Embed Size (px)
Citation preview

Sprinting as a clinical tool for the prevention of exercise-mediated
hypoglycaemia in Type 1 Diabetes Mellitus
Vanessa Anne Bussau
Bachelor of Science (Honours)
This Thesis is presented for the degree of Doctor of Philosophy at the University of Western Australia
School of Sport Science, Exercise and Health
2015

ii
Statement of Candidate Contribution
The work involved in designing and conducting the studies described in this thesis has
been carried out primarily by Vanessa Bussau (the candidate). The thesis outline and
experimental design of the studies was developed and planned by the candidate in
consultation with Professor Paul A. Fournier (the candidate’s primary supervisor) and
Professor Timothy W. Jones (co-supervisor). All participant recruitment and
management was carried out entirely by the candidate, along with the actual organisation,
implementation and performance of the experiments. In addition, the candidate was
responsible for all data analysis and original drafting of the thesis and peer-reviewed
publications. Professor Paul Fournier (and Dr Luis Ferreira) have provided feedback for
further drafts and completion of the thesis and manuscripts.
Signed:
Vanessa Bussau Paul Fournier
(Candidate) (Supervisor)

iii
Abstract
Despite the numerous physiological and psychological health benefits of a physically
active lifestyle for individuals with type 1 diabetes, the risk of hypoglycaemia increases
both during and after exercise. It is important to note, however, that not all types of
exercise result in an elevated risk of hypoglycaemia. For instance, prolonged high-
intensity aerobic exercise in these individuals result in an increase in glycaemia during
and after exercise. This raises the intriguing possibility that this type of exercise might be
beneficial if adopted to counter a fall in glycaemia in complication-free individuals with
type 1 diabetes, thus helping to prevent or delay hypoglycaemia if no carbohydrate is
readily available. Unfortunately, this type of exercise modality to prevent hypoglycaemia
is unlikely to be well tolerated by most individuals with type 1 diabetes due to the
impractical duration of this type of exercise. This raises the primary aim at the core of
this thesis to determine whether a much shorter bout of exercise lasting only 10 sec and
performed at maximal intensity could be adopted to prevent glycaemia from falling. For
this reason, the primary goal of this thesis was to determine whether a 10-sec maximal
sprint effort performed after (Chapter 3) or before (Chapter 4) moderate intensity exercise
provides a possible means other than carbohydrate intake to prevent glycaemia from
falling when exercise is performed under hyperinsulinaemic conditions by complication-
free individuals with type 1 diabetes. Also, given that for this type of study, it is common
practice to subject participants to a graded exercise test to set exercise intensity relative
to V O2peak, a secondary objective of this thesis was to determine whether the risk of
hypoglycaemia is increased early during recovery from this type of exercise protocol
(Chapter 2). Finally, since the counterregulatory response to sprinting has not been
examined in hyperinsulinaemic individuals with type 1 diabetes, thus making it difficult
to compare the findings of Chapters 3 and 4 with the literature, our last aim was to
examine the counterregulatory responses to sprinting in type 1 diabetic individuals under
hyperinsulinaemic conditions (Chapter 5). The first study of this thesis (Chapter 2) examines whether the risk of hypoglycaemia
increases in response to graded exercise testing in individuals with type 1 diabetes. Eight
non-diabetic male participants and seven complication-free type 1 diabetic male
individuals in good glycaemic control were recruited. On the morning of testing, the
diabetic participants followed their normal insulin regimen, and both groups ate their
usual breakfast. Then, participants were subjected to graded exercise testing

iv
approximately four hours later. We found that this type of exercise result in a rapid post-
exercise increase in blood glucose levels (> 2 mM), which remain elevated for the first
two hours of recovery. On clinical grounds, these findings suggest for the first time that
the early post-exercise risks of hypoglycaemia associated with graded exercise testing are
minimal when performed under near basal plasma insulin levels, with no carbohydrate
administration required soon before or after testing to prevent hypoglycaemia.
The primary goal of our next study (Chapter 3) was to determine whether a short 10-sec
maximal sprint effort is preferable to only resting as a means to counter a further fall in
glycaemia during recovery from moderate intensity exercise in hyperinsulinaemic
individuals with type 1 diabetes. To meet our objective, seven healthy complication-free
male participants with type 1 diabetes injected their normal insulin dose and ate their
usual breakfast. Then, when their postprandial glycaemia fell to ~11 mM they pedalled
at 40% peak 2OV for 20 min on a cycle ergometer followed immediately by either a
maximal 10-sec sprint or a rest. Our results show that, during exercise, blood glucose
levels fell rapidly. However, sprinting immediately after exercise opposes a further fall
in blood glucose levels for at least 120 min while glycaemia decreases significantly (p <
0.05) by ~ 3.5mM when no sprint was performed. We also found that sprinting is likely
to counter the exercise-mediated decrease in blood glucose levels through an increase in
catecholamine, lactate, and growth hormone levels. Interestingly, these glucoregulatory
benefits of sprinting are remarkable considering the sprint trial was performed when
insulin levels were elevated, a time when exercise is not usually recommended. On the
basis of these findings, one might tentatively recommend that in order to minimise the
risk of early hypoglycaemia post-moderate intensity exercise, it is preferable for
complication-free young individuals with type 1 diabetes to engage in a 10-sec maximal
sprint effort before resting than to only rest during recovery, particularly if a source of
dietary carbohydrate is not readily available.
Given the glycaemia stabilising effect of sprinting performed after moderate-intensity
exercise (Chapter 3), the study described in Chapter 4 examines whether performing a
short sprint effort immediately prior to moderate-intensity exercise may offer a novel way
of preventing glycaemia from falling both during and after moderate-intensity exercise.
To this end, seven complication-free type 1 diabetic males injected their normal morning
insulin dose and ate their usual breakfast. When post-meal glycaemia fell to ~11 mM,
they were asked to perform a 10 sec all-out sprint (sprint trial) or to rest (control trial)

v
immediately before cycling at 40% of peak rate of oxygen consumption for 20 min. We
found, against expectations, that sprinting for 10 sec immediately before moderate-
intensity exercise performed under hyperinsulinaemic conditions does not affect the rapid
decline in glycaemia during exercise. However, sprinting rather than resting before
moderate intensity exercise did prevent glycaemia from falling for at least the first 45 min
of recovery in individuals with type 1 diabetes. This suggests that including a short sprint
as part of the warm-up routine of individuals with type 1 diabetes before they engage in
sustained moderate-intensity exercise might provide another means of temporarily
stabilising glycaemia during early recovery.
Unfortunately, one difficulty with comparing the counterregulatory responses described
in Chapters 3 and 4 with the literature is the lack of information on the effect of short-
duration sprinting per se on the responses of counterregulatory hormones in
hyperinsulinaemic diabetic individuals. For this reason, the purpose of the study
described in Chapter 5 was to investigate the effect of a single 10-sec sprint on the levels
of the counterregulatory hormones in type 1 diabetic individuals under hyperinsulinaemic
conditions designed to approach those reported in Studies 3 and 4. In this study, we found
that performing a 10-sec maximal sprint resulted in patterns of change in plasma
catecholamines, growth hormone, cortisol and glucagon levels comparable to those
observed when a sprint is performed immediately after a bout of moderate intensity
exercise in individuals with type 1 diabetes (Study 2) and also comparable to those
observed in response to a sprint performed after an overnight fast (Fahey et al., 2012).
What remains to be established in future studies is the extent to which the changes in the
levels of these counterregulatory hormones contribute to the glucoregulatory benefits of
sprinting.
In conclusion, although a number of issues must be addressed before recommending the
adoption of short duration sprinting as a safe and reliable tool for the short-term
management of blood glucose levels in individuals with type 1 diabetes, this thesis shows
that sprinting has the potential to help individuals with type 1 diabetes to exercise more
safely and take advantage of the many physiological and psychological health benefits
of a physically active lifestyle.

vi
Acknowledgements
In writing my acknowledgements, I am overwhelmed with a sense of gratitude and
appreciation to all the people who have made this thesis possible. I have a huge smile as
I reminisce about the amazing people I have met and worked with along the way. Words
can not express how thankful I am for help and support of so many amazing people.
During the course of my PhD it seems like I have experienced almost every major life
event and the some of the absolute best and worst of times. Throughout the journey I
have had the support the best family anyone could wish for together with wonderful
colleagues who have become lifelong friends. I look forward to thanking you in person
but please realise how extremely grateful I am for your help, support and friendship
during the course of my PhD. In particular, I would like to sincerely thank the following
incredible people for their contribution towards my thesis:
My study participants - Thank you for your invaluable contribution. Recruitment was
such a challenge for this thesis so an extra huge thank you for your time and effort. It
was great to get to know you all and I wish you all the very best always. I hope you can
safely enjoy a more active life as a result of the research in this field.
Professor Paul Fournier (and Angeline) - for your passion, professionalism, infinite
knowledge and expertise, commitment, work ethic, endless enthusiasm and support (not
only with my PhD but with my career and life in general).
Dr Luis Ferreira (and Daniela) - for your research knowledge and expertise, balanced
advice, encouragement (together with your friendship and sharing my love of the Eagles,
sport and the important things in life).
Dr Kym Guelfi - for always being there throughout the journey with practical help,
advice, knowledge, friendship and huge morale support, you are an amazing friend and
researcher.
Alex D’Vauz (nee Baptista) - for your amazing friendship, encouragement, help, sense of
fun and for always bringing a smile to my face.

vii
Dr Ray Davey - from ‘Little Aths’ in Margaret River as kids to PhD buddies in the same
research team - thanks for your friendship, encouragement, advice and football banter.
Scott, Lian, Jen, Rob D, Hans (& Emma), Rob M, James (& Bree), Tim (& Sarah), Les,
Pete, Elisa, Tom, Jonas, Si, Nat, Stephen, Sani, Brad & the ‘Postgrad team’ - to each and
every one of you for your friendship, help, advice, camaraderie, fun times and great
memories (from corridor cricket & Friday tennis to our Annual Cocktail Party and so
many celebrations). Hans - it still seems surreal that you are not with us but your memory
will remind us to always cherish each day and we will never forget your love for your
family, friends and your passion for Exercise Physiology and Muscle Metabolism.
UWA Type 1 Diabetes Team – Paul, Luis, Kym, Alex, Tim, Avril, Katherine, Chee,
Harris & the PMH Team - Prof. Tim Jones, A/Prof. Liz Davis, Niru, Leanne, Michael,
Sarah, Vanessa. Thank you to a fantastic team of passionate researchers. It has been a
long journey but I have learnt so much from ‘being there at the start’ and seeing our
research team and field grow.
Our Nurses – Alisha, Nurse Bettye, Niru and Christy - thanks for your professionalism,
friendship and ability to make ‘testing days’ more enjoyable for all.
SSEH Team – Bec, Kerry, Prof. ‘Daws’, Karen, Danny, Brenda, Inga, Pat, Margaret,
Robin, Don & the Technical Staff for your help and support. Heads of Schools (Prof.
Brian Blanksby, Prof. Bruce Elliot, Prof. Tim Ackland) – for your support and exciting
career, research and teaching opportunities. To Prof. Tim Ackland, Sharon Gam and Jo
Francis for your huge help in this final phase.
Past Teachers, Lecturers and Mentors – from tiny Karridale Primary to Margaret River
High and UWA - to everyone who helped nurture my love of learning and science.
Inspiring Researchers, specialists, diabetes educators, endocrinologists and the incredible
people with type 1 diabetes I was privileged to meet. Hearing your thoughts and
discussing ideas at various conferences around the world was a huge privilege and
highlight of my PhD journey.

viii
To my awesome friends - a huge thank you for your friendship, inspiration and support
in so many ways. To Marshy & ‘The Marsh Family’, Michelle & Sean, Jas & Jamie,
Petrina & Rohan, Kym & Mark, Seaton, Amy & Dave, Jules & Matt, Carmen & Ross,
Ali & Mike, Alicia & Scott, Liv & Ian, Lise & Rod, Dan & Harriet, Fiona, Jaye, Krista
& Charles, Mia & Peter, Kylz & Dane, Erynne, Dale & Paul, Dyl, Joel, Waz, Col, Kat &
Guy, Sharne, Steph, K, Ames, Suz, Thalina, Alana & Ant, SuAnne, Bek, Mel, Paul,
Renae, Sarah, Anna-Mieke, Julia, Father Phillip & Our Guildford Grammar School &
Freeth House ‘Family’.
Mum, Dad, Daniel, Natalie - throughout life, school, ‘Undergrad’, Honours, PhD and
now life with my boys and business. Thanks for your unwavering love and support
always. Mum and Dad, thank you for always encouraging me to work hard and to pursue
my dreams. Thank you for all the opportunities you gave me throughout my education
and for encouraging me to follow my dreams and create a career in a field that I love. I
feel so incredibly grateful every time I see a patient and think how much I love helping
people and promoting exercise and health. Mum, Dad, Daniel (& Lucy) and Nat (&
Mike) – thank you for all your never-ending support along the journey – from school,
study, career and life. I love you all so unbelievably much and will never forget your
endless love and support always. Your contribution to this thesis is immense and will
always be cherished.
Dad - I miss you so much and wish you were here in person to firstly meet Marcus & my
‘boys’ and to see this PhD completed. I remember how proud you were when my 1st paper
was accepted for publication in your final days with us and hope you can enjoy a
celebration from ‘above’ with us.
To my extended family and friends, thank you to each and every one of you. In particular,
thank you to Ann, Noelle, Carrina for inspiring me in your own way. To Stu and Kay,
Rayma (& Geoff) and George for welcoming me into your family and for all your help
and support with our boys.
Finally, to Marcus, Owen, Max (and Lexie). Thanks for making me so unbelievably
happy and bringing so much joy and happiness into my life. I love you all more than
words can say. Marcus – thank you for your immense support. I will be eternally grateful
for the huge effort and sacrifices you and the boys have made so I could finish my PhD.

ix
Table of Contents
Statement of candidate contribution…………………………………………..
ii
Abstract………………………………………………………………………..
iii
Acknowledgements……………………………………………..…………….
vi
Table of contents………………………………………………………………
ix
List of figures.........……………………………………………………………
xiii
List of abbreviations…………………….........................………………….....
xiv
Publications arising from this thesis..………………………………………….
xvi
Chapter 1 - Introduction and Review of the Literature………… 1
1.1 Introduction and type 1 diabetes mellitus……….................................... 2
1.2 Treatment of type 1 diabetes mellitus and associated hypoglycaemia…... 3
1.3 Counterregulatory response to hypoglycaemia in non-diabetic individuals 5
1.3.1 Insulin………………………………………………………………. 5
1.3.2 Glucagon……………………………………………………...…….. 6
1.3.3 Catecholamines…………………………………………………...… 7
1.3.4 Growth hormone……………………………………………………. 8
1.3.5 Cortisol……………………………………………………………... 9
1.3.6 Other factors………………………………………………............ 10
1.4 Counterregulatory response to hypoglycaemia in type 1 diabetes……… 12
1.5 Glucoregulatory responses to moderate intensity exercise in non-diabetic
individuals...………………………………… ………………………...
15

x
1.6 Glucoregulatory responses to moderate intensity exercise in individuals
with type 1 diabetes…………………………………………..……………
20
1.7 Glucoregulatory responses to high-intensity aerobic exercise in non-
diabetic individuals…………………………………………………...……
23
1.8 Glucoregulatory responses to high-intensity aerobic exercise in
individuals with type 1diabetes……………………………………………
29
1.9 Glucoregulatory responses to intermittent high intensity exercise in non-
diabetic individuals. ………………………………………………..…
31
1.10 Glucoregulatory responses to intermittent high intensity exercise in
individuals with type 1 diabetes……………………………………….…
32
1.11 Glucoregulatory responses to a single short sprint in non-diabetic
individuals…………………………………………………….……………
34
1.12 Glucoregulatory response to a single short sprint in individuals with type
1 diabetes……………………………………………………..…..………..
40
1.13 Statement of the problem and aims………………………………...……... 41
1.14 Significance of the thesis………………………………...………...…...… 43
Chapter 2 – Glycaemic Response to Graded Exercise in
Individuals with Type 1 Diabetes………………………………...
44
2.1 Abstract……………………………………………………………….…. 45
2.2 Introduction…………………………………………………………….... 46
2.3 Research design and methods………………………………………..….. 48
2.3.1 Participants………………………………………………………… 48
2.3.2 Experimental trials and assays………………………………......… 48
2.3.3 Statistical analyses………………………………….……………… 50
2.4 Results……………………………………………………………………. 51
2.4.1 Blood metabolite response to graded exercise…………………...... 51
2.4.2 Hormonal response to graded exercise………………………..….... 53
2.5 Discussion…………………………………………………..……………. 55
2.6 Acknowledgements…………………………………………..…………... 59

xi
Chapter 3 – A 10-second Maximal Sprint Effort: A Novel
Approach to Counter an Exercise-Mediated Fall in Glycaemia in
Individuals with Type 1 Diabetes…………………………………
60 3.1 Abstract…………………………………………………………………… 61
3.2 Introduction………………………………………………………………. 62
3.3 Research design and methods…………………………………………….. 63
3.3.1 Participants………………………………………………………. 63
3.3.2 Experimental trials …………………………………………… 63
3.3.3 Hormones and metabolite assays……………………………… 65
3.3.4 Statistical analyses……………………………………………… 65
3.4 Results…………………………………………………………………... 66
3.4.1 Blood metabolite response……………………………………... 66
3.4.2 Hormonal response……………………………………………... 68
3.5 Discussion……………………………………………………………. 70
3.6 Acknowledgement………………………………………………………... 72
Chapter 4 – A 10-second Sprint Performed Prior to Moderate-
Intensity Exercise Prevents Early Post-Exercise Fall in
Glycaemia in Individuals with Type 1 Diabetes…………………
73
4.1 Abstract……………………………………………………………………. 74
4.2 Introduction……………………………………………………………... 75
4.3 Methods……………………………………………………………………. 77
4.3.1 Participants………………………………………………………. 77
4.3.2 Experimental Trials and Assays…………………………………. 77
4.3.3 Statistical Analyses……………………………………………… 78
4.4 Results ………………………………………………………. 79
4.4.1 Blood metabolite response 79
4.4.2 Hormonal response 79
4.5 Discussion………………………………………………………. 84
4.6 Acknowledgements………………………………………………… 86

xii
Chapter 5 – Counterregulatory Response to a 10-second Sprint
in Young Individuals with Type 1 Diabetes Mellitus ……………… 87
5.1 Abstract…………………………………………………………………. 88
5.2 Introduction……………………………………………………………... 89
5.3 Methods…………………………………………………………………. 91
5.3.1 Participants………………………………………………………. 91
5.3.2 Experimental Trials …………………………………………….. 91
5.3.3 Hormones and Metabolite Assays…………… 92
5.3.4 Statistical Analyses……………………………… 92
5.4 Results…………………………………………………………………... 93
5.4.1 Hormonal Response to a 10-sec Sprint………………………….. 93
5.4.2 Blood Metabolite Response to a 10-sec Sprint………………… 95
5.4.3 Work Load and Peak Power Associated with a 10-sec Sprint…... 95
5.5 Discussion………………………………………………………………. 97
5.6 Acknowledgements……………………………………………………... 100
Chapter 6 - General Discussion…………………………………. 101 6.1 General discussion……………………………………………………... 102
6.2 Clinical implications, limitations with our findings and direction for
future studies……………………………………………………………………
106
Chapter 7 – References…………………………………….......... 110

xiii
List of Figures
2.1 Effect of graded exercise on the levels of glucose (A), lactate (B), pH (C) and
free fatty acids (D)………………………………………………….................
52
2.2 Effect of graded exercise on the levels of epinephrine (A), norepinephrine (B),
growth hormone (C), cortisol (D), glucagon (E) and free insulin (F) ………....
54
3.1 Effect of a 10-sec sprint on blood glucose levels after moderate intensity
exercise………………………………………………………………...……...
67
3.2 Effect of a 10-sec sprint on the levels of lactate (A), free fatty acids (B),
norepinephrine (C), epinephrine (D), growth hormone (E), cortisol (F),
glucagon (G) and free insulin (H) after moderate intensity exercise ………….
69
4.1 Effect of a 10-sec sprint on blood glucose levels during and after moderate-
intensity exercise……………………………………………………...………
81
4.2 Effect of a 10-sec sprint on the levels of lactate (A), NEFA (B), norepinephrine
(C), epinephrine (D), growth hormone (E), cortisol (F), glucagon (G) and free
insulin (H) during and after moderate-intensity exercise……………………...
82
5.1 Effect of a single 10-sec sprint on the levels of norepinephrine (A), epinephrine
(B), growth hormone (C), cortisol (D), free insulin (E), and glucagon (F) ……
94
5.2 Effect of a single 10-sec sprint on blood glucose (A), free fatty acids (B), pH
(C) and blood lactate (D) levels………………………………………….........
96

xiv
List of Abbreviations
ADA American Diabetes Association
AIHW Australian Institute of Health and Welfare
ANOVA Analysis of variance
BMI Body mass index
CON Control
CGMS Continuous glucose monitoring system
DCCT Diabetes Control and Complications Trial
Direcnet Diabetes Research in Children Network
FFA Free fatty acids
GIR Glucose infusion rate
GH Growth hormone
GLUT Glucose transport protein
HbA1c Glycosylated haemoglobin
IHE Intermittent high-intensity exercise
IL-6 Interleukin 6
J Joule
JDRF Juvenile Diabetes Research Foundation
LOPEH Late onset post-exercise hypoglycaemia
Min Minute
mM Millimoles per litre
MOD Moderate-intensity exercise
n Number of participants
nmol/L Nanomoles per litre
N/A Not applicable
Ra Rate of appearance

xv
Rd Rate of disappearance
RIA Radioimmunoassay
SD Standard deviation
Sec Second
SEM Standard error of the mean
SPSS Statistical Package for the Social Sciences
T1DM Type 1 diabetes mellitus
µL Microlitres
O2 Rate of oxygen consumption
V O2max Maximal rate of oxygen consumption
V O2peak Peak rate of oxygen consumption

xvi
Publications Arising from this Thesis
Peer-Reviewed Publications
Bussau, V.A., Ferreira, L.D., Jones, T.W. & Fournier, P.A. (2007). A 10-s sprint
performed prior to moderate-intensity exercise prevents early post-exercise fall in
glycaemia in individuals with type 1 diabetes. Diabetologia 50 (9): 1815-1818.
Bussau, V.A., Ferreira, L.D., Jones, T.W. & Fournier, P.A. (2006). The 10-s Maximal
Sprint: A Novel Approach to Counter an Exercise-Mediated Fall in Glycemia in
Individuals with Type 1 Diabetes. Diabetes Care 29 (3): 601-606.
Conference Proceedings
Bussau, V.A., D’Vauz, A., Ferreira, L.D., Jones, T.W. & Fournier, P.A. (2010). Does
VO2 max testing increase risk of hypoglycaemia in individuals with type 1 diabetes?
Combined Biological Sciences Meeting, Perth, Australia.
Bussau, V.A., D’Vauz, A., Ferreira, L.D., Jones, T.W. & Fournier, P.A. (2009).
Glycaemic response to graded exercise in individuals with type 1 diabetes. Australian
Diabetes Society National Conference, Adelaide, Australia.
Bussau, V.A., D’Vauz, A., Ferreira, L.D., Jones, T.W. & Fournier, P.A. (2009).
Glycaemic response to graded exercise in individuals with type 1 diabetes. Australian
Diabetes Educators Association State Conference, Mandurah, Australia.
Fournier, P.A., Bussau, V.A., Fahey, A., Ferreira, L.D., Jones, T.W. (2008). Sprinting
as a novel approach to preventing exercise-mediated hypoglycemia. Diabetes,
Exercise & Sports Association International Conference, Toronto, Canada.
Bussau, V.A., Ferreira, L.D., Jones, T.W. & Fournier, P.A. (2006). A 10-second
sprint performed prior to moderate intensity exercise decreases the risk of post-
exercise hypoglycaemia in individuals with type 1 diabetes. International Diabetes
Federation Congress, Cape Town, South Africa.

xvii
Bussau, V.A., Ferreira, L.D., Youngs, L.M., Jones, T.W. & Fournier, P.A. (2005). A
10-second sprint performed prior to moderate intensity exercise decreases the risk of
post-exercise hypoglycaemia in individuals with type 1 diabetes. Australian Diabetes
Society Conference, Perth.
Bussau, V.A., Ferreira, L.D., Youngs, L.M., Jones, T.W. & Fournier, P.A. (2004). A
10-second sprint acutely prevents an exercise-mediated decrease in glycaemia in
individuals with type 1 diabetes mellitus. European College of Sports Science
Conference, Clermont- Ferrand, France.
Bussau, V.A., Ferreira, L.D., Youngs, L.M., Jones, T.W. & Fournier, P.A. (2004). A
10-second sprint acutely prevents an exercise-mediated decrease in glycemia in
individuals with type 1 diabetes. American Diabetes Association Conference,
Orlando, USA.

Chapter 1
Introduction and Review of the Literature

2
1.1 Introduction and Type 1 Diabetes Mellitus
Type 1 diabetes mellitus, formerly known as insulin-dependent or juvenile-onset diabetes
mellitus, is an endocrine disease that occurs commonly in childhood and adolescence, but
can be recognised and become symptomatic at any age (Dinneen and Rizza, 2001). Type
1 diabetes affects millions of people worldwide including approximately 140 000
Australians (IDF, 2009). The International Diabetes Federation found Australia to be one
of the top ten countries with the highest incidence of type 1 diabetes in children (Soltesz
et al., 2009), with over 8,000 new cases between 2000 and 2008, an average of two new
cases every day (AIHW, 2010). In individuals over 15 years, 9000 new cases of type 1
diabetes were diagnosed in Australia between 2000-2006 or three new cases per day
(AIHW, 2008).
Type 1 diabetes is characterised by the absence of insulin secretion due to the autoimmune
destruction of the beta cells of the Islets of Langerhans of the pancreas (Rizza et al., 2001).
The small proportion of individuals that appear not to have an autoimmune basis for their
beta cell destruction have a sub-type of diabetes referred to as idiopathic type 1 diabetes
(Dinneen and Rizza, 2001). As a consequence of the destruction of the beta cells, the
body loses its capacity to produce insulin. Given that insulin promotes the storage of
carbohydrates and fat, inhibits ketone body production, and stimulates a decrease in blood
glucose levels by both inhibiting hepatic glucose production and stimulating peripheral
glucose uptake, it is not surprising that the absence of insulin leads to an increase in blood
glucose (hyperglycaemia) and ketone body levels (Lernmark, 2001). This results in the
many symptoms typical of untreated type 1 diabetes including glucose and ketone body
loss in the urine (glucosuria and ketonuria), excessive urine production (polyurea),
extreme thirst and consumption of large quantities of water (polydipsia), excessive
consumption of food (polyphagia), and rapid weight loss (Eisenbarth et al., 2008). Other
symptoms may include nausea, vomiting, blurred vision, confusion, shortness of breath
and extreme fatigue (Eisenbarth et al., 2008). Severe hyperglycaemia and elevated ketone
body levels increase markedly the risk of septicaemia and ketoacidosis, respectively. This
explains why, without treatment, death generally occurs within 1-2 years of the onset of
type 1 diabetes (Campaigne and Lampman, 1994).
The discovery of insulin in 1921 and the initiation of treatments based on regular insulin
injections have extended considerably the life expectancy of individuals with type 1

3
diabetes. Currently, insulin is delivered by injection or constant subcutaneous infusion
via a pump (Sherr et al., 2009; Bergenstal et al., 2010). A number of types of insulin have
been developed, each with different absorption rates, onset, time to peak action and
duration of action. The dosage of insulin is often adjusted in response to self-monitored
blood glucose levels, nutritional intake and physical activity (Eisenbarth et al., 2008).
1.2 Treatment of Type 1 diabetes Mellitus and Associated Hypoglycaemia
The main challenge in the treatment of type 1 diabetes is to maintain blood glucose levels
close to a normal physiological range as this reduces the risk of developing a number of
severe diabetic complications including microvascular and macrovascular diseases,
nephropathy (kidney disease), retinopathy (damage to the retina of the eye) and
neuropathy (disease of the nervous system) (Brownlee et al., 2008; Eisenbarth et al.,
2008). This link between low risk of developing such complications and the importance
of maintaining blood glucose levels as close to normal as possible were demonstrated by
The Diabetes Control and Complications Trial in 1993 (Diabetes Control
and Complications Trial Research Group, 1993) and more recent studies (Nathan et al.,
2009). There is also evidence that, irrespective of glycaemic control, severe glycaemic
excursions also contribute to the development of these complications (Soupal et al.,
2014).
Unfortunately, the treatment of type 1 diabetes to maintain blood glucose levels within
the narrow physiological range found in non-diabetic individuals increases considerably
the risk of hypoglycaemic episodes (Cryer, 2010a). This is because of the absence of
feedback mechanisms between blood glucose levels and insulin secretion normally found
in non-diabetic individuals (Galassetti and Riddell, 2013). If too much insulin is
administered, the state of relative hyperinsulinaemia that ensues decreases hepatic blood
glucose production and increases glucose uptake, thus causing glucose utilisation to
exceed glucose production rate and, as a result, blood glucose levels decrease below
normal levels, a condition referred to as hypoglycaemia (Gerich, 2001).
In general, when blood glucose levels fall below 3 mM, warning symptoms alert an
individual to the presence of hypoglycaemia (McAulay et al., 2001). These include
“neuroglycopaenic” symptoms due to insufficient glucose reaching the brain (Cryer,
1999) and may include a loss in concentration, fatigue, weakness, confusion, difficulty in

4
thinking and speaking (Hepburn et al., 1991; Towler et al., 1993). Hypoglycaemia-
induced stimulation of the sympatho-adrenal system also triggers neurogenic (autonomic)
symptoms. These include “adrenergic” symptoms including tremor, heart palpitations
and anxiety as well as “cholinergic” symptoms such as sweating, hunger and paraesthesia
(Towler et al., 1993). Perception of these neurogenic symptoms is essential in one’s
awareness of hypoglycaemia and self-recognition that blood glucose levels are low
(Towler et al., 1993). Unfortunately, some individuals, diagnosed as “hypoglycaemia
unaware” experience minimal symptoms when their blood glucose levels fall to
dangerously low levels, with a late response to warning symptoms increasing their
possibility of experiencing severe hypoglycaemic episodes (Gold et al., 1994; McAuley
et al., 2001; Cryer et al., 2009; Candace et al., 2013).
The increased risk of hypoglycaemia associated with insulin-based therapy is a major
concern because severe hypoglycaemia can lead to central nervous system (CNS)
damage, and in extreme cases coma and even death (Ben-Ami et al., 1999; Cryer, 2007).
This is because a continuous supply of glucose is essential for normal cerebral function,
since the brain uses glucose as its main source of fuel under normal conditions (Gerich,
2000). Moreover, the brain stores little carbohydrate as glycogen (Herzog et al., 2010).
Given the importance of supporting the glucose requirements of the brain, it comes as no
surprise that most of the glucose synthesised in the body is utilised by this organ.
Unfortunately, hypoglycaemia is a common problem in the everyday life of most
individuals with type 1 diabetes. In fact, individuals with type 1 diabetes experience on
average two episodes of symptomatic hypoglycaemia per week, which equates to
thousands of hypoglycaemic episodes during their lifetime (Alsahli and Gerich, 2008). It
is important to remember that these rates are likely to underestimate the true number of
hypoglycaemic events due to many missed asymptomatic and symptomatic episodes
together with incorrect reporting (Cryer et al., 2009). In contrast, rates of severe episodes
of hypoglycaemia that require extended intervention to treat are more reliable, with
individuals with type 1 diabetes likely to suffer one such event per year often involving
coma or seizures (Cryer, 2008a, 2010b). Even more alarming is the fact that people with
type 1 diabetes die from hypoglycaemia (Laing et al., 1999). For these reasons, it is
understandable that hypoglycaemia is the primary and most feared complication of
insulin therapy (Amiel, 2009). It is not surprising, therefore, that such a therapy-induced
hypoglycaemia, or iatrogenic hypoglycaemia, is considered as a major limiting factor in

5
the glycaemic management of type 1 diabetes as it causes recurrent morbidity in most
individuals with type 1 diabetes (Cryer, 2005; 2010b). For this reason, any strategy aimed
at reducing the risk of hypoglycaemia is likely to be well received by individuals with
type 1 diabetes.
1.3 Counterregulatory Response to Hypoglycaemia in Non-Diabetic Individuals
Fortunately, the body possesses a range of mechanisms to counter hypoglycaemia. In
order to best appreciate how this is achieved in individuals with type 1 diabetes, it is
important in the first instance to examine how hypoglycaemia is countered in non-diabetic
individuals. Firstly, a number of highly specialised regions in the body including the beta
cells of the pancreas, ventromedial hypothalamus, and glucose sensing neurons in the
mouth, gut, portal/mesenteric vein and carotid body detect glucose changes or
hypoglycaemia (McCrimmon, 2009; Watts and Donovan., 2010). In healthy non-diabetic
individuals, a fall in blood glucose levels triggers complex but highly effective
physiological mechanisms known as counterregulatory responses that aim to reverse
falling blood glucose levels to restore euglycaemia (Cryer, 1993, 2008b). The magnitude
of such counterregulatory responses is determined in part by the depth and duration of
hypoglycaemia as well as by other factors including age, gender, rate of decline in blood
glucose before the onset of hypoglycaemia, antecedent exercise (Galassetti et al., 2001a),
and antecedent hypoglycaemia (Davis et al., 1991, 2000b).
1.3.1 Insulin
The first line of defence against hypoglycaemia is suppression of insulin secretion (Bolli,
1999) when blood glucose falls to a level of approximately 4.5 mM (Cryer et al., 2003).
The resultant decrease in circulating insulin levels stimulates an increase in hepatic
glucose production (Cherrington et al., 1998) and a decrease in glucose utilisation rate
(Cryer, 2001), which as a result counter declining glycaemia. However, if blood glucose
levels continue to fall to between 3.6 to 3.9 mM, the body responds by increasing the
secretion of counterregulatory hormones (Cryer et al., 2003). These hormones act to
restore glucose levels within a normal, safe physiological range by increasing glucose
production and inhibiting peripheral glucose uptake. It is important to note that the
contribution of each counterregulatory hormone is different, with some hormones being
more important than others as discussed below (Rizza et al., 1979b; Schwartz et al., 1987;
Mitrakou et al., 1991; Fanelli et al., 1994).

6
1.3.2 Glucagon
One of the key counterregulatory hormones is glucagon. Glucagon is secreted by the
alpha cells of the Islets of Langerhans of the pancreas (Quesada et al., 2008) when blood
glucose levels are low, with this hormone having an immediate effect on glucose kinetics
(Alsahli and Gerich, 2008). The secretion of glucagon by the alpha cells of the pancreas
is mainly influenced by blood glucose levels, with an increased secretion rate when blood
glucose levels are low (Jiang and Zhang, 2003; Porcellati et al., 2003). Other factors that
stimulate glucagon secretion are catecholamines (Gerich et al., 1973a), amino acids
(Rocha et al., 1972; Schmid et al., 1989) and short-term exposure to fatty acids (Hong et
al., 2005). In contrast, high levels of insulin and somatostatin inhibit glucagon secretion
(Ito et al., 1995; Hauge-Evans et al., 2009). Acting independently of these factors,
autonomic neural activation of the islet is thought to influence glucagon response to
hypoglycaemia (Havel et al., 1993).
Glucagon stimulates an increase in glucose levels firstly by activating hepatic
glycogenolysis and gluconeogenesis (Lecavalier et al., 1989; Roden et al., 1996;
Camacho et al., 2005). Even small changes in glucagon levels can increase glucose
production (Myers et al., 1991). Glucagon may also prevent blood glucose levels from
falling by inhibiting glycogen synthesis in the liver (Jiang and Zhang, 2003). Glucagon
is considered the most important counterregulatory hormone as demonstrated by studies
where recovery from hypoglycaemia has been shown to be impaired by a deficiency in
glucagon (Rizza et al., 1979b; Boyle et al., 1989). In particular, the importance of
glucagon is best shown by the observation that when glucagon response to hypoglycaemia
is prevented, this results in a blunted compensatory increase in endogenous glucose
production despite an increase in epinephrine secretion (De Feo et al., 1991b). In contrast,
glucagon does not affect renal glucose production or utilisation (Stumvoll et al., 1998a).
It is important to note that it is the level of portal glucagon relative to that of insulin as
expressed by the portal venous glucagon to insulin ratio that plays a central role in the
control of glycaemia (Quesada et al., 2008) as these hormones acutely increase or
decrease glucose production and utilisation to maintain blood glucose levels within a
narrow physiological range (Cryer, 2008b). A number of studies have highlighted the
importance of the portal venous glucagon-to insulin ratio in the regulation of hepatic
glucose production (Ferrannini et al., 1982; Lins et al., 1983; Steiner et al., 1990). When

7
this ratio is high, blood glucose levels increase as a result of decreased glucose utilisation
and increased glucose production (Boyle et al., 1989). The opposite occurs when this
ratio is low (Boyle et al., 1989).
1.3.3 Catecholamines
Catecholamines, including epinephrine and norepinephrine, are also thought to play a
major role in counterregulation in non-diabetic individuals. Epinephrine is released by
the chromaffin cells of the adrenal medulla, while norepinephrine is released by both the
adrenal medulla and the sympathetic nerve endings (Deschenes et al., 1991; Zouhal et al.,
2008). Hypoglycaemia, in its early stage, induces an elevation in sympathetic activity in
non-diabetic individuals, which results in an increase in catecholamine levels (Sotsky et
al., 1989; Bolli, 1999; Davis et al., 2000b). Although this increase in epinephrine and
norepinephrine levels is well established, there has been a long standing controversy
regarding the relative importance of catecholamines versus glucagon in counterregulation
(Cryer, 1981; Bolli, 1999). An important role for catecholamines in the stimulation of
glucose production during prolonged hypoglycaemia is highlighted by the observation
that pharmacological blockage of catecholamine action results in severe hypoglycaemia
despite increases in other counterregulatory hormones including glucagon (De Feo et al.,
1991a). In contrast, others have concluded that when glucagon response to
hypoglycaemia is normal, catecholamines play only a minor role (Rizza et al., 1979b;
Cryer et al., 2003). However, when glucagon response is impaired or absent,
catecholamines, in particular epinephrine, play a critical role in counterregulation (Rizza
et al., 1979b; Boyle et al., 1989; Cryer et al., 2003).
Catecholamines increase glycaemia by acting on multiple organs (Meyer et al., 1999;
Alsahli and Gerich, 2008). An increase in epinephrine and norepinephrine levels results
in an elevation in blood glucose levels due to an increase in hepatic (Sacca et al., 1980)
and renal (Stumvoll et al., 1998b) glucose production together with a fall in insulin-
mediated stimulation of glucose utilisation (Rizza et al., 1979a; Nonogaki, 2000; Coker
& Kjaer, 2005). Catecholamines increase the rate of hepatic glucose production by
stimulating an increase in both glycogenolysis and gluconeogenesis (Barth et al., 2007).
This increase in hepatic glucose production is initially due to the stimulation of
glycogenolysis, while hepatic gluconeogenesis later becomes the predominant
contributor to sustained hepatic glucose production (Sacca et al., 1983). Catecholamines
also decrease the rate of glucose utilisation with both epinephrine (Rizza et al., 1979a;

8
Deibert and Defronzo, 1980; Lager et al., 1986) and norepinephrine (Lembo et al., 1994)
inhibiting insulin-mediated stimulation of glucose uptake by skeletal muscles. As
discussed later, however, there are conditions where catecholamines and other
adrenoceptor agonists have been reported to increase muscle glucose uptake (Abe et al.,
1993; Nonogaki, 2000; Ngala et al., 2013).
Catecholamines also indirectly increase blood glucose levels. High levels of
catecholamines suppress endogenous insulin release (Clutter et al., 1980; Sherwin et al.,
1980; Sacca et al., 1983) and stimulate the supply of gluconeogenic substrates (Sacca et
al., 1983; Stumvoll et al., 1998b) including lactate from resting muscles (Sacca et al.,
1983). Catecholamines also stimulate glucagon (Gerich et al., 1973a) and growth
hormone release (Blackard and Heidingsfelder, 1968). Finally, catecholamines stimulate
lipolysis (Clutter et al., 1980) and therefore increase the levels of plasma free fatty acids
(Sherwin et al., 1980; Sacca et al., 1983), decreasing carbohydrate utilisation and
increasing hepatic glucose production.
1.3.4 Growth hormone
Hypoglycaemia also results in the activation and release of GH, a group of
counterregulatory hormones produced by the pituitary gland and which also plays some
role in the defence against hypoglycaemia. Indeed, growth hormone is a heterogeneous
class of protein hormones consisting of a series of related isoforms (Baumann, 2009).
When the hypothalamus senses hypoglycaemia, growth hormone releasing hormone and
somatostatin are thought to be released together with other growth hormone-releasing
factors resulting in the pulsatile secretion of growth hormone from the anterior pituitary
gland (Reiter and Rosenfeld, 2008).
Growth hormone is not considered important in opposing acute hypoglycaemia, but it
may play a role during prolonged hypoglycaemia. This is based on the observation that
the pharmacological blockage of growth hormone release or action during prolonged
hypoglycaemia impairs the increase in glucose production, thus resulting in more severe
hypoglycaemia (De Feo et al., 1989b; Boyle and Cryer, 1991). This effect of growth
hormone on glucose turnover takes several hours to take place (De Feo et al., 1989b;
Boyle and Cryer, 1991). Growth hormone has also an indirect effect on counterregulation
by stimulating lipolysis. The resulting increase in glycerol and free fatty acids levels
provide gluconeogenic substrates and fuels, respectively, with the capacity to spare

9
glucose by inhibiting glucose utilisation (De Feo et al., 1989b). Prolonged growth
hormone exposure has been shown to have insulin-antagonistic effects (Fowelin et al.,
1991) whereas in contrast short duration growth hormone infusion does not invoke insulin
resistance (Djurhuus et al., 2004).
It is noteworthy that some studies have shown that growth hormone has an acute effect
on glucose metabolism. Indeed, local growth hormone exposure results in a rapid
decrease in forearm glucose uptake (Zierler and Rabinowitz, 1963; Rabinowitz et al.,
1965; Gibney et al., 2007). Furthermore, the administration of a physiological growth
hormone pulse in non-diabetic individuals has been reported to result in a rapid fall in
muscle glucose uptake (Moller et al., 1990, 1992b, 2003) and a 1-2 hour delayed increase
in lipolysis, circulating free fatty acid levels, and fat oxidation rates (Moller et al., 1990,
1992b; Gravholt et al., 1999; Møller et al., 2003; Djurhuus et al., 2004), which altogether
could contribute further to lowering glucose utilisation rates (Møller et al., 1992b) and
stimulating hepatic glucose production.
1.3.5 Cortisol
Another counterregulatory hormone implicated in prevention of hypoglycaemia is
cortisol. When the hypothalamus senses hypoglycaemia, this results in the release of
corticotrophin-releasing factor in the pituitary portal vessels which in turn stimulates the
secretion of adrenocorticotrophic (ACTH) hormone by the anterior pituitary gland.
ACTH then activates the secretion of cortisol by the cortex of the adrenal glands
(Macdonald and King, 2007), which acts indirectly in the acute counterregulatory
response by further stimulating catecholamine secretion by an intra-adrenal effect.
Like GH, cortisol is not considered important in the acute response to hypoglycaemia
(Heller, 2011). Despite this, cortisol plays an important role in long term
counterregulation when hypoglycaemia is prolonged (De Feo et al., 1989a; Boyle and
Cryer, 1991). Under these conditions, cortisol increases glucose production and inhibit
glucose utilisation after approximately 3 hours (De Feo et al., 1989a; Boyle and Cryer,
1991). Prolonged infusion of cortisol increases glycaemia and reduces the required
glucose infusion rate to maintain euglycaemia by increasing glucose production and
decreasing glucose uptake by the peripheral tissues, partly by inducing hepatic and
peripheral insulin resistance (Rizza et al., 1982; Rooney et al., 1993).

