Embed Size (px)
Citation preview

Pathology Research and Practice 200 (2004) 41–46
ARTICLE IN PRESS
*Correspondin
E-mail addres
0344-0338/$ - see
doi:10.1016/j.prp
www.elsevier.de/prp
ANIMAL AND INVITRO MODELS IN HUMAN DISEASES
Vacuole formation in the endothelium of rat extremity vessels depends on
fixation techniques and vessel type
G!abor Raffaia,*, Erzs!ebet Feh!erb, Gy .orgy N!adasya, S!andor Pakuc, G!abor Pog!anyc,Ferenc T!ım!arc, B!ela Szendec, Emil Monosa
a Institute of Human Physiology and Clinical Experimental Research, Semmelweis University Budapest, .Ull *oi !ut 78a, P.O. Box 448,
Budapest 1082, HungarybDepartment of Anatomy, Histology and Embryology, Semmelweis University Budapest, Budapest 1082, Hungaryc1st Department of Pathology and Experimental Cancer Research and Molecular Pathology Research Group, Joint Research Orga-
nization of Hungarian Academy of Sciences and Semmelweis University, Semmelweis University Budapest, Budapest 1082, Hungary
Received 11 April 2003; accepted 8 December 2003
Abstract
Applying immersion fixation for electron microscopy, huge clear endothelial membrane-bound vacuoles of 0.1–3 mmdiameter were noted in the extremity veins of Sprague–Dawley rats. Histological and electron microscopichistochemical methods were applied to determine whether they were the product of programmed cell death or anyother kind of cell damage. Image analyzer was used to measure the total area of the vacuoles in the endothelium cells.Neither lipid content nor acidic phosphatase activity could be identified in the vacuoles. In saphenous and brachialveins, the vacuoles occupied 20.672.21% and 1872.45% of the endothelium, respectively. Venous endothelium oftwo different strains of rat also contained the vacuoles. No such structures appeared in extremity arteries. Long-termtilting did not influence vacuolization. Using in vivo whole body fixation, only pinocytotic and dense microvesicles, butno vacuoles were noted. In conclusion, the clear vacuolar structures represent neither lipid inclusions nor secondarylysosomes. The method of tissue fixation is critical when venous endothelial vesicles are investigated. It is presumedthat the vacuoles originated from intra- or intercellular microstructures, and that in case of the collapsible veinsegments, their size is increased under the pathological—hypoxic and low-pressure—conditions of in vitro fixation.r 2004 Elsevier GmbH. All rights reserved.
Keywords: Extremity vessels; Venous endothelium; Vascular histology; Endothelial vacuole; Histology artefact
Introduction
Coordinated operation of physiological mechanismssupporting orthostatic tolerance is an essential conditionfor normal human activity. Orthostatic intolerance mayinvolve serious pathological components such as ortho-static edema, orthostatic hypo- or hypertension, etc.[3,24]. Understanding the adaptive responses of thehighly compliant extremity veins to orthostatic chal-
g author. Tel.: +1-210-6038; fax: +1-334-3162.
s: [email protected] (G. Raffai).
front matter r 2004 Elsevier GmbH. All rights reserved.
.2003.12.005
lenges is especially important given the erect position ofthe human body during many conditions of activity.Previously, we found evidence for significant functionaland structural remodeling processes in the saphenousvein (SV) of rats tilted 45� head-up in special tube cagesfor 2 weeks [14,15,17,25]. Recently, applying quantita-tive electron microscopy, immunohistochemistry, andthe tilt model, we found that a 2-week exposure tothe experimental orthostatic body position causes astriking increase in sympathetic innervation densityof the adventitia of SV [16]. During these studies,when immersion fixation was applied, we accidentally

ARTICLE IN PRESSG. Raffai et al. / Pathology Research and Practice 200 (2004) 41–4642
observed clear vacuole-like vesicles (0.1–3 mm in dia-meter) of unidentified origin and content on the electronmicrographs of the SV endothelium cells [22].The literature provides relevant information about
such kind of endothelial vacuolization in post-ischemicmyocardial reperfused arteries of swine, dog, and rat[7,8,10,20] in microvessels of hypoxic heart of rat[26,27], and in rat coronary endothelial cells afteradministration of endotoxin [6]. Similar endothelialalterations were noted in hypoxic brain capillaries afterocclusion of middle cerebral artery of rat [1,4], in carotidarteries of rabbit [5], and in hypoxic cultured pig brainendothelial cells [12]. Pulmonary arterioles under acutehypoxic conditions, asphyxial pulmonary veins [9,23], aswell as hypoxic and reoxigenated cultured humanumbilical vein endothelial cells [11] also exhibited suchtype of endothelial cell injury.None of these studies provided quantitative data on
the size and frequency of vacuoles. Regarding vacuoleformation in extremity veins and arteries, no informa-tion is available in the literature. Therefore, the generalpurpose of this study was to characterize quantitativelythe vacuoles we accidentally observed and to investigatetheir possible function, as well as the circumstancesunder which they appear. The specific aims of theexperiments included:
* Identification and quantification of the amount ofvacuolar structures in the endothelium of extremityveins of Sprague–Dawley (SD) rats when usingimmersion (in vitro) tissue fixation.
