Embed Size (px)
Citation preview

Journal
of
Vector EcologyJournal of the Society for Vector Ecology
Volume 21, No. 1 June 1996
NE 21( 1): 1- 104 ISSN 1081- 1710

Journal of Vector EcologyVolume 21 - Numberl June 1996
Published by the Society for Vector Ecology
Marc J. Klowden, EditorDivision of Entomology
University of IdahoMoscow, ID 83844- 2339
Phone: ( 208) 885- 7546
Fax: ( 208) 885- 7760
E-mail: mklowden @uidaho.edu
EDITORIAL BOARD
M. S. Mulla, Chair W. J. Crans R. S. Nasci
University of California Rutgers University CDC-DVBID
Riverside, CA, USA New Brunswick, NJ, USA Fort Collins, CO, USA
R. C. Axtell C. I. Dahl M. W. Service
North Carolina State University University of Uppsala Liverpool School of TropicalRaleigh, NC, USA Uppsala, Sweden Medicine, Liverpool, England
N. Becker F. R. Holbrook E. D. Walker
KABS, Waldsee, Germany USDA- ARS Michigan State UniversityLaramie, WY, USA East Lansing, MI, USA
D. R. Barnard
Medical & Veterinary R. S. Lane S. K. Wikel
Entomolology Laboratory University of California Oklahoma State UniversityGainesville, FL, USA Berkeley, CA, USA Stillwater, OK, USA
H. Briegel G. G. Marten S. C. Williams
University ofZurich New Orleans MCD San Francisco State UniversityCH-8057, Zurich, Switzerland New Orleans, LA, USA San Francisco, CA, USA
E. P. Catts L. S. Mian
Washington State University County Vector ControlPullman, WA, USA San Bernardino, CA, USA
The Journal of Vector Ecology is published biannually in June and December. Authors agree to transfer thecopyright for their article to the publisher when the article is accepted for publication. Authorization to photocopyarticles is granted by the Society for Vector Ecology provided the indicated fee is sent to the Copyright ClearanceCenter, Inc., 222 Rosewood Drive, Danvers, MA 01923, USA. Individuals may make single copies of articleswithout charge. Communications relating to editorial matters and manuscripts should be addressed to the Editor.Communications relating to galley proofs, reprints, subscriptions, SOVE membership, change of address, andother matters should be addressed to the Business Office.
Publications and Business Office: Society for Vector Ecology, P. O. Box 87, Santa Ana, California 92702 USA.
Subscription Rates: Membership, including the Journal ofVector Ecology,$ 50.00, Student membership$ 25. 00,Institutional subscription$ 50.00, Sustaining membership$ 100.00

SOCIETY FOR VECTOR ECOLOGY
BOARD OF DIRECTORS
1996 OFFICERS
PresidentB. Fred Beams
President-ElectMichael W. Service
Vice-PresidentRex E. Thomas
Past-PresidentBruce F. Eldridge
Secretary- TreasurerGilbert L. Challet
REGIONAL DIRECTORS
SouthwesternMinoo B. Madon
NorthwesternSteven V. Romney
North CentralThomas R. Wilmot
South CentralCluff E. Hopla
NortheasternDurland Fish
SoutheasternJonathan F. Day
EuropeanMarija Zgomba

JOURNAL OF VECTOR ECOLOGY
Volume 21 JUNE, 1996 Number 1
CONTENTS
Guidelines for Contributors ii
George Brownlee Craig, Jr. Memoriam iii
Submitted Papers
The Blood-Feeding Habits of Aedes Sollicitans( Walker) in Relation to Eastern Equine Encephalitis Virusin Coastal Areas of New Jersey II. Results of Experiments with Caged Mosquitoes and the Effects ofTemperature and Physiological Age on Host Selection .. W. J. Crans, D. A. Sprenger, and F. Mahmood 1
The Blood-Feeding Habits ofAedes Sollicitans( Walker) in Relation to Eastern Equine Encephalitis Virusin Coastal Areas of New Jersey III. Habitat Preference, Vertical Distribution, and Diel Periodicity of Host-Seeking Adults W. J. Crans and D. A. Sprenger 6
Susceptibility of DDT, Dieldrin, and the Development of Malathion Resistance in Anopheles culicifaciesDiptera: Culicidae) in Ambala District of Haryana, India
R. S. Sharma, S. N. Sharma, and R. S. Sharma 14
Patterns in Aedes albopictus ( Skuse) Population Density, Host- Seeking, and Oviposition Behavior inPenang, Malaysia. A. Abu Hassan, C. R. Adanan, and W. A. Rahman 17
Serological Identification of the Predators of Adult Aedes albopictus ( Skuse) ( Diptera: Culicidae) in
Rubber Plantations and a Cemetery in MalaysiaS. Sulaiman, Z. A. Pawanchee, M. A. Karim, J. Jeffery, V. Busparani, and A. Wahab 22
Impact of Microorganisms and Water Quality on the Efficacy of Bacillus Sphaericus Neide Against CulexPipiens Larvae in The Laboratory M. Beck, M. Ludwig, and N. Becker 26
Seasonal Occurrence of Fleas ( Siphonaptera) on Deer Mice ( Peromyscus Maniculatus) in NorthernCalifornia O. R. Larson, R. G. Schwab, and A. Fairbrother 31
Downstream Carry ofLarvicides Used in the Control of Pest Black Flies( Diptera: Simuliidae) in the OrangeRiver, South Africa R. W. Palmer, M. Edwardes, and E. M. Nevill 37
Timing of Larvicide Treatments for the Control of Pest Black Flies( Diptera: Simuliidae) in a Semi-AridEnvironment in South Africa R. W. Palmer, M. Edwardes, and E. M. Nevill 48
Larvicidal Oils Modify the Oviposition Behavior of Culex Mosquitoes .. J. W. Beehler and M. S. Mulla 60
Species Concurrence and the Structure of a Community of Aquatic Insects in Tree Holes R. Barrera 66
Prevalence of Ehrlichia chaffeensis, Rickettsia spp., and Borrelia spp. Infections in Ticks and Rodents atFort Bragg, North Carolina V. B. Solberg, J. G. Olson, L. R. Boobar, J. R. Burge, and P.G. Lawyer 81
The Residual Effects of Alphacypermethrin and Permethrin Against the Dengue Vector Aedes albopictusSkuse) in Wooden Huts in Malaysia
S. Sulaiman, Z. A. Pawanchee, I. Ghauth, A. Wahab, B. Vadiveloo, J. Jeffery, and A. F. Mansor 85
Distribution of Lambornella stegomyiae in Malaysia and its Potential for the Control of Mosquitoes ofPublic Health Importance... I. Vythilingam, S. Mahadevan, K. K. Ong, Abdullah Ghani, and Y. F. Ong 89
Blood Feeding Success of Aedes aegypti and Culex nigripalpus ( Diptera: Culicidae) in Relation toDefensive Behavior of Japanese Quail( Coturnix japonica) in the Laboratory
R. A. Anderson and R. A. Brust 94

JOURNAL OF VECTOR ECOLOGY
Guidelines for Contributors
The Journal of Vector Ecology is an international journal published by the Society forVector Ecology. It is concerned with all aspects of the biology, ecology, and control of arthropodvectors and the interrelationships between the vectors and the disease agents they transmit. Thejournal publishes original research articles and research notes, as well as comprehensive reviewsof vector biology based on presentations at Society meetings. All papers are reviewed by at leasttwo referees who are qualified scientists and who recommend their suitability for publication.Acceptance ofmanuscripts is based on their scientific merit and is the final decision of the editor,but these decisions may be appealed to the editorial board.
Manuscripts intended for publication should be sent to Dr. Marc J. Klowden, Editor,Division of Entomology, University of Idaho, Moscow, Idaho 83844- 2339, U.S. A. Manuscriptsmust be double spaced on a single side of bond paper with 25 mm margins. An original and twoclear copies are required. Draft mode dot matrix type should not be used. Submission of text ona 3- 1/ 2" computer diskette formatted in MS- DOS is encouraged. Microsoft Word, Word Perfect,or Wordstar formats are acceptable, as well as unformatted text files. Please indicate the type offormat on the diskette label. Papers must be organized under the following headings, each on aseparate page, in order: Title page, abstract, text, acknowledgments( if appropriate), referencescited, tables, figure legends, and figures. The title page should contain the names of all authors,their affiliations and the identification and address of the corresponding author. It should alsoinclude a keyword index containing no more than five words that best describe the paper. Pagesshould be numbered consecutively starting with the title page. References should conform to thestyle in recent volumes. Illustrations that are submitted must be ofhigh quality and remain legibleafter reduction.
Page charges, which partially defray the cost of publication, are $ 35 per printed page.
SOVE members who are unable to pay page charges may apply for a limited number of waivers.Reprint charges are shown in the table below.
Pages 1- 4 5- 8 9- 12 13- 16 17- 20
50 copies 60.00 105. 00 150.00 195.00 240.00or less
Each add' l 20.00 38. 00 56.00 74.00 92.0050 copies
Same order
Communications relating to editorial matters and manuscripts should be addressed to theEditor. Communications concerning galley proofs reprints, subscriptions, SOVEmembership, and change of address should be addressed to the Business Office.
PUBLICATIONS AND BUSINESS OFFICE
Society for Vector EcologyP.O. Box 87
Santa Ana, CA 92702
ii

NUMBER 1 JUNE, 1996 VOLUME 21
IN MEMORIAM
George Brownlee Craig, Jr.
1930- 1995
70. 1,N*Via:
41 7 7' w'
0100
101
The world of medical entomology and vector in 1951. Thereupon he entered the University of Illinois
biology lost one of its brightest and most influential to study medical entomology under Dr.William Horsfall.stars recently when Dr. George B. Craig, Jr., Clark During the summers he was an entomologist with theProfessor of Biology at the University of Notre Dame DesPlaines Valley Mosquito Abatement District, and inand a member of SOVE, died of a heart attack while 1952 he earned an M.S. in entomology. At this point,
attending the annual meeting of the Entomological George went on active duty with the U.S. Army, serving
Society ofAmerica in Las Vegas, Nevada. It is difficult with a Preventive Medicine Detachment at Fort Meade,
to imagine any other individual who had a greater MD. He emerged in 1954 as a first lieutenant, and
impact in this field than did George Craig, and he will be resumed his studies under Dr. Horsfall at the University
sorely missed for his scientific accomplishments, his of Illinois, and at the same time held a civil service
teaching and guidance ofstudents, and his great courage position at the U.S. Army Chemical Center atEdgewood,and persistence in championing the cause of public MD. George received a Ph.D. degree in 1956, and in
health entomology at the national level. 1957 joined the faculty at the University ofNotre Dame,George Craig was born in Chicago on July 8, 1930. where he remained for the rest of his life.
He attended school there, including two years at the George Craig had a remarkable career in vectorUniversity of Chicago before entering Indiana biology, resulting in numerous awards and well overUniversity, where he received aB.A. degree in zoology 500 scientific publications. It began with his
iii

VOLUME 21 JUNE, 1996 NUMBER 1
understanding that mosquito biology and control needed with Carlos Finlay, Paul Russell, Karl Meyer, Albertto be more firmly grounded in fundamental studies of Sabin, Fred Soper, Harry Hoogstraal and Will Downs,genetics and reproduction. He established the Vector among others. In October of 1995 he was awarded theBiology Laboratory at Notre Dame in 1957 and Distinguished Achievement Award by SOVE.assembled an illustrious team of workers to address George was a legendary supporter of amateurthese fundamental biological questions. Over the next athletics at all levels, and he held the distinction ofbeingfew years George and his group established an impressive one of the few spectators ever to receive a penalty duringbody of work on the formal genetics of Aedes aegypti a hockey game for unruly conduct. He would drive longwhich will probably never be equaled for any other distances to attend athletic events. In 1984, he drovespecies of insect other than Drosophila melanogaster. non- stop to Los Angeles to attend the Olympic Games.In 1969, George became, in addition to his faculty I once asked George what he considered to be hisresponsibilities, director of the Mosquito Biology Unit greatest accomplishment. He didn' t hesitate. A fewat the International Center for Insect Physiology and years ago, when the Entomological Society of AmericaEcology in Nairobi, Kenya. He held this position until ruled that no person could be senior author on more than1977, and while at ICIPE, planned and executed a one submitted paper, he submitted a second paper heresearch program that extended many of his genetics wanted to give with S. Loren as senior author. It wasfindings into the field in the form of a large series of accepted, but Sophia couldn' t make the meeting, andexperiments on genetic control of Aedes aegypti. George presented it for her.
In the area of mosquito reproduction, George was Through his research, teaching, and dogged pursuitthe discoverer ofmatrone, the first mosquito pheromone. of causes which he believed in, George Craig was anIn the area ofpopulation ecology, George was one of the important force in the lives of all scientists involved inearly participants in what eventually became an entire solving public health problems. He was an activesubdiscipline in the study of tree- hole mosquitoes and participant in nearly all of the important scientific andtheir responses to competition and available resources. government committees and boards that consideredThis was an important field because of the discovery in policy questions relating to public health entomologythe 1960s that Aedes triseriatus was the primary vector for nearly 40 years.of the virus causing LaCrosse encephalitis, one of the Few people that I know can truly be calledmost serious mosquito-borne virus diseases in the United " legendary." However, George was such a person. TheStates. legend grew out of dozens of enormously entertaining
In the mid- 1980' s the Asian tiger mosquito invaded stories about him as a student, as a teacher on a field trip,the United States, and another phase of George' s career in class addressing students, or as an outraged scientistbegin. Not only did he discover the source of the sending a sharply- barbed letter to Washington. In theinfestation through a series of carefully performed long run, though, his status as legendary came fromexperiments on diapause induction, he also became the nearly boundless enthusiasm for entomology, and hisconscience of federal public health officials at a time eagerness to share this enthusiasm with countless friendswhen there was little evidence that many people and colleagues. It is for this that he will be missed theappreciated the threat to public health that this mosquito most.
posed here. Professor Craig is survived by his wife of 42 years,George Craig' s contribution to teaching and training Elizabeth Pflum Craig, a son, James, and two daughters,
of vector biologists equaled in importance his research Patricia Craig and Sarah Craig Peterek. He also leavescontributions. It is safe to say that the majority of all the six grandchildren. The George B. Craig, Jr. Visitingmedical entomologists who are active today have passed Lecture Series in Biological Sciences has beenthrough the Vector Biology Laboratory as a student, established at the University of Notre Dame George' spostdoctoral fellow, or visiting scientist. In recognition honor. Contributions to this memorial series can beofhis teaching, George was given the first distinguished directed to the Department of Biological Sciences,teaching award at Notre Dame. University ofNotre Dame, Notre Dame, IN 46556, or to
George was a fellow of the National Academy of Mrs. George B. Craig, Jr., 19645 Glendale Ave., SouthSciences, the American Academy ofArts and Sciences, Bend, IN 46337.
the Entomological Society of America, and the IndianaAcademy of Sciences. In 1993 he was the recipient ofthe Walter Reed Medal by the American Society of Bruce F. EldridgeTropical Medicine and Hygiene, an award he shared University of California
iv

Hits.
ii
Itw.
Misses.
ii;, y, ' `,
1(''' a
d akJm
xr
rs
I
0Stop the bad guys. Save the good guys. That' s your job, right?And if you don' t succeed, no one wins. Not your
community. Not the environment. Definitely not you. With Altosid®products, everyone wins. That' s because
Altosid hits mosquitoes hard without harming non-target species. By working specifically on fourth instar mosquito
larvae, Altosid offers consistent, highly effective control without disrupting the food chain. Choose from a wide
variety of formulations with up to 150 days of larvicidal control. Just think how cost-effective it will be to spend
less on labor, equipment and adulticides.
For complete details, k- k--. ._
AItosidcall 1- 800-248- 7763 today.
of The smart way to fight mosquitoes.11111111111111
I:. SANDOZ Always read the label before using the product. Sandoz Agro, Inc.. 1300 E. Touhy Ave., Des Plaines, IL 60018. Altosid is a trademark of Sandoz Ltd.© 1995 Sandoz Agro, Inc.

Journal of Vector Ecology 21( 1): 1- 5
The Blood-Feeding Habits ofAedes sollicitans (Walker) in Relation to
Eastern Equine Encephalitis Virus in Coastal Areas of New Jersey!
II. Results of Experiments with Caged Mosquitoes and the Effectsof Temperature and Physiological Age on Host Selection.
W. J. Crans2, D. A. Sprenger3, and F. Mahmood2
New Jersey Agricultural Experiment Station Publication No. D-40101- 02- 95supported by U.S. Hatch Act funds with partial support from the
New Jersey State Mosquito Control Commission.2Department ofEntomology, Cook College, Rutgers University,
P.O. Box 231, New Brunswick, NJ 08903.
Corpus Christi Health Department, Vector Control Division,
3041 Morgan Avenue, Corpus Christi, TX 78404.
Received 27 December 1990; Accepted 24 October 1995
ABSTRACT: Because of the extremely low incidence of avian feedings reported from field collectedAedes sollicitans( Walker), host-seeking mosquitoes were collected in the field, placed into outdoor cagesand offered a restrained bird ( Japanese quail) and restrained mammal ( guinea pig) to determine if thespecies would accept or reject an avian host. No significant difference in feeding response was observedbetween groups ofmosquitoes offered only the bird or the mammal. A majority of the mosquitoes acceptedthe host that was offered suggesting that Ae. sollicitans is an opportunistic blood- feeder rather than a fixedmammalophilic species. When mosquitoes were offered a choice of a mammal or bird, one- third of the
mosquitoes accepted the bird as the blood-meal host when the mammal was present as an alternative.
Physiological age dissections showed that both nulliparous and parous mosquitoes fed preferentially on themammalian host but significantly more parous mosquitoes accepted the guinea pig when a choice wasavailable. Reduced temperatures decreased the number of mosquitoes that attempted to blood- feed under
laboratory conditions but did not alter host selection in these tests. Results are discussed in relation toacquisition of eastern equine encephalitis virus and the role ofAe. sollicitans as an epidemic vector of the
disease.
Keyword Index: Aedes sollicitans, host preference, bird, eastern equine encephalitis.
INTRODUCTION have shown that this species only rarely feeds on avianhosts in nature ( Thompson et al. 1963, Crans 1964,
Considerable evidence has been accumulated to Edman and Downe 1964, Schaefer and Steelman 1969,
incriminate Aedes sollicitans( Walker) as the epidemic Edman 1971). Crans et al.( 1990) tested numerous wild-
vector of eastern equine encephalitis ( EEE) virus in caught specimens collected from study sites in NewNew Jersey( Kandle 1960, Hayes et al. 1962, Goldfield Jersey where EEE virus was known to be enzootic. Theet al. 1966, Crans 1977, Crans et al. 1986). Birds, results indicated that 98. 2% of the specimens tested had
however, are considered to be the primary amplification fed on a mammalian host and only 1. 4% fed on birds.
hosts for the virus( Stamm et al. 1962, Stamm 1963) and B lood- feeding patterns determined solely byblood-meal identification studies with Ae. sollicitans precipitin tests on wild-caught specimens may not be

2 Journal of Vector Ecology June, 1996
indicative of the true host preference for a given species. hosts in 1 ft3 cages that were set on a portable table in the
Host availability within collection sites strongly old field where the mosquitoes had been collected. Four
influences the range ofpotential hosts that are ultimately cages were used in each feeding trial. The experimentalexhibited in the results. The methods used to collect the design included a guinea pig( representing amammalianengorged specimens and the habitat that is sampled host) in one cage, a Japanese quail ( representing an
could further bias the sample. Nayer and Sauerman avian host) in another, and a combination of both
1977) successfully fed Ae. sollicitans on a chicken, animals in the remaining two cages. A purposeful effortdove, and owl during experiments designed to test the was made to eliminate defensive behavior of the host as
effects of blood source on oocyte development. Their a variable during the trials. As a result, the animals werestudies suggest that Ae. sollicitans may be more of a restrained separately in nylon stockings and were firmlycatholic feeder than the literature indicates. taped to the floor of the cage to minimize movement.
In an effort to explore avian feeding by Ae. This design was repeated on 11 separate dates.
sollicitans, a series of experiments were conducted to Two hours before sunset, 100 mosquitoes were
determine 1) if the mosquito would accept a bird as a released into each cage and allowed to engorge on the
blood-meal host when no other host was present, 2) if restrained animals throughout the twilight period. After
the mosquito would accept a bird when a mammal was four hours of exposure, the animals were removed and
present as an alternative, and 3) if the ratio ofmammalian the mosquitoes were anesthetized, placed into vials, and
to avian feedings approached the results obtained by frozen on dry ice. Individual mosquitoes were examinedprecipitin testing. The animals were restrained to for physiological age and host selection at a later date.
eliminate any variability in defensive behavior by the Blood meals were identified using the agar gel diffusionhosts which may have interfered with the success of technique described by Crans ( 1969). Parity wasengorgement once the mosquitoes initiated blood determined by examining tracheolation of the ovaries asfeeding. In addition, the effects of temperature and described by Detinova( 1962).physiological age were measured to determine if these
factors influenced host selection during the experiments. Host Selection at Controlled Temperatures
Since the experiments were designed to investigate The F1 progeny ofwild-caught Ae. sollicitans wereavian acceptance by Ae. sollicitans, unavoidable biases used to determine host selection at controlled
in methodology were purposely shifted in favor of the temperatures. The mosquitoes were collected by themammalian host( host initially used to attract specimens methods described earlier, given a blood meal on a
for the tests, comparative size of the caged animals used guinea pig in the laboratory, and allowed to lay eggsin the experiments, and source of the blood meal that on moist cotton. Adult mosquitoes for the experi-
produced F1 progeny). ments were then reared from the eggs by the methodsof Khoo and Sutherland ( 1983). A 10% sucrose
MATERIALS AND METHODS solution was made available as a source of nutrition
while the specimens were being held for blood-feedingHost Selection at Ambient Temperatures tests.
Once each week from July to October 1982 and Host selection experiments were conducted in an
again in July and August 1983, 400 Ae. sollicitans of incubator at three controlled temperatures ( 15°, 20°,
unknown physiological age were collected by aspiration and 27° C). Each temperature regime was replicated
as they attempted to bite the authors at a coastal site in five times. On the day of each test, a guinea pig and aNew Jersey. Collections were made during the late Japanese quail were restrained separately in a nylonafternoon in an old field in Belleplain State Forest, Cape stocking and taped, side by side, to the floor of a 1 ft3May County, New Jersey, where EEE virus had been cage, duplicating the methodology used in the fieldisolated from field-collected specimens in 1982( Crans trials. Fifty 4- 6 day old Ae. sollicitans were placed in aet al. 1986). Care was taken to assure that none of the paper pint carton just prior to each test. The restrained
specimens had successfully penetrated skin prior to animals and the mosquitoes were then allowed to
capture. The host-seeking mosquitoes were placed, 100 acclimate to the test temperature for two hours. Pans
each, in paper pint cartons and sealed in plastic bags of water were placed in the incubator where the tests
with moist paper toweling to maintain humidity until the were conducted and the cages were draped with cheese
tests were run. In no cases were the mosquitoes held cloth to maximize humidity in the artificial environ-longer than two hours from collection to exposure to the ment.
test animals. After acclimation, 50 mosquitoes were introduced
Mosquitoes were exposed to potential blood-meal into each cage with the restrained animals and allowed
June, 1996 Journal of Vector Ecology 3
to engorge over a four-hour period. When the test was TABLE 2 lists the results of 22 tests wherecompleted, the animals were released, the entire cage mosquitoes were exposed simultaneously to both animalswas placed in a freezer, and the mosquitoes were and then dissected to correlate physiological age withseparated into vials. Instead ofusing antisera to determine host selection. Of those mosquitoes that accepted athe host, each blood meal was smeared on a microscope host, 66.4% fed on the guinea pig and 33. 6% fed on theslide, stained with giemsa, and identified to host by the quail. Ovarian dissections showed that out of the totalpresence or absence of nuclei in the red blood cells. 981 mosquitoes that accepted a blood meal from one of
the hosts, 343 ( 35%) were nulliparous and 638 ( 65%)RESULTS were parous. To determine ifphysiological age affected
host selection, nulliparous and parous mosquitoes wereHost Selection at Ambient Temperatures tested by a 2x2 Yates x2 contingency test. The results
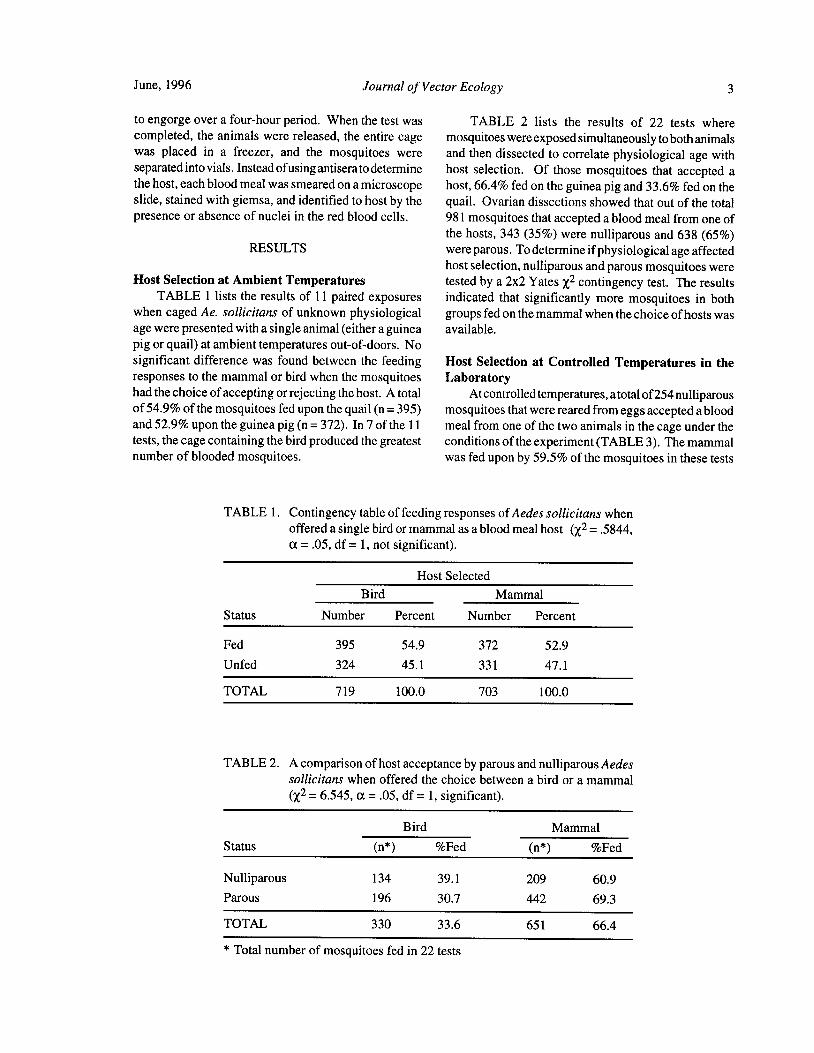
TABLE 1 lists the results of 11 paired exposures indicated that significantly more mosquitoes in bothwhen caged Ae. sollicitans of unknown physiological groups fed on the mammal when the choice ofhosts wasage were presented with a single animal( either a guinea available.
pig or quail) at ambient temperatures out-of-doors. Nosignificant difference was found between the feeding Host Selection at Controlled Temperatures in theresponses to the mammal or bird when the mosquitoes Laboratoryhad the choice ofaccepting or rejecting the host. A total At controlled temperatures, a total of254 nulliparousof54.9% of the mosquitoes fed upon the quail( n= 395) mosquitoes that were reared from eggs accepted a bloodand 52. 9% upon the guinea pig( n= 372). In 7 of the 11 meal from one of the two animals in the cage under thetests, the cage containing the bird produced the greatest conditions of the experiment( TABLE 3). The mammalnumber of blooded mosquitoes. was fed upon by 59.5% of the mosquitoes in these tests
TABLE 1. Contingency table of feeding responses ofAedes sollicitans whenoffered a single bird or mammal as a blood meal host ( x2=. 5844,
a=. 05, df= 1, not significant).
Host Selected
Bird Mammal
Status Number Percent Number Percent
Fed 395 54.9 372 52. 9
Unfed 324 45. 1 331 47. 1
TOTAL 719 100.0 703 100.0
TABLE 2. A comparison of host acceptance by parous and nulliparous Aedessollicitans when offered the choice between a bird or a mammalx2= 6.545, a= . 05, df= 1, significant).
Bird Mammal
Status n*) Fed n*) Fed
Nulliparous 134 39. 1 209 60.9
Parous 196 30.7 442 69. 3
TOTAL 330 33. 6 651 66.4
Total number of mosquitoes fed in 22 tests
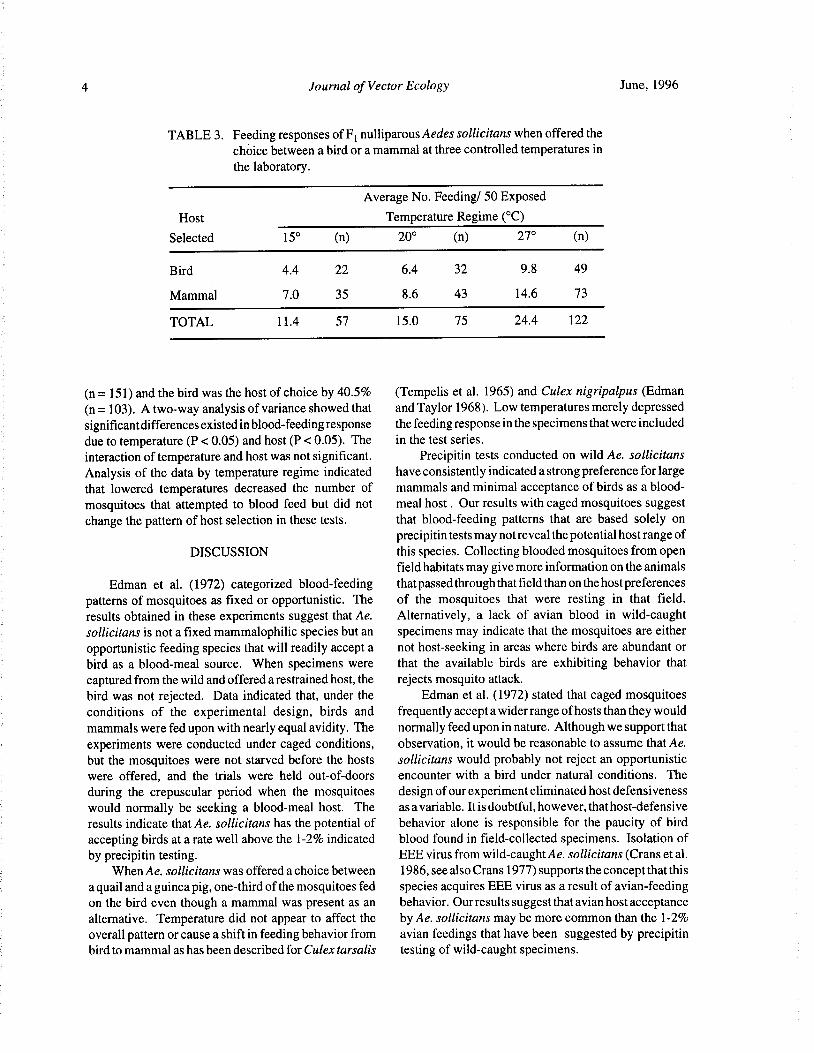
4 Journal of Vector Ecology June, 1996
TABLE 3. Feeding responses of Fl nulliparous Aedes sollicitans when offered thechoice between a bird or a mammal at three controlled temperatures in
the laboratory.
Average No. Feeding/ 50 Exposed
Host Temperature Regime(° C)
Selected 15° ( n) 20° n) 27° n)
Bird 4. 4 22 6.4 32 9. 8 49
Mammal 7. 0 35 8.6 43 14.6 73
TOTAL 11. 4 57 15. 0 75 24.4 122
n= 151) and the bird was the host of choice by 40.5% ( Tempelis et al. 1965) and Culex nigripalpus (Edman
n= 103). A two-way analysis of variance showed that and Taylor 1968). Low temperatures merely depressedsignificant differences existed in blood-feeding response the feeding response in the specimens that were includeddue to temperature( P< 0.05) and host( P< 0. 05). The in the test series.
interaction of temperature and host was not significant. Precipitin tests conducted on wild Ae. sollicitans
Analysis of the data by temperature regime indicated have consistently indicated a strong preference for largethat lowered temperatures decreased the number of mammals and minimal acceptance of birds as a blood-
mosquitoes that attempted to blood feed but did not meal host. Our results with caged mosquitoes suggest
change the pattern of host selection in these tests. that blood- feeding patterns that are based solely onprecipitin tests may not reveal the potential host range of
DISCUSSION this species. Collecting blooded mosquitoes from openfield habitats may give more information on the animals
Edman et al. ( 1972) categorized blood-feeding that passed through that field than on the host preferences
patterns of mosquitoes as fixed or opportunistic. The of the mosquitoes that were resting in that field.results obtained in these experiments suggest that Ae. Alternatively, a lack of avian blood in wild-caughtsollicitans is not a fixed mammalophilic species but an specimens may indicate that the mosquitoes are eitheropportunistic feeding species that will readily accept a not host-seeking in areas where birds are abundant orbird as a blood-meal source. When specimens were that the available birds are exhibiting behavior thatcaptured from the wild and offered a restrained host, the rejects mosquito attack.
bird was not rejected. Data indicated that, under the Edman et al. ( 1972) stated that caged mosquitoes
conditions of the experimental design, birds and frequently accept a wider range of hosts than they wouldmammals were fed upon with nearly equal avidity. The normally feed upon in nature. Although we support thatexperiments were conducted under caged conditions, observation, it would be reasonable to assume that Ae.
but the mosquitoes were not starved before the hosts sollicitans would probably not reject an opportunistic
were offered, and the trials were held out-of-doors encounter with a bird under natural conditions. The
during the crepuscular period when the mosquitoes design of our experiment eliminated host defensiveness
would normally be seeking a blood-meal host. The asavariable. It is doubtful, however, that host-defensive
results indicate that Ae. sollicitans has the potential of behavior alone is responsible for the paucity of birdaccepting birds at a rate well above the 1- 2% indicated blood found in field-collected specimens. Isolation of
by precipitin testing. EEE virus from wild-caught Ae. sollicitans( Crans et al.
When Ae. sollicitans was offered a choice between 1986, see also Crans 1977) supports the concept that this
a quail and a guinea pig, one- third of the mosquitoes fed species acquires EEE virus as a result of avian- feedingon the bird even though a mammal was present as an behavior. Our results suggest that avian host acceptance
alternative. Temperature did not appear to affect the by Ae. sollicitans may be more common than the 1- 2%overall pattern or cause a shift in feeding behavior from avian feedings that have been suggested by precipitinbird to mammal as has been described for Culex tarsalis testing of wild-caught specimens.

June, 1996 Journal of Vector Ecology 5
Acknowledgments Edman, J. D. and D. J. Taylor. 1968. Culex nigripalpus:
Seasonal shift in the bird-mammal feeding ratio inSincere appreciation is extended to the Cape May a mosquito vector of human encephalitis. Science
County Mosquito Control Commission and their staff 161: 67- 68.for providing storage space for field equipment and care Edman, J. D., L. A. Webber, and H. W. Kale. 1972.for the animals used in these experiments. Without their Host- feeding patterns of Florida mosquitoes. II.help and interest, the field aspects of this study would Culiseta. J. Med. Entomol. 9: 429- 434.have been difficult to conduct on a weekly basis. Goldfield, M., O. Sussman, and R. P. Kandle. 1966. A
progress report on arbovirus studies in New Jersey.REFERENCES CITED Proc. N. J. Mosq. Exterm. Assoc. 53: 47- 51.
Hayes, R. 0., L. D. Beadle, A. D. Hess, O. Sussman,Crans, W. J. 1964. Continued host preference studies and M. J. Bonese. 1962. Entomological aspects
with New Jersey mosquitoes. Proc. N. J. Mosq. of the 1959 outbreak of eastern encephalitis inExterm. Assoc. 51: 50-58. New Jersey. Amer. J. Trop. Med. and Hyg. 11: 115-
Crans, W. J. 1969. An agar diffusion method for the 121.
identification of mosquito blood meals. Mosq. Kandle, R. P. 1960. Eastern encephalitis in New Jersey.News 29: 563- 566. Proc. N.J. Mosq. Exterm. Assoc. 47: 11- 15.
Crans, W. J. 1977. The status ofAedes sollicitans as an Khoo, B. K. and D. J. Sutherland. 1983. The suscepti-epidemic vector of eastern equine encephalitis in bility status ofAedes sollicitans adults to topicallyNew Jersey. Mosq. News 37: 85- 89. applied malathion. Mosq. News 43: 441- 444.
Crans, W. J., J. McNelly, T.L. Schulze, and A. J. Main. Nayar, J. K. and D. M. Sauerman. 1977. The effects of1986. Isolation ofeastern equine encephalitis virus nutrition on survival and fecundity in Floridafrom Aedes sollicitans during an epizootic in mosquitoes. Part 4. Effects of blood source onsouthern New Jersey. J. Amer. Mosq. Control oocycte development. J. Med. Entomol. 14: 167-Assoc. 2: 68- 72. 174.
Crans, W. J., L. J. McCuiston, and D. A Sprenger. 1990. Schaefer, R. E. and C. D. Steelman. 1969. DeterminationThe blood- feeding habits of Aedes sollicitans of mosquito hosts in salt marsh areas of Louisiana.Walker) in relation to eastern equine encephalitis J. Med. Entomol. 6: 131- 134.
in New Jersey. I. Host selection in nature determined Stamm, D. D. 1963. Susceptibility of bird populationsby precipitin tests on wild-caught specimens. Bull. to eastern, western and St.Louis encephalitis viruses.
Soc. Vector Ecol. 15( 2): 144- 148. Proc. XIII Intern. Ornithol. Congr. 1: 591- 603.Detinova, T. S. 1962. Age-grouping methods in Diptera Stamm, D. D., R. W. Chamberlain, and W. D. Sudia.
of medical importance with special reference to 1962. Arbovirus studies in south Alabama, 1957-
some vectors ofmalaria. WHO Monograph no. 47, 1958. Am. J. Hyg. 76: 61- 81.Geneva, 216 pp. Tempelis, C. H., W. C. Reeves, R. E. Bellamy, and M.
Edman, J. D. 1971. Host-feeding patterns of Florida F. Lofy. 1965. A three-year study of the feedingmosquitoes. I. Aedes, Anopheles, Coguillettidia, habits of Culex tarsalis in Kern County, California.Mansonia, and Psorophora. J. Med. Entomol. 18: Amer. J. Trop. Med Hyg. 21: 238- 245.687- 695. Thompson, E. G., D. E. Hayes, and K. W. Ludham.
Edman, J. D. and A. R. Downe. 1964. Host-blood 1963. Notes on the feeding habits of Aedessources and multiple feeding habits of mosquitoes sollicitans in the Chincoteaque- Assateaque Islandin Kansas. Mosq. News 24: 154- 160. area of Virginia. Mosq. News 23: 297- 298.

Journal of Vector Ecology 21( 1): 6- 13
The Blood-Feeding Habits ofAedes sollicitans (Walker) in Relation toEastern Equine Encephalitis Virus in Coastal Areas of New Jersey!
III. Habitat Preference, Vertical Distribution, and
Diel Periodicity of Host-Seeking Adults
W. J. Crans2 and D. A. Sprenger3
1New Jersey Agricultural Experiment Station Publication No. D-40101- 03-95supported by U.S. Hatch Act funds with partial support from the
New Jersey State Mosquito Control Commission.2Department ofEntomology, Cook College, Rutgers University,
P.O. Box 231, New Brunswick, NJ 08903.
3Corpus Christi Health Department, Vector Control Division,
3041 Morgan Avenue, Corpus Christi, TX 78404.
Received 27 December 1990; Accepted 3 April 1992
ABSTRACT: The relative abundance of host-seeking Aedes sollicitans( Walker) was compared amonghabitats that included a deciduous forest, a coniferous forest, a cedar swamp, and an abandoned field insouthern New Jersey. Collections were made using dry ice baited CDC traps operated from dusk to dawnwithout a light source. Results indicated that significantly more mosquitoes were collected from the field;collections in wooded habitats and roadways were considerably lower. Fewest mosquitoes were collectedin the cedar bog where eastern equine encephalitis ( EEE) virus is believed to be most prevalent. Thetemporal and vertical distribution ofhost- seeking Ae. sollicitans were determined using dry ice baited CDCtraps placed at 0.5, 2.5, and 6.0 m above ground level on the edge of an open field and a deciduous forest.Over the course of the investigation, 82% of the total collection was taken at the lowest level, 17% at the
mid-level, and only 1% at the highest level. Host-seeking activity occurred in bimodal crepuscular peaksduring the summer and early fall. Later in the season, host-seeking was reduced to a single peak prior tosunset, presumably the result of decreasing temperatures. Results suggest that habitat preference andvertical distribution strongly influence the mammalian blood-feeding habits of this species in nature. Thereported paucity of avian blood meals in this mosquito may be due to a spatial disassociation between themosquito and tree- roosting birds. The results are discussed in relation to Ae. sollicitans potential as a vectorof EEE virus.
Keyword Index: Aedes sollicitans, host preference, vertical distribution, eastern equine encephalitis
INTRODUCTION showed that Ae. sollicitans is an opportunistic feeder
and will readily accept a bird as a blood-meal host whenBlood-meal identification studies with Aedes given the opportunity.
sollicitans (Walker) based on precipitin tests of wild- Host availability can affect the range of hosts thatcaught adults indicate that the species is strongly oriented are utilized by a particular mosquito species in naturetoward mammals and only rarely feeds on birds in and result in a feeding pattern that is determined bynature( Thompson et al. 1963, Crans 1964, Edman and repeated contact with a particular host rather than fixed
Downe 1964, Schaefer and Steelman 1969, Edman feeding behavior ( Edman et al. 1972). Availability1971, Crans et al. 1990). Crans et al.( 1996), however, requires spatial and temporal synchrony between the

June, 1996 Journal of Vector Ecology 7
potential hosts and the host- seeking mosquitoes. Spatial transformation was utilized to meet the requirements of
relationships can occur within horizontal zones or normality, additivity, and homoscedasticity in the test.habitats and in vertical strata or heights within those Differences in the means were tested using Fischer' shabitats. Temporal relationships may be either diel or protected LSD method at P< 0.05.
seasonal. This paper reports the results of a study toinvestigate the habitat, vertical distribution, and diel Vertical Distribution and Diel Periodicity of Host-rhythms ofhost- seeking Ae. sollicitans in relation to the Seeking Adultsavailability of hosts. Particular attention has been Aedes sollicitans were sampled weekly from latefocused on birds that roost in the forest canopy because July until mid-October, 1982, on the edge of a 1 ha fieldof their role as reservoirs of eastern equine encephalitis in the Belleplain State Forest where large populations ofEEE) virus( Stamm et al. 1962, Stamm 1963). Ae. sollicitans were observed to rest. The wooded
border of the field was composed ofMyrica heterophylla,MATERIALS AND METHODS Prunus serotina, Viburnum dentatum, Rhus copallina,
Sassafras albidum, and Diospyros virginiana. Six
Habitat Distribution of Host-Seeking Adults miniature CDC traps baited with dry ice were operatedStudies were conducted in the southern-most portion each week. The traps were randomly assigned to three
of the Belleplain State Forest, Cape May County, New height levels( 0.5, 2. 5, and 6. 0 m) in two replicate plots
Jersey. The area represents a peninsula composed of along the edge of the field and the deciduous forest.pine- oak forest bordered on its southern extremes by Traps were suspended from the branches of trees directlydeciduous, lowland tree species. The lowland forest on the ecotone. A pulley system was used to hoist theadjoins a tidal salt marsh which extends south, east, and traps operated at the 6.0 m level.
west of the study site. Large populations ofAe. sollicitans Each trap was baited with a 2. 5 kg block of dry icemove into the area from adjacent marshland that has at the onset of the collection period, and fresh blocksbeen diked for the production of salt hay. were added over the 25- hour collection period as needed.
Mosquitoes were collected using miniature CDC At one- hour intervals, collection sleeves were retrievedtraps ( Sudia and Chamberlain 1962), baited with and immediately replaced with empty duplicates. Theapproximately 3 kg of dry ice, and operated throughout mosquitoes were frozen on dry ice and placed in vialsthe collection period without a light source. Nine for later identification and sorting. Temperature andcollections were made at weekly intervals from mid- relative humidity were recorded for each hourlyJune to September 1983. On each collection date, nine collection using a Bendix model 566 psychrometer.traps were suspended at fixed locations at a height of0.5 The first hour' s collections were excluded from them and operated from one hour before sunset to one hour analysis because of the initial disturbance and attractionafter dawn. Two intersecting roads served as transects of resting mosquitoes during the erection of the traps.for the experiment. The exact time for beginning the hourly collections
The habitats sampled in this study included: 1) a was determined by the time at which civil twilightdeciduous forest composed primarily of Liquidambar occurred on that day. In this way, collection periods onstyraciflua, Acer rubrum, and Nyssa sylvatica; 2) a different collection dates could be compared in relationconiferous forest, dominated by Pinus strobus; 3) a to mosquito activity at sunrise and sunset.dense cedar swamp where Chamaecyparis thyoides was A three way analysis of variance was performed tothe main tree species; and 4) an upland field composed test the effects of height, time of day, and temperatureofnumerous grasses and herbaceous plants. Four of the upon trap catch. Because the length of scotophase
traps used in this study were placed along the shoulders increased during the course ofthis study, hourly samplesof the intersecting roads to represent a ubiquitous habitat could not be meaningfully compared by using recordedand determine if significant differences existed within times. Rather, collections were grouped into diel periods
the study area that could bias the comparison among ( PM of day 1, sunset, night, sunrise, and AM of day 2),habitats. A trap was also placed at the end of the and a mean was taken for the hourly catch within eachtransect on the edge of the nonbreeding Spartina diel period. Similarly, samples were grouped into onealterniflora salt marsh that surrounded the upland of three temperature ranges, determined by thepeninsula to determine if proximity to the salt marsh temperature at which the collection was taken. The
biased the results. groupings( 10- 15°, 16- 20°, and 21- 32° C) were based, in
Data were analyzed using an analysis of variance part, on the relative levels of activity for Ae. sollicitanson the log transformed data, log( x+ 1). One was added as observed by Rudolfs ( 1923). Trap data wereto each catch to avoid log transformations of 0. Log transformed to loge( x+ 1) to approximate the conditions
8 Journal of Vector Ecology June, 1996
ofnormality, additivity, and homoscedasticity required Vertical Distribution of Host-Seeking Adultsby the analysis. Catch was increased by 1 to avoid log A total of 8, 875 Ae. sollicitans was collected in
transformation of 0. Means are reported as modified 1, 572 separate samples from the traps operated at defined
geometric means. Differences between means were levels along the ecotone of a field and a forest. A three-tested using Fisher' s protected LSD method at P< 0.05. way analysis of variance indicated that the effects of
height, time of day, temperature, and all interactionsRESULTS were significant( P< 0.001). Differences in catch among
heights were great. Over the course of the investigation,
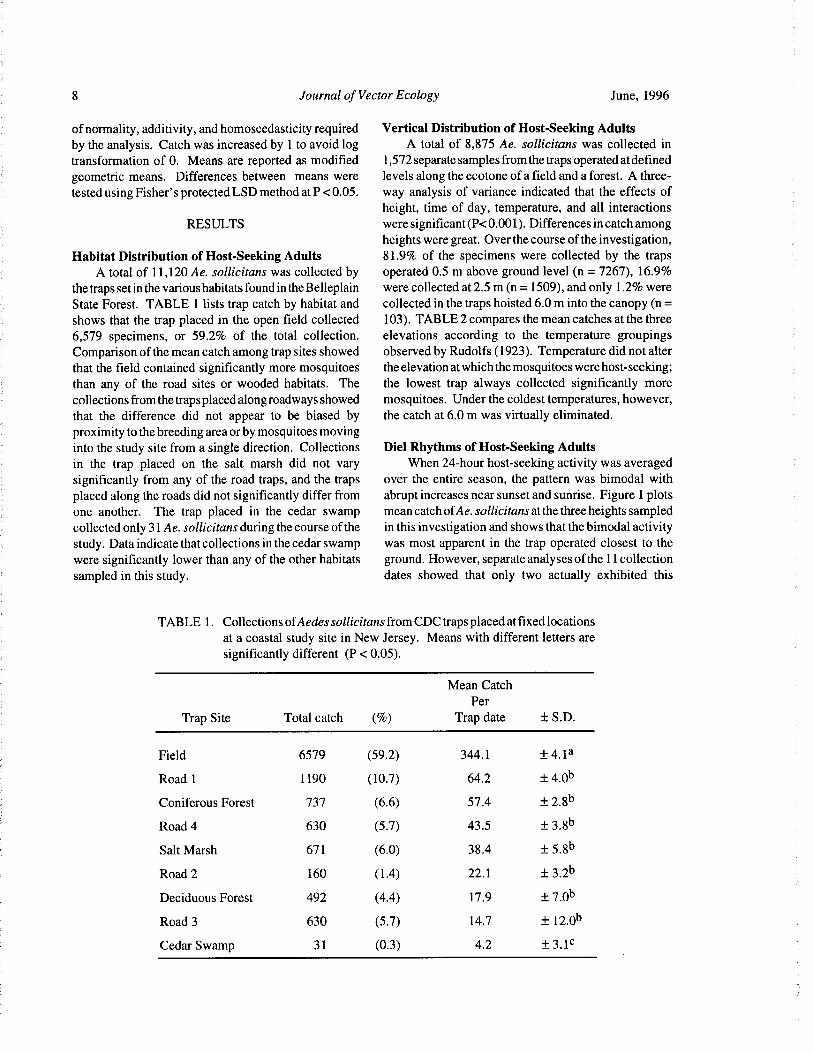
Habitat Distribution of Host-Seeking Adults 81. 9% of the specimens were collected by the trapsA total of 11, 120 Ae. sollicitans was collected by operated 0. 5 m above ground level ( n= 7267), 16.9%
the traps set in the various habitats found in the Belleplain were collected at 2. 5 m( n= 1509), and only 1. 2% were
State Forest. TABLE 1 lists trap catch by habitat and collected in the traps hoisted 6. 0 m into the canopy( n=shows that the trap placed in the open field collected 103). TABLE 2 compares the mean catches at the three
6, 579 specimens, or 59.2% of the total collection. elevations according to the temperature groupingsComparison of the mean catch among trap sites showed observed by Rudolfs( 1923). Temperature did not alter
that the field contained significantly more mosquitoes the elevation at which the mosquitoes were host-seeking;than any of the road sites or wooded habitats. The the lowest trap always collected significantly morecollections from the traps placed along roadways showed mosquitoes. Under the coldest temperatures, however,
that the difference did not appear to be biased by the catch at 6. 0 m was virtually eliminated.
proximity to the breeding area or by mosquitoes movinginto the study site from a single direction. Collections Diel Rhythms of Host-Seeking Adultsin the trap placed on the salt marsh did not vary When 24- hour host- seeking activity was averaged
significantly from any of the road traps, and the traps over the entire season, the pattern was bimodal with
placed along the roads did not significantly differ from abrupt increases near sunset and sunrise. Figure 1 plots
one another. The trap placed in the cedar swamp mean catch ofAe. sollicitans at the three heights sampled
collected only 31 Ae. sollicitans during the course of the in this investigation and shows that the bimodal activity
study. Data indicate that collections in the cedar swamp was most apparent in the trap operated closest to thewere significantly lower than any of the other habitats ground. However, separate analyses of the 11 collection
sampled in this study. dates showed that only two actually exhibited this
TABLE 1. Collections ofAedes sollicitans from CDC traps placed at fixed locations
at a coastal study site in New Jersey. Means with different letters aresignificantly different ( P< 0.05).
Mean Catch
Per
Trap Site Total catch (%) Trap date ± S. D.
Field 6579 59. 2) 344. 1 4. 1a
Road 1 1190 10.7) 64.2 4.0b
Coniferous Forest 737 6.6) 57.4 2. 8b
Road 4 630 5. 7) 43. 5 3. 8b
Salt Marsh 671 6.0) 38. 4 5. 8b
Road 2 160 1. 4) 22. 1 3. 2b
Deciduous Forest 492 4.4) 17. 9 7. 0b
Road 3 630 5. 7) 14.7 12.0b
Cedar Swamp 31 0. 3) 4.2 3. 1c
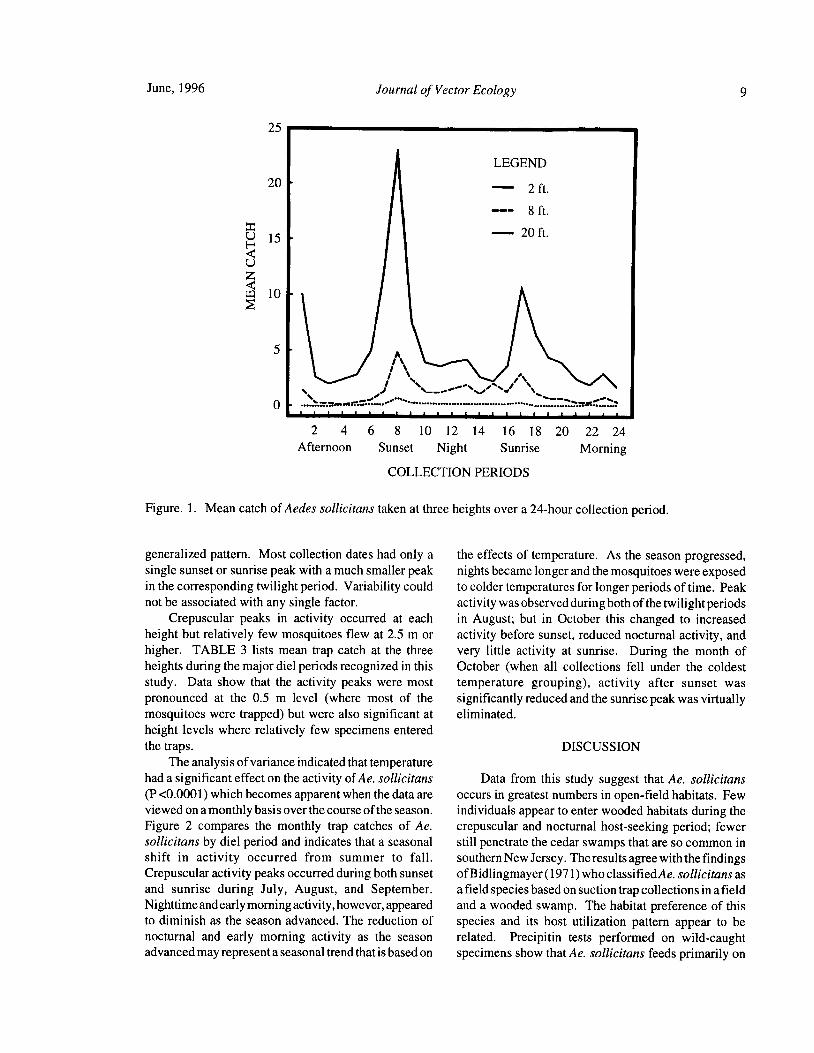
June, 1996 Journal of Vector Ecology 9
25
LEGEND
20 - 2 ft.
8 ft.
H15 - 20 ft.
W 10
5 -
A
2 4 6 8 10 12 14 16 18 20 22 24
Afternoon Sunset Night Sunrise Morning
COLLECTION PERIODS
Figure. 1. Mean catch of Aedes sollicitans taken at three heights over a 24- hour collection period.
generalized pattern. Most collection dates had only a the effects of temperature. As the season progressed,
single sunset or sunrise peak with a much smaller peak nights became longer and the mosquitoes were exposed
in the corresponding twilight period. Variability could to colder temperatures for longer periods of time. Peak
not be associated with any single factor. activity was observed during both of the twilight periodsCrepuscular peaks in activity occurred at each in August; but in October this changed to increased
height but relatively few mosquitoes flew at 2.5 m or activity before sunset, reduced nocturnal activity, andhigher. TABLE 3 lists mean trap catch at the three very little activity at sunrise. During the month ofheights during the major diel periods recognized in this October ( when all collections fell under the coldest
study. Data show that the activity peaks were most temperature grouping), activity after sunset waspronounced at the 0.5 m level ( where most of the significantly reduced and the sunrise peak was virtuallymosquitoes were trapped) but were also significant at eliminated.
height levels where relatively few specimens enteredthe traps. DISCUSSION
The analysis of variance indicated that temperature
had a significant effect on the activity ofAe. sollicitans Data from this study suggest that Ae. sollicitansP< 0.0001) which becomes apparent when the data are occurs in greatest numbers in open- field habitats. Few
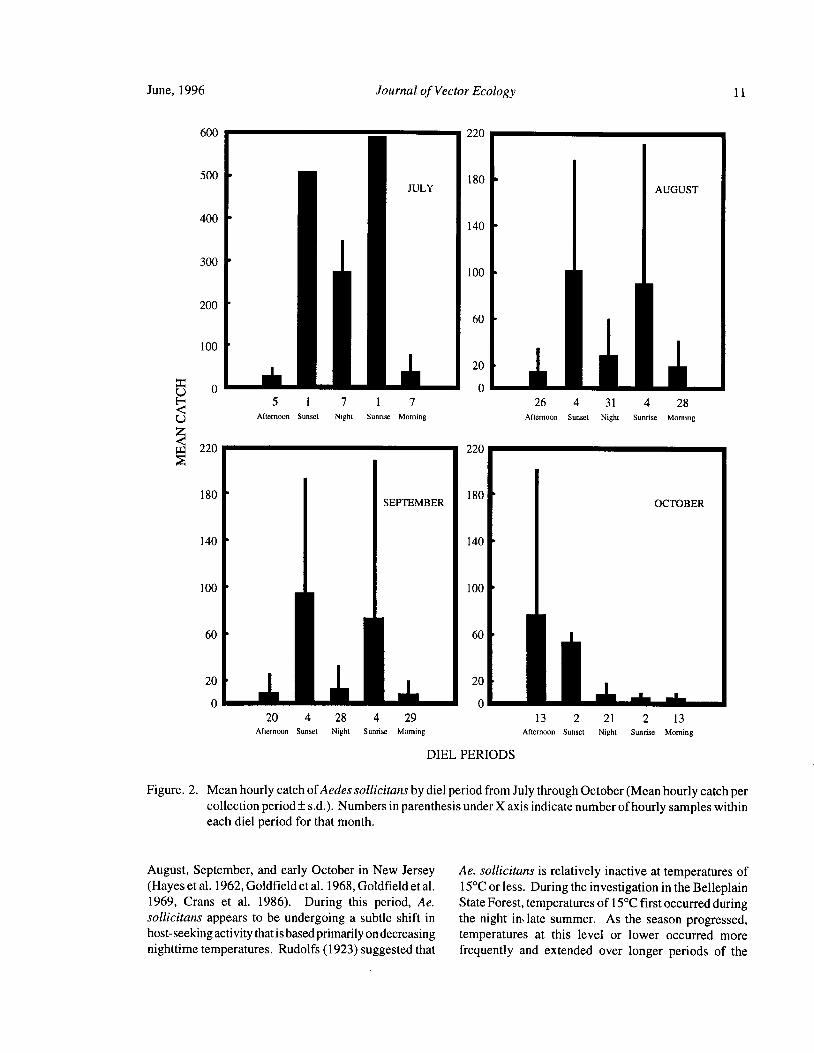
viewed on a monthly basis over the course of the season. individuals appear to enter wooded habitats during theFigure 2 compares the monthly trap catches of Ae. crepuscular and nocturnal host-seeking period; fewersollicitans by diel period and indicates that a seasonal still penetrate the cedar swamps that are so common in
shift in activity occurred from summer to fall. southern New Jersey. The results agree with the findingsCrepuscular activity peaks occurred during both sunset ofBidlingmayer( 1971) who classifiedAe. sollicitans as
and sunrise during July, August, and September. a field species based on suction trap collections in a fieldNighttime and early morning activity, however, appeared and a wooded swamp. The habitat preference of thisto diminish as the season advanced. The reduction of species and its host utilization pattern appear to be
nocturnal and early morning activity as the season related. Precipitin tests performed on wild-caught
advanced may represent a seasonal trend that is based on specimens show that Ae. sollicitans feeds primarily on
10 Journal of Vector Ecology June, 1996
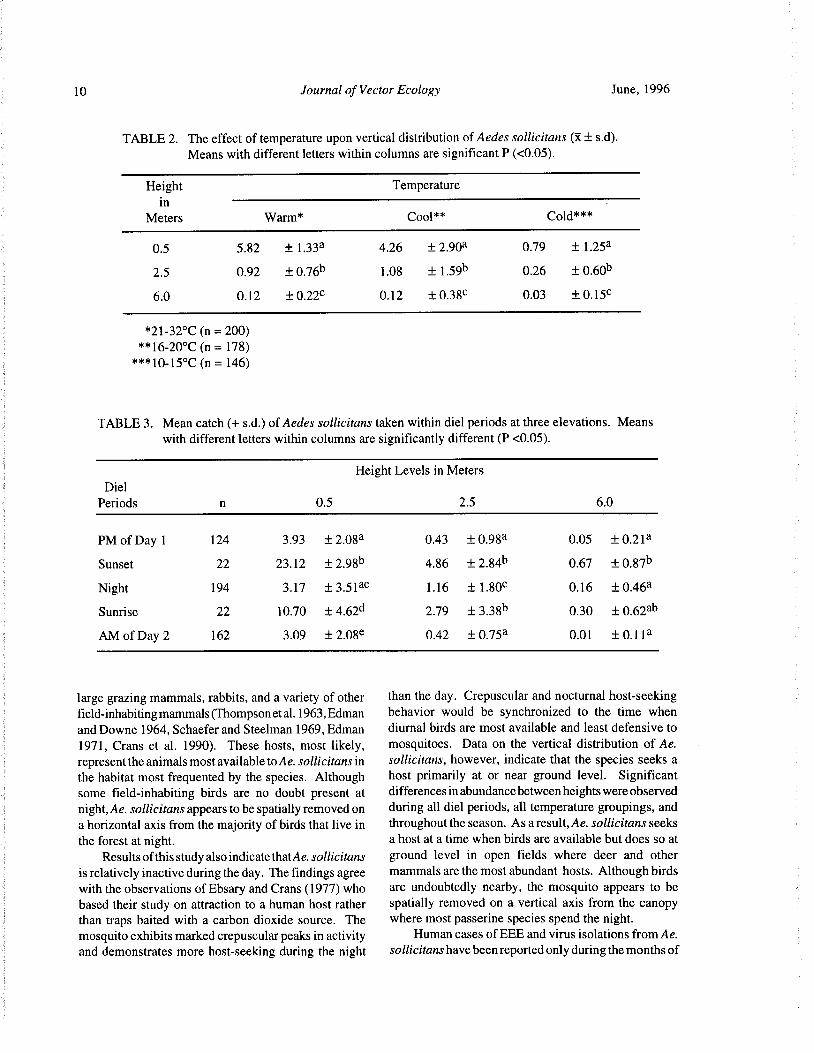
TABLE 2. The effect of temperature upon vertical distribution of Aedes sollicitans( x± s. d).
Means with different letters within columns are significant P(< 0.05).
Height Temperature
in
Meters Warm* Cool** Cold***
0. 5 5. 82 ± 1. 33a 4.26 ± 2. 903 0.79 ± 1. 25a
2. 5 0.92 ± 0.76b 1. 08 ± 1. 59b 0.26 ± 0.60b
6. 0 0. 12 ± 0. 22c 0. 12 ± 0.38c 0.03 ± 0. 15c
21- 32° C( n= 200)
16- 20°C( n= 178)
10- 15° C( n= 146)
TABLE 3. Mean catch(+ s. d.) of Aedes sollicitans taken within diel periods at three elevations. Means
with different letters within columns are significantly different( P< 0.05).
Height Levels in Meters
Diel
Periods n 0.5 2. 5 6.0
PM of Day 1 124 3. 93 ± 2. 08a 0.43 ± 0. 983 0.05 ± 0.213
Sunset 22 23. 12 ± 2.98b 4. 86 + 2. 84b 0.67 ± 0.87b
Night 194 3. 17 ± 3. 513e 1. 16 ± 1. 80e 0. 16 ± 0.46a
Sunrise 22 10.70 + 4.62d 2. 79 ± 3. 38b 0. 30 ± 0.62ab
AM of Day 2 162 3. 09 ± 2.08e 0. 42 ± 0.753 0. 01 ± 0. 113
large grazing mammals, rabbits, and a variety of other than the day. Crepuscular and nocturnal host-seekingfield-inhabiting mammals( Thompson et al. 1963, Edman behavior would be synchronized to the time when
and Downe 1964, Schaefer and Steelman 1969, Edman diurnal birds are most available and least defensive to
1971, Crans et al. 1990). These hosts, most likely, mosquitoes. Data on the vertical distribution of Ae.
represent the animals most available to Ae. sollicitans in sollicitans, however, indicate that the species seeks a
the habitat most frequented by the species. Although host primarily at or near ground level. Significant
some field- inhabiting birds are no doubt present at differences in abundance between heights were observed
night, Ae. sollicitans appears to be spatially removed on during all diel periods, all temperature groupings, anda horizontal axis from the majority of birds that live in throughout the season. As a result, Ae. sollicitans seeks
the forest at night. a host at a time when birds are available but does so at
Results of this study also indicate that Ae. sollicitans ground level in open fields where deer and other
is relatively inactive during the day. The findings agree mammals are the most abundant hosts. Although birds
with the observations of Ebsary and Crans( 1977) who are undoubtedly nearby, the mosquito appears to bebased their study on attraction to a human host rather spatially removed on a vertical axis from the canopythan traps baited with a carbon dioxide source. The where most passerine species spend the night.
mosquito exhibits marked crepuscular peaks in activity Human cases of EEE and virus isolations from Ae.
and demonstrates more host-seeking during the night sollicitans have been reported only during the months of
June, 1996 Journal of Vector Ecology 11
600 220
500 • 180 •JULY AUGUST
400 • 140 •
300100 •
200 '
60
ijkimum,•
100 '
016 20 •
Q 5 1 7 1 7 26 4 31 4 28Afternoon Sunset Night Sunrise Morning Afternoon Sunset Night Sunrise Morning
W 220 ` 220
SEPTEMBER180
OCTOBER
140 ' 140 •
100 ' 100 •
60 60 •
20 20
0 0
20 4 28 4 29 13 2 21 2 13Afternoon Sunset Night Sunrise Morning Afternoon Sunset Night Sunrise Morning
DIEL PERIODS
Figure. 2. Mean hourly catch of Aedes sollicitans by diel period from July through October( Mean hourly catch percollection period± s. d.). Numbers in parenthesis under X axis indicate number ofhourly samples withineach diel period for that month.
August, September, and early October in New Jersey Ae. sollicitans is relatively inactive at temperatures ofHayes et al. 1962, Goldfield et al. 1968, Goldfield et al. 15° C or less. During the investigation in the Belleplain
1969, Crans et al. 1986). During this period, Ae. State Forest, temperatures of 15° C first occurred duringsollicitans appears to be undergoing a subtle shift in the night in, late summer. As the season progressed,host-seeking activity that is based primarily on decreasing temperatures at this level or lower occurred morenighttime temperatures. Rudolfs( 1923) suggested that frequently and extended over longer periods of the

