Embed Size (px)
Citation preview

EGYETEMI DOKTORI (Ph.D) ÉRTEKEZÉS
Pécsi Tudományegyetem Doktori Iskola
(Elméleti orvostudományok)
A TRPV1 KAPSZAICIN RECEPTOR FARMAKOLÓGIAI VIZSGÁLAT A
Varga Angelika
Neurofarmakológia
címő program
Programvezetı: Dr. Szolcsányi János, egyetemi tanár
Témavezetı: Dr. Czéh Gábor és
Dr. Sándor Zoltán
Pécsi Tudományegyetem
Farmakológiai és Farmakoterápiai Intézet
MTA Neurofarmakológiai Kutatócsoport, Pécs
2006

2
TARTALOMJEGYZÉK
TARTALOMJEGYZÉK 2
Rövidítések 4
1. ÁLTALÁNOS BEVEZETÉS 5
1.1. Mérföldkövek a kapszaicin-kutatásban 5
1.2. A kapszaicin, mint neurofarmakológiai eszköz 6
1.3. A TRPV1 kapszaicin receptor bemutatása, jelentısége 7
1.4. Népi gyógyászat vagy modern farmakológia? 11
1.5. Az értekezés célkitőzései és tematikai felosztása 13
2. MÓDSZERTAN 16
2.1. Sejttípusok 16
2.1.1. Patkány trigeminális ganglionsejt tenyészet 16
2.1.2. TRPV1 receptort stabilan expresszáló HT5-1 sejtvonal 16
2.2. Mikrofluorimetriás technikák 16
2.2.1. Intracelluláris kalciumionszint-mérés fluoreszcens módszerrel 16
2.2.2. A relatív intracelluláris szabad kalciumionszint és a
mitokondriális membránpotenciál változás szimultán mérése 18
2.3. Kalciumizotóppal végzett kísérletek 19
2.4. P-anyag felszabadulásának vizsgálata in vitro tracheapreparátumon 19
2.5. In vivo kísérletek 20
2.5.1. Az SB366791 hatása a kapszaicinnel kiváltott térdízületi
vérátáramlás-fokozódásra 20
2.5.2. Az SB366791 hatása a kapszaicinnel kiváltott testhımérséklet
csökkenésre 20
2.5.3. Kapszaicinnel kiváltott szemtörlési reflex vizsgálata 21
2.5.4. Nociceptív hıküszöb mérése emelkedı hımérséklető forró lap
módszerrel 21
2.6. Alkalmazott hatóanyagok 22
3. EREDMÉNYEK 23 3.1. Az új TRPV1 receptor antagonista SB366791 hatásainak in vitro és
in vivo farmakológiai vizsgálata 23
Bevezetés 23

3
Eredmények 26
Következtetések 30
3.2. Az N-oleoildopamin és analógjainak in vitro hatáselemzése 32
Bevezetés 32
Eredmények 34
Következtetések 38
3.3. A protein kináz A és C relatív szerepének vizsgálata a TRPV1
receptor érzékenységének modulálásában 39
Bevezetés 39
Eredmények 42
Következtetések 53
3.4. A TRPV1 receptor-mediált intracelluláris kalciumionszint-emelkedés
vizsgálata extracelluláris kalcium hiányában 56
Bevezetés 56
Eredmények 57
Következtetések 61
3.5. A TRPV1 kapszaicin receptor aktiválás és a mitokondrium funkció
közötti viszony vizsgálata 63
Bevezetés 63
Eredmények 65
Következtetések 73
4. LEGFONTOSABB ÚJ EREDMÉNYEK ÖSSZEFOGLALÁSA 76
FELHASZNÁLT IRODALOM 78
KÖSZÖNETNYILVÁNÍTÁS 86
AZ ÉRTEKEZÉS ALAPJÁUL SZOLGÁLÓ KÖZLEMÉNYEK
JEGYZÉKE 87
Közlemények 87
Hazai angol nyelvő folyóiratban megjelent idézhetı elıadáskivonatok 87
Az értekezés alapját képezı elıadások 88
Az értekezés alapját képezı poszterek 89
AZ ÉRTEKEZÉSBEN FEL NEM HASZNÁLT TUDOMÁNYOS
MUNKÁK JEGYZÉKE 90
Közlemények 90
Idézhetı kivonatok, Poszterek 91

4
Rövidítések
ASIC proton-szenzitív ioncsatorna
BK bradikinin
CaMKII kalcium-kalmodulindependens kináz II
CPM count per minute (beütés/perc)
CZP kapszazepin
DAG diacil-glicerol
DbcAMP dibutiril-ciklikus-adenozin-5’-monofoszfát
DMEM Dulbecco′s Modified Eagle Medium
DRG hátsógyöki ganglion
ECS extracelluláris oldat
EGTA etilén glikol-bisz(β-aminoetil éter)-N,N,N,N΄-tetraecetsav
12-HPETE 12-(S)-hidroxi-peroxi-eikozanoidsav
IP3 inozitol-1,4,5-triszfoszfát
I-RTX iodo-reziniferatoxin
3-MOLDA 3-metoxi-oleoildopamin
4-MOLDA 4-metoxi-oleoildopamin
NADA N-arachidonildopamin
NGF idegnövekedési faktor
PAR2 proteáz aktivált receptor 2-es típus
PBS foszfát puffer oldat
PGE2 prosztaglandin E2
PI3K foszfatidil-inozitol-3 kináz
PIP2 foszfatidil-inozitol-trifoszfát
PKA cAMP-dependens protein kináz
PKC protein kináz C
PLC foszfolipáz C
PMA forbol 12-mirisztát 13-acetát
PP2B protein foszfatáz 2B
P2X3 ionotróp purinerg receptor X3-as típus
OLDA N-oleoildopamin
rTRPV1-HT5-1 patkány TRPV1 receptorral transzfektált HT1080-as sejtvonal
RTX reziniferatoxin
SDS nátrium-dodecil-szulfát
SP P-anyag
TRG trigeminális ganglion
TRP tranziens receptor potenciál
TRPV1 transiens receptor potenciál vanilloid 1-es típus
VR1 vanilloid receptor 1-es típus

5
1. ÁLTALÁNOS BEVEZETÉS
1.1. Mérföldkövek a kapszaicin-kutatásban
A Pécsi Tudományegyetem Farmakológiai és Farmakoterápiai Intézetében Dr. Szolcsányi János
akadémikus munkásságának köszönhetıen a kapszaicinnel (a paprika csípıanyaga, N-vanillil-8-
metil-6-nonamid) foglalkozó kutatások jelentıs múltra tekintenek vissza (Szolcsányi 2004, 2005).
Még mielıtt rátérnék a TRPV1 kapszaicin receptor és Ph.D munkám illetve eredményeim
bemutatására, érdemesnek vélem felvázolni, hogyan jutott el a tudomány a mai ismeretekig, hogy
egy átfogóbb képet kaphassanak a kapszaicinnel és receptorával kapcsolatos kutatások
jelentıségérıl.
1494 – a paprika növényt Columbus orvosa, Chanca behozta az Újvilágból Európába.
1748 – az elsı paprika feljegyzés Szegedrıl. Neve állítólag a „paprígató” szóból származik.
Az ottani ferences kolostorból terjedt el Magyarországon. Akkoriban a paprikaörleményt
hideglelés ellen alkalmazták.
1816 – Bucholtz megfigyelte, hogy a csípısségért felelıs anyago(ka)t szerves oldószerekkel
ki lehet vonni a paprikából.
1846 – Thresh a hatóanyagot kristályos állapotban extrahálta és ı nevezte el kapszaicinnek.
1878 – Hıgyes Endre a kivont kapszaicin számos élettani tulajdonságát jegyezte le, például,
hogy fokozza a gyomornedv szekrécióját és, hogy izzadás révén hıveszteséget vált ki.
1912 – Wilbur Scoville kémikus egy általa kifejlesztett „hígítás-ízlelés” módszer segítségével
az úgynevezett Scoville-féle organoleptikus teszttel mérte a Capsicum nemzetségbe tartozó
fajok csípısségét és az egyes specieszek erısségét egységekben fejezte ki (S.U). (Módszerét
napjainkra a HPLC váltotta fel.) A habanero egyik variánsa, a Red Savina ilymódon került be
a Guiness rekordok közé (350-577ezer S.U erısséggel).
1930 – Spath és Darling elsıként szintetizáltak kapszaicint.
1949 – Jancsó Miklós beszámolt a „kapszaicin deszenzibilizáció” jelenségérıl.
1967 – Jancsó Miklós, Jancsóné Gábor Aranka és Szolcsányi János elsıként publikáltak
nemzetközi folyóíratban teljes terjedelemben a „kapszaicin deszenzibilizációról” és a
kapszaicin neuroszelektív blokkoló hatásáról.
1975 –Szolcsányi János és Jancsóné Gábor Aranka a szerkezet-hatás összefüggések
alapján egy specifikus, kapszaicin-érzékeny receptor létezését posztulálták.
1982 – Szolcsányi János a primér szenzoros neuronokat kapszaicin-érzékenységük szerint
osztályozta („kapszaicin-szenzitív szenzoros neuronok”).

6
1992 – Bevan és mtsai: egy kapszaicin-analóg antagonista, a kapszazepin kifejlesztésével a
kapszaicin receptor 1975-ös koncepciója megerısítést nyert.
1997 – Caterina és mtsai: a patkány kapszaicin (vanilloid) receptor (VR1) sikeres
klónozásától a kapszaicin-kutatás virágkorát éli.
Innentıl exponenciálisan nı a TRPV1/VR1 receptor irodalma (Szolcsányi, 2005), és sorra
klónozzák meg a különbözı emlısfajok, a humán (Hayes és mtsai, 2000), a tengerimalac
(Savidge és mtsai, 2002), a nyúl (Gavva és mtsai, 2004), az egér (Correll és mtsai, 2004)
valamint a kutya (Phelps és mtsai, 2005) TRPV1 receptor ortológjait.
2000 – Caterina és mtsai valamint Davis és mtsai: VR1 génhiányos egerekben a gyulladás-
valamint a vanilloid-indukált fájdalomreakció károsodik, de a mechanotranszdukció érintetlen
marad.
1.2. A kapszaicin, mint neurofarmakológiai eszköz
A kapszaicin egy bevált neurofarmakológiai eszköz, melynek célmolekulája egy olyan 1997-
ben klónozott receptor-ioncsatorna komplex (Caterina és mtsai, 1997), ami a szenzoros
neuronok közül csak a forró ingerekkel aktiválható polimodális nociceptorok sejttestében
expresszálódik. Azóta bebizonyosodott, hogy az érzıneuronokon kívül kapszaicinre érzékeny
csatornafehérjét bizonyos agyrészek neuronjai (Mezey és mtsai, 2000), epidermális
keratinociták (Southall és mtsai, 2003), urotéliumsejtek (Szállási és mtsai, 1993) és
makrofágok (Chen és mtsai, 2003) is expresszálják. Nem-neuronális sejtekben fiziológiás
funkciójuk még nem tisztázott (Caterina, 2003).
A pszeudounipoláris szenzoros neuronok sejtteste a hátsógyöki (DRG) és a trigeminális
dúcokban (TRG) helyezkedik el. A fájdalom illetve a hıdetektáló polimodális és a
kimondottan hıre speciálizálódott termoszenzitív, unimodális nociceptorok centrálisan a
hátsószarv interneuronjaival alkotnak szinapszist, a perifériás axonterminálisuk pedig a bır
dermiszében/epidermiszében szabadon végzıdik. A fájdalomkeltı termikus és kémiai
stimulusokkal keltett ingerületet Aδ- illetıleg C-rostok közvetítik. A legtöbb kapszaicin-
szenzitív nociceptor C-rostú. A szenzoros neuronokat szövettani valamint neurobiokémiai
tulajdonságaik alapján két fı csoportba (A és B típusú sejtek) lehet sorolni. A sötéten festıdı,
kisebb átmérıjő, vékonyabb axonú B-típusú sejtek expresszálják a kapszaicin receptorát.
A kapszaicin (és analógjai) a kapszaicin-szenzitív primer afferens neuronokon azonnali
excitátoros (algézia), nagyobb koncentrációban alkalmazva deszenzitizáló (analgetikus),
illetve irreverzibilis neurotoxikus hatást fejtenek ki (Szolcsányi 1993, 2002, 2005; Sándor &
Szállási, 2005).

7
Napjainkban már több olyan szintetikus (pl: olvanil, rinvanil) valamint a természetben is
elıforduló kapszaicin-analógot is ismerünk (pl: a fekete bors és a gyömbér hatóanyaga, a
piperin és a gingerol), melyek a fájdalomérzékelı idegvégzıdéseket a kapszaicin csatorna
kinyitásával izgatják, de mind izgató, mind pedig deszenzitizáló hatásuk mértékében
különböznek egymástól. További példaként említeném az ultrapotens kapszaicin-analóg
reziniferatoxint (RTX; a Marokkóban ıshonos kaktuszszerő növény, az Euphorbia rezinifera
aktív hatóanyaga), mely a kapszaicinnél több nagyságrenddel hatásosabb izgató és aránylag
kis koncentrációban már neurotoxikus (Appendino & Szállási, 1997).
A deszenzitizáció világszerte intenzív kutatások tárgya a szenzoros neuronokra szelektíven
ható fájdalomcsillapító illetve gyulladáscsökkentı gyógyszerek kifejlesztése céljából. A
deszenzitizáció fogalmát kétféle értelemben szokás használni. Egyiknél a receptormolekula
agonista iránti érzékenysége csökken (farmakológiai deszenzitizáció), a másiknál pedig a
TRPV1 receptort hordozó idegvégzıdés lesz természetes ingereire érzéketlen (szenzoros vagy
funkcionális deszenzitizáció). Mivel a funkcionális változásokat morfológiai tünetek kísérhetik
(az enyhe mitokondriális duzzadástól a neurondegenerációig), a szenzoros afferensen ható
fájdalomcsillapítók kifejlesztésére irányuló próbálkozásoknál lényeges szempont, hogy az
analgetikus potenciállal bíró molekula milyen mértékben izgat, deszenzitizál, illetve
neurotoxikus.
1.3. A TRPV1/VR1 kapszaicin receptor bemutatása, jelentısége
Az eredetileg VR1-nek keresztelt kapszaicin-érzékeny receptormolekula (vanilloid receptor 1-
es típus; Caterina és mtsai, 1997) olyan ligandfüggı, nem szelektív kalciumcsatorna
(PCa:PNa=9:1), melynek direkt aktivátorai a különbözı vanilloid származékok, a fájdalmas
hıinger (Szállási & Blumberg, 1999) és az extracelluláris pH savas irányba történı eltolódása
(Tominaga és mtsai, 1998). A legújabb IUPHAR (International Union of Pharmacology)
nomenklatúra szerint a kapszaicin receptorát TRPV1 névvel illetik (transient receptor
potential vanilloid 1-es típus; Clapham és mtsai, 2003).
A TRPV1 kapszaicin receptor eddig felderített funkciója egyrészt a fájdalmas termikus és
kémiai stimulusok detektálása polimodális nociceptorokban (Caterina és mtsai, 2000),
másrészt szerepe van a gyulladás által kiváltott termális hiperalgéziában (Davis és mtsai,
2000). A hipotalamusz preoptikus areájában található TRPV1 receptorok a hıszabályozás
termodetektorai (Jancsó-Gábor és mtsai, 1970; Szolcsányi és mtsai, 1971).

8
A TRPV1 receptor olyan polimodális integratív funkciójú membránprotein, mely szerkezeti
hasonlóságot mutat az ecetmuslica (Drosophila Melanogaster) TRP ioncsatorna család
tagjaival. A TRP elnevezés a „tranziens receptor potenciál” kezdıbetőibıl kirakott
mozaikszó. Az 1970-es években megfigyelték, hogy egy mutáns Drosophila retinájában a
kalcium permeábilis receptormolekula folyamatos fényexpozícióra foszfolipáz C aktiválásán
keresztül folytonos receptorpotenciált mutat. Innen származik az elnevezés a késıbb megtalált
molekuláris-szerkezeti rokonság alapján (Clapham és mtsai, 2001; Montell és mtsai, 2002).
A TRPV1 receptor 9 évvel ezelıtti klónozása igazi tudományos áttörésnek bizonyult
(Caterina és mtsai, 1997) és egyhamar a nemzetközi kutatások élvonalába került. Mivel a
TRPV1 génhiányos egerekben a normális mechanikai érzékenység mellett némi termális
érzekenység is megmaradt, gyaníthatóvá vált más hımérsékleti tartományokban érzékeny
hıszenzorok létezése (Caterina és mtsai, 2000; Davis és mtsai, 2000). Ez a megfigyelés
nyitotta meg az utat a hıingerekkel aktiválható, szekvencia homológiát mutató, újabb
ioncsatornák felfedezéséhez (Gunthorpe és mtsai, 2002; Patapoutian és mtsai, 2003; Montell
és mtsai, 2003; Jordt és mtsai, 2003). A TRPV alcsalád jelenleg ismert hat tagja közül négy
(TRPV1-4) hıre is érzékeny kationcsatorna, melyek hımérsékleti/aktiválhatósági tartománya
részleges átfedést mutat. A TRPV5 és a TRPV6 pedig a kalcium abszorbcióban/reszorbcióban
közremőködnek vese és duodénum epitéliumban.
A robbanásszerő fejlıdést mi sem bizonyítja jobban, hogy már az egyes TRP molekulák
közötti filogenetikai kapcsolatokat is feltérképezték. Két további alcsalád (TRPC, TRPM)
külön-külön 6-8 génjét is azonosították és 2004-ben újabb homológ géncsaládokat
identifikáltak, a mukolipineket (TRPML) és a policisztineket (TRPP), melyek változatos
funkciókat töltenek be a Caenohabditis eleganstól a Homo sapiensig (pl.
feromonszignalizáció, hímfertilitás, Mg2+ homeosztázis, hıszabályozás, ízérzékelés és egyéb
szenzoros funkciók). Ma már a TRP receptorcsalád mintegy 30 tagja ismeretes.
A sajátos hımérsékleti profillal bíró termo-TRPV proteinek kivétel nélkül 6 transzmembrán
doménból szervezıdnek, intracellulárisan elhelyezkedı amino- és karboxi-terminálissal (1.
ábra).
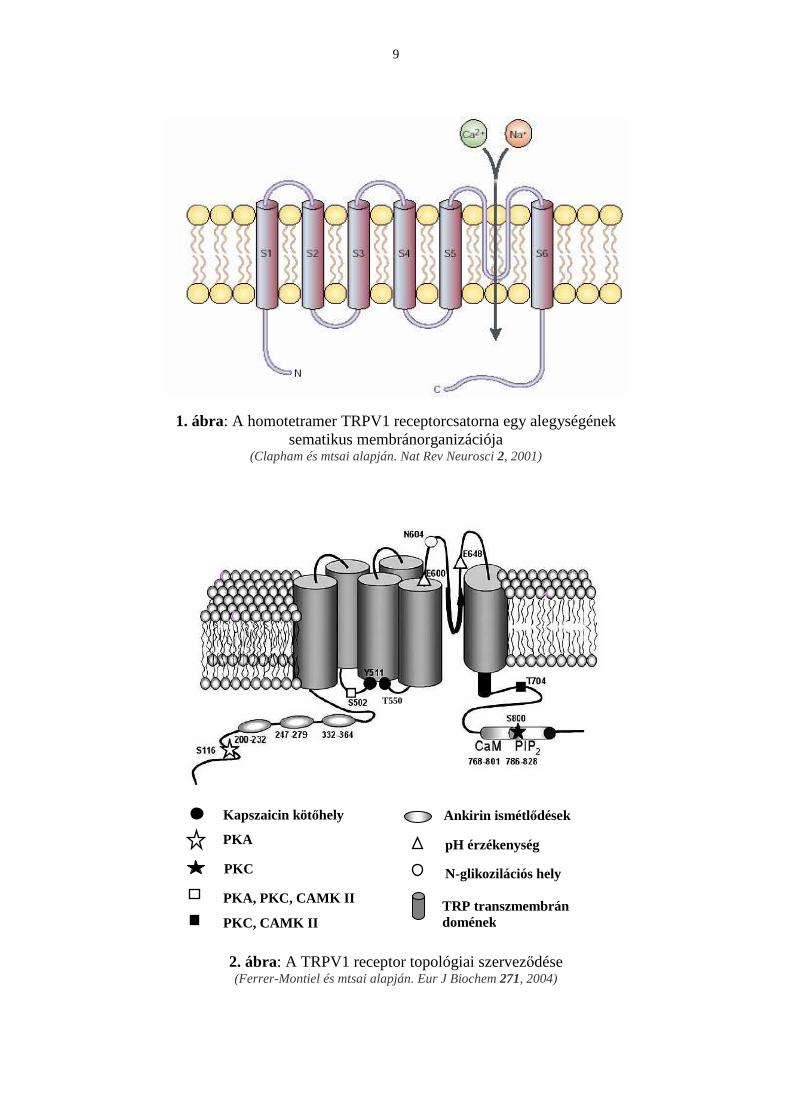
9
1. ábra: A homotetramer TRPV1 receptorcsatorna egy alegységének sematikus membránorganizációja
(Clapham és mtsai alapján. Nat Rev Neurosci 2, 2001)
2. ábra: A TRPV1 receptor topológiai szervezıdése (Ferrer-Montiel és mtsai alapján. Eur J Biochem 271, 2004)
Kapszaicin kötıhely Ankirin ismétl ıdések
PKC
PKA, PKC, CAMK II
N-glikozilációs hely
PKA pH érzékenység
PKC, CAMK II TRP transzmembrán domének
T550

10
A homotetramer szerkezető TRPV1 receptor modulálásában résztvevı kitüntetett
aminosavcsoportokat deléciókkal, pontmutációkkal határozták meg. A humán és a patkány
TRPV1 közötti különbségek elsısorban az N- és C-terminális régiókra esnek, míg az erısen
konzervált régiókban (transzmembrán domének, ankirin ismétlıdések, foszforilációs helyek;
2. ábra) nagy a hasonlóság.
A kapszaicin intracelluláris kötıdése a 3. és a 4. transzmembrán domén meghatározott
aminosavjaihoz (Y511 és T550) csatorna megnyílást okoz (Jordt & Julius, 2002; 2. ábra). Az
ultrapotens analógként ismert reziniferatoxin (RTX) kötéséhez az 547-es pozícióban lévı
metionin is szükséges (Gavva és mtsai, 2004). Ezek az aminosavcsoportok vesznek részt az
endogén agonisták és a kompetitív antagonisták kötıdésében is.
A TRPV1 receptor két extracelluláris, negatívan töltött glutamát csoportja felelıs a receptor
proton-érzékenységéért (E600 és E648; Jordt és mtsai, 2000; 2. ábra). Az E600-as aminosav
a protonnal kiváltott TRPV1 receptor szenzitizációjában lényeges, az E648-as pedig a TRPV1
allosztérikus aktivációjában és a kötıdés által megvalósított kalcium-influxban (Holzer,
2004). Az extracelluláris proton koncentrációjának növekedése a hıaktivált áramokat is
potencírozza (Tominaga és mtsai, 1998). A fiziológiás pH 6.4-re illetve 5.9-re csökkentése a
TRPV1 receptor hıküszöbét 43 ºC-ról 37 ºC-ra illetve 22 ºC-ra módosítja.
A TRPV1 csatorna hıküszöbe fiziológiás körülmények között 42-43 °C, azonban
gyulladáskor, szöveti sérüléskor szintetizálódó és/vagy liberalizálódó endogén
proinflammatorikus peptidek, bioaktív anyagok (bradikinin, hisztamin, ATP, tripszin, stb)
különbözı érzékenyítı mechanizmusok bevonásával (protein kináz stimulálás, foszfatidil-
inozitol-trifoszfát (PIP2) hidrolízis, TRPV1-kalmodulin interakció gátlás) csökkentik a
TRPV1 csatorna hıküszöbét.
A TRPV1 receptor C-terminális doménjének lehasításával (~ 160 aminosavnyi darab) a
csatorna elvesztette fı funkcióját, specifikus termális érzékenységét (Vlachova és mtsai,
2003). A TRPV1 receptor karboxi-végén kalmodulin-, PIP2- és nukleotidkötı-domén
található (Kwak és mtsai, 2000; 2. ábra). A PIP2 hidrolízise felszabadítja a TRPV1 receptort a
PIP2-mediált gátlás alól (Chuang és mtsai, 2001; Liu és mtsai, 2005; Prescott & Julius, 2003).
A bradikinin (BK) és az idegnövekedési faktor (NGF) szenzitizáló hatása részben ilymódon
érvényesül.
A kalmodulin (CaM) kalciumdependens módon inaktiválja a receptort azáltal, hogy kötıdik a
csatorna C-terminálisának megfelelı szegmenséhez (2. ábra). E kötıdés blokkolásakor a
receptor deszenzitizáció is gátlódik (Numazaki és mtsai, 2002).

11
Érdekes, hogy emberben állítólag az 585-ös pozícióban lévı valin/izoleucin polimorfizmus
felelıs a genetikusan öröklödı, eltérı fájdalomtőrı képességért, de ez a szubsztitúció nem
befolyásolja a receptor struktúrát/funkciót (Hayes és mtsai, 2000).
A teljesség kedvéért még megemlíteném, hogy léteznek más hıszenzorok is a TRPV
proteineken kívül, melyek szintén fontosak a termális érzékenységben. A meleg érzetek
közvetítésben a P2X3 ionotróp purinoceptor (Souslova és mtsai, 2000), míg a Na/K ATPáz
(Pierau és mtsai, 1974), a TRPA1 (ANKTM1; McKemy, 2005), a TRPM8 (McKemy, 2005),
valamint a degenerin családba tartozó proton-szenzitív csatornák (ASICs; Askwith és mtsai,
2001) alapfunkciójukon kívül a hidegingerek detektálásban is közremőködnek.
1.4. Népi gyógyászat vagy modern farmakológia?
Meglepı módon ma még nem rendelkezünk nociceptor-szelektív fájdalomcsillapítóval. A
fájdalomcsillapításra használt szerek a több mint százéves acetilszalicilsav és rokonai,
valamint a morfin és származékai (Szolcsányi, 2004, 2005). A csípıs paprika hatóanyagát
jelenleg még elsısorban a fájdalomérzés mechanizmusainak kutatásában alkalmazzák, de a
neuropathiás fájdalom csillapítására helyileg alkalmazott kapszaicinnel már több tanulmány is
ígéretes eredményt kapott (Bley & Malmberg, 2005).
A népi gyógyászat már régóta elixírként tartja számon. Évszázados megfigyelés, hogy a
pirospaprika rágcsálása enyhíti az izom és ízületi, valamint az övsömör-okozta fájdalmakat. A
cayenne paprika tea fogyasztása kitömíti az eldugult orrot, hatékonynak bizonyult
teniszkönyökre, derékbántalmakra. Az úgynevezett Capsadermától posztherpetikus
neuralgéziában remélhetünk enyhülést. Újabban léteznek kapszaicin tartalmú gyógyhatású
készítmények (orrspray formájában) migrénre, allergiára.
A kapszaicin termogén hatását is régóta kiaknázzák. A turisták és állítólag régen a cowboyok
is cipıjükbe szórták a cayenne/gyömbér/mustármagpor keverékét, lábuk melegen tartása
végett. Mivel a kapszaicin és a többi csípıs vegyület izzadás révén hıveszteséget vált ki, ezért
melegebb égövön szívesebben fogyasztanak erısebben főszerezett ételeket. A Recongem
paprikás pakolást sikeresen alkalmazzák cellulitisz kezelésére, valamint ránctalanításra, mely
a kapszaicin vérkeringést fokozó hatásának tulajdonítható. A kapszaicin a test metabolikus
rátájának növelése által fogyasztó hatású, így potenciális testsúlycsökkentı terápiaként is
kecsegtet. Utóbbi tulajdonságát kihasználva a „Master Cleaner” elnevezéső kapszaicin
tartalmú ital Hollywoodban hódít. Az USA-ban kapható egy olyan paprikaırleménybıl
készült édesítıtabletta, mely állítólag hatékony keringési, emésztési próblémák orvoslásában.

12
A kapszaicin TRPV1-et kifejezı sejtvonalak elıállítása megnyitotta az utat a nagyüzemi
gyógyszerkutatások elıtt, mivel e sejteken a kapszaicinnel kiváltott válaszok (többnyire
kalciumion-akkumuláció) vagy a kötıdés blokkolása in vitro sorozatvizsgálatokat tesz
lehetıvé. Ezzel az un. nagy hatékonyságú sorozatvizsgálattal (high throughput screening,
HTS) napi több tízezer kapszaicin antagonista/agonista vegyület hatásának tesztelése
valósulhat meg. Terápiásan egyaránt ígéretesnek bizonyulhat a különbözı TRPV1
antagonisták identifikálása, illetve azon parciális agonisták alkalmazása, melyek a TRPV1
csatorna megnyílását csak olyan mértékben váltják ki, mely akciós potenciálokat még nem
generál, de az intracelluláris másodlagos hatásaik következtében (kalcium beáramlás,
mitokondriális effektus) az érzıidegvégzıdés funkcionális blokkolását idézik elı.
A kifejlesztendı gyógyszer remélt fájdalomcsillapító és gyulladáscsökkentı hatását tehát a
nociceptorokon ható blokkolástól várják (ld. fentebb). Eddig a kapszaicint használták erre a
célra.
Topikális alkalmazása krónikus fájdalmak csillapítására ma már bizonyítottnak tekinthetı. A
húgyhólyagba adott kapszaicin illetve RTX a neurogén hólyag szenzoros afferentációjának
deszenzitizálásával ígéretes gyógyírnek tőnik inkontinencia kezelésében. Külföldön már
kétféle magas potenciájú kapszaicin krém is beszerezhetı, a 0.025% kapszaicin tartalmú
Zostrix és a 0.075%-os Axazin. (Összehasonlításként megemlíteném, hogy a kutyariasztó
gázspray aktív hatóanyag tartalma 0.35%.)
Kapszaicin és helyi érzéstelenítı kombinációval alkalmazható bırtapasz és intraartikulárisan
adható injekciós készítmény. Klinikai vizsgálatára két független amerikai cég eddig már 120
millió USA dollárt költött.
Napjainkra már nyilvánvaló, hogy a kapszaicin-érzékeny elsıdleges érzıneuronok aktivációja
illetve a következményes perifériás neuropeptid felszabadulás eredményeként kialakuló
neurogén gyulladásos komponens számos betegség patomechanizmusában játszik szerepet
(asztma, reumatoid artritis, allergiás rhinitis, migrén).

13
1.5. Az értekezés célkitőzései és tematikai felosztása
Az értekezésben bemutatott munka fı célja a TRPV1 receptorok által közvetített
intracelluláris kalciumkoncentráció növekedéssel járó különbözı szabályozó mechanizmusok
mélyebb feltárása és a TRPV1 csatornafehérje farmakológiai tanulmányozása, egyrészt
újszülött patkányok trigeminális neurontenyészetén, másrészt patkány TRPV1 receptorral
transzfektált HT1080-as fibroszarkóma sejteken, valamint különbözı in vivo
állatmodellekben.
Munkám 5 fı téma köré csoportosítható:
I. Az új TRPV1 receptor antagonista SB366791 hatásainak in vitro és in vivo
farmakológiai vizsgálata
I./1. A. Varga, J. Németh, Á. Szabó, J. J. McDougall, C. Zhang, K. Elekes, E. Pintér, J.
Szolcsányi, Zs. Helyes: Effects of the novel TRPV1 receptor antagonist SB366791 in vitro
and in vivo in the rat
Neuroscience Letters 385, pp.137-142, 2005
I./2. K. Sándor, Zs. Helyes, A. Varga, Á. Szabó, K. Bölcskei, J. Németh, K. Elekes, G.
Pozsgai, E. Pintér, J.J. McDougall, C. Zhang, G. Pethı and J. Szolcsányi: Examination of
novel TRPV1 receptor antagonists in vitro and in vivo in the rat
Inflammopharmacology 2005 (elıadáskivonat)
II. Az N-oleoildopamin és analógjainak in vitro hatáselemzése
II./1. J. Szolcsányi, Z. Sándor, G. Pethı, A. Varga, K. Bölcskei, R. Almási, Z. Riedl, G.
Hajós, G. Czéh: Direct evidence for activation and desensitization of the capsaicin receptor by
N-oleoildopamine on TRPV1-transfected cell line in gene deleted mice and in the rat
Neuroscience Letters 361, pp. 155-158, 2004

14
II./2. A. Varga, K. Bölcskei, Z. Sándor, R. Almási, G. Pethı, G. Czéh, Z. Riedl, G. Hajós, J.
Szolcsányi: Studying of OLDA as endogenous ligand for TRPV1 capsaicin receptor in vitro
and in vivo
Acta Physiologica Hungarica 91 (3-4), pp. 372, 2004 (elıadáskivonat)
II./3. Z. Sándor, A. Varga, K. Bölcskei, P. Horváth., Z. Riedl, G. Hajós, J. Szolcsányi:
Activation and desensitization of the TRPV1 capsaicin receptor by substituted N-
oleoylphenylethylamines
Clinical Neuroscience 2005 (elıadáskivonat)
II./4. A. Varga, K. Bölcskei, Z. Sándor, R. Almási, G. Pethı, Z. Riedl, G. Hajós, G. Czéh, J.
Szolcsányi: Effects of the substituted N-oleoylphenylethylamines on activation and
desensitization of the TRPV1 capsaicin receptor
Inflammopharmacology 2005 (elıadáskivonat)
III. A protein kináz A és C relatív szerepének vizsgálata a TRPV1 receptor
érzékenységének modulálásában
A. Varga, K. Bölcskei, É. Szıke, R. Almási, G. Czéh, J. Szolcsányi, G. Pethı: Relative roles
of protein kinase A and protein kinase C in modulation of TRPV1 receptor responsiveness in
rat sensory neurons in vitro and peripheral nociceptors in vivo
Neuroscience 2006 (elfogadva)
IV. A TRPV1 receptor-mediált intracelluláris kalciu mionszint emelkedés vizsgálata
extracelluláris kalcium hiányában
Z. Sándor, A. Varga, P. Horváth, B. Nagy, Z. Szilvássy, J. Szolcsányi: Construction and
analysis of a new stable cell line expressing the rat TRPV1 receptor
Cellular and Molecular Biology Letters 10, pp. 499-514, 2005
V. A TRPV1 kapszaicin receptor aktiválás és a mitokondrium funkció közötti viszony
vizsgálata

15
V./1. A. Varga, Z. Sándor, Z. Balla, G. Czéh, J. Szolcsányi: Relative changes of intracellular
calcium level and mitochondrium membrane potential simultaneously induced by capsaicin
(közlésre elıkészített kézirat)
V./2. Varga A., Szıke É., Balla Z., Czéh G., Szolcsányi J.: Parallel measurement of
rhodamine 123 and fura-2 in capsaicin-stimulated sensory neurones
Clinical Neuroscience 56 (2), pp. 93-94, 2003 (elıadáskivonat)
V./3. Czéh G., Varga A., Sándor Z., Szolcsányi J.: Calcium, mitokondrium, capsaicin
Acta Physiologica Hungarica 91 (3-4), pp. 284-285, 2004 (elıadáskivonat)

