Embed Size (px)
Citation preview

Research in Veterinary Science 90 (2011) 432–438
Contents lists available at ScienceDirect
Research in Veterinary Science
journal homepage: www.elsevier .com/locate / rvsc
Variation in the expression of Hsp27, aB-crystallin mRNA and protein in heartand liver of pigs exposed to different transport times
Miao Zhang a,b, Lingxiang Xin a, Endong Bao a,*, Jörg Hartung c, Zhenhua Yue a
a College of Veterinary Medicine, Nanjing Agricultural University, Nanjing 210095, Chinab College of Animal Science and Technology, Jinling Institute of Technology, Nanjing 210038, Chinac Institute for Animal Hygiene, Animal Welfare and Farm Animal Behaviour, University of Veterinary Medicine Hannover, Foundation, Hannover, Germany
a r t i c l e i n f o a b s t r a c t
Article history:Received 13 November 2009Accepted 28 June 2010
Keywords:Hsp27aB-crystallinHeartLiverTransportationPig
0034-5288/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.rvsc.2010.06.028
* Corresponding author. Address: College of Vetericultural University, Weigang 1, Nanjing 210095, Chin+86 2584398669.
E-mail address: [email protected] (E. Bao).
Twenty pigs were randomly divided into four groups of five pigs each (not transported – control, 1, 2 and4 h of transportation). A significant increase of ALT, AST and CK in the blood serum and acute parenchymacell lesions were observed and those were characterized by acute degenerations in the heart and liver.Hsp27 expression levels increased significantly in the heart after 2 h and in the liver after 4 h of transpor-tation, accompanying with the hsp27 mRNA increasing significantly in the heart and liver after 1 h oftransportation. aB-crystallin expression levels were fluctuant (not significantly) in the heart and liverduring transporting, however, aB-crystallin mRNA increase notably in the heart after 1 h and decreasesignificantly in the liver at 1 and 2 h of transportation, respectively. In conclusion, the cellular damageto the heart and liver is highest after 1 h of transportation, Hsp27 and aB-crystallin play dissimilar rolesand show tissue-specific response in different tissues during transportation.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
The swine industry in China has gradually moved to intensivemanagement systems that resulted in longer transportation ofpigs. It is known that transport can induce severe stress in pigs pre-dominantly from factors such as loading and unloading, heatstress, poor ventilation, high animal densities, mixing of unfamiliargroups, vibration, noise, and deprivation of water (Pineiro et al.,2007). Such multiple factors can be associated with biochemicaland structural changes in heart, liver, kidney, and other organs oftransported animals (Bao et al., 2008a,b; Yu et al., 2007). High lev-els of serum enzymes such as alanine aminotransferase (ALT),aspartate transaminase (AST) and creatine kinase (CK) often indi-cate liver and heart damage in pigs (Bao et al., 2008a; Dudleyet al., 1982; Li et al., 2007). Cellular injury and damage of musclemembranes can lead to poor meat quality such as pale, soft, andexudative (PSE) meat (Cheah et al., 1995; Gray et al., 1996; Pliquettet al., 2003). Stressful transports are also responsible for eventuallyshock and sudden death in transported pigs from organ failure,most of these occur due to sudden heart failure preceded by car-diovascular damage (Lee et al., 1996). DOA (dead on arrival) ratesof pigs being transported from the farm to a slaughterhouse range
ll rights reserved.
nary Medicine, Nanjing Agri-a. Tel.: +86 2584395316; fax:
from 0.1% to 1.0% (Allen et al., 1974; Averos et al., 2008; Warrisset al., 1989). Sains (1980) reported that 70% of pigs found deadon arrival, died due to heart failure in Great Britain. Patho-morpho-logical examinations of the myocardium of 100 pigs, those diedduring transport revealed that in 73 animals, degenerative or nec-robiotic changes of certain myocardial fibers, mostly (60%) were inthe form of granular degeneration (Bergmann et al., 1990).
Exposure of cells to stressful conditions can lead to the expres-sion of specific proteins called heat shock proteins (Hsps) (Arrigoet al., 1988). Hsps are a group of well-conserved protein familiesthat exert housekeeping, and protective functions in normal cells(Ritossa, 1962). The Hsps contain several protein groups and maybe divided into five major families on the basis of their size, struc-ture, and function: (1) Hsp110; (2) Hsp90; (3) Hsp70; (4) Hsp60;and (5) small Hsp (sHsp) families (Lindquist and Craig, 1988).The molecular mass of these sHsps range in from 15 to 30 kDa;they represent the least conserved group of Hsps. Hsp27 is a mam-malian sHsp family member that is expressed in a variety of tissuesin the presence or absence of stress (Hickey et al., 1986). It func-tions as a molecular chaperone to aid in refolding of inactive pro-teins under normal conditions and it can be overexpressed toincrease the stability of F-actin microfilaments during exposureto stressors such as hyperthermia, oxidants or cytochalasin D (Con-cannon et al., 2003). Efthymiou et al. (2004) reported that miceover-expressing Hsp27 were more protected from lethal ische-mia–reperfusion injury as compared to control littermates. VanderHeide (2002) also reported that increased expression of Hsp27

M. Zhang et al. / Research in Veterinary Science 90 (2011) 432–438 433
could protect canine ventricular myocytes from simulated ische-mia–reperfusion injury. Hsp27 has been found to be associatedwith the cytoskeleton and the regulation of actin filament dynam-ics via different signaling pathways (Arrigo, 2000; Lavoie et al.,1995). A shortage of Hsps and associated lesions in the myocar-dium may cause electromechanical dissociation, resulting in acuteheart failure and can lead under poor environmental and manage-ment conditions such as a stressful transport and may lead todeath.