10
During prolonged hypoglycaemia, cortisol has also an indirect effect on blood glucose
levels by stimulating systemic and regional lipolysis (Djurhuus et al., 2002, 2004) . If
growth hormone is present, the lipolytic effect of both hormones is additive (Djurhuus et
al., 2004). This increase in lipolysis, in turn, has independent insulin-resistant or glucose
sparing effects (De Feo et al., 1989b; Corral et al., 1998).
1.3.6 Other factors
In recent years, the potential role that the cytokine interleukin-6 (IL-6) plays in the
regulation of blood glucose levels and hepatic glucose production has received some
attention (Glund and Krook, 2008; Hoene and Weigert, 2008; Pedersen, 2009). IL-6 is a
pro-inflammatory cytokine that is involved in mediating many inflammatory processes,
brain function, fatigue and immune response (Glund and Krook, 2008). The relatively
recent discovery that skeletal muscle can produce and release cytokines such as IL-6 is at
the origin of the term, myokine. This class of proteins expressed and released by skeletal
muscle is associated with endocrine and/or paracrine effects (Pedersen, 2009). In fact,
IL-6 was the first discovered muscle contraction-induced “exercise factor” or myokine
(Pedersen, 2009).
Hypoglycaemia is associated with an increase in plasma IL-6 levels (Dotson et al., 2008).
IL-6 infusion in vivo increases glucose uptake potentially due to increased translocation
of GLUT4 from intracellular compartments to the plasma membrane (Carey et al., 2006).
Indeed, in healthy men subjected to a hyperinsulinaemic, euglycaemic clamp,
recombinant human IL-6 infusion increases the glucose infusion rate without affecting
the total suppression of endogenous glucose production (Carey et al., 2006), thus
suggesting an increase in glucose uptake. Although these effects of IL-6 would not be
expected to prevent hypoglycaemia, IL-6 might be indirectly beneficial given that
exogenous IL-6 administration has been shown to increase cortisol and glucagon levels
together with a rise in blood glucose levels, thus probably helping with opposing
hypoglycaemia (Dotson et al., 2008; Glund & Krook, 2008).
Other than IL-6, the levels of circulating lactate, glycerol and amino acids may also play
some role in counterregulation. These metabolites act as substrates for gluconeogenesis
to increase glucose production and modulate the secretion of hormones from the pancreas
(Roden and Bernroider, 2003). They may also act as fuels for peripheral tissues to spare
glucose or inhibit insulin-mediated glucose uptake. Indeed, both oral (Rossetti et al.,

11
2008) and intravenous (Porcellati et al., 2007) amino acids enhance the response of
glucagon to hypoglycaemia (Porcellati et al., 2007). Since lactate is a significant fuel
source, gluconeogenic precursor (Gerich, 1988), and may have a role in increasing insulin
resistance (Harmer et al., 2008), it has the potential to play a role in both increasing
glucose production and decreasing glucose utilisation. Some have proposed that
increased muscle lactate utilisation together with decreased muscle glucose uptake make
major contributions to glucose counterregulation in response to hypoglycaemia humans
(Meyer et al., 2005).
There is evidence that changes in circulating blood glucose concentrations have the
capacity to affect hepatic glucose production and thus contribute to the counterregulatory
response to hypoglycaemia. Such a non-hormonal autoregulatory mechanism accounts
for approximately 25% of the rise in net hepatic glucose production during
hypoglycaemia (Connolly et al., 1992). On the other hand, hyperglycaemia exerts a direct
inhibitory effect on endogenous glucose production via a glycogen phosphorylase-
mediated inhibition of glycogenolysis (Tonelli et al., 2005; Yki-Järvinen, 1993). In
addition, animal models have shown that net hepatic gluconeogenesis is reduced under
hyperglycaemic conditions when glycogen levels are depleted (Tonelli et al., 2005). The
mechanisms underlying the aforementioned autoregulation are only partly understood
(Moore et al., 1998; Tonelli et al., 2005).
1.4 Counterregulatory Response to Hypoglycaemia in Individuals with Type 1
Diabetes
In individuals with type 1 diabetes, many of the counterregulatory responses described
above are either absent or impaired, increasing the risk of severe and potentially life-
threatening episodes of hypoglycaemia (Galassetti and Riddell, 2013). Firstly, when
blood glucose levels begin to fall, the levels of insulin do not decrease, which is the initial
response to a decrease in glycaemia in non-diabetic individuals (Briscoe et al., 2007).
This is because the level of circulating insulin in insulin-treated individuals with type 1
diabetes is determined primarily by the rate of passive absorption from the site of insulin
injection together with the pharmokinetics of the particular type of insulin administered
(Cryer et al., 2003), with no feedback existing between blood glucose levels and insulin
release.

12
Early after the onset of type 1 diabetes, patients generally have normal insulin-
independent counter-regulatory responses to hypoglycaemia. However, this defence
mechanism against hypoglycaemia deteriorates thereafter (Richter and Galbo, 1986;
Gerich, 1988). Reduced glucagon response to a fall in glycaemia is the first and possibly
most important counterregulatory response to be impaired (Gerich et al., 1973b) despite
normal glucagon secretion in response to other stimuli such as exercise (Cryer et al.,
1989; Shilo et al., 1990). It is well established that this deficient glucagon response to
hypoglycaemia is strongly correlated to duration of diabetes (Bolli et al., 1983); however
the mechanism(s) involved is still unclear. Recent studies have provided evidence of beta
cell regulation of alpha cell glucagon secretion (Cooperberg and Cryer, 2009), with an
increase in insulin level signalling a decrease in glucagon secretion in response to
hypoglycaemia (Banarer et al., 2002; Cooperberg and Cryer, 2010). Therefore, current
evidence suggests defective glucagon response may be due to beta cell failure.
Furthermore, there may also be a central nervous system component to the impaired or
absent glucagon response since insulin's inhibitory effect on glucagon release is partly
mediated by the ventromedial hypothalamus under both normoglycaemic and
hypoglycaemic conditions (Paranjape et al., 2010).
As a result of their attenuated or lack of glucagon response to hypoglycaemia (Gerich et
al., 1973b; Bolli et al., 1983; Hirsch and Shamoon, 1987; Meyer et al., 1998), individuals
with type 1 diabetes become more dependent on catecholamines (Marker et al., 1991), in
particular epinephrine (Bolli et al., 1982; Cryer et al., 1989), together with other
counterregulatory hormones to overcome hypoglycaemia (De Feo et al., 1983).
Unfortunately, individuals with type 1 diabetes have an attenuated epinephrine (Bolli et
al., 1983; Amiel et al., 1988; Dagogo-Jack et al., 1993; Meyer et al., 1998) and
norepinephrine (Meyer et al., 1998) response to hypoglycaemia compared to non-diabetic
individuals. This is exacerbated by the fact that the rate of fall of glycaemia can affect
epinephrine response to hypoglycaemia, with a rapid rate of fall resulting in a smaller
epinephrine response compared with a slower rate of fall. In diabetic individuals with
autonomic neuropathy, catecholamine response to hypoglycaemia is further impaired
(Bolli et al., 1983; Bottini et al., 1997; Meyer et al., 1998). In summary, the three key
normal responses that play an important role in the defence against hypoglycaemia when
glycaemia is falling (decreased insulin levels, increased glucagon and increased
epinephrine levels) are often deficient in individuals with type 1 diabetes (Cryer, 2003).

13
This is highly problematic for those individuals with both glucagon and epinephrine
deficiency as they are at an increased risk of severe hypoglycaemia (White et al., 1983).
The aforementioned impaired counterregulatory responses to hypoglycaemia also
implicate the glycaemic threshold for the release of counterregulatory hormones, which
is often different in type 1 diabetic individuals compared to non-diabetic individuals
(Cryer et al., 2003). The glycaemic threshold for individuals with poorly controlled type
1 diabetes is higher (Amiel et al., 1988; Boyle et al., 1988) than in non-diabetic
individuals. In contrast, the plasma glucose level threshold is generally lower in tightly
controlled type 1 diabetes (Amiel et al., 1988). The degree of attenuation, absence or
lowering of the glycaemic threshold for these counterregulatory responses appears to be
associated with a longer duration of type 1 diabetes (Bolli et al., 1983). Strict glycaemic
control and intensive insulin therapy (Amiel et al., 1987; 1998) are associated with
impaired counterregulation, thus increasing the risk of severe hypoglycaemia (DCCT,
1997).
One key factor that can alter the counterregulatory response to hypoglycaemia in
individuals with type 1 diabetes is a recent previous episode of hypoglycaemia. Recent
antecedent hypoglycaemia reduces the glycaemic thresholds for the activation of
counterregulatory responses to subsequent hypoglycaemia (Amiel et al., 1988). This is
because recurrent hypoglycaemia interferes with glucose sensors and neural networks that
detect hypoglycaemia (McCrimmon, 2009). Antecedent hypoglycaemia also blunt the
magnitude of the response of counterregulatory hormones to subsequent hypoglycaemia
and increases the risk of future episodes of hypoglycaemia (Davis and Shamoon, 1991;
Heller and Cryer, 1991; Widom and Simonson, 1992; Dagogo-Jack et al., 1993; Hvidberg
et al., 1996; Davis et al., 1997, 2000b, 2000c; Shum et al., 2001; Cryer et al., 2003). Even
mild episodes of hypoglycaemia can attenuate this counterregulatory response; however
the depth of hypoglycaemia is an important determinant of the impaired
counterregulatory response to a subsequent hypoglycaemic episode with a dose-response
effect of antecedent hypoglycaemia (Davis et al., 1997). For instance, mild
hypoglycaemia of ~3.9 mM can reduce epinephrine, muscle sympathetic nervous system
and glucagon responses to hypoglycaemia by ~30% the next day (Davis et al., 1997).
Lower glycaemia of ~3.3 mM also reduces the response of these glucoregulatory
hormones together with blunting norepinephrine, growth hormone, endogenous glucose
production, pancreatic polypeptide and lipolytic responses to hypoglycaemia (Davis et

14
al., 1997). Interestingly, an even lower antecedent blood glucose level of ~ 2.9 mM
elicits a similar impaired counterregulation to ~3.3 mM (Davis et al., 1997). Together
with this dose-response effect of antecedent hypoglycaemia on subsequent
counterregulatory response to hypoglycaemia, the duration and frequency of
hypoglycaemic episodes can also influence this counterregulatory response. Indeed, there
seems to be a hierarchical effect of the duration of antecedent hypoglycaemia on the
reduction of the counterregulatory responses to subsequent hypoglycaemic episodes
(Davis et al., 2000b).
Recurring episodes of hypoglycaemia are also associated with marked attenuation of the
warning symptoms of hypoglycaemia (Bolli, 2003). This leads to a situation where
individuals with type 1 diabetes often suffer from a reduced ability or failure to recognise
hypoglycaemia, a condition referred to as hypoglycaemia unawareness (Cryer et a., 2003;
Alsahli and Gerich, 2008). As a result, these patients are unable to detect and therefore
correct hypoglycaemia (Bolli, 2003), increasing markedly their risk of severe
hypoglycaemia (Gold et al., 1994).
Antecedent exercise can also impair one’s capacity to counter a subsequent episode of
hypoglycaemia in individuals with type 1 diabetes as exercise impairs the
counterregulatory response to subsequent hypoglycaemia (Galassetti et al., 2001a;
Sandoval et al., 2004, 2006). Although exercise and hypoglycaemia can blunt
counterregulation to a similar level (Sandoval et al., 2006), the increase in insulin
sensitivity that occurs as a result of exercise can elevate the risk for hypoglycaemia during
subsequent hypoglycaemic episodes (Briscoe et al., 2007, Galassetti and Riddell, 2013).
Finally, one important factor that can aggravate the risk of hypoglycaemia in individuals
with type 1 diabetes is physical activity. Participation in regular physical activity
provides numerous health benefits for individuals with type 1 diabetes, including weight
control, improvement of muscle strength, lowering atherosclerosis risk factors, and
overall improvement in cardiovascular function (Chimen et al., 2012). Unfortunately,
exercise for these individuals increases the risk of hypoglycaemia during and for several
hours after exercise (MacDonald, 1987; Tsalikian et al., 2005; McMahon et al., 2007).
For this reason, many individuals with type 1 diabetes are often reluctant to be physically
active and thus miss out on the many physical and psychological benefits of an active
lifestyle (Ludvigsson et al., 1980). In order to fully appreciate how exercise increases the

15
risk of hypoglycaemia, we must examine how blood glucose levels are regulated during
exercise in healthy individuals. Here we will focus on sustained aerobic exercise of
moderate intensity, aerobic/anaerobic exercise of high intensity, intermittent high
intensity exercise, and maximal sprint effort, but not on other forms of exercise such as
resistance exercise.
1.5 Glucoregulatory Responses to Moderate Intensity Exercise in Non-Diabetic
Individuals
During moderate intensity exercise in healthy non-diabetic individuals, stable blood
glucose levels are maintained by feedback mechanisms that allow the increase in glucose
utilisation by exercising muscles to be precisely matched by an equal increase in glucose
production rate (Richter and Galbo, 1986; Marliss and Vranic, 2002). What is still a
source of some debate is the mechanisms that coordinate liver and muscle responses to
exercise and the role of insulin, glucagon, catecholamines together with other
counterregulatory hormones and metabolites in this precise matching of glucose kinetics.
At the start of moderate intensity exercise, it is well accepted that the contraction of
skeletal muscle induces a rapid increase in glucose uptake (Wahren and Ekberg, 2007).
At rest, only 15-20% of peripheral glucose utilisation is attributed to skeletal muscle, but
during moderate intensity exercise at 55-60% V O2max skeletal muscle accounts for up to
80-85% of whole body glucose disposal (Hargreaves and Spriet, 2006). This marked
increase in muscle glucose uptake during exercise (Camacho et al., 2005; Hargreaves and
Spriet, 2006) is explained by an exercise-mediated stimulation of the translocation of a
non-insulin responsive pool of the glucose transporter GLUT4 (Douen et al., 1989;
Coderre et al., 1995), together with increased blood flow, capillary recruitment, glucose
extraction and improved glucose delivery to skeletal muscle.
Although there is a significant increase in the rate of glucose removal from the blood
during moderate intensity exercise, non-diabetic individuals do not become
hypoglycaemic. Instead, their blood glucose levels remain stable as a result of a matched
increase in the rate of hepatic glucose production (Camacho et al., 2005). The activation
of hepatic glycogenolysis and gluconeogenesis increases hepatic glucose production, with
the relative contributions of each metabolic pathway changing with exercise duration

16
(Trimmer et al., 2002; Wahren and Ekberg, 2007) and intensity (MacRae et al., 1995;
Staehr et al., 2007) and dietary state (Staehr et al., 2007), with both prolonged fasting and
exercise being favourable to hepatic gluconeogenesis (Trimmer et al., 2002; Staehr et al.,
2007). At the start of exercise, hepatic glycogenolysis accounts for most of the increase
in glucose produced by the liver. As time progresses, the rate of glycogenolysis declines
with decreasing hepatic glycogen stores, and gluconeogenesis becomes increasingly more
important (Richter & Galbo, 1986; Camacho et al., 2005; Hargreaves & Spriet, 2006;
Wahren & Ekberg, 2007). If the duration of exercise is more than two hours, the
depletion of hepatic glycogen stores together with an inadequate compensatory increase
in gluconeogenesis can result in declining glycaemia and even hypoglycaemia (Trimmer
et al., 2002; Camacho et al., 2005).
The increase in hepatic glucose production during exercise results to some extent from
the combined fall in insulin level and rise in glucagon concentration (Marliss and Vranic,
2002; Camacho et al., 2005). A fall in insulin level is required for a full increase in hepatic
glycogenolysis (Wasserman et al., 1989b), whereas elevation in glucagon level is
necessary for both increased hepatic glycogenolysis and gluconeogenesis (Wasserman et
al., 1989a). Although such a fall in insulin and increase in glucagon levels are often
observed during moderate intensity exercise, some studies have reported that the levels
of these hormones, in particular glucagon levels, do not change or only change slightly
(Wasserman et al., 1993; Wasserman, 2009). It is important to note, however, that the
observation that the levels of these hormones change little or not at all (Wasserman et al.,
1993; Wasserman, 2009) informs us little about their importance as ultimately it is the
glucagon/insulin ratio (Richter & Galbo, 1986) and the portal levels of these hormones
that determine their effects on hepatic glucose production (Wasserman et al., 1989a). The
small changes in peripheral glucagon levels during exercise are due to the hepatic
extraction of glucagon released by the pancreas, leading to lesser rise in peripheral
glucagon levels and the underestimation of the physiological importance of glucagon in
blood glucose regulation (Wasserman et al., 1993). It has been shown that even a small
increase in glucagon level can have a marked effect on hepatic glucose production as the
potency of a given glucagon level is enhanced considerably during exercise compared to
rest (Wasserman et al., 1989a; Wasserman, 2009).
One powerful approach to evaluate the relative contributions of insulin and glucagon in
the activation of glucose production during exercise without the confounding effect of the

17
counterregulatory response to falling blood glucose level is to manipulate portal glucagon
and insulin levels by infusing an inhibitor (e.g. somatostatin, octreotide) of their
pancreatic release together with the infusion of insulin, glucagon and glucose to maintain
euglycaemia, a technique known as pancreatic islet clamp. Using this technique, insulin
and glucagon have been reported to account for ~55% and ~60% of the exercise-mediated
increase in hepatic glucose production in dogs, respectively (Wasserman et al., 1989a,
1989b). Similar techniques in humans have shown both hormones play an important role
in increasing hepatic glucose production during exercise since the absence of changes in
insulin and glucagon concentrations has been shown by many to prevent glucose
production rate from increasing (Wolfe et al., 1986; Hirsch et al., 1991; Kjaer et al.,
1993a; Lavoie et al., 1997). However, others have reported that the increase in hepatic
glucose production during moderate exercise is little or not affected when plasma insulin
and glucagon levels are held constant in humans (Bjorkman et al., 1983; Coker et al.,
2001). Therefore, despite playing some role, changes in glucagon and insulin do not
totally explain the increase in hepatic glucose production that takes place during moderate
intensity exercise in humans, thus indicating that other factors are involved (Hargreaves
and Spriet, 2006).
Catecholamines have been proposed to play some role in the activation of hepatic glucose
production during moderate intensity exercise. However, most studies have failed to
show a significant role for the sympathoadrenergic system although the increase in
epinephrine and norepinephrine levels has led some researchers to suggest the opposite.
Indeed, there is a strong correlation in humans between the exercise-induced rise in
hepatic glucose production and increases in catecholamines levels, with the amount of
active muscle mass influencing the magnitude of the rise in catecholamines (Kjaer et al.,
1991). Hypoxic exercise is another condition which results in a greater rise in both
hepatic glucose production and catecholamines (Cooper et al., 1986). Adrenal medulla
removal in rats decreases hepatic glycogenolysis (Richter et al., 1981) and hepatic glucose
production (Sonne et al., 1985) during exercise. However, some studies in rats have
shown no effect of epinephrine on hepatic glycogen breakdown during exercise
(Hargreaves and Spriet, 2006).
The small contribution of plasma catecholamines in mediating the increase in glucose
production during exercise is suggested by the studies which have shown that alpha- and
beta-adrenergic blockades have little or no effect on the rise in hepatic glucose production

18
during moderate exercise in dogs (Coker et al., 1997) and humans (Simonson et al, 1984;
Marker et al., 1991). Also, direct adrenergic stimulation at physiological dose has little
effect on the rate of hepatic glucose production during exercise even in the absence of
changes in glucagon and insulin levels (Coker et al., 2002). The small role played by
circulating catecholamines is not this surprising when one considers that epinephrine
levels in the portal circulation are much lower than in peripheral blood where
measurements are normally performed. This is because the gut extracts a large proportion
of blood epinephrine before it reaches portal circulation (Coker and Kjaer, 2005).
The role of hepatic nerves in the activation of hepatic glucose production has also been
investigated, with most studies suggesting that stimulation of hepatic nerves plays a
minimal role. For instance, hepatic denervation does not affect the rise in hepatic glucose
production during exercise in rats (Richter et al., 1980; Sonne et al., 1985) or dogs
(Wasserman et al., 1990). Similarly, blockade of sympathoadrenergic activity in healthy
males during exercise by local anaesthesia of the celiac ganglion that usually innervate
the liver and adrenal medulla, together with the infusion of glucagon and insulin
hormones and physiological doses of epinephrine to mimic usual responses to exercise,
has no effect on the increase in hepatic glucose production rate during exercise, indicating
that sympathetic liver nerve activity is unlikely to be involved (Kjaer et al., 1993a).
Likewise, patients with denervated liver transplants experience similar increases in
hepatic glucose production during exercise to those of control individuals, indicating
hepatic nerve activity is not an important glucoregulatory factor during exercise (Kjaer et
al., 1995).
Other than the sympathoadrenal system and pancreatic hormones, it has been proposed
that glucose itself plays some role in the regulation of hepatic glucose production during
exercise. In support of this view, a fall in glucose level during moderate intensity exercise
stimulates hepatic glucose production without changes in pancreatic hormone levels or
catecholamine release (Coker et al., 2002). These results support the view that
decrements in glycaemia may stimulate hepatic glucose production during moderate
exercise and therefore maintain euglycaemia. A small decline in glycaemia can also
indirectly stimulate hepatic glucose production by stimulating counterregulatory
hormone response (Wasserman et al., 1984, 1991), with an increase in growth hormone
(Shilo and Shamoon, 1990; Davis et al., 2000e) and cortisol levels being observed (Shilo
and Shamoon, 1990; Davis et al., 2000e; Horton et al., 2002). However, growth hormone

19
and cortisol are unlikely to play an important role as the levels of these hormones change
little during moderate intensity exercise, and experimentally induced growth hormone or
cortisol deficiency does not affect blood glucose levels during exercise (Hoelzser et al.,
1986a, 1986b; Wasserman, 1995). It is important to note, however, that when insulin and
glucagon levels are kept constant at basal levels during mild to moderate intensity
exercise, glucose production increases rapidly with a decrease in blood glucose level
before plateauing despite a decrease in glycaemia (Kjaer et al., 1993a). Hepatic glucose
production after prolonged low blood glucose levels thus appears reasonably insensitive
to small deceases in glycaemia (Kjaer et al., 1993a). In contrast, exogenous glucose
infusion or glucose ingestion affects insulin and glucagon secretion and markedly inhibits
hepatic glucose production during exercise (Manzon et al., 1998; Jeukendrup et al., 1999)
Jenkins et al., 1985 even in responses to very small changes in blood glucose levels
(Berger et al., 1994). This shows that hepatic glucose production is very sensitive to a
rise in blood glucose levels.
There is evidence that afferent neural reflex activity originating from exercising muscles
may be important for increasing hepatic glucose production during exercise. This is
illustrated by the observation that electrical stimulation of cut muscle branches of the
femoral nerves increases hepatic glucose production and glycaemia in cats (Vissing et al.,
1994). However, in humans, epidural blockade to evaluate the effect of neural feedback
does not change hepatic glucose production during moderate intensity exercise (Kjaer et
al., 1989). Therefore, although afferent neural reflex activity can increase glucose
mobilisation during exercise, it is likely to be secondary or only play a minor role
comparative to other mechanisms in healthy individuals.
There are other important factors that influence the glucoregulatory response to exercise.
In particular, counterregulatory responses to moderate intensity exercise are attenuated
by prior hypoglycaemia (Davis et al., 2000d), with blunting of glucagon, catecholamines,
GH, cortisol, endogenous glucose production, ketogenesis and lipolytic responses (Davis
et al., 2000d). Similarly, antecedent morning exercise of moderate intensity can
significantly impair metabolic and neuroendocrine responses during moderate exercise
performed three hours later in a gender-specific manner (Galassetti et al., 2001b).
Given that skeletal muscles release IL-6 during exercise, it has been proposed that this
might provide a means whereby muscle activity and associated fuel utilisation is related

20
to the regulation of hepatic glucose production. In support of this view, IL-6 increases
hepatic glucose production during exercise (Febbraio et al., 2004), and IL-6 levels
increase during exercise, following an almost exponential pattern reaching peak
concentration of up to 100-fold from basal levels immediately following exercise
(Pedersen and Fischer, 2007). The magnitude of IL-6 response is influenced by the
muscle mass recruited during exercise, endurance capacity, and both exercise intensity
and duration (Pedersen, 2009). IL-6 also increases both whole-body and intramuscular
fatty acid oxidation, thus contributing to the sparing of carbohydrate (Pedersen and
Febbraio, 2008). It must be stressed, however, that IL-6 also promotes fuel uptake and
utilisation in working muscle, and as a result increases whole body glucose disposal
during exercise (Febbraio et al., 2004), thus raising the question of its importance in
counterregulation.
1.6 Glucoregulatory Responses to Moderate Intensity Exercise in Individuals with
Type 1 Diabetes
In insulin-treated individuals with type 1 diabetes, moderate intensity exercise stimulates
an increase in muscle glucose utilisation that is similar in magnitude to non-diabetic
individuals (Richter and Galbo, 1986), but that increases with plasma insulin levels
(Chokkalingam et al., 2007). However, most but not all studies have reported that hepatic
glucose production does not rise to the same degree (Simonson et al., 1984; Richter and
Galbo, 1986), resulting in a decrease in blood glucose levels and a rise in hypoglycaemia
risk (Schiffrin et al., 1984; Simonson et al., 1984; Zinman et al., 1984; Hübinger et al.,
1985; Campaigne et al., 1987; Sonnenberg et al., 1990; Oskarsson et al., 1999; Riddell et
al., 1999; Francescato et al., 2004; Petersen et al., 2004). Furthermore, during recovery
from moderate intensity exercise in type 1 diabetic individuals, blood glucose levels
continue to decrease (Hübinger et al., 1985; Campaigne et al., 1987) or stabilise
(Simonson et al., 1984; Oskarsson et al., 1999) while both hepatic glucose production and
glucose utilisation decreasing back to basal levels.
The inability of hepatic glucose production rate to rise to a level that matches glucose
utilisation rate during moderate intensity exercise in individuals with type 1 diabetes is
due in part to the absence of a fall in circulating insulin levels. As discussed earlier, non-
diabetic individuals have the capacity to decrease the secretion of insulin during exercise,
thus favouring the stimulation of hepatic glucose production. In contrast, individuals with

21
type 1 diabetes are unable to control the rate of passive absorption of the insulin from the
previously administered insulin bolus, thus increasing their likelihood of exercising in a
hyperinsulinaemic state. Hyperinsulinaemia has been reported to increase the rate of
exercise-mediated glucose utilisation in type 1 diabetic individuals while having no effect
on hepatic glycogen breakdown (Chokkalingam et al., 2007).
Another factor which increases the mismatch between glucose production and utilisation
and thus increase further the risk of hypoglycaemia during exercise is the
hyperinsulinaemia that may occur as a result of increased insulin mobilisation from
subcutaneous depots, especially if insulin is injected near an exercising muscle (Koivisto
and Felig, 1978), with the rates of insulin absorption often increasing during exercise
(Richter & Galbo, 1986). Significant increases in insulin concentrations is also observed
during exercise in insulin-infused diabetic individuals, thus suggesting reduced insulin
clearance (Chokkalingam et al., 2007). Overall, since exercise not only increases insulin
absorption from subcutaneously administered injection depots, but also decreases insulin
clearance, these factors lead to elevated circulating insulin levels during exercise. This
state of over-insulinisation combined with exercise itself results in a profound increase in
insulin action (Camacho et al., 2005). Indeed, increased insulin level and action can
negate the effect of glucagon on hepatic glucose production and amplify glucose
utilisation to greater than required levels, thus resulting in a drop in blood glucose level
(Camacho et al., 2005). Whether the resulting fall in glycaemia reaches dangerous
hypoglycaemic levels greatly depends on pre-exercise blood glucose levels (Richter and
Galbo, 1986). Thus, in general, if insulin concentration is elevated, the lack of exercise-
induced decrease in insulin levels results in a marked activation of glucose utilisation
(Camacho et al., 2005). This together with decreased hepatic glucose production results
in a fall in blood glucose levels (Zinman et al., 1977) and a rise in risk of hypoglycaemia.
This risk is further exacerbated by the symptoms of hypoglycaemia that may alert an
individual to an impending episode such as sweating and tachycardia being difficult to
notice as they may be similar to normal responses to exercise.
Aside from the marked difference in insulin response to moderate intensity exercise
between diabetic and non-diabetic individuals, the response of counterregulatory
hormones to moderate intensity exercise in insulin-treated individuals with type 1
diabetes is relatively similar to that of non-diabetic individuals. Glucagon levels are
increased (Oskarsson et al., 1999; Galassetti et al., 2002, 2003, 2004, 2006) or unchanged

22
(Simonson et al., 1984; Hübinger et al., 1985; Sonnenberg et al., 1990), catecholamines
levels are increased (Simonson et al., 1984; Hübinger et al., 1985; Sonnenberg et al.,
1990; Oskarsson et al., 1999; Galassetti et al., 2002, 2003, 2004, 2006), and growth
hormone levels are elevated (Hübinger et al., 1985; Shilo and Shamoon, 1990;
Sonnenberg et al., 1990; Oskarsson et al., 1999; Galassetti et al., 2002, 2004). In contrast,
cortisol levels either remain stable (Hübinger et al., 1985; Sonnenberg et al., 1990) or
increase later (Oskarsson et al., 1999; Galassetti et al., 2002, 2003, 2004, 2006) during
moderate intensity exercise. It is noteworthy that despite the exercise-induced increase
in the levels of the circulating counterregulatory hormones being similar between diabetic
and non-diabetic individuals, hypoglycaemia risk is increased in insulin-treated diabetic
individuals, thus highlighting the glucoregulatory importance of insulin in glucose
homeostasis during moderate intensity exercise (Galassetti, et al., 2006).
There are a number of important factors that affect the counterregulatory response to
exercise in individuals with type 1 diabetes. In particular, this response is attenuated by
prior hypoglycaemia (Galassetti et al., 2003, 2006). This blunting effect appears to be
dose-dependent, with hypoglycaemia of increasing depth progressively inducing more
acute counterregulatory failure, with a reduction in glucagon, catecholamine, cortisol,
endogenous glucose production, and lipolytic responses (Galassetti et al., 2006). Prior
aerobic exercise also impairs the counterregulatory response associated with a subsequent
bout of moderate intensity exercise (Galassetti et al., 2001b).
Another important factor that influences glycaemic response to moderate intensity
exercise and associated risk of hypoglycaemia is the increase in insulin sensitivity that
occurs in response to exercise (Richter et al., 1982; Holloszy, 2005; Jensen and Richter,
2012). This together with the exercise-induced increase in glucose utilisation in insulin-
treated type 1 diabetic individuals can lead to a rapid decline in blood glucose levels not
only during exercise, but also for several hours afterwards (MacDonald, 1987b; Hirsch et
al., 1991; Tsalikian et al., 2005; McMahon et al., 2007). For this reason, it is generally
recommended that insulin dose should be reduced and carbohydrate intake increased prior
to and after exercise to reduce the aforementioned risk of hypoglycaemia (ADA, 2007;
Dube et al., 2005; Zinman et al., 2004).
It must be stressed, however, that in cases of severe insulin deficiency exercise provokes
a rise in blood glucose levels that increase further during recovery (Wahren et al., 1975).

23
The increase in blood glucose levels is explained in part on the basis that the magnitude
of the increase in muscle glucose utilisation is reduced due to the small additive effect of
exercise and insulin. In addition, low insulin is conducive to increased hepatic glucose
production at rates greater than rates of glucose utilisation, thus resulting in
hyperglycaemia (Riddell and Perkins, 2006). Finally, severe insulin deficiency is
accompanied by elevated glucagon levels, thus favouring the activation of hepatic glucose
production. Therefore, exercising in a state of severe insulin deficiency is not
recommended as the resulting lower insulin to glucagon ratio not only increases
glycaemia, but also ketone body levels, thus substantially increasing the risk of severe
ketoacidosis (Berger et al., 1980; Richter and Galbo, 1986) and ketoacidotic coma.
1.7 Glucoregulatory Responses to High-Intensity Aerobic Exercise in Non-Diabetic
Individuals
Although moderate intensity exercise increases risk of hypoglycaemia in insulin-treated
type 1 diabetic individuals, it is important to note that not all types of exercise increase
this risk. In fact, there are conditions where exercise is actually conducive to
hyperglycaemia not only in insulin-treated individuals with diabetes but also in non-
diabetic individuals. For instance, blood glucose levels in non-diabetic (Stokes et al.,
2013) and diabetic individuals (Turner et al., 2014) increase in response to resistance
exercise or when exercise is performed for at least 10 min at an intensity greater than 80%
of maximum rate of oxygen consumption ( V O2max; Mitchell et al., 1988; Marliss et al.,
1991,1992b; Purdon et al., 1993; Sigal et al., 1994b; Kreisman et al., 2000a; Marliss et
al., 2000; Sigal et al., 2000; Harmer et al., 2008; Manzon et al., 1998; Marliss & Vranic,
2002; Wahren & Ekberg, 2007). This rise is glycaemia is followed by a period of
hyperglycaemia that can persist for up to an hour post-exercise (Marliss et al., 1991, 2000;
Sigal et al., 2000).
The glycaemia-rising effect of intense aerobic exercise has been explained on the basis
that this type of exercise results in a seven- to eight-fold increase in glucose production
(Marliss et al., 1991,1992b; Purdon et al., 1993; Sigal et al., 1994b), the greatest increase
in glucose production observed under any physiological or pathophysiological condition
(Marliss and Vranic, 2002). This is accompanied by a lesser increase in glucose
utilisation rates of approximately 3-4 fold (Marliss et al., 1991, 1992b; Purdon et al.,
1993; Sigal et al., 1994b). This greater rate of glucose production in comparison to

24
glucose utilisation results in an increase in plasma glucose levels. A further increase in
glycaemia also occur immediately after the cessation of prolonged intense aerobic
exercise in non-diabetic individuals (Mitchell et al., 1988; Marliss et al., 1991, 1992b;
Sigal et al., 1994a, 1994b; Kreisman et al., 2000a; Marliss et al., 2000; Sigal et al., 2000),
before blood glucose levels return to pre-exercise levels. This is due to glucose utilisation
rate initially decreasing more rapidly than glucose production rates at the onset of
recovery, thus contributing further to greater hyperglycaemia (Marliss et al., 1992b; Sigal
et al., 1994b). This increase in glycaemia is only of short duration, with blood glucose
returning to pre-exercise levels by 60 min post-exercise.
Unlike moderate intensity exercise, the significant increase in hepatic glucose production
during high intensity exercise is unlikely to be due to changes in pancreatic hormone
levels. Insulin levels actually remain unchanged (Kjaer et al., 1986; Mitchell et al., 1988;
Marliss et al., 1991, 1992b; Kreisman et al., 2000a), or decline only slightly (Sigal et al.,
1994b; Marliss et al., 2000; Sigal et al., 2000), while glucagon levels increase minimally
(Marliss et al., 1991, 1992b; Purdon et al., 1993; Sigal et al., 1994b), resulting in a small
increase in glucagon to insulin ratio (Sigal et al., 1996; Manzon et al., 1998; Sigal et al.,
2000). As mentioned previously, it is important to note that changes in plasma glucagon
levels may underestimate the portal levels of this hormone due to hepatic extraction thus
suggesting glucagon could play an important role than that suggested by the
aforementioned studies. However, experiments involving islet clamps where insulin and
glucagon are kept at stable and basal levels show that this has no effect on hepatic glucose
production during intense aerobic exercise, thus indicating that these hormones are
unlikely to play a major role in increasing hepatic glucose production during high
intensity exercise (Sigal et al., 1996; Marliss and Vranic, 2002). A minor role for
pancreatic hormones is also suggested by the observation that the pattern of rise in glucose
production in response to high intensity exercise persists in glucose-infused individuals
despite high plasma insulin levels and insulin/glucagon ratios (Manzon et al., 1998).
The marked catecholamine response to high intensity exercise is thought to play a major
role in the activation of hepatic glucose production and rise in blood glucose levels in
non-diabetic individuals (Marliss and Vranic, 2002). This is supported by the observation
that the increase in glucose production associated with high intensity aerobic exercise is
accompanied by a 14- to 18-fold increase in both epinephrine and norepinephrine levels
(Kjaer et al., 1986; Marliss et al., 1991, 1992b; Sigal et al., 1994b; Manzon et al., 1998;

25
Kreisman et al., 2000a; Marliss et al., 2000; Sigal et al., 2000). However, this relationship
between catecholamine levels and an increase in hepatic glucose production during high
intensity exercise only suggests causality.
One approach to establish causality is to determine whether adrenergic blockade inhibits
the rise in hepatic glucose production during intense exercise. Although there is a well
demonstrated relationship between exercise-induced changes in glucose output and
circulating catecholamine levels, a role for catecholamines is challenged by the
observation that the administration of alpha or beta adrenergic blockers during high
intensity exercise has no inhibitory effect on the rise in hepatic glucose production (Sigal
et al., 1994b; Coker et al., 1997; Sigal et al., 1999, 2000). Indeed, the infusion of
propanolol (beta-adrenergic blocker) in non-diabetic individuals (Sigal et al., 1994a)
results in an unexpected higher rise rather than a lesser increase in hepatic glucose
production, and only a minor inhibition of glucose production is achieved in response to
phentolamine infusion (alpha-adrenergic blocker) during intense exercise in humans
(Sigal et al., 2000). Moreover, intraportal infusion of propanol and phentolamine to
selectively block hepatic and adrenergic receptors in dogs has no effect on endogenous
glucose production or net hepatic glucose output during intense exercise (Coker et al.,
1997). Overall, these findings do not exclude the possibility that other factors may
contribute to the regulation of glucose production during intense exercise, particularly
considering the lack of any effect of adrenergic blockade on glucose production (Sigal et
al., 1994a, 1999, 2000)” One limitation with these studies, however, is that it remains to
be determined whether the dose of adrenergic blockers administered was sufficient to
counter the effect of the large rise in catecholamines associated with intense exercise.
One of the most convincing pieces of evidence that catecholamines play a role in
stimulating hepatic glucose production during intense aerobic exercise is the fact that
when epinephrine and norepinephrine are infused during moderate intensity exercise to
mimic the levels attained during higher intensity exercise there is a further increase in
hepatic glucose production and rise in blood glucose levels that matches these observed
during high intensity exercise, therefore suggesting an important role for epinephrine and
norepinephrine. When infused alone, epinephrine (Howlett et al., 1999a; Kreisman et al.,
2000b) and norepinephrine (Kreisman et al., 2001) are each considered capable of
producing a portion of the increase in hepatic glucose production associated with intense
exercise. Moreover, blockade of sympathetic nerve activity to liver and adrenal medulla

26
by local anaesthesia of the celiac ganglion followed by infusion of high physiological
doses of epinephrine during exercise resulted in enhanced glucose production in healthy
males (Kjaer et al., 1993a). Also, when both catecholamines are infused together to mimic
the usual epinephrine and norepinephrine response to high intensity exercise in healthy
men, there is a marked and progressive increase in hepatic glucose production and
increase in glycaemia, suggesting a role for catecholamines in the regulation of hepatic
glucose production during high intensity exercise (Kreisman et al., 2003). However, it is
noteworthy that in epinephrine-deficient, bilaterally adrenalectomised humans, the
absence of epinephrine does not impair glucose production during high intensity exercise
(Howlett et al., 1999b), thus suggesting that norepinephrine might play a more important
role here and that other glucoregulatory factors might be involved.
Acute changes in the levels of other glucoregulatory hormones such as growth hormones
and cortisol are unlikely to have any effect on the acute increase in blood glucose levels
associated with high intensity exercise. This is, in part, because the levels of these
hormones change little during a short bout of intense exercise, with their levels increasing
mainly during recovery (Marliss & Vranic, 2002). Also, the infusion of octreotide to
inhibit insulin, glucagon and growth hormones secretion while glucagon and insulin
levels are replaced and maintained at stable levels has no effect on the increase in glucose
production during intense exercise (Marlis & Vranic, 2002; Sigal et al., 1996).
In order to understand how intense exercise increases blood glucose level it is important
to focus not only on the potential effect of catecholamines and other factors on hepatic
glucose production, but also on the factors that influence the rates of glucose utilisation
during exercise. On the basis of the findings by some that catecholamines inhibit insulin-
stimulated glucose utilisation in resting skeletal muscles (Aftab-Guy et al., 2005;
Wasserman, 1995) and glucose Rd during exercise (Howlett et al., 1999; Watt &
Hargreaves, 2002; Watt et al., 2001), in part via a glycogenolytically-mediated increase
in glucose-6-phosphate level (Watt et al., 2001), a potent inhibitor of hexokinase
(Nonogaki, 2000), it is possible that the lesser rise in glucose utilisation relative to glucose
production during intense exercise is due to a catecholamine-mediated inhibition of the
stimulatory effect of muscle contraction on peripheral glucose utilisation. This issue was
investigated in epinephrine-deficient, bilaterally adrenalectomised humans with
epinephrine infusion inhibiting glucose clearance during exercise (Howlett et al., 1999b).
Similarly, studies in rodents have also shown that epinephrine infusion decreases muscle

27
glucose transport despite an elevation in glucose transporter (GLUT4) translocation to
the plasma membrane, which suggests a reduction in GLUT4 intrinsic activity (Bonen et
al., 1992). Despite these findings, other researchers have found that elevated
catecholamines in humans stimulate rather than inhibit glucose utilisation (Kreisman et
al., 2000b, 2001, 2003), and these investigators proposed that the increase in blood
glucose levels during intense aerobic exercise could simply be the result of a greater
stimulation of glucose production by catecholamines compared to glucose utilisation rates
(Kreisman et al., 2003).
The apparent contradiction with respect to the role of catecholamines on glucose
utilisation rate is not surprising given the evidence that the stimulation of muscle
adrenoceptors by catecholamines and other adrenoceptor agonists can stimulate glucose
transport in skeletal muscle (Abe et al., 1993; Ngala et al., 2013), with the effect of
adrenoceptor antagonists having little or a marked effect depending on muscle fibre
compositions (Liu et al., 1995). Also, predicting the effect of catecholamines on
peripheral glucose utilisation rate is further complicated by the observation that
circulating epinephrine inhibits glucose uptake in skeletal muscle by insulin-dependent
mechanisms, whereas norepinephrine released from sympathetic nerves increases glucose
uptake via insulin-independent mechanisms (Nonogaki, 2000).
The post-exercise transient increase in blood glucose levels and its subsequent fall are
explained on the basis of the pattern of change in plasma insulin, glucose and
catecholamines levels. Immediately after intense exercise, catecholamine levels are
elevated and inhibit insulin secretion, thus contributing to a further transient increase in
blood glucose levels. Afterwards, the rapid decrease in catecholamine levels and high
blood glucose levels result in a marked rise in plasma insulin level (Marliss et al., 1991;
Purdon et al., 1993; Sigal et al., 1994a, 1994b; Kreisman et al., 2000a; Marliss et al.,
2000) that helps to decrease glucose back to basal level (Marliss and Vranic, 2002).
Indeed, elevated blood glucose and insulin levels enhance the rate of glucose transport
during recovery from intense exercise while inhibiting hepatic glucose production. It is
noteworthy that this rapid decline in epinephrine and norepinephrine levels during the
first few minutes of recovery (Marliss et al., 1991; Kreisman et al., 2000a; Marliss et al.,
2000; Sigal et al., 2000) is closely matched with the post-exercise decrease in hepatic
glucose production (Marliss et al., 2000; Marliss and Vranic, 2002), supporting the
opinion that declining catecholamine levels may potentially mediate the post-exercise fall

28
in glycaemia in non-diabetic individuals. Other factors might contribute to the fall in
hepatic glucose production during recovery. During recovery from high intensity aerobic
exercise, lactate (Marliss et al., 1991; Sigal et al., 1994b; Kreisman et al., 2000a; Marliss
et al., 2000) levels decline. Since lactate is a potential glucoregulatory precursor, lower
lactate levels would be expected to result in a decrease in hepatic gluconeogenesis.
Likewise, glucagon levels decline (Marliss et al., 1991; Sigal et al., 1994b) or remain
relatively stable during recovery is taking place during high intensity exercise (Marliss et
al., 2000), thus producing a further fall in hepatic glucose production. Other
counterregulatory hormones such as growth hormone and cortisol are unlikely to be
important in the fall in glycaemia observed during recovery from prolonged high intensity
exercise.
It is noteworthy that the glucoregulatory response to prolonged high intensity exercise is
affected by a number of factors. Firstly, the state of feeding seems to be one of these
factors, with a smaller increase in glycaemia observed during high intensity exercise
performed postprandially as opposed to in a postabsorptive state (Kreisman et al., 2000a).
Unlike moderate intensity exercise, glucose production during high intensity exercise is
not as sensitive to the inhibitory effect of hyperglycaemia (Manzon et al., 1998; Wiersma
et al., 1993). Gender may also play a role, since women experience a greater rise in
glycaemia in the post-exercise period due to a lesser increment in glucose utilisation rate
(Marliss et al., 2000) despite men and women experiencing a similar glucoregulatory
response to high-intensity exercise. This is also associated with higher insulin levels in
women compared to men post-exercise, although glucose levels return to basal within a
similar time-frame (Marliss et al., 2000). Finally, training may influence the
glucoregulatory response to high intensity exercise, with an increase in exercise-induced
epinephrine levels contributing to a greater increase in blood glucose levels due to
increased hepatic glucose production in exercise-trained individuals (Kjaer et al., 1986).
However, recently it has been reported that the rise in blood glucose levels in response to
intense exercise is less in sprint-trained individuals (Harmer et al., 2007).

