Embed Size (px)
Citation preview

GENERAL AND COMPARATIVE ENDOCRINOLOGY 79, 12-22 (1990)
Vasopressor and Depressor Effects of Native Angiotensins and Inhibition of These Effects in the Japanese Quail
YOSHIO TAKEI AND YUKIO HASEGAWA*
Department of Physiology, Kitasato Universib School of Medicine, 1-15-1 Kitasato, Sagamihara, Kanagawa 228, and *Department of Clinical Chemistry, School of Pharmaceutical Sciences, Toho University, 2-2-l
Miyama, Funabashi, Chiba 274, Japan
Accepted July 25, 1989
The amino acid sequence of angiotensin I (ANG I) from the Japanese quail, Coturnix coturnixjaponica, obtained from incubation of homologous plasma and kidney extract was determined as H-Asp-Arg-Val-Tyr-Val-His-Pro-PheSer-Leu-OH. A bolus, intravenous injection of native ANG I or of ANG II induced an immediate vasodepressor response and a subsequent vasopressor response in quail which has been lightly anesthetized with ure- thane (0.75 gikg). The values for ED,, for the vasopressor and depressor effects of ANG II were 85 and 113 pmoVlO0 g body weight, respectively. The extent of the hypotension was dependent on the arterial pressure prior to injection. The effects of ANG I and II on heart rate were variable. Human [Asp’, Ile5, Hiss] ANG I and II were almost as potent as their quail counterparts with respect to the cardiovascular effects, but eel [Asn’, Val’, Gly? ANG I and II were less than half as potent. Human ANG III had little effect on arterial pressure in the quail. A bolus injection (108 l.&30 g) or infusion (1 ~&100 g/min) of [Sar’, Be*] ANG II almost abolished the cardiovascular effects of ANG I and II, but [Sar’, Alas] ANG II and [Sar’, Thrs] ANG II, which are effective inhibitors in mammals, had no inhibitory effects. The vasopressor and depressor effects of ANG I were abolished, while those of ANG II were slightly enhanced, after injection (100 l&O0 g) or infusion (1 l@OO g/min) of SQl4225, whereas des-Pro’ bradykinin and bradykinin potentiator B, which are effective inhibitors of ANG I converting enzyme in mammals, failed to inhibit the effect of ANG I. These results indicate that vascular ANG II receptors and ANG I converting enzyme in the quail may be different from those in mammals. o 1990 Academic press, I~C.
Angiotensin II (ANG II) given exoge- nously increases blood pressure in repre- sentative species of all classes of verte- brates (Nishimura, 1980). However, ANG II causes an immediate vasodepressor re- sponse prior to the vasopressor response in fowl (Moore er al., 1981a; Nishimura et al., 1982; Nakamura et al., 1982). Since only a vasopressor response has been observed after intravenous injection of ANG II in the duck (Wilson and Butler, 1983) and the pi- geon (Evered and Fitzsimons 1981), it is not known whether or not the vasodepressor action of ANG II is a general phenomenon in avian species. In recent years, variants of ANG I from representative species of all classes of nonmammalian vertebrates have been sequenced and synthesized (Khosla et
al., 1983, 1985; Hasegawa et al., 1984), but, in the case of birds, only fowl ANG I has been sequenced. Thus, it appears to be of interest to determine the amino acid se- quence and synthesize ANG I from another avian species, and, using the native an- giotensins, to examine whether a vasode- pressor response is a unique phenomenon in fowl or not.
A number of blockers of the renin- angiotensin system have been utilized in mammals to evaluate the physiological role of this system (Regoli er al., 1974; Peach, 1977). Those blockers that are frequently used include competitive inhibitors of ANG II and inhibitors of ANG I converting en- zyme. These inhibitors have been used in nonmammalian vertebrates with the as-
12 0016~6480/90 $1.50 Copyright Q 1990 by Academic Press, Inc. All rights of reproduction in any form reserved

CARDIOVASCULAR EFFECTS OF ANGIOTENSINS 13
sumption that they are also effective in these animals. However, since ANG II re- ceptors and ANG I converting enzyme of nonmammalian species may differ from those of mammals (Nishimura et al., 1978, 1982; Harper and Stephens, 1985; Takei, 1987; Takei et al., 1988), it remains to be determined whether or not these inhibitors which are active in mammals are actually effective in nonmammalian species.
In the first experiment of this study, we attempted to determine the amino acid se- quence of quail ANG I. Since it was found that quail ANG I had the same amino acid sequence as fowl ANG I, the effects of quail (native), human, and eel ANG I and II on arterial pressure and heart rate were ex- amined in the anesthetized quail. We used a small dose of urethane as an anesthetic, be- cause this drug has been shown to exert a minimal influence on cardiovascular pa- rameters (Maggi and Meli, 1986). Finally we analyzed the effects of competitive in- hibitors of ANG II and inhibitors of ANG I converting enzyme on the cardiovascular effects of ANG I and II. These experiments will help to characterize the ANG II recep- tors and converting enzyme of the quail, and they will provide information on effec- tive inhibitors that will be useful for future experiments designed to evaluate the phys- iological role of the renin-angiotensin sys- tem in this bird.
MATERIALS AND METHODS
Animals
Male Japanese quail, Coturnix coturnix japonica, aged 4 weeks, were purchased from a local dealer (Su- zukei, Toyohashi). They were kept individually in small wire cages (21 X 12 x 17 cm3) under a 8L:16D photocycle at 25 + 1” for more than 2 weeks before use. Quail diet containing 150 meq/kg of Na (w/w) (Nippon Haigo Shiryo, Yokohama) and tap water were freely available until the day of experiment. The birds weighed 107 f 2mg (mean 2 SD, n = 42) at the time of experiment. In addition, more than 25 birds were used as a source of blood and kidney tissues for de- termination of the amino acid sequence of quail ANG I.