The protein a-crystallin was originally identified in the verte-brate lens (de Jong et al., 1998). Two forms exist: aF-crystallinand aB-crystallin (aB-c), which are closely related to Hsp27 in bothstructure and function (Caspers et al., 1995; Merck et al., 1993; VanDer Ouderaa et al., 1974). aB-crystallin, in contrast to aA-crystal-lin, is normally expressed in many non-lenticular tissues such asthe heart, liver, kidney and stomach, and shows molecular chaper-one and Hsp properties similar to Hsp27 (Atomi et al., 1991; Iwakiet al., 1990). Because of the similarities between Hsp27 and aB-crystallin, many studies concerned with these two proteins havebeen conducted under a variety of conditions. Martin et al.(1997) demonstrated that both Hsp27 and aB-crystallin were ableto protect cardiac myocytes from the effect of simulated ischemia.However, Bao et al. (2002) reported that the level of Hsp27 de-creased in the liver and heart of pigs after 6 h of transportationwhich suggested that the protection of Hsp27 to the heart maybe limited after longer transportation times. The dynamic changesin the levels of Hsp27, aB-crystallin proteins and their correspond-ing mRNA in the tissues of pigs during the transportation period,can help to understand their response to transport conditionsand their role in cellular protection. Therefore, the aim of the pres-ent study is to examine the dynamic changes of the expression ofHsp27, aB-crystallin proteins and their corresponding mRNA inheart and liver of young pigs, exposed to different transport times.
2. Materials and methods
2.1. Animals and experimental design
Twenty hybrid pigs from Erhualian and Pietrain strains were se-lected and raised in individual pens (2.5 � 3.0 m2) at the Academyof Agricultural Sciences (Nanjing City, China) following usual pro-cedures, necessary to meet the requirements of pigs.
The pigs were randomly divided into four groups, each groupcontained five pigs. The mean weight of the pigs was approxi-mately 50 ± 2 kg (Mean ± SD). On the day of the transport trial,one group was maintained under normal housing conditions andserved as control (not transported animals), while the other threegroups were transported for 1, 2 and 4 h, respectively. The route in-cluded an equivalent mix of local roads, including town traffic,state roads and highways. The average speed was 30–40 km perhour.
Immediately after the end of the transport, all animals wereeuthanized by jugular injection with 10 mg/kg of 3% sodium pen-tobarbital while the animals were on the truck or in the animalhouse. Blood samples for serum enzyme activities were collectedby exsanguination, and the sera obtained were frozen at �20 �Cand stored until further analysis. Tissue specimens were takenfrom heart and liver and fixed in paraformaldehyde solution forhistopathological examination. Additional samples were placedinto 1.5 ml tubes and were frozen in the liquid nitrogen for laterevaluation of Hsp27, aB-crystallin protein expressions and theircorresponding mRNA transcriptions.
All experiments were performed and approved by the Institu-tional Animal Care and Use Committee of Nanjing AgriculturalUniversity.
2.2. Determination of serum enzyme activities
Serum ALT, AST and CK were measured according to the instruc-tions given in the commercial kits (Nanjing Jiancheng BiochemicalReagent Co., Nanjing, China), using a clinical biochemical indicatorautoanalyzer (Vital Scientific NV, The Netherlands).
2.3. Histopathological examination
Paraformaldehyde-fixed and paraffin-embedded heart and livertissues were sliced serially into 4-lm sections, and the sectionswere routinely stained with hematoxylin and eosin (H&E) andexamined by using light microscopy.
2.4. Semi-quantitative detection of Hsp27 and aB-crystallin byenzyme-linked immunosorbent assay (ELISA)
After complete washing in ice-cold physiological saline, theheart and liver samples were homogenized on ice in 10 volumesof homogenization buffer [0.15 M NaCl, 20 mM Tris–HCl (pH 8.0),1 mM EDTA, 1 mM PMSF, 0.1 lM E-46, 0.08 lM aprotinin, 0.1 lMleupeptin, and 0.1% NP-40] (Shaila et al., 2005) using an Ultra-Tur-rax homogenizer (Scientz, China). The homogenates were centri-fuged at 12,000 g for 20 min at 4 �C to remove cellular debris.The supernatants were collected and stored at �20 �C for later pro-tein quantification.
The levels of Hsp27 and aB-crystallin in the heart and liversamples of both transported and not transported animals weremeasured using commercially available ELISA kits (QRCT-30123011 EIAnUTL, goat anti-porcine Heat Shock Protein 27,Adlitteram Diagnostic Laboratories, USA; QRCT-3013321030011EIAnUTL, goat anti-porcine aB-crystallin, Adlitteram DiagnosticLaboratories, USA). Quantification of samples was performed byusing a standard curve. b-Actin (QRCT-3222211 EIAnUTL, goatanti-porcine b-Actin, Adlitteram Diagnostic Laboratories, USA)was used to control for bias caused by the protein extraction pro-cedure. The assay was performed following the manufacturer’sprotocol and the sensitivity was 0.01 ng/mL. The quantity ofHsp27 and aB-crystallin in each sample was normalized usingthe following formula:
Relative quantity of Hsp27¼quantity of Hsp27=quantity of b-Actin
Relative quantity of aB-crystallin
¼ quantity of aB-crystallin=quantity of b-Actin
2.5. Detection of hsp27 and aB-crystallin mRNA by fluorescencequantitative real time PCR (FQ�RT-PCR)
2.5.1. Isolation of total RNA and RT-PCRHalf gram of the tissue stored at �70 �C was ground in liquid
nitrogen after being washed in ice-cold physiological saline. TotalRNA was isolated from the ground tissue using TRIZOL reagent(Invitrogen, USA) according to the manufacturer’s instructions.The concentration of RNA was determined by a spectrophotometer(Mx3000P, Stratagene) at 260 nm. Serial dilutions of RNA were pre-pared with ribonuclease-free water and 2 lg of each sample wassynthesized into DNA using the TRANScript M-MLV kit (Qiagen,Germany) following the manufacturer’s protocol, and then storedat �20 �C until use.