29
1.8 Glucoregulatory Responses to High-Intensity Aerobic Exercise in Individuals
with Type 1 Diabetes
Not all forms of exercise result in an acute fall in blood glucose levels. For instance,
resistance exercise has been shown in some studies to result in a post-exercise increase in
blood glucose level (Turner et al., 2014). Also, when exercise is performed at high
intensity (>80% V O2max) for at least 10 min in insulin-treated patients with type 1
diabetes, blood glucose levels increase (Mitchell et al., 1988; Purdon et al., 1993; Sigal
et al., 1994a) in a manner similar to non-diabetic individuals. One major difference,
however, is that during recovery, blood glucose level rises and remains elevated for up to
several hours post-exercise. This is in marked contrast to non-diabetic individuals where
blood glucose levels return to pre-exercise levels within one hour.
The increase in blood glucose levels during intense aerobic exercise in diabetic
individuals is due to the approximately 7-fod increase in hepatic glucose production rate
(Purdon et al., 1993) being higher than the approximately 4-fold increase in glucose
utilisation (Purdon et al., 1993; Sigal et al., 1994a). During early recovery, blood glucose
levels continue to rise for a few minutes in these individuals due to glucose production
rate being comparatively higher than glucose utilisation (Purdon et al., 1993; Sigal et al.,
1994a). Then glucose production and utilisation rate decline, with these processes
matching each other as hyperglycaemia remains for at least an hour (Sigal et al., 1999)
and up to several hours after the cessation of exercise (Mitchell et al., 1988; Purdon et al.,
1993; Sigal et al., 1994a).
The glucoregulatory mechanisms responsible for the increase in blood glucose level
during high intensity exercise are similar in both diabetic and non-diabetic individuals.
A large catecholamine response to high intensity exercise is also reported in individuals
with diabetes, with a similar relative increase of approximately 14-fold for both
epinephrine and norepinephrine levels (Purdon et al., 1993). Likewise, glucagon
response to high intensity exercise is similar in both diabetic and non-diabetic individuals,
together with similar increase in lactate (Mitchell et al., 1988) and cortisol levels (Purdon
et al., 1993). However, the pattern of free fatty acid response is different, with an increase
during recovery from high intensity exercise in individuals with diabetes, while free fatty
levels decrease in non-diabetic individuals (Mitchell et al., 1988).

30
The sustained rise in glycaemia throughout recovery in diabetic compared to non-diabetic
individuals results mainly from the absence of a post-exercise increase in insulin levels
(Mitchell et al., 1988; Purdon et al., 1993; Sigal et al., 1994a, 1999). Such a role for
insulin is suggested by the observation that the administration of insulin during recovery
from high intensity exercise accelerates the return of blood glucose to pre-exercise levels
and prevents prolonged hyperglycaemia (Purdon et al., 1993; Sigal et al., 1994a). Due to
the absence of a post-exercise increase in insulin release, there is no stimulus to promote
glucose uptake by skeletal muscles or to inhibit hepatic glucose production by the liver.
For these reasons, blood glucose levels in individuals with type 1 diabetes remain elevated
or continues to rise well after the cessation of exercise (Mitchell et al., 1988; Purdon et
al., 1993; Sigal et al., 1994a, 1999; Marliss and Vranic, 2002).
1.9 Glucoregulatory Responses to Intermittent High Intensity Exercise in Non-
Diabetic Individuals
Although the glucoregulatory responses to both moderate and high intensity exercise have
been researched comprehensively as described above, this is not the case for intermittent
high intensity, a pattern of activity where moderate intensity is combined with multiple
sprints performed in succession. This is unfortunate given that this pattern of physical
activity characterises most team sports such as soccer, rugby, Australian Rules football,
netball and basketball (Docherty et al., 1988; Bangsbo et al., 1991; Spencer et al., 2004;
Spencer et al., 2005; Bishop and Wright, 2006; Davidson and Trewartha, 2008).
This combination of multiple sprints interspersed with active or passive recovery
generally results in an increase in blood glucose levels and glucoregulatory responses
similar to high intensity exercise. In particular, multiple prolonged sprints of 45-60 sec
duration each interspersed with 3-5 min periods result in a progressive rise in glycaemia
after each work period in non-diabetic individuals (Hermansen et al., 1970). Blood
glucose often continues to rise following the cessation of exercise (Hermansen et al.,
1970; Näveri et al., 1985) with a peak in glycaemia reported ~5 min into recovery
(Hermansen et al., 1970) before blood glucose declines to pre-exercise levels. Such
multiple sprints of 45-60 sec duration are associated with a several-fold increase in the
levels of epinephrine, norepinephrine, lactate, glucagon, growth hormone and insulin
levels in non-diabetic individuals (Näveri et al., 1985). It is important to note, however,
that these prolonged sprints are physically demanding and are not representative of those

31
observed in team sports where each sprint lasts generally less that 10 sec (Spencer et al.,
2005). Sports such as soccer, rugby, hockey, netball, basketball and Australian Rules
football have average sprint duration of 2-4 sec (Docherty et al., 1988; Bangsbo et al.,
1991; Spencer et al., 2004, 2005; Bishop and Wright, 2006; Davidson and Trewartha,
2008).
Several studies have investigated on the glucoregulatory response to multiple sprint
protocols that more closely mimic team sport with shorter duration sprints of 10 sec or
less (Brooks et al., 1990; Hamilton et al., 1991; Gaitanos and Williams, 1993; Nevill et
al., 1993; Goto et al., 2007; Trapp et al., 2007; Bracken et al., 2009). These patterns of
exercise result in an overall increase in glycaemia post-exercise in non-diabetic
individuals (Holmyard et al., 1988; Brooks et al., 1990; Hamilton et al., 1991; Gaitanos
and Williams, 1993; Nevill et al., 1993). Blood glucose levels typically peak early during
recovery (3-5 min; Brooks et al., 1990; Gaitanos and Williams, 1993), with blood glucose
levels increasing by approximately 1-2.5 mM (Brooks et al., 1990; Gaitanos and
Williams, 1993; Nevill et al., 1993).
This increase in glycaemia in non-diabetic individuals after multiple short sprints is
associated with a several-fold increase in epinephrine (Brooks et al., 1990; Gaitanos and
Williams, 1993; Goto et al., 2007; Trapp et al., 2007; Bracken et al., 2009),
norepinephrine (Brooks et al., 1990; Gaitanos and Williams, 1993; Goto et al., 2007;
Trapp et al., 2007; Bracken et al., 2009) and blood lactate levels (Brooks et al., 1990;
Hamilton et al., 1991; Gaitanos and Williams, 1993; Nevill et al., 1993; Goto et al., 2007;
Trapp et al., 2007; Bracken et al., 2009). When multiple short sprints of less than 10 sec
duration are performed in non-diabetic individuals, a decrease in pH is also observed
(Brooks et al., 1990; Gaitanos and Williams, 1993; Goto et al., 2007; Bracken et al., 2009)
together with an increase in growth hormone (Goto et al., 2007), and glycerol levels (Goto
et al., 2007; Trapp et al., 2007).
1.10 Glucoregulatory Responses to Intermittent High Intensity Exercise in
Individuals with Type 1 Diabetes
Until quite recently, there has been very limited information on the glucoregulatory
responses to intermittent high intensity exercise in individuals with type 1 diabetes. The
information that was available came from studies focusing on multiple long sprints of 20

32
to 60 sec duration. One study involved 30 min of cycling interspersed with successive
one-min periods of high intensity cycling at 100% of maximal oxygen uptake, each
separated by a one-min rest between (Sills and Cerny, 1983). This resulted in a decline
in blood glucose levels during exercise in individuals with type 1 diabetes, but glycaemia
stabilised during early recovery (Sills and Cerny, 1983). An increase in growth hormone
was also observed while glucagon and cortisol levels remained stable in those individuals
(Sills and Cerny, 1983). Another study investigating the glycaemic response of type 1
diabetic individuals to multiple 20-sec sprints at 120% V O2max performed every 2 min,
with active recovery in between high intensity bouts, shows no significant change in
blood glucose levels during the multiple sprints or recovery (Ford et al., 1999). As
mentioned above, multiple high intensity bouts of prolonged duration are both physically
demanding and do not reflect the activity patterns of repeated sprints in most sports where
a typical sprint is usually very short in duration.
Recently, some studies have been performed in individuals with type 1 diabetes to
investigate their responses to the multiple short duration sprints typical of many team
sports (Guelfi et al., 2007a). Twenty min of repeated 4-sec sprints each interspersed with
2 min of passive recovery result in a decline in glycaemia, but blood glucose levels
stabilise during recovery (Guelfi et al., 2005c). This stabilisation was associated with
elevated catecholamine and growth hormone levels (Guelfi et al., 2005c). In another
study, 30 min of repeated 4-sec sprint cycling were once again performed, each separated
by 2 min of active recovery at 40% V O2peak was compared to constant moderate intensity
cycling at 40% V O2max (Guelfi et al., 2005a). In this study, the rate of decline in
glycaemia was lower during the multiple sprint protocol compared to sustained exercise,
with no further decrease during early recovery. In contrast, blood glucose levels declined
during early recovery from sustained exercise (Guelfi et al., 2005a). The lesser rate of
fall in blood glucose levels during intermittent compared to sustained exercise was
associated with increased catecholamine and growth hormone levels (Guelfi et al.,
2005a). Such a multiple short sprint protocol induces a greater increase in glucose
production rate during exercise than moderate intensity alone, with glucose utilisation
rate being similar in response to both exercise protocols (Guelfi et al., 2007b). The
findings of the aforementioned studies and of more recent ones (Dube et al., 2013;
Campbell et al., 2014) suggest that the risk of exercise-mediated hypoglycaemia during
exercise are lower if moderate intensity exercise is interspersed with repeated short sprints

33
(Guelfi et al., 2005a). It is noteworthy that resistance exercise also results in a lesser initial
rate of fall in blood glucose level than aerobic exercise (Yardley et al., 2012, 2013).
However, the effect of intermittent high intensity exercise on the risk of late onset
hypoglycaemia was not examined in these studies.
Two recent studies performed on individuals with type 1 diabetes have investigated the
risk of late-onset post-exercise hypoglycaemia (LOPEH) following intermittent high
intensity exercise (Maran et al., 2010; Iscoe and Riddell, 2011), but with conflicting
findings. One study reported an increased risk of LOPEH with repeated 5-sec sprints
compared to moderate intensity exercise (Maran et al., 2010). However, since energy
intake post-exercise was not measured and high intensity exercise can suppress appetite
(King et al., 1994), it is possible these findings could be explained by a lower
carbohydrate intake post-exercise as opposed to the exercise protocol itself. In contrast,
another study found that repeated 15-sec sprints as part of an intermittent high intensity
exercise protocol was associated with a lower risk of LOPEH in athletes with type 1
diabetes (Iscoe and Riddell, 2011). The discrepancy between the two studies may be
explained by differences in fitness levels and training status of the participants together
with different sprint durations and the nature of the exercise protocol being performed.
1.11 Glucoregulatory Responses to a Single Short Sprint in Non-Diabetic
Individuals
In non-diabetic individuals, the performance of a single short-duration maximal sprint
effort generally results in a post-exercise increase in blood glucose level (Hermansen et
al., 1970; Schnabel et al., 1983, 1984; Cheetham et al., 1985; Lavoie et al., 1987; Brooks
et al., 1988; Nevill et al., 1989; Allsop et al., 1990; Langfort and Zarzeczny, 1997; Bell
et al., 2001; Moussa et al., 2003; Vincent et al., 2004; Saraslanidis et al., 2009; Zouhal et
al., 2009) across a broad range of sprint duration. For instance, a 6-sec sprint in untrained
males results in an increase in glycaemia of approximately 1 to 2 mM, peaking 20 to 30
min after sprinting (Moussa et al., 2003). Similarly, a longer 30-sec sprint results in an
increase in blood glucose levels of approximately 1 to 2 mM, but with maximal levels
attained approximately 5 to 10 min after the sprint (Cheetham et al., 1985; Brooks et al.,
1988; Nevill and Boobis, 1989; Allsop et al., 1990; Langfort and Zarzeczny, 1997; Bell
et al., 2001; Moussa et al., 2003; Vincent et al., 2004; Zouhal et al., 2009). Sprinting of
even longer duration (40 to 60 sec) also increases blood glucose by approximately 1 to 2

34
mM, with maximal levels attained 5 to 10 min post-sprint with an increase of (Schnabel
et al., 1983, 1984; Saraslanidis, 2009). Likewise, a 90-sec sprint increases glycaemia by
approximately 1.5 to 2 mM (Lavoie et al., 1987). Although most studies show that a short
sprint increases blood glucose level, one study has reported that glycaemia remains stable
during recovery from a single short sprint (Vincent et al., 2004).
Interestingly, Moussa and colleagues (2003) reported significantly higher blood glucose
levels following a 6-sec sprint compared to a 30-sec sprint at both 20 and 30 min of
recovery in untrained non-diabetic males. The authors could not explain these findings
on the basis of changes in the levels of the glucoregulatory hormones measured in their
study, so they hypothesised a higher rate of muscle glucose utilisation following the 30-
sec sprint than the 6-sec sprint to replenish muscle glycogen stores may explain the
smaller increase in glycaemia during recovery (Moussa et al., 2003).
The glucoregulatory responses associated with a single maximal sprint efforts are
qualitatively comparable to those associated with high-intensity aerobic exercise. A short
duration maximal sprint effort results in a strong sympatho-adrenal response resulting in
a several-fold increase in epinephrine and norepinephrine levels in non-diabetic
individuals (Kindermann et al., 1982; Schnabel et al., 1983, 1984; Lavoie et al., 1987;
Brooks et al., 1988; Nevill et al., 1989; Allsop et al., 1990; Langfort et al., 1997; Zouhal
et al., 1998; Bell et al., 2001; Zouhal et al., 2001; Jacob et al., 2002; Moussa et al., 2003;
Vincent et al., 2003; Jacob et al., 2004; Vincent et al., 2004; Botcazou et al., 2007;
Bracken et al., 2009; Zouhal et al., 2009). This large increase in blood catecholamine
levels is observed irrespective of sprint duration. Epinephrine levels double immediately
following a 6-sec sprint, with maximal levels of approximately 1.2 nmol/l and 0.6 nmol/l
in men and women, respectively (Moussa et al., 2003; Botcazou et al., 2006, 2007;
Bracken et al., 2009). The duration of the sprint is one factor influencing the
sympathoadrenal activity with significantly higher epinephrine levels recorded after a 30-
sec sprint than a 6-sec sprint in untrained males (Moussa et al., 2003). In response to a
30-sec sprint, most studies report a 3 to 7 fold increase in epinephrine levels from resting
values (Brooks et al., 1988; Nevill et al., 1989; Langfort et al., 1997; Moussa et al., 2003;
Jacob et al., 2004; Vincent et al., 2004) with a peak concentration of approximately 1.4
to 4.5 nmol/l (Brooks et al., 1988; Nevill et al., 1989; Langfort et al., 1997; Zouhal et al.,
1998, 2001; Jacob et al., 2002; Moussa et al., 2003; Vincent et al., 2003; Jacob et al.,
2004; Vincent et al., 2004; Zouhal et al., 2009). Trained individuals attain even higher

35
peak epinephrine levels, ranging from approximately 7 to 8 nmol/l (an ~ 8 to 10 fold
increase), or at least double those of untrained individuals (Zouhal et al., 1998, 2001).
One study in active individuals reported an approximately 25-fold increase in epinephrine
levels, peaking at 10.2 nmol/l immediately after a 30-sec treadmill sprint (Allsop et al.,
1990). Longer sprint lasting 40 to 60 sec also results in a large increase in epinephrine
levels (Schnabel et al., 1983, 1984; Ohkuwa et al., 1984; Schwarz et al., 1990) as is the
case for a 90-sec sprint (Kindermann et al., 1982; Lavoie et al., 1987).
Plasma norepinephrine levels also increase markedly in response to a single short sprint,
with the magnitude of the increase linked to sprint duration. A single 6-sec sprint results
in an approximately 2-fold elevation in norepinephrine levels in men (Moussa et al., 2003;
Botcazou et al., 2006, 2007; Bracken et al., 2009) and women (Botcazou et al., 2007),
with maximal levels of approximately 5 nmol/l. Significantly higher norepinephrine
levels are recorded after a 30-sec sprint, with a 5-fold increase (Moussa et al., 2003) to
approximately 13.5 nmol/l in untrained males (Moussa et al., 2003). Numerous other
studies have shown that a 30-sec sprint is associated with a 5-6 fold increase in
norepinephrine levels (Brooks et al., 1988; Nevill et al., 1989; Zouhal et al., 1998, 2001;
Moussa et al., 2003; Vincent et al., 2003, 2004; Jacob et al., 2004; Zouhal et al., 2009) to
levels around 11 to 15 nmol/l (Brooks et al., 1988; Nevill et al., 1989; Langfort et al.,
1997; Zouhal et al., 1998, 2001; Moussa et al., 2003; Vincent et al., 2003; Jacob et al.,
2004; Vincent et al., 2004; Zouhal et al., 2009) in untrained individuals. Trained
individuals attain even higher peak norepinephrine levels, ranging from approximately
16 to 18 nmol/l (~ 7 to 10 fold increase) in endurance-trained individuals (Zouhal et al.,
2001, 2009) and up to 30 nmol/l (~ 14 fold increase) in sprint-trained individuals (Jacob
et al., 2002). One study of active individuals reported an approximately 30-fold increase
in norepinephrine levels, which reached 37.1 nmol/l immediately after a treadmill-sprint
(Allsop et al., 1990). Longer sprints lasting 40-60 sec (Schnabel et al., 1983, 1984;
Ohkuwa et al., 1984; Schwarz et al., 1990) and 90 sec (Kindermann et al., 1982; Lavoie
et al., 1987) also result in a large increase in norepinephrine levels. This large rise in
catecholamines immediately after short sprints of different durations has been proposed
to activate hepatic glucose production and inhibit insulin-mediated glucose uptake in
skeletal muscles (Nonogaki, 2000).
The pattern of change in plasma insulin levels in response to a sprint varies across studies
in non-diabetic individuals. Plasma insulin levels are unchanged following 6-sec and 30-

36
sec sprints in one study of untrained males (Moussa et al., 2003). Other studies have also
reported no change in insulin levels after a single 30-sec sprint in young active male
participants (Vincent et al., 2004; Zouhal et al., 2009). In contrast, a 30-sec sprint has
been reported to increase insulin levels during recovery in young active female
participants (Vincent et al., 2004), untrained 21 year old males, and trained 34 year old
males (Zouhal et al., 2009). Longer duration 45 to 50-sec sprints also result in an increase
in insulin levels (Schnabel et al., 1984) as do 90-sec sprints (Kindermann et al., 1982;
Lavoie et al., 1987). More recently, a decrease in insulin after a 30-sec sprint was reported
in young trained participants (Zouhal et al., 2009). The lack of increase in insulin in some
studies in response to the rise in blood glucose levels post-sprint may be due to the
inhibition of glucose-mediated stimulation of insulin secretion by the elevated levels of
circulating catecholamines (Marliss and Vranic, 2002). In most studies where plasma
insulin level increases, insulin helps to bring glucose back to pre-exercise levels by
stimulating glucose uptake by skeletal muscles and/or inhibiting hepatic glucose
production (Marliss and Vranic, 2002).
Changes in growth hormone level also occur in response to sprinting. Growth hormone
levels increases after a sprint, peaking approximately 20 to 60 min post- print (Nevill et
al., 1996; Stokes et al., 2002, 2003, 2004, 2005, 2006; Gilbert et al., 2008). Growth
hormone levels after sprinting remain elevated for at least 60 min (Nevill et al., 1996;
Stokes et al., 2002, 2004, 2005) and even for up to 90 to 120 min in some individuals
(Stokes et al., 2002). It should be noted that many studies have reported marked inter-
individual variability in growth hormone response to sprinting (Stokes et al., 2002; Stokes
et al., 2003). Sprint duration may be a factor influencing growth hormone response
(Stokes et al., 2003), with the increase in growth hormone levels being significantly less
following a 6-sec compared to a 30-sec sprint (Stokes et al., 2002). A sprint duration of
30 to 90-sec also increases markedly growth hormone levels (Gordon et al., 1994; Nevill
et al., 1996; Stokes et al., 2002, 2003, 2005; Gilbert et al., 2008).
There are a number of factors that may influence growth hormone response to a single
sprint effort. Firstly, the mode of exercise may influence growth hormone response with
the larger muscle mass recruited during treadmill sprinting resulting in greater increases
in growth hormone than sprint cycling (Stokes et al., 2002). There is also a trend towards
faster pedaling rates in cycling at a given workload resulting in greater growth hormone
concentrations (Stokes et al., 2002). The increase in growth hormone in response to

37
sprinting is also larger in sprint-trained compared to endurance-trained athletes (Nevill et
al., 1996). However, it is noteworthy that 6 weeks of combined speed- and speed-
endurance training blunts the growth hormone response to a single sprint (Stokes et al.,
2004). Finally, age may also influence the magnitude of growth hormone response, with
the rise in growth hormone levels after a 30-sec sprint being attenuated in older males
(Stokes et al., 2006; Gilbert et al., 2008).
There is evidence that the rise in growth hormone level may contribute to the acute
increase in blood glucose level post-sprint. Indeed, the administration of a physiological
growth hormone pulse in non-exercised individuals results in a rapid fall in muscle
glucose uptake (Moller et al., 1990, 1992b, 2003). If this were to be the case in resting
individuals recovering from a sprint, growth hormone could potentially contribute to the
post-sprint rise in blood glucose levels. However, evidence against this view is the
finding that growth hormone levels increase mainly during recovery (Kinderman, et al.,
1982; Schnabel et al., 1984; Stokes et al., 2002) and the infusion of octreotide (a
somatostatin analogue), an inhibitor of growth hormone release, has no acute effects on
the magnitude of the hyperglycaemic effect of high intensity exercise (Sigal et al., 1996).
Another counterregulatory hormone affected by sprinting is cortisol. Some studies have
shown that a sprint of 30 to 90-sec results in an increase in plasma cortisol levels which
peak approximately 15 to 20-min after sprinting (Kindermann et al., 1982; Buono et al.,
1986; Nevill et al., 1996). In contrast, 45 to 60 sec (Schnabel et al., 1984; Schwarz and
Kindermann, 1990) sprints have been reported not to change cortisol levels. Although
there is some published evidence that a rise in cortisol levels may play a role in stabilising
glycaemia due to its potential acute inhibitory effect on glucose utilisation in skeletal
muscles (Shamoon et al., 1980), this hormone is unlikely to contribute to early changes
in glycaemia post-sprint because cortisol levels increase after the post-exercise rise in
blood glucose level and the effects of cortisol on hepatic glucose production and blood
glucose levels require several hours to take place (De Feo et al., 1989a; Heller and Cryer,
1991; Marker et al., 1991; Mitrakou et al., 1991).
Another key glucoregulatory factor that increases after a single sprint effort in non-
diabetic individuals is lactate. Sprinting, irrespective of its duration, results in an increase
in blood lactate levels that peak approximately 5 to 7 min into recovery. A 6-sec sprint
results in an increase in blood lactate levels (Stokes et al., 2002; Moussa et al., 2003;

38
Botcazou et al., 2006; Bracken et al., 2009). Likewise, a 30-sec sprint causes a large
increase in blood lactate levels (Cheetham et al., 1985; Jones et al., 1985; Hardman et al.,
1986; Brooks et al., 1988; Nevill et al., 1989; Allsop et al., 1990; Bogdanis et al., 1994,
1995; Granier and et al, 1995; Bogdanis et al., 1996; Nevill et al., 1996; Langfort et al.,
1997; Weinstein et al., 1998; Bell et al., 2001; Zouhal et al., 2001; Jacob et al., 2002;
Stokes et al., 2002; Moussa et al., 2003; Vincent et al., 2003; Jacob et al., 2004; Gilbert
et al., 2008; Chiappa et al. 2009; Zouhal et al., 2009). This increase in lactate is of greater
magnitude after a 30-sec sprint, peaking at around 12 to 16 mM (Brooks et al., 1988;
Moussa et al., 2003). Longer sprints of 40 to 60 sec (Fujitsuka et al., 1982; Schnabel et
al., 1983; Ohkuwa et al., 1984; Schnabel et al., 1984; Saraslanidis et al., 2009) and up to
90 sec (Kindermann et al., 1982; Lavoie et al., 1987) are also associated with very large
increases in blood lactate levels. Not surprisingly, the increase in lactate is more
pronounced in sprint-trained athletes (Cheetham et al., 1985; Granier et al., 1995).
Middle distance runners who complete approximately three anaerobic training sessions
per week reach significantly higher peak lactate concentrations of approximately 15 mM
after a single sprint than long distance runners and untrained individuals (Jacob et al.,
2004). This increase in lactate levels may contribute to the early post-sprint increase in
glycaemia by providing gluconeogenic precursors for hepatic glucose production (Miller
et al., 2002) and by increasing peripheral insulin resistance (Vettor et al., 1997).
A short duration maximal sprint effort also results in a decrease in blood pH in non-
diabetic individuals (Brooks et al., 1988; Nevill et al., 1989; Allsop et al., 1990; Bogdanis
et al., 1995; Bogdanis et al., 1996; Stokes et al., 2002; Bracken et al., 2009). This decrease
in blood pH occurs after a single sprint of both 6 sec (Stokes et al., 2002; Bracken et al.,
2009) and 30 sec (Allsop et al., 1990; Bogdanis et al, 1995; Bogdanis et al., 1996; Stokes
et al., 2002). There is evidence that low blood pH can inhibit glucose utilisation by
skeletal muscles (Gordon et al., 1994), thus contributing to the early rise in blood glucose
levels post-sprint.
Changes in free fatty acid levels are also associated with sprinting. An increase in free
fatty acid levels has been reported to continue for at least 4 hours after a single 30-sec
cycle sprint (Stokes et al., 2005, 2008). In contrast, a decrease in FFA was recorded early
immediately after a longer approximately 45-sec treadmill sprint (Schnabel et al., 1984).
Since increases in free fatty acid levels occur hours after a sprint, they are unlikely to play
a role in the increase in glycaemia early in recovery from a short maximal sprint effort.

39
The mechanisms underlying the glycaemic rising effect of a short sprint differ markedly
from those associated with high intensity aerobic exercise. Recently, it has been shown
that the rise in blood glucose level following a 10-sec sprint results from a transient
decline in the rate of glucose utilisation together with the absence of changes in
endogenous rates of glucose production (Fahey et al., 2012). This is in marked contrast
to the mechanism underlying the post-exercise increase in blood glucose level following
a bout of intense aerobic exercise where the rise in blood glucose level results from a
disproportionate increase in endogenous glucose production relative to that of glucose
utilisation rate (Marliss & Vranic, 2002). Maybe the increases in plasma catecholamines
and growth hormone levels associated with short duration sprinting are too small to affect
endogenous glucose production rate, whereas the transient fall in peripheral glucose
utilisation may result from an exercise-mediated intramuscular rise in glucose 6-
phosphate levels resulting in an inhibition of glucose utilisation via a glucose 6-
phosphate-meidated inhibition of hexokinase (Fahey et al., 2013).
1.12 Glucoregulatory Response to a Single Short Sprint in Individuals with Type 1
Diabetes
Despite the large amount of research described above on the glucoregulatory responses
to a single sprint in non-diabetic individuals, there is a lack of information on the effect
of sprinting in individuals with type 1 diabetes. To the best of our knowledge, only a
handful of studies have examined the effect of sprinting on blood glucose levels in
insulin-treated individuals with type 1 diabetes (Harmer et al., 2006; Fahey et al., 2012).
One of these studies showed that a constant load sprint to exhaustion at 130% V O2peak
lasting approximately 60 to 80 sec increases blood glucose levels, with a peak change in
plasma glucose of approximately 3.8 mM after 60 min of recovery in individuals with
type 1 diabetes (Harmer et al., 2006). The performance of such a prolonged sprint in
these individuals is associated with a sharp elevation in norepinephrine (Harmer et al.,
2000, 2006), epinephrine (Harmer et al., 2000, 2006) and plasma lactate (Harmer et al.,
2000, 2006, 2008) levels immediately post-sprint, whereas immunoreactive free insulin
and glucagon levels did not significantly change during recovery (Harmer et al., 2006).
Interestingly, seven weeks of sprint-training did not prevent the rise in blood glucose
levels, but reduced its magnitude, and had little effect on catecholamine, insulin or

40
glucagon response to an identical 60 to 80 sec sprint matched for total work (Harmer et
al., 2006).
To the best of our knowledge, only one study other than those described in this thesis has
examined the effect of maximal sprint effort on blood glucose levels in individuals with
type 1 diabetes (Fahey et al., 2012). This study shows that sprinting for 10 sec in these
individuals under basal insulinaemic conditions results in a 1.2 mM increase in blood
glucose levels within 30 minutes of recovery, with blood glucose remaining at stable and
elevated levels afterwards. The mechanisms underlying this rise in blood glucose levels
differ markedly from those associated with prolonged high intensity aerobic exercise
(Marliss and Vranic, 2002). Indeed, the rise in blood glucose level in response to
sprinting results from a transient decline in the rate of glucose utilisation, with no change
in endogenous rate of glucose production (Fahey et al., 2012). This is markedly different
from intense aerobic exercise where the post-exercise rise in blood glucose level in type
1 diabetic individuals results from a disproportionate increase in endogenous glucose
production relative to the increase in glucose utilisation rate (Marliss and Vranic, 2002).
Since the levels of plasma epinephrine, norepinephrine and growth hormone rose
transiently and to only a small extent post-sprinting compared to intense aerobic exercise,
this suggests they reached levels insufficient to stimulate hepatic glucose production.
Interestingly, the effects of such a short sprint on blood glucose levels and
counterregulatory hormones are not affected by an episode of antecedent hypoglycaemia
(Davey et al., 2014).
1.13 Statement of the Problem and Aims
Individuals with Type 1 diabetes are encouraged to participate in regular physical activity
(Zinman et al., 2004) due to the numerous physiological and psychological health benefits
associated with a physically active lifestyle (Norris et al., 1990; Moy et al., 1993;
Laaksonen et al., 2000; Riddell and Iscoe, 2006; Chimen et al., 2012). Unfortunately,
exercise increases the risk of hypoglycaemia due, in part, to a contraction-mediated
activation of glucose utilisation rate by skeletal muscle (Peirce, 1999) and an increase in
insulin sensitivity (Wasserman and Zinman, 1994). This increased risk occurs both
during exercise (Riddell et al., 1999; Tuominen et al., 1995; Rabasa-Lhoret et al., 2001),
and for several hours during recovery (MacDonald, 1987a; Tsalikian et al., 2005;
McMahon et al., 2007; Maran et al., 2010; Iscoe and Riddell, 2011; Davey et al., 2013a).

41
This is of concern because severe hypoglycaemia can cause brain damage (Suh et al.,
2007; Puente et al., 2010), cognitive dysfunction (Northam et al., 2009; Asvold et al.,
2010) and even sudden death (Tanenberg et al., 2010). Approximately 6-10% of patients
with type 1 diabetes die from hypoglycaemia (Skrivarhau et al., 2006; Jacobson et al.,
2007; Feltbower et al., 2008). Consequently, it is not surprising that one of the biggest
barriers to regular physical activity in individuals with type 1 diabetes is the fear of
hypoglycaemia (Brazeau and et al., 2008). As a result of this legitimate fear, many
individuals with type 1 diabetes have been known to avoid physical activity (Ludvigsson
et al., 1980; Guelfi et al., 2007a; Brazeau et al., 2008), with ~60-65% of individuals with
type 1 diabetes reported to be inactive (Thomas et al., 2004; Plotnikoff et al., 2006).
It is important to remember, however, that not all types of exercise result in elevated risk
of hypoglycaemia. In fact, although exercise of low to moderate intensity increases the
risk of hypoglycaemia both during and after exercise (MacDonald, 1987a; Tuominen et
al., 1995; Riddell et al., 1999; Rabasa-Lhoret et al., 2001), exercise performed at high
intensity (>80% of V O2max) for approximately 10-15 min is accompanied by an increase
in glycaemia during and after exercise (Mitchell et al., 1988; Marliss et al., 1992a; Purdon
et al., 1993; Sigal et al., 1994c, 1999; Marliss and Vranic, 2002). The hyperglycaemic
effect of prolonged high intensity exercise in individuals with Type 1 diabetes raises the
intriguing possibility that this type of exercise might be beneficial if adopted to counter a
fall in glycaemia in complication-free individuals with type 1 diabetes, and thus might
help to prevent or delay hypoglycaemia if no carbohydrate is readily available. The
problem here is that 10-15 min of exercise at intensities above 80% V O2max would be
unlikely to be well tolerated by most individuals with type 1 diabetes due to the very
intense nature of such exercise combined with the impractical duration of the exercise
bout. This raises the question of whether a much shorter bout of exercise performed at a
higher intensity could be adopted to prevent glycaemia from falling. Although, as
discussed above, a 30-90-sec sprint can increase blood glucose levels in individuals with
type 1 diabetes, sprint efforts lasting 30-sec or more are associated with physical
discomfort (Marquardt et al., 1993; Laurent et al., 2007), undesirable physiological
consequences such as nausea, vomiting and dizziness (Inbar et al., 1996; 1998; Laurent
et al., 2007) and therefore may be unsafe or impractical for many individuals (Little et
al., 2010). However, a maximal sprint effort lasting only 10 sec is known to be well
tolerated. For this reason, it was the primary goal of this thesis to determine for the first
time whether a 10-sec maximal sprint effort provides a potential means other than

42
carbohydrate intake to oppose an insulin- or exercise- mediated fall in glycaemia, thus
decreasing the risk of hypoglycaemia in complication-free individuals with type 1
diabetes. Also, given that studies concerned with investigating response of individuals
with type 1 diabetes to exercise typically determine the V O2max of their participants using
graded exercise testing, another objective of this thesis is to determine whether risk of
hypoglycaemia is increased with this type of exercise protocol.
More specifically, our aims are to test the following hypotheses;
1) Graded exercise testing in complication-free individuals with type 1 diabetes will
result in a post-exercise increase in blood glucose level, therefore not increasing
the risk of hypoglycaemia during early recovery.
2) The performance of a single 10-sec maximal sprint effort after 20 min of moderate
intensity exercise performed under hyperinsulinaemic conditions will oppose the
exercise-mediated fall in glycaemia, thus decreasing the risk of hypoglycaemia
during early recovery in complication-free individuals with type 1 diabetes.
3) The performance of a single 10-sec maximal sprint effort before 20 min of
moderate intensity exercise performed under hyperinsulinaemic conditions will
lessen the subsequent exercise-mediated fall in glycaemia, thus helping to
decrease the risk of hypoglycaemia during exercise in complication-free
individuals with type 1 diabetes.
4) The performance of a single 10-sec maximal sprint under hyperinsulinaemic
conditions will result in a counterregulatory response typical of sprinting.

43
1.14 Significance of the Thesis
Unfortunately, individuals with type 1 diabetes are on average less physically active than
recommended due to their fear of hypoglycaemia and a lack of information and guidelines
on how to safely participate in various sports and recreational activities. It is important
for these individuals to enjoy the numerous health and lifestyle benefits of regular
physical exercise. Regrettably, there is little research in this area, resulting in a lack of
comprehensive guidelines and practical advice on how to safely manage blood glucose
levels while performing various types of physical activity. Our findings might identify
sprinting as a novel tool for the prevention of hypoglycaemia in individuals with type 1
diabetes.

44
Chapter 2
Glycaemic Response to Graded Exercise in
Individuals with Type 1 Diabetes

45
2.1 Abstract
Objective: To investigate whether the risk of hypoglycaemia in individuals with type 1
diabetes increases in response to the type of graded exercise testing used to determine
V O2peak
Research Design and Methods: Eight non-diabetic male participants (CON) and seven
complication-free type 1 diabetic male individuals in good to moderate glycaemic control
(HbA1c = 7.6 ± 0.5%) were recruited for this study. On the morning of testing, the type
1 diabetic participants followed their normal insulin regimen, and both groups ate their
usual breakfast to transiently increase their blood glucose levels. Approximately four
hours later, graded exercise was commenced on a cycle ergometer. Blood metabolites
and hormones were sampled at rest and regularly during recovery.
Results: During graded exercise, there were no significant changes in glycaemia in either
group, but blood glucose levels in type 1 diabetic participants increased by more than 2
mM during recovery (p < 0.05) and remained elevated for 120 min, while remaining at
stable levels in the control group. In both groups, catecholamine and lactate levels
increased significantly early in recovery and fell thereafter (p < 0.05). Glucagon and
cortisol levels also increased during recovery in both groups, while free fatty acid levels
increased only in the type 1 diabetic participants (p < 0.05). In contrast, the levels of
insulin remained stable in both groups (p > 0.05).
Conclusions: Our graded exercise protocol performed in the post-absorptive state while
plasma insulin levels are low increases glycaemia early post-exercise, with no
carbohydrate ingestion required for hypoglycaemia prevention early during that time.
This type of exercise, therefore, does not increase the risk of early post-exercise
hypoglycaemia in individuals with type 1 diabetes. The mechanisms underlying the post-
exercise increase in glycaemia remain to be determined.

46
2.2 Introduction
Regular participation in physical activity is an important component in type 1 diabetes
management because it is associated with numerous physiological and psychological
health benefits (Norris et al., 1990; Moy et al., 1993; Laaksonen et al., 2000; Steppel and
Horton, 2004; Zinman et al., 2004; Riddell and Iscoe, 2006; Chimen et al., 2012, Tonoli
et al., 2012). Unfortunately, an active lifestyle may make acute management of blood
glucose levels more difficult (Steppel and Horton, 2004; Camacho et al., 2005; Riddell
and Perkins, 2006; Guelfi et al., 2007c; Younk and Davis, 2012). For instance, exercise
of low to moderate intensity increases the risk of hypoglycaemia both during and after
exercise (Camacho et al., 2005; Guelfi et al., 2007c) due, in part, to a contraction-
mediated activation of glucose utilisation rate by skeletal muscle and an increase in
insulin sensitivity (Steppel and Horton, 2004; Camacho et al., 2005). In contrast, exercise
performed at high intensity increases blood glucose levels (Marliss and Vranic, 2002).
Given the numerous benefits of an active lifestyle for individuals with type 1 diabetes
together with the difficulties and challenges associated with managing blood glucose
levels during and after exercise, it is not surprising that these issues have attracted a large
volume of research (Chapter 1).
One feature shared by most of the research on exercise in diabetes is the common practice
of standardising exercise intensity relative to maximal or peak rate of oxygen
consumption ( V O2 max or V O2peak) or lactate threshold (Chapters 3, 4, and 5 of this thesis;
Fremion et al., 1987; Wanke et al., 1992; Purdon et al., 1993; Sigal et al., 1994c, 1996;
Nugent et al., 1997; Ford et al., 1999b; Galassetti et al., 2001b; Rabasa-Lhoret et al., 2001;
Galassetti et al., 2003; Heyman et al., 2007). The determination of V O2peak or lactate
threshold requires the performance of a graded exercise test whereby exercise intensity
increases in a stepwise fashion or continuously until maximal intensity is achieved
(volitional exhaustion or attainment of criteria for V O2max/peak).
Despite the importance and prevalence of graded exercise testing for V O2/peak
determination, it is unclear whether this testing protocol increases the risk of
hypoglycaemia in type 1 diabetic individuals, and whether insulin reduction as well as
increasing food intake before or immediately after exercise is required to prevent blood
glucose levels from falling post-exercise. All studies addressing this issue have performed
blood glucose measurements before and immediately after V O2peak testing (Fremion et

47
al., 1987; Wanke et al., 1992; Nugent et al., 1997; Ford et al., 1999; Heyman et al., 2007),
with some reporting either the absence of changes in blood glucose levels (Fremion et al.,
1987; Wanke et al., 1992; Nugent et al., 1997; Ford et al., 1999) or a decrease (Heyman
et al., 2007). Since there are conditions where blood glucose level can change markedly
after exercise (Purdon et al., 1993; Sigal et al., 1996; Marliss and Vranic, 2002) it is
important to assess whether the risk of hypoglycaemia increases during recovery from
graded exercise testing. To the best of our knowledge, only one study has examined this
issue, but found no significant changes in blood glucose levels 20 min post-exercise (Ford
et al., 1999b). However, as pointed out by the authors themselves, the main limitation
with their study is that insulin dosage was adjusted and food was ingested one hour prior
to exercise (Ford et al., 1999b), thus leaving unanswered the important question of
whether blood glucose levels would have fallen during recovery had no food been
ingested.
Given that a short bout of intense aerobic exercise under basal insulinaemic conditions
can increase blood glucose levels (Marliss and Vranic, 2002), this raises the possibility
that graded exercise testing performed when plasma insulin is at near basal level might
also be associated with a post-exercise increase in glycaemia, with no carbohydrate
ingestion required prior to or after exercise to prevent hypoglycaemia. The primary
objective of this study is to test this hypothesis by examining the glycaemic and hormonal
response to the type of graded exercise test used for V O2peak determination under
conditions where food is not ingested soon prior to testing and without adjusting insulin
dosage. This is an important issue to address given the widespread use of graded exercise
testing in diabetes research and fitness testing, and the absence of information about the
risk of hypoglycaemia associated with this exercise protocol.

48
2.3 Research Design and Methods
2.3.1 Participants
Seven young males with type 1 diabetes (aged 20.4 ± 1.5 years; BMI 27.4 ± 1.0 kg/m2;
peak 2OV 45.9 ± 2.4 ml/kg/min; duration of diabetes 5.8 ± 1.8 years) and eight age-matched
non-diabetic male (CON; aged 20.1 ± 0.8 years; BMI 25.3 ± 1.5 kg/m2; peak 2OV 44.6 ±
3.6 ml/kg/min) participants were recruited from Princess Margaret Hospital and the
University of Western Australia, respectively. All participants with type 1 diabetes were
in moderate glycaemic control (HbA1c = 7.6 ± 0.5%), free from diabetic complications,
hypoglycaemia aware, had undetectable levels of C-peptide and were not taking any
prescribed medication other than insulin. Participants with type 1 diabetes were on a
multiple-injection insulin regime that had not changed for at least three months prior to
testing. Following a familiarisation session during which their informed consent was
obtained together with anthropometric measurements, participants were required to attend
our laboratory on another occasion for testing. Both the University of Western Australia
and the Princess Margaret Hospital Ethics Committees approved all the procedures
described in this study.
2.3.2 Experimental trials and assays
Participants were not allowed to exercise for 48 h prior to the experimental trial since
antecedent exercise has the potential to affect the endocrine response to a subsequent bout
of exercise (Galassetti et al., 2001b). Also, testing was rescheduled if they had
experienced a hypoglycaemic episode over the previous 48 h because prior
hypoglycaemia can also affect the counter-regulatory response to exercise (Galassetti et
al., 2003). Each participant was also required to maintain a normal diet and to avoid
alcohol for 24 h prior to testing. On the morning of testing, participants were instructed
to monitor their blood glucose regularly. They arrived in the laboratory at ~ 7:30 am, and
all type 1 diabetic participants were instructed to self-administer their usual dose of
morning insulin into their abdomen. Both the control and type 1 diabetes groups were
then asked to consume a breakfast, with the meal choice and nutritional content reflecting
that normally ingested by the participant.

