Embed Size (px)
Citation preview

Institute for Microbiology
Department of Infectious Diseases
University of Veterinary Medicine Hannover
Virulence mechanisms of Streptococcus suis:
Molecular characterisation of the
biological activities of suilysin
THESIS
Submitted in partial fulfilment of the requirements for the degree
DOCTOR OF PHILOSOPHY (PhD)
awarded by the University of Veterinary Medicine Hannover
by
Maren Seitz
(Hannover)
Hannover 2011

Supervisor: Prof. Dr. Peter Valentin-Weigand
Supervision Group: Prof. Dr. Silke Rautenschlein PD Dr. Manfred Rohde
1st Evaluation: Prof. Dr. Peter Valentin-Weigand
Institute for Microbiology
Department of Infectious Diseases
University of Veterinary Medicine Hannover
Prof. Dr. Silke Rautenschlein Clinic for Poultry
University of Veterinary Medicine Hannover
PD Dr. Manfred Rohde Department of Medical Microbiology
Helmholtz Centre for Infectious Research, Braunschweig
2nd Evaluation: Prof. Dr. Barbara Spellerberg
Institute of Medical Microbiology and Hygiene
University of Ulm
Date of final exam: 9th November 2011
This work was financially supported by the Deutsche Forschungsgemeinschaft
(DFG), Bonn, Germany (SFB 587).

Meiner Familie


Table of contents Chapter 1 General introduction ..........................................................................13
1. Streptococcus (S.) suis ................................................................15 1.1. S. suis infections .......................................................................................15
1.2. Pathogenesis and virulence mechanisms ................................................17
2. Cholesterol-dependent pore-forming cytolysins (CDC) ............19 2.1. The structure of CDC................................................................................19
2.2. The mechanism of pore-formation by CDC ..............................................21
2.3. The functional role of cholesterol and membrane recognition..................22
2.4. Suilysin......................................................................................................23
2.4.1. Prevalence, diversity, and regulation of the sly gene ..................23
2.4.2. Role of suilysin in host-pathogen interaction ...............................25
2.4.3. Role of suilysin in virulence and pathogenesis............................27
3. Outline of the present study.........................................................28
Chapter 2 Material and methods.........................................................................39 1. Bacterial strains and growth conditions.....................................41 2. Molecular biology and protein biochemical methods................42
2.1. Construction of mutated suilysin W461F and SVD...................................42
2.2. Expression of recombinant proteins .........................................................43
2.3. Heterologous expression of SLY, W461F and SVD in L. lactis ................43
2.4. Immunoblot analysis .................................................................................44
3. Cell culture methods.....................................................................44 3.1. Epithelial cells ...........................................................................................44
3.2. Antibiotic protection assay ........................................................................45
3.3. Labelling of bacteria for flow cytometry experiments................................46
3.4. Bacteria-cell association ...........................................................................46
3.5. Immunofluorescence microscopy .............................................................47
3.6. Double immunofluorescence microscopy (DIF)........................................48
3.7. Colocalisation experiments.......................................................................48
3.8. Field emission scanning electron microscopy (FESEM) ..........................49
3.9. Cell-permeability and macropore-formation assay ...................................49
3.10. Detection of α5β1 integrin expression on HEp-2 cells ...............................50
3.11. Haemolysis assay .....................................................................................50

3.12. Cytotoxicity assay .....................................................................................50
3.13. Pull down experiments..............................................................................51
3.14. GLISA .......................................................................................................52
4. Mouse infection model .................................................................52 4.1. Preparation of infection culture .................................................................52
4.2. Intranasal infection of mice .......................................................................52
4.3. Intravenous infection of mice ....................................................................53
4.4. Clinical score.............................................................................................53
4.5. Histopathological screening......................................................................54
4.6. Reisolation of S. suis from tissue and tracheo-nasal lavage (TNL)..........54
5. Statistical analyses .......................................................................55 Chapter 3 Results, part I: ....................................................................................57
Subcytolytic suilysin promotes invasion of Streptococcus suis in HEp-2
epithelial cells by Rac1-dependent activation of the actin cytoskeleton
Chapter 4 Results, part II:....................................................................................91
Identification of a RGD-motif in suilysin possibly involved in host cell
binding, Rac1-activation and macropore-formation
Chapter 5 Results, part III: ................................................................................115
Establishment of an intranasal CD1 mouse infection model for
colonization and invasion of Streptococcus suis serotype 2
Chapter 6 General discussion ..........................................................................153 Chapter 7 Summary ...........................................................................................163 Chapter 8 Zusammenfassung...........................................................................167 Chapter 9 Literature...........................................................................................171

List of abbreviations Δ Delta % Percent A. bidest. Aqua bidestillata A. dest. Aqua destillata ANOVA ANalysis Of VAriance ATCC American Type Culture Collection, Manassas, USA BALB/c Bagg ALBino; color locus c/c BBB Blood Brain Barrier BMEC Brain Microvascular Endothelial Cells bp Base pair(s) BSA Bovine Serum Albumin °C Degree Celsius CcpA Catabolite control protein A CDC Cholesterol-Dependent pore-forming Cytolysins Cdc42 Cell division control protein 42 homolog cf. Conferre CFSE CarboxyFluorescein Succinimidyl Ester CFU Colony Forming Units cm Centimetre CNS Central Nervous System CO2 Carbon dioxide CPS Capsule PolySaccharide Crl:CD1 (ICR) Charles River:CD1 (Institute for Cancer Research) CRM Cholesterol Recognition Motif C3 C3 toxin of Clostridium limosum Da Dalton DAPI 4',6-DiAmidino-2-PhenylIndole CD Cluster of Differentiation DMEM Dulbecco’s Modified Eagle’s Medium DNA DeoxyriboNucleic Acid DTT DiThioThreitol EDTA EthyleneDiamineTetraacetic Acid EF Extracellular Factor e.g. Exampli gratia ECM ExtraCellular Matrix et al. Et alii etc. Et cetera FBPS Fibronectin and Fibrinogen Binding Protein of S. suis FCS Fetal Calf Serum FESEM Field Emission Scanning Electron Microscopy Fig. Figure FITC Fluorescein IsoThioCyanate FL FLuorescent FSC Forward SCatter g Gravitational constant

g Gram(s) GLISA G-Protein-Linked ImmunoSorbent Assay GM17 M17 medium supplemented with 5% Glucose G-protein Guanine nucleotide-binding protein GST Glutathione S-Transferase h Hour(s) HE Hematoxylin and Eosin stain Hsd:ICR (CD1®) Harlan:Institute for Cancer Research (CD1®) HU Haemolytic Unit IgG Immunoglobulin G IPTG IsoPropylThioGalactoside IL InterLeukin ILY Intermedilysin of Streptococcus intermedius k Kilo kb Kilo base pair(s) kDa KiloDalton l Litre LB Luria Bertani LDH Lactate-Dehydrogenase LLO Listeriolysin O of Listeria monocytogenes LTA LipoTeichonic Acid M Molar (mol/l) MCP-1 Monocyte Chemotractic Protein-1 mF Milli Farad MFI Mean Fluorescent Intensity mg Milligram ml Millilitre mM Millimolar μg Microgram μl Microlitre µM Micromolar min Minute(s) MOI Multiplicity Of Infection MRP Muramidase-Released Protein ng Nano gram(s) nm Nanometer Ω Ohm O2 Oxygen ODXXX Optical Density at xxx nanometres OFS Opacity Factor of S. suis ORF Open Reading Frame PAGE PolyAcrylamide Gel Electrophoresis PBMEC Porcine Brain Microvascular Endothelial Cells PBS Phosphate Buffered Saline PCPEC Porcine Choroid Plexus Epithelial Cells PCR Polymerase Chain Reaction PFO Perfringolysin O of Clostridium perfringens

pH Power of Hydrogen PLY Pneumolysin of Streptococcus pneumoniae PMN Polymorphonuclear Neutrophil PTS PhosphoTransferase System PVDF PolyVinyliDene Fluoride Rac Ras-related C3 botulinum toxin substrate Rho Ras homolog gene family RNA RiboNucleic Acid rSLY Recombinant suilysin rSVD Recombinant RGD-SVD mutant of suilysin rW461F Recombinant W461F mutant of suilysin rpm Rounds per minute RT Room Temperature RT-PCR Reverse Transcriptase PCR ® Registered trademark s Second(s) SDS Sodium Dodecyl Sulphate SLY Suilysin SSC Sideward SCatter ST Sequence Type STSS Streptococcal Toxic Shock like Syndrome TACY Thiol-Activated CYtolysin TcdB Toxin B of Clostridium difficile TcdB-F Toxin B of Clostridium difficile Serotype F strain 1470 THB Todd Hewitt Broth Tig Trigger factor from S. suis TLR Toll-Like Receptor TM Trade mark TMH TransMembrane Hairpin TNL Tracheo-Nasal Lavage TNF-α Tumor Necrosis Factor alpha Tris Tris-(hydroxymethyl-) aminomethane TRITC TetramethylRhodamine-IsoThioCyanate U Unit V Volt wt wild type × Multiply Abbreviations of bacterial strains mentioned C. limosum Clostridium limosum C. defficile Clostridium difficile E. coli Escherischia coli L. lactis Lactococcus lactis St. aureus Staphylococcus aureus S. suis Streptococcus suis

List of figures and tables Figure 1-1: Crystal structure of suilysin and mechanism of pore-formation by CDC. ....................21
Figure 3-1: Suilysin promotes invasion of S. suis in HEp-2 epithelial cells. ...................................64
Figure 3-2: Suilysin-mediated invasion of S. suis in epithelial cells involves the actin
cytoskeleton and Rho-GTPases. .................................................................................66
Figure 3-3: Treatment of HEp-2 cells with recombinant suilysin (rSLY) leads to activation
of Rac1, but not RhoA. ................................................................................................67
Figure 3-4: Recombinant suilysin (rSLY) binds to HEp-2 cell membrane and is located in
association with F-actin and Rac1. ..............................................................................68
Figure 3-5: Substitution of an amino acid in the Trp-rich undecapeptide of domain 4 of
recombinant suilysin leads to abolishment of its haemolytic and cytolytic
activity. .........................................................................................................................70
Figure 3-6: Substitution of an amino acid in the Trp-rich undecapeptide of domain 4 of
recombinant suilysin does not affect its ability to promote HEp-2 cell
association of S. suis or to activate Rac1. ...................................................................72
Figure 4-1: Alignment of amino acid sequences of different CDC. ................................................98
Figure 4-2: Substitution of the RGD-motif of recombinant suilysin leads to abolishment
of its haemolytic and cytotoxic activity. .........................................................................99
Figure 4-3: Heterologous expression of SLY, but not W461F or SVD in L. lactis, confers
a haemolytic and cytotoxic phenotype. ..................................................................... 100
Figure 4-4: Substitution of the RGD-motif of recombinant suilysin leads to a loss in
membrane binding. . .................................................................................................. 104
Figure 4-5: Substitution of the RGD-motif leads to loss in Rac1-activation and
macropore-formation. ............................................................................................... 105

Figure 5-1: Histological findings of the nose and lung of mice infected intranasally with
5 x 109 CFU of S. suis after predisposition with 1% acetic acid. .............................. 122
Figure 6-1: Hypothetical model of suilysin-promoted interaction of S. suis with epithelial
cells............................................................................................................................ 159
Table 5-1: Clinical score sheet for health monitoring of mice after infection with S. suis. ......... 123
Table 5-2: Clinical course of CD1 mice intranasally infected with 5 x 109 CFU of S. suis
strain 10 (mrp+ epf+ sly+ cps2). ............................................................................... 124
Table 5-3: Scoring of fibrinosuppurative and purulent necrotizing lesions of mice
intranasally infected with 5 x 109 CFU of S. suis (mrp+ epf+ sly+ cps2). .................. 125
Table 5-4: Reisolation of the challenge strain S. suis strain 10 (mrp+ epf+ sly+ cps2) in
intranasally infected mice. ........................................................................................ 126
Table 5-5: Clinical course of Hsd:ICR (CD-1®) mice intranasally infected with 5 x 109
CFU of S. suis strain 10 (sly+, cps2), 10Δsly (sly-, cps2), 10cpsΔEF (sly+,
cps2-), and 10cpsΔEFΔsly (sly-, cps2-) after predisposition with 1% acetic
acid. ........................................................................................................................... 128
Table 5-6: Scoring of fibrinosuppurative and purulent necrotizing lesions of Hsd:ICR
(CD-1®) mice intranasally infected with 5 x 109 CFU of S. suis strain 10 (sly+,
cps2), 10Δsly (sly-, cps2), 10cpsΔEF (sly+, cps2-), and 10cpsΔEFΔsly (sly-,
cps2-) after predisposition with 1% acetic acid. ........................................................ 129
Table 5-7: Reisolation of the challenge strains S. suis strain 10 (sly+, cps2), 10Δsly
(sly-, cps2), 10cpsΔEF (sly+, cps2-), and 10cpsΔEFΔsly (sly-, cps2-) in
intranasally infected Hsd:ICR (CD-1®) after predisposition with 1% acetic. . ............ 130
Table 5-S1: Register of all infected mice including the clinical course of infection, scoring
of fibrinosuppurative and purulent necrotizing lesions and reisolation of the
challenge strain (table includes intravenously infected mice). ................................. 140


Chapter 1
General introduction


General introduction Chapter 1
15
1. Streptococcus (S.) suis S. suis is a gram-positive, facultative anaerobic bacterium causing invasive diseases
in swine worldwide, associated with meningitis, septicaemia, arthritis, endocarditis
and bronchopneumonia. In the last years S. suis has been considered as an
important human pathogen leading to bacterial meningitis and the life-threatening
streptococcal toxic shock like syndrome (STSS) in humans (Gottschalk et al., 2010;
Tang et al., 2006). The bacterium shows α-haemolysis on sheep blood agar plates
and α- and β-haemolysis on horse blood agar plates.
On the basis of the capsule polysaccharides, 33 different serotypes have been
described so far, of which serotype 2 is worldwide most frequently isolated from
diseased pigs and humans in Europe and Asia (Gottschalk et al., 2010; Silva et al.,
2006; Wei et al., 2009). Distribution of serotypes differs between geographical
regions. Serotype 9 has emerged as the most common pig isolate in Germany and
The Netherlands, serotype 7 is most prevalent in Scandinavia and Germany, and
serotype 1 and 14 in the United Kingdom (Baums and Valentin-Weigand, 2009;
Perch et al., 1983; Tian et al., 2004; Wisselink et al., 2000). In contrast, in Canada
and the USA serotypes 2, 1/2 and 3 are most frequently associated with disease
(Messier et al., 2008). Noteworthy, a specific sequence type (ST), namely ST7,
evolved from the highly pathogenic ST1 type of a serotype 2 strain, which was found
to be responsible for human outbreaks in china and directly associated with the
STSS (Ye et al., 2006; Ye et al., 2009). The ST7 carries a putative pathogenicity
island (designated 89K), possibly involved in development of STSS (Chen et al.,
2007; Zhao et al., 2011).
1.1. S. suis infections S. suis can infect pigs of each age group, including suckling and weaning piglets as
well as growers. The natural habitat of S. suis is the upper respiratory tract. S. suis
colonizes the nasopharynx, in particular the tonsils and the nasal cavities, as well as
other mucosal surfaces like the intestinal and genital tract asymptomatically, resulting
in a high carrier rate of healthy pigs of up to 100% (Arends et al., 1984; Clifton-
Hadley et al., 1986; Higgins and Gottschalk, 1990; Lowe et al., 2011; O'Sullivan et

General introduction Chapter 1
16
al., 2011). Such carrier-pigs are the most important source of infection (Clifton-
Hadley and Alexander, 1980). Horizontal transmission of disease occurs most
frequently via the direct 'nose-to-nose' contact between healthy and infected pigs,
besides airborne transmission of S. suis has also been shown in experimental
infected piglets (Berthelot-Herault et al., 2001). Oral infection (feed) and transmission
via skin wounds is also possible. Vertical transmission of S. suis via the navel or the
genital tract is another relevant route of infection (Amass et al., 1997; Robertson et
al., 1991; Staats et al., 1997). Furthermore, insects (houseflies) as potential vectors
have been discussed (Enright et al., 1987).
Although the morbidity rate of pigs is less than 5% due to prophylaxis with antibiotics
(Clifton-Hadley et al., 1986), in the case of disease the mortality rate can reach 20%
in the absence of treatment. Several forms of streptococcal disease in pigs are
known. The occurrence of sudden death due to a peracute septicaemia as well as
the development of severe meningitis, polyarthritis and bronchopneumonia can be
observed. Acute infections are indicated by high fever (>40˚C), persistent anorexia
followed by lameness and central nervous failure (Sanford and Tilker, 1982; Windsor
and Elliott, 1975). Typical histopathological lesions are characterized by an acute
fibrinosuppurative inflammation of the respective tissue (Beineke et al., 2008;
Williams and Blakemore, 1990).
As an important emerging zoonotic agent S. suis has gained public interest due to
increased reports on human infections (Gottschalk et al., 2007; Trottier et al., 1991;
Wertheim et al., 2009). To date human cases of S. suis infections are mainly
reported in Asia. In Vietnam S. suis is considered as the most common causative
pathogen of bacterial meningitis (second most common in Thailand and third most
frequent in Hong Kong) (Mai et al., 2008; Petersen et al., 2011; Sriskandan and
Slater, 2006; Wangkaew et al., 2006). Particularly humans exposed to infected pigs
or contaminated pig-products are at risk (Arends and Zanen, 1988). An additional
potential source of infection for humans are wild boars (Baums et al., 2007).
Therefore, S. suis is an occupational disease of people in close contact with swine,
like farmers, butcher, hunters and veterinarians (high-risk group). Hence, the
nasopharyngeal carrier rate of S. suis serotype 2 strains of persons belonging to the

General introduction Chapter 1
17
high-risk group in Germany was 5.3% (Strangmann et al., 2002). For humans without
swine contact oral transmission via raw pork or contaminated pig-products is
possible, whereas a 'human-to-human' infection has not been proven so far
(Wertheim et al., 2009). After an incubation period ranging from a few hours to five
days post infectionem, purulent meningitis, septicaemia and arthritis associated with
leukocytosis and neutrophilia are the most common manifestations in humans
(Arends and Zanen, 1988; Fongcom et al., 2001). A serious consequence following
S. suis meningitis is chronic deafness (Navacharoen et al., 2009). Two human
outbreaks in China in 1998 and 2005 were associated with increased severeness of
clinical symptoms. A noticeable high incidence of the STSS, which is characterized
by high fever, erythoderma and multi organ failure (liver, heart, kidney, CNS) was
observed, resulting in a high mortality rate of more than 20% (Tang et al., 2006; Yu et
al., 2006).
1.2. Pathogenesis and virulence mechanisms The mechanisms underlying pathogenesis of S. suis infections are only poorly
known. A hypothetical model of pathogenesis includes three main steps. Firstly, after
colonisation the mucosal surface of the upper respiratory tract, S. suis invades into
deeper tissues of the epithelium. Secondly, the bacterium disseminates within the
bloodstream to finally cross the endothelium of target tissues, such as the blood brain
barrier (BBB) of the central nervous system (CNS) to cause meningitis (Chanter et
al., 1993; Gottschalk and Segura, 2000). S. suis produces a wide array of virulence
and virulence-associated factors, either secreted or surface-associated, involved in
this process. A comprehensive review of bacterial factors expressed by S. suis was
recently published (Baums and Valentin-Weigand, 2009).
Bacterial factors such as the fibronectin and fibrinogen binding protein (FBPS) (de
Greeff et al., 2002) or the cell wall component lipoteichonic acid (LTA) (Fittipaldi et
al., 2007; Vanier et al., 2007) mediate adherence of bacteria to target cells for initial
colonisation. Another bacterial mechanism for sufficient colonisation is the formation
of biofilms probably enhancing bacterial resistance to innate and adaptive host
defence mechanisms and treatment with antibiotics (Bonifait et al., 2008). To get

General introduction Chapter 1
18
access into deeper tissues bacteria might invade the respiratory epithelium. Suilysin,
the haemolysin of S. suis, is discussed to play a role in interaction of S. suis with
epithelial cells and disruption of these cells due to its cytolytic function (cf. 2.4.2.).
Furthermore, the capsule is assumed to be involved in host cell interaction. Since its
main function is protection against phagocytosis after entering the bloodstream
(Benga et al., 2008; Chabot-Roy et al., 2006; Charland et al., 1998; Segura and
Gottschalk, 2002), it has been proposed that the capsule is down-regulated during
colonisation of the mucosal epithelium to allow adherence and invasion of the
bacterium to overcome this first barrier within the host (Gottschalk and Segura, 2000;
Okamoto et al., 2004; Willenborg et al., 2011). In accordance, unencapsulated
S. suis stains showed higher adhesion and invasion rates, indicating a negative
correlation between encapsulation and interaction with host cells (Benga et al., 2004;
Gottschalk et al., 1991). A possible explanation for this phenotype is the masking
effect of the capsule, whereby potentially involved surface associated proteins or cell
wall components might be hidden (Lalonde et al., 2000; Tenenbaum et al., 2008;
Vanier et al., 2007). Moreover, a direct uptake of S. suis by monocytes for crossing
the epithelium as well as for entering the bloodstream within circulating cells, known
as the 'Trojan horse theory', is controversially discussed due to the protecting effect
of the capsule. A 'travelling' of either free bacteria or monocyte-associated bacteria
('modified Trojan horse theory') is more likely (Gottschalk and Segura, 2000).
Circulation of bacteria within the bloodstream may lead to onset of acute bacteraemia
or septicaemia and the release of several pro-inflammatory cytokines by cells of the
innate immune system to control acute infection or to contribute to immunopathology
(Segura et al., 1999; Segura et al., 2002; Segura et al., 2006).
Nevertheless, for induction of meningitis S. suis has to penetrate the BBB to reach
the CNS. S. suis has the ability to adhere to and invade into brain microvascular
endothelial cells (BMEC) and porcine choroid plexus epithelial cells (PCPEC), the
main components of the BBB (Benga et al., 2005; Charland et al., 2000; Tenenbaum
et al., 2005; Tenenbaum et al., 2008; Vanier et al., 2004). Moreover, an increase in
tight junction permeability and loss of barrier function is ascribed to direct cytotoxic
effects of suilysin (Charland et al., 1998; Vanier et al., 2004). Apart from suilysin

General introduction Chapter 1
19
S. suis can stimulate the production of pro-inflammatory cytokines like interleukin-6
(IL-6), IL-8 and monocyte chemotactic protein-1 (MCP-1) by BMEC, which in turn
alters BBB permeability (Vadeboncoeur and Pelletier, 1997). However, Tenenbaum
et al. (2009) described the entry S. suis into the CNS as a transcellular translocation
without destruction of PCPEC lining of the BBB.
2. Cholesterol-dependent pore-forming cytolysins (CDC) Suilysin, the secreted haemolysin of S. suis, belongs to the family of cholesterol-
dependent pore-forming cytolysins (CDC) and was considered as a virulence-
associated factor, which contributes to pathogenesis of S. suis. CDC are a large
family of membrane-damaging toxins produced by more than 20 gram-positive
bacteria (Tweten, 2005), including the genera Streptococcus, Listeria, Bacillus,
Clostridum, Paenibacillus, Arcanobacterium and most recently Gardnerella (Gelber et
al., 2008) and Lactobacillus (Rampersaud et al., 2011).
2.1. The structure of CDC All common CDC show a high similarity in primary amino acid sequence varying
between 40-70% identity. CDC are single stranded peptides showing an elongated
rod-like three-dimensional structure, which was first described for the prototype of
CDC, perfringolysin O of Clostrium perfringens.
CDC were previously named thiol-activated cytolysins (TACY) due to the fact, that
most members carry a single cysteine residue within the undecapeptide at the
carboxy-terminus of the molecule. This cysteine residue is believed to be essential
for membrane binding and the cytolytic function. It is known that oxidation of carbon-
bonded sulfohydryl groups (formation of disulfide bonds) inhibits CDC. The name
'thiol-activated' derives from the properties of thiols, which are able to reactivate the
toxin via the cleavage of disulfide bonds of the cysteine residue. However, two
members of the toxin family, pyolysin (Arcanobacterium pyogenes) (Billington et al.,
1997) and intermedilysin (Streptococcus intermedius) carry an alanin residue instead
of the cysteine without any deficiency in lysis activity (Herbert and Todd, 1941).
Moreover, mutation studies targeting the cysteine residue did not alter the cytolysis

General introduction Chapter 1
20
process (Michel et al., 1990; Pinkney et al., 1989; Saunders et al., 1989; Stachowiak
et al., 2009). In contrast to the cytolytic activity, cysteine substitution of listeriolysin O
decreased invasion of Listieria monocytogenes (Stachowiak et al., 2009).
Nevertheless, these findings clearly show, that the cysteine residue is not required
for cytolytic function of the toxin, therefore the name 'thiol-activated' is no longer
appropriate.
The monomeric protein contains four domains (D1-D4, Figure 1-1A): D1 and D3 are
located at the N-terminus of the protein, linked to D4 via the connecting domain D2
(Rossjohn et al., 1997). The main part of the protein is formed by β-strands (43%),
whereas in particular D3 also contains a couple of α-helixes (14% α-helices of the
whole structure), playing a special role in pore-formation (Xu et al., 2010). The most
conserved regions are the C-terminal tryptophan-rich undecapeptide (Figure 1-1A),
consisting of eleven amino acids, (ECTGLAWEWWR; c.f. Chapter 4, Figure 4-2) of
D4 and the hydrophobic core of D1. The undecapeptide is considered to be
responsible for the initial membrane binding and triggers the formation of functional
pores (Rossjohn et al., 1997). Supportively, it has been shown, that recombinant D4
alone is still able to bind to membranes of erythrocytes (Weis and Palmer, 2001). In a
recent study the crystal structure and D4-folding of CDC, including suilysin,
intermedilysin, perfringolysin O and anthrolysin (Bacillus anthracis), was analyzed in
more detail. Bending-degree of the angle between D1 and D4 as well as the
conformations of the tryptophan-rich loop on the tip of the undecapeptide were
compared. Predictions of intermedilysin and suilysin revealed a more extended
structure (Figure 1-1A), whereas the undecapeptide of perfringolysin O and
anthrolysin was folded back into a hydrophobic pocket (Xu et al., 2010).