12 Journal of Vector Ecology June, 1996
night. Low temperatures appeared to gradually reduce The results of the studies conducted in the Belleplain
the nocturnal activity of Ae. sollicitans and also lower State Forest agree with that observation and suggest that
the size of the activity peak in early morning. The trend ground-dwelling birds rather than perching birds shouldwas obvious throughout the month of September. By be more fully investigated as a source for infection ofOctober, host-seeking at dawn was virtually eliminated Ae. sollicitans with EEE virus.
and the peak at dusk shifted from sunset to earlyafternoon. Blaustein et al. ( 1980) made a similar Acknowledgments
observation in New Jersey when flight activity wasbeing measured to determine the best timing for aerial Appreciation is expressed to the Cape May Countyapplications for the control of Ae. sollicitans. Mosquito Control Commission and their staff for the
The combination of habitat preference, vertical assistance provided in this study. Without their help,distribution ofhost-seeking, and seasonal diel periodicity interest, and support, weekly collections would haveofAe. sollicitans may be important factors limiting the been difficult; and preparing the equipment for 25- hourspecies' potential as a vector of EEE virus. Forest- sampling periods would have been a monumental task.roosting birds are believed to function as the main The authors extend their sincere thanks to each member
disease reservoir ( Stamm 1963) and two hypotheses of the Cape May County Mosquito Commission and tohave been proposed regarding the relationship ofhabitat the Superintendent, Judy A. Hansen. They are to beand prevalence of EEE virus in birds and mosquitoes. commended for supporting basic research andWilliams et al.( 1972) suggestedthatEEEviruscirculates recognizing that research, in turn, benefits mosquitoprimarily in birds deep within cedar bogs and wooded control.
swamps. Morris et al.( 1980) and Howard et al.( 1983)
felt that virus transmission to birds might be greatest on REFERENCES CITED
the edges of bogs, swamps, woodlands, or open fields.
Data from the study conducted in the Belleplain State Bidlingmayer, W. L. 1971. Mosquito flight path in
Forest of New Jersey suggest that in either case, Ae. relation to the environment. 1. Illumination levels,
sollicitans would have limited contact with the orientation, and resting areas. Ann. Entomol. Soc.amplification source ofEEE virus. If the virus circulates Amer. 64: 1121- 1131.
in birds in dense swamps and bogs, Ae. sollicitans Blaustein, L., W. J. Crans, M. Slaff, and R. Kent. 1980.
would have little opportunity to make contact with the The influence of decreasing evening temperaturesvirus since it rarely enters this type of habitat. If virus on the activity of Aedes sollicitans. Proc. N.J.
occurs primarily in birds that inhabit ecotonal areas, Ae. Mosq. Control. Assoc. 67: 99.sollicitans would be spatially removed from the reservoir Crans, W. J. 1964. Continued host preference studies
source that roosted in the canopy surrounding the field with New Jersey mosquitoes. Proc. N.J. Mosq.habitat and be dependent upon field inhabiting birds as Exterm. Assoc. 51: 50-58.
a virus source during enzootic and epizootic periods. Crans, W. J., J. McNelly, T. L. Schulze, and A. J Main.Consequently, considerable virus activity might be 1986. Isolation ofeastern equine encephalitis virus
required to result in the infection of this highly susceptible from Aedes sollicitans during an epizootic inmosquito, even though the mosquito is abundant within southern New Jersey. J. Amer. Mosq. Controlthe salt marsh/ cedar swamp focus where EEE virus is Assoc. 2: 68- 72.
most common. Crans, W. J., L. J. McCuiston, and D. A. Sprenger. 1990.
Decreasing nighttime temperatures appear to The blood- feeding habits of Aedes sollicitansincrease the spatial disassociation betweenAe. sollicitans Walker) in relation to eastern equine encephalitis
and tree- roosting birds by reducing the numbers of in New Jersey. I. Host selection in nature determinedmosquitoes that enter the canopy and shifting crepuscular by precipitin tests on wild-caught specimens. Bull.and nocturnal host- seeking behavior to midday and late Soc. Vector Ecol. 15( 2): 144- 148.
afternoon. Since low nighttime temperatures occur Crans, W. J., D. A. Sprenger, and F. Mahmood. 1996.
during late summer and fall when EEE virus is most The blood- feeding habits of Aedes sollicitansabundant in the passerine population, opportunistic Walker) in relation to eastern equine encephalitis
feeding would increase at ground level where mammals, virus in coastal areas of New Jersey. II. Results ofrather than birds, would be the most likely hosts. Crans experiments with caged mosquitoes and the effects
et al. ( 1990) suggested that nonpasserine bird species, of temperature and physiological age on host
particularly ciconiiforms, may have special epidemio- selection. J. Vector Ecol. 21( 1): 1- 5.
logical significance during periods ofvirus amplification. Ebsary, B. A. and W. J. Crans. 1977. The biting activity

June, 1996 Journal of Vector Ecology 13
of Aedes sollicitans in New Jersey. Mosq. News 1980. Epizootiology of eastern equine encepha-37: 721- 724. lomyelitis in upstate New York, U.S. A. II.
Edman, J. D. 1971. Host- feeding patterns of Florida Population dynamics and vector potential of adultmosquitoes. I. Aedes, Anopheles, Coquillettidia, Culiseta melanura in relation to distance from
Mansonia and Psorophora. J. Med. Entomol. 8: the breeding habitat. J. Med. Entomol. 17: 453- 465.687- 695. Rudolfs, W. 1923. Observations on the relations
Edman, J. D. and A. R. Downe. 1964. Host-blood between atmospheric conditions and the behavior
sources and multiple feeding habits of mosquitoes of mosquitoes. Bull. N.J. Agr. Exp. Sta. No.33, 32in Kansas. Mosq. News 24: 154- 160. pp.
Edman, J. D., L. A. Webber, and H. W. Kale. 1972. Schaefer, R. E. and C. D. Steelman. 1969. Determination
Host- feeding patterns of Florida mosquitoes. II. of mosquito hosts in salt marsh areas ofLouisiana.
Culiseta. J. Med Entomol. 9: 429- 434. J. Med. Entomol. 6: 131- 134.
Goldfield, M., O. Sussman, W. Gusciora, R. Kerlin, W. Stamm, D. D. 1963. Susceptibility of bird populationsCarter, and R. P. Kandle. 1968. Arbovirus activity to eastern, western, and St. Louis encephalitis
in New Jersey during 1967. Proc. N.J. Mosq. viruses. Proc. XIII Intern. Congr. Vol. 1: 591- 603.
Exterm. Assoc. 55: 14- 19. Stamm, D. D., D. W. Chamberlain, and W. D. Sudia.
Goldfield, M., O. Sussman, R. Altman, and R. P. Kandle. 1962. Arbovirus studies in south Alabama, 1957-
1969. Eastern encephalitis in New Jersey during 1958. Amer. J. Hyg. 76: 61- 81.1968. Proc. N.J. Mosq. Exterm. Assoc. 56: 56- 63. Sudia, W. D. and R. W. Chamberlain. 1962. Battery-
Hayes, R. O., L. D. Beadle, A. D. Hess, O. Sussman, and operated light trap, an improved model. Mosq.M. J. Bonese. 1962. Entomological aspects of the News 22: 126- 129.
1959 outbreak of Eastern Equine Encephalitis in Thompson, E. G., D. E. Hayes, and K. W. Ludham.
New Jersey. Amer. J. Trop. Med.& Hyg. 11: 115- 1963. Notes on the feeding habits of Aedes121. sollicitans in the Chincoteaque-Assateaque Island
Howard, J. J., D. E. Emord and C. D. Morris. 1983. area of Virginia. Mosq. News 23: 297- 298.Epizootiology of eastern equine encephalomyelitis Williams, W. E., D. M. Watts, O. P. Young, and T. J.virus in upstate New York, U.S. A. V. Habitat Reed. 1972. Transmission of eastern ( EEE) and
preference of host-seeking mosquitoes ( Diptera: western( WEE) encephalitis to sentinels in relation
Culicidae). J. Med. Entomol. 20: 62- 69. to density ofCulisetamelanura mosquitoes. Mosq.Morris, C. D., R. H. Zimmerman, and J. D. Edman. News 32: 188- 192.

Journal of Vector Ecology 21( 1): 14- 16
Susceptibility of DDT, Dieldrin, and the Developmentof Malathion Resistance in Anopheles culicifacies
Diptera: Culicidae) in Ambala District of Haryana, India
R. S. Sharma, S. N. Sharma, and R. S. Sharma
National Malaria Eradication Programme, 22, Sham Nath Marg, Delhi-110054, India
Received 10 September 1994; Accepted 2 February 1996
ABSTRACT: The susceptibility of the malaria vector Anopheles culicifacies to DDT, dieldrin, andmalathion was determined in Nada Sahib of Ambala District, India. The vector population was resistant
to DDT, dieldrin, and malathion. The mortality ofAnopheles culicifacies to malathion ranged between 20to 80% at different times. Malathion coverage during spray operations was between 62. 1% in human
dwellings and 91. 6% in cattle sheds.
Keyword Index: Mosquitoes, insecticide resistance, malaria.
INTRODUCTION MATERIALS AND METHODS
The large scale use of insecticides for public health The field study was carried out in the Nada Sahiband agriculture, deforestation, extension of irrigation area of Ambala District ofHaryana State, where duringfacilities to bring more areas under cultivation, and lack the last few years the incidence of malaria was
of water management have all changed the ecology of extremely high. The susceptibility of the vectormalaria vectors in India. Anopheles culicifacies is an population was determined with the WHO Kit ( WHO
important malaria vector in rural areas of India, and its 1975). Gravid mosquitoes were exposed to malathion
resistance to DDT and dieldrin has been found to be 5% impregnated papers. The exposure period was one
widespread. This resistance may have resulted from a hour, and mortality counts were made after a 24-hourchange in ecological conditions, and there is an urgent holding period. The method reported by Brown and Palneed for entomological studies in the malarious areas to ( 1971) was adopted in the study. Adult female
assess vector susceptibility to different insecticides. mosquitoes were collected with aspirator tubes from the
Anopheles culicifacies has shown varying levels of field and transferred to a plastic holding tube. Tests wereresistance to DDT, dieldrin, and malathion. The carried out in different months within the study area.resistance tomalathion in An. culicifacies was identified Three cycles( rounds) of malathion spray in the studyin Gujarat State( Rajagopal 1977). Among the malaria area were carried out since 1983. Two rounds of
vectors, Anopheles stephensi was first to have malathion spray were undertaken in 1990 and 1992 duedeveloped resistance to DDT in 1945( Rajagopal et. al to nonavailability of insecticide.1956). Subha Rao et. al ( 1984) identified the
development ofmalathion resistance in An. stephensi in RESULTS AND DISCUSSION
Haryana State. In western India, most parts of Gujarat,
Maharshtra, and bordering districts ofMadhya Pradesh, The malariometric indices of the study area areAn. culicifacies has become resistant to malathion ( Das shown in TABLE 1. The annual blood examination rate
et. al 1986, Vittal and Deshpande 1983). This study was was more than 10% in all years. The annual parasiteundertaken to determine the level of susceptibility of incidence was highest at 6. 1 in 1990. The slide
An. culicifacies populations to diagnostic concen- positivity rates were highest in 1990 and 1991.trations of DDT, dieldrin, and malathion. Susceptibility tests were carried out by exposing field
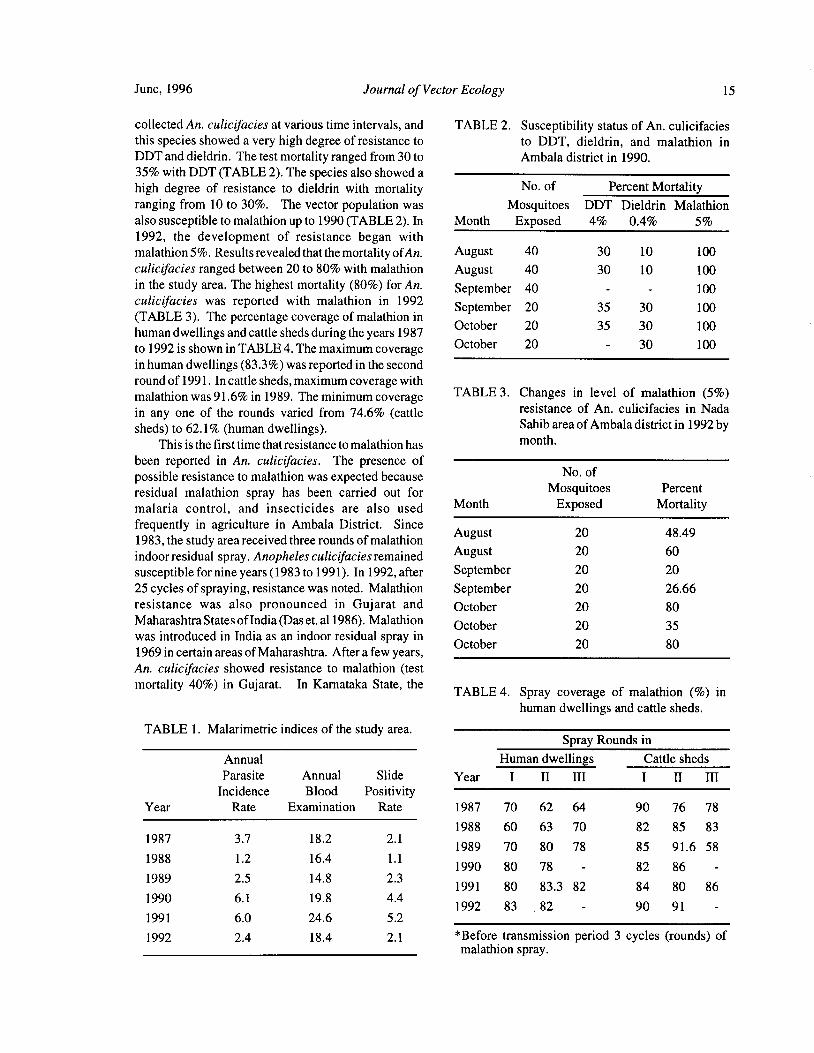
June, 1996 Journal of Vector Ecology 15
collected An. culicifacies at various time intervals, and TABLE 2. Susceptibility status of An. culicifaciesthis species showed a very high degree of resistance to to DDT, dieldrin, and malathion inDDT and dieldrin. The test mortality ranged from 30 to Ambala district in 1990.
35% with DDT( TABLE 2). The species also showed a
high degree of resistance to dieldrin with mortalityNo. of Percent Mortality
ranging from 10 to 30%. The vector population was Mosquitoes DDT Dieldrin Malathion
also susceptible to malathion up to 1990( TABLE 2). In Month Exposed 4% 0.4% 5%
1992, the development of resistance began with
malathion 5%. Results revealed that the mortality ofAn. August 40 30 10 100
culicifacies ranged between 20 to 80% with malathion August 40 30 10 100
in the study area. The highest mortality ( 80%) for An. September 40 100culicifacies was reported with malathion in 1992
September 20 35 30 100TABLE 3). The percentage coverage of malathion in
October 20 35 30 100human dwellings and cattle sheds during the years 1987to 1992 is shown in TABLE 4. The maximum coverage
October 20 30 100
in human dwellings( 83. 3%) was reported in the second
round of 1991. In cattle sheds, maximum coverage with
malathion was 91. 6% in 1989. The minimum coverageTABLE 3. Changes in level of malathion ( 5%)
in any one of the rounds varied from 74.6% ( cattleresistance of An. culicifacies in Nada
sheds) to 62. 1%( human dwellings). Sahib area of Ambala district in 1992 by
This is the first time that resistance to malathion has month.
been reported in An. culicifacies. The presence of
possible resistance to malathion was expected because No. of
residual malathion spray has been carried out for Mosquitoes Percent
malaria control, and insecticides are also usedMonth Exposed Mortality
frequently in agriculture in Ambala District. SinceAugust 20 48.49
1983, the study area received three rounds of malathionindoor residual spray. Anopheles culicifacies remained
August 20 60
susceptible for nine years( 1983 to 1991). In 1992, after September 20 20
25 cycles of spraying, resistance was noted. Malathion September 20 26.66
resistance was also pronounced in Gujarat and October 20 80Maharashtra States of India( Das et. al 1986). Malathion October 20 35was introduced in India as an indoor residual spray in
October 20 801969 in certain areas of Maharashtra. After a few years,
An. culicifacies showed resistance to malathion ( test
mortality 40%) in Gujarat. In Karnataka State, theTABLE 4. Spray coverage of malathion (%) in
human dwellings and cattle sheds.
TABLE 1. Malarimetric indices of the study area.Spray Rounds in
Annual Human dwellings Cattle sheds
Parasite Annual Slide Year I II III I II III
Incidence Blood PositivityYear Rate Examination Rate 1987 70 62 64 90 76 78
1988 60 63 70 82 85 831987 3. 7 18. 2 2. 1
1989 70 80 78 85 91. 6 581988 1. 2 16.4 1. 1
1990 80 78 - 82 86 -1989 2. 5 14. 8 2.3
1991 80 83. 3 82 84 80 861990 6. 1 19. 8 4.4
1992 83 . 82 - 90 91 -1991 6.0 24.6 5. 2
1992 2.4 18. 4 2. 1 Before transmission period 3 cycles ( rounds) ofmalathion spray.

16 Journal of Vector Ecology June, 1996
mortality to malathion ofAn. culicifacies was 74. 2%. In populations to deltamethrin. J. Am. Mosq. Contr.Thane district of Maharashtra, An. culicifacies Assoc. 2: 553- 555.
developed resistance after 13 cycles of spray( Vittal and Rajgopal, N., J. C. Vedamanikkam, and H. C. Ramoo.
Deshpande 1983). The development of resistance in An. 1956. A preliminary note on the development ofculicifacies is due to prolonged selection pressure resistance to DDT by larvae ofAnopheles stephensimaintained by the control program and its indoor resting type in Erode Urban, South India. Bull. Malar.
and biting habits. The noncooperative attitude of Soc. Mosq. Dis. 4: 126- 128.villagers to residual spray and resistance to An. Rajagopal, R. 1977. Malathion resistance in Anopheles
culicifacies to malathion necessitated the development culicifacies in Gujarat. Ind. J. Med. Res. 66: 27- 28.
of an alternative control strategy. Sharma, V. P., H. C. Uprety, Nutan Nanda, V. K. Raina,S. R. Parida, and V. K. Gupta. 1982. Impact of
Acknowledgments DDT spraying on malaria transmission in villageswith resistant Anopheles culicifacies. Ind. J.
We wish to acknowledge our gratitude to Mr. P. K. Malariol. 19: 5- 12.
Jena and Mr. Y. S. Parihar for collecting the mosquitoes Subba Rao, S. K. Sharma, V.P. Sharma, K. Vasantha,
and Mr. Jarnail Singh, Mr. S. K. Katyal, and Mr. Ashok and T. Adak. 1984. Effect of malathion sprayingKumar Sharma for secretarial assistance. in four anopheline species and the development of
resistance in Anopheles stephensi in Mandora,
REFERENCES CITED Haryana. Ind. J. Malariol. 21: 109- 114.
Vittal, M. and L. B. Deshpande. 1983. Development of
Brown, A.W. A and R. Pal. 1971. Insecticide resistance malathion resistance in a DDT, HCH resistant
in Arthropods. World Health Organization, Anopheles culicifacies population in Thane district
Geneva, 491 pp. Maharashtra). J. Com. Dis. 15: 144- 145.
Das, M. S., P. Srivastava, J. S. Khamre, and L. B. World Health Organization. 1975. Manual on practical
Deshpande. 1986. Susceptibility ofDDT, dieldrin, entomology in malaria. Part- II. Methods andand malathion resistant Anopheles culicifacies techniques. Pp. 141- 147.

Journal of Vector Ecology 21( 1): 17- 21
Patterns in Aedes albopictus ( Skuse) Population Density,Host-Seeking, and Oviposition Behavior in Penang, Malaysia.
A. Abu Hassan, C. R. Adanan, and W. A. Rahman
School ofBiological Sciences, Universiti Sains Malaysia, 11800 Penang, Malaysia
Received 30 September 1994; Accepted 13 July 1995
ABSTRACT: This study examined the ovipositional rhythm, biting cycle, density ofadult and larvae, andthe relationship between wing length and parity ofAedes albopictus adult mosquitoes attracted to humanbait. The mosquitoes showed a peak of oviposition activity at 1600 h and two peaks during their diurnalbiting cycle. The density ofadults and larvae were correlated with rainfall. The wing length of the parouspopulation was significantly longer than the wing length of the nulliparous population.
Keyword Index: Ovipositional rhythm, biting cycle, density, wing length, parity.
INTRODUCTION MATERIALS AND METHODS
Aedes albopictus has been implicated as a vector of Location
dengue fever in endemic areas in Southeast Asia( Chan The study was conducted at the campus ofUniversitiet al. 1971). In the United States, Ae. albopictus has the Sains Malaysia, Penang, Malaysia, from July 1990 topotential to become an important arbovirus vector( Willis January 1991. The study site was in a wooded areaand Nasci 1994). Rudnick( 1965) isolated the dengue surrounded by buildings.virus from field-collected Ae. albopictus mosquitoes.
In Malaysia, Ae. albopictus is usually found at forest Oviposition Rhythm
fringes, in secondary forests, and in green areas in towns Tin cans ( 105 mm high and 772 mm in diameter)
Abu Hassan 1994, Macdonald 1957). It is one of the painted black inside and out were used as ovitraps. A
most common anthrophilic mosquitoes in Malaysia, hole on the side of the can at the height 90 mm
and it breeds in man- made containers, tree holes, and controlled the water level. Each trap was filled withbamboo stumps. about 200 ml of water and a paddle made of cardboard
Age and longevity of the larvae and adult of Ae. with a rough surface on one side was placed inside as an
albopictus depend on food supply and living longer if oviposition substrate. A total of 10 ovitraps was placed
there is enough food. Aedes albopictus females survive 10 m apart at 1 m height in the woods of the selected site
over a broad range of temperatures and relative at the campus. Every hour for six 24- hour periods thehumidities. Aedes albopictus adults aggresively bite paddles were removed from the ovitraps and brought to
any exposed skin of a motionless observer( McClelland laboratory for egg counting. Fresh paddles were replacedet al. 1973), but they prefer to feed around the ankles and in the ovitraps.
knees ( Robertson and Hu 1935) and to bite outdoors
rather than indoors( Reid 1961). During the day, adults Larval Population Studyrest near the ground in undergrowth( Chan et al. 1971). A total of 20 tins was set out in five rows with a
Willis and Nasci( 1994) found that parous host- seeking distance of 30 m between rows. Four cans were placed
females had longer wings than nulliparous host-seeking in a row with a distance of 10 m apart. Each tin was filled
females. with about 200 ml of water. The number of larvae in
Because dengue is becoming important in Malaysia each container was recorded weekly for 30 weeks.and Ae. albopictus is an important vector, we examined
the oviposition, biting activities, and distribution of this Biting Adultsmosquito in an urban area in Penang, Malaysia. Mosquitoes were sampled weekly for 12 hours
18 Journal of Vector Ecology June, 1996
from sunrise to sunset by using human bait ( Service equipped with an ocularmicrometer( Harbach and Knight
1977) from July 1990 to January 1991. Human bait 1980).
catches were carried out by two collectors, one sitting The parity of each specimen was determined asunder a tree with legs outstretched and the second described by Detinova ( 1962). The ovaries of each
collecting the mosquitoes in the test tubes from the host. specimen were removed, placed in a drop of distilledTemperatures and relative humidities ( RH) were water, and transferred to a clean slide to dry. The
recorded. The biting cycle was computed as William' s condition of the ovarian tracheoles was observed with a
Mean( MW) expressed as a percentage, which gives the compound microscope. The wing length and parity databest measure of central tendency( Haddow 1964). The were analyzed by Student' s t- test.MW is a modified geometric mean which allows for
zero values in the data. RESULTS
Collected mosquitoes were identified and the left
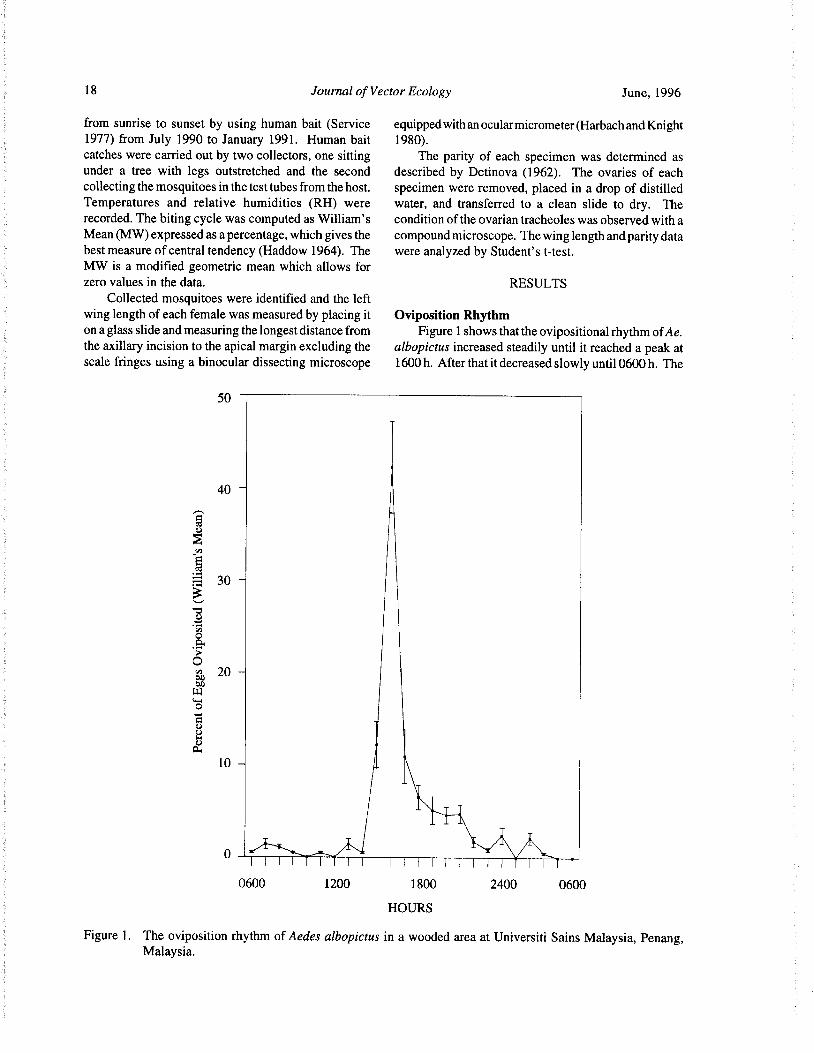
wing length of each female was measured by placing it Oviposition Rhythm
on a glass slide and measuring the longest distance from Figure 1 shows that the ovipositional rhythm ofAe.the axillary incision to the apical margin excluding the albopictus increased steadily until it reached a peak atscale fringes using a binocular dissecting microscope 1600 h. After that it decreased slowly until 0600 h. The
50
40 —
a
30 —
3
O
C
a 20 —w
0
a
U i
1-.
a
10 -
1 -
01 1 1 1 I 1 1 1 1 1 1 1 1 1 1 1 1 ' I T T
0600 1200 1800 2400 0600
HOURS
Figure 1. The oviposition rhythm of Aedes albopictus in a wooded area at Universiti Sains Malaysia, Penang,Malaysia.
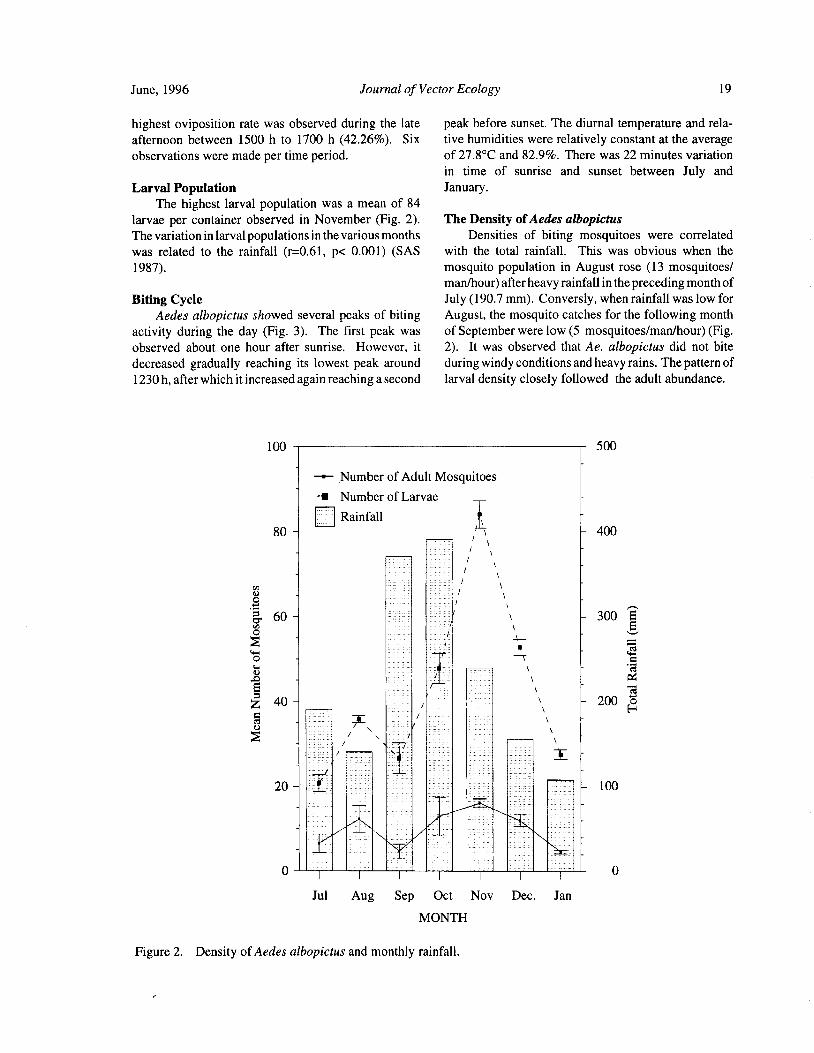
June, 1996 Journal of Vector Ecology 19
highest oviposition rate was observed during the late peak before sunset. The diurnal temperature and rela-
afternoon between 1500 h to 1700 h ( 42.26%). Six tive humidities were relatively constant at the averageobservations were made per time period. of 27. 8° C and 82.9%. There was 22 minutes variation
in time of sunrise and sunset between July andLarval Population January.
The highest larval population was a mean of 84
larvae per container observed in November ( Fig. 2). The Density ofAedes albopictusThe variation in larval populations in the various months Densities of biting mosquitoes were correlatedwas related to the rainfall ( r=0.61, p< 0.001) ( SAS with the total rainfall. This was obvious when the
1987). mosquito population in August rose ( 13 mosquitoes/
man/ hour) after heavy rainfall in the preceding month of
Biting Cycle July( 190. 7 mm). Conversly, when rainfall was low forAedes albopictus showed several peaks of biting August, the mosquito catches for the following month
activity during the day ( Fig. 3). The first peak was of September were low( 5 mosquitoes/ man/ hour)( Fig.observed about one hour after sunrise. However, it 2). It was observed that Ae. albopictus did not bite
decreased gradually reaching its lowest peak around during windy conditions and heavy rains. The pattern of1230 h, after which it increased again reaching a second larval density closely followed the adult abundance.
100 500
Number of Adult Mosquitoes
Number of Larvae
Rainfall I\
80 - r ' 400
rI
r
r
N _ r
0
c° 60 - I 300 Eo
r
5o - r
az _ 1
1
o
Z 40 200 FI -
2 i
r
d
20 - 100
0 1 I I I I I I":.0
Jul Aug Sep Oct Nov Dec. Jan
MONTH
Figure 2. Density of Aedes albopictus and monthly rainfall.
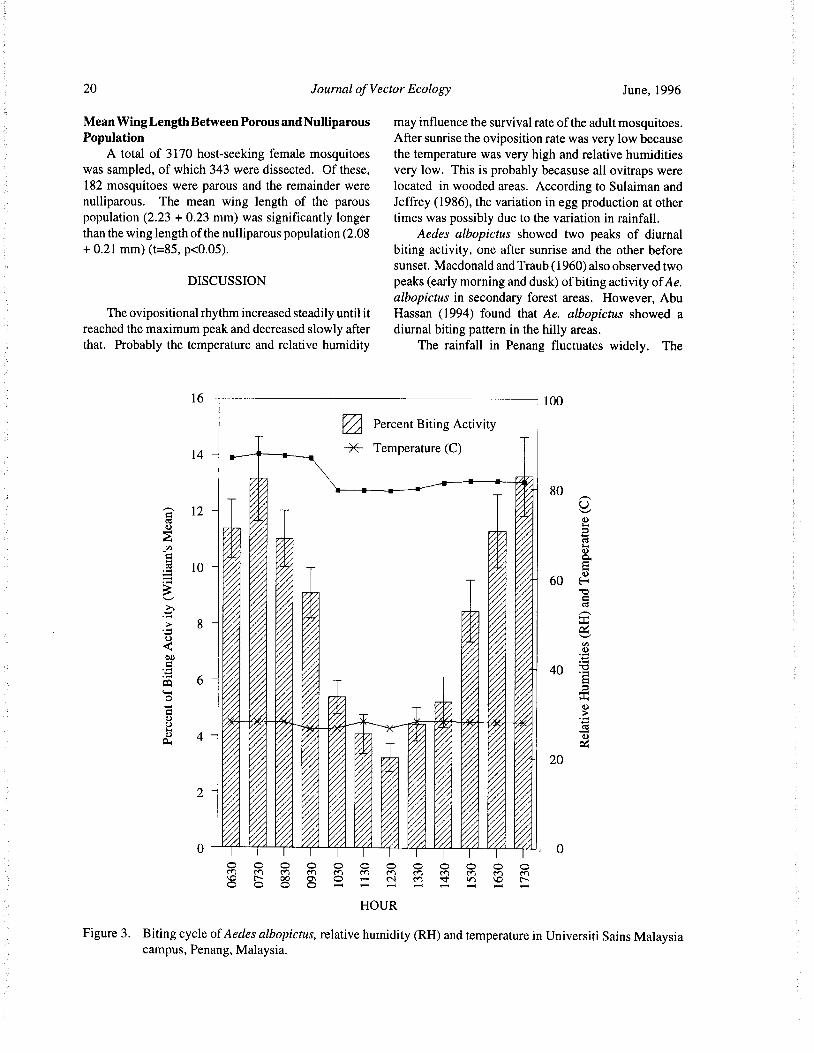
20 Journal of Vector Ecology June, 1996
Mean Wing Length Between Porous and Nulliparous may influence the survival rate of the adult mosquitoes.Population After sunrise the oviposition rate was very low because
A total of 3170 host-seeking female mosquitoes the temperature was very high and relative humiditieswas sampled, of which 343 were dissected. Of these, very low. This is probably becasuse all ovitraps were182 mosquitoes were parous and the remainder were located in wooded areas. According to Sulaiman andnulliparous. The mean wing length of the parous Jeffrey( 1986), the variation in egg production at otherpopulation ( 2.23 + 0.23 mm) was significantly longer times was possibly due to the variation in rainfall.than the wing length of the nulliparous population( 2.08 Aedes albopictus showed two peaks of diurnal
0.21 mm)( t= 85, p< 0.05). biting activity, one after sunrise and the other beforesunset. Macdonald and Traub( 1960) also observed two
DISCUSSION peaks( early morning and dusk) ofbiting activity ofAe.albopictus in secondary forest areas. However, Abu
The ovipositional rhythm increased steadily until it Hassan ( 1994) found that Ae. albopictus showed a
reached the maximum peak and decreased slowly after diurnal biting pattern in the hilly areas.that. Probably the temperature and relative humidity The rainfall in Penang fluctuates widely. The
16 100
Percent Biting Activity
14 — X Temperature( C)
Fr 80
12 — %/cl
r
FO
jr0
07: 3
0op71
40 b
0
VP0 A' 4 , . 4 ,
40.0 al, . 4 ., r -' ei
pir-, P.. P.- -
r"
0A A A A A 4 A A. A , A Acl000 ° 00000aoo
O O O OO - N M vD r
HOUR
Figure 3. Biting cycle of Aedes albopictus, relative humidity( RH) and temperature in Universiti Sains Malaysiacampus, Penang, Malaysia.

June, 1996 Journal of Vector Ecology 21
number of larvae and adults caught were correlated with Ho, B. C., K. L. Chan, and Y. C. Chan. 1971. Aedes
rainfall. The high larval population observed in aegypti ( L.) and Aedes albopictus( Skuse) in
November( Fig. 2) was possibly due to the increase in Singapore City 3. Population fluctuations. Bull.rainfall in September and October. Ho et al.( 1971) also WHO 44: 635- 641.
found that population ofAe. aegypti and Ae. albopictus Macdonald, W. W. 1957. An interim review of the non
populations fluctuated in close association with rainfall Anopheline mosquitoes of Malaya. Studies from
in Singapore. the Institute for Medical Research 50: 1- 34.
Wing length was found to be proportional to body Macdonald, W. W. and R. Traub. 1960. An introduction
weight in many mosquitoes( Shelton 1972, Parker and to the ecology of mosquitoes of the lowlandCorbet 1989). It has been used an indicator ofbody size. dipterocarp forest of Selangor, Malaya. StudiesThis implies that parous mosquitoes are bigger in size from the Institute of Medical Research 29: 79-
and have longer wing lengths. Large individuals were 109.
more successful at obtaining blood meals( Nasci 1986). McClelland, G. A. H., W. K. Hartberg, and C. M.If a population of Ae. albopictus has a high percentage Courtois. 1973. Aedes mascarensis MacGregor on
of parous mosquitoes, the transmission of disease will Mauritius. 1. Ecology in relation toAedes albopictusbe higher because parous mosquitoes are more successful Skuse) ( Diptera: Culicidae). J. Med. Ento. 10:
in getting blood meals. 570- 577.
In conclusion, the information made available will Nasci, R. S. 1986. Relationship between adultbe useful in Ae. albopictus control programs and elucidate mosquitoes ( Diptera: Culicidae) body size andits potential epidemiological importance in arbovirus parity in field population. Environ. Entomol. 15:transmission in many parts of the world. For example, 874- 876.
knowledge on the biting activity of the mosquitoes is Parker, M. J. and P. S. Corbet. 1989. Seasonal
important in determining periods of potential disease emergence, host seeking activity, age compositiontransmission. and reproductive biology of the mosquito Aedes
punctor. Ecol. Entomol. 14: 433- 442.
Acknowledgments Reid, J. A. 1961. The attraction of mosquito by humanor animal bait in relation to the transmission of
We thank the School of Biological Sciences, disease. Bull. Entomol. Res. 52: 43- 62.
Universiti Sains Malaysia, for providing the working Robertson, R. C. and S. M. K. Hu. 1935. The tiger
space. mosquito in Shanghai. The China Journal 23: 299-
306.
REFERENCES CITED Rudnick, A. 1965. Studies on ecology of dengue inMalaysia: a preliminary report. J. Med. Ento. 2:
Abu Hassan, A. 1994. Studies on the mosquito fauna 203- 208.
of Kerian district with emphasis on the mangrove SAS. 1987. SAS system for elementary statistical
swamp ecotype. PH.D thesis, University ofMalaya, analysis. Cary, NC: SAS Institute Inc., 256 pp.Kuala Lumpur, 185 pp. Service, M. W. 1977. A critical review of procedure for
Chan, Y. C., K. L. Chan, and B. C. Ho. 1971. Aedes sampling population of adult mosquitoes. Bull.aegypti ( L) and Aedes albopictus in Singapore Entomol. Res. 67: 343- 382.
City. 1. Distribution and density. Bull. WHO 44: Shelton, R. M. 1972. The effects blood source and
617- 627. quantity on production of eggs by Culex salinariusDetinova, T. S. 1962. Age grouping methods in Diptera Coquillett( Diptera: Culicidae). Mosq. News 32:
of medical importance. World Health Organisation 31- 37
Monograph 47. Geneva, 216 pp. Sulaiman, S. and J. Jeffery. 1986. The ecology ofAedesHaddow, A. J. 1964. Observations on the biting-habits albopictus( Skuse)( Diptera: Culicidae) in a rubber
ofmosquitoes in the forest canopy at Zika, Uganda, estate in Malaysia. Bull. Entomol. Res. 76: 553-
with special reference to the crepuscular periods. 557.
Bull. Entomol. Res. 55: 589- 608. Willis, F. S. and R. S. Nasci. 1994. Aedes albopictus
Harbach, R. E. and K. L. Knight. 1980. Taxonomists Diptera:Culicidae) population density and structure
glossary ofmosquito anatomy. Plexus, New Jersey, in Southwest Louisiana. J. Med. Entomol. 31: 594-
415 pp. 599.

Journal of Vector Ecology 21( 1): 22- 25
Serological Identification of the Predators of AdultAedes albopictus (Skuse) ( Diptera: Culicidae) in
Rubber Plantations and a Cemetery in Malaysia
S. Sulaiman', Z. A. Pawanchee2, M. A. Karim2
J. Jeffery', V. Busparani2, and A. Wahab2
Department ofParasitology and Medical Entomology, Faculty of Medicine,Universiti Kebangsaan Malaysia, Jalan Raja Muda Abdul Aziz,
50300 Kuala Lumpur, MALAYSIA
2Vector Control Unit, Department ofHealth, Municipality of Kuala Lumpur,Jalan Tun Razak, Kuala Lumpur, MALAYSIA
Received 13 December 1994; Accepted 26 May 1995
ABSTRACT: In Malaysia, cemeteries and rubber plantations are the preferential habitats of Aedes
albopictus( Skuse), the principal vector of dengue. Natural predators ofAe. albopictus were identified bya precipitin test. Positive reactions with anti-albopictus sera were shown in four species of spiders,
comprising Heteropoda venatoria( 76.6%), Heteropoda sp.( 42.9%), Pardosa sp.( 75.9%), and Leucauge
decorata( 60%), collected from the cemetery, and five species comprising Passiena sp.( 41. 8%), Leucauge
grata( 46.2%), Heteropoda venatoria( 90%), Cyclosa insulana( 33. 3%), and Nephila maculata( 27. 8%)
collected from rubber plantations. All of the spider species represent new host records as predators ofAe.albopictus.
Keyword Index: Aedes albopictus, predators, spiders, rubber plantations, cemetery.
INTRODUCTION MATERIALS AND METHODS
The use of the precipitin test for the detection of Localities and Collecting Methodsmosquito predators was first tested by Brooke and All samples of suspected predators were collectedProske( 1946) and then by Service( 1973a,b; 1977 a,b) from rubber plantations in Hulu Klang and Gombak,who used this technique to study the arthropod predators about 10 km and 15 km from Kuala Lumpur and from aofAedes cantans( Meigen) and the Anopheles gambiae cemetery of about 5 ha in Kuala Lumpur City. EachGiles) complex. Studies on the predators of the suspected predator was placed in a plastic container
Simulium damnosum Theobald complex ( Service and measuring 32 mm x 72 mm. Each suspected predatorLyle 1975, Service and Elouard 1980), Culex pipiens was dissected individually and the gut content wasand Culex torrentium ( Onyeka 1983), and Culex smeared on Whatman filter paper and then kept in atritaeniorhynchus ( Urabe et al. 1994) have also been desiccator with phosphorus pentoxide for rapid drying.successful. Preparations of antigen, antiserum, and the precipitin
Due to the likely importance of predators in test were conducted according to Service ( 1973a,b;regulating the populations of the dengue vector Aedes 1977a,b) with some modifications.albopictus ( Sulaiman and Jeffery 1986), we under-
took this study to identify the predators in two rubber Antigen Preparationplantations and a cemetery where Ae. albopictus Aedes albopictus larvae and adults caught from thebreeds. rubber plantations, the cemetery, and from the colony in

June, 1996 Journal of Vector Ecology 23
our insectarium were used for antigen preparation. 0. 2 ml, and large sized gut smear in 0.3 ml normal salineAfter keeping the larvae for about 24 hours in clean tap for 20 hours at room temperature. The soaked smearswater to empty their guts, the larvae were dried on filter were centrifuged for 15 min at 500 g. A small quantitypaper, weighed together with the adults, and ground in of 0.02 ml to 0.04 ml of supernatant fluid was drawn upnormal saline( 0.85%) in a tissue grinder at the rate of 2 into a small capillary tube of bore size 2.0 mm followedg Ae. albopictus per 10 ml normal saline. The extract by about an equal volume of antiserum. The capillarywas placed in 15 ml centrifuge tubes and left for about tube was then pushed into a tray of plasticine. The tube20 hours at 4°C before centrifuging at 700 G for 15 min. was left at room temperature for one to two hours andThe extract was then filtered through sterile Sartorius then examined. The presence of a distinct precipitinvacuum filtration with a pore size of 0.2 µ m and flow ring at the interface of the two fluids indicated that Ae.rate of200 ml/min. Sodium azide( 0.01%) was added as albopictus was consumed by a predator.a preservative. Quantities of 2. 5 ml of sterile antigen
mainly proteins) were placed in small sterile glass RESULTS AND DISCUSSIONbottles and kept at- 20° C.
Based on the results of this study, spiders wereProduction of Antiserum determined as predators ofAe. albopictus in the rubber
The proteins of the antigen were precipitated with plantations as well as in the cemetery( TABLES 1 and1 ml of0.4% potassium alum and the pH adjusted to 6. 8 2). Comparing the two habitats where Ae. albopictusby sterile 4% HC1 or sterile 4% NaOH. The suspension breeds in large numbers in Malaysia, there werewas then injected into the hind leg muscle of two New differences in the occurrence of species of spiders inZealand white rabbits weighing about 4 kg each. Each both habitats. From the cemetery, large numbers ofrabbit received four injections. The rabbits were test Heteropoda venatoria were collected and this speciesbled from the marginal ear vein 10 days after the second, showed the highest positive reaction with anti-third, and fourth injections. The antiserum produced in albopictus sera compared to the other species of spiders
both rabbits reacted against sterile antigen at the highest available. Although H. venatoria also showed thetiter of 1/ 1024, 10 days after the fourth injection. highest positive reaction with anti- albopictus sera in the
rubber plantations, the number collected was not highThe Precipitin Test compared to that from the cemetery. Thus, H.venatoria
Gut content smears were cut from Whatman No. 1 could play a vital role in suppressing Ae. albopictus infilter paper, placed in 2 ml centrifuge tubes. A small gut the cemetery but presumably not in the rubbersmear was soaked in 0. 1 ml, medium sized gut smear in plantations. Pardosa sp., another predator in the
TABLE 1. List of predators from a cemetery that showed positive reactions with anti-albopictus sera using the precipitin test.
Precipitin Reaction
Species No. Tested No. Positive Positive
Araneae
Sparassidae
Heteropoda venatoria 222 170 76.6
Heteropoda sp. 7 3 42.9
Lycosidae
Pardosa sp. 58 44 75.9
Tetragnathidae
Leucauge decorata 5 3 60
Total 292 220 75. 3

24 Journal of Vector Ecology June, 1996
cemetery, also showed a high positive reaction with addition to H. venatoria, Cyclosa insulana, Leucauge
anti- albopictus sera. This species could also play a grata, Nephila maculata, and Passiena sp. weremajor role in vector population suppression. However, demonstrated to be predators ( TABLE 2). Except for
Pardosa sp. thus far has not been collected in the rubber Passiena sp., the numbers of the other predator speciesplantation, indicating the habitat-specific nature of this collected in the rubber plantations were comparativelyspecies. Both H. venatoria and Pardosa sp. were low. Although Passiena sp. was the dominant predatorusually seen at ground level around the tombs and in the rubber plantations, the precipitin reaction was not
flower pots in the cemetery. The other species found in that high ( TABLE 2) compared to H. venatoria and
the cemetery included Heteropoda sp. and Leucauge Pardosa sp. in the cemetery ( TABLE 1). Thus, H.
decorata. Although both species were proven as venatoria is more cosmopolitan in its distribution,
predators serologically, their populations were too low occurring both in the rubber plantations and theto have the potential for suppressing vector populations. cemetery. Sulaiman and Jeffery( 1986) also observedAll three latter species viz., Pardosa sp., Heteropoda the dytiscid, Lacconectus punctipennis, eating Ae.
sp., and Leucauge decorata constitute new records as albopictus larvae in containers in a rubber plantation in
predators of Ae. albopictus in the field. Malaysia. However, beetle larvae were not found in the
Studies in the rubber plantations indicated that, in containers in the cemetery.
TABLE 2. List of predators from rubber estates that showed positive reactions with anti-
albopictus sera using the precipitin test.
Precipitin Reaction
Species No. Tested No. Positive % Positive
Araneae
Araneidae
Cyclosa bifida( Doleschall) 5 0 0
Cyclosa insulana( 0. Costa) 18 6 33. 3
Nephila maculata ( Fabricius) 18 5 27. 8
Lycosidae
Passiena sp. 98 41 41. 8
Sparassidae
Heteropoda venatoria( L) 10 9 90
Tetragnathidae
Leucauge grata( Guerin) 39 18 46.2
Orthoptera
Acridiidae
Atractomorpha sp. 13 0 0
Garyanda sp. 12 0 0
Oxya hyla intricata( Stal) 5 0 0
Phlaeoba antennata( B.V. Matt) 10 0 0
Gryllidae
Gryllus mitratus( Brum) 20 0 0
Total 248 79 31. 9
Data from Sulaiman et al. 1990.