49
After breakfast, a catheter was inserted in one of the superficial veins at the back of the
subject’s hand prior to heating the hand for the sampling of arterialised venous blood. No
physical activity was allowed and blood glucose levels were measured every 15 min. In
an effort to comply with the recommendations that exercise should be avoided if insulin
levels are peaking, commencement of testing was delayed until four hours after insulin
injection, at a time when insulin was near basal levels. During that time, post-prandial
blood glucose levels fell, and when glycaemia approached 7 mM (7.24 ± 0.45 mM in the
type 1 diabetic participants; 5.27 ± 0.20 mM CON), the graded exercise test to determine
V O2peak was initiated on an Evolution cycle ergometer (Evolution, Geelong, Australia).
The graded exercise test described here required participants to initiate cycling at an
intensity of 50 watts while breathing through a mouthpiece into an oxygen analyser
system. Briefly, every 3 min the exercise intensity was raised progressively by 50 watts
until the subject reached his maximal rate of oxygen consumption. A plateau in oxygen
consumption (an increase of < 150 mL·min-1) and/or a respiratory exchange ratio greater
than 1.15 during the last min of exercise were the criteria for the achievement of V O2peak.
The participant’s expired air was monitored continuously, and both oxygen uptake and
carbon dioxide production were calculated every 15 sec using a computerized on-line gas
analysis system. This comprised of a Morgan Ventilation Monitor (Morgan, Reinham,
Kent, U.K.), Ametek S3A Oxygen Analyser and Ametek CD3A Carbon Dioxide
Analyser (Ametek, Paoli, PA).
Blood was sampled prior to exercise and then at 0, 5, 10, 15, 30, 60, 90 and 120 min post-
exercise. Some of the blood was assayed immediately for glucose, pO2, pH and lactate
levels using an ABL 625 blood gas system (Radiometer, Copenhagen) and the remainder
of the blood was combined with sodium metabisulphite, polyethylene glycol or aprotinin
(Trasylol) for the assays of catecholamines, insulin and glucagon, respectively (Bussau
et al., 2006). All samples were centrifuged at 720g for 5 min and the plasma stored at -
80C for later analyses of catecholamines, free fatty acids, insulin, glucagon, cortisol, GH
and C-peptide levels.
Heparinized plasma treated with sodium metabisulphate was used for the determination
of catecholamine levels by reverse phase high-performance liquid chromatography using
a Waters Novapak C18 reverse phase column and a model 5200A Coulochem detector
(ESA Biosciences Inc., Chelmsford, MA, USA). Free fatty acids levels were measured

50
in EDTA-treated plasma using the Roche Half Micro Test Free Fatty Acids Assay Kit
(Roche Diagnostic, Mannheim, Germany). Heparinized plasma treated with polyethylene
glycol was assayed for free insulin using the Coat-a-Count Insulin Kit (Diagnostic
Products Corporation, Los Angeles, CA, USA). Glucagon levels in plasma collected with
trasylol were measured from EDTA-treated plasma by radioimmunoassay using a Linco
Glucagon RIA Kit (Linco Research, St Charles, Missouri, USA). Cortisol levels were
assayed from venous serum by competitive immunoassay on an Immulite 2000 Analyser
using the Immulite Cortisol Assay Kit (Diagnostic Products Corporation, Los Angeles,
CA, USA). Growth hormone levels were determined from serum by immunometric assay
on an Immulite 2000 Analyser using the Immulite Growth Hormone Assay Kit
(Diagnostic Products Corporation, Los Angeles, CA, USA). Finally, C-peptide levels
were determined by solid-phase competitive chemiluminescent enzyme immunoassay on
an Immulite 2000 analyser using the Immulite C-peptide Assay Kit (Diagnostic Products
Corporation, Los Angeles, CA, USA).
2.3.3 Statistical analyses
The results within each experimental group were analysed using a mixed model repeated
measure analysis of variance (ANOVA) and Fisher’s least significant differences test for
a posteriori analysis using SPSS 13.0 for Windows software (SPSS, Chicago, IL, USA).
Statistical significance was accepted at p < 0.05. Participants’ characteristics are
expressed as means ± S.D. whereas all other results are expressed as means ± S.E.M.

51
2.4 Results
2.4.1 Blood metabolite response to graded exercise
Before the graded exercise test, blood glucose levels in both experimental groups fell
significantly (p < 0.05) (Fig. 2.1). When glycaemia reached ~7 mM in diabetic
participants (7.24 ± 0.45 mM in type 1 diabetic participants; 5.27 ± 0.20 mM control
participants), the test was initiated. In response to graded exercise (exercise duration ~
12 min), there were no significant changes in glycaemia in either experimental group.
Immediately following exercise, glycaemia increased significantly (p < 0.05) and
remained elevated for the whole duration of the 120-min recovery period (p < 0.05) in the
type 1 diabetic participants (Fig. 2.1). In contrast, blood glucose levels remained
relatively stable in the control group (Fig. 2.1). Blood lactate levels reached maximal
levels at 5 min post-exercise in both experimental groups and decreased throughout
recovery (Fig. 2.1). In contrast, blood pH reached minimal levels after 5 min of recovery
in both experimental groups before returning to pre-exercise levels within 60 min of
recovery (Fig. 2.1). During recovery, free fatty acid levels increased gradually over time
and reached maximal levels after 90-120 min of recovery (Fig. 2.1).
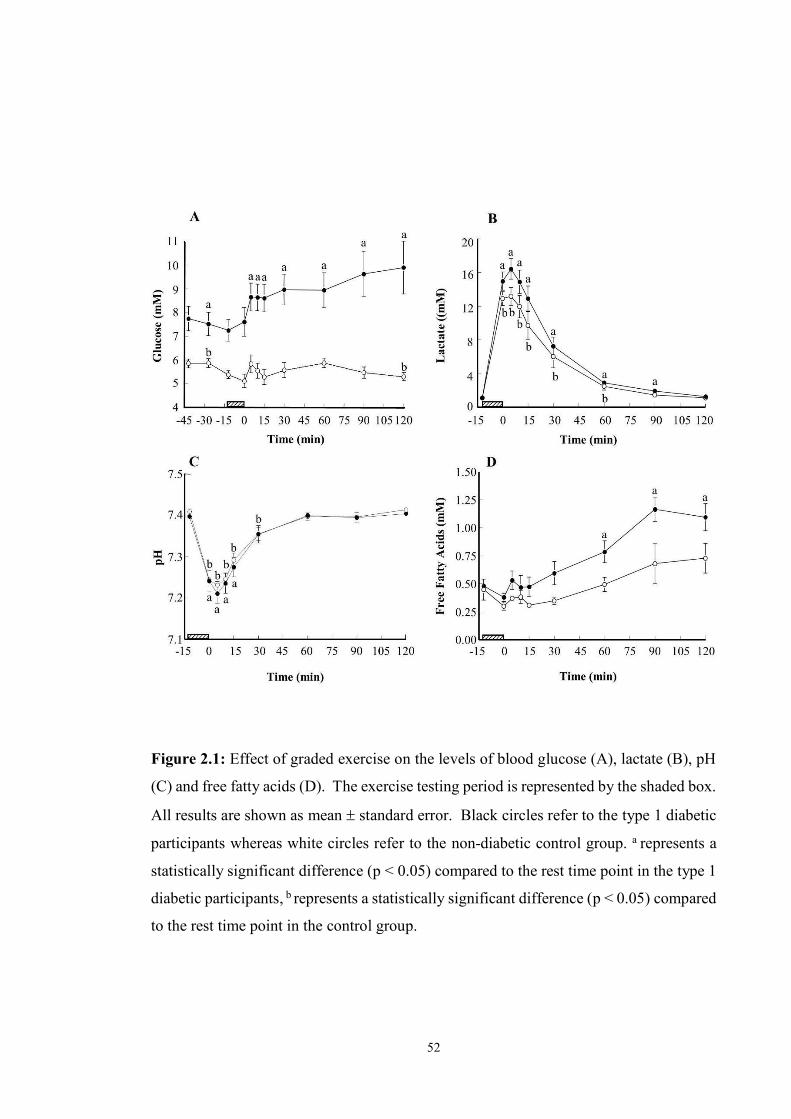
52
Figure 2.1: Effect of graded exercise on the levels of blood glucose (A), lactate (B), pH
(C) and free fatty acids (D). The exercise testing period is represented by the shaded box.
All results are shown as mean standard error. Black circles refer to the type 1 diabetic
participants whereas white circles refer to the non-diabetic control group. a represents a
statistically significant difference (p < 0.05) compared to the rest time point in the type 1
diabetic participants, b represents a statistically significant difference (p < 0.05) compared
to the rest time point in the control group.

53
2.4.2 Hormonal response to graded exercise
In response to graded exercise, catecholamines increased to maximal levels at the onset
of recovery in both experimental groups (p < 0.05; Fig. 2.2), with the levels attained being
not significantly different between experimental groups (p > 0.05). Norepinephrine levels
returned to pre-exercise levels within 15 and 30 min after exercise in the control and type
1 diabetic groups, respectively (Fig. 2.2), whereas epinephrine returned to pre-exercise
levels within 5 min post-exercise in both experimental groups (Fig. 2.2). Growth
hormone (GH) levels displayed a non-significant transient increase in the type 1 diabetic
and control groups, and reached significantly elevated levels in the control group after 15
min of recovery (p < 0.05; Fig. 2.2). Following graded exercise, plasma cortisol levels
increased significantly (p < 0.05) and reached maximal levels within 10-30 min post-
exercise in both groups (Fig. 2.2). Likewise, glucagon levels increased significantly in
the type 1 diabetic group by 30 min post-exercise and by 90 min in the control group (p
< 0.05; Fig. 2.2). During exercise, plasma insulin levels remained at relatively stable low
levels in both experimental groups (Fig. 2.2).
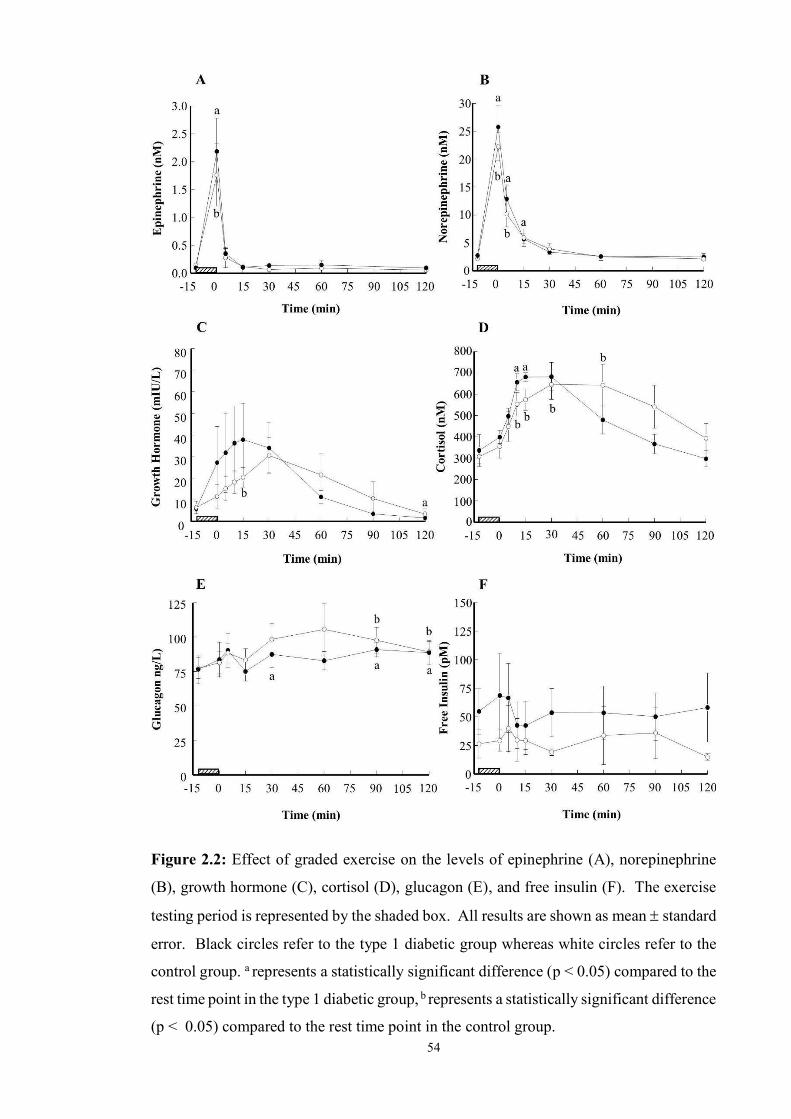
54
Figure 2.2: Effect of graded exercise on the levels of epinephrine (A), norepinephrine
(B), growth hormone (C), cortisol (D), glucagon (E), and free insulin (F). The exercise
testing period is represented by the shaded box. All results are shown as mean standard
error. Black circles refer to the type 1 diabetic group whereas white circles refer to the
control group. a represents a statistically significant difference (p < 0.05) compared to the
rest time point in the type 1 diabetic group, b represents a statistically significant difference
(p < 0.05) compared to the rest time point in the control group.

55
2.5 Discussion
Graded exercise testing is integral to V O2max/peak and lactate threshold determination in
basic, applied and clinical research. What is still unclear, however, is whether the risk of
hypoglycaemia increases in response to this type of exercise in individuals with type 1
diabetes. This is an important issue given that most laboratory-based studies on exercise
in diabetes involve the performance of graded exercise for V O2max/peak determination.
Here we show for the first time that graded exercise performed when plasma insulin is at
near basal level in young adult males with type 1 diabetes increases blood glucose level
post-exercise and thus does not increase the risk of early hypoglycaemia. In particular,
graded exercise under these conditions results in a rapid post-exercise increase in blood
glucose levels (> 2 mM) which remains elevated for the first two hours of recovery (Fig.
2.1), thus suggesting that no carbohydrate ingestion before or after this type of exercise
is required to prevent hypoglycaemia. Our findings thus suggest this exercise protocol is
safe provided it is performed some time after the injection of rapid acting insulin.
The absence of a significant fall in glycaemia observed here during graded exercise
corroborates the findings of most (Fremion et al., 1987; Wanke et al., 1992; Nugent et al.,
1997; Ford et al., 1999), but not all studies (Heyman et al., 2007) on graded exercise in
individuals with type 1 diabetes, whereas the post-exercise increase in blood glucose
levels is described here for the first time. Although differences in experimental design
make it difficult to compare our findings with those of others, most earlier studies reported
that graded exercise does not affect blood glucose levels, but participants were fasted and
omitted their morning insulin to reduce the risk of exercise-induced hypoglycaemia in
one of these studies (Nugent et al., 1997) or were fed with excess of food before exercise
with (Ford et al., 1999) or without (Fremion et al., 1987) adjusting their insulin dosage
prior to exercise, thus making it less likely for blood glucose levels to fall. Although one
study reported a fall in glycaemia (Heyman et al., 2007) during V O2peak testing, exercise
in that study was performed when plasma insulin levels were near maximal, a time when
exercise is usually not recommended for individuals with type 1 diabetes (Rabasa-Lhoret
et al., 2001; Steppel and Horton, 2004).
The finding here that blood glucose levels increase during recovery from graded exercise
has never been reported before. To the best of our knowledge, only one other study has
examined how blood glucose levels change during recovery from graded exercise testing

56
in individuals with type 1 diabetes, but it only measured glycaemia at 0 and 20 min post-
exercise and found no significant change in blood glucose levels (Ford et al., 1999b).
However, the main limitation with that study is that insulin was adjusted and food was
ingested one hour prior to exercise, thus limiting the relevance of these findings and
leaving unanswered the question of whether graded exercise increases the risk of early
hypoglycaemia post-exercise if insulin dosage is not reduced or if no carbohydrate is
ingested soon before exercise (Ford et al., 1999b). Our findings are consistent with those
examining blood glucose response to intense aerobic exercise (>80% of V O2peak)
performed under basal insulinaemia. Indeed, 10-15 min of sustained high intensity
exercise at near 80% of V O2peak in individuals with type 1 diabetes results in a sustained
post-exercise increase in blood glucose levels (Purdon et al., 1993; Sigal et al., 1994c;
Marliss and Vranic, 2002) similar to that observed here where blood glucose levels
increase and remain elevated or continue to rise well after the cessation of exercise
(Purdon et al., 1993; Sigal et al., 1994c; Marliss and Vranic, 2002). Similarly, the
relatively stable blood glucose levels during graded exercise in the control group (Fig.
2.1) were similar to those reported in previous studies (Nugent et al., 1997; Heyman et
al., 2007).
The marked rise in epinephrine and norepinephrine levels immediately following graded
exercise might explain, at least in part, the rapid increase in glycaemia during early
recovery in the type 1 diabetic group. In support of this view, the increased catecholamine
levels during that time coincided with the rapid initial rise in blood glucose levels, with
epinephrine and norepinephrine levels increasing by ~10-fold in the type 1 diabetic group
(Fig. 2.2). A comparable increase in catecholamines levels (14-18-fold) has also been
reported to occur in response to sustained high intensity aerobic exercise in individuals
with type 1 diabetes (Purdon et al., 1993; Marliss and Vranic, 2002). It is important to
note, however, that others have reported that catecholamines increase by less than 5-fold
immediately following graded exercise (Nugent et al., 1997). Although elevated
catecholamine levels are acknowledged to oppose insulin-mediated fall in glycaemia
(Marliss and Vranic, 2002) via activation of hepatic glucose production and inhibition of
insulin-mediated glucose uptake in skeletal muscles (Marliss and Vranic, 2002;
Nonogaki, 2000), it is important to note that the importance of catecholamines in the
activation of hepatic glucose production in response to intense exercise has been the
object of some dispute (Camacho et al., 2005; Coker and Kjaer, 2005). Also, and more
importantly, the fact that blood glucose levels remained stable in the control group despite

57
experiencing a comparable rise in catecholamine levels and no changes in plasma insulin
levels in response to graded exercise suggests that other mechanisms both mediate the
post-exercise rise in glycaemia in type 1 diabetic participants and explain the different
glycaemic responses between the two experimental groups.
During recovery from graded exercise in the type 1 diabetic group, the absence of a post-
exercise increase in insulin levels (Fig. 2.2) probably explains, at least in part, the early
post-exercise rise in blood glucose level and sustained increased glycaemia as is the case
following sustained high intensity aerobic exercise in type 1 diabetic individuals (Purdon
et al., 1993; Sigal et al., 1994c; Marliss and Vranic, 2002). Here, however, not only
plasma insulin levels did not differ between treatment groups, there was no increase in
plasma insulin post-graded exercise test in the control group. This is consistent with the
absence of post-exercise increase in blood glucose level in the control group. However,
our findings with the control group differ from the transient increase in plasma insulin
levels that occurs in non-diabetic individuals recovering from intense aerobic exercise
and which have been proposed to explain their lesser post-exercise rise in glycaemia
compared to diabetic individuals (Sigal et al., 1994c; Marliss and Vranic, 2002). Our
findings thus suggest that other mechanisms are likely to explain the different patterns of
glucose response between the control and type 1 diabetic participants.
Glucagon and cortisol are unlikely to play a major role in increasing glycaemia during
recovery from graded exercise testing in individuals with type 1 diabetes. The levels of
plasma glucagon, a potent activator of hepatic gluconeogenesis (Marliss and Vranic,
2002; Camacho et al., 2005), did not increase significantly during early recovery in type
1 diabetes or control participants although they did rise later during recovery (Fig. 2.2).
This pattern of response in control participants differs from the increase in glucagon
observed in response to sustained high intensity aerobic exercise (Purdon et al., 1993;
Sigal et al., 1994c). Assuming that portal glucagon and insulin levels follow patterns of
change similar to those of peripheral blood glucose and insulin levels, this suggests that
both glucagon and glucagon/insulin ratio play little role in mediating the post-exercise
rise in blood glucose levels. Although there is some published evidence that a rise in
cortisol levels may play a role in stabilising glycaemia due to its potential acute inhibitory
effect on glucose utilisation in skeletal muscles (Shamoon et al., 1980), this hormone is
unlikely to play a major role here firstly because the effects of cortisol on hepatic glucose
production and blood glucose levels require several hours to take place (McMahon et al.,

58
1988), and secondly because the increase in glycaemia observed here in type 1 diabetic
participants preceded that of cortisol.
Growth hormone is also unlikely to play a major role in the increase in glycaemia during
recovery from graded exercise as suggested by the absence of significant changes in the
levels of this hormone in type 1 diabetic individuals (Fig. 2.2). It is important to stress,
however, that a close inspection of Fig. 3c suggests that significant changes in growth
hormone levels in response to graded exercise might have been undetected due to the
large inter-individual variability in growth hormone levels. A trend for not only an
increase in growth hormone levels in both experimental groups, but also a more
pronounced early rise in growth hormone levels in the diabetic compared to non-diabetic
group is consistent with the findings of others that graded exercise test results in a lesser
post-exercise increase in growth hormone levels in non-diabetic compared to diabetic
participants (Coiro et al., 2004). These responses of growth hormone to graded exercise
suggest that these hormones could mediate the early increase in glycaemia post-graded
exercise. In support of this interpretation, the administration of a physiological growth
hormone pulse in non-exercised non-diabetic individuals results in a rapid fall in muscle
glucose uptake (Moller et al., 1990). However, even if the increase in growth hormone
levels had been significant, it would have probably played a role of lesser importance in
increasing glycaemia post-exercise in the type 1 diabetic group. This is because the
administration of a growth hormone pulse has been shown not to affect peripheral glucose
uptake in insulin-treated individuals with type 1 diabetes (Møller et al., 1992a) and also
because the infusion of octreotide, an inhibitor of growth hormone release, has no acute
effect on the magnitude of the hyperglycaemic effect of high intensity exercise (Sigal et
al., 1996).
Finally, the elevated lactate level after graded exercise (Fig. 2.1) is another factor that
may have contributed to the stabilisation of glycaemia early in recovery by providing
gluconeogenic precursors for hepatic glucose production (Miller et al., 2002) and by
increasing peripheral insulin resistance (Vettor et al., 1997). However, the fact that
lactate reached similar levels post-exercise in the type 1 diabetic and control participants
suggests that this factor alone is unlikely to explain the post-exercise increase in
glycaemia in type 1 diabetes since no change in glycaemia took place in the control
participants despite both groups experiencing a similar increase in plasma lactate levels.

59
Overall, despite the potential role for catecholamines and maybe growth hormone and
lactate in mediating, at least in part, the increase in glycaemia during recovery in the type
1 diabetic group, the fact that the patterns of change in plasma insulin and counter-
regulatory hormones were similar between the type 1 diabetic and control groups suggest
that other factors explain both the different blood glucose responses between these groups
and the absence of increase in blood glucose levels in the control group during recovery
from graded exercise. The counterregulatory hormone(s) responsible for increasing
glycaemia post-exercise in type 1 diabetes thus remain to be identified.
In conclusion, on clinical grounds, our findings suggest that the risks of hypoglycaemia
associated with graded exercise testing are minimal when performed under basal/near
basal insulin levels, with no carbohydrate administration required just before or early after
testing to prevent hypoglycaemia in individuals with type 1 diabetes. This is because this
type of exercise protocol increases glycaemia during recovery if performed when insulin
levels are not elevated. However, the risk of late onset post-exercise hypoglycaemia
during recovery from graded exercise testing needs to be investigated in future studies.
In addition, it is our view that health professionals should still regularly monitor
glycaemia before, during and after testing since it remains to be established whether
similar findings would have been reported with different graded exercise protocols and
individuals of different ages, circulating insulin levels or severity of diabetic
complications.
2.6 Acknowledgements
This research was funded by a Juvenile Diabetes Research Foundation/ National Health
Medical Research Council of Australia program grant to T.W. Jones and P.A. Fournier.
L.D. Ferreira is supported by a Juvenile Diabetes Research Fund International
Fellowship.

60
Chapter 3
A 10-second Maximal Sprint Effort: A Novel
Approach to Counter an Exercise-Mediated
Fall in Glycaemia in Individuals with Type 1
Diabetes
An amended version of this chapter has been published in Diabetes Care:
Bussau, V.A., Ferreira, L.D., Jones, T.W. & Fournier, P.A. (2006). The 10-s Maximal
Sprint: A Novel Approach to Counter an Exercise-Mediated Fall in Glycemia in
Individuals with Type 1 Diabetes. Diabetes Care 29 (3): 601-606.

61
3.1 Abstract
Objective: Our aim is to investigate whether a short maximal sprint effort can provide
another means to counter the rapid fall in glycaemia associated with moderate intensity
exercise in individuals with type 1 diabetes, and therefore decrease the risk of early post-
exercise hypoglycaemia.
Research Design and Methods: Seven healthy male participants with type 1 diabetes
injected their normal insulin dose and ate their usual breakfast. When their postprandial
glycaemia fell to ~11 mM, they pedalled at 40% peak 2OV for 20 min on a cycle ergometer
then immediately engaged in a maximal 10-sec cycling sprint (sprint trial) or rested
(control trial). Both trials were administered in a counterbalanced order.
Results: Moderate intensity exercise resulted in a significant fall (p < 0.05) in glycaemia
in both trials (mean ± S.E.M.: 3.6 ± 0.5 vs 3.1 ± 0.5 mM for sprint and control,
respectively). The subsequent short sprint opposed a further fall in glycaemia for 120 min,
whereas in the absence of a sprint, glycaemia decreased further (3.6 ± 1.22 mM; p < 0.05)
after exercise. The stabilisation of glycaemia in the sprint trial was associated with
elevated levels of catecholamines, growth hormone, and cortisol. In contrast, these
hormones remained at stable or near stable levels in the control trial. Changes in insulin
and free fatty acid levels were similar in the sprint and control trials.
Conclusions: These results suggest that following moderate intensity exercise, it is
preferable for young individuals with insulin-treated, complication-free type 1 diabetes
to engage in a 10-sec maximal sprint effort to acutely oppose a further fall in glycaemia
than to only rest, thus providing another means to reduce risk of hypoglycaemia in active
individuals with type 1 diabetes.

62
3.2 Introduction
It is well established that exercise of moderate intensity increases the risk of
hypoglycaemia both during and after exercise in individuals with type 1 diabetes
(Campaigne et al., 1987; Riddell et al., 1999) due, in part, to a contraction-mediated
activation of glucose utilisation rate in skeletal muscle (Peirce, 1999) and an increase in
insulin sensitivity (Wasserman and Zinman, 1994). In contrast, 10-15 min of high
intensity exercise (>80% of maximal rate of oxygen consumption [ peak 2OV ]) causes a
post-exercise increase in blood glucose levels in insulin-treated individuals with type 1
diabetes, irrespective of their glycaemic control (Mitchell et al., 1988; Marliss et al.,
1992a; Purdon et al., 1993; Sigal et al., 1994c, 1999; Marliss and Vranic, 2002). This
hyperglycaemic effect of prolonged high intensity exercise raises the intriguing
possibility that this type of exercise might provide a means other than carbohydrate intake
to counter a fall in glycaemia post-exercise in complication-free individuals with type 1
diabetes, and thus acutely reduce their risk of hypoglycaemia. Unfortunately, 10-15 min
of exercise at high intensity is unlikely to be well tolerated by most individuals with type
1 diabetes due to the very intense nature of such exercise combined with its impractical
duration.
A more practical way of using intense exercise as a means to prevent glycaemia from
falling might be to engage in a much shorter bout of exercise performed at maximal
intensity. The main difficulty with this suggestion is that a maximal sprint effort that lasts
in excess of 30-sec is associated with unpleasant consequences such as nausea, vomiting
and dizziness (Inbar et al., 1996; Van Praagh, 1998). This raises the question of whether
a shorter sprint could provide a potential means other than carbohydrate intake to oppose
a post-exercise fall in glycaemia. Although individuals at rest are unlikely to engage in a
sprint to stabilise their glycaemia, doing so might prove to be an effective and convenient
way to counteract a rapid fall in blood glucose level in response to moderate intensity
exercise. Because this possibility has never been examined before, it was the primary goal
of this study to determine whether a 10-sec maximal sprint effort is preferable to only
resting as a means to counter a further fall in glycaemia during recovery from moderate
intensity exercise in individuals with type 1 diabetes.

63
3.3 Research Design and Methods
3.3.1 Participants
Seven young men with type 1 diabetes were recruited to this study (aged 21.0 ± 3.5 years;
BMI 26.9 ± 4.0 kg/m2; c 44.5 ± 4.2 ml/kg/min; duration of type 1 diabetes 9.1 ± 3.6 years;
HbA1c 7.4 ± 0.8% ranging between 6.6 to 9.0 %). All participants were free from any
diabetic complications, had undetectable levels of C-peptide, and were not taking any
prescribed medication other than insulin. They had all been treated with a stable insulin
regimen composed of a combination of slow or intermediary acting insulin (eg NPH), and
fast acting insulin analogues for at least three months prior to the study. All participants
were required to attend our laboratory on three occasions. The first visit was a
familiarisation session during which their informed consent was obtained together with
anthropometric measurements, and their maximal rate of oxygen consumption was
determined as previously described (Fairchild et al., 2002b). The next two visits were the
sprint and control rest trials administered in a random counterbalanced order. The
Institutional Ethics Committee approved all the procedures described in this study.
3.3.2 Experimental trials
The participants were not permitted to exercise for 24 h prior to the experimental trial,
and testing was rescheduled if they had experienced a hypoglycaemic episode over the
previous 48 h. Participants were also instructed to maintain a similar diet and avoid
alcohol for 24 h before each testing session. On the morning of testing, the participants
were required to monitor their blood glucose regularly. They arrived in the laboratory at
around 8 A.M. and were instructed to self-administer their usual dose of morning insulin
and eat their breakfast as per normal before a catheter was inserted for blood sampling.
Participants’ insulin dose and breakfast content (1833 ± 224 kJ total energy; 57 ± 3%
carbohydrate; 17 ± 1% protein; 26 ± 4% fat) were kept identical for both trials. Glycaemia
was then measured every 15 min, and once blood glucose levels fell after peaking
postprandially, no mid-morning snack was allowed so as not to counter the fall in
glycaemia. If participants’ blood glucose levels were not decreasing at the
commencement of exercise, the testing session was cancelled because the purpose of this
study was to determine whether a short sprint could counter an exercise-mediated fall in
glycaemia.

64
When falling blood glucose levels reached ~ 11 mM (approximately 121 ± 15 and 109 ±
10 min after insulin injection in the sprint and control trials), they engage in 20 min of
moderate intensity exercise (40% peak 2OV ) on an air-braked Repco front access cycle
ergometer (Repco, Sydney, Australia), with the resistance to cycling increasing with
cycling rate. An intensity of 40% peak 2OV was adopted because it more closely mimics
the intensity of most activity patterns performed under ‘real life’ conditions for the
general population. Also, because all participants were tested at a time close to peak
plasma insulin levels and insulin-treated individuals with type 1 diabetes are discouraged
from engaging in any intense exercise during that time (Zinman et al., 2004), the
information gathered from exercising our participants at higher intensity would have been
of little practical relevance to most individuals with type 1 diabetes. The 20-min duration
was adopted because some preliminary work in our laboratory revealed that had the
exercise duration been longer, a large proportion of our participants would have reached
hypoglycaemic levels because of the rapid fall in glycaemia when exercising at 40%
peak 2OV , and testing would have had to end prematurely to avoid a hypoglycaemia-
mediated counterregulatory response. On completion of this moderate intensity cycling,
participants were instructed to rest or perform a 10-sec sprint, depending on their
experimental trial. All participants were instructed to cycle as hard as possible and not to
pace themselves for the whole duration of the 10-sec sprint. Venous blood from the arm
and arterialised capillary blood from the earlobe were sampled before the moderate
intensity exercise, immediately before the cycling sprint (in the sprint trial) and then at 0,
5, 10, 15, 30, 45, 60, 75, 90, 105 and 120 min of recovery or until blood glucose levels
declined to 3.5 mM, in which case the trial was ended and the participant was immediately
given carbohydrates to prevent hypoglycaemia.
At each sampling point, 35 l of arterialised capillary blood was taken from the earlobe
and assayed immediately for glucose and lactate levels using an ABL 625 blood gas
system (Radiometer, Copenhagen). A 15 ml blood sample was also removed from the
catheter for measuring hormones. Some of the blood was combined with sodium
metabisulphite, poly-ethylene glycol or aprotinin (Trasylol) for the assay of
catecholamines, insulin, and glucagon, respectively. All samples were then centrifuged
at 720g for 5 min, and the plasma was stored at -80C for later analysis of catecholamines,
free fatty acids, insulin, glucagon, cortisol, growth hormone and C-peptide levels.

65
3.3.3 Hormones and metabolite assays
Glucose and lactate were analysed using an ABL 625 Blood Gas System (Radiometer,
Copenhagen, Denmark). Heparinized plasma treated with sodium metabisulphate was
used to determine catecholamine levels by reverse phase high-performance liquid
chromatography using a Waters Novapak C18 reverse phase column and a model 5200A
Coulochem detector (ESA Biosciences Inc, Chelmsford, MA, USA). Free fatty acids
levels were measured in EDTA-treated plasma using the Roche Half Micro Test Free
Fatty Acids Assay kit (Roche, Mannheim, Germany). Heparinized plasma treated with
polyethylene glycol was assayed for free insulin using the Coat-a-Count Insulin kit
(Diagnostic Products Corporation, Los Angeles, CA, USA). Glucagon levels in plasma
collected with aprotinin (Trasylol) were measured from EDTA-treated plasma by
radioimmunoassay using a Linco glucagon radioimmunoassay kit (Linco Research, St
Charles, Missouri, USA). Cortisol levels were assayed from venous serum by
competitive immunoassay on an Immulite 2000 Analyser using the Immulite Cortisol
Assay kit (Diagnostic Products Corporation, Los Angeles, CA, USA). Growth hormone
levels were determined from serum by immunometric assay on an Immulite 2000
Analyser using the Immulite Growth Hormone Assay kit (Diagnostic Products
Corporation, Los Angeles, CA, USA). Finally, C-peptide levels were determined by
solid-phase competitive chemiluminescent enzyme immunoassay on an Immulite 2000
Analyser using the Immulite C-peptide Assay kit (Diagnostic Products Corporation, Los
Angeles, CA, USA).
3.3.4 Statistical analyses
The results were analysed using a two-way (time trial) repeated-measures analysis of
variance (ANOVA) and Fisher least significant differences (LSD) test for a posteriori
analysis using SPSS 11.0 software. Statistical significance was accepted at p < 0.05.
Participants’ characteristics are expressed as means ± S.D, whereas all other results are
expressed as means ± S.E.M.

66
3.4 Results
3.4.1 Blood metabolite response
Before the bout of moderate intensity exercise, blood glucose levels in both experimental
trials fell significantly (p < 0.05; Fig. 1). When glycaemia reached ~11 mM (11.2 ± 0.4
vs 11.9 ± 0.4 mM in the sprint and control trial, respectively), 20 min of cycling at 40%
peak 2OV was initiated, with the total workload being identical between treatments for each
participant (total work of 1176 ± 105 and 1178 ± 104 kJ/kg for sprint and control trials,
respectively). This resulted in a further rapid significant decrease in glycaemia in both
experimental trials (sprint: 3.6 ± 0.5 mM, p < 0.05; control: 3.1 ± 0.5 mM, p < 0.05; Fig.
3.1). When a 10-sec maximal sprint effort was performed immediately after the moderate
intensity exercise, the sprint opposed a further fall in blood glucose levels for the
following 120 min. In contrast, blood glucose levels decreased further (3.6 ± 1.22 mM;
p < 0.05) in the control trial (Fig. 3.1).
The response of free fatty acid levels to the sprint and control trials was similar, with
stable levels observed during moderate intensity exercise followed by a marginal increase
later in recovery (Fig. 3.2). In response to moderate intensity exercise, lactate levels
increased moderately and rose to a greater extent in response to the sprint, reaching
maximal levels after 5 min (p < 0.05) before decreasing to basal levels within 45 min after
the sprint. In contrast, in the absence of a short sprint, lactate levels remained at stable
basal levels throughout the recovery period.

67
Figure 3.1: Effect of a 10-sec sprint on blood glucose levels after moderate intensity
exercise. The moderate intensity exercise commenced at time-point -20 min. The black
line represents the sprint whereas the shaded box represents moderate intensity exercise.
Blood glucose levels are expressed relative to those immediately after moderate intensity
exercise (time-point= 0). All data are means standard error. Black circles refer to the
sprint trial whereas white circles refer to the control trial. b represents a statistically
significant difference (p < 0.05) compared to the 0 min time point after moderate intensity
exercise, c represents a statistically significant difference (p < 0.05) compared to the 0 min
time point after moderate intensity exercise.

68
3.4.2 Hormonal response
In response to the 10-sec maximal sprint effort initiated immediately after moderate
intensity exercise, epinephrine and norepinephrine reached maximal levels at the onset of
recovery (p < 0.05 for each) and returned to pre-exercise levels within 5 min after the
sprint (Fig. 3.2). In contrast, catecholamine levels remained stable in the control trial
(Fig. 3.2). Likewise, the response of growth hormone differed between the sprint and
control trials, with growth hormone levels increasing progressively after the sprint to
reach maximal levels after 15 min of recovery (Fig. 3.2). The response of plasma cortisol
levels in the sprint trial also differed from that of the control trial, with cortisol levels
increasing significantly during recovery to reach maximal levels 30 min after the sprint
(p < 0.05), but remained at stable levels in the control trial (Fig. 3.2). Glucagon increased
early in the recovery period in the control trial (p < 0.05) but did not change significantly
in the sprint trial. Finally, the pattern of insulin response to exercise was similar in the
sprint and control trials, with insulin levels remaining relatively stable throughout
exercise and recovery (Fig. 3.2). It is important to note that both trials were performed at
a time of the day when insulin levels were elevated (121 ± 15 and 109 ± 10 min after
insulin injection in the sprint and control trials, respectively), and plasma insulin levels
were similar between trials (Fig. 3.2).
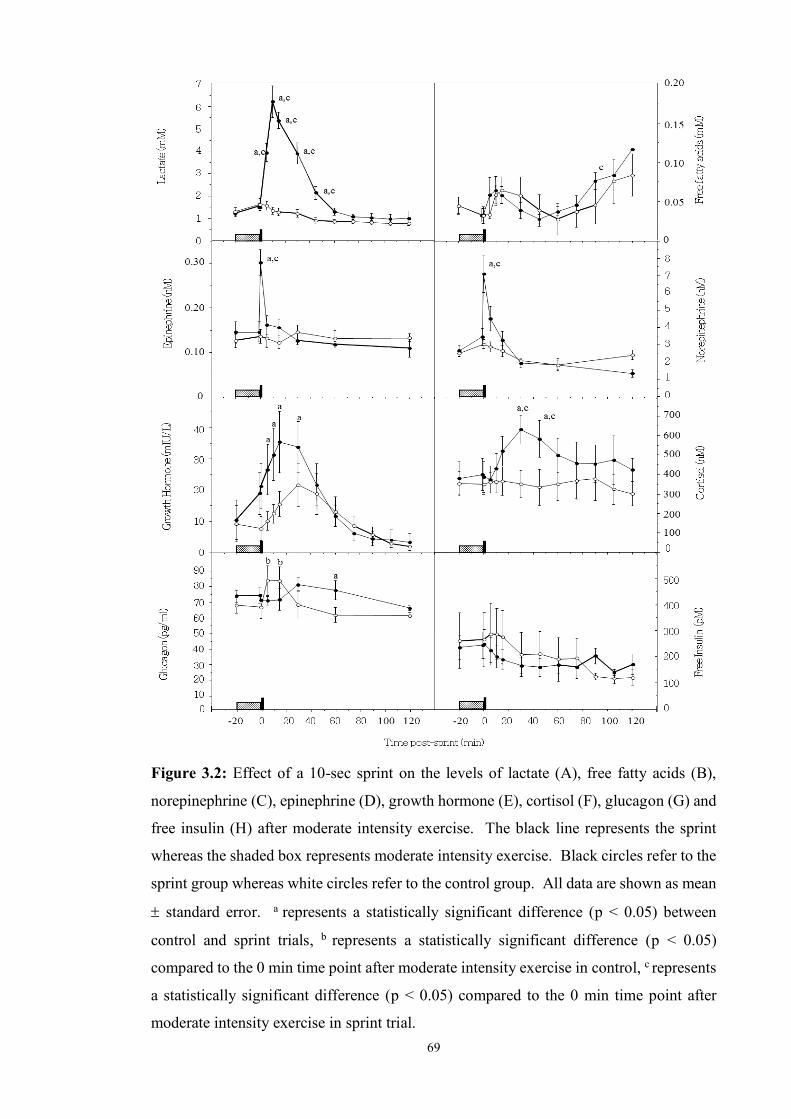
69
Figure 3.2: Effect of a 10-sec sprint on the levels of lactate (A), free fatty acids (B),
norepinephrine (C), epinephrine (D), growth hormone (E), cortisol (F), glucagon (G) and
free insulin (H) after moderate intensity exercise. The black line represents the sprint
whereas the shaded box represents moderate intensity exercise. Black circles refer to the
sprint group whereas white circles refer to the control group. All data are shown as mean
standard error. a represents a statistically significant difference (p < 0.05) between
control and sprint trials, b represents a statistically significant difference (p < 0.05)
compared to the 0 min time point after moderate intensity exercise in control, c represents
a statistically significant difference (p < 0.05) compared to the 0 min time point after
moderate intensity exercise in sprint trial.

70
3.5 Discussion
Current guidelines for minimising the risks of hypoglycaemia associated with exercise in
type 1 diabetes recommend a reduction in insulin dose or increased ingestion of
carbohydrates before exercise based on an individual’s previous glycaemic responses to
similar exercise (Zinman et al., 2004). This study investigated the intriguing possibility
of using a short bout of intense exercise as another means to counter an exercise-mediated
fall in glycaemia. In particular, it examined whether a short 10-sec cycling sprint could
acutely oppose an exercise-mediated fall in glycaemia during recovery from exercise in
individuals with insulin-treated, complication-free type 1 diabetes. Our results suggest
that to minimise the risk of a fall in glycaemia after a bout of moderate intensity exercise
in young individuals with complication-free type 1 diabetes, it is preferable to engage in
a 10-sec maximal sprint effort before resting than to only rest during recovery. Such a
sprint opposed a further fall in blood glucose levels for at least 120 min, whereas
glycaemia decreased significantly (p < 0.05) by ~ 3.5mM if no sprint was performed (Fig.
3.1). Sprinting is likely to counter the exercise-mediated decrease in blood glucose levels
through an increase in catecholamine, lactate, cortisol and growth hormone levels. The
ability of the sprint to oppose the fall in glycaemia was more remarkable considering the
sprint and control trials were performed when insulin levels were elevated, a time when
exercise is not usually recommended (Rabasa-Lhoret et al., 2001).
It is likely that the marked rise in catecholamine levels at the onset of recovery after the
sprint explain how such a sprint counters an exercise-mediated fall in glycaemia, as the
levels of the other counterregulatory hormones examined in this study did not change
significantly during this time (Fig. 3.2). It is generally acknowledged that high
catecholamine levels oppose an insulin-mediated fall in glycaemia (Marliss and Vranic,
2002) via their activation of hepatic glucose production and inhibition of insulin-mediated
glucose uptake in skeletal muscles (Nonogaki, 2000). Likewise, elevated lactate levels
(Fig. 3.2) may contribute to the stabilisation of glycaemia early in recovery by providing
gluconeogenic precursors for hepatic glucose production (Miller et al., 2002). The above
interpretation has to be taken with caution given recent findings from this laboratory that
a 10-sec sprint by itself has no effect on hepatic glucose production rate but decreases
peripheral glucose utilisation rate despite causing a rise in plasma catecholamines and
lactate levels comparable to those measured here (Fahey et al., 2012).

71
In this study, catecholamines returned rapidly to basal levels after the sprint, thus raising
the question of whether other hormones opposed the decrease in glycaemia as recovery
progressed in the sprint trial. Because insulin levels did not change significantly from pre-
exercise levels over the two hours of recovery (Fig. 3.2), they could not explain how
sprinting opposed the exercise-mediated fall in glycaemia. Likewise, the absence of an
increase in plasma glucagon levels in the sprint trial makes it unlikely that glucagon was
responsible for opposing the decrease in glycaemia (Fig. 3.2). Although there is some
evidence that the progressive rise in cortisol levels may play a role in stabilising
glycaemia due to cortisol’s potential acute inhibitory effect on glucose utilisation in
skeletal muscles (Shamoon et al., 1980), it is likely that this hormone played only a minor
role because the effects of cortisol on hepatic glucose production and blood glucose levels
have been shown to require several hours to take place (McMahon et al., 1988).
The elevated levels of growth hormone after exercise (Fig. 3.2) might play some role in
opposing the decrease in glycaemia later during recovery from sprinting. In support of
this view, the administration of a physiological growth hormone pulse in non-exercised
non-diabetic individuals has been reported to result in a rapid fall in muscle glucose
uptake (Moller et al., 1990, 1992b, 2003) and a 1-2 h delayed increase in lipolysis,
circulating free fatty acid levels, and fat oxidation rates (Moller et al., 1990, 1992b, 2003),
which could contribute further to lowering glucose utilisation rates (Møller et al., 1992b).
However, the aforementioned fall in muscle glucose uptake in response to a growth
hormone pulse does not occur in insulin-treated individuals with type 1 diabetes (Møller
et al., 1992a) and is not associated with a corresponding change in glycaemia and glucose
appearance (Ra) and disposal (Rd) rates (Møller et al., 2003), thereby making it unlikely
that growth hormone had a role in opposing the post-sprint fall in glycaemia. More
importantly, the administration of this hormone after a bout of moderate intensity exercise
in growth hormone-deficient individuals has no effect on glucose Ra and Rd (Kanaley et
al., 2004), and the administration of octreotide (an inhibitor of growth hormone secretion)
in non-diabetic individuals has also no acute effects on the magnitude of the
hyperglycaemic effect of high intensity exercise (Sigal et al., 1996). It is important to
stress, however, that no study so far has evaluated whether glucose metabolism in
response to a short sprint is affected by growth hormone levels. The identity of the
counterregulatory hormone(s) responsible for opposing the fall in glycaemia when a
sprint is performed following a bout of moderate intensity exercise remains to be
established.