General introduction Chapter 1
21
Figure 1-1: Crystal structure of suilysin and the mechanism of pore-formation by CDC. (A) Ribbon diagram of suilysin crystal structure modified from Xu et al., 2010. Each domain (D1-D4) is coloured individually and the location of the tryptophan-rich undecapeptide, the two transmembrane β-hairpins (THM1 and THM2) and the three loops (L1-3) are indicated by arrows. (B) The putative CDC pore-forming mechanism modified from Tweten et al., 2005. The CDC molecule is secreted as a soluble monomer consisting of 4 domains (D1-D4) and binds to cholesterol-containing target membranes via D4 (1). Several monomers interact to form the arc-shaped prepore complex triggered by structural changes within D3 (2), followed by formation of the preinsertion transmembrane β-barrel structure (TMH1 and 2) (3). Upon collapse of D2 the oligomer inserts into the membrane generating a permeable pore (4). (C) 3D-reconstruction of electronmicroscopy images of liposomes with the prepore complex and the permeable pore of pneumolysin modified from Tilley et al., 2005.
2.2. The mechanism of pore-formation by CDC All members of the CDC family are secreted as water soluble monomers due to a
signal sequence at the N-terminus of the protein, with the exception of pneumolysin
(Streptococcus pneumoniae), which is not released until lysis of the bacterial cell
(Walker et al., 1987). A common, but still putative, mechanism of pore-formation was
described by Tweten et al., 2005. Generation of channels is a 4-step process (Figure
1-1B) (Tweten, 2005). (1) Initially several monomers bind vertically to the target cell
membrane. Interaction between the CDC molecule and the cell is mediated via D4,
forming a hydrophobic 'danger' consisting of the undecapeptide, which superficially

General introduction Chapter 1
22
inserts into the membrane (Rossjohn et al., 1998). Binding of D4 to the membrane is
necessary for subsequent structural changes within the not directly connected D3,
called 'conformational coupling' (Rossjohn et al., 2007). Subsequently, cell-bound
monomers undergo a lateral shift to form an oligomer. (2) Further structural changes
within D3 lead to the formation of the oligomeric ring- or arc-shaped prepore complex
(Figure 1-1C). (3) The N-terminal D3 builds the inner structure of the transmembrane
pore. Therefore, six α-helices form two β-hairpins (Figure 1-1A; TMH1 and TMH2).
This ultimate β-barrel structure consists of several β-hairpins of agminated
monomers, which are connected through an intermolecular interaction of the
β1-strand and β4-strand of two individual molecules (Ramachandran et al., 2004). (4)
In the final step, the connecting domain D2 severs as a 'hinge-joint' to bring D3 in
close contact with the cell surface. After the 'collapse' of D2 the β-barrel structure
inserts into the membrane and forms the transmembrane pore. This pore, 30 nm in
size, consists of 35-50 monomers and is permeable for ions and macro molecules
(Figure 1-1C).
2.3. The functional role of cholesterol and membrane recognition The role of cholesterol in CDC function is not finally clarified (Hotze and Tweten,
2011). Nevertheless, a general observation is that cholesterol is required for pore-
formation and free cholesterol inhibits haemolysis of CDC (Alouf, 2000; Jacobs et al.,
1994; Watson et al., 1972). Furthermore, cholesterol-depletion studies using
cholesterol-containing liposomes showed that >30mol% of total membrane lipids has
to be cholesterol for efficient membrane binding (Flanagan et al., 2009; Heuck et al.,
2000). However, not all CDC use cholesterol as a membrane receptor. It has been
shown that the human complement regulator molecule CD59 is required for
membrane binding of intermedilysin and vaginolysin, thereby conferring a species
specificity. However, cell lysis activity of intermedilysin and vaginolysin depends on
cholesterol (Gelber et al., 2008; Giddings et al., 2004).
The cholesterol recognition motif (CRM) is most likely located within D4. Several
studies concerning the CRM defined the undecapeptide to be responsible for
cholesterol binding (Rossjohn et al., 1997; Rossjohn et al., 2007). In contrast, more

General introduction Chapter 1
23
recent studies revealed three highly conserved loops (Figure 1-1A; L1-3) located next
to the undecapeptide to mediate recognition of cholesterol (Soltani et al., 2007;
Soltani et al., 2007b). Furthermore, it has been suggested that CDC present only a
single binding side due to the fact that cholesterol binding activity is linear to CDC
concentration (Johnson et al., 1980).
2.4. Suilysin Suilysin was identified as a haemolysin of S. suis several years ago (Jacobs et al.,
1994) and confirmed to be a member of CDC by sequence analysis. The protein has
a molecular weight of 54 kDa and possesses an N-terminal signal sequence, thus
considered as a secreted exotoxin. Suilysin is most related to pneumolysin, sharing
52% amino acid identity (Segers et al., 1998).
2.4.1. Prevalence, diversity, and regulation of the sly gene Suilysin is expressed by many but not all S. suis strains. The sly gene has been
detected in 95% of European and Asian invasive serotype 2 strains, but only in 7% of
the North American strains (Segers et al., 1998). In various studies concerning the
prevalence of the sly gene in isolates, belonging to different S. suis serotypes,
obtained from diseased pigs in European countries, including Germany, Italy, Spain,
Poland, France, The Netherlands and The United Kingdom, the sly gene was
detectable in a range between 33.7% and 100% (Berthelot-Herault et al., 2000;
Blume et al., 2009; de Greeff et al., 2011; Fabisiak et al., 2005; King et al., 2001;
Princivalli et al., 2009; Silva et al., 2006; Tarradas et al., 2001). Approximately 80%
of these clinical isolates were obtained from porcine cases of invasive disease
associated with meningitis, septicaemia, and arthritis. Isolates derived from lung
samples (pneumonia) were positive tested for suilysin to a lesser degree of
approximately 50% (King et al., 2001; Silva et al., 2006). Furthermore, the sly gene is
prevalent in up to 90% of colonising S. suis strains isolated from the tonsils of healthy
pigs (Fabisiak et al., 2005; King et al., 2001). Besides, suilysin is present in wild
boars and domestic swine S. suis strains of Northwestern Germany in 39% and 66%,
respectively (Baums et al., 2007). In Asian S. suis strains, isolated from healthy

General introduction Chapter 1
24
(slaughtered) as well as from diseased swine, the prevalence of suilysin reaches a
maximum of 100% (Hoa et al., 2011; Kim et al., 2010; Padungtod et al., 2010; Wei et
al., 2009; Xiong et al., 2007). Equally, almost all isolates from human cases of a
S. suis infection carry a sly gene (de Greeff et al., 2011; King et al., 2001; Princivalli
et al., 2009). The situation is different for North American isolates, in which suilysin is
less frequently present (Fittipaldi et al., 2009; Gottschalk et al., 1998).
However, presence of the sly gene has been used for characterisation and
differentiation of S. suis strains (Gottschalk et al., 2007; Vecht et al., 1991), but
presence of the sly gene does not necessarily result in protein expression (de Greeff
et al., 2011).
Suilysin was detected as a highly conserved single copy gene within the S. suis
genome (Okwumabua et al., 1999). Furthermore, the genetic diversity as well as the
allelic variance of the sly gene appears to be low (King et al., 2001). The flanking
genes (2 open reading frames (ORF) upstream, orf100 and orf101; and 2 ORF
downstream, nanE and pstG) are also highly conserved. Sly-negative strains exhibit
an alternative gene (orf102) at the same position instead of the sly gene. Therefore,
genetic exchange via homologous recombination between different S. suis strain is
most likely (Takamatsu et al., 2002). Moreover, it is conceivable that the sly gene is
not organised as an operon and under the control of its own promoter, because of
large non coding region upstream and downstream of the sly gene.
Little is known about the regulation of the sly gene. The protein is expressed at late
logarithmic growth phase, possibly dependent on nutrient availability, pH and
bacterial density (Gottschalk et al., 1995). A hyper-haemolytic phenotype is
described for a manN-negative S. suis mutant strain, suggesting that the mannose
phosphotransferase system affects the suilysin promoter activity (Lun et al., 2003).
The global orphon response regulator CovR (control of virulence regulator) controls
the expression of about 200 genes, including the capsule biosynthesis and suilysin.
Inactivation of covR led to the production of a thicker capsule and slightly higher
haemolytic activity associated with increased adhesion to epithelial cells and reduced
phagocytosis and killing by polymorphonuclear neutrophils (PMN) (Pan et al., 2009).
In contrast to other CDC, like intermedilysin, which is under the transcriptional

General introduction Chapter 1
25
catabolite control protein A (CcpA), the homologous global regulator of S. suis is
most likely not involved in regulation of suilysin (Tomoyasu et al., 2010; Willenborg et
al., 2011). Besides, expression of the sly gene is influenced by the trigger factor from
S. suis (Tig) and the orphan response regulator RevSC21. Both, deletion of the tig
gene and the RevSC21 gene degraded expression of suilysin and resulted in a lack
of haemolytic activity of the respective S. suis strain (Wu et al., 2009; Wu et al.,
2011).
2.4.2. Role of suilysin in host-pathogen interaction Similar to other members of the CDC family suilysin can damage different types of
host cells by its cytolytic activity (Benga et al., 2004; Charland et al., 2000; Jacobs et
al., 1994; Lalonde et al., 2000; Norton et al., 1999; Segura and Gottschalk, 2002;
Tenenbaum et al., 2005; Tenenbaum et al., 2006; Vanier et al., 2004). A suilysin-
induced cell injury is associated with loss of cytoplasmic density, disruption of
cytoplasmic membranes and the release of cellular contents (Allen et al., 2001;
Segura and Gottschalk, 2002). Haemolysis of erythrocytes was first described by
Jacobs et al. (1994), whereas human red blood cells were the most susceptible,
followed by horse, sheep, and cow erythrocytes (Gottschalk et al., 1995).
A multifunctional role in pathogenesis was described for other members of the CDC-
family. These biological effects can be observed even at subcytolytic concentration of
the respective toxins (Billington et al., 2000). For pneumolysin, listeriolysin O, and
intermedilysin was reported that they may contribute to bacterial adherence and
invasion (Cockeran et al., 2002; Krawczyk-Balska and Bielecki, 2005; Rubins et al.,
1998; Sukeno et al., 2005). Likewise, suilysin has been described to be involved in
the modulation of S. suis host cell interaction, including endothelial cells, epithelial
cells, PMN and macrophages (Baums and Valentin-Weigand, 2009; Gottschalk and
Segura, 2000). In particular, it has been suggested that suilysin plays a role in
pathogenesis of S. suis (Norton et al., 1999) such as the crossing of the BBB by
interruption of intracellular junctions for entering the CNS (Charland et al., 2000;
Tenenbaum et al., 2005; Tenenbaum et al., 2008). However, the ability of several

General introduction Chapter 1
26
S. suis strains to interact with PBMEC does not correlate with suilysin production
(Vanier et al., 2004; Vanier et al., 2007).
More recently, it has been found that the toxin may be involved in cytokine release
and protection against opsonophagocytosis. Thus, a reduced survival time of a
sly-deficient mutant strain after co-incubation with polymorphonuclear neutrophils
(PMN) was observed by Benga et al. (2008). This was confirmed by using antisera
raised against purified suilysin, which increased the uptake of the wild type strain by
PMN. In addition, adding recombinant suilysin at subcytolytic concentration increased
the growth capacity of the sly-deficient mutant (Benga et al., 2008). Furthermore,
suilysin contributes to resistance of complement-dependent killing of S. suis by
neutrophils (Chabot-Roy et al., 2006), perhaps by impairing complement factor C3
deposition on the surface of S. suis (Lecours et al., 2011). For pneumolysin,
reduction of serum complement levels and decreased opsonisation of pneumococci
has been described as well (Alcantara et al., 2001). Although, S. suis was found to
be resistant to phagocytosis by murine astrocytes, suilysin was mainly responsible
for pro-inflammatory cytokine production and partially involved in toll-like receptor 2
(TLR2) expression of these cells (Zheng et al., 2011). Similarly, recognition of
pneumolysin via the TLR4 is critically involved in the innate immune response to
pneumococci (Dessing et al., 2009; Malley et al., 2003).
In general, interference of suilysin with different types of immune cells and induction
of cytokine release suggests the important role of suilysin in host innate defence
response. Suilysin is responsible for the release of IL-6 and IL-8 by BMEC
(Vadeboncoeur et al., 2003) and PBMEC (Vanier et al., 2008) as well as for the
production of tumour necrosis factor α (TNF-α) by human monocytes and IL-6 by
porcine pig pulmonary alveolar macrophages and monocytes (Lun et al., 2003). In
contrast, suilysin did not have a major impact on phagocytosis as well as on TNF-α
and MCP-1 production by murine microglia (Dominguez-Punaro et al., 2010).
Accordingly, suilysin failed to induce TNF-α and IL-6 in murine macrophage line J774
(Segura et al., 1999) and plays a limited role in modulation of cytokines and
chemokine response in a whole-blood system (Segura et al., 2006). An involvement
in cytokine production by bone marrow–derived dendritic cells was recently described

General introduction Chapter 1
27
by (Lecours et al., 2011). In inflammatory events increased recruitment of leucocytes
is linked to adhesion molecules. Stimulation of THP-1 monocytes with suilysin led to
an up-regulation of CD11a/CD18 and CD11c/CD18 (Al Numani et al., 2003). Most of
the other CDC have also been shown to display a role in modulation of immune
response mechanisms (Cockeran et al., 2002; Ratner et al., 2006; Tsuchiya et al.,
2005).
2.4.3 Role of suilysin in virulence and pathogenesis The importance of suilysin in the pathogenesis of S. suis is not finally clarified. While
suilysin has been shown to be associated with virulent strains, there are also virulent
strains that do not produce suilysin (Staats et al., 1999). Only few experimental infections of mice and piglets addressing the role of suilysin
were performed so far. Intraperitonally infection of BALB/c mice demonstrated
attenuation of a sly-deficient mutant in comparison to the highly virulent S. suis
serotype 2 strain P1/7. In contrast, the sly knock-out strain was only slightly
attenuated (reduced severeness of clinical signs and pathological findings) in an
intravenous porcine infection model (Allen et al., 2001), indicating that suilysin does
not play a major role in disease development after systemic administration. Similar
results were obtained by Lun et al. (2003) using three different sly-deficient strains in
an intrapharyngeal piglet infection model. All swine developed clinical symptoms
associated with septicaemia, arthritis, and meningitis regardless of the challenge
strain. However, vaccination containing purified suilysin was capable to induce
protection in BALB/c mice and piglets after challenge with a homologous strain
(S. suis P1/7). Furthermore, immunisation led to an increase in haemolysin
neutralisation antibody titre (Jacobs et al., 1994; Jacobs et al., 1996). Additionally,
intranasal immunisation of piglets with a S. suis live vaccine elicited most prominently
serum immunoglobulin G responses against suilysin (Kock et al., 2009). Likewise,
the highly related pneumolysin protects mice against homologues challenge and
moreover a pneumolysin-deficient mutant was attenuated in a BALB/c infection
model (Alexander et al., 1994; Orihuela et al., 2004).

General introduction Chapter 1
28
3. Outline of the present study As pointed out in the general introduction, knowledge on pathogenesis of S. suis
infection and potentially involved bacterial factors is still limited. The aim of this study
was to evaluate suilysin-mediated effects on S. suis-epithelial cell interaction
including investigations on the underlying mechanisms and the phenotypical
characterisation of putatively involved functional domains of the suilysin molecule.
Furthermore, the study focussed on the development of an intranasal mouse
infection model to further elucidate the role of suilysin in colonisation and invasion of
the upper respiratory tract in vivo.
According to these objectives, the results are divided into 3 chapters. In chapter 3
the role of suilysin in S. suis invasion in epithelial cells is investigated, revealing a
subcytolytic activity of suilysin in a Rac-dependent activation of the actin cytoskeleton
promoting invasion of S. suis. Chapter 4 comprises the characterisation of two
(functional) domains within the suilysin molecule. Site-directed amino acid
substitution and comparative functional analysis of the tryptophan-rich undecapeptide
and a putative integrin-binding RGD-motif indicate that both domains are required for
cytolytic function. Additionally, a functional RGD-motif is essential for membrane-
binding and activation of Rac. Finally, in chapter 5 the establishment of an intranasal
S. suis mouse infection model is described. The results are generally discussed in
chapter 6 with regard to relevance for S. suis pathogenesis. A short summary of this
thesis is provided in chapter 7 (English) and chapter 8 (German).

General introduction Chapter 1
29
References Al Numani, D., Segura, M., Dore, M., and Gottschalk, M. (2003) Up-regulation of ICAM-1, CD11a/CD18 and CD11c/CD18 on human THP-1 monocytes stimulated by Streptococcus suis serotype 2. Clin Exp Immunol 133: 67-77. Alcantara, R. B., Preheim, L. C., and Gentry-Nielsen, M. J. (2001) Pneumolysin-induced complement depletion during experimental pneumococcal bacteremia. Infect Immun 69: 3569-3575.
Alexander, J. E., Lock, R. A., Peeters, C. C., Poolman, J. T., Andrew, P. W., Mitchell, T. J., Hansman, D., and Paton, J. C. (1994) Immunization of mice with pneumolysin toxoid confers a significant degree of protection against at least nine serotypes of Streptococcus pneumoniae. Infect Immun 62: 5683-5688.
Allen, A. G., Bolitho, S., Lindsay, H., Khan, S., Bryant, C., Norton, P., Ward, P., Leigh, J., Morgan, J., Riches, H., Eastty, S., and Maskell, D. (2001) Generation and characterization of a defined mutant of Streptococcus suis lacking suilysin. Infect Immun 69: 2732-2735.
Alouf, J. E. (2000) Cholesterol-binding cytolytic protein toxins. Int J Med Microbiol 290: 351-356.
Amass, S. F., SanMiguel, P., and Clark, L. K. (1997) Demonstration of vertical transmission of Streptococcus suis in swine by genomic fingerprinting. J Clin Microbiol 35: 1595-1596.
Arends, J. P., Hartwig, N., Rudolphy, M., and Zanen, H. C. (1984) Carrier Rate of Streptococcus-Suis Capsular Type-2 in Palatine Tonsils of Slaughtered Pigs. Journal of Clinical Microbiology 20: 945-947.
Arends, J. P. and Zanen, H. C. (1988) Meningitis Caused by Streptococcus Suis in Humans. Reviews of Infectious Diseases 10: 131-137.
Baums, C. G. and Valentin-Weigand, P. (2009) Surface-associated and secreted factors of Streptococcus suis in epidemiology, pathogenesis and vaccine development. Anim Health Res Rev 10: 65-83.
Baums, C. G., Verkuhlen, G. J., Rehm, T., Silva, L. M., Beyerbach, M., Pohlmeyer, K., and Valentin- Weigand, P. (2007) Prevalence of Streptococcus suis genotypes in wild boars of Northwestern Germany. Appl Environ Microbiol 73: 711-717.
Beineke, A., Bennecke, K., Neis, C., Schroder, C., Waldmann, K. H., Baumgartner, W., Valentin- Weigand, P., and Baums, C. G. (2008) Comparative evaluation of virulence and pathology of Streptococcus suis serotypes 2 and 9 in experimentally infected growers. Vet Microbiol 128: 423-430.
Benga, L., Friedl, P., and Valentin-Weigand, P. (2005) Adherence of Streptococcus suis to porcine endothelial cells. J Vet Med B Infect Dis Vet Public Health 52: 392-395.
Benga, L., Fulde, M., Neis, C., Goethe, R., and Valentin-Weigand, P. (2008) Polysaccharide capsule and suilysin contribute to extracellular survival of Streptococcus suis co-cultivated with primary porcine phagocytes. Vet Microbiol 132: 211-219.
Benga, L., Goethe, R., Rohde, M., and Valentin-Weigand, P. (2004) Non-encapsulated strains reveal novel insights in invasion and survival of Streptococcus suis in epithelial cells. Cellular Microbiology 6: 867-881.
Berthelot-Herault, F., Gottschalk, M., Labbe, A., Cariolet, R., and Kobisch, M. (2001) Experimental airborne transmission of Streptococcus suis capsular type 2 in pigs. Vet Microbiol 82: 69-80.

General introduction Chapter 1
30
Berthelot-Herault, F., Morvan, H., Keribin, A. M., Gottschalk, M., and Kobisch, M. (2000) Production of Muraminidase-Released Protein (MRP), Extracellular Factor (EF) and Suilysin by field isolates of Streptococcus suis capsular types 2, 1/2, 9, 7 and 3 isolated from swine in France. Veterinary Research 31: 473-479.
Billington, S. J., Jost, B. H., Cuevas, W. A., Bright, K. R., and Songer, J. G. (1997) The Arcanobacterium (Actinomyces) pyogenes hemolysin, pyolysin, is a novel member of the thiol- activated cytolysin family. J Bacteriol 179: 6100-6106.
Billington, S. J., Jost, B. H., and Songer, J. G. (2000) Thiol-activated cytolysins: structure, function and role in pathogenesis. FEMS Microbiol Lett 182: 197-205.
Blume, V., Luque, I., Vela, A. I., Borge, C., Maldonado, A., Dominguez, L., Tarradas, C., and Fernandez-Garayzabal, J. F. (2009) Genetic and virulence-phenotype characterization of serotypes 2 and 9 of Streptococcus suis swine isolates. Int Microbiol 12: 161-166.
Bonifait, L., Grignon, L., and Grenier, D. (2008) Fibrinogen induces biofilm formation by Streptococcus suis and enhances its antibiotic resistance. Appl Environ Microbiol 74: 4969-4972.
Chabot-Roy, G., Willson, P., Segura, M., Lacouture, S., and Gottschalk, M. (2006) Phagocytosis and killing of Streptococcus suis by porcine neutrophils. Microb Pathog 41: 21-32.
Chanter, N., Jones, P. W., and Alexander, T. J. (1993) Meningitis in pigs caused by Streptococcus suis--a speculative review. Vet Microbiol 36: 39-55.
Charland, N., Harel, J., Kobisch, M., Lacasse, S., and Gottschalk, M. (1998) Streptococcus suis serotype 2 mutants deficient in capsular expression. Microbiology-Sgm 144: 325-332. Charland, N., Nizet, V., Rubens, C. E., Kim, K. S., Lacouture, S., and Gottschalk, M. (2000) Streptococcus suis serotype 2 interactions with human brain microvascular endothelial cells. Infect Immun 68: 637-643.
Chen, C., Tang, J., Dong, W., Wang, C., Feng, Y., Wang, J., Zheng, F., Pan, X., Liu, D., Li, M., Song, Y., Zhu, X., Sun, H., Feng, T., Guo, Z., Ju, A., Ge, J., Dong, Y., Sun, W., Jiang, Y., Wang, J., Yan, J., Yang, H., Wang, X., Gao, G. F., Yang, R., Wang, J., and Yu, J. (2007) A glimpse of streptococcal toxic shock syndrome from comparative genomics of S. suis 2 Chinese isolates. PLoS ONE 2: e315.
Clifton-Hadley, F. A. and Alexander, T. J. (1980) The carrier site and carrier rate of Streptococcus suis type II in pigs. Vet Rec 107: 40-41.
Clifton-Hadley, F. A., Alexander, T. J., and Enright, M. R. (1986) Monitoring herds for Streptococcus suis type 2: chance contamination of slaughter pigs. Vet Rec 118: 274.
Cockeran, R., Durandt, C., Feldman, C., Mitchell, T. J., and Anderson, R. (2002) Pneumolysin activates the synthesis and release of interleukin-8 by human neutrophils in vitro. J Infect Dis 186: 562-565.
de Greeff, A., Buys, H., Verhaar, R., Dijkstra, J., van Alphen, L., and Smith, H. E. (2002) Contribution of fibronectin-binding protein to pathogenesis of Streptococcus suis serotype 2. Infect Immun 70: 1319-1325.
de Greeff, A., Wisselink, H. J., de Bree, F. M., Schultsz, C., Baums, C. G., Thi, H. N., Stockhofe- Zurwieden, N., and Smith, H. E. (2011) Genetic diversity of Streptococcus suis isolates as determined by comparative genome hybridization. BMC Microbiol 11: 161.

General introduction Chapter 1
31
Dessing, M. C., Hirst, R. A., de Vos, A. F., and van der, Poll T. (2009) Role of Toll-like receptors 2 and 4 in pulmonary inflammation and injury induced by pneumolysin in mice. PLoS One 4: e7993.
Dominguez-Punaro, Mde L., Segura, M., Contreras, I., Lachance, C., Houde, M., Lecours, M. P., Olivier, M., and Gottschalk, M. (2010) In vitro characterization of the microglial inflammatory response to Streptococcus suis, an important emerging zoonotic agent of meningitis. Infect Immun 78: 5074-5085.
Enright, M. R., Alexander, T. J., and Clifton-Hadley, F. A. (1987) Role of houseflies (Musca domestica) in the epidemiology of Streptococcus suis type 2. Vet Rec 121: 132-133.
Fabisiak, M., Kita, J., Jedryczko, R., and Binek, M. (2005) Prevalence of the suilysin gene in Streptococcus suis strains isolated from diseased and healthy carrier pigs. Pol J Vet Sci 8: 141-145.
Feldman, C., Mitchell, T. J., Andrew, P. W., Boulnois, G. J., Read, R. C., Todd, H. C., Cole, P. J., and Wilson, R. (1990) The effect of Streptococcus pneumoniae pneumolysin on human respiratory epithelium in vitro. Microb Pathog 9: 275-284.
Fittipaldi, N., Fuller, T. E., Teel, J. F., Wilson, T. L., Wolfram, T. J., Lowery, D. E., and Gottschalk, M. (2009) Serotype distribution and production of muramidase-released protein, extracellular factor and suilysin by field strains of Streptococcus suis isolated in the United States. Vet Microbiol 139: 310-317.
Fittipaldi, N., Gottschalk, M., Vanier, G., Daigle, F., and Harel, J. (2007) Use of selective capture of transcribed sequences to identify genes preferentially expressed by Streptococcus suis upon interaction with porcine brain microvascular endothelial cells. Appl Environ Microbiol 73: 4359- 4364.
Flanagan, J. J., Tweten, R. K., Johnson, A. E., and Heuck, A. P. (2009) Cholesterol exposure at the membrane surface is necessary and sufficient to trigger perfringolysin O binding. Biochemistry 48: 3977-3987.
Fongcom, A., Pruksakorn, S., Mongkol, R., Tharavichitkul, P., and Yoonim, N. (2001) Streptococcus suis infection in northern Thailand. J Med Assoc Thai 84: 1502-1508.
Gelber, S. E., Aguilar, J. L., Lewis, K. L., and Ratner, A. J. (2008) Functional and phylogenetic characterization of Vaginolysin, the human-specific cytolysin from Gardnerella vaginalis. J Bacteriol 190: 3896-3903.
Giddings, K. S., Johnson, A. E., and Tweten, R. K. (2003) Redefining cholesterol's role in the mechanism of the cholesterol-dependent cytolysins. Proc Natl Acad Sci U S A 100: 11315- 11320.
Giddings, K. S., Zhao, J., Sims, P. J., and Tweten, R. K. (2004) Human CD59 is a receptor for the cholesterol-dependent cytolysin intermedilysin. Nat Struct Mol Biol 11: 1173-1178.
Gottschalk, M., Lebrun, A., Wisselink, H., Dubreuil, J. D., Smith, H., and Vecht, U. (1998) Production of virulence-related proteins by Canadian strains of Streptococcus suis capsular type 2. Canadian Journal of Veterinary Research-Revue Canadienne de Recherche Veterinaire 62: 75-79.
Gottschalk, M., Petitbois, S., Higgins, R., and Jacques, M. (1991) Adherence of Streptococcus suis capsular type 2 to porcine lung sections. Can J Vet Res 55: 302-304.