June, 1996 Journal of Vector Ecology 25
According to Riechert and Lockley( 1984), spiders of Culex pipiens L. and Cx. torrentium Martini
feed almost exclusively on insects, and yet little Diptera: Culicidae) in England. Bull. Ent. Res. 73:
attention has been given to their possible role in insect 185- 194.
pest suppression. The impact of predators in reducing Riechert, S. E. and T. Lockley. 1984. Spiders as
the vector population depends on the predator numbers biological control agents. Ann. Rev. Entomol. 29:
and the number of prey they are able to find and 299- 320.
consume( Service 1973b). According to Boreham and Service, M. W. 1973a. Mortalities of the larvae of the
Ohiagu ( 1978), serological tests have limitations in Anopheles gambiae Giles complex and detection of
differentiating taxonomically closely related prey predators by the precipitin test. Bull. Ent. Res. 62:species. However, from our experience, we found that 359- 369.
more than 95% of container breeding mosquitoes in Service, M. W. 1973b. Study of the natural predatorsboth habitats were Ae. albopictus and believe that the ofAedes cantans( Meigen) using the precipitin test.use of this serological test and the interpretation of the J. Med. Entomol. 10: 503- 510.
results are valid in the present situation. Service, M. W. 1977a. Ecological and biological
Despite all the mosquitoes that the spiders appear to studies on Aedes cantans ( Meigen) ( Diptera:
be consuming, there are still an abundance of Ae. Culicidae) in Southern England. J. Appl. Ecol. 14:
albopictus in the rubber plantations and the cemetery. 159- 196.
More extensive studies are needed to determine the Service, M. W. 1977b. Mortalities of the immature
population dynamics of Ae. albopictus in its natural stages of species B of the Anopheles gambiae
habitats. complex in Kenya: Comparison between rice
fields and temporary pools, identification ofAcknowledgments predators and effects of insecticidal spraying. J.
Med. Entomol. 13: 535- 545.
We wish to thank the Ministry of Science, Service, M. W. and J. M. Elouard. 1980. Serological
Technology, and Environment, Malaysia for awarding identification of the predators of the complex of
research grant IRPA 3- 07- 03- 095 to support this study. Simulium damnosum Theobald ( Diptera:
We also wish to thank the National University of Simuliidae) in the Ivory Coast. Bull. Ent. Res. 70:Malaysia for providing research facilities. We 657- 663.
appreciate the staff of the Vector Control Unit, Service, M. W. and P. Lyle. 1975. Detection of the
Municipality of Kuala Lumpur, for their technical predators of Simulium damnosum by the precipitinsupport. We thank the CAB International Institute of test. Ann. Trop. Med. Parasitol. 69: 105- 108.Entomology, London, for identifying the spiders. Last Sulaiman, S., B. Omar, S. Omar, I. Ghauth, and J.
but not least, we appreciate Mrs. Wirda Hassan for Jeffery. 1990. Detection of the predators ofAedestyping the manuscript. albopictus ( Skuse) ( Diptera: Culicidae) by the
precipitin test. Mosq. Borne Dis. Bull. 7: 1- 4.REFERENCES CITED Sulaiman, S. and J. Jeffery. 1986. The ecology of
Aedes albopictus( Skuse)( Diptera: Culicidae) in a
Boreham, P. F. L. and C. E. Ohiagu. 1978. The use of rubber estate in Malaysia. Bull. Ent. Res. 76: 553-
serology in evaluating invertebrate prey-predator 557.
relationships: areview. Bull. Ent. Res. 68: 171- 194. Urabe, K., Y. Sekijima, and K. Nakazawa. 1994. StudyBrooke, M. M. and H. O. Proske. 1946. Precipitin test of the natural predators of a mosquito Culex
for determining natural insect predators of tritaeniorhynchus in rice field areas by using pre-immature mosquitoes. J. Natl. Malariol. Soc. 5: 45- cipitin tests. 1. Laboratory tests for the detection56. of the antigens specific to Culex tritaeniorhynchus.
Onyeka, J. O. A. 1983. Studies on the natural predators Japn. J. Sanit. Zool. 45: 43- 51.

Journal of Vector Ecology 21( 1): 26- 30
Impact of Microorganisms and Water Quality on theEfficacy of Bacillus sphaericus Neide Against
Culex pipiens Larvae in the Laboratory
M. Beck, M. Ludwig, and N. Becker'
German Mosquito Control Association( KABS),
Ludwigstrasse 99, 67165 Waldsee, Germany
Received 28 December 1994; Accepted 25 April 1995
ABSTRACT: The influence of water quality on the efficacy of Bacillus sphaericus against Culex pipienslarvae was investigated. Bioassays were conducted with different dilutions of a filtrate of hay- infusion.Impact of microorganisms in the filtrate on mosquito larval mortality was studied by comparison ofbioassays and spore counts in sterilized and nonsterilized filtrates ofhay- infusion. Increasing concentrationsof the hay- infusion filtrate( 0, 1, 10, 100%) resulted in higher LC50 and LC90 values. There were significantdifferences between larval mortalities in distilled water and hay- infusion concentrations, resulting in 4 to5- fold higher LC50 and LC90 values in 1 to 100% hay- infusion. A linear relationship was observed betweenthe organic content of the various dilutions of the filtrate and the efficacy of B. sphaericus. The presenceof microorganisms resulted in a 2. 7- fold increase in the LC90 values of a hay- infusion filtrate compared toa sterilized hay- infusion filtrate at the beginning of the test. Seven days later the increase was as much asfive times. There was a significantly higher efficacy of B. sphaericus in the sterilized hay- infusion sevendays after incubation, which was not observed with the nonsterilized hay- infusion.
Keyword Index: Bacillus sphaericus, Culex pipiens, microorganisms, water quality.
INTRODUCTION et al. 1993, Lacey 1990), and water quality( Lacey 1990,Mian and Mulla 1983, Mulla et al. 1990, Mulligan et al.
Bacillus sphaericus Neide has been shown to be 1980) that may influence the efficacy of B. sphaericus.active against a large number ofmosquito genera( Mulla In an attempt to achieve economic control of Culexet al. 1984, Singer 1985), but species of Culex have been pipiens, one of the most common stagnant waterfound to be most sensitive to this bacterium( Davidson mosquitoes in the Upper Rhine Valley, a study was1985). In field situations, efficacy of B. sphaericus is undertaken to investigate the effect of different wateraffected by numerous bioecological factors ( Lacey qualities and the influence of microorganisms on the1990). It is therefore important from both ecological efficacy of B. sphaericus, using bioassays and sporeand economic viewpoints to understand these factors count methods, respectively.and choose appropriate dosages of B. sphaericus forroutine field treatments to control mosquito larvae under MATERIAL AND METHODSvarious environmental conditions.
Apart from a difference in the susceptibility of Preparation of the Hay-infusionvarious mosquito larval instars to B. sphaericus( Wraight Five hundred grams of dried and chopped-up hayet al. 1981), there are other factors, such as larval density were mixed with 50 liters of spring water in a 70-literBecker et al. 1993, Mulla et al. 1990), temperature plastic barrel. After one week this stock suspension wasBecker et al. 1993, Mulla et al. 1990), sunlight( Becker filtered using a net with a mesh size of 0. 1 mm.
Address for correspondence

June, 1996 Journal of Vector Ecology 27
Influence of Water Quality on the Efficacy of B. The samples were diluted with sterile distilled water tosphaericus achieve appropriate spore numbers and plated on BATS
Four test series were conducted in 5- liter glass medium( Yousten et al. 1985). To assess the total sporevessels, which contained either a 100% filtrate of hay- numbers of SPHERIMOS FC and B. sphaericus powderinfusion, 10% filtrate of hay- infusion, 1% filtrate of ( BSB 0003), asample ofeach formulation was suspendedhay- infusion, or distilled water. Three vessels of each in sterile distilled water and homogenized with 15 glassdilution were treated with B. sphaericus strain 2362 beads( 6 mm) on a Vortex shaker for 10 min at maximumSpherimos FC, Novo Nordisk, Denmark) at a rate of 10 speed. Dilutions of this stock solution were also heated
mg/ l. The same number of vessels were kept untreated for 12 min at 80°C and then plated on a nonselectiveas controls. complex medium( CASO-Agar, MERCK, Darmstadt)
and on BATS medium to determine the percentage ofImpact of Microorganisms on the Efficacy of B. retrieval.
sphaericus
Two test series containing either a filtrate of hay- Abiotic Parametersinfusion or a sterilized filtrate of hay- infusion were In order to define the water quality, the followingused. Three 5- liter vessels filled with filtrate of hay- chemical parameters were measured: pH, conduc-infusion and three vessels filled with sterilized filtrate of tivity, dissolved oxygen, total hardness, carbonatehay- infusion were treated with B. sphaericus strain hardness, chloride, phosphate, ammonium, nitrite, and2362 at a rate of 10 mg/ 1 ( BSB 0003 primary powder, nitrate.
Novo Nordisk, Denmark). These vessels and the samenumber of untreated controls were kept at 20± 1° C. RESULTS
Both series of the bioassay tests were conductedimmediately after B. sphaericus treatment as well as Influence of Water Quality on the Efficacy of B.seven days posttreatment. sphaericus
TABLE 1 displays the abiotic parameters of theBioassays hay- infusion filtrates. There were significant( p<_0.05)
The fate of the active ingredient was evaluated by differences between the LC50 and LC90 values of B.modified bioassays, using WHO guidelines ( World sphaericus against mosquito larvae tested at differentHealth Organization 1985). Depending on the dilutions of the filtrate of hay- infusion ( Fig. 1). Thereconcentration required, 0.5 to 150 ml of the treated was a direct correlation between the hay- infusion and B.dilutions of the filtrate of hay- infusion were added to sphaericus activity resulting in higher LC50 and LC90200 ml plastic cups. The cups were then filled to 150 ml values ( lower toxicity) at increasingly higherwith the appropriate untreated dilution. The dilutions concentrations of the filtrate of hay- infusion. Inwere thoroughly stirred before being added to the cups comparison with distilled water( LC50= 0.0513 mg/ 1)containing the samples. Twenty- five third-instar larvae the LC50 value of the 1% filtrate ofhay- infusion( 0.0865of Culex pipiens from a colony maintained in the mg/ I) was 1. 7- fold higher. Similarly the LCS0 values oflaboratory were added to each cup. Tests were run in the the 10% filtrate ofhay- infusion( 0. 1397 mg/ 1) and 100%laboratory at 20 ± 1° C, maintaining eleven different filtrate of hay- infusion( 0.2808 mg/ 1) showed 2. 7- andconcentrations and controls, three replicates each. 5. 5- fold increase over that ofdistilled water, respectively.
Data on the 48- hour larval mortality were corrected Comparison of the LC90 values showed similar trends.according to Abbott' s formula( Abbott 1925) prior to The LC90 value in distilled water( 0.2025 mg/ 1) was 1. 6,being subjected to log-probit analysis( Raymond 1985). 2. 3, and 4. 1 times lower than that of 1( 0.3280 mg/ 1), 10Comparison of LC means and correlation between ( 0.4733 mg/ 1), and 100% filtrate ofhay- infusion( 0.8835parameters used Duncan' s multiple range test and mg/ 1), respectively. There were significant differencesregression analysis( Kohler et al. 1984) p<_0.05, Duncan' s multiple range test) between all the
different dilutions, with the exception between distilledSpore Counts water and the 1% hay- infusion.
In addition to the bioassays, in order to investigatethe tendency for B. sphaericus to recycle, spore counts Impact of Microorganisms on the Efficacy of B.were carried out according to Yousten et al.( 1985) at the sphaericus
same time intervals as the bioassays. To eliminate the The presence of microorganisms in fermentingvegetative cells ofB. sphaericus, a sample of 10 ml was water had a significant impact on the efficacy of B.taken from each vessel and heated for 12 min at 80°C. sphaericus against third- instar larvae of Culexpipiens.

28 Journal of Vector Ecology June, 1996
TABLE 1. Abiotic parameters in different dilutions of the filtrate of hay- infusion.
Dilution in percent
Parameter 100 10 1 0
pH 7 6.5 6. 5 6.5
Conductivity inµ S/ cm 1043 198 28 0
Oxygen in mg/ 1 0 4.5 6. 5 8
Total hardness in° dH 118 8 4.5 0
Carbonic hardness in° dH 26 3 0.5 0
Ammonium in mg/ 1 10 3 1 0
Nitrite in mg/ 1 0 0 0 0
Nitrate in mg/ 1 60 10 2 0
Chloride in mg/ 1 140 20 20 0
Phosphate in mg/ 1 10 5 1 0
As shown in Figure 2, the LC90 values in nonsterilized larvae in polluted waters. In our standardized experiments
filtrate ofhay- infusion were 2.7 times higher than those we were able to show that there is a linear relationshipwith sterilized filtrate of hay- infusion. At seven days between the level of organic content of the suspension
posttreatment both LC50 and LC90 values in nonsterilized and the efficacy of B. sphaericus.filtrate of hay- infusion showed a 5- fold increase as The lowerefficacy ofB. sphaericus against mosquitocompared with those in sterilized filtrate ofhay- infusion. larvae in hay- infusion in contrast to distilled water wasAlso there were significant differences ( p <_ 0.001) due to the presence of suspended solid components of
between the LC50 and LC90 values of the two test series. the hay- infusion. This effect is due to the ingestion ofThe increase in activity of B. sphaericus( 3. 1 times microparticles instead of the B. sphaericus-spore-crystal-
the LC50 and 2.7 times the LC90 values) was significantly complex by mosquito larvae, which leads to a reducedconspicuous in sterilized hay- infusion at 0 and 7- days relative uptake of the toxin in hay- infusion than inposttreatment. No such significant increase was observed distilled water. In accordance with these findings, B.
with the nonsterilized hay- infusion. During the test sphaericus is more effective in sterilized hay- infusionperiod, the water quality did not change significantly in than in nonsterilized infusion. This effect may be due toeither test series. These results were also supported by the reduced number of microorganisms available as
the spore counts of B. sphaericus, indicating 63 times food particles following sterilization. It seems unlikelymore spores in the sterilized filtrate of hay- infusion than that the differences in effect are caused by the degradationin the nonsterilized filtrate of hay- infusion at 7- day of the toxins by microorganisms. Another factor whichposttreatment( Fig. 3). A significant increase( 45 times) needs to be considered is that in the unsterilized hay-was also observed when the numbers of spores in the infusion there is a rich protozoan fauna( for example,
sterilized hay- infusion were compared at 0 and 7 days ciliates) which can compete with mosquito larvae for
after treatment. feeding on B. sphaericus.The lower LC values in the sterilized hay- infusion
DISCUSSION after seven days, together with the increase in the spore
numbers, show that recycling of the bacterium may takeOur experiments demonstrate that the activity ofB. place under certain conditions. The absence of
sphaericus is significantly influenced by water quality. microorganisms following sterilization allows the B.This is in accordance with earlier studies in which the sphaericus spores to germinate and reproduce themselves
efficacy of B. sphaericus 1593 against floodwater without any competition for nutrients with other bacteriamosquito larvae was found to be reduced in raw sewage and fungi and the predation by ciliates. Furthermore, theeffluent( Mulligan et al. 1980) and in experimental pond decomposition processes are reduced. It was demon-
water enriched with chicken manure( Mian and Mulla strated during this series ofexperiments thatB. sphaericus1983). Mulla et al.( 1984, 1990) found that higher rates is able to recycle under certain environmental conditions.
of this biolarvicide were needed to control mosquito The composition of microorganisms in mosquito
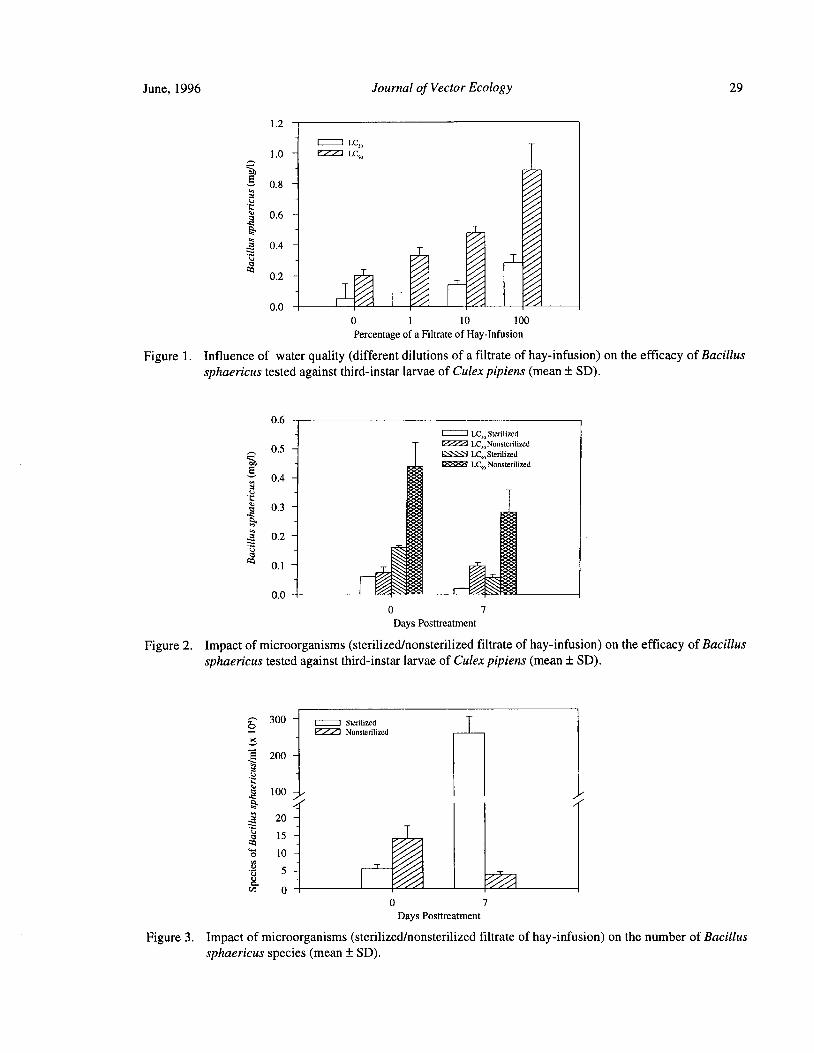
June, 1996 Journal of Vector Ecology 29
1. 2
I I LC,o1. 0 — ® LC,
ti0. 8 —
U
0.6 —
0.
0. 4
T
0. 2 =
0.0
0 1 10 100
Percentage of a Filtrate of Hay- Infusion
Figure 1. Influence of water quality( different dilutions of a filtrate of hay- infusion) on the efficacy of Bacillussphaericus tested against third- instar larvae of Culex pipiens( mean± SD).
0.6
I I LC,o SterilizedLC,0Nonsterilized
0' 5LC. Sterilized
LC„ Nonsterilized
h 0.4
1 0.3
0.2
co0. 1
ice= t0.0
0 7
Days Posttreatment
Figure 2. Impact of microorganisms( sterilized/nonsterilized filtrate of hay- infusion) on the efficacy of Bacillussphaericus tested against third- instar larvae of Culex pipiens( mean± SD).
s- 300 — I I Sterilized
Nonsterilized
5C
200 —h
U
100
ti 20 -
0 15 —
O 10
T5 —
vl o --
0 7
Days Posttreatment
Figure 3. Impact of microorganisms( sterilized/ nonsterilized filtrate of hay- infusion) on the number of Bacillussphaericus species( mean± SD).

30 Journal of Vector Ecology June, 1996
breeding sites will clearly influence the recycling of B. 128- 134.
sphaericus. Mulla, M. S., H. A. Darwazeh, E. W. Davidson, and H.
T. Dulmage. 1984. Efficacy and persistence of theAcknowledgments microbial agent Bacillus sphaericus against
mosquito larvae in organically enriched habitats.This investigation received financial support from Mosq. News 44: 166- 173.
the UNDP/World Bank/ WHO Special Programme for Mulla, M., H. A. Darwazeh, and M. Zgomba. 1990.
Research and Training in Tropical diseases. The study Effect of some environmental factors on the efficacywas also financially supported by the" Gesellschaft zur of Bacillus sphaericus 2362 and Bacillus thurin-
Forderung der Stechmuckenbekampfung e. V., GFS," giensis( H- 14) against mosquitoes. Bull. Soc. Vect.
Waldsee, Germany. Ecol. 15: 166- 176.
Mulligan, F. S., C. H. Schaefer, and W. H. Wilder.REFERENCES CITED 1980. Efficacy and persistence of Bacillus
sphaericus and Bacillus thuringiensis( H- 14) againstAbbott, W. S. 1925. A method of computing the mosquitoes under laboratory and field conditions.
effectiveness of an insecticide. J. Econ. Entomol. J. Econ. Entomol. 73: 684- 688.18: 265- 267. Raymond, M. 1985. Presentation d' un programme
Becker, N., M. Ludwig, M. Beck, and M. Zgomba. d' analyse log-probit pour micro-ordinateur. Cah.1993. The impact of environmental factors on the ORSTOM Ser. Entomol. Med. Parasitol. 22: 117-
efficacy of Bacillus sphaericus against Culex 121.
pipiens. Bull. Soc. Vector Ecol. 18: 61- 66. Singer, S. 1985. Bacillus sphaericus ( Bacteria). Pp.Davidson, E. W. 1985. Bacillus sphaericus as a 123- 131 in Biological control ofmosquitoes( H. C.
microbial control agent for mosquito larvae. Pp. Chapman, ed.). Am. Mosq. Cont. Assoc. Bull. 6,213- 226 in Integrated mosquito control 243 pp.methodologies Vol. 2 ( M. Laird and J. M. Miles, Wraight, S. P., D. Molloy, H. Jamnback, and P. McCoy.eds.). Academic Press, New York, 477 pp. 1981. Effects of temperature and instar on the
Kohler, W. G., G. Schachtel, and G. Voleske. 1984. efficacy of Bacillus thuringiensis var. israelensisBiometrie. Springerverlag, Berlin,Heidelberg, New and Bacillus sphaericus strain 1593 against AedesYork, 230 pp. simulans larvae. J. Invert. Pathol. 38: 78- 87.
Lacey, L. A. 1990. Persistence and formulation of World Health Organization. 1985. Informal consul-Bacillus sphaericus. Pp 284- 294 in Bacterial con- tation on the development of Bacillus sphaericustrol of mosquitoes and black flies( H. de Barjac and as a microbial larvicide. W.H.O./ TDR/ BCV/D. J. Sutherland, eds.). Rutgers Univ. Press, New SPHAERICUS/ 85: 3. 24 pp. ( mimeogr.). Geneva.
Brunswick, N. J., 349 pp. Yousten, A. A., S. B. Fretz, and S. A. Jelley. 1985.
Mian, L. S. and M. S. Mulla. 1983. Factors influencing Selective medium for mosquito-pathogenic strains
activity of the microbial agent Bacillus sphaericus of Bacillus sphaericus. Appl. Environ. Microbiol.against mosquito larvae. Bull. Soc. Vect. Ecol. 8: 49: 1532- 1533.

Journal of Vector Ecology 21( 1): 31- 36
Seasonal Occurrence of Fleas (Siphonaptera) on Deer Mice
Peromyscus maniculatus) in Northern California
0. R. Larson', R. G. Schwabe, and A. Fairbrother3
Department ofBiology, University ofNorth Dakota, Grand Forks, ND 58202, USA
2Division of Wildlife and Fisheries Biology, University of California, Davis, CA 95616, USA3Ecological Planning and Toxicology, Inc., 5010 S. W. Hout Street, Corvallis, OR 97333, USA
Received 27 February 1995; Accepted 17 July 1995
ABSTRACT: Fleas were collected from 657 of 1, 277 livetrapped Peromyscus maniculatus at Tule Lake
National Wildlife Refuge, Siskiyou County, California, from March, 1978, through February, 1979. Sevenof eight flea species( 1, 820 specimens) collected are characteristic ectoparasites of deer mice in western
North America. Of these, Aetheca wagneri, Malaraeus telchinus, and Opisodasys keeni were abundant and
comprised 94. 2% of all specimens. Although present throughout the entire year, their populations peaked
in May and June. Aetheca wagneri also exhibited a well-defined secondary peak in early October.Atyphloceras m. multidentatus, Catallagia mathesoni, Peromyscopsylla selenis, and Rhadinopsylla s.
sectilis were collected only during autumn and winter months, and then in low numbers.
Keyword Index: Siphonaptera, fleas, seasonality, Peromyscus maniculatus, California.
INTRODUCTION positive for the virus( Jay 1994), and with six fatalitiesamong 10 cases of hantavirus pulmonary syndrome in
Abundance, distribution, and seasonal aspects of the state( Jay and Madon 1995), caution is prudent. Theectoparasite populations on small mammals have been public health awareness of hantavirus may causestudied in many regions of the United States. Among nonsurveillance trapping of small rodents to becomesuch mammals, Peromyscus maniculatus, the deer less frequent. The present study conducted in northernmouse, is widely distributed throughout much of North California was a continuation of an earlier one, which
America. Three previous flea surveys of wild mammals examined ectoparasite reinfestation ofectoparasite- free
have been made in extreme northern California, all deer mice( Glicken and Schwab 1980).
motivated by the presence of epizootic plague ( Starkand Kinney 1969, Nelson and Smith 1976, Clover et al. MATERIALS AND METHODS
1989). Earlier collections and records of fleas from this
area and from the entire state, were compiled by Hubbard Deer mice examined in this study were collected1943). However, data concerning seasonal abundance from 9 March 1978 to 26 February 1979 in the Tule Lake
of fleas are limited or lacking in these studies. In National Wildlife Refuge, Siskiyou County, California,addition to being a reservoir host for plague, the deer at an elevation of 1, 220 m( Fig. 1). The habitat, a grassy,mouse has received recent attention because of its role weed- infested dike road, was bounded by Tule Lake toas a carrier of a hantavirus in the western half of the the north and by an irrigation ditch to the south. A
United States. Many mammalogists and parasitologists detailed description of the area and its vegetation was
who have trapped and studied deer mice with little provided by Glicken and Schwab( 1980).concern about infectious diseases, now regard P. Two hundred Sherman live traps( 7. 5 x 7. 5 x 25 cm)
maniculatus as a mammal to be avoided, or handled baited with rolled oats were placed at 10 m intervals.
with much caution ( CDC 1993, Fehrenbach 1994). Trapping commenced with the new moon phase of 13With 11% of deer mice surveyed in California sero- consecutive lunar cycles, and continued for one to three

32 Journal of Vector Ecology June, 1996
Tule Lake National
Wildlife Refuge
Lava Beds Nat'l Mon.
Stark and Kenney 1969)Nelson and Smith 1976)
Redwood Nat' l Park
Clover et al. 1989)
NEVADA
CALIFORNIA
150 KM
Figure 1. Current( Tule Lake) and previous collection sites for fleas in northern California.
nights until approximately 100 deer mice were captured. obtusata, a species typical of shrews, is an accidental
Traps were set one hour before dusk and checked two ectoparasite on other small mammals. If one excludes
hours after dark, and every two hours thereafter it because of its anomalous occurrence on deer mice,
throughout the night. The mice were lightly anesthetized then the remaining species fall into two seasonalwith chloroform and exsanguinated by cardiac puncture categories. Atyphloceras m. multidentatus, Catallagia
for a concurrent serology study. Mice were placed in mathesoni, Peromyscopsylla selenis, andRhadinopsylla
individual plastic bags and held in a freezer at- 20°C s. sectilis, although never abundant, are clearly cool
until pelts could be digested in a 0. 3% trypsin solution. weather, autumn or winter fleas. The other three species
Ectoparasites recovered from the digest were preserved ( Aetheca wagneri, Malaraeus telchinus, Opisodasys
in alcohol and later processed for mounting onto slides. keeni) comprised 94.2% of all specimens collected
Only the flea component of that parasite assemblage is ( TABLE 2). Although these were collected year-round,
reported in this paper. they showed marked population peaks in May and JuneFig. 2). In addition, A. wagneri exhibited a second peak
RESULTS AND DISCUSSION in the early autumn. The presence or absence of a
second seasonal peak requires verification since it mayThe 13 trapping efforts yielded 1, 277 deer mice, of be an artifact of greater or lesser collecting efforts
which 657( 51. 4%) were infested by fleas with intensities ( Benton and Kelly 1969, Durden 1980), or ecologicalranging from 1 to 12 per mouse ( TABLE 1). Of the variation from year-to-year. In our study, the latter is1, 888 flea specimens recovered, 68 were too damaged possible. The time needed to trap approximately 100by the digest technique for positive identification. The deer mice varied from three hours to three nights, with
remaining fleas ( 716 males, 1, 104 females) included winter collections requiring the longer time. Unfortu-eight species, seven of which are characteristic nately, most flea population estimates are based onectoparasites of deer mice in the western portion of adults removed from hosts, and little is known about the
North America, north of Mexico. Only Corrodopsylla abundance of the other stages. In a broad ecological
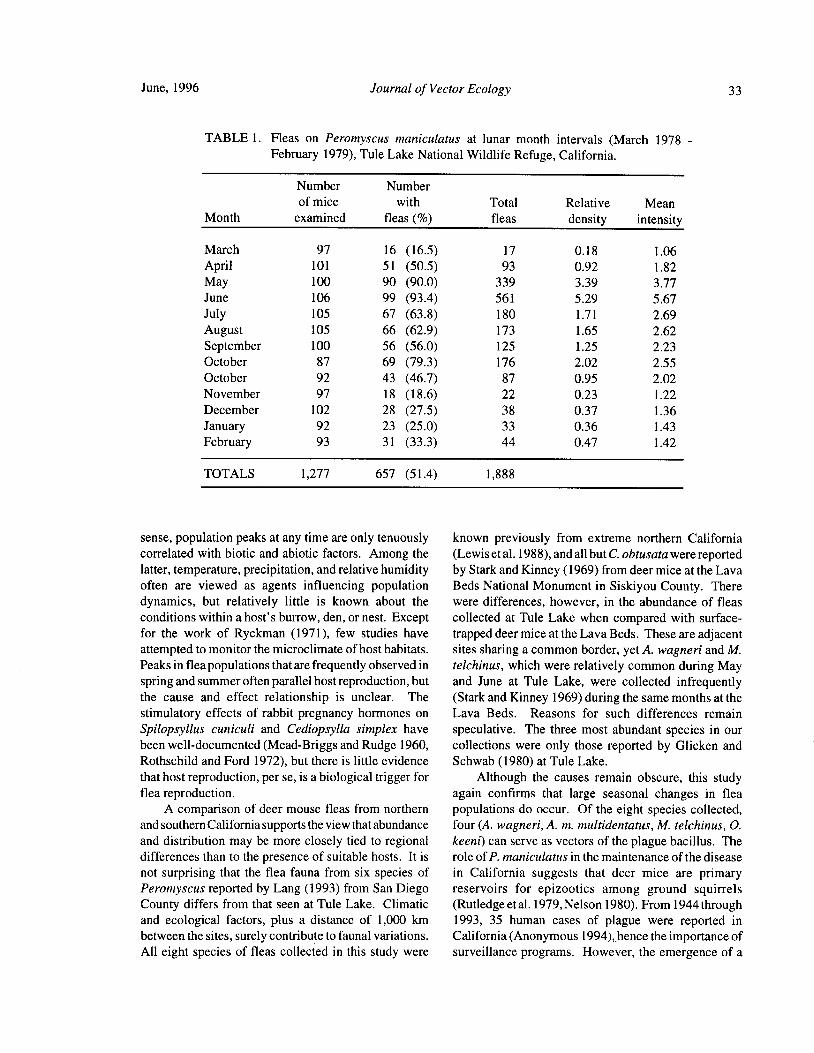
June, 1996 Journal of Vector Ecology 33
TABLE 1. Fleas on Peromyscus maniculatus at lunar month intervals ( March 1978 -
February 1979), Tule Lake National Wildlife Refuge, California.
Number Number
of mice with Total Relative Mean
Month examined fleas(%) fleas density intensity
March 97 16 ( 16.5) 17 0. 18 1. 06
April 101 51 ( 50.5) 93 0.92 1. 82
May 100 90 ( 90.0) 339 3. 39 3. 77
June 106 99 ( 93. 4) 561 5. 29 5. 67
July 105 67 ( 63. 8) 180 1. 71 2. 69
August 105 66 ( 62.9) 173 1. 65 2. 62
September 100 56 ( 56.0) 125 1. 25 2. 23
October 87 69 ( 79. 3) 176 2. 02 2. 55
October 92 43 ( 46. 7) 87 0.95 2. 02
November 97 18 ( 18. 6) 22 0.23 1. 22
December 102 28 ( 27. 5) 38 0.37 1. 36
January 92 23 ( 25. 0) 33 0.36 1. 43
February 93 31 ( 33. 3) 44 0.47 1. 42
TOTALS 1, 277 657 ( 51. 4) 1, 888
sense, population peaks at any time are only tenuously known previously from extreme northern Californiacorrelated with biotic and abiotic factors. Among the ( Lewis et al. 1988), and all but C.obtusata were reportedlatter, temperature, precipitation, and relative humidity by Stark and Kinney( 1969) from deer mice at the Lavaoften are viewed as agents influencing population Beds National Monument in Siskiyou County. Theredynamics, but relatively little is known about the were differences, however, in the abundance of fleas
conditions within a host' s burrow, den, or nest. Except collected at Tule Lake when compared with surface-
for the work of Ryckman ( 1971), few studies have trapped deer mice at the Lava Beds. These are adjacentattempted to monitor the microclimate of host habitats. sites sharing a common border, yet A. wagneri and M.Peaks in flea populations that are frequently observed in telchinus, which were relatively common during Mayspring and summer often parallel host reproduction, but and June at Tule Lake, were collected infrequentlythe cause and effect relationship is unclear. The ( Stark and Kinney 1969) during the same months at thestimulatory effects of rabbit pregnancy hormones on Lava Beds. Reasons for such differences remain
Spilopsyllus cuniculi and Cediopsylla simplex have speculative. The three most abundant species in our
been well-documented( Mead-Briggs and Rudge 1960, collections were only those reported by Glicken andRothschild and Ford 1972), but there is little evidence Schwab( 1980) at Tule Lake.
that host reproduction, per se, is a biological trigger for Although the causes remain obscure, this studyflea reproduction. again confirms that large seasonal changes in flea
A comparison of deer mouse fleas from northern populations do occur. Of the eight species collected,
and southern California supports the view that abundance four( A. wagneri, A. m. multidentatus, M. telchinus, O.
and distribution may be more closely tied to regional keeni) can serve as vectors of the plague bacillus. The
differences than to the presence of suitable hosts. It is role of P. maniculatus in the maintenance of the disease
not surprising that the flea fauna from six species of in California suggests that deer mice are primaryPeromyscus reported by Lang( 1993) from San Diego reservoirs for epizootics among ground squirrelsCounty differs from that seen at Tule Lake. Climatic ( Rutledge et al. 1979, Nelson 1980). From 1944 through
and ecological factors, plus a distance of 1, 000 km 1993, 35 human cases of plague were reported in
between the sites, surely contribute to faunal variations. California( Anonymous 1994), hence the importance of
All eight species of fleas collected in this study were surveillance programs. However, the emergence of a
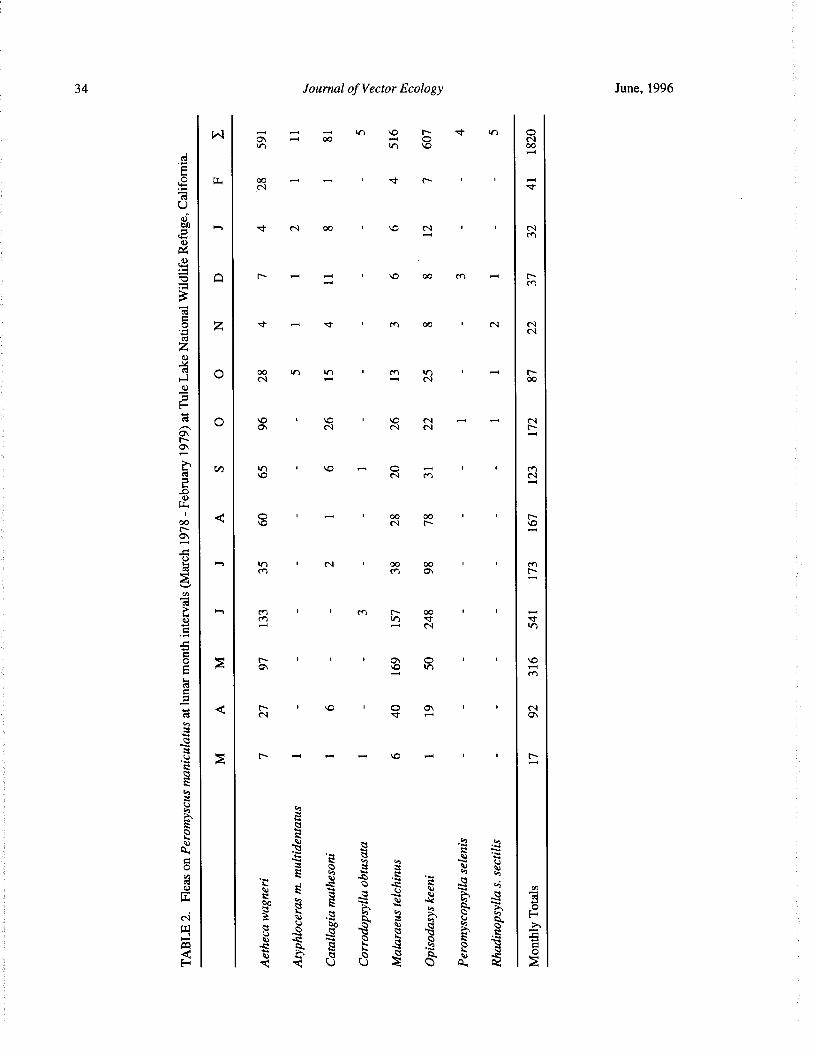
34 Journal of Vector Ecology June, 1996
W -, oo o No lin oo NIn In o0
a
EN 1-
U
b0 I N 00 ' VD N ' ' Nwa)
f1;
wbQ i .-+ 0 00 m •--' NM
a
0 Z M 00 ' N N7cd
z
00 In In en v NN 1 N 00
0
oF'
Vs 0-. O N N N ~ ~ NNON
0 N M N
Nw00
QN
00 . 8
hNCA
In ' N ' 00 00 ' ' MM M O N
ti
a>4. i
r-. M ' ' M N 00 ' . r-1
M to
A N In
r7
0 N . ' . ON 0 , .O\ O In M
a0N , ' i 0 CA I I N
aQ
N I C
h
w
tlU N .-- 8 ,, r, ' C .--. . . N
I of t
o h w h
00
y „N l
0' ooq,
oi o
Pitt a a0Q Q U U O) 0. c
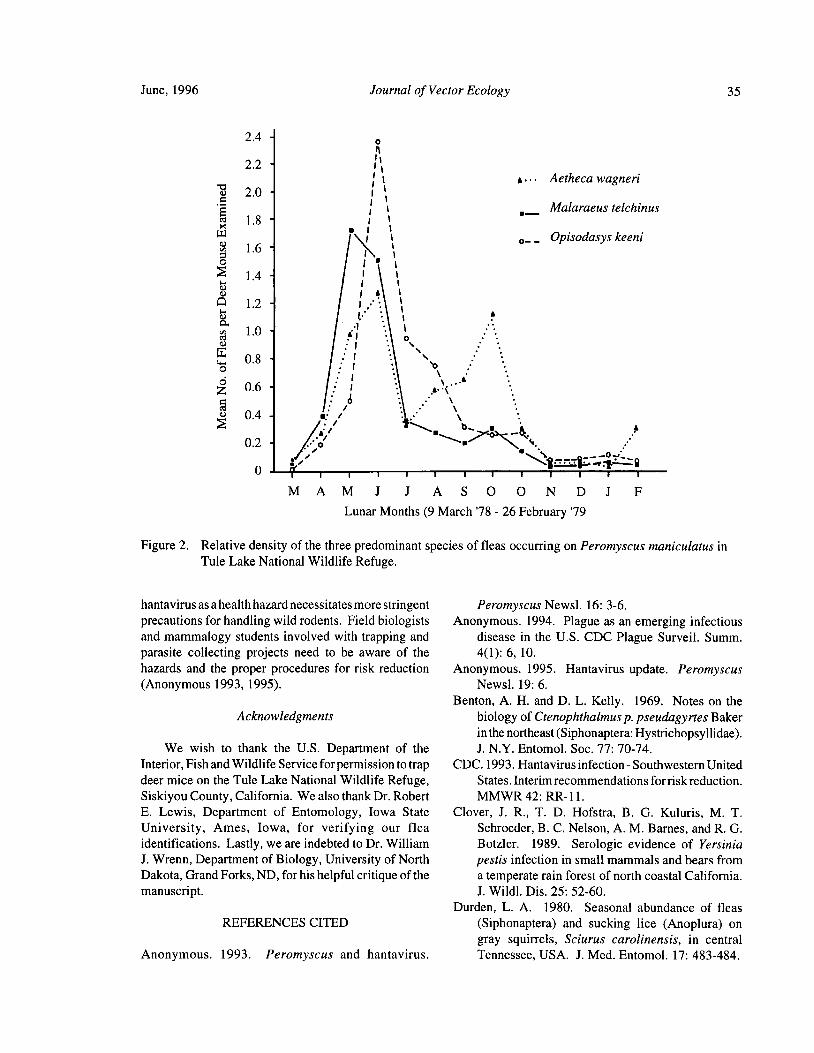
June, 1996 Journal of Vector Ecology 35
2.4c
St
2.2 11 a... Aetheca wagneri
a 11
Malaraeus telchinus
k1. 8
lil0__ Opisodasys keeni
1. 6I
o I
Q
I
1. 4
a) I1. 2 1 ,
ta• 1. 0
o
I
w, 0.8 I
o I b
1
z 0.6 1 i
cl ai b ai
0.2 oos.
cN
0
M A M J J A S O O N D J F
Lunar Months( 9 March' 78- 26 February' 79
Figure 2. Relative density of the three predominant species of fleas occurring on Peromyscus maniculatus inTule Lake National Wildlife Refuge.
hantavirus as a health hazard necessitates more stringent Peromyscus Newsl. 16: 3- 6.
precautions for handling wild rodents. Field biologists Anonymous. 1994. Plague as an emerging infectiousand mammalogy students involved with trapping and disease in the U.S. CDC Plague Surveil. Summ.parasite collecting projects need to be aware of the 4( 1): 6, 10.
hazards and the proper procedures for risk reduction Anonymous. 1995. Hantavirus update. Peromyscus
Anonymous 1993, 1995). Newsl. 19: 6.
Benton, A. H. and D. L. Kelly. 1969. Notes on the
Acknowledgments biology of Ctenophthalmusp. pseudagyrtes Bakerin the northeast( Siphonaptera: Hystrichopsyllidae).
We wish to thank the U.S. Department of the J. N.Y. Entomol. Soc. 77: 70- 74.
Interior, Fish and Wildlife Service forpermission to trap CDC. 1993. Hantavirus infection- Southwestern Uniteddeer mice on the Tule Lake National Wildlife Refuge, States. Interim recommendations for risk reduction.
Siskiyou County, California. We also thank Dr. Robert MMWR 42: RR- 11.
E. Lewis, Department of Entomology, Iowa State Clover, J. R., T. D. Hofstra, B. G. Kuluris, M. T.
University, Ames, Iowa, for verifying our flea Schroeder, B. C. Nelson, A. M. Barnes, and R. G.
identifications. Lastly, we are indebted to Dr. William Botzler. 1989. Serologic evidence of Yersinia
J. Wrenn, Department of Biology, University of North pestis infection in small mammals and bears from
Dakota, Grand Forks, ND, for his helpful critique of the a temperate rain forest of north coastal California.
manuscript. J. Wildl. Dis. 25: 52- 60.
Durden, L. A. 1980. Seasonal abundance of fleas
REFERENCES CITED Siphonaptera) and sucking lice ( Anoplura) ongray squirrels, Sciurus carolinensis, in central
Anonymous. 1993. Peromyscus and hantavirus. Tennessee, USA. J. Med. Entomol. 17: 483- 484.

36 Journal of Vector Ecology June, 1996
Fehrenbach, P. 1994. Hantavirus: are PCOs at risk? roles of various species ofsylvatic rodents in plague
Pest Contr. Technol. 22: 32- 34, 108. ecology in California. Proc. Vert. Pest Conf. 9: 89-Glicken, A. and R. G. Schwab. 1980. Modes of 96.
ectoparasitereinfestationsofdeermice( Peromyscus Nelson, B. C. and C. R. Smith. 1976. Ecological effects
maniculatus). J. Wildl. Dis. 16: 577- 586. of a plague epizootic on the activities of rodents
Hubbard, C. A. 1943. The fleas of California. Pacific inhabiting caves at Lava Beds National Monument,Univ. Bull. 39( 8): 1- 12. California. J. Med. Entomol. 13: 51- 61.
Jay, M. 1994. Hantavirus pulmonary syndrome: Rothschild, M. and B. Ford. 1972. Breeding cycle of theCalifornia update. Vector Ecol. Newsl. 25( 4): 12. flea Cediopsylla simplex is controlled by breeding
Jay, M. and M. B. Madon. 1995. Hantavirus pulmonary cycle of host. Science 178: 625- 626.
syndrome( HPS). Vector Ecol. Newsl. 26( 1): 16. Rutledge, L. C., M. A. Moussa, B. L. Zeller, and M. A.
Lang, J. D. 1993. Fleas found on mammals in San Diego Lawson. 1979. Field studies of reservoirs and
County, California. Bull. Soc. Vector Ecol. 18: vectors of sylvatic plague at Fort Hunter Liggett,
114- 124. California. J. Med. Entomol. 15: 452- 458.
Lewis, R. E., J. H. Lewis, and C. Maser. 1988. The fleas Ryckman, R. E. 1971. Plague vector studies. Part II.
of the Pacific Northwest. Oregon State Univ. Press, The role of climatic factors in determining seasonalCorvallis, 296 pp. fluctuations of flea species associated with the
Mead-Briggs, A. R. and A. J. B. Rudge. 1960. Breeding California ground squirrel. J. Med. Entomol. 8:
of the rabbit flea, Spilopsyllus cuniculi ( Dale): 541- 549.
requirement of a" factor" from a pregnant rabbit for Stark, H. E. and A. R. Kinney. 1969. Abundance ofovarian maturation. Nature, Lond. 187: 1136- rodents and fleas as related to plague in Lava Beds
1137. National Monument, California. J. Med. Entomol.
Nelson, B. C. 1980. Plague studies in California- the 6: 287- 294.

Journal of Vector Ecology 21( 1): 37- 47
Downstream Carry of Larvicides Used in the Control of Pest Black FliesDiptera: Simuliidae) in the Orange River, South Africa
R. W. Palmeri, M. Edwardesz, and E. M. Nevi112
IOnderstepoort Veterinary Institute, do Department of WaterAffairs and Forestry, Private Bag X5912,
Upington, 8800, SOUTH AFRICA
2Entomology Department, Onderstepoort Veterinary Institute,Private Bag X5, Onderstepoort, 0110, SOUTH AFRICA
Received 10 April 1995; Accepted 9 July 1995
ABSTRACT: The downstream carry of larvicides used in the control of the pest black fly Simuliumchutteri( Diptera: Simuliidae) was assessed during 55 field trials in the middle reaches of the Orange River,South Africa. Larvicides were applied in the vicinity ofUpington by means of a boat, a helicopter, and frombridges. Larvicidal carry was determined by assessing the abundance of black fly larvae on 10 to 30substrates( stones or trailing vegetation) at various sites before and after each application. Downstreamcarry was highly variable, but the main factors which affected carry were flow, the presence of pools andvegetation, algal blooms, high concentrations of suspended solids, dosage, and formulation. At medianflows( 100 m3/ s) the bacterium Bacillus thuringiensis var. israelensis( B. t.i.) was usually effective(> 80%
larval mortality) for 6- 9 km, and up to 20 km, downstream of the point of application. By contrast, theorganophosphate temephos was usually effective for 15- 50 km. Carry was further following multiple- sitecontrol) applications compared to single- site( experimental) applications. We conclude that considerable
savings can be made by spacing treatment intervals as far apart as possible.
Keyword Index: Simulium, temephos, Bacillus thuringiensis, black fly.
INTRODUCTION could be both financially and environmentally costlyJackson et al. 1994).
The distance between successive larvicide Two larvicides widely used to control pest blackapplications is an important consideration in the control flies are the bacterium Bacillus thuringiensis var.of pest black flies. This distance is determined by the israelensis( B. t.i.) and the organophosphate temephos.downstream carry of larvicides. Although numerous For B.t.i., carry is usually limited to a few hundredstudies have examined the downstream carry of black meters to a few kilometres ( Lacey and Undeen 1984,fly larvicides, most studies have been undertaken in Colbo and O' Brien 1984, Horosko and Noblet 1983,small, clear streams ( discharge < 10 m3/ s). In these Pistrang and Burger 1984, Knutti and Beck 1987, Merrittstreams, the major expense in the control of black flies et al. 1989). In 1983, B.t.i. was tested against the pestis labor ( Molloy and Struble 1989), and therefore black fly Simulium chutteri in the Orange River andminimizing the use of larvicides is not financially critical. found to be effective for 6 km at a flow of 38 m3/ s( deIn larger rivers, the major expense is larvicide, and it is Moor and Car 1986). Temephos, on the other hand,therefore critical that their use isminimized. Furthermore, carries further than B. t.i. and is effective for up to 40 tomost studies have been based on single-site ( experi- 50 km in large rivers( Kurtak et al. 1987). In general,mental) applications. Extrapolating such results to a small rivers have short carry because ofa lowdepth: widthcontrol program on a large, turbid river, in which several ratio ( Undeen et al. 1984) and usually require moresites are treated simultaneously, may invite errors which application points than large rivers. Likewise, the same

38 Journal of Vector Ecology June, 1996
river at low flow may require more application points mammalophilic blackfly Simuliumchutteri in the middlethan at high flow ( Colbo and O' Brien 1984). reaches of the Orange River, South Africa, was initiated.
Downstream carry is also affected by the physical and The aim of the control program was to reduce the
chemical conditions in the river. In particular, numbers of adult black flies to within acceptable levels.
downstream carry is affected by gradient( Frommer et The Orange River has a median discharge of 100 m3/ s,
al. 1981a) and reduced by the presence of pools( Molloy and the concentration of suspended solids is typically
and Jamnback 1981, Colbo and O' Brien 1984, Lacey 40- 60 mg/ 1 ( Palmer 1994a). The Orange River is
and Undeen 1986) and aquatic vegetation( Frommer et therefore large and turbid compared to rivers on which
al. 1981b, Undeen et al. 1984, Lacey and Undeen 1986). much of the black fly control research has beenFurthermore, larvicidal efficacy ( and presumably its undertaken. Furthermore, the control program needed
carry) is reduced in cold water ( Lacey et al. 1978, to operate within a limited financial budget, and it was
Undeen and Colbo 1980, Molloy et al. 1981, Colbo and therefore important to minimize the number of
O' Brien 1984, Rodrigues and Kaushik 1984, Olejnicek application points without affecting the overall efficacy
et al. 1985, Lacoursiere and Charpentier 1988, Morin et of the control program. The aim of this paper was to
al. 1989), polluted water( Car 1984), acidic water( Lacey determine the appropriate spacing ofsuccessive larvicideet al. 1978, Lacoursiere and Charpentier 1988), and applications for the control of S. chutteri in the middle
turbid water( Ramoska et al. 1982, Morin et al. 1989). reaches of the Orange River.
Larvicidal carry is also affected by the particularformulation used( Guillet and Escaffre 1979, Lacey and STUDY AREA
Undeen 1984, Lacey and Heitzman 1985, Lacey 1986).Given the large number of variables that may affect Physical Conditions
carry, a simple solution for predicting downstream carry Trials were conducted between Westerberg and the
is unlikely. Ideally, downstream carry should be Augrabies Falls, a distance of 345 km ( TABLE 1).
determined for each river in which larvicides are to be Rapids were interspersed by long stretches of pools, f-used. 2 m deep. Average distance between rapids was 3. 8 km
In 1991, a larvicidal control program against the (± 4. 1 SD; max= 33 km; n= 89). With the exception of
TABLE 1. Details of larvicide application sites in the Orange River in the vicinity of Upington.Width" refers to the approximate bank-full width. " Gradient" refers to the drop in
altitude over the first 10 km from the point of application. Sites are arranged in orderdownstream. LC=Left channel; RC=Right channel.
Grid Channel Gradient
Site South East Width( m) Type m/ 10 km)
Westerberg 29° 21' 22°22' 200 Single& braided 6. 5
Groblershoop 28° 53' 22° 59' 300 Single 4. 5
Sishen Bridge 28° 46' 21° 52' 280 Single 5. 5
Grootdrink 28° 33' 21° 45' 190 Single 1. 3
Swartkop 28° 25' 21° 41' 300 Single& braided 5. 6
Gifkloof 1 28° 26' 21° 23' 380 Single& braided 11
Gifkloof 2 as above but downstream of weir)
Upington Bridge 28° 27' 21° 15' 250 Single& braided 10
Sonesta 28° 28' 21° 14' 200 Braided 10
Kanon Island( LC) 28° 38' 21° 05' 140 Braided 11
Kanon Island( RC) 28° 39' 21° 05' 160 Braided 11
Keimoes 28° 43' 21° 59 150 Single& braided 11
Neus Island( LC) 28° 46' 20° 43' 100 Single& braided 24
Neus Island( RC) as above)
Kakamas 28° 45' 20° 37' 250 Single& braided 6.7
Augrabies Falls 28° 35' 20° 20 10 Single 82