72
In conclusion, this study provides the first evidence that a short maximal sprint effort
performed immediately after moderate intensity exercise is preferable to only resting as
a means to counter a further fall in glycaemia after exercise, thus decreasing the risk of
early post-exercise hypoglycaemia in individuals with type 1 diabetes. On this basis, one
might tentatively recommend that following exercise of moderate intensity, young
individuals with complication-free type 1 diabetes consider performing a short 10-sec
sprint before resting to counter a further fall in their blood glucose levels rather than only
resting, particularly if a source of dietary carbohydrate is not readily available. This
recommendation does not extend to intermittent high intensity exercise, as a study from
our laboratory has shown recently that blood glucose remains at stable levels for at least
one hour after this type of exercise (Guelfi et al., 2005a, 2005c). Although the long-term
health benefits of regular exercise are generally recognised, to the best of our knowledge
these findings provide the first example of a bout of exercise offering immediate short-
term benefits (stabilisation of glycaemia). It is important to stress, however, that different
results might have been obtained had sprinting been initiated after exercise of higher
intensity or longer duration, in younger or older individuals with reduced sprinting
capacity, or in individuals with impaired counterregulatory responses. For these reasons,
more studies of the kind described here will be required to identify the subpopulation of
type 1 diabetic individuals for whom a short maximal sprint effort can be recommended
as a safe approach for the short-term stabilisation of blood glucose levels.
3.6 Acknowledgements This research was funded jointly by a Juvenile Diabetes Research Foundation/National
Health Medical Research Council of Australia program grant to T. Jones and P.A.
Fournier. L. D. Ferreira is supported by a Juvenile Diabetes Research Fund International
Fellowship. The authors acknowledge the technical assistance of Leanne Youngs.

73
Chapter 4
A 10-second Sprint Performed Prior to
Moderate-Intensity Exercise Prevents Early
Post-Exercise Fall in Glycaemia in
Individuals with Type 1 Diabetes
An amended version of this chapter has been published in Diabetologia:
Bussau, V.A., Ferreira, L.D., Jones, T.W. & Fournier, P.A. (2007). A 10-s sprint
performed prior to moderate-intensity exercise prevents early post-exercise fall in
glycaemia in individuals with type 1 diabetes. Diabetologia 50 (9): 1815-1818.

74
4.1 Abstract
Objective: We investigated whether a 10-sec maximal sprint effort performed
immediately prior to moderate-intensity exercise provides another means to counter the
rapid fall in glycaemia associated with moderate-intensity exercise in individuals with
type 1 diabetes.
Research Design and Methods: Seven complication-free type 1 diabetic males (21.6 ±
3.6 years; mean ± SD) with HbA1c levels of 7.4 ± 0.7% injected their normal morning
insulin dose and ate their usual breakfast. When post-meal glycaemia fell to ~11 mM,
participants were asked to perform a 10 sec all-out sprint (sprint trial) or to rest (control
trial) immediately before cycling at 40% of peak rate of oxygen consumption for 20 min,
with both trials conducted in a random counterbalanced order.
Results: Sprinting did not affect the rapid fall in glycaemia during the subsequent bout
of moderate-intensity exercise (2.9 ± 0.4 mM fall in 20 min; p = 0.00; mean ± SE).
However, during the following 45 min of recovery, glycaemia in the control trial
decreased by 1.23 ± 0.60 mM (p = 0.04) while remaining stable during early recovery in
the sprint trial, subsequently decreasing in this latter trial at a rate similar to that in the
control trial. The large increase in norepinephrine (p = 0.005) and lactate levels (p =
0.0005) may have contributed to the early post-exercise stabilisation of glycaemia in the
sprint trial. During recovery, epinephrine and free fatty acid levels increased marginally
in the sprint trial, but other counterregulatory hormones did not change significantly (p <
0.05).
Conclusions: A 10-sec sprint performed immediately prior to moderate-intensity
exercise prevents glycaemia from falling during early recovery from moderate intensity
exercise in individuals with type 1 diabetes.

75
4.2 Introduction
Individuals with type 1 diabetes mellitus are faced with the daily challenge of maintaining
their blood glucose levels within a narrow physiological range in order to minimise the
risks of developing long-term diabetic complications associated with chronic
hyperglycaemia (DCCT, 1993). Unfortunately, the achievement of good glycaemic
control results in an increased risk of experiencing severe episodes of hypoglycaemia
(DCCT, 1993). This is further exacerbated in active individuals, since physical activity
of moderate intensity increases the risk of hypoglycaemia both during exercise
(Tuominen et al., 1995; Riddell et al., 1999; Rabasa-Lhoret et al., 2001) and for several
hours afterwards (MacDonald, 1987). For these reasons, current guidelines aimed at
minimising the risk of a fall in glycaemia associated with exercise recommend a reduction
in insulin dose or increased ingestion of carbohydrates prior to exercise based on the
individual’s previous glycaemic responses to similar exercise (Zinman et al., 2004)
It is well established that prolonged exercise of high intensity (>10 min at >80% peak rate
of oxygen consumption; O2peak) differs from moderate-intensity exercise in that it causes
an increase rather than a fall in glycaemia in insulin-treated individuals with type 1
diabetes (Marliss and Vranic, 2002). For this reason, our laboratory recently examined
whether the performance of one or several short maximal sprint efforts could offer an
alternative means to counter the fall in glycaemia associated with moderate-intensity
exercise or elevated plasma insulin levels in individuals with type 1 diabetes (Chapter 3;
Guelfi et al., 2005b, 2005c; Bussau et al., 2006). This research showed that the fall in
glycaemia during and after exercise of moderate intensity is not as marked if this type of
exercise is interspersed with several 4 sec sprints, despite more work being performed
(Guelfi et al., 2005b). Also, as described in Chapter 3 (Bussau et al., 2006), we found
that immediately after a bout of moderate-intensity exercise it is preferable to sprint for
10 sec than to only rest as a means of countering a decrease in glycaemia during recovery.
It is important to note that the sprints in these studies (Chapter 3; Guelfi et al., 2005b,
2005c; Bussau et al., 2006) were short enough (4–10 sec) to be well tolerated by all
participants.
Given the benefits of sprinting during or after moderate-intensity exercise (Chapter 3;
Guelfi et al., 2005b; Bussau et al., 2006), this raises the intriguing question of whether
performing a short sprint effort immediately prior to, rather than after, moderate-intensity
V

76
exercise may offer a novel way of preventing the rapid fall in glycaemia normally
observed during and after moderate-intensity exercise. In other words, this study
examines whether short-duration sprinting should be an integral part of the preparatory
routine of individuals with type 1 diabetes before they engage in sustained physical
activities of moderate intensity.

77
4.3 Methods
4.3.1 Participants
Seven males with type 1 diabetes, (age 21.6 ± 3.6 years; BMI 26.7 ± 4.3 kg/m2; O2peak
45.2 ± 5.0 ml kg–1 min –1; diabetes duration 9.5 ± 3.3 years; HbA1c 7.4 ± 0.7%; total daily
insulin dose 94 ± 38 units; all means ± SD), who were free from diabetic complications,
had undetectable levels of C-peptide and were hypoglycaemia-aware, gave informed
consent in accordance with both the University of Western Australia and Princess
Margaret Hospital Ethics Committees. Participants were not taking any prescribed
medication other than insulin and had not changed their insulin regimen for at least 3
months before the study. Following a familiarisation session, during which
anthropometric measurements were taken, O2peak was determined as described
previously (Fairchild et al., 2002b). The next two visits were the sprint and control rest
trials conducted in a random counterbalanced order.
4.3.2 Experimental trials and assays
In the 48 h prior to the experimental trial, participants were not allowed to exercise as
antecedent exercise could have affected the endocrine response to exercise (Galassetti et
al., 2001b). In addition, testing was rescheduled if participants had experienced a
hypoglycaemic episode during the 48 h pre-trial phase because prior hypoglycaemia can
also affect the counter-regulatory response during exercise (Galassetti et al., 2003).
Following their arrival in the laboratory at 08:00 hours, participants self-administered
their usual dose of morning insulin into their abdomen, with insulin dosage kept identical
between trials (mean dose 15 ± 2 units short-acting insulin, 16 ± 9 units intermediate-
acting insulin), and a catheter was inserted for blood sampling. Blood glucose levels were
similar (p = 0.843) before insulin injection, with levels of 10.1 ± 1.4 and 10.3 ± 1.5 mM
in the sprint and control trials, respectively. Participants then consumed breakfast to
increase glycaemia above 11 mM. Breakfast food choice and nutritional content reflected
that normally eaten by the participants, with breakfast kept the same (p > 0.05) for both
trials (1736 ± 193 kJ total energy, with relative energy content as follows; 56 ± 3%
carbohydrate; 17 ± 1% protein; 27 ± 4% fat). However, one of the control participants
consumed additional carbohydrate to achieve post-breakfast glycaemia above 11 mM.
After breakfast, no physical activity was allowed and glycaemia peaked at 13.8 ± 0.8 and
13.9 ± 0.50 mM in the sprint and control trials, respectively (p = 0.890). When glycaemia
V
V

78
post-breakfast fell back to ~ 11 mM (approximately 111 ± 10 and 106 ± 8 min after insulin
injection in the sprint and control trials, respectively; p = 0.564), participants either rested
or performed a 10-sec maximal sprint effort on an air-braked cycle ergometer (Repco,
Sydney, NSW, Australia). This was immediately followed by 20 min of moderate-
intensity exercise (40% O2peak), with the rationale underlying the intensity and duration
of this bout of moderate-intensity exercise similar to that discussed previously in Chapter
3 (Bussau et al., 2006). Blood sampling and assays of metabolites and hormones were
also performed as described in earlier studies from this laboratory (Chapter 3; Guelfi et
al., 2005b; Bussau et al., 2006).
4.3.3 Statistical analyses
Data were analysed using a two-way repeated-measures ANOVA and Fisher’s least
significant differences test for a posteriori analysis with SPSS 13.0 for Windows software
(SPSS, Chicago, IL, USA). Statistical significance was accepted at p < 0.05.
Participants’ characteristics are expressed as mean ± SD; all other results are mean ±
SEM.
V

79
4.4 Results
4.4.1 Blood metabolite response
Before the bout of moderate intensity exercise, blood glucose levels in both experimental
groups fell significantly (p = 0.00; Fig. 4.1). When glycaemia reached ~11 mmol/l (11.4
± 0.5 and 11.8 ± 0.5 mmol/l in the sprint and control trial, respectively; mean ± S.E., n=7,
p = 0.446), participants either rested or performed a 10-sec all-out sprint immediately
before cycling at 40% O2peak for 20 min. The total workload during the 20-min bout of
exercise did not differ between treatments (total work of 295 ± 18 and 281 ± 25 kcal/kg
for the sprint and control trials, respectively). Performing either a 10-s all-out sprint prior
to exercise or only resting did not affect the rapid fall in glycaemia during the subsequent
bout of moderate intensity exercise (2.9 ± 0.4 mmol/l in the sprint trial, p = 0.00; 3.2 ±
0.5 mmol/l in the control trial, p = 0.001; Fig. 4.1). During the initial 45 min of recovery,
glycaemia in the control trial decreased linearly by 1.23 ± 0.60 mmol/l (p = 0.04), and
fell by 3.80 ± 1.33 mmol/l after 120 min of recovery (p = 0.03; Fig. 4.1). In contrast,
glycaemia remained stable in the sprint trial during the initial 45 min of recovery before
eventually decreasing at a rate similar to that in the control trial, thus resulting in a delayed
fall in blood glucose levels post-exercise (Fig. 4.1).
In the sprint trial, blood lactate levels increased significantly immediately post-sprint (p
= 0.0005) and returned to pre-exercise levels within 30 min of recovery (Fig. 4.2). In
contrast, in the control trial, lactate levels only increased immediately post-exercise (p =
0.031) before returning to pre-exercise levels after 5 min of recovery (Fig. 4.2). The
responses of free fatty acid levels to the sprint and control trials were similar early in
recovery; however these levels increased significantly between 90 (p = 0.0005) and 120
min (p = 0.0005) post-exercise in the sprint trial (Fig. 4.2).
4.4.2 Hormonal response
In response to the 10-s maximal sprint effort initiated immediately before moderate
intensity exercise, norepinephrine (p = 0.005) reached maximal levels immediately after
the sprint before gradually decreasing during the bout of moderate intensity exercise (Fig.
4.2). During recovery, epinephrine and norepinephrine returned to basal levels within 5
and 30 min, respectively. In contrast, epinephrine levels in the control trial remained
relatively stable, whereas norepinephrine levels were only elevated immediately
V

80
following moderate intensity exercise (p = 0.013) and returned to basal levels within 5
min post-exercise (Fig. 4.2). The responses of growth hormone and cortisol in the sprint
differed markedly from those of the catecholamines in that growth hormone levels did
not change significantly and cortisol levels only increased marginally immediately after
the sprint (p = 0.026; Fig. 4.2). Glucagon levels also did not change significantly in
response to the sprint, but increased transiently (P < 0.05) at the onset of recovery from
exercise in the control trial (Fig. 4.2). Finally, insulin levels before and after exercise did
not change significantly in both trials (Fig. 4.2). It is noteworthy that both trials were
performed at a time when plasma insulin levels were elevated (111 ± 10 and 106 ± 8 min
in the sprint and control trials, respectively, post-insulin administration).
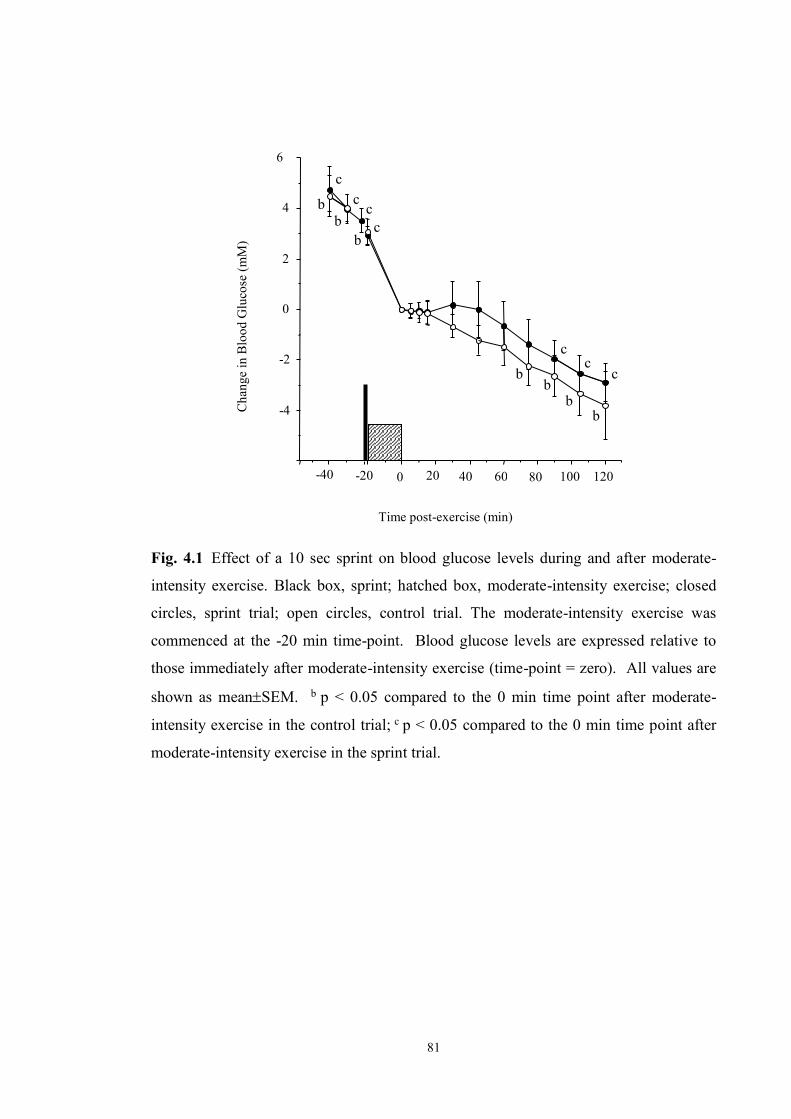
81
Fig. 4.1 Effect of a 10 sec sprint on blood glucose levels during and after moderate-
intensity exercise. Black box, sprint; hatched box, moderate-intensity exercise; closed
circles, sprint trial; open circles, control trial. The moderate-intensity exercise was
commenced at the -20 min time-point. Blood glucose levels are expressed relative to
those immediately after moderate-intensity exercise (time-point = zero). All values are
shown as meanSEM. b p < 0.05 compared to the 0 min time point after moderate-
intensity exercise in the control trial; c p < 0.05 compared to the 0 min time point after
moderate-intensity exercise in the sprint trial.
c c
c
b b
b Cha
nge
in B
lood
Glu
cose
(mM
)
120 100 80
Time post-exercise (min)
60 40 20 0 -20 -40
-4
-2
0
2
4
6
b
b
c c
c
b b
c
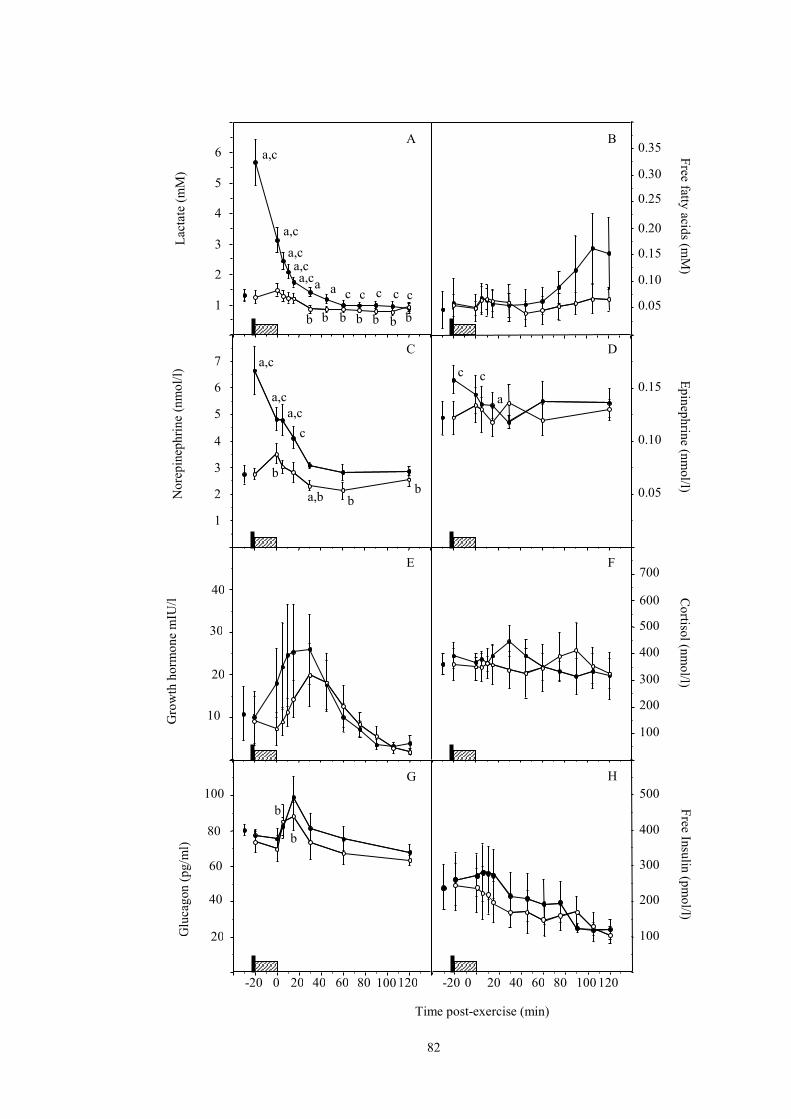
82
120 100 80 60 40 20 0 -20
1
2
3
4
5
6 a,c
a,c a,c a,c a,c a
c a c c c c 0.05
0.10
0.15
0.20
0.25
0.30
0.35 A B
20
40
60
80
100 G
0.05
0.10
0.15
D
1
2
3
4
5
6
7 a,c
b
a,c a,c
c
b
b a,b
C
10
20
30
40
Lact
ate
(mM
) G
luca
gon
(pg/
ml)
Nor
epin
ephr
ine
(nm
ol/
l)
Gro
wth
hor
mon
e m
IU/l
E
100
200
300
400
500
600
700 F
Free fatty
acids (m
M)
Epinephrine (nmol/l)
Cortisol (nm
ol/l) Free Insulin (pm
ol/l)
H
b b b b b b b
b
b
c
a
c
Time post-exercise (min)
100
400
300
200
500
120 100 80 60 20 40 0 -20

83
Fig. 4.2 Effect of a 10-sec sprint on the levels of lactate (A), free fatty acids (B),
norepinephrine (C), epinephrine (D), growth hormone (E), cortisol (F), glucagon (G) and
free insulin (H) during and after moderate-intensity exercise. Black boxes, sprint; hatched
boxes, moderate-intensity exercise; closed circles, sprint trial; open circles, control trial.
All values are shown as mean SEM. a p < 0.05 compared with control; b p < 0.05
compared to the 0 min time point after moderate-intensity exercise in the control trial; c p
< 0.05 compared to the 0 min time point after moderate-intensity exercise in the sprint
trial.

84
4.5 Discussion It is well established that exercise of moderate intensity increases the risk of
hypoglycaemia both during and after exercise in type 1 diabetes (MacDonald, 1987;
Tuominen et al., 1995; Riddell et al., 1999; Rabasa-Lhoret et al., 2001). However, in
Chapter 3 we reported that a single all-out 10-s sprint performed immediately after
moderate intensity exercise provides a simple means to oppose a decrease in glycaemia
during recovery (Bussau et al., 2006). Here we show that although a 10-sec sprint
initiated immediately before moderate-intensity exercise did not affect the rapid decline
in glycaemia during a subsequent bout of moderate-intensity exercise performed when
plasma insulin levels were elevated, it did prevent the fall in glycaemia for at least the
first 45 min of recovery (Fig. 4.1). This suggests that including a short sprint as part of
the preparatory routine of individuals with type 1 diabetes before they engage in sustained
moderate-intensity exercise might provide another means of temporarily stabilising
glycaemia during early recovery.
The marked rise in norepinephrine and lactate levels immediately after exercise in the
sprint trial might explain, in part, how sprinting before moderate-intensity exercise delays
the fall in glycaemia early into recovery. Norepinephrine levels during early recovery
were elevated in the sprint trial while blood glucose levels were stable, but soon after the
return of norepinephrine to basal pre-exercise levels, blood glucose levels in the sprint
and control trials fell at comparable rates (Fig. 4.2). Although high catecholamine levels
have been generally acknowledged to counter insulin-mediated falls in glycaemia
(Marliss and Vranic, 2002) via activation of hepatic glucose production and inhibition of
insulin-mediated glucose uptake in skeletal muscles (Nonogaki, 2000), their role as
counter-regulatory hormones during exercise in type 1 diabetic individuals has been a
controversial issue (Coker and Kjaer, 2005). Elevated lactate levels (Fig. 4.2) are another
factor that may have contributed to the stabilisation of glycaemia early in recovery by
providing gluconeogenic precursors for hepatic glucose production and by increasing
peripheral insulin resistance (Vettor et al., 1997). The above interpretation is challenged,
at least in part, by recent findings from this laboratory that sprinting for 10 sec has no
effect on hepatic glucose production rate but decreases peripheral glucose utilisation rate
despite causing a rise in plasma catecholamines and lactate levels comparable to those
measured here (Fahey et al., 2012).

85
The patterns of change in the levels of insulin, cortisol, glucagon and growth hormones
in the sprint and control trials suggest that these hormones are unlikely to explain the
delayed fall in glycaemia during recovery in the sprint trial. Insulin levels in both trials
did not change significantly during recovery (Fig. 4.2). Also, the levels of glucagon did
not increase significantly during and after exercise in the sprint trial. The question of
whether portal glucagon levels responded in a similar manner remains to be established
(Fig. 4.2). Although there is some published evidence that a rise in cortisol levels may
play a role in stabilising glycaemia due to its potential acute inhibitory effect on glucose
utilisation in skeletal muscles (Shamoon et al., 1980), this hormone is likely to play only
a minor role here not only because of the absence of changes in the levels of this hormone
during recovery in both trials, but also because the effects of cortisol on hepatic glucose
production and blood glucose levels require several hours to take place (McMahon et al.,
1988). Finally, growth hormones are also unlikely to play a role as no significant changes
in growth hormone levels were observed in either of the groups (Fig. 4.2). However, a
close inspection of Fig. 4.2e suggests that significant changes in growth hormone levels
might have been masked by the large inter-individual variability in growth hormone
levels. Nevertheless, even if growth hormone levels had increased significantly, this
would probably have been of lesser importance because it has been reported that (1)
administration of a growth hormone pulse does not affect peripheral glucose uptake in
insulin-treated individuals with type 1 diabetes (Møller et al., 1992a); and (2) the infusion
of octreotide, an inhibitor of growth hormone release, has no acute effects on the
magnitude of the hyperglycaemic effect of high-intensity exercise (Sigal et al., 1996).
There is evidence that the aforementioned factors proposed to explain the stabilising
effects of a short sprint on post-exercise glycaemia might be similar, irrespective of
whether sprinting is performed before, during or after moderate-intensity exercise.
Indeed, elevated lactate and catecholamines levels have also been proposed to stabilise
glycaemia at the onset of recovery from moderate-intensity exercise interspersed either
with several short sprints (Guelfi et al., 2005b) or followed by a 10-sec sprint (Chapter 3;
Bussau et al., 2006). It is less clear, however, whether GH is also involved under these
conditions because of a lack of consistencies across studies. Clearly, more research is
required to identify the factors responsible for the sprint-mediated stabilisation of
glycaemia after exercise.

86
On practical and clinical grounds, our findings suggest that before engaging in moderate-
intensity exercise, individuals with type 1 diabetes might consider incorporating into their
warm-up routine a short 10-sec maximal sprint effort, especially as a 10-sec sprint is well
tolerated (Chapter 3; Bussau et al., 2006). However, before recommending the use of
sprinting to prevent an early post-exercise fall in glycaemia, a number of issues must be
addressed such as the extent to which the efficacy of prior sprinting is affected by the
intensity and duration of the subsequent bout of exercise and the identification of any
subpopulations of type 1 diabetic individuals unlikely to benefit from sprinting, such as
very young children and older individuals who have limited capacity to engage in a
maximal sprint effort..
In conclusion, our findings show that sprinting before a bout of exercise of moderate
intensity could be a novel tool of clinical importance for preventing the fall in glycaemia
during early recovery in young healthy individuals with type 1 diabetes. These findings
provide one of the very few examples (Chapter 3; Guelfi et al., 2005b, 2005c; Bussau et
al., 2006) of exercise offering an immediate short-term clinical benefit (delayed fall in
glycaemia).
4.6 Acknowledgements This research was funded jointly by a Juvenile Diabetes Research Foundation
(JDRF)/National Health Medical Research Council of Australia program grant to T. W.
Jones and P. A. Fournier. L. D. Ferreira is supported by a JDRF Fellowship. The authors
acknowledge the technical assistance of L. Youngs and A. Thompson.

87
Chapter 5
Counterregulatory Response to a 10-second
Sprint in Individuals with Type 1 Diabetes
Mellitus

88
5.1 Abstract
Objective: A 10-sec sprint performed before or after moderate intensity exercise reduces
the risk of post-exercise hypoglycaemia in hyperinsulinaemic fed individuals with type 1
diabetes mellitus. Since the glucoregulatory response to such a short sprint in these
individuals has never been examined, this study investigates the response of the
counterregulatory hormones to a 10-sec sprint in individuals with type 1 diabetes under
hyperinsulinaemic fed conditions.
Research Design and Methods: Seven complication-free male individuals with type 1
diabetes (HbA1c = 7.0 ± 0.3%) were recruited to the study. On the morning of testing, the
participants followed their normal insulin regimen and ate their usual breakfast. Blood
glucose levels were determined regularly and allowed to fall to approximately 6.0 mM.
Then, participants pedalled for 10 sec at maximal intensity on a cycle ergometer.
Results: Immediately after sprinting, plasma norepinephrine and epinephrine levels rose
significantly (p < 0.05) to maximal levels (7.3 ± 1.0 and 0.66 ± 0.09 nmol/L, respectively,
p < 0.05). Growth hormone increased to maximal levels (33.1 ± 6.7 mIU/L) 30 min after
sprinting (p < 0.05), and remained elevated for the whole duration of the recovery period.
Glucagon levels remained at stable levels. Similarly, plasma insulin and cortisol levels
did not change significantly. Before sprinting, glycaemia fell significantly and remained
stable during recovery.
Conclusions: The response of the counterregulatory hormones to a 10-sec sprint in type
1 diabetic individuals in a moderate hyperinsulinaemic and fed state is characterised
mainly by marked changes in catecholamines and growth hormone levels.

89
5.2 Introduction
Individuals with type 1 diabetes mellitus are encouraged to participate in regular physical
activity due to the numerous physiological and psychological health benefits associated
with an active lifestyle (Norris et al., 1990; Moy et al., 1993; Laaksonen et al., 2000;
Zinman et al., 2004; Riddell and Iscoe, 2006; Chimen et al., 2012, Tonoli et al., 2012).
Unfortunately, the risk of hypoglycaemia is increased both during exercise (Riddell et al.,
1999; Tuominen et al., 1995; Rabasa-Lhoret et al., 2001) and for several hours during
recovery (MacDonald, 1987a; Tsalikian et al., 2005; McMahon et al., 2007; Maran et al.,
2010; Iscoe and Riddell, 2011; Davey et al., 2013a). Due to this increased risk of
hypoglycaemia, it is not surprising that fear of hypoglycaemia is a major barrier to regular
physical activity in individuals with type 1 diabetes (Ludvigsson et al., 1980; Guelfi et
al., 2007c; Brazeau et al., 2008).
As mentioned in Chapter 1, not all types of exercise increase the risk of hypoglycaemia.
Several studies have reported that 10-15 min of high-intensity aerobic exercise (>80% of
peak 2OV ) under basal insulinaemic conditions is accompanied by an increase in blood
glucose levels during and after exercise (Mitchell et al., 1988; Marliss et al., 1992a;
Purdon et al., 1993; Sigal et al., 1994c, 1999; Marliss and Vranic, 2002). This is also the
case in response to a 10-sec maximal sprint effort (Fahey et al., 2012). This glycaemia-
rising effect of aerobic short sprint is such that, in theory, this type of exercise might be
beneficial if adopted to prevent or delay hypoglycaemia when no carbohydrate is readily
available to exercising individuals with type 1 diabetes.
As indicated in Chapter 3 and 4, when blood glucose level in individuals with type 1
diabetes is decreasing rapidly during moderate intensity exercise, engaging in a maximal
10-sec sprint immediately before or after exercise causes blood glucose levels after
exercise to stabilise for 0.5-2 hours, thus decreasing the early risk of hypoglycaemia post-
exercise (Chapters 3 and 4; Bussau et al., 2006, 2007). It is noteworthy that sprinting in
these studies was beneficial despite elevated plasma insulin levels, a time when physical
activity is generally not recommended due to the increased risk of experiencing
hypoglycaemia (Rabasa-Lhoret et al., 2001).
In an attempt to identify the endocrine mechanisms likely to mediate the glucoregulatory
benefits of sprinting, the levels of several counterregulatory hormones were measured in

90
the aforementioned studies (Chapters 3 and 4; Bussau et al., 2006, 2007), with our
findings suggesting that a post-exercise increase in catecholamines and growth hormone
levels might contribute to the protective effect of sprinting in type 1 diabetic individuals
(Chapters 3 and 4; Bussau et al., 2006, 2007). However, one difficulty with comparing
these findings with the literature has been the lack of information on the effect of short-
duration sprinting per se on the levels of counterregulatory hormones in
hyperinsulinaemic diabetic individuals in a fed state. Indeed, only one study has examined
the effect of a 10-sec sprint on the levels of these hormones in individuals with type 1
diabetes (Fahey et al., 2012). However, this study was performed in overnight fasted
individuals and under basal insulinaemic conditions, thus leaving unanswered the
question of the effect of sprinting on the counterregulatory hormones responses to short-
duration sprinting under combined hyperinsulinaemic and fed conditions. Given the
clinical benefits of a 10-sec sprint for the prevention of hypoglycaemia in
hyperinsulinaemic fed individuals with type 1 diabetes, and the lack of information about
the counterregulatory response to this type of exercise, the purpose of this study was to
investigate the effect of a single 10-sec sprint on the levels of the counterregulatory
hormones under dietary and insulinaemic conditions mimicking those where sprinting has
been shown to be beneficial to individuals with type 1 diabetes. Given that our studies on
the benefits of sprinting were performed in hyperinsulinaemic fed individuals (Chapters
3 and 4; Bussau et al., 2006, 2007), this was also the physiological state examined here.
The information thus obtained should provide the basis for future studies aimed at
elucidating the mechanisms underlying the clinical benefits of sprinting in the prevention
of hypoglycaemia in type 1 diabetic individuals.

91
5.3 Methods
5.3.1 Participants
Seven young males with type 1 diabetes (aged 17.7 0.5 years; BMI 25.9 4.1kg/m2;
peak 2OV 43.5 ± 4.8 ml/kg/min; duration of diabetes 5.8 ± 1.8 years) were recruited from
Princess Margaret Hospital and the University of Western Australia, respectively. All
participants were in moderate glycaemic control (Hb A1c = 7.0 ± 0.3%), free from
diabetic complications, were hypoglycaemia aware, had undetectable levels of c-peptide,
and were not taking any prescribed medication other than insulin. Moreover, these
participants were on a multiple-injection insulin regime that had not changed for at least
three months prior to the study. All participants were subjected to a familiarisation session
during which their informed consent was obtained as well as their height, body mass and
maximal rate of oxygen consumption as described previously (Fairchild et al., 2002a).
Then, each participant was required to attend our laboratory on two occasions. Both the
University of Western Australia and the Princess Margaret Hospital Ethics Committees
approved the procedures described in this study.
5.3.2 Experimental trials
Participants were not allowed to exercise for 48 h prior to the experimental trial since
antecedent exercise has the potential to affect the endocrine response to exercise
(Galassetti et al., 2001b). Also, testing was rescheduled if they had experienced a
hypoglycaemic episode over the previous 48 h because prior hypoglycaemia can also
affect the counterregulatory response to exercise (Davis et al., 2000d; Galassetti et al.,
2003). Each participant was also required to maintain their normal diet and to avoid
alcohol for 24 h prior to testing. On the morning of testing, all participants were required
to self-monitor their blood glucose levels regularly before attending the laboratory at ~
07.30 h where they were instructed to self-administer their usual morning dose of insulin
into the abdomen (mean dose 34.9 ± 10.2 units). Then, they ingested their breakfast as
described in Chapters 3 and 4 (1933 ± 287 kJ total energy; 62 ± 3% carbohydrate; 16 ±
1% protein; 22 ± 4% fat), and a catheter was inserted for blood sampling. Following
breakfast and when glycaemia started falling, blood glucose levels were measured every
15 min. Then, a 10-sec maximal sprint effort was initiated on a Repco front access cycle
ergometer (Repco, Sydney, Australia) when blood glucose levels reached approximately
6.0 mM. The participants were then instructed to cycle as hard as possible for 10 sec and
not to pace themselves for the whole duration of the sprint. Blood was sampled prior to

92
exercise and then at 0, 5, 10, 15, 30, 45 and 60 min post-sprint. At each sampling point,
0.2 ml of blood was first removed for the measurement of blood glucose, pO2, pH and
lactate. A second sample of 15 ml was also removed for the measurement of hormones.
Some of the blood was combined with sodium metabisulphite, poly-ethylene glycol or
trasylol for the assays of catecholamines, insulin and glucagon, respectively. All samples
were then centrifuged at 720g for 5 min and the plasma stored at -80C for later analysis.
5.3.3 Hormones and metabolite assays
Glucose, lactate, pH and pO2 were analysed using an ABL 625 blood gas system
(Radiometer, Copenhagen, Denmark). Free fatty acids levels were measured using the
Roche half micro test free fatty acids assay kit (Roche Diagnostic, Germany). All
hormones were assayed as described previously (Chapters 3 and 4; Bussau et al., 2006,
2007). Plasma catecholamine levels were determined by reverse phase HPLC using a
Waters Novapak C18 reverse phase column and a model 5200A Coulochem detector
(ESA Inc, USA). Free insulin levels were measured by radioimmunoassay using the
Phadeseph insulin RIA kit (Pharmacia, Uppsala, Sweden). Glucagon levels were
measured by radioimmunoassay using a Linco glucagon RIA kit (Linco Research, St
Charles, Missouri, USA). Cortisol levels were assayed by competitive immunoassay on
an Immulite 2000 analyser using the Immulite cortisol assay kit (Diagnostic Products
Corporation, Los Angeles, CA, USA). Growth hormone (GH) levels were determined by
immunometric assay on an Immulite 2000 analyser using the Immulite growth hormone
assay kit (Diagnostic Products Corporation, Los Angeles, CA, USA). Finally, C-peptide
levels were determined by solid-phase competitive chemiluminescent enzyme
immunoassay on an Immulite 2000 analyser using the Immulite C-peptide assay kit
(Diagnostic Products Corporation, Los Angeles, CA, USA).
5.3.4 Statistical analyses
The results were analysed using a one-way repeated measure analysis of variance and
Fisher LSD test for a posteriori comparisons. Statistical significance was accepted at p
< 0.05 and all analyses were carried out using SPSS 17.0 software. Participants’
characteristics are expressed as means ± S.D whereas all other results are expressed as
means ± S.E.M.

93
5.4 Results
5.4.1 Hormonal response to a 10-sec sprint
In response to sprinting, plasma norepinephrine levels rose significantly (p < 0.05) and
reached maximal levels at the onset of recovery (7.3 ± 1.0 nM) before returning to pre-
exercise levels within 10 min after exercise (Fig. 5.1). Similarly, plasma epinephrine
reached maximal levels at the onset of recovery (0.66 ± 0.09 nM, p < 0.05) and remained
elevated above pre-sprint levels during the first 10 min of recovery before returning to
pre-sprint levels (Fig. 5.1). Plasma growth hormone levels increased progressively during
recovery from sprinting to reach maximal levels of 33.1 ± 6.7 mIU/L after 30 min of
recovery (p < 0.05; Fig 5.1), with these levels still remaining above pre-sprint levels 60
min after exercise. In response to sprinting, plasma glucagon, cortisol and insulin levels
did not change significantly (p > 0.05; Fig. 5.1). However, it must be noted that there
was a non-significant trend for cortisol levels to increase and reach maximal levels after
30 min of recovery.
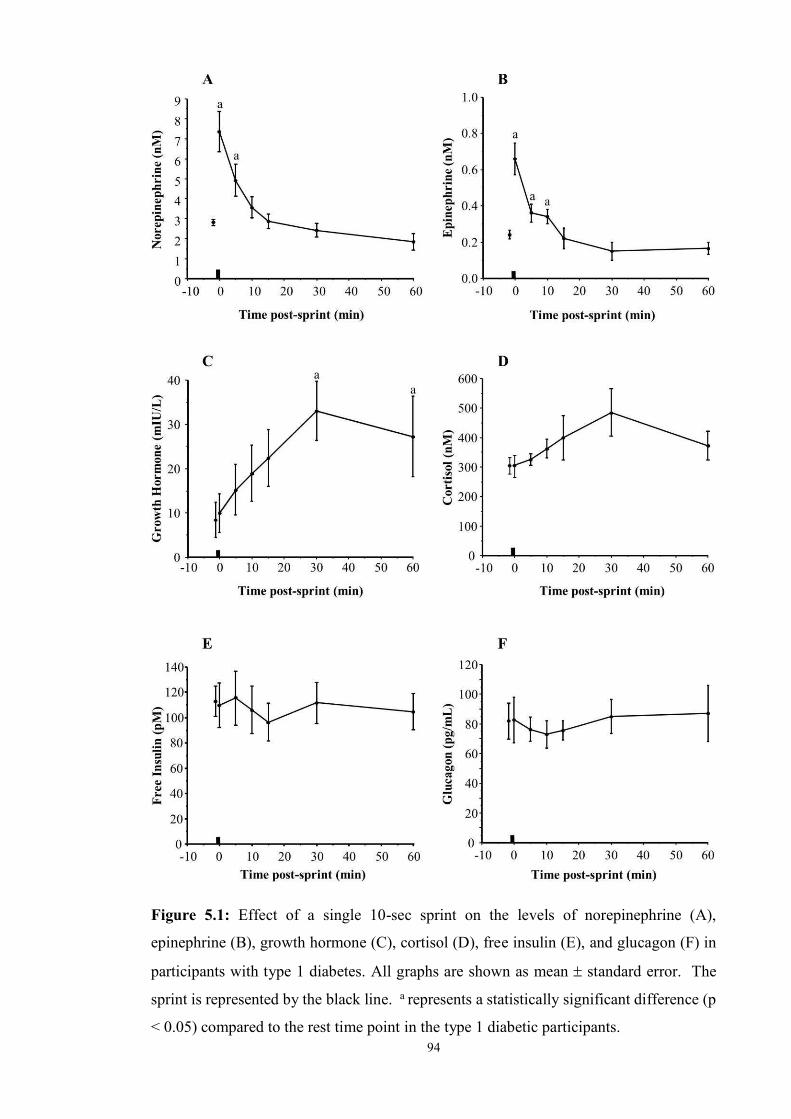
94
Figure 5.1: Effect of a single 10-sec sprint on the levels of norepinephrine (A),
epinephrine (B), growth hormone (C), cortisol (D), free insulin (E), and glucagon (F) in
participants with type 1 diabetes. All graphs are shown as mean standard error. The
sprint is represented by the black line. a represents a statistically significant difference (p
< 0.05) compared to the rest time point in the type 1 diabetic participants.

95
5.4.2 Blood metabolite response to a 10-sec sprint
In response to sprinting, blood lactate levels reached maximal levels after 5 min of
recovery (p < 0.05; Fig. 5.2) before decreasing to pre-sprint levels within 60 min after
exercise. Blood pH fell to minimal levels at 5 min after sprinting (p < 0.05; Fig. 5.2)
before returning to pre-sprint levels within 60 min of recovery. Free fatty acid levels
increased progressively after exercise to reach maximal levels after 60 min of recovery
(p < 0.05; Fig. 5.2). Before sprinting, blood glucose levels fell significantly (p < 0.05;
Fig. 5.2) and remained stable for 60 min after sprinting.
5.4.3 Work load and peak power associated with a 10-sec sprint
The total work performed during the 10-sec sprint was 82.8 8.4 kJ/kg and the peak
power achieved was 10.7 0.9 W/kg.
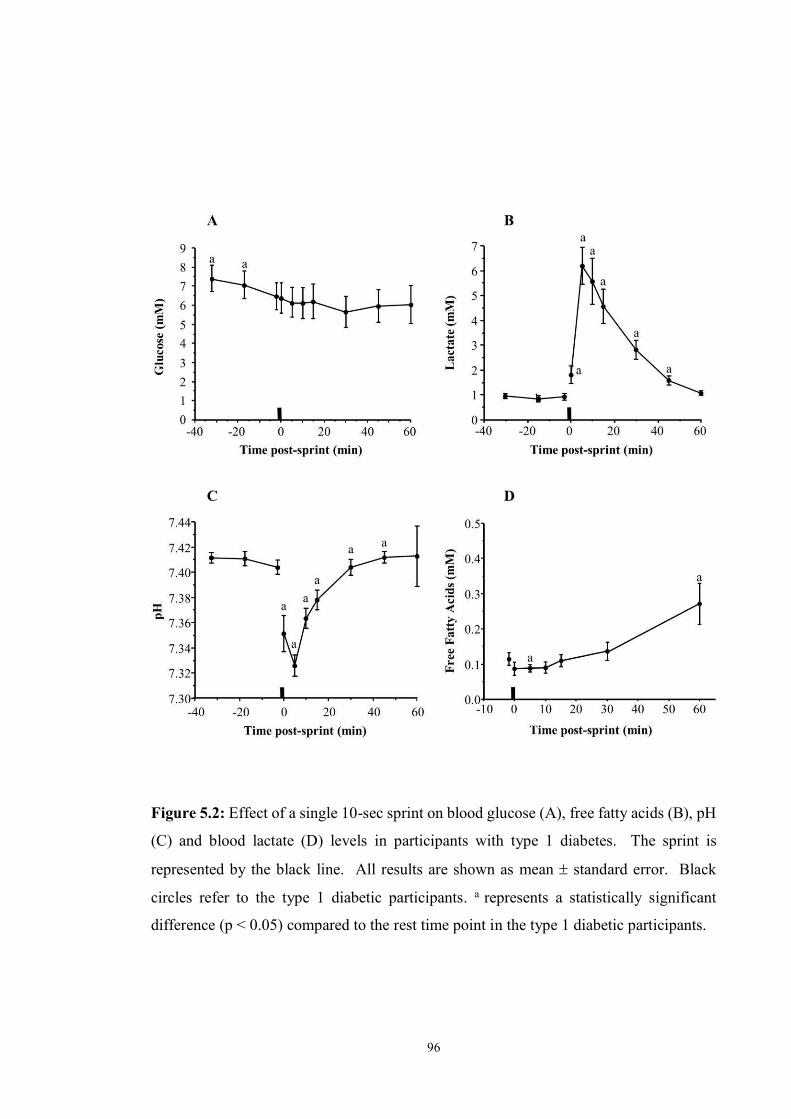
96
Figure 5.2: Effect of a single 10-sec sprint on blood glucose (A), free fatty acids (B), pH
(C) and blood lactate (D) levels in participants with type 1 diabetes. The sprint is
represented by the black line. All results are shown as mean standard error. Black
circles refer to the type 1 diabetic participants. a represents a statistically significant
difference (p < 0.05) compared to the rest time point in the type 1 diabetic participants.