2.5.2. Design of primers for hsp27 mRNA, aB-crystallin mRNA andglyceraldehyde-3-phosphate dehydrogenase mRNA
Primer sets were specifically designed to anneal to each targetmRNA. The sequences of hsp27 mRNA and GAPDH mRNA were ob-

434 M. Zhang et al. / Research in Veterinary Science 90 (2011) 432–438
tained from the National Center for Biotechnology Information’s(NCBI) GenBank (accession nos. NM_001007518 and AF017079,respectively) and aB-crystallin mRNA from Liao et al. (1998). Theprimers were designed using Primer premier 5.0 software for con-ventional and RT-PCR amplification. Highly purified salt-free prim-ers for the hsp27 gene (forward primer, 50-CCGGTGTTTCACTCGAAAATACA-30, reverse primer, 50-GCTTTTCCGACTTTCCAGCTTCT-30, 200 bp), aB-crystallin gene (forward primer, 50-CCGCCTCTTTGACCAGTT-30, reverse primer, 50-AGAGCCCAGTGTCAATCC-30,134 bp) and the GAPDH gene (forward primer, 50-GAAGGTCGGAGTGAACGGAT-30, reverse primer, 50-CATGGGT AGAATCATACTGGAACA-30, 149 bp) were synthesized by Invitrogen (Nanjing, Chi-na) and optimized to an equal annealing temperature of 60 �C.
2.5.3. FQ RT-PCREach DNA sample (2 lL, 25 times dilution) was suspended in
2 � SYBR Premix Ex Taq™ (DRR041A, TaKaRa, Japan) with primer(25 pmol of downstream and upstream primer, respectively), anddouble-distilled water to a total volume of 25 lL. QuantitativePCR was performed using an ABI 7300 RT-PCR thermocycler (Ap-plied Biosystems, USA). The thermal profile was establishedaccording to the manufacturer’s protocol. Briefly, this protocolwas 95 �C for 3 min for enzyme activation, followed by denatur-ing at 95 �C for 5 s, and annealing and elongation at 60 �C for 30s, for a total of 45 cycles. For each run, a negative control tubewithout DNA was run along with the experimental samples. A2-fold dilution series of the template was used in the FQ-PCRreactions to obtain three standard curves as follows: the target,hsp27 mRNA: Y = �3.299 Log(x) + 36.19, r2 = 0.998; aB-crystallinmRNA: Y = �3.316 Log(x) + 26.13, r2 = 0.998; and as a reference,GAPDH mRNA: Y = �3.310 Log(x) + 26.86, r2 = 0.997. The amplifi-cation efficiencies of the target and reference were approxi-mately equal. Therefore, the hsp27 mRNA and aB-crystallinmRNA of all samples could be normalized using the followingformula:
Relative quantity of �mRNA ¼ 2�DDCt:
DDCt ¼ ðCt�mRNA � CtGAPDHÞ control group� ðCt�
� CtGAPDHÞ test group
� represents either Hsp27 or aB-crystallin.
2.6. Statistical analysis
Statistical analysis of the differences between each group wascarried out using a one-way analysis of variance (ANOVA) usingthe Statistical Package for Social Sciences (SPSS version 11.5). Com-parison of the mean value of the control group with that of eachexperimental group was performed using the Duncan test for mul-tiple comparisons.
Table 1Activity levels of ALT, AST and CK in the serum of transported pigs (U/L).
Transport Control groupTime 0 h 1 h
ALT 34.75 ± 9.60 55.60 ± 12.62**
AST 50.50 ± 11.82 95.75 ± 11.87**
CK 999.67 ± 81.35 5416.5 ± 1133.28**
Values indicated are Mean ± SD; n = 5.* P < 0.05.** P < 0.01.
3. Results
3.1. The levels of ALT, AST and CK in serum
Table 1 shows the serum levels of ALT, AST and CK obtained fromthe three transported pig groups (1, 2, 4 h) and the not transportedpig group (control, 0 h). Highest ALT concentrations were observedin the group which was transported for 1 h (P < 0.01). In the groupwhich was transported for 2 h, the value was decreased. The ani-mals in the 4 h group showed similar levels of ALT to that of thecontrol group (0 h). AST levels displayed a similar tendency, butthe levels of the 4 h group did not fully return to normal. Levelsof CK in the animals who were transported for 1 and 2 h groupswere more than four times higher than that of the control group.
3.2. Histopathological examination
Histopathological changes of the liver tissues are illustrated inFig. 1. After 1 h transportation, the parenchyma cells of livershowed acute degeneration in all five pigs (Fig. 1b). Vacuolardegeneration was detected in the cytoplasm of hepatocytes, espe-cially at the centre of the hepatic lobule after 2 h transportation(shown in four of five pigs) (Fig. 1c). Hepatocyte necrosis charac-terized by karyolysis was also present after 4 h of transportation(shown in four of five pigs) (Fig. 1d). No obvious lesions were foundin the liver of control pigs (Fig. 1a).
After 2 h transportation, the acute degeneration of the myocar-dial cells was recognized by light pink staining and loss of stria-tions in the cytoplasm (shown in four of five pigs). Themyocardial fibers showed various thicknesses after 4 h transporta-tion (shown in three of five pigs).
3.3. The expression levels of hsp27 mRNA and protein in the heart andliver of transported pigs
As shown in Fig. 2, the levels of Hsp27 in the heart were in-creased significantly (P < 0.01) and reached peak levels after 2 hof transportation. After 4 h of transport, Hsp27 expression were de-creased to the control level. The transcription of hsp27 mRNA wassignificantly higher in all transported groups as compared to thecontrol. It seemed to increase with the length of the journey, atleast up to 4 h.
The levels of Hsp27 expression and hsp27 mRNA transcription inthe liver are shown in Fig. 3. While the increase of Hsp27 was inco-herent with the highest value in the 4 h group, the hsp27 mRNAshowed a steep increase (P < 0.01) with transport time.