June, 1996 Journal of Vector Ecology 39
Augrabies Falls, the river was mostly 100- 300 m wide unavailable, control operations were simulated bywhere it was a single channel; but in places it was applying larvicides at three sites at 5- 6 km intervals. Theextensively braided and up to 3. 5 km wide from the distance upstream of rapids that larvicides were to beextreme north ( right) bank to the extreme south ( left) applied was determined by observing the movement ofbank. Gradient ranged from 1. 3 m/ 10 km, at Grootdrink, dye ( Cartosol Red) through rapids after applying atto 82 m/ 10 km, at Augrabies Falls( measured over 10 km various distances upstream at different discharges. Tendownstream of application points from 1: 5000 scale minutes was regarded as an ideal time for larvicides tomaps). The stream bed consisted largely of granite pass the first rapids.boulders and sandbanks. At Neus Island, the right Larvicidal efficacy was determined according tochannel dropped 15 m over 2 km, whereas, the left the method described by Palmer( 1994b). The methodchannel dropped the same height within the first 0. 3 km. assesses the relative abundance of black fly larvae on aA deep ( 16 m) pool in the left channel separated the 10-point, semi- logarithmic, relative abundance scale atsecond sampling site from the third. three to seven sites downstream of the point of
Experimental application points were chosen based application, before and after each application. At eachon access and availability of sampling sites During site the abundance of larvae on 10 to 30 substrates( rocksmoderate to high flow (> 60 m3/ s) trials were often or trailing vegetation, depending on availability) wereconducted from the Upington road bridge because of the sampled from fast- flowing water. To reduce samplinglong distances between available sampling sites. At low variability, larval abundance was assessed in a 4 x 4 cmflow(< 60 m3/ s) the river at Kanon or Neus Islands was area of highest population density on each replicateusually used for experimental trials because of the short substrate. Rapids upstream of Upington were used as adistances between available sampling sites. Carry trials control. Carry was defined as the downstream distanceconducted during multiple-site ( control) applications that larvicides caused > 80% black fly larval mortality.were restricted to the river downstream of the lowest site Discharge data were obtained from weirs managedtreated. by the Department of Water Affairs and Forestry,
The most numerous riparian plants were reeds Upington. Water temperature and the concentration ofPhragmites spp.), which formed dense stands on the total suspended solids( TSS) were measured at the time
islands, and their trailing edges provided attachment of application. Total suspended solids was determinedsites for black fly larvae. The most common in-stream gravimetrically by filtering a measured volume ofwatermacrophyte was the fennel- leaved pondweed ( 100 to 300 ml depending on clogging) through a pre-Potamageton pectinatus), which formed large stands weighed Whatman GF/C( 0. 6µ m pore diameter) filter
in slow- flowing water(< 0.5 m/ s). Five black fly species and drying for at least 24 hrs at 60° C.were found in the main channel during the study period,the most common of which was the pest Simulium RESULTS AND DISCUSSIONchutteri, which usually exceeded 90% of the black flypopulation in fast- flowing water (> 0.8 m/ s). Other Observations on the movement of dye throughblack fly species present were Simulium mcmahoni, rapids indicated that each rapid is unique, and it isSimulium adersi, Simulium damnosum s. l., Simulium therefore difficult to prescribe recommended distancesruficorne, and Simulium gariepense. upstream of rapids to apply larvicides. Furthermore,
strong winds affected the movement ofdye, and in someMETHODS cases winds blew the dye upstream of the point of
application. As a general guideline, we suggest that inTrials were conducted between August 1990 and the Orange River larvicides should be applied directly
June 1994. The larvicides used were Abate® 200-EC, ( 1 m) upstream of deep (> 2 m) weirs, about 50 mTeknar® HP-D and Vectobac® 12AS. Larvicides were upstream of shallow weirs (< 2 m) and rapids at low-applied undiluted directly across the river from bridges, flow(< 59 m3/ s), 100- 300 m upstream at moderate- flowa boat, or a canoe. During multiple-site( control) appli- ( 60- 143 m3/ s), and > 400 m upstream at high-flowcations, a helicopter fitted with a Simplex® spray tank (> 144 m3/ s).
was used to apply larvicides in a z-pattern across theriver. The volume of larvicides applied was based on B.t.i.
river discharge over 10 minutes. The time taken for A total of 29 carry trials was conducted with B. t.ranged from less than 1 minute ( when of which 21 were single-site( experimental) applications
applied by helicopter) to over 10 minutes when applied and 7 were multiple-site( control) applications( TABLEby boat. On two occasions when the helicopter was 2; Fig. 1). Discharge during the trials ranged from an
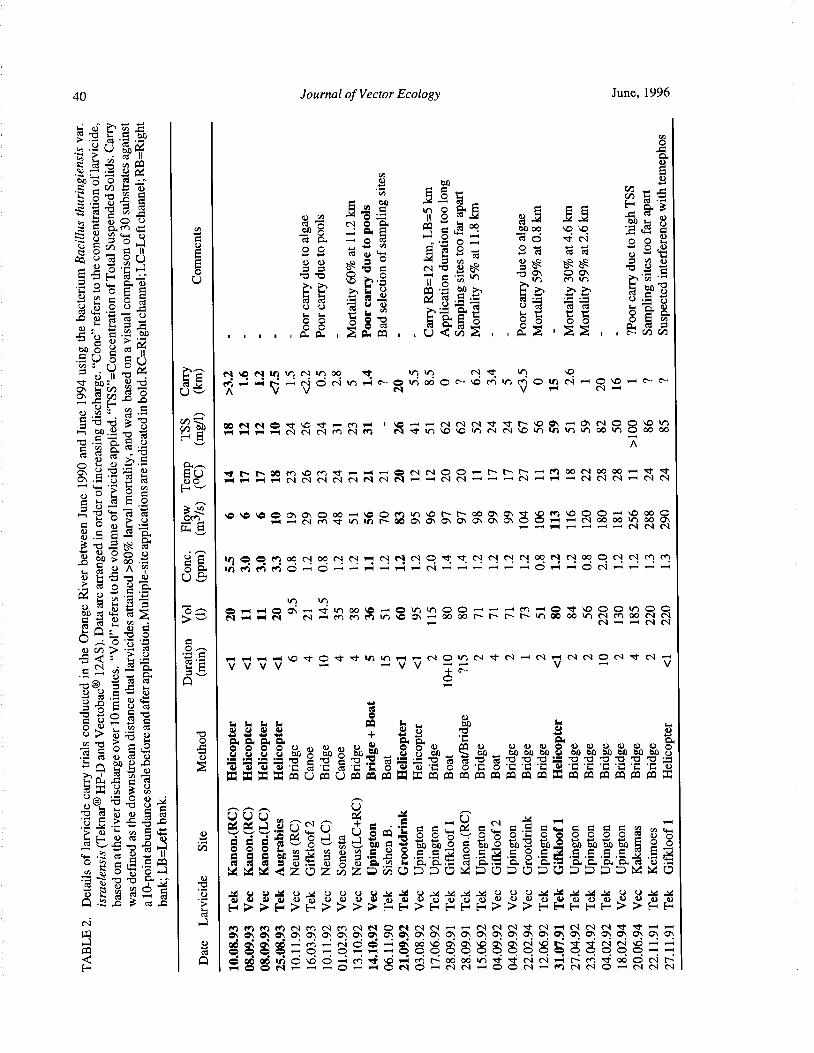
40 Journal of Vector Ecology June, 1996
0
a Cy
a0 0 O a te. bA w
a En
co
N ° NOO
oOA oo V0 , 0y O .ti U a o a o o 18,a a.)
PP 00
oE. 3 a b O'Lla .
CO
U OM
O a U
P p ", 4, y, bo b
ti a 0a
i C St)o U . 6 U U 9 d U i wG. 0VI
a) o a Co o ao0
DA
a
s,0 o asU
N VD N 11 V1 N V1 00 ' V V1 v1 N et V1 VD
b0AII c 'b e) •" 4 -. N O cV v1 4 c• O N oo O C• v6 M h M O kn N .. O VD .. h h
CA a p U.. A V V eV V - N .-ON ..
GO ' C 3 a) bD 00 N N O Tr VD 7r M rl • \ O .. N N N Tt Tt l VD Ch •. CAN O p V0 v1b0.?? . 0 rl rl re rl N N N M N M N 7t v1 V0 V0 v1 N N V0 V7 V) v1 v1 0o V1 800 00
a y a A
O H U 4"C C],
CT . p b OIN h 00 M s. 0 M d• .. IN.) .•••, O N N O O .. e— l h .. M 00 N 00 00 ..
rN rl rl r N N N N N N N N •-+ — N N r• •--- .--. N •--1 rl •--' N N N .. N N
ar. E 0
d 0 m m pen VD VD VDOCTCT 000 .. DOMv1VDNNoocrcr7tV0 f) ' CD CD .. 0000
a o 0 Er -4 N M d• v1 N 00 CT CT OT CT CT CT C'.
CD CD N 00 00 N 00 N0 a cad 0.0.
b E
0 bA >00 .=., g 5 vi O O M 09 N 00 N N r+ Cl N N O V• v, N ( 1 N N00N NOo O NN cc? Ma) A cn
O 04 eA M f7 e l O ...y O .-. .- rl .-. rl .. N .-. .. .- 4 .. .. T. O rl .. 0 N .-. .- 4 .. ..
a 47 • a a) U a
W 0
YOa ' Q
Mt : 1- 1
r.
In v1
a .-, a) C N . tiN N —enonrnv• Ov1 -
000000NNNNv100000000If)
Nen0000CA CA
a
cV w CI • M M e en OT '.
N - —
b oF. _
0is O U
aC o
N > ~
waV V V V
VD Tr Od' Tt v1V
VN0V1Na- N •--4N
VNNON -ctN
vr .--
a Ac-
0 a .0 o U w
0 CI D
o0 -
wa La c. 1. E. CA I. H bUO y a.
C O g w s o 0 o o a) 0 a) 0 a) Ort
0 0 0 a) a) a) N O N a) a) a) N N
CI a) 8 a) i .CJ .. Q .. V. 00 O O ' b .' V. ...c.) bA 0Q
YbA bA bA DO OA 00 bA 00 bU Q
AbA ,
0 2 a a a G a z 0 G ' C 0 v a) z 0 0 o a a G G c G a)aci: = c xxxxPP0PPUaaaar xXPPPP PaaaawaaaaxmPPPPPPP1PP =
x a
ti U
V. •> . b . D U U U y ,N .-• }' i U N .
s4 - 4
ao) o _ Q4" 1 U , . U U oOPC1
a s45 `—' 0 ' 45
p4 a a . E a w a a a a0 p4 0 0 0 0 0 0 0 0 o a) O
F y b a in o 0 0 0 " o .. 5 a a to V',o 0 d to 0 to b to 0 oA to o' Vo § 0 0
11o 'y a c a g l I V E w 8 o e ' a g ' a•a F g ' a 'a 0 ' a. • a•a' a' a'' 'v)
o 0 as WW dZC7Z ,nZDidoCD ) 6 4AC7A6 D0 DAA D W W 6U
o 3 O U y 44 a)) aa a) 0) N
aa)) N 0 • y 0 a) N ' tea) aa)) ) ) .) a) aa
ate) ate) a'a)) N a) 0) a)A • D 3 a
H > > F > E. > > > > E-. F > HE-. F+ F > > > HFFFF+ > > FH
N aM M M M N M N M N N O N N N .— .. N N N ' et N re N N N Tr Tr .. ..
W 0\ C. CS C'. co) C'. ChC'. C'. C'. C'. C; O'. C% a, cr, a., C\ 010\ 01 01CTC'. C; a; a\ C; cr1 0,) cofa, a; oe •-. M r• NOC -. pc; vOa; C vOC\ a; cV vGt: NN4D •-- •--,
tP a 00 CDCD .. 0 .. CD . .. .-. 000000000000000 •--x •--
A C 03 0C ui O O -. M 4 v6 4 M N 00 06 vi 4 N N rl N M oo O cV r`F rl O O N .. .. .. C) .. .. 001 O -- N N .. O O N •-+ M N N O •-. N N N
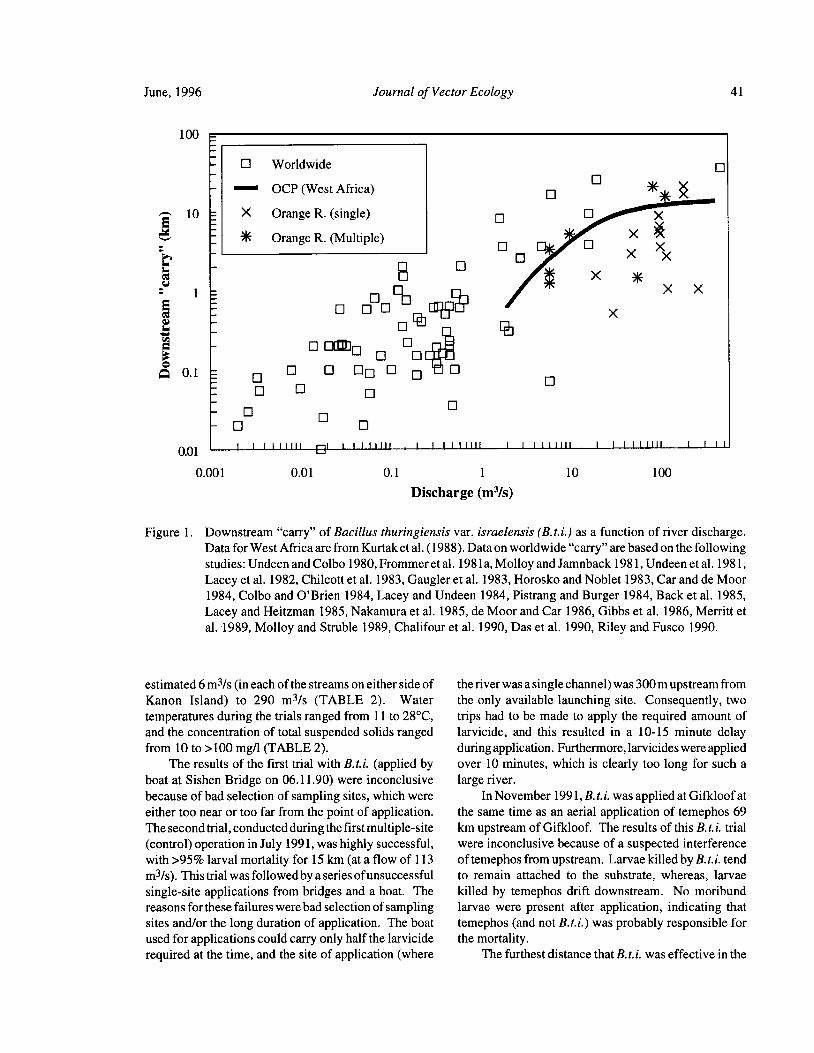
June, 1996 Journal of Vector Ecology 41
100 t
Worldwide
OCP( West Africa)
10 _ X Orange R. (single) X
Orange R. (Multiple) X kX XX
92u
EX
mat
X X
D X
3 ICM 0. 111
q 0. 1 = II
O
0.01I I i 1 1 1 1 1 DI i i I l l I I I . i i i I I I I I I I I I I 1 1 1 1 1 I I I I I 1 1 1 1 I I I I
0.001 0.01 0. 1 1 10 100
Discharge( m3/s)
Figure 1. Downstream" carry" of Bacillus thuringiensis var. israelensis( B.t.i.) as a function of river discharge.Data for West Africa are from Kurtak et al.( 1988). Data on worldwide" carry" are based on the followingstudies: Undeen and Colbo 1980, Frommer et al. 1981a, Molloy and Jamnback 1981, Undeen et al. 1981,Lacey et al. 1982, Chilcott et al. 1983, Gaugler et al. 1983, Horosko and Noblet 1983, Car and de Moor1984, Colbo and O' Brien 1984, Lacey and Undeen 1984, Pistrang and Burger 1984, Back et al. 1985,Lacey and Heitzman 1985, Nakamura et al. 1985, de Moor and Car 1986, Gibbs et al. 1986, Merritt etal. 1989, Molloy and Struble 1989, Chalifour et al. 1990, Das et al. 1990, Riley and Fusco 1990.
estimated 6 m3/ s( in each of the streams on either side of the river was a single channel) was 300 m upstream from
Kanon Island) to 290 m3/ s ( TABLE 2). Water the only available launching site. Consequently, twotemperatures during the trials ranged from 11 to 28° C, trips had to be made to apply the required amount ofand the concentration of total suspended solids ranged larvicide, and this resulted in a 10- 15 minute delayfrom 10 to> 100 mg/ 1( TABLE 2). during application. Furthermore, larvicides were applied
The results of the first trial with B.t.i. ( applied by over 10 minutes, which is clearly too long for such aboat at Sishen Bridge on 06. 11. 90) were inconclusive large river.
because of bad selection of sampling sites, which were In November 1991, B.t.i. was applied at Gifkloof at
either too near or too far from the point of application. the same time as an aerial application of temephos 69
The second trial, conducted during the first multiple-site km upstream of Gifkloof. The results of this B.t.i. trial
control) operation in July 1991, was highly successful, were inconclusive because of a suspected interference
with> 95% larval mortality for 15 km( at a flow of 113 of temephos from upstream. Larvae killed by B. t.i. tendm3/ s). This trial was followed by a series ofunsuccessful to remain attached to the substrate, whereas, larvae
single-site applications from bridges and a boat. The killed by temephos drift downstream. No moribund
reasons for these failures were bad selection of sampling larvae were present after application, indicating thatsites and/or the long duration of application. The boat temephos( and not B.t.i.) was probably responsible forused for applications could carry only half the larvicide the mortality.required at the time, and the site of application( where The furthest distance that B.t.i. was effective in the

42 Journal of Vector Ecology June, 1996
Orange River was 20 km. This was achieved following Discharge during the trials ranged from 10 to 298 m3/ shelicopter applications upstream of Grootdrink, at a ( TABLE 3). Water temperatures during the trials rangedflow of 83 m3/ s, as well as following a high-dosage( 2. 0 from 9 to 29° C, and the concentration of total suspendedppm) single-site application from the Upington Bridge solids ranged from 14 to 99 mg/ I ( TABLE 3).when the flow was high( 180 m3/ s). A subsequent high- The first trial, conducted at Sishen Bridge in Augustdosage trial at a lower flow( 96 m3/ s) was effective for 1990, showed a complete kill of black fly larvae at thebetween 5 km( left bank) and 12 km( right bank). At first sampling site, 0.3 km downstream of the applicationvery low flow( 6 to 10 m3/ s), high dosages ( 3. 0 to 5. 5 point, but no significant kill at the second site, 6 kmppm) were effective for 1. 2 to 3. 2 km. A low-dosage downstream. It is likely that the duration of applicationapplication( 0. 8 ppm) at moderate flows( 120 m3/ s) was ( 9 minutes) was too long for a river of this size. Theeffective for 1 km, with partial control( 59%) at 2. 6 km. second trial, conducted at Sishen Bridge in October
Subsequent low-dosage applications were effective for 1990, was inconclusive because of bad selection of0 to 1. 5 km. Although increasing dosages increases sampling sites. The following two trials, conductedcarry, the relationship is not proportional ( Lacey and from the Upington and Keimoes Bridges in NovemberUndeen 1984). Poor carry at Upington and Neus Island 1991, when flow was high ( 298 and 293 m3/ s,left channel) during low- flow(< 60 m3/ s) was attributed respectively) were highly successful. Effective carry
to pools. from the Upington Bridge was between 45. 1 and 56.2Larvicides were often ineffective on the edge of the km, while carry downstream of the Keimoes Bridge
river but worked well towards the middle. This was exceeded 37 km. Likewise, larvicides applied from the
particularly noticeable downstream of large stands of Kakamas Bridge at high flow( 181 m3/ s) were effectivePhragmites sp. reeds. Aquatic vegetation is well- known for 50 km. Multiple-site control operations upstream offor reducing larvicidal efficacy( Frommer et al. 1981b, Grootdrink during high flow( 290 m3/ s) were not intendedUndeen et al. 1984, Lacey and Undeen 1986). It is as a carry trial, but it was suspected that the temephostherefore essential that upstream aquatic vegetation be interfered with aB. t.i. trial 69 km downstream at Gifidoofavoided when taking larval pre- and post counts. Weir. Multiple-site applications of temephos during
In March 1993, there was a bloom of blue- green high- flow(> 95 m3/ s) in the Orange River carry for atalgae Microcystis sp. during which the number of algal least 50 km. During low-flow( 10 to 29 m3/ s) temephoscells exceeded 15, 000 per ml. A trial conducted under carried 2 to 3 km when applied at the recommendedthese conditions( 16. 03. 93) was unsuccessful, and caused dosage of 0.05 ppm.
only 52-66% mortality at 2.2 km downstream. Likewise, In May 1992, a trial from the Upington Bridge wasB. t.i. applied from Grootdrink Bridge in February 1994, conducted at a flow of 107 m3/ s. Mortality at the firstwhen the planktonic algal concentration was 12, 000 sampling site, 0. 8 km downstream, was low( 56%), but
cells/ ml, did not reach the first sampling site 3. 5 km reached> 99% at 4. 3 km downstream. Effective carrydownstream. In June 1994, the flow was high( 256 m3/ was about 13 km. A similar result was obtained ats), and a trial conducted from the Kakamas Bridge Westerberg in March 1993, in which temephos was notindicated a carry of 1 km only. The reason for the poor effective within the first 0.7 km. Thereafter, larval
carry was thought to be related to the high concentration mortality exceeded 99% for at least 7. 5 km. A suggestionof total suspended solids( TSS). Unfortunately, the TSS by Dr. Dan Kurtak to observe larvicides in a glasswas not measured, but the Secchi depth value at the time cylinder filled with water showed that the temephoswas 10 cm, indicating a TSS exceeding 100 mg/1( Palmer used was extremely buoyant and remained on the surface1994a). of the water for several days without mixing. This could
Although B.t.i. is less effective at low temperatures explain the poor efficacy within the first 0.8 km ofthan at high temperatures( Lacey et al. 1978, Molloy et application.
al. 1981, Morin et al. 1989), it worked well in the Orange Multiple-site applications conducted in winter whenRiver at 11° C ( TABLE 2) and is effective ( 70%) at spot water temperatures were 12° C and the flow 95 m3/temperatures as low as 3° C( Undeen and Colbo 1980). s, resulted in carry for at least 50 km. However, a single-However, increased dosages may be required at site application during cold conditions ( 9° C) was nottemperatures< 10° C( Colbo and O' Brien 1984). effective. Temephos has been shown to have no effect
at 7- 9° C( Fredeen 1987) and little effect at temperaturesTemephos less than 18° C( Back et al. 1979, Rodrigues and Kaushik
A total of 26 carry trials was conducted with 1984).
temephos, of which 5 were multiple-site applications A single-site application performed from theand 21 were single-site applications( TABLE 3; Fig. 2). Upington Bridge in September 1992, when the flow was
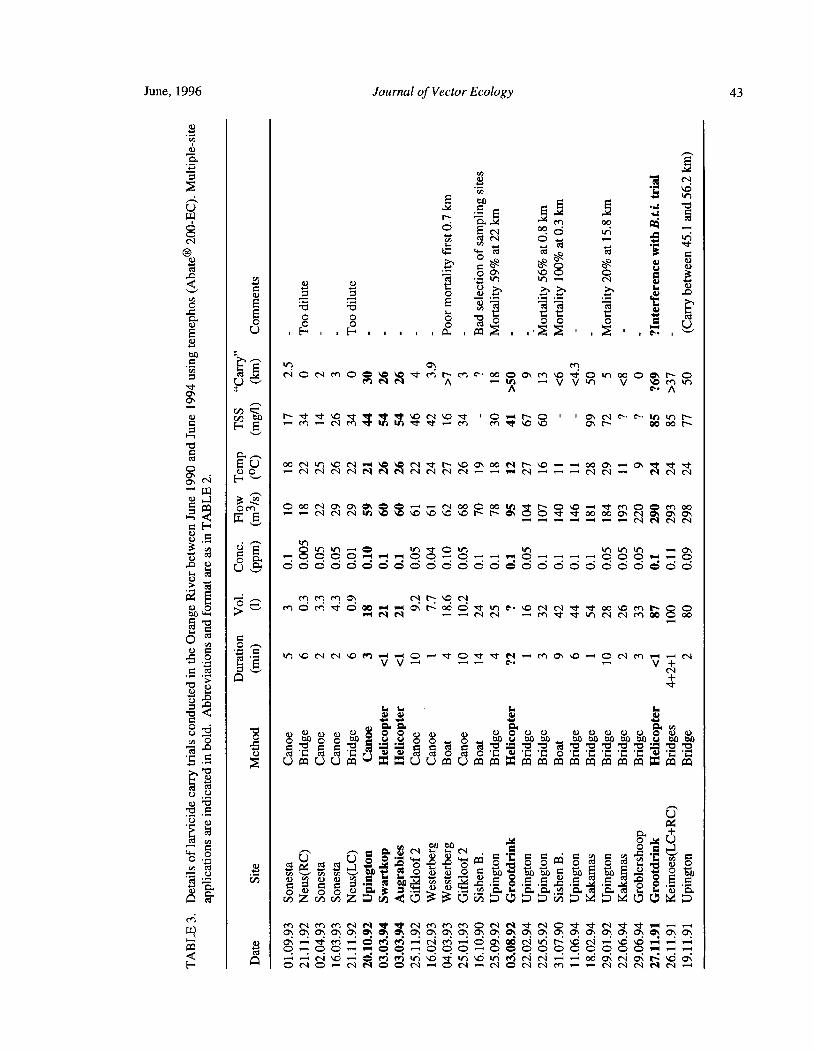
June, 1996 Journal of Vector Ecology 43
v)
0
w
M 00
8 O
aN to O
O v1N a a t
wo a ao
0 h VD 0 ON 0
a, 00 0 0 ' b o 0 0 0
EU , E2 , .
Fo- . . . , . a . G4 , 4 . I. 22 , . 2 . . e.. ,
u
au
v ON enFAa
UN 0 N en 0 M N N
M
nen e- 0000 © ON en v 0 v) o 0 0 M 0O
A C• AONO
Ca> bA N VD ' t Tr VO N O , 0 N 0 1 , OT N r• r• 1A v1 N
E„ a E - M a N M an an V a M M er vs s.O O\ N 00 00 N
o a
E U 0o N tr) VD N - , O \ O N 4 N vs O\ 0o N N '.O -. 0o ON •- 4 O` d• ..1'O a) O — N N N N N N N N N N N •--+ •-. .•1N .- i '- - N N — 7t N Nrn ( Ni F' ..
c aO M O 00 N a,
Nen O O '. O N 0o O oo N 0 O v)
000 M 0 CO en 00
g Q Ea a N N N 1n O O O O vO VD S S ON O 0 00 00 a1 N O Osi a1
CN N N N
3 n ci ^ n O v) 4 O v v in v v --- O0 0. 00. 0 ti . I O O ,-. 0 •-4 -+ I O ^ ,. ", ... O O O 1. 4 O0 aU a 6 6 6 0 6 0 0 0 6 6 6 6 6 6 0 0 6 6 6 6 0 6 6 0 6 6
No liia en en en O'. N N '.O N
a) oO '
M O Cr; 4 O 00 .-+ a T N 00 O h C• VD N N t t 00 " O en N p 0
Cw N N 1 — N N — en d N N en 00 8 00
N 0a _
r •• oa
in '. O N N '.O en V v 0 a 4 O 4 c ---- en O'. '. 0 .--. Oa.
N en
v + N
a c> O
A
6) wv
oa
w y oQ 4141.) a a)a
OU b 8 a) O a) a> 0 o 0 O a) a> 0 o 0 0 a) a) a) a> o a) O a) a)a a a a 0 0 ` b u
S d b - - 0 cd • 0 b b ct b 'b"0 b'.. 'O' Oa
0 n • C a cd • C U as a+ a n o n o • C as • C • C o • C • C • C • C • C as • C • CE 2 UaaUUOa x xUUaoUaaraxraaaaaaaa1MPC1faxwaa
o , c4
cub0
Ua> A~
a +
oo too ,g U
o ..-1- o U U a o ° w ) w a C 0 a a C y y 0
a) .5 C4 . 5 . 5 . i o r F2 00 a a b 01; n
o2a
o2g tt, '
0
nvs,g,
a• 0 . 0 o o a 3 0 ) a •— v 'o. $ a' c.,a a ' ' a ' ° $ 0 a
Men N cn en N N ' c!' N en M cn 0 N N N 0 N l .--, ,--.
W O Q\ ON O\ O), ON O• 0) O\ Oh 0) Os O1 at O, Os ON 0\ O\ ON 0) 01 O) O)
a) O: ,--, 4 M ^- C M M a N M r 6 O oo N cn N "0 N a v0 vs 4 . .O . O O - 1 , 4 O O . O O O . O O O O O O O O O O . .
H A O N O N Net.;
O N O N N O N N Mono O; N O: P . 0 0;
N N N N N •-+
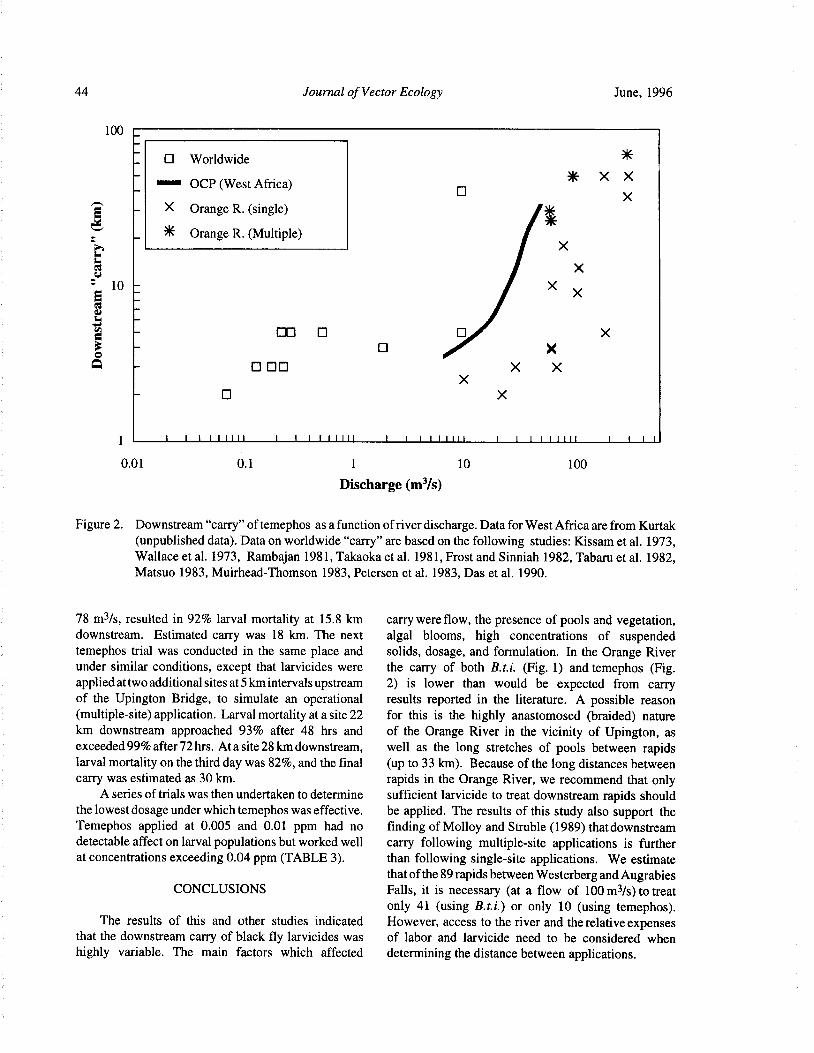
44 Journal of Vector Ecology June, 1996
100
Worldwide
OCP( West Africa) CIE X X
XX Orange R.( single)
Orange R.( Multiple)X
hoE: X
XX
X
XA - X X
X
X
1 1 1 1 1 1 1 1 1 1 I I 1 1 11111 I I 1 1 1 1 111 1 I 1 111111 1 I I I
0.01 0. 1 1 10 100
Discharge( m3/ s)
Figure 2. Downstream" carry" of temephos as a function of river discharge. Data for West Africa are from Kurtakunpublished data). Data on worldwide" carry" are based on the following studies: Kissam et al. 1973,
Wallace et al. 1973, Rambajan 1981, Takaoka et al. 1981, Frost and Sinniah 1982, Tabaru et al. 1982,Matsuo 1983, Muirhead-Thomson 1983, Petersen et al. 1983, Das et al. 1990.
78 m3/ s, resulted in 92% larval mortality at 15. 8 km carry were flow, the presence of pools and vegetation,downstream. Estimated carry was 18 km. The next algal blooms, high concentrations of suspendedtemephos trial was conducted in the same place and solids, dosage, and formulation. In the Orange Riverunder similar conditions, except that larvicides were the carry of both B.t.i. ( Fig. 1) and temephos ( Fig.applied at two additional sites at 5 km intervals upstream 2) is lower than would be expected from carryof the Upington Bridge, to simulate an operational results reported in the literature. A possible reasonmultiple-site) application. Larval mortality at a site 22 for this is the highly anastomosed ( braided) nature
km downstream approached 93% after 48 hrs and of the Orange River in the vicinity of Upington, asexceeded 99% after 72 hrs. At a site 28 km downstream, well as the long stretches of pools between rapidslarval mortality on the third day was 82%, and the final ( up to 33 km). Because of the long distances betweencarry was estimated as 30 km. rapids in the Orange River, we recommend that only
A series of trials was then undertaken to determine sufficient larvicide to treat downstream rapids shouldthe lowest dosage under which temephos was effective. be applied. The results of this study also support theTemephos applied at 0.005 and 0.01 ppm had no finding of Molloy and Struble( 1989) that downstreamdetectable affect on larval populations but worked well carry following multiple- site applications is furtherat concentrations exceeding 0.04 ppm( TABLE 3). than following single-site applications. We estimate
that of the 89 rapids between Westerberg and AugrabiesCONCLUSIONS Falls, it is necessary ( at a flow of 100 m3/ s) to treat
only 41 ( using B.t.i.) or only 10 ( using temephos).The results of this and other studies indicated However, access to the river and the relative expenses
that the downstream carry of black fly larvicides was of labor and larvicide need to be considered when
highly variable. The main factors which affected determining the distance between applications.

June, 1996 Journal of Vector Ecology 45
Acknowledgments Nematocera) in the middle Orange River.
Onderstepoort J. Vet. Res. 53: 43- 50.
We thank Morgan Scheepers and Danie de Klerk Fredeen, F. J. H. 1987. Black flies: approaches to
for help in the field, Dr. Dan Kurtak for unpublished population management in a large temperate- zone
data and useful suggestions regarding the control river system. Pp 295- 304 in Black Flies: Ecology,program, members of Resource Conservation, Depart- Population Management, and annotated world list
ment ofAgriculture, for their assistance during multiple- K. C. Kim and R. W. Merritt, eds.). Pennsylvania
site control operations, the Department ofWater Affairs State University, xv+ 528 pp.
and Forestry, Upington, for the use of their facilities, Frommer, R. L., S. C. Hembree, J. H. Nelson, M. P.
and the Water Research Commission for funding. Remington, and P. H. Gibbs. 1981a. The evaluation
ofBacillus thuringiensis var. israelensis in reducing
REFERENCES CITED Simulium vittatum( Diptera: Simuliidae) larvae intheir natural habitat with no extensive aquatic
Back, C., J. Boisvert, J. O. Lacoursiere, and G. vegetative growth. Mosq. News 41( 2): 339- 347Charpentier. 1985. High-dosage treatment of a Frommer, R. L., J. H. Nelson, M. P. Remington, and P.
Quebec stream with Bacillus thuringiensis serovar. H. Gibbs. 1981b. The effects of extensive aquatic
israelensis: efficacy against black fly larvae vegetative growth on the distribution of Bacillus
Diptera: Simuliidae) and impact on nontarget thuringiensis var. israelensis in flowing water.
insects. Can. Entomol. 117: 1523- 1534. Mosq. News 41( 4): 713- 724Back, C., J. G. Lanouette, and A. Aubin. 1979. Frost, S. and L. B. Sinniah. 1982. Effect of particulate
Preliminary tests on the use of temephos for the Abate insecticide on invertebrate stream drift
control of black flies ( Diptera: Simuliidae) in communities in Newfoundland. Int. J. Environ.
northern Quebec. Mosq. News 39: 762- 767. Studies 19( 3/ 4): 231- 243.
Car, M. 1984. Laboratory and field trials with two Gaugler, R., B. Kaplan, C. Alvarado, J. Montoya, and
Bacillus thuringiensis var. israelensis products for M. Ortega. 1983. Assessment of Bacillus
Simulium( Diptera: Nematocera) control in a small thuringiensis serotype 14 and Steinernema feltiae
polluted river in South Africa. Onderstepoort J. Nematoda: Steinernematidae] for control of the
Vet. Res. 51: 141- 144. Simulium vectors of Onchocerciasis in Mexico.
Car, M. and F. C. de Moor. 1984. The response of Vaal Entomophaga 28( 4): 309- 315
River drift and benthos to Simulium ( Diptera: Gibbs, K. E., F. C. Brautigam, C. S. Stubbs, and L. M.
Nematocera) control using Bacillus thuringiensis Zibilske. 1986. Experimental applications of B. t.i.
H- 14). Onderstepoort J. Vet. Res. 51: 155- 160. for larval black fly control: persistence and
Chalifour, A., J. Boisvert, and C. Back. 1990. downstream carry, efficacy, impact on nontargetOptimization of insecticide treatments in rivers: an invertebrates and fish feeding. Maine Agric. Exp.application of graph theory for planning a black fly Station Techn. Bull. 123: 1- 22.
larvae control program. Can. J. Fish. Aquat. Sci. Guillet, P. and H. Escaffre. 1979. Evaluation ofBacillus
47: 2049- 2056. thuringiensis israelensis Barjac for the control of
Chilcott, C. N., J. S. Pillai, and J. Kalmakoff. 1983. larvae of Simulium damnosum s. l. 11. Compared
Efficacy of Bacillus thuringiensis var. israelensis efficacy of three experimental preparations ( Inas a biocontrol agent against larvae of Simuliidae French). Unpublished document WHONBC/79
Diptera) in New Zealand. New Zealand J. Zool. 735 ( 1979), WHO Vector Biology and Control
10: 319- 326. Geneva Switzerland, 1- 7 pp.
Colbo, M. H. and H. O' Brien. 1984. A pilot black fly Horosko, S. III and R. Noblet. 1983. Efficacy ofBacillus
Diptera: Simuliidae) control program using thuringiensis var. israelensis for control of black
Bacillus thuringiensis var. israelensis in fly larvae in South Carolina. J. Georgia Entomol.Newfoundland. Can. Entomol. 116: 1085- 1096. Soc. 18( 4): 531- 537.
Das, S. C., D. R. Nath, M. Bhuyan, N. G. Gas, and S. Jackson, J. K.,B. W. Sweeney, T.L. Bott, J. D. Newbold,Hazarika. 1990. Field trial of Abate and Teknar for and L. A. Kaplan. 1994. Transport of Bacillus
Simulium( Diptera: Simuliidae) control in India. J. thuringiensis var. israelensis and its effect on drift
Am. Mosq. Cont. Assoc. 6( 1): 135- 137 and benthic densities of nontarget macro-
de Moor, F. C. and M. Car. 1986. A field evaluation of invertebrates in the Susquehanna River, northern
Bacillus thuringiensis var. israelensis as abiological Pennsylvania. Can. J. Fish. Aquat. Sci. 51: 295- 314
control agent for Simulium chutteri ( Diptera: Kissam, J. B., R. Noblet, and H. S. Moore, IV. 1973.

46 Journal of Vector Ecology June, 1996
Simulium: field evaluation of abate larvicide for against black fly larvae ( Diptera: Simuliidae). J.control in an area endemic forLeucocytozoon smithi Am. Mosq. Cont. Assoc. 4( 1): 64- 72.of turkeys. J. Econ. Entomol. 66( 2): 426-428. Matsuo, K. 1983. Aplicacion de larvicida solido de
Knutti, H. J. and W. R. Beck. 1987. The control of black temefos en el control de la onchocercosis enfly larvae with Teknar. Pp. 409-418 in Black Flies: Guatemala. Boletin de la Oficina SanitariaEcology, Population Management, and annotated Panamericana 95( 5): 412- 417world list ( K. C. Kim and R. W. Merritt, eds.). Merritt, R. W., E. D. Walker, M. A. Wilzbach, K. W.Pennsylvania State University, xv+ 528 pp. Cummins, and W. T. Morgan. 1989. A broad
Kurtak, D. M., J. Grunewald, and D. A. T. Baldry. 1987. evaluation ofB.t.i. for black fly( Diptera: Simuliidae)Control of black fly vectors of Onchocerciasis in control in a Michigan River: efficacy, carry andAfrica. Pp. 341- 362 in Black Flies: Ecology, nontarget effects on invertebrates and fish. J. Am.Population Management, and annotated world list Mosq. Cont. Assoc. 5( 3): 397-415K. C. Kim and R. W. Merritt, eds.). Pennsylvania Molloy, D. P., R. Gaugler, and H. Jamnback. 1981.
State University, xv+ 528 pp. Factors influencing efficacy of Bacillus thurin-Kurtak, D., M. Ocran and M. Meyer. 1988. Evaluation giensis var. israelensis as a biological control agent
ofB.t.H14 formulations as larvicides against larvae of black fly larvae. J. Econ. Entomol. 74( 1): 61- 64.of Simulium damnosum s. 1. in West Africa. WHO/ Molloy, D. P. and H. Jamnback. 1981. Field evaluationOCP report No 12/ 88. 3 pp+ tables and figures. ofBacillus thuringiensis var. israelensis as a black
Lacey, L. A. 1986. Development ofoperational formu- fly biocontrol agent and its effect on nontargetlations of Bacillus thuringiensis var. israelensis stream insects. J. Econ. Entomol. 74: 314- 318and Bacillus sphaericus for vector control. Pp. Molloy, D. P. and R. H. Struble. 1989. Investigation of497- 500 in Fundamental and applied aspects of the feasibility of the microbial control ofblack fliesinvertebrate pathology( R. A. Sampson, J. M. Vlak Diptera: Simuliidae) with Bacillus thuringiensisand D. Peters, eds.). Foundation of the fourth var. israelensis in the Adirondack Mountains ofInternational Colloquium of invertebrate pathology, New York. Bull. Soc. Vect. Ecol. 14( 2): 266-276Wageningen, The Netherlands. Morin, A., C. Back, J. Boisvert, and R. H. Peters. 1989.
Lacey, L. A., H. Escaffre, B. Philippon, A. Seketeli, and A conceptual model for the estimation of theP. Guillet. 1982. Large river treatment withBacillus sensitivity of black fly larvae to Bacillusthuringiensis ( H- 14) for the control of Simulium thuringiensis var. israelensis. Can. J. Fish. Aquat.damnosum s. l. in the Onchocerciasis Control Sci. 46: 1785- 1792Programme: preliminary trials with Sandoz 402 Muirhead-Thomson, R. C. 1983. Time/concentrationformulation. Tropenmedizin and Parasitologie 33: impact of the Simulium larvicide, Abate®, and its97- 101.
relevance to practical control programs. Mosq.Lacey, L. A. and C. M. Heitzman. 1985. Efficacy of News 43( 1): 73- 76.
flowable concentrate formulations of Bacillus Nakamura, Y., A. Kanayama, H. Sato, K. Fujita, Y.thuringiensis var. israelensis against black flies Tabaru, and A. Shimada. 1985. Field trials ofDiptera: Simuliidae). J. Am. Mosq. Cont. Assoc. Bacillus thuringiensis var. israelensis against black
1( 4): 493- 497 fly larvae in Japan. Jap. J. Sanit. Zool. 6( 4): 371-Lacey, L. A., M. S. Mulla, and H. T. Dulmage. 1978. 373.
Some factors affecting the pathogenicity ofBacillus Olejnicek, J., V. Matha, and J. Weiser. 1985. Thethuringiensis Berliner against black flies. Environ. efficacy of Bacillus thuringiensis var. israelensisEntomol. 7: 583- 588. against larvae of the black fly Odagmia ornata
Lacey, L. A. and A. H. Undeen. 1984. Effect of Simuliidae) at low temperatures. Folia Parasitolformulation, concentration, and application time Prague) 32: 271- 277on the efficacy of Bacillus thuringiensis ( H- 14) Palmer, R. W. 1994a. Biological and chemical controlagainst black fly( Diptera: Simuliidae) larvae under of blackflies ( Diptera: Simuliidae) in the Orangenatural conditions. J. Econ. Entomol. 77: 412-418 River. Water Research Commission Report No
Lacey, L. A. and A. H. Undeen. 1986. Microbial control 343/ 1/ 94, Pretoria. 106 pp.of black flies and mosquitoes. Ann. Rev. Entomol. Palmer, R. W. 1994b. A rapid method of estimating the31: 265- 296 abundance of immature blackflies( Diptera: Simu-
Lacoursiere, J. O. and G. Charpentier. 1988. Laboratory liidae). Onderstepoort J. Vet. Res. 61: 117- 126study of the influence of water temperature and pH Petersen, J. L., A. J. Adames, and L. de Leon. 1983.on Bacillus thuringiensis var. israelensis efficacy Bionomics and control of black flies ( Diptera:

June, 1996 Journal of Vector Ecology 47
Simuliidae) at the Fortuna Hydroelectric Project, vector control in Guatemala. Jap. J. Sanit. Zool. 33:Panama. J. Med. Entomol. 20: 399- 408 369- 377.
Pistrang, L. A. and J. F. Burger. 1984. Effects of Takaoka, H., J. O. Ochoa, M. Takahashi, and H.
Bacillus thuringiensis var. israelensis on a gene- Takahashi. 1981. Evaluation of temephos as a
tically-defined population of black flies( Diptera: larvicide against Simulium ochraceum ( Diptera:
Simuliidae) and associated insects in a montane Simuliidae) in Guatemala. J. Med. Entomol. 18:
New Hampshire stream. Can. Entomol. 116: 975- 145- 152
981 Undeen, A. H. and M. H. Colbo. 1980. The efficacy ofRambajan, I. 1981. Abate for the Control of Black fly in Bacillus thuringiensis var. israelensis against
the River Burru, Lethem Area, Rupununi, Guyana. blackfly larvae( Diptera: Simuliidae) in their naturalTropical Geogr. Med. 33( 1): 55- 57. habitat. Mosq. News 40: 181- 184.
Ramoska, W. A., S. Watts, and R. E. Rodriguez. 1982. Undeen, A. H., L. A. Lacey, and S. W. Avery. 1984. AInfluence of suspended particulates on the activity system for recommending dosage of Bacillusof Bacillus thuringiensis Serotype H- 14 against thuringiensis( H- 14) for control of Simuliid larvae
mosquito larvae. J. Econ. Entomol. 75: 1- 4. in small streams based upon stream width. Mosq.
Riley, C. M. and R. Fusco. 1990. Field efficacy of News 44: 553- 559
Vectobac - 12AS and Vectobac - 24AS against Undeen, A. H., H. Takaoka, and K. Hansen. 1981. A test
black fly larvae in New Brunwick streams( Diptera: of Bacillus thuringiensis var. israelensis de Barjac
Simuliidae). J. Am. Mosq. Cont. Assoc. 6: 43- 46. as a larvicide for Simulium ochraceum, the central
Rodrigues, C. S. and N. K. Kaushik. 1984. The effect of American vector of Onchocerciasis. Mosq. Newstemperature on the toxicity of Temephos to black 41: 37- 40.
fly ( Diptera: Simuliidae) larvae. Can. Entomol. Wallace, R. R., A. S. West, A. E. R. Downe, and H. B.
116: 451- 455 N.Hynes. 1973. The effect ofexperimental blackflyTabaru, Y., Y. Nakamura, A. J. Onofre Ochoa, P. A. Diptera: Simuliidae) larviciding with Abate®,
Molina, and H. Takahasi. 1982. Preliminary filed Dursban®, and Methoxychlor on stream
study on larvicide formulations for onchocerciasis invertebrates. Can. Entomol. 105( 6): 817- 831.

Journal of Vector Ecology 21( 1): 48- 59
Timing of Larvicide Treatments for the Control of Pest Black Flies
Diptera: Simuliidae) in a Semi-Arid Environment in South Africa
R. W. Palmer!, M. Edwardes2, and E. M. Nevi112
IOnderstepoort Veterinary Institute, c% Department of WaterAffairs and Forestry, Private Bag X5912,
Upington, 8800, SOUTH AFRICA
2Entomology Department, Onderstepoort Veterinary Institute, Private Bag X5,Onderstepoort, 0110, SOUTH AFRICA
Received 10 April 1995; Accepted 9 July 1995
ABSTRACT: Seasonal changes in the abundance of larvae, the rate of larval development, and the survivaladults of the pest black fly Simulium chutteri Lewis were used to determine the timing of larvicidetreatments in the middle and lower reaches of the Orange River, South Africa. Larval abundance washighest in late winter( July/August), low in late summer( January/ February), and dropped sharply duringblue- green algal blooms in autumn( March/April). The time taken for the first cohort of larvae to pupate,following larvicide treatment, ranged from seven days in midsummer ( water temperature 25- 29°C) to 37days in midwinter( 10- 16° C). Adult abundance was consistently high in spring( September and October)and sometimes in autumn ( April and May). Abundance dropped sharply in summer when evaporationexceeded 12 mm/ day. The estimated number ofgenerations per year was 11- 13, although adequate controlduring normal( non- flood) years was obtained with six" spring" and three" autumn" larvicide applications.It is recommended that applications to prevent a spring outbreak should start towards the end ofJuly, whenmost of the population is in the larval stage, and continue into late October. Applications to prevent anautumn outbreak, when necessary, should start in mid-March and continue to the end of April. Althoughwater temperatures provided reasonably accurate information for the correct timing of treatments, single-site applications underestimated the appropriate timing of subsequent large- scale control treatments.
Treatment intervals should be based on water temperature according to the function y= exp( 0.065t-4. 13),where y=larval development rate( in days- 1), and t= water temperature( in° C).
Keyword Index: Simulium, black fly, Orange River.
INTRODUCTION addition, food quality, parasitism, larval density,hydrological conditions, and the species of black fly
Correct timing of larvicide applications is critical may also be important( Colbo and Porter 1979, Rossfor effective control of pest black flies. Ideally, and Merritt 1987, Crosskey 1990: 135, Davis et al.applications should start when the majority of the 1992). For example, growth rate was found to be fasterpopulation is in the late larval stage( 4th to 6th in star) during the initial stages of high discharge, presumablyWestwood and Brust 1981, de Moor 1982a). Successive because of an increase in seston ( food) quality and
treatments should be spaced out as far as possible to quantity( Hauer and Bencke 1987). However, most pestreduce the cost of control ( Colbo and O' Brien 1984, black fly species need 200 to 350 degree- days for larvalDavis et al. 1992). The time interval between larvicidal development ( Davies et al. 1962, Ross and Merritttreatments depends on the rate of larval development, 1978, Brenner et al. 1981, Merritt et al. 1982, Shipp andwhich is strongly influenced by water temperature Procunier 1986, McCreadie and Colbo 1991).Merritt et al. 1982, Hauer and Benke 1987, Rempel and Consequently, black fly control programs throughout
Carter 1987, Ross and Merritt 1987, de Moor 1982b, the world have similar schedules, depending on their1994, McCreadie and Colbo 1991, Davis et al. 1992). In latitude. In arctic rivers, black fly larvae overwinter at

June, 1996 Journal of Vector Ecology 49
water temperatures close to freezing, and development tudinal changes in temperature were ofconcern because
proceeds as soon as temperatures increase with the warmer water in the lower reaches of the river was likely
spring melt( Anderson and Dicke 1960, Westwood and to increase larval development rates so that different
B rust 1981). Control ofblack flies in arctic and temperate reaches of the river would require treatments at different
rivers usually starts towards the end of winter and early times. The aim of this paper was to provide guidelines
spring and continues at intervals of7 to 14 days until mid for the appropriate timing of larvicide treatments for thesummer ( Westwood and Brust 1981, Chilcott et al. annual control of pest black flies along the middle1983, Petersen et al. 1983). In tropical areas there is no Orange River, South Africa.
definite starting time, and treatments are often requiredat weekly intervals throughout the year ( Elsen et al. METHODS
1981, Davis et al. 1992). In West Africa the developmenttimes of the pest Simulium damnosum s. l. are generally River Conditions
8 to 12 days in the wet season and 11 to 14 days in the Discharge data were supplied by the Department of
dry season( Crosskey 1990: 137). Not much is known Water Affairs and Forestry, Pretoria. Spot water
about black fly control in arid areas, but it has been noted temperatures were recorded at the time of application,
that the abundance of the pest black fly Simulium and maximum-minimum temperatures were measured
griseicollealongtheNileRiverwassignificantlyreduced weekly in a slow- flowing pool near Upington.
by hot, dry weather( de Meillon 1930). Along the Vaal Degree-day calculations were based on the product ofRiver, South Africa, adult black flies were inactive the number of days taken for the first pupae to appear
when air temperatures exceeded 33° C and the relative and the average ( maximum- minimum) water
humidity was less than 35%( Steenkamp 1972). temperature recorded over the same period( McCreadie
The middle and lower reaches of the Orange River, and Colbo 1991). Data on longitudinal changes in water
South Africa, have been plagued by pest outbreaks of temperature were not available because of the long
the mammalophilic black fly Simulium chutteri Lewis distances involved( TABLE 1). However, an indirect
Jordaan and van Ark 1990). The river runs through arid measure of downstream changes in water temperature
and semiarid conditions, and seasonal temperature was obtained from head-capsule lengths of final instar
fluctuations are extreme. Larval development rates were S. chutteri, which were used as " biological" thermo-
therefore expected to be seasonally variable. Further- meters. Head-capsule lengths were measured on the
more, high air temperatures and low humidities were postgena, between the mandibular phragma and the
likely to reduce adult survival during summer. post-occiput( Fredeen 1976).
Consequently, timing of larvicide applications was likely The abundance and species composition of
to be highly variable, and depend on both water planktonic algae was assessed weekly in 1993 and 1994.temperature and adult survival. Furthermore, longi- Water was sieved through a Whatman GF/C filter,
TABLE 1. Details of sampling sites in the Orange River, arranged in orderdownstream, and the average(± 1SD) head-capsule lengths of final
instar larvae of Simulium chutteri collected in summer( 15th- 18th
February 1992) and winter ( 13- 22nd July 1992). Sample size is
indicated in brackets.
Distance from
Lake v.d. Kloof Head- Capsule Lengths ( gm)
Site km) Summer Winter
Walaza 456 429± 18( 29)
Serfontein Bridge 91 360± 21 ( 40) 414± 24( 40)
Orania 45 421± 23 ( 12)
Prieska 379 349± 20( 44) 435± 21 ( 28)
Gifkloof 638 348± 22( 50) 437± 21 ( 40)
Onseeepkans 917 348± 21 ( 43) 401± 22( 40)
Vioolsdrift 1139 404± 18( 37)