General introduction Chapter 1
32
Gottschalk, M. and Segura, M. (2000) The pathogenesis of the meningitis caused by Streptococcus suis: the unresolved questions. Veterinary Microbiology 76: 259-272.
Gottschalk, M., Segura, M., and Xu, J. (2007) Streptococcus suis infections in humans: the Chinese experience and the situation in North America. Anim Health Res Rev 8: 29-45.
Gottschalk, M., Xu, J., Calzas, C., and Segura, M. (2010) Streptococcus suis: a new emerging or an old neglected zoonotic pathogen? 25. Future Microbiol 5: 371-391. Gottschalk, M. G., Lacouture, S., and Dubreuil, J. D. (1995) Characterization of Streptococcus suis capsular type 2 haemolysin. Microbiology 141 ( Pt 1): 189-195.
Herbert, D. and Todd, E. W. (1941) Purification and properties of a haemolysin produced by group A haemolytic streptococci (streptolysin O). Biochem J 35: 1124-1139.
Heuck, A. P., Hotze, E. M., Tweten, R. K., and Johnson, A. E. (2000) Mechanism of membrane insertion of a multimeric beta-barrel protein: perfringolysin O creates a pore using ordered and coupled conformational changes. Mol Cell 6: 1233-1242.
Higgins, R. and Gottschalk, M. (1990) An update on Streptococcus suis identification. J Vet Diagn Invest 2: 249-252.
Hoa, N. T., Chieu, T. T., Nghia, H. D., Mai, N. T., Anh, P. H., Wolbers, M., Baker, S., Campbell, J. I., Chau, N. V., Hien, T. T., Farrar, J., and Schultsz, C. (2011) The antimicrobial resistance patterns and associated determinants in Streptococcus suis isolated from humans in southern Vietnam, 1997-2008. BMC Infect Dis 11: 6.
Hotze, E. M. and Tweten, R. K. (31-7-2011a) Membrane assembly of the cholesterol-dependent cytolysin pore complex. Biochim Biophys Acta.
Jacobs, A. A. C., Loeffen, P. L. W., vandenBerg, A. J. G., and Storm, P. K. (1994) Identification, Purification, and Characterization of A Thiol-Activated Hemolysin (Suilysin) of Streptococcus Suis. Infection and Immunity 62: 1742-1748.
Jacobs, A. A. C., vandenBerg, A. J. G., and Loeffen, P. L. W. (1996) Protection of experimentally infected pigs by suilysin, the thiol-activated haemolysin of Streptococcus suis. Veterinary Record 139: 225-228.
Johnson, M. K., Geoffroy, C., and Alouf, J. E. (1980) Binding of cholesterol by sulfhydryl-activated cytolysins. Infect Immun 27: 97-101.
Kim, D., Han, K., Oh, Y., Kim, C. H., Kang, I., Lee, J., Gottschalk, M., and Chae, C. (2010) Distribution of capsular serotypes and virulence markers of Streptococcus suis isolated from pigs with polyserositis in Korea. Can J Vet Res 74: 314-316.
King, S. J., Heath, P. J., Luque, I., Tarradas, C., Dowson, C. G., and Whatmore, A. M. (2001) Distribution and genetic diversity of suilysin in Streptococcus suis isolated from different diseases of pigs and characterization of the genetic basis of suilysin absence. Infection and Immunity 69: 7572-7582.
Kock, C., Beineke, A., Seitz, M., Ganter, M., Waldmann, K. H., Valentin-Weigand, P., and Baums, C. G. (2009) Intranasal immunization with a live Streptococcus suis isogenic ofs mutant elicited suilysin-neutralization titers but failed to induce opsonizing antibodies and protection. Veterinary Immunology and Immunopathology 132: 135-145.

General introduction Chapter 1
33
Krawczyk-Balska, A. and Bielecki, J. (2005) Listeria monocytogenes listeriolysin O and phosphatidylinositol-specific phospholipase C affect adherence to epithelial cells. Can J Microbiol 51: 745-751.
Lalonde, M., Segura, M., Lacouture, S., and Gottschalk, M. (2000) Interactions between Streptococcus suis serotype 2 and different epithelial cell lines. Microbiology 146 ( Pt 8): 1913- 1921.
Lecours, M. P., Gottschalk, M., Houde, M., Lemire, P., Fittipaldi, N., and Segura, M. (2011) Critical Role for Streptococcus suis Cell Wall Modifications and Suilysin in Resistance to Complement-Dependent Killing by Dendritic Cells. J Infect Dis 204: 919-929.
Lowe, B. A., Marsh, T. L., Isaacs-Cosgrove, N., Kirkwood, R. N., Kiupel, M., and Mulks, M. H. (2011) Microbial communities in the tonsils of healthy pigs. Vet Microbiol 147: 346-357.
Lun, S. C., Perez-Casal, J., Connor, W., and Willson, P. J. (2003) Role of suilysin in pathogenesis of Streptococcus suis capsular serotype 2. Microbial Pathogenesis 34: 27-37.
Mai, N. T., Hoa, N. T., Nga, T. V., Linh, le D., Chau, T. T., Sinh, D. X., Phu, N. H., Chuong, L. V., Diep, T. S., Campbell, J., Nghia, H. D., Minh, T. N., Chau, N. V., de Jong, M. D., Chinh, N. T., Hien, T. T., Farrar, J., and Schultsz, C. (2008) Streptococcus suis meningitis in adults in Vietnam. Clin Infect Dis 46: 659-667.
Malley, R., Henneke, P., Morse, S. C., Cieslewicz, M. J., Lipsitch, M., Thompson, C. M., Kurt-Jones, E., Paton, J. C., Wessels, M. R., and Golenbock, D. T. (2003) Recognition of pneumolysin by Toll-like receptor 4 confers resistance to pneumococcal infection. Proc Natl Acad Sci U S A 100: 1966-1971.
Messier, S., Lacouture, S., and Gottschalk, M. (2008) Distribution of Streptococcus suis capsular types from 2001 to 2007. Can Vet J 49: 461-462.
Michel, E., Reich, K. A., Favier, R., Berche, P., and Cossart, P. (1990) Attenuated mutants of the intracellular bacterium Listeria monocytogenes obtained by single amino acid substitutions in listeriolysin O. Mol Microbiol 4: 2167-2178.
Navacharoen, N., Chantharochavong, V., Hanprasertpong, C., Kangsanarak, J., and Lekagul, S. (2009) Hearing and vestibular loss in Streptococcus suis infection from swine and traditional raw pork exposure in northern Thailand. J Laryngol Otol 123: 857-862.
Norton, P. M., Rolph, C., Ward, P. N., Bentley, R. W., and Leigh, J. A. (1999) Epithelial invasion and cell lysis by virulent strains of Streptococcus suis is enhanced by the presence of suilysin. Fems Immunology and Medical Microbiology 26: 25-35.
O'Sullivan, T., Friendship, R., Blackwell, T., Pearl, D., McEwen, B., Carman, S., Slavic, D., and Dewey, C. (2011) Microbiological identification and analysis of swine tonsils collected from carcasses at slaughter. Can J Vet Res 75: 106-111.
Okamoto, S., Kawabata, S., Terao, Y., Fujitaka, H., Okuno, Y., and Hamada, S. (2004) The Streptococcus pyogenes capsule is required for adhesion of bacteria to virus-infected alveolar epithelial cells and lethal bacterial-viral superinfection. Infect Immun 72: 6068-6075.
Okwumabua, O., Abdelmagid, O., and Chengappa, M. M. (1999) Hybridization analysis of the gene encoding a hemolysin (suilysin) of Streptococcus suis type 2: evidence for the absence of the gene in some isolates. FEMS Microbiol Lett 181: 113-121.

General introduction Chapter 1
34
Orihuela, C. J., Gao, G., Francis, K. P., Yu, J., and Tuomanen, E. I. (2004) Tissue-specific contributions of pneumococcal virulence factors to pathogenesis. J Infect Dis 190: 1661-1669.
Padungtod, P., Tharavichitkul, P., Junya, S., Chaisowong, W., Kadohira, M., Makino, S., and Sthitmatee, N. (2010) Incidence and presence of virulence factors of Streptococcus suis infection in slaughtered pigs from Chiang Mai, Thailand. Southeast Asian J Trop Med Public Health 41: 1454-1461.
Pan, X. Z., Ge, J. C., Li, M., Wu, B., Wang, C. J., Wang, J., Feng, Y. J., Yin, Z. M., Zheng, F., Cheng, G., Sun, W., Ji, H. F., Hu, D., Shi, P. J., Feng, X. D., Hao, X. N., Dong, R. P., Hu, F. Q., and Tang, J. Q. (2009) The Orphan Response Regulator CovR: a Globally Negative Modulator of Virulence in Streptococcus suis Serotype 2. Journal of Bacteriology 191: 2601-2612. Perch, B., Pedersen, K. B., and Henrichsen, J. (1983) Serology of capsulated streptococci pathogenic for pigs: six new serotypes of Streptococcus suis. J Clin Microbiol 17: 993-996.
Petersen, R., Hannerz, H., Tuchsen, F., and Egerton, J. R. (2011) Meningitis, sepsis and endocarditis among workers occupationally exposed to pigs. Occup Med (Lond).
Pinkney, M., Beachey, E., and Kehoe, M. (1989) The thiol-activated toxin streptolysin O does not require a thiol group for cytolytic activity. Infect Immun 57: 2553-2558.
Princivalli, M. S., Palmieri, C., Magi, G., Vignaroli, C., Manzin, A., Camporese, A., Barocci, S., Magistrali, C., and Facinelli, B. (2009) Genetic diversity of Streptococcus suis clinical isolates from pigs and humans in Italy (2003-2007). Euro Surveill 14.
Ramachandran, R., Tweten, R. K., and Johnson, A. E. (2004) Membrane-dependent conformational changes initiate cholesterol-dependent cytolysin oligomerization and intersubunit beta-strand alignment. Nat Struct Mol Biol 11: 697-705.
Rampersaud, R., Planet, P. J., Randis, T. M., Kulkarni, R., Aguilar, J. L., Lehrer, R. I., and Ratner, A. J. (2011) Inerolysin, a cholesterol-dependent cytolysin produced by Lactobacillus iners. J Bacteriol 193: 1034-1041.
Ratner, A. J., Hippe, K. R., Aguilar, J. L., Bender, M. H., Nelson, A. L., and Weiser, J. N. (2006) Epithelial cells are sensitive detectors of bacterial pore-forming toxins. J Biol Chem 281: 12994-12998.
Robertson, I. D., Blackmore, D. K., Hampson, D. J., and Fu, Z. F. (1991) A longitudinal study of natural Infection of piglets with Streptococcus suis types 1 and 2. Epidemiol Infect 107: 119- 126.
Rossjohn, J., Feil, S. C., McKinstry, W. J., Tweten, R. K., and Parker, M. W. (1997) Structure of a cholesterol-binding, thiol-activated cytolysin and a model of its membrane form. Cell 89: 685- 692.
Rossjohn, J., Gilbert, R. J., Crane, D., Morgan, P. J., Mitchell, T. J., Rowe, A. J., Andrew, P. W., Paton, J. C., Tweten, R. K., and Parker, M. W. (1998) The molecular mechanism of pneumolysin, a virulence factor from Streptococcus pneumoniae. J Mol Biol 284: 449-461.
Rossjohn, J., Polekhina, G., Feil, S. C., Morton, C. J., Tweten, R. K., and Parker, M. W. (2007) Structures of perfringolysin O suggest a pathway for activation of cholesterol-dependent cytolysins. J Mol Biol 367: 1227-1236.
Rubins, J. B., Paddock, A. H., Charboneau, D., Berry, A. M., Paton, J. C., and Janoff, E. N. (1998) Pneumolysin in pneumococcal adherence and colonization. Microb Pathog 25: 337-342.

General introduction Chapter 1
35
Sanford, S. E. and Tilker, M. E. (1982) Streptococcus suis type II-associated diseases in swine: observations of a one-year study. J Am Vet Med Assoc 181: 673-676.
Saunders, F. K., Mitchell, T. J., Walker, J. A., Andrew, P. W., and Boulnois, G. J. (1989) Pneumolysin, the thiol-activated toxin of Streptococcus pneumoniae, does not require a thiol group for in vitro activity. Infect Immun 57: 2547-2552.
Segers, R. P., Kenter, T., de Haan, L. A., and Jacobs, A. A. (1998) Characterisation of the gene encoding suilysin from Streptococcus suis and expression in field strains. FEMS Microbiol Lett 167: 255-261.
Segura, M. and Gottschalk, M. (2002) Streptococcus suis interactions with the murine macrophage cell line J774: adhesion and cytotoxicity. Infect Immun 70: 4312-4322.
Segura, M., Stankova, J., and Gottschalk, M. (1999) Heat-killed Streptococcus suis capsular type 2 strains stimulate tumor necrosis factor alpha and interleukin-6 production by murine macrophages. Infect Immun 67: 4646-4654.
Segura, M., Vadeboncoeur, N., and Gottschalk, M. (2002) CD14-dependent and -independent cytokine and chemokine production by human THP-1 monocytes stimulated by Streptococcus suis capsular type 2. Clin Exp Immunol 127: 243-254.
Segura, M., Vanier, G., Al Numani, D., Lacouture, S., Olivier, M., and Gottschalk, M. (2006) Proinflammatory cytokine and chemokine modulation by Streptococcus suis in a whole-blood culture system. FEMS Immunol Med Microbiol 47: 92-106.
Silva, L. M., Baums, C. G., Rehm, T., Wisselink, H. J., Goethe, R., and Valentin-Weigand, P. (2006) Virulence-associated gene profiling of Streptococcus suis isolates by PCR. Vet Microbiol 115: 117-127.
Soltani, C. E., Hotze, E. M., Johnson, A. E., and Tweten, R. K. (2007) Specific protein-membrane contacts are required for prepore and pore assembly by a cholesterol-dependent cytolysin. J Biol Chem 282: 15709-15716.
Soltani, C. E., Hotze, E. M., Johnson, A. E., and Tweten, R. K. (2007b) Structural elements of the cholesterol-dependent cytolysins that are responsible for their cholesterol-sensitive membrane interactions. Proc Natl Acad Sci U S A 104: 20226-20231.
Sriskandan, S. and Slater, J. D. (2006) Invasive disease and toxic shock due to zoonotic Streptococcus suis: an emerging infection in the East? PLoS Med 3: e187.
Staats, J. J., Feder, I., Okwumabua, O., and Chengappa, M. M. (1997) Streptococcus suis: past and present. Vet Res Commun 21: 381-407.
Staats, J. J., Plattner, B. L., Stewart, G. C., and Chengappa, M. M. (1999) Presence of the Streptococcus suis suilysin gene and expression of MRP and EF correlates with high virulence in Streptococcus suis type 2 isolates. Veterinary Microbiology 70: 201-211.
Stachowiak, R., Wisniewski, J., Osinska, O., and Bielecki, J. (2009) Contribution of cysteine residue to the properties of Listeria monocytogenes listeriolysin O. Can J Microbiol 55: 1153-1159.
Strangmann, E., Froleke, H., and Kohse, K. P. (2002) Septic shock caused by Streptococcus suis: case report and investigation of a risk group. Int J Hyg Environ Health 205: 385-392.

General introduction Chapter 1
36
Sukeno, A., Nagamune, H., Whiley, R. A., Jafar, S. I., Aduse-Opoku, J., Ohkura, K., Maeda, T., Hirota, K., Miyake, Y., and Kourai, H. (2005) Intermedilysin is essential for the invasion of hepatoma HepG2 cells by Streptococcus intermedius. Microbiol Immunol 49: 681-694.
Takamatsu, D., Osaki, M., and Sekizaki, T. (2002) Evidence for lateral transfer of the Suilysin gene region of Streptococcus suis. J Bacteriol 184: 2050-2057.
Tang, J., Wang, C., Feng, Y., Yang, W., Song, H., Chen, Z., Yu, H., Pan, X., Zhou, X., Wang, H., Wu, B., Wang, H., Zhao, H., Lin, Y., Yue, J., Wu, Z., He, X., Gao, F., Khan, A. H., Wang, J., Zhao, G. P., Wang, Y., Wang, X., Chen, Z., and Gao, G. F. (2006) Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med 3: e151.
Tarradas, C., Borge, C., Arenas, A., Maldonado, A., Astorga, R., Miranda, A., and Luque, I. (2001) Suilysin production by Streptococcus suis strains isolated from diseased and healthy carrier pigs in Spain. Vet Rec 148: 183-184.
Tenenbaum, T., Adam, R., Eggelnpohler, I., Matalon, D., Seibt, A., GE, K. Novotny, Galla, H. J., and Schroten, H. (2005) Strain-dependent disruption of blood-cerebrospinal fluid barrier by Streptoccocus suis in vitro. FEMS Immunol Med Microbiol 44: 25-34.
Tenenbaum, T., Essmann, F., Adam, R., Seibt, A., Janicke, R. U., Novotny, G. E., Galla, H. J., and Schroten, H. (2006) Cell death, caspase activation, and HMGB1 release of porcine choroid plexus epithelial cells during Streptococcus suis infection in vitro. Brain Res 1100: 1-12.
Tenenbaum, T., Papandreou, T., Gellrich, D., Friedrichs, U., Seibt, A., Adam, R., Wewer, C., Galla, H. J., Schwerk, C., and Schroten, H. (2008) Polar bacterial invasion and translocation of Streptococcus suis across the blood-cerebrospinal fluid barrier in vitro. Cell Microbiol.
Tian, Y., Aarestrup, F. M., and Lu, C. P. (2004) Characterization of Streptococcus suis serotype 7 isolates from diseased pigs in Denmark. Vet Microbiol 103: 55-62.
Tomoyasu, T., Tabata, A., Hiroshima, R., Imaki, H., Masuda, S., Whiley, R. A., Aduse-Opoku, J., Kikuchi, K., Hiramatsu, K., and Nagamune, H. (2010) Role of catabolite control protein A in the regulation of intermedilysin production by Streptococcus intermedius. Infect Immun 78: 4012- 4021.
Trottier, S., Higgins, R., Brochu, G., and Gottschalk, M. (1991) A Case of Human Endocarditis Due to Streptococcus Suis in North-America. Reviews of Infectious Diseases 13: 1251-1252.
Tsuchiya, K., Kawamura, I., Takahashi, A., Nomura, T., Kohda, C., and Mitsuyama, M. (2005) Listeriolysin O-induced membrane permeation mediates persistent interleukin-6 production in Caco-2 cells during Listeria monocytogenes infection in vitro. Infect Immun 73: 3869-3877.
Tweten, R. K. (2005) Cholesterol-dependent cytolysins, a family of versatile pore-forming toxins. Infection and Immunity 73: 6199-6209.
Vadeboncoeur, C. and Pelletier, M. (1997) The phosphoenolpyruvate:sugar phosphotransferase system of oral streptococci and its role in the control of sugar metabolism. Fems Microbiology Reviews 19: 187-207. Vadeboncoeur, N., Segura, M., Al Numani, D., Vanier, G., and Gottschalk, M. (2003) Pro-inflammatory cytokine and chemokine release by human brain microvascular endothelial cells stimulated by Streptococcus suis serotype 2. FEMS Immunol Med Microbiol 35: 49-58.
Vanier, G., Segura, M., Friedl, P., Lacouture, S., and Gottschalk, M. (2004) Invasion of porcine brain microvascular endothelial cells by Streptococcus suis serotype 2. Infection and Immunity 72: 1441-1449.

General introduction Chapter 1
37
Vanier, G., Segura, M., and Gottschalk, M. (2007) Characterization of the invasion of porcine endothelial cells by Streptococcus suis serotype 2. Can J Vet Res 71: 81-89.
Vanier, G., Segura, M., Lecours, M. P., Grenier, D., and Gottschalk, M. (2008) Porcine brain microvascular endothelial cell-derived interleukin-8 is first induced and then degraded by Streptococcus suis. Microb Pathog.
Vecht, U., Wisselink, H. J., Jellema, M. L., and Smith, H. E. (1991) Identification of 2 Proteins Associated with Virulence of Streptococcus Suis Type-2. Infection and Immunity 59: 3156- 3162.
Walker, J. A., Allen, R. L., Falmagne, P., Johnson, M. K., and Boulnois, G. J. (1987) Molecular cloning, characterization, and complete nucleotide sequence of the gene for pneumolysin, the sulfhydryl-activated toxin of Streptococcus pneumoniae. Infect Immun 55: 1184-1189.
Wangkaew, S., Chaiwarith, R., Tharavichitkul, P., and Supparatpinyo, K. (2006) Streptococcus suis infection: a series of 41 cases from Chiang Mai University Hospital. J Infect 52: 455-460.
Watson, K. C., Rose, T. P., and Kerr, E. J. (1972) Some factors influencing the effect of cholesterol on streptolysin O activity. J Clin Pathol 25: 885-891.
Wei, Z., Li, R., Zhang, A., He, H., Hua, Y., Xia, J., Cai, X., Chen, H., and Jin, M. (2009) Characterization of Streptococcus suis isolates from the diseased pigs in China between 2003 and 2007. Vet Microbiol 137: 196-201.
Weis, S. and Palmer, M. (2001) Streptolysin O: the C-terminal, tryptophan-rich domain carries functional sites for both membrane binding and self-interaction but not for stable oligomerization. Biochim Biophys Acta 1510: 292-299.
Wertheim, H. F., Nghia, H. D., Taylor, W., and Schultsz, C. (2009) Streptococcus suis: an emerging human pathogen. Clin Infect Dis 48: 617-625.
Willenborg, J., Fulde, M., de Greeff, A., Rohde, M., Smith, H. E., Valentin-Weigand, P., and Goethe, R. (2011) Role of glucose and CcpA in capsule expression and virulence of Streptococcus suis. Microbiology 157: 1823-1833.
Williams, A. E. and Blakemore, W. F. (1990) Pathogenesis of meningitis caused by Streptococcus suis type 2. J Infect Dis 162: 474-481.
Windsor, R. S. and Elliott, S. D. (1975) Streptococcal infection in young pigs. IV. An outbreak of streptococcal meningitis in weaned pigs. J Hyg (Lond) 75: 69-78.
Wisselink, H. J., Smith, H. E., Stockhofe-Zurwieden, N., Peperkamp, K., and Vecht, U. (2000) Distribution of capsular types and production of muramidase-released protein (MRP) and extracellular factor (EF) of Streptococcus suis strains isolated from diseased pigs in seven European countries. Veterinary Microbiology 74: 237-248.
Wu, T., Chang, H., Tan, C., Bei, W., and Chen, H. (2009) The orphan response regulator RevSC21 controls the attachment of Streptococcus suis serotype-2 to human laryngeal epithelial cells and the expression of virulence genes. FEMS Microbiol Lett 292: 170-181.
Wu, T., Zhao, Z., Zhang, L., Ma, H., Lu, K., Ren, W., Liu, Z., Chang, H., Bei, W., Qiu, Y., and Chen, H. (2011) Trigger factor of Streptococcus suis is involved in stress tolerance and virulence. Microb Pathog 51: 69-76.

General introduction Chapter 1
38
Xiong, Y., Liu, Q., Qin, F. Y., Bai, Y., Zhu, W., Li, H. M., Guo, J. G., Qin, L., Pan, J., Long, J. M., and Chen, L. (2007) [Study on the molecular epidemiology of Streptococcus suis type 2 from healthy pigs in Guangxi]. Zhonghua Liu Xing Bing Xue Za Zhi 28: 593-596.
Xu, L., Huang, B., Du, H., Zhang, X. C., Xu, J., Li, X., and Rao, Z. (2010) Crystal structure of cytotoxin protein suilysin from Streptococcus suis. Protein Cell 1: 96-105.
Ye, C., Zheng, H., Zhang, J., Jing, H., Wang, L., Xiong, Y., Wang, W., Zhou, Z., Sun, Q., Luo, X., Du, H., Gottschalk, M., and Xu, J. (2009) Clinical, Experimental, and Genomic Differences between Intermediately Pathogenic, Highly Pathogenic, and Epidemic Streptococcus suis. J Infect Dis 199: 97-107.
Ye, C., Zhu, X., Jing, H., Du, H., Segura, M., Zheng, H., Kan, B., Wang, L., Bai, X., Zhou, Y., Cui, Z., Zhang, S., Jin, D., Sun, N., Luo, X., Zhang, J., Gong, Z., Wang, X., Wang, L., Sun, H., Li, Z., Sun, Q., Liu, H., Dong, B., Ke, C., Yuan, H., Wang, H., Tian, K., Wang, Y., Gottschalk, M., and Xu, J. (2006) Streptococcus suis sequence type 7 outbreak, Sichuan, China. Emerg Infect Dis 12: 1203-1208.
Yu, H., Jing, H., Chen, Z., Zheng, H., Zhu, X., Wang, H., Wang, S., Liu, L., Zu, R., Luo, L., Xiang, N., Liu, H., Liu, X., Shu, Y., Lee, S. S., Chuang, S. K., Wang, Y., Xu, J., and Yang, W. (2006) Human Streptococcus suis outbreak, Sichuan, China. Emerg Infect Dis 12: 914-920.
Zhao, Y., Liu, G., Li, S., Wang, M., Song, J., Wang, J., Tang, J., Li, M., and Hu, F. (2011) Role of a type IV-like secretion system of Streptococcus suis 2 in the development of streptococcal toxic shock syndrome. J Infect Dis 204: 274-281.
Zheng, H., Punaro, M. C., Segura, M., Lachance, C., Rivest, S., Xu, J., Houde, M., and Gottschalk, M. (2011) Toll-like receptor 2 is partially involved in the activation of murine astrocytes by Streptococcus suis, an important zoonotic agent of meningitis. J Neuroimmunol 234: 71-83.

Chapter 2
Material and methods


Material and methods Chapter 2
41
If not stated otherwise all materials were purchased from Sigma (Muenchen,
Germany).
1. Bacterial strains and growth conditions In this study Streptococcus (S.) suis serotype 2 wild type strain 10, kindly provided by
H. Smith (Lelystad, NL), and its isogenic mutants were used. This strain expresses
EF, MRP, SLY, FBPS and OFS. It has been used by different groups successfully for
mutagenesis and experimental intranasal infections of pigs (Baums et al., 2006;
Smith et al., 1999; Vecht et al., 1997). The unencapsulated isogenic mutant strain
10cpsΔEF of the virulent Streptococcus (S.) suis serotype 2 wild type strain 10
(designated strain 10cpsΔEF) was kindly provided by H. Smith (Lelystad, NL). Strain
10cpsΔEF has been generated by insertional mutagenesis of the genes cps2E and
cps2F involved in biosynthesis of the capsule and has been demonstrated to be
severely attenuated in virulence, most likely due to increased opsonophagocytosis
(Smith et al., 1999). The corresponding suilysin deficient mutants of wild type strain
10 (designed 10Δsly) and 10cpsΔEF (designed 10cpsΔEFΔsly) were constructed by
insertion of an erythromycin cassette in the sly-gene of the unencapsulated strain
10cpsΔEF using the plasmid pBlue/sly/erm as previously described for generation of
strain 10Δsly (Benga et al., 2008). Mutagenesis was confirmed by PCR and Southern
Blot analysis. Streptococci were grown on Columbia agar supplemented with 7%
sheep blood (Oxoid, Wesel, Germany) or in Bacto Todd Hewitt broth (THB) overnight
under aerobic conditions at 37°C. If required, antibiotics were added to the media at
the following concentrations: spectinomycin 100 µg/ml for S. suis; erythromycin 1
µg/ml for S. suis.
Lactococcus lactis subspecies cremoris MG1363 (L. lactis), L. lactis heterologously
expressing SLY (L. lactis pORI-SLY, L. lactis pORI-W461F, L. lactis pORI-SVD) and
L. lactis carrying the shuttle vector pORI23 (L. lactis pORI23) were kindly provided by
D. Reinscheid (Fachhochschule Bonn-Rhein-Sieg, Germany). All L. lactis strains
were cultured on M17 agar plates (Oxoid) supplemented with 5% glucose (GM17)
overnight under aerobic conditions at 30°C. If required, 5 μg/ml of erythromycin was
added to the GM17 medium or plates.