16
2. MÓDSZERTAN
2.1. Sejttípusok
2.1.1. Patkány trigeminális ganglionsejt tenyészet készítése Szıke és mtsai leírása alapján
(2000).
A 2-5 napos újszülött Wistar patkányokat éteres altatás mellett dekapitáltuk, kétoldali
trigeminális ganglionjukat steril foszfát pufferben (PBS) kipreparáltuk, 1 mm3-es darabokra
vágtuk. A gangliondarabokat 30-35 percre 0.8 mg/ml XI. típusú kollagenázt tartalmazó PBS-
be tettük, majd 8 percen keresztül 600 egység/ml deoxiribonukleáz I-gyel inkubáltuk 37 °C-
on, 5% CO2-tartalom mellett. Triturálás után a sejteket steril, 24-lyukú tenyésztıedénybe
pipettáztuk, melyekbe elızıleg 100 µg/ml poli-D-lizinnel elıkezelt 12 mm-es átmérıjő, steril
üvegkorongokat helyeztünk (4 °C, 24 óra). A tenyésztımédium a következı összetevıkbıl
állt: 170 ml Dulbecco′s Modified Eagle Medium (DMEM), 10-10 ml ló szérum, magzati
szarvasmarha és újszülött borjú szérum, 100 µg/ml streptomicin, 100 NE/ml penicillin. A
tápoldathoz 7 ng/ml idegnövekedési faktort (NGF) adtunk a maximális TRPV1 receptor
expresszió céljából. A tápoldatot és az NGF-et kétnaponta cseréltük. A kísérletekhez 1-7
napos tenyészeteket használtunk.
2.1.2. A patkány TRPV1 receptort stabilan expresszáló HT5-1 sejtvonal (a továbbiakban:
rTRPV1-HT5-1) HT1080 humán fibroszarkóma sejtvonalból származik (Sándor és mtsai,
2005). A TRPV1 cDNS klónozása és a humán sejtvonal transzfekciója nem képezi Ph.D
disszertációm anyagát.
2.2. Mikrofluorimetriás technikák
2.2.1. Intracelluláris kalciumionszint-mérés fluoreszcens módszerrel
Módszertani szempontból kutatásaink egyik fı pillére a szelektív fluoreszcens festékek
használatára alapozott, relatív intracelluláris kalciumionszint-változás mérése (Szıke és mtsai,
2000). Ennek méréstechnikáját intézetünkben Dr. Szıke Éva állította be, de a kisebb-nagyobb
metodikai módosításaink az eljárás ismételt leírását indokolják.
Az 1-7 napos TRG tenyészeteket fura-2-acetoxy-metilészter (fura-2; 1 µM) fluoreszcens
kalciumindikátor festéket tartalmazó oldatban (összetételét lásd az 1. táblázatban) inkubáltuk
15 percig, 37 °C-on üzemeltetett termosztátban. A festési idıt rTRPV1-HT5-1 sejtek esetén
17
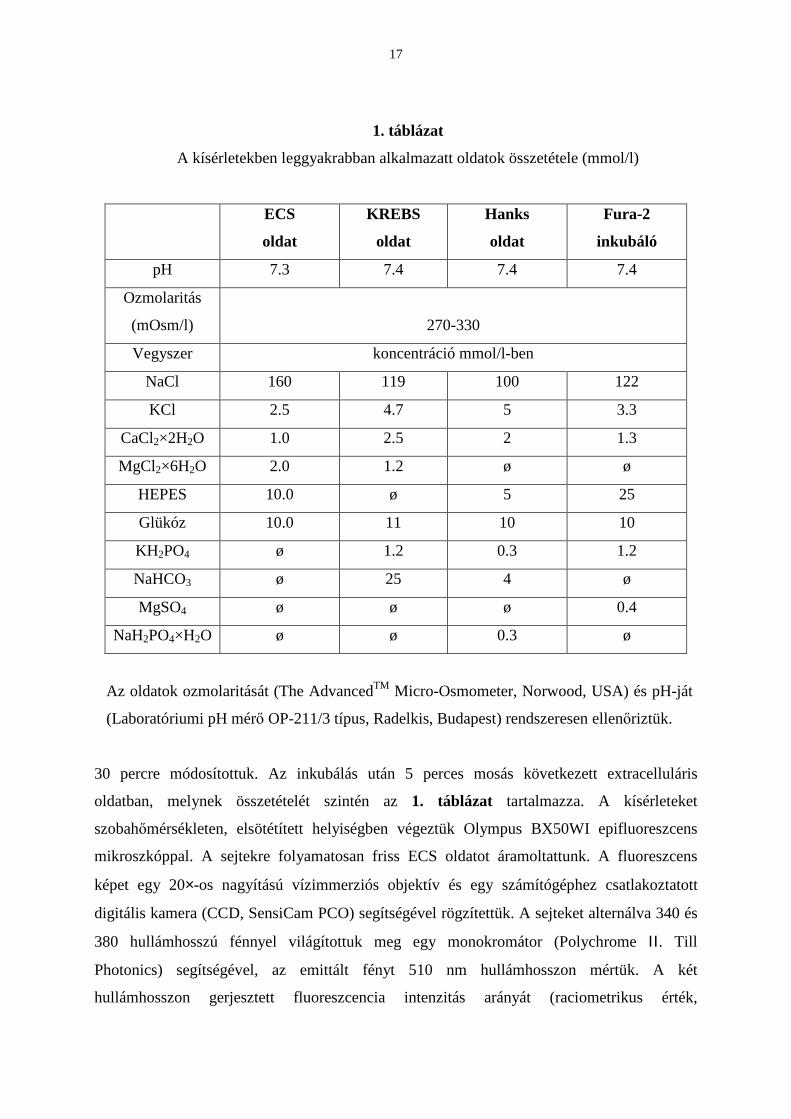
1. táblázat
A kísérletekben leggyakrabban alkalmazatt oldatok összetétele (mmol/l)
ECS
oldat
KREBS
oldat
Hanks
oldat
Fura-2
inkubáló
pH 7.3 7.4 7.4 7.4
Ozmolaritás
(mOsm/l)
270-330
Vegyszer koncentráció mmol/l-ben
NaCl 160 119 100 122
KCl 2.5 4.7 5 3.3
CaCl2×2H2O 1.0 2.5 2 1.3
MgCl2×6H2O 2.0 1.2 ø ø
HEPES 10.0 ø 5 25
Glükóz 10.0 11 10 10
KH2PO4 ø 1.2 0.3 1.2
NaHCO3 ø 25 4 ø
MgSO4 ø ø ø 0.4
NaH2PO4×H2O ø ø 0.3 ø
Az oldatok ozmolaritását (The AdvancedTM Micro-Osmometer, Norwood, USA) és pH-ját
(Laboratóriumi pH mérı OP-211/3 típus, Radelkis, Budapest) rendszeresen ellenıriztük.
30 percre módosítottuk. Az inkubálás után 5 perces mosás következett extracelluláris
oldatban, melynek összetételét szintén az 1. táblázat tartalmazza. A kísérleteket
szobahımérsékleten, elsötétített helyiségben végeztük Olympus BX50WI epifluoreszcens
mikroszkóppal. A sejtekre folyamatosan friss ECS oldatot áramoltattunk. A fluoreszcens
képet egy 20×-os nagyítású vízimmerziós objektív és egy számítógéphez csatlakoztatott
digitális kamera (CCD, SensiCam PCO) segítségével rögzítettük. A sejteket alternálva 340 és
380 hullámhosszú fénnyel világítottuk meg egy monokromátor (Polychrome ΙΙ. Till
Photonics) segítségével, az emittált fényt 510 nm hullámhosszon mértük. A két
hullámhosszon gerjesztett fluoreszcencia intenzitás arányát (raciometrikus érték,

18
R=F340/F380) az idı függvényében az Axon Imaging Workbench 2.1 szoftverrel
regisztráltuk, a kapott függvényeket a Microcal Origin 7.0 program segítségével értékeltük.
Az R értékbıl valós intracelluláris kalcium koncentrációt ([Ca2+] ic) nem számoltunk. Minden
egyes vonatkozó fejezetben külön részletezem a Microcal Origin 7.0 program alapján számolt
statisztikát és az egyes szignifikanciaszintek teljesülését.
2.2.2. A relatív intracelluláris szabad kalciumionszint és a mitokondriális
membránpotenciál változás szimultán mérése
Ennek a technikának a bevezetése, helyi viszonyainkra történı adaptálása saját feladatom
volt.
A kísérleteket szobahımérsékleten, elsötétített helyiségben végeztük TRG neuronokon és
rTRPV1-HT5-1 sejteken Olympus BX50WI fluoreszcens mikroszkóppal. Elızıleg az 1-7
napos TRG tenyészeteket valamint az 1 napos rTRPV1-HT5-1 sejteket a fluoreszcens
kalciumindikátorral (fura-2; 1 µM) és a pozitív töltéső rhodamin 123-mal (rhod-123; 26 µM)
egyidıben festettük meg 30 percig, 37 °C-on.
A lipofil kationfesték rhod-123 a mitokondriális depolarizációt nonraciometrikusan méri.
Pozitív töltései révén a negatív membránpotenciálú mitokondriumban kioltott
fluoreszcenciával akkumulálódik, majd egy depolarizáló impulzus alkalmával a festék
reverzibilisen, a mitokondriális membránpotenciál változását követve kikerül a citoszolba
(redisztribúció). A rhod-123 citoplazmába történı kiszabadulása (fluoreszcencia
intenzitásának emelkedése) a mitokondrium belsı membránjának depolarizációját jelzi
(Emaus és mtsai, 1986; White & Reynolds, 1995).
A fluoreszcens képet egy 20×-os nagyítású vízimmerziós objektív és egy számítógéphez
csatlakoztatott digitális kamera (CCD, SensiCam PCO) segítségével rögzítettük a 2.2.1
alfejezetnél ismertetett módon. Egy monokromátor (Polychrome ΙΙ. Till Photonics)
segítségével a sejteket három különbözı hullámhosszon világítottuk meg a festékmolekulák
kielégítı gerjesztése céljából. A fura-2 indikátor raciometrikus sajátsága miatt alternált
megvilágítást igényelt (340 és 380 nm), míg a nonraciometrikus rhod-123 expozíciója 488 nm
hullámhosszon történt. Az illumináció idıintervalluma a fura-2-nél 80 és 400 msec között
volt, míg a rhod-123-nál 80-200 msec-ig terjedt. Az emittált fényt 510 nm-nél 1.2
másodpercenként szimultán győjtöttük. A megközelítıen 1 Hz-es mintavételezés maximum
15 percig tartott. A gerjesztett fluoreszcencia intenzitások arányát (raciometrikus érték,
R=F340/F380), illetve a nonraciometrikus próba fluoreszcencia változását (∆F) az idı

19
függvényében az Imaging Workbench 4.0 szoftver automatikusan, szimultán regisztrálta. Az
R értékbıl valós intracelluláris kalcium koncentrációt ([Ca2+] ic) nem számoltunk. Fontosnak
tartom hangsúlyozni, hogy a deltaF érték gyakorlatilag nem kalibrálható a membránpotenciál
változás szempontjából, mivel a rhod-123 az egy-egy sejtben tömörült, összes mitokondrium
átlagolt membránpotenciál változását követi. A kapott függvényeket a Microcal Origin 7.0
program segítségével értékeltük.
2.3. A Kalciumizotóppal (45
Ca2+
) végzett kísérletek kivitelezését Wood és munkatársai
1988-as publikációja alapján végeztük és mindössze 2 éve vezettük be intézetünk in vitro
metodikáinak fegyvertárába. Az eljárás honosításában tevékenyen résztvettem. A
mikrofluorimetriás kísérletek hátránya, a sejtes válaszok nagymértékő heterogenitása
küszöbölhetı ki ezzel a technikával, mely egyszerre képes többezer sejt együttes reakcióját
CPM (count per minute) értékben detektálni.
Egy nappal az izotópos kísérletek elıtt 72-lyukú mini tenyésztıedénybe 15-15 µl-es térfogatú
DMEM-be kiplételtük a rTRPV1-et expresszáló HT5-1 sejteket (kb. 4000 sejt/lyuk). Másnap
a letapadt klónsejteket ötször mostuk 2 mM kalcium-kloridot tartalmazó Hank's oldattal
(összetételét lásd 1. táblázatban), majd a kívánt koncentrációjú TRPV1 receptor agonista és
200 µCi/ml 45
Ca2+ izotóp elegyét mértük a sejtekre. A 10 perces együttinkubálás 37 °C-on
történt. Az alapos, de óvatos ötszöri ECS mosás végeztével a sejteket tartalmazó mini
tenyésztıedényt kiszárítás céljából 75 °C-ra helyeztük, majd 10 µl 0.1%-os SDS-sel lizáltuk a
sejteket, hogy az általuk akkumulált 45
Ca2+ izotópot maradéktalanul összegyőjtsük. A lizátum
radioaktivitását 2 ml szcintillációs folyadékban Packard Tri-Carb 2800-as TR számlálóval
mértük.
A Delree-féle percoll-grádiensen alapuló szenzoros neuron dúsítási, tisztítási protokoll
eredménytelensége folytán a TRG sejtekkel végzett izotópos próbálkozásaink nem jártak
sikerrel a jelentıs sejtveszteség és az elégtelen neuron/gliasejt arány miatt (Delree és mtsai,
1989).
2.4. P-anyag felszabadulásának vizsgálata in vitro tracheapreparátumon
A nátrium-pentobarbitállal (40 mg/kg, i.p.) altatott patkányok tracheáit kimetszettük és
szervfürdınként (1.8 ml) 2-2 tracheát 37 °C-os oxigenizált Krebs oldattal (95% O2 és 5%
CO2) 1 órán át perfundáltuk (1 ml/perc; ld. 1. táblázat). Az átáramlás leállítása után a
kamrákban lévı oldatot 8 percenként lecserélve három frakciót győjtöttünk (ingerlés elıtti -

20
ingerelt - ingerlés utáni). A peptidfelszabadulást kémiai stimulációval (10-6 M kapszaicinnel)
vagy elektromos téringerléssel (paraméterek: 40 V, 0.1 ms, 10 Hz, 2 perc; 1200 impulzus)
váltottuk ki a második 8 perces periódus kezdetén. A TRPV1 antagonista SB366791-et (0.1
vagy 0.5 µM) minden nyolc perces periódus elején adtuk az inkubációs médiumhoz.
Értelemszerően, a kontroll kísérletekben az ingerlést az antagonista hiányában végeztük. A
mintákat jégbe hőtött csövekbe győjtöttük és a procedúra végén a légcsıdarabok nedves
súlyát lemértük. Az egyes frakciók P-anyag (SP) koncentrációit intézetünkben Németh József
és kollégái által kifejlesztett specifikus RIA módszer segítségével határoztuk meg (Németh és
mtsai, 1999), melynek érzékenysége 2 fmol/csı. Az SP mennyiségét fmol/mg egységben,
nedves szövetsúlyra vonatkoztatva fejeztük ki. Az eredmények statiszikai értékelése Student-
féle páratlan t-teszttel történt.
2.5. In vivo kísérletek
2.5.1. Az SB366791 hatása a kapszaicinnel kiváltott térdízületi vérátáramlás-
fokozódásra
Az uretános (2g/kg, i.p.) altatásban a lábelhúzási (flexor) reflex hiányában kanült vezettünk a
tracheába a kielégítı légzésbiztosítás valamint a jobb oldali artéria karotiszba a közvetlen
vérnyomásmonitorozás céljából. A jobb oldali térdízületet borító bırterületet leborotváltuk és
az alatta lévı kötıszöveti elemeket eltávolítottuk. A kapszaicin (100 µg/kg, 100 µl)
térdízületre történı lokális alkalmazását követıen 35 percen keresztül, 5 percenként
monitoroztuk a szinoviális véráramlást lézer Doppler áramlásmérı mőszerrel (kontroll
csoport). Az állatok másik csoportja 20 perccel a kapszaicin adása elıtt intraperitoneális
SB366791 (100 vagy 500 µg/kg) vagy a referencia vegyület kapszazepin (CZP, 2 mg/kg)
elıkezelésben részesült. Az eredmények statiszikai értékelésére egyszempontú ANOVA
tesztelést követıen Dunnett’s post próbát alkalmaztunk.
2.5.2. Az SB366791 hatása a kapszaicinnel kiváltott testhımérséklet csökkenésre
Régóta ismert a kapszaicin maghımérsékletet csökkentı hatása a perifériás és centrális
melegszenzorok stimulálásán keresztül (Jancsó-Gábor és mtsai, 1970). A kapszaicin (300
µg/kg, s.c.) okozta hipotermiát rektális hımérızéssel 10 percenként, összesen 6 alkalommal
(azaz 1 órán keresztül) mértük hím Wistar patkányokban (180-220 g), valamint Balb/c
egerekben (~20 g). Az állatok másik csoportját 20 perccel a kapszaicin adása elıtt
intraperitoneálisan elıkezeltük az SB366791 (100 vagy 500 µg/kg, egereknél: 1 mg/kg) vagy
a CZP (2 mg/kg, illetve egereknél: 5 mg/kg) megfelelı oldatával. A kontroll csoport egyedei

21
az antagonisták oldószerét ugyanolyan térfogatban i.p. kapták. Az eredmények statiszikai
értékelése Mann-Whitney U-teszttel történt.
2.5.3. Kapszaicinnel kiváltott szemtörlési reflex vizsgálata
A bal szembe történı kapszaicin cseppentést (10 µg/ml, 50 µl) követıen 3 percen keresztül
számoltuk a mellsı lábakkal végzett szemtörlı mozdulatok számát (Szolcsányi és mtsai,
1975). A szemtörlési reflex egy olyan megbízható akut kemonociceptív teszt, mely ugyan
fájdalmas az állatnak, de fájdalomkeltı hatása az alkalmazott meglehetısen alacsony
koncentrációtartományban gyorsan megszőnik (<1 perc). Az SB366791 (100 vagy 500 µg/kg)
vagy a CZP (2 mg/kg) elıkezelés a kapszaicin cseppentés elıtt 20, 60 vagy 120 perccel
korábban történt intraperitoneálisan. A kontroll csoport egyedei az antagonisták oldószerét
ugyanolyan térfogatban, szintén i.p. kapták. Az eredmények statiszikai értékelésére a Mann-
Whitney U-teszt szolgált.
2.5.4. Nociceptív hıküszöb mérése emelkedı hımérséklető forró lap módszerrel
A TRG sejttenyészeten kapott in vitro eredményeinket intraplantáris adást követı nocicepciós
teszttel kívántuk megerısíteni. Ennek egyik lehetséges módja az Almási és kollégái (2003)
által kidolgozott ún. „hot plate” módszer, melynek lényege, hogy a számítógép-vezérelt
fémlap hımérsékletét 30 ºC-ról egyenletes ütemben (12 ºC/perc) emeltük mindaddig, amíg az
állat (120-160 gramm nıstény Wistar patkány) fájdalomra utaló magatartását, azaz nocifenzív
viselkedését nem tapasztaltuk (lábemelés ill. nyalás). RTX intraplantáris injektálásával
váltottunk ki hıküszöbcsökkenést, mert az RTX rövidebb ideig tartó fájdalomreakciót okoz
(173.6±11.5 sec; n=8), mint a kapszaicin. Az RTX által indukált, kb. 20 percig tartó
hıküszöbcsökkenés a talpba adást követı 5. percben éri el a maximumát (ekkora már az RTX
okozta fájdalomreakció lecseng). Az intraplantáris elıkezelés alkalmazásának nagy elınye,
hogy a tesztanyagok hatása lokálisan, a perifériás nociceptorok szintjén érvényesül, így a
lehetséges szisztémás hatás elkerülhetı. A különbözı protein kináz inhibitorok hatásának
tesztelésére az RTX nagyobb (0.5 µM, 100 µl), míg a protein kináz aktivátorok szerepének
tanulmányozására az RTX tízszer alacsonyabb dózisát választottuk (0.05 µM, 100 µl). Az
RTX beadása elıtt az állatok egyik csoportját 5 percre intraplantárisan elıkezeltük a PKC
vagy a PKA modulátorral, míg a másik csoport az enzimmodulátor szolvensét (50 µl) kapta.
Az RTX szolvense sem fájdalomreakciót, sem köszöbcsökkenést nem produkált. Minden
egyes állatot csak egyszer, 1-1 kísérletben használtunk. Az 5. percben mért RTX-indukált
hıköszöbcsökkenést Student-féle páratlan t-teszttel hasonlítottuk össze az enzimmodulátort

22
illetıleg a szolvenst kapott állatokban. Statisztikailag szignifikánsnak a p<0.05 adatokat
tartottuk. Az enzimmodulátorok inhibitoros vagy facilitáló jellegét az RTX-szel kiváltott
küszöbcsökkenésre a következı egyenlet alapján százalékban fejeztük ki: (szolvenssel
kiváltott hıköszöbcsökkenés−enzimmodulátorral kiváltott hıköszöbcsökkenés)/szolvenssel
kiváltott hıköszöbcsökkenés×100, ahol a szolvens ill. az enzimmodulátor által okozott
hıköszöbcsökkenés valójában az 5. percben mért RTX-szel indukált átlagos
küszöbcsökkenésre utal a kezelt illetve a szolvenst kapott állatban.
2.6. Alkalmazott hatóanyagok
Az in vitro kísérletekhez a kapszaicin 10 mM-os törzsoldatát DMSO-val készítettük, in vivo
pedig 1%-os törzsoldatát használtuk (80%-a 0.9%-os NaCl, 10%-a Tween80, a maradék 10%
etanol), melyet fiziológiás sóoldattal, in vitro pedig ECS-sel hígítottunk tovább. A kapszaicin
végkoncentrációja kísérletenként változott.
A forbol 12-mirisztát 13-acetátot (a törzsoldat koncentrációja: 5mg/ml), a forskolint
(1mg/ml), a KT5720-at (50 µg/ml), az SB366791-et (2 mg/ml) és az N-oleoilfeniletilaminokat
(10 mM) DMSO-ban oldottuk fel és vagy ECS-sel vagy fiziológiás sóoldattal hígítottuk
tovább. A staurosporin (100 µg/ml) és a reziniferatoxin törzsoldatát (1 mg/ml) etanollal
készítettük. A vízoldékony dibutiril-ciklikusAMP-t (5 mg/ml), az FCCP-t (2.5 mg/ml) illetve
a chelerithrin-kloridot (5mg/ml) ECS-ben oldottuk. Az 50 mM KCl tartalmú oldatot
módosított összetételő ECS-sel készítettük (20×KCl koncentrációval). A legtöbb vegyszerünk
a Sigmától (St. Louis, MO, USA) származott, kivéve a fluoreszcens festékeket, melyeket a
Molecular Probes cégtıl (Eugene, Oregon, USA), az SB366791-et és az OLDA-t a Tocris
Cookson Ltd-tıl (UK), valamint a KT5720-at, a forskolint, a staurosporint és a chelerithrin-
kloridot pedig az Alomone Labs-tól (Jerusalem, Israel) vásároltuk.

23
3. EREDMÉNYEK
3.1. AZ ÚJ TRPV1 RECEPTOR ANTAGONISTA SB366791 HATÁSAINAK IN VITRO
ÉS IN VIVO FARMAKOLÓGIAI VIZSGÁLATA
Bevezetés
A gyulladáscsökkentı és fájdalomcsillapító vegyületek új generációjának talán legígéretesebb
targetmolekulája a TRPV1 kapszaicin receptor (Szállási & Blumberg, 1999; Szolcsányi,
2002). A C-rostú szenzoros neuronok deszenzitizálása az alapkutatásokon túlmenıen az egyik
legintenzívebben kutatott fájdalomcsillapítási stratégia számos gyógyszergyárban (Novartis,
Bayer, SmithKline, Johnson & Johnson, stb).
Biológiai illetve gyógyászati szempontból a fájdalomcsillapítás célja a nociceptorok
érzékenységének csökkentése. Erre két lehetıség is kínálkozik. Kívánatos volna egy olyan
vegyület kifejlesztése, mely elızetes izgatás nélkül deszenzitizálná az idegvégzıdést, hiszen
az izgalmi fázis hiánya (vagy csekély mértéke) egyben a fájdalmas (égetı) érzés kiiktatását
jelenti. A másik lehetıség a TRPV1 receptormolekula antagonistákkal történı, szelektív
farmakológiai gátlása. Ez utóbbi folyamat potenciálisan új típusú analgetikus
hatásmechanizmus alapja.
Jelenleg csak kevés adat áll rendelkezésünkre a TRPV1 antagonisták in vivo hatásaival
kapcsolatosan, pedig egy potens és szelektív blokkolóra nagy szükség lenne az alapkutatáshoz
és a gyógyszerfejlesztéshez egyaránt.
A TRPV1 funkció vizsgálatára ötféle antagonistát használtunk (I./1. ábra), de ezek közül
három távolról sem kifogástalan, a két újabb (ld. lentebb 4. és 5. pont) hatásspektruma pedig
még nem teljesen tisztázott.
1. A sandozi fejlesztéső (most Novartis) kapszazepint (CZP) 1992 óta használják a TRPV1
receptor kutatásában (Bevan és mtsai, 1992; Walpole és mtsai, 1994). A CZP, mely a
kapszaicin szintetikus szerkezeti analógja, egy gyenge kompetitív antagonista (IC50
tartománya kapszaicin ellenében 0.2−5 µM sejttípustól, fajtól függıen).
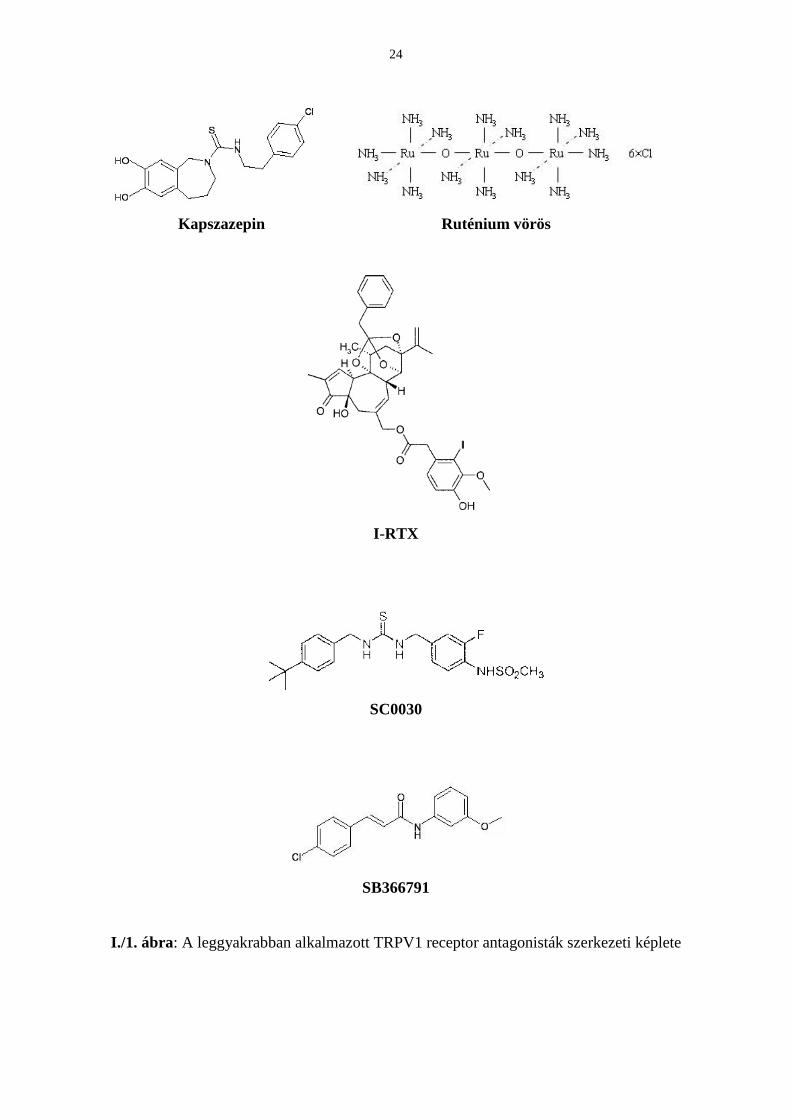
24
Kapszazepin Ruténium vörös
I-RTX
SC0030
SB366791
I./1. ábra: A leggyakrabban alkalmazott TRPV1 receptor antagonisták szerkezeti képlete