Drugs
The ANG I and II used were [Asp’, Va15, SeP] ANG I, and its ANG II, which have been shown to be native to the quail in this study, human [Asp’, Be’, Hiss] ANG I and its ANG II and III, and eel [Asn’, Va15, Gly? ANG I and its ANG II (Hasegawa ef al., 1983a; Khosla et al., 1985). Eel ANG I was synthesized and generously provided by Dr. Mahesh C. Khosla, De- partment of Brain and Vascular Research, Research Institute, Cleveland Clinic Foundation, Cleveland, Ohio. Competitive inhibitors of ANG II used were [Sar’, Ala*] ANG II, [Sari, Thrs] ANG II, and [Sar’, Ilea] ANG II. Inhibitors of ANG I converting enzyme used were des-Pro’ bradykinin (BK) (Naruse et al., 1981) and BK potentiator B, which is an undecapep tide obtained from the venom of Agkistrodon halys blomhofii (Kato and Suzuki, 1971), and SQ14225. SQ14225 (Captopril) was generously donated by San- kyo Pharmaceutical Co. Ltd., Tokyo. All peptides were purchased from the Peptide Institute Inc., Osaka, and other drugs were purchased from Wako Pure Chemicals, Tokyo. All peptides were dissolved in 0.9% NaCl, and stock solutions (100 nmol/ml for an- giotensins and 2 mg/ml for inhibitors) were stored at - 25” until use.
Measurement of Blood Pressure
The birds were lightly anesthetized by an intramus- cular injection of urethane (0.75 g/kg) in the breast region and placed on their side on operating boards. The neck of each bird was lightly fuced between nails, and the body was wrapped gently with a cylindrical cloth. After tracheotomy, a polyethylene catheter (PElO, Clay Adams) was inserted into the right atrium through the right, external jugular vein for injections. Another polyethylene catheter was inserted into the right, common carotid artery for measurement of ar- terial pressure. Care was taken not to damage the va- gus nerve and other nerves that innervate the carotid sinus. Blood loss was slight during the surgery. Be- cause the anesthesia was so light, the birds moved occasionally during measurements. Thus, we utilized a system for measurement of blood pressure that al- lowed the birds to move a little without the cannula slipping out of the artery. The details of the apparatus for the measurements have been reported previously (Takei and Watanabe, 1990).
Determination of Amino Acid Sequence of Quail ANG Z
Collection of blood and kidney tissue. Two millili- ters of blood was withdrawn directly from the jugular vein of conscious quail into a chilled syringe which contained 20 pJ of 15% ammonium EDTA. Five days thereafter, 2 ml of blood was again withdrawn in the same way. Soon after the second withdrawal of blood, birds were decapitated and the kidneys were removed,

14 TAKE1 AND HASEGAWA
weighed, and stored frozen at -25” until use. Col- lected blood was centrifuged at 2700s for 15 min at 4”, and the plasma was stored at - 25” until use.
Preparation of crude angiotensin. Crude angioten- sin was prepared by the modiied version of Boucher’s procedure, as reported previously (Hasegawa ef al., 1983b). Kidney tissue (13.7 g) was homogenized with 14 ml of 5.8 mM 2Na-EDTA and centrifuged at 18,OOOg for 15 min at 2-4”. The supematant was dia- lyzed against 5.8 mM 2Na-EDTA at 4” for 24 hr. The dialysate was then acidified to pH 3.0 with 1 M HCl, and kept at 4” for 30 min. After neutralization, the kidney extract was centrifuged at 18,OOOg as above. The supematant (14 ml), plasma dialyzed against 5.8 mM 2Na-EDTA (55 ml), and 2.5% phenylmethyl sul- fonyl fluoride (0.5 ml) were incubated together with Dowex 5OW-X2 resin (PH 7.4, NH,-form) at 20” for 20 hr and loaded onto a column that contained Dowex 5OW-X2 resin (PH 6.0, NH,-form). The column was washed with 0.2 M ammonium acetate-acetic acid (pH 6.0), 10% acetic acid, and water. The active principle was eluted with 0.1 M diethylamine and 0.5 M ammo- nium hydroxide. The eluate was evaporated to dry- ness, and the activity was determined by the vasopres- sor bioassay in the quail with synthetic [Asp’, Val’, SeP] ANG I as a standard.
Purification and analysis of quail angiotensin. Crude angiotensin was suspended in 0.1% formic acid and passed through a Sep-Pak C,, cartridge (Waters Associates) for desalting. The active principle was eluted with 5% methanol, dried, and bioassayed as above to assess recovery. The dried residue was then dissolved in 0.05 M ammonium fonnate-formic acid (pH 3.0) and loaded onto a column of SP-Sephadex C-25 (26.5 x 0.9 cm). The active principle was eluted with a linear gradient from 0.05 M ammonium for- mate-formic acid (pH 3.0) to 0.5 M ammonium for- mate @H 6.5), and each fraction was bioassayed as above. The fraction with vasoactivity was desalted on a Sep-Pak C,s cartridge and eluted with 8% methanol that contained 0.05% tritluoroacetic acid (TFA). The dried material was further purified by high-per- formance liquid chromatography (HPLC) on a column of TSKgel ODS 80 TM (4.6 mm i.d. x 30 cm, Tosoh, Tokyo) using an eluent composed of methanol, water, and TFA (430:570:1) at a flow rate of 0.5 ml/mitt, and each fraction was bioassayed. The active principle gave the same retention time (52.8 mitt) as that of syn- thetic [Asp’, Val’, SeP] ANG I and was found to be homogeneous by high-performance thin-layer chroma- tography on a plate of silica gel 60 (Merck). Finally, the amino acid sequence of the quail angiotensin was analyzed by stepwise Edman degradation using a gas- phase automated protein sequencer (Applied Biosys- terns, Model 470A) coupled to an HPLC system for identification of the phenylthiohydantoin-derivatized amino acids produced (Applied Biosystems, Model 120 PTH analyzer).