Material and methods Chapter 2
42
Escherichia (E.) coli strains BL21, BL21 (DE3) and DH5α were used for molecular
cloning and protein expression experiments. E. coli were grown in Luria Bertani (LB)
medium overnight under aerobic conditions at 37°C with vigorous shaking. In
appropriate cases, 100 μg/ml of ampicillin was added to the LB medium or plates.
For infection of HEp-2 cells, streptococci were grown in Todd-Hewitt broth (THB,
Difco, Detroit, USA) overnight at 37°C under aerobic conditions, adjusted to an
optical density (OD600) of 0.02 in pre-warmed media the next day and grown to late
exponential growth phase (OD600 0.8).
2. Molecular biology and protein biochemical methods Routine molecular biology techniques including restriction endonuclease digestion,
DNA ligations, agarose gel electrophoresis, transformation of E. coli and plasmid
isolation were performed according to standard procedures (Sambrook et al., 1989).
Restriction enzymes were purchased from New England Biolabs (Frankfurt am Main,
Germany). Plasmid preparations were performed with kits from Machery-Nagel
(Dueren, Germany).
2.1. Construction of mutated suilysin W461F und SVD W461F and RGD-SVD substitutions were constructed using site-directed-
mutagenesis according to the instruction manual of QuikChange® Site-Directed
Mutagenesis Kit (Stratagene, Ja Jolla, CA). As template DNA pET45bslynew was
used (Kock et al., 2009). Oligonucleotide primers encoding the W461 to F461
substitution, namely slyTrp-Phefornew (TACAGGATTAGCATTTGAGTGGTGGAG
AAC) and slyTrp-Pherevnew (GTTCTCCACCACTCAAATGCTAATCCTGTAC), and
oligonucleotide primers encoding R124-G125-D126 to S124-V125-D126, namely
slyRGDMutfor (CAGTATTGCGTCGGTAGATCTGACGCTTAG) and slyRGDMutrev
(CTAAGCGTCAGATCTACCGACGCAATACTG) were used. After digestion of the
methylated non-mutated parental DNA template with DpnI, mutated plasmid
pET45bslynewW461F was electroporated (2.5 kV, 200 Ω, 25 mF) into E. coli BL21
(DE3) and then purified. Mutation sites of each suilysin derivate were sequenced

Material and methods Chapter 2
43
using the primers slyseqfor (GGATCATTCAGGTGCTTATG) and pET45seqrev
(TGCTGGCGTTCAAATTTCGC).
2.2. Expression of recombinant proteins Recombinant His-tagged suilysin (rSLY) was expressed in E. coli BL21
pET45bslynew (Kock et al., 2009). In accordance, recombinant point-mutated
suilysin W461F (rW461F) was expressed in E. coli BL21 pET45bslynewW461F and
recombinant RGD-SVD (rSVD) in E. coli BL21 pET45bslynewRGD-SVD. Proteins
were expressed after induction with 0.4 mM isopropyl-β-D-thiogalactoside for 4 h.
His-tagged proteins were purified under native conditions with Protino® Ni-TED 2000
packed columns as recommended by the manufacturer (Macherey-Nagel) with some
modification. rSLY and rW461F were eluted with a 1:10 dilution of the elution buffer
(containing 10 mM imidazole). The eluted proteins were dialysed against 100 mM
TrisHCl [pH 7.0] with 1 mM DTT. Protein concentrations were measured according to
microplate assay protocol (Bio-Rad, Muenchen, Germany) and the haemolytic
activity was tested as described by Takamatsu et al. (2001). Purification of proteins
were verified by immunoblot analysis with anti-Penta His antibodies (Qiagen, Hilden,
Germany) as recommended. Silver staining was used to control the purity grade. The
protein was stored at -20°C.
Suilysin was completely sequenced with standard oligonucleotide primers T7
(TAATACGACTCACTATAGGG) and T7 term (CTAGTTATTGCTCAGCGGT;
Eurofins MWG operon, Ebersberg, Germany).
2.3. Heterologous expression of SLY, W461F and SVD in L. lactis
The sly gene was amplified from chromosomal DNA of S. suis by PCR with
oligonucleotide primers slyPstI (TAGTCTGCAGCTCCTAGCCTCTGTGGCTAA) and
slyBamHIoptRBS (CAGAGGATCCAGGAGAAAACTTATGAGAAAAAG). After
digestion of the PCR product with BamHI and PstI the fragment containing the whole
sly-gene was cloned into the BamHI/PstI-digested shuttle vector pOri23 (Que et al.,
2000). The resulting plasmid was named pOri23-SLY and transferred in L. lactis as
described previously (Holo and Nes, 1989). Transformants were screened by plating

Material and methods Chapter 2
44
on GM17 agar plates containing 5 µg/ml erythromycin. For heterologous expression
of W461F and SVD site-directed mutagenesis was performed as described above
using purified plasmid DNA pOri23sly and the respective oligonucleotide primers
slyTrp-Phefornew and slyTrp-Pherewnew as well as slyRGDMutfor and
slyRGDMutrev. Resulting constructs pOri23-W461F and pOri23-SVD were
electroporated in L. lactis.
2.4. Immunoblot analysis Recombinant proteins or supernatants of infected cells were separated by SDS-
polyacrylamid gel electrophoresis with a 4% stacking and a 10% separating gel
under denaturing conditions and transferred to a PVDF-membrane (Serva,
Heidelberg, Germany). For suilysin immunoblot, membrane-blocking was performed
overnight with 3% milk powder in TBS with 0.5% Tween. Polyclonal antiserum raised
against rSLY (Benga et al., 2008) diluted 1:1300 in 1% milk powder was used to
detect either rSLY, rW461F, rSVD or suilysin and its mutated derivates expressed by
the respective L. lactis strains (L. lactis pORI-SLY, L. lactis pORI-W461F and L. lactis
pORI-SVD) present in culture supernatants. For detection of His-tagged recombinant
proteins membranes were blocked with 3% BSA in TBS with 0.5% Tween overnight
and incubated with a monoclonal anti-Penta His antibody diluted 1:2000 in 3% BSA.
Membranes were developed with horseradish peroxidase-conjugated anti-rabbit or
anti-mouse IgG antiserum diluted 1:10000 in 1% milk powder (Amersham, Freiburg,
Germany) followed by using SuperSignal® West Pico Chemiluminescent Substrate
(Pierce, Rockford, USA) according to the manufactures protocol.
3. Cell culture methods 3.1. Epithelial cells The human laryngeal epithelial cell line HEp-2 (ATCC CCL 23) was used. Cells were
maintained in Dulbecco’s modified Eagle’s medium (DMEM, Gibco-Invitrogen,
Groningen, The Netherlands) supplemented with 10% fetal calf serum (FCS) and 5
mM glutamine at 37°C and 8% CO2. The cells were subcultured every 2-3 days after
detachment with 0.25% trypsin and 1 mM Na-EDTA (trypsin-EDTA, Gibco-

Material and methods Chapter 2
45
Invitrogen). For antibiotic protection assays, approximately 1.8 x 105 cells per well
were seeded on 24 well tissue culture plates, for immunofluorescence and
electronmicroscopy 0.5 x 105 and 1.5 x 105 cells per well were seeded on a 12 mm
diameter glass cover slips placed in 24 well plates. For flow cytometry analysis
approximately 6 x 105 cells per well were seeded on a 6 well plate and for cellular
cytotoxicity assay 0.2 x 105 cells were seeded on a 96 well plate. The cells were
grown over-night and then used for experiments.
3.2. Antibiotic protection assay The number of adherent and invasive streptococci was quantified using a gentamicin
protection assay as described with the following modifications (Valentin-Weigand et
al., 1996). Adhesion and invasion assays were performed with fresh grown
streptococci in THB harvested at an OD600 of 0.8 by centrifugation. Subsequently,
bacteria were resuspended in PBS containing 0.9 mM CaCl2/0.5 mM MgCl26H2O
[pH 7.3] (PBS+; Gibco-Invitrogen) and adjusted to an OD600 0.6. Then, diluted 1:10 in
PBS+ and used for adherence and invasion assay. Confluent HEp-2 cell monolayers
were inoculated with 100 bacteria per cell. The number of colony forming units (CFU)
inoculated per well was determined by serial platings on Columbia agar
supplemented with 7% sheep blood. The cells were incubated for 2 h at 37°C with
8% CO2 in order to allow adherence and invasion of the bacteria. Supernatants of
infected cells were collected 2 hours post infection for immunoblot analysis of
suilysin. To determine adherence rates cells were washed three times with PBS
immediately after supernatant collection, and 100 μl trypsin-EDTA solution was
added to each well. After 5 min, cells were lysed by adding 400 μl 0.025%
Triton X-100. Serial dilutions of these lysates were plated in triplicates on Columbia
agar and incubated at 37°C for 24 h. For invasion assay, cells were washed 3 times
with PBS 2 h post infection and incubated with 500 μl per well of DMEM containing
100 μg gentamicin and 5 μg penicillin per ml for 2 additional hours at 37°C with 8%
CO2 to kill extracellular bacteria. The cells were washed and lysed as described for
adherence and lysates were plated for viable counts of streptococci. Results for

Material and methods Chapter 2
46
adherence and invasion were expressed as percentage of the inoculum used for
infection.
In some adherence and invasion assays, HEp-2 cells were treated before infection
with the following inhibitors: 1 µg/ml Latrunculin B for 30 min, 10 ng/ml toxin B of
Clostridium (C.) difficile (TcdB), 10 ng/ml toxin B of C. difficile serotype F strain 1470
(TcdB-F) and 1 µg/ml C3 toxin of C. limosum for 3 h. C3 toxin was always
reconstituted freshly by mixing the binding domain (C2IIa) with the catalytic domain
(FT) in a ratio of 1:1 as described (Just et al., 1995). After preincubation with
inhibitors cells were washed and adherence and invasion assays were performed as
described above. Latrunculin B was left on to the cells during the whole infection
experiment because its effect is reversible up to 1 h.
3.3. Labelling of bacteria for flow cytometry experiments CellTrace™ CFSE Cell Proliferation Kit (Invitrogen) was used for fluorescent staining
of bacteria. Labelling of bacteria with CFSE was done as described by Logan et al.
(1998) with some modifications. Streptococcal strains 10cpsΔEF and 10cpsΔEFΔsly
were grown to an OD600 of 0.8, pelleted by centrifugation, washed once with PBS
(Gibco-Invitrogen) and resuspended in PBS containing 5 µM carboxyfluorescein
succinimidyl ester (CFSE)-reagent (prepared from a CFSE stock solution as
recommended by the manufacturer). Staining procedures were carried out at 37°C
for 20 min on an end-over-end rotator. After washing (removal of unbound
fluorescent dye) culture stocks were prepared as described above. Labelling
efficiency (>90%) was confirmed by fluorescent microscopy and measurement of
fluorescent intensity by flow cytometry (side-ward-scatter [ssc] versus fluorescence
channel FL-1) in comparison to unlabeled streptococci (data not shown).
3.4. Bacteria-cell association HEp-2 cell monolayers were infected with CFSE-labelled streptococci at a multiplicity
of infection [moi] of 125:1 bacteria per cell, followed by centrifugation (250 x g for
5 min) and incubation at 37°C for 2 h. After incubation supernatants of infected cell
were collected for determination of suilysin expression by immunoblot analysis. Cells

Material and methods Chapter 2
47
were detached with trypsin-EDTA solution, washed with PBS and fixed by
resuspension in PBS with 0.375% formaldehyde in PBS. In complementation
experiments, rSLY and rW461F were added to strain 10cpsΔEFΔsly in a protein
concentration of 100 ng/ml during the experiment. Bacteria-cell association was
measured using FACScan® (Becton Dickinson, 488 nm Argon laser). Further analysis
was performed with the software WinMDI (version 2.9.). For each determination at
least 10,000 events were acquired and initial analysis of infected cells was carried
out by dot plot analysis (forward scatter [fsc] versus sideward scatter [ssc]) to define
the cell population of interest (data not shown). Subsequently, fluorescent cells were
detected at channel FL-1. The numerical results were expressed as mean
fluorescent intensity [mfi] values of cell population. Noninfected cells served as
background control. Means and standard deviations refer to eight independent
experiments. To exclude cytotoxic effects of SLY during the experiment cells were
stained afterwards with propidium iodid (4 µg/ml, DNA-intercalating dye) for 2 min at
room temperature to discriminate between viable and nonviable cells. Red
fluorescent cells (dead cells) were determined by dot plot analysis (ssc versus FL-2).
In some assays, HEp-2 cells were treated before infection with 1µg/ml latrunculin B
for 30 min and latrunculin B was also added to the cells during the whole experiment
because its effect is reversible during 1 h. After preincubation with latrunculin B cells
were washed and flow cytometry was performed as described above.
3.5. Immunofluorescence microscopy Membrane binding of rSLY, rW461F and rSVD was determined by
immunofluorescent microscopy. HEp-2 cells were incubated with the recombinant
suilysin derivates at subcytolytic (100 ng/ml) or lytic (400 ng/ml) concentrations for 30
min at 37°C. Cells were washed three times to remove unbound suilysin, fixed with
2% parafolmaldehyde and permeabilised with 0.1% Triton X-100 in PBS (Bio-Rad,
Muenchen, Germany). Blocking was performed for 30 min at room temperature with
PBS containing 10% FCS. Suilysin was stained using a polyclonal antiserum raised
against rSLY (Benga et al., 2008) diluted 1:1,300 in 1% milk powder for 1 h at room
temperature followed by an incubation with FITC-conjugated goat anti-rabbit antibody

Material and methods Chapter 2
48
(1:1,000 in 1% FCS in PBS, Dianova, Hamburg, Germany). F-actin was labelled with
TRITC-conjugated Phalloidin (20 µg/ml, Invitrogen) for 40 min at room temperature.
After final washing DAPI (Invitrogen) was used for staining the nuclei. Antifading
reagent DABCO (Sigma) was used for sealing of the samples. Mounted samples
were examined using inverted immunofluorescence microscope Nikon Eclipse Ti-S
equipped with a 40×, 0.6 S Plan Fluor objective (Nikon, Duesseldorf, Germany)
driven by NIS Elements software BR 3.2..
3.6. Double immunofluorescence microscopy (DIF) Determination of adherence and invasion of streptococci by DIF was done as
described by Benga et al. (2004) with the following modifications. Semiconfluent
HEp-2 cells were incubated with streptococci at a moi of 100 bacteria per epithelial
cell. ProLong® Gold antifade reagent with DAPI (Invitrogen) was used for sealing and
staining of the nucleus. Mounted samples were examined using inverted
immunofluorescence microscope Nikon Eclipse Ti-S. equipped with a 40×, 0.6 S Plan
Fluor objective (Nikon, Duesseldorf, Germany) driven by NIS Elements software BR
3.2..
3.7. Colocalisation experiments For suilysin localization experiments GFP-Rac1-expressing HEp-2 cells were fixed
after incubation with rSLY (protein concentration of 100 ng/ml) for 30 min at 37°C
with 3% PFA in modified cytoskeleton buffer (10 mM MES [pH 7.0], 150 mM NaCl, 5
mM EGTA, 5 mM MgCl2, 5 mM sucrose) for 20 min and quenched with 10 mM glycin
in PBS and permeabelised with 0.1% Triton X-100 in PBS. Permeabilised cells were
blocked in PBS with 5% horse serum and 1% BSA (blocking buffer) for 30 min at
room temperature and subsequently incubated with polyclonal antiserum raised
against rSLY (Benga et al., 2008) diluted 1:1,200 in blocking buffer for 45 min at
room temperature. Samples were washed twice in PBS and incubated with Cy5-
conjugated goat anti-rabbit IgG (Millipore, Schwalbach/Ts, Germany) for 30 min at
room temperature. F-actin was labelled with Alexa® Fluor 568-conjugated Phalloidin
(Invitrogen) for 20 min at room temperature. Coverslips were washed three times in

Material and methods Chapter 2
49
PBS and then mounted using ProLong® Gold antifade reagent with DAPI (Invitrogen).
Mounted samples were examined using a LSM 510 Meta confocal microscope
equipped with a 63×, 1.2NA Plan-NEOFLUAR oil immersion objective (Zeiss, Jena,
Germany) driven by LSM software v3.2. For 3D z-stack acquisition the pinholes were
set to one Airy unit and confocal planes were acquired every 340 nm. Images were
deconvolved and maximum intensity projection were rendered using Huygens®
Essential (Hilversum, The Netherlands).
3.8. Field emission scanning electron microscopy (FESEM) Association of HEp-2 cell monolayers with either 10cpsΔEF or 10cpsΔEFΔsly at 100
bacteria per cell were investigated with FESEM as described previously (Benga et
al., 2004).
3.9. Cell-permeability and macropore-formation assay Experiments were performed in PBS containing 0.9 mM CaCl2/0.5 mM MgCl26H2O
[pH 7.3] (Gibco-Invitrogen). Cell permeability was determined after co-incubation of
HEp-2 cells (4 x 105 cells/ml) with recombinant proteins (rSLY, rW461F, and rSVD) at
subcytolytic (100 ng/ml) or lytic (400 ng/ml) concentrations and the fluorescent
marker calcein (2 µg/ml). Incubation was performed for 2 h at 37°C on an end-over-
end rotator. After two washing steps with PBS to remove extracellular calcein, cells
were fixed with 0.37% formaldehyde (for 10 min at room temperature and afterwards
resuspended in 500 µl PBS for measurement of fluorescent cells using FACScan®.
Further analysis was performed with the software WinMDI. For each determination at
least 10,000 events were measured. Initial analysis of fluorescent cells was carried
out by dot plot analysis (forward scatter [fsc] versus sideward scatter [ssc]) to define
the cell population of interest (data not shown). Subsequently, fluorescent cells were
detected at channel FL-1. Data were expressed in comparison to cells only treated
with calcein (background control).

Material and methods Chapter 2
50
3.10. Detection of α5β1 integrin expression on HEp-2 cells HEp-2 cell-suspension was incubated with either a monoclonal anti-human CD29
antibody or a monoclonal anti-human CD49e (Bio Legend) both used at a
concentration of 500 ng per 1.5 x 106 cells (Bio Legend, Fell, Germany). All
antibodies were diluted in blocking buffer (PBS [pH 7.3] containing 3% BSA) and
incubation was performed for 30 min at room temperature with gently agitation. After
three washing steps with blocking buffer, cells were incubated with a TRITC-
conjugated goat anti-mouse antibody (1:1,000, Dianova, Hamburg, Germany).
Stained cells were washed three times, fixed with 0.37% formaldehyde and
resuspended in blocking buffer for subsequent measurement. Flow cytometry
measurement was performed as described above. Fluorescent cells were counted at
channel FL-1 and results were expressed in comparison to unstained cells
(background control).
3.11. Haemolysis assay The haemolytic activities of rSLY, rW461F or rSVD were determined by a haemolysis
assay, performed essentially as described by Takamatsu et al. (2001). Titration of
haemolytic activity was performed in a 96-well vee bottom microplate. Twofold
dilutions of the toxins were incubated with a 2% sheep erythrocyte suspension in
0.9% NaCl for 2 h at 37°C mixed gently. Unlysed red blood cells were allowed to
pellet by centrifugation and 100 µl of the supernatant was transferred into a new flat
bottom microplate. Subsequently, absorption was measured at 550 nm in a
microplate reader (GENios Pro, TECAN AUSTRIA GMBH). Determination of
haemolytic activity was done in three independent experiments. One haemolytic unit
(HU) was defined as the concentration of toxin causing 50% haemolysis of a 2%
sheep blood suspension.
3.12. Cytotoxicity assays Cytotoxicity was detected by an LDH-release assay as described previously (Benga
et al., 2004). HEp-2 cells were either incubated with rSLY or rW461F. Supernatants
were removed 2 h after treatment and their LDH activities were determined using the

Material and methods Chapter 2
51
Cytotox® 96 assay kit (Promega, Mannheim, Germany). The experiments were
performed in triplicates and repeated at least three times. To quantify relative cellular
damage, results were expressed relative to LDH release observed after Triton X-100
(Biorad, Muenchen, Germany) lysis of non-infected cells.
To determine haemolytic and cytotoxic activity of heterologous expressed SLY,
W461F or SVD in L. lactis, respective lactococci (L. lactis pORI-SLY, -W461F and -
SVD) were grown in GM17 medium overnight, adjusted to an optical density (OD600)
of 0.02 in pre-warmed media the next day and then grown to late exponential growth
phase (OD600 0.8). Subsequently bacteria were removed by centrifugation and
culture supernatants were concentrated and purified using Amicon centrifHugal filter
devices 30 kDa (Millipore, Schwalbach/Ts., Germany) to remove remaining medium
components. Concentrated culture supernatants diluted 1:5 in 0.9% NaCl were used
as test samples.
3.13. Pull down experiments Experiments were done as described previously with some modifications (Reid et al.,
1996). HEp-2 cells were incubated with rSLY or rW461F (100 ng/ml) for indicated
time periods. Then cells were rinsed twice with ice cold PBS to stop incubation. Lysis
of cells was done by addition of 1 ml of ice cold lysis buffer (50 mM NaCl, 20 mM
Tris-HCl [pH 7.4], 3 mM MgCl2, 1% Nonidet-P 40, 0.25% Triton X-100, 5 mM
dithiothreitol, 100 µM PMSF). Cells were scraped off, and the lysates were
centrifuged at 14,000 rpm. The supernatant was split and used for pull down
experiments of activated RhoA and Rac1 in parallel. For this, 20 µl of beads slurry
bearing approximately 20 µg protein of the GST-fusion protein of either the Rho-
binding domain C21 or the PAK-GBD, respectively, were added to 500 µl sample and
incubated on a rotator at 4°C for 30 min. Beads were collected by centrifugation at
10,000 rpm, washed twice with lysis buffer and subjected to immunoblot analysis
using monoclonal antibodies against Rac-1 (cloe 102; BD Pharmingen, Heidelberg,
Germany) and against RhoA (clone 26C4; Santa Cruz, Heidelberg, Germany),
respectively. Amounts of (activated and total) GTPases were quantified by
densitometric measurement of three independent immunoblot analyses.

Material and methods Chapter 2
52
3.14. GLISA HEp-2 cells were treated with rSLY, rW461F or rSVD at subcytolytic concentration
(100 ng/ml) for 15 min and rinsed twice with ice cold PBS to stop incubation. Lysis of
cells was achieved by addition of 1 ml of ice cold lysis buffer (50 mM NaCl, 20 mM
Tris-HCl, pH 7.4, 3 mM MgCl2, 1% Nonidet-P 40, 0.25% Triton X-100, 5 mM
dithiothreitol, 100 µM PMSF). Cells were scraped off, and the lysates were
centrifuged at 14,000 rpm. The supernatant was used for a pull down-based GLISA
(Cytoskeleton, CO, USA) to determine activation of Rac1, RhoA, and Cdc42
according to the protocol by the supplier.
4. Mouse infection model 4.1. Preparation of infection culture For infection of mice, streptococci were grown in Todd-Hewitt broth (THB, Difco,
Detroit, USA) overnight at 37°C under aerobic conditions, adjusted to an optical
density (OD600) of 0.02 in pre-warmed media the next day and then grown to late
exponential growth phase (OD600 0.8). Streptococci were harvested by centrifugation,
resuspended in sterile PBS (pH 7.4) and adjusted to the final concentration of
5 x 1011 CFU/ml for intranasal infection. Inoculum concentrations were verified by
platting 10-fold serial dilutions on Colombia agar plates with 7% sheep blood after
infection.
4.2. Intranasal infection of mice Four (expt. 1 to 4) or six (expt. 5) week old specific-pathogen-free female mice of the
outbreed strain Crl:CD1 (ICR) were obtained from Charles River Laboratories
(Sulzfeld, Germany). Mice of the Staphylococcus (St.) aureus free outbred strain
Hsd:ICR (CD1®) were purchased from Harlan Laboratories (AN Venray, The
Netherlands). Animals were randomly divided into groups consisting of five to six
animals each. Mice were allowed to acclimate for one week and cared for in
accordance with the principles outlined in the European Convention for the Protection
of Vertebrate Animal Used for Experimental and Other Scientific Purposes [European
Treaty Series, no. 123: http://conventions.coe.int/treaty/EN/Menuprincipal.htm; permit

Material and methods Chapter 2
53
no.33.9-42502-04-08/1589]. Before infection mice were anesthetized via inhalation of
isofluran (IsoFlo®, Albrecht, Germany). In experiment 2 to 5 mice were pre-treated
with 12.5 µl 1% acetic acid [pH 4.0] placed in each nostril 1 h prior intranasal
infection. After a controlled recovery phase and re-anaesthesia per isofluran
inhalation, mice were infected with 5 x 109 CFU of either S. suis wild type strain 10,
10Δsly, 10cpsΔEF or 10cpsΔEFΔsly. The dose was applied in two drops of 12.5 µl
volume placed in front of the nostrils.
4.3. Intravenous infection of mice Five- to seven-week-old specific-pathogen-free female inbreed strain C57/BL6 mice
and outbreed strain Crl:CD1 (ICR) mice were purchased from Charles River
(Sulzfeld, Germany), randomly divided into groups (five to six mice each) and kept in
conformity with the European conventions. After an adjustment period of one week,
mice were infected via the lateral tail vein with ~6 x 105 CFU, 1x108 CFU, 5 x 108
CFU, and 1 x 109 CFU per individual (total volume of 100 µl per ventral tail vein).
4.4. Clinical score Animals were clinically scored every 8 h. The health status was rated using a clinical
score sheet (Table 5-1), including weight development, clinical signs of general
sickness (rough coat, rapid breathing, dehydration), clinical signs indicating
meningitis (apathy, apraxia), and septicaemia (swollen eyes, depression), developed
on the basis of mouse clinical monitoring score described by the Research office of
the Australian National University [http://www.anu.edu.au/ro/ORI/Animal/AEEC001_
MouseMonitoringSOP.doc]. A cumulative score of 3 to 4 indicated mild clinical
symptoms, a score of 5 to 6 moderate clinical symptoms and a score greater than 6
severe clinical symptoms, in particular persistent anorexia, apathy, and/or neural
disorder. Mice with a cumulative score equal or greater than 3 were classified as
diseased (calculation of morbidity). In the case of severe weight loss (> 20%) and/or
enduring severe clinical signs, mice were euthanized for reasons of animal welfare
by inhalation of CO2 and cervical dislocation.

Material and methods Chapter 2
54
4.5. Histological screening Immediately after euthanasia, necropsy was conducted and the organs were
aseptically removed and split for histological and bacteriological screenings, including
spleen, liver, kidney, heart, lung, brain, spinal cord, and nose. The histological
screenings were carried out as blind experiments. Findings were scored as described
for piglets (Baums et al., 2006). In contrast to piglets, in addition to fibrinosuppurative
lesions, purulent necrotizing lesions associated with the challenge strain were scored
as well (see results). Rhinitis was not included in the general score ω designed to
reflect lesions caused by invading streptococci.
4.6. Reisolation of S. suis strains from tissue and tracheo-nasal lavage (TNL) One half of each organ was suspended in 5 ml PBS [pH 7.4] and weighed. All organs
were homogenized with an Ultra Turrax (IKA, Staufen, Germany). Ten-fold serial
dilutions of samples were plated on blood agar plates. Colony forming units (CFU)
were counted the next day after incubation at 37°C for 24 h and CFU per mg organ
was determined.
For sampling TNL the trachea was opened and a retrograde irrigation of the nasal
cavity with 300 µl PBS was collected. Number of typical α-haemolytic streptococci
per µl TNL was determined by serial plating on blood agar. Isolated α-haemolytic
streptococci were investigated in a S. suis multiplex PCR for the detection of mrp,
epf, sly, arcA, gdh, cps1, cps2, cps7, and cps9 (Silva et al., 2006). Isolates received
from mice challenged either with strain 10Δsly, 10cpsΔEF or 10cpsΔsly were
additionally tested in a cps2E-specific PCR (with oligonucleotide primers cps2Efor
(TTTCGCACTTTCAAGACGTG) and cps2Erev (GGACGGGTACCGACTAGACTC)
and sly-specific PCR using primers slyAgefor (TGTACCGGTGATTCCAAACAAGA
TATTAA) and slyAge3new (TTAACCGGTTACTCTATCACCTCATCCG). Based on
CFU per mg organ or per µl TNL, bacterial loads were classified as mild (+; <100),
moderate (++; >100 - <1000) or severe (+++; >1000).