97
5.5 Discussion
In chapters 3 and 4, we showed that even when plasma insulin levels are elevated, a 10-
sec sprint performed either immediately before or after moderate intensity exercise can
prevent blood glucose levels from falling after exercise (Bussau et al., 2006, 2007), thus
providing a novel clinical tool to acutely decrease the risk of post-exercise
hypoglycaemia. Although, the combined effects of sprinting and moderate intensity
exercise on counterregulatory hormones levels were examined in these studies (Chapters
3 and 4; Bussau et al., 2006, 2007), surprisingly the responses of these hormones to
sprinting per se in type 1 diabetic individuals who are fed and under hyperinsulinaemic
conditions has never been investigated before, thus making it impossible to compare our
earlier findings with those of others. Here, for the first time, the effect of short-duration
sprinting per se on the levels of several counterregulatory hormones was examined in
hyperinsulinaemic fed individuals with type 1 diabetes. We found that a 10-sec maximal
sprint in those individuals resulted in marked changes in plasma catecholamines and
growth hormone levels, with no significant effect on plasma cortisol, insulin and glucagon
levels. In addition, the responses of growth hormones and cortisol levels differed from
those observed when a sprint is performed immediately before or after a bout of moderate
intensity exercise in individuals with type 1 diabetes (Chapters 3 and 4; Bussau et al.,
2006, 2007).
Our findings show that a 10-sec sprint results in an early rise in plasma epinephrine and
norepinephrine levels in hyperinsulinaemic fed individuals with type 1 diabetes. The
magnitude of this rise, however, was submaximal and comparable to that observed when
a 10-sec sprint is performed immediately before or after a bout of moderate intensity
exercise under hyperinsulinaemic fed conditions (Chapters 3 and 4; Bussau et al., 2006,
2007). Only one other study, published from this laboratory, has investigated the effect
of a 10-sec sprint per se on plasma catecholamines levels in type 1 diabetic individuals
and also reported an increase in the levels of these hormones, but all participants were in
an overnight fasted state and under basal insulinaemic conditions (Fahey et al., 2012).
Interestingly, the rise in catecholamines levels reported here was more pronounced than
those reported by others in non-diabetic individuals subjected to a 6-sec sprint (Moussa
et al., 2003; Botcazou et al., 2007; Bracken et al., 2009), but generally of a lesser
magnitude compared to longer duration sprinting (Brooks et al., 1988; Nevill et al., 1989;

98
Langfort et al., 1997; Zouhal et al., 1998, 2001; Jacob et al., 2002; Moussa et al., 2003;
Vincent et al., 2003; Jacob et al., 2004; Vincent et al., 2004; Zouhal et al., 2009).
The pattern of change in growth hormone levels in response to sprinting in
hyperinsulinaemic type 1 diabetic individuals in the fed state differed in some respects
from that when sprinting is performed immediately before or after a bout of moderate
intensity exercise (Chapters 3 and 4; Bussau et al., 2006, 2007). Indeed, growth
hormones levels remained elevated for longer in response to sprinting alone. Only one
other study has investigated the effect of a 10-sec sprint per se on plasma growth
hormones levels in type 1 diabetic individuals, and showed that the levels of this hormone
increased post-sprinting (Fahey et al., 2012). However, as mentioned earlier, the
participants involved in that study were in an overnight fasted state and under basal
insulinaemic conditions (Fahey et al., 2012) rather than being in a hyperinsulinaemic fed
state. It is noteworthy that sprinting for as little as 6 sec has been found to result in a small
significant increase in growth hormone levels in non-diabetic individuals (Stokes et al.,
2002), with the magnitude of growth hormone response increasing with sprint duration
(Stokes et al., 2002). Other factors are also likely to affect the sprint-mediated increase
in growth hormone levels. For instance, GH secretion increases in response to both high
catecholamines levels and acidosis (Stokes, 2003), but is opposed by elevated plasma
insulin (Lanzi et al., 1997; Frystyk, 2004) and fatty acids levels (Godfrey et al., 2003).
Although sprinting for 10 sec did not change significantly plasma cortisol levels, there
was a trend towards a small increase after exercise. This trend is consistent with earlier
work from our laboratory where sprinting performed immediately before or after a bout
of moderate intensity exercise has been shown to result in a significant but small increase
in cortisol levels in hyperinsulinaemic fed individuals with type 1 diabetes (Chapters 3
and 4; Bussau et al., 2006, 2007). Only one other study has investigated the effect of a
10-sec sprint per se on plasma cortisol levels in type 1 diabetic individuals and shown
that sprinting results in a small increase in plasma cortisol levels after exercise (Fahey et
al., 2012). However, as mentioned earlier, the participants in that study were in an
overnight fasted state and under basal insulinaemic conditions (Fahey et al., 2012) rather
than being in a hyperinsulinaemic fed state.
The absence of any effect of a 10-sec sprint on plasma glucagon levels in
hyperinsulinaemic fed participants with type 1 diabetes is consistent with earlier

99
observations from our laboratory that a 10-sec sprint performed immediately before or
after moderate intensity exercise has no effect on plasma glucagon levels (Chapters 3 and
4; Bussau et al., 2006, 2007). In the absence of any change in plasma insulin levels, our
findings also suggests that the plasma glucagon/insulin ratio is not affected by a sprint of
short duration. The main limitation with this interpretation, however, is that it assumes
that portal glucagon and insulin levels follow patterns of change similar to those of
peripheral blood glucose and insulin levels. The only other study that has examined the
effect of a 10-sec sprint on plasma glucagon levels has also reported no changes in plasma
glucagon levels, but all participants were in an overnight fasted state and under basal
insulinaemic conditions (Fahey et al., 2012). Since catecholamines and insulin stimulate
and inhibit glucagon secretion (Ahren, 2000; Gromada et al., 2007) respectively, maybe
the rise in epinephrine levels was not sufficient to stimulate glucagon release.
The post-exercise rises in plasma lactate and free fatty acids levels in response to sprinting
in hyperinsulinaemic type 1 diabetic individuals in the fed state were similar to those
observed when a 10-sec sprint is performed immediately before or after a bout of
moderate intensity exercise (Chapters 3 and 4; Bussau et al., 2006, 2007). These findings
are also comparable to those obtained in a study examining the effect of a 10-sec sprint
on plasma lactate and free fatty acid levels, but all participants were in an overnight fasted
state and under basal insulinaemic conditions in that study (Fahey et al., 2012). Finally,
others have reported similar plasma lactate levels in response to a 6- or 10-sec sprint in
non-diabetic individuals (Zajac et al., 1999; Moussa et al., 2003). Plasma pH, another
potential glucoregulatory variable with the capacity to affect hepatic glucose production
and muscle glucose transport (Kashiwagura et al., 1985; Kristiansen et al., 1994) fell in
response to the 10-sec sprint, consistent with the decrease in blood pH that occur in non-
diabetic individuals after a sprint lasting 6 sec or more (Allsop et al., 1990; Bogdanis,
1995; Bogdanis and Nevill, 1996; Stokes et al., 2002, 2005).
It is important to note that the comparison made here between the findings of this study
and those obtained in participants were sprinting was performed before or after moderate
intensity exercise (Chapters 3 and 4; Bussau et al., 2006, 2007) should be performed with
caution. Although the work load was well matched between these studies, as indicated by
their similar total work and peak power, and blood glucose levels were falling, it is
important to note that pre-sprint plasma insulin levels were not fully matched at the time
of the sprint. Indeed, the hyperinsulinaemic state of our participants in this study was not

100
as marked as in those other studies. Since the release of growth hormone and glucagon
are opposed by elevated plasma insulin levels (Lanzi et al., 1997; Ahren, 2000; Frystyk,
2004; Gromada et al., 2007), it is possible that a less marked increase in plasma growth
hormone level would have occurred under more pronounced hyperinsulinaemic
conditions.
In conclusion, this study is the first one to examine the counterregulatory responses to a
maximal sprint effort of short duration in fed type 1 diabetic individuals under
hyperinsulinaemic conditions. What remains to be established in future studies is the
extent to which the changes in the levels of each of the counterregulatory hormones
examined here affects blood glucose levels, hepatic glucose production and peripheral
glucose utilisation in response to sprinting. Overall, this study provides the basis for more
research to understand better the mechanisms underlying the clinical benefits of sprinting
in hypoglycaemia prevention.
5.6 Acknowledgements This research was funded by a Juvenile Diabetes Research Foundation/National Health
Medical Research Council of Australia program grant to T. Jones and P.A. Fournier. L.
D. Ferreira was supported by a Juvenile Diabetes Research Foundation International
Fellowship.

101
Chapter 6
General Discussion

102
6.1 General Discussion
Maintaining blood glucose levels within physiological range is a constant challenge for
individuals with type 1 diabetes. As a result of the lack of an acute response of plasma
insulin levels to changes in glycaemia, blood glucose levels are usually managed by
combining daily exogenous insulin administration, carbohydrate intake, and regular
glucose monitoring. Unfortunately, despite the numerous physiological and
psychological health benefits of exercise (Norris et al., 1990; Moy et al., 1993; Laaksonen
et al., 2000; Zinman et al., 2004; Riddell and Iscoe, 2006; Chimen et al.,2012), physical
activity can make the acute management of blood glucose levels even more difficult for
individuals with type 1 diabetes (Riddell and Perkins, 2006; Guelfi et al., 2007c; Younk
and Davis, 2012). In fact, the risk of hypoglycaemia is increased both during (Tuominen
et al., 1995; Riddell et al., 1999; Rabasa-Lhoret et al., 2001) and after exercise
(MacDonald, 1987a; Tsalikian et al., 2005; McMahon et al., 2007; Maran et al., 2010;
Iscoe and Riddell, 2011; Davey et al., 2013). Consequently, it is not surprising that the
biggest barrier to regular physical activity in individuals with type 1 diabetes is fear of
hypoglycaemia (Brazeau et al., 2008). As a result, many people with type 1 diabetes
avoid physical activity (Ludvigsson et al., 1980; Fremion et al., 1987; Guelfi et al., 2007c;
Brazeau et al., 2008) with ~60-65% of individuals with type 1 diabetes found to be
inactive (Thomas et al., 2004; Plotnikoff et al., 2006). It is also concerning that young
individuals are discouraged from performing vigorous physical activity by physicians,
school staff or parents due to their fear of exercise-induced hypoglycaemia (Fremion et
al., 1987). As discussed at length in Chapter 1 of this thesis, not all types of exercise result in an
elevated risk of hypoglycaemia. Aerobic exercise performed at high intensity (>80% of
V O2max) as well as sprinting or intermittent high intensity exercise are all associated with
an increase in glycaemia during and after exercise. This raises the intriguing possibility
that these types of exercise might be beneficial if adopted to counter a fall in glycaemia
in complication-free individuals with type 1 diabetes, and thus might help to prevent or
delay hypoglycaemia if no carbohydrate is readily available. The problem here is that
adopting such a strategy to prevent hypoglycaemia is unlikely to be well tolerated by most
individuals with type 1 diabetes due to the very intense nature and impractical duration
of these types of exercise. This raises the primary aim at the core of this thesis which was
to determine whether a much shorter bout of exercise performed at maximal intensity

103
could be adopted to prevent glycaemia from falling. Since a single maximal intensity
sprint effort lasting 30-sec or more is associated with undesirable physiological
consequences such as nausea, vomiting and dizziness (Inbar et al., 1996; 1998; Laurent
et al., 2007; Stickley et al., 2008; Little et al., 2010), we undertook to explore the
glucoregulatory benefits of a maximal sprint effort lasting only 10 sec. For this reason,
the primary goal of this thesis was to determine whether a 10-sec maximal sprint effort
performed after (Chapter 3) or before (Chapter 4) moderate intensity exercise provides a
possible means other than carbohydrate intake to prevent glycaemia from falling when
exercise is performed under hyperinsulinaemic conditions by complication-free
individuals with type 1 diabetes, thus decreasing acutely their risk of hypoglycaemia.
Also, given that for this type of study, it is common practice to subject participants to a
graded exercise test to determine their V O2peak not only to determine their aerobic fitness,
but also to set exercise intensity relative to V O2peak, a secondary objective of this thesis
was to determine whether the risk of hypoglycaemia is increased early during recovery
from this exercise protocol (Chapter 2). Finally, given that the counterregulatory response
to sprinting has not been examined in hyperinsulinaemic fed individuals with type 1
diabetes, thus making it difficult to compare the findings of Chapters 3 and 4 with the
literature, our last aim was to examine the counterregulatory responses to sprinting in type
1 diabetic individuals under hyperinsulinaemic conditions (Chapter 5).
Given that most laboratory-based studies on exercise in diabetes involve the performance
of graded exercise for V O2peak and lactate threshold determination, the primary aim of the
study described in Chapter 2 was to determine whether the risk of hypoglycaemia
increases in response to graded exercise in individuals with type 1 diabetes. For this study,
eight non-diabetic control male participants and seven complication-free type 1 diabetic
male individuals in good glycaemic control were recruited. On the morning of testing,
the type 1 diabetic participants followed their normal insulin regimen, and both groups
ate their usual breakfast. Then, participants were subjected to graded exercise testing
approximately four hours later. We found that this type of exercise result in a rapid post-
exercise increase in blood glucose levels (> 2 mM), which remain elevated for the first
two hours of recovery. On clinical grounds, these findings suggest for the first time that
the early post-exercise risks of hypoglycaemia associated with graded exercise testing are
minimal when performed under near basal plasma insulin levels, with no carbohydrate
administration required after testing to prevent hypoglycaemia. However, it is important

104
to stress that the risk of late onset post-exercise hypoglycaemia associated with this type
of exercise was not examined and should be investigated in future studies. In addition, it
is our view that health professionals should still regularly monitor glycaemia before,
during and after testing since it remains to be established whether similar findings would
have been obtained with different graded exercise protocols and individuals with different
circulating insulin levels.
The primary goal of the study described in Chapter 3 was to determine whether a short
10-sec maximal sprint effort is preferable to only resting as a means to counter a further
fall in glycaemia during recovery from moderate intensity exercise in hyperinsulinaemic
individuals with type 1 diabetes, thus providing another clinical tool for the prevention of
early post-exercise hypoglycaemia. To meet our objective, seven healthy complication-
free male participants with type 1 diabetes injected their normal insulin dose and ate their
usual breakfast. Then, when their postprandial glycaemia fell to ~11 mM they pedalled
at 40% peak 2OV for 20 min on a cycle ergometer followed immediately by either a
maximal 10-sec sprint or a rest. Our results show that during exercise blood glucose levels
fell rapidly. However, sprinting immediately after exercise opposes a further fall in blood
glucose levels for at least 120 min while glycaemia decreases significantly (p < 0.05) by
~ 3.5mM when no sprint was performed. Our findings also suggest that sprinting is likely
to counter the exercise-mediated decrease in blood glucose levels through an increase in
catecholamine, lactate, and growth hormone levels. These glucoregulatory benefits of
sprinting are remarkable considering the sprint trial was performed when insulin levels
were elevated, a time when exercise is not usually recommended. On the basis of these
findings, one might tentatively recommend that in order to minimise the risk of early
hypoglycaemia post-moderate intensity exercise, it is preferable for complication-free
young individuals with type 1 diabetes to engage in a 10-sec maximal sprint effort before
resting than to only rest during recovery, particularly if a source of dietary carbohydrate
is not readily available. It is important to stress, however, that our findings do not exclude
the possibility that different results might have been obtained if sprinting had been
initiated after a bout of exercise of higher intensity or longer duration, in younger or older
individuals with reduced sprinting capacity, or in individuals with impaired
counterregulatory responses. For these reasons, more studies of the kind described here
are required in order to identify the conditions and subpopulation of individuals with type
1 diabetes for whom a short maximal sprint effort can be recommended as a safe approach
for the short-term stabilisation of blood glucose levels post-exercise.

105
Given the glycaemia stabilising effect of sprinting performed after moderate-intensity
exercise (Chapter 3), this raises the issue examined in the study described in Chapter 4 of
whether performing a short sprint effort immediately prior to, rather than after, moderate-
intensity exercise may offer a novel way of preventing the rapid fall in glycaemia
normally observed both during and after moderate-intensity exercise performed under
hyperinsulinaemic conditions. In other words, this study examined whether short-
duration sprinting should be an integral part of the preparatory routine of individuals with
type 1 diabetes before they engage in sustained physical activities of moderate intensity.
To this end, seven complication-free type 1 diabetic males injected their normal morning
insulin dose and ate their usual breakfast. When post-meal glycaemia fell to ~11 mM,
they were asked to perform a 10 sec all-out sprint (sprint trial) or to rest (control trial)
immediately before cycling at 40% of peak rate of oxygen consumption for 20 min. We
found, against expectations, that sprinting for 10 sec immediately before moderate-
intensity exercise performed under hyperinsulinaemic conditions does not affect the rapid
decline in glycaemia during exercise. However, sprinting rather than resting before
moderate intensity exercise did prevent glycaemia from falling for at least the first 45 min
of recovery in individuals with type 1 diabetes. This suggests that including a short sprint
as part of the warm-up routine of individuals with type 1 diabetes before they engage in
sustained moderate-intensity exercise might provide another means of temporarily
stabilising glycaemia during early recovery. However, before recommending such a use
of sprinting, a number of issues must be addressed such as the extent to which the efficacy
of prior sprinting is affected by the intensity and duration of the subsequent bout of
exercise and the identification of any subpopulations of type 1 diabetic individuals
unlikely to benefit from sprinting.
In our attempt to identify the endocrine mechanisms mediating the glucoregulatory
benefits of sprinting, the levels of several counterregulatory hormones were measured in
the studies described in Chapters 3 and 4. Unfortunately, one difficulty with comparing
these findings with the literature has been the lack of information on the effect of short-
duration sprinting per se on the responses of counterregulatory hormones in diabetic
individuals in a hyperinsulinaemic fed state. For this reason, the purpose of the study
described in Chapter 5 was to investigate the effect of a single 10-sec sprint on the levels
of the counterregulatory hormones in type 1 diabetic individuals under hyperinsulinaemic
fed conditions approaching those reported in Studies 3 and 4. In this final study, we found

106
that performing a 10-sec maximal sprint resulted in patterns of change in plasma
catecholamines, growth hormone, cortisol and glucagon levels comparable to those
observed when a sprint is performed immediately after a bout of moderate intensity
exercise in individuals with type 1 diabetes (Study 2) and also comparable to those
observed in response to a sprint performed after an overnight fast (Fahey et al., 2012).
What remains to be established in future studies is the extent to which the changes in the
levels of each of the counterregulatory hormones examined here affects blood glucose
levels, hepatic glucose production and peripheral glucose utilisation in response to
sprinting. Overall, this final study provides the basis for more research to understand
better the mechanisms underlying the clinical benefits of sprinting in hypoglycaemia
prevention.
6.2 Clinical Implications, Limitations with our Findings and Direction for Future
Studies
The counter-intuitive findings described in this thesis that the risk of hypoglycaemia
associated with exercise can be decreased by sprinting before or after moderate intensity
exercise have the potential to cause a significant shift in the way blood glucose levels are
managed in active individuals with type 1 diabetes. Some authors have mentioned the
potential of incorporating sprints to help oppose an exercise-mediated decline in
glycaemia in young healthy adults with type 1 diabetes (Tonoli et al., 2012; Canadian
Diabetes Association Clinical Practice Guidelines Expert Committee, 2013; Robertson et
al., 2014; Yardley & Sigal, 2015). It is important to stress, however, that before
recommending the general adoption of short-duration sprinting as a safe and reliable
clinical tool for the short-term stabilisation of blood glucose levels in exercising type 1
diabetic individuals, a number of issues must be addressed as in our view it would be
premature and irresponsible at this stage to advocate its widespread adoption. Indeed, it
is of paramount importance to determine whether there are conditions likely to impair the
efficacy of sprinting in hypoglycaemia prevention. One must remember that the findings
arising from this thesis are based on data obtained from a very specific population of
healthy complication-free young adults with type 1 diabetes, and for this reason may not
apply to the wider population of individuals with type 1 diabetes. For instance, it would
be ill advised to recommend the use of a short sprint as a means to reduce the risk of
exercise-mediated hypoglycaemia for individuals with an impaired capacity to engage in
a maximal sprint effort, such as very young children or old sedentary individuals with or

107
without pre-existing medical conditions, or for diabetic individuals with advanced
neuropathy, or for individuals with diabetic complications for whom intense exercise is
contra-indicated. But what about the large population of complication-free type 1 diabetic
individuals who have the capacity to engage in an all-out sprint effort?
Even with the population of type 1 diabetic individuals who would be responsive to the
benefits of sprinting, a number of important issues must also be addressed before
recommending the adoption of short-duration sprint as a safe and reliable clinical tool for
the short-term management of blood glucose levels. For instance, it remains to be
determined whether our findings obtained on a cycle ergometer in a well-controlled
laboratory environment would extend to all modes of all-out sprinting including running
and swimming. It is also unclear whether performing an ‘all out’ sprint and reaching
maximal power output early in the sprint is important as opposed to ‘pacing’ for the
duration of the sprint. The effect of the fitness and training status of the participants on
the glucoregulatory response to sprinting also remains to be investigated. Since the rate
of fall in glycaemia during moderate intensity exercise increases with both exercise
intensity and plasma insulin levels, it is possible that the efficacy of sprinting at
preventing hypoglycaemia post-moderate intensity exercise might be reduced in response
to more intense aerobic exercise or more severe hyperinsulinaemic conditions than those
tested in this thesis. Given our findings that catecholamines and growth hormones might
mediate the effect of sprinting on glycaemia (Chapters 3, 4), this raises the possibility that
a defect in the secretion of these hormones could impair the ability of a sprint to counter
glycaemia in exercising individuals. Moreover, it remains to be established whether the
glucoregulatory benefits of sprinting increase with one’s maximal power output since the
catecholamine response to exercise increases with exercise intensity. Finally, it would be
interesting to investigate the effect of gender and phase of menstrual cycle on the
glucoregulatory responses to a single 10-sec sprint.
Several questions related to the counterregulatory status of diabetic individuals prior to
exercise must also be answered before recommending the adoption of short duration
sprinting as a safe and reliable tool for the short-term management of blood glucose
levels. For instance, the depletion of hepatic glycogen stores by a prolonged fast, and
associated changes in counterregulatory hormone levels prior to exercise might impair
the glucoregulatory benefit of sprinting. However, this prediction is not supported by the
finding that intense exercise results in comparable hyperglycaemic effect in fed and

108
fasted non-diabetic individuals (Lavoie et al., 1987). Given that the counterregulatory
response to exercise is reduced following antecedent hypoglycaemia (Galassetti et al.,
2003, 2004) or antecedent exercise performed hours earlier (Galassetti et al., 2001a), this
raises the intriguing possibility that the efficacy of sprinting as a means to prevent
glycaemia from falling post-exercise might be impaired if preceded hours earlier by
either an episode of hypoglycaemia or exercise. Such a potential detrimental effect of
antecedent hypoglycaemia on the glycaemia-stabilising effect of sprinting might also be
more pronounced in men than in women given that antecedent hypoglycaemia has been
reported to result in a relatively greater fall in the responses of counterregulatory
hormones (catecholamines, growth hormone, glucagon) and glucose production in men
compared with women in response to a subsequent prolonged bout of moderate intensity
exercise (Galassetti et al., 2004). However, the possibility that antecedent hypoglycaemia
impairs the benefit of sprinting is not supported by a study recently published from this
laboratory where a one-hour hypoglycaemia period was reported to have no effect on the
glycaemia-rising effect and counterregulatory response of a sprint performed hours later
(Davey et al., 2014).
Finally, another important clinical issue to address is whether sprinting impairs the
counterregulatory responses to a subsequent hypoglycaemia episode, thus increasing the
risk of late onset hypoglycaemia. That this might be an important issue is suggested by
the work of Galassetti and colleagues who showed that the rise in catecholamines, growth
hormone and cortisol levels in response to hypoglycaemia is impaired if preceded several
hours earlier by a bout of moderate intensity exercise, with more glucose being required
to prevent hypoglycaemia (Galassetti et al., 2001b; Sandoval et al., 2004). In addition,
this impairment is more pronounced in male compared to female participants. However,
given the short duration of the sprint investigated in this thesis and the small amount of
carbohydrate expected to be mobilised by during such a sprint, we predict that the effect
of sprinting on the counterregulatory responses to a subsequent episode of
hypoglycaemia should be minimal. This prediction is supported by recent findings from
this laboratory that a 10-sec sprint does not affect the glucose demands to maintain blood
glucose at stable levels for several hours post-exercise (Davey et al., 2013b). Overall, it is important to answer the aforementioned questions before recommending
the adoption of short duration sprinting as a safe and reliable tool for the short-term
management of blood glucose levels in individuals with type 1 diabetes as this clinical

109
tool has the potential to be integral to future glucose management guidelines aimed at
helping diabetic individuals exercise more safely and take advantage of the many
physiological and psychological health benefits of a physically active lifestyle.

110
Chapter 7
References

111
ABE, H., MINOKOSHI, Y. & SHIMAZU, T. 1993. Effect of a -adrenergic agonist,
brl35135a, on glucose uptake in rat skeletal muscle in vivo and in vitro. Journal
of Endocrinology, 139, 479-486.
AFTAB GUY, D., SANDOVAL, D., RICHARDSON, M.A., TATE, D., FLAKOLL, P.J.
& DAVIS, S.N. (2005). Differing physiological effects of epinephrine in type 1
diabetes and nondiabetic humans. American Journal of Physiology,
Endocrinology and Metabolism. 288, E178-E186.
AHREN, B. 2000. Autonomic regulation of islet hormone secretion–implications for
health and disease. Diabetologia, 43, 393-410.
AIHW 2008. Incidence of Type 1 diabetes in Australia: first results. Diabetes series 9
Cat. no. CVD 42. Canberra: Australian Institute of Health and Welfare (AIHW).
AIHW 2010. Incidence of Type 1 diabetes in Australian children 2000–2008. Diabetes
series No. 13 Cat. no. CVD 51. Canberra: Australian Institute of Health and
Welfare (AIHW).
ALLSOP, P., CHEETHAM, M., BROOKS, S., HALL, G. M. & WILLIAMS, C. 1990.
Continuous intramuscular pH measurement during the recovery from brief,
maximal exercise in man. European Journal of Applied Physiology, 59, 465-70.
ALSAHLI, M. & GERICH, J. E. 2008. Hypoglycemia in Diabetes Mellitus. In:
PORETSKY, L. (ed.) Principles of Diabetes Mellitus. 2 ed. New York: Springer.
AMERICAN DIABETES ASSOCIATION. Nutrition recommendations and
interventions for diabetes: a position statement of the American Diabetes
Association. Diabetes Care, 30 Suppl 1: S48-65, 2007.
AMIEL, S., TAMBORLANE, W., SIMONSON, D. & SHERWIN, R. 1987. Defective
glucose counterregulation after strict glycemic control of insulin-dependent
diabetes mellitus. New England Journal of Medicine, 316, 1376-1383.
AMIEL, S., SHERWIN, R., SIMONSON, D. & TAMBORLANE, W. 1988. Effect of
intensive insulin therapy on glycemic thresholds for counterregulatory hormone
release. Diabetes, 37, 901-907.
AMIEL, S.A. 2009. Hypoglycemia: From the laboratory to the clinic. Diabetes Care, 32,
1364-1371.
ASVOLD, B.O., SAND, T., HESTAD, K. & BJORGASS, M.R. 2010. Cognitive function
in type 1 diabetic adults with early exposure to severe hypoglycemia: a 16-year
follow-up study. Diabetes Care, 33, 1945-1947.

112
BANARER, S., MCGREGOR, V. P. & CRYER, P. E. 2002. Intraislet hyperinsulinemia
prevents the glucagon response to hypoglycemia despite an intact autonomic
response. Diabetes, 51, 958-965.
BANGSBO, J., NØRREGAARD, L. & THORSOE, F. 1991. Activity profile of
competition soccer. Canadian Journal of Sport Sciences, 16, 110-116.
BARTH, E., ALBUSZIES, G., BAUMGART, K., MATEJOVIC, M., WACHTER, U.,
VOGT, J., RADERMACHER, P. & CALZIA, E. 2007. Glucose metabolism and
catecholamines. Critical Care Medicine, 35, S508-S518.
BAUMANN, G. 2009. Growth hormone isoforms. Growth Hormone & IGF Research,
19, 333-340.
BELL, D. G., JACOBS, I. & ELLERINGTON, K. 2001. Effect of caffeine and ephedrine
ingestion on anaerobic exercise performance. Medicine and Science in Sports and
Exercise, 33 (8), 1399-1403.
BEN-AMI, H., NAGACHANDRAN, P., MENDELSON, A. & EDOUTE, Y. 1999.
Drug-induced hypoglycemic coma in 102 diabetic patients. Archives of Internal
Medicine, 159, 281-284.
BERGENSTAL, R., TAMBORLANE, W., AHMANN, A., BUSE, J., DAILEY, G.,
DAVIS, S., JOYCE, C., PEOPLES, T., PERKINS, B. & WELSH, J. 2010.
Effectiveness of sensor-augmented insulin-pump therapy in type 1 diabetes. New
England Journal of Medicine, 363, 311-320.
BERGER, C.M., SHARIS, P.J., BRACY, D.P., LACY, D.B. & WASSERMAN, D. H.
1994. Sensitivity of exercise-induced increase in hepatic glucose production to
glucose supply and demand. American Journal of Physiology, Endocrinology and
Metabolism. 267, E411-E421.
BERGER, M., ASSAL, J.P. & JORGENS, V. 1980. Physical exercise in the diabetic. The
importance of understanding endocrine and metabolic responses. Diabete
Metabolism, 6, 59-69.
BISHOP, D. & WRIGHT, C. 2006. A time-motion analysis of professional basketball to
determine the relationship between three activity profiles: high, medium and low
intensity and the length of the time spent on court. International Journal of
Performance Analysis in Sport, 6, 130-139.
BJORKMAN, O. & ERIKSSON, L. 1983. Splanchnic glucose metabolism during leg
exercise in 60-hour-fasted human subjects. American Journal of Physiology-
Endocrinology and Metabolism, 245, E443-E448.

113
BLACKARD, W. & HEIDINGSFELDER, S. 1968. Adrenergic receptor control
mechanism for growth hormone secretion. Journal of Clinical Investigation, 47,
1407-1414.
BOGDANIS, G.C., NEVILL, M.E. & LAKOMY, H.K. 1994. Effects of previous
dynamic arm exercise on power output during repeated maximal sprint cycling.
Journal of Sports Science, 12, 363-370.
BOGDANIS, G.C., BOOBIS, L.H., LAKOMY, H.K. & NEVILL, M.E. 1995. Recovery
of power output and muscle metabolites following 30 s of maximal sprint cycling
in man. Journal of Physiology, 482, 467-480.
BOGDANIS, G.C., NEVILL M.E, LAKOMY, H.K., GRAHAM, C.M. & LOUIS, G.
1996. Effects of active recovery on power output during repeated maximal sprint
cycling. European Journal of Applied Physiology and Occupational Physiology.
74(5), 461-469.
BOLLI, G., DE FEO, P., COMPAGNUCCI, P., CARTECHINI, M., ANGELETTI, G.,
SANTEUSANIO, F. & BRUNETTI, P. 1982. Important role of adrenergic
mechanisms in acute glucose counterregulation following insulin-induced
hypoglycemia in type I diabetes. Evidence for an effect mediated by beta-
adrenoreceptors. Diabetes, 31, 641-647.
BOLLI, G., DE FEO, P., COMPAGNUCCI, P., CARTECHINI, M., ANGELETTI, G.,
SANTEUSANIO, F., BRUNETTI, P. & GERICH, J. 1983. Abnormal glucose
counterregulation in insulin-dependent diabetes mellitus. Interaction of anti-
insulin antibodies and impaired glucagon and epinephrine secretion. Diabetes, 32,
134-141.
BOLLI, G., DE FEO, P., DE COSMO, S., PERRIELLO, G., VENTURA, M.,
BENEDETTI, M., SANTEUSANIO, F., GERICH, J. & BRUNETTI, P. 1984. A
reliable and reproducible test for adequate glucose counterregulation in type I
diabetes mellitus. Diabetes, 33, 732-737.
BOLLI, G. B. 1999. Physiology of glucose counterregulation to hypoglycemia. 1999.
Endocrinology and Metabolism Clinics of North America, 28, 467-493.
BOLLI, G. 2003. Treatment and prevention of hypoglycemia and its unawareness in type
1 diabetes mellitus. Reviews in Endocrine & Metabolic Disorders, 4, 335-341.
BONEN, A., MEGENEY, L., MCCARTHY, S., MCDERMOTT, J. & TAN, M. 1992.
Epinephrine administration stimulates GLUT4 translocation but reduces glucose
transport in muscle. Biochemical and Biophysical Research Communications,
187, 685-691.

114
BOTCAZOU, M., ZOUHAL, H., JACOB, C., GRATAS-DELAMARCHE, A.,
BERTHON, P., BENTUÉ-FERRER, D. & DELAMARCHE, P. 2006. Effect of
training and detraining on catecholamine responses to sprint exercise in
adolescent girls. European Journal of Applied Physiology, 97, 68-75.
BOTCAZOU, M., JACOB, C., GRATAS-DELAMARCHE, A., VINCENT, S.,
BENTUÉ-FERRER, D., DELAMARCHE, P. & ZOUHAL, H. 2007. Sex effect
on catecholamine responses to sprint exercise in adolescents and adults. Pediatric
Exercise Science. 19(2),132-144.
BOTTINI, P., BOSCHETTI, E., PAMPANELLI, S., CIOFETTA, M., DEL SINDACO,
P., SCIONTI, L., BRUNETTI, P. & BOLLI, G.B. 1997. Contribution of
autonomic neuropathy to reduced plasma adrenaline responses to hypoglycemia
in IDDM: evidence for a nonselective defect. Diabetes, 46, 814-823.
BOULTON, A. 2005. Management of diabetic peripheral neuropathy. Clinical Diabetes,
23, 9-15.
BOYLE, P., SCHWARTZ, N., SHAH, S., CLUTTER, W. & CRYER, P. 1988. Plasma
glucose concentrations at the onset of hypoglycemic symptoms in patients with
poorly controlled diabetes and in nondiabetics. New England Journal of Medicine,
318, 1487-1492.
BOYLE, P.J., SHAH, S.D. & CRYER, P.E. 1989. Insulin, glucagon, and catecholamines
in prevention of hypoglycemia during fasting. American Journal of Physiology-
Endocrinology and Metabolism, 256, E651-E661.
BOYLE, P.J. & CRYER, P.E. 1991. Growth hormone, cortisol, or both are involved in
defense against, but are not critical to recovery from, hypoglycemia. American
Journal of Physiology- Endocrinology and Metabolism, 260, E395-E402.
BRACKEN, R.M., LINNANE, D.M. & BROOKS, S. 2009. Plasma catecholamine and
nephrine responses to brief intermittent maximal intensity exercise. Amino Acids.
36(2), 209-217.
BRAZEAU, A.S, RABASA-LHORET, R., STRYCHAR, I. & MIRCESCU, H. 2008.
Barriers to physical activity among patients with type 1 diabetes. Diabetes Care.
31(11), 2108-2109.
BRISCOE, V.J., TATE, D.B. & DAVIS, S.N. 2007. Type 1 diabetes: exercise and
hypoglycemia. Applied Physiology Nutrition Metabolism, 32, 576-582.

115
BROOKS, S., BURRIN, J., CHEETHAM, M., HALL, G., YEO, T. & WILLIAMS, C.
1988. The responses of the catecholamines and beta-endorphin to brief maximal
exercise in man. European Journal of Applied Physiology and Occupational
Physiology, 57, 230-234.
BROOKS, S., NEVILL, M.E., MELEAGROS, L., LAKOMY, H.K., HALL, G.M.,
BLOOM, S.R. & WILLIAMS, C. 1990. The hormonal responses to repetitive
brief maximal exercise in humans. European Journal of Applied Physiology and
Occupational Physiology, 60, 144-148.
BROWNLEE, M., AIELLO, L.P., COOPER, M.E., VINIK, A.I., NESTO, R.W. &
BOULTON, A.J.M. 2008. Complications of Diabetes Mellitus. In:
KRONENBERG, H. M. (ed.) Williams Textbook of Endocrinology. 11th ed.
Philadelphia: Saunders Elsevier.
BUONO, M.J., YEAGER, J.E. & HODGDON, J.A. 1986. Plasma adrenocorticotropin
and cortisol responses to brief high-intensity exercise in humans. Journal of
Applied Physiology, 61, 1337-1339.
BUSSAU, V.A., FERREIRA, L.D., JONES, T.W. & FOURNIER, P.A. 2006. The 10-s
maximal sprint: a novel approach to counter an exercise-mediated fall in glycemia
in individuals with type 1 diabetes. Diabetes Care, 29, 601-606.
BUSSAU, V.A., FERREIRA, L.D., JONES, T.W. & FOURNIER, P.A. 2007. A 10-s
sprint performed prior to moderate-intensity exercise prevents early post-exercise
fall in glycaemia in individuals with type 1 diabetes. Diabetologia, 50, 1815-1818.
CAMACHO, R.C., GALASSETTI, P., DAVIS, S.N. & WASSERMAN, D.H. 2005.
Glucoregulation during and after exercise in health and insulin-dependent
diabetes. Exercise and Sport Sciences Reviews, 33, 17-23.
CAMPBELL, M.D., WEST, D.J., BAIN, S.C., KINGSLEY, M.I., FOLEY, P.,
KILDUFF, L., TURNER, D., GRAY, B., STEPHENS, J.W. & BRACKEN,
R.M. 2014. Simulated games activity vs continuous running exercise: A novel
comparison of the glycemic and metabolic responses in T1DM patients. Mar 4.
doi: 10.1111/sms.12192. [Epub ahead of print]
CAMPAIGNE, B.N., WALLBERG-HENRIKSSON, H. & GUNNARSSON, R. 1987.
Glucose and insulin responses in relation to insulin dose and caloric intake 12h
after acute physical exercise in men with IDDM. Diabetes Care, 10, 716-721.
CAMPAIGNE, B.N. & LAMPMAN, R.M. 1994. Exercise in the Clinical Management
of Diabetes. Champaign, IL, Human Kinetics.

116
CANADIAN DIABETES ASSOCIATION CLINICAL PRACTICE GUIDELINES
EXPERT COMMITTEE, SIGAL, R.J., ARMSTRONG, M.J., COLBY, P.,
KENNY, G.P., PLOTNIKOFF, R.C., REICHERT, S.M. & RIDDELL, M.C.
2013. Physical activity and diabetes. Canadian Journal of Diabetes, 37(1), S40-
S44.
CANDACE, M., LITVIN, M., CLARK, A.L., FISHER, S.J., 2013. Defective
counterregulation and hypoglycemia unawareness in diabetes mechanisms and
emerging treatments. Endocrinology and Metabolism Clinics of North America
42, 15–38.
CAREY, A., STEINBERG, G., MACAULAY, S., THOMAS, W., HOLMES, A.,
RAMM, G., PRELOVSEK, O., HOHNEN-BEHRENS, C., WATT, M. &
JAMES, D. 2006. Interleukin-6 increases insulin-stimulated glucose disposal in
humans and glucose uptake and fatty acid oxidation in vitro via AMP-activated
protein kinase. Diabetes, 55, 2688-2697.
CHEETHAM, M.E., WILLIAMS, C. & LAKOMY, H.K. 1985. A laboratory running
test: metabolic responses of sprint and endurance trained athletes. British Journal
of Sports Medicine,19(2):81-84.
CHERRINGTON, A., EDGERTON, D. & SINDELAR, D. 1998. The direct and indirect
effects of insulin on hepatic glucose production in vivo. Diabetologia, 41, 987-
996.
CHIAPPA, G.R., RIBEIRO, J.P, ALVES, C.N., VIEIRA, P.J., DUBAS, J., QUEIROGA,
F., BATISTA, L.D., SILVA, A.C. & NEDER, J.A. 2009. Inspiratory resistive
loading after all-out exercise improves subsequent performance European
Journal of Applied Physiology, 106, 297-303.
CHIMEN, M, KENNEDY A., NIRANTHARAKUMAR, K., PANG, T.T., ANDREWS,
R., NARENDRAN, P. 2012. What are the health benefits of physical activity in
type 1 diabetes mellitus? A literature review Diabetologia 55:542–551.
CHOKKALINGAM, K., TSINTZAS, K., SNAAR, J.E.M., NORTON, L., SOLANKY,
B., LEVERTON, E., MORRIS, P., MANSELL, P. & MACDONALD, I.A. 2007.
Hyperinsulinaemia during exercise does not suppress hepatic glycogen
concentrations in patients with type 1 diabetes: a magnetic resonance spectroscopy
study. Diabetologia 50:1921–1929.
CLUTTER, W., BIER, D., SHAH, S. & CRYER, P. 1980. Epinephrine plasma metabolic
clearance rates and physiologic thresholds for metabolic and hemodynamic
actions in man. Journal of Clinical Investigation, 66, 94-101.

117
CODERRE, L., KANDROR, K., VALLEGA, G. & PILCH, P. 1995. Identification and
characterization of an exercise-sensitive pool of glucose transporters in skeletal
muscle. Journal of Biological Chemistry, 270, 27584-27588.
COIRO, V., JOTTI, G.S., VOLPI, R., MAGOTTI, M., GALLI, P., FINARDI, L.,
MAFFEI, M., CHIODERA, P. & CASTI, A. 2004. Difference between diabetic
and nondiabetic smokers in the pituitary response to physical exercise.
Metabolism, 53, 1140-1144.
COKER, R.H., KRISHNA, M.G., LACY, D.B., BRACY, D.P. & WASSERMAN, D. H.
1997. Role of hepatic - and-adrenergic receptor stimulation on hepatic glucose
production during heavy exercise. American Journal of Physiology-
Endocrinology and Metabolism, 273, E831 - E838.
COKER, R., SIMONSEN, L., BULOW, J., WASSERMAN, D. & KJAR, M. 2001.
Stimulation of splanchnic glucose production during exercise in humans contains
a glucagon-independent component. American Journal of Physiology-
Endocrinology and Metabolism, 280, E918-E927.
COKER, R., KOYAMA, Y., DENNY, J., CAMACHO, R., LACY, D. & WASSERMAN,
D. 2002. Prevention of overt hypoglycemia during exercise. Diabetes, 51, 1310-
1318.
COKER, R.H. & KJAER, M. 2005. Glucoregulation during exercise: the role of the
neuroendocrine system. Sports Medicine, 35, 575-583.
CONNOLLY, C.C., ADKINS-MARSHALL, B.A., NEAL, D.W., PUGH, W., JASPAN,
J.B. & CHERRINGTON, A.D. (1992). Relationship between decrements in
glucose level and metabolic responses to hypoglycemia in absence of
counterregulatory hormones in the conscious dog. Diabetes. 41, 1308-1319.
COOPER, D., WASSERMAN, D., VRANIC, M. & WASSERMAN, K. 1986. Glucose
turnover in response to exercise during high-and low-FIO2 breathing in man.
American Journal of Physiology- Endocrinology and Metabolism, 251, E209-
E214.
COOPERBERG, B.A. & CRYER, P.E. 2009. Beta-cell–mediated signaling predominates
over direct alpha-cell signaling in the regulation of glucagon secretion in humans.
Diabetes Care, 32, 2275-2280.
COOPERBERG, B.A. & CRYER, P.E. 2010. Insulin reciprocally regulates glucagon
secretion in humans. Diabetes, 59, 2936-2940.