Experimental Protocols for Physiological Studies
Relative pofency of angiotensins. [Asp’, Val’] ANG II, which is native to the quail, was used as the stan- dard angiotensin. In order to examine their relative potencies, quail, human, and eel ANG I and II and human ANG III were injected into birds over 10 set at doses of 0, 5, 10, 20, 50, 100, 200, and 500 pmol/lOO g in a volume of 50 pl. Each injection was followed by flushing of the cannula (dead space: 10 pl) with 30 ~1 of 0.9% NaCl. Each bird received different doses of an- giotensins once, and the order of injection of angioten- sins and doses was determined by the Latin squares method. The injections were repeated after the arterial pressure returned to the level recorded prior to the injections, because repeated injections of 100 pmol/lOO g of quail ANG II in this way generated reproducible effects. BK was also injected after the last injection of angiotensin at a dose of 500 pmoYlO0 g. At the begin- ning and end of injections, 100 pmoYlO0 g of quail ANG II was injected to confii the reproducibility of the response. The average decrease in the response at the end was 4.5 2 2.1% (n = 9).
Effect of inhibitors. The values of ED, of quail ANG I and II determined in the above experiment were used to examine the effects of inhibitors. The injection volume of angiotensin and inhibitors was 50 ~1. The preliminary experiment indicated that [Sari, Iles] ANG II and SQ14225 appear to be the only effec- tive inhibitors of the vasopressor and depressor effects of ANG II and ANG I, respectively. Therefore, in the first experiment, quail ANG II was injected 5 min be- fore and 5 min after injection of 0.9% NaCl or 100 cLg/lOO g of the inhibitors in the following order: [Sari, Ala*] ANG II, [Sar’, Thrs] ANG II, [Sar’, Ile’] ANG II, and SQ14225, in six quail. After injection of [Sar’, Ile’] ANG II, injection of quail ANG II was repeated every 5 min until its effect was fully restored. In the next experiment, quail ANG I was injected 5 min be- fore and 5 min after injection of 0.9% NaCl or 100 pg/lOO g of [Sari, Ile’] ANG II, des-Pro* BK, BK po- tentiator B, and SQ14225, in that order, in another six quail. des-Pro* BK was injected 30 min after injection of [Sar’, Ile’] ANG II, and the injection of ANG I was given every 30 min for up to 2 hr after injection of SQ14225.
We also examined the effects of slow infusion of the inhibitors at a rate of 1 ug/lOO g/l0 pl/min following a priming injection of 5 pg/lOO g in 50 ~1. The injection of quail ANG II was made more than 15 min after the start of infusion of 0.9% NaCl, [Sari, Ala’] ANG II, [Sar’, Th?] ANG II, [Sar’, Ile’] ANG II, or SQ14225 in six birds, and the injection of quail ANG I was made after infusion of 0.9% NaCl, l&r’, Ile’] ANG II, des- Pro* BK, BK potentiator B, or SQ14225 in another six birds. If no inhibition was observed following the in-

CARDIOVASCULAR EFFECTS OF ANGIOTENSINS 15
fusion of an inhibitor, the next infusion was initiated 15 min after the termination of the previous infusion, but in the case of [Sari, Iles] ANG II, 1 hr was allowed to pass before the start of the next infusion.
Analyses of Responses
The ED, of each angiotensin was calculated from a logistic-log transformation based on the changes in ar- terial pressure of all birds at doses between 5 and 500 pmol. The fitting to the logistic curve was executed using the Newton-Raphson algorithm for estimates of maximum likelihood. Since the maximal change, which is necessary for calculation of the logistic-log transformation, did not appear to be attained even at 500 pmol, it was estimated as follows: the change in arterial pressure after 500 pmol of quail ANG II was increased by l-10 mm Hg, and using each value as a maximal change, curve fitting was executed. Then, the value that produced the best fit, judged by the x2 test, was selected as the maximal change of a bird. The ratio of potency of each angiotensin to the standard quail ANG II was calculated from the ratio of values of ED, after the parallelism of the logistic curves was confirmed. In the case of human ANG III, the potency ratio was calculated from the ratio of doses of quail ANG II and human ANG III that produced the same change. The relationship between two variables was examined by linear regression analysis. All other sta- tistical comparisons were made by the paired t test. Statistical significance was determined at P < 0.05. All results are expressed as means f SE of the mean. However, 95% confidence intervals are attached to the
ED, because the mean and the variance are given as logarithms. All computations were carried out on an IBM 4341 computer using an APL interpreter.
RESULTS
Amino Acid Sequence of Quail Angiotensin
The total angiotensin-like vasoactivity af- ter the incubation of kidney extract and plasma was equivalent to 24 nmol of [Asp’, Val’, SeP] ANG I. Thus, the yield was 436 pmol/ml plasma. After the Sep-Pak treat- ment of the crude extract, SP-Sephadex chromatography and HPLC, the total level fell to 16, 14, and 10 nmol, respectively. The analysis by sequential Edman degrada- tion of 2 nmol of the final product showed that the amino acid sequence of quail ANG I is H-Asp-Arg-Val-Tyr-Val-His-Pro- Phe-Ser-Leu-OH.