50 Journal of Vector Ecology June, 1996
which was cut to size and placed on a glass slide daily. Fly abundance was quantified using a 4- pointcontaining a drop of cane- sugar syrup. Algal cells were ranked abundance scale ( 0= no flies; 1= present;counted in each of 10 fields at 300 times magnification. 2= common; 3= abundant). These data were averaged to
provide an overall index of black fly abundanceLarvae throughout the study area. Climatic data for the region
Larval and pupal abundance was assessed weekly were obtained from the Weather Bureau, Pretoria.
in rapids upstream of Upington ( 28° 27' S; 21° 15' E)between January 1992 and December 1994. RESULTS AND DISCUSSION
Assessments were based on the visual comparison oflarvae within a 4 x 4 cm quadrat of highest larval River Conditions
population density on 30 stones, to ten diagrammatically Water temperature during the trials ranged from 8prepared abundance classes, according to the method of to 29° C ( TABLE 2). Weekly temperatures usuallyPalmer 1994). Abundance classes for larvae ranged, on fluctuated by 2 to 3° C. The highest recorded weekly
a semilog10 scale, from no individuals ( Class 1) to an fluctuation occurred in the second week of April 1993,excess of 500,000 per m2( Class 10). when Boegoeberg Dam( 143 km upstream) was drained.
Larvicides were applied at various localities in the Temperatures dropped from 23 to 15° C. TemperaturesOrange River between July 1991 and June 1994. were reasonably consistent from year-to- year, althoughLarvicides used were either the protein toxins produced winter temperatures in 1993 were warmer and in 1994by the bacterium Bacillus thuringiensis var. israelensis colder than in 1992( Fig. 1). Estimates ofannual degree-Teknar® HP-D or Vectobac® 12AS) or the organo- days were 6,986 in 1992, 7,288 in 1993, and 8,008 in
phosphate temephos( Abate® 200EC). Larval develop- 1994.
ment was monitored on 34 occasions following both Head-capsule lengths of final instar larvae weresingle-site ( experimental) and multiple- site ( control) larger among larvae collected in cold water than inapplications. Single- site applications were administered warm water( Fig. 2). Larvae collected from Serfonteinfrom bridges or a canoe, whereas, multiple-site Bridge ( upstream of Lake van der Kloof) in summerapplications were applied by helicopter. To reduce the ( 15. 02.92) were significantly larger than those collectedlikelihood of colonization from upstream, sites were downstream of Lake van der Kloof (Student' s t-test;chosen at rapids which had long stretches of pool t=31, P< 0. 03; TABLE 1). There was no significantupstream. Despite this precaution, mixed populations difference between head-capsule lengths of larvaewere often found within a few days of application. One collected from three sites downstream of Lake van derreason for this was that larvicide applications were not Kloof( P> 0. 8), indicating similar water temperatures atalways 100% effective. However, although the post- these sites. However, larvae collected from the lowerapplication populations were often mixed, there was reaches at Onseepkans and Vioolsdrift in winter wereusually a cohort of similar-sized individuals, which significantly smaller than larvae collected near Upington.marked the timing of application. Sites were visited Likewise, larvae collected from Orania were slightlyevery one to three days following application, and smaller than those collected from Prieska and Upingtondevelopment time" was defined as the time required ( TABLE 1). Larval development in the lower reaches
for the first cohort of larvae to pupate following larvicide of the Orange River was therefore faster than upstream,application. The " recommended treatment interval" and more applications would be required to adequatelywas defined as the number of days taken for the first treat the lower reaches than the middle reaches.cohort to reach 5- 6th instar. The rapids upstream of In both 1993 and 1994 there was a bloom of theUpington were twice treated for black fly control in blue- green algae Microcystis sp. in autumn ( Fig. 3).1991, and once in 1992. Thereafter, these rapids were High densities of these algae ( up to 48,000 cells/ml)left untreated as a control area. develop during thermal mixing in Lake van der Kloof,
situated 656 km upstream ofUpington. Thermal mixingAdults in the reservoir takes place towards the end of summer
Estimates of the abundance of adult black flies ( March/April) each year( Allanson and Jackson 1983).along the Orange River started in August 1984.Abundance was assessed by interested farmers in the Larvae and Pupae
vicinity of livestock between Boegoeberg Dam and Natural fluctuations in larval abundance rangedAugrabies Falls, a river distance of 280 km. Initially from zero, in March 1993, to an excess of 500,000/m2,
1984- 1987), the assessments were made monthly. Later in July 1992( Fig. 4). The abundance of pupae showedNovember 1989 to 1995), assessments were made similar trends to the larvae, although there was a
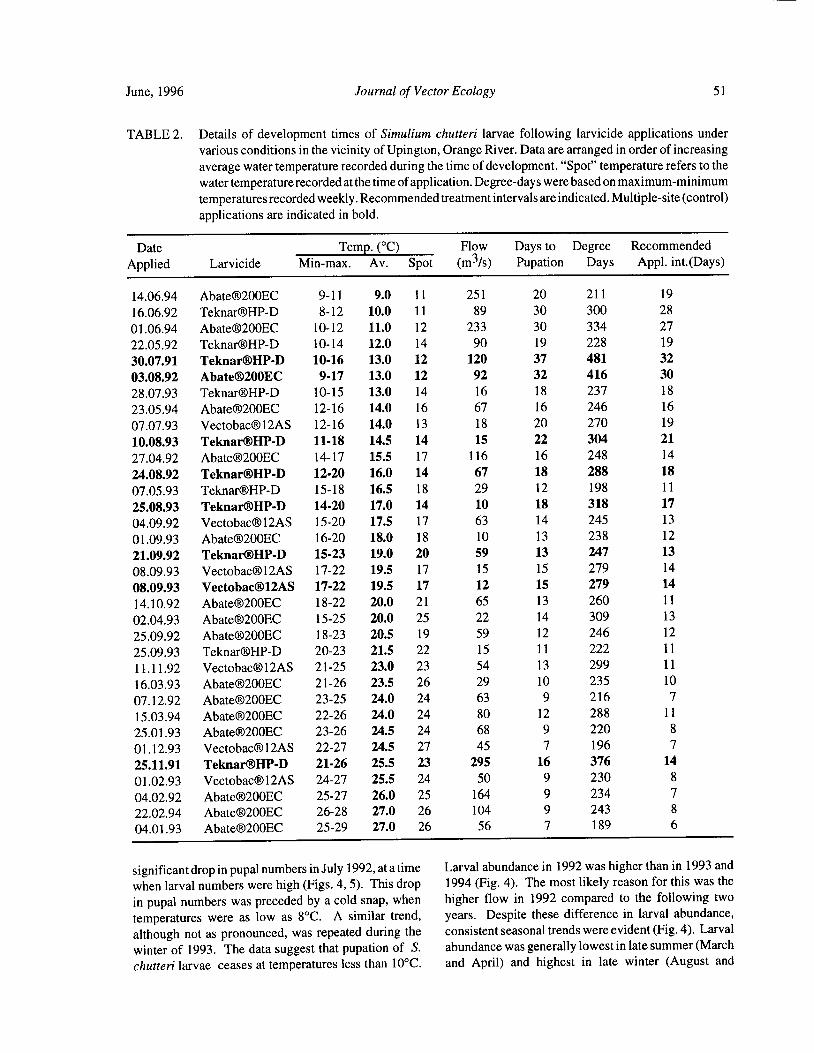
June, 1996 Journal of Vector Ecology 51
TABLE 2. Details of development times of Simulium chutteri larvae following larvicide applications undervarious conditions in the vicinity of Upington, Orange River. Data are arranged in order of increasingaverage water temperature recorded during the time of development." Spot" temperature refers to thewater temperature recorded at the time ofapplication. Degree- days were based on maximum-minimum
temperatures recorded weekly. Recommended treatment intervals are indicated. Multiple-site( control)applications are indicated in bold.
Date Temp. (° C) Flow Days to Degree Recommended
Applied Larvicide Min-max. Av. Spot ( m3/
s) Pupation Days Appl. int.(Days)
14.06.94 Abate® 200EC 9- 11 9.0 11 251 20 211 19
16.06.92 Teknar® HP-D 8- 12 10.0 11 89 30 300 28
01. 06.94 Abate® 200EC 10- 12 11.0 12 233 30 334 27
22.05. 92 Teknar® HP-D 10- 14 12.0 14 90 19 228 19
30.07.91 Teknar®HP-D 10- 16 13.0 12 120 37 481 32
03.08.92 Abate® 200EC 9- 17 13.0 12 92 32 416 30
28.07. 93 Teknar® HP-D 10- 15 13.0 14 16 18 237 18
23. 05. 94 Abate® 200EC 12- 16 14.0 16 67 16 246 16
07.07.93 Vectobac® 12AS 12- 16 14.0 13 18 20 270 19
10. 08.93 Teknar®HP-D 11- 18 14.5 14 15 22 304 21
27. 04.92 Abate® 200EC 14- 17 15.5 17 116 16 248 14
24.08.92 Teknar®HP-D 12- 20 16.0 14 67 18 288 18
07. 05.93 Teknar® HP-D 15- 18 16.5 18 29 12 198 11
25.08.93 Teknar®HP-D 14- 20 17.0 14 10 18 318 17
04.09.92 Vectobac® 12AS 15- 20 17.5 17 63 14 245 13
01. 09.93 Abate® 200EC 16- 20 18.0 18 10 13 238 12
21.09.92 Teknar®HP-D 15- 23 19.0 20 59 13 247 13
08.09.93 Vectobac® 12AS 17- 22 19.5 17 15 15 279 14
08.09.93 Vectobac® 12AS 17- 22 19.5 17 12 15 279 14
14. 10.92 Abate® 200EC 18- 22 20.0 21 65 13 260 11
02.04.93 Abate® 200EC 15- 25 20.0 25 22 14 309 13
25. 09.92 Abate® 200EC 18- 23 20.5 19 59 12 246 12
25. 09.93 Teknar® HP-D 20-23 21. 5 22 15 11 222 11
11. 11. 92 Vectobac® 12A5 21- 25 23.0 23 54 13 299 11
16.03.93 Abate® 200EC 21- 26 23.5 26 29 10 235 10
07. 12.92 Abate® 200EC 23- 25 24.0 24 63 9 216 7
15. 03.94 Abate® 200EC 22- 26 24.0 24 80 12 288 11
25.01. 93 Abate® 200EC 23- 26 24. 5 24 68 9 220 8
01. 12.93 Vectobac® 12AS 22-27 24. 5 27 45 7 196 7
25. 11. 91 Teknar®HP-D 21- 26 25. 5 23 295 16 376 14
01. 02.93 Vectobac® 12AS 24-27 25.5 24 50 9 230 8
04.02.92 Abate® 200EC 25- 27 26.0 25 164 9 234 7
22. 02.94 Abate® 200EC 26- 28 27.0 26 104 9 243 8
04.01. 93 Abate® 200EC 25- 29 27.0 26 56 7 189 6
significant drop in pupal numbers in July 1992, at a time Larval abundance in 1992 was higher than in 1993 and
when larval numbers were high( Figs. 4, 5). This drop 1994( Fig. 4). The most likely reason for this was the
in pupal numbers was preceded by a cold snap, when higher flow in 1992 compared to the following two
temperatures were as low as 8° C. A similar trend, years. Despite these difference in larval abundance,
although not as pronounced, was repeated during the consistent seasonal trends were evident( Fig. 4). Larval
winter of 1993. The data suggest that pupation of S. abundance was generally lowest in late summer( Marchchutteri larvae ceases at temperatures less than 10°C. and April) and highest in late winter ( August and
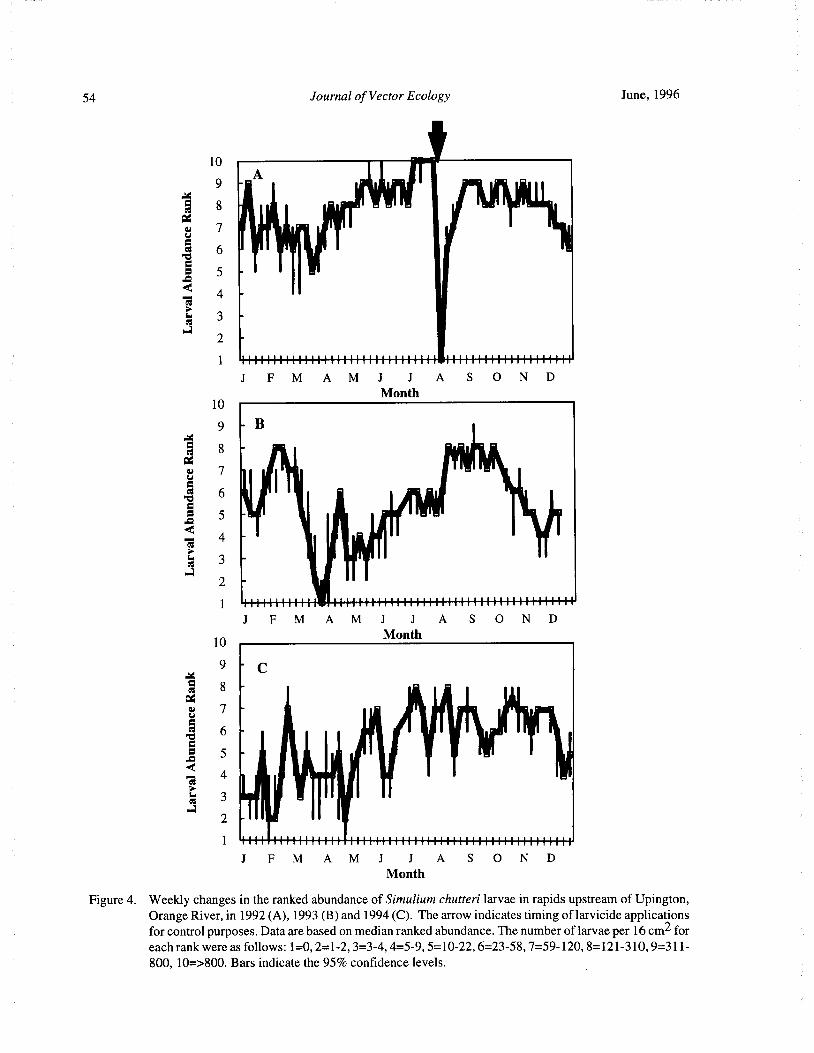
54 Journal of Vector Ecology June, 1996
10
9A
g 8A4a.) 7
b 6 WSWa 5
4
et3 -
2 -
1 1111111111111111111111111111111 11111111111111111111
J F M A M J J ASO N D
Month10
9 - B
8 -
v_r%7
IeQe 6b
5
d4
3 -1. 4
2 -
1 111111111111 11111111111111I111111111111111111111II
J FM A M J J A S O N D
10Month
9 - C
g 8 -
u 7 -
6 -
5 -
4
es 3 - a
2 -
1 1111 IIII1111111 1111111111111111111111111111II11111
J FM A M J J A S O N D
Month
Figure 4. Weekly changes in the ranked abundance of Simulium chutteri larvae in rapids upstream of Upington,Orange River, in 1992( A), 1993( B) and 1994( C). The arrow indicates timing of larvicide applicationsfor control purposes. Data are based on median ranked abundance. The number of larvae per 16 cm2 for
each rank were as follows: 1= 0, 2= 1- 2, 3= 3- 4, 4= 5- 9, 5= 10- 22, 6= 23- 58, 7= 59- 120, 8= 121- 310, 9= 311-
800, 10=> 800. Bars indicate the 95% confidence levels.
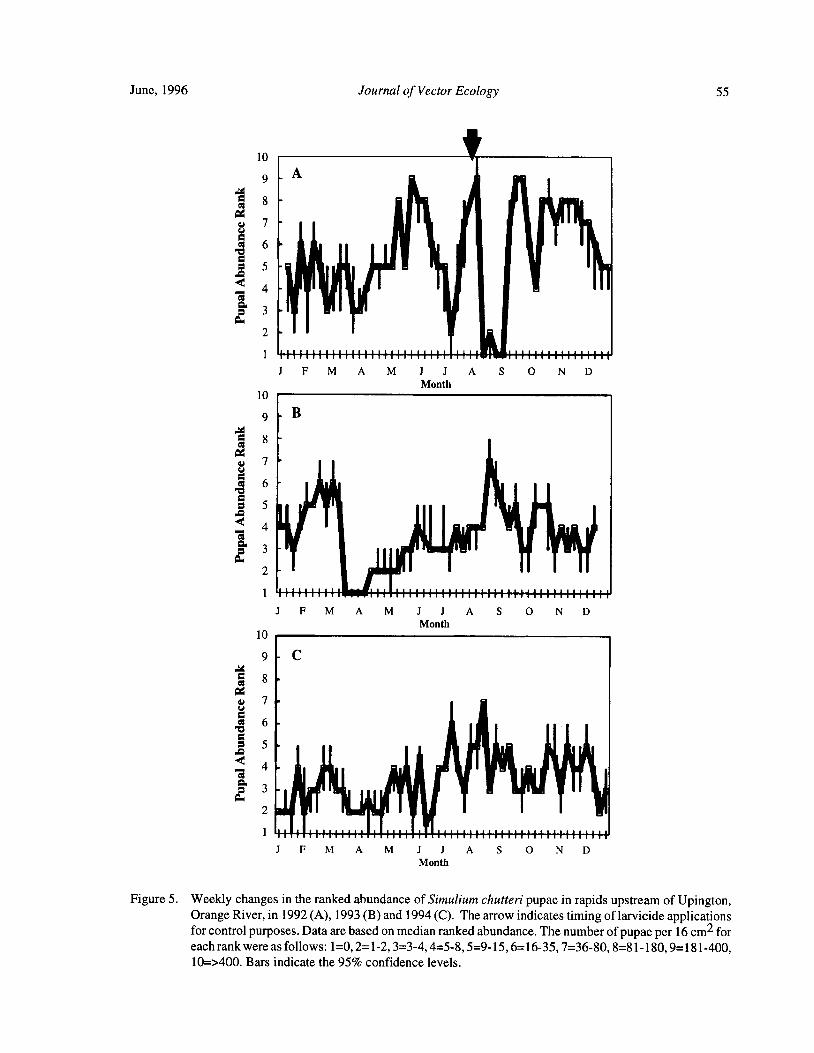
June, 1996 Journal of Vector Ecology 55
10
9 - A
8
a7 -
0 5 -
4 -
4 3
2 -
1 Iiliil111111111111111i1111 iiii II1 H t11111IIIII
J F M A M J J A S 0 ND
Month
10
9 B
8 -
a7 r
E
4 6 -
0 5
d4
E 3
2 -
1 1114111111 111 11III11IIII111I111I11III11III1I11
J FM A M J J A S 0 ND
Month
10
9 - C
8
a>• 7
0 6
0 5 •
4
3
2
1 Illilllll l, l ll11111111 M IIIIIIJ F M A M J J A S O ND
Month
Figure 5. Weekly changes in the ranked abundance of Simulium chutteri pupae in rapids upstream of Upington,Orange River, in 1992( A), 1993( B) and 1994( C). The arrow indicates timing of larvicide applicationsfor control purposes. Data are based on median ranked abundance. The number of pupae per 16 cm2 foreach rank were as follows: 1= 0, 2= 1- 2, 3= 3- 4, 4= 5- 8, 5= 9- 15, 6= 16- 35, 7= 36- 80, 8= 81- 180, 9= 181- 400,10=> 400. Bars indicate the 95% confidence levels.
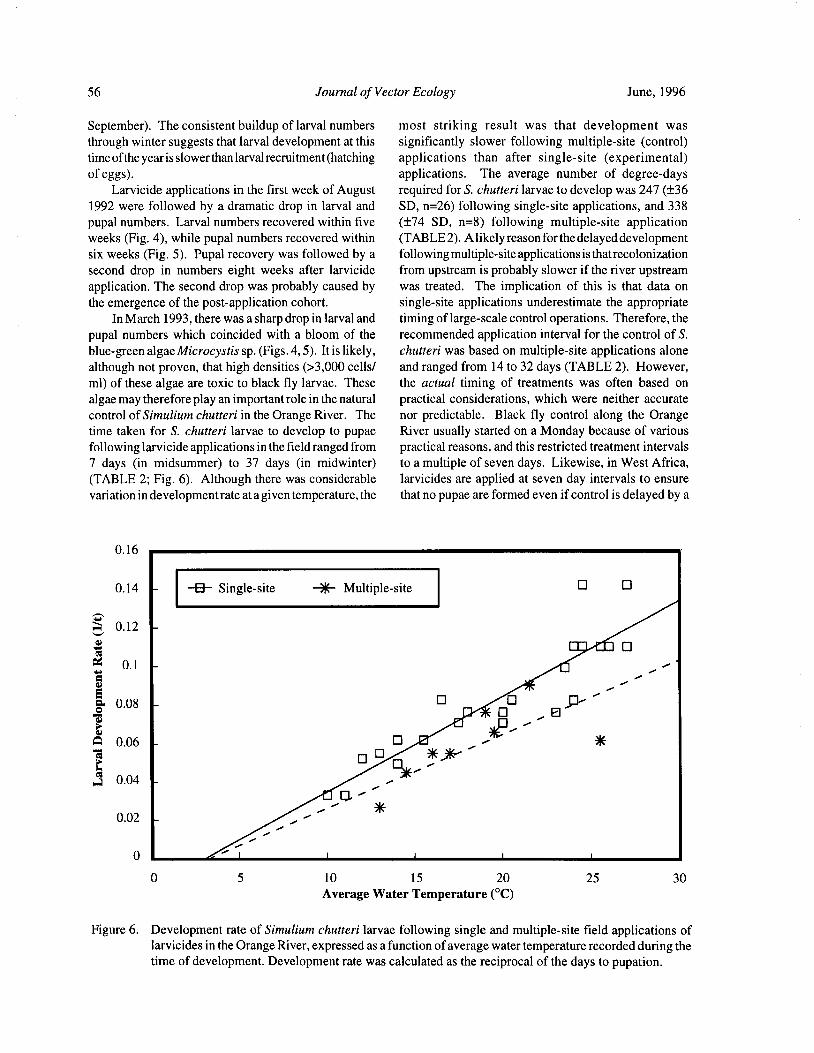
56 Journal of Vector Ecology June, 1996
September). The consistent buildup of larval numbers most striking result was that development wasthrough winter suggests that larval development at this significantly slower following multiple-site ( control)time ofthe year is slower than larval recruitment( hatching applications than after single- site ( experimental)
of eggs). applications. The average number of degree-days
Larvicide applications in the first week of August required for S. chutteri larvae to develop was 247(± 36
1992 were followed by a dramatic drop in larval and SD, n= 26) following single- site applications, and 338pupal numbers. Larval numbers recovered within five (± 74 SD, n= 8) following multiple-site applicationweeks( Fig. 4), while pupal numbers recovered within ( TABLE 2). A likely reason for the delayed developmentsix weeks ( Fig. 5). Pupal recovery was followed by a following multiple-site applications is thatrecolonizationsecond drop in numbers eight weeks after larvicide from upstream is probably slower if the river upstreamapplication. The second drop was probably caused by was treated. The implication of this is that data on
the emergence of the post-application cohort. single- site applications underestimate the appropriate
In March 1993, there was a sharp drop in larval and timing of large- scale control operations. Therefore, thepupal numbers which coincided with a bloom of the recommended application interval for the control of S.
blue-green algae Microcystis sp.( Figs. 4, 5). It is likely, chutteri was based on multiple-site applications alone
although not proven, that high densities(> 3, 000 cells/ and ranged from 14 to 32 days( TABLE 2). However,
ml) of these algae are toxic to black fly larvae. These the actual timing of treatments was often based onalgae may therefore play an important role in the natural practical considerations, which were neither accurate
control of Simulium chutteri in the Orange River. The nor predictable. Black fly control along the Orangetime taken for S. chutteri larvae to develop to pupae River usually started on a Monday because of variousfollowing larvicide applications in the field ranged from practical reasons, and this restricted treatment intervals
7 days ( in midsummer) to 37 days ( in midwinter) to a multiple of seven days. Likewise, in West Africa,
TABLE 2; Fig. 6). Although there was considerable larvicides are applied at seven day intervals to ensurevariation in development rate at a given temperature, the that no pupae are formed even if control is delayed by a
0. 16
0. 14 - - B- Single- site Multiple-site
0. 12 -
a 0. 1 - a
0 0.08 - off
A 0.06 -
0.04 -
Q
0.02 _
0 5 10 15 20 25 30
Average Water Temperature(° C)
Figure 6. Development rate of Simulium chutteri larvae following single and multiple- site field applications oflarvicides in the Orange River, expressed as a function ofaverage water temperature recorded during thetime of development. Development rate was calculated as the reciprocal of the days to pupation.

June, 1996 Journal of Vector Ecology 57
few days because of technical or logistical problems 1978, Shipp and Procunier 1986, McCreadie and ColboKurtak pers. comm.). 1991). Therefore, degree- days provides a useful first
approximation for estimating appropriate treatmentAdults intervals for black fly control. However, degree-days
Assessment of seasonal changes in the abundance should be used with caution because more degree-daysof adult S. chutteri along the Orange River before black are required for development at colder temperaturesfly control was initiated in 1991 shows a consistent peak ( McCreadie and Colbo 1991).in spring( August-October)( Fig. 7). The worst year on Considering the time required for the developmentrecord for black fly annoyance was in 1990, when there ofS. chutteri eggs( Begemann 1986), larvae( this study),were annoyance peaks in both spring and autumn( April pupae( de Moor 1982a), and the survival ofadults in theand May) ( Fig. 7). A possible reason for the autumn field ( this study), it is estimated that S. chutteri in theoutbreak in 1990 is that floods in the preceding two middle Orange River undergo 11 to 13 generations peryears had flushed the reservoirs, and consequently, year. However, adequate control during normal( non-there may not have been a Microcystis sp. bloom in flood) years may be obtained with nine applications( six1990. spring" and three" autumn"). Although it is tempting
In most years adult populations dropped to use longer rather than shorter treatment intervals, adramatically towards the end of October ( Fig. 7). This single adult female S. chutteri lays an estimated 500 tois when average A-pan evaporation rates start to exceed 1, 000 eggs in a lifetime( unpublished data), and the extra12 mm/day. In 1992, there was a localized outbreak of days thus gained do not compensate for the reducedblack flies in the Upington area. The river was treated efficacy of control because ofemerged( escaped) adults.the first week in August, and the flies disappeared after It is therefore recommended that shorter rather than15 days. Air temperatures during this period ranged longer treatment intervals are used even though this mayfrom- 3. 5 to 29.6°C, and evaporation ranged from 2 to necessitate extra treatment.
6 mm/day. Adult black flies were often found in veryhigh densities in shady areas near the river on hot days, Acknowledgments
indicating that adult black flies actively seek favorablemicroenvironments where evaporation rates are low. We would like to thank Mr. Morgan Scheepers forThe absence of a summer outbreak therefore appears to assistance in the field and laboratory, Dr. Henk Van Arkbe related to high air temperatures (> 38° C) and for comments on an earlier draft of this paper, Dr.evaporation(> 12 mm/ day), both of which are likely to Leonie Jordaan for data collected on adult abundancecause rapid adult mortality. Black fly control operations between 1984 and 1987, and the many farmers whoalong the Orange River are therefore unlikely to be provided data on adult abundance. The project wasrequired between the months ofNovember and February. funded by the Water Research Commission.
CONCLUSIONS REFERENCES CITED
Larvicidal applications to prevent spring outbreaks Allanson, B. R and P. B. N. Jackson. 1983. Limnologyof the pest black fly Simulium chutteri along the middle and fisheries potential of Lake le Roux. SouthOrange River should start towards the end of July when African National Scientific Programmes Reportmost of the population is in the larval stage. Subsequent No. 77, 182 pp.treatments should be made at 14 to 32 day intervals, Anderson, J. R. and R. J. Dicke. 1960. Ecology of thedepending on water temperature. Treatments should immature stages of some Wisconsin black fliescontinue until late October, after which outbreaks are Simuliidae: Diptera). Ann. Entomol. Soc. Am. 53:unlikely because of the high rate of evaporation (> 12 386-404.mm/ day). High rates of evaporation continue until the Begemann, G. J. 1986. Die bionomie van sekere suid-end of February, after which a second population afrikaanse Simuliidae species ( Diptera). M.Sc.outbreak may be expected. However, blooms of blue- thesis, University of Pretoria, Pretoria. 127 pp.green algae following thermal overturn in Lake van der Brenner, R. J., E. W. Cupp, and M. J. Bernado. 1981.Kloof in late summer may reduce larval populations, Growth and development of geographic andmaking it unnecessary to apply larvicides in autumn. crossbred strains of colonized Simulium decorumThe number ofdegree-days required for the development Diptera: Simuliidae). Can. J. Zool. 59: 2072- 2079.of S. chutteri is similar to that reported for many other Chilcott, C. N., J. S. Pillai, and J. Kalmakof. 1983.black fly species( Davies et al. 1962, Ross and Merritt Efficacy of Bacillus thuringiensis var. israelensis
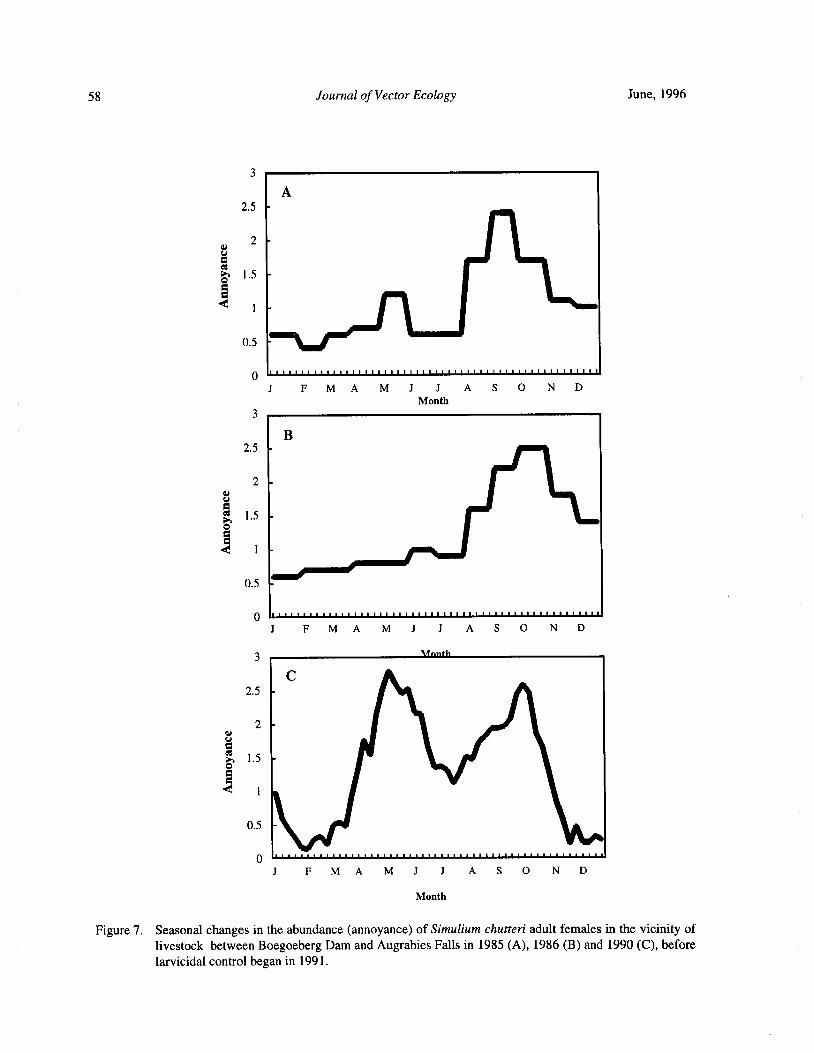
58 Journal of Vector Ecology June, 1996
3
A2.5
u
11Lorefteli
2 -
c 1. 5d 1 -
0.5 -
0J F M A M J J A S O ND
Month
3
2. 5 -
2 -u
1. 5 -0
d 1 -
0.5 -
0J F M A M J J A S O ND
3Mnnth
2.5 -
2 -u
1. 5 -0
1
0.5 -
0J F M A M J J A SO ND
Month
Figure 7. Seasonal changes in the abundance ( annoyance) of Simulium chutteri adult females in the vicinity oflivestock between Boegoeberg Dam and Augrabies Falls in 1985( A), 1986( B) and 1990( C), beforelarvicidal control began in 1991.

June, 1996 Journal of Vector Ecology 59
as a biocontrol agent against larvae of Simuliidae annoyance of livestock by Simulium chutteri LewisDiptera) in New Zealand. New Zealand J. Zool. along the Orange River, South Africa ( Diptera:
10: 319- 326. Simuliidae). Onderstepoort J. vet. Res. 57: 189- 195Colbo, M. H. and H. O' Brien. 1984. A pilot black fly McCreadie, J. W. and M. H. Colbo. 1991. The influence
Diptera: Simuliidae) control program using of temperature on the survival, development,Bacillus thuringiensis var. israelensis in growth, and chromosomes preparation quality ofNewfoundland. Can. Ent. 116: 1085- 1096. the EFG/C, ACD, and AA cytotypes ofthe Simulium
Colbo, M. H. and G. N. Porter. 1979. Effects of the food venustum- verecundum complex ( Diptera:supply on the life history of Simuliidae( Diptera). Simuliidae). Can. J. Zool. 69: 1356- 1365Can. J. Zool. 57: 301- 306 Merritt, R. W., D. H. Ross, and G. J. Larson. 1982.
Crosskey, R. W. 1990. The natural history of blackflies. Influence of stream temperature and seston on theChichester, England, John Wiley& Sons, ix+ 711 growth and production ofoverwintering larval blackpp. flies( Diptera: Simuliidae). Ecology 63: 1322- 1331
Davies, D. M., B. V. Peterson, and D. M. Wood. 1962. Palmer, R. W. 1994. A rapid method of estimating theThe black flies ( Diptera; Simuliidae) of Ontario, abundance of immature blackflies ( Diptera:Part 1. Adult identification and distribution with Simuliidae). Onderstepoort J. Vet. Res. 61: 117-descriptions ofsix new species. Proc. Ent. Soc. Ont. 126.
92: 70- 154. Petersen, J. L., A. J. Adames, and L. de Leon. 1983.Davis, J. R., V. K. Bariero, and M. Trpis. 1992. Duration Bionomics and control of black flies ( Diptera:
of larval development ofSimuliumyahense( Diptera: Simuliidae) at the Fortuna Hydroelectric Project,Simuliidae) under natural conditions. J. Med. Panama. J. Med. Entomol. 20(4): 399- 408.Entomol. 29: 108- 110. Rempel, R. S. and J. C. H. Carter. 1987. Temperature
de Meillon, B. 1930. On the Ethiopean Simuliidae. Bull. influences on adult size, development, andEnt. Res. 21: 185- 200 reproductive potential of aquatic diptera. Can. J.
de Moor, F. C. 1982a. A community of Simulium Fish. Aquat. Sci. 44: 1743- 1752.species in the Vaal River near Warrenton. Ph.D. Ross, D. H. and R. W. Merritt. 1978. The larval instarsthesis, University of the Witwatersrand, and population dynamics of five species of black
Johannesburg, 317 pp. flies ( Diptera: Simuliidae) and their responses tode Moor, F. C. 1982b. Determination of the number of selected environmental factors. Can. J. Zool. 56:
instars and size variation in the larvae and pupae of 1633- 1642
Simulium chutteri Lewis ( 1965)( Diptera: Ross, D. H. and R. W. Merritt. 1987. Factors affectingSimuliidae) and some possible bionomical larval black fly distributions and populationimplications. Can. J. Zool. 60: 1374- 1382. dynamics. Pp. 90- 108 in Black flies: ecology,
de Moor, F. C. 1994. Aspects of the life history of population management, and annotated world list,Simulium chutteri andS. bovis( Diptera; Simuliidae) The Pennsylvania State University( Kim, K. C. andin relation to changing environmental conditions in R. W. Merritt, eds.), 722 pp.SouthAfricanrivers. Verb. Internat. Verein. Limnol. Shipp, J. L. and W. S. Procunier. 1986. Seasonal25: 1817- 1821 occurrence of,development of,and the influence of
Elsen, P., C. Bellec, and G. Hebrard. 1981. Speed of selected environmental factors on the larvae ofrecolonization of an Ivory Coast breeding site by Prosimulium and Simulium species of blackfliesSimuliumdamnosums. l.( Diptera: Simuliidae) after Diptera: Simuliidae) found in the rivers ofexperimental suspending of larvicide treatments southwestern Alberta. Can. J. Zool. 64: 1491- 1499and its consequence on strategy of fight against this Steenkamp, J. A. 1972. ` n Ondersoek na dievector of Onchocerciasis. ( In French). Cah. wisselwerking tussen sommige ekologiese faktoreORSTOM Ser. Entomol. Med. Parasit. 19: 5- 9. en die bevolkings van Simulium damnosum
Fredeen, F. J. H. 1976. The seven larval instars of Theobald en S. nigritarsis Coquillett( Simuliidae:Simuliumarcticum( Diptera; Simuliidae). Can. Ent. Diptera) in die Vaalrivier by Parys. D.Sc. thesis,108: 591- 600. Potchefstroom University, Potchefstroom, South
Hauer, F. R. and A. C. Benke. 1987. Influence of Africa, 156 pp.temperature and river hydrograph on black fly Westwood, A. R. and R. A. Brust. 1981. Ecology ofgrowth rates in a subtropical blackwater river. J. N. black flies( Diptera: Simuliidae) of the Souris River,Am. Benthol. Soc. 6: 251- 162. Manitoba as a basis for control strategy. Can.
Jordaan, L. C. and H. van Ark. 1990. A survey of Entomol. 113: 223- 234

Journal of Vector Ecology 21( 1): 60-65
Larvicidal Oils Modify the OvipositionBehavior of Culex Mosquitoes
J. W. Beehlerl and M. S. Mulla
Department of Entomology, University of California, Riverside, CA 92521 USACurrent Address: Jenkins, Sanders and Associates,
4320 Atlantic Avenue, Suite 216, Long Beach, CA 90807 USA
Received 18 April 1995; Accepted 27 August 1995
ABSTRACT: Larvicidal oils were shown in laboratory and field studies to be repellent to ovipositingCulex quinquefasciatus, Culex tarsalis and Culex stigmatosoma. In laboratory two-choice ovipositionassays, open cups in which water was treated with Golden Bear 1356 and 1111, Rodspray MFSTS, andOrchex parafinic oil received fewer Culex quinquefasciatus egg rafts than did untreated cups. Cups inwhich water was treated with Golden Bear 1111 and Rodspray MFSTS also received reduced Culextarsalis oviposition when compared to untreated cups. Cups treated with Golden Bear 1111 or Rodspray
MFSTS received fewer egg rafts than untreated cups in Culex quinquefasciatus paired oviposition testsfor three to seven days post- treatment. In two field studies, Golden Bear 1111 and Rodspray MFSTSreduced egg deposition by Culex stigmatosoma and Culex quinquefasciatus for at least seven days post-treatment at operational rates.
Keyword Index: Oviposition attractants, mosquitoes, Culex, larvicidal oils.
INTRODUCTION studies( Mather and DeFoliart 1983). Our objective was
to determine if commercially available petroleumPetroleum hydrocarbon larvicidal oils have been hydrocarbon larvicidal oils had any repellent/deterrent
used to effectively control mosquito larvae for nearly effect on the ovipositional success of Culex mosquitoes.
100 years. In California, petroleum oils are the onlylabeled materials effective against pupae. Moreover, no MATERIALS AND METHODS
emergence of resistance in larvae to petroleum oils has
been reported to date. Due to these advantages, oils Laboratory Studieshave been widely used for mosquito control in a number The repellency of several larvicidal oils to Culexof habitats. quinquefasciatus and Culex tarsalis was first assessed
On the other hand, the use of oils has been recently using choice tests in small cages. The first species haslimited by adverse effects on some nontarget macro- been under colonization for almost 25 years while the
invertebrates. However, Mulla et al.( 1979) concluded second species has been colonized for about five years.
that petroleum hydrocarbons used in mosquito control Both colonies have been infused from time to time with
have little effect on benthic fauna. Detrimental effects, larvae from the field. Two waxed paper cups( 5 x 7.5 cm
however, were shown to be more pronounced on those diam) were placed in the rear corners of a small cage( 23
organisms which surface to obtain oxygen. These x 23 x 32 cm). One cup contained 80 ml of distilledgroups usually recover one to three weeks post-treatment. water treated with the larvicidal oil to be tested for
Larvicidal oils are widely used, but like many other repellency and the other cup contained distilled waterinsecticidal agents little is known about their effects on only. Gravid females ( 25 Cx. quinquefasciatus or 20the ovipositing fraction of the mosquito population. Culex tarsalis) were placed in the cage at 16: 00 h and
The emulsifiers and petroleum solvents in several left overnight. Cages were kept in a photoperiod( 14: 10
organophosphate insecticides were shown to be highly L:D including a 2 h dusk/dawn period) and temperaturerepellent to ovipositing Aedes triseriatus( Say) in field ( 25± 3° C) controlled insectary. The following morning

June, 1996 Journal of Vector Ecology 61
8: 00 h) the numbers of egg rafts deposited in the treated on Culex oviposition. On June 13, 1994, 12 wood-sided
and control cups were counted. Egg raft numbers were ponds ( 27 m2) were flooded to a mean depth of 30
transformed to the square root(' Vx) and a paired t-test cm with canal water. Each pond was supplemented with
analysis was used to compare treatments with controls. 4 kg of rabbit pellets( Brookhurst Mill, Riverside, CA).Each treatment was tested in six cages for two nights Two days post- flooding before any oviposition, pondsfor a total of 12 replications. Golden Bear 1111 and were treated with Rodspray MFSTS and Golden Bear1356 ( Witco Corp., Oildale, CA) Rodspray MFSTS 1111 ( 12. 5 liters/ ha) or left untreated. Egg rafts wereRod Products Co., Marina del Rey, CA), and a para- counted along a transect ( 5. 5 x 14 m) in each of the
finic oil mixture were tested for their repellency to ponds ( Beehler and Mulla 1993) on day 1 ( beforeCx. quinquefasciatus at 1, 2.5, 5, and 10 p1/ cup. The treatment), 3, 5, 7, and 9 days post- treatment. Subsamples
parafinic oil stock solution consisted of 40 ml Orchex of Culex egg rafts were taken to the laboratory, rearedExxon Co., Houston, TX), 10 ml acetone as cosolvent, individually to the 4th- instar, and identified.
and 0.25 ml Pluronic 121 ( Wyandotte Chemical Co., The second field experiment was designed to
Wyandotte, MI). These rates approximate those used simulate an operational situation where oil applications
operationally in mosquito control. Golden Bear 1111 were made when late instar larvae were present. Twelve
and Rodspray MFSTS were tested for repellency to Cx. ponds were flooded on August 8, 1994 with canal water
tarsalis at rates ranging from 1 to 5 pl/ cup. and supplemented with rabbit pellets( 4 kg/pond). ThreeThe effects of aging of oil on water surface were days post- flooding the ponds were all treated with
considered in tests using Cx. quinquefasciatus. Distilled bifenthrin( 0.5 g/ ha AI) to control tadpole shrimp( Triopswater in test cups was treated with the larvicidal oil longicaudatus ( Notostraca: Triopsidae) which are
Golden Bear 1111 or Rodspray MFSTS) and held with predaceous on mosquito larvae. Bifenthrin at the rate
the untreated cups in the insectary until use in a two- used has no effect on mosquito larvae when used before
choice test. Both Golden Bear and Rodspray MFSTS significant oviposition by mosquitoes occurred( Mullawere subjected to oviposition tests every two days post- et al. 1992). Culex rafts were sampled along a transecttreatment until repellency was no longer evident. as described above 3, 6, 8, 10, 12, 14, 16, and 18 days
Another type of experimental design was used to post- flooding. Subsamples of the rafts were returned tomore directly measure repellency. In these experiments the laboratory, reared individually to the 4th-instar, and50 Cx. quinquefasciatus were placed in each of four identified to species.
screen cages( 45 cm on a side). In each cage two glass Larval samples were taken using a 350 ml dipperjars ( 16. 5 x 7. 6 cm diam) holding 100 ml of distilled from the corners of each pond until approximately 50water were placed in the rear corners. The water in one percent of the ponds contained 3rd- instar Culex larvae.
jar was treated with larvicidal oil and the other was left Four ponds were then treated with Golden Bear 1111 at
untreated as a control. A funnel- shaped plastic cup( No. 12. 5 liters/ ha, four ponds treated at 6. 3 liters/ ha, and
806A, Solo Cup Company, Urbana, IL) was inserted four left untreated as controls using a completelyinto the top of each jar. A hole had been placed in the randomized block design. The number of egg raftsbottom ofeach cup using a# 9 cork borer( 1. 5 cm). The counted along the transects were transformed to the
resulting experimental set- up consisted of a jar with a square root( I)for analysis( Box et al. 1978). Egg raftfunnel in the top with a 9 cm airspace between the numbers from treated ponds were compared with the
bottom of the funnel and the water' s surface. Mosquitoes untreated ponds using quadratic multiple regression
entering the cone to approach the water were retained in with dummy variables to account for the time periodthe jar. Golden Bear 1111 and Rodspray MFSTS were before treatment( Chatterjee and Price 1977).
tested at rates ranging from 2.5 to 10 ill/jar. Cages wereheld overnight at 21± 2° C and were exposed to natural RESULTS AND CONCLUSIONS
twilight. The data were analyzed as above, but in this
case the number of gravid females, rather than the egg Laboratory Studiesrafts, collected in each container was considered. Each All larvicidal oils tested ( Golden Bear 1111 and
experiment was replicated over two nights for a total of 1356, Rodspray MFSTS, and the Orchex parafinic oileight replicates per rate. formulation) showed high levels of repellency to Cx.
quinquefasciatus in two- choice comparisons when
Field Experiments applied at rates ranging from 1. 0 p1 to 10. 0 µ 1/ cupTwo field experiments were conducted at the ( TABLE 1). These rates corresponding to 2. 5 to 25.0
Aquatic and Vector Control Research Facility in liters/ ha are well within the range of those used in
Riverside, CA to determine the effect of larvicidal oil mosquito control operations suggesting that these oils

62 Journal of Vector Ecology June, 1996
may also have significant repellency under field con-ditions. Similar repellency was also noted with Cx.
of) tarsalis using Golden Bear 1111 and Rodspray MFSTSo a. at rates ranging from 1. 0 to 5 µ l/ cup( TABLE 1).k I + I + I + I v Petroleum hydrocarbon larvicides contain
G, U 4 c 4 surfactants which aid in spreading of the oil on theV surface of the water. These surfactants greatly reduce
CM M 0 0 the surface tension of the water, which in the field results
a I ++° I + ++° I 0 in the sinking of Culex egg rafts present before treatmentE M 00 M o M.S. Mulla, unpublisheddata). Subsequent oviposition
E0 0 0 o
Y o cannot occur as gravid females are not able to rest on the
30 3 surface film to oviposit. Gravid females are unable to
4..'-' o detect the surfactant alone( Isoe et al. 1995) and quicklyo
drown if they land on the oviposition water. In thesev, o" v? canA o
C. -
d — ° ,-• studies, we did not note significantly greater numbers ofb1pI + I + I + I + I + I
0 0 omen ,--, s 00 0 gravid females drowning in the oil treated cups compareda U 3 to the control cups. This indicates that reduced
p oviposition was due to deterrency and/ or repellencyo a
Ti" o"
4? 't
cc` N o ,,; rather than the drowning of gravid females in the oil
E0, I + I + I °
o treated containers.
E cu No o - o 0 o b y Additional laboratory studies determined that at
F" operational rates the duration of repellency lasted atact
0 yleast three days post- treatment. Golden Bear 1111 was
o3 repellent for seven days post- treatment at the 12.5 liters/
or 0) °• O0• .
t1v1 C.) A. ha rate and repellent for nine days at 25 liters/ ha( Fig.
a 0 I + I + I + I + I + I oNU C m N — oo . s r
1). Rodspray MFSTS was repellent for three days post-
j i . treatment at the lower rate and 7 days post- treatment at•
3 v the higher rate( Fig. 2).M co v U a= The last series of laboratory experiments provided
oo N• tn
I + I +° IC +°
I o a more direct measure of ovipositional repellency.v .. U
oo — oo ry Cr a .ti Gravid females had to enter an inverted cone and wereC.) ' d o ri o
a °' not able to contact the surface of thewater until trappedo o° r
in the glass container. Both Golden Bear 1111 and0) 0•
o ,. V Rodspray MFSTS showed activity similar to that in the
0o\ N M M
Si3cu
egg raft counting experiments using open cups. At rates
o E b I + I + I + I + I + I ' a b„ o of 2.5 to 10µ l/ container, both oils were repellent to Cx.N.'
CN 4 0O c`• 00 00 in " quinquefasciatus with significantly more gravid femalesU 4 4 ,--, 00 0o vi N •++ p
a r. " r" b u collected in the untreated containers compared to the
o 'o o .. 3 9. oil- treated containers( TABLE 2). Benzon and Apperson
g o 00 °` N 3 ( 1988) noted that assays which directly measure the0 3 1 + 1 ++ I ++ I ° + 1 ct > effect of volatiles on oviposition behavior must be0
0.) M ° ok C • y designed where the female is not able to contact theas F1 i N N rn O • cA
w3a. surface of the water before ovipositing and used wire
b o screen cones in their attractant assays. This experimental
r? b, o _. design offers direct evidence of oil repellency as gravida, a M H H ? Y females were able to detect volatiles from at least 9 cm
g •e 8 w w A o and avoid those containers.
bGQ a
El3 to
W 0 4 q ty ' o g w Field Experiments1 a> a) O. v a) GL „ p o N
ccqq a o 0 0 o o c vi ,3, In the first field experiment, both Golden Bear 1111
a p u; u1 a and Rodspray MFSTS( 12. 51iters/ ha) showed repellency
throughout much of the sampling period( Fig. 3A). Theuntreated ponds showed the usual pattern of Culex
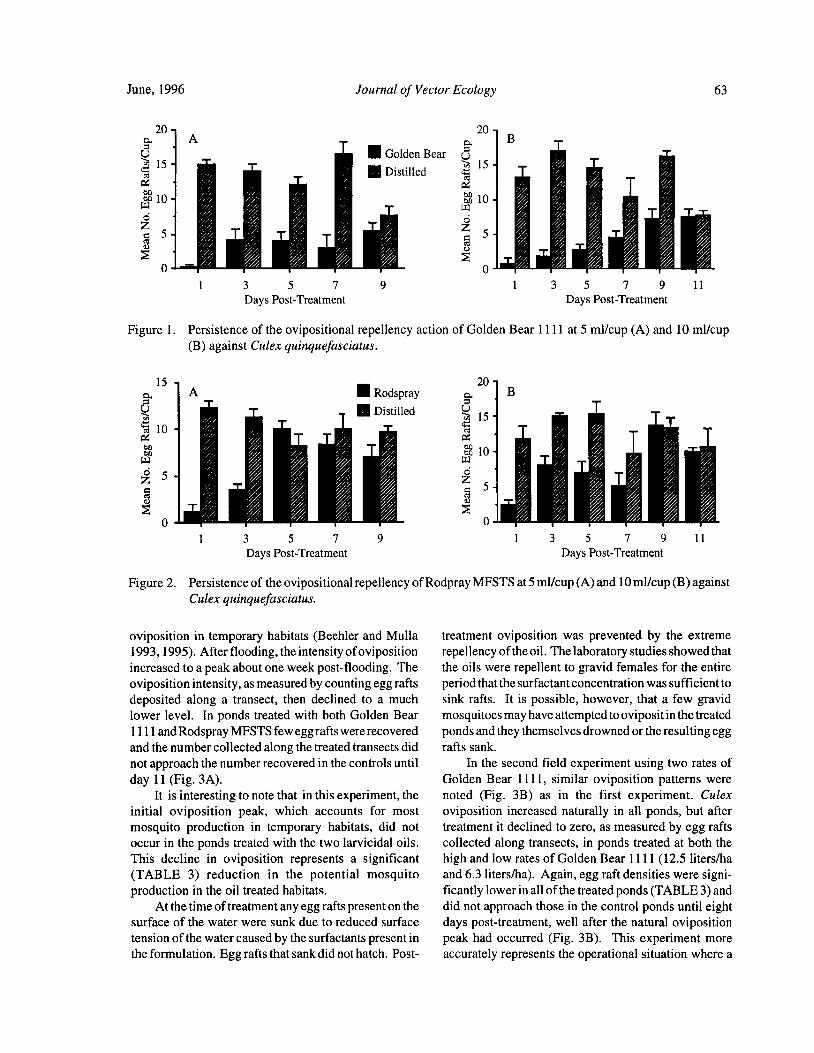
June, 1996 Journal of Vector Ecology 63
20- 20-a A
a B
U U Golden Bear ° U15 -
Distilled 4 15 -oG rx
bo 10- 10- /-; /
s;
i
5 _ Z g '
1 3 5 7 9 1 3 5 7 9 11
Days Post-Treatment Days Post- Treatment
Figure 1. Persistence of the ovipositional repellency action of Golden Bear 1111 at 5 ml/cup( A) and 10 ml/cupB) against Culex quinquefasciatus.
15 - 20-
a. A Rodspray a • B
U Distilled15 -
g geu
o 5 -
W10
jZ
i z i;
j 5
1 3 5 7 9 1 3 5 7 9 11
Days Post-Treatment Days Post-Treatment
Figure 2. Persistence of the ovipositional repellency ofRodpray MFSTS at 5 ml/cup( A) and 10 ml/cup( B) againstCulex quinquefasciatus.
oviposition in temporary habitats ( Beehler and Mulla treatment oviposition was prevented by the extreme1993, 1995). After flooding, the intensity ofoviposition repellency of the oil. The laboratory studies showed thatincreased to a peak about one week post- flooding. The the oils were repellent to gravid females for the entire
oviposition intensity, as measured by counting egg rafts period that the surfactant concentration was sufficient to
deposited along a transect, then declined to a much sink rafts. It is possible, however, that a few gravid
lower level. In ponds treated with both Golden Bear mosquitoes may have attempted to oviposit in the treated1111 and Rodspray MFSTS few egg rafts were recovered ponds and they themselves drowned or the resulting eggand the number collected along the treated transects did rafts sank.
not approach the number recovered in the controls until In the second field experiment using two rates of
day 11 ( Fig. 3A). Golden Bear 1111, similar oviposition patterns were
It is interesting to note that in this experiment, the noted ( Fig. 3B) as in the first experiment. Culexinitial oviposition peak, which accounts for most oviposition increased naturally in all ponds, but aftermosquito production in temporary habitats, did not treatment it declined to zero, as measured by egg raftsoccur in the ponds treated with the two larvicidal oils. collected along transects, in ponds treated at both theThis decline in oviposition represents a significant high and low rates of Golden Bear 1111 ( 12.5 liters/ha
TABLE 3) reduction in the potential mosquito and 6. 3 liters/ha). Again, egg raft densities were signi-production in the oil treated habitats. ficantly lower in all of the treated ponds( TABLE 3) and
At the time of treatment any egg rafts present on the did not approach those in the control ponds until eight
surface of the water were sunk due to reduced surface days post- treatment, well after the natural oviposition
tension of the water caused by the surfactants present in peak had occurred ( Fig. 3B). This experiment more
the formulation. Egg rafts that sank did not hatch. Post- accurately represents the operational situation where a

64 Journal of Vector Ecology June, 1996
habitat found to have larvae is treated with oil. The
subsequent reduction of mosquito production in that
v habitat is the sum of larval mortality and the gravidfemales which do not utilize the breeding source while
u 0 0 it is unattractive.0 0 3 All larvae identified from egg rafts reared
4.) individually in the laboratory were again identified to be0 3 N a 65: 35 mixture of Cx. quinquefasciatus and Cx.
aa b stigmatosoma. The species composition did not change
c . in either the treated or untreated ponds, nor did it changeo•
o q pre- and post treatment.
aoa
U N N In conclusion, these studies demonstrate that
c a larvicidal oils used in mosquito control programs exerto :
a strong repellent action against gravid Culex. This00
rs 0 v-, ,-, o effect may aid mosquito control operations in temporary
B + i - Ti 3 habitats for a number of reasons. First, egg deposition
qo 0 c
M in the habitat can be blocked at the early period ofE — o flooding when the larval food source is nutritionally
ac„ most rich and oviposition intensity is high. Second, the
N
y b ntural peak of oviposition in temporary habitats,V b r`
g occurring in the first two to three weeks of flooding,0
G C
I + I 3 resulting in most mosquito production can be eliminated.o o M ; o Third, the early oviposition by gravids in a habitat
3 a
U rn oo O which allows resulting larvae to escape predator0 ...... 0
VI
pressure can be delayed until the body of water agesa 1. and supports an abundant predator population. On the
w1r, 3 other hand, the use of oils may delay the establishment
0 S TI T y of macroinvertebrate predators through mortality and/
M W u 3 or repellency. This aspect requires further investigation.
a o
a = °o Aknowledgments
o E0 V This research was supported by the Orange County
o ' b 3 Vector Control District, Southeast and Coachella Valleyz 0 N
o 00 t3 gq Mosquito Abatement Districts, Northwest Mosquitoa.•a U M d- 0 a and Vector Control District, and the University ofx v a in a) Y
b S o California Special Funds for Mosquito Research.r o q 3 0
o N 09 N p
REFERENCES CITED
0 0 = I + I ° aio
a~ ; o Q o ` 6 Beehler, J. W. and M. S. Mulla. 1993. The effect of
o o F" m
organic enrichment and flooding duration on theoo
w 0 a0i oviposition behavior of Culex mosquitoes. Proc.
a) E , y o Calif. Mosq. Vector Control Assoc. 61: 121- 124.0 0 0 2 u
Beehler, J. W. and M. S. Mulla. 1995. Effects of organicz
o H enrichment on temporal distribution and abundancea4 5
o L 3 ° ofCulicine egg rafts. J. Am. Mosq. Control Assoc.NI 71 cq 0 8 11: 167- 171.
a ca 0 o Benzon, G. L. and C. S. Apperson. 1988. Reexamination
d o o C a of chemically mediated oviposition behavior inF•4 a C7 ( X
NAedes aegypti ( L.) ( Diptera: Culicidae) J. Med.Entomol. 25: 158- 164.
Box, G. E. P., W. G. Hunter, and J. S. Hunter. 1978.
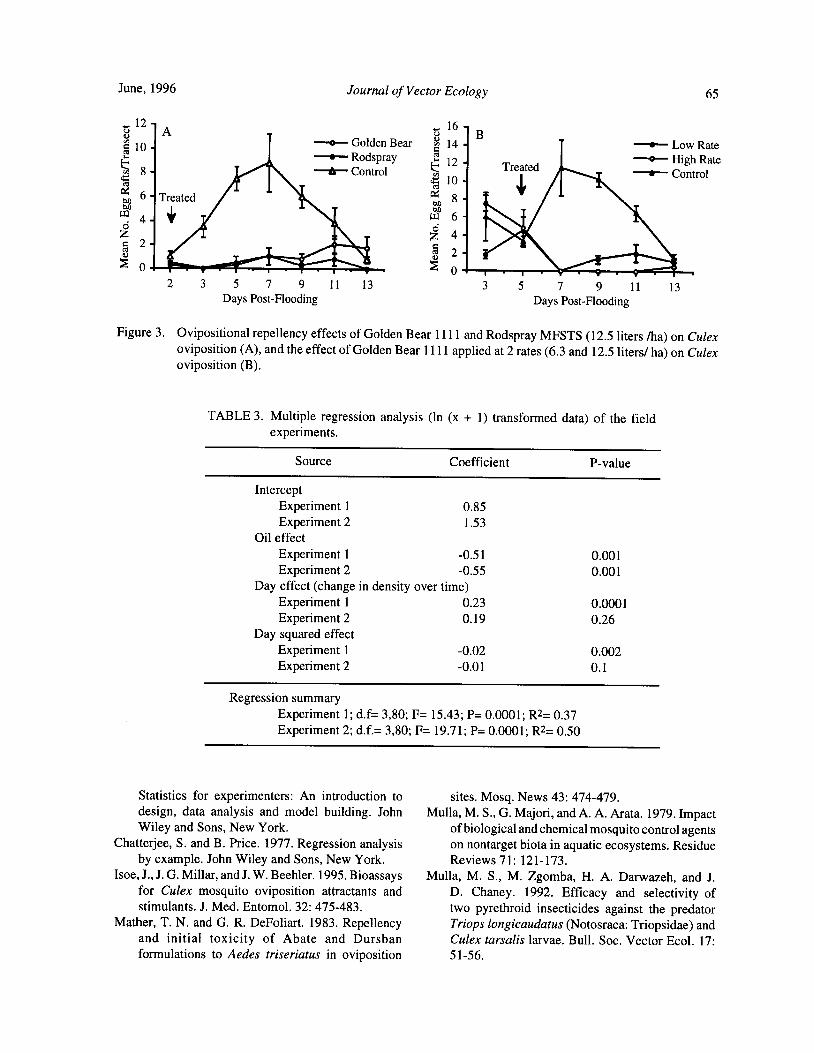
June, 1996 Journal of Vector Ecology 65
12A
16B
i 10o— Golden Bear N 14 Low Rate
Rodspray 12 0— High Rate
4 8 at—Control10
Treedt-- Control
a: 6 Treated 8bil
W4 W 6
0Z Z 4eg 2
2
0
2 3 5 7 9 11 13 3 5 7 9 11 13Days Post- Flooding Days Post- Flooding
Figure 3. Ovipositional repellency effects of Golden Bear 1111 and Rodspray MFSTS( 12. 5 liters/ ha) on Culexoviposition( A), and the effect of Golden Bear 1111 applied at 2 rates( 6. 3 and 12.5 liters/ ha) on Culexoviposition( B).
TABLE 3. Multiple regression analysis ( In ( x + 1) transformed data) of the fieldexperiments.
Source Coefficient P- value
Intercept
Experiment 1 0.85
Experiment 2 1. 53
Oil effect
Experiment 1 0.51 0.001
Experiment 2 0.55 0.001
Day effect( change in density over time)Experiment 1 0.23 0.0001
Experiment 2 0. 19 0.26
Day squared effectExperiment 1 0.02 0.002
Experiment 2 0.01 0. 1
Regression summaryExperiment 1; d.f=3, 80; F. 15. 43; P. 0.0001; R2= 0.37Experiment 2; d.f.=3, 80; F. 19. 71; P. 0.0001; R2= 0.50
Statistics for experimenters: An introduction to sites. Mosq. News 43: 474-479.design, data analysis and model building. John Mulla, M. S., G. Majori, and A. A. Arata. 1979. Impact
Wiley and Sons, New York. ofbiological and chemical mosquito control agentsChatterjee, S. and B. Price. 1977. Regression analysis on nontarget biota in aquatic ecosystems. Residue
by example. John Wiley and Sons, New York. Reviews 71: 121- 173.Isoe, J., J. G. Millar, and J. W. Beehler. 1995. Bioassays Mulla, M. S., M. Zgomba, H. A. Darwazeh, and J.
for Culex mosquito oviposition attractants and D. Chaney. 1992. Efficacy and selectivity ofstimulants. J. Med. Entomol. 32: 475- 483. two pyrethroid insecticides against the predator
Mather, T. N. and G. R. DeFoliart. 1983. Repellency Triops longicaudatus( Notosraca: Triopsidae) andand initial toxicity of Abate and Dursban Culex tarsalis larvae. Bull. Soc. Vector Ecol. 17:formulations to Aedes triseriatus in oviposition 51- 56.