25
Hátránya, hogy a hatékony koncentrációtartományban mind a purinerg (Wang és mtsai, 2002)
és a kolinerg receptorokat (Liu & Simon, 1997), mind pedig a feszültségdependens
kalciumcsatornákat gátolja (Docherty és mtsai, 1997). Aktivitása erısen faj- és
modalitásfüggı (Savidge és mtsai, 2001; McIntyre és mtsai, 2001).
2. Az anionos hisztokémiai festékkomplex ruténium vörös egyben potens TRPV1 receptor
antagonista (Dray és mtsai, 1990). Mivel nem szelektív pórusblokkoló, ezért a többi TRPV
receptort, a mitokondriális kalcium-uniportert és egyéb csatornákat is hatékonyan blokkol
(Amann & Maggi, 1991; Gunthorpe és mtsai, 2002).
3. A nem kompetitív iodo-reziniferatoxin (I-RTX) affinitása a TRPV1 receptor iránt az összes
eddig ismert antagonista közül a legnagyobb (Wahl és mtsai, 2001; IC50 = 3.8 nM vs 1 µM
kapszaicin). In vivo szisztémás alkalmazásakor azonban dejodinálódhat és a keletkezı
molekula RTX agonistaként viselkedik (Seabrook és mtsai, 2002).
4. 2002-ben a Szöuli Nemzeti Egyetem kutatói által kifejlesztett SC0030 (JYL1421) kódjellel
jelzett, vanilloid szerkezető kompetítiv antagonista irodalmi adatok alapján 60-szor potensebb
a CZP-nél és a pH, a hı, és a kapszaicin ellen egyaránt hatásos (IC50 = 9 nM versus 50 nM
kapszaicin; Wang és mtsai, 2002; Suh és mtsai, 2003; Jakab és mtsai, 2005).
5. Végül a GlaxoSmithKline által szintetizált kismolekulájú, nem vanilloid szerkezető TRPV1
receptor antagonista, az SB366791 került érdeklıdésünk középpontjába, mely ígéretes
inhibitornak bizonyult TRPV1 receptorral transzfektált sejtvonalban (Gunthorpe és mtsai,
2004; Fowler és mtsai, 2003; Rami és mtsai, 2004).
Értekezésemben ez utóbbi vegyület hatásait mutatnám be a kapszaicinnel kiváltott válaszokra
különféle in vitro és in vivo kísérleti modellekben.
26
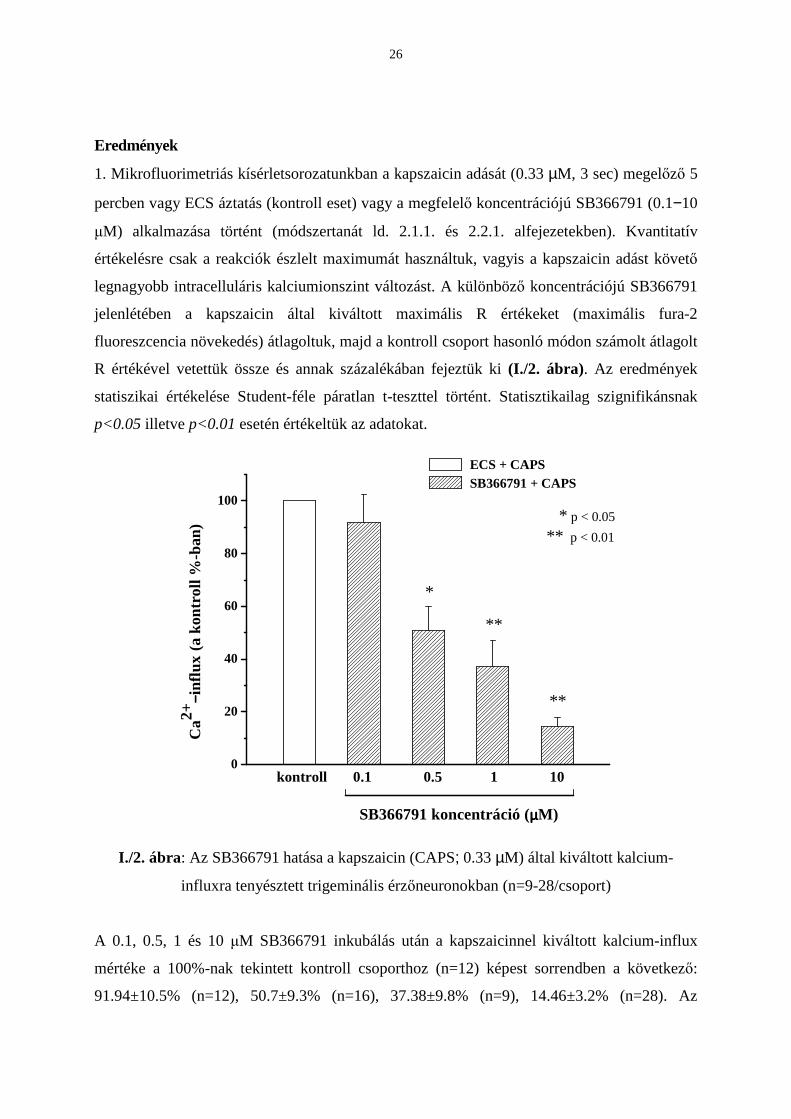
Eredmények
1. Mikrofluorimetriás kísérletsorozatunkban a kapszaicin adását (0.33 µM, 3 sec) megelızı 5
percben vagy ECS áztatás (kontroll eset) vagy a megfelelı koncentrációjú SB366791 (0.1−10
µM) alkalmazása történt (módszertanát ld. 2.1.1. és 2.2.1. alfejezetekben). Kvantitatív
értékelésre csak a reakciók észlelt maximumát használtuk, vagyis a kapszaicin adást követı
legnagyobb intracelluláris kalciumionszint változást. A különbözı koncentrációjú SB366791
jelenlétében a kapszaicin által kiváltott maximális R értékeket (maximális fura-2
fluoreszcencia növekedés) átlagoltuk, majd a kontroll csoport hasonló módon számolt átlagolt
R értékével vetettük össze és annak százalékában fejeztük ki (I./2. ábra). Az eredmények
statiszikai értékelése Student-féle páratlan t-teszttel történt. Statisztikailag szignifikánsnak
p<0.05 illetve p<0.01 esetén értékeltük az adatokat.
0
20
40
60
80
100
**
*
**
ECS + CAPS SB366791 + CAPS
SB366791 koncentráció (µµµµM)
kontroll 0.1 0.5 1 10
Ca2+
−− −−inf
lux
(a k
ontr
oll %
-ban
)
* p < 0.05
** p < 0.01
I./2. ábra: Az SB366791 hatása a kapszaicin (CAPS; 0.33 µM) által kiváltott kalcium-
influxra tenyésztett trigeminális érzıneuronokban (n=9-28/csoport)
A 0.1, 0.5, 1 és 10 µM SB366791 inkubálás után a kapszaicinnel kiváltott kalcium-influx
mértéke a 100%-nak tekintett kontroll csoporthoz (n=12) képest sorrendben a következı:
91.94±10.5% (n=12), 50.7±9.3% (n=16), 37.38±9.8% (n=9), 14.46±3.2% (n=28). Az
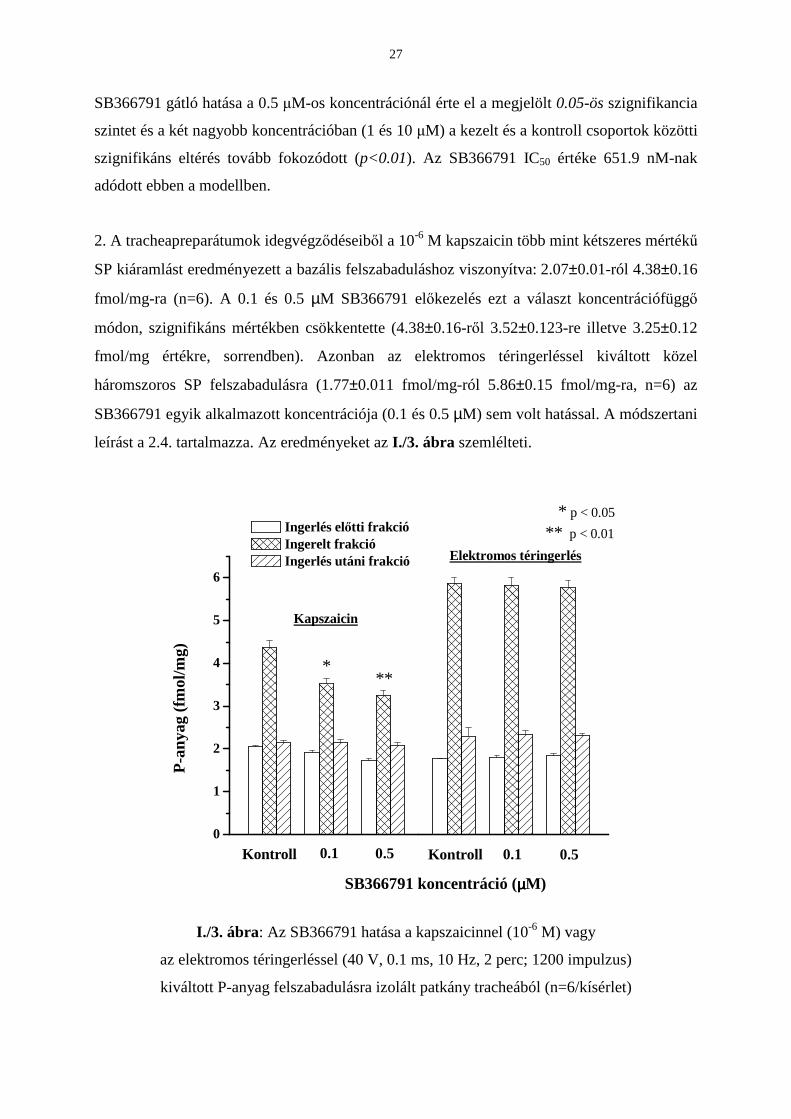
27
SB366791 gátló hatása a 0.5 µM-os koncentrációnál érte el a megjelölt 0.05-ös szignifikancia
szintet és a két nagyobb koncentrációban (1 és 10 µM) a kezelt és a kontroll csoportok közötti
szignifikáns eltérés tovább fokozódott (p<0.01). Az SB366791 IC50 értéke 651.9 nM-nak
adódott ebben a modellben.
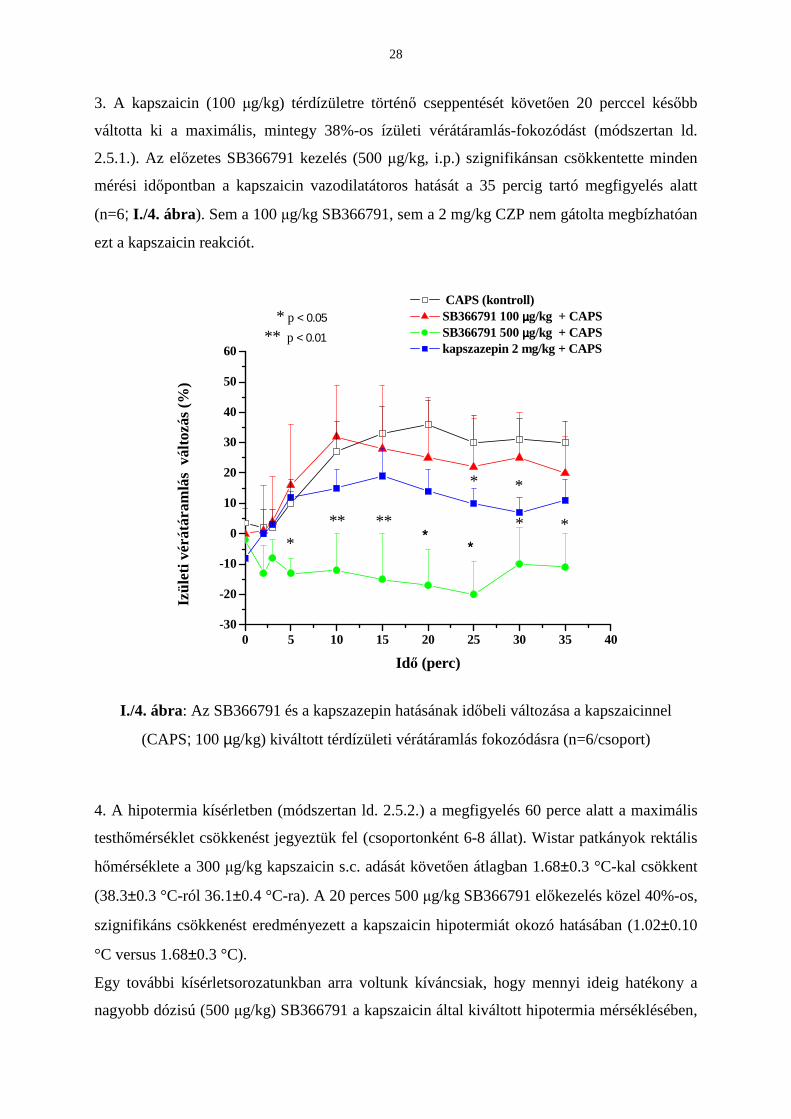
2. A tracheapreparátumok idegvégzıdéseibıl a 10-6 M kapszaicin több mint kétszeres mértékő
SP kiáramlást eredményezett a bazális felszabaduláshoz viszonyítva: 2.07±0.01-ról 4.38±0.16
fmol/mg-ra (n=6). A 0.1 és 0.5 µM SB366791 elıkezelés ezt a választ koncentrációfüggı
módon, szignifikáns mértékben csökkentette (4.38±0.16-rıl 3.52±0.123-re illetve 3.25±0.12
fmol/mg értékre, sorrendben). Azonban az elektromos téringerléssel kiváltott közel
háromszoros SP felszabadulásra (1.77±0.011 fmol/mg-ról 5.86±0.15 fmol/mg-ra, n=6) az
SB366791 egyik alkalmazott koncentrációja (0.1 és 0.5 µM) sem volt hatással. A módszertani
leírást a 2.4. tartalmazza. Az eredményeket az I./3. ábra szemlélteti.
0
1
2
3
4
5
6
Ingerlés elıtti frakció Ingerelt frakció Ingerlés utáni frakció
P-a
nyag
(fm
ol/m
g)
Kontroll 0.1 0.5
SB366791 koncentráció (µµµµM)
Kontroll 0.1 0.5
Kapszaicin
Elektromos téringerlés
* p < 0.05
** p < 0.01
***
I./3. ábra: Az SB366791 hatása a kapszaicinnel (10-6 M) vagy
az elektromos téringerléssel (40 V, 0.1 ms, 10 Hz, 2 perc; 1200 impulzus)
kiváltott P-anyag felszabadulásra izolált patkány tracheából (n=6/kísérlet)
28
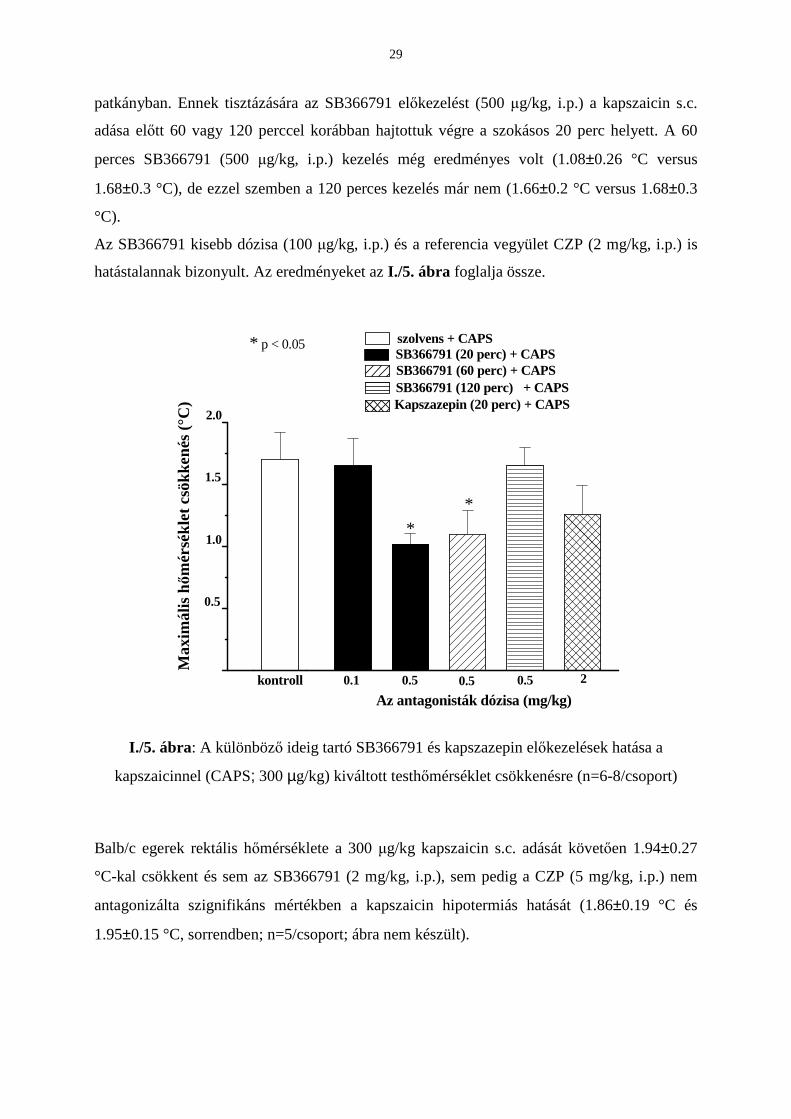
3. A kapszaicin (100 µg/kg) térdízületre történı cseppentését követıen 20 perccel késıbb
váltotta ki a maximális, mintegy 38%-os ízületi vérátáramlás-fokozódást (módszertan ld.
2.5.1.). Az elızetes SB366791 kezelés (500 µg/kg, i.p.) szignifikánsan csökkentette minden
mérési idıpontban a kapszaicin vazodilatátoros hatását a 35 percig tartó megfigyelés alatt
(n=6; I./4. ábra). Sem a 100 µg/kg SB366791, sem a 2 mg/kg CZP nem gátolta megbízhatóan
ezt a kapszaicin reakciót.
0 5 10 15 20 25 30 35 40-30
-20
-10
0
10
20
30
40
50
60
* p < 0.05
** p < 0.01
* *
**
∗∗
**∗∗
**
Izül
eti v
érát
áram
lás
vál
tozá
s (%
)
Idı (perc)
CAPS (kontroll) SB366791 100 µµµµg/kg + CAPS SB366791 500 µµµµg/kg + CAPS kapszazepin 2 mg/kg + CAPS
*
I./4. ábra: Az SB366791 és a kapszazepin hatásának idıbeli változása a kapszaicinnel
(CAPS; 100 µg/kg) kiváltott térdízületi vérátáramlás fokozódásra (n=6/csoport)
4. A hipotermia kísérletben (módszertan ld. 2.5.2.) a megfigyelés 60 perce alatt a maximális
testhımérséklet csökkenést jegyeztük fel (csoportonként 6-8 állat). Wistar patkányok rektális
hımérséklete a 300 µg/kg kapszaicin s.c. adását követıen átlagban 1.68±0.3 °C-kal csökkent
(38.3±0.3 °C-ról 36.1±0.4 °C-ra). A 20 perces 500 µg/kg SB366791 elıkezelés közel 40%-os,
szignifikáns csökkenést eredményezett a kapszaicin hipotermiát okozó hatásában (1.02±0.10
°C versus 1.68±0.3 °C).
Egy további kísérletsorozatunkban arra voltunk kíváncsiak, hogy mennyi ideig hatékony a
nagyobb dózisú (500 µg/kg) SB366791 a kapszaicin által kiváltott hipotermia mérséklésében,
29
patkányban. Ennek tisztázására az SB366791 elıkezelést (500 µg/kg, i.p.) a kapszaicin s.c.
adása elıtt 60 vagy 120 perccel korábban hajtottuk végre a szokásos 20 perc helyett. A 60
perces SB366791 (500 µg/kg, i.p.) kezelés még eredményes volt (1.08±0.26 °C versus
1.68±0.3 °C), de ezzel szemben a 120 perces kezelés már nem (1.66±0.2 °C versus 1.68±0.3
°C).
Az SB366791 kisebb dózisa (100 µg/kg, i.p.) és a referencia vegyület CZP (2 mg/kg, i.p.) is
hatástalannak bizonyult. Az eredményeket az I./5. ábra foglalja össze.
* p < 0.05
2.0
20.50.5
*
1.5
0.5
1.0
0.5
Az antagonisták dózisa (mg/kg)
*
Max
imál
is hım
érsé
klet
csö
kken
és (
°C)
0.1
SB366791 (60 perc) + CAPSSB366791 (120 perc) + CAPSKapszazepin (20 perc) + CAPS
szolvens + CAPSSB366791 (20 perc) + CAPS
kontroll
I./5. ábra: A különbözı ideig tartó SB366791 és kapszazepin elıkezelések hatása a
kapszaicinnel (CAPS; 300 µg/kg) kiváltott testhımérséklet csökkenésre (n=6-8/csoport)
Balb/c egerek rektális hımérséklete a 300 µg/kg kapszaicin s.c. adását követıen 1.94±0.27
°C-kal csökkent és sem az SB366791 (2 mg/kg, i.p.), sem pedig a CZP (5 mg/kg, i.p.) nem
antagonizálta szignifikáns mértékben a kapszaicin hipotermiás hatását (1.86±0.19 °C és
1.95±0.15 °C, sorrendben; n=5/csoport; ábra nem készült).

30
5. A kapszaicin (10 µg/ml, 50 µl) szembe cseppentése 3 percen belül 16.6±2.13 védekezı,
szemtörlı mozdulatot eredményezett (n=8). Sem a 100 µg/kg SB366791 (17.8±1.92), sem
pedig a 2 mg/kg CZP (14.26±1.53) 20 perccel korábban történı alkalmazása nem befolyásolta
szignifikánsan a szemtörlési reflexet (I./6. ábra). Viszont az 500 µg/kg SB366791 20, 60 és
120 perces elıkezelés jelentısen, sorrendben 58.9, 57.7 és 35.5%-kal mérsékelte a
kapszaicinnel kiváltott szemtörlı mozdulatok számát (6.82±0.78, 7.02±0.83, 10.7±2.10; n=6-
8/csoport). A kísérlet módszertanát a 2.5.3. fejezet ismerteti.
0
2
4
6
8
10
12
14
16
18
20
20.5
* p < 0.05
*
SB366791 (60 perc) + CAPSSB366791 (120 perc) + CAPSkapszazepin (20 perc) + CAPS
0.5
*
Sze
mtö
rlı m
ozdu
lato
k sz
áma
Az antagonista dózisa (mg/kg)0.50.1
szolvens + CAPS SB366791 (20 perc) + CAPS
kontroll
I./6. ábra: A különbözı ideig tartó SB366791 és kapszazepin elıkezelések hatása a
kapszaicinnel (CAPS; 10 µg/ml) kiváltott szemtörlési reflexre (n=6-8/csoport)
Következtetések
Az SB366791 koncentrációfüggı módon, szignifikánsan gátolta a kapszaicin által kiváltott
kalciumion belépést TRG sejttenyészetben és a P-anyag felszabadulást izolált patkány
tracheából, habár ebben a koncentrációtartományban nem befolyásolta az elektromos
téringerléssel keltett neuropeptid liberalizációt. Az SB366791 IC50 értéke 651.9 nM-nak
adódott a mikrofluorimetriás in vitro kísérleti modellben.
Elsıként bizonyítottuk, hogy a kapszazepinnel ellentétben a nagyobb dózisban alkalmazott
SB366791 in vivo kivédte a kapszaicinnel kiváltott hipotermiát, a szemtörlési reflexet és a

31
térdízületi vazodilatációt is. Hatása az in vivo eredmények tükrében erısen idıdependens. A
hipotermiás kísérletsorozatban az SB366791 fajfüggı tulajdonságát tapasztaltuk, mivel a
Balb/c egerek rektális hımérsékletét nagy dózisban sem mérsékelte.
Mindezek alapján az SB366791 a kapszazepinnél hatékonyabb TRPV1 receptor
antagonista in vivo és in vitro patkány modellekben egyaránt, így a különbözı fiziológiás
illetve patofiziológiás körülményekben szereplı TRPV1 receptor funkciójának alaposabb
tisztázására alkalmas vegyület.

32
3.2. AZ N-OLEOILDOPAMIN ÉS ANALÓGJAINAK IN VITRO HATÁSELEMZÉSE
Bevezetés
A fájdalmas ingerek integrátoraként funkciónáló TRPV1 kapszaicin receptor egyes aktivátorai
(vanilloid származékok, savas pH, fájdalmas hıinger) jól definiáltak, de endogén ligandját, az
úgynevezett „endovanilloidot” még nem sikerült teljes bizonyossággal azonosítani.
A korábban endogén ligandnak vélt endokannabinoid anandamid (Zygmunt és mtsai, 1999),
az egyes lipoxigenáz termékek, mint a 12-(S)-hidroxi-peroxi-eikozanoidsav (12-HPETE;
Hwang és mtsai, 2000) valamint az N-arachidonildopamin (NADA; Huang és mtsai, 2002;
II./1. ábra) mellett, a legújabb feltételezett jelölt a szarvasmarhaagyból 2003-ban izolált N-
oleoildopamin (OLDA; Chu és mtsai, 2003; II./2. ábra), mely a noradrenalin metabolitjának
oleinsavval képzett savamidja.
A struktúra-aktivitás kutatások alapján az látszik, hogy a vanillilamin csoport és a hosszú
telítetlen zsírsavrész szükséges az optimális TRPV1 interakcióhoz (Szolcsányi & Jancsó-
Gábor, 1975; II./1. és 2. ábrák). Mivel sem az anandamid, sem a lipoxigenáz produktumok
nem rendelkeznek vanillilamin csoporttal, ezért bizonyultak csak gyenge TRPV1 receptor
agonistáknak. Ugyanakkor az anandamid alacsonyabb koncentrációtartományban (<1 µM) az
RTX-indukált szenzoros neuropeptid felszabadulást és a neuropátiás hiperalgéziát egyaránt
gátolta patkányban (Helyes és mtsai, 2003; Németh és mtsai, 2003), így endogén ligand
szerepét jogosan vonták kétségbe (Szolcsányi, 2000). A NADA TRPV1 endogén ligand
funkciója pedig azért kérdıjelezhetı meg, mert legnagyobb mennyiségben a hippocampusban
mutatták ki, ahol bizonyítottan nincs TRPV1 receptor, viszont detektálható kannabinoid
(CB1) receptor expresszió (Huang és mtsai, 2002). Az OLDA azért is érdemel kitüntetett
figyelmet, mert mindamellett, hogy ötvenszer potensebb humán TRPV1 receptoron, mint
patkány CB1-en, I-RTX-szel gátolható termális hiperalgéziát okozott viselkedési tesztben
(Chu és mtsai, 2003). Ezek a megfigyelések megerısítik, hogy az OLDA szelektívebb és
hatékonyabb aktivátora a TRPV1 receptornak, mint az említett vegyületek.
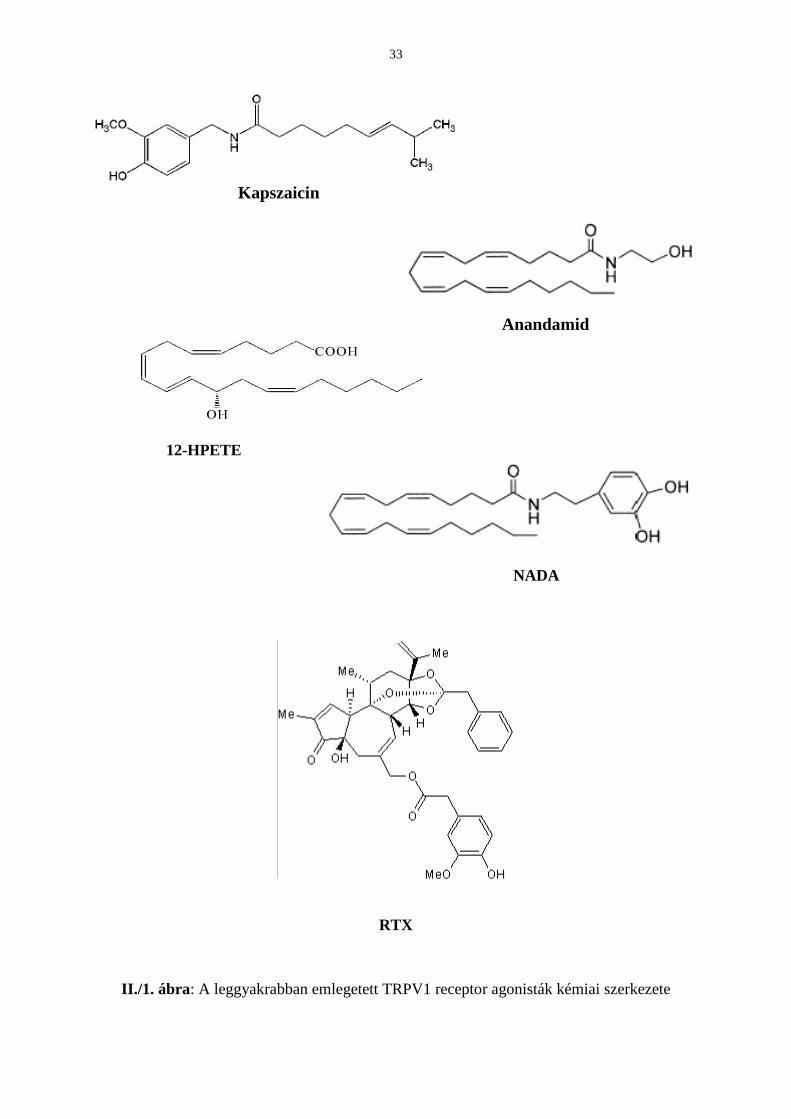
33
Kapszaicin
Anandamid
12-HPETE
NADA
RTX
II./1. ábra: A leggyakrabban emlegetett TRPV1 receptor agonisták kémiai szerkezete

34
OLDA
3-MOLDA
4-MOLDA
II./2. ábra: A szintetizált N-oleoildopamin (OLDA) és szubsztituált analógjainak (3- és 4- MOLDA) kémiai szerkezete
A Magyar Tudományos Akadémia Központi Kémiai Kutató Intézetének közremőködésével
célunk volt a szerkezet-hatás összefüggések alapján elıállított OLDA és származékai
(összefoglaló néven: N-oleoilfeniletilaminok; II./2. ábra) TRPV1 kapszaicin receptorra
gyakorolt hatásainak in vitro és in vivo farmakológiai elemzése. Tekintettel azonban arra,
hogy Dr. Almási Róbert kollégámmal megosztott közleményt birtoklunk, értekezésemben
csak az in vitro kapott eredményeket ismertetném, míg az in vivo adatok Dr. Almási Róbert
Ph.D téziseinek anyagát képezik.
Eredmények
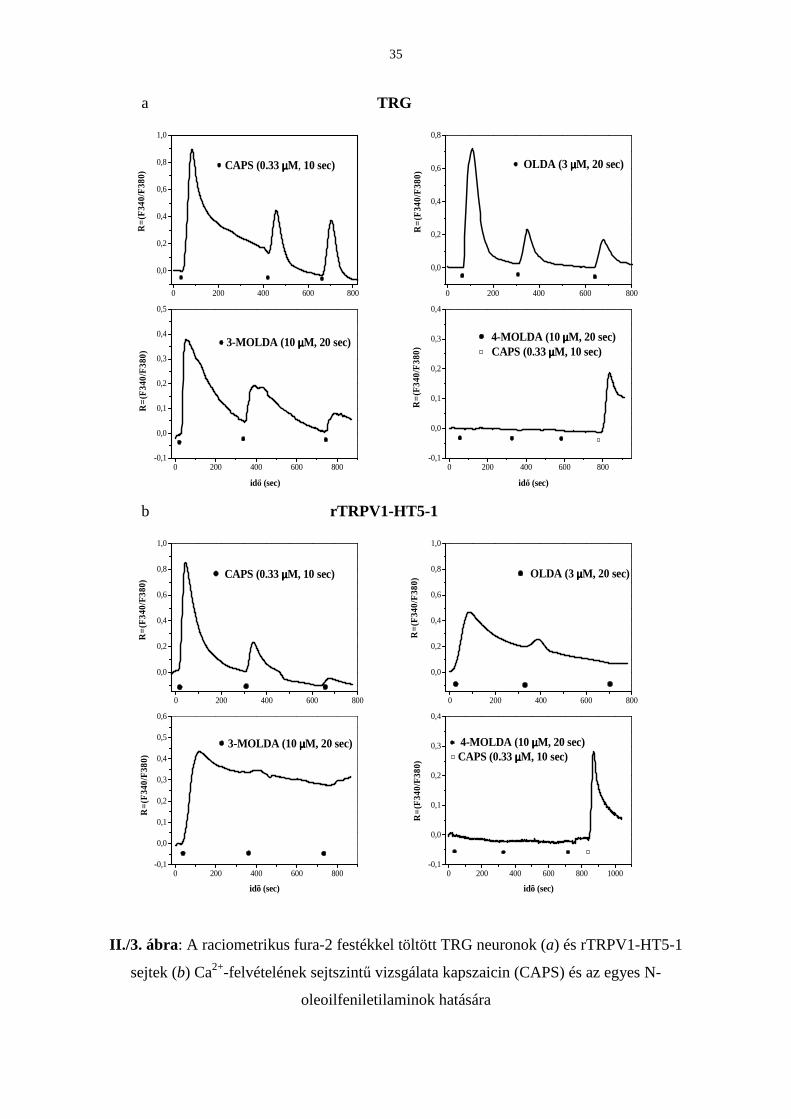
1. A raciometrikus fura-2 festékkel töltött TRG neuronok és rTRPV1-HT5-1 sejtek Ca2+-
felvételének sejtszintő vizsgálatára fluoreszcens, mikrofluorimetriás technikát alkalmaztunk
(módszertan ld. 2.2.1.). Minden egyes méréskor csak egyfajta vegyületet adtuk egymás után
háromszor, 3-5 perces mosási periódusok közbeiktatásával. Kvantitatív kiértékelés helyett
csak a (regisztrátumok kapcsán észlelt) deszenzitizáció mértékét és a reakciók kinetikai
eltéréseit kívántuk demonstrálni.
35
a TRG
0,0
0,2
0,4
0,6
0,8
OLDA (3 µµµµM, 20 sec)
0 200 400 600 800
R=
(F34
0/F
380)
0 200 400 600 800-0,1
0,0
0,1
0,2
0,3
0,4
4-MOLDA (10 µµµµM, 20 sec) CAPS (0.33 µµµµM, 10 sec)
R=
(F34
0/F
380)
idı (sec)
0 200 400 600 800-0,1
0,0
0,1
0,2
0,3
0,4
0,5
R=
(F34
0/F
380)
3-MOLDA (10 µµµµM, 20 sec)
idı (sec)
0 200 400 600 800
0,0
0,2
0,4
0,6
0,8
1,0
CAPS (0.33 µµµµM , 10 sec)
R=
(F34
0/F
380)
b rTRPV1-HT5-1
0 200 400 600 800 1000-0,1
0,0
0,1
0,2
0,3
0,4
idõ (sec)
4-MOLDA (10 µµµµM, 20 sec) CAPS (0.33 µµµµM, 10 sec)
R=
(F34
0/F
380)
0 200 400 600 800-0,1
0,0
0,1
0,2
0,3
0,4
0,5
0,6
3-MOLDA (10 µµµµM, 20 sec)
idõ (sec)
R=(
F34
0/F
380)
0 200 400 600 800
0,0
0,2
0,4
0,6
0,8
1,0
OLDA (3 µµµµM, 20 sec)
R=(
F34
0/F
380)
0 200 400 600 800
0,0
0,2
0,4
0,6
0,8
1,0
R
=(F
340/
F38
0)
CAPS (0.33 µµµµM, 10 sec)
II./3. ábra: A raciometrikus fura-2 festékkel töltött TRG neuronok (a) és rTRPV1-HT5-1
sejtek (b) Ca2+-felvételének sejtszintő vizsgálata kapszaicin (CAPS) és az egyes N-
oleoilfeniletilaminok hatására
36
TRG neuronokban és rTRPV1-expresszáló sejtvonalban a kapszaicin (0.33 µM, 10 sec), az
OLDA (3 µM, 20 sec) és a 3-MOLDA (10 µM, 20 sec) kiváltott kalcium-tranziensek (fura-2
fluoreszcencia növekedés) nagysága ismételt (háromszori), rövididejő applikációkra egyre
csökkent (II./3. ábra).
A 4-MOLDA (10 µM, 20 sec) nem okozott kalcium belépést egyik sejttípusban sem (II./3.
ábra). A sejtek kapszaicin-érzékenységét, azaz a TRPV1 receptor meglétét egy
soronkövetkezı kapszaicin-teszttel (0.33 µM, 10 sec) ezekben a kísérletekben ellenıríztük.
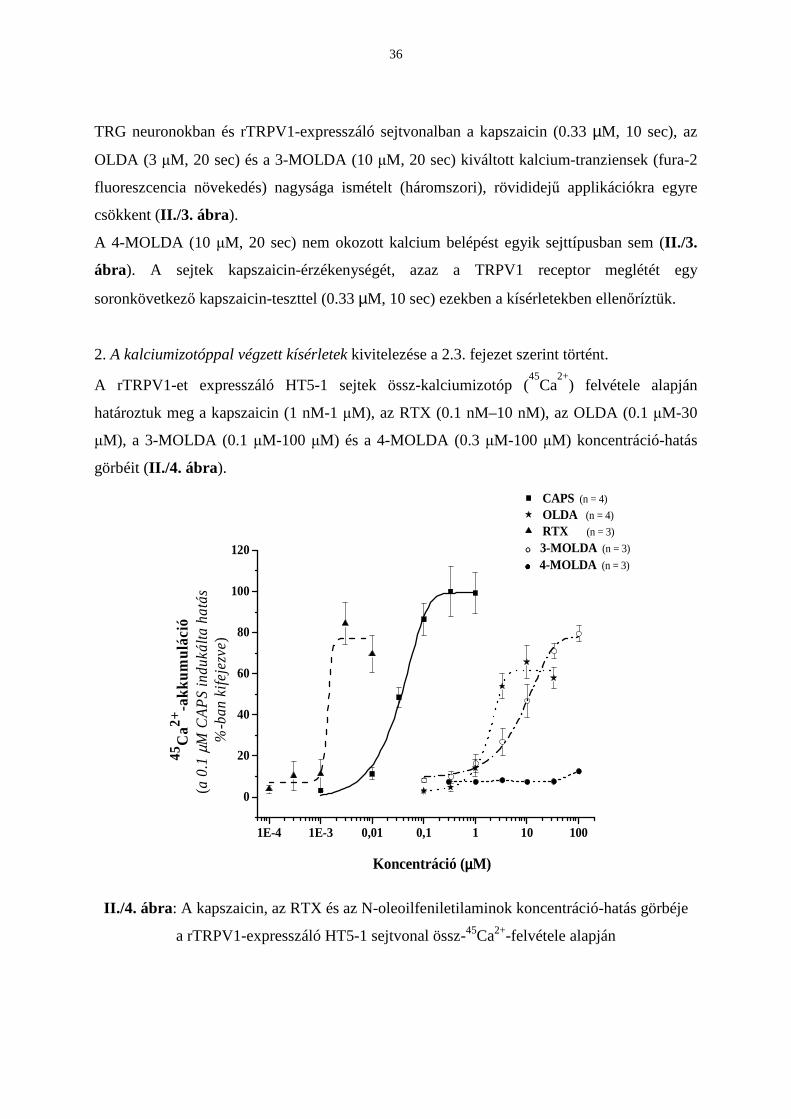
2. A kalciumizotóppal végzett kísérletek kivitelezése a 2.3. fejezet szerint történt.
A rTRPV1-et expresszáló HT5-1 sejtek össz-kalciumizotóp (45
Ca2+
) felvétele alapján
határoztuk meg a kapszaicin (1 nM-1 µM), az RTX (0.1 nM–10 nM), az OLDA (0.1 µM-30
µM), a 3-MOLDA (0.1 µM-100 µM) és a 4-MOLDA (0.3 µM-100 µM) koncentráció-hatás
görbéit (II./4. ábra).
1E-4 1E-3 0,01 0,1 1 10 100
0
20
40
60
80
100
120
Koncentráció (µµµµM)
CAPS (n = 4)
OLDA (n = 4)
RTX (n = 3)
���� 3-MOLDA (n = 3)
���� 4-MOLDA (n = 3)
45C
a2+-a
kkum
ulác
ió
(a 0
.1 µM
CA
PS
ind
uká
lta h
atá
s %
-ba
n k
ifeje
zve)
II./4. ábra: A kapszaicin, az RTX és az N-oleoilfeniletilaminok koncentráció-hatás görbéje
a rTRPV1-expresszáló HT5-1 sejtvonal össz-45Ca2+-felvétele alapján
37
Patkány TRPV1 receptort expresszáló HT5-1 sejteken az RTX, a kapszaicin, az OLDA és a 3-
MOLDA EC50 értékek sorrendben: 1.5 nM, 36 nM, 1.8 µM és 9 µM. Az OLDA ötször
potensebb a 3-MOLDA-nál, de 50-szer kevésbé hatásos, mint a kapszaicin. Az OLDA és a 3-
MOLDA a kapszaicinre adott maximális válasznak csupán 60%-t keltette.
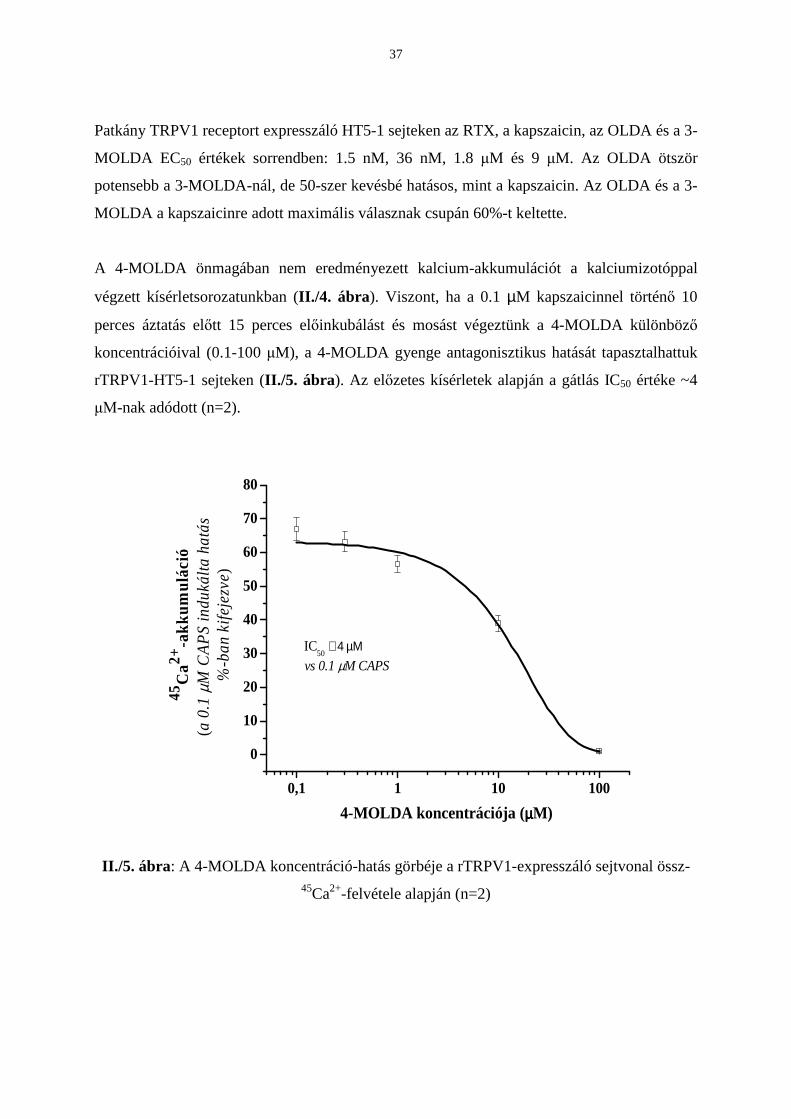
A 4-MOLDA önmagában nem eredményezett kalcium-akkumulációt a kalciumizotóppal
végzett kísérletsorozatunkban (II./4. ábra). Viszont, ha a 0.1 µM kapszaicinnel történı 10
perces áztatás elıtt 15 perces elıinkubálást és mosást végeztünk a 4-MOLDA különbözı
koncentrációival (0.1-100 µM), a 4-MOLDA gyenge antagonisztikus hatását tapasztalhattuk
rTRPV1-HT5-1 sejteken (II./5. ábra). Az elızetes kísérletek alapján a gátlás IC50 értéke ~4
µΜ-nak adódott (n=2).
0,1 1 10 100
0
10
20
30
40
50
60
70
80
45C
a2+-a
kkum
ulác
ió
(a 0
.1 µM
CA
PS
indu
kálta
ha
tás
%-b
an
kife
jezv
e)
vs 0.1 µM CAPS
IC50
∼ 4 µΜ
4-MOLDA koncentrációja (µµµµM)
II./5. ábra: A 4-MOLDA koncentráció-hatás görbéje a rTRPV1-expresszáló sejtvonal össz-
45Ca2+-felvétele alapján (n=2)

38
Következtetések
Patkány TRPV1 receptort expresszáló HT5-1 sejtvonalon valamint TRG neuronokon
raciometrikus, mikrofluorimetriás kalciumionszint méréssel és 45Ca2+-akkumuláción alapuló
szcintillográfiás módszerrel vizsgáltuk a különbözı TRPV1 receptor agonisták és az N-
oleoilfeniletilaminok in vitro hatásait.
A transzfektált sejtekben felvett koncentráció-hatás görbék alapján az RTX, a kapszaicin, az
OLDA és a 3-MOLDA EC50 értékek 1.5 nM, 36 nM, 1.8 µM és 9 µM, sorrendben.
Általánosságban elmondható, hogy az OLDA és a 3-MOLDA reakciók kinetikája lassúbb,
mint a kapszaiciné mindkét sejttípusban. Viszont rTRPV1-HT5-1 sejtekben az általuk
indukált tachyfilaxis (gyengülı válaszok) mértéke kifejezettebb, mint szenzoros neuronok
esetén.
Habár a 3- és a 4-MOLDA szerkezetileg csupán az aromás rész 4. helyzetben lévı hidroxil-
csoportjának meglétében különbözik egymástól, ez az adottságuk szabja meg
farmakológiailag eltérı hatásukat.
A 4-MOLDA egyik tesztünkben sem okozott kalciumion belépést. Azonban a kapszaicin
kalciumakkumuláló hatásának csökkenését tapasztaltuk a 4-MOLDA elıinkubálás hatására,
mely a vanilloid jellegő vegyületek 4-es pozícióban lévı aromás hidroxilcsoportjának
jelentıségét hangsúlyozza a receptorfunkcióban.
Az N-oleoilfeniletilaminok fájdalomkeltı hatását patkányok nociceptív hıküszöbe, valamint
vad típusú és TRPV1 génhiányos egerek nocifenzív viselkedése alapján Dr. Almási Róbert
tesztelte. Az in vivo eredmények jól korrelálnak az általam ismertetett in vitro adatokkal,
melyekkel a TRPV1 legígéretesebb endogén ligandjának szerepét bizonyítottuk.