Vasopressor and Depressor Actions of Angiotensins
A bolus injection of quail ANG II caused
PA
Dose (picomoles/lOO g) FIG. 1. Changes in arterial pressure (PA), mean arterial pressure (MA), and heart rate (HR) after a
bolus injection of native [Asp’, Val’] ANG II in a quail which has been lightly anesthetized with urethane (0.75 g/kg). A vasodepressor response and a subsequent vasopressor response were always observed after injection but the response of the heart rate was not consistent.

16 TAKE1 AND HASEGAWA
an immediate, transitory decrease and a arterial pressure, even at high doses. BK subsequent increase in arterial pressure in caused only a small effect on arterial pres- the quail (Fig. 1). The extent of these sure, even at high doses. BK caused only a changes was dose dependent. The effect of small increase in blood pressure (2.5 + 2.1 ANG II on heart rate was variable and de- mm Hg, n = 6) at 500 pmoYlO0 g. Human pended on dosage and other conditions angiotensins were as potent as their quail (Fig. 1). However, there was an tendency counterparts in terms of both vasopressor for tachycardia to be induced in birds and depressor effects, but eel angiotensins whose normal heart rate was low, while were much less potent (Fig. 2, Table 1). bradycardia occurred in birds whose heart ANG II was generally more potent than rate was high. All variants of ANG I and II ANG I when compared on a molar basis. tested induced both decreases and in- The potency of human ANG III relative to creases in arterial pressure in a dose- quail ANG II was only 1.8 + 0.6% (n = 9). dependent manner (Fig. 2). However, hu- The extent of the hypotension immediately man ANG III had only a small effect on after injection was significantly correlated
Increase
0 IAsp: Val’l ANG ,I . [ASP: 118’1 ANG I,
g) 20-
5 Decrease
5 10 ,
0 [ASP: Val: Ser’l A& ,
. iAS!3: Ile: HIS’, ANG ,
A [Asn: Val: Gly’l ANG ,
5 10 20 50 100 200 500
Dose (picomoles/ 100 g) FIG. 2. Dose-response relationships for the vasopressor (upper) and depressor (lower) effects of
angiotensins in the urethane-anesthetized quail (n = 9). Changes in arterial pressure are corrected for the changes in controls injected with 0.9% NaCl. The values of ED, calculated from the logistic-log transformation are given in Table 1. The resting arterial pressure was 93.0 -’ 10.3 mm Hg (n = 9) before the start of injections and 86.9 k 8.3 mm Hg (n = 9) after the last injection. The vertical bars show SE of the mean. *Significantly (P < 0.05) diierent from the control level.

CARDIOVASCULAR EFFECTS OF ANGIOTENSINS 17
TABLE 1 THE RELATIVE POTENCIES AND VALUES OF EDH, FOR THE VASOPRESSOR AND DEPRESSOR EFFECTS OF
ANGIOTENSIN I AND II (ANG I AND II) IN THE QUAIL (n = 9)
Potency on molar basis (%) EDso (pmol/lOO g)
Angiotensins Pressor Depressor
[Asp’,Val’] ANG II 100 100 85.4 (84.4-86.4) 112.7 (110.7-114.6)
[Asp1,11e5] ANG II 86.6 + 7.4 120.9 f 9.1 104.9 (103.5-106.3) 100.7 (98.6-102.8)
[Asn’,Val’] ANG II 22.3 k 4.3* 37.2 k 5.7* 374.1 (369.6-378.7) 382.9 (375.7-390.2)
[Asp’,Val’,SeP] ANG I 68.9 2 13.3 99.4 2 26.3 121.5 (120.4-122.7) 240.5 (236.2-244.9)
[Asp1,11e5,His~ ANG I 55.5 +- 11.5* 86.0 k 23.9 154.7 (153.2-156.3) 194.3 (191.2-197.5)
[Asnl,ValS,GlyT ANG I 17.4 2 3.5* 39.4 k 9.3* 488.3 (482.M94.2) 407.8 (402.2-413.4)
Note. The relative potency is expressed as a mean k SE of the mean. Since the ED, is given as a logarithm after logistic-log transformation, the variation of the mean is expressed in terms of 95% confidence intervals. *P < 0.05 compared with 100%.
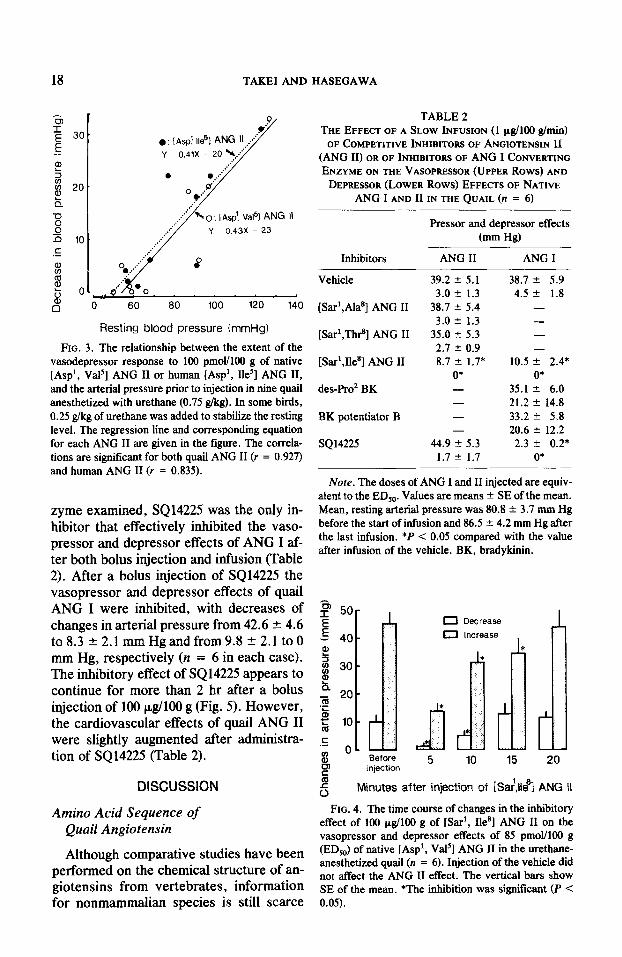
with the arterial pressure prior to injection (Fig. 3).