Journal of Vector Ecology 21( 1): 66- 80
Species Concurrence and the Structure of a
Community of Aquatic Insects in Tree Holes
R. Barrera'
Department ofBiology, The Pennsylvania State UniversityUniversity Park, PA 16802, USA.
Permanent Address: Instituto de Zoologia Tropical, Facultad deCiencias, Universidad Central de Venezuela, Apartado 47058,
Caracas 1041-A, VENEZUELA., E-mail: rbarrera @dino.conicit.ve.
Received 8 May 1995; Accepted 13 November 1995
ABSTRACT: Habitat and temporal partitioning, species concurrence, and community structure of theaquatic insects of tree holes were investigated in central Pennsylvania, USA. Tree holes were sampled
completely for the immature stages and observations on locomotion, gas exchange, location, feeding mode,and food type were used to classify the species into guilds. A total of 20 species was observed to exploitthe aquatic habitats, although the mean number of species observed simultaneously was four( range 2- 7).The classification analyses reported four guilds of aquatic insects: predators, suspension- feeders, deposit-feeders, and scrapers. The numeric structure showed that predators were uncommon, and that only one
species was numerically dominant in each guild: Aedes triseriatus ( suspension- feeders), Culicoides
guttipennis ( deposit- feeders), and Elodes pulchella ( scrapers). These species represented 95% of all
insects. The analysis of concurrence revealed that most species encountered each other independently andin proportion to their occurrence. This pattern ofencounters determines asymmetrical potential interactionswith the frequent species at a disproportionate advantage. Temporal patterns in habitat partitioning were
determined by adaptations for the winter at this latitude. Predation and desiccation are not frequentecological factors, but freezing and perhaps food limitation are. There is convergence in communitystructure with those of neighboring states, but lack of convergence with assemblages located at differentlatitudes or biogeographical regions.
Keyword Index: Ecology, community structure, aquatic insects, mosquitoes, Aedes.
INTRODUCTION of small bodies of aquatic habitats contained in plants
collectively called phytotelmata( Varga 1928; KitchingCavities in trees that accumulate water and sustain 1971; Maguire 1971, Frank and Lounibos 1983), such
communities ofarthropods with aquatic immature stages as leaf axils of tank-bromeliads, bamboo internodes,
are widespread throughout the world( Fish 1983). Tree and pitcher plants. Worldwide over 1, 500 species of
holes are particularly attractive for studies ofcommunity plants contain phytotelmata( Fish 1983), most of them
organization because of their relative simplicity in habitat tropical. From a faunistic point of view, tree holes and
structure, small size, discrete boundaries, and well- phytotelmata communities show similar patterns in the
defined communities. Tree- hole assemblages can be major types oforganisms, particularly insects and aquaticused as model communities for experimental study of mites that inhabit them( Thienemann 1934; Fish 1983).
factors determining the structure of the community Most faunistic studies of tree holes have con-
along altitudinal and latitudinal gradients, islands vs. centrated on mosquitoes because of their medical
continents, and in general, to determine the relative importance, generally well- known natural histories, andimportance of local vs. regional factors on community the ease of experimental manipulations. Few authors
organization( Pearson 1986; Ricklefs 1987). Ecologi- have studied the other aquatic insects found in tree holes
cally, tree holes are part of a more general classification ( Thienemann 1934, Rohnert 1950 in Germany; Kitching

June, 1996 Journal of Vector Ecology 67
1971, 1983, 1986, Kitching and Callaghan 1982 in co-occur and what combination of factors are mostEngland and Australia; Bradshaw and Holzapfel 1992 likely to influence their coexistence in tree holes inin England; Snow 1949, Park et al. 1950, Woodward et Central Pennsylvania; and( 2) To explore whether thereal. 1988 in North America). Tree holes in different is ecological convergence in the community structure ofgeographical regions contain similar aquatic insects, tree holes in different biogeographical regions.most of them Diptera and Coleoptera. Analogous foodresources ( decomposing leaf litter), substrate ( water), MATERIALS AND METHODSand internal physical structure of the basin( walls, freewater, sediment, littoral zones of submerged objects, Tree holes and their communities were investigatedand beach zone) may partly explain the similarity in the in deciduous forests in central Pennsylvania, specificallycomposition of the major types of organisms exploiting in Rothrock State Forest in Centre and Huntingdonthe tree-hole resources in various parts of the world. counties. This area belongs to the Ridge and ValleyTree-hole insect species in England and Australia physiographic Province( Westerfeld 1959) and consistspartition food resources by particle size, with a group of of limestone valleys separated by parallel ridges ofspecies feeding directly on submerged leaves, and other sandstone. Elevations range from 304 m( 1, 000 feet) togroups feeding on smaller pieces and particles coming 734 m( 2, 407 feet) above sea level, and samples werefrom the original leaf litter ( Kitching 1983). North taken between 310-650 m. Approximate latitude of theAmerican tree- hole insects partition the space within study area is between 40° 37' and 40° 45' N. The area isthe tree hole( Snow 1949). Observations on the mosquito described as an oak-chestnut forest region ( Hui- linsub- communities in Europe ( Bradshaw and Holzapfel 1972), although chestnut is no longer present. Most of1986, 1991), North America( Bradshaw and Holzapfel the tree holes studied were in oaks( Quercus spp.) and1983, 1985, 1988; Copeland and Craig 1990; Lounibos maples( Ater spp.), the most common trees. A few tree1983, 1985) and Africa( Lounibos 1981) indicate within- holes were studied in beech ( Fagus sp.) and yellow
and between- tree-hole segregation among mosquito birch( Betula lutea Michaux).species. Coexistence in the spatially- limited habitat of Mean annual temperature and mean accumulatedtree holes could be partly explained by resource( food, precipitation for Pennsylvania are 9. 7° C and 1, 027. 5space) partitioning. However, observations on the gut mm( 1931- 1977; National Climatic Center, 1978). To
content of tree- hole insects in western North America determine the probability of finding tree holes withshows overlap in food items ( detritus particles, given levels of water, 22 tree holes randomly selected inprotozoans, fungal hyphae; Woodward et al., 1988). the study area were visited during 20 weeks in 1985.Potential interactions among species could be minimized Levels of water were: dry, wet, low, intermediate, andif there is spatial partitioning, or if the species members high level of standing water. The probability of findingin a community do not encounter each other most of the tree holes with a given level of water in the study sitestime( Grubb 1986). These tree- hole communities exhibit was calculated by dividing the total number of tree holespatterns of niche partitioning among coexisting species observed for this purpose with a given value, by the totalsimilar to those observed in many other natural number of observations ( 22 tree holes x 20 weeks =
communities( Schoener 1974; Toft 1985). Ecological 440). The probabilities calculated were: 4%( dry), 12%factors other than inter-specific competition, such as ( wet), 17%( low level), 33. 5%( intermediate level), andpredation, mutualism, abiotic factors, and migration can 33. 5%( high level).
also promote resource partitioning ( Schoener 1986;Bradshaw and Holzapfel 1983). Sampling the Community
Bradshaw and Holzapfel ( 1985) predicted that at Each tree hole was taken as the sample unit and nohigh northern latitudes freezing and competition should subsampling within a single tree hole was feasiblebecome the important factors organizing the guild of because of the high variation in tree- hole shape andtree- hole mosquitoes, because desiccation should structure, and because insect species required usingdecrease in importance, and because the main predator different techniques for extraction. A preliminaryToxorhynchites rutilus( Coquillett) is rare or absent at sampling of seven tree holes showed that when the leafhigh latitudes. The factors structuring the tree-hole litter was removed from the cavity, it contained Aedesmosquito assemblages at higher latitudes in North mosquitoes( 47%), ceratopogonids( 28%), and scirtids
America have not been studied quantitatively, nor has ( 25%). Pipeting( diameter 5 mm) the free water reportedthe role of other aquatic insect species in structuring the more mosquito larvae( 86%) than ceratopogonids( 6%)
community. The objectives of this study are: ( 1) To or scirtids ( 8%), whereas in the sediment, the insectdetermine the extent to which the component species composition was similar to that of the leaf litter. So I

68 Journal of Vector Ecology June, 1996
decided to sample each tree hole completely. The and( 5) Food type more likely used( leaflitter,particulatecontents were sampled by first collecting the litter and organic matter deposited, particulate organic matter
coarse materials that would interfere with the siphoning suspended in the water, aquatic insects; Merritt and
of the tree- hole water. After the standing water was Cummins 1978).
siphoned, the sediment accumulated at the bottom of the The attributes( TABLE 1) were used to classify the
cavity was collected to a depth of at least 3 cm. Tree insects into groups using a cluster analysis ( BMDPholes were flushed at least three times with aerated tap Statistical Software 1982). The variables were assigned
water, and the fluid collected with the pipette until no discrete ranks at each level( e.g., for the tree- hole zone,more insects were apparent in the tree hole. Tree-hole beach= 1, littoral= 2, etc.). Euclidean distance or the
contents were placed in plastic bags and transferred to square root of the sum of squares of the differences
the laboratory in ice boxes for their immediate processing. between the values of the standardized variables for the
Coarse material was washed in pans in the laboratory to species was used as the criterion for similarity. Singledetach the insects, and the water was then passed through linkage algorithm was used to create the clusters where
a series of sieves to sort out the sample. The smallest the species were assigned to clusters with members
pore diameter used was 0.25 mm. Counting was closest to the species.
performed on live insects because they were easier todetect than dead or preserved insects due to the large Interspecific Encounters
content of small particles in the samples. A stereo- To determine how frequently tree- hole organismsmicroscope was used to search for small insects and encountered each other in natural tree holes, I calculated
early instars. A total of 160 tree holes( from 0 to 2 m the frequency of intraspecific and interspecificabove ground level) was located and marked in the study encounters. The frequency of intraspecific encounterssites, and a random sample of 78 tree holes was or percentage of occurrence is the percentage of tree
destructively collected between April and September of holes where the species occurred. Interspecific encounter
1985 and 1986. of species" A" with species" B" is the percentage of tree
The most common tree- hole insects were reared in holes where both species occurred divided by the numberthe laboratory to make observations on behavior and of tree holes where only species" A" was present. Thesedevelopment and to obtain adults for identification values of interspecific encounters between a pair of
purposes. Field observations of undisturbed tree holes species are not symmetrical, unless the number of tree
were periodically made to record behavior and stage of holes occupied by each species was the same. Assumingdevelopment of the more conspicuous insects. independence, the percentage with which species A
encounters species B is equal to the frequency of B
Determining Functional Roles of Species occurrence). To test whether species A and B were
Five aspects of the tree-hole organisms were independently distributed, 2 x 2 Contingency Tablesobserved or investigated from the literature available to ( with continuity correction) were calculated using
report on functional roles and trophic structure of the presence- absence data, along with Fisher' s tests( SASassemblage ( Acarina: Fashing 1975, 1976, pers. Institute Inc. 1987) if the expected value of one or more
commun.; Staphylinidae: J. H. Frank pers. commun.; cells was less than five. Lloyd' s( 1967) indices of mean
Scirtidae: Thienemann 1934, Snow 1949, Fish 1983, intra- and interspecific crowding were calculatedFrank 1983, Kitching 1983: Ceratopogonidae: Snow following Bradshaw and Holzapfel' s ( 1983)
1949, Foote and Pratt 1954, Hair and Turner 1966, Hair modification to express crowding of immatures per literet al. 1966; Culicidae: Dyar 1928, Jenkins and Carpenter of tree- hole water. Intraspecific crowding is the mean1946, Breland 1960, Truman and Craig 1968, Zavortink number of congeners encountered by an individual per1968, 1970, 1972, Petersen et al. 1969, Grimstad et al. liter of tree-hole water, whereas interspecific crowding1974, Lunt 1977, Scholl and DeFoliart 1977, Sinsko and is the mean number of individuals of a species
Grimstad 1977; Psychodidae and Syrphidae: Snow encountered by an individual of another species.1949). These aspects were: ( 1) Tree-hole zone where
organisms spend most of the time ( beach, littoral, RESULTS
profundal, limnetic);( 2) Means of locomotion( crawlers,
burrower- crawler, burrower- swimmer, climber, Trophic Structure
swimmer); ( 3) Gas exchange structure ( short siphon, Eighteen species of aquatic arthropods and two
long siphon, spiracles, cutaneous);( 4) Functional feeding terrestrial species were collected from tree holes in the
group ( scrapers, deposit feeders, suspension- feeder, study area ( TABLE 1). The classification analysis
suspension- feeder/ browser, predator of other insects); ( Fig. 1) shows that mosquito species( Ae. triseriatus,Ae.
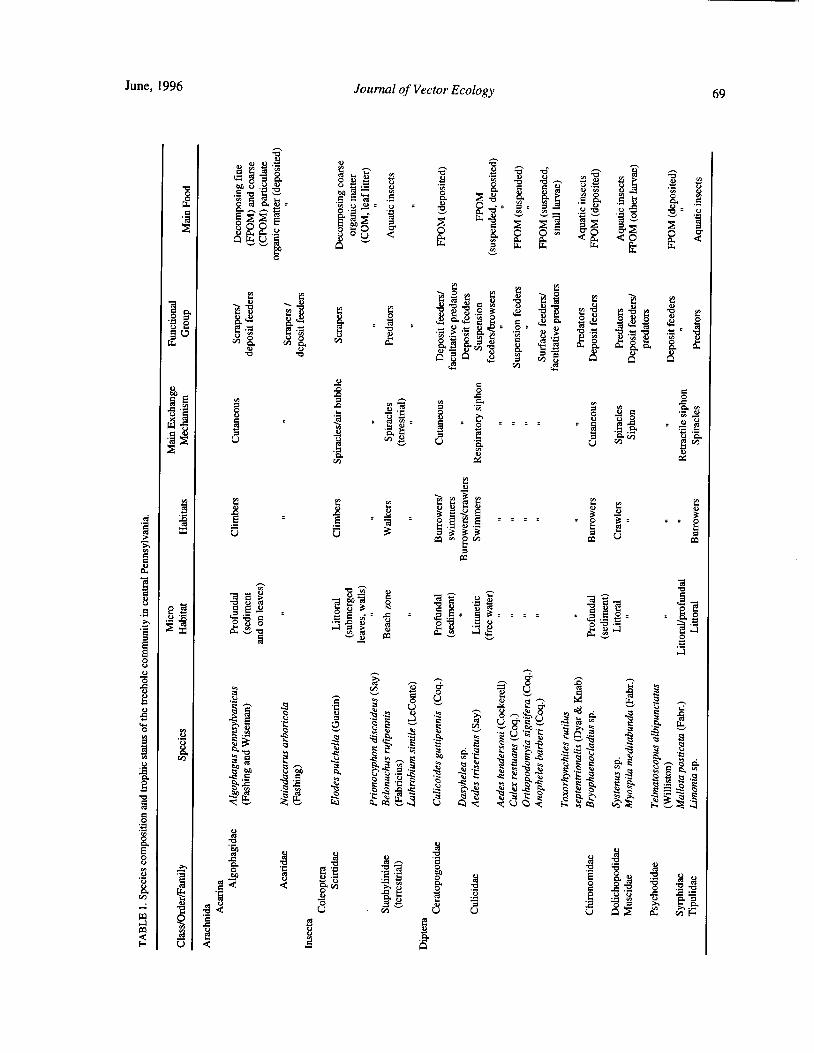
June, 1996 Journal of Vector Ecology 69
ts
8w O 0 4 U L"
U O 'o ' d
U U0.
3U
N
4 .
4
4 0.
U y N y 0
EU h y y
4. o A 4i : 5
u :? ° U n O - - g : g . U EL a • c
oE 2 4 E Pn a
we 3 = o2 2 2 2 12 .
1 o " O O 0 0 h °' O ° a
0
O 2
o da y aui
a Q Y 8. 4-
4y&i . ; _
w y w a ° _
0 = w 4 ; w
0 0 0 O O U N . A N 4 y 0 0 Vl 0 ' y • yIL, i' n at g. o a ; a a, cC $, 4 $, at49.
Aw
A y52
A A L
00 0
0.0
GOD.
E
Q00 o ' 1 o .°' 0 y 0. H
LI f0O
U
O _ - - N U ° ..
u ' id
12 U8
Cl U a V AN
4 U0
rn a
4
6
ifl 0
0 3 . . . 0x U U 3 y 0 vi
mgU
ogo
0c o
00aN o
0a
u
E : 53 : : 11! ::k.'
z..' -0 0 , so0 ga
OUU
y2UA0 q —
0O
o
Jy 9
G 0 U
u yd
E0
0t.J b • ^ a, o ` h
g _ ° a 1 0w C 0 0 0 t% U e U ' cl y
C ,0 rn y O y cl , v y
O oo. 0 A0•
L7 U v 3 C b v O 4, a -O yQb 0 C CR o
0°°
0 C i Cpip "
ycy
i .~ Op 04 C .`
U'443-
O. OA O Q a. T V ..=.
e
b N .w - C i 0 - C . O obi y c' O R. 00 2 d o o r
R Q, o R q = o
QEo
ig
w ov dbo C o R 5 0 E ' 3
g z w a.. c:0 v Ae < 0Q E2 5w h' ti3
a0pN 038 8
H O c
0 O
c0
yN 0,0 5 y y' 2.
w7
cda R7
ti C• U 7w 4 v
IP, y ° o y .?Q U i U U U A a r4 F
o A 2. Ea4
FF U A
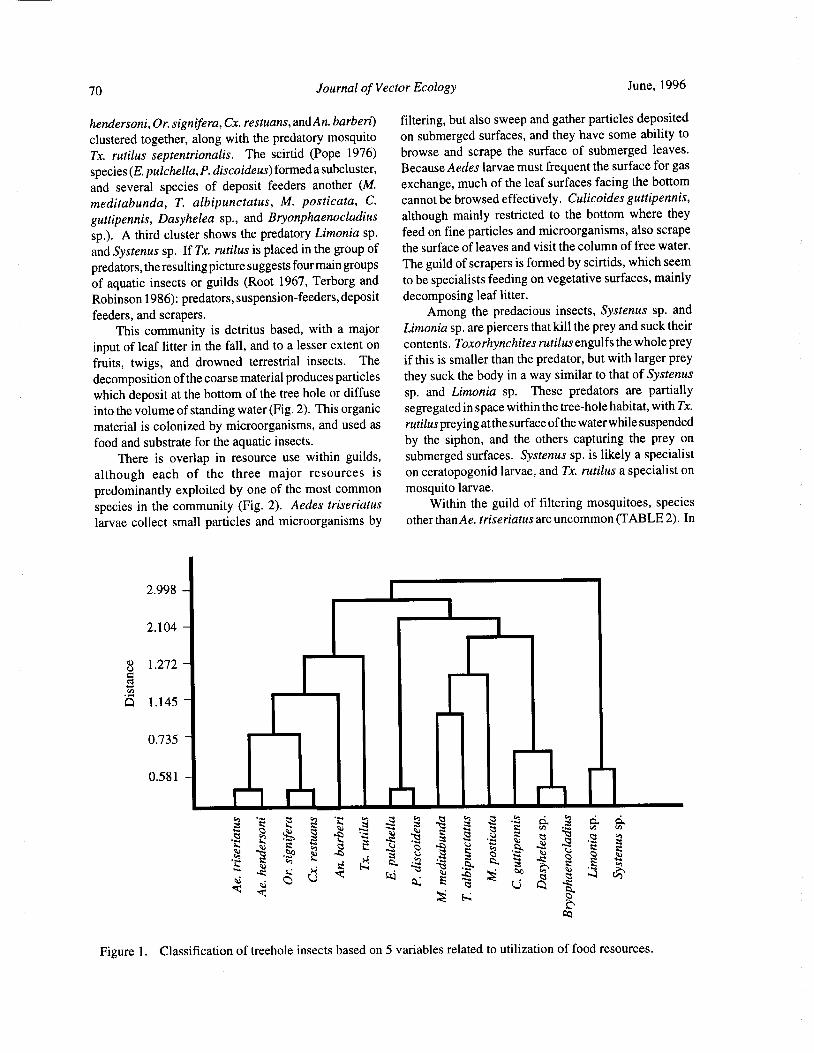
70 Journal of Vector Ecology June, 1996
hendersoni, Or.signifera, Cx. restuans, andAn. barberi) filtering, but also sweep and gather particles depositedclustered together, along with the predatory mosquito on submerged surfaces, and they have some ability to
Tx. rutilus septentrionalis. The scirtid ( Pope 1976) browse and scrape the surface of submerged leaves.
species( E.pulchella,P. discoideus) formed a subcluster, Because Aedes larvae must frequent the surface for gas
and several species of deposit feeders another ( M. exchange, much of the leaf surfaces facing the bottom
meditabunda, T. albipunctatus, M. posticata, C. cannot be browsed effectively. Culicoides guttipennis,
guttipennis, Dasyhelea sp., and Bryonphaenocladius although mainly restricted to the bottom where they
sp.). A third cluster shows the predatory Limonia sp. feed on fine particles and microorganisms, also scrape
and Systenus sp. If Tx. rutilus is placed in the group of the surface of leaves and visit the column of free water.
predators, the resulting picture suggests fourmain groups The guild of scrapers is formed by scirtids, which seem
of aquatic insects or guilds ( Root 1967, Terborg and to be specialists feeding on vegetative surfaces, mainly
Robinson 1986): predators, suspension- feeders, deposit decomposing leaf litter.feeders, and scrapers. Among the predacious insects, Systenus sp. and
This community is detritus based, with a major Limonia sp. are piercers that kill the prey and suck theirinput of leaf litter in the fall, and to a lesser extent on contents. Toxorhynchites rutilus engulfs the whole prey
fruits, twigs, and drowned terrestrial insects. The if this is smaller than the predator, but with larger prey
decomposition of the coarse material produces particles they suck the body in a way similar to that of Systenuswhich deposit at the bottom of the tree hole or diffuse sp. and Limonia sp. These predators are partially
into the volume of standing water( Fig. 2). This organic segregated in space within the tree-hole habitat, with Tx.
material is colonized by microorganisms, and used as rutilus preying at the surface ofthe water while suspendedfood and substrate for the aquatic insects. by the siphon, and the others capturing the prey on
There is overlap in resource use within guilds, submerged surfaces. Systenus sp. is likely a specialistalthough each of the three major resources is on ceratopogonid larvae, and Tx. rutilus a specialist on
predominantly exploited by one of the most common mosquito larvae.
species in the community ( Fig. 2). Aedes triseriatus Within the guild of filtering mosquitoes, species
larvae collect small particles and microorganisms by other than Ae. triseriatus are uncommon( TABLE 2). In
2.998 —
2. 104 —
1. 272 —
IQ 1. 145 —
0.735 —
0.581 — nI I
ti a. `Q a•
cs E
ti 4iQ
c
Figure 1. Classification of treehole insects based on 5 variables related to utilization of food resources.
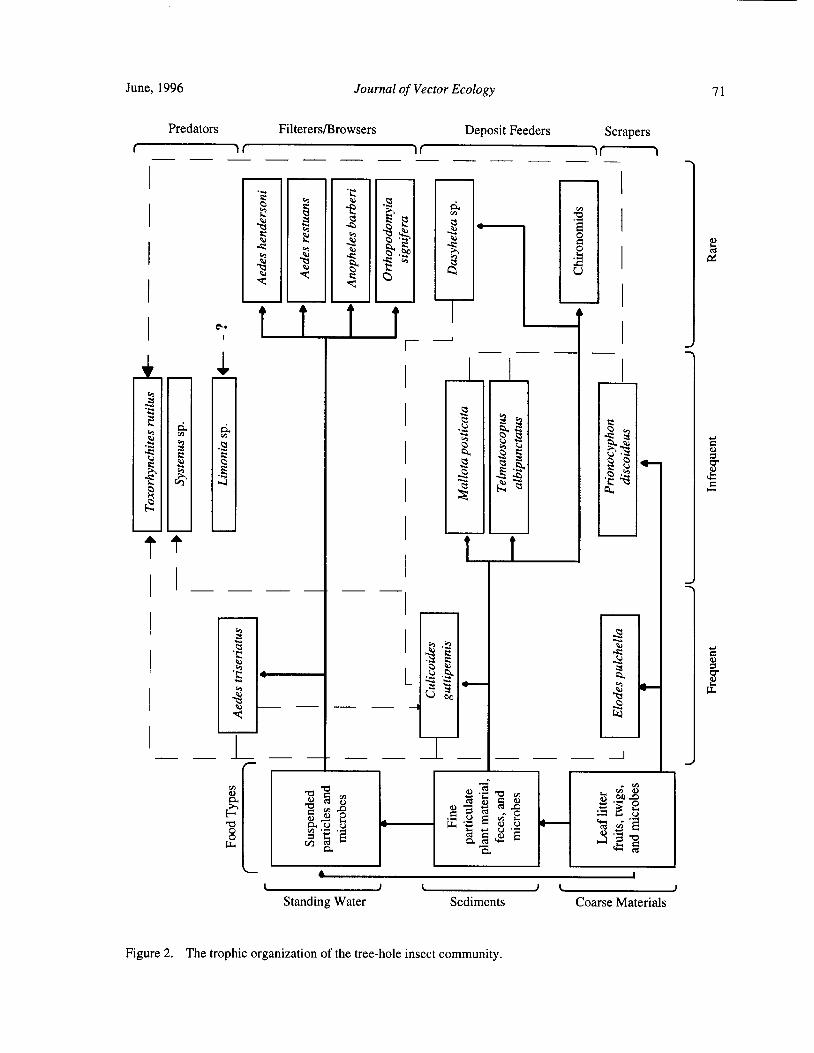
June, 1996 Journal of Vector Ecology 71
Predators FilterersBrowsers Deposit Feeders Scrapers
1 1( if
b y o E40 i O
a
a)5 01 O0.. p0 -L 0
a)
a0 c:1 U
fl__ I tlir 1 _ _
0
6. y
o o zC oi., y O CZ) v
0 0
O
O
1 ? L 1
1 1
1U C
cr
y w - wV °° ool
Q W
Lb r. aa) o ti 6
o oy
O
w czw a E),acl,•
I
t t J
Standing Water Sediments Coarse Materials
Figure 2. The trophic organization of the tree-hole insect community.

72 Journal of Vector Ecology June, 1996
this guild, segregation within a tree hole is limited aquatic mites were observed on submerged leaves and
because mosquito species feed on suspended particulate sediment material in the laboratory. Locomotion is bymatter and microorganisms contained in the volume of walking on submerged surfaces and gas exchange isfree-water. However, some segregation occurs between cutaneous( TABLE 1). Between hole dispersal is phoretic
An. barberi and the other mosquitoes because Anopheles on adult females of the tree- hole syrphids, Mallota
larvae feed upon particles in the film of water at the air- posticata and M. bautias ( Walker), and dispersion is
water interphase. Aedes triseriatus occasionally feeds carried out only in the immature hypopus stage( Fashingat this layer, usually when there is little suspended 1975, 1976). Naiadacarus arboricola can feed directly
material in the water. Culex restuans, Or. signifera, and on leaves, whereas A. pennsylvanicusismainly a scraper,
An. barberi occurred in large, permanent tree holes, and feeding on fungal mats growing on the decomposingAe. hendersoni occurred in a few elevated tree holes. leaves( Fashing, pers. commun.), although presumably
Within the guild of deposit feeders, C. guttipennis, both use fine particulate material accumulated on the
Dasyhelea sp., Bryophaenocladius sp., and T. bottom of the cavity.albipunctatus seem to feed in the layer of sediment. The other two species not included in the guild
However, C. guttipennis larvae have access to other analysis were the terrestrial predatory beetles Belonochuszones of the tree hole by active swimming, and T. rufipennis and Lathrobium simile ( TABLE 1).
albipunctatus occurs in shallow areas. Syrphids have a Belonochus rufipennis is a predator on aquatic insects
different mechanism of feeding from the rest of the and has a wide distribution from Canada to Brazil( J. H.
guild, and have relative independence of movement Frank, pers. commun.). This rove beetle species is
because of their retractile siphon. The most restricted common in the aquatic floral bracts ofHeliconia caribaea
species in relation to locomotion are the uncommon Lamarck( Zingiberales: Heliconiaceae) and cacao husks
Dasyhelea sp. and Bryophaenocladius sp. in Venezuela ( pers. observ.), where these predators
The aquatic mites Algophagus pennsylvanicus and capture aquatic insects. Lathrobium simile has narrower
Naiadacarus arboricola are the most common mite distribution and is apparently restricted to eastern Northspecies in tree holes of eastern North America( Fashing America ( J. H. Frank, pers. commun.). The relative
1975; N. Fashing, pers. commun.). They were not abundance of these predators could not be determined
included in the classification because of uncertainty by the present method because they commonly hunt forabout their local distribution within natural tree holes. aquatic insects from the beach or littoral area. Their
However, their high frequency and abundance make impact on the aquatic community of tree holes has notthem important components of the community. The been evaluated.
TABLE 2 Mean and standard error of species abundance per tree hole and per liter of water, mean and
range of the relative frequency of species per tree hole, and percentage of tree holes occupiedor ocurrence.
Relative
Individuals Frequency of Species
Individuals per Liter of Individual per Occurrence
Species per Tree Hole Tree Hole water Tree Hole(%)
Aedes 298.2 64.9 818. 9 135. 3 43. 0 98. 8 96.2
Scirtids 198.7 33. 6 777. 3 159. 1 36.7 94.5 94.9
Culicoides 81. 7 12. 8 395. 8 101. 7 25. 0 61. 9 91. 0
Mallota 4.4 1. 5 21. 6 6. 1 2.7 88.2 37.2
Systenus 1. 7 0. 6 7. 3 2. 1 0.7 8. 3 48. 7
Telmatoscopus 1. 4 0.4 6.5 2.4 0.5 13. 3 26.9
Chironomids 0.7 0.7 2.8 2.6 0.2 11. 8 2. 6
Limonia 0. 5 0.2 1. 5 0.7 0.9 50.0 15. 4
Culex 0.4 0.3 0.6 0.5 0. 1 2.2 5. 1
Dasyhelea 0.2 0. 1 0. 8 0. 6 0. 1 0.6 3. 9
Anopheles 0. 1 < 0. 1 0.4 0. 2 0. 1 1. 2 6.4
Toxorhynchites 0. 1 < 0. 1 0. 1 0. 1 0. 1 2. 6 2. 6

June, 1996 Journal of Vector Ecology 73
Numeric Structurespecies commonly, encountered the infrequent species
The mean abundance of species( TABLE 2) can be uncommonly, and seldom encountered the rare species.separated into three groups: those with abundances in By comparing the intraspecific and interspecificthe hundreds( Ae. triseriatus, scirtids, and C.guttipennis), encounters(%) it was apparent that most of the speciesthose around the order of ten ( Mallota, Systenus, in the community were encountering other species inTelmatoscopus, Limonia), and those with mean values proportion to the relative abundance of the encounteredbelow unity( the rest of species, includingAe. hendersoni, species( e.g., independently). The rare species encoun-and M. meditabunda). Orthopodomyia signifera was tered the frequent species virtually always and encoun-observed in the study area but it did not appear in the 78 tered other rare species on few occasions or not at alltree holes sampled. The same division results, although ( i.e., the rare species occurred in only a few tree holesfor different scales, on the basis of the mean relative where the frequent species were nearly always present).frequencies and occurrence. These three classes of Only five out of 66 pairwise analyses of associationaquatic insects are termed frequent, infrequent, and rare rejected the null hypothesis of independence( P< 0.05).in the order just described. The frequent insects are Four of these cases involved the psychodids ( T.represented by only three species, the infrequent insects albipunctatus), which had positive associations withby five species, and the rare insects by seven species( not syrphids (M. posticata), dolichopodids ( Systenus sp.),all species shown in TABLE 2). The data were expressed and ceratopogonids ( Dasyhelea sp.), and a negativeas mean number oforganisms per liter of tree- hole water association with Aedes spp. The other significant testto allow comparisons with other studies. was a positive association between syrphids and
The mean relative frequency of species found per Dasyhelea sp. ( TABLE 3). The fact that most speciesTABLE 2) estimates the proportion of each aquatic were independently distributed across the tree holes
insect species in the community. It was calculated by does not mean that there was no concurrence. Indeed,averaging the proportion of each species per tree hole the most frequent species encountered each other inover all samples( n= 76). Aedes spp. represented 43% of most of the tree holes.all inhabitants per tree hole, scirtids 37%, and The mean number of congeners encountered, orceratopogonids 15%( 95% of all aquatic insects). The intraspecific crowding, exceeded the mean density( ind./range of values of the relative frequency shows that, for 1.) in every species ( TABLE 3), meaning that theat least one tree hole, Aedes spp. were 99% of the insects. dispersion of immatures per liter of tree-hole water wasOther species such as scirtids ( 95%), syrphids ( 88%), aggregated or patched( Lloyd 1967). Mean interspecifcand ceratopogonids( 62%) also became the most frequent crowding followed the same general tendencies observedspecies in single tree holes. with the percentage ofencounters( TABLE 3). However,
this measure stressed the asymmetry of the interspecificConcurrence and Species Associations encounters, particularly between the less abundant and
Mean and standard error ofspecies richness per tree frequent species and the more ubiquitous ones. Forhole ( number of species) were 4. 3 ± 0. 1 ( n= 78; example, An. barberi showed a low mean density( 0.4CV=28. 6%; range 2- 7). Only a few species out of the ind./ 1) and mean crowding ( 4.9 ind./ l), whereas thetotal sampled in the community( TABLE 1) concurred mean number of individuals of Ae. triseriatusin the same tree holes. encountered was rather large( 919.2 ind./ 1). On the other
The percentage of intraspecific ( TABLE 3; main hand, Ae. triseriatus encountered 1, 783. 2 congeners butdiagonal) and interspecific encounters between pairs of only 0.4 immatures of An. barberi( TABLE 3).species were analyzed in order of abundance per treehole ( TABLE 2). The rows ( TABLE 3) indicate the Seasonal Changes in Community Structurefrequency of encounters of the species listed with each The community was dominated numerically byof the other species on top ofeach column. For example, three species over time, with Aedes spp. beingAedes spp. encountered scirtids in 95% of the tree holes numerically dominant until June, scirtids becomingoccupied by this mosquito, C. guttipennis in 91%, etc. If proportionally more abundant later on, and
TABLE 3 is read from top to bottom the converse ceratopogonids keeping a relatively constant numericoutcome results, indicating the frequency with which a proportion ( Fig. 3). The relative frequencies of lessgiven species was encountered by the other species. For abundant species were not represented because theirexample, Aedes spp. was encountered by 96% of the combined relative frequency was 5% or less of the totalscirtids; the proportion of tree holes occupied by scirtids community. Only in June did syrphids become relativelythat also contained Aedes spp. abundant when many small larvae appeared in the tree
The frequent species encountered other frequent holes, likely as a result of recent oviposition.
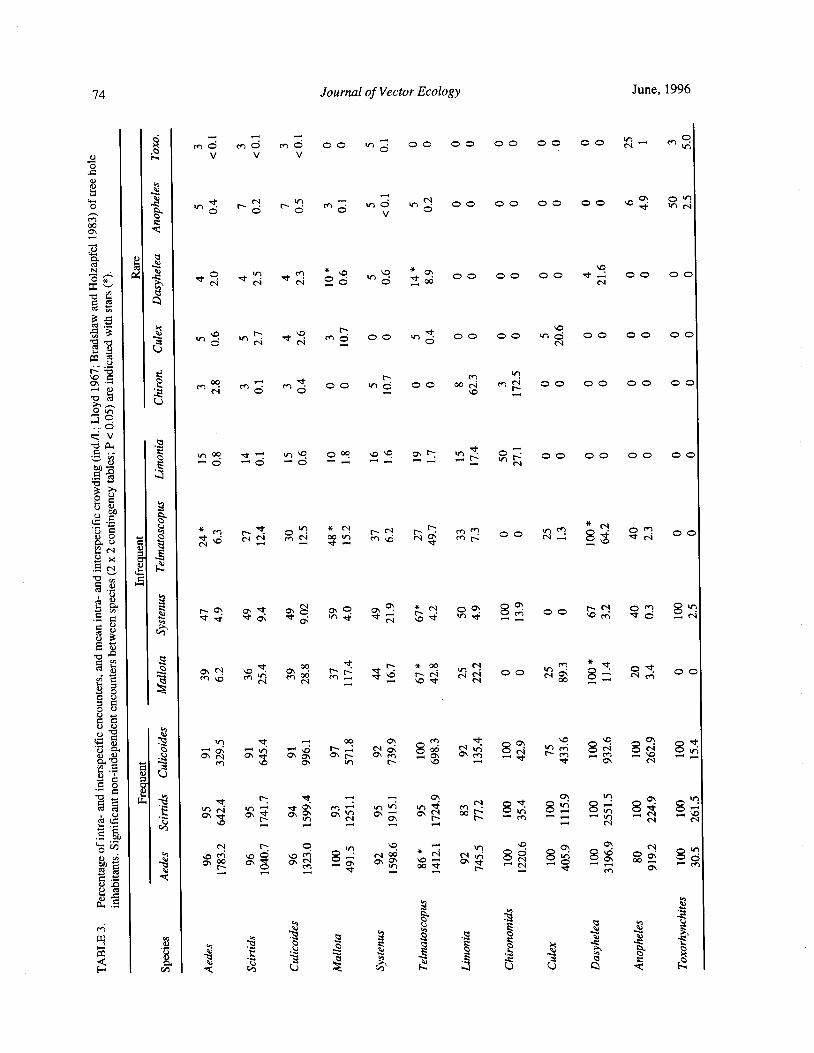
74 Journal of Vector Ecology June, 1996
M O en O M O 0 0 to . 0 0 0 0 0 0 0 0 0 0 N — M
y Er V V V
0
cuh
OO NO NC MO tn0 viC 00 00 00 00
X00 0 N
M0 V
003
ON Q
W
Nr
0- * N eri et • 0O• O• d' o000 00 00 fir. 0O 00
N
C Qc hN"+
cs
3 et
b
w U
en O 0 0 to O O O O p O O O A O O
C:64 1UvD C N M
h
Mao cn '.• M 00 tnp 00 00 c- MN 00 00 00 00
bN O O r k.0
o(A'U
4 C2p
V
O0 O O O O O O O O
eu O ^ O ^' O N
b VJr
O
U NU C R
0 0 h * en 'in * N N N
Nen en h M *
N0 M
y 0G
O qj N OM
dam' In M N0 MN
00 N ,--i 83 e} t,j 00
NN ok
CV )
LI4_,
Er
N H
rI• LL N O * N O 8
0! N 0 M 8 v1d - o 't N vpd' in4 , 0 M 4 O .-. N
etO 3v'
E 48b a
r-.) vcE
M MNON M et - NN NN 00 NON 8 - OM 00
0O
C
G U
O• 0
0C. 00N U
U b 014:
0y 00 ON en d o\ •
op
op
ONp
0 & O O: •.-. .-. N O\ 8 oó N to g , ,,; 8 c"i 8 i g •d N v O, N oN cA O\ O, N O, en .-. ON OA M . r
N M ... r en .-. ' 0 .. r „'_,
O. M ON v N kD .--, d' ON Ni
0. 'o O U
p ppen to to to 4 en
p p pc,
0N ON ON CA ON N 0 3 D el oo •N g M g - p I p N
O
NCW
Ip
vOp
D\ ON N
in
cd
c Oh00 O\ g O\ oO
O\ 000 -
a\ 80 80 8M 00a 80
r enen
ON
C,'Nw as
LL G m.
y
O Ven
so.) 5 Y , 0
0
O C O CC
0 C1: e N c
O -
Q a v v U o c
E. cn Q o- a [
a U U QO
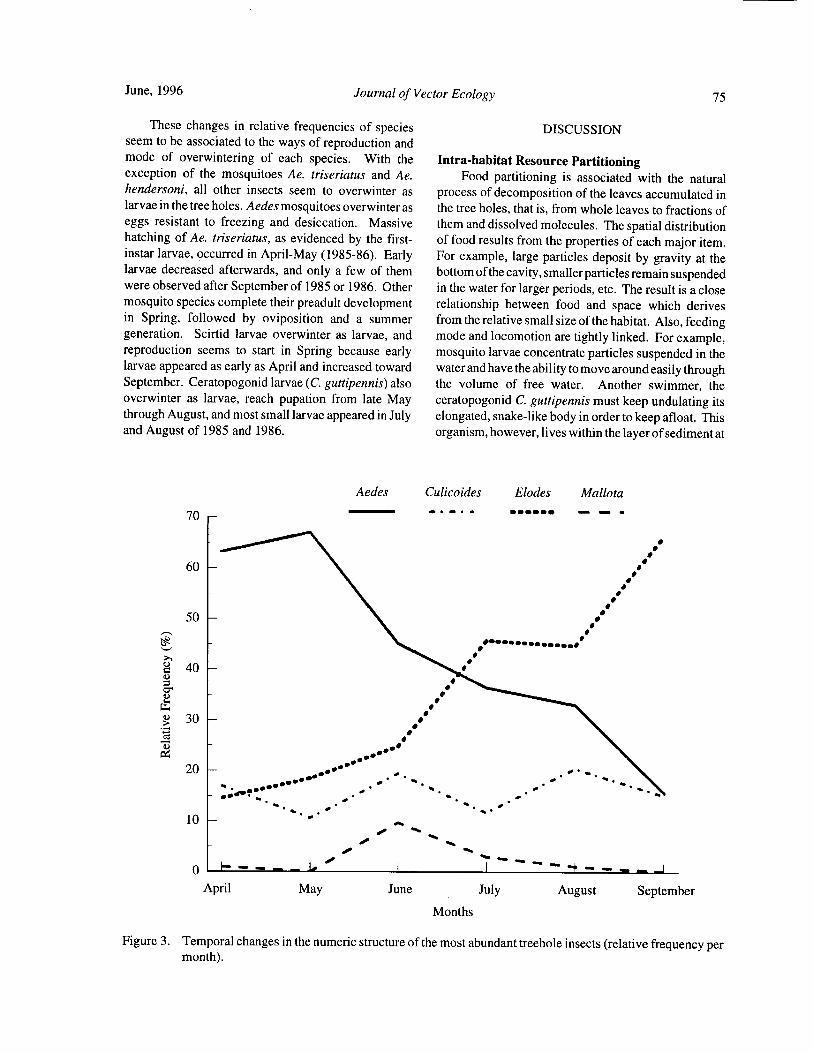
June, 1996 Journal of Vector Ecology 75
These changes in relative frequencies of species DISCUSSIONseem to be associated to the ways of reproduction andmode of overwintering of each species. With the Intra-habitat Resource Partitioningexception of the mosquitoes Ae. triseriatus and Ae. Food partitioning is associated with the naturalhendersoni, all other insects seem to overwinter as process of decomposition of the leaves accumulated inlarvae in the tree holes. Aedes mosquitoes overwinter as the tree holes, that is, from whole leaves to fractions ofeggs resistant to freezing and desiccation. Massive them and dissolved molecules. The spatial distributionhatching of Ae. triseriatus, as evidenced by the first- of food results from the properties of each major item.instar larvae, occurred in April-May( 1985- 86). Early For example, large particles deposit by gravity at thelarvae decreased afterwards, and only a few of them bottom of the cavity, smaller particles remain suspendedwere observed after September of 1985 or 1986. Other in the water for larger periods, etc. The result is a closemosquito species complete their preadult development relationship between food and space which derivesin Spring, followed by oviposition and a summer from the relative small size of the habitat. Also, feedinggeneration. Scirtid larvae overwinter as larvae, and mode and locomotion are tightly linked. For example,reproduction seems to start in Spring because early mosquito larvae concentrate particles suspended in thelarvae appeared as early as April and increased toward water and have the ability to move around easily throughSeptember. Ceratopogonid larvae( C. guttipennis) also the volume of free water. Another swimmer, theoverwinter as larvae, reach pupation from late May ceratopogonid C. guttipennis must keep undulating itsthrough August, and most small larvae appeared in July elongated, snake- like body in order to keep afloat. Thisand August of 1985 and 1986. organism, however, lives within the layer ofsediment at
Aedes Culicoides Elodes Mallota
70 —
60 —
50 —
S°
c 40 —
w
30 —
p4
20 —
w•'•••
I'
10 —
ti
0 "' - - - L
April May June July August September
Months
Figure 3. Temporal changes in the numeric structure of the most abundant treehole insects( relative frequency permonth).

76 Journal of Vector Ecology June, 1996
the bottom where its means of locomotion are more microorganisms involved in leaf litter decomposition.
adequate for moving around in a dense medium. Scirtid Kurihara and Kikkawa ( 1986) explained that
beetles use their legs to attach to firm substrate such as microorganisms generally enrich the quality ofdetritus.the leaves on which they feed. Other organisms, most However, bacteria and other organisms that can digest
of them deposit feeders, are burrowers and/ or crawlers cellulose, lignin, and other complex molecules occur at
TABLE 1). Syrphids crawl by extending and low densities and may not be an abundant source of food
compressing their soft body on surfaces, but cannot for detritivorous insects. Their energy is concentratedattach firmly, or move easily in a viscose medium, or through protozoa and other lower predators.
swim. They feed by sweeping particles accumulated on Consumption of microorganisms by one guild within
the substrate on which they move. the restricted space of tree holes might reduce the
There is another dimension that seems to intervene number of microbes available to colonize and grow on
in resource partitioning, which is the way of gas ex- other component substrata. Observations on insect gut
change. Tree-hole mosquitoes keep frequent contact contents by Woodward et al.( 1988) seem to indicate thewith the air through their siphons, either by visiting the use of similar resources by various aquatic insects. Insurface such as Aedes spp., or by remaining in contact practice, however, the number of insect species
with the surface most of the time while feeding, such as simultaneously occupying a tree hole in the study areaCx. restuans, Or. signifera, and An. barberi. The cera- was low ( 3 to 7 species) in relation to all species
topogonid C. guttipennis does not seem to visit the sur- encountered during the study. The trophic structure offace for gas exchange, and Snow( 1949) observed that the community ( Fig. 2) rarely might converge on athese organisms can exist for long in low oxygen water. single, discrete habitat as to trigger many interactions
It seems that at the level of the four guilds identified among most species. In fact,only three species concurredin the tree-hole community, organisms partition resources in most tree holes, each one being the most frequent andbecause they have well-defined differences in feeding, abundant of its respective guild( scrapers: E.pulchella,
locomotion, and gas exchange mechanisms. Resource suspension- feeders: Ae. triseriatus, deposit- feeders: C.
partitioning, however, does not seem to be complete, guttipennis).
and potential interactions might exist within and between
guilds. Concurrence and Coexistence
The majority ofencounters between pairs ofspeciesPotential Interactions was independent, leading to the generalization that
Within the guild ofmosquitoes exists much potential species in this community encountered each other with
overlap because they feed by concentrating particles. a frequency that was proportional to the abundance ofHowever, Aedes triseriatus can also scrape micro- the encountered species. The impact of species with
organisms ( Fish and Carpenter 1982), and leaf cells high frequency and abundance on the less frequentpers. observ.). In laboratory microcosms, Ae. triseriatus species would depend on whether the abundant species
has been shown to be a superior competitor to Or. actually saturate the habitat and its resources and on thesignifera, which is mainly a suspension- feeder degree of overlap in resource use. Aedes triseriatusChambers 1985). The ability ofAedes spp. to use food saturated the habitat at this study site ( Barrera, in
resources normally not available for other tree- hole preparation), and in other sites at similar latitudes( Fish
mosquitoes may confer them competitive advantage and Carpenter 1982; Livdahl 1982, Carpenter 1983).
under conditions of scarcity of suspended particles. Intraspecific effects also exist in the scirtid E.pulchella,
Within the guild of scrapers, scirtids predominate, and to a lesser extent in C. guttipennis ( Barrera, in
and E. pulchella and P. discoideus might overlap preparation). Several lines of evidence indicate that
extensively. Within the guild of deposit feeders, the many tree holes in a given area do not receive muchoverlap is less clear because of uncertainty about what litter. Fashing ( 1975) noted that much of the litterfood is used by each species. The sediment of tree holes accumulated during the previous fall was exhausted incontains leaf skeletons, feces, chitin, remains of bark, natural tree holes by mid-summer. Fish( pers. commun.;and microorganisms. Culicoides guttipennis seems to Fish and Carpenter 1982) estimated the mean amount of
feed actively on microorganisms( Williams and Turner litter entering tree holes in Indiana and reported a mean1976), but also on other insects, probably through ofabout 1 gram per tree hole per season, which has been
facultative predation( Hair and Turner 1966, Clark and demonstrated to produce density-dependent effects inFukuda 1967), and on dead insects( pers. observ.). Ae. triseriatus in simulated tree holes( Fish and Carpenter
Interactions between members of different guilds 1982). Additionally, in my study area 60% of the tree
may come also from overlap in food use, probably holes contained little or no litter because the diameter

June, 1996 Journal of Vector Ecology 77
and angle of the cavities prevented the accumulation of however, may have a positive impact on other species ofleaves ( Barrera, in preparation). If exploitative mosquitoes that remain active as larvae, because oftheircompetition exists between infrequent and frequent expected overlap in food use, but also because foodspecies, niche or habitat partitioning should promote resources may become scarcer in the summer. This typecoexistence in this community. Currently, there is a of temporal segregation would contribute to thewealth of evidence showing intra- and inter-tree- hole maintenance of a larger pool of mosquito species inpartitioning among tree- hole mosquitoes ( Bradshaw the community if resource depletion or interspecificand Holzapfel 1983, 1985, 1986, 1988, 1991; Lounibos competition limit the other species.
1981, 1983, 1985; Copeland and Craig 1990).The data also showed that a small fraction of the Community Structure
frequent species is exposed to the less frequent species. The profile of relative frequencies of the mostLet us consider the strong negative effect of the predator common species in the tree-hole community showed theTx. rutilus on the frequent species. Given the low numerical dominance of the mosquito Ae. triseriatusoccurrence and density of this predator( TABLE 2) only ( 43%), the ceratopogonid C. guttipennis( 15%), and the
a fraction of the frequent species ( proportional to the scirtid E. pulchella ( 37%). This numeric structure ispercentage of occurrence of Tx. rutilus) will be exposed similar to that observed by D. Fish( pers. commun.) into predation. Since most other infrequent species in the Indiana, USA( 41° 40' N; 86° 15' W) where Ae. triseriatus
community are not predators, their effects on the frequent was the most abundant species( 41%), shortly followedspecies should be less important than that of Tx. rutilus. by the scirtid Elodes sp. ( 35%), and the ceratopogonid
Therefore, the populations of the frequent species are C. guttipennis ( 22%). The similarity in the numericrelatively protected from significant interactions with structure of the tree- hole insect communities in central
the less frequent species. For Tx. rutilus, freezing, Pennsylvania and Indiana can most simply be explainedcannibalism, and the scarcity of large and permanent by their geographical proximity and their likely similartree holes may all be important reasons why this predator species pool. From data on adult emergence from treeis so scarce at this latitude ( Bradshaw and Holzapfel holes in northern California (Woodward et al. 1988),1985). Less is known about the factors that keep other Aedes sierrensis was the most common species( 82%),rare aquatic insects at such low frequencies ofoccurrence followed by chironomids ( 8%), Culicoides spp. ( 7%),in the study area. Lastly, the rare species encounter and scirtids ( 1%). The data available for Wytham
other rare species uncommonly; but when they do, their Woods in England( Kitching 1986) show that mosquitoespotential interactions may be significant because a ( Aedes) made up only 1% of the community, scirtidssignificant fraction of their populations overlap. If the ( Prionocyphon) 2%, and ceratopogonids ( Dasyhelea)
interactions involve strong negative effects, such as 18%. This community was dominated by a chironomidpredation, these low frequency populations would face ( MetriocnemusmartiniiThien.) with arelative frequencyimportant local reductions in population size. of 78%. Based on the available data on intraspecific
The pattern of interspecific encounters in this crowding, the insect community in Blacknest, England,community is highly predictable because of the was also dominated by the chironomid M. martinii. Theprevalence of null associations between species, and as data for Lamington Park in Australia( Kitching 1986)the data showed, it is frequency- dependent. Species show Aedes mosquitoes with a relative frequency ofhave asymmetrical potential interactions with the 3%, and scirtids with 74%. It seems, therefore, that the
frequent species being at a disproportionate advantage structures of aquatic communities in different biogeo-
given their high frequencies and the discrete nature of graphical regions do not converge. It is probably impor-the tree-hole habitat. tant to consider the different sampling methods, the
biology ofthe component species and their local environ-Time Partitioning ment, and the role of component species in determining
Changes in the relative frequency of the most the structure of the community. The recent study ofcommon tree-hole insects( Fig. 3) showed two distinct Copeland and Craig( 1990), who made special emphasistrends, with Aedes decreasing in numbers toward in sampling elevated tree holes in Indiana, showed thatsummer, and the opposite tendency of scirtids to some species otherwise rare, such as Orthopodomyia
accumulate in time. This separation of phenology alba Baker were rather common in protected tree holes
between Aedes and scirtids seems to result from their high in the canopy(> 2 m). Therefore, to obtain fair corn-
different biologies, in particular, their ways of facing the parisons ofcommunity structure, future studies may de-prolonged winter at this latitude. Disappearance ofAe. termine the availability of tree-hole types at differenttriseriatus larvae from tree holes during late summer, heights and use a standardized sampling methodology.