39
3.3. A PROTEIN KINÁZ A ÉS C RELATÍV SZEREPE A TRPV1 RECEPTOR
ÉRZÉKENYSÉGÉNEK MODULÁLÁSÁBAN
Bevezetés
A polimodális nociceptorok a TRPV1 kapszaicin receptorral többféle környezeti inger
detektálására képesek. A fehérjék, köztük a ligand-vezérelt ioncsatornák,
foszforilált/defoszforilált állapota mérhetıen befolyásolja azok biokémiai-fiziológiai
funkcióit. Fiziológiásan dinamikus egyensúlyt tételeznek fel a TRPV1 receptor protein
kinázok (pl. PKA és PKC) általi foszforilálása (aktiválás) és a kalciumdependens
foszfatázokkal (pl. protein foszfatáz 2B azaz kalcineurin) keltett defoszforilálása (deaktiválás)
viszonyában. Kínálkozik a feltevés, hogy a foszforiláltság mértéke a
szenzitizálás/deszenzitizálás mérlegét döntıen befolyásolja. Rövid (óra-perc) dimenzióban ez
döntheti el a farmakológiai receptor valamint a nociceptor funkcionális reaktivitását, de más
tényezık szerepe sem elhanyagolható (például az expresszált TRPV1 receptorok száma vagy
azok kinetikai, ionkonduktancia tulajdonságai).
A TRPV1 receptor integrátor molekula szerepe gyulladásos hiperalgéziában jól ismert
(Caterina és mtsai, 2000; Davis és mtsai, 2000; Szolcsányi, 2002). Patológiás körülmények
között sokféle endogén algogén szintetizálódik és/vagy szabadul fel a környezı kötıszöveti
sejtekbıl valamint a neuronális sejtek végzıdéseibıl (Aley & Levine, 1999, Aley és mtsai,
2000), melyek többek között PKA-t (pl. PGE2) vagy PKC-t (pl. bradikinin, hisztamin, ATP) is
aktiválhatnak. A TRPV1 receptormolekula hét cAMP-függı protein kináz A (PKA;
Mohapatra & Nau, 2003), hat protein kináz C (PKC; Bhave és mtsai, 2003) és hat kalcium-
kalmodulindependens kináz II (CaMKII; Jung és mtsai, 2004) foszforilálható szekvenciával
rendelkezik. Mutáns receptorok elektrofiziológiai analízisével megállapították, hogy PKA-
dependens foszforilációs hely a 116-os szerin (Bhave és mtsai, 2002), kizárólagos PKC
foszforilációs hely a S800-as (Numazaki és mtsai, 2002; ld. Általános Bevezetés 2. ábra). A
S502-es aminosav többféle kináz támadáspontja (2. ábra).
Ellentmondásos az irodalom abban a tekintetben, hogy melyik PKC izoforma felelıs a
TRPV1 receptor szenzitizációjáért. Premkumar és Ahern (2000) valamint Numazaki és
munkatársai (2002) szerint már szobahımérsékleten is lehetséges a TRPV1 receptor
aktiválása akár elızetes agonista adás nélkül is a kalcium-inszenzitív PKCε közvetítésével.

40
Mások a konvencionális, Ca-szenzitív PKCα szerepét hangsúlyozzák a gyulladásos és az
ischémiás fájdalmak közvetítésében szenzoros neuronban (Oláh és mtsai, 2002). A
kapszaicinnel kiváltott deszenzitizáció forbol 12-mirisztát 13-acetát (PMA) általi kivédése a
S502-es és a S800-as alanin-szubsztituált variánsaiban nem figyelhetı meg (Mandadi és
mtsai, 2004). A cAMP-függı PKA aktiválás nemcsak fokozza a receptor
vanilloidérzékenységét, hanem az ismételt adásokra kialakuló kalciumdependens
deszenzitizációt (tachyfilaxis) is regulálja a S116 és a T370 csoportok direkt foszforilációjával
(Mohapatra & Nau, 2003, 2005).
Szenzoros neuronban a mGluR1 és a mGluR5 glutamát (Hu és mtsai, 2002), a P2X3 purinerg
(Tominaga és mtsai, 2001) és a TRPV1 kapszaicin receptorok is aktiválódásuk esetén kvázi
tízszer több kalciumot engednek be, mint nátriumot. A kalcium, mint intracelluláris
másodlagos hírvívımolekula kinázok, nitrogénoxid szintáz, foszfatázok enziminduktora
illetve kulcsregulátora, melyek aktiválódása újabb, a neuronfunkcióban lényeges
folyamatokat mőködtetı mechanizmusokat kapcsolnak be vagy éppen ki. Újabban a parakrin
módon szabályozó nonapeptid bradikinin (BK), az ATP, az autokrin hatású NGF, a protein
kináz C-t stimuláló PMA, valamint a proteáz-aktivált receptort (PAR2) hasító tripszin
nociceptív hatásairól is bebizonyosodott, hogy a TRPV1 receptort indirekt módon, foszfolipáz
C- (PLC) illetve protein kináz-dependens mechanizmusokkal szenzitizálják természetes
agonistái iránt (Vellani és mtsai, 2001; Numazaki és mtsai, 2002; Tominaga és mtsai, 2001;
Cesare & McNaughton, 1996; Sugiura és mtsai, 2002; Shu & Mendell, 2001; Mandadi és
mtsai, 2004; Amadesi és mtsai, 2004). A proalgogén BK a Gq/11 fehérje-kapcsolt
heptahelikális BK2 receptorán keresztül indukálja a PLCβ jelátviteli útvonalat, mely elsı
lépésben a membrán PIP2 hidrolízisét katalizálja diacil-glicerollá (DAG) és inozitol-1,4,5-
triszfoszfáttá (IP3). A membránkötött DAG molekulák közvetlen PKCε aktiválásra (Cesare és
mtsai, 1999), a szolubilis IP3 pedig kalcium mobilizására képes az IP3-szenzitív intracelluláris
raktárakból. Mindkét mechanizmus fokozza az agonista-indukált válaszokat TRPV1 receptort
expresszáló sejtekben (Premkumar & Ahern, 2000). Az NGF hatásának kivitelezésében − a
magas affinitású TrkA receptorának dimerizációját és (transz)autofoszforilációját követıen −
számos enzim és csatornafehérje, mint például a foszfatidil-inozitol-3 kináz (PI3K), a
CAMKII, a PLCγ, egyes PKC izoenzimek és a TRPV1 receptor egyaránt érintett (Bonnington
& McNaughton, 2003). Az NGF centrális szenzitizációt eredményezı tartós hatása a TRPV1,
a GABA és a BK2 receptorok mRNS-ének poszttranszlációs módosítása révén érvényesül p38

41
mitogén-aktivált (szerin/treonin illetve tirozin) protein kinázok kaszkádján keresztül (Snider
& McMahon, 1998). Egyes prosztaglandinok (PGE2), az anandamid, a forskolin és a glutamát
is cAMP-dependens PKA aktiválással befolyásolja a TRPV1 receptor funkcióját (Lopshire &
Nicol, 1998; Gu és mtsai, 2003; De Petrocellis és mtsai, 2001; Rathee és mtsai, 2002; Hu és
mtsai, 2002). Említenek BK/ATP/NGF/PGE2-indukált, a TRPV1 receptorra konvergálódó
„fájdalom útvonalakat”, melyek többnyire foszfolipáz C- és protein kináz-kapcsolt
mechanizmusok bevonásával valósítják meg a TRPV1 termális érzékenységének fentebb
vázolt, összetett finomszabályozását (Tominaga és mtsai, 2001; Chuang és mtsai, 2001;
Vellani és mtsai, 2001).
A fent említett tanulmányok többségében in vitro megfigyeléseket végeztek TRPV1-
transzfektált nem-neuronális sejtekben vagy szenzoros neuronok szómáján, figyelmen kívül
hagyva a perifériás nociceptorok in vivo jelentıségét. Ugyanakkor kevés publikációban
hasonlították össze szisztematikusan a PKC illetıleg a PKA hatásait a TRPV1 receptor
érzékenységére ugyanabban a modellben (Hu és mtsai, 2002; Mohapatra & Nau, 2003),
valamint az alapaktivitású (azaz a nem stimulált) PKC és PKA szerepét együttesen is ritkán
elemezték.
Ráadásul az érintett témakörben számos ellentmondásos adat áll rendelkezésünkre. Például az
NGF által kiváltott TRPV1 receptor facilitációt patkány hátsógyöki neuronban (DRG) PKA
(Shu & Mendell, 2001), míg egér DRG-ben PKC modulálja (Bonnington & McNaughton,
2003). A PKA aktivitásának serkentése Vulcu és mtsai szerint (2003) hatékonyan
szenzitizálta a Xenopus petesejtbe transzfektált TRPV1 receptort természetes agonistái iránt,
ugyanakkor Lee és kollégái (2000) ezzel ellentétes eredményt közöltek.
Célunk a stimulált illetve az alapaktivitású PKC és a PKA relatív jelentıségének
összehasonlítása a TRPV1 receptor funkcióra in vitro és in vivo patkány modellekben.
Vizsgáltuk az egyes protein kináz modulátorok (inhibitorok és aktivátorok) hatását a
kapszaicin, a reziniferatoxin (RTX) és a KCl által keltett kalcium-tranziensekre újszülött
patkány trigeminális szenzoros neurontenyészeten (TRG sejttest), valamint az RTX-szel
kiváltott fájdalmas hıküszöbcsökkenésre a perifériás idegvégzıdések szintjén is (DRG
végzıdés).

42
Eredmények
Patkány trigeminális ganglionsejt tenyészet készítését és az intracelluláris kalciumionszint-
mérést a 2.1.1. és a 2.2.1. alfejezeteknél ismertetett módon végeztük.
Kvantitatív értékelésre csak az R>0.1 (fura-2 fluoreszcencia növekedés > 0.1) reakciók
maximumait használtuk, mely a kapszaicin, az RTX és a KCl applikáció elsı 3
másodpercében mindig kialakult.
Kísérletünk kivitelezése úgy zajlott, hogy a két rövid ideig alkalmazott kapszaicin (0.33 µM,
3 sec), RTX (1 nM, 3 sec) vagy KCl (50 mM, 3 sec) adás között vagy ECS mosás (5 perc,
kontroll eset) vagy a megfelelı protein kináz inhibitor (5 perc) vagy aktivátor (10-30 sec)
alkalmazása történt. Az egyes sejtek normalizált fluoreszcencia arányát úgy számoltuk, hogy
a második kapszaicin/RTX/KCl teszt R értékét (maximális kalciumion akkumulációjának
mértékét) osztottuk az elsı teszt R értékével, és a sejtenkénti hányadosokat felszoroztuk
százzal. A kapszaicin/RTX/KCl ismétlések által, az enzimmodulátorok hiányában
megvalósított tachyfilaxis (deszenzitizáció) statisztikai kiértékelésére Student-féle páros t-
próbát alkalmaztunk. A különbözı enzimmodulátorok jelenlétében kapott normalizált
válaszokat egyszempontú Anova-féle Tukey-teszttel vetettük össze az össz-kontroll R2/R1
értékekkel (az 5 percben ECS mosás).
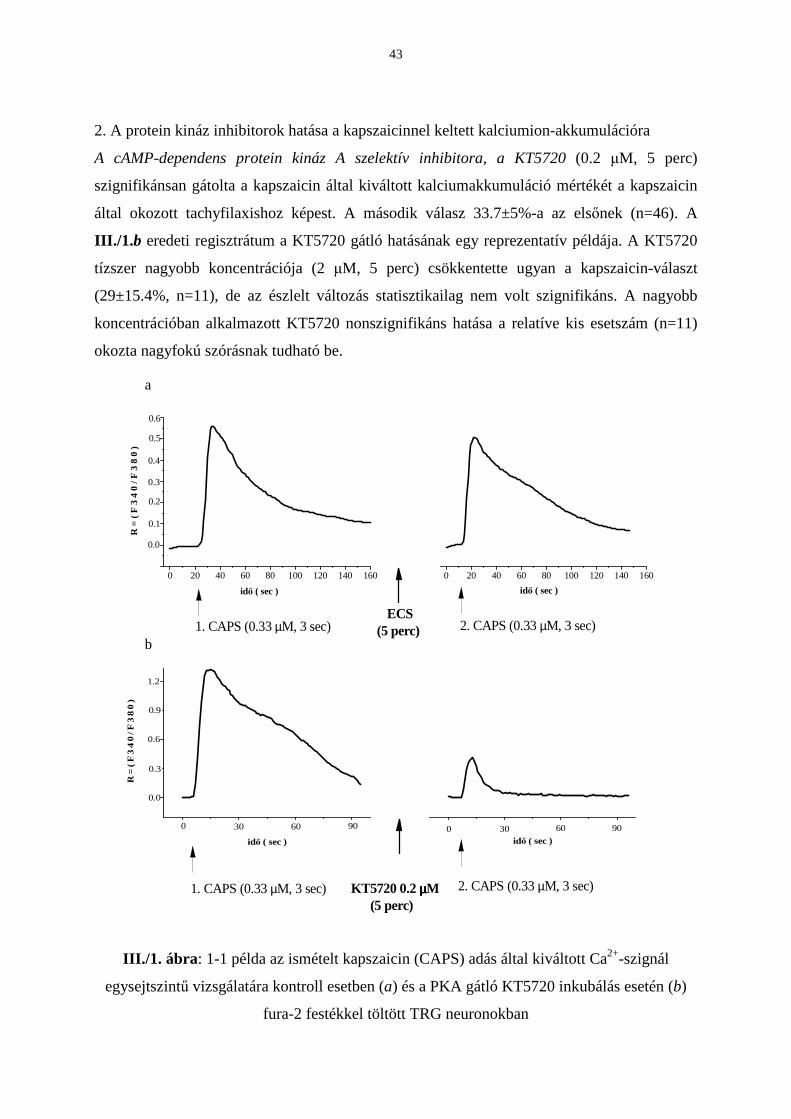
1. Elöljáróban a kapszaicinnel keltett kalcium-szignál néhány jellemzı tulajdonságát
ismertetném tenyésztett TRG neuronokban.
A kapszaicinnel (0.33 µM, 3 sec) kiváltott kalcium-tranziens idıtartama 20 másodperctıl
egészen 3 percig terjedt. A vizsgált kismérető szenzoros neuronok (n=392) 77.5%-a bizonyult
kapszaicin-érzékenynek. A kontroll csoport 186 neuronjában az elsı kapszaicin-teszt
maximális fluoreszcencia növekedése (R) 1.37±0.08. Az 5 perccel késıbb alkalmazott
második kapszaicin által kiváltott kalcium-válaszban relatíve kismértékő tachyfilaxist
tapasztaltunk (R=1.08±0.07). A III./1. a ábra egy ilyen kontroll reakciópárt szemléltet. A
második válasz 92.6±4.1%-a az elsınek. Ez az érték a hatásszámítások összehasonlítási alapja
a kapszaicinnel keltett válaszok esetén (III./2. és 6. ábrák).
43
2. A protein kináz inhibitorok hatása a kapszaicinnel keltett kalciumion-akkumulációra
A cAMP-dependens protein kináz A szelektív inhibitora, a KT5720 (0.2 µM, 5 perc)
szignifikánsan gátolta a kapszaicin által kiváltott kalciumakkumuláció mértékét a kapszaicin
által okozott tachyfilaxishoz képest. A második válasz 33.7±5%-a az elsınek (n=46). A
III./1. b eredeti regisztrátum a KT5720 gátló hatásának egy reprezentatív példája. A KT5720
tízszer nagyobb koncentrációja (2 µM, 5 perc) csökkentette ugyan a kapszaicin-választ
(29±15.4%, n=11), de az észlelt változás statisztikailag nem volt szignifikáns. A nagyobb
koncentrációban alkalmazott KT5720 nonszignifikáns hatása a relatíve kis esetszám (n=11)
okozta nagyfokú szórásnak tudható be.
ECS(5 perc)
b
0 20 40 60 80 100 120 140 160
idı ( sec )
0 20 40 60 80 100 120 140 160
0.6
0.5
R =
( F
3 4
0 /
F 3
8 0
)
idı ( sec )
0.4
0.3
0.2
0.1
0.0
idı ( sec )0 30 9060
1.2
0.9
0.6
R =
( F
3 4
0 /
F 3
8 0
)
idı ( sec )
0 30 60 90
0.0
0.3
1. CAPS (0.33 µM, 3 sec) 2. CAPS (0.33 µM, 3 sec)
KT5720 0.2 µµµµM(5 perc)
1. CAPS (0.33 µM, 3 sec) 2. CAPS (0.33 µM, 3 sec)
a
III./1. ábra : 1-1 példa az ismételt kapszaicin (CAPS) adás által kiváltott Ca2+-szignál
egysejtszintő vizsgálatára kontroll esetben (a) és a PKA gátló KT5720 inkubálás esetén (b)
fura-2 festékkel töltött TRG neuronokban
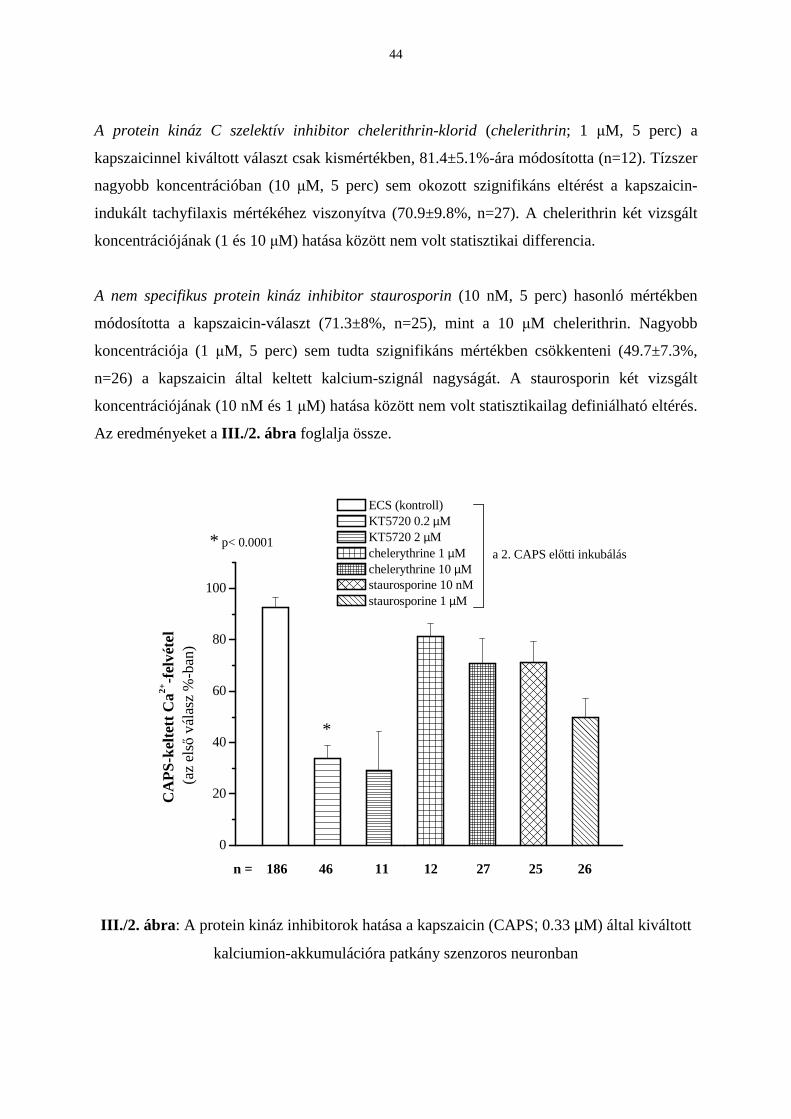
44
A protein kináz C szelektív inhibitor chelerithrin-klorid (chelerithrin; 1 µM, 5 perc) a
kapszaicinnel kiváltott választ csak kismértékben, 81.4±5.1%-ára módosította (n=12). Tízszer
nagyobb koncentrációban (10 µM, 5 perc) sem okozott szignifikáns eltérést a kapszaicin-
indukált tachyfilaxis mértékéhez viszonyítva (70.9±9.8%, n=27). A chelerithrin két vizsgált
koncentrációjának (1 és 10 µM) hatása között nem volt statisztikai differencia.
A nem specifikus protein kináz inhibitor staurosporin (10 nM, 5 perc) hasonló mértékben
módosította a kapszaicin-választ (71.3±8%, n=25), mint a 10 µM chelerithrin. Nagyobb
koncentrációja (1 µM, 5 perc) sem tudta szignifikáns mértékben csökkenteni (49.7±7.3%,
n=26) a kapszaicin által keltett kalcium-szignál nagyságát. A staurosporin két vizsgált
koncentrációjának (10 nM és 1 µM) hatása között nem volt statisztikailag definiálható eltérés.
Az eredményeket a III./2. ábra foglalja össze.
0
20
40
60
80
100
*
C
AP
S-k
elte
tt C
a2+-f
elvé
tel
(az
elsı
vál
asz
%-b
an)
* p< 0.0001
n = 186 46 11 12 27 25 26
ECS (kontroll) KT5720 0.2 µM KT5720 2 µM chelerythrine 1 µM chelerythrine 10 µM staurosporine 10 nM staurosporine 1 µM
a 2. CAPS elıtti inkubálás
III./2. ábra : A protein kináz inhibitorok hatása a kapszaicin (CAPS; 0.33 µM) által kiváltott
kalciumion-akkumulációra patkány szenzoros neuronban
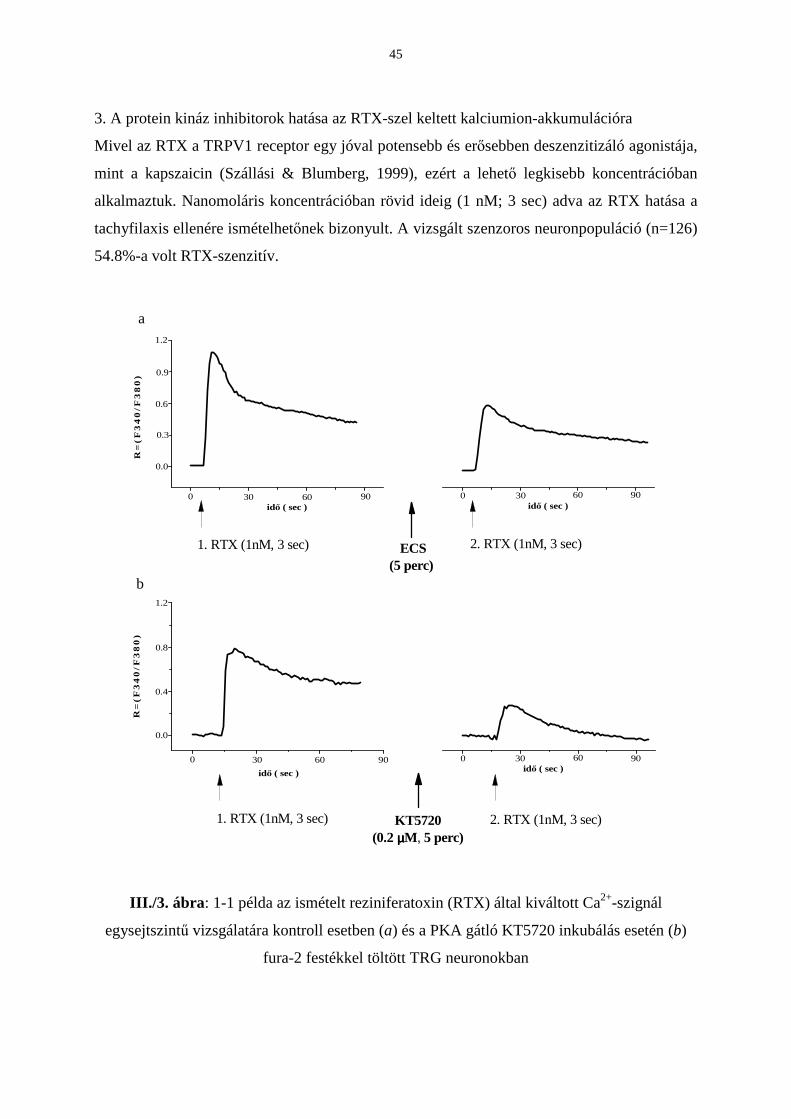
45
3. A protein kináz inhibitorok hatása az RTX-szel keltett kalciumion-akkumulációra
Mivel az RTX a TRPV1 receptor egy jóval potensebb és erısebben deszenzitizáló agonistája,
mint a kapszaicin (Szállási & Blumberg, 1999), ezért a lehetı legkisebb koncentrációban
alkalmaztuk. Nanomoláris koncentrációban rövid ideig (1 nM; 3 sec) adva az RTX hatása a
tachyfilaxis ellenére ismételhetınek bizonyult. A vizsgált szenzoros neuronpopuláció (n=126)
54.8%-a volt RTX-szenzitív.
0 30 60 90idı ( sec )
0 30 60 90
0.0
0.4
0.8
1.2
R =
( F
3 4
0 /
F 3
8 0
)
idı ( sec )
0 30 9060idı ( sec )
1.2
0 30 60 90
R =
( F
3 4
0 /
F 3
8 0
)
idı ( sec )
0.9
0.6
0.3
0.0
ECS(5 perc)
1. RTX (1nM, 3 sec) 2. RTX (1nM, 3 sec)
KT5720 (0.2 µµµµM, 5 perc)
1. RTX (1nM, 3 sec) 2. RTX (1nM, 3 sec)
a
b
III./3. ábra : 1-1 példa az ismételt reziniferatoxin (RTX) által kiváltott Ca2+-szignál
egysejtszintő vizsgálatára kontroll esetben (a) és a PKA gátló KT5720 inkubálás esetén (b)
fura-2 festékkel töltött TRG neuronokban
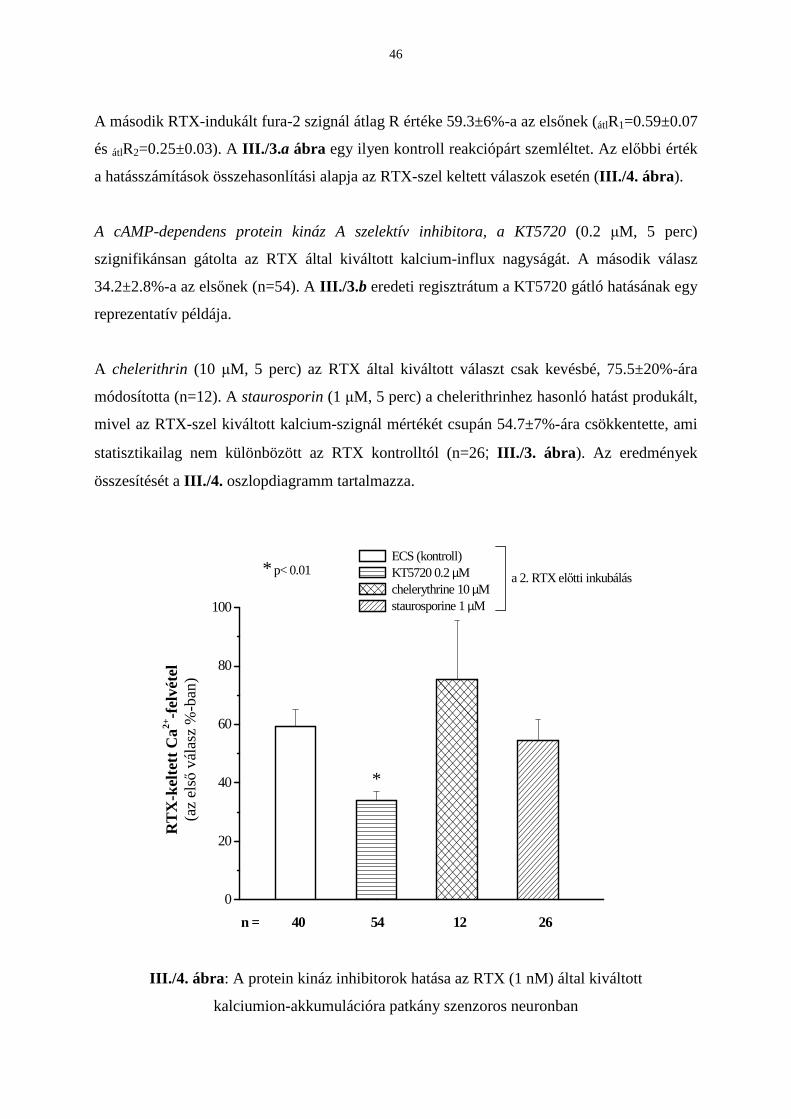
46
A második RTX-indukált fura-2 szignál átlag R értéke 59.3±6%-a az elsınek (átlR1=0.59±0.07
és átlR2=0.25±0.03). A III./3. a ábra egy ilyen kontroll reakciópárt szemléltet. Az elıbbi érték
a hatásszámítások összehasonlítási alapja az RTX-szel keltett válaszok esetén (III./4. ábra ).
A cAMP-dependens protein kináz A szelektív inhibitora, a KT5720 (0.2 µM, 5 perc)
szignifikánsan gátolta az RTX által kiváltott kalcium-influx nagyságát. A második válasz
34.2±2.8%-a az elsınek (n=54). A III./3. b eredeti regisztrátum a KT5720 gátló hatásának egy
reprezentatív példája.
A chelerithrin (10 µM, 5 perc) az RTX által kiváltott választ csak kevésbé, 75.5±20%-ára
módosította (n=12). A staurosporin (1 µM, 5 perc) a chelerithrinhez hasonló hatást produkált,
mivel az RTX-szel kiváltott kalcium-szignál mértékét csupán 54.7±7%-ára csökkentette, ami
statisztikailag nem különbözött az RTX kontrolltól (n=26; III./3. ábra ). Az eredmények
összesítését a III./4. oszlopdiagramm tartalmazza.
0
20
40
60
80
100
RT
X-k
elte
tt C
a2+
-fel
véte
l(a
z el
sı v
álas
z %
-ban
)
n = 40 54 12 26
ECS (kontroll) KT5720 0.2 µM chelerythrine 10 µM staurosporine 1 µM
*
* p< 0.01 a 2. RTX elıtti inkubálás
III./4. ábra : A protein kináz inhibitorok hatása az RTX (1 nM) által kiváltott
kalciumion-akkumulációra patkány szenzoros neuronban
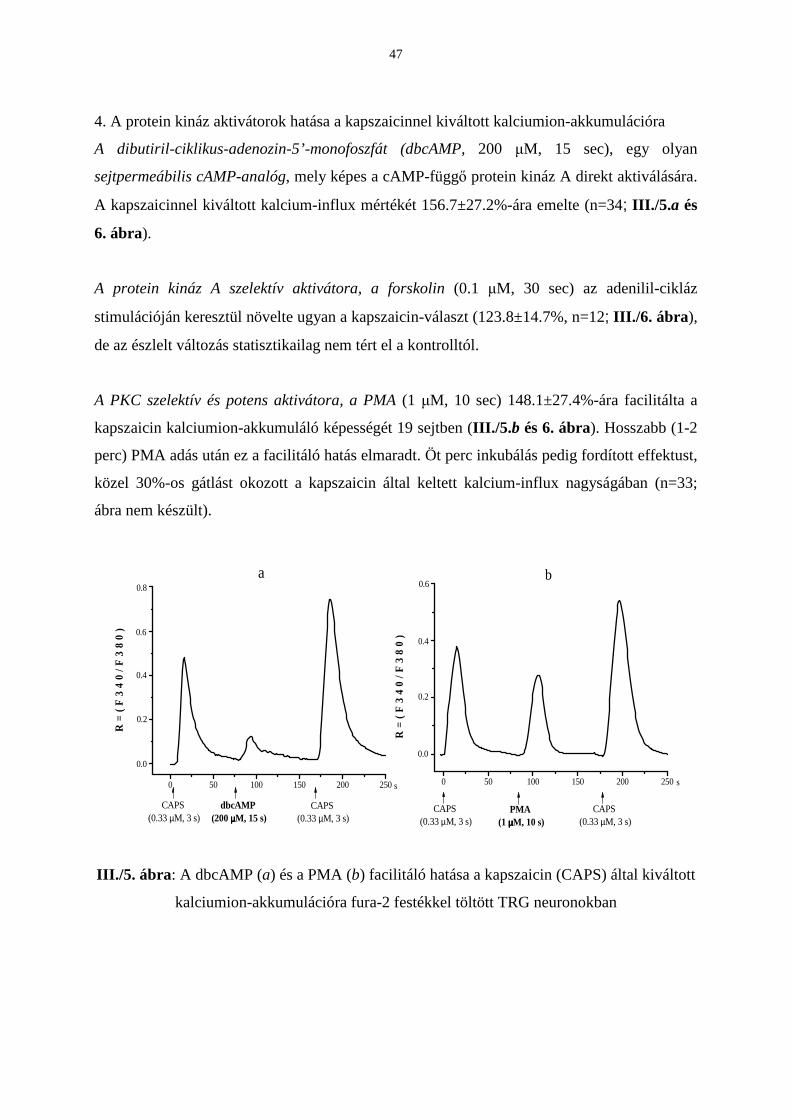
47
4. A protein kináz aktivátorok hatása a kapszaicinnel kiváltott kalciumion-akkumulációra
A dibutiril-ciklikus-adenozin-5’-monofoszfát (dbcAMP, 200 µM, 15 sec), egy olyan
sejtpermeábilis cAMP-analóg, mely képes a cAMP-függı protein kináz A direkt aktiválására.
A kapszaicinnel kiváltott kalcium-influx mértékét 156.7±27.2%-ára emelte (n=34; III./5. a és
6. ábra).
A protein kináz A szelektív aktivátora, a forskolin (0.1 µM, 30 sec) az adenilil-cikláz
stimulációján keresztül növelte ugyan a kapszaicin-választ (123.8±14.7%, n=12; III./6. ábra ),
de az észlelt változás statisztikailag nem tért el a kontrolltól.
A PKC szelektív és potens aktivátora, a PMA (1 µM, 10 sec) 148.1±27.4%-ára facilitálta a
kapszaicin kalciumion-akkumuláló képességét 19 sejtben (III./5. b és 6. ábra). Hosszabb (1-2
perc) PMA adás után ez a facilitáló hatás elmaradt. Öt perc inkubálás pedig fordított effektust,
közel 30%-os gátlást okozott a kapszaicin által keltett kalcium-influx nagyságában (n=33;
ábra nem készült).
a
0 50 100 150 200 250s
dbcAMP (200 µµµµM, 15 s)
R =
( F
3 4
0 /
F 3
8 0
)
0.0
0.2
0.4
0.6
0.8
CAPS
(0.33 µM, 3 s)CAPS
(0.33 µM, 3 s)
b
0 50 100 150 200 250
PMA (1 µµµµM, 10 s)
R =
( F
3 4
0 /
F 3
8 0
)
0.0
0.2
0.4
0.6
CAPS (0.33 µM, 3 s)
CAPS (0.33 µM, 3 s)
s
III./5. ábra : A dbcAMP (a) és a PMA (b) facilitáló hatása a kapszaicin (CAPS) által kiváltott
kalciumion-akkumulációra fura-2 festékkel töltött TRG neuronokban
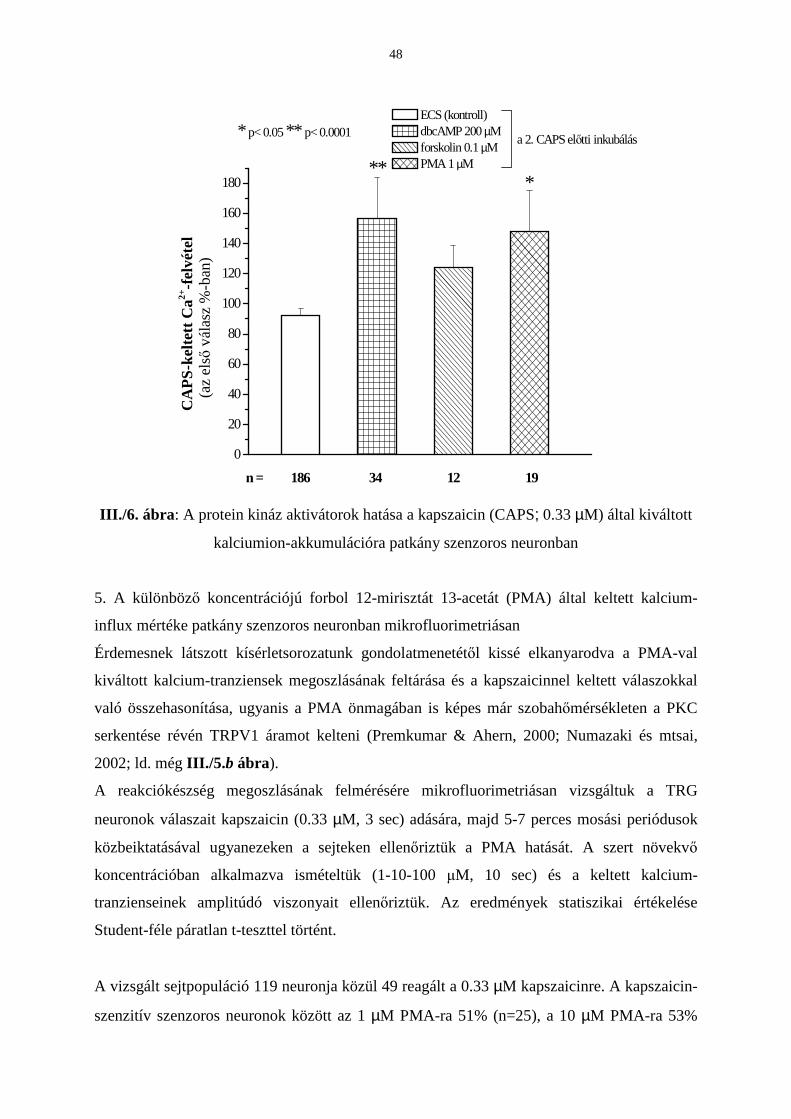
48
0
20
40
60
80
100
120
140
160
180 * **
* p< 0.05 ** p< 0.0001
C
AP
S-k
elte
tt C
a2+-f
elvé
tel
(az
elsı
vála
sz %
-ban
)
a 2. CAPS elıtti inkubálás
n = 186 34 12 19
ECS (kontroll) dbcAMP 200 µM forskolin 0.1 µM PMA 1 µM
III./6. ábra : A protein kináz aktivátorok hatása a kapszaicin (CAPS; 0.33 µM) által kiváltott
kalciumion-akkumulációra patkány szenzoros neuronban
5. A különbözı koncentrációjú forbol 12-mirisztát 13-acetát (PMA) által keltett kalcium-
influx mértéke patkány szenzoros neuronban mikrofluorimetriásan
Érdemesnek látszott kísérletsorozatunk gondolatmenetétıl kissé elkanyarodva a PMA-val
kiváltott kalcium-tranziensek megoszlásának feltárása és a kapszaicinnel keltett válaszokkal
való összehasonítása, ugyanis a PMA önmagában is képes már szobahımérsékleten a PKC
serkentése révén TRPV1 áramot kelteni (Premkumar & Ahern, 2000; Numazaki és mtsai,
2002; ld. még III./5. b ábra).
A reakciókészség megoszlásának felmérésére mikrofluorimetriásan vizsgáltuk a TRG
neuronok válaszait kapszaicin (0.33 µM, 3 sec) adására, majd 5-7 perces mosási periódusok
közbeiktatásával ugyanezeken a sejteken ellenıriztük a PMA hatását. A szert növekvı
koncentrációban alkalmazva ismételtük (1-10-100 µM, 10 sec) és a keltett kalcium-
tranzienseinek amplitúdó viszonyait ellenıriztük. Az eredmények statiszikai értékelése
Student-féle páratlan t-teszttel történt.
A vizsgált sejtpopuláció 119 neuronja közül 49 reagált a 0.33 µM kapszaicinre. A kapszaicin-
szenzitív szenzoros neuronok között az 1 µM PMA-ra 51% (n=25), a 10 µM PMA-ra 53%
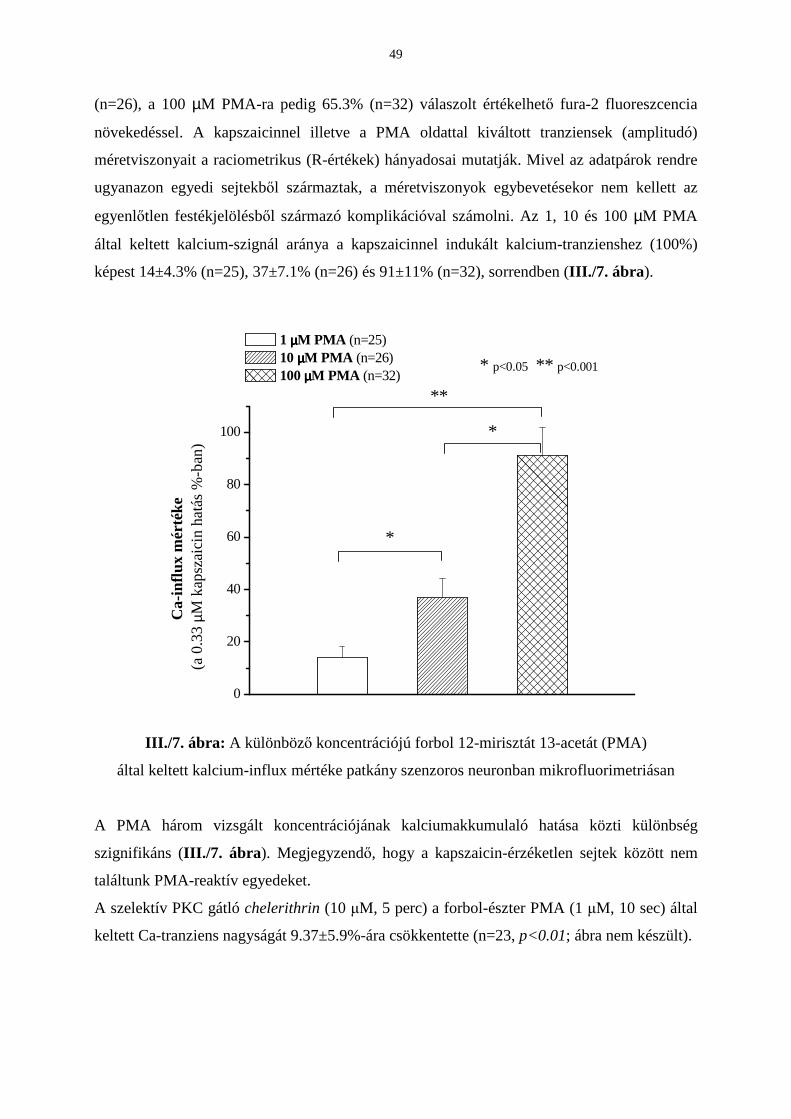
49
(n=26), a 100 µM PMA-ra pedig 65.3% (n=32) válaszolt értékelhetı fura-2 fluoreszcencia
növekedéssel. A kapszaicinnel illetve a PMA oldattal kiváltott tranziensek (amplitudó)
méretviszonyait a raciometrikus (R-értékek) hányadosai mutatják. Mivel az adatpárok rendre
ugyanazon egyedi sejtekbıl származtak, a méretviszonyok egybevetésekor nem kellett az
egyenlıtlen festékjelölésbıl származó komplikációval számolni. Az 1, 10 és 100 µM PMA
által keltett kalcium-szignál aránya a kapszaicinnel indukált kalcium-tranzienshez (100%)
képest 14±4.3% (n=25), 37±7.1% (n=26) és 91±11% (n=32), sorrendben (III./7. ábra ).
0
20
40
60
80
100 *
1 µµµµM PMA (n=25) 10 µµµµM PMA (n=26) 100 µµµµM PMA (n=32)
Ca-
influ
x m
érté
ke
(a 0
.33
µM k
apsz
aici
n ha
tás
%-b
an)
**
* p<0.05 ** p<0.001
*
III./7. ábra: A különbözı koncentrációjú forbol 12-mirisztát 13-acetát (PMA)
által keltett kalcium-influx mértéke patkány szenzoros neuronban mikrofluorimetriásan
A PMA három vizsgált koncentrációjának kalciumakkumulaló hatása közti különbség
szignifikáns (III./7. ábra ). Megjegyzendı, hogy a kapszaicin-érzéketlen sejtek között nem
találtunk PMA-reaktív egyedeket.
A szelektív PKC gátló chelerithrin (10 µM, 5 perc) a forbol-észter PMA (1 µM, 10 sec) által
keltett Ca-tranziens nagyságát 9.37±5.9%-ára csökkentette (n=23, p<0.01; ábra nem készült).
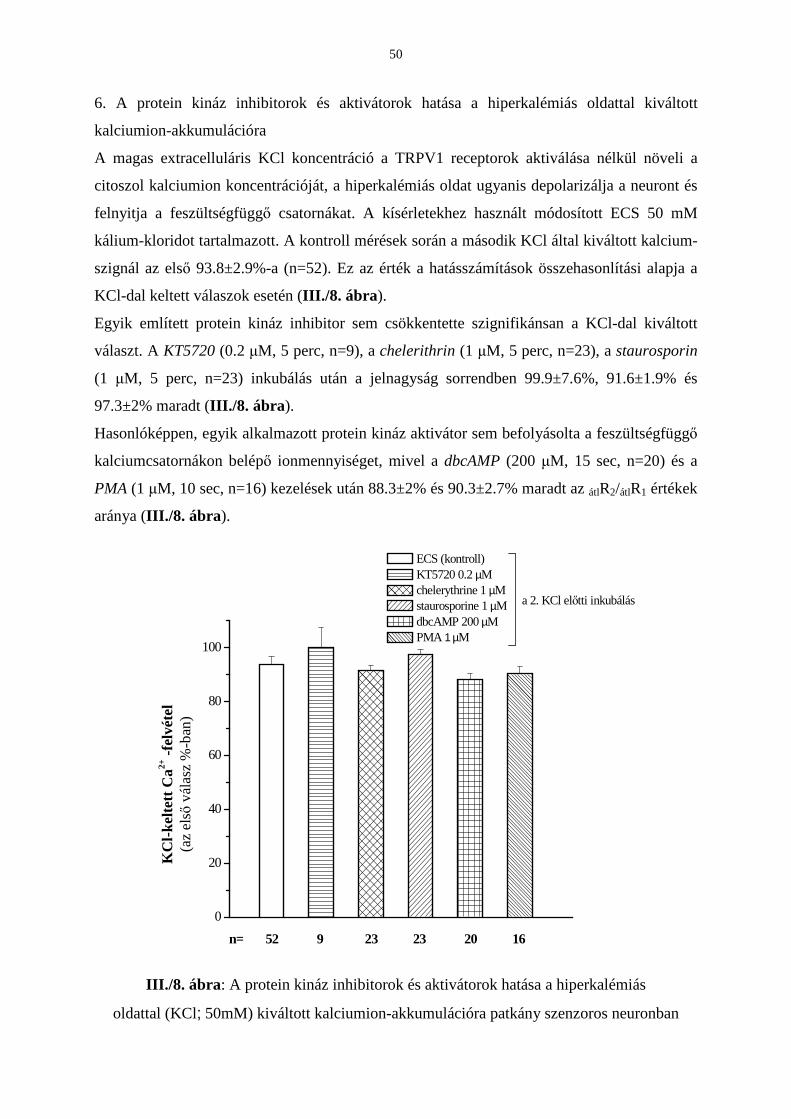
50
6. A protein kináz inhibitorok és aktivátorok hatása a hiperkalémiás oldattal kiváltott
kalciumion-akkumulációra
A magas extracelluláris KCl koncentráció a TRPV1 receptorok aktiválása nélkül növeli a
citoszol kalciumion koncentrációját, a hiperkalémiás oldat ugyanis depolarizálja a neuront és
felnyitja a feszültségfüggı csatornákat. A kísérletekhez használt módosított ECS 50 mM
kálium-kloridot tartalmazott. A kontroll mérések során a második KCl által kiváltott kalcium-
szignál az elsı 93.8±2.9%-a (n=52). Ez az érték a hatásszámítások összehasonlítási alapja a
KCl-dal keltett válaszok esetén (III./8. ábra ).
Egyik említett protein kináz inhibitor sem csökkentette szignifikánsan a KCl-dal kiváltott
választ. A KT5720 (0.2 µM, 5 perc, n=9), a chelerithrin (1 µM, 5 perc, n=23), a staurosporin
(1 µM, 5 perc, n=23) inkubálás után a jelnagyság sorrendben 99.9±7.6%, 91.6±1.9% és
97.3±2% maradt (III./8. ábra ).
Hasonlóképpen, egyik alkalmazott protein kináz aktivátor sem befolyásolta a feszültségfüggı
kalciumcsatornákon belépı ionmennyiséget, mivel a dbcAMP (200 µM, 15 sec, n=20) és a
PMA (1 µM, 10 sec, n=16) kezelések után 88.3±2% és 90.3±2.7% maradt az átlR2/átlR1 értékek
aránya (III./8. ábra ).
0
20
40
60
80
100
KC
l-kel
tett
Ca
2+ -
felv
étel
(az
elsı
vál
asz
%-b
an)
n= 52 9 23 23 20 16
ECS (kontroll) KT5720 0.2 µM chelerythrine 1 µM staurosporine 1 µM dbcAMP 200 µM PMA 1 µM
a 2. KCl elıtti inkubálás
III./8. ábra : A protein kináz inhibitorok és aktivátorok hatása a hiperkalémiás
oldattal (KCl; 50mM) kiváltott kalciumion-akkumulációra patkány szenzoros neuronban
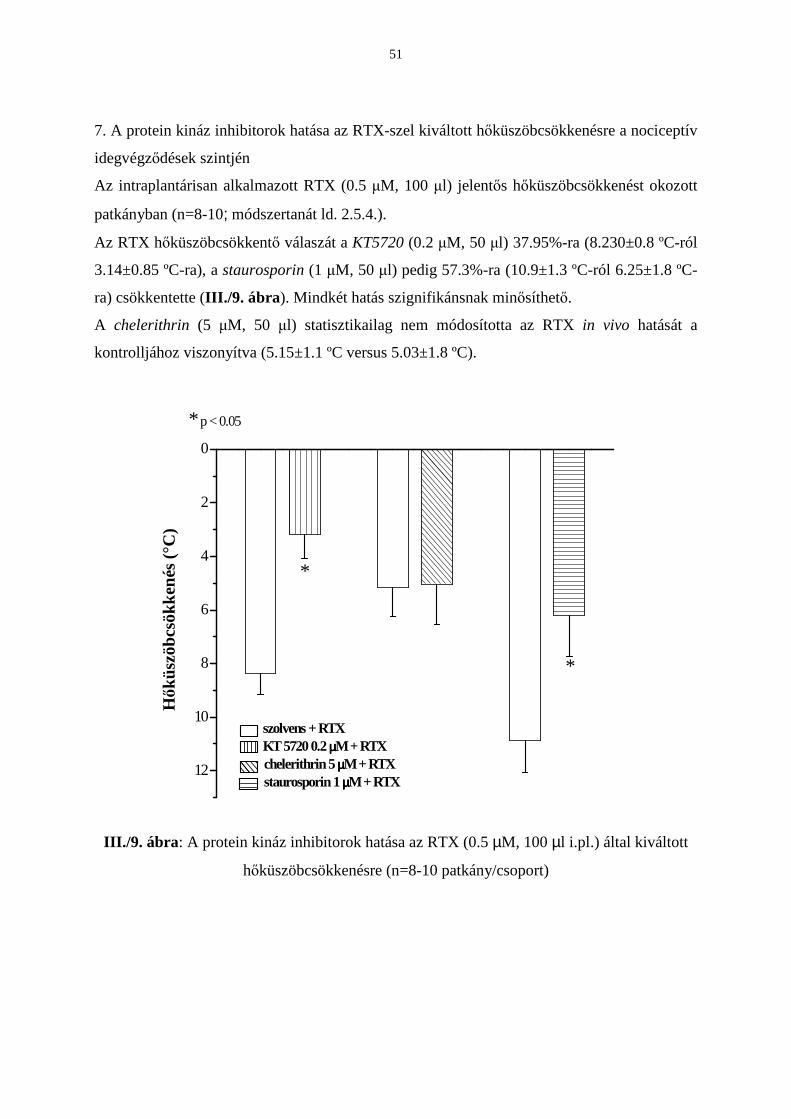
51
7. A protein kináz inhibitorok hatása az RTX-szel kiváltott hıküszöbcsökkenésre a nociceptív
idegvégzıdések szintjén
Az intraplantárisan alkalmazott RTX (0.5 µM, 100 µl) jelentıs hıküszöbcsökkenést okozott
patkányban (n=8-10; módszertanát ld. 2.5.4.).
Az RTX hıküszöbcsökkentı válaszát a KT5720 (0.2 µM, 50 µl) 37.95%-ra (8.230±0.8 ºC-ról
3.14±0.85 ºC-ra), a staurosporin (1 µM, 50 µl) pedig 57.3%-ra (10.9±1.3 ºC-ról 6.25±1.8 ºC-
ra) csökkentette (III./9. ábra ). Mindkét hatás szignifikánsnak minısíthetı.
A chelerithrin (5 µM, 50 µl) statisztikailag nem módosította az RTX in vivo hatását a
kontrolljához viszonyítva (5.15±1.1 ºC versus 5.03±1.8 ºC).
12
10
8
6
4
2
0
*
chelerithrin 5 µµµµM + RTX staurosporin 1 µµµµM + RTX
szolvens + RTX KT 5720 0.2 µµµµM + RTX
Hıkü
szöb
csök
kené
s (°
C)
* p < 0.05
*
III./9. ábra : A protein kináz inhibitorok hatása az RTX (0.5 µM, 100 µl i.pl.) által kiváltott
hıküszöbcsökkenésre (n=8-10 patkány/csoport)
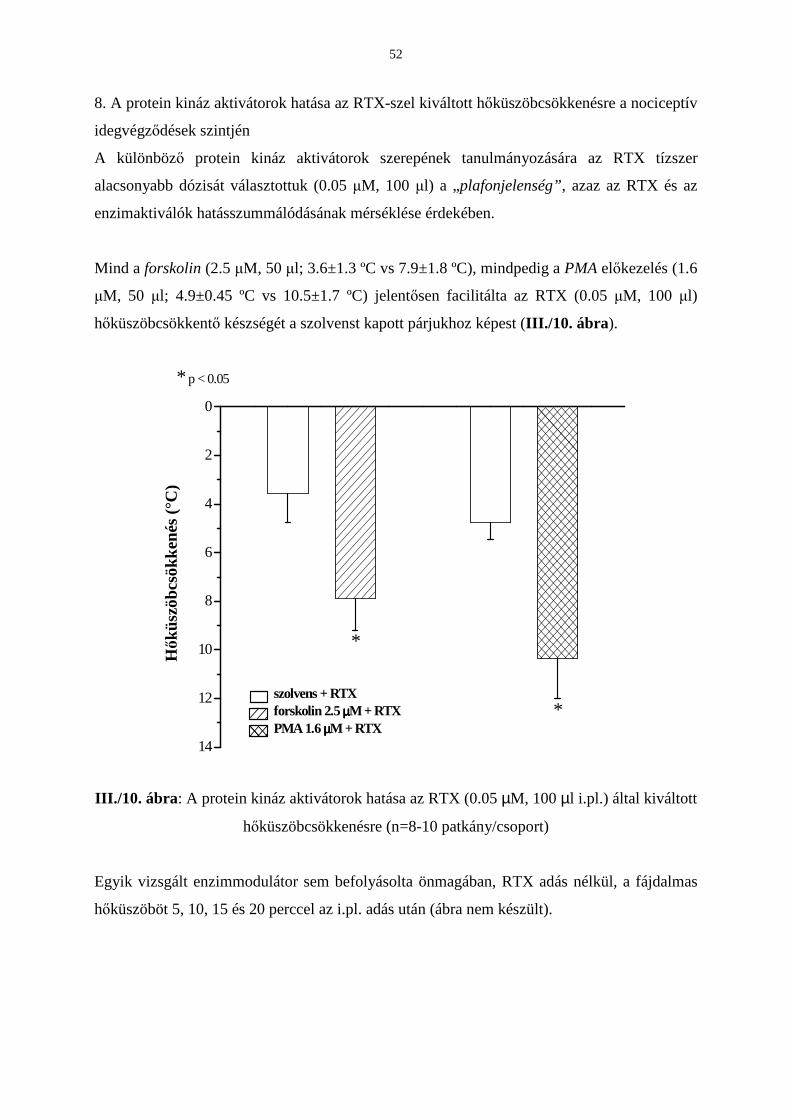
52
8. A protein kináz aktivátorok hatása az RTX-szel kiváltott hıküszöbcsökkenésre a nociceptív
idegvégzıdések szintjén
A különbözı protein kináz aktivátorok szerepének tanulmányozására az RTX tízszer
alacsonyabb dózisát választottuk (0.05 µM, 100 µl) a „plafonjelenség”, azaz az RTX és az
enzimaktiválók hatásszummálódásának mérséklése érdekében.
Mind a forskolin (2.5 µM, 50 µl; 3.6±1.3 ºC vs 7.9±1.8 ºC), mindpedig a PMA elıkezelés (1.6
µM, 50 µl; 4.9±0.45 ºC vs 10.5±1.7 ºC) jelentısen facilitálta az RTX (0.05 µM, 100 µl)
hıküszöbcsökkentı készségét a szolvenst kapott párjukhoz képest (III./10. ábra ).
14
12
10
8
6
4
2
0
*
szolvens + RTX forskolin 2.5 µµµµM + RTX PMA 1.6 µµµµM + RTX
*
Hıkü
szöb
csök
kené
s (°
C)
* p < 0.05
III./10. ábra : A protein kináz aktivátorok hatása az RTX (0.05 µM, 100 µl i.pl.) által kiváltott
hıküszöbcsökkenésre (n=8-10 patkány/csoport)
Egyik vizsgált enzimmodulátor sem befolyásolta önmagában, RTX adás nélkül, a fájdalmas
hıküszöböt 5, 10, 15 és 20 perccel az i.pl. adás után (ábra nem készült).