Effects of Inhibitors of the Renin-Angiotensin System
A bolus injection of competitive inhibi- tors of ANG II (100 pg/lOO g) caused a tran- sitory increase in arterial pressure: 6.4 + 1.1 mm Hg for [Sat-‘, Ala*] ANG II, 8.4 + 1.5 mm Hg for [Sar’, Thr8] ANG II, and 16.9 k 2.3 mm Hg for [Sar’, Ile8] ANG II (n = 6 in each case). The increase disap- peared within 2 min. Injection of the same dose of des-Pro’ BK or of BK potentiator B caused immediate decreases (33.3 + 3.7 and 11.0 k 2.6 mm Hg, respectively) and subsequent increases (13.0 k 4.5 and 42.0 + 4.4 mm Hg, respectively) in arterial pres- sure, whereas SQ14225 caused only a small increase (4.9 + 1.2 mm Hg) in arterial pres- sure (n = 6 in each case). The arterial pres- sure returned to the control level within 5 min after injection of these inhibitors. [Sar’, Ile8] ANG II increased the arterial pressure by 10.7 + 2.4 mm Hg (n = 6) during infu- sion at the rate of 1 pg/lOO g/mm, while all
other inhibitors tested had no effects on ar- terial pressure during infusion.
A bolus injection or infusion of the vehi- cle (0.9% NaCl) did not affect vasopressor and depressor effects of any angiotensin in- jected. Among three competitive inhibitors of ANG II, only [Sar’, Ile81 ANG II inhib- ited vasopressor and depressor effects of ANG II after both bolus injection and infu- sion (Table 2). After a bolus injection, the vasopressor response to quail ANG II was decreased from 44.9 + 4.6 to 13.9 + 2.9 mm Hg and the vasodepressor response from 10.1 k 2.5 to 1.3 2 1.3 mmHg (n = 6 in each case). [Sar’, Ile81 ANG II was also the sole inhibitor capable of blocking the car- diovascular effects of quail ANG I after a bolus injection or an infusion (Table 2). Af- ter a bolus injection of [Sari, Ile8] ANG II, the vasopressor response was decreased from 42.6 + 4.6 to 10.9 + 2.1 mm Hg, and the vasodepressor response from 9.8 + 2.1 to 0 mm Hg (n = 6 in each case). The in- hibitory effect of [Sar’, Ile8] ANG II con- tinued for 15-20 min after a bolus injection of 100 &OO g (Fig. 4).
Among three inhibitors of converting en-
18 TAKEI AND HASEGAWA
4 Y 0.43X - 23 n 10.
.E
8
E 8
0
Resting blood pressure (mmHg)
FIG. 3. The relationship between the extent of the vasodepressor response to 100 pmol/lOO g of native [Asp’, Va15] ANG II or human [Asp’, Be’] ANG II, and the arterial pressure prior to injection in nine quail anesthetized with urethane (0.75 g/kg). In some birds, 0.25 glkg of urethane was added to stabilize the resting level. The regression line and corresponding equation for each ANG II are given in the figure. The correla- tions are signiticant for both quail ANG II (r = 0.927) and human ANG II (r = 0.835).
zyme examined, SQ14225 was the only in- hibitor that effectively inhibited the vaso- pressor and depressor effects of ANG I af- ter both bolus injection and infusion (Table 2). After a bolus injection of SQ14225 the vasopressor and depressor effects of quail ANG I were inhibited, with decreases of changes in arterial pressure from 42.6 2 4.6 to 8.3 2 2.1 mm Hg and from 9.8 2 2.1 to 0 mm Hg, respectively (n = 6 in each case). The inhibitory effect of SQ14225 appears to continue for more than 2 hr after a bolus injection of 100 p&100 g (Fig. 5). However, the cardiovascular effects of quail ANG II were slightly augmented after administra- tion of SQ14225 (Table 2).
DISCUSSION
Amino Acid Sequence of Quail Angiotensin
Although comparative studies have been performed on the chemical structure of an- giotensins from vertebrates, information for nonmammalian species is still scarce
TABLE 2 THE EFFECT OF A SLOW INFUSION (1 p&O0 g/min)
OF COMPETITIVE INHIBITOIS OF ANGIOTENSIN II (ANG II) OR OF INHIBITORS OF ANG I CONVERTING ENZYME ON THE VASOPRESSOR (UPPER Rows) AND
DEPRESSOR (LOWER Rows) EFFECTS OF NATIVE ANG I AND II IN THE QUAIL (n = 6)
Pressor and depressor effects (mm W
Inhibitors ANG II ANG I
Vehicle 39.2 + 5.1 38.7 + 5.9 3.0 k 1.3 4.5 + 1.8
[Sar’,Ala*] ANG II 38.7 f 5.4 -
3.0 k 1.3 -
[Sar’,Thrs] ANG II 35.0 f. 5.3 -
2.7 -t 0.9 -
[Sar1,11e8] ANG II 8.7 f 1.74 10.5 k 2.4* 0* 0*
des-Pro’ BK - 35.1 2 6.0 - 21.2 f 14.8
BK potentiator B - 33.2 2 5.8 - 20.6 2 12.2
SQ14225 44.9 _' 5.3 2.3 -t- 0.2* 1.7 2 1.7 0*
Note. The doses of ANG I and II injected are equiv- alent to the ED,. Values are means -C SE of the mean. Mean, resting arterial pressure was 80.8 + 3.7 mm Hg before the start of infusion and 86.5 + 4.2 mm Hg after the last infusion. *P < 0.05 compared with the value after infusion of the vehicle. BK, bradykinin.