3.4. The expression levels of aB-crystallin mRNA and protein in theheart and liver of transported pigs
Fig. 4 shows that the concentrations of aB-crystallin mRNA inthe heart of transported pigs were increased significantly with
Transported pig groups2 h 4 h
53.60 ± 7.09** 30.60 ± 5.1875.60 ± 12.93* 66.75 ± 15.264564.75 ± 1669.89** 1769.75 ± 376.81
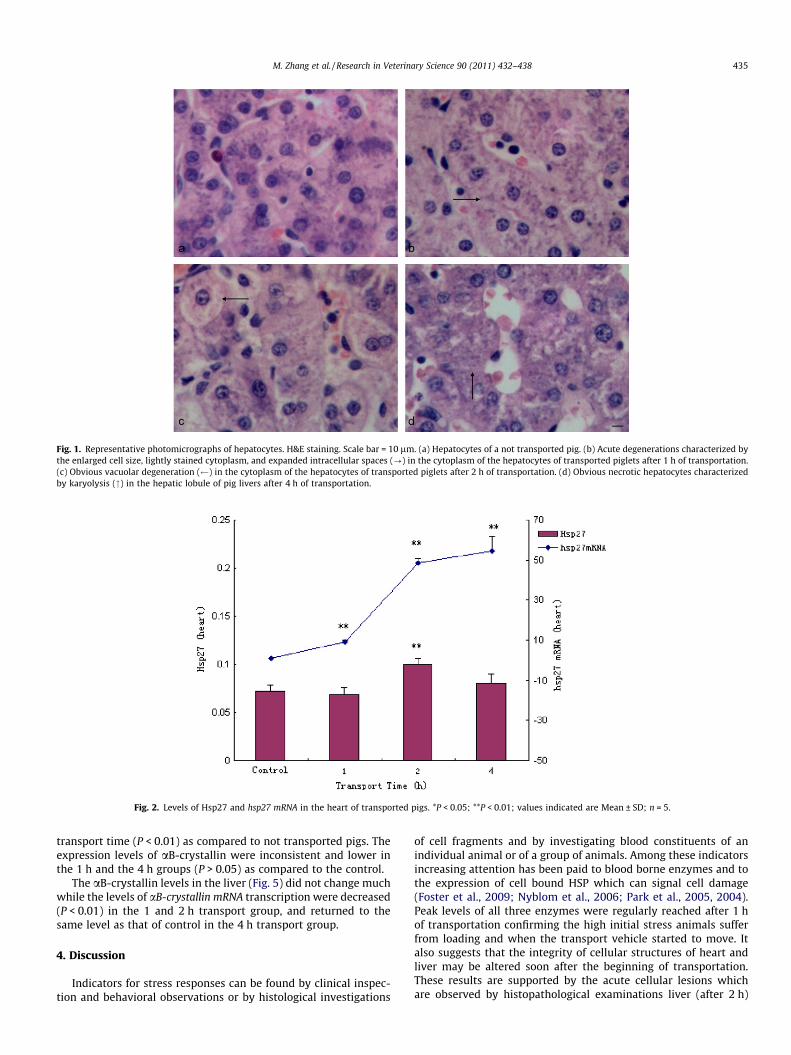
Fig. 1. Representative photomicrographs of hepatocytes. H&E staining. Scale bar = 10 lm. (a) Hepatocytes of a not transported pig. (b) Acute degenerations characterized bythe enlarged cell size, lightly stained cytoplasm, and expanded intracellular spaces (?) in the cytoplasm of the hepatocytes of transported piglets after 1 h of transportation.(c) Obvious vacuolar degeneration ( ) in the cytoplasm of the hepatocytes of transported piglets after 2 h of transportation. (d) Obvious necrotic hepatocytes characterizedby karyolysis (") in the hepatic lobule of pig livers after 4 h of transportation.
Fig. 2. Levels of Hsp27 and hsp27 mRNA in the heart of transported pigs. *P < 0.05; **P < 0.01; values indicated are Mean ± SD; n = 5.
M. Zhang et al. / Research in Veterinary Science 90 (2011) 432–438 435
transport time (P < 0.01) as compared to not transported pigs. Theexpression levels of aB-crystallin were inconsistent and lower inthe 1 h and the 4 h groups (P > 0.05) as compared to the control.
The aB-crystallin levels in the liver (Fig. 5) did not change muchwhile the levels of aB-crystallin mRNA transcription were decreased(P < 0.01) in the 1 and 2 h transport group, and returned to thesame level as that of control in the 4 h transport group.
4. Discussion
Indicators for stress responses can be found by clinical inspec-tion and behavioral observations or by histological investigations
of cell fragments and by investigating blood constituents of anindividual animal or of a group of animals. Among these indicatorsincreasing attention has been paid to blood borne enzymes and tothe expression of cell bound HSP which can signal cell damage(Foster et al., 2009; Nyblom et al., 2006; Park et al., 2005, 2004).Peak levels of all three enzymes were regularly reached after 1 hof transportation confirming the high initial stress animals sufferfrom loading and when the transport vehicle started to move. Italso suggests that the integrity of cellular structures of heart andliver may be altered soon after the beginning of transportation.These results are supported by the acute cellular lesions whichare observed by histopathological examinations liver (after 2 h)
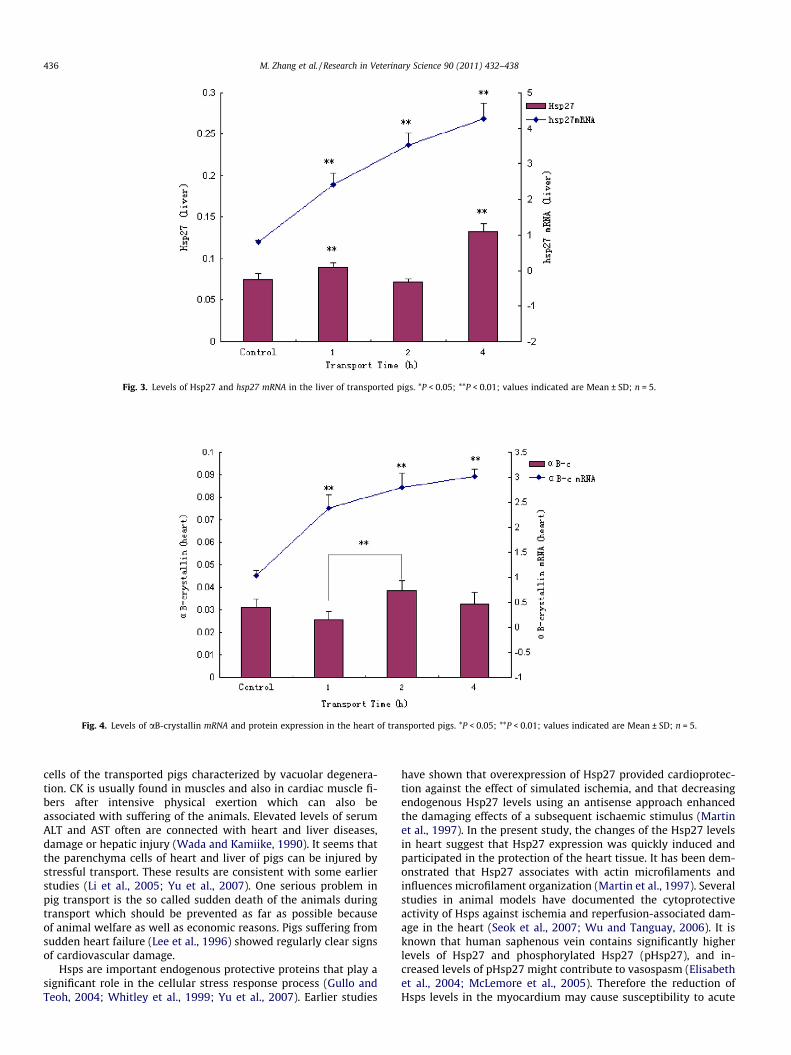
Fig. 3. Levels of Hsp27 and hsp27 mRNA in the liver of transported pigs. *P < 0.05; **P < 0.01; values indicated are Mean ± SD; n = 5.