53
9. További kiegészítı vizsgálatok
Mivel a receptor foszforiláltsági/funkcionális állapotának fontos tényezıje a kalcium-
kalmodulinfüggı kalcineurin általi defoszforilálás (Numazaki és mtsai, 2003), próbálkoztunk
a különbözı foszfatázgátlók (a fenvalerát, a neuroprotektív immunszuppresszáns ciklosporin
A és a cipermetrin) hatásainak vizsgálatával (Docherty és mtsai, 1996; Enan & Matsumura,
1992), de a korábbi megfigyelésekkel összhangban (Fakata és mtsai, 1998) egyik vegyület
sem tudta kivédeni vagy legalábbis mérsékelni a kalcineurin-okozta TRPV1 receptor
defoszforilációt/deszenzitizációt.
Következtetések
Célul tőztük ki az alapaktivitású és a stimulált protein kináz A valamint a protein kináz C
moduláló szerepének szisztematikus összehasonlítását a TRPV1 receptor érzékenységére in
vitro és in vivo patkány modellekben.
Sikerült igazolnunk, hogy a TRPV1 receptor foszforilálásán keresztül módosítani lehet a
csatornafehérje érzékenységét. Eredményeink azonban a protein kinázok szelektív gátlására,
illetve serkentésére érvényes kvantitatív adatokat is tartalmaznak külön a PKA illetve a PKC
vonatkozásában.
Szemben a szelektív PKC inhibitor chelerithrinnel és a nem specifikus protein kináz blokkoló
staurosporinnal, egyedül a cAMP-dependens protein kináz A szelektív inhibitora, a KT5720
csökkentette szignifikánsan a kapszaicin és az RTX által kiváltott kalcium-tranzienst
szenzoros neuronban. A nagyobb koncentrációban alkalmazott KT5720 nonszignifikáns
hatása a relatíve kis esetszám (n=11) okozta nagyfokú szórásnak tudható be.
Hogy a nociceptor végzıdéseken a KT5720 mellett a staurosporin is szignifikánsan
befolyásolta az RTX hıküszöbcsökkentı hatását, egyrészt az eltérı sejtes milliınek
tulajdonítható, másrészt a staurosporin PKA gátló hatásának in vivo érvényesülésével
magyarázható. Mindezekbıl azt a következtetést vonhatjuk le, hogy a TRPV1 receptor
vanilloid-érzékenységét alapszinten elsısorban a protein kináz A tartja fent.
Ugyanakkor mind a PKA, illetıleg mind a PKC aktivátorai (dbcAMP, PMA) facilitálták a
kapszaicin válaszát mikrofluorimetriásan és a PMA illetve a forskolin az RTX-ét az in vivo
viselkedési modellben. Tehát a TRPV1 receptor akár PKC, akár PKA aktiválás esetén képes
szenzibilizálódni, azaz a sejttestbe több kalciumion lép be TRPV1 agonista stimuláció

54
alkalmával, valamint a perifériás végzıdésekben az enzimstimuláció hatására az RTX-
indukált fájdalomreakció is potencírozódik (plafonjelenség).
Eredményeink azonban ellentmondanak néhány korábbi megfigyelésnek, ahol a PKC
inhibitorral történı elıkezelés gátolta a kapszaicinnel keltett kalcium-influxot TRPV1
receptort expresszáló HEK293 sejtekben és mérsékelte a kapszaicin által kiváltott inward
áramot DRG-ben (Zhou és mtsai, 2001; Amadesi és mtsai, 2004). Az ellentmondás lehetséges
oka talán az lehet, hogy velük ellentétben nem DRG sejttesteket, hanem trigeminális
érzıneuronokat használtunk in vitro kísérleteinkben.
Egyik típusú enzimmodulátor sem befolyásolta a hiperkalémiás oldattal kiváltott
kalcium-szignált szenzoros neuronban, mely arra enged következtetni, hogy a vizsgált
enzimmodulátorok hatása kizárólag a TRPV1 receptor szintjén mediálódik, így
nemspecifikus reakciójukkal – az adott körülmények között – nem kellett számolnunk.
A PMA/kapszaicin-érzékeny sejtek aránya a PMA koncentrációjának növelésével nem
szignifikáns mértékben ugyan, de enyhén emelkedett, viszont a PMA által kiváltott jelnagyság
statisztikailag módosult a töménység függvényében. A PMA-val keltett kalcium-szignált a
chelerithrin hatékonyan antagonizálta, mely arra utal, hogy a PMA kalciumakkumuláló
hatása valóban PKC stimuláción keresztül érvényesül.
Irodalomból ismert, hogy a cAMP-PKA jelátviteli útvonal aktiválása a TRPV1 receptor
közvetlen foszforilálásával csökkenti a receptor deszenzibilizációjának mértékét (Bhave és
mtsai, 2002; Mohapatra & Nau, 2003; ld. 3.3. Bevezetés). Ezért a PKA gátlás alatt észlelt
csökkent TRPV1 agonista válasz egy erısebben érvényesülı TRPV1 deszenzibilizáció
következményeként értelmezhetı. Míg a PKA/PKC enzimek aktivitásának fokozása (i)
érzékenyíti a receptort agonistái iránt és/vagy (ii) egy kevésbé deszenzitizált állapotot
eredményez.
A jelenlegi in vitro és in vivo eredményeink tükrében kijelenthetjük, hogy fiziológiásan a
TRPV1 receptor érzékenységének fenntartásában döntıen a protein kináz A
alapaktivitása játszik meghatározó szerepet, míg a protein kináz C nem vagy csak kissé
befolyásolja a receptor nyugalmi státuszát. Mivel az alkalmazott enzimaktivátorok
eredményesen érzékenyítették a receptort agonistái iránt bizonyítást nyert, hogy mindkét
vizsgált protein kináz stimulálása hatékony a receptor szenzitizált állapotának

55
kialakításában mind centrálisan (TRG sejttest), mind pedig a perifériás
érzıidegvégzıdések szintjén (DRG végzıdés).

56
3.4. A TRPV1 RECEPTOR-MEDIÁLT INTRACELLULÁRIS KALCI UMIONSZINT
EMELKEDÉS VIZSGÁLATA EXTRACELLULÁRIS KALCIUM HIÁNYÁ BAN
Bevezetés
A kapszaicinnel foglalkozó terjedelmes irodalom régóta egyetért abban, hogy normál
körülmények között a vanilloidokkal megnyitott TRPV1 receptorcsatorna komplexen át a
kalciumionok a koncentrációgradienst követve az extracelluláris folyadékból lépnek be a
citoszolba. Mint a korábbi fejezetekben láttuk, mikrofluorimetriás módszerünk e reakció
közvetlen mérésére alkalmas, de nem tájékoztat az ionváltozás forrásairól. Az intracelluláris
szabad kalciumion koncentráció átmeneti emelkedésének egyik fiziológiás következménye a
szenzoros neuronok kapszaicin válaszainak deszenzitizációja illetve tachyfilaxisa, melynek
extracelluláris kalciumtól függı természetét többen megerısítették (Cholewinski és mtsai,
1993; Garcia-Hirshfield és mtsai, 1995; Koplas és mtsai, 1997; Mandadi és mtsai, 2004).
Érzékeny méréstechnika segítségével bizonyítható az is, hogy a belépett kalcium a sejten
belüli raktárakból további szabad kalciumionokat mobilizál, különösen az endoplazmatikus
retikulumból (ER) igen gyorsan és jelentıs mennyiségben, miközben különbözı
pufferrendszerek védik a sejtet a kalciumionok káros hatásaitól illetve biztosítják a sejt
kalcium homeosztázisát (Nicholls, 1986; Duchen, 2000a és 2000b).
Mivel a kapszaicin zsírokban jól oldódik és a lipidmembránon át akadálymentesen jut a
neuronok belsejébe, így felmerül a lehetıség, hogy az ER-ben található, kapszaicin által
aktivált TRPV1 molekulák közvetlenül hozzájárulnak az intracelluláris kalciumionszint
emelkedéséhez az ER-ben található kalcium mobilizációjával. Ebbıl a szempontból a
szenzoros neuronok és a klónozással létrehozott sejtmodellek viselkedése eltérı lehet (Lázár
és mtsai, 2003). Ebben a tekintetben meglehetısen ellentmondásos a TRPV1 receptor
irodalma.
Feltételezik, hogy az intracelluláris raktárak membránjában található TRPV1 receptor képes
az ER kiürítésével az intracelluláris szabad kalciumion koncentráció növelésére extracelluláris
kalcium hiányában is. Ilyen megfigyelést tettek TRPV1-vel transzfektált HEK293 sejtekben
(Oláh és mtsai, 2001; Marshall és mtsai, 2003) és hátsógyöki neuronban (DRG) egyaránt
(Eun és mtsai, 2001; Oláh és mtsai, 2001; Liu és mtsai, 2003). Mások viszont az ER-lokalizált
TRPV1 receptor szerepét megkérdıjelezték szenzoros neuronban (Cholewinski és mtsai,
1993; Garcia-Hirshfield és mtsai, 1995).

57
Érdemes hangsúlyt fektetni a koncentráció viszonyokra. Extracelluláris kalcium hiányában a
kapszaicinnel kiváltott kalcium felszabadulás EC50 értéke Eun és munkatársai szerint 13.5 µM
(2001), míg Sf9 rovarsejtben expresszált TRPV1 receptor esetén ~ 10 µM (Wisnoskey és
mtsai, 2003). Liu és munkatársai (2003) kalciummentes közegben a kapszaicinnel keltett
kalcium felszabadulás EC50 értékét csak 133 nM-nak találták DRG-ben, viszont ık
meglehetısen alacsony, csupán 0.5 mM EGTA tartalmú oldatot használtak a Ca2+ ionok
keletálására. Marshall és kollégái arra a következtetésre jutottak (2003), hogy a potensebb
reziniferatoxinnal ellentétben a 100 µM kapszaicin sem okozott kalciumion felszabadulást
hTRPV1-HEK293 sejtek endoplazmás retikulumjaiból. Ezzel szemben Eun és munkatársai
2001-es cikkében az olvasható, hogy már az 1 µM kapszaicin is képes az intracelluláris
raktárak kiürítésére patkány DRG-ben.
Mindebbıl arra lehet következtetni, hogy kalciummentes közegben többnyire csak a sokkal
magasabb kapszaicin koncentráció vagy a potensebb agonista RTX képes a TRPV1 receptor
izgatása útján csekély mértékő intracelluláris kalciumion mobilizációt okozni. (Itt vissza
szeretnék utalni a 3.2 fejezetben taglalt saját eredményre, miszerint a kapszaicin EC50 értéke
36 nM a rTRPV1-expresszáló HT5-1 sejtvonal össz-45Ca2+-felvétele alapján.)
Saját lehetıségeink korlátozottak ugyan, de azt ellenırizni tudjuk, hogy vajon a tenyésztett
TRG neuronok és a rTRPV1-HT5-1 sejtek reakciókészsége hogyan változik meg
kalciummentes közegben. Ezt a vizsgálatot feltétlenül érdemesnek véltük elvégezni, hiszen
ezt a sejtmodellt korábban nem használták ilyen jellegő vizsgálatokra.
Mindezek alapján tehát fontosnak tartottuk megvizsgálni, egyrészt, hogy az általunk használt
sejttípusok produkálnak-e extracelluláris kalciumtól független, TRPV1-mediált választ,
másrészt, hogy mennyire hasonlítanak egymásra e tekintetben.
Eredmények
Az intracelluláris kalciumionszint-mérést fluoreszcens módszerrel az elızıekben már
ismertetett módon végeztük, azzal a különbséggel, hogy az 1 µM fura-2 fluoreszcens,
kalciumindikátor festéket tartalmazó inkubációs oldatból és az ECS-bıl is kihagytuk az 1 mM
kalciumkloridot (CaCl2). Ezt neveztük el 1-es típusú oldatnak.
Egy másik kísérletsorozatunkban azt vizsgáltuk, hogyha az 1-es típusú oldatot kiegészítjük 3
mM EGTA-val a maradék kalciumionok lekötése céljából (2-es típusú oldat), tapasztalunk-e
szignifikáns eltérést az 1-es illetve a 2-es típusú oldattal kapott eredményeink között.
Tudomásunk szerint az impermeábilis EGTA nem befolyásolja közvetlenül a sejten belüli