B 50-
i 40- ?
2 I 30-
h 2 20-
$ 5 lo-
t .G
01 _$ ~~~ * % Before
z 5 10 15 20
injection
i
0 Decrease
0 Increase 1 * * *
m
5 Minutes after injection of tSa/,ii& ANG II
FIG. 4. The time course of changes in the inhibitory effect of 100 pg/lOO g of [Sar’, Be81 ANG II on the vasopressor and depressor effects of 85 pmol/lOD g (ED& of native [Asp’, Val’] ANG II in the urethane- anesthetized quail (n = 6). Injection of the vehicle did not affect the ANG II effect. The vertical bars show SE of the mean. *The inhibition was significant (P < 0.05).

CARDIOVASCULAR EFFECTS OF ANGIOTENSINS 19
c3 Decrease 0 Increase * * * * * All &kNl * * *
Before 5 30 60 90 120 Injection
Minutes after injection of XI14225
FIG. 5. The time course of changes in the inhibitory effect of 100 pg/lOO g of SQ14225 on the vasopressor and depressor effects of 113 pmoYlO0 g (ED& of na- tive [Asp’, Val’, Ser9] ANG I in the urethane- anesthetized quail (n = 6). Injection of the vehicle did not affect the ANG I effect. The vertical bars repre- sent SE of the mean. *The inhibition was significant (P < 0.05).
compared to that for mammalian species (Khosla et al., 1983). The paucity of infor- mation is especially apparent in the case of birds since only fowl ANG I has been se- quenced (Nakayama et al., 1973). Compar- ative studies of ANG I show that variations in amino acids occur at positions 1,5, and 9 (Nishimura, 1980). The nature of the amino acids at these positions may affect the af- finity of ANG I for the converting enzyme and the binding affinity of ANG II for its receptors in each species. Thus, it is impor- tant to sequence, synthesize, and use na- tive angiotensins in physiological studies of the renin-angiotensin system in nonmam- malian vertebrates. We have identified quail ANG I as [Asp’, Val’, Se?] ANG I in the present study and, thus, quail ANG I is identical to fowl ANG I. Since quail and fowl belong to the same order of classitica- tion (Galliformes), the biochemical nature also appears to reflect taxonomical proxim- ity which is based on the anatomy.
Vasodepressor Eflect of Angiotensin
Moore et al. (1981a) reported that the va- sodepressor effect of human ANG II which
was observed in the conscious fowl disap- peared after urethane anesthesia. It has been shown that urethane anesthesia is characterized by the preservation of a num- ber of cardiovascular reflexes, but high doses often produce adverse reactions such as hypotension (Maggi and Meli, 1986). Moore et al. (1981a) injected fowl with twice the amount of urethane per body weight that we used for the quail and their dose decreased the blood pressure of the fowl. Thus, the difference in the vasode- pressor response between the quail and fowl may be due to the difference in the levels of anesthesia.
The vasodepressor effect of angiotensin was observed in the fowl (Moore et al., 1981a; Nishimura et al., 1982) and in the quail in this study, but it was not observed in the pigeon (Evered and Fitzsimons, 1981) or the duck (Wilson and Butler, 1983). Because the extent of the vasode- pressor response was significantly corre- lated with the level of resting arterial pres- sure, as observed in this study, it is possible that the difference in the responses of these avian species is due to the difference in the resting level of arterial pressure. The mean, resting arterial pressure before injection was 115 + 5 mm Hg (n = 6) in the con- scious fowl (Nakamura et al., 1982); 93 + 10 mm Hg (n = 9) in the anesthetized quail in this study; 131 + 6 mm Hg (n = 8) in the conscious pigeon (Evered and Fitzsimons, 1981); and approximately 158 mm Hg in the conscious duck (Wilson and Butler, 1983). Thus, the absence of a vasodepressor re- sponse in the pigeon and duck is not due to the low resting blood pressure. Since quail and fowl belong to the order Galliformes, while pigeons (Columbiformes) and ducks (Anseriformes) belong to different orders, it is possible that the vasodepressor response to angiotensin is unique to the gallinacean birds.
Relative Potency of Angiotensins
The present study has shown that eel ANG I and II are much less potent than

20 TAKE1 AND HASEGAWA
their quail and human counterparts in terms of their vasopressor and depressor effects in the quail. Human ANG III had only 2% of the activity of quail ANG II. These re- sults are quite similar to those obtained in the fowl (Nishimura et al., 1982; Table 3). However, the relative potency of angioten- sins in the rat is different from those found with quail and fowl: eel ANG II and human (or rat) ANG III have greater activities in the rat than in these two avian species (Ta- ble 3). Only Watanabe et al. (1977) ob- served a comparably low vasopressor ac- tivity of eel ANG II in the rat. Recently, we obtained preliminary results showing that eel ANG I and II are more effective than human and quail counterparts in terms of vasopressor effect in the eel (Takei, 1987). These results show that native ANG II is most effective in each class of vertebrates, and further suggest that ANG II receptors have evolved in parallel with the evolution of their ligand molecules.