Fig. 4. Levels of aB-crystallin mRNA and protein expression in the heart of transported pigs. *P < 0.05; **P < 0.01; values indicated are Mean ± SD; n = 5.
436 M. Zhang et al. / Research in Veterinary Science 90 (2011) 432–438
cells of the transported pigs characterized by vacuolar degenera-tion. CK is usually found in muscles and also in cardiac muscle fi-bers after intensive physical exertion which can also beassociated with suffering of the animals. Elevated levels of serumALT and AST often are connected with heart and liver diseases,damage or hepatic injury (Wada and Kamiike, 1990). It seems thatthe parenchyma cells of heart and liver of pigs can be injured bystressful transport. These results are consistent with some earlierstudies (Li et al., 2005; Yu et al., 2007). One serious problem inpig transport is the so called sudden death of the animals duringtransport which should be prevented as far as possible becauseof animal welfare as well as economic reasons. Pigs suffering fromsudden heart failure (Lee et al., 1996) showed regularly clear signsof cardiovascular damage.
Hsps are important endogenous protective proteins that play asignificant role in the cellular stress response process (Gullo andTeoh, 2004; Whitley et al., 1999; Yu et al., 2007). Earlier studies
have shown that overexpression of Hsp27 provided cardioprotec-tion against the effect of simulated ischemia, and that decreasingendogenous Hsp27 levels using an antisense approach enhancedthe damaging effects of a subsequent ischaemic stimulus (Martinet al., 1997). In the present study, the changes of the Hsp27 levelsin heart suggest that Hsp27 expression was quickly induced andparticipated in the protection of the heart tissue. It has been dem-onstrated that Hsp27 associates with actin microfilaments andinfluences microfilament organization (Martin et al., 1997). Severalstudies in animal models have documented the cytoprotectiveactivity of Hsps against ischemia and reperfusion-associated dam-age in the heart (Seok et al., 2007; Wu and Tanguay, 2006). It isknown that human saphenous vein contains significantly higherlevels of Hsp27 and phosphorylated Hsp27 (pHsp27), and in-creased levels of pHsp27 might contribute to vasospasm (Elisabethet al., 2004; McLemore et al., 2005). Therefore the reduction ofHsps levels in the myocardium may cause susceptibility to acute
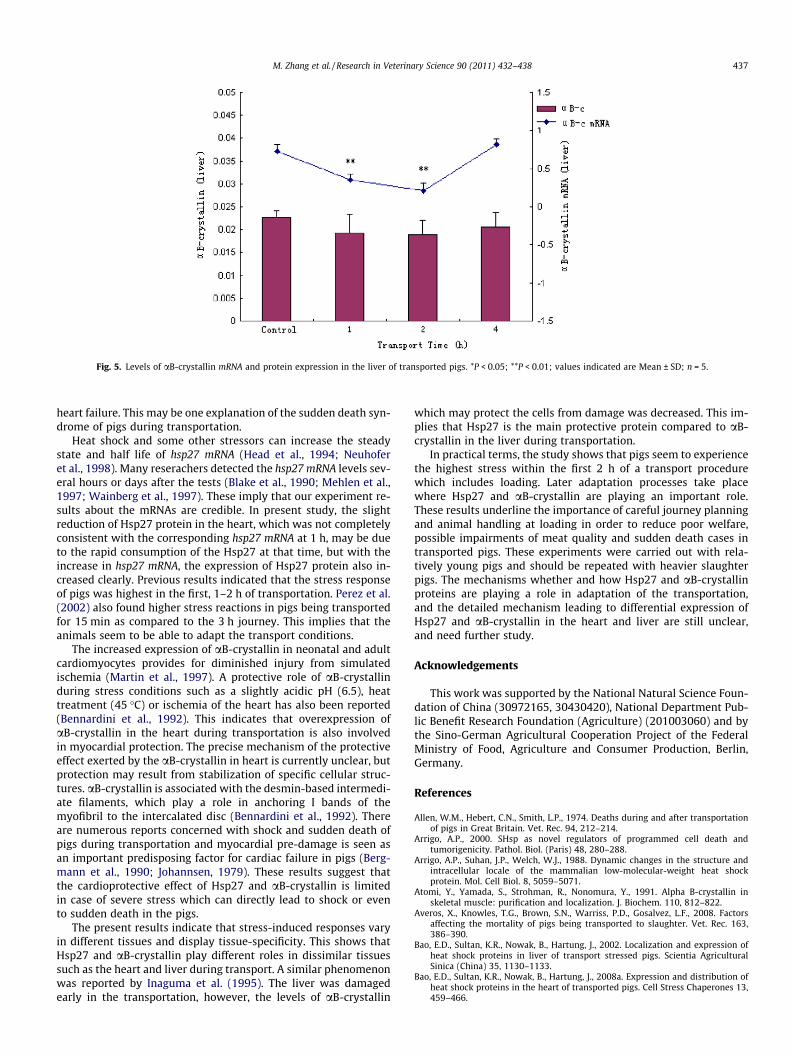
Fig. 5. Levels of aB-crystallin mRNA and protein expression in the liver of transported pigs. *P < 0.05; **P < 0.01; values indicated are Mean ± SD; n = 5.