58
folyamatokat. A kísérletek kivitelezése a korábbiakhoz képest ilymódon csak az ECS
összetételében módosult.
Normál ECS helyett vagy az 1-es vagy a 2-es típusú kalciummentes ECS oldatokat
használtuk. A kapszaicin 1 µM-os oldatának készítéséhez a megfelelı kalciummentes ECS
oldatot következetesen alkalmaztuk. A kísérletek elıtt a 4 beömlıtartályt (melyek valójában
10-20 ml-es fecskendık) alaposan kimostuk 70%-os alkohollal, majd, amelyik tartályba a
kalciummentes oldat került (akár ECS, akár kapszaicin oldat), azt elızıleg a megfelelı típusú
kalciummentes oldattal öblítettük. A beömlık egyikébe a sejtek kapszaicin-érzékenységének
tesztelése céljából normál, 1 mM CaCl2-ot tartalmazó ECS-ben hígított kapszaicint (1 µM)
mértünk.
A kontroll kísérletekben a kalcium-tranziensek kiváltásához a kapszaicint (1 µM) 10
másodpercig adtuk a sejtekre, míg a kalciummentes oldatok alkalmazása esetén a kapszaicin
expozíció (1 µM) 1 percig tartott.
A raciometrikus értéket (R=F340/F380) az idı függvényében az Axon Imaging Workbench
2.1 szoftverrel regisztráltuk, a kapott függvényeket a Microcal Origin 7.0 program
segítségével elemeztük. Kiértékelésre csak a reakciók észlelt csúcsértékeit (maximális R
érték) használtuk.
A reagáló sejtek arányát sejttípus szerint kétféleképpen számoltuk. Az áramlási citométerrel
szortírozott HT5-1 sejtek esetén viszonylag egyszerő dolgunk volt, mivel gyakorlatilag
homogén sejtpopulációval dolgoztunk. A látómezıben lévı reagáló sejtek számát
viszonyítottuk a látómezıben lévı összsejt számhoz. Mivel TRG neuronok tenyészetében
gliasejtek is szép számmal vannak, – de ezek a gyakorlott szem számára jól elkülöníthetık a
szenzoros neuronoktól – a reagáló neuronok számát osztottuk a kijelölt, neuronnak vélt sejtek
számával (szemben a látómezıben lévı összsejt számmal) és felszoroztuk 100-zal. Az adatok
kiértékelése után megvizsgáltuk, hogy tapasztalunk-e szignifikáns eltérést az alkalmazott
oldatokkal kapott eredmények között. A statiszikai értékelés Student-féle páratlan t-próbával
illetve Fischer statisztikai tesztje alapján történt.
A normál összetételő ECS-sel végzett kontroll kísérleteinkben szignifikáns különbséget
tapasztaltunk a kapszaicinnel kiváltott (1 µM, 10 sec) kalcium-tranziensek nagyságában a
rTRPV1-HT5-1 sejtek és a TRG neuronok között (#p<0.0001). A maximális fluoreszcencia
növekedések átlaga (R) az elıbbiben 0.62±0.03 (n=19), az utóbbiban pedig 0.377±0.02
(n=14). A mintában észlelt legkisebb, minimális fluoreszcencia növekedés a rTRPV1-HT5-1
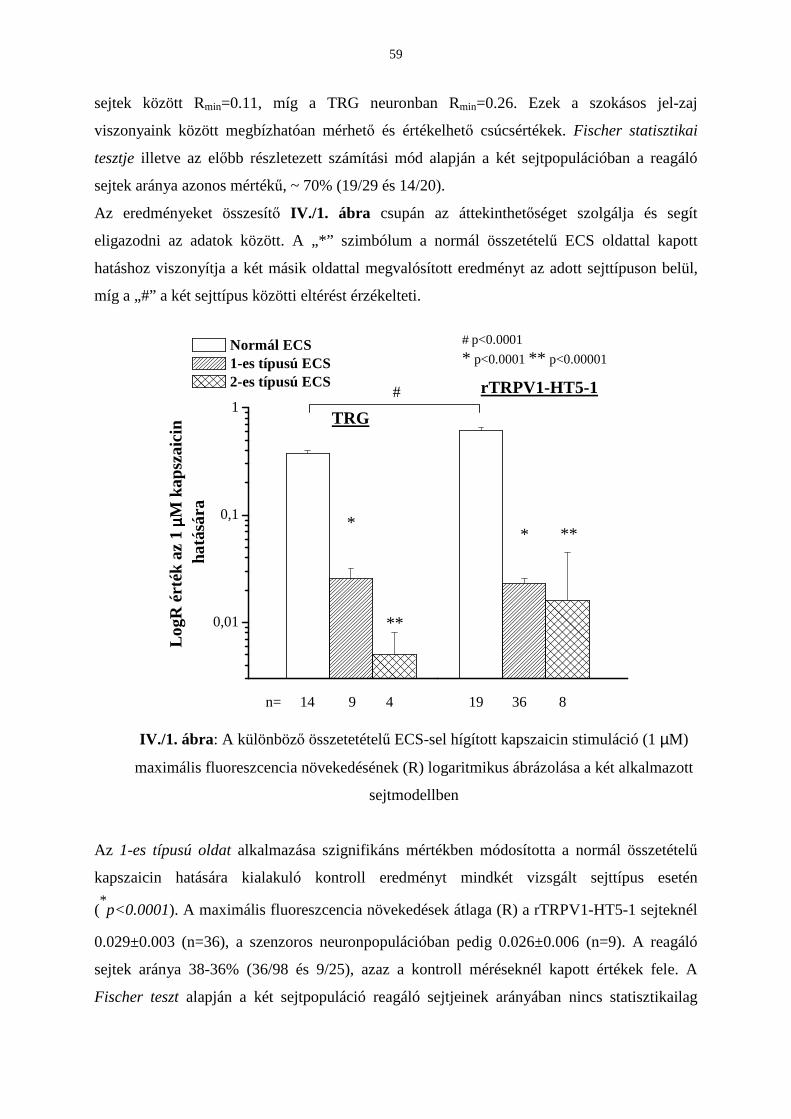
59
sejtek között Rmin=0.11, míg a TRG neuronban Rmin=0.26. Ezek a szokásos jel-zaj
viszonyaink között megbízhatóan mérhetı és értékelhetı csúcsértékek. Fischer statisztikai
tesztje illetve az elıbb részletezett számítási mód alapján a két sejtpopulációban a reagáló
sejtek aránya azonos mértékő, ~ 70% (19/29 és 14/20).
Az eredményeket összesítı IV./1. ábra csupán az áttekinthetıséget szolgálja és segít
eligazodni az adatok között. A „*” szimbólum a normál összetételő ECS oldattal kapott
hatáshoz viszonyítja a két másik oldattal megvalósított eredményt az adott sejttípuson belül,
míg a „#” a két sejttípus közötti eltérést érzékelteti.
0,01
0,1
1
**
***
# p<0.0001
* p<0.0001 ** p<0.00001
Normál ECS 1-es típusú ECS 2-es típusú ECS
TRG
rTRPV1-HT5-1
n= 14 9 4 19 36 8
#
*
LogR
ért
ék a
z 1 µµ µµ
M k
apsz
aici
n ha
tásá
ra
IV./1. ábra: A különbözı összetetételő ECS-sel hígított kapszaicin stimuláció (1 µM)
maximális fluoreszcencia növekedésének (R) logaritmikus ábrázolása a két alkalmazott
sejtmodellben
Az 1-es típusú oldat alkalmazása szignifikáns mértékben módosította a normál összetételő
kapszaicin hatására kialakuló kontroll eredményt mindkét vizsgált sejttípus esetén
(*p<0.0001). A maximális fluoreszcencia növekedések átlaga (R) a rTRPV1-HT5-1 sejteknél
0.029±0.003 (n=36), a szenzoros neuronpopulációban pedig 0.026±0.006 (n=9). A reagáló
sejtek aránya 38-36% (36/98 és 9/25), azaz a kontroll méréseknél kapott értékek fele. A
Fischer teszt alapján a két sejtpopuláció reagáló sejtjeinek arányában nincs statisztikailag
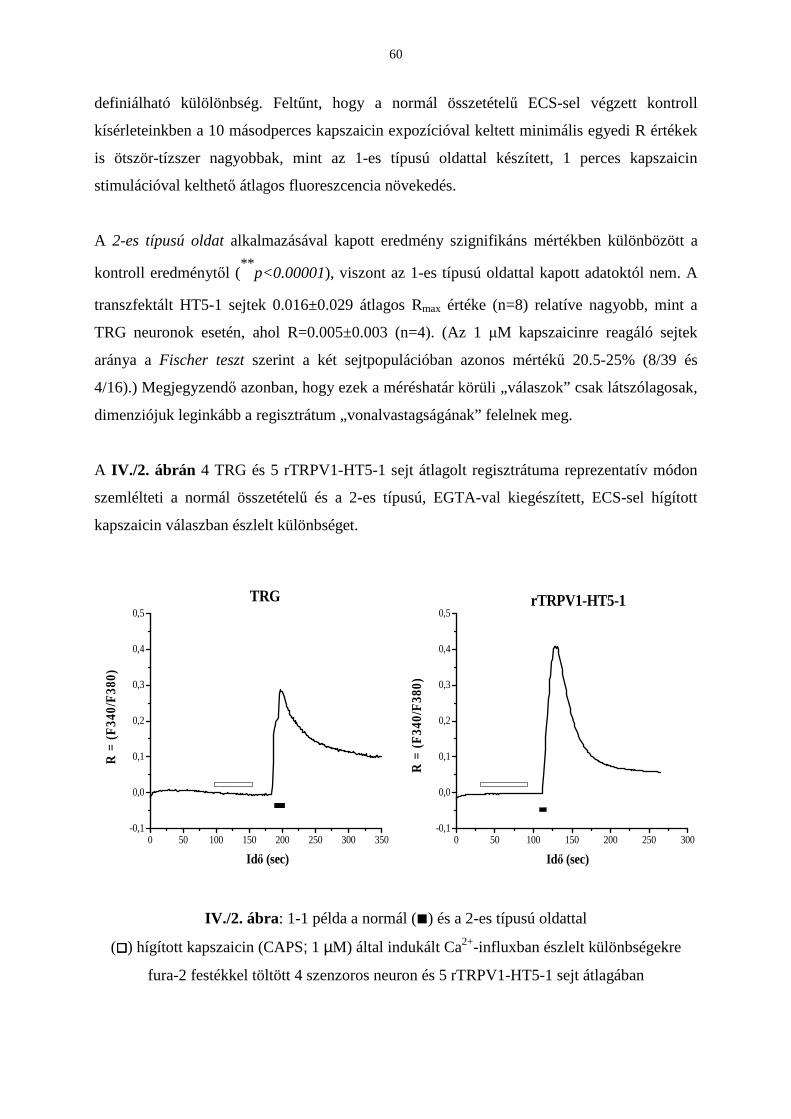
60
definiálható külölönbség. Feltőnt, hogy a normál összetételő ECS-sel végzett kontroll
kísérleteinkben a 10 másodperces kapszaicin expozícióval keltett minimális egyedi R értékek
is ötször-tízszer nagyobbak, mint az 1-es típusú oldattal készített, 1 perces kapszaicin
stimulációval kelthetı átlagos fluoreszcencia növekedés.
A 2-es típusú oldat alkalmazásával kapott eredmény szignifikáns mértékben különbözött a
kontroll eredménytıl (**
p<0.00001), viszont az 1-es típusú oldattal kapott adatoktól nem. A
transzfektált HT5-1 sejtek 0.016±0.029 átlagos Rmax értéke (n=8) relatíve nagyobb, mint a
TRG neuronok esetén, ahol R=0.005±0.003 (n=4). (Az 1 µM kapszaicinre reagáló sejtek
aránya a Fischer teszt szerint a két sejtpopulációban azonos mértékő 20.5-25% (8/39 és
4/16).) Megjegyzendı azonban, hogy ezek a méréshatár körüli „válaszok” csak látszólagosak,
dimenziójuk leginkább a regisztrátum „vonalvastagságának” felelnek meg.
A IV./2. ábrán 4 TRG és 5 rTRPV1-HT5-1 sejt átlagolt regisztrátuma reprezentatív módon
szemlélteti a normál összetételő és a 2-es típusú, EGTA-val kiegészített, ECS-sel hígított
kapszaicin válaszban észlelt különbséget.
0 50 100 150 200 250 300-0,1
0,0
0,1
0,2
0,3
0,4
0,5
R =
(F
340/
F38
0)
Idı (sec)
rTRPV1-HT5-1
0 50 100 150 200 250 300 350-0,1
0,0
0,1
0,2
0,3
0,4
0,5
TRG
R =
(F
340/
F38
0)
Idı (sec)
IV./2. ábra: 1-1 példa a normál (�) és a 2-es típusú oldattal
(����) hígított kapszaicin (CAPS; 1 µM) által indukált Ca2+-influxban észlelt különbségekre
fura-2 festékkel töltött 4 szenzoros neuron és 5 rTRPV1-HT5-1 sejt átlagában

61
Az egyedi sejtek regisztrátumain gyakori „zajszintet” az alkalmazott jelátlagolás elsimította,
de értékelhetı reakciónak semmi nyoma. Érdemes megfigyelni a normál összetételő ECS-ben
oldott kapszaicin stimulációjakor jelentkezı azonnali markáns fluoreszcencia növekedést.
Következtetések
Valamely sejtben az észlelt fura-2 jel nagyságát döntıen három tényezı határozza meg.
Közülük természetesen a legrelevánsabb a szabad kalciumion koncentráció illetve annak
változása. Nem szabad elhanyagolni azonban a sejtbe került és kalciumkötésre alkalmas
festékmolekulák mennyiségét és a reakcióképes TRPV1 receptorok számát sem, melyek
sejtrıl sejtre kisebb-nagyobb mértékben befolyásolják a detektált fura-2 szignál
méretviszonyait. Egyrészt e járulákos komponensek biológiai fontossága aligha vitatható,
másrészt sajnos nincsen mód annak becslésére, hogy közremőködésük mennyire „torzítja” a
keresett kalciumionszint változás rovására írható komponenst.
A normál összetételő ECS-sel hígított 1 µM kapszaicin a rTRPV1-HT5-1 sejtekben közel
kétszer akkora R értéket (maximális fura-2 fluoreszcencia növekedések átlaga) produkált,
mint szenzoros neuronban. Ez valószínőleg az elıbbi sejttípus TRPV1 receptorainak
overexpressziójával magyarázható. Extracelluláris kalcium hiányában a rTRPV1-HT5-1
sejtvonal a TRG neuronokhoz hasonlóan nem eredményezett jelentıs (átlR<0.03)
intracelluláris kalciumszint növekedést. Az 1-es típusú oldat nyomokban bizonyosan
tartalmaz kalciumionokat, melyekhez valószínőleg a sejtek fehérjéirıl ledisszociálodó
kalciummennyiség is társul. Az 1-es típusú oldat alkalmazásakor a kismértékő, de még
detektálható kapszaicin indukálta fura-2 intenzitásfokozódás (mely a kontroll mindössze
5-7%-a) leginkább az oldatok kalciumionokkal történı szennyezıdésének rovására írható,
nem pedig az intracelluláris kalciumraktárak kiürítésének tulajdonítható . Az EGTA
tartalmú kalciummentes közeg (2-es oldat) gyakorlatilag nullára csökkentette a kontroll
válaszokat mindkét sejtféleségben, de az 1-es illetve a 2-es típusú oldattal kapott
eredményeink között nem kaptunk szignifikáns mértékben kimutatható különbséget. A
módszerek fejezetben taglalt számítási mód illetve a Fischer-féle statisztika alapján a reagáló
sejtek százalékos aránya a különbözı összetetételő ECS oldatban hígított kapszaicin hatására
a két alkalmazott sejtmodellben közel azonos mértékő.
Eredményeink több laboratórium korábbi megfigyeléseit erısítik meg, akik velünk
egybehangzóan szenzoros neuronok kapszaicin válaszainak hiányát illetve erıs redukcióját

62
tapasztalták kalciummentes közegben (Cholewinski és mtsai, 1993; Koplas és mtsai, 1997;
Liu & Simon, 1998), míg mások az érzıneuronok endoplazmás retikulumjaiból származónak
vélik a regisztrált kismértékő kalciumion felszabadulást kalciummentes kísérleti körülmények
között (Eun és mtsai, 2001; Oláh és mtsai, 2001; Liu és mtsai, 2003). Elképzelhetı, hogy
transzfektált sejtek esetén a nagymértékő TRPV1 receptor expresszióval (overexpresszió),
míg natív sejteknél a több napig tartó tenyésztéssel magyarázható az utóbbi jelenség, melynek
eredményeként a feleslegben termelıdı TRPV1 membránproteinek intracelluláis
membránokban is fixálódnak. Lehetséges továbbá, hogy az alkalmazott sejttípus, az agonista
valamint az EGTA koncentrációjának különbségében keresendı az ellentmondás oka.
Nem tekintjük ugyan lezártnak a témakört, hiszen további metodikák bevonásával (például
konfokális mikroszkóp, immuncitokémia) szeretnénk álláspontunkat alátámasztani, de eddigi
eredményeink tükrében mindenképpen kijelenthetı, hogy a rTRPV1-HT5-1 sejtvonal ebben
a tekintetben a TRG neuronok megbízható modelljének tőnik , így alkalmas a TRPV1
receptor további funkcionális és farmakológiai tanulmányozására.

63
3.5. A TRPV1 KAPSZAICIN RECEPTOR AKTIVÁLÁS ÉS A MITOKOND RIUM
FUNKCIÓ KÖZÖTTI VISZONY VIZSGÁLATA
Bevezetés
Régóta ismert a mitokondriumok kitüntetett szerepe a sejtek kalcium homeosztázisának
szabályozásában (Nicholls, 1986; Duchen, 1992). Az is régóta ismert, hogy felnıtt patkányok
trigeminális B-típusú neuronjaiban hónapokig tartó mitokondrium-duzzadás jön létre
szisztémás kapszaicin kezelést követıen (Szolcsányi és mtsai, 1975). Szıke és munkatársai
(2002a és b) kísérleteikben kimutatták, hogyha újszülött patkányokat kapszaicinnel (50
mg/kg) vagy a TRPV1 receptor egyik feltételezett endogén ligandjával (ld. 3.2. fejezet), az
anandamiddal (1 mg/kg) s.c. elıkezelték, a 3. hétre antinocicepciót, a neurogén gyulladás
megszőnését, a B-típusú szenzoros neuronok számbeli csökkenését tapasztalták.
Elektronmikroszkópos felvételeken a B-típusú sejtek egy részében szelektíven, hetekre
fenntartott tünet a mitokondriumok krisztáinak eltőnése („duzzadt mitokondriumok”), de se
apoptózis, se nekrózis jelei nem mutatkoztak. Az eleinte elenyészı, de idıvel fokozódó
sejtpusztulás feltételezhetı oka a károsodott sejtek csökkent NGF felvétele, hiszen a
kemonociceptorok differenciálódásához, valamint fenotípusmintázatuk fenntartásához NGF
jelenléte szükséges (Shu & Mendell, 2001). Érthetı módon a TRPV1-negatív A-típusú
neuronok sem morfológiai, sem sejtszámbeli elváltozást nem mutattak.
Dedov és Roufogalis is megerısítették Szolcsányi és mtsai megfigyelését (1975) és
kimutatták, hogy a hosszantartó, funkcionális TRPV1/VR1 deszenzitizációhoz a
mitokondriumok károsodása is hozzájárul, a mitokondriumokat találóan a „fájdalom
neuronális memóriája”-ként emlegetik (2000).
Feltételezésünk, hogy a TRPV1 receptor aktiváció intracelluláris kalciumkoncentráció
növekedés közvetítésével változtathatja meg a mitokondriumok szerkezetét, hiszen ezen
organellumok jelentıs szerepet játszanak az intracelluláris Ca2+-szint változásainak térbeli és
idıbeli összehangolásában, szabályozásában (Duchen, 2000a és 2000b; Hajnóczky és mtsai,
2000; Rizzuto és mtsai, 2000). Ugyanakkor a mitokondriális kalcium felvétel a mitokondriális
mátrix kalcium-regulált enzimeinek szabályozásával hozzájárul a sejt energiaegyenlegének
beállításához is. A központi mitokondriális bioenergetikai paraméter, a proton elektrokémiai
potenciál (∆p~180 mV), mely miközben a protonokat a belsı mitokondrium membránon át a
mátrixba hajtja ATP molekulákat termel. A proton elektrokémiai potenciál komponensei a
mitokondrium belseje felé negatív 150 mV körüli membránpotenciál (∆Ψm) és egy -0.5

64
egységnyi pH grádiens (a mátrix alkalikusabb) (Nicholls & Budd, 2000; Nicholls & Ward,
2000). A ∆p a mitokondriális ATP szintézis kontrollálásában, a ∆Ψm pedig a kalcium
szekvesztrálásában vesz részt. Bármelyik folyamat elégtelensége az energiatermelés
romlásához, a kalcium homeosztázisának zavarához és következményes sejtelhaláshoz vezet.
Werth és Thayer megfigyelése alapján (1994) a relatíve kismértékő kalcium beáramlást az
idegsejt nem-mitokondriális mechanizmussal, a kalcium kipumpálásával ellensúlyozza.
Viszont a nagy kalciumterhelést a kalciumionok gyors mitokondriális felvételével igyekszik
kompenzálni. A belsı mitokondriális membránon át a kalciumionokat egy ruténium vörös-
szenzitív, divalens kationokra szelektív uniporter juttatja keresztül, mely folyamat hajtóereje a
belsı membrán két oldala közötti potenciálkülönbség (Khodorov és mtsai, 1999; Duchen,
2000a és b). Kismértékő kalcium felvétel a mitokondriumba a mitokondriális
membránpotenciál reverzibilis csökkenéséhez vezet. Nagymértékő kalciumion beáramlás
viszont a membránpotenciál teljes mértékő és végleges összeomlását vonhatja maga után.
Szolcsányi a primér szenzoros neuronok kapszaicinnel kiváltott tartós deszenzitizációjában
jelentıs szerepet tulajdonít az in vivo kísérletekkel igazolt morfológiai változásoknak, melyek
talán a funkcionális történések mögötti károsodásnak tekinthetık, bár ezen összefüggés ok-
okozati viszonya tisztázatlan (Szolcsányi és mtsai, 1975; Szolcsányi 1993, 2005; Sándor &
Szállási, 2005).
Célszerőnek véltük in vitro elemzéssel feltárni a TRPV1 receptor aktiválás és a mitokondrium
funkció közötti kapcsolatot kettıs fluoreszcens jelölés alkalmazásával. A mitokondrium funkció
becslésére egy redisztribúciós, pozitív töltéső, lipofil kationfestéket (rhodamin 123, röviden rhod-
123) választottunk. E festék optikai tulajdonságai megengedik a mitokondriális membránpotentiál
(∆Ψm vagy deltapszi-m) mérés kombinálását a raciometrikus, kalciumindikátor fura-2 festékre
alapozott citoszolikus kalciumionszint változásának ([Ca2+] ic) egyidejő detektálásával.
A kettıs fluoreszcens jelölések alkalmazása a szigorú technikai követelmények miatt ritka az
irodalomban, a közölt munkák legtöbbje vagy a glutamát toxicitásával foglalkozik (Budd &
Nicholls, 1996a és b; Vergun és mtsai, 1999), vagy másfajta fluorofórral dolgoztak (Krieger &
Duchen, 2002). Fontos metodikai újításunk a korábbi, idıben lassúbb, széthúzott illetve felváltva
végzett mérésekhez képest (Duchen, 1992; Budd & Nicholls, 1996a és b; Dedov és mtsai, 2001),
hogy az együtt és dinamikusan változó két faktort relatíve gyors, szimultán, lehetıségeink
határáig feszített tér- és idıbeli feloldással próbáltuk megfigyelni.

65
Eredmények
A relatív intracelluláris szabad kalciumionszint és a mitokondriális membránpotenciál
változás szimultán méréstechnikájának kivitelezését kettıs fluoreszcens festéssel már vázoltam
a 2.2.2. fejezetben. A citoplazmatikus szabad kalciumkoncentráció ([Ca2+] ic) idıbeli változása
és a deltapszi-m (∆Ψm) közti összefüggések feltárására újszülött patkányok tenyésztett
trigeminális neuronjait (TRG) és rTRPV1-el transzfektált HT1080 sejteket (rTRPV1-HT5-1)
használtunk.
A H+-ionofór FCCP (karbonil-cianid 4-[trifluorometoxi]-fenilhidrazon, 5 µM) a
mitokondriális membránpotenciált reverzibilisen felfüggeszti, ezért a mitokondriumban tárolt
kalcium a citoplazmába jut. A kapszaicinnel kiváltott szignálokat az FCCP által keltett (5 µM,
20 sec) rhod-123 és fura-2 referenciajelekhez viszonyítottuk.
Kísérleteinkben a kapszaicint 0.33 és 3.3 µM koncentrációban 10-10 szekundumig
alkalmaztuk. Minden egyes méréskor a kapszaicin csak egyféle koncentrációját adtuk egymás
után háromszor, 3-5 perces mosási periódusok közbeiktatásával. Az eredmények elsı három
pontjában csak az elsı kapszaicin-teszt reakciói kerültek kiértékelésre. A leíró statisztikák, a
hisztogrammok a Shapiro és Wilk próbák szerint készültek. A regressziós és exponenciális
görbeillesztéseket, az idıállandókat és az eredmények statiszikai értékelését a Student-féle
páros illetve páratlan t-teszttel az Origin 7.0 alapján (Microcal Software Inc., MA) végeztük.
1. Az V./1. ábra „a” jelő szenzoros neuronjának példáján keresztül elemzünk és értelmezünk
egy jellemzı szimultán reakciópárt. Arteficiális színskála szemlélteti a két szignál
fluoreszcenciájában észlelhetı változásokat a két alkalmazott stimulus hatására (V./1.a ábra).
Az intenzitás fokozódásakor egyre inkább a vörös szín válik dominánssá. Az idı
függvényében regisztrált adatpontok a két fluoreszcens szignál idıbeli változását 1 Hz-es
mintavétellel hően követik (V./1.b ábra). Ez a feloldás a viszonylag lassú jelek
dokumentálására bıven elég. A fura-2 szignál felfelé történı kitérése a citoszolikus
kalciumkoncentráció emelkedésére utal. A rhod-123 szignál emelkedése a belsı
mitokondriummembrán depolarizációjára (membránpotenciál csökkenés), míg csökkenése a
repolarizációra vezethetı vissza.
Az „a” jelzéső, kapszaicin-szenzitív érzıneuronban kapszaicin hatására (0.33 µM, 10 sec)
nem történt rhod-123 intenzitásfokozódás a fura-2-szignál fluoreszcenciájában tapasztalható
jelentıs növekedéshez képest (V./1.a & b ábrák).
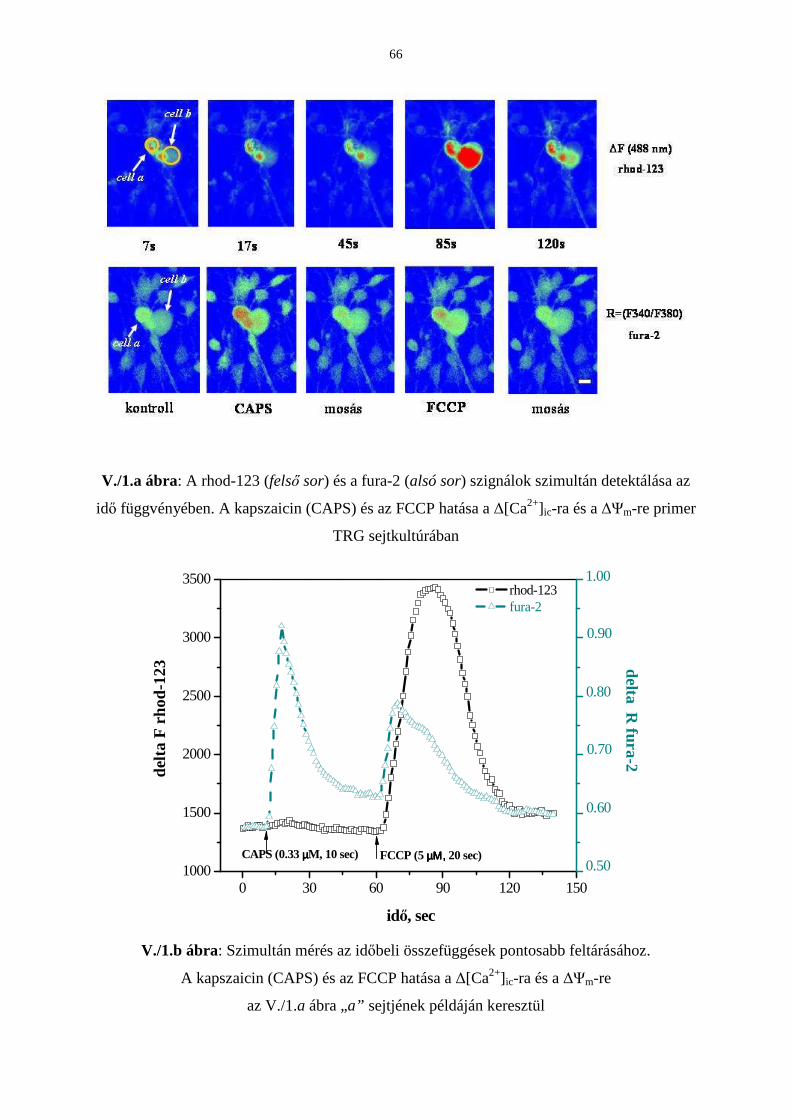
66
V./1.a ábra: A rhod-123 (felsı sor) és a fura-2 (alsó sor) szignálok szimultán detektálása az
idı függvényében. A kapszaicin (CAPS) és az FCCP hatása a ∆[Ca2+] ic-ra és a ∆Ψm-re primer
TRG sejtkultúrában
0 30 60 90 120 1501000
1500
2000
2500
3000
3500
rhod-123 fura-2
idı, sec
delta
F r
hod-
123
CAPS (0.33 µµµµM, 10 sec) FCCP (5 µΜ, µΜ, µΜ, µΜ, 20 sec)
1.00
0.90
0.70
0.80
0.60
delta R fura-2
0.50
V./1.b ábra: Szimultán mérés az idıbeli összefüggések pontosabb feltárásához.
A kapszaicin (CAPS) és az FCCP hatása a ∆[Ca2+] ic-ra és a ∆Ψm-re
az V./1.a ábra „a” sejtjének példáján keresztül
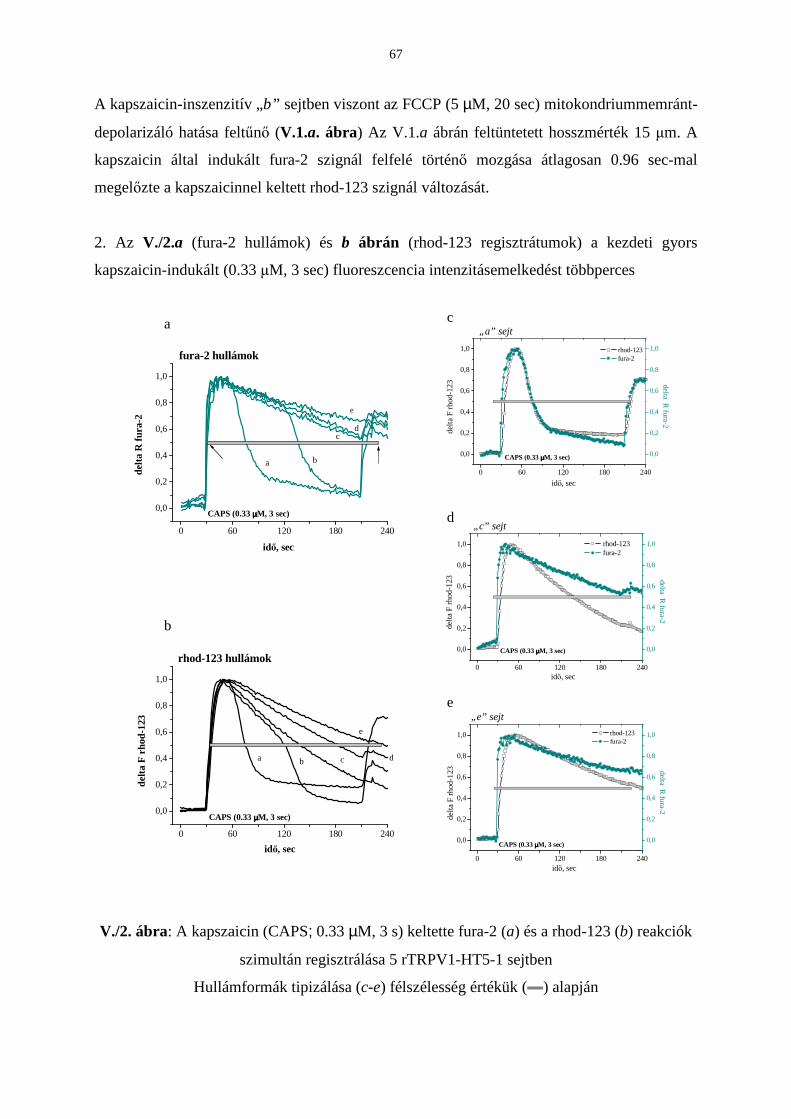
67
A kapszaicin-inszenzitív „b” sejtben viszont az FCCP (5 µM, 20 sec) mitokondriummemránt-
depolarizáló hatása feltőnı (V.1.a. ábra) Az V.1.a ábrán feltüntetett hosszmérték 15 µm. A
kapszaicin által indukált fura-2 szignál felfelé történı mozgása átlagosan 0.96 sec-mal
megelızte a kapszaicinnel keltett rhod-123 szignál változását.
2. Az V./2.a (fura-2 hullámok) és b ábrán (rhod-123 regisztrátumok) a kezdeti gyors
kapszaicin-indukált (0.33 µM, 3 sec) fluoreszcencia intenzitásemelkedést többperces
e
d
c
b
0 60 120 180 240
0,0
0,2
0,4
0,6
0,8
1,0
idı, sec
rhod-123 fura-2
delta
F r
hod
-123
CAPS (0.33 µµµµM, 3 sec)0,0
0,2
0,4
0,6
0,8
1,0
delta
R fura
-2
„e” sejt
0 60 120 180 240
0,0
0,2
0,4
0,6
0,8
1,0
„c” sejt
idı, sec
rhod-123 fura-2
delta
F r
hod
-123
CAPS (0.33 µµµµM, 3 sec) 0,0
0,2
0,4
0,6
0,8
1,0
delta
R fura
-2
0 60 120 180 240
0,0
0,2
0,4
0,6
0,8
1,0
„a” sejt
idı, sec
rhod-123 fura-2
delta
F r
hod
-12
3CAPS (0.33 µµµµM, 3 sec) 0,0
0,2
0,4
0,6
0,8
1,0
delta
R fura
-2
0 60 120 180 240
0,0
0,2
0,4
0,6
0,8
1,0
del
ta F
rho
d-12
3
idı, sec
rhod-123 hullámok
a b c d
e
CAPS (0.33 µµµµM, 3 sec)
0 60 120 180 240
0,0
0,2
0,4
0,6
0,8
1,0
del
ta R
fura
-2
CAPS (0.33 µµµµM, 3 sec)
idı, sec
fura-2 hullámok
a b
c
e
d
a
V./2. ábra: A kapszaicin (CAPS; 0.33 µM, 3 s) keltette fura-2 (a) és a rhod-123 (b) reakciók
szimultán regisztrálása 5 rTRPV1-HT5-1 sejtben
Hullámformák tipizálása (c-e) félszélesség értékük (▬) alapján

68
nagyságrendő visszatérési periódus követ mindkét jel esetén, melyek többnyire közel
párhuzamos lefutásúak az öt rTRPV1-HT5-1 sejt mindegyikében.
A két vizsgált szignál leszálló szárainak kinetikája azonban meglehetısen komplex és
nagymértékő variabilitást mutat sejtrıl sejtre. A leszálló szár kezdeti, csúcs utáni részét vagy
exponenciális vagy lineáris hanyatlás követi, néhány esetben pedig egy gyors, majd egy lassú
exponenciális komponens alakul ki.
A görbékre illesztett függvények összetevıi (meredekség, exponenciális változás, stb)
kvantitatív becslésre illetve a számszerő adatok feldolgozására alkalmasak. Görbeillesztésre
azonban a regisztrátumok csak egy töredékét lehet elég kicsi hibafaktorral használni. Ennek a
feldolgozásnak ismertetéséhez sok részletkérdésre ki kellene térnünk, melyektıl most
eltekintenénk.
A csúcsamplitúdók 50%-os szintjénél mért idıparaméterünk, a „félszélesség érték” szolgált az
adatok kvantitatív kiértékelésére, továbbá alkalmasnak bizonyult kvalitatív csoportosításukra
is (V./2. és 3.). Méréstechnikai okokból a félszélesség idıtartamát 200 másodpercre
korlátoztuk. Így e két paraméter alapján (a csúcsmagasság 50%-nál mért 200 sec-os idıkorlát)
ugyanis lehetségessé vált a hullámformák tranziens illetve elhúzódó jellegük szerinti
besorolása.
A tranziens jelzıvel illetett reakció arra utal, hogy a hullámforma 200 másodperc alatt 50%
alá visszatér. Mindkét szignál ebbe a típusba tartozik az „a” sejt esetén (V./2.c ábra).
Az elhúzódó leszálló szár jellegzetessége, hogy 200 sec alatt nem tér vissza a
csúcsamplitudójának 50%-os szintjére. Az elhúzódó leszálló szár reprezentatív formája az
V./2.e sejt mindkét tagja illetve a „c” sejt fura-2 jele (V./2.d). A „c” sejt reprezentatív példája
a reakciópárok közötti kinetikai variabilitásnak; míg az egyik vizsgált paraméter tranziens
természető, a másik elhúzódó jellegő.
A félszélesség adatok frekvencia eloszlása hisztogrammal ábrázolva kétcsúcsú eloszlást mutat
sejttípustól és a kapszaicin koncentrációjától függetlenül (V./3. ábra). A tranziens és az
elhúzódó reakciók százalékos megoszlása a két sejttípusban meglehetısen hasonló kis
kapszaicin koncentráció esetén (0.33 µM; V./3.).
A hisztogramm adatai alapján azonban TRG neuronokban a 3.3 µM kapszaicin
alkalmazásakor (n=54) az elhúzódó fura-2 reakciók aránya 7%-kal nıtt (24%-ról 31%-ra), az
elhúzódó rhod-123 reakciók aránya viszont 23%-kal csökkent (44-rıl 21%-ra) a 0.33 µM
kapszaicinnel kiváltott válaszokhoz képest (n=50).

69
50 100 150 2000
5
10
15
20
rTRPV1-HT5-1 sejtek, CAPS 3.3 µM (n=34)
fura-2 átlag=83.44SEM=10.25
félszélesség érték, sec
18%
20 40 60 80 100 120 140 160 180 200 2200
5
10
15
20S
ejts
zám
rhod-123
átlag=82.03 SEM=11,63
17%
20 40 60 80 100 120 140 160 180 200 2200
5
10
15
20
Sej
tszá
m
átlag=106.09SEM=9.96
fura-2
félszélességérték, sec
40%
20 40 60 80 100 120 140 160 180 200 2200
5
10
15
20
27%
átlag=113.34SEM=12.68
rhod-123
rTRPV1-HT5-1 sejtek, CAPS 0.33 µM (n=44)
0 20 40 60 80 100 120 140 160 180 200 2200
5
10
15
20
átlag=115.85 SEM=9.29
fura-2
félszélességérték, sec
21%
0 20 40 60 80 100 120 140 160 180 200 2200
5
10
15
20
31%
Sej
tszá
m
átlag=103.39 SEM=11.22
TRG sejtek, CAPS 3.3 µM (n=54)
rhod-123
20 40 60 80 100 120 140 160 180 200 2200
5
10
15
20
átlag=109.18 SEM=9.16
fura-2
44%Sej
tszá
m
20 40 60 80 100 120 140 160 180 200 2200
5
10
15
20
TRG sejtek, CAPS 0.33 µM (n=50)
félszélességérték, sec
átlag=123.22 SEM=13.16
rhod-123
24%
V./3. ábra: Az elsı kapszaicin applikációra (CAPS; 0.33 vagy 3.3 µM) megvalósuló kalcium-
válasz félszélesség értékeinek frekvencia megoszlása a két vizsgált sejtmodellben
Míg a rTRPV1-HT5-1 sejtekben nagyobb kapszaicin koncentrációnál (n=34) mindkét szignál
esetén szignifikáns csökkenés tapasztalható az elhúzódó hullámformák arányában (27%-ról
17%-re a fura-2-nél és 40-rıl 18%-ra a rhod-123-nál) a 0.33 µM kapszaicin hatáshoz
viszonyítva (n=44).
A 0.33 µM kapszaicin által kiváltott fura-2 és rhod-123 válaszok félszélesség értékei a két
sejtféleségben nem különböztek egymástól szignifikánsan: TRG neuronokban illetve
rTRPV1-HT5-1 sejtekben a fura-2 szignálok átlagos félszélesség értéke 109.18±9.16 valamint
106.09±9.96 sec. A rhod-123 szignálok átlagos félszélesség adata sorrendben 123.22±13.16
és 113.34±12.68 sec (V./3. ábra).