* Investigation of such endothelial vacuoles of extre-mity veins that develop in other rat strains (F-344 andMSD rats) during in vitro fixation.
* Examination of the endothelium of extremity arterieswhether it also contains vacuoles under the sameconditions as veins do.
* Investigation of long-term experimental orthostaticgravitational stress for its influence on the quantity ofthe vacuoles in the endothelium of extremity vessels.
* Identification of lipid content and ultrastructuralacidic phosphatase activity of endothelial vacuoles,or any endothelial cytoplasmatic signs, to determinewhether they are the product of an apoptotic process.
* Investigation of in vivo whole body fixation for itsinfluence on the formation of vacuolar structures invascular endothelium.
Methods
Experimental animals and chronic tilt
Adult male SD (n ¼ 39), as well as Fischer 344 (F-344, n ¼ 3) and AO/ola/MSD (MSD, n ¼ 2) ratsweighing 300–400 and 120–260 g, respectively, were
used. To study the effects of long-term experimentalorthostatic body position, SD rats were placed in tube-like, tiltable transparent cages set in horizontal (HOR,n ¼ 9), or in 45
�
head-up tilt (HUT, n ¼ 15) position for2 weeks, as described previously [14,16]. SD rats kept intraditional cages served as parallel controls (NC,n ¼ 15). Body weight, food, and water intake weremeasured daily. Studies were carried out according tothe guidelines of the Hungarian law on animal protec-tion (243/1998). The protocols were approved by theSemmelweis University Committee on the Ethical Useof Experimental Animals (590/99 Rh).
Vessel preparation, lipid content and acidic
phosphatase activity
To identify the content of the endothelial vacuolarstructures, 8–12mm long segments of saphenous andbrachial arteries and veins (SA, SV, BA, BV) werecarefully dissected from anesthetized SD rats (pento-barbital sodium, 50mg/kg bw, ip).For light and transmission electron microscopy, tissue
samples were fixed by immersion in a mixture of 1%glutaraldehyde and 4% formalin diluted in 0.15Mphosphate buffer (pH 7.4), washed in 0.15M phosphatebuffer, post-fixed in 1% OsO4, dissolved in 0.15Mphosphate buffer, dehydrated, penetrated with 1:1mixture of EPON 812 and propylenoxide, then em-bedded into Poly Bed 812. Semithin sections (1 mm) werecut and stained with toluidine blue, and also withhematoxylin and eosin. These sections were examinedusing a light microscope (Zeiss Axioplan). Ultrathinsections were cut, stained with uranyl acetate and leadcitrate, and analyzed using Philips CM10 electronmicroscope. Lipid droplets appear in such ultrathinsections as electron-dense granules.A modified method of Novikoff et al. [18] was used
for identifying ultrastructural acidic phosphatase activ-ity, characteristic of lysosomal and non-lysosomalcytoplasmic vacuolar programmed cell death [2]. Inbrief, vascular tissue samples were fixed in vitro byimmersion in 2.5% glutaraldehyde-cacodylate for 2 h,then washed in sodium acetate–sacharose solution (pH5), incubated in sodium acetate–sacharose supplementedwith sodium glicerophosphate substrate at 37�C for45min, and embedded into EPON 812. Ultrathinsections were cut for electron microscopic analysis.Acidic phosphatase positivity appears as dark electron-dense areas in such ultrathin sections.In another part of the study, whole body fixation
was applied for transmission electron microscopy. Theanesthetized animals were perfused at 100 cm H2Operfusion pressure with physiological salt solution(37�C) via the aorta, followed by a fixative solutioncontaining 3% paraformaldehyde and 0.1% glutaralde-
ARTICLE IN PRESSG. Raffai et al. / Pathology Research and Practice 200 (2004) 41–46 43
hyde in 0.2M phosphate buffer (pH 7.4). Small pieces ofthe extremity vessels were then dissected and immersedinto the same fixative for 3–5 h. All the other steps of theprocedure were identical with that described above.