Material and methods Chapter 2
55
5. Statistical analyses If not stated otherwise, experiments were performed at least three times and results
were expressed as means and standard deviations. Data were analyzed by t-test.
Pull down analysis was carried out using an unpaired t-test with no assumption of
Gaussian distribution (Mann-Whitney). A P-value <0.05 was considered significant.

Material and methods Chapter 2
56
References
Baums, C. G., Kaim, U., Fulde, M., Ramachandran, G., Goethe, R., and Valentin-Weigand, P. (2006) Identification of a novel virulence determinant with serum opacification activity in Streptococcus suis. Infect Immun 74: 6154-6162.
Benga, L., Fulde, M., Neis, C., Goethe, R., and Valentin-Weigand, P. (2008) Polysaccharide capsule and suilysin contribute to extracellular survival of Streptococcus suis co-cultivated with primary porcine phagocytes. Vet Microbiol 132: 211-219.
Benga, L., Goethe, R., Rohde, M., and Valentin-Weigand, P. (2004) Non-encapsulated strains reveal novel insights in invasion and survival of Streptococcus suis in epithelial cells. Cellular Microbiology 6: 867-881.
Holo, H. and Nes, I. F. (1989) High-Frequency Transformation, by Electroporation, of Lactococcus lactis subsp. cremoris Grown with Glycine in Osmotically Stabilized Media. Appl Environ Microbiol 55: 3119-3123.
Just, I., Selzer, J., Wilm, M., Eichel-Streiber, C., Mann, M., and Aktories, K. (1995) Glucosylation of Rho proteins by Clostridium difficile toxin B. Nature 375: 500-503.
Kock, C., Beineke, A., Seitz, M., Ganter, M., Waldmann, K. H., Valentin-Weigand, P., and Baums, C. G. (2009) Intranasal immunization with a live Streptococcus suis isogenic ofs mutant elicited suilysin-neutralization titers but failed to induce opsonizing antibodies and protection. Veterinary Immunology and Immunopathology 132: 135-145.
Logan, R. P., Robins, A., Turner, G. A., Cockayne, A., Borriello, S. P., and Hawkey, C. J. (1998) A novel flow cytometric assay for quantitating adherence of Helicobacter pylori to gastric epithelial cells. J Immunol Methods 213: 19-30.
Que, Y. A., Haefliger, J. A., Francioli, P., and Moreillon, P. (2000) Expression of Staphylococcus aureus clumping factor A in Lactococcus lactis subsp. cremoris using a new shuttle vector. Infect Immun 68: 3516-3522.
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N Y.
Silva, L. M., Baums, C. G., Rehm, T., Wisselink, H. J., Goethe, R., and Valentin-Weigand, P. (2006) Virulence-associated gene profiling of Streptococcus suis isolates by PCR. Vet Microbiol 115: 117-127.
Smith, H. E., Damman, M., van der Velde, J., Wagenaar, F., Wisselink, H. J., Stockhofe-Zurwieden, N., and Smits, M. A. (1999) Identification and characterization of the cps locus of Streptococcus suis serotype 2: the capsule protects against phagocytosis and is an important virulence factor. Infect Immun 67: 1750-1756.
Takamatsu, D., Osaki, M., and Sekizaki, T. (2001) Thermosensitive suicide vectors for gene replacement in Streptococcus suis. Plasmid 46: 140-148.
Vecht, U., StockhofeZurwieden, N., Tetenburg, B. J., Wisselink, H. J., and Smith, H. E. (1997) Virulence of Streptococcus suis type 2 for mice and pigs appeared host-specific. Veterinary Microbiology 58: 53-60.

Chapter 3
Results, part I:
Subcytolytic suilysin promotes invasion of Streptococcus suis in HEp-2
epithelial cells by Rac1- dependent activation of the actin cytoskeleton
M. Seitz, C. G. Baums, R. Gerhard, I. Just, C. Neis, L. Benga, M. Fulde, A. Nerlich,
M. Rohde, R. Goethe and P. Valentin-Weigand
Running title: Subcytoytic suilysin promotes invasion of S. suis
(Manuscript submitted)


Results, part I Chapter 3
59
Abstract Suilysin is a cholesterol-dependent pore-forming cytolysin secreted by Streptococcus
suis, an important swine and zoonotic pathogen. The role of suilysin in S. suis-host
cell interaction is still unclear. In the present study we found a higher invasion rate in
epithelial cells of an unencapsulated sly-positive strain compared to its sly-negative
mutant. Electron microscopy revealed that uptake-accompanying membrane ruffling
was abolished in the sly-negative mutant. Since invasion of S. suis depended on
actin polymerization, we used specific inhibitors of Rho-GTPases and could show
that Rac, but not Rho was involved in suilysin-mediated uptake. Accordingly, pull
down analysis with specific ligands demonstrated activation of Rac, but not Rho in
suilysin-treated cells. Furthermore, confocal microscopy revealed colocalisation of
suilysin with F-actin and Rac1. To further dissect the relevance of cytolytic pore-
formation we produced recombinant suilysin in which the protein domain responsible
for membrane binding and pore-formation was inactivated via substitution of a
tryptophan by phenylalanine. The mutated suilysin had lost haemolytic and cytolytic
activity but retained its ability to mediate S. suis invasion and activation of Rac.
Concluding, our results indicate that subcytolytic suilysin promotes S. suis invasion in
epithelial cells by Rac-dependent activation of the actin cytoskeleton.

Results, part I Chapter 3
60
Introduction Streptococcus (S.) suis is one of the most important swine pathogens worldwide
causing meningitis, arthritis, septicaemia, bronchopneumonia, and other pathologies.
Furthermore, S. suis colonizes the nasopharynx and other mucosal surfaces,
resulting in high carrier rates of healthy pigs (Arends et al., 1984; O'Sullivan et al.,
2011). S. suis is also an important zoonotic agent (Trottier et al., 1991; Gottschalk et
al., 2007). Meningitis and septicaemia are the most common manifestations in
humans, occurring particularly in people exposed to pigs or pig-products (Arends and
Zanen, 1988). Two human outbreaks in China in 1998 and 2005 were associated
with increased severeness of clinical symptoms, a high rate of mortality, and
streptococcal toxic shock-like syndrome (Tang et al., 2006). Among the 33 serotypes
of S. suis, serotype 2 is worldwide most frequently isolated from diseased pigs
(Wisselink et al., 2000) and humans (Gottschalk et al., 2010). Mechanisms
underlying pathogenesis of S. suis infections, however, are only poorly known.
Suilysin was identified as haemolysin of S. suis several years ago (Jacobs et al.,
1994). It is a member of the cholesterol-dependent pore-forming cytolysins (CDC)
family and expressed by many S. suis strains. The sly gene has been detected in
95% of European and Asian invasive serotype 2 strains (Segers et al., 1998), and it
was found in 69.4% of isolates from 10 different serotypes. Though these isolates
were mainly obtained from porcine cases of meningitis and septicaemia (King et al.,
2001), experimental infections demonstrated attenuation of a sly knock-out mutant
only in mice, but not in piglets (Allen et al., 2001; Lun et al., 2003).
Similar to other members of the CDC family suilysin can damage host cells by its
cytolytic activity (Norton et al., 1999; Charland et al., 2000; Lalonde et al., 2000;
Segura and Gottschalk, 2002; Vanier et al., 2004; Tenenbaum et al., 2005;
Tenenbaum et al., 2006). It has also been suggested that suilysin plays a role in
invasion and pathogenesis of S. suis (Norton et al., 1999). More recently, it has been
found that the toxin may be involved in cytokine release and protection against
opsonophagocytosis (Benga et al., 2008; Lecours et al., 2011). Some of the other
CDC have been shown to display biological effects at sublytic concentrations, e. g.

Results, part I Chapter 3
61
phosphorylation of p38 mitogen-activated protein kinase (MAPK) in epithelial cells,
which is crucial for local production of IL-8 and subsequent recruitment of neutrophils
to the site of infection (Ratner et al., 2006). For the CDC of S. pneumoniae
(pneumolysin), Listeria monocytogenes (listeriolysin O), and S. intermedius
(intermedilysin) it was suggested that they contribute to bacterial adherence and
invasion (Rubins et al., 1998; Sukeno et al., 2005; Krawczyk-Balska and Bielecki,
2005). These findings indicate that CDC express subcytolytic activities that may
modify host cell responses to infection. For suilysin, however, such activities and
their possible biological relevance still await to be elucidated in detail.
The objective of this study was to evaluate the possible role of suilysin in host cell
interaction of S. suis, in particular its impact on association of S. suis with respiratory
epithelial cells, i. e. adherence and invasion. Using respective mutant strains and
HEp-2 epithelial cells we identified a suilysin-dependent invasive phenotype of an
unencapsulated S. suis serotype 2 strain and determined host cell GTPases involved
in suilysin-mediated effects. Furthermore, we showed that these effects did not
require formation of a cytolytic pore.

Results, part I Chapter 3
62
Results Suilysin promotes adherence and invasion of S. suis. The present study was
undertaken to investigate the possible role of the cytolytic toxin suilysin, since its role
in virulence of S. suis is poorly understood. For this, we constructed a suilysin-
negative mutant of the unencapsulated strain 10cpsΔEF (kindly provided by H. E.
Smith, Lelystad, NL). We used this strain because it originated from a highly virulent
serotype 2 strain and displays a much better epithelial cell adherence and invasion
than encapsulated strains, as observed in our previous studies (Benga et al., 2004).
We compared the sly-positive strain with its sly-negative mutant (10cpsΔEFΔsly) in
adherence and invasion using the respiratory epithelial cell line HEp-2. Under our
experimental conditions we could exclude cytotoxic effects of suilysin, as confirmed
by viability staining of HEp-2 cells with propidium iodide (data not shown). First we
determined bacterial association with HEp-2 cells by FACS of CFSE-labelled
streptococci, which had been incubated with epithelial cells for 2 h. The
unencapsulated mutant showed a high association with HEp-2 cells (mean
fluorescence intensity [mfi] of 10.4 as compared to 3.1 of uninfected cells). The
sly-negative mutant of this strain demonstrated significantly lower association (mfi of
7.2, P = 0.0092; Figure 3-1A). Secondly, we performed antibiotic protection assays to
differentiate adherence and invasion of both S. suis strains. Results revealed that the
sly-negative strain adhered to a much lower degree and showed a significantly
reduced invasion as compared to the sly-positive strain (Figure 3-1B and C). Notably,
differences in adherence were not significant due to high standard deviations, but
were significant with respect to invasion. Thus, we assume that differences seen in
FACS analyses (Figure 3-1A) reflect differences in invasion capacity rather than in
adherence. Detection of suilysin by immunoblot analysis confirmed that suilysin was
expressed under these conditions only by strain 10cpsΔEF, but not by its sly-
negative mutant (Figure 3-1C, inlay). Thirdly, we compared both strains by double
immunofluorescent microscopy (DIF) for differentiation of extra- and intracellular
streptococci. Results confirmed that the sly-positive strain adhered and invaded HEp-
2 cells much better than the sly-negative mutant, which could be detected only

Results, part I Chapter 3
63
extracellularly (Figure 3-1D and E). Quantification of microscopic examinations by
counting of intracellular bacteria per HEp-2 cell confirmed that these differences were
significant (P = 0.042; Figure 3-1F). Finally, we analysed adherence and invasion
process of both strains by field emission scanning electron microscopy (FESEM).
Similar to what we have found in previous studies (Benga et al., 2004), uptake of
unencapsulated sly-positive S. suis was accompanied by formation of membrane
ruffles in close vicinity to the streptococci (Figure 3-1G and H). Interestingly, this was
observed only for the sly-positive strain, but not for the sly-negative strain, which
showed adherence to cells but no association with ruffles or other cell morphologies
reminiscent of invasion (Figure 3-1I). These findings suggest that suilysin contributes
to S. suis induced membrane ruffling and uptake by epithelial cells.
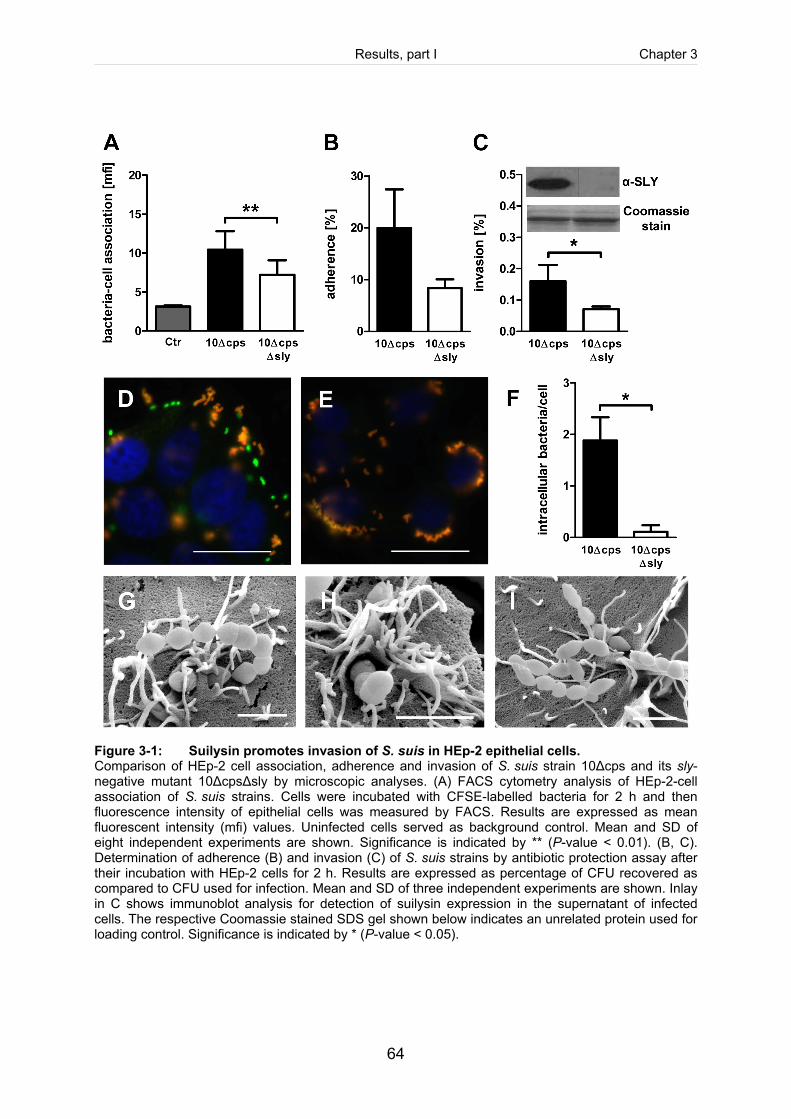
Results, part I Chapter 3
64
Figure 3-1: Suilysin promotes invasion of S. suis in HEp-2 epithelial cells. Comparison of HEp-2 cell association, adherence and invasion of S. suis strain 10Δcps and its sly-negative mutant 10ΔcpsΔsly by microscopic analyses. (A) FACS cytometry analysis of HEp-2-cell association of S. suis strains. Cells were incubated with CFSE-labelled bacteria for 2 h and then fluorescence intensity of epithelial cells was measured by FACS. Results are expressed as mean fluorescent intensity (mfi) values. Uninfected cells served as background control. Mean and SD of eight independent experiments are shown. Significance is indicated by ** (P-value < 0.01). (B, C). Determination of adherence (B) and invasion (C) of S. suis strains by antibiotic protection assay after their incubation with HEp-2 cells for 2 h. Results are expressed as percentage of CFU recovered as compared to CFU used for infection. Mean and SD of three independent experiments are shown. Inlay in C shows immunoblot analysis for detection of suilysin expression in the supernatant of infected cells. The respective Coomassie stained SDS gel shown below indicates an unrelated protein used for loading control. Significance is indicated by * (P-value < 0.05).

Results, part I Chapter 3
65
(D-F). Determination of extra- and intracellular bacteria by double immunofluorescence (DIF) microscopy after incubation of S. suis strains with HEp-2 cells. In (D) cells were infected with strain 10Δcps, in (E) strain 10ΔcpsΔsly was used for infection. Extracellular bacteria are shown in orange-red and intracellular bacteria are stained in green. The nucleus was labelled by DAPI (blue). Bars represent 15 µm. (F) shows results of quantification of intracellular streptococci per cell by counting green (intracellular) bacteria in 50 epithelial cells (F). Mean and SD of two independent experiments are shown. Significance is indicated by * (P-value < 0.05). (G-I). Analysis of association of S. suis strains with HEp-2 cells by field emission scanning electron microscopy (FESEM). In (G) and (H) strain 10Δcps is shown to be associated with the formation of membrane ruffles. Strain 10ΔcpsΔsly adhered to the cell surface, but did not induce membrane ruffling or uptake by HEp-2 cells (I). Bars represent 1 µm. Invasion-promoting activity of suilysin involves Rac-dependent activation of the actin cytoskeleton. Based on our results described above we hypothesized that
suilysin promotes invasion of S. suis by manipulating the host cell cytoskeleton.
Since actin is the major component involved in formation of lammellipodia and
membrane ruffles, we first performed experiments with latrunculin B, a specific
inhibitor of actin polymerisation (Coue et al., 1987; Greenwood et al., 2006). As
shown in Figure 3-2A, FACS analysis revealed that bacteria-cell association of the
sly-positive strain was significantly reduced in the presence of latrunculin B as
compared to control cells without inhibitor. Reduced cell-association reached a level
which was almost similar to that of the sly-negative mutant (Figure 3-2A).
Next we studied the role of the small GTPases Rho, Rac, and Cdc42. These belong
to the family of Rho-GTPases which are molecular switches signalling to the actin
cytoskeleton, thereby translating environmental signals into cellular morphological
responses (Hall, 1998). We performed bacterial invasion experiments with HEp-2
cells, which were pretreated with different inhibitors of Rho-GTPases. First we used
toxin B of Clostridium (C.) difficile (TcdB), which inhibits all Rho-GTPases by
monoglycosylation (Just et al., 1995). The treatment of cells with TcdB resulted in a
significant reduction of invasion of strain 10cpsΔEF as determined by gentamicin
protection assays (Figure 3-2B). In contrast, the invasion of the isogenic sly-negative
strain 10cpsΔEFΔsly was not affected. Next we tested invasion of HEp-2 cells
pretreated with C3 exoenzyme, an ADP-ribosyltransferase of C. limosum, which
specifically inactivates Rho-, but not Rac- and Cdc42-GTPase. This treatment had no
effect on invasion of either of the two S. suis strains (Figure 3-2C). Thirdly, we used a
specific inhibitor of Rac1-GTPase, TcdB toxin of C. difficile serotype F strain 1470
(TcdB-F). TcdB-F pretreatment of epithelial cells resulted in a significant inhibition of
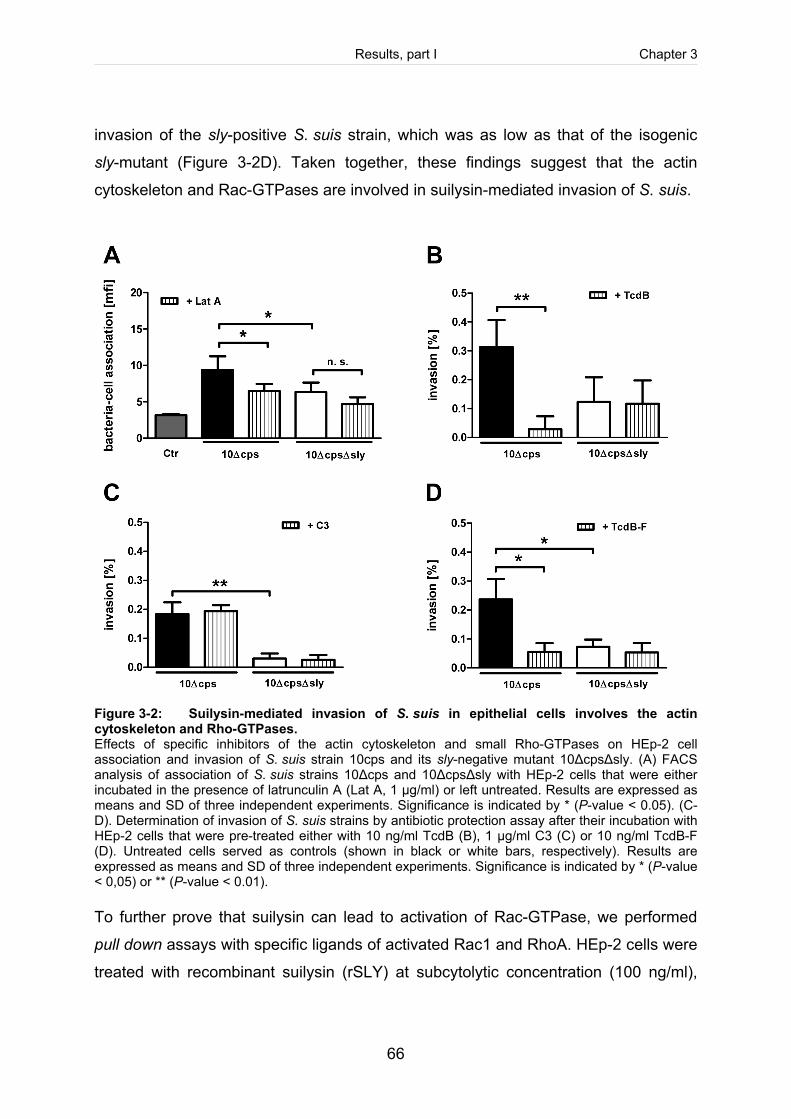
Results, part I Chapter 3
66
invasion of the sly-positive S. suis strain, which was as low as that of the isogenic
sly-mutant (Figure 3-2D). Taken together, these findings suggest that the actin
cytoskeleton and Rac-GTPases are involved in suilysin-mediated invasion of S. suis.
Figure 3-2: Suilysin-mediated invasion of S. suis in epithelial cells involves the actin cytoskeleton and Rho-GTPases. Effects of specific inhibitors of the actin cytoskeleton and small Rho-GTPases on HEp-2 cell association and invasion of S. suis strain 10cps and its sly-negative mutant 10ΔcpsΔsly. (A) FACS analysis of association of S. suis strains 10Δcps and 10ΔcpsΔsly with HEp-2 cells that were either incubated in the presence of latrunculin A (Lat A, 1 µg/ml) or left untreated. Results are expressed as means and SD of three independent experiments. Significance is indicated by * (P-value < 0.05). (C-D). Determination of invasion of S. suis strains by antibiotic protection assay after their incubation with HEp-2 cells that were pre-treated either with 10 ng/ml TcdB (B), 1 µg/ml C3 (C) or 10 ng/ml TcdB-F (D). Untreated cells served as controls (shown in black or white bars, respectively). Results are expressed as means and SD of three independent experiments. Significance is indicated by * (P-value < 0,05) or ** (P-value < 0.01). To further prove that suilysin can lead to activation of Rac-GTPase, we performed
pull down assays with specific ligands of activated Rac1 and RhoA. HEp-2 cells were
treated with recombinant suilysin (rSLY) at subcytolytic concentration (100 ng/ml),
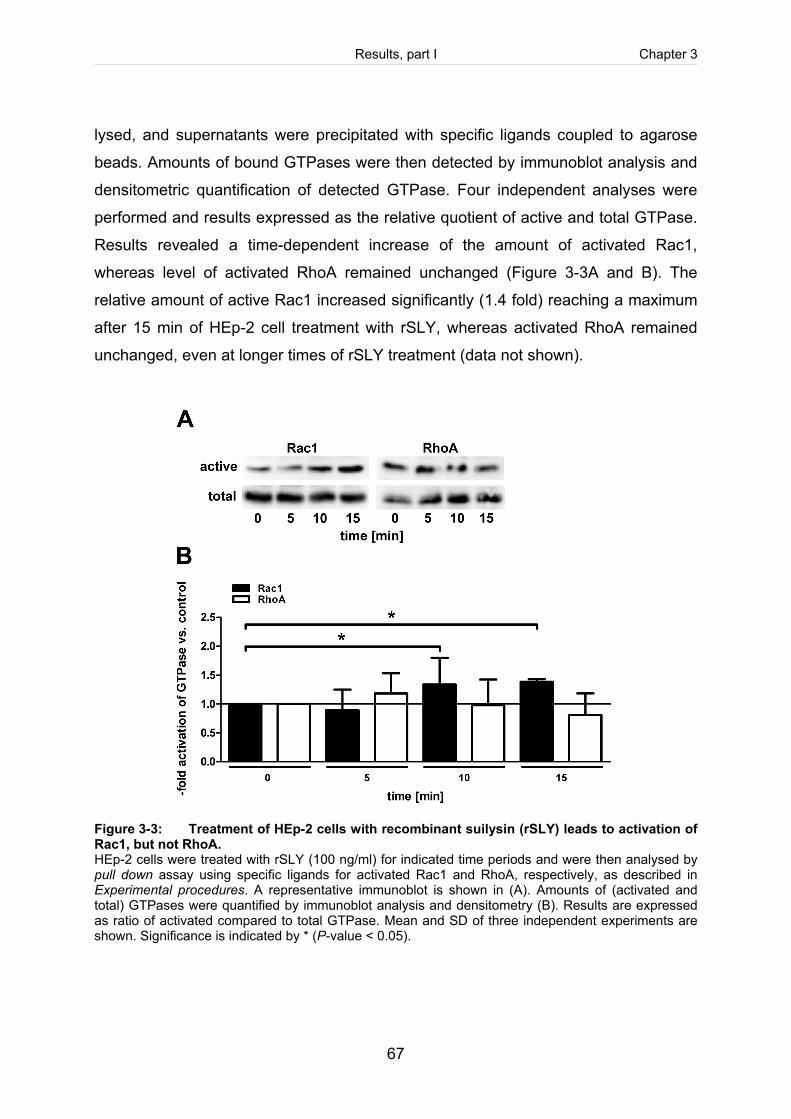
Results, part I Chapter 3
67
lysed, and supernatants were precipitated with specific ligands coupled to agarose
beads. Amounts of bound GTPases were then detected by immunoblot analysis and
densitometric quantification of detected GTPase. Four independent analyses were
performed and results expressed as the relative quotient of active and total GTPase.
Results revealed a time-dependent increase of the amount of activated Rac1,
whereas level of activated RhoA remained unchanged (Figure 3-3A and B). The
relative amount of active Rac1 increased significantly (1.4 fold) reaching a maximum
after 15 min of HEp-2 cell treatment with rSLY, whereas activated RhoA remained
unchanged, even at longer times of rSLY treatment (data not shown).
Figure 3-3: Treatment of HEp-2 cells with recombinant suilysin (rSLY) leads to activation of Rac1, but not RhoA. HEp-2 cells were treated with rSLY (100 ng/ml) for indicated time periods and were then analysed by pull down assay using specific ligands for activated Rac1 and RhoA, respectively, as described in Experimental procedures. A representative immunoblot is shown in (A). Amounts of (activated and total) GTPases were quantified by immunoblot analysis and densitometry (B). Results are expressed as ratio of activated compared to total GTPase. Mean and SD of three independent experiments are shown. Significance is indicated by * (P-value < 0.05).
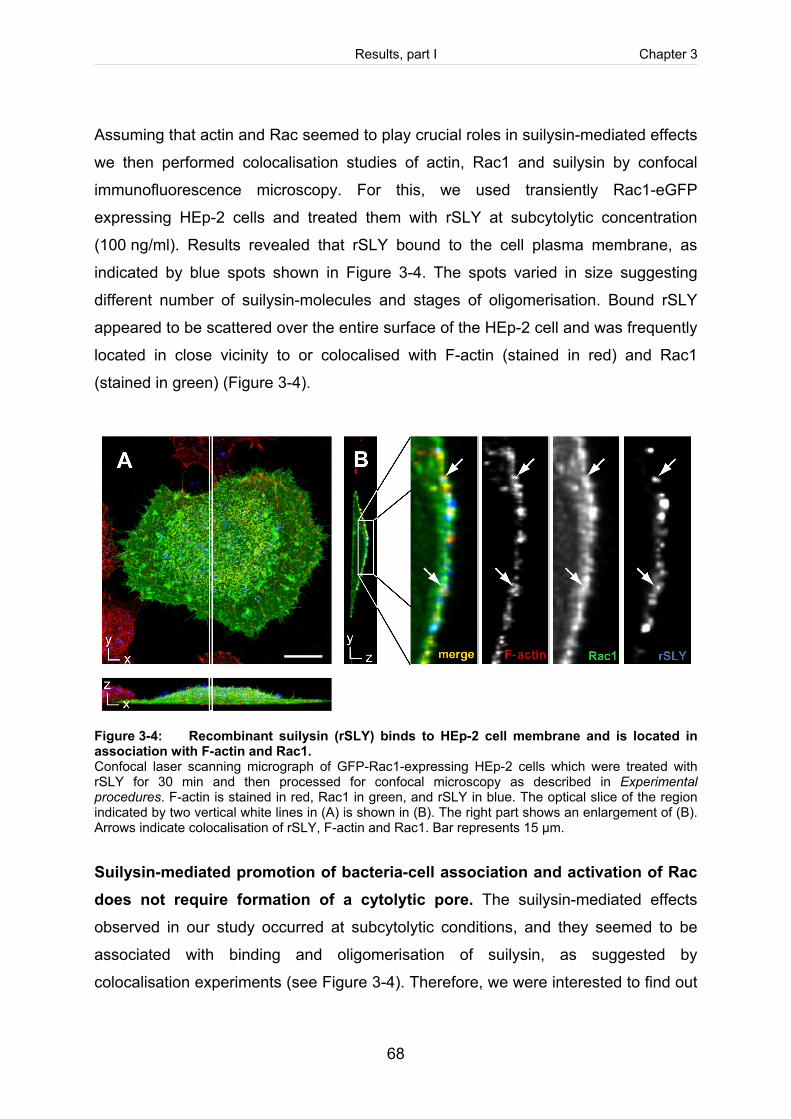
Results, part I Chapter 3
68
Assuming that actin and Rac seemed to play crucial roles in suilysin-mediated effects
we then performed colocalisation studies of actin, Rac1 and suilysin by confocal
immunofluorescence microscopy. For this, we used transiently Rac1-eGFP
expressing HEp-2 cells and treated them with rSLY at subcytolytic concentration
(100 ng/ml). Results revealed that rSLY bound to the cell plasma membrane, as
indicated by blue spots shown in Figure 3-4. The spots varied in size suggesting
different number of suilysin-molecules and stages of oligomerisation. Bound rSLY
appeared to be scattered over the entire surface of the HEp-2 cell and was frequently
located in close vicinity to or colocalised with F-actin (stained in red) and Rac1
(stained in green) (Figure 3-4).
Figure 3-4: Recombinant suilysin (rSLY) binds to HEp-2 cell membrane and is located in association with F-actin and Rac1. Confocal laser scanning micrograph of GFP-Rac1-expressing HEp-2 cells which were treated with rSLY for 30 min and then processed for confocal microscopy as described in Experimental procedures. F-actin is stained in red, Rac1 in green, and rSLY in blue. The optical slice of the region indicated by two vertical white lines in (A) is shown in (B). The right part shows an enlargement of (B). Arrows indicate colocalisation of rSLY, F-actin and Rac1. Bar represents 15 µm.
Suilysin-mediated promotion of bacteria-cell association and activation of Rac does not require formation of a cytolytic pore. The suilysin-mediated effects
observed in our study occurred at subcytolytic conditions, and they seemed to be
associated with binding and oligomerisation of suilysin, as suggested by
colocalisation experiments (see Figure 3-4). Therefore, we were interested to find out