78 Journal of Vector Ecology June, 1996
Structure of the Guild of Mosquitoes in Eastern Lastly, the impact of tree- hole desiccation in the studyNorth America area is less frequent than in Florida ( Bradshaw and
The species composition of the guild ofmosquitoes Holzapfel 1983, Lounibos 1983), but it should continue
in central Pennsylvania was similar to that of northern to exert a negative impact on mosquito species which do
Florida ( Bradshaw and Holzapfel 1983), with the not possess eggs resistant to desiccation. The mecha-
exception of Corethrella appendiculata Grabham, nisms that promote habitat partitioning among tree-holewhich was not present in this study because it is a mosquitoes in Florida( predation, desiccation, tree-hole
neotropical species ( Lounibos 1983). The structure of characteristics) exist in central Pennsylvania, but they
the guild oftree-hole mosquitoes in central Pennsylvania do not appear to affect the dominance ofAe. triseriatus.
is, however, different from that of northern Floridabecause Ae. triseriatus was extremely common and the Acknowledgments
other mosquito species were rare. Predation by Tx.rutilus on Ae. triseriatus must be unimportant because I thank Drs. D. L. Pearson, J. H. Frank, D. Fish, W.
the predator occurs at low density, but also because most E. Bradshaw, and L. P. Lounibos forvaluable suggestions
individuals of Ae. triseriatus disappear from the tree and commentaries. I acknowledge Dr. L. Knutson,
holes after August due to arrested hatching at the time Biosystematics and Beneficial Insects Institute, U.S.
when most eggs of the predator were observed. Department of Agriculture, who facilitated the
Therefore, the larval development of Tx. rutilus from identification of tree-hole insects, and the entomologists
September until the freezing of the tree holes must not who identified the organisms: Drs. W. L. Grogan, Jr.
rely on Ae. triseriatus as an important source of food. ( Ceratopogonidae), S. S. Roback( Chironomidae), N. J.
The other mosquitoes, such as An. barberi, Or. Fashing ( Acarina), R. J. Gagne ( Muscidae), R. V.
signifera, and Cx. restuans were predominantly observed Peterson( Psychodidae), and J. H. Frank( Staphylinidae).
as larvae afterJuly, probably as aresultofan amplification This study was possible thanks to a fellowship from theof the population bytheovipositionoffemalessurviving Consejo de Desarrollo Cientifico y Humanisticoto the summer generation. Nonetheless, these mosquitoes ( CDCH), Universidad Central de Venezuela.
occurred in low numbers in a few large and permanent
tree holes with Tx. rutilus. The decline in Ae. triseriatus REFERENCES CITED
larvae in late Summer( Fig. 3) should increase the risksof predation by Tx. rutilus on the other mosquito species. BMDP. 1982. BMDP Statistical Software, University
The structure of the mosquito guild at the study of California Press, Berkeley, California.sites is similar to that reported by Mitchell and Rockett Bradshaw, W. E. and C. M. Holzapfel. 1983. Predator-
1981) in Ohio( 41° 30' N; 83° 30' W). They observed mediated, non-equilibrium coexistence of tree-hole
that Ae. triseriatus was the most abundant and frequent mosquitoes in southeastern North America.
mosquito species, and Or. signifera and An. barberi Oecologia 57: 239- 256.
became more frequent in late summer when they Bradshaw, W. E. and C. M. Holzapfel. 1985. The
occupied a few large tree holes. Toxorhynchites rutilus distribution and abundance of tree-hole mosquitoes
was not reported as present in the Ohio locality, and it is in eastern North America: perspectives from north
also absent in Indiana( Fish,pers. comm.). The similarity Florida. Pp. 3- 23 in Ecology of Mosquitoesin mosquito guild structure in Ohio and Pennsylvania, Lounibos, L. P., J. R. Rey, and J. H. Frank, eds.).and of the insect community in Indiana where Tx. rutilus Florida Medical Entomology Laboratory. Verois absent, suggests that this predator has little or no Beach, FL, USA, 579 pp.influence on the structure of the tree-hole community. Bradshaw, W. E. and C. M. Holzapfel. 1986. Habitat
The low abundance of Tx. rutilus, Or. signifera, segregation among European tree-hole mosquitoes.and An. barberi in the study area is probably due in part Nat. Geogr. Res. 2: 167- 178.
to freezing ( Bradshaw and Holzapfel 1985); but also Bradshaw, W. E. and C. M. Holzapfel. 1988. Drought
these mosquito species must encounter large numbers and the organization of tree- hole mosquito
ofAe. triseriatus in spring after ice-melting, which may communities. Oecologia 74: 507- 514.
have a negative impact on the other mosquitoes. Bradshaw, W. E. and C. M. Holzapfel. 1991. Fitness and
However, the densities of Or. signifera and An. barberi habitat segregation ofBritish tree-hole mosquitoes.
may have been subestimated if these species were as Ecol. Entomol. 16: 133- 144.
abundant in central Pennsylvania as in Indiana, where Bradshaw, W. E. and C. M. Holzapfel. 1992. Resource
Copeland and Craig( 1990) found that these species and limitation, habitat segregation, and species
Or.alba were more common in elevated, deep rot holes. interactions of British tree- hole mosquitoes in

June, 1996 Journal of Vector Ecology 79
nature. Oecologia 90: 227- 237. Communities. Plexus Publishing Inc., Medford,Breland, O. P. 1960. Restoration of the name, Aedes NJ, USA, 293 pp.
hendersoni Cockerell, and its elevation to full Grimstad, P. R., C. E. Garry, and G. R. DeFoliart. 1974.specific rank( Diptera: Culicidae). Ann. Entomol. Aedes hendersoni and Aedes triseriatus ( Diptera:Soc. Am. 53: 600-606. Culicidae) in Wisconsin: Characterization of larvae,
Carpenter, S. R. 1983. Resource limitation of larval larval hybrids, and comparison of adult and hybridtree-hole mosquitoes subsisting on beech detritus. mesoscutal patterns. Ann. Entomol. Soc. Amer.
Ecology 64: 219- 223. 67: 795- 804.Chambers, R. C. 1985. Competition and predation Grubb, P. J. 1986. Problems posed by sparse and
among larvae of three species of tree- hole breeding patchily distributed species in species- rich plantmosquitoes. Pp. 25- 53 in Ecology of Mosquitoes communities. Pg. 207- 225 in Community Ecology.Lounibos, L. P., J. R. Rey, and J. H. Frank, eds.). Diamond, J. and T. J. Case, eds). Harper and Row
Florida Medical Entomology Laboratory. Vero Publ., NY, USA, 665 pp.Beach, FL, USA, 579 pp. Hair, J. A. and E. C. Turner. 1966. Laboratory
Clark, T. B. and T.Fukuda. 1967. Predation of Culicoides colonization and mass-production procedures forcavaticus Wirth and Jones larvae on Aedes sierrensis Culicoides guttipennis. Mosq. News 26: 429- 433.Ludlow). Mosq. News 27: 424. Hair, J. A., E. C. Turner, and D. H. Messersmith. 1966.
Copeland, R. S. and G. B. Craig. 1990. Habitat Larval habitats of some Virginia Culicoidessegregation among tree-hole mosquitoes( Diptera: Diptera: Ceratopogonidae). Mosq. News 26: 195-Culicidae) in the Great Lakes region of the United 204.
States. Ann. Entomol. Soc. Am. 83: 1063- 1073. Hui- lin, L. 1972. Trees of Pennsylvania, the Atlantic
Dyar, H. G. 1928. The mosquitoes of the Americas. states and the Lake states. University ofWashington D.C.: Carnegie Institute Washington. Pennsylvania Press, PA, USA, 276 pp.Publ. No. 387: 1- 616. Jenkins, D. W. and S. J. Carpenter. 1946. Ecology of the
Fashing, N. J. 1975. Life history and general biology of tree hole breeding mosquitoes of Nearctic NorthNaiadacarus arboricola Fashing, a mite inhabiting America. Ecol. Monog. 16: 31- 47.water- filled tree holes( Acarina: Acaridae). J. Nat. Kitching, R. L. 1971. An ecological study of water-Hist. 9: 413- 424. filled tree-holes and their position in the woodland
Fashing, N. J. 1976. The evolutionary modification of ecosystem. J. Anim. Ecol. 40: 281- 302.
dispersal in Naiadacarus arboricola Fashing, a Kitching, R. L. 1983. Community structure in water-mite restricted to water-filled tree holes( Acarina: filled tree holes in Europe and Australia -
Acaridae). Amer. Midl. Nat. 95: 337- 346. Comparisons and speculations. Pg. 205- 222 inFish, D. 1983. Phytotelmata: Flora and Fauna. Pg. 1- 27 Phytotelmata: Terrestrial Plants as Hosts forAquatic
in Phytotelmata: Terrestrial Plants as Hosts for Insect Communities. ( Frank, J. H. and L. P.
Aquatic Insect Communities.( Frank, J. H. and L. P. Lounibos, eds.). Plexus Publishing Inc., Medford,Lounibos, eds.). Plexus Publishing Inc., Medford, NJ, USA, 293 pp.NJ, USA, 293 pp. Kitching, R. L. 1986. Prey- predator interactions in
Fish, D. and S. R. Carpenter. 1982. Leaf litter and larval Community Ecology: Pattern and process
mosquito dynamics in tree- hole ecosystems. Kikkawa, J. and D. J. Anderson, eds). Blackwell
Ecology 63: 283- 288. Scientific Publications. Carlton, Australia, 432 pp.Foote, R. H. and H. D. Pratt. 1954. The Culicoides of the Kitching, R. L. and C. Callaghan. 1982. The fauna of
eastern United States ( Diptera: Heleidae). Public water-filled tree holes in Box Forest in south- east
Health Monog. No. 18, Public Health Reports, U. S. Queensland. Australian Entomol. Magaz. 8: 61- 70.
Department of Health, Education, and Welfare, Kurihara, Y. and J. Kikkawa. 1986. Trophic relations of
Washington. decomposers. Pg. 91- 126 in Community Ecology:Frank, J. H. 1983. Bromeliad phytotelmata and their Pattern and process( Kikkawa, J. and D. J. Anderson,
biota, especially mosquitoes. Pg. 101- 128 in eds). Blackwell Scientific Publications. Carlton,
Phytotelmata:Terrestrial Plants as Hosts for Aquatic Australia, 432 pp.Insect Communities. ( Frank, J. H. and L. P. Livdahl, T. P. 1982. Competition within and betweenLounibos, eds.). Plexus Publishing Inc., Medford, hatching cohorts of a tree- hole mosquito. EcologyNJ, USA, 293 pp. 63: 1751- 1760.
Frank, J. H. and L. P. Lounibos. 1983. Phytotelmata: Lloyd, M. 1967. Mean crowding. J. Anim. Ecol. 36: 1-Terrestrial Plants as Hosts for Aquatic Insect 30.

80 Journal of Vector Ecology June, 1996
Lounibos, L. P. 1981. Habitat segregation among 126 in Community Ecology: Pattern and processAfrican tree- hole mosquitoes. Ecol. Entomol. 6: Kikawa, J. and D. J. Anderson, eds.). Blackwell
129- 154. Scientific Publications. Carlton, Australia, 432 pp.
Lounibos, L. P. 1983. The mosquito community of tree- Scholl, P. J. and G. R. DeFoliart. 1977. Aedes triseriatus
holes in subtropical Florida. Pg. 223- 246 in and Aedes hendersoni: vertical and temporal
Phytotelmata: Terrestrial Plants as Hosts forAquatic distribution as measured by oviposition. Env.Insect Communities. ( Frank, J. H. and L. P. Entomol. 6: 355- 358.
Lounibos, eds.). Plexus Publishing Inc., Medford, Sinsko, M. J. and P. R. Grimstad. 1977. Habitat
NJ, USA, 293 pp. separation by differential vertical oviposition ofLounibos, L. P. 1985. Interactions influencing produc- two tree hole Aedes in Indiana. Env. Entomol. 6:
tion of treehole mosquitoes in South Florida. Pp. 485- 487.
65- 67 in Ecology of Mosquitoes( Lounibos, L. P., Snow, W. E. 1949. The Arthropoda of wet tree holes.
J. R. Rey, and J. H. Frank, eds.). Florida Medical Ph. D. thesis. University of Illinois. Urbana, IL,
Entomology Laboratory. Vero Beach, FL, USA, USA.
579 pp. Terborgh, J. and S. Robinson. 1986. Guilds and their
Lunt, S. R. 1977. Morphological characteristics of the utility in ecology. Pg. 65- 90 in Community Ecology:larvae ofAedes triseriatus and Aedes hendersoni in Pattern and process( Kikawa, J. and D. J. Anderson
Nebraska. Mosq. News 37: 654- 656. eds.). Blackwell Scientific Publications. Carlton,
Maguire, B. 1971. Phytotelmata: Biota and community Australia., 432 pp.structure determination in plant-held waters. Ann. Thienemann, A. 1934. Die Tierwelt der tropischen
Rev. Ecol. Syst. 2: 439-464. Pflanzengewasser. Arch. Hydrobiol. Suppl. 13: 1-
Merritt, R. W. and K. W. Cummins. 1978. An 91.
introduction to the aquatic insects ofNorth America. Toft, C. A. 1985. Resource partitioning in amphibiansKendall/ Hunt, Dubuque, IA, 441 pp. and reptiles. Copeia 1985: 1- 21.
Mitchell, L. and C. L. Rockett. 1981. An investigation Truman, J. W. and G. B. Craig. 1968. Hybridizationon the larval habitat of five species of tree- hole between Aedes hendersoni and Aedes triseriatus.
breeding mosquitoes ( Diptera: Culicidae). Great Ann. Entomol. Soc. Am. 61: 1020- 1025.
Lakes Entomol. 14: 123- 129. Varga, L. 1928. Ein interessanter Biotop der BioconosePark, 0., S. Auerbach, and G. Corley. 1950. The tree- von Wasser-Organismen. Biologisches Zentralblatt
hole habitat with emphasis on the pselaphid beetle 48: 143- 162.
fauna. Bull. Chicago Acad. Sci. 9: 19- 57. Westerfeld, W.F. 1959. Flora ofCentre and Huntingdon
Pearson, D. L. 1986. Community structure and species Counties with related historical, geological, and
co-occurrence: a basis for developing broader physiographic features. Agriculture Experimental
generalizations. Oikos 46: 419- 423. Station, University Park, Pennsylvania. Bull. 647.Petersen, J. J., H. C. Chapman, and O. R. Willis. 1969. Williams, R. E. and E. C. Turner. 1976. An improved
Predation of Anopheles barberi Coquillett on first laboratory larval medium for Culicoides guttipennisinstar mosquito larvae. Mosq. News 29: 134- 135. Coq.) ( Diptera: Ceratopogonidae). J. Med.
Pope, R. D. 1976. Nomenclatural notes on the British Entomol. 13: 157- 161.
Scirtidae (= Helodidae) ( Col.). Entomol. Monthly Woodward, D. L., A. E. Colwell, and N. L. Anderson.
Magaz. 111: 186- 187. 1988. The aquatic insect community of tree holesRicklefs, R. E. 1987. Community diversity: Relative in northern California oak woodlands. Bull. Soc.
roles of local and regional processes. Science N.Y. Vector Ecol. 13: 221- 234.
235: 167- 171. Zavortink, T. J. 1968. Mosquito studies ( Diptera:
Rohnert, U. 1950. Wassererfullte Baumhohlen and ihre Culicidae). VIII. A prodrome of the genus
Besiedlung. Ein Beitrag zur Fauna Dendrolimnetica. Orthopodomyia. Contrib. Am. Entomol. Inst. 3: 1-
Arch. Hydrobiol. 44: 472- 516. 221.
Root, R. 1967. The niche exploitation pattern of the Zavortink, T. J. 1970. Mosquito studies ( Diptera:
blue- grey gnatcatcher. Ecol. Monog. 37: 317- 350. Culicidae). XIX. The tree hole Anopheles of the
SAS Institute 1987. SAS user' s guide: statistics. SAS New World. Contrib. Am. Entomol. Inst. 5: 1- 35.
Institute, Cary, North Carolina. Zavortink, T. J. 1972. Mosquito studies ( Diptera:
Schoener, T. W. 1974. Resource partitioning in Culicidae). XXVIII. The New World species
ecological communities. Science N.Y. 185: 27- 39. formerly placed in Aedes( Finlaya). Contrib. Am.
Schoener, T. W. 1986. Resource partitioning. Pg. 91- Entomol. Inst. 8: 1- 206.

Journal of Vector Ecology 21( 1): 81- 84
Prevalence of Ehrlichia chaffeensis, Spotted Fever GroupRickettsia, and Borrelia spp. Infections in Ticks and
Rodents at Fort Bragg, North Carolina
V. B. Solberg', J. G. Olson2, L. R. Boobar3,J. R. Burge', and P.G. Lawyer" 4
1Departments ofEntomology and Biometrics, Walter Reed ArmyInstitute ofResearch, Washington, DC 20307-5100
2Centers for Disease Control and Prevention, Atlanta, GA 303333Department ofEntomology, University ofMaine, Orono, ME 04469
Received 8 May 1995; Accepted 6 June 1995
ABSTRACT: The prevalence of Ehrlichia chaffeensis, spotted fever group rickettsia, and Borrelia spp.was determined in ticks that had been collected using two methods at Fort Bragg, North Carolina. Of 159tick midgut samples examined using direct fluorescent assays, 3% were positive for spotted fever groupRickettsiae and 5% for Borrelia spp. Sera from 1 Peromyscus leucopus( Gloger), 3 Mus musculus( L.), and
25 Sigmodon hispidus( Say and Ord) were negative for antibodies to Ehrlichia chaffeensis. Dry ice- baitedtick traps caught significantly more Ixodes scapularis( Say), A mblyomma americanum( L.),and Dermacentorvariabilis( Packard)( total of 158 ticks) compared to collections from rodent hosts( total of one tick).
Keyword Index: Ehrlichia, spotted fever group rickettsia, Borrelia, Ixodes scapularis, Dermacentorvariabilis, Amblyomma americanun, Sigmodon hispidus.
INTRODUCTION MATERIAL AND METHODS
A recent retrospective epidemiological study Fort Bragg Military Reservation, 55,000 ha, isrevealed that 38% of soldiers who had trained at Fort located in central North Carolina, USA. PreliminaryChaffee, Arkansas, exhibited serological evidence of trapping throughout the base indicated that smallrecent infections with spotted fever group rickettsiae mammals were scarce. Mice ( Peromyscus leucopusSFGR) and Ehrlichia chaffeensis ( Sanchez et al. [ Gloger] and Mus musculus [ L.]) were only found in
1992). Many of these soldiers had also trained at Fort areas where water was available; however, cotton ratsBragg, North Carolina. Infection prevalences for ticks ( Sigmodon hispidus [ Say and Ord]) were found bothat Fort Chaffee were 4. 8% for SFGR, 0. 1% for Borrelia around lakes and in fields. Five sites were selected thatspp., and 0. 3% for Ehrlichia ( Kardatzke et al. 1992); rodents inhabited. Two of the sites were field locationshowever, it was unknown if these organisms posed a where rye grass ( Secale cereale), catbrier ( Smilaxsimilar disease threat at Fort Bragg. This study was glauca), and immature longleaf pine trees ( Pinusconducted at Fort Bragg to determine the relative palustris) predominated. The other three sites wereabundance of vector tick species and their infection shoreline habitats surrounding 0.4 to 4,000 ha lakes,frequencies with Ehrlichia chaffeensis, SFGR, and where blueberry ( Vaccinuum spp.), red bay ( PerseaBorrelia spp. In addition, a small sample of rodents was borbonia), and bitter gallberry ( Ilex glabra)surveyed for the presence of antibodies to Ehrlichia predominated with an overstory mainly of longleaf pinechaffeensis. P. palustris) and turkey oak( Quercus laevis).
4The views of the authors do not purport to reflect the position of the U.S. Department of the Army or the U.S.Department of Defense.

82 Journal of Vector Ecology June, 1996
Sherman" live capture" box traps( H. B. Sherman Viral Disease, Atlanta, GA), and B. burgdorferi( obtained
Traps, Tallahassee, FL) or Mohawk small mammal from CDC, Division of Vector- Borne Infectious
traps ( Tomahawk Trap Company, Tomahawk, WI)5, Diseases, FortCollins, CO) were used as positive control
baited with both peanut butter mixed with bird seed and antigen slides. All immunostain reagents were negative
a slice of apple, were set in the late afternoon ( 1600- for cross- reactivity.
2000 hours) from March 23- 26, June 9- 12, and July 28- Indirect immunofluorescent assays ( IFA) were
31, 1992. Six to 12 traps were set each night at performed ( Dawson et al. 1990) on the 29 rodent
alternating sites and checked 12- 16 hours later. The sera to detect IgG antibody to E. chaffeensis.
location where each rodent was captured was marked Ehrlichia chaffeensis was cultivated in DH82 cells.
with a flag and considered as the center of a circular 7 m Serum samples were tested in two- fold dilutions
diameter test plot. beginning at 1: 64 using FITC conjugated goat anti-
Each trapped rodent was anesthetized, a 0. 5 ml rat IgG ([ H+L], Kirkegaard and Perry Laboratories,
blood sample was taken by retroorbital bleeding or heart Gaithersburg, MD) for Sigmodon hispidus sera orpuncture, and attached ticks were collected. After FITC-conjugated goat anti-mouse IgG ([ H+L],
recovery from anesthesia, the rodent was released at Biotech, Birmingham, AL). The criterion for a
the place of capture. Immediately after release of positive sample was the observation of specific
each rodent, four dry ice-baited tick traps, containing fluorescence in dilutions of 1: 128 or greater.
2. 7 kg of solid CO2 each ( Solberg et al. 1992), wereplaced equidistant on the circumference of a 7 m diameter RESULTS AND DISCUSSION
circle centered on each mammal trap site. Ticks captured
by the tick traps were removed 24 hours later, Of the 159 tick midgut samples tested by DFA, five
identified, and stored at or below ambient temperature ( 3%) were positive for SFGR and eight ( 5%) were
until assayed. positive for Borrelia spp. ( TABLE 1). All tick
Ticks were examined for Ehrlichia spp., SFGR, hemolymph samples were negative for Ehrlichia spp.
and Borrelia spp. using direct fluorescent anti-body All rodent sera were negative for antibodies to E.
DFA) assays( Kardatzke et al. 1992). Tick hemolymph chaffeensis.
and guts were collected from all live ticks and fixed on Significantly more ticks were collected using tick
slides according to the method described by Burgdorfer traps( n= 158) than the rodent host survey method( n= 1)
1970). The hemolymph was immuno- stained with a ( Wilcoxon signed rank test, P< 0. 001, n= 159; March:
mouse polyclonal, fluorescein- conjugated anti- P= 0.25, n= 8, Wilcoxon statistic= 34. 5; June: P= 0.014,
Ehrlichia canis serum( 1: 50, obtained from University n= 8, Wilcoxon statistic= 36; July: P= 0.009, n= 12,
of Maryland, Baltimore, MD) and counter-stained Wilcoxon statistic= 45.0). Most ticks caught by tick
with Evans blue or eriochrome black ( 1: 1000). Tick traps were Amblyomma americanum( n= 146), followed
gut samples were stained with a 1: 1 mixture of anti-B. by Dermacentorvariabilis( n= 9), and Ixodes scapularisburgdorferi antibodies to anti-Rickettsia rickettsii ( n= 3). Amblyomma americanum nymphs were more
antibodies. The anti-B. burgdorferi antibodies were numerous ( n= 95) than adults ( n= 50). Three adult I.
conjugated with fluorescein isothiocyanate ( FITC) at scapularis and eight adult and one nymphal D. variabilis
1: 50 dilution( Barbour et al. 1983) and the anti-Rickettsia were also collected. There were no significant differences
rickettsii antibodies were conjugated with FITC at a in the number of ticks collected at lake and field sites.
1: 10 dilution( University of Maryland), counterstained One nymphal D. variabilis collected from one cotton rat
as above, and examined for spirochetes and SFGR was negative for Ehrlichia spp., SFGR, and Borrelia
400x). Slides were considered positive for Borrelia spp.
spp. ifmore than one characteristic fluorescing spirochete A total of 1 Peromyscus leucopus, 3 Mus
was observed; for SFGR if characteristic fluorescing musculus, and 25 Sigmodon hispidus was captured at
rod- shaped bacteria were observed; and for Ehrlichia five test sites. The number of ticks caught parasitizing
spp. if fluorescing bacteria in morulas were observed small rodents at Fort Bragg( 1 D. variabilis nymph) was
in hemolymph cells. Cultured Rickettsia rickettsii much lower than that observed by Gage et al. ( 1992),
University of Maryland), E. chaffeensis ( Centers for who reported a mean of 0.7- 17. 4 larval and 0.44- 8. 29
Disease Control and Prevention [ CDC], Division of nymphal D. variabilis attached to cotton rats at similar
5Use of trade names is for identification only and does not imply endorsement by the Public Health Service, the U.S.Department of Health and Human Services, or the U.S. Army.
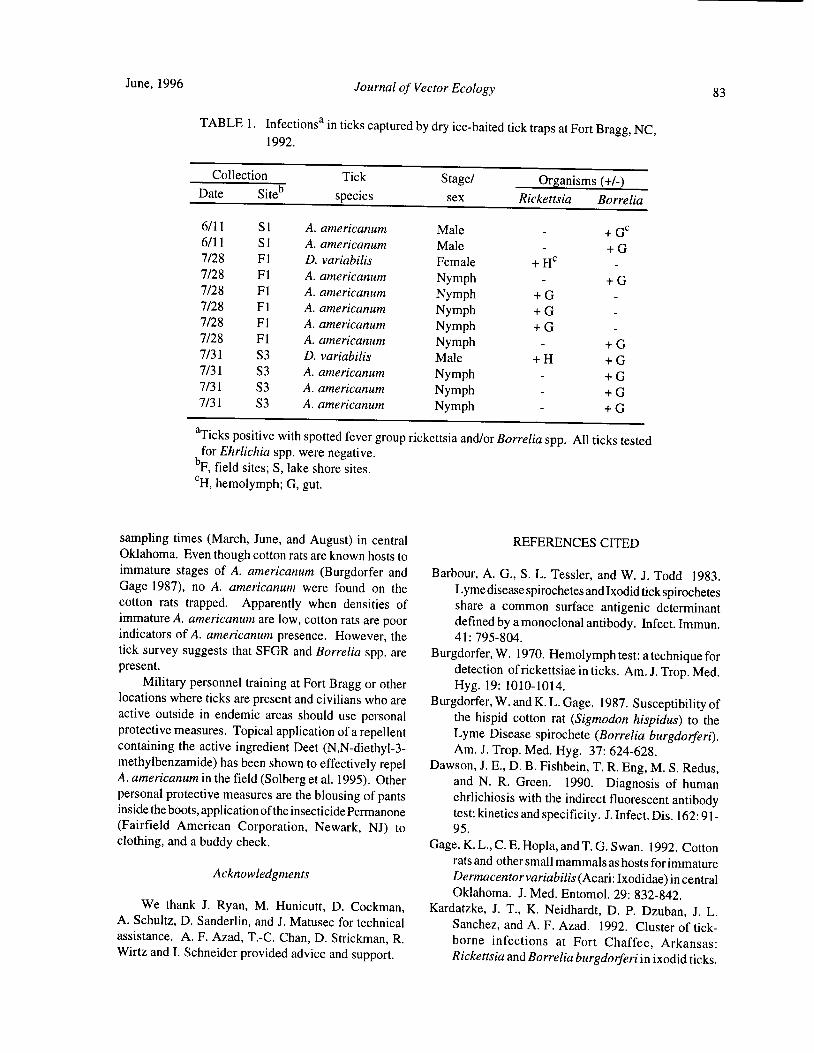
June, 1996 Journal of Vector Ecology 83
TABLE 1. Infectionsa in ticks captured by dry ice-baited tick traps at Fort Bragg, NC,1992.
Collection Tick Stage/ Organisms (+/-)Date
Siteb
species sex Rickettsia Borrelia
6/ 11 51 A. americanum Male G°6/ 11 S 1 A. americanum Male G7/ 28 Fl D. variabilis Female H` 7/ 28 Fl A. americanum Nymph G7/ 28 Fl A. americanum Nymph G7/ 28 Fl A. americanum Nymph G7/ 28 Fl A. americanum Nymph G7/ 28 Fl A. americanum Nymph G7/ 31 S3 D. variabilis Male H G7/ 31 S3 A. americanum Nymph G7/ 31 S3 A. americanum Nymph G7/ 31 S3 A. americanum Nymph G
aTicks positive with spotted fever group rickettsia and/ or Borrelia spp. All ticks testedfor Ehrlichia spp. were negative.
bF, field sites; S, lake shore sites.H, hemolymph; G, gut.
sampling times ( March, June, and August) in central REFERENCES CITEDOklahoma. Even though cotton rats are known hosts toimmature stages of A. americanum ( Burgdorfer and Barbour, A. G., S. L. Tessler, and W. J. Todd 1983.Gage 1987), no A. americanum were found on the Lyme disease spirochetes and Ixodid tick spirochetescotton rats trapped. Apparently when densities of share a common surface antigenic determinantimmature A. americanum are low, cotton rats are poor defined by a monoclonal antibody. Infect. Immun.indicators of A. americanum presence. However, the 41: 795- 804.tick survey suggests that SFGR and Borrelia spp. are Burgdorfer, W. 1970. Hemolymph test: a technique forpresent.
detection of rickettsiae in ticks. Am. J. Trop. Med.Military personnel training at Fort Bragg or other Hyg. 19: 1010- 1014.
locations where ticks are present and civilians who are Burgdorfer, W. and K. L. Gage. 1987. Susceptibility ofactive outside in endemic areas should use personal the hispid cotton rat ( Sigmodon hispidus) to theprotective measures. Topical application of a repellent Lyme Disease spirochete ( Borrelia burgdorferi).containing the active ingredient Deet ( N,N-diethyl- 3- Am. J. Trop. Med. Hyg. 37: 624-628.methylbenzamide) has been shown to effectively repel Dawson, J. E., D. B. Fishbein, T. R. Eng, M. S. Redus,A. americanum in the field( Solberg et al. 1995). Other and N. R. Green. 1990. Diagnosis of humanpersonal protective measures are the blousing of pants ehrlichiosis with the indirect fluorescent antibodyinside the boots, application of the insecticide Permanone test: kinetics and specificity. J. Infect. Dis. 162: 91-Fairfield American Corporation, Newark, NJ) to 95.
clothing, and a buddy check. Gage, K. L., C. E. Hopla, and T. G. Swan. 1992. Cottonrats and other small mammals as hosts for immature
Acknowledgments Dermacentorvariabilis( Acari: Ixodidae) in centralOklahoma. J. Med. Entomol. 29: 832- 842.
We thank J. Ryan, M. Hunicutt, D. Cockman, Kardatzke, J. T., K. Neidhardt, D. P. Dzuban, J. L.A. Schultz, D. Sanderlin, and J. Matusec for technical Sanchez, and A. F. Azad. 1992. Cluster of tick-assistance. A. F. Azad, T.-C. Chan, D. Strickman, R. borne infections at Fort Chaffee, Arkansas:Wirtz and I. Schneider provided advice and support. Rickettsia and Borrelia burgdorferi in ixodid ticks.

84 Journal of Vector Ecology June, 1996
J. Med. Entomol. 29: 669- 672. 1992. Quantitative evaluation ofsampling methods
Sanchez, J. L., W. H. Candler, D. B. Fishbien, C. R. for Ixodes dammini and Amblyomma americanum
Green, T. R. Cote, D. J. Kelly, D. P. Driggers, and Acari: Ixodidae). J. Med. Entomol. 29: 451- 456.
B. J. B. Johnson. 1992. A cluster of tick-borne Solberg, V. B., R. A. Wirtz, K. R. McPherson, B. A.infections: association with military training and Bradford, J. R. Burge, and T. A. Klein. 1995. Field
asymptomatic infections due to Rickettsia rickettsii. evaluation of Deet and a piperidine repellent, AI3-
Trans. R. Soc. Trop. Med. Hyg. 86: 321- 325. 37220, against the lone star tick, Amblyomma
Solberg, V. B., K. Neidhardt, M. R. Sardelis, J. americanum( Acari: Ixodidae). J. Med. Entomol.
Hildebrandt, F. J. Hoffmann, and L. R. Boobar. 32: 870- 875.

Journal of Vector Ecology 21( 1): 85- 88
The Residual Effects ofAlphacypermethrin and Permethrin
Against the Dengue Vector Aedes albopictus ( Skuse)in Wooden Huts in Malaysia
S. Sulaiman', Z. A. Pawanchee2, I. Ghauth', A. Wahab2B. Vadiveloo', J. Jeffery', and A. F. Mansor'
Department ofParasitology and Medical Entomology, Faculty ofMedicine,Universiti Kebangsaan Malaysia, Jalan Raja Muda Abdul Aziz,
50300 Kuala Lumpur, MALAYSIA
Vector Control Unit, Department ofHealth, Municipality ofKuala Lumpur,KM 4 Jalan Ceras 56100 Kuala Lumpur, MALAYSIA
Received 16 June 1995; Accepted 25 August 1995
ABSTRACT: The pyrethroids alphacypermethrin and permethrin were compared for residual activityagainst the dengue vector Aedes albopictus( Skuse) in wooden huts in Kuala Lumpur. Two huts wereindividually sprayed inside with one of the pyrethroids and the third hut served as a control. At an appliedconcentration of 0.01 g a. i./ m2, both pyrethroids were equally effective in causing more than 80%mortality ofAe. albopictus one week after spraying. The Ae. albopictus exposed to a higher concentrationof 0.05 g a. i./m2
on alphacypermethrin treated walls showed 100% mortality one week post- spraying thatdeclined to 91. 3% mortality eight weeks post-spraying. In contrast, the permethrin- treated walls caused87.5% mortality one week after spraying and 75% mortality eight weeks after spraying. Thus,
alphacypermethrin showed a greater residual efficacy than permethrin in controlling the dengue vector.
Keyword Index: Alphacypermethrin, Permethrin, Aedes albopictus, wooden huts.
INTRODUCTION represent a small percentage of the original larval
population. It was the purpose of this study to determineIn Southeast Asia, Aedes albopictus ( Skuse) has by field bioassays the, residual effectiveness of
been incriminated as a secondary vector ofdengue fever alphacypermethrin and permethrin on the dengue vectorand Aedes aegypti ( Linn.) as the principal vector of Ae. albopictus in wooden huts in Malaysia.dengue viruses ( Russell et al. 1969, Chan et al. 1971,Jumali et a1. 1979, Harinasuta 1984). From our previous MATERIALS AND METHODSstudy it was shown thatAe. albopictus was more dominantthan Ae. aegypti as a domestic mosquito species inside Three experimental huts each measuring 2. 93 mpremises ofhouses, shop houses, and factories in Bandar long, 2.29 m wide, and 2. 12 m high were placed in dif-Tun Razak, a satellite town ofKuala Lumpur( Sulaiman ferent areas of Kuala Lumpur city. Each hut was madeet al. 1991).
ofwood, wire gauze, and zinc roofand all were essentiallyPermethrin treated fabrics, such as clothing, bednets, identical in design. The first two huts were individuallyand other materials have been shown to be effective in sprayed with a specific insecticide, and the third hutprotecting against mosquito bites( Schreck et al. 1978, served as a control( sprayed with tap water only). TapSchreck 1991, Anonymous 1989). Alphacypermethrin water was the diluent for both insecticide formulations.has been shown to be effective against dengue vectors Suspension concentrate ( SC) formulation of alpha-under field conditions in Malaysia demonstrating both cypermethrin( Fendona, Shell Public Health, UK) andadulticidal and larvicidal effects( Sulaiman et al. 1995). emulsion concentrate( EC) ofpermethrin( Talcord, ShellThe advantage ofkilling adult dengue vectors instead of Public Health, UK) were diluted with tap water andexhaustive larval control is that the adult infected females applied with knapsack sprayer( 20 strokes per minute)
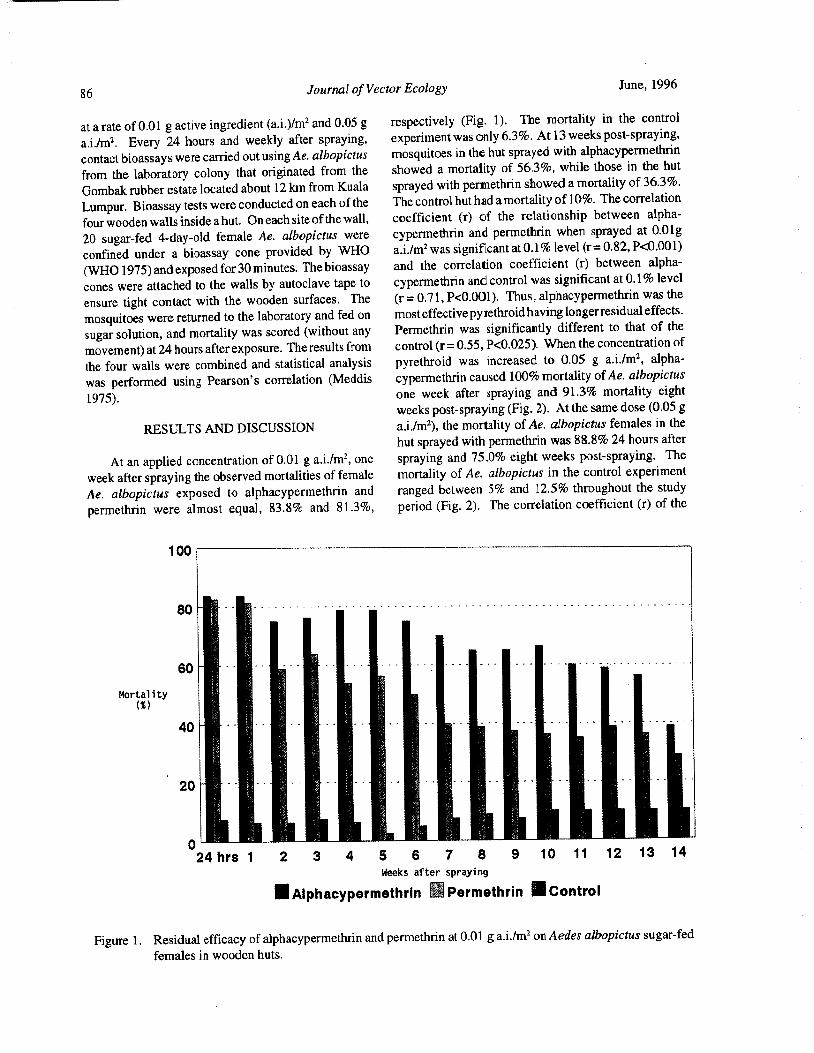
86 Journal of Vector Ecology June, 1996
at a rate of 0.01 g active ingredient( a. i.)/ m2
and 0.05 g respectively ( Fig. 1). The mortality in the control
a. i./m2. Every 24 hours and weekly after spraying, experiment was only 6.3%. At 13 weeks post-spraying,
contact bioassays were carried out using Ae. albopictus mosquitoes in the hut sprayed with alphacypermethrin
from the laboratory colony that originated from the showed a mortality of 56.3%, while those in the hut
Gombak rubber estate located about 12 km from Kuala sprayed with permethrin showed a mortality of 36.3%.
Lumpur. Bioassay tests were conducted on each of the The control hut had a mortality of 10%. The correlation
four wooden walls inside a hut. On each site of the wall, coefficient ( r) of the relationship between alpha-
2.0 sugar- fed 4- day- old female Ae. albopictus were cypermethrin and permethrin when sprayed at 0.Olg
confined under a bioassay cone provided by WHO a. i./ m2
was significant at 0. 1% level( r= 0.82, P< 0.001)
WHO 1975) and exposed for 30 minutes. The bioassay and the correlation coefficient ( r) between alpha-
cones were attached to the walls by autoclave tape to cypermethrin and control was significant at 0. 1% level
ensure tight contact with the wooden surfaces. The ( r= 0.71, P< 0.001). Thus, alphacypermethrin was the
mosquitoes were returned to the laboratory and fed on most effective pyrethroid having longer residual effects.
sugar solution, and mortality was scored( without any Permethrin was significantly different to that of the
movement) at 24 hours after exposure. The results from control( r= 0.55, P< 0.025). When the concentration of
the four walls were combined and statistical analysis pyrethroid was increased to 0.05 g a. i./m2, alpha-
was performed using Pearson' s correlation ( Meddis cypermethrin caused 100% mortality of Ae. albopictus
1975). one week after spraying and 91. 3% mortality eight
weeks post- spraying( Fig. 2). At the same dose( 0.05 g
RESULTS AND DISCUSSION a. i./ m2), the mortality of Ae. albopictus females in thehut sprayed with permethrin was 88. 8% 24 hours after
At an applied concentration of 0.01 g a. i./m2,
one spraying and 75. 0% eight weeks post-spraying. The
week after spraying the observed mortalities of female mortality of Ae. albopictus in the control experimentAe, albopictus exposed to alphacypermethrin and ranged between 5% and 12. 5% throughout the study
permethrin were almost equal, 83. 8% and 81. 3%, period ( Fig. 2). The correlation coefficient( r) of the
100
80 , - -
60
i iMortality
1%)
40
024 hrs 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Weeks after spraying
Alphacypermethrin Permethrin "' Control
Figure 1. Residual efficacy of alphacypermethrin and permethrin at 0.01 g a.i./ m2 on Aedes albopictus sugar- fedfemales in wooden huts.
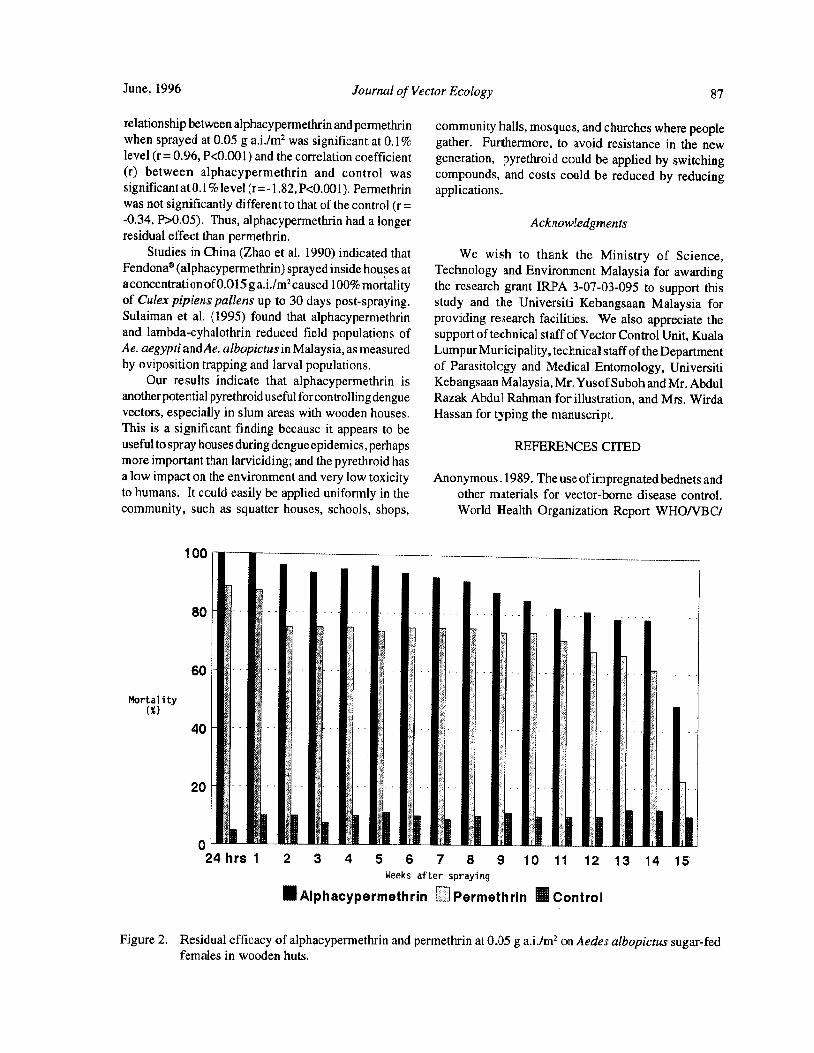
June, 1996 Journal of Vector Ecology 87
relationship between alphacypermethrin and permethrin community halls, mosques, and churches where peoplewhen sprayed at 0.05 g a. i./ m2
was significant at 0. 1% gather. Furthermore, to avoid resistance in the newlevel( r= 0.96, P< 0.001) and the correlation coefficient generation, pyrethroid could be applied by switchingr) between alphacypermethrin and control was compounds, and costs could be reduced by reducing
significant at 0. 1% level( r=- 1. 82, P< 0.001). Permethrin applications.
was not significantly different to that of the control( r=0.34, P> 0.05). Thus, alphacypermethrin had a longer Acknowledgments
residual effect than permethrin.
Studies in China( Zhao et al. 1990) indicated that We wish to thank the Ministry of Science,Fendona®(alphacypermethrin) sprayed inside houses at Technology and Environment Malaysia for awardingaconcentrationof0.015ga.i./m2caused 100% mortality the research grant IRPA 3- 07- 03- 095 to support thisof Culex pipiens pallens up to 30 days post-spraying. study and the Universiti Kebangsaan Malaysia forSulaiman et al. ( 1995) found that alphacypermethrin providing research facilities. We also appreciate theand lambda-cyhalothrin reduced field populations of support of technical staff of Vector Control Unit, KualaAe. aegypti and Ae. albopictus in Malaysia, as measured Lumpur Municipality, technical staffof the Departmentby oviposition trapping and larval populations. of Parasitology and Medical Entomology, Universiti
Our results indicate that alphacypermethrin is Kebangsaan Malaysia, Mr.YusofSuboh and Mr. Abdulanotherpotentialpyrethroidusefulforcontrollingdengue Razak Abdul Rahman for illustration, and Mrs. Wirdavectors, especially in slum areas with wooden houses. Hassan for typing the manuscript.This is a significant finding because it appears to beuseful to spray houses during dengue epidemics, perhaps REFERENCES CITED
more important than larviciding; and the pyrethroid hasa low impact on the environment and very low toxicity Anonymous. 1989. The use of impregnated bednets andto humans. It could easily be applied uniformly in the other materials for vector-borne disease control.community, such as squatter houses, schools, shops, World Health Organization Report WHONBC/
100
80 - - -
t
4t'
60 . . . . FMortality
40
20
0I I L t 1 1 I.
24 hrs 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Weeks after spraying
UAlphacypermethrin E Permethrin Control
Figure 2. Residual efficacy of alphacypermethrin and permethrin at 0. 05 g a. i./ m2 on Aedes albopictus sugar- fedfemales in wooden huts.

88 Journal of Vector Ecology June, 1996
89.981. 71: 397- 400.
Chan, Y. C., B. C. Ho, and K. L. Chan. 1971. Aedes Schreck, C. E. 1991. Permethrin and dimethyl phthalate
aegypti ( L.) and Aedes albopictus ( Skuse) in as tent fabric treatments against Aedes aegypti. J.
Singapore City. 5. Observations in relation to Am. Mosq. Contr. Assoc. 7: 533- 535.dengue haemorrhagic fever. Bull WHO 44: 651- Sulaiman, S., Z.A. Pawanchee, J. Jeffery, I. Ghauth, and658. V. Busparani. 1991. Studies on the distribution and
Harinasuta, C. 1984. Mosquito-borne diseases in abundance of Aedes aegypti ( L.) and Aedes
Southeast Asia. Mosq. Borne Dis. Bull. 1: 1- 11. albopictus ( Skuse) ( Diptera: Culicidae) in an
Jumali, S., D. J. Gubler, S. Nalim, S. Eram, and J. S. endemic area ofdengue/dengue haemorrhagic fever
Saroso. 1979. Endemic dengue haemorrhagic fever in Kuala Lumpur. Mosq. Borne Dis. Bull. 8: 35- 39.in rural Indonesia III. Entomological studies. Am. Sulaiman, S., M. A. Karim, B. Omar, and S. Omar.
J. Trop. Med. Hyg. 28: 717- 724. 1995. Field evaluation of alphacypermethrin and
Meddis, R. 1975. Statistical handbook for non- lambda-cyhalothrin againstAedesaegyptiandAedes
statisticians. McGraw-Hill Book Company ( UK) albopictus in Malaysia. J. Am. Mosq. Contr. Assoc.Ltd., 162 pp. 11: 54- 58.
Russell, P. K., D. J. Gould, T. M. Yuill, A. Nisalak, and WHO. 1975. Manual on practical entomology inmalariaP. E. Winter. 1969. Recovery of dengue-4 viruses Part II. World Health Organization, Geneva, 191
from mosquito vectors and patients during an pp.
epidemic of dengue haemorrhagic fever. Am. J. Zhao Zuezhong, Jiang Zhikuan, and Zhong Yingkuo.Trop. Med. Hyg. 18: 580-583. 1990. Observations on the efficacy of Fendona
Schreck, C. E., K. Posey, and D. Smith. 1978. Durability against mosquitoes and flies. Pest Control, vol. 6
of permethrin as a clothing treatment to protect No. 1, 1990 ( supplement). Assoc. Vector Biol.
against blood- feeding arthropods. J. Econ. Entomol. Contr. China Assoc. Prevent. Med. 6( 1): 75- 79.

Journal of Vector Ecology 21( 1): 89- 93
Distribution of Lambornella stegomyiae in Malaysia and its Potentialfor the Control of Mosquitoes of Public Health Importance
I. Vythilingam, S. Mahadevan, K. K. Ong, Abdullah Ghani, and Y. F. Ong
Division ofMedical Entomology, Institute for Medical Research,Jalan Pahang, 50588 Kuala Lumpur, MALAYSIA
Received 9 July 1995; Accepted 12 January 1996
ABSTRACT: Very little work has been conducted on the parasitic ciliate, Lambornella stegomyiae, sinceit was first found in Aedes albopictus larvae breeding in an earthenware pot in Kajang, Malaysia, in 1920.The present study was carried out to determine the natural distribution of this organism in PeninsularMalaysia and to study its potential as a biological control agent. The central zone had the highest percentage
of larvae infected with L. stegomyiae, followed closely by the northwestern zone. The only state that didnot yield any infected larvae was Trengganu. Most of the L. stegomyiae were found in rubber cups.Lambornella stegomyiae were found mostly in Ae. albopictus and in Armigeres spp. Laboratoryexperiments showed that L. stegomyiae did not show potential as a good biological control agent. Theparasite was only able to infect first-instar larvae and it was not possible to culture the parasites in vitro.
Keyword Index: Lambornella stegomyiae, Aedes albopictus, rubber cups, distribution.
INTRODUCTION the mosquito larva in the form of a round transparent
structure. The ciliate then gains entry into the hemocoelThe ciliated protozoan, Lambornella stegomyiae, and rapid multiplication takes place which causes death
is a facultative parasite belonging to the family to the larva. The cuticle of the larva ruptures and theTetrahymenidae. Lambornella stegomyiae was first ciliates escape into the water and are able to infect newfound in a batch of 29 Aedes albopictus larvae breeding larvae.
in an earthenware pot in Kajang, Malaysia, in 1920 by Although it appears that Lambornella could be aLamborn ( Lamborn 1921). The parasitic ciliate, potential biological control agent, very little work hasLambornella, has been reported from widespread been carried out in this area. Lambornella stegomyiae
geographical areas, such as Africa ( Muspratt 1945, was first found in Malaysia in the early twenties, but it1947), Europe( MacArthur 1922), Southeast Asia( Laird was not until 1990 that it was reported again in Penang,1959, Corliss 1961), South Korea ( Yu et al. 1978), Malaysia ( Sulaiman, unpublished data). With this
USSR( Dzerhinsky et al. 1976), and USA( Corliss 1954, paucity ofdata we decided to survey Peninsular MalaysiaChapman 1974, Clark and Brandl 1976). Although to determine the natural distribution of this organismmost of the infections were found inAedes species, there and to study its feasibility as a biological control agent.have been reports of its breeding in culicine species ofArmigeres ( Corliss 1961), Culiseta( MacArthur 1922) MATERIALS AND METHODS
and Wyeomyia( Corliss 1954). It has also been reported
in Anopheles( Yu et al. 1978). Larval Surveys
In addition to L. stegomyiae, Lambornella clarki, a Larval surveys were carried out in all states of
species that has been found in Aedes sierrensis breeding Peninsular Malaysia with the exception of Penang.in tree holes in California( Clark and Brandl 1976), is the Surveys were carried out over a period of two years from
only other Lambornella species known. The life cycle 1992 to 1993. Each state was surveyed at different timesof the free living forms and the resting cyst stages ofL. of the year. All types of artificial containers, bamboostegomyiae has not been extensively studied. However, stumps, and tree holes were examined for larvae both in
it is known that the cyst attaches itself on the surface of urban and rural areas. Larvae from each container were

90 Journal of Vector Ecology June, 1996
collected into a plastic collection bottle and brought in the various states of Malaysia. The central zone had
back to the laboratory for study. the highest percentage positive( 6. 6), followed closelyLarval collections were examined in the laboratory by the northwestern zone. However, it must be noted
under a compound microscope( 40X) for infections. All that L. stegomyiae was present in all states in Peninsular
larvae that were positive for L. stegomyiae were Malaysia with the exception of Trengganu. Besides
transferred into small containers and reared in the Lambornella, the fungal pathogen Coelomomyces was
laboratory. also present. This will be reported elsewhere.
The different types of containers examined are
In vivo Culture shown in TABLE 2. Most of the L. stegomyiae were
Lambornella stegomyiae that were obtained from found in rubber cups; a total of 2,506 rubber cups was
field collected larvae were maintained in vivo in examined, and of these 130 were positive( 5%). Onlylaboratory colonized Aedes albopictus larvae. When two breedings were found in other artificial containers
the larva became fully infected with the endoparasitic and one in a coconut shell. None were found in tree
forms, the larval cuticle was punctured to release the holes. From the 2, 896 collections, 112 Ae. albopictus,
parasites. Larval mortality naturally occurs between 29 Armigeres spp., and one Anopheles subpictus werefour and six days. The parasites were then transferred to positive. Most of the Armigeres collections were found
a new container and after four hours newly hatched first- in Selangor.
instar larvae were added. Larval food was added and the TABLE 3 shows the different species of larvae that
larvae were maintained as before at room temperature. were examined and found positive for L. stegomyiae.
Water obtained from the containers in the field were Aedes albopictus was the predominant species infected,
autoclaved and mixed with distilled water. This water followed by Annigeres spp. Several Toxorynchites
was used for maintaining the larvae throughout our larvae and 1, 367 Culex larvae were also examined, but
study. none were positive.
Laboratory infection studies carried out on An.Susceptibility ofLambornella stegomyiaetoMosquito maculatus and Cx. quinquefasciatus with L. stegomyiae
Larvae were not successful. The larvae were not infected by L.Four species of mosquitoes were tested for their stegomyiae. TABLE 4 shows the results of exposure of
susceptibility in the laboratory: Aedes albopictus, Aedes Ae. albopictus and Ae. aegypti to L. stegomyiae. Aedes
aegypti, Anopheles maculatus, and Culex quinque- albopictus and Ae. aegypti larvae appeared to be
fasciatus. Two fourth- instar larvae, which were full uninfected by L. stegomyiae. On days 1 and 2
with L. stegomyiae, were teased apart and their contents postexposure the mean was less than 1, and on day 7 thetransferred to 200 ml ofautoclaved water. The container mean was 13. 8 for Ae. albopictus and 3. 1 for Ae.
was then placed on a magnetic stirrer and 10 ml of this aegypti. It was also observed that many of the larvaesolution was then added to 20 containers containing 100 were not infected.
ml of distilled water each. This was left aside for four
to six hours at room temperature. Then, 20- 25 first- DISCUSSION
instar larvae were added to each container. Food was
provided 24 hours later. Most L. stegomyiae were found in Ae. albopictus
Approximately 20 larvae were examined at 24, 48, inhabiting rubber cups. A large number of rubber cupsand 72 hours postexposure and thereafter every day was surveyed compared to other containers because in
until day 7. Larvae that were examined at 24 and 48 our initial surveys larvae from many other types ofhours postexposure were stained with amido black dye, artificial containers were not infected with L. stegomyiae.
which killed the larvae and stained the cyst blue. The Hence, most efforts in collecting were concentrated innumber of cysts on the larvae were counted. Larvae rubber estates. Our findings suggest that L. stegomyiae
examined on day 3 postexposure and thereafter were not will survive only in undisturbed habitats. It was foundstained but the number of L. stegomyiae present was more in rural areas than urban areas. Aedes albopictus
counted. is also more rural in its habitat compared to Ae. aegypti
Sulaiman and Jeffrey 1986). This may explain why itRESULTS was found in Ae. albopictus and not in Ae. aegypti.
Lambornella stegomyiae was also found in Ae.
A total of 2,896 containers was examined and of albopictus in larger numbers compared to other container
these 133 ( 4. 6%) were found to be positive for L. breeding mosquitoes. In some containers there werestegomyiae. TABLE 1 shows the collections carried out mixed populations of Ae. albopictus, Annigeres spp.,
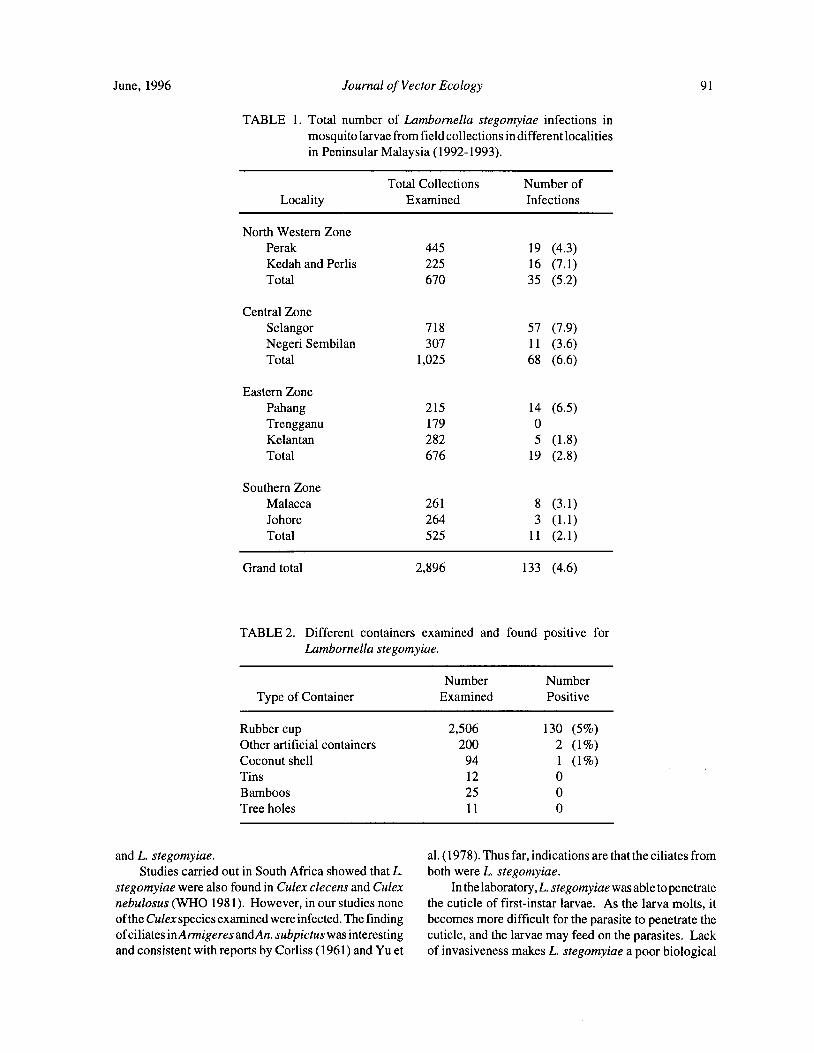
June, 1996 Journal of Vector Ecology 91
TABLE 1. Total number of Lambornella stegomyiae infections in
mosquito larvae from field collections in different localities
in Peninsular Malaysia( 1992- 1993).
Total Collections Number of
Locality Examined Infections
North Western Zone
Perak 445 19 ( 4.3)
Kedah and Perlis 225 16 ( 7. 1)
Total 670 35 ( 5. 2)
Central Zone
Selangor 718 57 ( 7. 9)
Negeri Sembilan 307 11 ( 3. 6)
Total 1, 025 68 ( 6.6)
Eastern Zone
Pahang 215 14 ( 6.5)
Trengganu 179 0
Kelantan 282 5 ( 1. 8)
Total 676 19 ( 2. 8)
Southern Zone
Malacca 261 8 ( 3. 1)
Johore 264 3 ( 1. 1)
Total 525 11 ( 2. 1)
Grand total 2, 896 133 ( 4.6)
TABLE 2. Different containers examined and found positive for
Lambornella stegomyiae.
Number Number
Type of Container Examined Positive
Rubber cup 2, 506 130 ( 5%)
Other artificial containers 200 2 ( 1%)
Coconut shell 94 1 ( 1%)
Tins 12 0
Bamboos 25 0
Tree holes 11 0
and L. stegomyiae. al.( 1978). Thus far, indications are that the ciliates from
Studies carried out in South Africa showed that L. both were L. stegomyiae.
stegomyiae were also found in Culex clecens and Culex In the laboratory, L.stegomyiae was able to penetratenebulosus( WHO 1981). However, in our studies none the cuticle of first-instar larvae. As the larva molts, itof the Culex species examined were infected. The finding becomes more difficult for the parasite to penetrate theofciliates inArmigeres andAn. subpictus was interesting cuticle, and the larvae may feed on the parasites. Lackand consistent with reports by Corliss( 1961) and Yu et of invasiveness makes L. stegomyiae a poor biological
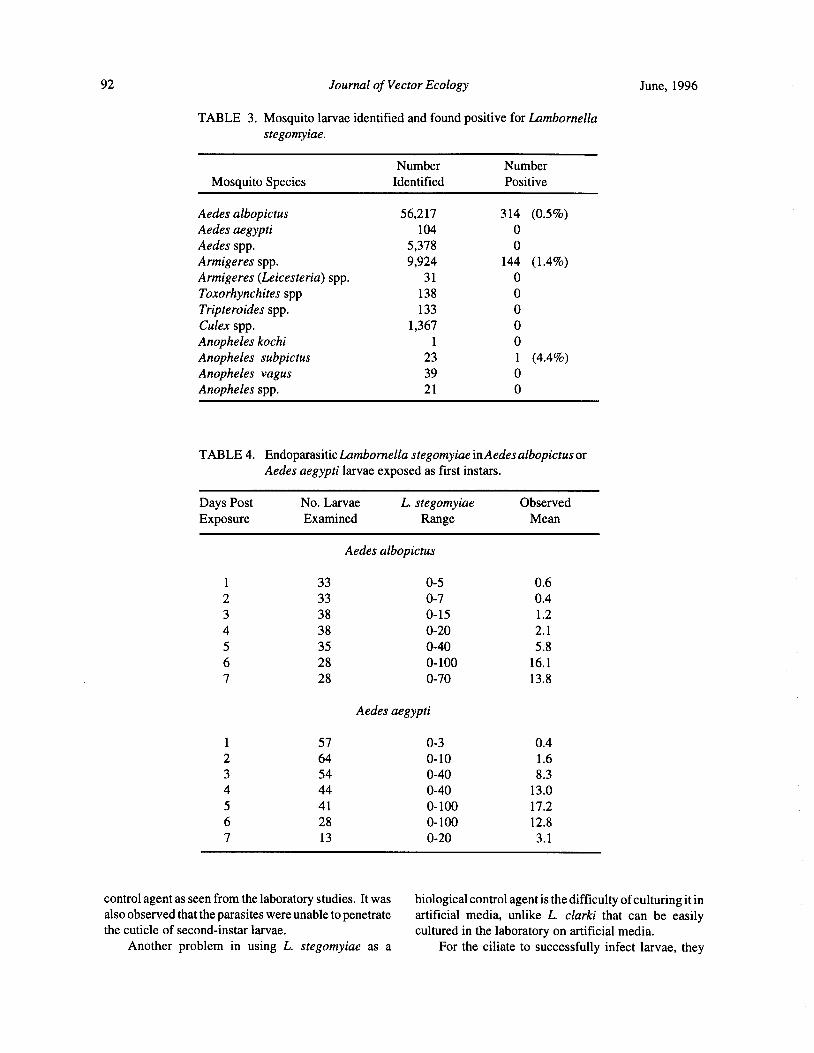
92 Journal of Vector Ecology June, 1996
TABLE 3. Mosquito larvae identified and found positive for Lambornella
stegomyiae.
Number Number
Mosquito Species Identified Positive
Aedes albopictus 56,217 314 ( 0. 5%)
Aedes aegypti 104 0
Aedes spp. 5, 378 0
Armigeres spp. 9,924 144 ( 1. 4%)
Armigeres( Leicesteria) spp. 31 0
Toxorhynchites spp 138 0
Tripteroides spp. 133 0
Culex spp. 1, 367 0
Anopheles kochi 1 0
Anopheles subpictus 23 1 ( 4.4%)
Anopheles vagus 39 0
Anopheles spp. 21 0
TABLE 4. Endoparasitic Lambornella stegomyiae inAedes albopictus or
Aedes aegypti larvae exposed as first instars.
Days Post No. Larvae L. stegomyiae Observed
Exposure Examined Range Mean
Aedes albopictus
1 33 0- 5 0.6
2 33 0- 7 0.4
3 38 0- 15 1. 2
4 38 0- 20 2. 1
5 35 0- 40 5. 8
6 28 0- 100 16. 1
7 28 0-70 13. 8
Aedes aegypti
1 57 0- 3 0.4
2 64 0- 10 1. 6
3 54 0-40 8. 3
4 44 0-40 13. 0
5 41 0- 100 17. 2
6 28 0- 100 12. 8
7 13 0- 20 3. 1
control agent as seen from the laboratory studies. It was biological control agent is the difficulty ofculturing it inalso observed that the parasites were unable to penetrate artificial media, unlike L. clarki that can be easilythe cuticle of second- instar larvae. cultured in the laboratory on artificial media.
Another problem in using L. stegomyiae as a For the ciliate to successfully infect larvae, they