Morphometric analysis
To determine the size of large vesicular structures,semithin sections were made and stained with toluidineblue. Light-microscopic pictures were taken and digita-lized with an Olympus digital camera. The digitalizedimages were analyzed with a PC-based image analyzingsoftware IMAN (beta) 2.0 (MFA, Budapest, Hungary).Analysis was carried out by two investigators as adouble blind trial. Specifically, the luminal and ablum-inal contours of the endothelial cell layer were outlined.Within this area, total size of the vacuoles was measuredand expressed as percentage of the whole endothelialcross-sectional area.
Statistical analysis
Results are given as means7SEM. One-way analysisof variance and pairwise multiple comparison procedure(Student–Newman–Keul method) were used for the finalstatistical evaluation of the experimental data. Po0:05was considered statistically significant.
Results
Endothelium cells of both SV and BV obtained fromnormal control SD rats contained a huge amount oflarge clear vacuoles in 0.1–3 mm+ range, when in vitroimmersion fixation of the tissue specimens was applied
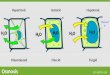
Fig. 1. Representative light microscopic picture of (a) an SV and (
(immersion) fixation technique. Arrows point to the large vacuol
scale=100 mm.
(Figs. 1a and 2a). These vacuoles were surroundedindividually by a membrane.The vacuoles in NC SD rats occupied 20.672.21% of
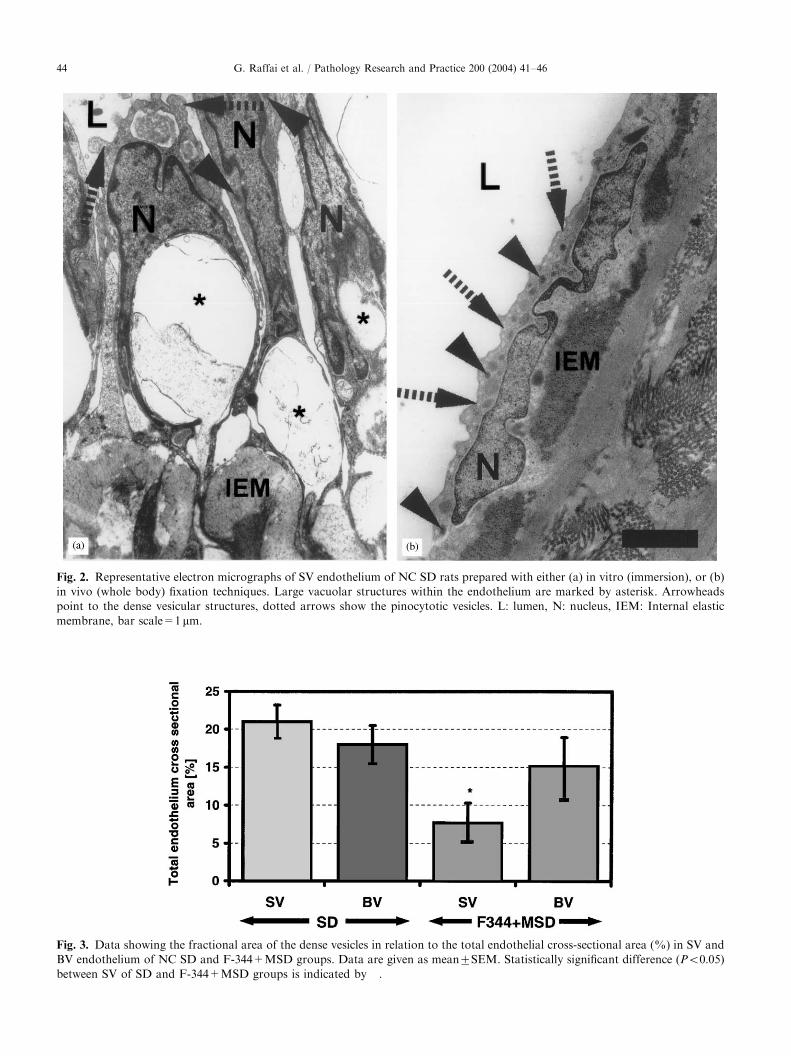
the total surface area of SV endothelium and 1872.45%of that of BV endothelium (Fig. 3). The SV endotheliumand BV endothelium of the other two rat strains, F-344and MSD, also contained such vacuoles in a stillremarkable quantity (7.7472.55% and 15.1373.88%,respectively) (Fig. 3).No such endothelial vacuoles appeared in the micro-
graphs of extremity arteries (SA and BA) of SD, F-344,or MSD rats (Fig. 1b).Long-term experimental orthostatic body position did
not influence significantly the production of such clearendothelial vacuoles in the SV and BV of SD rats (SV:HOR=17.1373.21%, HUT=17.5271.92%; BV:HOR=20.6374.37%, HUT=18.3872,15%).Light and electron microscopic analysis of the venous
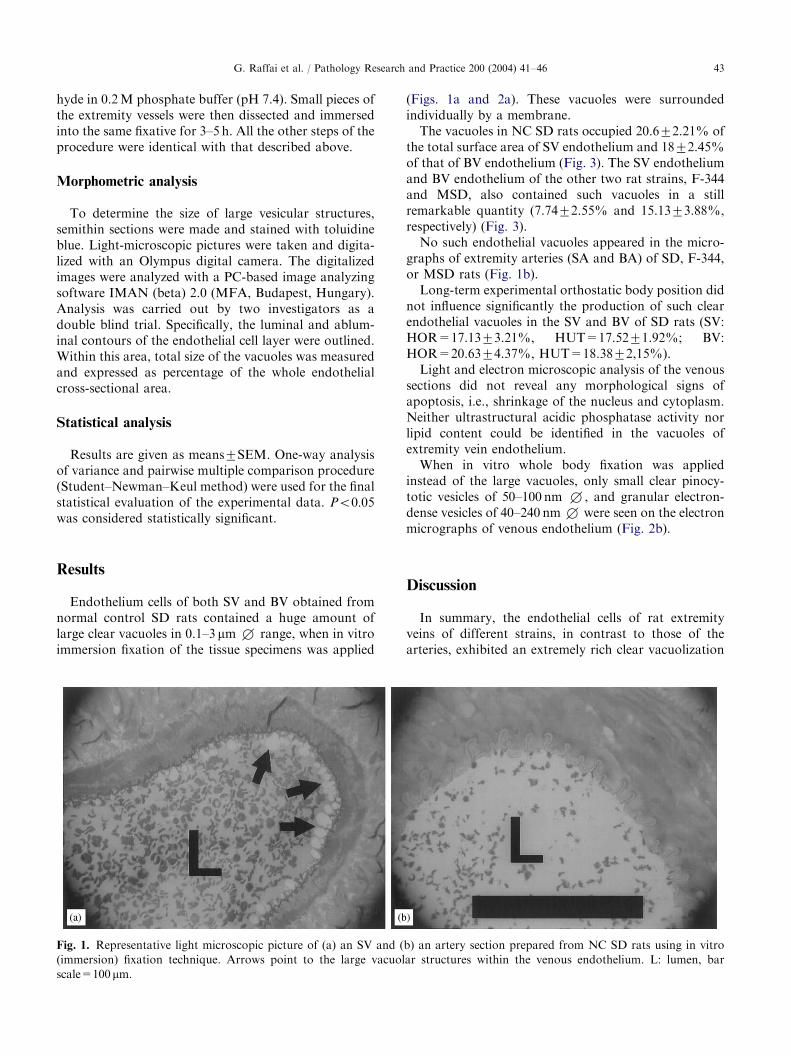
sections did not reveal any morphological signs ofapoptosis, i.e., shrinkage of the nucleus and cytoplasm.Neither ultrastructural acidic phosphatase activity norlipid content could be identified in the vacuoles ofextremity vein endothelium.When in vitro whole body fixation was applied
instead of the large vacuoles, only small clear pinocy-totic vesicles of 50–100 nm +, and granular electron-dense vesicles of 40–240 nm+ were seen on the electronmicrographs of venous endothelium (Fig. 2b).