118
CORRAL, P.D., HOWLEY, E.T., HARTSELL, M., ASHRAF, M. & YOUNGER, M.S.
(1998). Metabolic effects of low cortisol during exercise in humans. Journal of
Applied Physiology. 84, 939-947.
CRYER, P. 1981. Glucose counterregulation in man. Diabetes, 30, 261-264.
CRYER, P., BINDER, C., BOLLI, G., CHERRINGTON, A., GALE, E., GERICH, J. &
SHERWIN, R. 1989. Hypoglycemia in IDDM. Diabetes, 38, 1193-1199.
CRYER, P. 1999. Symptoms of hypoglycemia, thresholds for their occurrence, and
hypoglycemia unawareness. Endocrinology & Metabolism Clinics of North
America, 28, 495-500.
CRYER, P.E. 2001. The prevention and correction of hypoglycemia. In: JEFFERSON,
L.S.C. (ed.) The Endocrine Pancreas and Regulation of Metabolism. New York:
Oxford University Press.
CRYER, P.E., DAVIS, S.N. & SHAMOON, H. 2003. Hypoglycemia in diabetes.
Diabetes Care, 26, 1902-1912.
CRYER, P.E. 2004. Diverse causes of hypoglycemia-associated autonomic failure in
diabetes. New England Journal of Medicine, 350, 2272-2279.
CRYER, P.E. 2005. Mechanisms of hypoglycemia-associated autonomic failure and its
component syndromes in diabetes. Diabetes, 54, 3592-3601.
CRYER, P.E. 2007. Hypoglycemia, functional brain failure, and brain death. Journal of
Clinical Investigation, 117, 868-870.
CRYER, P.E. 2008a. The barrier of hypoglycemia in diabetes. Diabetes, 57, 3169-3176.
CRYER, P.E. 2008b. Glucose homeostasis and hypoglycaemia. In: KRONENBERG,
H.M. (ed.) Williams Textbook of Endocrinology. 11th ed. Philadelphia: Saunders
Elsevier.
CRYER, P., AXELROD, L., GROSSMAN, A., HELLER, S., MONTORI, V. &
SEAQUIST, E. 2009. Evaluation and management of adult hypoglycemic
disorders: an Endocrine Society Clinical Practice Guideline. Journal of Clinical
Endocrinology & Metabolism, 94, 709-728.
CRYER, P.E. 2010a. Hypoglycemia in Type 1 Diabetes Mellitus. Endocrinology
&Metabolism Clinics of North America, 39, 641-654.
CRYER, P.E. 2010b. Hypoglycemia in Type 1 Diabetes Mellitus. Endocrinology &
Metabolism Clinics of North America, 39, 641-654.

119
DAGOGO-JACK, S., CRAFT, S. & CRYER, P.E. 1993. Hypoglycemia-associated
autonomic failure in insulin-dependent diabetes mellitus. Recent antecedent
hypoglycemia reduces autonomic responses to, symptoms of, and defense against
subsequent hypoglycemia. Journal of Clinical Investigation, 91, 819-828.
DAVEY, R.J., PARAMALINGAM, N., RETTERATH, A.J.. LIM, E.M., DAVIS, E.A.,
JONES, T.W. 2014. Antecedent hypoglycaemia does not diminish the glycaemia-
increasing effect and glucoregulatory responses of a 10 s sprint in people with
type 1 diabetes. Diabetologia, 57(6), 1111-1118.
DAVEY, R.J., HOWE, W., PARAMALINGAM, N. FERREIRA, L.D., DAVIS, E.A.,
FOURNIER, P.A. & JONES, T.W. 2013a. The effect of midday moderate-
intensity exercise on postexercise hypoglycemia risk in individuals with type 1
diabetes. Journal of Clinical Endocrinology and Metabolism 98(7), 2908-2914.
DAVEY, R.J., BUSSAU, V.A., PARAMALINGAM, N., FERREIRA, L.D., LIM, E.M.,
DAVIS, E.A., JONES, T.W. & FOURNIER, P.A. 2013b. A 10-s sprint performed
after moderate-intensity exercise neither increases nor decreases the glucose
requirement to prevent late-onset hypoglycemia in individuals with type 1
diabetes. Diabetes Care 36(12), 4163-4165.
DAVIDSON, A. & TREWARTHA, G. 2008. Understanding the Physiological Demands
of Netball: a time-motion investigation. International Journal of Performance
Analysis in Sport, 8, 1-17.
DAVIS, M. & SHAMOON, H. 1991. Counterregulatory adaptation to recurrent
hypoglycemia in normal humans. Journal of Clinical Endocrinology &
Metabolism, 73, 995-1001.
DAVIS, S.N., SHAVERS, C., MOSQUEDA-GARCIA, R. & COSTA, F. 1997. Effects
of differing antecedent hypoglycemia on subsequent counterregulation in normal
humans. Diabetes, 46, 1328-1335.
DAVIS, S.N., FOWLER, S. & COSTA, F. 2000a. Hypoglycemic counterregulatory
responses differ between men and women with type 1 diabetes. Diabetes, 49, 65-
72.
DAVIS, S.N., MANN, S., GALASSETTI, P., NEILL, R., TATE, D., ERTL, A. &
COSTA, F. 2000b. Effects of differing durations of antecedent hypoglycemia on
counterregulatory responses to subsequent hypoglycemia in normal humans.
Diabetes, 49, 1897-1903.

120
DAVIS, S.N., SHAVERS, C. & COSTA, F. 2000c. Differential gender responses to
hypoglycemia are due to alterations in CNS drive and not glycemic thresholds.
American Journal of Physiology- Endocrinology and Metabolism, 279, E1054-
E1063.
DAVIS, S.N., GALASSETTI, P., WASSERMAN, D.H. & TATE, D. 2000d. Effect of
antecedent hypoglycemia on subsequent counterregulatory responses to exercise.
Diabetes, 49, 73-81.
DAVIS, S.N., GALASSETTI, P., WASSERMAN, D.H. & TATE, D. 2000e. Effects of
gender on neuroendocrine and metabolic counterregulatory responses to exercise
in normal man. Journal of Clinical Endocrinology & Metabolism, 85, 224-230.
DAVIS, S.N., SHAVERS, C. & COSTA, F. (2000f). Gender-related differences in
counterregulatory responses to antecedent hypoglycaemia in normal humans.
Journal of Clinical Endocrinology and Metabolism. 85, 2148-2157.
DCCT 1993. The Diabetes Control and Complications Trial Research Group: The effect
of intensive treatment of diabetes on the development and progression of long
term complications in insulin-dependent diabetes mellitus. The New England
Journal of Medicine, 329, 977-986.
DCCT 1997. The Diabetes Control and Complications Trial Research Group:
Hypoglycemia in the diabetes control and complications trial. Diabetes, 46, 271-
286.
DE FEO, P., BOLLI, G., PERRIELLO, G., DE COSMO, S., COMPAGNUCCI, P.,
ANGELETTI, G., SANTEUSANIO, F., GERICH, J., MOTOLESE, M. &
BRUNETTI, P. 1983. The adrenergic contribution to glucose counterregulation
in type I diabetes mellitus. Dependency on A-cell function and mediation through
beta 2-adrenergic receptors. Diabetes, 32, 887-893.
DE FEO, P., PERRIELLO, G., TORLONE, E., VENTURA, M.M., FANELLI, C.,
SANTEUSANIO, F., BRUNETTI, P., GERICH, J. E. & BOLLI, G. B. 1989a.
Contribution of cortisol to glucose counterregulation in humans. American
Journal of Physiology, Endocrinology and Metabolism, 257, E35-E42.
DE FEO, P., PERRIELLO, G., TORLONE, E., VENTURA, M.M., SANTEUSANIO, F.,
BRUNETTI, P., GERICH, J.E. & BOLLI, G.B. 1989b. Demonstration of a role
for growth hormone in glucose counterregulation American Journal of
Physiology, Endocrinology and Metabolism, 256, E835-E843.

121
DE FEO, P., PERRIELLO, G., TORLONE, E., FANELLI, C., VENTURA, M.M.,
SANTEUSANIO, F., BRUNETTI, P., GERICH, J.E. & BOLLI, G.B. 1991a.
Contribution of adrenergic mechanisms to glucose counterregulation in humans.
American Journal of Physiology- Endocrinology and Metabolism, 261, E725-
E736.
DE FEO, P., PERRIELLO, G., TORLONE, E., FANELLI, C., VENTURA, M.M.,
SANTEUSANIO, F., BRUNETTI, P., GERICH, J.E. & BOLLI, G.B. 1991b.
Evidence against important catecholamine compensation for absent glucagon
counterregulation. American Journal of Physiology- Endocrinology and
Metabolism, 260, E203-12.
DEIBERT, D. & DEFRONZO, R. 1980. Epinephrine-induced insulin resistance in man.
Journal of Clinical Investigation, 65, 717-721.
DINNEEN, S.F. & RIZZA, R.A. 2001. Classification and Diagnosis of Diabetes Mellitus.
In: DEGROOT, L.J. & JAMESON, L. (eds.) Endocrinology. 4th ed. Philadelphia:
W.B. Saunders Company.
DJURHUUS, C., GRAVHOLT. C., NIELSEN, S., MENGEL, A., CHRISTIANSEN, J.,
SCHMITZ, O. & MOLLER, N. 2002. Effects of cortisol on lipolysis and regional
interstitial glycerol levels in humans. American Journal of Physiology-
Endocrinology and Metabolism, 283, E172-E177.
DJURHUUS, C., GRAVHOLT, C., NIELSEN, S., PEDERSEN, S., MOLLER, N. &
SCHMITZ, O. 2004. Additive effects of cortisol and growth hormone on regional
and systemic lipolysis in humans. American Journal of Physiology-
Endocrinology and Metabolism, 286, E488-E494.
DOCHERTY, D., WENGER, H. & NEARY, P. 1988. Time-motion analysis related to
the physiological demands of rugby. Journal of Human Movement Studies, 14,
269–277.
DOTSON, S., FREEMAN, R., FAILING, H.J. & ADLER, G.K. 2008. Hypoglycemia
increases serum interleukin-6 levels in healthy men and women. Diabetes Care,
31, 1222-1223.
DOUEN, A., RAMLAL, T., KLIP, A., YOUNG, D., CARTEE, G. & HOLLOSZY, J.
1989. Exercise-induced increase in glucose transporters in plasma membranes of
rat skeletal muscle. Endocrinology, 124, 449-454.
DUBE, M., WIESNAGEL, S.J., PRUD’HOMME, D. & LAVOIE, C. (2005). Exercise
and newer insulin: How much glucose supplement to avoid hypoglycemia.
Medicine and Science in Sports and Exercise. 37, 1276-1282.

122
DUBÉ, M.-C., LAVOIE, C. & WEISNAGEL, S.J. 2013. Glucose or intermittent high-
intensity exercise in glargine/glulisine users with T1DM. Medicine and Science
in Sports and Exercise, 45(1), 3–7.
EISENBARTH, G.S., POLONSKY, K.S. & BUSE, J.B. 2008. Type 1 Diabetes Mellitus.
In: KRONENBERG, H.M. (ed.) Williams Textbook of Endocrinology. 11th ed.
Philadelphia: Saunders Elsevier.
ESBJÖRNSSON, M., NORMAN, B., SUCHDEV. S., VIRU, M., LINDHGREN, A. &
JANSSON, E. 2009. Greater growth hormone and insulin response in women than
in men during repeated bouts of sprint exercise. Acta Physiologica (Oxf), 197,
107-115.
EVANS, M., MATYKA, K., LOMAS, J., PERNET, A., CRANSTON, I.,
MACDONALD, I. & AMIEL, S. 1998. Reduced counterregulation during
hypoglycemia with raised circulating nonglucose lipid substrates: evidence for
regional differences in metabolic capacity in the human brain? Journal of Clinical
Endocrinology & Metabolism, 83, 2952-2959.
EVANS, M., HOPKINS, D., MACDONALD, I. & AMIEL, S. 2004. Alanine infusion
during hypoglycaemia partly supports cognitive performance in healthy human
subjects. Diabetic Medicine, 21, 440-446.
FAIRCHILD, T.J., FLETCHER, S., STEELE, P., GOODMAN, C., DAWSON, B. &
FOURNIER, P.A. 2002a. Rapid carbohydrate loading after a short bout of near
maximal-intensity exercise. Medicine and Science in Sports and Exercise, 34,
980-986.
FANELLI C.G., PAMPANELLI S., EPIFANO L., RAMBOTTI A., CIOFETTA M.,
MODARELLI F., DI VINCENZO A., ANNIBALE B., LEPORE M., LALLI C.,
DEL SINDACO P., BRUNETTI P. & BOLLI G.B. 1994. Relative roles of insulin
and hypoglycaemia on induction of neuroendocrine responses to, symptoms of,
and deterioration of cognitive function in hypoglycaemia in male and female
humans. Diabetologia 37, 797–807.
FANELLI, C.G., PAMPANELLI, S., PORCELLATI, F., BARTOCII, L., SCIONTI, L.,
ROSSETTO, P. & BOLLI, G.B. 2003. Rate of fall of blood glucose and
physiological responses of counterregulatory hormones, clinical symptoms and
cognitive function to hypoglycaemia in Type 1 diabetes mellitus in the
postprandial state. Diabetologia. 46, 53-64.

123
FAHEY, A.J., PARAMALINGAM, N., DAVEY, R.J., DAVIS, E.A., JONES, T.W. &
FOURNIER, P.A. 2012. The effect of a short sprint on postexercise whole-body
glucose production and utilization rates in individuals with type 1
diabetesmellitus. Journal of Clinical Endocrinology and Metaolismb 97, 4193-
4200.
FEBBRAIO, M., HISCOCK, N., SACCHETTI, M., FISCHER, C. & PEDERSEN, B.
2004. Interleukin-6 is a novel factor mediating glucose homeostasis during
skeletal muscle contraction. Diabetes, 53, 1643-1646.
FELTBOWER. R.G., BODANSKY, H.J., PATTERSON, C.C., PARSLOW, R.C.,
STEPHENSON, C.R., REYNOLDS, C. & MCKINNEY, P.A. 2008. Acute
complications and drug misuse are important causes of death for children and young
adults with type 1 diabetes - Results from the Yorkshire Register of Diabetes in
Children and Young Adults. Diabetes Care 31:922-926.
FERRANNINI, E., DEFRONZO, R. & SHERWIN, R. 1982. Transient hepatic response
to glucagon in man: role of insulin and hyperglycemia. American Journal of
Physiology- Endocrinology and Metabolism, 242, E73-E81.
FORD, T., BERG, K., LATIN, R. & THOMAS, L. 1999. The effect of exercise intensity
on blood glucose in persons with type 1 diabetes. International Sports Journal, 3,
91-100.
FOWELIN, J., ATTVALL, S., SCHENCK, H., SMITH, U. & LAGER, I. 1991.
Characterization of the insulin-antagonistic effect of growth hormone in man.
Diabetologia, 34, 500-506.
FRANCESCATO, M., GEAT, M., FUSI, S., STUPAR, G., NOACCO, C. & CATTIN,
L. 2004. Carbohydrate requirement and insulin concentration during moderate
exercise in type 1 diabetic patients 1. Metabolism, 53, 1126-1130.
FREMION, A.S., MARRERO, D.G. & GOLDEN, M.P. 1987. Maximum oxygen uptake
determination in insulin-dependent diabetes mellitus. The Physician and Sports
Medicine, 15, 119-126.
FRYSTYK, J. 2004. Free insulin-like growth factors-measurements and relationships to
growth hormone secretion and glucose homeostasis. Growth Hormone & IGF
Research, 14, 337-375.
FUJITSUKA, N., YAMAMOTO, T., OHKUWA,T. SAITO, M. & MIYAMURA, M.
1982. Peak blood lactate after short periods of maximal treadmill running.
European Journal of Applied Physiology and Occupational Physiology.
48(3):289-296.

124
GAITANOS, G.C., WILLIAMS, C., BOOBIS, L.H. & BROOKS, S. 1993. Human
muscle metabolism during intermittent maximal exercise. Journal of Applied
Physiology. 75(2):712-719.
GALASSETTI, P., MANN, S., TATE, D., NEILL, R.A., COSTA, F., WASSERMAN,
D.H. & DAVIS, S.N. 2001a. Effects of antecedent prolonged exercise on
subsequent counterregulatory responses to hypoglycemia. American Journal of
Physiology-Endocrinology and Metabolism, 280, E908-E917.
GALASSETTI, P. & RIDDELL, M. 2013. Exercise and Type 1 Diabetes (T1DM).
Comprehensive Physiology, 3, 1309-1336.
GALASSETTI, P., MANN, S., TATE, D., NEILL, R.A., WASSERMAN, D.H. &
DAVIS, S.N. 2001b. Effect of morning exercise on counterregulatory responses
to subsequent, afternoon exercise. Journal of Applied Physiology, 91, 91-99.
GALASSETTI, P., TATE, D., NEILL, R., MORREY, S. & DAVIS, S. 2002. Effect of
gender on counterregulatory responses to euglycemic exercise in type 1 diabetes.
Journal of Clinical Endocrinology & Metabolism, 87, 5144-5150.
GALASSETTI, P., TATE, D., NEILL, R.A., MORREY, S., WASSERMAN, D.H. &
DAVIS, S.N. 2003. Effect of antecedent hypoglycemia on counterregulatory
responses to subsequent euglycemic exercise in type 1 diabetes. Diabetes, 52,
1761-1769.
GALASSETTI, P., TATE, D., NEILL, R.A., MORREY, S., WASSERMAN, D.H. &
DAVIS, S.N. 2004. Effect of sex on counterregulatory responses to exercise after
antecedent hypoglycemia in type 1 diabetes. American Journal of Physiology-
Endocrinology and Metabolism, 287, E16-E24.
GALASSETTI, P., TATE, D., NEILL, R.A., RICHARDSON, A., LEU, S.Y. & DAVIS,
S.N. 2006. Effect of differing antecedent hypoglycemia on counterregulatory
responses to exercise in type 1 diabetes. American Journal of Physiology-
Endocrinology and Metabolism, 290, E1109-E1117.
GERICH, J., KARAM, J. & FORSHAM, P. 1973a. Stimulation of glucagon secretion by
epinephrine in man. Journal of Clinical Endocrinology & Metabolism, 37, 479-
481.
GERICH, J., LANGLOIS, M., NOACCO, C., KARAM, J. & FORSHAM, P. 1973b. Lack
of glucagon response to hypoglycemia in diabetes: evidence for an intrinsic
pancreatic alpha cell defect. Science, 182, 171-3.
GERICH, J.E. 1988. Lilly lecture 1988. Glucose counterregulation and its impact on
diabetes mellitus. Diabetes, 37, 1608-1617.

125
GERICH, J.E. 2000. Physiology of glucose homeostasis. Diabetes, Obesity &
Metabolism, 2, 345-350.
GERICH, J.E. 2001. Hypoglycemia. In: DEGROOT, L.J. & JAMESON, L. (eds.)
Endocrinology. 4th ed. Philadelphia: W.B. Saunders Company.
GIBNEY, J., HEALY, M-L. & SONKSEN, P.H. (2007). The growth hormone/insulin-
like growth factor-I axis in exercise and sport. Endocrine Reviews. 28, 603-624.
GILBERT, K.L., STOKES, K.A., HALL, G.M. & THOMPSON, D. 2008. Growth
hormone responses to 3 different exercise bouts in 18- to 25- and 40- to 50-year-
old men. Applied Physiology Nutrition Metabolism. 33(4),706-712.
GLUND, S. & KROOK, A. 2008. Role of interleukin-6 signalling in glucose and lipid
metabolism. Acta Physiologica, 192, 37-48.
GODFREY, R.J., MADGWICK, Z. & WHYTE, G.P. 2003. The exercise-induced growth
hormone response in athletes. Sports Medicine, 33, 599-613.
GOLD, A., MACLEOD, K. & FRIER, B. 1994. Frequency of severe hypoglycemia in
patients with type I diabetes with impaired awareness of hypoglycemia. Diabetes
Care, 17, 697-703.
GORDON, S.E., KRAEMER, W.J., VOS, N.H., LYNCH, J.M. & KNUTTGEN, H.G.
1994. Effect of acid-base balance on the growth hormone response to acute high-
intensity cycle exercise. Journal of Applied Physiology, 76, 821-829.
GOTO, K., ISHII, N., KUROKAWA, K. & TAKAMATSU, K. 2007. Attenuated growth
hormone response to resistance exercise with prior sprint exercise. Medicine &
Science in Sports & Exercise, 39, 108-115.
GRANIER, P. MERCIER, B., MERCIER, J., ANSELME, F. & PRÉFAUT C. 1995.
Aerobic and anerobic contribution to Wingate test performance in sprint and
middle-distance runners. European Journal of Applied Physiology and
Occupational Physiology. 70 (1), 58-65. GRAVHOLT, C., SCHMITZ, O., SIMONSEN, L., BULOW, J., CHRISTIANSEN, J. &
MOLLER, N. 1999. Effects of a physiological GH pulse on interstitial glycerol in
abdominal and femoral adipose tissue. American Journal of Physiology-
Endocrinology and Metabolism, 277, E848-E854.
GROMADA, J., FRANKLIN, I. & WOLLHEIM, C. B. 2007. Alpha-cells of the
endocrine pancreas: 35 years of research but the enigma remains. Endocrine
reviews, 28, 84-116.

126
GUELFI, K.J., JONES, T.W. & FOURNIER, P.A. 2005a. The decline in blood glucose
levels is less with intermittent high-intensity compared with moderate exercise in
individuals with type 1 diabetes. Diabetes Care, 28, 1289-1294.
GUELFI, K. J., JONES, T. W. & FOURNIER, P. A. 2005b. Intermittent high-intensity
exercise does not increase the risk of early postexercise hypoglycemia in
individuals with type 1 diabetes. Diabetes Care, 28, 416-418.
GUELFI, K.J., JONES, T.W. & FOURNIER, P.A. 2007a. New insights into managing
the risk of hypoglycaemia associated with intermittent high-intensity exercise in
individuals with type 1 diabetes mellitus: implications for existing guidelines.
Sports Medicine, 37, 937-946.
GUELFI, K.J., RATNAM, N., SMYTHE, G., JONES, T.W. & FOURNIER, P.A. 2007b.
Effect of intermittent high-intensity compared with continuous moderate exercise
on glucose production and utilization in individuals with type 1 diabetes.
American Journal of Physiology- Endocrinology and Metabolism, 292, E865-
E870.
HAMILTON, A., NEVILL, M., BROOKS, S. & WILLIAMS, C. 1991. Physiological
responses to maximal intermittent exercise: differences between endurance-
trained runners and games players. Journal of Sports Sciences, 9, 371-382.
HARDMAN, A.E., WILLIAMS, C. & WOOTTON, S.A. 1986. The influence of short-
term endurance training on maximum oxygen uptake, submaximum endurance
and the ability to perform brief, maximal exercise. Journal of Sports Science., 4,
109-16.
HARGREAVES, M. & SPRIET, L. 2006. Exercise Metabolism. 2nd edition ed.
Champaign, IL: Human Kinetics.
HARMER, A. R. & McKENNA, M.J, SUTTON, J.R.,SNOW, R.J., RUELL, P.A.,
BOOTH, J., THOMPSON, M.W., MACKAY, N.A., STATHIS, C.G.,
CRAMERI, R.M., CAREY, M.F. & EAGER, D.M. 2000. Skeletal muscle
metabolic and ionic adaptations during intense exercise following sprint training
in humans. Journal of Applied Physiology. 89(5),1793-1803.
HARMER, A. R., RUELL PA, McKENNA, M.J., CHISHOLM, D.J., HUNTER, S.K.,
THOM, J.M., MORRIS, N.R. & FLACK, J.R. 2006. Effects of sprint training on
extrarenal potassium regulation with intense exercise in Type 1 diabetes. Journal
of Applied Physiology, 100(1):26-34.

127
HARMER, A.R., CHISHOLM, D.J., McKENNA, M.J., MORRIS, N.R., THOM, J.M.,
BENNETT, G., & FLACK, J.R. (2007). High-intensity training improves plasma
glucose and acid-base regulation during intermittent maximal exercise in type 1
diabetes. Diabetes Care. 30, 1269-1271.
HARMER, A. R., CHISHOLM, D.J., McKENNA, M.J., HUMTER, S.K., RUELL, P.A.,
NAYLOR, J.M., MAXWELL, L.J. & FLACK, J.R. 2008. Sprint training
increases muscle oxidative metabolism during high-intensity exercise in patients
with type 1 diabetes. Diabetes Care, 31 (11), 2097-3102.
HAUGE-EVANS, A., KING, A., CARMIGNAC, D., RICHARDSON, C., ROBINSON,
I., LOW, M., CHRISTIE, M., PERSAUD, S. & JONES, P. 2009. Somatostatin
secreted by islet-cells fulfills multiple roles as a paracrine regulator of islet
function. Diabetes, 58, 403-411.
HAVEL, P., AKPAN, J., CURRY, D., STERN, J., GINGERICH, R. & AHREN, B. 1993.
Autonomic control of pancreatic polypeptide and glucagon secretion during
neuroglucopenia and hypoglycemia in mice. American Journal of Physiology-
Regulatory, Integrative and Comparative Physiology, 265, 246-256.
HAYWOOD, S., BREE, A., PUENTE, E., DAPHNA-IKEN, D. & FISHER, S. 2009.
Central but not systemic lipid infusion augments the counterregulatory response
to hypoglycemia. American Journal of Physiology- Endocrinology and
Metabolism, 297, E50-E56.
HELLER, S.R. & CRYER, P.E. 1991. Reduced neuroendocrine and symptomatic
responses to subsequent hypoglycemia after 1 episode of hypoglycemia in
nondiabetic humans. Diabetes, 40, 223-226.
HELLER, S. R. 2011. Hypoglycaemia: Its pathophysiology in insulin treated diabetes and
hypoglycaemia unawareness. The British Journal of Diabetes & Vascular
Disease, 11, S6-S9.
HEPBURN, D., DEARY, I., FRIER, B., PATRICK, A., QUINN, J. & FISHER, B. 1991.
Symptoms of acute insulin-induced hypoglycemia in humans with and without
IDDM. Factor-analysis approach. Diabetes Care, 14, 949-957.
HERMANSEN, L., PRUETT, E.D., OSNES, J.B. & GIERE, F.A. 1970. Blood glucose
and plasma insulin in response to maximal exercise and glucose infusion. Journal
of Applied Physiology. 29(1),13-16.

128
HERZOG, R.I., MCNAY, E.C., MCCRIMMON, R., YECKEL, C.W., ZHU, W.L.,
DING, Y.Y., WANG, A.J., FAN, X.N. & SHERWIN, R.S. 2006. Effect of acute
and recurrent hypoglycemia on regional changes in brain glycogen concentration.
Diabetes 55: A149-149.
HEYMAN, E. DELAMARCHE, P., BERTHON, P., MEEUSEN, R., BRIARD, D.,
VINCENT, S., DEKERDANET, M. & DELAMARCHE, A. 2007. Alteration in
sympathoadrenergic activity at rest and during intense exercise despite normal
aerobic fitness in late pubertal adolesecent girls with type 1 diabetes. Diabetes
and Metabolism, 33, 422-429.
HIRSCH, B. & SHAMOON, H. 1987. Defective epinephrine and growth hormone
responses in type I diabetes are stimulus specific. Diabetes, 36, 20-26.
HIRSCH, I., MARKER, J., SMITH, L., SPINA, R., PARVIN, C., HOLLOSZY, J. &
CRYER, P. 1991. Insulin and glucagon in prevention of hypoglycemia during
exercise in humans. American Journal of Physiology- Endocrinology and
Metabolism, 260, E695-E704.
HOELZER, D., DALSKY, G., CLUTTER, W., SHAH, S., HOLLOSZY, J. & CRYER,
P. 1986a. Glucoregulation during exercise: hypoglycemia is prevented by
redundant glucoregulatory systems, sympathochromaffin activation, and changes
in islet hormone secretion. Journal of Clinical Investigation, 77, 212-221.
HOELZER, D.R., DALSKY, G.P., SCHWARTZ, N.S., CLUTTER, W.E., SHAH, S.D.,
HOLLOSZY, J.O. & CRYER, P.E. 1986b. Epinephrine is not critical to
prevention of hypoglycemia during exercise in humans. American Journal of
Physiology- Endocrinology and Metabolism, 251, E104-E110.
HOENE, M. & WEIGERT, C. 2008. The role of interleukin-6 in insulin resistance, body
fat distribution and energy balance. Obesity Reviews, 9, 20-29.
HOLLOSZY, J.O. 2005. Exercise-induced increase in muscle insulin sensitivity. Journal
of Applied Physiology, 99, 338-343.
HOLMYARD, D., CHEETHAM, M., LAKOMY, H. & WILLIAMS, C. 1988. Effect of
recovery duration on performance during multiple treadmill sprints. In: T. ReiUy,
A. Lees, K. Davids, and W.J, Murphy (eds.). Science and Football, London/New
York: E. & F.N. Spon, Science and Football, 134-141.
HONG, J., ABUDULA, R., CHEN, J., JEPPESEN, P., DYRSKOG, S., XIAO, J.,
COLOMBO, M. & HERMANSEN, K. 2005. The short-term effect of fatty acids
on glucagon secretion is influenced by their chain length, spatial configuration,
and degree of unsaturation: studies in vitro. Metabolism, 54, 1329-1336.

129
HORTON, T.J., MILLER, E.K., GLUECK, D. & TENCH, K. 2002. No effect of
menstrual cycle phase on glucose kinetics and fuel oxidation during moderate-
intensity exercise. American Journal of Physiology-Endocrinology and
Metabolism, 282, E752-E762.
HOWLETT, K., FEBBRAIO, M. & HARGREAVES, M. 1999a. Glucose production
during strenuous exercise in humans: role of epinephrine. American Journal of
Physiology, 276, E1130 - E1135.
HOWLETT, K., GALBO, H., LORENTSEN, J., BERGERON, R., ZIMMERMAN
BELSING, T., BÜLOW, J., FELDT RASMUSSEN, U. & KJAER, M. 1999b.
Effect of adrenaline on glucose kinetics during exercise in adrenalectomised
humans. The Journal of Physiology, 519, 911-921.
HÜBINGER, A., RIDDERSKAMP, I., LEHMANN, E. & GRIES, F. 1985. Metabolic
response to different forms of physical exercise in type I diabetics and the duration
of the glucose lowering effect. European Journal of Clinical Investigation, 15,
197-203.
HVIDBERG, A., FANELLI, C., HERSHEY, T., TERKAMP, C., CRAFT, S. & CRYER,
P. 1996. Impact of recent antecedent hypoglycemia on hypoglycemic cognitive
dysfunction in nondiabetic humans. Diabetes, 45, 1030-1036.
IDF. 2009. Diabetes atlas 4th edition. Brussels: International Diabetes Federation.
INBAR, O., BAR-OR, O. & SKINNER, J.S. 1996. The Wingate anaerobic test,
Champaign, Human Kinetics.
ISCOE, K. & RIDDELL, M. 2011. Continuous moderate intensity exercise with or
without intermittent high intensity work: effects on acute and late glycaemia in
athletes with Type 1 diabetes mellitus. Diabetic Medicine, 28, 824-832.
ITO, K., MARUYAMA, H., HIROSE, H., KIDO, K., KOYAMA, K., KATAOKA, K. &
SARUTA, T. 1995. Exogenous insulin dose-dependently suppresses glucopenia-
induced glucagon secretion from perfused rat pancreas. Metabolism, Clinical and
Experimental, 44, 358-362.
JACOB, C., ZOUHAL, H, GRATAS-DELAMARCHE, A., BERTHON, P.M.,
BENTUÉ-FERRER, D. & DELAMARCHE, P. 2002. Training status (endurance
or sprint) and catecholamine response to the Wingate-test in women. International
Journal of Sports Medicine. 23(5):342-7.

130
JACOB, C., ZOUHAL, H., PRIOUZ, J., GRATAS-DELAMARCHE, A, BENTUÉ-
FERRER, D. & DELAMARCHE, P. 2004. Effect of the intensity of training on
catecholamine responses to supramaximal exercise in endurance-trained men.
European Journal of Applied Physiology. 91(1),35-40.
JACOBSON, A.M., MUSEN, G., RYAN, C.M., SILVERS, N., CLEARY, P.,
WABERSKI, B., BURWOOD, A., WEINGER, K., BAYLESS, M., DAHMS, W.
& HARTH, J. 2007. Long-term effect of diabetes and its treatment on cognitive
function. New England Journal of Medicine, 356:1842-1852.
JENKINS, A., CHISHOLM, D., JAMES, D., HO, K. & KRAEGEN, E. 1985. Exercise-
induced hepatic glucose output is precisely sensitive to the rate of systemic
glucose supply. Metabolism, 34, 431-436.
JENSEN, T.E. & RICHTER, E.A. 2012. Regulation of glucose and glycogen metabolism
during and after exercise. The Journal of Physiology, 590, 1069-1076.
JEUKENDRUP, A., WAGENMAKERS, A., STEGEN, J., GIJSEN, A., BROUNS, F. &
SARIS, W. 1999. Carbohydrate ingestion can completely suppress endogenous
glucose production during exercise. American Journal of Physiology-
Endocrinology and Metabolism, 276, E672-E683.
JIANG, G. & ZHANG, B. 2003. Glucagon and regulation of glucose metabolism.
American Journal of Physiology-Endocrinology and Metabolism, 284, E671-678.
JONES, N. L. MCCARTNEY, N., GRAHAM, T., SPRIET, L.L., KOWALCHUK, J.M.,
HEIGENHAUSER, G.J. & SUTTON, J.R.1985. Muscle performance and
metabolism in maximal isokinetic cycling at slow and fast speeds. Journal of
Applied Physiology 59(1),132-136.
KANALEY, J. A., DALL, R., MØLLER, N., NIELSEN, S.C., CHRISTIANSEN, J.S.,
JENSEN, M.D. & JØRGENSEN, J.O.L. 2004. Acute exposure to GH during
exercise stimulates the turnover of free fatty acids in GH-deficient men. Journal
of Applied Physiology, 96, 747–753.
KASHIWAGURA, T., ERECINSKA, M. & WILSON, D. 1985. pH dependence of
hormonal regulation of gluconeogenesis and urea synthesis from glutamine in
suspensions of hepatocytes. Journal of Biological Chemistry, 260, 407-414.
KINDERMANN, W. , SCHNABEL A, SCHMITT, W.M., BIRO, G., CASSENS, J. &
WEBER, F. 1982. Catecholamines, growth hormone, cortisol, insulin, and sex
hormones in anaerobic and aerobic exercise. European Journal of Applied
Physiology and Occupational Physiology. 49(3), 389-399.

131
KING, N., BURLEY, V. & BLUNDELL, J. 1994. Exercise-induced suppression of
appetite: effects on food intake and implications for energy balance. European
Journal of Clinical Nutrition, 48, 715-724.
KJAER, M., FARRELL, P.A., CHRISTENSEN, N.J. & GALBO, H. 1986. Increased
epinephrine response and inaccurate glucoregulation in exercising athletes.
Journal of Applied Physiology, 61, 1693-1700.
KJAER, M., SECHER, N.H., BACH, F.W., SHEIKH, S. & GALBO, H. 1989. Hormonal
and metabolic responses to exercise in humans: effect of sensory nervous
blockade. American Journal of Physiology- Endocrinology and Metabolism, 257,
E95-E101.
KJAER, M., KIENS, B., HARGREAVES, M. & RICHTER, E. 1991. Influence of active
muscle mass on glucose homeostasis during exercise in humans. Journal of
Applied Physiology, 71, 552-557.
KJAER, M., ENGFRED, K., FERNANDES, A., SECHER, N. & GALBO, H. 1993a.
Regulation of hepatic glucose production during exercise in humans: role of
sympathoadrenergic activity. American Journal of Physiology- Endocrinology
and Metabolism, 265, E275-E283.
KJAER, M., ENGFRED, K., FERNANDES, A., SECHER, N.H. & GALBO, H. 1993b.
Regulation of hepatic glucose production during exercise in humans: role of
sympathoadrenergic activity. American Journal of Physiology, 265, E275-283.
KJAER, M., KEIDING, S., ENGFRED, K., RASMUSSEN, K., SONNE, B.,
KIRKEGARD, P. & GALBO, H. 1995. Glucose homeostasis during exercise in
humans with a liver or kidney transplant. American Journal of Physiology-
Endocrinology and Metabolism, 268, E636-E644.
KJÆR, M., POLLACK, S., MOHR, T., WEISS, H., GLEIM, G., BACH, F.,
NICOLAISEN, T., GALBO, H. & RAGNARSSON, K. 1996. Regulation of
glucose turnover and hormonal responses during electrical cycling in tetraplegic
humans. American Journal of Physiology-Regulatory, Integrative and
Comparative Physiology, 271, R191-R199.
KOIVISTO, V.A. & FELIG, P. 1978. Effects of leg exercise on insulin absorption in
diabetic patients. New England Journal of Medicine, 298, 79-83.
KREISMAN, S.H., MANZON, A., NESSIM, S.J., MORAIS, J.A., GOUGEON, R.,
FISHER, S.J., VRANIC, M. & MARLISS, E.B. 2000a. Glucoregulatory
responses to intense exercise performed in the postprandial state. American
Journal of Physiology-Endocrinology and Metabolism, 278, E786 - E793.

132
KREISMAN, S. H., MEW, N. A. H., ARSENAULT, M., NESSIM, S. J., HALTER, J.
B., VRANIC, M. & MARLISS, E. B. 2000b. Epinephrine infusion during
moderate intensity exercise increases glucose production and uptake. American
Journal of Physiology-Endocrinology and Metabolism, 278, E949 - E957.
KREISMAN, S.H., MEW, N.A.H., HALTER, J.B., VRANIC, M. & MARLISS, E. B.
2001. Norepinephrine infusion during moderate-intensity exercise increases
glucose production and uptake. Journal of Clinical Endocrinology & Metabolism,
86, 2118-2124.
KREISMAN, S., HALTER, J., VRANIC, M. & MARLISS, E. 2003. Combined infusion
of epinephrine and norepinephrine during moderate exercise reproduces the
glucoregulatory response of intense exercise. Diabetes, 52, 1347-1354.
KRISTIANSEN, S., WOJTASZEWSKI, J.F., JUEL, C. & RICHTER, E.A. 1994. Effect
of glucose-6-phosphate and pH on glucose transport in skeletal muscle plasma
membrane giant vesicles. Acta Physiologica Scandinavica, 150, 227-233.
LAAKSONEN, D.E., ATALAY, M., NISKANEN, L.K., MUSTONEN, J., SEN, C. K.,
LAKKA, T.A. & UUSITUPA, M.I. 2000. Aerobic exercise and the lipid profile
in type 1 diabetic men: a randomized controlled trial. Medicine and Science in
Sports and Exercise, 32, 1541-1548.
LAGER, I., ATTVALL, S., ERIKSSON, B., SCHENK, H. & SMITH, U. 1986. Studies
on the insulin-antagonistic effect of catecholamines in normal man. Diabetologia,
29, 409-416.
LAING, S., SWERDLOW, A., SLATER, S., BOTHA, J., BURDEN, A., WAUGH, N.,
SMITH, A., HILL, R., BINGLEY, P. & PATTERSON, C. 1999. The British
Diabetic Association Cohort Study, I: all cause mortality in patients with insulin
treated diabetes mellitus. Diabetic Medicine, 16, 459-465.
LAM, T., POCAI, A., GUTIERREZ-JUAREZ, R., OBICI, S., BRYAN, J., AGUILAR-
BRYAN, L., SCHWARTZ, G. & ROSSETTI, L. 2005. Hypothalamic sensing of
circulating fatty acids is required for glucose homeostasis. Nature Medicine, 11,
320-327.
LANGFORT, J., ZARZECZNY, R., PILIS, W., NAZAR, K. & KACIUBA-UŚCITKO,
H. 1997. The effect of a low-carbohydrate diet on performance, hormonal and
metabolic responses to a 30-s bout of supramaximal exercise. European Journal
of Applied Physiology, 76, 128-133.

133
LANZI, R., MANZONI, M., ANDREOTTI, A., MALIGHETTI, M., BIANCHI, E.,
SERENI, L. P., CAUMO, A., LUZI, L. & PONTIROLI, A. 1997. Evidence for
an inhibitory effect of physiological levels of insulin on the growth hormone (GH)
response to GH-releasing hormone in healthy subjects. Journal of Clinical
Endocrinology & Metabolism, 82, 2239-2243.
LAURENT, C.M., MEYERS, M.C., ROBINSON, C.A. & GREEN, J.M. 2007. Cross-
validation of the 20- versus 30-s Wingate anaerobic test. European Journal of
Applied Physiology, 100, 645-651.
LAVOIE, J.M., BONNEAU, M.C., ROY, J.Y., BRISSON, G.R. & HÉLIE, R. 1987.
Effects of dietary manipulations on blood glucose and hormonal responses
following supramaximal exercise. Europoean Journal of Physiology and
Occupational Physiology, 56, 109-114.
LAVOIE, C., DUCROS, F., BOURQUE, J., LANGELIER, H. & CHIASSON, J.L. 1997.
Glucose metabolism during exercise in man: the role of insulin and glucagon in
the regulation of hepatic glucose production and gluconeogenesis. Canadian
Journal of Physiology and Pharmacology. 75(1), 36-43.
LECAVALIER, L., BOLLI, G., CRYER, P. & GERICH, J. 1989. Contributions of
gluconeogenesis and glycogenolysis during glucose counterregulation in normal
humans. American Journal of Physiology- Endocrinology and Metabolism, 256,
E844-851.
LEMBO, G., CAPALDO, B., RENDINA, V., IACCARINO, G., NAPOLI, R., GUIDA,
R., TRIMARCO, B. & SACCA, L. 1994. Acute noradrenergic activation induces
insulin resistance in human skeletal muscle. American Journal of Physiology-
Endocrinology and Metabolism, 266, E242-E247.
LERNMARK, A. 2001. Type 1 (Insulin-Dependent) Diabetes Mellitus: Etiology,
Pathogenesis, and Natural History. In: DEGROOT, L.J. & JAMESON, L. (eds.)
Endocrinology. Philadelphia: W.B. Saunders Company.
LINS, P., WAJNGOT, A., ADAMSON, U., VRANIC, M. & EFENDI , S. 1983. Minimal
increases in glucagon levels enhance glucose production in man with partial
hypoinsulinemia. Diabetes, 32, 633-636.
LITTLE, J.P., SAFDAR, A.S., WILKIN, G.P., TARNOPOLSKY, M.A. & GIBALA,
M.J. 2010. A practical model of low-volume high-intensity interval training
induces mitochondrial biogenesis in human skeletal muscle: potential
mechanisms. Journal of Physiology, 588, 1011-1022.

134
LUDVIGSSON, J., LARSSON, Y. & SVENSSON, P.G. 1980. Attitudes towards
physical exercise in juvenile diabetes. Acta Paediatrica Scandinavica, 283, Suppl.
106-111.
LIU, Y. & STOCK, M.J. Acute effect of the -adrenoceptor agonist, BRL35135, on
tissue glucose utilisation. 1995. British Journal of Pharmacology, 114, 888-894.
MACDONALD, M.J. 1987a. Postexercise late-onset hypoglycemia in insulin-dependent
diabetic patients. Diabetes Care, 10, 584-588.
MACDONALD, I. & KING, P. (eds.) 2007. Normal glucose metabolism and response to
hypoglycaemia, Chichester, UK: John Wiley & Sons Ltd.
MACRAE, H.H., NOAKES, T.D. & DENNIS, S.C. 1995. Effects of endurance training
on lactate removal by oxidation and gluconeogenesis during exercise. Pflügers
Archives 430: 964–970.
MANZON, A., FISHER, S.J., MORAIS, J.A., LIPSCOMBE, L., GUIMOND, M.C.,
NESSIM, S.J., SIGAL, R.J., HALTER, J.B., VRANIC, M. & MARLISS, E.B.
1998. Glucose infusion partially attenuates glucose production and increases
uptake during intense exercise. Journal of Applied Physiology, 85, 511-524.
MARAN, A., CRANSTON, I., LOMAS, J., AMIEL, S. & MACDONALD, I. 1994.
Protection by lactate of cerebral function during hypoglycaemia. The Lancet, 343,
16-20.
MARAN, A., PAVAN, P., BONSEMBIANTE, B., BRUGIN, E., ERMOLAO, A.,
AVOGARO, A. & ZACCARIA, M. 2010. Continuous glucose monitoring reveals
delayed nocturnal hypoglycemia after intermittent high-intensity exercise in
nontrained patients with type 1 diabetes. Diabetes Technology & Therapeutics,
12, 763-768.
MARKER, J., HIRSCH, I., SMITH, L., PARVIN, C., HOLLOSZY, J. & CRYER, P.
1991. Catecholamines in prevention of hypoglycemia during exercise in humans.
American Journal of Physiology- Endocrinology and Metabolism, 260, E705-
E712.
MARLISS, E.B., SIMANTIRAKIS, E., MILES, P.D.G., PURDON, C., GOUGEON, R.,
FIELD, C.J., HALTER, J.B. & VRANIC, M. 1991. Glucoregulatory and
hormonal responses to repeated bouts of intense exercise in normal male subjects.
Journal of Applied Physiology, 71, 924 –933.