M. Zhang et al. / Research in Veterinary Science 90 (2011) 432–438 437
heart failure. This may be one explanation of the sudden death syn-drome of pigs during transportation.
Heat shock and some other stressors can increase the steadystate and half life of hsp27 mRNA (Head et al., 1994; Neuhoferet al., 1998). Many reserachers detected the hsp27 mRNA levels sev-eral hours or days after the tests (Blake et al., 1990; Mehlen et al.,1997; Wainberg et al., 1997). These imply that our experiment re-sults about the mRNAs are credible. In present study, the slightreduction of Hsp27 protein in the heart, which was not completelyconsistent with the corresponding hsp27 mRNA at 1 h, may be dueto the rapid consumption of the Hsp27 at that time, but with theincrease in hsp27 mRNA, the expression of Hsp27 protein also in-creased clearly. Previous results indicated that the stress responseof pigs was highest in the first, 1–2 h of transportation. Perez et al.(2002) also found higher stress reactions in pigs being transportedfor 15 min as compared to the 3 h journey. This implies that theanimals seem to be able to adapt the transport conditions.
The increased expression of aB-crystallin in neonatal and adultcardiomyocytes provides for diminished injury from simulatedischemia (Martin et al., 1997). A protective role of aB-crystallinduring stress conditions such as a slightly acidic pH (6.5), heattreatment (45 �C) or ischemia of the heart has also been reported(Bennardini et al., 1992). This indicates that overexpression ofaB-crystallin in the heart during transportation is also involvedin myocardial protection. The precise mechanism of the protectiveeffect exerted by the aB-crystallin in heart is currently unclear, butprotection may result from stabilization of specific cellular struc-tures. aB-crystallin is associated with the desmin-based intermedi-ate filaments, which play a role in anchoring I bands of themyofibril to the intercalated disc (Bennardini et al., 1992). Thereare numerous reports concerned with shock and sudden death ofpigs during transportation and myocardial pre-damage is seen asan important predisposing factor for cardiac failure in pigs (Berg-mann et al., 1990; Johannsen, 1979). These results suggest thatthe cardioprotective effect of Hsp27 and aB-crystallin is limitedin case of severe stress which can directly lead to shock or evento sudden death in the pigs.
The present results indicate that stress-induced responses varyin different tissues and display tissue-specificity. This shows thatHsp27 and aB-crystallin play different roles in dissimilar tissuessuch as the heart and liver during transport. A similar phenomenonwas reported by Inaguma et al. (1995). The liver was damagedearly in the transportation, however, the levels of aB-crystallin
which may protect the cells from damage was decreased. This im-plies that Hsp27 is the main protective protein compared to aB-crystallin in the liver during transportation.
In practical terms, the study shows that pigs seem to experiencethe highest stress within the first 2 h of a transport procedurewhich includes loading. Later adaptation processes take placewhere Hsp27 and aB-crystallin are playing an important role.These results underline the importance of careful journey planningand animal handling at loading in order to reduce poor welfare,possible impairments of meat quality and sudden death cases intransported pigs. These experiments were carried out with rela-tively young pigs and should be repeated with heavier slaughterpigs. The mechanisms whether and how Hsp27 and aB-crystallinproteins are playing a role in adaptation of the transportation,and the detailed mechanism leading to differential expression ofHsp27 and aB-crystallin in the heart and liver are still unclear,and need further study.
Acknowledgements
This work was supported by the National Natural Science Foun-dation of China (30972165, 30430420), National Department Pub-lic Benefit Research Foundation (Agriculture) (201003060) and bythe Sino-German Agricultural Cooperation Project of the FederalMinistry of Food, Agriculture and Consumer Production, Berlin,Germany.
References
Allen, W.M., Hebert, C.N., Smith, L.P., 1974. Deaths during and after transportationof pigs in Great Britain. Vet. Rec. 94, 212–214.
Arrigo, A.P., 2000. SHsp as novel regulators of programmed cell death andtumorigenicity. Pathol. Biol. (Paris) 48, 280–288.
Arrigo, A.P., Suhan, J.P., Welch, W.J., 1988. Dynamic changes in the structure andintracellular locale of the mammalian low-molecular-weight heat shockprotein. Mol. Cell Biol. 8, 5059–5071.
Atomi, Y., Yamada, S., Strohman, R., Nonomura, Y., 1991. Alpha B-crystallin inskeletal muscle: purification and localization. J. Biochem. 110, 812–822.
Averos, X., Knowles, T.G., Brown, S.N., Warriss, P.D., Gosalvez, L.F., 2008. Factorsaffecting the mortality of pigs being transported to slaughter. Vet. Rec. 163,386–390.
Bao, E.D., Sultan, K.R., Nowak, B., Hartung, J., 2002. Localization and expression ofheat shock proteins in liver of transport stressed pigs. Scientia AgriculturalSinica (China) 35, 1130–1133.
Bao, E.D., Sultan, K.R., Nowak, B., Hartung, J., 2008a. Expression and distribution ofheat shock proteins in the heart of transported pigs. Cell Stress Chaperones 13,459–466.

438 M. Zhang et al. / Research in Veterinary Science 90 (2011) 432–438
Bao, E.D., Sultan, K.R., Nowak, B., Hartung, J., 2008b. Localization of heat shockproteins and histopathological changes in the kidneys of transported pigs.Livestock Sci. 118, 231–237.