Discussion
In summary, the endothelial cells of rat extremityveins of different strains, in contrast to those of thearteries, exhibited an extremely rich clear vacuolization
b) an artery section prepared from NC SD rats using in vitro
ar structures within the venous endothelium. L: lumen, bar
ARTICLE IN PRESS
Fig. 2. Representative electron micrographs of SV endothelium of NC SD rats prepared with either (a) in vitro (immersion), or (b)
in vivo (whole body) fixation techniques. Large vacuolar structures within the endothelium are marked by asterisk. Arrowheads
point to the dense vesicular structures, dotted arrows show the pinocytotic vesicles. L: lumen, N: nucleus, IEM: Internal elastic
membrane, bar scale=1 mm.
Fig. 3. Data showing the fractional area of the dense vesicles in relation to the total endothelial cross-sectional area (%) in SV and
BV endothelium of NC SD and F-344+MSD groups. Data are given as mean7SEM. Statistically significant difference (Po0:05)between SV of SD and F-344+MSD groups is indicated by �.
G. Raffai et al. / Pathology Research and Practice 200 (2004) 41–4644

ARTICLE IN PRESSG. Raffai et al. / Pathology Research and Practice 200 (2004) 41–46 45
when in vitro immersion fixation of the tissue wasapplied.The endothelium of the extremity vessels did not show
any shrinkage of nucleus and cytoplasm. The content ofthese vacuoles did not give any positive histochemicalreactions characteristic of acidic phosphatase activity orlipid content. Thus, the possibility of being an apoptoticproduct or any kind of endothelial cell damage can beexcluded. In addition to venous segments of SD rats, theendothelial vacuoles were also present in the extremityveins of adult F-344 and MSD rats. The content of thesevacuoles was probably water solution—accumulated inthe intra- or intercellular space. The total area of thevacuoles was not influenced by long-term orthostatic-type gravitational load.When in vivo whole body fixation was applied at
physiological pressure, the histological picture of theendothelium was normal, without the presence ofvacuoles. The appearance of the large vacuoles isobviously an artificial product of immersion fixation.However, the process should have been more complexthan a simple artifact formation, because the endothe-lium of corresponding arteries, under the same condi-tions, did not show any vacuolation. The vacuolizationresembles a pathological alteration of the transendothe-lial fluid transport with increased endothelial perme-ability, or an initial phase of endothelial injury.It remains to be answered why these endothelial
vacuoles appear only in veins. We presume that thefixative solution in vitro can reach the endothelium cellsof the collapsed cylindrical vein segments more slowlythan that of the arteries that do not collapse. Thus,despite abnormal hypoxic and low-pressure conditions,the vein endothelium cells probably survive longerin vitro than the endothelium of arterial segments. Wepresume that the vacuoles developed from a micro-vesicular transport system, probably from the miniatureclear pinocytotic vesicles, or they were produced by theswelling of some intracellular microstructures. It is alsopossible that elimination of intercellular contacts canresult in the vacuoles. The size of these—originallyminiature—structures is increased and probably furtheraugmented under extreme physiological or pathologicalconditions in a way that makes them grow to largevacuoles.This hypothesis is also supported by the studies of
Michel and Neal [13,19], who concluded that transcel-lular openings may contribute to the increased perme-ability of the venular endothelium, e.g. under hightransmural pressure, and these openings (channels)develop from fusion of intracellular vesicles. Data inthe literature are consistent with these assumptions.Enhanced appearance of endothelial vacuoles was foundin myocardial capillaries of endurance trained rats [21],in coronary endothelium of isolated rat heart after60min of global ischemia [7], in ischemic pig coronary
arteries after 5–60min of occlusion [20], and in ratcoronary vessels after administration of endotoxin [6].Similar cellular injuries were described in rat cardiaccapillaries subjected to hypoxia. In that experiment,luminal vesicles decreased in number but increased insize and density, while the endothelial cross-sectionalarea did not change [26]. Localized endothelial swelling,perivascular edema, and also a decrease in the number ofplasmalemmal vesicles and an increase in the number ofvacuoles characterized the rat myocardial microvesselsafter hypoxic insult [27]. Canine ischemic and reperfusedheart after 30min of ischemia exhibited endothelialvaculation, swelling of cytoplasmic organelles, anddetachment of endothelium [8]. In rat coronary en-dothelium, after 60min of ischemia and 60min ofreperfusion, the intercellular junctions disrupted [10].These structures were present not only in the afore-