70
Azonban a nagy kapszaicin koncentráció (3.3 µM) által kiváltott fura-2 és rhod-123 reakciók
félszélesség értékei szignifikánsan eltértek egymástól a két sejttípusban: TRG neuronokban
115.85±9.29 sec, rTRPV1-HT5-1 sejtekben pedig 83.44±10.25 sec a fura-2 szignálok átlagos
félszélesség értéke (p<0.01). A rhod-123 szignálok átlagos félszélesség adata TRG-nél
103.39±11.22 sec és rTRPV1-HT5-1 sejtek esetén 82.03±11.63 sec (p<0.05;V./3. ábra).
3. TRG neuronokban a kapszaicin koncentrációdependens módon hatott: minél nagyobb a
koncentráció, annál nagyobb 1R és 1∆F maximális csúcsértékeket mértünk (2. táblázat). (Az
R jelölés a fura2 jel változását jelenti, a ∆F pedig a rhod123 fluoreszcencia változását jelöli. A
jelölésekben a bal oldali alsó index a kapszaicin applikáció sorszámát jelzi, míg a jobb oldali
alsó index a kapszaicin alkalmazott koncentrációjára utal.)
A 3.3 µM kapszaicin (n=15) majdnem háromszor nagyobb 1R-t (1R0.33 = 0.147±0.05 versus
1R3.3 = 0.37±0.07) és több mint kétszeres 1∆F-et produkált a 0.33 µM kapszaicinnel keltett
eredményekhez képest (1∆F0.33 = 172.7±53.8 vs 1∆F3.3 = 360.7±85.7; n=7).
2. táblázat
A két sejttípus összehasonlítása a ∆[Ca 2+] ic és a ∆Ψm válaszaik alapján 0.33
valamint 3.3 µM kapszaicin (CAPS) egyszeri adásának hatására
TRG rTRPV1-HT5-1
Kapszaicin
koncentrációja
(µM)
0.33
(n=7)
3.3
(n=15)
0.33
(n=15)
3.3
(n=12)
1R értékek 0.147±0.05 0.37±0.07 0.84±0.10 0.56±0.06
1∆∆∆∆F értékek 172.7±53.8 360.7±85.7 587.5±105 600±91.4
1CAPS/FCCP
(fura-2) 166% 518% ø ø
1CAPS/FCCP
(rhod-123) 26% 70% ø ø
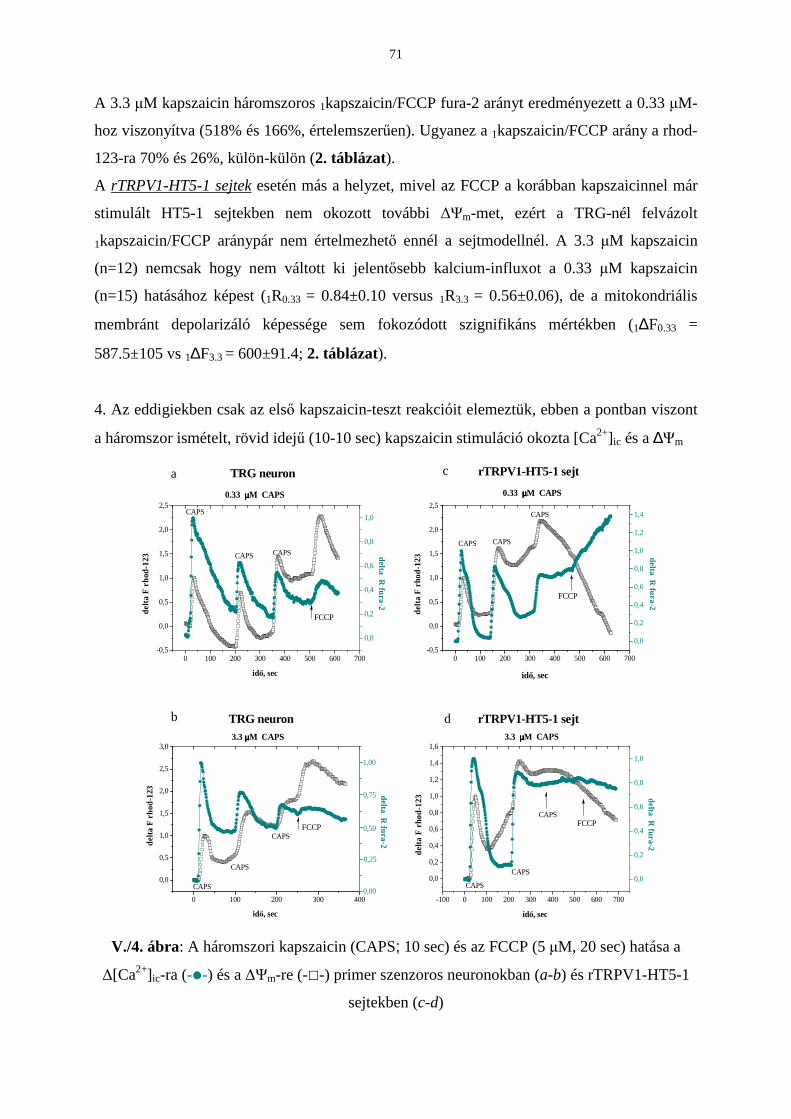
71
A 3.3 µM kapszaicin háromszoros 1kapszaicin/FCCP fura-2 arányt eredményezett a 0.33 µM-
hoz viszonyítva (518% és 166%, értelemszerően). Ugyanez a 1kapszaicin/FCCP arány a rhod-
123-ra 70% és 26%, külön-külön (2. táblázat).
A rTRPV1-HT5-1 sejtek esetén más a helyzet, mivel az FCCP a korábban kapszaicinnel már
stimulált HT5-1 sejtekben nem okozott további ∆Ψm-met, ezért a TRG-nél felvázolt
1kapszaicin/FCCP aránypár nem értelmezhetı ennél a sejtmodellnél. A 3.3 µM kapszaicin
(n=12) nemcsak hogy nem váltott ki jelentısebb kalcium-influxot a 0.33 µM kapszaicin
(n=15) hatásához képest (1R0.33 = 0.84±0.10 versus 1R3.3 = 0.56±0.06), de a mitokondriális
membránt depolarizáló képessége sem fokozódott szignifikáns mértékben (1∆F0.33 =
587.5±105 vs 1∆F3.3 = 600±91.4; 2. táblázat).
4. Az eddigiekben csak az elsı kapszaicin-teszt reakcióit elemeztük, ebben a pontban viszont
a háromszor ismételt, rövid idejő (10-10 sec) kapszaicin stimuláció okozta [Ca2+] ic és a ∆Ψm
d
c
b rTRPV1-HT5-1 sejtTRG neuron
0 100 200 300 400 500 600 700-0,5
0,0
0,5
1,0
1,5
2,0
2,5CAPS
CAPSCAPS
idı, sec
delta
F r
hod-
123
0.33 µµµµM CAPS
FCCP
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
delta R fura-2
-100 0 100 200 300 400 500 600 700
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
CAPS
CAPS
CAPS
idı, sec
delta
F r
hod-
123
0,0
0,2
0,4
0,6
0,8
1,0
delta R fura-2
3.3 µµµµM CAPS
FCCP
0 100 200 300 400
0,0
0,5
1,0
1,5
2,0
2,5
3,0
CAPS
CAPS
CAPS
idı, sec
delta
F r
hod-
123
0,00
0,50
0,75 delta R fura-2
3.3 µµµµM CAPS
FCCP
0,25
1,00
0 100 200 300 400 500 600 700-0,5
0,0
0,5
1,0
1,5
2,0
2,5
CAPSCAPS
idı, sec
delta
F r
hod-
123
0,0
0,2
0,4
0,6
0,8
1,0
delta R fura-2
0.33 µµµµM CAPS
CAPS
FCCP
TRG neuron rTRPV1-HT5-1 sejta
V./4. ábra: A háromszori kapszaicin (CAPS; 10 sec) és az FCCP (5 µM, 20 sec) hatása a
∆[Ca2+] ic-ra (-�-) és a ∆Ψm-re (-�-) primer szenzoros neuronokban (a-b) és rTRPV1-HT5-1
sejtekben (c-d)
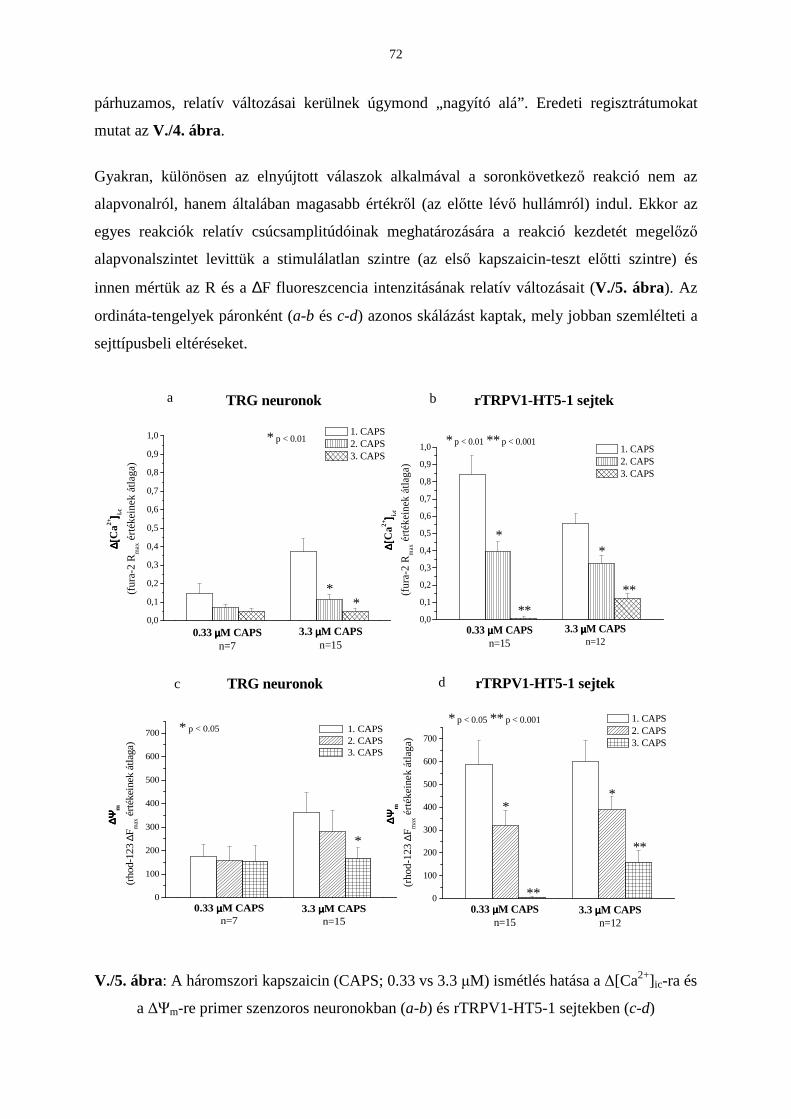
72
párhuzamos, relatív változásai kerülnek úgymond „nagyító alá”. Eredeti regisztrátumokat
mutat az V./4. ábra.
Gyakran, különösen az elnyújtott válaszok alkalmával a soronkövetkezı reakció nem az
alapvonalról, hanem általában magasabb értékrıl (az elıtte lévı hullámról) indul. Ekkor az
egyes reakciók relatív csúcsamplitúdóinak meghatározására a reakció kezdetét megelızı
alapvonalszintet levittük a stimulálatlan szintre (az elsı kapszaicin-teszt elıtti szintre) és
innen mértük az R és a ∆F fluoreszcencia intenzitásának relatív változásait (V./5. ábra). Az
ordináta-tengelyek páronként (a-b és c-d) azonos skálázást kaptak, mely jobban szemlélteti a
sejttípusbeli eltéréseket.
d
b
c
0
100
200
300
400
500
600
700
**
*
* p < 0.05 ** p < 0.001
**
*
0.33 µµµµM CAPSn=15
1. CAPS 2. CAPS 3. CAPS
3.3 µµµµM CAPSn=12
∆Ψ∆Ψ ∆Ψ∆Ψ
m
(rho
d-12
3 ∆F m
ax é
rték
eine
k át
laga
)
0
100
200
300
400
500
600
700
∆Ψ∆Ψ ∆Ψ∆Ψ
m
(rhod-
123 ∆
F max é
rtéke
inek
átla
ga) * p < 0.05 1. CAPS
2. CAPS 3. CAPS
*
0.33 µµµµM CAPSn=7
3.3 µµµµM CAPSn=15
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0
∆[∆[ ∆[∆[C
a2+]] ]] i.c
(fur
a-2
R max é
rté
kein
ek á
tlaga
)
*
**
**
0.33 µµµµM CAPS
n=153.3 µµµµM CAPS
n=12
1. CAPS 2. CAPS 3. CAPS
* p < 0.01 ** p < 0.001
*
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0 * p < 0.01
*
3.3 µµµµM CAPSn=15
1. CAPS 2. CAPS 3. CAPS
0.33 µµµµM CAPSn=7
*
∆[∆[ ∆[∆[C
a2+]] ]] i.c
(fur
a-2
R ma
x ért
ékei
nek
átla
ga)
TRG neuronok rTRPV1-HT5-1 sejtek
TRG neuronok rTRPV1-HT5-1 sejtek
a
V./5. ábra: A háromszori kapszaicin (CAPS; 0.33 vs 3.3 µM) ismétlés hatása a ∆[Ca2+] ic-ra és
a ∆Ψm-re primer szenzoros neuronokban (a-b) és rTRPV1-HT5-1 sejtekben (c-d)

73
TRG neuronokban a háromszor ismételt, rövid idejő (10-10 sec) kapszaicin stimuláció
(koncentrációtól függetlenül) szignifikánsan kisebb intracelluláris kalciumionszint növekedést
eredményezett (1R0.33=0.147±0.05, 2R0.33=0.07±0.017, 3R0.33=0.059±0.015; 1R3.3=0.37±0.069,
2R0.33=0.112±0.029, 3R0.33=0.048±0.014; V./5.a) a rTRPV1-HT5-1 sejtekben kapott
adatokhoz képest (1R0.33=0.84±0.11, 2R0.33=0.39±0.06, 3R0.33=0.009±0.007; 1R3.3=
0.557±0.06, 2R3.3=0.329±0.046, 3R3.3=0.121±0.03; V./5.b). (A jobb oldali alsó indexben jelölt
szám a kapszaicin koncentrációját, míg a bal oldali a kapszaicin applikáció sorszámát jelzi.)
A kis koncentrációjú kapszaicin (0.33 µM) alkalmazásakor a receptor deszenzitizáció hasonló
mértékő a két sejtmodellben, de csak a transzfektált sejtekben szignifikáns. Szenzoros
neuronban a deszenzitizáció mértéke a 3.3 µM kapszaicin esetén éri el a megjelölt
szignifikancia szintet, ekkor ugyanis a második válasz csupán 30.2%-a, a harmadik pedig
12.9%-a az elsınek. TRG neuronban a nanomoláris koncentrációban adott kapszaicin gyors és
reverzibilis intracelluláris kalciumszint növekedése az esetek döntı többségében nem társult
statisztikailag jelentıs mitokondriális depolarizációval (1∆F0.33=172.7±53.8,
2∆F0.33=159.7±56.01, 3∆F0.33=155.5±65.2; 1∆F3.3=360.7±85.7, 2∆F3.3=281.3±89.5,
3∆F3.3=167.3±46.9; V./5.c).
Azonban TRPV1-HT5-1 sejtekben a 0.33 µM kapszaicin elsı applikációja kétszer markánsabb
fura-2 és rhod-123 szignált produkált (V./5.b & d), mint TRG neuronokban a mikromolos
koncentrációjú kapszaicin és további membránpotenciál csökkenést már nem tapasztaltunk a
kísérlet végén alkalmazott FCCP adásakor (V./4.c & d és 2. táblázat). Meglepetésünkre a
rTRPV1-HT5-1 sejteknél a nagyobb koncentrációjú kapszaicin (3.3 µM) nem a várt
eredményre vezetett (vö. V./3. ábra), ugyanis kisebb kalciumakkumulációt produkált, amit
közel ugyanakkora ∆Ψm követett, mint 0.33 µM kapszaicin esetén (1∆F0.33=587.5±105.3,
2∆F0.33=322.06±62.3, 3∆F0.33=5.7±2.7; 1∆F3.3=600.6±91.4, 2∆F3.3=390.5±56.6,
3∆F3.3=160.08±52.66; V./5.d).
Következtetések
Tenyésztett TRG neuronokat és patkány TRPV1 kapszaicin receptorral transzfektált humán
fibroszarkóma sejteket fura-2-vel és rhod-123-mal festettünk meg, a relatív citoszolikus
szabad kalciumionszint ([Ca2+] ic) és a mitokondriális membránpotenciál (∆Ψm, deltapszi-m)
változásainak szimultán detektálása céljából.

74
Az elızı (3.4) fejezet egyik fontos következtetése a feltőnı hasonlóság a kétféle sejtmodell
válaszkészségében. Ezt a hasonlóságot tenyésztett TRG neuronokban és rTRPV1-HT5-1
sejtekben a relatív ∆[Ca2+] ic és a ∆Ψm szimultán mérése során is megtaláltuk. Azonos
viselkedésrıl azonban az alább részletezendı kisebb eltérések miatt, csak bizonyos
körülmények között beszélhetünk. A korábban taglalt ionszint változások méréseit
befolyásoló faktorokhoz (festékfelvétel és az expresszált receptor mennyiség) talán biológiai
szempontból sem elhanyagolható eltérés járul a mitokondriumok reakciókészsége, illetve
tágabb értelemben a kalcium homeosztázis regulációja vonatkozásában.
A két vizsgált fluoreszcens szignál idıparaméterei és csúcsamplitúdói alapján egy komplex
kapcsolat tételezhetı fel a ∆[Ca2+] ic és a ∆Ψm viszonyában. A reakciók leszálló szárának
tranziens illetıleg elhúzódó csoportba történı kategorizálása egyszerősíti az összehasonlítást
és ez a tulajdonság függetlennek tőnik a sejt természetétıl, az alkalmazott kapszaicin
koncentrációjától és a kapszaicin inzultusok sorszámától. A TRG neuronok és a TRPV1
kapszaicin receptort transzfektáló HT5-1 sejtek ebbıl a szempontból is – különösen a kis
koncentrációjú kapszaicin adásakor – hasonló módon reagáltak.
A kapszaicin (0.33 és 3.3 µM) a TRPV1 receptor aktivációján keresztül [Ca2+] ic emelkedést
eredményez, ami többnyire mérhetı, de nem mindig jelentıs mitokondriális membrán
depolarizációval társulhat. Kapszaicin adásakor a citoszolikus szabad kalcium
koncentrációjának növelése mindig megelızte a mitokondriális depolarizációt. A
nanomoláris koncentrációban adott kapszaicin ismételt expozíciója receptor
deszenzibilizációt okozott kifejezetebb mitokondriális funkcióvesztés jelei nélkül TRG
sejtekben. Szenzoros neuronok esetén a kapszaicin koncentrációjának növelésekor (3.3
µΜ) a kapszaicin receptort deszenzibilizáló, valamint mitokondriális membránt
depolarizáló képessége is kifejezetebb volt. Trigeminális neuronokban a 0.33 és 3.3 µΜ
kapszaicin az FCCP-vel kiváltott rhod-123 jelnek csupán 26 illetve 70%-át érte el. Ebben a
koncentrációtartományban tehát a TRG reakciók észlelhetı fokú agonista koncentrációfüggést
mutattak.
A rTRPV1-HT5-1 sejtekben a 0.33 µΜ kapszaicin elsı applikációja markáns és
komplett ∆Ψm esést produkált, melyet a kapszaicin koncentrációjának 3.3 µM -ra történı
emelése már nem tudott szignifikáns mértékben tovább fokozni, sıt a félszélesség értékek és a
csúcsamplitúdók adatai alapján gyanítható, hogy ez a koncentráció a transzfektált sejteken

75
már toxikus (a reakciókészség nyilvánvaló csökkenése értelmében). A rTRPV1-HT5-1
sejtekben már a kis koncentrációjú kapszaicin is legfeljebb kétszer keltett aránylag
reprodukálható válaszokat, az FCCP pedig – az esetek döntı hányadában – képtelen volt
további membránpotenciál csökkenést elıidézni. A fenti jelenségek lehetséges okai között
említendık a TRPV1 receptor overexpressziója, a sejt kalciumpufferoló kapacitásának
deficitje, más membrán tulajdonságok (plazmamembrán fluiditás) és az ebbıl adódó esetleges
festékszivárgás.
A kapszaicin által kiváltott mitokondriális károsodás nem direkt, közvetlen hatása a
mitokondriumokra, mivel sem A-típusú TRG neuronokban, sem pedig rTRPV1-negatív
HT1080 sejtekben nem tapasztaltunk mitokondriális membránpotenciál csökkenést
kapszaicin hatására.
A fentiekhez hasonló összehasonlító méréseket kapszaicin-érzékeny trigeminális neuronokon
és rTRPV1 receptort expresszáló HT1080 sejteken még nem végeztek. Közvetlen mérésekkel
igazoltuk az extracelluláris kalcium belépést a kapszaicinnel aktivált TRPV1 receptoron
keresztül a velejáró funkcionális mitokondriális membránpotenciál változással.

76
4. LEGFONTOSABB ÚJ EREDMÉNYEK ÖSSZEFOGLALÁSA
Eredményeink alapján megállapítható, hogy
1. az SB366791 ígéretes és megbízható TRPV1 receptor antagonistának és ezáltal egy
potenciális fájdalomcsillapító kifejlesztésére alkalmas molekulának bizonyult in vitro és in
vivo egyaránt, patkányban.
2. az OLDA kellı hatékonysággal aktiválta és deszenzibilizálta a TRPV1 receptort, hogy
endogén ligand szerepét kijelenthessük. Szubsztituált származékai (3- és 4-MOLDA)
ellentétes farmakológiai hatást gyakoroltak a TRPV1 receptorra, mely arra enged
következteni, hogy a vanilloid jellegő vegyületek aromás részének 4-es pozíciójában lévı
hidroxilcsoportja különösen fontos a receptor aktiválása szempontjából. Egy kémiai csoport
szubsztitúciója azt eredményezte, hogy a 3-MOLDA (az OLDA-nál ötször gyengébb) TRPV1
agonistának, a 4-MOLDA pedig receptor antagonistának bizonyult.
3. a TRPV1 receptor foszforiláltságának szintje valóban lényeges szerepet játszik az agonisták
aktiváló hatásában. Kísérleteinkkel bizonyítottuk, hogy mindkét típusú protein kináz (A és C)
stimulálása a nyugalmi szint fölé fokozott TRPV1 receptor aktivációt eredményez, míg a
TRPV1 receptor érzékenységének fenntartásában a protein kináz A alapaktivitását találtuk
meghatározó fontosságúnak trigeminális érzıneuronok sejttestjeiben és hátsógyöki szenzoros
neuronok perifériás végzıdéseiben egyaránt.
4. az általunk használt egyik sejttípus sem produkált kalcium-independens TRPV1-mediált
választ kalciummentes kísérleti körülmények között. Véleményünk szerint, az endoplazmás
retikulumban lévı TRPV1 receptorok mennyisége – az általunk alkalmazott sejtmodellekben
– nem jelentıs és csak olyan nagy koncentrációjú kapszaicinnel lenne aktiválható, mely már
nemcsak a receptoriális hatások kiváltására alkalmas.
5. összefüggést találtunk a kapszaicin koncentrációjának növelése (0.33 és 3.3 µM) és a
receptor deszenzitizáció mértéke között, valamint a koncentráció és az indukált mitokondriális
membránpotenciál-változás vonatkozásában TRG neuronokban. Ezek a korrelációk viszont
nem érvényesek maradéktalanul a transzfektált sejtekre. Közismert a kapszaicin TRPV1

77
receptort deszenzitizáció hatása, mely tartós ugyan, de reverzibilis. Észrevételeink alapján a
deszenzitizáció mechanizmusában a mitokondriális történések lényeges szerepet játszanak, de
a jelen kísérleteinkben trigeminális neuronokban a kapszaicin nem okozott olyan mértékő és
jellegő változást, melybıl a mitokondrium funkcionális károsodására lehetne következtetni.
6. a 3.2., a 3.4. és a 3.5. témakörök eredményei alapján a rTRPV1 receptort expresszáló
HT1080 sejtvonal a szenzoros neuronok kitőnı modelljének tekinthetı (bizonyos feltételek
mellett) és ezeken a transzfektált sejteken a kapszaicin TRPV1 funkcióra gyakorolt akut
illetve tartós hatásai in vitro megbízhatóan és eredményesen tanulmányozhatóak.
Folyamatban van intézetünkben a humán TRPV1 receptor ortológ HT1080 és CHO sejtekbe
történı klónozása, mely újabb perspektívákat teremt a TRPV1 funkciójában szereplı
mechanizmusok pontosabb, mélyebb megértéséhez.
7. a fenti eredmények módszereinek bevezetése a PTE Farmakológiai és Farmakoterápiai
Intézetébe megnyitotta a lehetıséget a TRPV1 receptoron ható gyógyszerek fejlesztéséhez. A
nagy hatékonyságú (high throughput screening, HTS) in vitro módszerek elindításához az
értekezés eredményei kitünı kiinduló- és támpontot jelentenek.

78
FELHASZNÁLT IRODALOM
Aley KO, Levine JD.: Role of protein kinase A in the maintenance of inflammatory pain. J Neurosci 19:2181-2186, 1999 Aley KO, Messing RO, Mochly-Rosen D, Levine JD.: Chronic hypersensitivity for inflammatory nociceptor sensitization mediated by the epsilon isozyme of protein kinase C. J Neurosci 20:4680-4685, 2000 Almási R, Pethı G, Bölcskei K, Szolcsányi J.: Effect of resiniferatoxin on the noxious heat threshold temperature in the rat: a novel heat allodynia model sensitive to analgesics. Br. J. Pharmacol. 139:49-58, 2003 Amadesi S, Nie J, Vergnolle N, Cottrell GS, Grady EF, Trevisani M, Manni C, Geppetti P, McRoberts JA, Ennes H, Davis JB, Mayer EA, Bunnett NW.: Protease-activated receptor 2 sensitizes the capsaicin receptor transient receptor potential vanilloid receptor 1 to induce hyperalgesia. J Neurosci 24:4300-4312, 2004 Amann R, Maggi CA.: Ruthenium red as a capsaicin antagonist. Life Sci. 49:849-856, 1991 review Appendino G, Szállási A.: Euphorbium: modern research on its active principle, resiniferatoxin, revives an ancient medicine. Life Sci 60:681-696, 1997 review Askwith CC, Benson CJ, Welsh MJ, Snyder PM.: DEG/ENaC ion channels involved in sensory transduction are modulated by cold temperature. Proc Natl Acad Sci U S A 98:6459-63, 2001 Bevan S, Hothi S, Hughes G, James IF, Rang HP, Shah K, Walpole CS, Yeats JC.: Kapszazepine: a competitive antagonist of the sensory neurone excitant capsaicin. Br J Pharmacol 107:544-552, 1992 Bhave G, Zhu W, Wang H, Brasier DJ, Oxford GS, Gereau RW 4th.: cAMP-dependent protein kinase regulates desensitization of the capsaicin receptor (VR1) by direct phosphorylation. Neuron 35:721-731, 2002 Bhave G, Hu HJ, Glauner KS, Zhu W, Wang H, Brasier DJ, Oxford GS, Gereau RW 4th.: Protein kinase C phosphorylation sensitizes but does not activate the capsaicin receptor transient receptor potential vanilloid 1 (TRPV1). Proc Natl Acad Sci USA 100:12480-12485, 2003 Bley KR and Malmberg AB.: TRPV1 agonist-based therapies: mechanism of action and clinical prospects. In: Malmberg AB and Bley KR (eds.), Turning the heat on pain: TRPV1 receptor in pain and inflammation. Birkhäuser Verlag. Basel, pp. 191-209, 2005 Bonnington JK, McNaughton PA.: Signalling pathways involved in the sensitisation of mouse nociceptive neurones by nerve growth factor. J Physiol 551:433-446, 2003 Budd SL, Nicholls DG.: A reevaluation of the role of mitochondria in neuronal Ca2+ homeostasis. J Neurochem 66:403-11, 1996a Budd SL, Nicholls DG.: Mitochondria, calcium regulation, and acute glutamate excitotoxicity in cultured cerebellar granule cells. J Neurochem 67:2282-2291, 1996b Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D.: The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 389:816-824, 1997 Caterina MJ, Leffler A, Malmberg AB, Martin WJ, Trafton J, Petersen-Zeitz KR, Koltzenburg M, Basbaum AI, Julius D.: Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 288:306-313, 2000 Caterina MJ.: Vanilloid receptors take a TRP beyond the sensory afferent. Pain 105:5-9, 2003 review Cesare P, McNaughton P.: A novel heat-activated current in nociceptive neurons and its sensitization by bradykinin. Proc Natl Acad Sci USA 93:15435-15439, 1996

79
Cesare P, Dekker LV, Sardini A, Parker PJ, McNaughton PA.: Specific involvement of PKC-epsilon in sensitization of the neuronal response to painful heat. Neuron 23:617-624, 1999 Chen CW, Lee ST, Wu WT, Fu WM, Ho FM, Lin WW.: Signal transduction for inhibition of inducible nitric oxide synthase and cyclooxygenase-2 induction by capsaicin and related analogs in macrophages. Br J Pharmacol 140:1077-1087, 2003 Cholewinski, A., Burgess, G.M. and Bevan, S.: The role of calcium in capsaicin-induced desensitization in rat cultured dorsal root ganglion neurons. Neuroscience 55:1015-1023, 1993 Chuang HH, Prescott ED, Kong H, Shields S, Jordt SE, Basbaum AI, Chao MV, Julius D.: Bradykinin and nerve growth factor release the capsaicin receptor from PtdIns(4,5)P2-mediated inhibition. Nature 411:957-62, 2001 Chu CJ, Huang SM, De Petrocellis L, Bisogno T, Ewing SA, Miller JD, Zipkin RE, Daddario N, Appendino G, Di Marzo V, Walker JM.: N-oleoildopamine, a novel endogenous capsaicin-like lipid that produces hyperalgesia. J Biol Chem 278:13633-13639, 2003 Clapham DE, Runnels LW, Strubing C.: The TRP ion channel family. Nat Rev Neurosci 2:387-396, 2001 Clapham DE, Montell C, Schultz G, Julius D; International Union of Pharmacology.: International Union of Pharmacology. XLIII. Compendium of voltage-gated ion channels: transient receptor potential channels. Pharmacol Rev 55:591-596, 2003 review Correll CC, Phelps PT, Anthes JC, Umland S, Greenfeder S.Cloning and pharmacological characterization of mouse TRPV1. Neurosci Lett 370:55-60, 2004 Davis JB, Gray J, Gunthorpe MJ, Hatcher JP, Davey PT, Overend P, Harries MH, Latcham J, Clapham C, Atkinson K, Hughes SA, Rance K, Grau E, Harper AJ, Pugh PL, Rogers DC, Bingham S, Randall A, Sheardown SA.: Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 405:183-187, 2000 Dedov VN, Roufogalis BD.: Mitochondrial calcium accumulation following activation of vanilloid (VR1) receptors by capsaicin in dorsal root ganglion neurons. Neuroscience 95:183-188, 2000 Dedov VN, Mandadi S, Armati PJ, Verkhratsky A.: Capsaicin-induced depolarisation of mitochondria in dorsal root ganglion neurons is enhanced by vanilloid receptors. Neuroscience 103:219-226, 2001 Delree P, Leprince P, Schoenen J, Moonen G.: Purification and culture of adult rat dorsal root ganglia neurons. J Neurosci Res 23:198-206, 1989 De Petrocellis L, Harrison S, Bisogno T, Tognetto M, Brandi I, Smith GD, Creminon C, Davis JB, Geppetti P, Di Marzo V.: The vanilloid receptor (VR1)-mediated effects of anandamide are potently enhanced by the cAMP-dependent protein kinase. J Neurochem 77:1660-1663, 2001 Docherty RJ, Yeats JC, Bevan S, Boddeke HW.: Inhibition of calcineurin inhibits the desensitization of capsaicin-evoked currents in cultured dorsal root ganglion neurones from adult rats. Pflugers Arch 431:828-37, 1996 Docherty RJ, Yeats JC, Piper AS.: Kapszazepine block of voltage-activated calcium channels in adult rat dorsal root ganglion neurones in culture. Br J Pharmacol 121: 1461-1467, 1997 Dray A, Forbes CA, Burgess GM.: Ruthenium red blocks the capsaicin-induced increase in intracellular calcium and activation of membrane currents in sensory neurones as well as the activation of peripheral nociceptors in vitro. Neurosci Lett 110:52-9, 1990 Duchen MR.: Ca2+-dependent changes in the mitochondrial energetics in single dissociated mouse sensory neurons. Biochem J 283:41-50, 1992 Duchen MR.: Mitochondria and Ca2+ in cell physiology and pathophysiology. Cell Calcium 28:339-48, 2000a review

80
Duchen MR.: Mitochondria and calcium: from cell signalling to cell death. J Physiol 529:57-68, 2000b review Emaus RK, Grunwald R, Lemasters JJ.: Rhodamine 123 as a probe of transmembrane potential in isolated rat-liver mitochondria: spectral and metabolic properties. Biochim Biophys Acta 850:436-448, 1986 Enan E, Matsumura F.: Specific inhibition of calcineurin by type II synthetic pyrethroid insecticides. Biochem Pharmacol 43:1777-84, 1992 Eun SY, Jung SJ, Park YK, Kwak J, Kim SJ, Kim J.: Effects of capsaicin on Ca2+-release from the intracellular Ca2+ stores in the dorsal root ganglion cells of adult rats. Biochem Biophys Res Commun 285:1114-1120, 2001 Fakata KL, Swanson SA, Vorce RL, Stemmer PM.: Pyrethroid insecticides as phosphatase inhibitors. Biochem Pharmacol 55:2017-2022, 1998 Ferrer-Montiel A, Garcia-Martinez C, Morenilla-Palao C, Garcia-Sanz N, Fernandez-Carvajal A, Fernandez-Ballester G, Planells-Cases R.: Molecular architecture of the vanilloid receptor. Insights for drug design. Eur J Biochem 271:1820-1826, 2004 review Fowler CJ, Jonsson KO, Andersson A, Juntunen J, Jarvinen T, Vandevoorde S, Lambert DM, Jerman JC, Smart D.: Inhibition of C6 glioma cell proliferation by anandamide, 1-arachidonoylglycerol, and by a water soluble phosphate ester of anandamide: variability in response and involvement of arachidonic acid. Biochem Pharmacol 66:757-67, 2003 García-Hirschfeld, J., Lopez-Briones, L.G., Belmonte, C. and Valdeolmillos, M.: Intracellular free calcium responses to protons and capsaicin in cultured trigeminal neurons. Neuroscience 67:235-243, 1995 Gavva NR, Klionsky L, Qu Y, Shi L, Tamir R, Edenson S, Zhang TJ, Viswanadhan VN, Toth A, Pearce LV, Vanderah TW, Porreca F, Blumberg PM, Lile J, Sun Y, Wild K, Louis JC, Treanor JJ.: Molecular determinants of vanilloid sensitivity in TRPV1. J Biol Chem 279:20283-20295, 2004 Gu Q, Kwong K, Lee LY.: Ca2+ transient evoked by chemical stimulation is enhanced by PGE2 in vagal sensory neurons: role of cAMP/PKA signaling pathway. J Neurophysiol 89:1985-1993, 2003 Gunthorpe MJ, Benham CD, Randall A, Davis JB.: The diversity in the vanilloid (TRPV) receptor family of ion channels. Trends Pharmacol Sci 23:183-191, 2002 review Gunthorpe MJ, Rami HK, Jerman JC, Smart D, Gill CH, Soffin EM, Luis Hannan S, Lappin SC, Egerton J, Smith GD, Worby A, Howett L, Owen D, Nasir S, Davies CH, Thompson M, Wyman PA, Randall AD, Davis JB.: Identification and characterisation of SB-366791, a potent and selective vanilloid receptor (VR1/TRPV1) antagonist. Neuropharmacology 46:133-49, 2004 Hajnoczky G, Csordas G, Madesh M, Pacher P.: The machinery of local Ca2+ signalling between sarco-endoplasmic reticulum and mitochondria. J Physiol 529:69-81, 2000 review Hayes P, Meadows HJ, Gunthorpe MJ, Harries MH, Duckworth DM, Cairns W, Harrison DC, Clarke CE, Ellington K, Prinjha RK, Barton AJ, Medhurst AD, Smith GD, Topp S, Murdock P, Sanger GJ, Terrett J, Jenkins O, Benham CD, Randall AD, Gloger IS, Davis JB.: Cloning and functional expression of a human orthologue of rat vanilloid receptor-1. Pain 88:205-215, 2000 Helyes Zs, Németh J, Thán M, Bölcskei K, Pintér E, Szolcsányi J.: Inhibitory effect of anandamide on resiniferatoxin-induced sensory neuropeptide release in vivo and neuropathic hyperalgesia in the rat. Life Sci 73:2345-2353, 2003 Holzer P.: TRPV1 and the gut: from a tasty receptor for a painful vanilloid to a key player in hyperalgesia. Eur J Pharmacol 500:231-241, 2004 review Hu HJ, Bhave G, Gereau RW 4th.: Prostaglandin and protein kinase A-dependent modulation of vanilloid receptor function by metabotropic glutamate receptor 5: potential mechanism for thermal hyperalgesia. J Neurosci 22:7444-7452, 2002