Inhibitors of the Renin-Angiotensin System
In the quail, [Sar’, Ile*] ANG II is an ef- fective inhibitor which antagonizes the vas- cular effects of ANG II, while [Sar’, Ala*] ANG II and [Sar’, Thr*] ANG II, which are potent inhibitors in mammals (Khosla et al., 1974), are ineffective. These results in- dicate that vascular ANG II receptors in the quail may differ from those in mam-
mals, as has already been suggested to be the case in the fowl (Nishimura et al., 1982). In the fowl, however, [Sar’, Ala81 ANG II and [Sar’, Thr*] ANG II appear to be weak but effective inhibitors: [Sar’, Ala81 ANG II infused at a rate of 12.5 pg/kg/min inhibits the vasodepressor action of human ANG II (Moore et al., 198Ib); and [Sar’, The*] ANG II infused at a rate of 5 pg/kg/min decreases by 50% the vaso- pressor response to native ANG II (Ni- shimura et al., 1982). The inhibitory effects of [Sar’, Ala’] ANG II and of [Sar’, Thr*] ANG II are, respectively, 10% and less than 4% of that of [Sar’, Ile’] ANG II in the fowl. In this study, however, both inhibi- tors failed to inhibit the vasopressor and depressor effects of native ANG II when infused at a rate of 10 p,g/kg/min after a priming injection of 50 &kg. It is possible that vascular ANG II receptors in the quail also differ from those in.the fowl.
The effects of ANG II competitive inhib- itors have been examined in other nonmam- malian vertebrates, and the results appear to vary among species. Both [Sar’, Ile’] ANG II and [Sar’, Ala81 ANG II are effec- tive inhibitors in the turtle, Pseudemys scriptu (Stephens, 1981), [Sar’, Ileg] ANG II is effective but [Sar’, Ala*] ANG II and [Sar’, Thr’] ANG II are ineffective in the bullfrog (Harper and Stephens, 1985), and neither [Sar’, Ile*] ANG II nor [Sar’, Th$] ANG II appears to be effective in the eel (Nishimura et al., 1978). Together with the
TABLE 3 THE RELATIVE POTENCIES OF ANGIOTENSINS IN THE RAT, FOWL, AND QUAIL WITH RESPECT TO THE
VASOPRESSOR EFFECT
Animals
Angiotensins Rat” Ratb Rat’ Fowld Qd [Asp’,Val’] ANG II 100 100 100 100 100 [Asp’,Ile5] ANG II 100 74 50 117 87 [Asn’,Val’] ANG II 93 - 25 27 22 [Asp’,Va15,SeP] ANG I - 85 69 56 69 [Asp’,Ile’,Hisg] ANG I 50 54 38 - 56 [Ile’] ANG III 38 - - 2 2
D Regoli et al. (1974); bKhosla et al. (1977); ‘Watanabe et al. (1977); dNisbimura et nl. (1982).

CARDIOVASCULAR EFFECTS OF ANGIOTENSINS 21
results of varying pressor potencies of an- giotensins in different classes of verte- brates, it is suggested that vascular ANG II receptors have evolved during vertebrate phylogeny.
The present study also showed that SQ14225 is a potent inhibitor of the cardio- vascular actions of ANG I, but des-Pro2 BK and BK potentiator B, which are effective inhibitors in mammals, are ineffective. Since the effect of ANG II is not inhibited by SQ14225, ANG I may be active after its conversion to ANG II. These results indi- cate that the converting enzyme of the quail differs from that of mammals. Since amino acid of ANG I at position 9 differs in all classes of vertebrates ever examined (Ni- shimura, 1980), it is possible that convert- ing enzyme, which cleaves ANG II be- tween the eighth and ninth position, is also different among these animals. However, the enzyme may retain common character- istics throughout the vertebrate phylogeny because SQ14225 is an effective inhibitor in all mammalian and nonmammalian verte- brates so far examined (fowl, Nishimura et al., 1982; rat snake, Ho et al., 1984; and bullfrog, Harper and Stephens, 1985).
ACKNOWLEDGMENTS
The authors are very grateful to Dr. Mahesh C. Khosla, Department of Brain and Vascular Research, Research Institute, Cleveland Clinic Foundation, Cleveland, Ohio, for his generous gift of eel angioten- sin I, and to Sankyo Pharmaceutical Co. Ltd. for pro- viding SQ14225. They also thank Dr. Haruo Suzuki and Ms. Etsuko B. Mukouyama, Department of Bio- physical Chemistry, Kitasato University School of Medicine, for their help in operating a protein se- quencer. This investigation was supported in part by a grant-in-aid from the Ministry of Education, Science and Culture of Japan (62748449).
REFERENCES
Evered, M. D., and Fitzsimons, J. T. (1981). Drinking and changes in blood pressure in response to an- giotensin II in the pigeon, Columba livia. J. Phys- iol. (London) 310, 337-352.
Harper, R. A., and Stephens, G. A. (1985). Blockade of the pressor response to angiotensin I and II in
the bullfrog, Rana catesbeiana. Gen. Comp. En- docrinol. 60, 227-235.
Hasegawa Y., Cipolle, M., Watanabe, T. X., Naka- jima, T., Sokabe, H., and Zehr, J. F. (1984). Chemical structure of angiotensin in the turtle, Pseudemys scripta. Gen. Camp. Endocrinol. 53, 159-162.
Hasegawa, Y., Nakajima, T., and Sokabe, H. (1983a). Chemical structure of angiotensin formed with kidney renin in the Japanese eel, Anguilla japo- nica. Biomed. Res. 4, 417-420.