Results, part I Chapter 3
69
whether formation of a cytolytic pore was required for suilysin-mediated bacteria-cell
association and Rac-activation. In some CDC it has been shown by structural or
functional analyses that the tryptophan (Trp)-rich motif at the tip of the C-terminal of
domain 4 is crucial for cytolytic activity (Billington et al., 2000). Hence, we introduced
a point mutation in the sly gene by site-directed mutagenesis, resulting in a single
amino acid substitution of a conserved Trp residue at position 461 (W was replaced
by F). An alignment of the homologous Trp-rich motif of suilysin, the mutated suilysin
derivative W461F, and pneumolysin (PLY) is shown in Figure 3-5A. We first
compared haemolytic and cytolytic activities of rSLY and rW461F. Determination of
haemolytic activities showed that the point-mutated toxin rW461F almost completely
lost its capacity to lyse sheep erythrocytes. Compared to rSLY, of which 256 (28)
ng/ml were sufficient for 50 % haemolysis, rW461F had to be applied at 4096 (212)
ng/ml to cause 50 % haemolysis (Figure 3-5B). Calculation of haemolytic units (HU)
revealed 0.53 x 105 HU per mg of rSLY as compared to 0.25 x 104 HU/mg for
rW461F. In good agreement, cytotoxic activities determined by LDH release assay
using HEp-2 cells were abolished in rW461F, reaching only 2.1% of that of rSLY.
Figure 3-5C shows the respective dose-response curve and estimation of
concentrations needed for 50% cytotoxicity. Based on these findings, for the
following experiments we used rSLY and rW461F at 100 ng/ml, which was clearly
below the (rSLY) protein concentrations causing 50% cytotoxicity. These data
indicate that substitution of one amino acid in the conserved Trp-rich motif of domain
4 completely inactivates haemolytic and cytolytic activity of suilysin. To our
knowledge, this is the first experimental proof that the Trp-rich motif of suilysin is
crucial for formation of a functional (i. e. cytolytic) pore, which corresponds well to the
high similarities of this region within the CDC family (as shown for pneumolysin in
Figure 3-5A) as well as to predictions based on the crystal structure (Xu et al., 2010).
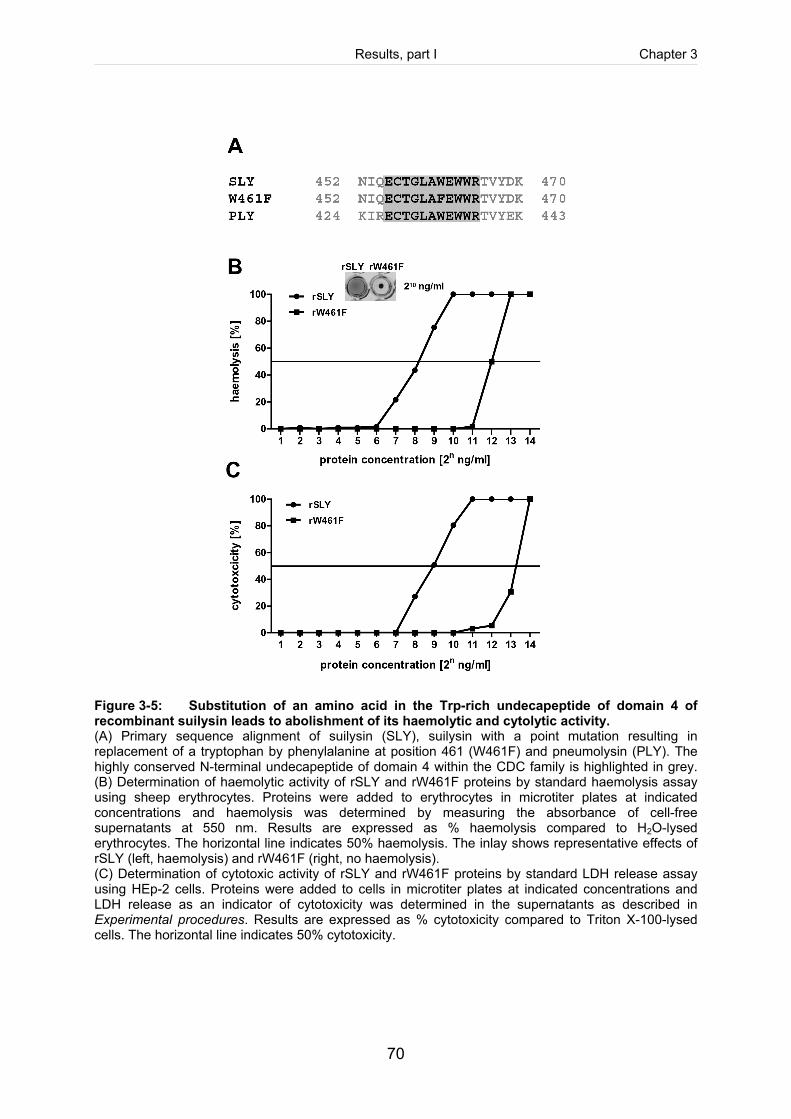
Results, part I Chapter 3
70
Figure 3-5: Substitution of an amino acid in the Trp-rich undecapeptide of domain 4 of recombinant suilysin leads to abolishment of its haemolytic and cytolytic activity. (A) Primary sequence alignment of suilysin (SLY), suilysin with a point mutation resulting in replacement of a tryptophan by phenylalanine at position 461 (W461F) and pneumolysin (PLY). The highly conserved N-terminal undecapeptide of domain 4 within the CDC family is highlighted in grey. (B) Determination of haemolytic activity of rSLY and rW461F proteins by standard haemolysis assay using sheep erythrocytes. Proteins were added to erythrocytes in microtiter plates at indicated concentrations and haemolysis was determined by measuring the absorbance of cell-free supernatants at 550 nm. Results are expressed as % haemolysis compared to H2O-lysed erythrocytes. The horizontal line indicates 50% haemolysis. The inlay shows representative effects of rSLY (left, haemolysis) and rW461F (right, no haemolysis). (C) Determination of cytotoxic activity of rSLY and rW461F proteins by standard LDH release assay using HEp-2 cells. Proteins were added to cells in microtiter plates at indicated concentrations and LDH release as an indicator of cytotoxicity was determined in the supernatants as described in Experimental procedures. Results are expressed as % cytotoxicity compared to Triton X-100-lysed cells. The horizontal line indicates 50% cytotoxicity.

Results, part I Chapter 3
71
Next we tested whether addition of both recombinant proteins to the sly-negative
S. suis mutant could reconstitute the phenotype of the sly-positive parental strain. For
this, we determined HEp-2 cell-association of S. suis strains by FACS in the
presence and absence of both recombinant proteins. Results revealed that addition
of rSLY and rW461F both resulted in regain of function of the sly-negative strain, i. e.
the mutant reached cell-association levels comparable to that of the sly-positive
strain when proteins were present (Figure 3-6A). Immunoblot analysis for detection of
recombinant suilysin protein in the supernatants of infected cells showed that rSLY
and rW461F were present in equal amounts and were comparable to suilysin
expressed by the sly-positive strain (Figure 3-6A, lower part). Finally, we tested
whether rW461F was still able to activate host cell GTPase Rac. For this, we
performed pull down experiments as described above. Results clearly showed that
rW461F treatment of HEp-2 cells led to an increase of the amount of activated Rac1
comparable to that induced by rSLY (Figure 3-6B).
Concluding, our results revealed that suilysin can promote invasion of S. suis in
epithelial cells by Rac-dependent activation of the actin cytoskeleton. Furthermore,
we showed that these effects occur at subcytolytic concentrations and do not require
formation of a functional (cytolytic) pore. Whether this can be considered as a
common mechanism of CDC producing streptococci remains to be an interesting
question for future studies.
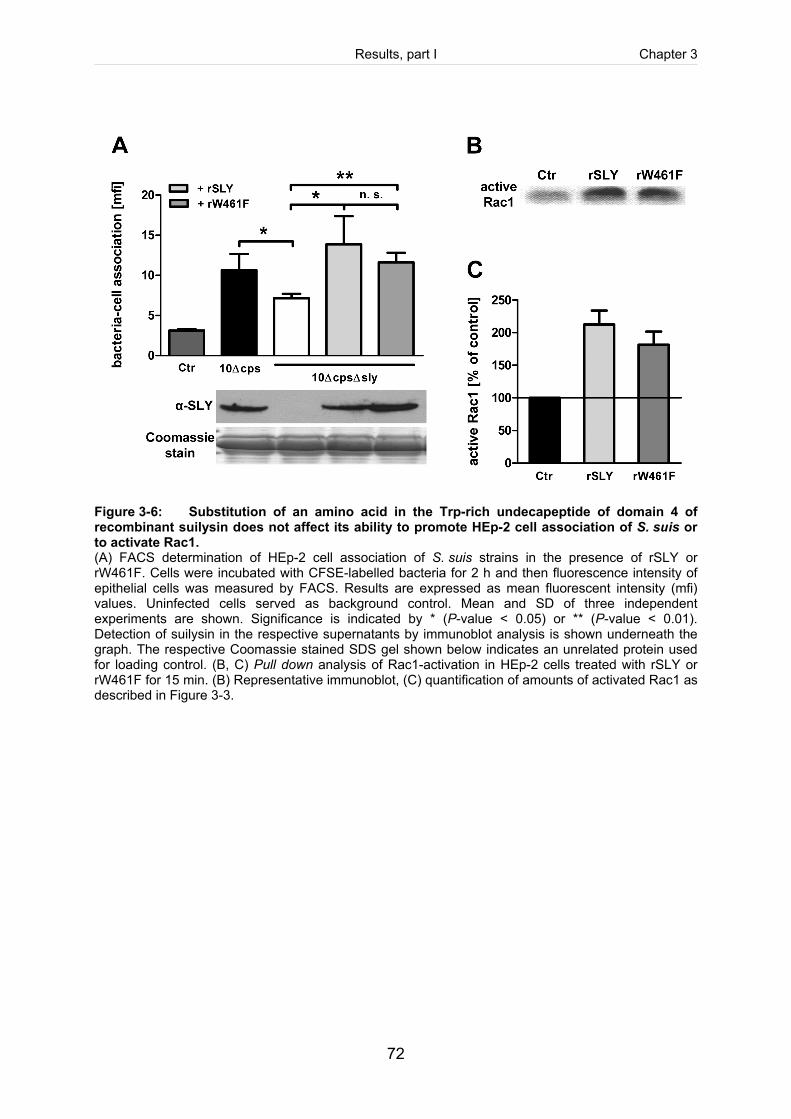
Results, part I Chapter 3
72
Figure 3-6: Substitution of an amino acid in the Trp-rich undecapeptide of domain 4 of recombinant suilysin does not affect its ability to promote HEp-2 cell association of S. suis or to activate Rac1. (A) FACS determination of HEp-2 cell association of S. suis strains in the presence of rSLY or rW461F. Cells were incubated with CFSE-labelled bacteria for 2 h and then fluorescence intensity of epithelial cells was measured by FACS. Results are expressed as mean fluorescent intensity (mfi) values. Uninfected cells served as background control. Mean and SD of three independent experiments are shown. Significance is indicated by * (P-value < 0.05) or ** (P-value < 0.01). Detection of suilysin in the respective supernatants by immunoblot analysis is shown underneath the graph. The respective Coomassie stained SDS gel shown below indicates an unrelated protein used for loading control. (B, C) Pull down analysis of Rac1-activation in HEp-2 cells treated with rSLY or rW461F for 15 min. (B) Representative immunoblot, (C) quantification of amounts of activated Rac1 as described in Figure 3-3.

Results, part I Chapter 3
73
Discussion Suilysin has been identified as a secreted toxin produced by many virulent S. suis
strains (Jacobs et al., 1995; Segers et al., 1998). It belongs to the pore-forming
cholesterol-dependent cytolysin (CDC) family, as recently confirmed by determination
of its crystal structure (Xu et al., 2010). In vitro and in vivo experiments suggest that
suilysin, though it is not essential for virulence of S. suis, most likely contributes to
pathogenesis by modification of host-pathogen interactions (Norton et al., 1999; Allen
et al., 2001; Vadeboncoeur et al., 2003; Lun et al., 2003; Benga et al., 2008). The
current concept of the biological role of suilysin is a matter of ongoing discussions. A
possible role of suilysin in host cell invasion of S. suis has been suggested by Norton
et al. (1999). Furthermore, we and others have observed that suilysin may be
involved in protection against opsonophagocytosis and cytokine release by host cells
(Benga et al., 2008; Lecours et al., 2011). However, the molecular mechanisms of
these effects are poorly understood. Considering that suilysin may in vivo often not
reach concentrations sufficient for lysis of host cell membranes, it is plausible to
speculate that the toxin expresses biological activities below cytotoxicity. Subcytolytic
effects of suilysin have not been investigated in detail, but for some other CDC such
activities have been shown to modulate host cell responses (Tsuchiya et al., 2005;
Krawczyk-Balska and Bielecki, 2005; Ratner et al., 2006; Iliev et al., 2009).
This prompted us to analyse effects of suilysin on epithelial cell-interactions of S. suis
at subcytolytic conditions. Based on preliminary studies to establish such conditions,
we then used 100 ng/ml (recombinant) suilysin in all experiments, which are clearly
below cytolytic effects (see also Figure 3-5). Accordingly, in studies on other CDC
nanomolar concentrations were also found to be far beyond that needed for cytolytic
pore-formation (Ratner et al., 2006).
An unencapsulated S. suis serotype 2 strain was used since in our previous studies
unencapsulated S. suis adhered to and invaded epithelial cells much better than
encapsulated strains (Benga et al., 2004). One might argue that an unencapsulated
strain does not represent a virulent phenotype. However, capsule expression is
highly regulated, most likely in response to environmental signals such as glucose

Results, part I Chapter 3
74
availability (Willenborg et al., 2011). Thus, by using this strain we mimic an in vivo
situation of low capsule expression which is relevant mainly during adherence and
invasion of host cells. Our comparison of this strain with its sly-negative mutant
revealed that inactivation of the sly gene resulted in a significant reduction of
bacteria-cell association, as determined by FACS analysis, and of invasion, as
determined by gentamicin-protection assay and microscopy. Furthermore, we
observed that cholesterol-depletion of epithelial cells significantly reduced uptake of
streptococci (data not shown). These results suggest that suilysin contributes to
invasion of S. suis.
Our findings in this and a previous study (Benga et al., 2004) revealed that S. suis
invasion of epithelial cells was accompanied by formation of membrane ruffles. A
major component of ruffles is actin. Thus, we hypothesized that the invasion-
mediating effects of suilysin may be due to activation of the actin host cell
cytoskeleton after binding of the (secreted) toxin to the host cell membrane.
Involvement of the actin cytoskeleton in invasion process could be shown using
latrunculin B as a specific inhibitor. It is known to complex G-actin and prevents actin
polymerisation (Coue et al., 1987), and, in our study, caused a significantly reduced
HEp-2 cell association of S. suis. Membrane ruffling and involvement of actin is also
seen in the trigger-like uptake mechanism described for other pathogenic bacteria,
e. g. Salmonella (Finlay and Cossart, 1997). Noteworthy, our results suggest that in
S. suis a toxin can promote membrane ruffling and uptake of bacteria.
It has been shown for other pathogens that activation of GTPases is involved in
bacterial invasion of host cells (Finlay, 2005). The underlying mechanisms seem to
be rather complex and differ between pathogenic bacteria. For instance,
Rho-GTPases Rac, Rho and Cdc42 are necessary for invasion of group A
streptococci (Burnham et al., 2007). In contrast, invasion of Salmonella Typhimurium
requires only Rac1 and Cdc42 (Criss and Casanova, 2003). Hence, to analyse the
molecular basis of suilysin-mediated effects on actin remodelling and streptococcal
uptake in more depth, we focussed on involvement of small Rho-GTPases. These
proteins are known to signal to the actin cytoskeleton, thereby regulating
morphological responses of the cell to environmental signals (Mackay and Hall,

Results, part I Chapter 3
75
1998). Using clostridial toxins as specific inhibitors we could show that the
recruitment of Rac-GTPase, but not Rho is crucial for invasion-mediating effects of
suilysin. This corresponds to the well-established role of Rac in polymerization of
actin fibres involved in formation of lamellipodia and membrane ruffles (Ridley et al.,
1992), whereas Rho is considered a regulator mainly of stress fibre formation.
Accordingly, our colocalisation experiments revealed that suilysin bound to the
epithelial cell membranes and was spatially associated with Rac1 and F-actin. This
supports our assumption that suilysin can bind to epithelial cells and activate Rac1,
which leads to activation of the actin cytoskeleton and formation of membrane ruffles.
To prove that activated GTP-ases were involved we treated HEp-2 cells with suilysin
and then detected activated Rac1 and RhoA by pull down analysis using specific
ligands coupled to agarose beads. Results showed that Rac1 was activated within
15 min after addition of suilysin, whereas activation of RhoA seemed not to be
affected. Involvement of Cdc42 in streptococcal internalisation is unlikely, because
activation of Cdc42 triggers mainly the formation of filopodia and microspikes
(Mackay and Hall, 1998). Accordingly, in preliminary experiments we found that
Cdc42 is not significantly activated by suilysin (data not shown).
Among other members of CDC an interaction with G-proteins has been described for
pneumolysin. Iliev et al. (2007) reported formation of stress fibres, filopodia, and
lamellipodia as a consequence of RhoA and Rac1 recruitment after treatment of
human neuroblastoma cells with pneumolysin. Strikingly, suilysin seems to activate
only Rac1, but not RhoA. However, it has to be considered that we used respiratory
epithelial cells, and that both host cells play different roles during streptococcal
infection process.
The CDC share two basic functions, which is reflected by more than 40% sequence
identity over their common domain regions 1-4 (Xu et al., 2010). First, CDC bind to
host cell membrane via specific cholesterol-dependent interaction between the
C-terminal region of domain 4 and the cell membrane. Second, after binding toxin
monomers oligomerise and change conformation in the N-terminal region, which
finally leads to formation of transmembrane pores and lysis of the target cell (Tweten,
2005). Structural-functional analyses revealed that domain 4 carries a highly

Results, part I Chapter 3
76
conserved tryptophan-rich motif consisting of 11 amino acids, which is crucial for
cytolytic activity of CDC (Billington et al., 2000). An alignment of the homologous
region of pneumolysin and suilysin is shown in Figure 3-5A. Point mutations within
this undecapeptide of pneumolysin (Feldman et al., 1990), perfringolysin O of
Clostridium perfringens (Polekhina et al., 2005) or listeriolysin O of Listeria
monocytogenes (Michel et al., 1990) lead to an attenuated ability to cause membrane
damage. It is known that such mutated toxins are still able to bind to the cell plasma
membrane, but the formation of a functional permeable transmembrane pore is
inhibited (Korchev et al., 1998). For suilysin, predictions based on its crystal structure
suggest that it shares common features with other CDC and, concerning the
undecapeptide, particularly with intermedilysin (Xu et al., 2010). However, these
predictions have yet to been proven by functional experiments.
Interactions of pneumolysin with Rho-GTPases (see above) seem to occur
independent of (macro) pore-formation (Iliev et al., 2007). This prompted us to further
dissect the relevance of pore-formation in suilysin-mediated effects on epithelial cells.
For this, we took advantage of the high similarities in primary amino acid sequence
and three-dimensional secondary structure among members of CDC family (Segers
et al., 1998b; Xu et al., 2010) as described above. Hence we produced recombinant
suilysin in which the undecapeptide was inactivated by substitution of the first
tryptophan (W461) by phenylalanine (see also alignment in Figure 3-5A). Haemolytic
and cytolytic activity of the mutated recombinant suilysin (rW461F) were almost
completely abolished in comparison with the suilysin (rSLY), indicating a loss in
transmembrane pore-forming activity. This was further confirmed in experiments
using fluorescent calcein to prove macropore-formation (data not shown).
Remarkably, the mutated suilysin had retained its ability to promote epithelial cell
invasion of S. suis and to activate Rac1 comparable to rSLY.
To our knowledge, this is the first experimental evidence for the function of this
domain in suilysin-mediated cytolysis according to the predictions based on the
crystal structure (Segers et al., 1998b; Xu et al., 2010). Furthermore, our results
indicate that a functional undecapeptide motif in domain 4 is not required for
CDC-mediated GTPase-activation and invasion of bacteria.

Results, part I Chapter 3
77
Concluding, this study revealed that suilysin can promote invasion of S. suis in
epithelial cells by Rac-dependent activation of the actin cytoskeleton at subcytolytic
conditions. Thus, we propose that subcytolytic activities of suilysin play a role in host-
pathogen interactions by modulating different host cell functions, such as
morphological responses, as shown here, or release of cytokines, as shown by
others (Lun et al., 2003; Chen et al., 2007; Vanier et al., 2009; Zheng et al., 2011;
Lecours et al., 2011). Further studies will show which domains of suilysin are
involved in this process and which significance this might have for virulence and
pathogenesis.