June, 1996 Journal of Vector Ecology 93
must avoid being eaten by potential hosts, must attach to 55: 149- 152.the host where dislodgment is unlikely, and finally Dzerhinsky, V. A., E. A. Nam and A. M. Dubitsky.invading ciliates must not become trapped between the 1976. The finding of Lankesteria culicis andcuticle and epidermis. Ciliates should also be able to Tetrahymena stegomyiae in larvae ofAedesaegypti.escape from dead hosts. Parasitologija 10: 381- 382.
Lambornella clarki, which parasitizes the western Laird M. 1959. Parasites of Singapore mosquitoes withtree-hole mosquito, Aedes sierrensis, appears to be a particular reference to the significance of larval
promising candidate as a biological agent ( Washburn epibionts as an index of habitat pollution. Ecologyand Anderson 1986). The present study has clearly 40: 206- 221.
demonstrated that L. stegomyiae is widespread in Lamborn, W. A. 1921. A protozoon pathogenic tonature especially in Ae. albopictus larvae in rubber cups mosquito larvae. Parasitology 13: 213- 215.in rubber estates in most of Peninsular Malaysia. MacArthur,W. P. 1922. A holotrichus ciliate pathogenicHowever the apparent lack of invasiveness of L. to Theobaldia annulata. J. Army. Med. Corps. 38:stegomyiae in the present study is difficult to explain 83- 92.
and more work is necessary to evaluate its use as a Muspratt, J. 1945. Observation on the larvae of treeholebiological control agent. breeding Culicini ( Diptera: Culicidae) and two of
their parasites. J. Entomol. Soc. S. Africa. 8: 13- 20.Acknowledgments Muspratt, J. 1947. Notes on a ciliate protozoan, probably
Glaucoma pyriformis parasitic in culicine mosquitoThe authors wish to thank the Director, Institute for larvae. Parasitology 38: 107- 110.
Medical Research for his permission to publish this Sulaiman, S. and J. Jeffery. 1986. The ecology ofAedespaper and John Jeffery for his constructive comments. albopictus( Skuse)( Diptera: Culicidae) in a rubberThis project was funded by Malaysian Government estate in Malaysia. Bull. Entomol. Res. 76: 553-R& D Grant No. IMR 92- 7. 557.
Washburn, J. O. and Anderson, J. R. 1986. DistributionREFERENCES CITED of Lambornella clarki ( Ciliophora: Tetra-
hymenidae) and other mosquito parasites inChapman, H. C. 1974. Biological control of mosquito California Treeholes. J. Invert. Pathol. 48: 296-
larvae. Ann. Rev. Entomol. 19: 33- 59. 309.
Clark, T. B and D. G. Brandl. 1976. Observations on the World Health Organization. 1981. Data sheet on theinfection of Aedes sierrensis by a tetrahymenine Biological Control agents. Unpublished documentciliate. J. Invert Pathol. 23: 341- 349. WHO/ VBC 81. 803.
Corliss, J. O. 1954. The literature on Tetrahymena: its Yu, H. S., H. W. Cho, and J. S. Pillai. 1978. Field
history, growth, and recent trends. J. Protozool. 1: survey of mosquito pathogens in South Korea and156- 169. infection ofAnopheles, Aedes, and Culex larvae by
Corliss, J. O. 1961. Natural infection of tropical protozoan parasites Tetrahymena spp. andmosquitoes by ciliated protozoa of the genus Lankesteria culicis ( Ross). Rpt. Natl. Inst. Hlth.Tetrahymena. Trans. Roy. Soc. Trop. Med. Hyg. Korea. 15: 283- 290.

Journal of Vector Ecology 21( 1): 94- 104
Blood Feeding Success ofAedes aegypti and Culex nigripalpusDiptera: Culicidae) in Relation to Defensive Behavior of
Japanese Quail (Coturnix japonica) in the Laboratory
R. A. Anderson and R. A. Brust
Department ofEntomology, 214 Animal Science/ Entomology,University of Manitoba, Winnipeg, MB, R3T 2N2, CANADA.
Received 9 November 1995; Accepted 29 December 1995
ABSTRACT: Laboratory studies were conducted to determine the degree to which individual Japanesequail vary with respect to intensity and relative prevalence of several categories of anti-mosquito behaviorwhen exposed to Aedes aegypti( L.) or Culex nigripalpus Theobald in the laboratory. Quail were exposedin pairs and their behavior recorded by videotape. Five categories of behavior, including head shakes, footstamps, pecks, feather fluffs, and changes in body position, were observed before the quail were exposedto mosquitoes. In response to mosquito attack, the frequency of each of these behaviors increased. Theintensity of each of the three dominant behaviors( pecks, foot stamps, and head shakes) varied significantlyamong individual birds. The overall intensity of anti- mosquito activity, as measured by all categoriescombined, varied significantly between birds in each pair and was negatively correlated with the proportionof blood-fed mosquitoes in each cage that had fed on the corresponding bird.
Key Words: Blood-feeding, mosquitoes, Culex, defensive behavior.
INTRODUCTION cases, behavioral differences among otherwise similarhost individuals may influence feeding success and thus
Host-seeking mosquitoes face a great deal of vectorial capacity of attacking mosquitoes( Smith 1987,uncertainty in locating vertebrate hosts in complex Dye 1992).
habitats. Once located, not all vertebrates are necessarily During a field study of blood feeding success ofequally available because some are less tolerant than several species ofwild mosquitoes on pairs ofapparently
others ofmosquito attack( Edman et al. 1972, Kale et al. identical birds, we observed that blood feeding often1972). Edman and Scott( 1987) listed host species, size, was distributed unevenly between the two hostsage, health, and individuality as factors that they ( Anderson and Brust, unpublished data). We assumed
considered to account for variation in mosquito feeding that differences in the probability ofeach bird being fedsuccess on different hosts. Individuality ( differences upon were due to corresponding differences in thenot related to species, size, age, and health status) was intensity of defensive behavior exhibited by each of theaccorded the least importance. However, published bait hosts ( quail) as has been demonstrated for other
studies on differences in defensive behavior among avian species( Edman et al. 1972, Kale et al. 1972). We
species may not have included enough replicate hypothesized that the probability of any one bird beinganimals to thoroughly document the degree of variation fed on was inversely proportional to the degree ofamong hosts of the same physical status( Edman et al. defensive behavior of that bird relative to the other bird
1972, Kale et al. 1972, Webber and Edman 1972). in the cage. The purpose of this paper is to report the
Ornithophilic mosquito species, including Culex tarsalis results of experiments conducted to evaluate the
Coquillett, may vector arboviruses, such as Western relationship between defensive behavior and bloodEquine Encephalomyelitis Virus ( WEEV) among feeding success, and to present information on therelatively homogeneous populations of birds, for degree of variability in several anti-mosquito behaviorsexample, house sparrows( Holden et al. 1973). In these among individual quail.

June, 1996 Journal of Vector Ecology 95
MATERIALS AND METHODS quail was placed into each of two compartments in the
top half of the cage. The quail were separated by oneWe conducted laboratory experiments with two layer ofmesh to prevent them from interacting directly
species of mosquitoes. First, Japanese quail', Coturnix during the experiment, but they were otherwisejaponica Temminck and Schlegel,( Saskatchewan wild unrestrained. The remaining volume ofthe cage( bottomtype, Quail Genetic Stock Centre, University of British half) provided a refuge for mosquitoes from the quail.Columbia) were exposed to Aedes aegypti( L.)( Rocke- Quail behavior was recorded on video tape prior tofeller strain). Fifteen pair of quail were used; six pair and during exposure to the mosquitoes. All experimentswere exposed to 100 mosquitoes and nine pair were were conducted under red light. The light and darkexposed to 200 mosquitoes. Each pair was exposed periods of each species of mosquito were adjusted so
twice, each time to the same number of mosquitoes. that experiments were carried out during the photophaseFirst and second exposures were separated by no more ofAe. aegypti and the scotophase of Cx. nigripalpus tothan 8 hours to minimize variation in the hunger status match the active period of each mosquito. Theofmosquitoes drawn from the same cohort. Female Ae. experiments were conducted during the quail scotophaseaegypti were 3 to 5 days old and maintained on 3% when they are normally inactive. Mosquitoes weresucrose until 48 hours prior to use, when water only was placed in an acrylic tube separated from the main cageprovided. The photoperiod ofAe. aegypti was adjusted by a remotely controlled sliding door. The quail wereso that they were allowed to feed during their photophase placed in the cage approximately 30 minutes beforeas the peak feeding activity of this species is during the video taping began, to allow them to habituate to theday. In the second experiment, 20 pair of Japanese quail cage. All manipulations performed by the experimenter,of the same line were exposed to Culex nigripalpus including activation of video recorders, were done fromTheobald. The photoperiod of Cx. nigripalpus was behind a black cloth screen to minimize disturbance toadjusted so that experimental exposure to quail coincided the quail. Experiments were conducted in well- insulated,with the scotophase of this mosquito species, the time controlled-environment chambers to minimize externalwhen it is naturally blood feeding in the wild. Each pair noise disturbance. Mosquitoes were released into theof quail was exposed once to 100 Cx. nigripalpus( F13), cage five minutes after the cameras were activated.a Vero Beach strain colonized from wild females After 55 minutes of exposure to the mosquitoes, thecollected at the Florida Medical Entomology Laboratory quail were removed from the cage. The number ofdeadin 1992. The colony was not sufficiently productive to mosquitoes in the cages and their blood- fed status wereexamine the effect of density and exposure. Female Cx. recorded. The cages were then placed in a freezer to killnigripalpus were 7 to 12 days old, but otherwise main- the remaining mosquitoes. After freezing, mosquitoestamed on the same sugar and water regimen as Ae. were counted and sorted according to the presence ofaegypti in the first experiment. blood and then analyzed for rubidium and cesium
For the first experiment, 15 quail were injected with ( Anderson et al. 1990).
rubidium and 15 others with cesium so that we could The proportion of blood- fed mosquitoes in each
identify the blood source( s) of each engorged mosquito cage was calculated as the number of mosquitoes with
Anderson et al. 1990). Each injected bird was marked blood from at least one quail divided by the number ofwith a leg label that denoted its marker and a bird mosquitoes released into the cage. The proportion of
number. Quail were exposed in pairs. The birds in each mosquitoes with multiple blood meals was calculated aspair were selected randomly by reaching into the flock the number of mosquitoes with blood from both quail
cage and catching each individual by hand without divided by the number of blood- fed mosquitoes.regard to the label. Rubidium- injected quail were Comparisons of entomological data ( proportion ofpaired with cesium- injected quail, but without regard to engorged mosquitoes and proportion of mosquitoes
bird number. The bird number was used to keep the bird with multiple meals) were made in the following manner.pairs consistent from the first exposure to the second. A possible effect of mosquito species was examined byThe quail were placed in 30 X 30 X 30 cm clear, acrylic comparing results from trials with Cx. nigripalpus onlycages( Fig. 1). The cages were divided with galvanized with results from Ae. aegypti cages in which there weremetal screening( mesh size, 13 X 13 mm) such that one 100 mosquitoes/cage ( first exposure). The effect of
Animal Use; all experiments which involve the use of animals conform to guidelines contained in the Guide to theCare and Use ofExperimental Animals, Vol. 1., Canadian Council on Animal Care and experimental protocol# C-91- 46 has been approved by the University of Manitoba Animal Care Committee.

96 Journal of Vector Ecology June, 1996
AIIII
I III t f,. t
II ! 1 I
fI!r 1.. 1. t1. i!Ill.„,„,„„, „;
rt III !-; I+ t itllIi, ,;-lki i} 1 h P 1; 1 I I I L l¢ 4
I!,,{I
Ii l ii
t
fit+ 't.St t ,,! i i
l -
1JII r- It 1 1— k ti
tI1II 111'.
1 1 1 1 h i! 4I t I{
1l1 1 Fri 1 19—i t+
iceI II
I i 1 I' 77FFi # t i.1-.
1ryry11 EI'..1...i..
jIf'
iIm$iyfi{ i i i;
1
Ii ilil1 i11 r . I.. iii•'T'' LLI'rllitii I liyl. i ,
rEPIIIH6_Figure 1. Experimental cage used to expose quail to Ae. aegypti or Cx. nigripalpus. The mosquitoes were released
by remote control from the cylinder at the bottom- front of the cage. The quail were separated from eachother by hardware cloth( 13 X 13 mm mesh). The clear acrylic front and sides of the cages permitted videorecording of quail behavior.
exposure was examined only for Ae. aegypti at each was done for entomological data, comparisons of quail
density. The effect ofdensity was examined, again only behavior during mosquito exposure among groups offorAe. aegypti at each exposure. Thus, five comparisons cages with different species of mosquitoes, different
were made using the 2-sample t- test and the rank sum numbers of the same species( Ae. aegypti), or from the
test( Snedecor and Cochran 1976). first exposure to the second( for each of the two densities
Quail behavioral counts were transcribed from the ofAe. aegypti) were made by the 2- sample t- test and thevideo tapes to data sheets. All activity during the five rank sum test( Snedecor and Cochran 1976). An overall
minute interval prior to mosquito release was counted. index ofactivity foreach quail in each cage was calculatedFor each tape, a 10 minute interval from the mosquito by dividing the total number ofmovements( all categoriesexposure period( from 5 to 15 minutes after mosquito combined) by one quail by the total of all movements ofrelease) was selected for transcription. The length of the both quail. Thus, the activity index for each quail wasinterval sampled was determined from preliminary a proportion of all activity for the corresponding cage.analysis to be sufficient to predict the overall pattern of This index was compared by regression to the proportionbehavior for each bird. Head shakes, pecks, feather of blood meals attributable to the corresponding quailfluffs, change of location, and foot stamps were counted for each of the two species of mosquito.
for analysis. Categories ofbehavior were decided upon
from trial observations of quail before and during RESULTS
exposure to mosquitoes prior to the actual experiments.
Quail used for the experiments had not been exposed The mean proportion of blood-fed mosquitoes was
previously to mosquitoes. The categories listed above highest( 0.302) for cages with 100Ae. aegypti( exposure
were those that accounted for most of the quail activity 1) and lowest (0.078) for cages with 200 Ae. aegypti
and that changed most markedly in intensity during ( exposure 2). The mean proportion of blood- fed
exposure to mosquitoes. mosquitoes decreased significantly from exposure 1 toThe rates of each type of activity per minute were exposure 2 for cages with 200 Ae. aegypti. The mean
calculated. The rates ofeach behavior before and during proportion of multiple meals was highest ( 0. 14) for
exposure to mosquitoes were compared by the paired t- cages with 200 Ae. aegypti ( exposure 2) and lowest
test and the sign test( Snedecor and Cochran 1976). As ( 0.046) for cages with 100 Ae. aegypti ( exposure 1),
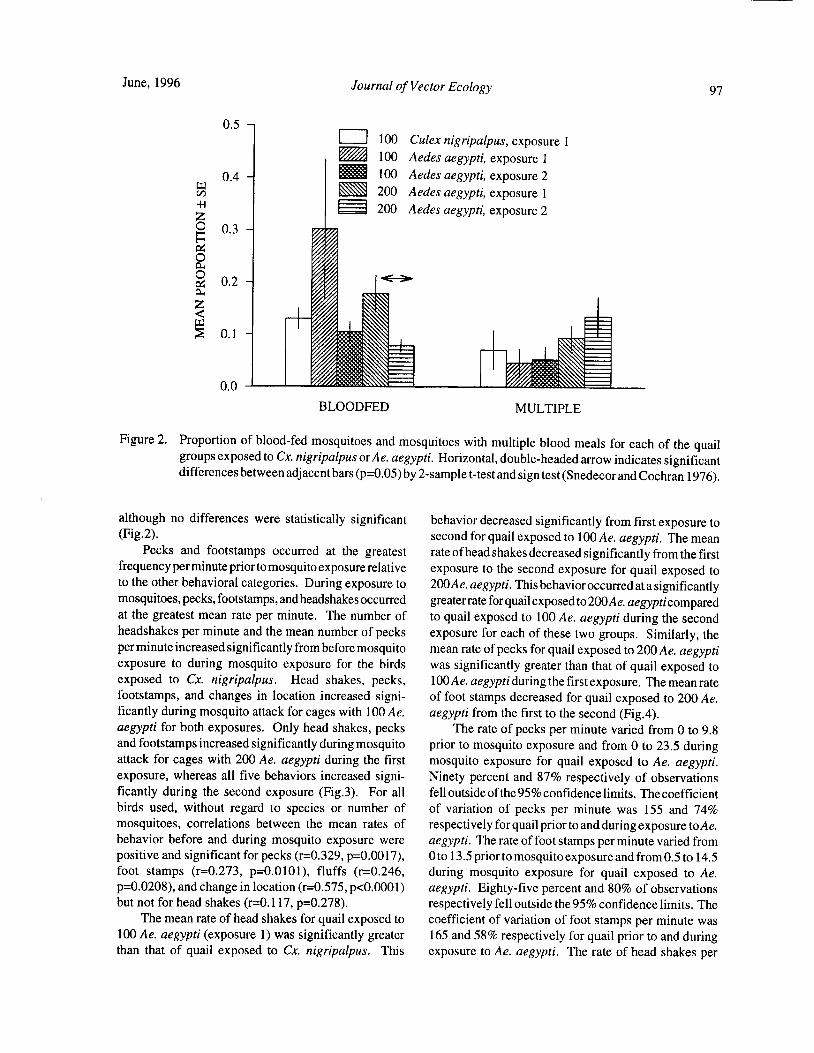
June, 1996 Journal of Vector Ecology 97
0. 5
100 Culex nigripalpus, exposure 1
A 100 Aedes aegypti, exposure 10.4 100 Aedes aegypti, exposure 2
200 Aedes aegypti, exposure 1I
200 Aedes aegypti, exposure 2
0. 3
o
a 0. 2 F
mom
low. mom
BLOODFED MULTIPLE
Figure 2. Proportion of blood-fed mosquitoes and mosquitoes with multiple blood meals for each of the quailgroups exposed to Cx. nigripalpus orAe. aegypti. Horizontal, double- headed arrow indicates significantdifferences between adjacent bars( p= 0.05) by 2- sample t- test and sign test( Snedecor and Cochran 1976).
although no differences were statistically significant behavior decreased significantly from first exposure toFig.2). second for quail exposed to 100 Ae. aegypti. The mean
Pecks and footstamps occurred at the greatest rate ofhead shakes decreased significantly from the firstfrequency perminute prior to mosquito exposure relative exposure to the second exposure for quail exposed toto the other behavioral categories. During exposure to 200Ae. aegypti. This behavior occurred at a significantlymosquitoes, pecks, footstamps, and headshakes occurred greater rate forquail exposed to200Ae. aegypti comparedat the greatest mean rate per minute. The number of to quail exposed to 100 Ae. aegypti during the secondheadshakes per minute and the mean number of pecks exposure for each of these two groups. Similarly, theper minute increased significantly from before mosquito mean rate of pecks for quail exposed to 200 Ae. aegyptiexposure to during mosquito exposure for the birds was significantly greater than that of quail exposed toexposed to Cx. nigripalpus. Head shakes, pecks, 100Ae. aegypti during the firstexposure. The mean ratefootstamps, and changes in location increased signi- of foot stamps decreased for quail exposed to 200 Ae.ficantly during mosquito attack for cages with 100 Ae. aegypti from the first to the second( Fig. 4).aegypti for both exposures. Only head shakes, pecks The rate of pecks per minute varied from 0 to 9. 8and footstamps increased significantly during mosquito prior to mosquito exposure and from 0 to 23. 5 duringattack for cages with 200 Ae. aegypti during the first mosquito exposure for quail exposed to Ae. aegypti.exposure, whereas all five behaviors increased signi- Ninety percent and 87% respectively of observationsficantly during the second exposure ( Fig.3). For all fell outside of the 95% confidence limits. The coefficientbirds used, without regard to species or number of of variation of pecks per minute was 155 and 74%mosquitoes, correlations between the mean rates of respectively for quail prior to and during exposure to Ae.behavior before and during mosquito exposure were aegypti. The rate of foot stamps per minute varied frompositive and significant for pecks( r=0.329, p= 0.0017), 0 to 13. 5 prior to mosquito exposure and from 0.5 to 14. 5foot stamps ( r=0. 273, p= 0. 0101), fluffs ( r=0.246, during mosquito exposure for quail exposed to Ae.p= 0.0208), and change in location( r=0. 575, p< 0.0001) aegypti. Eighty- five percent and 80% of observations
but not for head shakes( r=0. 117, p= 0. 278). respectively fell outside the 95% confidence limits. The
The mean rate of head shakes for quail exposed to coefficient of variation of foot stamps per minute was100 Ae. aegypti( exposure 1) was significantly greater 165 and 58% respectively for quail prior to and duringthan that of quail exposed to Cx. nigripalpus. This exposure to Ae. aegypti. The rate of head shakes per
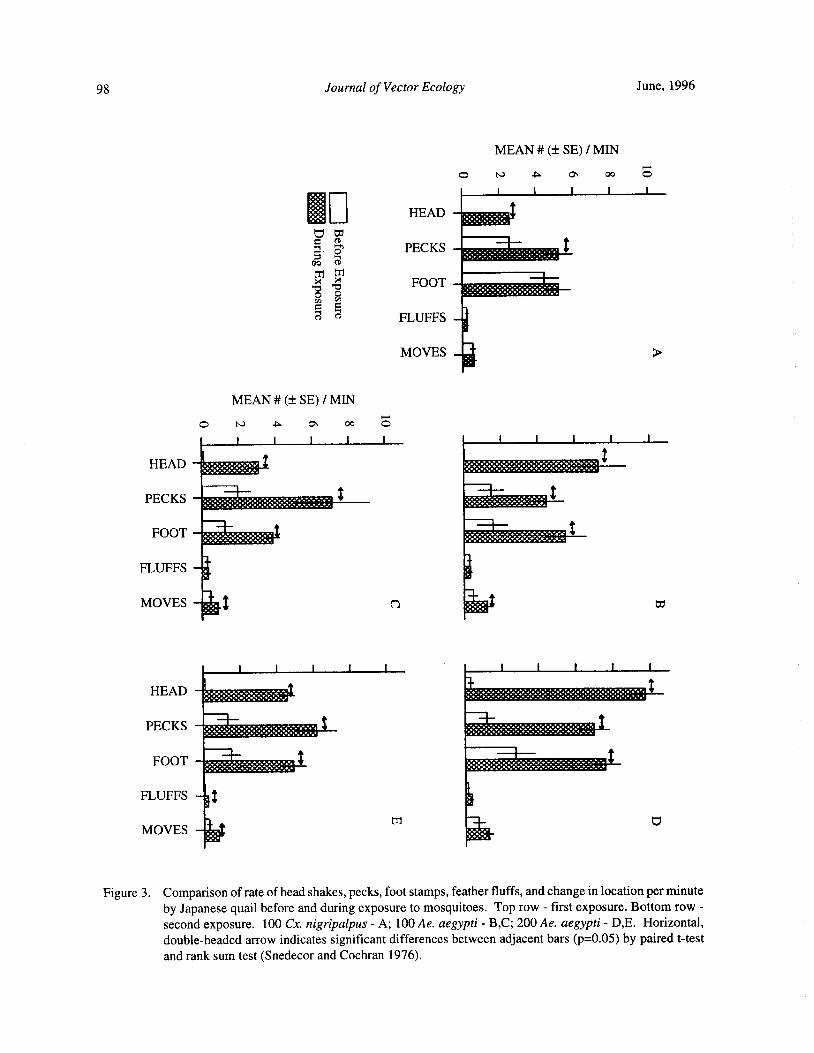
98 Journal of Vector Ecology June, 1996
MEAN#( t SE)/ MIN
N . CT CC O
HEAD
d
PECKS MI= t
n FOOT
cD
0 0
FLUFFS
MOVES
MEAN#(± SE)/ MIN
O N 4 CA 00 O
HEAD
PECKS j C
FOOT C
FLUFFS
MOVES C- bd
HEADt
PECKS C C
FOOT C Z
FLUFFS
MOVES Im
Figure 3. Comparison of rate of head shakes, pecks, foot stamps, feather fluffs, and change in location per minute
by Japanese quail before and during exposure to mosquitoes. Top row- first exposure. Bottom row-second exposure. 100 Cx. nigripalpus- A; 100 Ae. aegypti- B,C; 200 Ae. aegypti- D,E. Horizontal,
double-headed arrow indicates significant differences between adjacent bars( p= 0.05) by paired t-testand rank sum test( Snedecor and Cochran 1976).
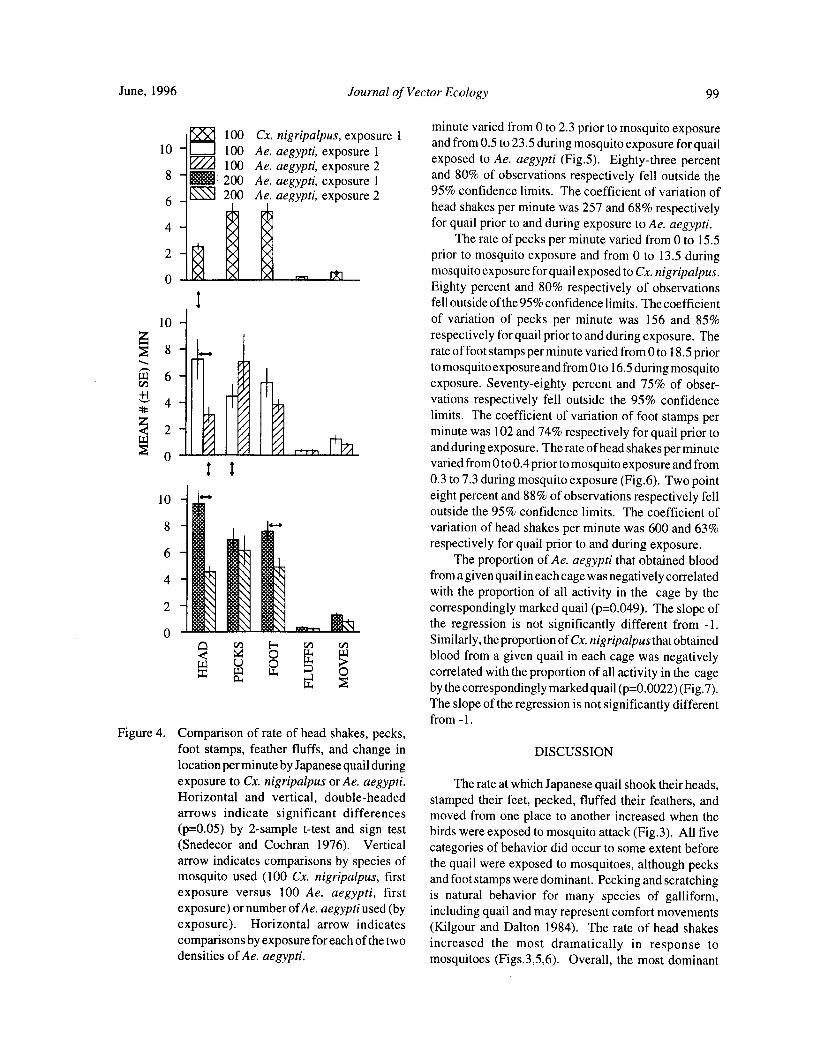
June, 1996 Journal of Vector Ecology 99
minute varied from 0 to 2. 3 prior to mosquito exposure100 Cx. nigripalpus, exposure 1
and from 0. 5 to 23. 5 during mosquito exposure for quail10 LJ 100 Ae. aegypti, exposure 1exposed to Ae. aegypti ( Fi ' 5 . Eighty- three percent®
100 Ae. aegypti, exposure 28 200 Ae. aegypti, exposure 1
and 80% of observations respectively fell outside the
200 Ae. aegypti, exposure 2 95% confidence limits. The coefficient of variation of6 head shakes per minute was 257 and 68% respectively
4 for quail prior to and during exposure to Ae. aegypti.The rate of pecks per minute varied from 0 to 15. 5
2 P prior to mosquito exposure and from 0 to 13. 5 during
0 • m mosquito exposure for quail exposed to Cx. nigripalpus.
Eighty percent and 80% respectively of observationsIfell outside of the 95% confidence limits. The coefficient
10 of variation of pecks per minute was 156 and 85%
4 respectively for quail prior to and during exposure. The2 8 rate of foot stamps per minute varied from 0 to 18. 5 prior
n to mosquito exposure and from 0 to 16. 5 during mosquito6
0% exposure. Seventy- eighty percent and 75% of obser-
14 vations respectively fell outside the 95% confidence
V limits. The coefficient of variation of foot stamps per2 V minute was 102 and 74% respectively for quail prior to
4 /, /, N91 and during exposure. The rate ofhead shakes per minute0
varied from 0 to 0.4 prior to mosquito exposure and fromI0. 3 to 7. 3 during mosquito exposure( Fig.6). Two point
10 — eight percent and 88% of observations respectively felloutside the 95% confidence limits. The coefficient of
8 — F-. variation of head shakes per minute was 600 and 63%
respectively for quail prior to and during exposure.6
1 The proportion of Ae. aegypti that obtained blood
4 — \ ': ::: from a given quail in each cage was negatively correlatedwith the proportion of all activity in the cage by the
2 — ::: correspondingly marked quail( p= 0.049). The slope of
0 ::.\ II i:M the regression is not significantly different from - 1.
AW
Similarly, the proportion of Cx. nigripalpus that obtainedQ U 0 blood from a given quail in each cage was negatively
We. w 0 correlated with the proportion of all activity in the cageby the correspondingly marked quail( p= 0.0022)( Fig.7).The slope of the regression is not significantly differentfrom- 1.
Figure 4. Comparison of rate of head shakes, pecks,
foot stamps, feather fluffs, and change in DISCUSSION
location per minute by Japanese quail duringexposure to Cx. nigripalpus or Ae. aegypti. The rate at which Japanese quail shook their heads,Horizontal and vertical, double- headed stamped their feet, pecked, fluffed their feathers, andarrows indicate significant differences moved from one place to another increased when thep=0.05) by 2- sample t- test and sign test birds were exposed to mosquito attack( Fig. 3). All fiveSnedecor and Cochran 1976). Vertical categories of behavior did occur to some extent before
arrow indicates comparisons by species of the quail were exposed to mosquitoes, although pecksmosquito used ( 100 Cx. nigripalpus, first and foot stamps were dominant. Pecking and scratchingexposure versus 100 Ae. aegypti, first is natural behavior for many species of galliform,exposure) or number ofAe. aegypti used( by including quail and may represent comfort movementsexposure). Horizontal arrow indicates ( Kilgour and Dalton 1984). The rate of head shakescomparisons by exposure for each of the two increased the most dramatically in response todensities of Ae. aegypti. mosquitoes ( Figs. 3, 5, 6). Overall, the most dominant
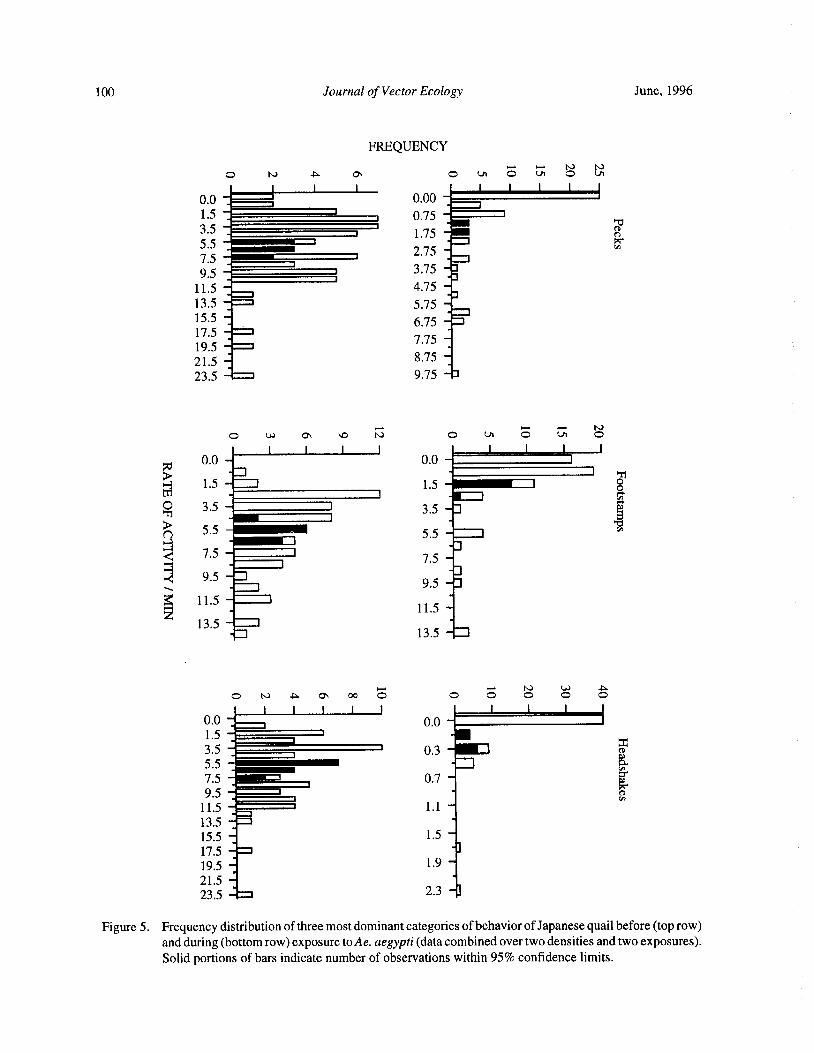
100 Journal of Vector Ecology June, 1996
FREQUENCY
O N - P. as O U O Ui N NVi
I 1 I I I 1
0.0r 0.00 - I
1. 5 - I1 0. 75 -
3. 5 - b
1. 75
7. 5 - 2. 75
9.5 - 3. 75 -
11. 5 -= I
4.75
13. 5 _° 5. 75
15. 5 - 6.75 -17. 5 :=
7.75 -19. 5 -:_c==
21. 5 - 8. 75 =
23. 5 - 9.75 - Q
O W o N O U - - N
l I I I0.0 - 0.0
m 11 1111N11111111111 111111111• 1 II
m 1. 5 - I 1. 5 0I N G
OO 3. 5 - I 3. 5 •ri I 5
5,3n 5. 5 5. 5
7. 5 - I7.5
i
9. 5 -- L7
9. 5 ••I
11. 5 I
11. 5
13. 5 -' I
13. 5
O N - IP C\ 00 O O ON W
O O
I I I I I
0.0 - 0.01. 5 - I
3. 5 - l 0. 3
5. 5 EN a
7. 5 - w= 0.7•
9.5 - t CD
11. 5 fl 1. 1y
13. 5 -<= 1
15. 5 - 1. 5
17. 5 - C= 3 I
19. 5 - 1. 9
21. 5 -
23.5 .. 2. 3 I
Figure 5. Frequency distribution of three most dominant categories ofbehavior of Japanese quail before( top row)and during( bottom row) exposure to Ae. aegypti( data combined over two densities and two exposures).Solid portions of bars indicate number of observations within 95% confidence limits.
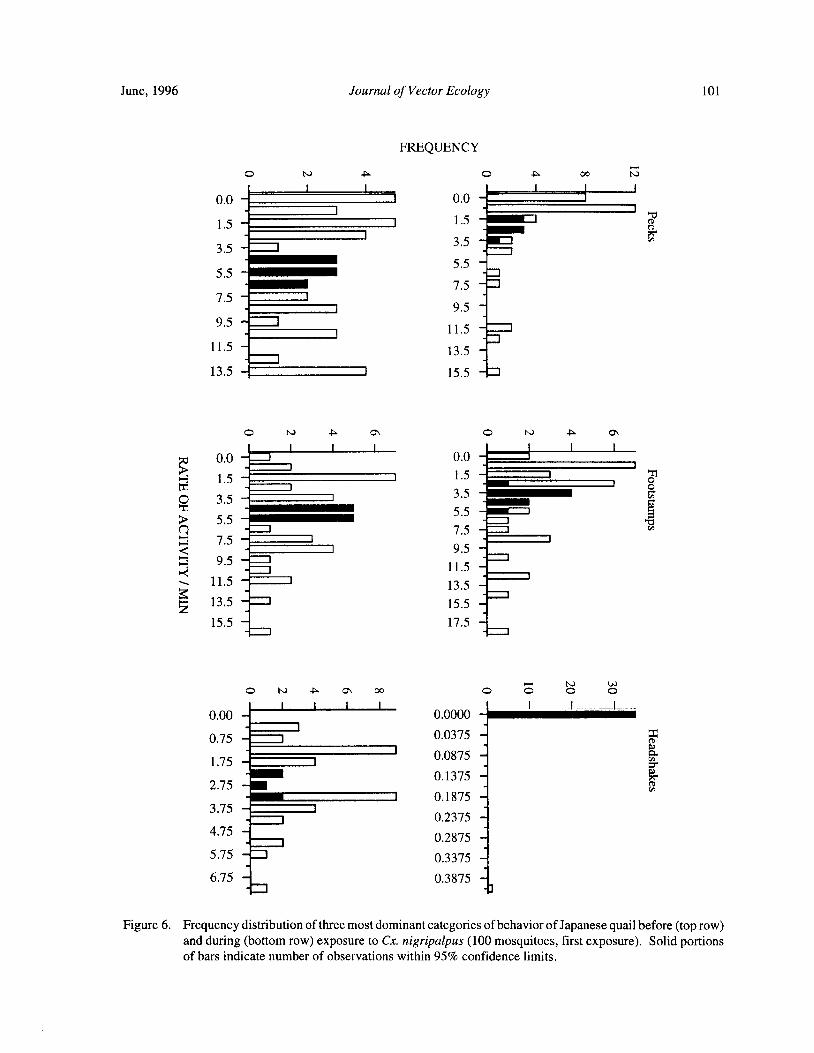
June, 1996 Journal of Vector Ecology 101
FREQUENCY
O N O 00 N
I L0.0 - I 0. 0
1 b1. 5 - I 1. 5 No cco
3. 5 -, I
I3. 5 mil
5. 55. 5
7. 5 •7.5 - 1
1 9. 5
9.5 - 111. 5 E
1mu
11. 5 - 13. 5I
13. 5 - 1 15. 5 No
O N . ON O N ? ON
0.0 -I I I
0.01 I
M 1. 5 - 1 1. 5 - 1
0I
1 o
O3. 5 -
3. 5 -111111111• 111111
5. 5
5 .5 -,--
J 7. 5 - 1 cn
7.5 - 11 9. 5 -
1
y 9.5 - I 11. 5 -
11. 5 - 113. 5 -
1
13. 5 15. 5 715. 5 - 17. 5 -
I ti_ 1
N WO N 4,. O\ 00 O O O O
0.00 - I I I I
0. 0000I I._-.... - 1
0.75 J I
I0.0375 c
1. 75 - Ii
0.0875
0. 1375cp2.75 All rn
I 0. 18753. 75 - I
1 0.2375
4.75 -1
0.2875
5. 75 0.3375
6.75 - 0.3875
Figure 6. Frequency distribution of three most dominant categories of behavior of Japanese quail before( top row)and during( bottom row) exposure to Cx. nigripalpus( 100 mosquitoes, first exposure). Solid portions
of bars indicate number of observations within 95% confidence limits.
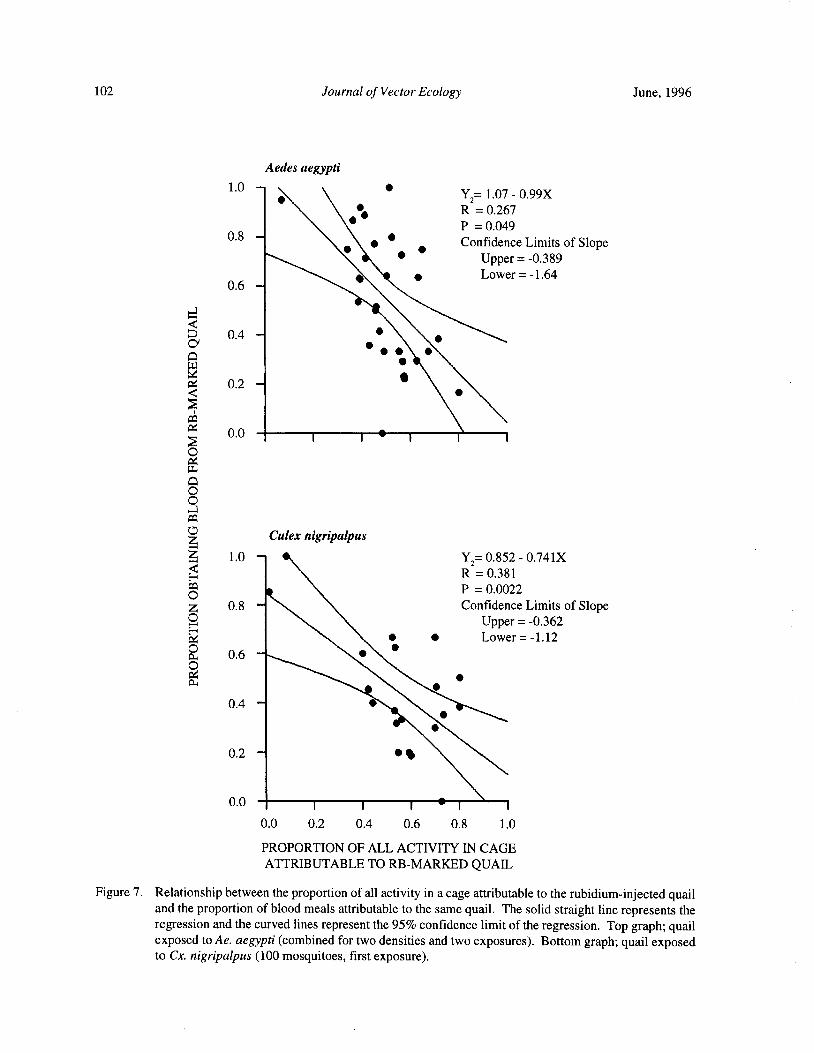
102 Journal of Vector Ecology June, 1996
Aedes aegypti
1. 0 • Y2= 1. 07- 0.99XR = 0.267
P = 0.0490. 8
Confidence Limits of Slope
Upper=- 0.389
Lower=- 1. 640.6
0.4
WQW
Q 0. 2
0.0
O
00
Culex nigripalpus
4 1. 0 YZ 0.852- 0. 741XR = 0.381
P = 0.0022
z 0. 8 Confidence Limits of Slope
O Upper=- 0.362
Lower=- 1. 12O
0.60
a0.4
0.2
0.0
0.0 0.2 0.4 0. 6 0. 8 1. 0
PROPORTION OF ALL ACTIVITY IN CAGE
ATTRIBUTABLE TO RB-MARKED QUAIL
Figure 7. Relationship between the proportion of all activity in a cage attributable to the rubidium-injected quailand the proportion of blood meals attributable to the same quail. The solid straight line represents theregression and the curved lines represent the 95% confidence limit of the regression. Top graph; quailexposed to Ae. aegypti( combined for two densities and two exposures). Bottom graph; quail exposedto Cx. nigripalpus( 100 mosquitoes, first exposure).

June, 1996 Journal of Vector Ecology 103
anti-mosquito behaviors were exaggerations of pre- evidence of extraordinary variation in the rates of
existing behavioral patterns. Although we could not behavior among birds may be derived from the coefficientunequivocally determine the number of mosquitoes of variation which ranged upwards from 58%. Another
attacking different anatomical areas of the quail, we did way to view the extent of relative variation between anyobserve relatively high attack rates on the heads and 2 quail in defensive behavior may be seen in Figure 7.feet. This may explain why foot stamps also were one The proportion of all activity in a given cage attributableof the dominant categories during mosquito exposure to 1 quail varied from near 0 to near 1 and this was
Figs. 3, 5, 6). Most of the dead mosquitoes, about one- negatively correlated with the proportion ofblood mealshalf of all Ae. aegypti and about one- fourth of Cx. attributable to the corresponding quail. These data
nigripalpus, were eaten( Fig.2). This was likely because support our assumption that relative variation in blood
pecks were a relatively dominant category of behavior feeding success of mosquitoes on individual quail hostsFigs. 3, 5, 6) during mosquito exposure. The high is due, at least in part, to differences in tolerance of
probability of mortality observed in this study may be mosquito attack( Anderson and Brust unpublished data).
interpreted as evidence that blood feeding under the Other authors have noted variation in the engorged
laboratory conditions described is risky behavior. proportion of mosquitoes attracted to different
Although the mean number and proportion ofblood- individuals of the same avian species ( Blackmore and
fed mosquitoes decreased significantly from exposure 1 Dow 1958, Dow et al. 1957). In the context of our
to exposure 2 only in cages with 200Ae. aegypti( Fig.2), experiments with Japanese quail and those ofEdman et
the same trend is apparent for cages with 100 Ae. aegypti al. ( 1972) with several ciconiiform species, it is likely
Fig.2). It appears that blood feeding success of Ae. that the variation in engorgement rates on different
aegypti was negatively affected by the previous exposure individuals of the same avian species observed byof the host quail, however these results are difficult to Blackmore and Dow ( 1958) and by Dow et al. ( 1957)
explain in the context of the behavioral data obtained were due to differences in the intensity of anti- mosquitofrom the quail ( Fig.4). The rates of all categories of behavior.
behavior for quail exposed to 200Ae. aegypti decreased It was difficult for us to characterize an average
from exposure 1 to 2, significantly so in the case of quail with respect to the most dominant behavior because
headshakes and footstamps. The same trend is noted for of the variation among individuals( Figs.5, 6). Kale et al.behavior of quail exposed to 100 Ae. aegypti except for ( 1972) observed considerable variation among somethe rate ofpecks which increased( albeit insignificantly) individuals of the same ciconiiform species in the
from exposure 1 to exposure 2. We would have expected proportion of activity in each behavioral category.that decreases in blood feeding success should be Significant variation in defensive behavior amongcorrelated with increased rates of defensive behavior as individual avian hosts of the same species, size, age, sex,
was observed for several ciconiiform species( Edman et and health status may influence feeding success andal. 1972). It is possible that the quail learned to deter thus vectorial capacity of mosquitoes to a larger extentmosquitoes more effectively with fewer, more efficient than previously thought( Edman and Scott 1987). Furthermovements from the first to the second exposure. The research in this area with other avian hosts such as
rate of multiple feeding by Ae. aegypti observed in this passerine birds and different species of mosquitoes
study ( Fig.2) is similar to the rate of multiple feeding adapted to greater or lesser extent to feeding on birdsobserved for this species by Scott et al.( 1993). The rate may produce insight into those behaviors most importantof multiple feeding by Cx. nigripalpus observed in this in limiting blood feeding success of mosquitoes.
study ( Fig.2) is consistent with the rate of multiplefeeding observed for Cx. nigripalpus collected in the Acknowledgements
field in cages baited with two quail( Anderson and B rust
1995). We thank Terry Galloway( University of Manitoba)Japanese quail were extremely variable with respect and two anonymous reviewers for helpful comments on
to the rates at which they shook their heads, stamped the manuscript. We thank the staff of the University oftheir feet, and pecked, the three dominant categories of Manitoba Field Station and the Florida Medical
behavior( Figs.5, 6). The spread in the distribution of the Entomology Laboratory for logistical and materialrates of these 3 behaviors and the fact that most support and Robert Hunt of the Fresh Water Institute for
observations actually fall outside of the 95% confidence use of a flame spectrophotometer. We gratefullylimits may be interpreted as evidence of significant acknowledge financial assistance in the form of a
intrinsic variation in the intensity of defensive behavior University of Manitoba Graduate Fellowship and anagainst mosquitoes by individual quail. Additional Entomological Society of Canada research travel grant

104 Journal of Vector Ecology June, 1996
to RAA. We also acknowledge operating grants from Am. J. Trop. Med. Hyg. 21: 487- 491the Canadian Shield Foundation and the Natural Sciences Edman, J. D. and T. W. Scott. 1987. Host defensiveand Engineering Research Council of Canada ( grant behaviour and the feeding success of mosquitoes.No. A2545) to RAB. Insect Sci. Applic. 8: 617- 622.
This is publication No. 259 of the University of Holden, P., R. O. Hayes, C. J. Mitchell, D. B. Francy, J.Manitoba Field Station, Delta Marsh. S. Lazuick, and T.B. Hughes. 1973. House sparrows,
Passer domesticus ( L.) as hosts of arboviruses inREFERENCES CITED Hale County, Texas. I. Field studies, 1965- 1969.
Am. J. Trop. Med. Hyg. 22: 244- 253.Anderson, R. A. and R. A. Brust. 1995. Field evidence Kale, H. W., J. D. Edman, and L. A. Webber. 1972.
for multiple host contacts during blood feeding by Effect ofbehavior and age of individual ciconiiformCulex tarsalis, Cx. restuans, and Cx. nigripalpus birds on mosquito feeding success. Mosq. NewsDiptera: Culicidae). J. Med. Entomol. 32: 705- 32: 343- 350.
710. Kilgour, R. and C. Dalton. 1984. Livestock Behaviour.Anderson, R. A., J. D. Edman, and T. W. Scott. 1990. Westview Press, Boulder, CO, 320 pp.
Rubidium and cesium as host blood-markers to Scott, T. W.,E. Chow, D. Strickman, P. Kittayapong, R.study multiple blood feeding by mosquitoes A. Wirtz, L. H. Lorenz, and J. D. Edman. 1993.Diptera: Culicidae). J. Med. Entomol. 27: 999- Blood feeding patterns of Aedes aegypti1001. Diptera:Culicidae) collected in a rural Thai village.
Blackmore, J. S. and R. P. Dow. 1958. Differential J. Med. Entomol. 30: 922- 927.
feeding of Culex tarsalis on nestling and adult Smith, C.E.G. 1987. Factors influencing the transmissionbirds. Mosq. News 18: 15- 17. of western equine encephalomyelitis virus between
Dow, R. P., W. C. Reeves, and R. E. Bellamy. 1957. its vertebrate maintenance hosts and from them toField tests ofavian host preference ofCulextarsalis humans. Am. J. Trop. Med. Hyg. 37:( Suppl.) 33s-Coq. Am. J. Trop. Med. Hyg. 6: 294- 303. 39s.
Dye, C. 1992. The analysis of parasite transmission by Snedecor, G. W. and W. G. Cochran. 1976. Statistical
bloodsucking insects. Annu. Rev. Entomol. 37: 1- Methods. 6th edition. The Iowa State University19. Press. IA, 593 pp.
Edman, J. D., L. A. Webber, and H. W. Kale II. 1972. Webber, L. A. and J. D. Edman. 1972. Anti-mosquitoEffect of mosquito density on the interrelationship behaviour of ciconiiform birds. Animal Behay. 20:of host behavior and mosquito feeding success. 228- 232.

ADVERTISING
Commercial advertising space is available in the Journal;full page (black and white) at $ 150.00 per issue, half page
black and white) at $ 90.00 per issue. Inquiries may beaddressed to H. B. Munns, Advertising Manager, SOVE,P. O. Box 338, Pioneer, California 95666, telephone ( 209)
295- 3540 or Fax ( 209) 295- 3563.
The publisher reserves the right to approve or refuse anyadvertisement. The publisher is not responsible for anyclaims, litigations, or expenses resulting from theadvertiser's unauthorized use of any name, photograph,
sketch, or words protected by registered trademark orcopyright.





