81
Huang SM, Bisogno T, Trevisani M, Al-Hayani A, De Petrocellis L, Fezza F, Tognetto M, Petros TJ, Krey JF, Chu CJ, Miller JD, Davies SN, Geppetti P, Walker JM, Di Marzo V.: An endogenous capsaicin-like substance with high potency at recombinant and native vanilloid VR1 receptors. Proc Natl Acad Sci U S A 99:8400-5, 2002 Hwang SW, Cho H, Kwak J, Lee SY, Kang CJ, Jung J, Cho S, Min KH, Suh YG, Kim D, Oh U.: Direct activation of capsaicin receptors by products of lipoxygenases: endogenous capsaicin-like substances. Proc Natl Acad Sci 97:6155-6160, 2000 Jakab B, Helyes Z, Varga A, Bolcskei K, Szabo A, Sandor K, Elekes K, Borzsei R, Keszthelyi D, Pinter E, Petho G, Németh J, Szolcsányi J.: Pharmacological characterization of the TRPV1 receptor antagonist JYL1421 (SC0030) in vitro and in vivo in the rat. Eur J Pharmacol 517:35-44, 2005 Jancsó-Gabor A, Szolcsányi J, Jancsó N.: Irreversible impairment of thermoregulation induced by capsaicin and similar pungent substances in rats and guinea-pigs. J Physiol 206:495-507, 1970 Jordt SE, Tominaga M, Julius D.: Acid potentiation of the capsaicin receptor determined by a key extracellular site. Proc Natl Acad Sci U S A 97:8134-8139, 2000 Jordt SE, Julius D.: Molecular basis for species-specific sensitivity to "hot" chili peppers. Cell 108:421-30, 2002 Jordt SE, McKemy DD, Julius D.: Lessons from peppers and peppermint: the molecular logic of thermosensation. Curr Opin Neurobiol 13:487-492, 2003 review Jung J, Shin JS, Lee SY, Hwang SW, Koo J, Cho H, Oh U.: Phosphorylation of vanilloid receptor 1 by Ca2+/calmodulin-dependent kinase II regulates its vanilloid binding. J Biol Chem 279:7048-7054, 2004 Khodorov B, Pinelis V, Storozhevykh T, Yuravichus A, Khaspekhov L.: Blockade of mitochondrial Ca2+ uptake by mitochondrial inhibitors amplifies the glutamate-induced calcium response in cultured cerebellar granule cells. FEBS Lett 458:162-166, 1999 Koplas PA, Rosenberg RL and Oxford, GS.: The role of calcium in the desensitization of capsaicin responses in rat dorsal root ganglion neurons. J Neurosci 17:3525-3537, 1997 Krieger C, Duchen MR.: Mitochondria, Ca2+ and neurodegenerative disease. Eur J Pharmacol 447:177-188, 2002 review Kwak J, Wang MH, Hwang SW, Kim TY, Lee SY, Oh U.: Intracellular ATP increases capsaicin-activated channel activity by interacting with nucleotide-binding domains. J Neurosci 20:8298-304, 2000 Lazar J, Szabo T, Kovacs L, Blumberg PM, Biro T.: Distinct features of recombinant rat vanilloid receptor-1 expressed in various expression systems. Cell Mol Life Sci 60:2228-2240, 2003 Lee YS, Lee JA, Jung J, Oh U, Kaang BK.: The cAMP-dependent kinase pathway does not sensitize the cloned vanilloid receptor type 1 expressed in xenopus oocytes or Aplysia neurons. Neurosci Lett 288:57-60, 2000 Liu L, Simon SA.: Capsazepine, a vanilloid receptor antagonist, inhibits nicotinic acetylcholine receptors in rat trigeminal ganglia. Neurosci Lett 228:29-32, 1997 Liu, L. and Simon, S.: The influence of removing extracellular Ca2+ in the tachyfilaxis responses to capsaicin, zingerone and olvanil in rat trigeminal ganglion neurons. Brain Res 809:246-262, 1998 Liu, M., Liu, M.C., Magoulas, C., Priestley, J.V. and Willmott, N.J.: Versatile regulation of cytosolic Ca2+ by vanilloid receptor I in rat dorsal root ganglion neurons. J Biol Chem 278:5462-5472, 2003 Liu B, Zhang C, Qin F.: Functional recovery from desensitization of vanilloid receptor TRPV1 requires resynthesis of phosphatidylinositol 4,5-bisphosphate. J Neurosci 25:4835-4843, 2005

82
Lopshire JC, Nicol GD.: The cAMP transduction cascade mediates the prostaglandin E2 enhancement of the capsaicin-elicited current in rat sensory neurons: whole-cell and single-channel studies. J Neurosci 18:6081-6092, 1998 Mandadi S, Numazaki M, Tominaga M, Bhat MB, Armati PJ, Roufogalis BD.: Activation of protein kinase C reverses capsaicin-induced calcium-dependent desensitization of TRPV1 ion channels. Cell Calcium 35:471-478, 2004 Marshall IC, Owen DE, Cripps TV, Davis JB, McNulty S, Smart D.: Activation of vanilloid receptor 1 by resiniferatoxin mobilizes calcium from inositol 1,4,5-trisphosphate-sensitive stores. Br J Pharmacol 138:172-176, 2003 McIntyre P, McLatchie LM, Chambers A., Phillips E, Clarke M, Savidge J, Toms C, Peacock M, Shah K, Winter J, Weerasakera N, Webb M, Rang HP, Bevan S, James IF.: Pharmacological differences between the human and rat vanilloid receptor 1 (VR1). Br J Pharmacol 132:1084-1094, 2001 McKemy DD.: How cold is it? TRPM8 and TRPA1 in the molecular logic of cold sensation. Mol Pain 1:16 2005 review Mezey E, Toth ZE, Cortright DN, Arzubi MK, Krause JE, Elde R, Guo A, Blumberg PM, Szállási A.: Distribution of mRNA for vanilloid receptor subtype 1 (VR1), and VR1-like immunoreactivity, in the central nervous system of the rat and human. Proc Natl Acad Sci U S A 97:3655-3660, 2000 Mohapatra DP, Nau C.: Desensitization of capsaicin-activated currents in the vanilloid receptor TRPV1 is decreased by the cyclic AMP-dependent protein kinase pathway. J Biol Chem 278:50080-50090, 2003 Mohapatra DP, Nau C.: Regulation of Ca2+-dependent desensitization in the vanilloid receptor TRPV1 by calcineurin and cAMP-dependent protein kinase. J Biol Chem 280:13424-13432, 2005 Montell C, Birnbaumer L, Flockerzi V, Bindels RJ, Bruford EA, Caterina MJ, Clapham DE, Harteneck C, Heller S, Julius D, Kojima I, Mori Y, Penner R, Prawitt D, Scharenberg AM, Schultz G, Shimizu N, Zhu MX.: A unified nomenclature for the superfamily of TRP cation channels. Mol Cell 9:229-31, 2002 Montell C.: Thermosensation: hot findings make TRPNs very cool. Curr Biol 13:476-478, 2003 review Németh J, Oroszi G, Than M, Helyes Zs, Pinter E, Farkas B, Szolcsányi J.: Substance P radioimmunoassay for quantitative characterization of sensory neurotransmitter release. Neurobiology 7:437-444, 1999 Németh J, Helyes Zs, Thán M, Jakab B, Pintér E, Szolcsányi J.: Concentration-dependent dual effect of anandamice on sensory neuropeptide release from isolated rat tracheae. Neurosci Lett 336:89-92 2003 Nicholls DG.: Intracellular calcium homeostasis. Br Med Bull 42:353-8, 1986 review Nicholls DG, Budd SL.: Mitochondria and neuronal survival. Physiol Rev 80:315-60, 2000 review Nicholls DG, Ward MW.: Mitochondrial membrane potential and neuronal glutamate excitotoxicity: mortality and millivolts. Trends Neurosci 23:166-74, 2000 review Numazaki M, Tominaga T, Toyooka H, Tominaga M.: Direct phosphorylation of capsaicin receptor VR1 by protein kinase Cepsilon and identification of two target serine residues. J Biol Chem 277:13375-13378, 2002 Numazaki M, Tominaga T, Takeuchi K, Murayama N, Toyooka H, Tominaga M.: Structural determinant of TRPV1 desensitization interacts with calmodulin. Proc Natl Acad Sci U S A 100:8002-8006, 2003 Oláh, Z., Szabo, T., Karai, L., Hough, C., Fields, R.D., Caudle, R.M., Blumberg, P.M. and Iadarola, M.J.: Ligand-induced dynamic membrane changes and cell deletion conferred by vanilloid receptor 1. J Biol Chem 276:11021-11030, 2001 Oláh Z, Karai L, Iadarola MJ.: Protein kinase C(alpha) is required for vanilloid receptor 1 activation. Evidence for multiple signaling pathways. J Biol Chem 277:35752-35759, 2002

83
Patapoutian A, Peier AM, Story GM, Viswanath V.: ThermoTRP channels and beyond: mechanisms of temperature sensation. Nat Rev Neurosci 4:529-39, 2003 review Phelps PT, Anthes JC, Correll CC.: Cloning and functional characterization of dog transient receptor potential vanilloid receptor-1 (TRPV1). Eur J Pharmacol 513:57-66. 2005 Premkumar LS, Ahern GP.: Induction of vanilloid receptor channel activity by protein kinase C. Nature 408:985-990, 2000 Pierau FK, Torrey P, Carpenter DO.: Mammalian cold receptor afferents: role of an electrogenic sodium pump in sensory transduction. Brain Res 73:156-60, 1974 Prescott ED, Julius D.: A modular PIP2 binding site as a determinant of capsaicin receptor sensitivity. Science 300:1284-1288, 2003 Rami HK, Thompson M, Wyman P, Jerman JC, Egerton J, Brough S, Stevens AJ, Randall AD, Smart D, Gunthorpe MJ, Davis JB.: Discovery of small molecule antagonists of TRPV1. Bioorg Med Chem Lett 14:3631-3634, 2004 Rathee PK, Distler C, Obreja O, Neuhuber W, Wang GK, Wang SY, Nau C, Kress M.: PKA/AKAP/VR-1 module: A common link of Gs-mediated signaling to thermal hyperalgesia. J Neurosci 22:4740-4745, 2002 Rizzuto R, Bernardi P, Pozzan T.: Mitochondria as all-round players of the calcium game. J Physiol 529:37-47, 2000 review Sándor Z, and Szállási Á.: Vanilloid receptor-mediated hyperalgesia and desensitization. In: Malmberg AB and Bley KR (eds.), Turning the heat on pain: TRPV1 receptor in pain and inflammation. Birkhäuser Verlag. Basel, pp. 95-115, 2005 Savidge JR, Ranasinghe SP, Rang HP.: Comparison of intracellular calcium signals evoked by heat and capsaicin in cultured rat dorsal root ganglion neurons and in a cell line expressing the rat vanilloid receptor, VR1. Neuroscience 102: 177-184, 2001 Savidge J, Davis C, Shah K, Colley S, Phillips E, Ranasinghe S, Winter J, Kotsonis P, Rang H, McIntyre P.: Cloning and functional characterization of the guinea pig vanilloid receptor 1. Neuropharmacology 43:450-456, 2002 Seabrook GR, Sutton KG, Jarolimek W, Hollingworth GJ, Teague S, Webb J, Clark N, Boyce S, Kerby J, Ali Z, Chou M, Middleton R, Kaczorowski G, Jones AB.: Functional properties of the high-affinity TRPV1 (VR1) vanilloid receptor antagonist (4-hydroxy-5-iodo-3-methoxyphenylacetate ester) iodo-resiniferatoxin. J Pharmacol Exp Ther 303:1052-1060, 2002 Shin J, Cho H, Hwang SW, Jung J, Shin CY, Lee SY, Kim SH, Lee MG, Choi YH, Kim J, Haber NA, Reichling DB, Khasar S, Levine JD, Oh U.: Bradykinin-12-lipoxygenase-VR1 signaling pathway for inflammatory hyperalgesia. Proc Natl Acad Sci U S A 99:10150-5, 2002 Shu X, Mendell LM.: Acute sensitization by NGF of the response of small-diameter sensory neurons to capsaicin. J Neurophysiol 86:2931-2938, 2001 Snider WD and McMahon SB.: Tackling pain at the source: new ideas about nociceptors. Neuron 20:629-632, 1998 Souslova V, Cesare P, Ding Y, Akopian AN, Stanfa L, Suzuki R, Carpenter K, Dickenson A, Boyce S, Hill R, Nebenuis-Oosthuizen D, Smith AJ, Kidd EJ, Wood JN.: Warm-coding deficits and aberrant inflammatory pain in mice lacking P2X3 receptors. Nature 407:1015-7, 2000 Southall MD, Li T, Gharibova LS, Pei Y, Nicol GD, Travers JB.: Activation of epidermal vanilloid receptor-1 induces release of proinflammatory mediators in human keratinocytes. J Pharmacol Exp Ther 304:217-222, 2003

84
Sugiura T, Tominaga M, Katsuya H, Mizumura K.: Bradykinin lowers the threshold temperature for heat activation of vanilloid receptor 1. J Neurophysiol 88:544-548, 2002 Suh YG, Lee YS, Min KH, Park OH, Seung HS, Kim HD, Park HG, Choi J, Lee J, Kang SW, Oh UT, Koo JY, Joo YH, Kim SY, Kim JK, Park YH.Novel non-vanilloid VR1 antagonist of high analgesic effects and its structural requirement for VR1 antagonistic effects. Bioorg Med Chem Lett 13:4389-93, 2003 Szállási A, Conte B, Goso C, Blumberg PM, Manzini S.: Characterization of a peripheral vanilloid (capsaicin) receptor in the urinary bladder of the rat. Life Sci 52:221-226, 1993 Szállási A, Blumberg PM.: Vanilloid (Capsaicin) receptors and mechanisms. Pharmacol Rev 51:159-212, 1999 review Szoke E, Balla Z, Csernoch L, Czeh G, Szolcsányi J.: Interacting effects of capsaicin and anandamide on intracellular calcium in sensory neurones. Neuroreport 11:1949-1952, 2000 Szoke E, Seress L, Szolcsányi J.: Neonatal capsaicin treatment results in prolonged mitochondrial damage and delayed cell death of B cells in the rat trigeminal ganglia. Neuroscience 113:925-37, 2002a Szoke E, Czeh G, Szolcsányi J, Seress L.: Neonatal anandamide treatment results in prolonged mitochondrial damage in the vanilloid receptor type 1-immunoreactive B-type neurons of the rat trigeminal ganglion. Neuroscience 115:805-814, 2002b Szolcsányi J, Joo F, Jancsó-Gabor A.: Mitochondrial changes in preoptic neurons after capsaicin desensitization of the hypothalamic thermodetectors in rats. Nature 229:116-117, 1971 Szolcsányi J, Jancsó-Gábor A.: Sensory effect of capsaicin congeners. I. Relationship between chemical structure and pain-producing potency. Arzneim Forsch Drug Res 25:1877-1881, 1975 Szolcsányi J, Jancsó-Gabor A, Joo F.: Functional and fine structural characteristics of the sensory neuron blocking effect of capsaicin. Naunyn Schmiedebergs Arch Pharmacol 287:157-169, 1975 Szolcsányi J.: Actions of capsaicin on sensory receptors. In: Wood JN (ed.), Capsaicin in the study of pain. Academic Press London, pp. 1-26, 1993 Szolcsányi J.: Anandamide and the question of its functional role for activation of capsaicin receptors. Trends in Pharmacol Sci 21: 203-204 2000 Szolcsányi J.: Capsaicin receptors as target molecules on nociceptors for development of novel analgesics. In: Keri Gy, Toth I (eds.), Molecular Pathomechanisms and New Targets in Drug Research, Taylor and Francis, London and New York, pp. 319-333, 2002 Szolcsányi J.: Forty years in capsaicin research for sensory pharmacology and physiology. Neuropeptides 38:377-84, 2004 review Szolcsányi J.: Hot peppers, pain and analgesics. In: Malmberg AB and Bley KR (eds.), Turning the heat on pain: TRPV1 receptor in pain and inflammation. Birkhäuser Verlag. Basel, pp. 3-22, 2005 Tominaga M, Caterina MJ, Malmberg AB, Rosen TA, Gilbert H, Skinner K, Raumann BE, Basbaum AI, Julius D.: The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 21:531-543, 1998 Tominaga M, Wada M, Masu M.: Potentiation of capsaicin receptor activity by metabotropic ATP receptors as a possible mechanism for ATP-evoked pain and hyperalgesia. Proc Natl Acad Sci USA 98:6951-6956, 2001 Vellani V, Mapplebeck S, Moriondo A, Davis JB, McNaughton PA.: Protein kinase C activation potentiates gating of the vanilloid receptor VR1 by capsaicin, protons, heat and anandamide. J Physiol 534:813-825, 2001 Vellani V, Zachrisson O, McNaughton PA.: Functional bradykinin B1 receptors are expressed in nociceptive neurones and are upregulated by the neurotrophin GDNF. J Physiol 560:391-401, 2004

85
Vergun O, Keelan J, Khodorov BI, Duchen MR.: Glutamate-induced mitochondrial depolarisation and perturbation of calcium homeostasis in cultured rat hippocampal neurones. J Physiol 519:451-66, 1999 Vlachova V, Teisinger J, Susankova K, Lyfenko A, Ettrich R, Vyklicky L.: Functional role of C-terminal cytoplasmic tail of rat vanilloid receptor 1. J Neurosci 23:1340-1350, 2003 Vulcu SD, Rupp J, Wiwie C, Gillen C, Jostock R, Nawrath H.: The cAMP pathway sensitizes VR1 expressed in oocytes from Xenopus laevis and in CHO cells. Pharmacology 69:38-43, 2003 Wahl P, Foged C, Tullin S, Thomsen C.: Iodo-resiniferatoxin, a new potent vanilloid receptor antagonist. Mol Pharmacol 59:9-15, 2001 Walpole CS, Bevan S, Bovermann G, Boelsterli JJ, Breckenridge R, Davies JW, Hughes GA, James I, Oberer L, Winter J, et al.: The discovery of kapszazepine, the first competitive antagonist of the sensory neuron excitants capsaicin and resiniferatoxin. J Med Chem 37:1942-1954, 1994 Wang Y, Szabo T, Welter JD, Toth A, Tran R, Lee J, Kang SU, Suh YG, Blumberg PM, Lee J.: High affinity antagonists of the vanilloid receptor. Mol Pharmacol 62:947-56, 2002 Werth JL, Thayer SA.: Mitochondria buffer physiological calcium loads in cultured rat dorsal root ganglion neurons. J Neurosci 14:348-56, 1994 Wisnoskey BJ, Sinkins WG and Schilling WP.: Activation of vanilloid receptor type I in the endoplasmatic reticulum fails to activate store-operated Ca2+ entry. Biochem J 372:517-528, 2003 White RJ, and Reynolds IJ.: Mitochondria and Na+/Ca2+ exchange buffer glutamate-induced calcium loads in cultured cortical neurons. J. Neurosci 15:1318-1328, 1995 Wood JN, Winter J, James IF, Rang HP, Yeats J, Bevan S.: Capsaicin-induced ion fluxes in dorsal root ganglion cells in culture. J Neurosci 8:3208-3220, 1988 Zhou Y, Zhou ZS, Zhao, ZQ.: PKC regulates capsaicin-induced currents of dorsal root ganglion neurons in rats. Neuropharmacology 41:601-608, 2001 Zygmunt PM, Petersson J, Andersson PA, Chuang HH, Sorgard M, DiMarzo V, Julius D and Högestätt ED.: Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 400:452-457, 1999

86
KÖSZÖNETNYILVÁNÍTÁS
Mindenekelıtt Dr. Barthó Loránd Professzor Úrnak szeretném hálámat kifejezni, hogy
lehetıvé tette számomra, hogy intézetében végezhessem Ph.D tanulmányaimat és Dr.
Szolcsányi János akadémikusnak, a Neurofarmakológia Program vezetıjének, amiért
ösztönzött munkám elvégzésében.
Köszönettel tartozom témavezetıimnek Dr. Czéh Gábor Professzor Úrnak és Dr. Sándor
Zoltánnak munkám során nyújtott szakmai irányításukért és baráti támogatásukért.
Szerzıtársaimnak, Dr. Helyes Zsuzsannának és Dr. Pethı Gábornak a PTE Farmakológiai és
Farmakoterápiai Intézet munkatársainak ıszintén köszönöm hasznos tanácsaikat és segítı
pártfogásukat.
A patkány trigeminális ganglionsejt tenyészet készítésének, valamint az intracelluláris
kalciumionszint-változás technikájának alapos elsajátítását Dr. Szıke Évának köszönhetem. A
kettıs fluoreszcens festés szimultán méréstechnikájának sikeres kivitelezésében Dr. Balla
Zsolt egykori kollégám tevékenyen közremőködött. Külön köszönet illeti Dr. Bölcskei Katát,
aki számos felmerülı probléma megoldásában nélkülözhetetlen segítségemre volt.
Szeretném köszönetemet kifejezni közvetlen munkatársaimnak, Buzásiné Annának és Disztl
Cecíliának, hogy az elmúlt 4 év során kifogástalan asszisztenciájukkal járultak hozzá munkám
kiteljesedéséhez. Hálás vagyok az intézet valamennyi dolgozójának, hogy bíztosították
számomra a nyugodt, oldott légkört, melyben könnyebben haladt a munka.
Végül, de nem utolsó sorban hálával tartozom páromnak és családomnak, türelmükért,
megértésükért, amiért végig mellettem álltak minden nehézségben és gondoskodó
szeretetükkel biztosították tanulmányaimat.

87
AZ ÉRTEKEZÉS ALAPJÁUL SZOLGÁLÓ KÖZLEMÉNYEK JEGYZÉKE Közlemények
Angelika Varga, József Németh, Árpád Szabó, Jason J. McDougall, Chunfen Zhang, Krisztián Elekes, Erika Pintér, János Szolcsányi, Zsuzsanna Helyes: Effects of the novel TRPV1 receptor antagonist SB366791 in vitro and in vivo in the rat Neurosci. Lett. 385, pp. 137-142, 2005 (IF: 2.019) Angelika Varga, Kata Bölcskei, Éva Szıke, Róbert Almási, Gábor Czéh, János Szolcsányi, Gábor Pethı:: Relative roles of protein kinase A and protein kinase C in modulation of TRPV1 receptor responsiveness in rat sensory neurons in vitro and peripheral nociceptors in vivo Neuroscience 140, pp. 645-657, 2006 ( IF: 3.456) J. Szolcsányi, Z. Sándor, G. Pethı, A. Varga, K. Bölcskei, R. Almási, Z. Riedl, G. Hajos, G. Czéh: Direct evidence for activation and desensitization of the capsaicin receptor by N-oleoildopamine on TRPV1-transfected cell, line in gene deleted mice and in the rat Neurosci. Lett. 361, pp. 155-158, 2004 (IF: 2.019/2=1.0095) Zoltán Sándor, Angelika Varga, Péter Horváth, Barbara Nagy, Zoltán Szilvássy, János Szolcsányi.: Construction and analysis of a new stable cell line expressing the rat TRPV1 receptor Cellular and Molecular Biology Letters 10, pp. 499-514, 2005 (IF: 0.495) Angelika Varga, Zoltán Sándor, Zsolt Balla, Gábor Czéh, János Szolcsányi.: Relative changes of intracellular calcium level and mitochondrium membrane potential simultaneously induced by capsaicin (közlésre elıkészített kézirat) Hazai angol nyelvő folyóiratban megjelent idézhetı elıadáskivonatok Varga A., Szıke É., Balla Zs., Czéh G., Szolcsányi J.: Parallel measurement of rhodamine 123 and fura-2 in capsaicin-stimulated sensory neurones Clinical Neuroscience 56 (2), pp. 93-94, 2003 Varga A., Bölcskei K., Sándor Z., Almási R., Pethı G., Czéh G., Riedl Zs., Hajós G., Szolcsányi J.: Studying of OLDA as endogenous ligand for TRPV1 capsaicin receptor in vitro and in vivo Acta Physiologica Hungarica 91 (3-4), pp. 372, 2004 Czéh G., Varga A., Sándor Zoltán., Szolcsányi János.: Calcium, mitokondrium, capsaicin Acta Physiologica Hungarica 91 (3-4), pp. 284-285. 2004 K. Sándor, Zs. Helyes, A. Varga, Á. Szabó, K. Bölcskei, J. Németh, K. Elekes, G. Pozsgai, E. Pintér, J.J. McDougall, C. Zhang, G. Pethı and J. Szolcsányi: Examination of novel TRPV1 receptor antagonists in vitro and in vivo in the rat Inflammopharmacology 2005, közlésre elfogadva

88
A. Varga, K. Bölcskei, Z. Sándor, R. Almási, G. Pethı, Zs. Riedl, Gy. Hajós, G. Czéh, J. Szolcsányi.: Effects of the substituted N-oleoylphenylethylamines on activation and desensitization of the TRPV1 capsaicin receptor Inflammopharmacology 2005, közlésre elfogadva Z. Sándor, A. Varga, K. Bölcskei, P. Horváth., Zsuzsanna Riedl, György Hajós, J. Szolcsányi.: Activation and desensitization of the TRPV1 capsaicin receptor by substituted N-oleoylphenylethylamines Clinical Neuroscience 2005, közlésre elfogadva Az értekezés alapját képezı elıadások Varga A., Disztl Cecília., Czéh G., Szolcsányi J.: A VR1 receptor foszforiláltságának szerepe a capsaicin aktiváló hatásában Pécsi Akadémiai Bizottság Sejtbiológiai PhD konferencia, 2002, Pécs Varga A., Disztl Cecília., Sándor Z., Czéh G., Szolcsányi J.: Az intracelluláris kalciumszint és a mitokondrium membránpotenciál párhuzamos mérése capsaicin-szenzitív sejtekben Richter Gedeon Rt, Kisfaludy Lajos Alapítvány tudományos elıadóülése, 2004, Budapest Varga A., Sándor Z., Disztl Cecília., Czéh G., Szolcsányi J.: A kalcium homeosztázis vizsgálata TRPV1 capsaicin receptort expresszáló sejtekben XII. Sejt- és fejlıdésbiológiai napok, 2004, Pécs Czéh G., Varga A., Sándor Zoltán., Szolcsányi János.: Kalcium, capsaicin, mitokondrium Magyar Élettani Társaság LXVIII. Vándorgyûlése 2004, Debrecen G. Czéh., A. Varga., Z. Sándor., J. Szolcsányi.: Calcium, mitochondria, toxicity IBRO Advanced School of Neuroscience 2004, Jalta, Ukrajna A. Varga, K. Bölcskei, G. Pethı, Z. Sándor, G. Czéh, R. Almási, Z. Riedl, Gy. Hajós, J. Szolcsányi: Evidence that N-oleoildopamine is an endogenous ligand for TRPV1 capsaicin receptor IBRO Advanced School of Neuroscience 2004, Jalta, Ukrajna Varga Angelika, Bölcskei Kata, Sándor Z., Almási R., Pethı G., Riedl Zs., Hajós Gy.,Czéh G., Szolcsányi J. Az N-oleoilfeniletilaminok szerepe a TRPV1 capsaicin receptor aktiválásában és deszenzitizálásában Neurobiológus Doktoranduszok Fóruma, 2005, Pécs Zoltán Sándor, Angelika Varga, K. Kvell and J. Szolcsányi.: Construction of cell lines expressing the TRPV1 receptor Magyar experimentális farmakológia tavaszi szimpóziuma, 2005, Budapest Z. Helyes, A. Varga, J. Németh, Á. Szabó, Hungary; J.J. McDougall, C. Zhang, Canada; K.Elekes, E. Pintér and J. Szolcsányi.: Effects of the novel TRPV1 receptor antagonist SB366791 in vitro and in vivo in the rat Summer Neuropeptide 2005 Conference, Miami Beach, Florida, USA

89
Az értekezés alapját képezı poszterek A. Varga, É. Szıke, G. Czéh, J. Szolcsányi: Effect of protein kinase inhibitors of capsaicin-evoked desensitization IBRO International Workshop on Signalling Mechanisms in the Central and Peripheral Nervous System, 2002, Debrecen Varga A., Disztl Cecília., Czéh G., Szolcsányi J.: Proteinkináz modulátorok hatása a VR-1 receptor aktiválással keltett kalcium-tranziensekre Magyar Kísérletes és Klinikai Farmakológiai Társaság V. Kongresszusa, 2002, Debrecen Varga A., Szıke É., Balla Zsolt., Czéh G., Szolcsányi J.: A rhodamin 123 és a fura-2 párhuzamos mérése capsaicin-szenzitív szenzoros neuronokban Magyar Idegtudományi Társaság (MITT) IX. Konferenciája, 2003, Balatonfüred J. Szolcsányi., L. Seress., A. Varga., Z. Balla., Z. Sándor.,G. Czeh.: Changes of mitochondrial membrane potential associated with rise of cytosolic free calcium in capsaicin-sensitive sensory neurons Society for Neuroscience, 33rd annual meeting, 2003, New Orleans, Luisiana, USA Varga A., Bölcskei K.,Disztl Cecília., Sándor Z., Almási R., Pethı G., Czéh G., Riedl Zs., Hajós G., Szolcsányi J.: Az N-oleoildopamine szerepe a TRPV1 capsaicin receptor aktiválásában és deszenzibilizálásában Magyar Klinikai Farmakológusok V. Kongresszusa és MFT Immunfarmakológiai Szekció Ülése, 2003, Debrecen A. Varga, Z. Sándor, G. Czéh, J. Szolcsányi.: Changes of mitochondrial membrane potential associated with rise of cytosolic free calcium in capsaicin-sensitive sensory neurons IBRO International Workshop on Neuronal Circuits: from Elementary to Complex Functions, 2004, Budapest K. Bölcskei, A. Varga, R Almási., G Pethı., Z. Sándor, G Czéh., Zs Riedl., G Hajós., J Szolcsányi.: Activation and desensitization of the TRPV1 capsaicin receptor by N-oleoildopamine IBRO International Workshop on Neuronal Circuits: from Elementary to Complex Functions, 2004, Budapest Sándor Z., Varga A., Szolcsányi J.: Comparison of TRPV1 receptor activation and desensitization in TRG neurons and transfected HT1080 cells IBRO International Workshop on Neuronal Circuits: from Elementary to Complex Functions, 2004, Budapest Varga A., Bölcskei K., Sándor Z., Almási R., Pethı G., Czéh G., Riedl Zs., Hajós G., Szolcsányi J.: Az OLDA TRPV1 endogén ligand szerepének in vitro és in vivo vizsgálata Magyar Élettani Társaság LXVIII. Vándorgyûlése 2004, Debrecen Horváth P., Sándor Z., Varga A., Szıke É., Seress L., Szolcsányi J.: Capsaicin kiváltotta mitokondriális károsodás vizsgálata patkány trigeminusz szenzoros neuronban és TRPV1 receptort expresszáló stabil sejtvonalban

90
Magyar Élettani Társaság LXVIII. Vándorgyûlése 2004, Debrecen Z. Sándor., P. Horváth., A. Varga., J. Szolcsányi.: Construction of a new stable cell line expressing the TRPV1 receptor to study longterm receptor activation and desensitization EPHAR conference (Federation of European Pharmacological Societies) 2004, Porto, Portugália P. Horváth, Z. Sándor, A. Varga, É. Szıke, L. Seress and J. Szolcsányi: Analysis of capsaicin induced mitochondrial damage in rat trigeminal sensory neurons and in a stable cell line expressing the TRPV1 receptor IBRO Summer school 2004, Moszkva, Oroszország A. Varga, K. Bölcskei, G. Pethı, Z. Sándor, G. Czéh, R. Almási, Z. Riedl, Gy. Hajós, J. Szolcsányi: Evidence that N-oleoildopamine is an endogenous ligand for TRPV1 capsaicin receptor IBRO Advanced School of Neuroscience 2004, Jalta, Ukrajna Z. Sándor, K. Bölcskei, A. Varga, R. Almási, G. Pethı, G. Czéh, Z. Riedl, Gy. Hajós, J. Szolcsányi: Activation and desensitization of the TRPV1 capsaicin receptor by substituted N-oleoylphenylethylamines Society for Neuroscience, 34rd annual meeting of Society of Neuroscience, 2004, San Diego, USA Z. Sándor, A. Varga, K. Bölcskei, P. Horváth., Zsuzsanna Riedl, György Hajós, J. Szolcsányi: Activation and desensitization of the TRPV1 capsaicin receptor by substituted N-oleoylphenylethylamines Magyar Idegtudományi Társaság (MITT) XI. Kongresszusa, 2005, Pécs A. Varga, K. Bölcskei, Z. Sándor, R. Almási, G. Pethı, Zs. Riedl, Gy. Hajós, G. Czéh, J. Szolcsányi: Effects of the substituted N-oleoylphenylethylamines on activation and desensitization of the TRPV1 capsaicin receptor Magyar experimentális farmakológia tavaszi szimpóziuma, 2005, Budapest K. Sándor, Zs. Helyes, A. Varga, Á. Szabó, K. Bölcskei, J. Németh, K. Elekes, G. Pozsgai, E. Pintér, J.J. McDougall, C. Zhang, G. Pethı and J. Szolcsányi: Examination of novel TRPV1 receptor antagonists in vitro and in vivo in the rat Magyar experimentális farmakológia tavaszi szimpóziuma, 2005, Budapest AZ ÉRTEKEZÉSBEN FEL NEM HASZNÁLT TUDOMÁNYOS MUNKÁK JEGYZÉKE Közlemények Balázs Jakab, Zsuzsanna Helyes, Kata Bölcskei, Angelika Varga, Árpád Szabó, Katalin Sándor, Krisztián Elekes, Rita Börzsei, Erika Pintér, József Németh, János Szolcsányi: Examination of the novel TRPV1 receptor antagonist JYL1421 (SC0030) in vitro and in vivo in the rat Eur J Pharmacol. 517, pp. 35-44. 2005 (IF: 2.432)

91
R. Gesztelyi, J. Zsuga, A. Cseppento, A. Bajza, A. Varga, JZ. Szabo, AJ. Szentmiklosi: Special sensitization pattern in adenisine-induced myocardial responses after thyroxine-treatment J. Pharmacol. Sci. 91, pp. 295-304, 2003 (IF: 1.419) Idézhetı kivonatok Horváth P., Sándor Z., Varga A., Szıke É., Seress L., Szolcsányi J.: Analysis of capsaicin induced mitochondrial damage in rat trigeminal sensory neurons and in a stable cell line expressing the TRPV1 receptor Acta Physiologica Hungarica 91, pp. 306-307, 2004 Németh J., Jakab B., Helyes Zs., Varga A., Bölcskei K., Szabó Á., Sándor K., Elekes K., Börzsei K., Pintér E., Pethı G., Szolcsányi J.: Examination of the novel TRPV1 receptor antagonist JYL1421 (SC0030) in vitro and in vivo in the rat Clinical Neuroscience 2005, közlésre elfogadva Poszterek Sándor Z., Horváth P., Varga A., Szolcsányi J.: Patkány TRPV1 receptort termelı stabil sejtvonal elıállítása Magyar Klinikai Farmakológusok V. Kongresszusa és MFT Immunfarmakológiai Szekció Ülése, 2003, Debrecen P. Horváth, Z. Sándor, A. Varga, Z. Szilvassy and J. Szolcsányi: Capsaicin induced enhanced apoptotic gene expression IBRO Advanced School of Neuroscience 2004, Jalta, Ukrajna Jakab B., Varga A., Bölcskei K., Szabó Á., Elekes K., Helyes Zs., Pintér E., Németh J., Szolcsányi J.: A kapszaicin receptor (TRPV1) antagonista SC0030 hatásainak vizsgálata in vitro és in vivo Magyar Élettani Társaság LXVIII. Vándorgyûlése 2004, Debrecen Németh J., Jakab B., Helyes Zs., Varga A., Bölcskei K., Szabó Á., Sándor K., Elekes K., Börzsei K., Pintér E., Pethı G., Szolcsányi J.: Examination of the novel TRPV1 receptor antagonist JYL1421 (SC0030) in vitro and in vivo in the rat Magyar Idegtudományi Társaság (MITT) XI. Kongresszusa, 2005, Pécs
Az értekezés közleményeinek összesített impakt faktora: 6.9795
Egyéb közlemények impakt faktora: 3.851
Összesen: 10.8305





