Results, part I Chapter 3
78
Experimental procedures If not stated otherwise all materials were purchased from Sigma (Muenchen,
Germany).
Bacterial strains and growth conditions. The unencapsulated isogenic mutant
strain 10cpsΔEF was generated by insertional mutagenesis of the virulent
Streptococcus (S.) suis serotype 2 strain 10 (both strains were kindly provided by H.
Smith, Lelystad, NL) as described previously (Smith et al., 1999). The corresponding
suilysin deficient mutant of 10cpsΔEF, strain 10cpsΔEFΔsly, was constructed as
described below. Escherichia (E.) coli strains BL21, BL21 (DE3) and DH5α were
used for molecular cloning and protein expression experiments. Streptococci were
grown on Columbia agar supplemented with 7% sheep blood (Oxoid, Wesel,
Germany) overnight under aerobic conditions at 37°C. For infection of HEp-2 cells,
streptococci were grown in Todd-Hewitt broth (THB, Difco, Detroit, USA) overnight at
37°C under aerobic conditions, adjusted to an optical density (OD600) of 0.02 in pre-
warmed media the next day and grown to late exponential growth phase (OD600 0.8).
E. coli strains were cultured on Luria Bertani (LB) agar overnight at 37°C under
aerobic conditions. When necessary, antibiotics were added to the media at the
following concentrations: spectinomycin 100 µg/ml for S. suis; erythromycin 1 µg/ml
for S. suis; ampicillin 100 µg/ml for E. coli.
DNA techniques. Routine molecular biology techniques including restriction
endonuclease digestion, DNA ligations, agarose gel electrophoresis, Southern Blot
analysis, transformation of E. coli and plasmid isolation were performed according to
standard procedures (Sambrook et al., 1989). Restriction enzymes were purchased
from New England Biolabs (Frankfurt am Main, Germany). Plasmid preparations
were performed with kits from Machery-Nagel (Dueren, Germany).
Construction of the suilysin deficient mutant strain 10cpsΔEFΔsly. The mutant
10cpsΔEFΔsly was constructed by insertion of an erythromycin cassette in the sly

Results, part I Chapter 3
79
gene of the unencapsulated strain 10cpsΔEF using the plasmid pBlue/sly/erm as
previously described (Benga et al., 2008). Mutants were controlled by PCR and
Southern Blot analysis.
Expression of recombinant wild type suilysin proteins. Recombinant His-tagged
suilysin (rSLY) and point-mutated suilysin W461F (rW461F) were expressed in BL21
pET45bslynew (Kock et al., 2009) and BL21 pET45bslynewW461F, respectively. The
plasmid pET45bslynewW461F was constructed by site-directed-mutagenesis
according to the instruction manual of QuikChange® Site-Directed Mutagenesis Kit
(Stratagene, Ja Jolla, CA) with some modifications. Purified plasmid DNA carrying
the sly gene (pET45bslynew) was amplified using oligonucleotide primers encoding
the W461 to F461 substitution (slyTrp-Phefornew [TACAGGATTAGCAT
TTGAGTGGTGGAGAAC], slyTrp-Pherevnew [GTTCTCCACCACTCAAATGCTAATC
CTGTAC]). The resulting plasmid pET45bslynewW461F was electroporated into E.
coli BL21 (DE3), purified and controlled by sequencing. Expression and purification
of rSLY and rW461F were performed as described previously (Willenborg et al.,
2011). The quality of purified proteins was controlled by separation on SDS-
polyacrylamide gels and immunoblot analysis using with anti-Penta His antibodies
(Qiagen, Hilden, Germany).
Immunoblot analysis. Recombinant proteins or supernatants of infected cells were
separated by SDS-polyacrylamide gel electrophoresis with a 4% stacking and a 10%
separating gel under denaturing conditions and transferred to a PVDF-membrane
(Serva, Heidelberg, Germany). Membrane-blocking was performed overnight with 1%
milk powder in TBS with 0.5% Tween. For detection of His-tagged recombinant
proteins membranes were blocked with 3% BSA and incubated with a monoclonal
anti-Penta His antibody (Qiagen) diluted 1:2,000 in 3% BSA. Polyclonal antiserum
raised against rSLY (Benga et al., 2008) diluted 1:1,300 in 1% milk powder was used
to detect either rSLY, rW461F or secreted suilysin in supernatants of infected cells.
Membranes were developed with horseradish peroxidase-conjugated anti-mouse or
anti-rabbit IgG antiserum diluted 1:10,000 in 1% milk powder (Amersham, Freiburg,

Results, part I Chapter 3
80
Germany) followed by using SuperSignal® West Pico Chemiluminescent Substrate
(Pierce, Rockford, USA) according to the manufactures protocol.
HEp-2 epithelial cell culture. The human laryngeal epithelial cell line HEp-2 (ATCC
CCL 23) was used as described previously (Benga et al., 2004). For adherence and
invasion assays, approximately 1.8 x 105 cells per well were seeded on 24 well tissue
culture plates. For immunofluorescence and electron microscopy 0.5 x 105 and
1.5 x 105 cells per well were seeded on 12 mm diameter glass cover slips placed in
24 well plates. For FACS analysis approximately 6 x 105 cells per well were seeded
on a 6 well plate, and for cellular cytotoxicity assay 0.2 x 105 cells per well were
seeded on a 96 well plate. The cells were grown overnight and then used for
experiments. For colocalisation experiments HEp-2 cells were transfected with
GFP-tagged wild type Rac1 (GFP-Rac1) using the Amaxa nucleofection system
(Amaxa, Cologne, Germany) applying the standard protocol as specified by the
manufacturer. Plasmids were purified with the EndoFreeTM plasmid purification kit
from Qiagen.
In some assays, HEp-2 cells were treated before infection with the following
inhibitors: 1 µg/ml latrunculin B for 30 min, 10 ng/ml toxin B of Clostridium (C.) difficile
(TcdB), 10 ng/ml toxin B of C. difficile serotype F strain 1470 (TcdB-F) and 1 µg/ml
C3 toxin of C. limosum for 3 h. Toxins were purified as described previously
(Huelsenbeck et al., 2007). C3 toxin was always reconstituted freshly by mixing the
binding domain (C2IIa) with the catalytic domain (FT) in a ratio of 1:1 as described
(Just et al., 1995). After preincubation with inhibitors cells were washed and used for
further experiments as described below, except for latrunculin B, which was left on to
the cells during the whole infection experiment because its effect is reversible up to
1 h.
Determination of haemolytic activity and cytotoxicity. Haemolytic activities of
rSLY and rW461F were determined in 96-well vee bottom microplates as described
by Takamatsu et al. (2001) with some modifications. Briefly, twofold dilutions of the
toxins were incubated with a 2% sheep erythrocyte suspension in 0.9% NaCl for 2 h

Results, part I Chapter 3
81
at 37°C. Unlysed red blood cells were allowed to pellet by centrifugation, and 100 µl
of the supernatant was transferred into a new flat bottom microplate. Subsequently,
absorption was measured at 550 nm in a microplate reader (GENios Pro, TECAN
AUSTRIA GMBH). Determination of haemolytic activity was done in three
independent experiments. One haemolytic unit (HU) was defined as the
concentration of toxin causing 50% haemolysis of a 2% sheep blood suspension
(Takamatsu et al., 2001).
Cytotoxicity was detected by LDH-release assay as described previously (Benga et
al., 2004). Briefly, HEp-2 cells were either incubated with rSLY or rW461F at
indicated concentrations. Supernatants were removed 2 h after treatment and LDH
activities were determined using the Cytotox® 96 assay kit (Promega, Mannheim,
Germany). All experiments were performed in triplicates and repeated at least three
times. Results were expressed as % LDH-release compared that of Triton X-100
lysed non-infected cells.
Antibiotic protection assay. The number of adherent and invasive streptococci was
quantified using a gentamicin protection assay as described earlier (Valentin-
Weigand et al., 1996) with the following modifications. Adherence and invasion
assays were performed with streptococci grown in THB to an OD600 of 0.8.
Subsequently, bacteria were resuspended in Dulbecco’s PBS containing 0.9 mM
CaCl2 and 0.5 mM MgCl26H2O, pH 7.3 (PBS+; Gibco-Invitrogen) and adjusted to an
OD600 0.6. Then, streptococcal suspensions were diluted 1:10 in PBS+ and used for
adherence and invasion assay. Confluent HEp-2 cell monolayers were inoculated
with 100 bacteria per cell. The number of colony forming units (CFU) inoculated per
well was determined by serial platings. After incubation infected cells for 2 h the
supernatants were collected for detection of suilysin expression by immunoblot
analysis. To determine adherence cells were washed three times with PBS
immediately after supernatant collection and lysed by 0.025% Triton X-100. Serial
dilutions of these lysates were plated in triplicates on Columbia agar for
determination of CFU. For invasion assay, after PBS washes cells were incubated for
2 h with 100 μg gentamicin and 5 μg penicillin per ml to kill extracellular bacteria.

Results, part I Chapter 3
82
Then lysates were prepared and plated as described for adherence. Results were
expressed as % adherence/invasion compared to the inoculum used for infection.
FACS analysis of bacteria-cell association. For this, streptococcal strains were
labelled with the carboxyfluorescein succinimidyl ester (CFSE) using CellTrace™
CFSE Cell Proliferation Kit (Invitrogen). Labelling was performed as described by
(Logan et al., 1998) with some modifications. Briefly, S. suis strains were grown to an
OD600 of 0.8, pelleted by centrifugation, washed once with PBS+ and resuspended in
PBS+ containing 5 µM CFSE-reagent (prepared from a CFSE stock solution as
recommended by the manufacturer). Staining procedures were carried out at 37°C
for 20 min on an end-over-end rotator. After removal of unbound fluorescent dye by
washing labelling efficiency was confirmed to be > 90%.
HEp-2 cell monolayers were infected with CFSE-labelled streptococci at a multiplicity
of infection [moi] of 125:1 bacteria per cell. For this, streptococci were centrifuged on
the epithelial cells (250 x g for 5 min) and then incubated with the cells at 37°C for
2 h. Then supernatants were collected for determination of suilysin expression by
immunoblot analysis. Epithelial cells were then detached with trypsin-EDTA, washed
with PBS and fixed with 0.375% formaldehyde in PBS. In some experiments, rSLY or
rW461F were added to epithelial cells together with streptococcal strains (100 ng/ml,
final concentration). To exclude cytotoxic effects of suilysin during the experiment
cells were stained afterwards with the DNA-intercalating dye propidium iodide
(4 µg/ml) for 2 min at room temperature to discriminate between viable and nonviable
cells. Bacteria-cell association was measured using FACScan® (Becton Dickinson,
488 nm Argon laser) and analysed using the software WinMDI (version 2.9.). For
each determination at least 10,000 events were measured. Results were expressed
as mean fluorescent intensity [mfi] values of the cell population. Uninfected cells
served as background control. Means and standard deviations were determined from
eight independent experiments. Fluorescence and electron microscopy. Determination of adherence and invasion
of streptococci by double immunofluorescence microscopy (DIF) was performed as

Results, part I Chapter 3
83
described previously (Benga et al., 2004) with the following modifications.
Semiconfluent HEp-2 cells were incubated with streptococci at a moi of 100 bacteria
per epithelial cell. ProLong® Gold antifade reagent with DAPI (Invitrogen) was used
for sealing and staining of the nucleus. Mounted samples were examined using
inverted immunofluorescence microscope Nikon Eclipse Ti-S equipped with a 40×,
0.6 S Plan Fluor objective (Nikon, Duesseldorf, Germany) driven by NIS Elements
software BR 3.2.. Intracellular streptococci were quantified by counting green
(intracellular) bacteria in at least 50 cells, and results were expressed as intracellular
bacteria per epithelial cell.
For colocalisation experiments HEp-2 cells transiently expressing GFP-Rac1 were
fixed after incubation with rSLY (100 ng/ml) for 30 min at 37°C with 3%
paraformaldehyde in modified cytoskeleton buffer (10 mM MES [pH 7.0], 150 mM
NaCl, 5 mM EGTA, 5 mM MgCl2, 5 mM sucrose) for 20 min, quenched with 10 mM
glycin in PBS and then permeabelised with 0.1% Triton X-100 in PBS. Permeabilised
cells were blocked in PBS with 5% horse serum and 1% BSA (blocking buffer) for
30 min at room temperature and subsequently incubated with polyclonal antiserum
raised against rSLY (Benga et al., 2008) diluted 1:1,200 in blocking buffer for 45 min
at room temperature. Samples were washed twice with PBS and incubated with
Cy5-conjugated goat anti-rabbit IgG (Millipore, Schwalbach/Ts, Germany) for 30 min
at room temperature. F-actin was labelled with Alexa® Fluor 568-conjugated
Phalloidin (Invitrogen) for 20 min at room temperature. Coverslips were washed three
times in PBS and then mounted using ProLong® Gold antifade reagent with DAPI
(Invitrogen). Mounted samples were examined using a LSM 510 Meta confocal
microscope equipped with a 63×, 1.2NA Plan-NEOFLUAR oil immersion objective
(Zeiss, Jena, Germany) driven by LSM software v3.2. For 3D z-stack acquisition the
pinholes were set to one Airy unit and confocal planes were acquired every 340 nm.
Images were deconvolved and maximum intensity projection were rendered using
Huygens® Essential (Hilversum, The Netherlands).
Field emission scanning electron microscopy (FESEM) was performed as described
previously (Benga et al., 2004) using HEp-2 cells infected as described for DIF.

Results, part I Chapter 3
84
Pull down experiments. Experiments were done as described previously with some
modifications (Reid et al., 1996). HEp-2 cells were incubated with rSLY or rW461F
(100 ng/ml) for indicated time periods. Then cells were rinsed twice with ice cold PBS
to stop incubation. Lysis of cells was done by addition of 1 ml of ice cold lysis buffer
(50 mM NaCl, 20 mM Tris-HCl [pH 7.4], 3 mM MgCl2, 1% Nonidet-P 40,
0.25% Triton X-100, 5 mM dithiothreitol, 100 µM PMSF). Cells were scraped off, and
the lysates were centrifuged at 14,000 rpm. The supernatant was split and used for
pull down experiments of activated RhoA and Rac1 in parallel. For this, 20 µl of
beads slurry bearing approximately 20 µg protein of the GST-fusion protein of either
the Rho-binding domain C21 or the PAK-GBD, respectively, were added to 500 µl
sample and incubated on a rotator at 4°C for 30 min. Beads were collected by
centrifugation at 10,000 rpm, washed twice with lysis buffer and subjected to
immunoblot analysis using monoclonal antibodies against Rac-1 (cloe 102; BD
Pharmingen, Heidelberg, Germany) and against RhoA (clone 26C4; Santa Cruz,
Heidelberg, Germany), respectively. Amounts of (activated and total) GTPases were
quantified by densitometric measurement of three independent immunoblot analyses.
Statistical analyses. If not stated otherwise, experiments were performed at least
three times and results were expressed as means and standard deviations. Data
were analyzed by t-test. Pull down analysis was carried out using an unpaired t-test
with no assumption of Gaussian distribution (Mann-Whitney). A P-value <0.05 was
considered significant.
Acknowledgements We thank Hilde E. Smith (Lelystad, NL) for providing S. suis strain 10cpsΔEF. This
work was financially supported by the Deutsche Forschungsgemeinschaft (DFG),
Bonn, Germany (SFB587, GRK745).

Results, part I Chapter 3
85
References Allen, A. G., Bolitho, S., Lindsay, H., Khan, S., Bryant, C., Norton, P., Ward, P., Leigh, J., Morgan, J., Riches, H., Eastty, S., and Maskell, D. (2001) Generation and characterization of a defined mutant of Streptococcus suis lacking suilysin. Infect Immun 69: 2732-2735.
Arends, J. P., Hartwig, N., Rudolphy, M., and Zanen, H. C. (1984) Carrier Rate of Streptococcus suis Capsular Type-2 in Palatine Tonsils of Slaughtered Pigs. J Clin Microbiol 20: 945-947.
Arends, J. P. and Zanen, H. C. (1988) Meningitis Caused by Streptococcus suis in Humans. Rev Infect Dis 10: 131-137.
Benga, L., Fulde, M., Neis, C., Goethe, R., and Valentin-Weigand, P. (2008) Polysaccharide capsule and suilysin contribute to extracellular survival of Streptococcus suis co-cultivated with primary porcine phagocytes. Vet Microbiol 132: 211-219.
Benga, L., Goethe, R., Rohde, M., and Valentin-Weigand, P. (2004) Non-encapsulated strains reveal novel insights in invasion and survival of Streptococcus suis in epithelial cells. Cell Microbiol 6: 867-881.
Billington, S. J., Jost, B. H., and Songer, J. G. (2000) Thiol-activated cytolysins: structure, function and role in pathogenesis. FEMS Microbiol Lett 182: 197-205.
Burnham, C. A., Shokoples, S. E., and Tyrrell, G. J. (2007) Rac1, RhoA, and Cdc42 participate in HeLa cell invasion by group B streptococcus. FEMS Microbiol Lett 272: 8-14.
Charland, N., Nizet, V., Rubens, C. E., Kim, K. S., Lacouture, S., and Gottschalk, M. (2000) Streptococcus suis serotype 2 interactions with human brain microvascular endothelial cells. Infect Immun 68: 637-643.
Chen, C., Tang, J., Dong, W., Wang, C., Feng, Y., Wang, J., Zheng, F., Pan, X., Liu, D., Li, M., Song, Y., Zhu, X., Sun, H., Feng, T., Guo, Z., Ju, A., Ge, J., Dong, Y., Sun, W., Jiang, Y., Wang, J., Yan, J., Yang, H., Wang, X., Gao, G. F., Yang, R., Wang, J., and Yu, J. (2007) A glimpse of streptococcal toxic shock syndrome from comparative genomics of S. suis serotype 2 Chinese isolates. PLoS ONE 2: e315.
Coue, M., Brenner, S. L., Spector, I., and Korn, E. D. (1987) Inhibition of actin polymerization by latrunculin A. FEBS Lett 213: 316-318.
Criss, A. K. and Casanova, J. E. (2003) Coordinate regulation of Salmonella enterica serovar Typhimurium invasion of epithelial cells by the Arp2/3 complex and Rho GTPases. Infect Immun 71: 2885-2891.
Feldman, C., Mitchell, T. J., Andrew, P. W., Boulnois, G. J., Read, R. C., Todd, H. C., Cole, P. J., and Wilson, R. (1990) The effect of Streptococcus pneumoniae pneumolysin on human respiratory epithelium in vitro. Microb Pathog 9: 275-284.
Finlay, B. B. (2005) Bacterial virulence strategies that utilize Rho GTPases. Curr Top Microbiol Immunol 291: 1-10.
Finlay, B. B. and Cossart, P. (1997) Exploitation of mammalian host cell functions by bacterial pathogens. Science 276: 718-725
Gottschalk, M., Segura, M., and Xu, J. (2007) Streptococcus suis infections in humans: the Chinese experience and the situation in North America. Anim Health Res Rev 8: 29-45.

Results, part I Chapter 3
86
Gottschalk, M., Xu, J., Calzas, C., and Segura, M. (2010) Streptococcus suis: a new emerging or an old neglected zoonotic pathogen? Fut Microbiol 5: 371-391. Greenwood, J., Steinman, L., and Zamvil, S. S. (2006) Statin therapy and autoimmune disease: from protein prenylation to immunomodulation. Nat Rev Immunol 6: 358-370.
Hall, A. (1998) Rho GTPases and the actin cytoskeleton. Science 279: 509-514.
Huelsenbeck, J., Dreger, S., Gerhard, R., Barth, H., Just, I., Genth, H. (2007) Difference in the cytotoxic effects of toxin B from Clostridium difficile strain VPI 10463 and toxin B from variant Clostridium difficile strain 1470. Infect Immun 75(2): 801-9.
Iliev, A. I., Djannatian, J. R., Nau, R., Mitchell, T. J., and Wouters, F. S. (2007) Cholesterol-dependent actin remodeling via RhoA and Rac1 activation by the Streptococcus pneumoniae toxin pneumolysin. Proc Natl Acad Sci U S A 104: 2897-2902.
Iliev, A. I., Djannatian, J. R., Opazo, F., Gerber, J., Nau, R., Mitchell, T. J., and Wouters, F. S. (2009) Rapid microtubule bundling and stabilization by the Streptococcus pneumoniae neurotoxin pneumolysin in a cholesterol-dependent, non-lytic and Src-kinase dependent manner inhibits intracellular trafficking. Mol Microbiol 71: 461-477.
Jacobs, A. A. C., Loeffen, P. L. W., vandenBerg, A. J. G., and Storm, P. K. (1994) Identification, Purification, and Characterization of A Thiol-Activated Hemolysin (Suilysin) of Streptococcus suis. Infect Immun 62: 1742-1748.
Jacobs, A. A. C., vandenBerg, A. J. G., and Loeffen, P. L. W. (1996) Protection of experimentally infected pigs by suilysin, the thiol-activated haemolysin of Streptococcus suis. Veterinary Record 139: 225-228.
Just, I., Selzer, J., Wilm, M., Eichel-Streiber, C., Mann, M., and Aktories, K. (1995) Glucosylation of Rho proteins by Clostridium difficile toxin B. Nature 375: 500-503.
King, S. J., Heath, P. J., Luque, I., Tarradas, C., Dowson, C. G., and Whatmore, A. M. (2001) Distribution and genetic diversity of suilysin in Streptococcus suis isolated from different diseases of pigs and characterization of the genetic basis of suilysin absence. Infect Immun 69: 7572-7582.
Kock, C., Beineke, A., Seitz, M., Ganter, M., Waldmann, K. H., Valentin-Weigand, P., and Baums, C. G. (2009) Intranasal immunization with a live Streptococcus suis isogenic ofs mutant elicited suilysin-neutralization titers but failed to induce opsonizing antibodies and protection. Vet Immuno Immunopathol 132: 135-145.
Korchev, Y. E., Bashford, C. L., Pederzolli, C., Pasternak, C. A., Morgan, P. J., Andrew, P. W., and Mitchell, T. J. (1998) A conserved tryptophan in pneumolysin is a determinant of the characteristics of channels formed by pneumolysin in cells and planar lipid bilayers. Biochem J 329 ( Pt 3): 571-577.
Krawczyk-Balska, A. and Bielecki, J. (2005) Listeria monocytogenes listeriolysin O and phosphatidylinositol-specific phospholipase C affect adherence to epithelial cells. Can J Microbiol 51: 745-751.
Lalonde, M., Segura, M., Lacouture, S., and Gottschalk, M. (2000) Interactions between Streptococcus suis serotype 2 and different epithelial cell lines. Microbiology 146 ( Pt 8): 1913- 1921.

Results, part I Chapter 3
87
Lecours, M. P., Gottschalk, M., Houde, M., Lemire, P., Fittipaldi, N., and Segura, M. (2011) Critical Role for Streptococcus suis Cell Wall Modifications and Suilysin in Resistance to Complement-Dependent Killing by Dendritic Cells. J Infect Dis 204: 919-929.
Logan, R. P., Robins, A., Turner, G. A., Cockayne, A., Borriello, S. P., and Hawkey, C. J. (1998) A novel flow cytometric assay for quantitating adherence of Helicobacter pylori to gastric epithelial cells. J Immunol Methods 213: 19-30.
Lun, S. C., Perez-Casal, J., Connor, W., and Willson, P. J. (2003) Role of suilysin in pathogenesis of Streptococcus suis capsular serotype 2. Microb Pathogen 34: 27-37.
Mackay, D. J. and Hall, A. (1998) Rho GTPases. J Biol Chem 273: 20685-20688.
Michel, E., Reich, K. A., Favier, R., Berche, P., and Cossart, P. (1990) Attenuated mutants of the intracellular bacterium Listeria monocytogenes obtained by single amino acid substitutions in listeriolysin O. Mol Microbiol 4: 2167-2178.
Norton, P. M., Rolph, C., Ward, P. N., Bentley, R. W., and Leigh, J. A. (1999) Epithelial invasion and cell lysis by virulent strains of Streptococcus suis is enhanced by the presence of suilysin. FEMS Immunol Med Microbiol 26: 25-35.
O'Sullivan, T., Friendship, R., Blackwell, T., Pearl, D., McEwen, B., Carman, S., Slavic, D., and Dewey, C. (2011) Microbiological identification and analysis of swine tonsils collected from carcasses at slaughter. Can J Vet Res 75: 106-111.
Polekhina, G., Giddings, K. S., Tweten, R. K., and Parker, M. W. (2005) Insights into the action of the superfamily of cholesterol-dependent cytolysins from studies of intermedilysin. Proc Natl Acad Sci U S A 102: 600-605.
Ratner, A. J., Hippe, K. R., Aguilar, J. L., Bender, M. H., Nelson, A. L., and Weiser, J. N. (2006) Epithelial cells are sensitive detectors of bacterial pore-forming toxins. J Biol Chem 281: 12994-12998.
Reid, T., Furuyashiki, T., Ishizaki, T., Watanabe, G., Watanabe, N., Fujisawa, K., Morii, N., Madaule, P., and Narumiya, S. (1996) Rhotekin, a new putative target for Rho bearing homology to a serine/threonine kinase, PKN, and rhophilin in the rho-binding domain. J Biol Chem 271: 13556-13560.
Ridley, A. J., Paterson, H. F., Johnston, C. L., Diekmann, D., and Hall, A. (1992) The small GTP- binding protein rac regulates growth factor-induced membrane ruffling. Cell 70: 401-410.
Rubins, J. B., Paddock, A. H., Charboneau, D., Berry, A. M., Paton, J. C., and Janoff, E. N. (1998) Pneumolysin in pneumococcal adherence and colonization. Microb Pathog 25: 337-342.
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N Y.
Segers, R. P., Kenter, T., de Haan, L. A., and Jacobs, A. A. (1998) Characterisation of the gene encoding suilysin from Streptococcus suis and expression in field strains. FEMS Microbiol Lett 167: 255-261.
Segura, M. and Gottschalk, M. (2002) Streptococcus suis interactions with the murine macrophage cell line J774: adhesion and cytotoxicity. Infect Immun 70: 4312-4322.

Results, part I Chapter 3
88
Smith, H. E., Damman, M., van der Velde, J., Wagenaar, F., Wisselink, H. J., Stockhofe-Zurwieden, N., and Smits, M. A. (1999) Identification and characterization of the cps locus of Streptococcus suis serotype 2: the capsule protects against phagocytosis and is an important virulence factor. Infect Immun 67: 1750-1756.
Sukeno, A., Nagamune, H., Whiley, R. A., Jafar, S. I., Aduse-Opoku, J., Ohkura, K., Maeda, T., Hirota, K., Miyake, Y., and Kourai, H. (2005) Intermedilysin is essential for the invasion of hepatoma HepG2 cells by Streptococcus intermedius. Microbiol Immunol 49: 681-694.
Takamatsu, D., Osaki, M., and Sekizaki, T. (2001) Thermosensitive suicide vectors for gene replacement in Streptococcus suis. Plasmid 46: 140-148.
Tang, J., Wang, C., Feng, Y., Yang, W., Song, H., Chen, Z., Yu, H., Pan, X., Zhou, X., Wang, H., Wu, B., Wang, H., Zhao, H., Lin, Y., Yue, J., Wu, Z., He, X., Gao, F., Khan, A. H., Wang, J., Zhao, G. P., Wang, Y., Wang, X., Chen, Z., and Gao, G. F. (2006) Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med 3: e151.
Tenenbaum, T., Adam, R., Eggelnpohler, I., Matalon, D., Seibt, A., GE, K. Novotny, Galla, H. J., and Schroten, H. (2005) Strain-dependent disruption of blood-cerebrospinal fluid barrier by Streptoccocus suis in vitro. FEMS Immunol Med Microbiol 44: 25-34.
Tenenbaum, T., Essmann, F., Adam, R., Seibt, A., Janicke, R. U., Novotny, G. E., Galla, H. J., and Schroten, H. (2006) Cell death, caspase activation, and HMGB1 release of porcine choroid plexus epithelial cells during Streptococcus suis infection in vitro. Brain Res 1100: 1-12.
Trottier, S., Higgins, R., Brochu, G., and Gottschalk, M. (1991) A Case of Human Endocarditis Due to Streptococcus Suis in North-America. Reviews of Infectious Diseases 13: 1251-1252.
Tsuchiya, K., Kawamura, I., Takahashi, A., Nomura, T., Kohda, C., and Mitsuyama, M. (2005) Listeriolysin O-induced membrane permeation mediates persistent interleukin-6 production in Caco-2 cells during Listeria monocytogenes infection in vitro. Infect Immun 73: 3869-3877.
Tweten, R. K. (2005) Cholesterol-dependent cytolysins, a family of versatile pore-forming toxins. Infection and Immunity 73: 6199-6209.
Vadeboncoeur, N., Segura, M., Al Numani, D., Vanier, G., and Gottschalk, M. (2003) Pro-inflammatory cytokine and chemokine release by human brain microvascular endothelial cells stimulated by Streptococcus suis serotype 2. FEMS Immunol Med Microbiol 35: 49-58.
Valentin-Weigand, P., Benkel, P., Rohde, M., and Chhatwal, G. S. (1996) Entry and intracellular survival of group B streptococci in J774 macrophages. Infect Immun 64: 2467-2473.
Vanier, G., Fittipaldi, N., Slater, J. D., Dominguez-Punaro, Mde L., Rycroft, A. N., Segura, M., Maskell, D. J., and Gottschalk, M. (2009) New putative virulence factors of Streptococcus suis involved in invasion of porcine brain microvascular endothelial cells. Microb Pathog 46: 13-20.
Vanier, G., Segura, M., Friedl, P., Lacouture, S., and Gottschalk, M. (2004) Invasion of porcine brain microvascular endothelial cells by Streptococcus suis serotype 2. Infection and Immunity 72: 1441-1449.
Willenborg, J., Fulde, M., de Greeff, A., Rohde, M., Smith, H. E., Valentin-Weigand, P., and Goethe, R. (2011) Role of glucose and CcpA in capsule expression and virulence of Streptococcus suis. Microbiology 157: 1823-1833.