Bennardini, F., Wrzosek, A., Chiesi, M., 1992. Alpha B-crystallin in cardiac tissue.Association with actin and desmin filaments. Circ. Res. 71, 288–294.
Bergmann, V., Grafe, A., Seifert, H., 1990. The pathomorphology and pathogenesis ofacute cardiovascular failure in swine. 3. Histopathologic findings in themyocardium of swine with transport-related cardiovascular insufficiency.Arch. Exp. Veterinarmed. 44, 521–532.
Blake, M.J., Gershon, D., Fargnoli, J., Holbrook, N.J., 1990. Discordant expression ofheat shock protein mRNAs in tissues of heat-stressed rats. J. Biol. Chem. 265,15275–15279.
Caspers, G.J., Leunissen, J.A., de Jong, W.W., 1995. The expanding small heat-shockprotein family, and structure predictions of the conserved ‘‘alpha-crystallindomain”. J. Mol. Evol. 40, 238–248.
Cheah, K.S., Levy, A., Trainor, P.A., Wai, A.W., Kuffner, T., So, C.L., Leung, K.K., Lovell-Badge, R.H., Tam, P.P., 1995. Human COL2A1-directed SV40 T antigenexpression in transgenic and chimeric mice results in abnormal skeletaldevelopment. J. Cell Biol. 128, 223–237.
Concannon, C.G., Gorman, A.M., Samali, A., 2003. On the role of Hsp27 in regulatingapoptosis. Apoptosis 8, 61–70.
de Jong, W.W., Caspers, G.J., Leunissen, J.A., 1998. Genealogy of the alpha-crystallin–small heat-shock protein superfamily. Int. J. Biol. Macromol. 22, 151–162.
Dudley, R.E., Svoboda, D.J., Klaassen, C., 1982. Acute exposure to cadmium causessevere liver injury in rats. Toxicol. Appl. Pharmacol. 65, 302–313.
Efthymiou, C.A., Mocanu, M.M., de Belleroche, J., Wells, D.J., Latchmann, D.S., Yellon,D.M., 2004. Heat shock protein 27 protects the heart against myocardialinfarction. Basic Res. Cardiol. 99, 392–394.
Elisabeth, C.M., Deron, T., Padmini, K., Jeffrey, T., Colleen, B., 2004. The yin and yangof vascular smooth muscle tone. J. Am. Coll. Surg. 199, 106–107.
Foster, C.S., Dodson, A.R., Ambroisine, L., Fisher, G., Moller, H., Clark, J., Attard, G., De-Bono, J., Scardino, P., Reuter, V.E., Cooper, C.S., Berney, D.M., Cuzick, J., 2009.Hsp-27 expression at diagnosis predicts poor clinical outcome in prostatecancer independent of ETS-gene rearrangement. Br. J. Cancer 101, 1137–1144.
Gray, J.I., Gomaa, E.A., Buckley, D.J., 1996. Oxidative quality and shelf life of meats.Meat Sci. 43, 111–112.
Gullo, C.A., Teoh, G., 2004. Heat shock proteins: to present or not, that is thequestion. Immunol. Lett. 94, 1–10.
Head, M.W., Corbin, E., Goldman, J.E., 1994. Coordinate and independent regulationof alpha B-crystallin and hsp27 expression in response to physiological stress. J.Cell Physiol. 159, 41–50.
Hickey, E., Brandon, S.E., Potter, R., Stein, G., Stein, J., Weber, L.A., 1986. Sequenceand organization of genes encoding the human 27 kDa heat shock protein.Nucleic Acids Res. 14, 4127–4145.
Inaguma, Y., Hasegawa, K., Goto, S., Ito, H., Kato, K., 1995. Induction of the synthesisof hsp27 and alpha B crystallin in tissues of heat-stressed rats and itssuppression by ethanol or an alpha 1-adrenergic antagonist. J. Biochem. 117,1238–1243.
Iwaki, T., Kume-Iwaki, A., Goldman, J.E., 1990. Cellular distribution of alpha B-crystallin in non-lenticular tissues. J. Histochem. Cytochem. 38, 31–39.
Johannsen, U., 1979. Studies of cardiac pathomorphology in swine which diedduring transport. Arch. Exp. Veterinarmed. 33, 377–391.
Lavoie, J.N., Lambert, H., Hickey, E., Weber, L.A., Landry, J., 1995. Modulation ofcellular thermoresistance and actin filament stability accompaniesphosphorylation-induced changes in the oligomeric structure of heat shockprotein 27. Mol. Cell Biol. 15, 505–516.
Lee, W.C., Lin, K.Y., Chiu, Y.T., Lin, J.H., Cheng, H.C., Huang, H.C., Yang, P.C., Liu, S.K.,Mao, S.J., 1996. Substantial decrease of heat shock protein 90 in ventriculartissues of two sudden-death pigs with hypertrophic cardiomyopathy. FASEB J.10, 1198–1204.
Li, Y.B., Bao, E.D., Wang, Z.L., Zhao, R.Q., 2007. Detection of HSP mRNA transcriptionin transport stressed pigs by fluorescence quantitative RT-PCR. Agric. Sci. China6, 361–367.
Li, Y.B., Sun, F.M., Wang, Z.L., 2005. Relationship between pathological lesion anddistribution of HSP27 in transport stressed pigs. J. Nanjing Agric. University 28,83–87.
Liao, J.H., Hung, C.C., Lee, J.S., Wu, S.H., Chiou, S.H., 1998. Characterization, cloning,and expression of porcine alpha B crystallin. Biochem. Biophys. Res. Commun.244, 131–137.
Lindquist, S., Craig, E.A., 1988. The heat-shock proteins. Annu. Rev. Genet. 22, 631–677.
Martin, J.L., Mestril, R., Hilal-Dandan, R., Brunton, L.L., Dillmann, W.H., 1997. Smallheat shock proteins and protection against ischemic injury in cardiac myocytes.Circulation 96, 4343–4348.