Hasegawa, Y., Watanabe, T. X., Sokabe, H., and Na- kajima, T. (1983b). Chemical structure of an- giotensin in the bullfrog, Rana catesbeiana. Gen. Comp. Endocrinol. 50, 75-80.
Ho, B. M. Y., Sham, J. S. K., and Chiew, K. W. (1984). The vasopressor action of the renin- angiotensin system in the rat snake, Ptyas korros. Gen. Comp. Endocrinol. 56, 313-320.
Kato, H., and Suzuki, T. (1971). Bradykinin- potentiating peptides from the venom of Agkis- trodon halys blomhofJii: Isolation of five bradyki- nin potentiators and the amino acid sequences of two of them, potentiators B and C. Biochemistry 10,972-980.
Khosla, M. C., Bumpus, F. M., Hayashi, Y., Naka- jima, T., Watanabe, T. X., and Sokabe, H. (1977). Synthesis and specific pressor activity of [I-aspartic acid, 5-valine, Pserine] angiotensin I (“fowl angiotensin I”). .I. Med. Chem. 20, 315- 316.
Khosla, M. C., Bumpus, F. M., Nishimura, H., Op- dyke, D. F., and Coviello, A. (1983). Synthesis of nonmammalian angiotensins and their compara- tive pressor properties in dogfish shark, domestic chicken and rat. Hypertension 5, Suppl. V22- V28.
Khosla, M. C., Leese, R. A., Maloy, W. L., Ferreira, A. T., Smeby, R. R., and Burnpus, F. M. (1972). Synthesis of some analogs of angiotensin II as specific antagonists of the parent hormone. J. Med. Chem. 15, 792-795.
Khosla, M. C., Nishimura, H., Hasegawa, Y., and Bumpus, F. M. (1985). Identification and synthe- sis of [I-asparagine, 5-valine, 9-glycine] angioten- sin I produced from plasma of American eel An- guilla rostrata. Gen. Camp. Endocrinol. 57, 223- 233.
Maggi, C. A., and Meli, A. (1986). Suitability of ure- thane anesthesia for physiopharmacological in- vestigations in various systems. Part 2. Cardio- vascular system. Experientia 42, 292-297.
Moore, A. F., Strong, J. H., and Buckley, J. P. (1981a). Cardiovascular actions of angiotensin in the fowl (Gallus domesticus). I. Analysis. Res. Commun. Chem. Pathol. Pharmacol. 32, 423- 445.

22 TAKE1 AND HASEGAWA
Moore, A. F., Strong, J. H., and Buckley, J. P. (1981b). Cardiovascular actions of angiotensin in the fowl (Callus domesricus). II. Angiotensin an- alog agonists and antagonists. Res. Commun. Chem. Pathol. Pharmacol. 32, 447-457.
Nakamura, Y., Nishimura, H., and Khosla, M. C. (1982). Vasodepressor action of angiotensin in conscious chickens. Amer. J. Physiol. 243, H456 H462.
Nakayama, T., Nakajima, T., and Sokabe, H. (1973). Comparative studies on angiotensins. III. Struc- ture of fowl angiotensin and its identification by DNS-method. Chem. Pharm. Bull. 21,2085-2087.
Naruse, M., Yamanami, S., Shuto, K., Sakakibara, S., and Kimura, T. (1981). Potentiating effects of bradykinin-related substances on bradykinin- induced contraction of guinea-pig ileum and hy- potension in rats. Chem. Pharm. Bull. 29, 3369- 3374.
Nishimura, H. (1980). Comparative endocrinology of renin and angiotensin. In “The Renin-Angiotensin System” (J. A. Johnson and R. R. Anderson, Eds.), pp. 29-77. Plenum, New York.
Nishimura, H. Nakamura, Y., Sumner, R. P., and Khosla, M. C. (1982). Vasopressor and depressor actions of angiotensin in the anesthetized fowl. Amer. J. Physiol. 242, H314-H324.
Nishimura H., Norton, V. M., and Bumpus, F. M. (1978). Lack of specific inhibition of angiotensin
II in eels by angiotensin antagonists. Amer. J. Physiol. 235, H95-H103.
Peach, M. J. (1977). Renin-angiotensin system: Bio- chemistry and mechanisms of action. Physiol. Rev. 57, 313-370.
Regoli, D., Park, W. K., and Rouix, F. (1974). Phar- macology of angiotensin. Pharmacol. Rev. 26,59- 123.
Stephens, G. A. (1981). Blockade of angiotensin pres- sor activity in the freshwater turtle. Gen. Camp. Endocrinol. 45, 364-371.
Takei, Y. (1987). In vivo characterization of angioten- sin II receptors and converting enzymes in the quail and eel. 2001. Sci. 4, 1088.
Takei, Y., Stallone, J. N., Nishimura, H., and Cam- panile, C. P. (1988). Angiotensin II receptors in the fowl aorta. Gen. Comp. Endocrinol. 69, 205- 216.
Takei, Y., and Watanabe, T. X. (1990). Vasodepres- sor effect of atrial natriuretic peptides in the quail: Structure-activity relationships. Zool. Sci., 6, in press.
Watanabe, T. X., Sokabe, H., Honda, I., Sakakibara, S., Nakayama, T., and Nakajima, T. (1977). Spe- cific pressor activity and stability of synthetic an- giotensins. Japan. .I. Pharmacol. 27, 137-144.
Wilson, J. X., and Butler, D. G. (1983). Catechol- amine-mediated pressor responses to angiotensin II in the Pekin duck, Anas platyrhynchos. Gen. Comp. Endocrinol. 51, 477489.





