mentioned cardiac endothelium, but also in braincapillaries of rats that showed ultrastructural alterations,including vacuole formation after 3 h of hypoxia [1], aswell as in cultured pig brain capillary endothelial cellsafter hypoxia in contrast to normoxic cells [12].Cerebrovascular injuries after occlusion of carotid arteryin rabbit or in middle cerebral artery of rat caused eitherappearance of intracytoplasmic vacuoles [5] or accumu-lation of vesicles [4]. In the pulmonary circulation,asphyxic pulmonary veins were probably associated withan enhancement of the endothelial permeability [9], or inacute hypoxic pulmonary hypertension, endothelial cellsshowed edematic vacuolation and were separated fromthe basal lamina [23]. Vacuole formation also occurred incultured hypoxic and reoxigenated human umbilical veinendothelial cells [11].In summary, in our study we found that the venous
endothelium is sensitive to the type of fixation (in vitrovs. in vivo) in different strains of rat. Immersion fixationresulted in vacuole formation exclusively in the venousendothelium. They were not formed when in vivofixation was applied. The degree of vacuolation wasnot influenced by long-term orthostasis. We provedneither apoptotic nor any other type of endothelial celldamage.The exact mechanism of vacuole formation needs to
be verified in further studies. It is also important to gointo the question of whether the miniature clear vesiclespresent in the endothelium cells of saphenous vesselsconstitute one of those adaptation mechanisms that maysupport orthostatic tolerance during long-term gravita-tional stress. In another study, we gave evidence that thedense secretory-type vesicles react to long-lastingorthostatic stress with diminution in total size [22].In conclusion, this study accentuates the critical
significance of methods chosen for tissue fixation,particularly when venous endothelial vesicular systemsare studied. We presume that vacuolar formation in theendothelium of extremity vein preparations after in vitro

ARTICLE IN PRESSG. Raffai et al. / Pathology Research and Practice 200 (2004) 41–4646
immersion fixation can serve as a model regardingdisturbed endothelial function, or can provide the basisfor a new experimental hypoxic model.
Acknowledgements
We thank !Eva Burka, Ferencn!e Hajd!u, and Ildik !oMur!anyi for their technical assistance. This work wassupported by Hungarian Grants OTKA T-030245/1999,OTKA T-042670/2003, ETT-240/2003, and TP-163/2003.
References
[1] A. Binovsky, T. Remis, J. Benuska, M. Masarova,
Ultrastructural changes in the brain capillaries after
hypoxia, Folia Morph. 38 (1990) 236–240.
[2] P.G. Clarke, Developmental cell death: morphological
diversity and multiple mechanisms, Anat. Embryol.
(Berl.) 181 (1990) 195–213.
[3] V.A. Convertino, G-factor as a tool in basic research:
mechanisms of orthostatic tolerance, J. Grav. Physiol. 6
(1999) 73–76.
[4] W.D. Dietrich, H. Nakayama, B.D. Watson, H. Kanemitsu,
Morphological consequences of early reperfusion following
thrombotic or mechanical occlusion of the rat middle
cerebral artery, Acta Neuropathol. 78 (1989) 605–614.
[5] T. Fujimoto, H. Suzuki, K. Tanoue, Y. Fukushima, H.
Yamazaki, Cerebrovascular injuries induced by activation
of platelets in vivo, Stroke 16 (1985) 245–250.
[6] M. Fukui, Y. Qiao, F. Guo, G. Asano, Cell damage and
liberation of nitric oxide synthase in rat heart induced by
endotoxin administration, Nippon Ika Daigaku Zasshi—
J. Nippon Med. School 62 (1995) 469–481.
[7] A.C. Hegstad, K. Ytrehus, S. Lindal, R. Myklebust, L.
Jorgensen, The initial phase of myocardial reperfusion is
not associated with aggravation of ischemia-induced
ultrastructural alterations in isolated rat hearts exposed
to prolonged global ischemia, Ultrastruct. Pathol. 23
(1999) 93–105.
[8] Y.D. Kim, J.S. Fomsgaard, K.F. Heim, P.W. Ramwell,
G. Thomas, E. Kagan, S.P. Moore, S.S. Coughlin, M.
Kuwahara, A. Analouei, et al., Brief ischemia-reperfusion
induces stunning of endothelium in canine coronary
artery, Circulation 85 (1992) 1473–1482.
[9] T. Kita, Y. Furuya, Histamine effects on pulmonary
blood vessels in strangulation, Z. Rechtsmed.—J. Legal
Med. 103 (1980) 85–91.