McLemore, E.C., Tessier, D.J., Thresher, J., Komalavilas, P., Brophy, C.M., 2005. Role ofthe small heat shock proteins in regulating vascular smooth muscle tone. J. Am.Coll. Surg. 201, 30–36.
Mehlen, P., Mehlen, A., Godet, J., Arrigo, A.P., 1997. Hsp27 as a switch betweendifferentiation and apoptosis in murine embryonic stem cells. J. Biol. Chem. 272,31657–31665.
Merck, K.B., Groenen, P.J., Voorter, C.E., de Haard-Hoekman, W.A., Horwitz, J.,Bloemendal, H., de Jong, W.W., 1993. Structural and functional similarities ofbovine alpha-crystallin and mouse small heat-shock protein. A family ofchaperones. J. Biol. Chem. 268, 1046–1052.
Neuhofer, W., Muller, E., Burger-Kentischer, A., Beck, F.X., 1998. Hypertonicityaffects heat shock protein 27 and F-actin localization in Madin–Darby caninekidney cells. Kidney Int. Suppl. 67, S165–167.
Nyblom, H., Bjornsson, E., Simren, M., Aldenborg, F., Almer, S., Olsson, R., 2006. TheAST/ALT ratio as an indicator of cirrhosis in patients with PBC. Liver Int. 26,840–845.
Park, G., Jones, D.B., Katelaris, P., 2005. Value of AST/ALT ratio as fibrotic predictor inchronic hepatitis C. Am. J. Gastroenterol. 100, 1623–1624 (author reply 1624).
Park, S.Y., Kang, K.H., Park, J.H., Lee, J.H., Cho, C.M., Tak, W.Y., Kweon, Y.O., Kim, S.K.,Choi, Y.H., 2004. Clinical efficacy of AST/ALT ratio and platelet counts aspredictors of degree of fibrosis in HBV infected patients without clinicallyevident liver cirrhosis. Korean J. Gastroenterol. 43, 246–251.
Perez, M.P., Palacio, J., Santolaria, M.P., del Acena, M.C., Chacon, G., Verde, M.T.,Calvo, J.H., Zaragoza, M.P., Gascon, M., Garcia-Belenguer, S., 2002. Influence oflairage time on some welfare and meat quality parameters in pigs. Vet. Res. 33,239–250.
Pineiro, M., Pineiro, C., Carpintero, R., Morales, J., Campbell, F.M., Eckersall, P.D.,Toussaint, M.J., Lampreave, F., 2007. Characterisation of the pig acute phaseprotein response to road transport. Vet. J. 173, 669–674.
Pliquett, U., Altmann, M., Pliquett, F., Schöberlein, L., 2003. Py-a parameter for meatquality. Meat Sci. 65, 1429–1437.
Ritossa, F., 1962. A new puffing pattern induced by temperature shock and DNP inDrosophila. Cell. Mol. Life Sci. 18, 571–573.
Sains, A., 1980. Deaths in transit: what British surveys show. Pig Farming 28, 40–41.Seok, Y.M., Kim, J., Choi, K.C., Yoon, C.H., Boo, Y.C., Park, Y., Park, K.M., 2007. Wen-pi-
tang-Hab-Wu-ling-san attenuates kidney ischemia/reperfusion injury in mice.A role for antioxidant enzymes and heat-shock proteins. J. Ethnopharmacol.112, 333–340.
Shaila, S., Angshuman, S., Abhijeet, K., Samindranath, M., Jayanta, K.P., 2005.Flufenoxuron, an acylurea insect growth regulator, alters development ofTribolium castaneum (Herbst) (Coleoptera: Tenebrionidae) by modulatinglevels of chitin, soluble protein content, and HSP70 and p34cdc2 in the larvaltissues. Pestic. Biochem. Physiol. 85, 84–90.
Van Der Ouderaa, F.J., De Jong, W.W., Hilderink, A., Bloemendal, H., 1974. Theamino-acids sequence of the alphaB2 chain of bovine alpha-crystallin. Eur. J.Biochem. 49, 157–168.
Vander Heide, R.S., 2002. Increased expression of HSP27 protects canine myocytesfrom simulated ischemia-reperfusion injury. Am. J. Physiol. Heart Circ. Physiol.282, H935–941.
Wada, H., Kamiike, W., 1990. Aspartate aminotransferase isozymes and theirclinical significance. Prog. Clin. Biol. Res. 344, 853–875.
Wainberg, Z., Oliveira, M., Lerner, S., Tao, Y., Brenner, B.G., 1997. Modulation ofstress protein (hsp27 and hsp70) expression in CD4+ lymphocytic cellsfollowing acute infection with human immunodeficiency virus type-1.Virology 233, 364–373.
Warriss, P.D., Bevis, E.A., Ekins, P.J., 1989. The relationships between glycogen storesand muscle ultimate pH in commercially slaughtered pigs. Br. Vet. J. 145, 378–383.
Whitley, D., Goldberg, S.P., Jordan, W.D., 1999. Heat shock proteins: a review of themolecular chaperones. J. Vasc. Surg. 29, 748–751.
Wu, T., Tanguay, R.M., 2006. Antibodies against heat shock proteins inenvironmental stresses and diseases: friend or foe? Cell Stress Chaperones 11,1–12.
Yu, H., Bao, E.D., Zhao, R.Q., Lv, Q.X., 2007. Effect of transportation stress on heatshock protein 70 concentration and mRNA expression in heart and kidneytissues and serum enzyme activities and hormone concentrations of pigs. Am. J.Vet. Res. 68, 1145–1150.







![Characterization of an antibody that recognizes peptides ... · in αA-crystallin (Asp 58 and Asp 151) [3], αB-crystallin (Asp 36 and Asp 62) [4], and βB2-crsytallin (Asp 4) [5]](https://img.pdfslide.net/doc/110x75/5ff1e68e89243b57b64135f8/characterization-of-an-antibody-that-recognizes-peptides-in-a-crystallin-asp.jpg)





