135
MARLISS, E.B., PURDON, C., MILES, P.D.G., HALTER, J.B., SIGAL, R.J. &
VRANIC, M. 1992a. Glucoregulation during and after intense exercise in control
and diabetic subjects. In: DELVIN J. (ed.) Diabetes Mellitus and Exercise.
London: Smith Gordon.
MARLISS, E.B., SIMANTIRAKIS, E., MILES, P.D.G., HUNT, R., GOUGEON, R.,
PURDON, C., HALTER, J.B. & VRANIC, M. 1992b. Glucose turnover and its
regulation during intense exercise and recovery in normal male subjects. Clinical
and Investigative Medicine., 15, 406-419.
MARLISS, E.B., KREISMAN, S.H., MANZON, A., HALTER, J.B., VRANIC, M. &
NESSIM, S.J. 2000. Gender differences in glucoregulatory responses to intense
exercise. Journal of Applied Physiology, 88, 457-466.
MARLISS, E.B. & VRANIC, M. 2002. Intense exercise has unique effects on both insulin
release and its roles in glucoregulation: implications for diabetes. Diabetes, 51,
S271- S283.
MARQUARDT, J., BACHARACH, D. & KOTLY, J. 1993. Predicting 30 Second
Minimum Power From A 20 Second Wingate Test. Medicine & Science in Sports
& Exercise, 25, S108.
MCAULAY, V., DEARY, I. & FRIER, B. 2001. Symptoms of hypoglycaemia in people
with diabetes. Diabetic Medicine, 18, 690-705.
MCCRIMMON, R. 2009. Glucose sensing during hypoglycemia: lessons from the lab.
Diabetes Care, 32, 1357-1363.
MCMAHON, M., GERICH, J. & RIZZA, R. 1988. Effects of glucocorticoids on
carbohydrate metabolism. Diabetes/Metabolism Reviews, 4, 17-30.
MCMAHON, S., FERREIRA, L., RATNAM, N., DAVEY, R., YOUNGS, L., DAVIS,
E., FOURNIER, P.A. & JONES, T.W. 2007. Glucose requirements to maintain
euglycemia after moderate-intensity afternoon exercise in adolescents with type 1
diabetes are increased in a biphasic manner. Journal of Clinical Endocrinology &
Metabolism, 92, 963-968.
MEYER, C., GROSSMANN, R., MITRAKOU, A., MAHLER, R., VENEMAN, T.,
GERICH, J. & BRETZEL, R. 1998. Effects of autonomic neuropathy on
counterregulation and awareness of hypoglycemia in type 1 diabetic patients.
Diabetes Care, 21, 1960-1966.
MEYER, C., DOSTOU, J. & GERICH, J. 1999. Role of the human kidney in glucose
counterregulation. Diabetes, 48, 943-948.

136
MEYER, C., SAAR, P., SOYDAN, N., ECKHARD, M., BRETZEL, R., GERICH, J. &
LINN, T. 2005. A potential important role of skeletal muscle in human
counterregulation of hypoglycemia. Journal of Clinical Endocrinology &
Metabolism, 90, 6244-6250.
MIGRENNE, S., MAGNAN, C. & CRUCIANI-GUGLIELMACCI, C. 2007. Fatty acid
sensing and nervous control of energy homeostasis. Diabetes & metabolism, 33,
177-182.
MILLER, B.F., FATTOR, J.A., JACOBS, K.A., HORNING, M.A., NAVAZIO, F.,
LINDINGER, M.I. & BROOKS, G.A. 2002. Lactate and glucose interactions
during rest and exercise in men: effect of exogenous lactate infusion. Journal of
Physiology, 544, 963-975.
MITCHELL, T.H., ABRAHAM, G., SCHIFFRIN, A., LEITER, L.A. & MARLISS, E.
B. 1988. Hypoglycemia after intense exercise in IDDM subjects during
continuous subcutaneous insulin infusion. Diabetes Care, 11, 311 - 317.
MITRAKOU, A., RYAN, C., VENEMAN, T., MOKAN, M., JENSSEN, T., KISS, I.,
DURRANT, J., CRYER, P. & GERICH, J. 1991. Hierarchy of glycemic
thresholds for counterregulatory hormone secretion, symptoms, and cerebral
dysfunction. American Journal of Physiology- Endocrinology and Metabolism,
260, E67-74.
MOLLER, N., JØRGENSEN, J.O.L., SCHMITZ, O., MØLLER, J., CHRISTIANSEN, J.
S., ALBERTI, K.G.M.M. & ØRSKOV, H. 1990. Effects of a growth hormone
pulse on total and forearm substrate fluxes in humans. American Journal of
Physiology-Endocrinology and Metabolism, 258, E86-E91.
MØLLER, N., SCHMITZ, O., MØLLER, J. & BUTLER, P.C. 1992a. Effects of a
physiological growth hormone pulse on substrate metabolism in insulin-
dependent (Type 1) diabetic subjects. Journal of Clinical Endocrinology and
Metabolism, 75, 432-436.
MØLLER, N., SCHMITZ, O., PØRKSEN, N., MØLLER, J. & JØRGENSEN, J.O.L.
1992b. Does-response studies on the metabolic effects of a growth hormone pulse
in humans. Metabolism, 41, 172-175.
MØLLER, N., GJEDSTED, J., GORMSEN, L., FUGLSANG, J. & DJURHUUS, C.
2003. Effects of growth hormone on lipid metabolism in humans. Growth
Hormone & IGF Research, 13, S18-S21.
MOORE, M.C., CONNOLLY, C.C. & CHERRINGTON, A.D. 1998. Autoregulation of
hepatic glucose production. European Journal of Endocrinology, 138, 240-248.

137
MOUSSA, E., ZOUHAL, H., VINCENT, S., PROIUX, J., DELAMARCHE, P. &
GRATAS-DELAMARCHE, A. 2003. Effect of sprint duration (6 s or 30 s) on
plasma glucose regulation in untrained male subjects. Journal of Sports Medicine
and Physical Fitness, 43, 546-53.
MOY, C.S., SONGER, T.J., LAPORTE, R.E., DORMAN, J.S., KRISKA, A.M.,
ORCHARD, T.J., BECKER, D.J. & DRASH, A.L. 1993. Insulin-dependent
diabetes mellitus, physical activity, and death. American Journal of
Epidemiology, 137, 74-81.
MYERS, S.R., DIAMOND, M.P., ADKINS-MARSHALL, B.A., WILLIAMS, P.E.,
STINSEN, R. & CHERRINGTON, A.D. 1991. Effects of small changes in
glucagon on glucose production during a euglycemic, hyperinsulinemic clamp.
Metabolism, 40, 66-71.
NATHAN, D., ZINMAN, B., CLEARY, P., BACKLUND, J., GENUTH, S., MILLER,
R. & ORCHARD, T. 2009. Modern-day clinical course of type 1 diabetes mellitus
after 30 years’ duration: the diabetes control and complications trial/epidemiology
of diabetes interventions and complications and Pittsburgh epidemiology of
diabetes complications experience (1983-2005). Archives of Internal Medicine,
169, 1307-1316.
NÄVERI, H., KUOPPASALMI, K. & HÄRKÖNEN, M. 1985. Plasma glucagon and
catecholamines during exhaustive short-term exercise. European Journal of
Applied Physiology and Occupational Physiology, 53, 308-311.
NEVILL, M.E., BOOBIS, L.H, BROOKS, S. & WILLIAMS, C. 1989. Effect of training
on muscle metabolism during treadmill sprinting. Journal of Applied Physiology.
67(6),2376-2382.
NEVILL, M.E., WILLIAMS, C., ROPER, D., SLATER, C. & NEVILL, A.M. 1993.
Effect of diet on performance during recovery from intermittent sprint exercise. J
Sports Sci, 11, 119-126.
NEVILL, M.E., HOLMYARD, D.J., HALL, G.M., ALLSOP, P., VAN OOSTERHOUT,
A., BURRIN, J.M. & NEVILL, A.M. 1996. Growth hormone responses to
treadmill sprinting in sprint- and endurance-trained athletes. European Journal of
Applied Physiology and Occupational Physiology, 72, 460-467.
NGALA, R.A. & O'DOWD, J.F., STOCKER, C.J., CAWTHORME, M.A. & ARCH
J.R.S. 2013. -adrenoceptor agonists can both stimulate and inhibit glucose
uptake in mouse soleus muscle through ligand-directed signalling. Naunyn-
Schmiedeberg's Archives of Pharmacology, 386, 761-773.

138
NONOGAKI, K. 2000. New insights into sympathetic regulation of glucose and fat
metabolism. Diabetologia, 43, 533-549.
NORRIS, R., CARROLL, D. & COCHRANE, R. 1990. The effects of aerobic and
anaerobic training on fitness, blood pressure, and psychological stress and well-
being. Journal of Psychosomatic Research, 34, 367-375.
NORTHAM, E.A., RANKINS, D., LIN, A., WELLARD, R.M., PELL, G.S., FINCH,
S.J., WERTHER, G.A. & CAMERON FJ. 2009. Central nervous system function
in youth with type 1 diabetes 12 years after disease onset. Diabetes Care. 32, 445-
450.
NUGENT, A. M., STEELE, I. C., AL-MODARIS, F., VALLELY, S., MOORE, A.,
CAMPBELL, N.P.S., BELL, P.M., BUCHANAN, K.D., TRIMBLE, E.R. &
NICHOLLS, D.P. 1997. Exercise responses in patients with IDDM. Diabetes
Care, 20, 1814 -1821.
OBICI, S., FENG, Z., ARDUINI, A., CONTI, R. & ROSSETTI, L. 2003. Inhibition of
hypothalamic carnitine palmitoyltransferase-1 decreases food intake and glucose
production. Nature Medicine, 9, 756-761.
OHKUWA,T., KATO Y, KATSUMATA, K., NAKAO, T. & MIYAMURA, M. 1984.
Blood lactate and glycerol after 400-m and 3,000-m runs in sprint and long
distance runners. European Journal of Applied Physiology and Occupational
Physiology. 53(3), 213-218.
OSKARSSON, P.R., LINS, P.E., WALLBERG HENRIKSSON, H. & ADAMSON, U.C.
1999. Metabolic and hormonal responses to exercise in type 1 diabetic patients
during continuous subcutaneous, as compared to continuous intraperitoneal,
insulin infusion. Diabetes Metabolism., 25, 491-497.
PAGE, K., WILLIAMSON, A., YU, N., MCNAY, E., DZUIRA, J., MCCRIMMON, R.
& SHERWIN, R. 2009. Medium-chain fatty acids improve cognitive function in
intensively treated type 1 diabetic patients and support in vitro synaptic
transmission during acute hypoglycemia. Diabetes, 58, 1237-1244.
PARANJAPE, S.A., CHAN, O., ZHU, W., HORBLITT, A.M., MCNAY, E.C.,
CRESSWELL, J.A., BOGAN, J.S., MCCRIMMON, R.J. & SHERWIN, R.S.
2010. Influence of insulin in the ventromedial hypothalamus on pancreatic
glucagon secretion in vivo. Diabetes, 59, 1521-1527.
PEDERSEN, B.K. & FISCHER, C.P. 2007. Beneficial health effects of exercise - the role
of IL-6 as a myokine. Trends in Pharmacological Sciences, 28, 152-156.

139
PEDERSEN, B. & FEBBRAIO, M. 2008. Muscle as an endocrine organ: focus on
muscle-derived interleukin-6. Physiological Reviews, 88, 1379-1406.
PEDERSEN, B.K. 2009. Edward F. Adolph Distinguished Lecture: Muscle as an
endocrine organ: IL-6 and other myokines. Journal of Applied Physiology.
107(4),1006-1014.
PEIRCE, N.S. 1999. Diabetes and Exercise. British Journal of Sports Medicine, 33, 161-
173.
PERRY, E. & GALLEN, I. 2009. Guidelines on the current best practice for the
management of type 1 diabetes, sport and exercise. Practical Diabetes
International, 26, 116-123.
PETERSEN, K.F., PRICE, T.B. & BERGERON, R. 2004). Regulation of net hepatic
glycogenolysis and gluconeogenesis during exercise: impact of type 1 diabetes.
Journal of Clinical Endocrinology and Metabolism, 89, 4656–4664.
PLOTNIKOFF, R.C., TAYLOR, L.M., WILSON, P.M., COURNEYA, K.S., SIGAL,
R.J., BIRKETT, N., RAINE, K. & SVENSON, L.W. 2006. Factors associated
with physical activity in Canadian adults with diabetes. Medicine & Science in
Sports & Exercise, 38, 1526-1534.
PORCELLATI, F., PAMPANELLI, S., ROSSETTI, P., CORDONI, C., MARZOTTI, S.,
SCIONTI, L., BOLLI, G. & FANELLI, C. 2003. Counterregulatory hormone and
symptom responses to insulin-induced hypoglycemia in the postprandial state in
humans. Diabetes, 52, 2774-2783.
PORCELLATI, F., PAMPANELLI, S., ROSSETTI, P., BUSCIANTELLA RICCI, N.,
MARZOTTI, S., LUCIDI, P., SANTEUSANIO, F., BOLLI, G. & FANELLI, C.
2007. Effect of the amino acid alanine on glucagon secretion in non-diabetic and
type 1 diabetic subjects during hyperinsulinaemic euglycaemia, hypoglycaemia
and post-hypoglycaemic hyperglycaemia. Diabetologia, 50, 422-430.
PUENTE, E.C., SILVERSTEIN, J., & BREE, A.J., MUSIKANTOW, D.R.,
WOZNIAK, D.F., MALONEY, S., DAPHNA-IKEN, D. & FISHER, S.J. 2010.
Recurrent moderate hypoglycaemia ameliorates brain damage and cognitive
dysfunction induced by severe hypoglycemia. Diabetes, 59:1055–1062.
PURDON, C., BROUSSON, M., NYVEEN, S.L., MILES, P.D.G., HALTER, J.B.,
VRANIC, M. & MARLISS, E.B. 1993. The roles of insulin and catecholamines
in the glucoregulatory response during intense exercise and early recovery in
insulin-dependent diabetic and control subjects. Journal of Clinical
Endocrinology and Metabolism, 76, 566-573.

140
QUESADA, I., TUDURI, E., RIPOLL, C. & NADAL, A. 2008. Physiology of the
pancreatic {alpha}-cell and glucagon secretion: role in glucose homeostasis and
diabetes. Journal of Endocrinology, 199, 5-19
RABASA-LHORET, R., BOURQUE, J., DUCROS, F. & CHIASSON, J.L. 2001.
Guidelines for premeal insulin dose reduction for postprandial exercise of
different intensities and durations in type 1 diabetic subjects treated intensively
with a basal-bolus insulin regimen (ultralente-lispro). Diabetes Care, 24, 625-630.
RABINOWITZ, D., KLASSEN, G. & ZIERLER, K. 1965. Effect of human growth
hormone on muscle and adipose tissue metabolism in the forearm of man. Journal
of Clinical Investigation, 44, 51-61.
REITER, E.O. & ROSENFELD, R.G. 2008. Normal and aberrant growth. In:
KRONENBERG, H.M. (ed.) Williams Textbook of Endocrinology. 11 ed.
Philadelphia: Saunders Elsevier.
RICHTER, E., GALBO, H. & CHRISTENSEN, N. 1981. Control of exercise-induced
muscular glycogenolysis by adrenal medullary hormones in rats. Journal of
Applied Physiology, 50, 21-26.
RICHTER, E., GALBO, H., SONNE, B., HOLST, J. & CHRISTENSEN, N. 1980.
Adrenal medullary control of muscular and hepatic glycogenolysis and of
pancreatic hormonal secretion in exercising rats. Acta Physiologica Scandinavica,
108, 235-242.
RICHTER, E.A., GARETTO, L.P., GOODMAN, M.N. & RUDERMAN, N.B. 1982.
Muscle glucose metabolism following exercise in the rat: increased sensitivity to
insulin. Journal of Clinical Investigation, 69, 785-793.
RICHTER, E.A. & GALBO, H. 1986. Diabetes, insulin and exercise. Sports Medicine, 3,
275-88.
RIDDELL, M.C., BAR-OR, O., AYUB, B.V., CALVERT, R.E. & HEIGENHAUSER,
G.J. 1999. Glucose ingestion matched with total carbohydrate utilization
attenuates hypoglycemia during exercise in adolescents with IDDM. International
Journal of Sports Nutrition, 9, 24-34.
RIDDELL, M.C. & ISCOE, K.E. 2006. Physical activity, sport, and pediatric diabetes.
Pediatric Diabetes. 7(1), 60-70.
RIDDELL, M.C. & PERKINS, B.A. 2006. Type 1 Diabetes and Vigorous Exercise:
Applications of Exercise Physiology to Patient Management. Canadian Journal
of Diabetes, 30, 63-71.

141
RIZZA, R., HAYMOND, M., CRYER, P. & GERICH, J. 1979a. Differential effects of
epinephrine on glucose production and disposal in man. American Journal of
Physiology- Endocrinology and Metabolism, 237, E356-E362.
RIZZA, R.A., CRYER, P.E. & GERICH, J.E. 1979b. Role of glucagon, catecholamines,
and growth hormone in human glucose counterregulation: effects of somatostatin
and combined -and -adrenergic blockade on plasma glucose recovery and glucose
flux rates after insulin-induced hypoglycemia. Journal of Clinical Investigation,
64, 62-71.
RIZZA, R.A., MANDARINO, L.J. & GERICH, J.E. 1982. Cortisol-induced insulin
resistance in man: impaired suppression of glucose production and stimulation of
glucose utilization due to a postreceptor defect of insulin action. Journal of
Clinical Endocrinology & Metabolism, 54, 131-138.
RIZZA, R., JENSEN, L.S. & SREEKUMARAN NAIR, K. 2001. Type 1 diabetes
mellitus. In: JEFFERSON (ed.) The endocrine pancreas and regulation of
metabolism. New York: Oxford University Press.
ROBERTSON, K., ADOLFSSON, P., RIDDELL, M.C., SCHEINER, G. & HANAS, R.
2008. Exercise in children and adolescents with diabetes. Pediatric Diabetes, 9,
65-77.
ROBERTSON, K., RIDDELL, M.C., GUINHOUYA, B.C., ADOLFSSON, P. &
HANAS, R. 2014. Exercise in children and adolescents with diabetes. Pediatric
Diabetes, 15, 203-223.
ROCHA, D., FALOONA, G. & UNGER, R. 1972. Glucagon-stimulating activity of 20
amino acids in dogs. Journal of Clinical Investigation, 51, 2346-2351.
RODEN, M. & BERNROIDER, E. 2003. Hepatic glucose metabolism in humans--its role
in health and disease. Best Practice & Research Clinical Endocrinology &
Metabolism, 17, 365-383.
ROONEY, D., NEELY, R., CULLEN, C., ENNIS, C., SHERIDAN, B., ATKINSON, A.,
TRIMBLE, E. & BELL, P. 1993. The effect of cortisol on glucose/glucose-6-
phosphate cycle activity and insulin action. Journal of Clinical Endocrinology &
Metabolism, 77, 1180-1183.
ROSSETTI, P., PORCELLATI, F., BUSCIANTELLA RICCI, N., CANDELORO, P.,
CIOLI, P., NAIR, K., SANTEUSANIO, F., BOLLI, G. & FANELLI, C. 2008.
Effect of oral amino acids on counterregulatory responses and cognitive function
during insulin-induced hypoglycemia in nondiabetic and type 1 diabetic people.
Diabetes, 57, 1905-1917.

142
SACCA, L., MORRONE, G., CICALA, M., CORSO, G. & UNGARO, B. 1980.
Influence of epinephrine, norepinephrine, and isoproterenol on glucose
homeostasis in normal man. Journal of Clinical Endocrinology & Metabolism,
50, 680-684.
SACCA, L., VIGORITO, C., CICALA, M., CORSO, G. & SHERWIN, R. 1983. Role of
gluconeogenesis in epinephrine-stimulated hepatic glucose production in humans.
American Journal of Physiology- Endocrinology and Metabolism, 245, E294-
E302.
SANDOVAL, D., GUY, D., RICHARDSON, M., ERTL, A. & DAVIS, S. 2004. Effects
of low and moderate antecedent exercise on counterregulatory responses to
subsequent hypoglycemia in type 1 diabetes. Diabetes, 53, 1798-1806.
SANDOVAL, D.A., AFTAB GUY, D.L., RICHARDSON, M.A., ERTL, A.C. & DAVIS,
S.N. 2006. Acute, same-day effects of antecedent exercise on counterregulatory
responses to subsequent hypoglycemia in type 1 diabetes mellitus. American
Journal of Physiology-Endocrinology and Metabolism, 290, E1331-1338.
SARASLANIDIS, P.J., MANETZIS, C.G., TSALIS, G.A., ZAFEIRIDIS, A.S.,
MOUGIOS, V.G. & KELLIS, S.E. 2009. Biochemical evaluation of running
workouts used in training for the 400-m sprint. Journal of Strength and
Conditioning Research, 23, 2266-2271.
SCHIFFRIN, A., PARIKH, S., MARLISS, E.B. & DESROSIERS, M.M. 1984.
Metabolic response to fasting exercise in adolescent insulin-dependent diabetic
subjects treated with continuous subcutaneous insulin infusion and intensive
conventional therapy. Diabetes Care, 7, 255 - 260.
SCHMID, R., SCHUSDZIARRA, V., SCHULTE-FROHLINDE, E., MAIER, V. &
CLASSEN, M. 1989. Role of amino acids in stimulation of postprandial insulin,
glucagon, and pancreatic polypeptide in humans. Pancreas, 4, 305-314.
SCHNABEL, A. & KINDERMANN W., CASSENS, J., STEINKRAUS, V. 1983. Effect
of beta-adrenergic blockade on supramaximal exercise capacity. International
Journal of Sports Medicine. 4(4), 278-81.
SCHNABEL, A., KINDERMAN, W., STEINKRAUS, V., SALAS-FRAIRE, O. &
BIRO, G. 1984. Metabolic and hormonal response to exhaustive supramaximal
running with and without b-adrenergic blockade. European Journal of Applied
Physiology . 52, 214-218.

143
SCHWARTZ, N., CLUTTER, W., SHAH, S. & CRYER, P. 1987. Glycemic thresholds
for activation of glucose counterregulatory systems are higher than the threshold
for symptoms. Journal of Clinical Investigation, 79, 777-781.
SCHWARZ, L. & KINDERMANN, W. 1990. Endorphin, adrenocorticotropic hormone,
cortisol and catecholamines during aerobic and anaerobic exercise. European
Journal of Applied Physiology and Occupational Physiology, 61, 165-171.
SHAMOON, H., SOMAN, V. & SHERWIN, R.S. 1980. The influence of acute
physiological increments of cortisol on fuel metabolism and insulin binding to
monocytes in normal humans. Journal of Clinical Endocrinology and
Metabolism, 50, 495-501.
SHERR, J., CENGIZ, E. & TAMBORLANE, W. 2009. From pumps to prevention: recent
advances in the treatment of type 1 diabetes. Drug Discovery Today, 14, 973-981.
SHERWIN, R., SHAMOON, H., HENDLER, R., SACC , L., EIGLER, N. &
WALESKY, M. 1980. Epinephrine and the regulation of glucose metabolism:
Effect of diabetes and hormonal interactions 1. Metabolism, 29, 1146-1154.
SHILO, S. & SHAMOON, H. 1990a. Abnormal growth hormone responses to
hypoglycemia and exercise in adults with type I diabetes. Israel Journal of
Medical Science., 26, 136-141.
SHILO, S., SOTSKY, M. & SHAMOON, H. 1990b. Islet hormonal regulation of glucose
turnover during exercise in type 1 diabetes. Journal of Clinical Endocrinology
and Metabolism, 70, 162 - 172.
SHUM, K., INOUYE, K., CHAN, O., MATHOO, J., BILINSKI, D., MATTHEWS, S. &
VRANIC, M. 2001. Effects of antecedent hypoglycemia, hyperinsulinemia, and
excess corticosterone on hypoglycemic counterregulation. American Journal of
Physiology- Endocrinology and Metabolism, 281, E455-E465.
SIGAL, R., PURDON, C., FISHER, S., HALTER, J., VRANIC, M. & MARLISS, E.
1994a. Hyperinsulinemia prevents prolonged hyperglycemia after intense
exercise in insulin-dependent diabetic subjects. Journal of Clinical Endocrinology
& Metabolism, 79, 1049-1057.
SIGAL, R.J., PURDON, C., BILINSKI, D., VRANIC, M., HALTER, J.B. & MARLISS,
E.B. 1994b. Glucoregulation during and after intense exercise: Effects of -
blockade. Journal of Clinical Endocrinology & Metabolism, 78, 359-366.
SIGAL, R.J., FISHER, S., HALTER, J.B., VRANIC, M. & MARLISS, E.B. 1996. The
roles of catecholamines in glucoregulation in intense exercise as defined by the
islet cell clamp technique. Diabetes, 45, 148-156.

144
SIGAL, R.J., FISHER, S.J., HALTER, J.B., VRANIC, M. & MARLISS, E.B. 1999.
Glucoregulation during and after intense exercise: effects of -adrenergic
blockade in subjects with type 1 diabetes mellitus. Journal of Clinical
Endocrinology & Metabolism, 84, 3961 - 3971.
SIGAL, R. J., FISHER, S.J., MANZON, A., MORAIS, J.A., HALTER, J.B., VRANIC,
M. & MARLISS, E.B. 2000. Glucoregulation during and after intense exercise:
Effects of adrenergic blockade. Metab., 49, 386-394.
SILLS, I. & CERNY, F. 1983. Responses to continuous and intermittent exercise in
healthy and insulin-dependent diabetic children. Medicine & Science in Sports &
Exercise, 15, 450-454.
SIMONSON, D., KOIVISTO, V., SHERWIN, R., FERRANNINI, E., HENDLER, R.,
JUHLIN-DANNFELT, A. & DEFRONZO, R. 1984. Adrenergic blockade alters
glucose kinetics during exercise in insulin-dependent diabetics. Journal of
Clinical Investigation, 73, 1648-1658.
SKRIVARHAUG, T., BANGSTAD, H.J., STENE, L.C., SANDVIK, L., HANSSEN,
K.F. & JONER, G.. 2006. Long-term mortality in a nationwide cohort of
childhood-onset type 1 diabetic patients in Norway. Diabetologia, 49:298–305.
SOLTESZ, G., PATTERSON, C. & DAHLQUIST, G. 2009. Diabetes in the Young: a
global perspective; global trends in childhood type 1 diabetes. IDF diabetes atlas.
4th edn. Brussels: International Diabetes Federation.
SONNE, B., MIKINES, K., RICHTER, E., CHRISTENSEN, N. & GALBO, H. 1985.
Role of liver nerves and adrenal medulla in glucose turnover of running rats.
Journal of Applied Physiology, 59, 1640-1646.
SONNENBERG, G.E., KEMMER, F.W. & BERGER, M. 1990. Exercise in type 1
(insulin-dependent) diabetic patients treated with continuous subcutaneous insulin
infusion - Prevention of exercise induced hypoglycemia. Diabetologia, 33, 696-
703.
SOTSKY, M.J., SHILO, S. & SHAMOON, H. 1989. Regulation of counterregulatory
hormone secretion in man during exercise and hypoglycaemia. Journal of Clinical
Endocrinology and Metabolism. 68, 9-16.
SOUPAL, J., SKRHA, J., FAJMON, M., HOROV, Á.E., MRÁZ, M., SKRHA, J. &
PRÁZNÝ, M. 2014. Glycemic variability is higher in type 1 diabetes patients with
microvascular complications irrespective of glycemic control. Diabetes
Technology and Therapeuthics 16(4), 198-203.

145
SPENCER, M., BISHOP, D., DAWSON, B. & GOODMAN, C. 2005. Physiological and
metabolic responses of repeated-sprint activities: specific to field-based team
sports. Sports Medicine, 35, 1025-1044.
SPENCER, M., LAWRENCE, S., RECHICHI, C., BISHOP, D., DAWSON, B. &
GOODMAN, C. 2004. Time–motion analysis of elite field hockey, with special
reference to repeated-sprint activity. Journal of Sports Sciences, 22, 843-850.
STAEHR, P., HOTHER-NIELSEN, O., BECK-NIELSEN, H., RODEN, M., STING, L
H., HOLST, J.J., JONES, P.K., CHANDRAMOULI, V. & LANDAU, B.R. 2007.
Hepatic autoregulation:response of glucose production and gluconeogenesis to
increased glycogenolysis. American Journal of Physiology- Endocrinology and
Metabolism, 292, E1265–E1269.
STEINER, K., WILLIAMS, P., LACY, W. & CHERRINGTON, A. 1990. Effects of
insulin on glucagon-stimulated glucose production in the conscious dog.
Metabolism, 39, 1325-1333.
STEPPEL, J.H.H. & HORTON, E.S. (eds.) 2004. Exercise for the patient with type 1
diabetes mellitus. Lippincott Williams and Wilkins.
STICKLEY, C.D., HETZLER, R.K. & KIMURA, I.F. 2008. Prediction of anaerobic
power values from an abbreviated WAnT protocol. Journal of Strength and
Conditioning Research, 22, 958-965.
STOKES, K.A., NEVILL, M.E., HALL, G.M. & LAKOMY, H.K. 2002a. The time
course of the human growth hormone response to a 6 s and a 30 s cycle ergometer
sprint. Journal of Applied Physiology, 92, 602-608.
STOKES, K.A. & NEVILL, M.E. & LAKOMY, H.K. 2002b. Growth hormone responses
to repeated maximal cycle ergometer exercise at different pedaling rates. Journal
of Applied Physiology, 92(2), 602-608.
STOKES, K. 2003. Growth hormone responses to sub-maximal and sprint exercise.
Growth Hormone & IGF Research, 13, 225-231.
STOKES, K.A. & NEVILL, M.E. & HALL, G.M. 2003. Reproducibility of the growth
hormone response to sprint exercise. Growth Hormone and IGF Research.
13(6),336-340.
STOKES, K.A., NEVILL, M.E., CHERRY, P.W., LAKOMY, H.K. & HALL, G.M.
2004. Effect of 6 weeks of sprint training on growth hormone responses to
sprinting. European Journal of Applied Physiology, 92, 26-32.

146
STOKES, K., NEVILL, M., FRYSTYK, J. LAKOMY, H. & HALL, G. 2005. Human
growth hormone responses to repeated bouts of sprint exercise with different
recovery periods between bouts. Journal of Applied Physiology, 99(4), 1254-1261
STOKES, K., NEVILL, M. & HALL, G. 2006. Age is an important determinant of the
growth hormone response to sprint exercise in non-obese young men. Hormones
Research. 65(2), 57-61.
STOKES, K.A., TYLER, C. & GILBERT, K.L. 2008. The growth hormone response to
repeated bouts of sprint exercise with and without suppression of lipolysis in men.
Journal of Applied Physiology, 104, 724-8.
STOKES, K.A., GILBERT, K.L., HALL, G.M., ANDREWS, R.C. & THOMPSON, D.
2013. Different responses of selected hormones to three types of exercise in young
men. European Journal of Applied Physiology. 113(3),775-783.
STUMVOLL, M., MEYER, C., KREIDER, M., PERRIELLO, G. & GERICH, J. 1998a.
Effects of glucagon on renal and hepatic glutamine gluconeogenesis in normal
postabsorptive humans 1. Metabolism, 47, 1227-1232.
STUMVOLL, M., MEYER, C., PERRIELLO, G., KREIDER, M., WELLE, S. &
GERICH, J. 1998b. Human kidney and liver gluconeogenesis: evidence for organ
substrate selectivity. American Journal of Physiology- Endocrinology and
Metabolism, 274, E817-E826.
SUH, S.W., HAMBY, A.M., & SWANSON, R.A. 2007. Hypoglycemia, brain energetics,
and hypoglycaemic neuronal death. Glia, 2007, 55:1280–1286.
TANENBERG, R.J., NEWTON, C.A. & DRAKE, A.J. 2010. Confirmation of
hypoglycemia in the “dead-in-bed” syndrome, as captured by a retrospective
continuous glucose monitoring system. Endocrine Pathology, 16, 244-248.
TELFORD, R.D., MINIKIN, B.R., HOOPER, L.A., HAHN, A.G. & TUMILTY, D.
1987. The tri-level fitness profile. Excel, 4, 11-13.
TESFAYE, S., BOULTON, A., DYCK, P., FREEMAN, R., HOROWITZ, M.,
KEMPLER, P., LAURIA, G., MALIK, R., SPALLONE, V. & VINIK, A. 2010.
Diabetic neuropathies: update on definitions, diagnostic criteria, estimation of
severity, and treatments. Diabetes Care, 33, 2285-2293.
THOMAS, N., ALDER, E. & LEESE, G. 2004. Barriers to physical activity in patients
with diabetes. Postgraduate Medical Journal, 80, 287-291.
TONELLI, J., KISHORE, P., LEE, D-E. & HAWKINS, M. (2005). The regulation of
glucose effectiveness: how glucose modulates its own production. Current
Opinion in Clinical Nutrition and Metabolic Care. 8, 450-456.

147
TONOLI, C., HEYMAN, E., ROELANDS, B., BUYSE, L., CHEUNG, S.S.,
BERTHOIN, S. & MEEUSEN, R. 2012. Effects of different types of acute and
chronic (training) exercise on glycaemic control in type 1 diabetes mellitus: a
meta-analysis. Tonoli C, Heyman E, Roelands B, Buyse L, Cheung SS, Berthoin
S, Meeusen R. Sports Medicine, 42(12), 1059-1080.
TOWLER, D., HAVLIN, C., CRAFT, S. & CRYER, P. 1993. Mechanism of awareness
of hypoglycemia. Perception of neurogenic (predominantly cholinergic) rather
than neuroglycopenic symptoms. Diabetes, 42, 1791-1798.
TRAPP, E., CHISHOLM, D. & BOUTCHER, S. 2007. Metabolic response of trained and
untrained women during high-intensity intermittent cycle exercise. American
Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 293,
R2370-R2375.
TRIMMER, J.K., SCHWARZ, J.M., CASAZZA, G.A., HORNING, M.A.,
RODRIGUEZ, N. & BROOKS, G.A. 2002. Measurement of gluconeogenesis in
exercising men by mass isotopomer distribution analysis. Journal of Applied
Physiology, 93, 233-241.
TURNER, D., LUZIO, S., GRAY, B.J., DUNSEATH, G., REES, E.D., KILDUFF,
L.P., CAMPBELL, M.D., WEST, D.J., BAIN, S.C. & BRACKEN, R.M. 2015.
Impact of single and multiple sets of resistance exercise in type 1 diabetes.
Scandinavian Journal of Medicine and Science in Sports, 25(1), 99-109.
TSALIKIAN, E., MAURAS, N., BECK, R., TAMBORLANE, W., JANZ, K., CHASE,
H., WYSOCKI, T., WEINZIMER, S., BUCKINGHAM, B., KOLLMAN, C.,
XING, D., RUEDY, K.-DIABETES RESEARCH IN CHILDREN NETWORK
DIRECNET STUDY GROUP. 2005. Impact of exercise on overnight glycemic
control in children with type 1 diabetes mellitus. The Journal of Pediatrics, 147,
528-534.
TUOMINEN, J.A., KARONEN, S.L., MELAMIES, L., BOLLI, G. & KOIVISTO, V. A.
1995. Exercise-induced hypoglycaemia in IDDM patients treated with a short-
acting insulin analogue. Diabetologia, 38, 106 –111.
VAN PRAAGH, E. 1998. Pediatric Anaerobic Performance, Champaign, Human
Kinetics.
VENEMAN, T., MITRAKOU, A., MOKAN, M., CRYER, P. & GERICH, J. 1994. Effect
of hyperketonemia and hyperlacticacidemia on symptoms, cognitive dysfunction,
and counterregulatory hormone responses during hypoglycemia in normal
humans. Diabetes, 43, 1311-1317.

148
VETTOR, R., LOMBARDI, A.M., FABRIS, R., PAGANO, C., CUSIN, I., ROHNER-
JEANRENAUD, F., FEDERSPIL, G. & JEANRENAUD, B. 1997. Lactate
infusion in anesthetized rats produces insulin resistance in heart and skeletal
muscles. Metabolism, 46, 684-690.
VINCENT, S., BERTHON, P., ZOUHAL, H., MOUSSA, E., CATHELINE, M.,
BENTUÉ-FERRER, D. & GRATAS-DELAMARCHE, A. 2004. Plasma glucose,
insulin and catecholamine responses to a Wingate test in physically active women
and men. European Journal of Applied Physiology, 91(1), 15-21.
VINCENT, S. & GRATAS-DELAMARCHE A, BERTHON, P.M., ZOUHAL, H.,
JACOB, C., BENTUE-FERRER, D. & DELAMARCHE, P. 2003. Catecholamine
response to the Wingate test in untrained women. Canadian Journal of Applied
Physiology, 28(5), 685-698.
VISSING, J., IWAMOTO, G., FUCHS, I., GALBO, H. & MITCHELL, J. 1994. Reflex
control of glucoregulatory exercise responses by group III and IV muscle
afferents. American Journal of Physiology- Regulatory, Integrative and
Comparative Physiology, 266, R824-R830.
WAHREN, J. & EKBERG, K. 2007. Splanchnic regulation of glucose production.
Nutrition, 27, 329-345.
WAHREN, J., HAGENFELDT, L. & FELIG, P. 1975. SPLANCHNIC AND LEG
EXCHANGE OF glucose, amino acids, and free fatty acids during exercise in
diabetes mellitus. Journal of Clinical Investigation, 55, 1303-1314.
WANKE, T., FORMANEK, D., AUINGER, M., ZWICK, H. & IRSIGLER, K. 1992.
Pulmonary gas exchange and oxygen uptake during exercise in patients with type
1 diabetes mellitus. Diabetic Medicine, 9, 252-257.
WASSERMAN, D., LICKLEY, H. & VRANIC, M. 1984. Interactions between glucagon
and other counterregulatory hormones during normoglycemic and hypoglycemic
exercise in dogs. Journal of Clinical Investigation, 74, 1404-1413.
WASSERMAN, D., SPALDING, J., LACY, D., COLBURN, C., GOLDSTEIN, R. &
CHERRINGTON, A. 1989a. Glucagon is a primary controller of hepatic
glycogenolysis and gluconeogenesis during muscular work. American Journal of
Physiology- Endocrinology and Metabolism, 257, E108-E117.
WASSERMAN, D., WILLIAMS, P., LACY, D., GOLDSTEIN, R. & CHERRINGTON,
A. 1989b. Exercise-induced fall in insulin and hepatic carbohydrate metabolism
during muscular work. American Journal of Physiology- Endocrinology and
Metabolism, 256, E500-E509.

149
WASSERMAN, D., WILLIAMS, P., LACY, D., BRACY, D. & CHERRINGTON, A.
1990. Hepatic nerves are not essential to the increase in hepatic glucose
production during muscular work. American Journal of Physiology-
Endocrinology end Metabolism, 259, E195-203.
WASSERMAN, D., LACY, D., COLBURN, C., BRACY, D. & CHERRINGTON, A.
1991. Efficiency of compensation for absence of fall in insulin during exercise.
American Journal of Physiology- Endocrinology and Metabolism, 261, E587-
E597.
WASSERMAN, D., LACY, D. & BRACY, D. 1993. Relationship between arterial and
portal vein immunoreactive glucagon during exercise. Journal of Applied
Physiology, 75, 724-729.
WASSERMAN, D.H. & ZINMAN, B. 1994. Exercise in individuals with IDDM.
Diabetes Care, 17, 924-937.
WASSERMAN, D.H. (1995). Regulation of glucose fluxes during exercise in the post-
absorptive state. Annual Review of Physiology. 57, 191-218.
WASSERMAN, D.H. 2009. Four grams of glucose. American Journal of Physiology -
Endocrinology And Metabolism, 296, E11-21.
WATT, M.J. & HARGREAVES, M. 2002. Effect of epinephrine on glucose disposal
during exercise in humans.: Role of muscle glycogen. American Journal of
Physiology, Endocrinology and Metabolism. 283, E578-E583.
WATT, M.J., HOWLETT, K.F., FEBBRAIO, M.A., SPRIET, L.L. & HARGREAVES,
M. 2001. Adrenaline increases skeletal muscle glycogenolysis, PDH activation
and carbohydrate oxidation during moderate exercise in humans. Journal of
Physiology (London). 534, 269-278.
WATTS, A.G. & DONOVAN, C.M. 2010. Sweet talk in the brain: Glucosensing, neural
networks, and hypoglycemic counterregulation. Frontiers in Neuroendocrinology
31, 32-43.
WEINSTEIN, Y., BEDIZ, C., DOTAN, R. & FALK, B. 1998. Reliability of peak-lactate,
heart rate, and plasma volume following the Wingate test. Medicine Science in
Sports and Exercise. 30(9),1456-1460.
WHITE, N., SKOR, D., CRYER, P., LEVANDOSKI, L., BIER, D. & SANTIAGO, J.
1983. Identification of type I diabetic patients at increased risk for hypoglycemia
during intensive therapy. New England Journal of Medicine, 308, 485-491.
WIDOM, B. & SIMONSON, D. 1992. Intermittent hypoglycemia impairs glucose
counterregulation. Diabetes, 41, 1597-1602.

150
WIERSMA, M.M., VISSING, J., STEFFENS, A.B. & GALBO, H. (1993). Effects of
glucose infusion on hormone secretion and hepatic glucose production during
heavy exercise. American Journal of Physiology, Regulatory, Integrative and
Comparative Physiology. 265, R1333-R1338.
WOLFE, R., NADEL, E., SHAW, J., STEPHENSON, L. & WOLFE, M. 1986. Role of
changes in insulin and glucagon in glucose homeostasis in exercise. Journal of
Clinical Investigation, 77, 900-907.
YARDLEY, J.E., KENNY, G.P., PERKINS, B.A. 2012. Effects of performing
resistance exercise before versus after aerobic exercise on glycemia in type 1
diabetes. Diabetes Care, 35(4), 669–675.
YARDLEY, J.E., KENNY, G.P., PERKINS, B.A., RIDDELL, M.C., BALAA, N.,
MALCOLM, J., BOULAY, P., KHANDWALA, F. & SIGAL, R.J. 2013.
Resistance versus aerobic exercise: acute effects on glycemia in type 1 diabetes.
Diabetes Care, 36(3), 537-542.
YARDLEY, J.E. & SIGAL, R.J. 2015. Exercise strategies for hypoglycemia prevention
in individuals with type 1 diabetes. Diabetes Spectrum, 28(1), 32-38.
YOUNK, L.M. & DAVIS, S.N. 2012. Hypoglycemia and hypoglycemia unawareness
during and following exercise. In: GALLEN, I. (ed.) Type 1 Diabetes: Clinical
Management of the Athlete.\
ZAJAC, A., JARZABEK, R. & WASKIEWICZ, Z. 1999. The diagnostic value of the 10-
and 30-second Wingate test for competitive athletes. The Journal of Strength and
Conditioning Research, 13, 16-19.
ZIERLER, K. & RABINOWITZ, D. 1963. Roles of insulin and growth hormone, based
on studies on forearm metabolism in man. Medicine, 42, 385-402.
ZINMAN, B., MURRAY, F.T., VRANIC, M., ALBISSER, A.M., LEIBEL, B.S.,
MCCLEAN, P.A. & MARLISS, E.B. 1977. Glucoregulation during moderate
exercise in insulin treated diabetics. Journal of Clinical Endocrinology and
Metabolism, 45, 641 - 652.
ZINMAN, B., ZUNIGA-GUAJARDO, S. & KELLY, D. 1984. Comparison of the acute
and long-term effects of exercise on glucose control in type I diabetes. Diabetes
Care, 7, 515-519.
ZINMAN, B., RUDERMAN, N., CAMPAIGNE, B.N., DEVLIN, J.T., SCHNEIDER,
S.H. & ASSOCIATION, A.D. 2004. Physical activity/exercise and diabetes
(Position Statement). Diabetes Care, 27, S58 –S62.

151
ZOUHAL, H., JACOB, C., DELAMARCHE, P. & GRATAS-DELAMARCHE, A. 2008.
Catecholamines and the effects of exercise, training and gender. Sports Medicine,
38, 401-423.
ZOUHAL, H., JACOB, C., RANNOU, F., GRATAS-DELAMARCHE, A., BENTUÉ-
FERRER, D. & DEL, P. 2001. Effect of training status on the sympathoadrenal
activity during a supramaximal exercise in human. Journal of Sports Medicine
Physiology Fitness. 41(3):330-336.
ZOUHAL, H., RANNOU, F., GRATAS-DELAMARCHE, A., MONNIER, M.,
BENTUÉ-FERRER, D. & DELAMARCHE, P. 1998. Adrenal medulla
responsiveness to the sympathetic nervous activity in sprinters and untrained
subjects during a supramaximal exercise. International Journal of Sports
Medicine. 19(3), 172-176.
ZOUHAL, H., VINCENT, S., MOUSSA, E., BOTCAZOU, M., DELAMARCHE, P. &
GRATAS-DELAMARCHE, A. 2009. Early advancing age alters plasma glucose
and glucoregulatory hormones in response to supramaximal exercise. Journal of
Science Medicine Sport. 12(6), 652-6.


![INDEX [cornerstone.ac.za]€¦ · Bachelor of Arts Honours in Community Development 158 Bachelor of Arts Honours in Psychology (Academic) 163 Bachelor of Arts Honours in Psychology](https://img.pdfslide.net/doc/110x75/5fc6aa834c59c236646cfc46/index-bachelor-of-arts-honours-in-community-development-158-bachelor-of-arts.jpg)


