[10] P.J. Lin, C.H. Chang, Y.S. Lee, Y.Y. Chou, J.J. Chu, J.P.
Chang, M.J. Hsieh, Acute endothelial reperfusion injury
after coronary artery bypass grafting, Ann. Thorac. Surg.
58 (1994) 782–788.
[11] H. Mataki, T. Inagaki, M. Yokoyama, S. Maeda, ICAM-
1 expression and cellular injury in cultured endothelial
cells under hypoxia/reoxygenation, Kobe J. Med. Sci. 40
(1994) 49–63.
[12] K. Mertsch, T. Grune, W.G. Siems, A. Ladhoff, N.
Saupe, I.E. Blasig, Hypoxia and reoxygenation of brain
endothelial cells in vitro: a comparison of biochemical
and morphological response, Cell. Mol. Biol. 41 (1995)
243–253.
[13] C.C. Michel, C.R. Neal, Openings through endothelial
cells associated with increased microvascular permeabil-
ity, Microcirculation 6 (1999) 45–54.
[14] E. Monos, S.J. Contney, A.W. Cowley Jr., W.J. Stekiel,
Effect of long-term tilt on mechanical and electrical
properties of rat saphenous vein, Am. J. Physiol. 256
(1989) H1185–H1191.
[15] E. Monos, K. Kauser, S.J. Contney, A.W. Cowley Jr.,
W.J Stekiel, Biomechanical and electrical responses of
normal and hypertensive veins to short-term pressure
increases, in: G. Bruschi, A. Borghetti (Eds.), Cellular
Aspects of Hypertension, Springer, Heidelberg, 1991,
pp. 51–57.
[16] E. Monos, .M. L !or!ant, .E. Feh!er, Influence of long-term
experimental orthostatic body position on innervation
density in extremity vessels, Am. J. Physiol. 281 (2001)
H1606–H1612.
[17] E. Monos, V. B!erczi, G.L. N!adasy, Local control of veins:
biomechanical, metabolic, and humoral aspects, Physiol.
Rev. 75 (1995) 611–666.
[18] P.M. Novikoff, A.B. Novikoff, N. Quintana, J.J. Hauw,
Golgi apparatus, GERL, and lysosomes of neurons in rat
dorsal root ganglia, studied by thick section and thin
section cytochemistry, J. Cell Biol. 50 (1971) 859–886.
[19] K. Ogawa, M. Imai, T. Ogawa, Y. Tsukamoto, F. Sasaki,
Caveolar and intercellular channels provide major trans-
port pathways of macromolecules across vascular en-
dothelial cells, Anat. Rec. 264 (2001) 32–42.
[20] W.H. Park, M. Ashraf, R.W. Millard, J. Erickson,
Morphological changes in the coronary circulation
following experimental myocardial ischemia in swine,
Artery 12 (1985) 286–300.
[21] X. Qu, Morphological study of myocardial capillaries in
endurance trained rats, Br. J. Sports Med. 24 (1990)
113–116.
[22] G. Raffai, E. Feh!er, G. N!adasy, S. Paku, F. T!ım!ar, B.
Szende, E. Monos, Does gravitational stress affect the
dense vesicular and clear vacuolar endothelial structures
in the extremity veins of the rat? Acta Phys. Hung.
(Abstract) 89 (2002) 44.
[23] S.S. Sobin, P.C. Chen, Ultrastructural changes in the
pulmonary arterioles in acute hypoxic pulmonary hyper-
tension in the rat, High Altitude Med. Biol. 1 (2000)
311–322.
[24] D.H.P. Streeten, Orthostatic Disorders of the Circulation,
Plenum Medical Book Company, New York, 1987.
[25] M. Szentiv!anyi, V. B!erczi, T. H .uttl, R.S. Reneman, E.
Monos, Venous myogenic tone and its regulation through
K+ channels depend on chronic intravascular pressure,
Circ. Res. 81 (1997) 988–995.
[26] B.J. Ward, J.L Donnelly, The endothelial vesicular system
in cardiac capillaries subjected to hypoxia, Microvasc.
Res. 48 (1994) 143–150.
[27] K. Welt, L. Schaffranietz, H. Lukas, G. Wassilev, G.
Fitzl, N2O-induced hypoxia as a model of hypoxic stress
of rat myocardium. An ultrastructural-morphometric
study, Exp. Toxicol. Pathol. 50 (1998) 229–237.