. LEUTNER, Herausgeber: J. R. Geigy AG, Pharma, Basel.
Acad. Sci. 119; 1109-1132. 9. FORBATH. N.. A. B. KENSHOLE. and G. HETENYI ir.. 1967: Turnover of lactic acid in
normal' and diabetic dogs calculated by two tiac'er methods. Am. J. Physiol. 2f2, 1179-1184.
10. GIESECKE, D., C. BARTELMUS und M. STANGASSINGER, 1976: Untersuchungen zur Genese und Biochemie der Pansenacidose. 3. Stoffwechseldynamik von D(-) und L( +)Milch- saure nach Verabreichung von Zuckern und nach Oberfiitterung mit Srarke. Zbl. Vet. Med. A, 23, 353-363.
11. HOHORST, H . J., GAWEHN und H. U. BERGMEYER, 1970: L-Lactat: Bestimmung mit L D H und N A D , S. 1452. In: H. M. Bergmeyer: Methoden der enzymatischen Analyse. 1. Auflage, Band 11, Verlag Chemie, Weinheim/BergstraRe.
12. HUBBARD, J. L., 1973: The effect of exercise on lactate metabolism. J. Physiol. 231, 1-18. 13. JUDSON, G. J., and R. A. LENG, 1972: Estimation of the total entry rate and resynthesis
of glucose in sheep using glucose uniformly labelled with 3H. Aust. J. Biol. Sci. 25, 1313-1332.
14. KREISBERG, R. A,, J. F. PENNINGTON, and C. R. BASHELL, 1970: Lactate turnover and gluconeogenesis in normal and obese humans. Effect of starvation. Diabetes 19, 53-63.
15. OGIMOTO, K., und D. GIESECKE, 1974: Untersuchungen zur Genese und Biochemie der Pansenacidose. 2. Mitteilung. Zbl. Vet. Med. A, 21, 532-538.
16. Roos, A,, 1975: Intracellular p H and distribution of weak acids across cell membranes. A study of D- and L-lactate and of DMO in rat diaphragm. J. Physiol. 249, 1-25.
17. SACHS, L., 1972: Statistische Auswertungsmethoden. 3. Auflage, Springer-Verlag, Berlin- Heidelberg-New York.
18. SEARLE, G. L , and R. R. CAVALIBRI, 1972: Determination of lactate kinetics in the human; analysis of data from single injection vs. continuous infusion methods. Proc. SOC. Exp. Biol. Med. 239, 1002-1006.
19. SHICHIRI, H., M. MILLER, and M. WOODWARD jr , 1967: Disappearance of DL-Iactate-2-l4C from blood in normal and diabetic rats. Proc. Soc. Exp. Biol. (N.Y.) 125, 1238-1242.
20. TELLE, P. P., and R. L. PRESTON, 1971 : Ovine lactic acidosis intraruminal and systemic. J. Animal Sci. 33, 698-705.
21. WATERHOUSE, CH., and J. KEILPON, 1969: Cory cycle activity in man. J. d i n . Invest.
22. WHITE, R. G., J. W. STEEL, R. A. LENG, and J. R. LUICK, 1969: Evaluation of three isotope-dilution techniques for studying the kinetics of glucose metabolism in sheep. Biochem. J. 214, 203-214.
Anschrift der Verfasser: Institut fur Physiologie, Physiologishe Chemie und Ernah-
48, 2359-2366.
rungsphysiologie, VeterinlrstraBe 13, D-8000 Miinchen 22.

![2011-02 ParasitenKlWdk [Kompatibilit-344tsmodus] · Interzol-orale Suspension für Tiere Rind, Schaf Oxfendazol Panacur 2,5%-Suspension zum Eingeben für Schafe Schaf Fenbendazol](https://img.pdfslide.net/doc/110x75/5e9d7e8c87b9b270c63998f9/2011-02-parasitenklwdk-kompatibilit-344tsmodus-interzol-orale-suspension-fr.jpg)



