Results, part I Chapter 3
89
Wisselink, H. J., Smith, H. E., Stockhofe-Zurwieden, N., Peperkamp, K., and Vecht, U. (2000) Distribution of capsular types and production of muramidase-released protein (MRP) and extracellular factor (EF) of Streptococcus suis strains isolated from diseased pigs in seven European countries. Veterinary Microbiology 74: 237-248.
Xu, L., Huang, B., Du, H., Zhang, X. C., Xu, J., Li, X., and Rao, Z. (2010) Crystal structure of cytotoxin protein suilysin from Streptococcus suis. Protein Cell 1: 96-105.
Zheng, H., Punaro, M. C., Segura, M., Lachance, C., Rivest, S., Xu, J., Houde, M., and Gottschalk, M. (2011) Toll-like receptor 2 is partially involved in the activation of murine astrocytes by Streptococcus suis, an important zoonotic agent of meningitis. J Neuroimmunol 234: 71-83.


Chapter 4
Results, part II:
Identification of a RGD-motif in suilysin possibly involved in host-cell binding,
Rac1-activation and macropore-formation
M. Seitz, R. Gerhard, R. Goethe and P. Valentin-Weigand
Running title: RGD-motif in suilysin
(Manuscript in preparation)


Results, part II Chapter 4
93
Abstract Suilysin is a cholesterol-dependent pore-forming cytolysin secreted by Streptococcus
suis, an important causative agent of meningitis in swine and human. The role of
suilysin in host cell interaction is still unclear. Here, we showed that suilysin contains
structural elements involved in specific functions of the toxin. Using oligonucleotide
directed mutagenesis we produced recombinant suilysin in which either the highly
conserved tryptophan-rich undecapeptide of the fourth domain or the RGD-motif of
domain one was inactivated. Measurement of haemolytic and cytotoxic activity
revealed that both domains are required for full lytic function of suilysin. In contrast to
the tryptophan-mutant, the RGD-mutant showed also a deficiency in membrane
binding to HEp-2 cells. To further resolve if subsequent cellular signalling in host cells
is affected by amino acid substitution, we used a pull down based GLISA to study the
activation of small Rho-GTPases. As anticipated from our previous studies,
unmodified suilysin and the tryptophan-mutant activated Rac1. In contrast, mutation
of the RGD-motif abolished ability to induce G-proteins. Furthermore, studies with the
fluorescent dye calcein revealed that both mutated derivates of suilysin had lost their
ability to form lytic macropores in host cells. Thus, we suggest that the RGD-motif is
required for membrane binding of suilysin as well as for an outside-in signalling
mediated by the G-protein kinase Rac1 and the induction of lytic macropores in
HEp-2 cells.

Results, part II Chapter 4
94
Introduction Streptococcus (S.) suis is one of the most important swine pathogens worldwide
causing high economical losses in pig husbandry due to invasive diseases
associated with meningitis, arthritis, septicaemia, and bronchopneumonia.
Furthermore, S. suis is a zoonotic bacterium causing meningitis and the life-
threatening streptococcal toxic shock like syndrome in humans (Gottschalk et al.,
2007; Gottschalk et al., 2010; Tang et al., 2006; Trottier et al., 1991).
Suilysin, the secreted haemolysin of S. suis, belongs to the family of cholesterol-
dependent pore-forming cytolysins (CDC) (Jacobs et al., 1994). This large family of
toxins generate complex channels in target cells, causing membrane damage of host
cells and lysis of erythrocytes (Lalonde et al., 2000; Norton et al., 1999; Tenenbaum
et al., 2006; Vanier et al., 2004). The primary and secondary structure of CDC is
highly conserved. They share the same monomeric structure, comprised of four
domains (Rossjohn et al., 1997; Xu et al., 2010). A mechanism of pore-formation has
been proposed by Tweten (2005). Several monomers bind vertically to the
cholesterol-containing membrane of target cells via the fourth domain, followed by an
oligomerisation step resulting in formation of the prepore complex. The second
domain builds a 'hinge-like' region allowing the insertion of the oligomer into the
membrane, whereas the third domain mainly serves as a transmembrane domain,
representing the internal structure of the permeable pore (Czajkowsky et al., 2004;
Ramachandran et al., 2002; Shepard et al., 2000). The fourth domain contains the
tryptophan-rich undecapeptide, which is highly conserved and known to be
responsible for formation of a functional pore (Feldman et al., 1990; Korchev et al.,
1998; Michel et al., 1990; Polekhina et al., 2005). We recently confirmed the
predicted 'pore-forming' function of the undecapeptide for suilysin using a point-
mutated derivate of suilysin W461F (Seitz et al., submitted; cf. chapter 3).
Furthermore, we showed that suilysin promotes invasion of S. suis independent of
the formation of a functional pore. However, the specific molecular mechanism
underlying suilysin-induced activation of host cells is still unknown.

Results, part II Chapter 4
95
The objective of this study was to further characterize possible functional domains of
suilysin to understand the molecular mechanism underlying suilysin-induced
activation of host cells. We identified a RGD (Arg-Gly-Asp)-motif within the sequence
of the first domain of the molecule. This is a special feature of suilysin, since none of
the other well-characterized CDC posses this sequence. Bacterial proteins containing
a RGD-sequence are known to mediate adherence and entry of pathogens into host
cells (Scibelli et al., 2007), most likely by binding host cell surface proteins
recognizing RGD-sequences, known as integrins. Using target mutagenesis we
constructed recombinant suilysin in which the putative integrin-binding side
(RGD-motif) is inactivated by substitution. This abolished cytolytic and haemolytic
function of suilysin. Detailed analysis revealed that the RGD-motif is most likely
required for membrane binding of suilysin, subsequent activation of Rac1 as well as
for the formation of lytic macropores.

Results, part II Chapter 4
96
Results and Discussion
Inactivation of the RGD-motif leads to almost complete loss of haemolytic and cytotoxic activity of suilysin. The purpose of this study was to characterise a
second putative functional region, a RGD-motif identified within the first domain of
suilysin. Hence, we introduced a point-mutation in the sly gene by site-directed
mutagenesis, resulting in amino substitution of RGD by SVD.
Full length suilysin lacking the signal sequence (rSLY) and the point-mutated
derivates (rW461F and rSVD) were overexpressed in E. coli and purified. Silver
straining of respective recombinant proteins showed no difference in size and a high
purity grade, as indicated by a single clear band at approximately 58 kDa (Figure
4-1). To exclude frameshifts, rSLY was sequenced in total. Moreover, both point-
mutations (rW461F and rSVD) were confirmed by sequencing of the respective
regions within the molecule. Alignment of amino acid sequences of suilysin (NCBI
accession # 253753945), sequenced rSLY, in silico analyzed rW461F and rSVD and
four other members of the CDC-family, including pneumolysin (PLY), perfringolysin O
(PFO), intermedilysin (ILY), and listeriolysin O (LLO), is shown in Figure 4-1.
Sequence alignment confirmed the high similarity of the undecapeptide of the fourth
domain, termed tryptophan-rich motif (coloured in light green; blue similarity bar
below sequences indicates 100% homology). The RGD-motif within domain one
(coloured in light red) is shown to be only present within the suilysin molecule. The
substitution of the undecapeptide at position W461 (rW461F, highlighted in white)
and the RGD-sequence (rSVD, highlighted in white), respectively, was confirmed.
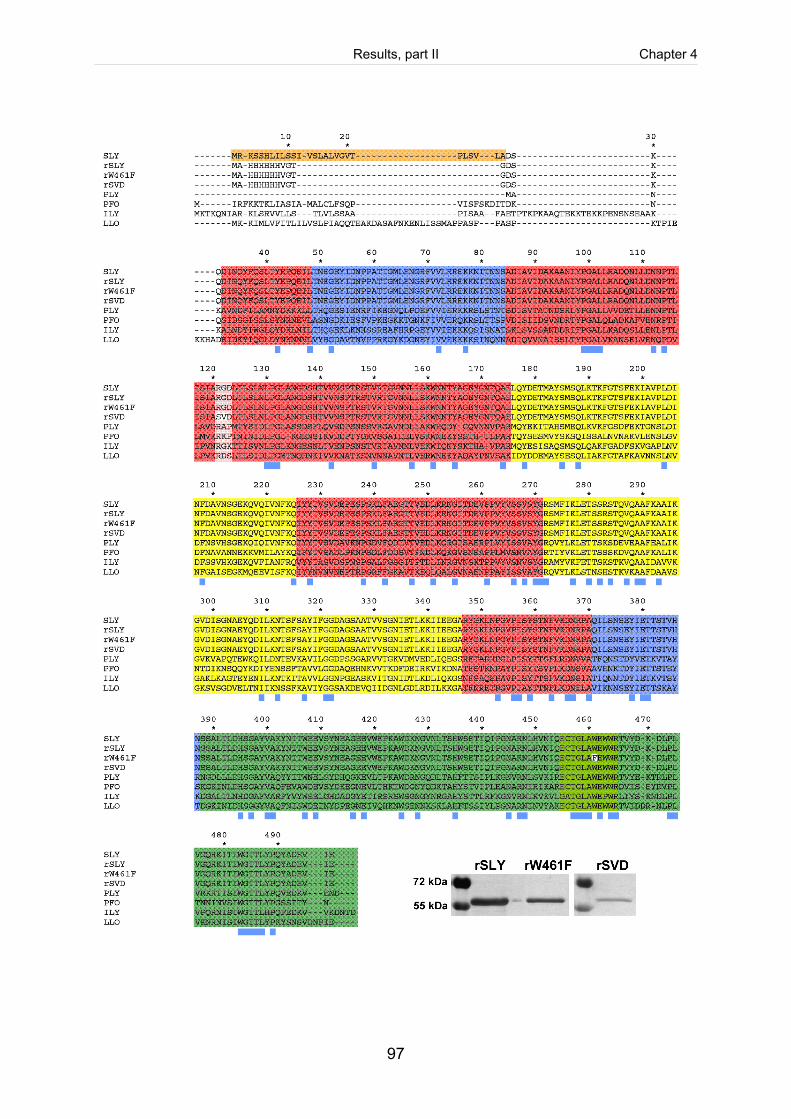
Results, part II Chapter 4
97

Results, part II Chapter 4
98
Figure 4-1: Alignment of amino acid sequences of different CDC. Suilysin (NCBI accession # 253753945), sequenced rSLY, in silico analyzed rW461F and rSVD and four other members of the CDC family, including pneumolysin (PLY, # 294652455), perfringolysin O (PFO, # 144884), intermedilysin (ILY, # 6729344) and listeriolysin O (LLO, # 44112) are shown. Subunits of the suilysin molecule are highlighted, including domain 1 (red; RGD-motif in light red), domain 2 (blue), domain 3 (yellow) and domain 4 (green; tryptophan-rich undecapetide in light green). The same colour code was used for the crystal structure of the suilysin molecule modified from (Xu et al., 2010a) (cf. chapter 1). Substitutions of the RGD-motif and W461 of the undecapeptide are marked in white. The signal-sequence of SLY is marked in orange. Similarity bars (light blue) below sequences indicate a similarity of 100% of all analyzed CDC. Alignment was performed with the program Clone Manager 9. The inlay shows a silver staining of recombinant suilysin rSLY, rW461F, and rSVD (~58kDa) separated by SDS-polyacrylamid gel electrophoresis.
We have previously shown that substitution of the tryptophan-rich undecapeptide
resulted in loss of lytic function (Seitz et al., submitted; cf. Chapter 3). Therefore,
haemolytic and cytotoxic activity tests of the mutated suilysin rSVD were carried out
in comparison to rSLY and rW461F. Determination of haemolytic capacity showed
that the point-mutated toxin rSVD almost completely lost its capacity to lyse sheep
erythrocytes. Compared to rSLY, of which 256 (28) ng/ml were sufficient for 50%
haemolysis, rSVD had to applied at 2048 (211) ng/ml and rW461F at 4096 (212) ng/ml
to cause 50% haemolysis (Figure 4-2A). Calculation of haemolytic units revealed
0.53 x 105 HU/mg for rSLY as compared to 0.3 x 104 HU/mg for rSVD and 0.25 x 104
HU/mg for rW461F. In addition, membrane damage of HEp-2 cells by suilysin and its
mutated derivates was determined using a LDH release test (Figure 4-2B). Testing
the specific cytotoxic activity of rSVD revealed a loss of cytotoxic function (2.1% of
the rSLY), similar to the rW461F (8.1% of rSLY).
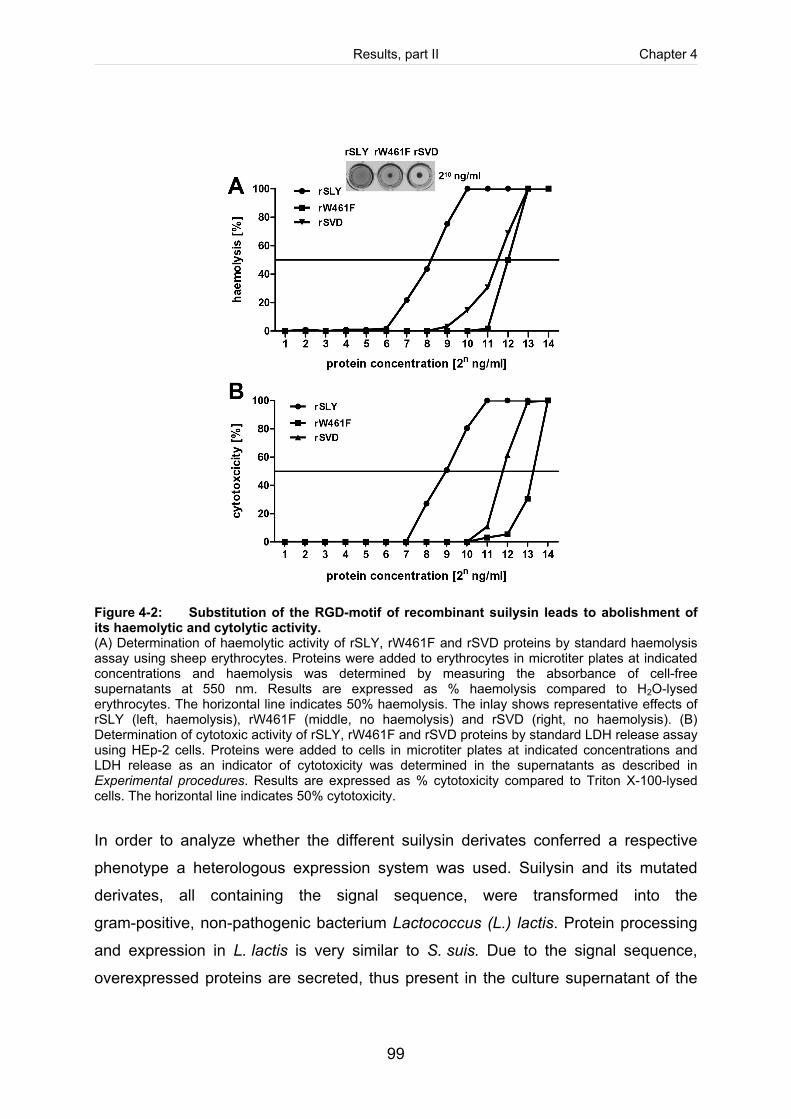
Results, part II Chapter 4
99
Figure 4-2: Substitution of the RGD-motif of recombinant suilysin leads to abolishment of its haemolytic and cytolytic activity. (A) Determination of haemolytic activity of rSLY, rW461F and rSVD proteins by standard haemolysis assay using sheep erythrocytes. Proteins were added to erythrocytes in microtiter plates at indicated concentrations and haemolysis was determined by measuring the absorbance of cell-free supernatants at 550 nm. Results are expressed as % haemolysis compared to H2O-lysed erythrocytes. The horizontal line indicates 50% haemolysis. The inlay shows representative effects of rSLY (left, haemolysis), rW461F (middle, no haemolysis) and rSVD (right, no haemolysis). (B) Determination of cytotoxic activity of rSLY, rW461F and rSVD proteins by standard LDH release assay using HEp-2 cells. Proteins were added to cells in microtiter plates at indicated concentrations and LDH release as an indicator of cytotoxicity was determined in the supernatants as described in Experimental procedures. Results are expressed as % cytotoxicity compared to Triton X-100-lysed cells. The horizontal line indicates 50% cytotoxicity.
In order to analyze whether the different suilysin derivates conferred a respective
phenotype a heterologous expression system was used. Suilysin and its mutated
derivates, all containing the signal sequence, were transformed into the
gram-positive, non-pathogenic bacterium Lactococcus (L.) lactis. Protein processing
and expression in L. lactis is very similar to S. suis. Due to the signal sequence,
overexpressed proteins are secreted, thus present in the culture supernatant of the
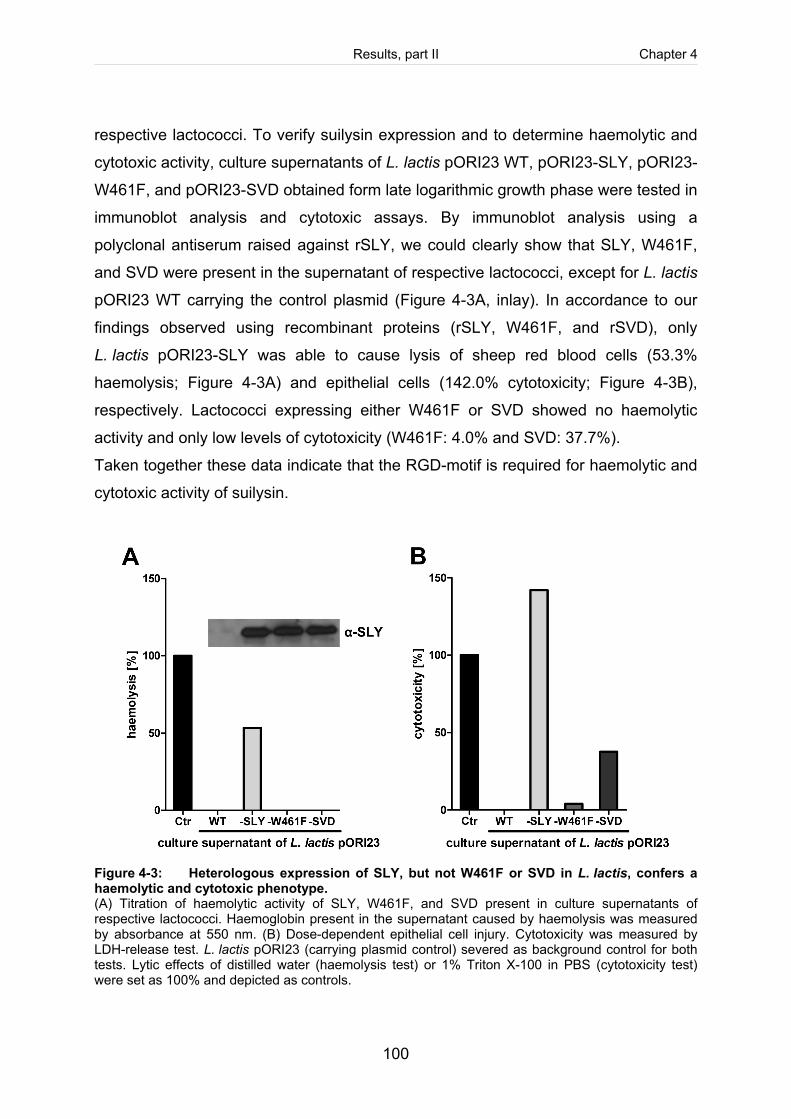
Results, part II Chapter 4
100
respective lactococci. To verify suilysin expression and to determine haemolytic and
cytotoxic activity, culture supernatants of L. lactis pORI23 WT, pORI23-SLY, pORI23-
W461F, and pORI23-SVD obtained form late logarithmic growth phase were tested in
immunoblot analysis and cytotoxic assays. By immunoblot analysis using a
polyclonal antiserum raised against rSLY, we could clearly show that SLY, W461F,
and SVD were present in the supernatant of respective lactococci, except for L. lactis
pORI23 WT carrying the control plasmid (Figure 4-3A, inlay). In accordance to our
findings observed using recombinant proteins (rSLY, W461F, and rSVD), only
L. lactis pORI23-SLY was able to cause lysis of sheep red blood cells (53.3%
haemolysis; Figure 4-3A) and epithelial cells (142.0% cytotoxicity; Figure 4-3B),
respectively. Lactococci expressing either W461F or SVD showed no haemolytic
activity and only low levels of cytotoxicity (W461F: 4.0% and SVD: 37.7%).
Taken together these data indicate that the RGD-motif is required for haemolytic and
cytotoxic activity of suilysin.
Figure 4-3: Heterologous expression of SLY, but not W461F or SVD in L. lactis, confers a haemolytic and cytotoxic phenotype. (A) Titration of haemolytic activity of SLY, W461F, and SVD present in culture supernatants of respective lactococci. Haemoglobin present in the supernatant caused by haemolysis was measured by absorbance at 550 nm. (B) Dose-dependent epithelial cell injury. Cytotoxicity was measured by LDH-release test. L. lactis pORI23 (carrying plasmid control) severed as background control for both tests. Lytic effects of distilled water (haemolysis test) or 1% Triton X-100 in PBS (cytotoxicity test) were set as 100% and depicted as controls.

Results, part II Chapter 4
101
The RGD-motif is required for membrane binding, Rac1-activation and macropore-formation in HEp-2 cells. Next we analysed in more detail, in which
step the RGD-motif might be involved. In general, RGD-containing proteins can act
as connecting molecules between pathogens and host cells to allow adherence and
uptake of pathogens or in order to induce cell signalling pathways (Scibelli et al.,
2005). Therefore, we first analysed the initial membrane binding capacity of suilysin
and its mutated derivates using immunofluorescent staining. HEp-2 cells were
treated with rSLY, rW461F or rSVD at subcytolytic (100 ng/ml; Figure 4-4B-D) or lytic
(400 ng/ml; Figure 4-4E-G) concentration. Cell-bound suilysin was detected using a
polyclonal antiserum raised against rSLY. Unmodified rSLY was able to bind to the
cell surface at subcytolytic and lytic concentration (green spots in Figure 4-4B and E).
The size of individual spots (most likely due to different numbers of molecules or
oligomerisation stages) increased at higher concentration. Likewise, rW461F bound
to cell membranes at lytic concentration (green spots in Figure 4-4G). This
phenotype was described before for homologous tryptophan-mutants of the highly
related pneumolysin as well as for intermedilysin (Polekhina et al., 2005). It has been
shown, that loss of lytic function did not include absence of oligomerisation and
formation of the prepore complex (Boulnois et al., 1991; Korchev et al., 1998;
Polekhina et al., 2005). In contrast, we showed that substitution of the RGD-motif
affected membrane binding capacity drastically, so that rSVD was unable to bind to
HEp-2 cells (Figure 4-4C and F).
Possible interaction partners of RGD-containing proteins are the heterodimeric
transmembrane integrins. The integrin family is subdivided into groups, whereas one
subgroup is specified on recognizing RGD-containing proteins (RGD-receptors), such
as α5β1 integrins (Scibelli et al., 2005; Scibelli et al., 2007). Two mechanisms of
integrin-bacteria interactions are known (Scibelli et al., 2007). An indirect interaction
is mediated by extracellular matrix components (ECM) containing a RGD-sequence,
such as fibronectin or vitronectin. These 'bridging molecules' connect bacterial
ECM-binding proteins with integrins (Cue et al., 2000; Jonsson et al., 1991). An
alternative way is the direct linking of microbial RGD-exhibiting proteins to integrins
(Hamzaoui et al., 2004; Tran et al., 1999). We hypothesized that the latter is

Results, part II Chapter 4
102
plausible for suilysin-integrin interaction. Therefore, we analyzed by flow cytometry, if
α5β1 integrins were expressed on the cellular surface of unstimulated HEp-2 cells.
The results clearly showed that α5β1 integrins were present on HEp-2 cells (Figure
4-4H), thus available as putative interaction partners for suilysin. Further studies will
have to proof whether they in fact function as receptors.
In principle, CDC bind preferentially to cholesterol-rich microdomains (lipid rafts)
(Gekara and Weiss, 2004; Shimada et al., 2002; Waheed et al., 2001), but
membrane components required for CDC binding have not been characterized in
detail. The impact of cholesterol on membrane binding of CDC and CDC function is
still controversially discussed (Hotze and Tweten, 2011). First assumed as the
cellular receptor, recognized by the fourth domain of the CDC monomer (Sekino-
Suzuki et al., 1996), a recent study pointed out that cholesterol plays a role
downstream of membrane binding by triggering the conversion of the prepore
complex to a functional transmembrane pore (Giddings et al., 2003). Nevertheless,
lipid rafts are known to function as a 'platform' for bacteria-host cell interaction,
enhancing bacterial adherence and invasion (Lafont and van der Goot, 2005). It is
proposed that this 'triggering effect' is mediated by clustering of cell surface receptors
within cholesterol enriched microdomains. Receptors associated with lipid rafts are
among other integrins. We have previously demonstrated that cholesterol is essential
for cytolytic activity. Future studies will have to show how cholesterol contributes to
cell-activation by suilysin.
Integrins can also transmit extracellular signals into the cell. The small family of Rho-
GTPases, namely Rac, Rho and Cdc42, play a key role in integrin-mediated outside-
in signalling. Integrin-ligand interaction triggers either a direct activation of GTPases
or regulates GTPase effector proteins. Furthermore, activation of G-proteins in turn
induces integrin-clustering and a higher ligand-affinity. This bidirectional interaction is
known to regulate cytoskeletal rearrangements (Schwartz and Shattil, 2000)
putatively involved in bacteria-cell interaction.
As described in our previous study suilysin promotes invasion of S. suis by a
Rac-dependent activation of the actin cytoskeleton (Seitz et al., submitted; cf. chapter
3). We showed that rSLY specifically activated Rac1, a member of small

Results, part II Chapter 4
103
Rho-GTPases, in HEp-2 cells at subcytolytic concentration (100 ng/ml), thus
suggesting a pore-independent mechanism. Moreover, the mutated suilysin rW461F,
deficient in functional pore-formation, still retained its ability to activate Rac1. To
elucidate, if the RGD-motif is required for activation of Rho signalling G-proteins, we
performed pull down based GLISA analysis. HEp-2 cells were treated with rSVD in
comparison to rSLY and rW461F used at subcytolytic concentration (100 ng/ml) for
15 min. Results revealed an activation of Rac1 after stimulation of HEp-2 cells with
either rSLY or W461F (Figure 4-5A), confirming previous results (Seitz et al.,
submitted; cf. chapter 3). As anticipated, neither RhoA nor Cdc42 were inducible.
Interestingly, in contrast to rSLY and W461F, rSVD failed to activate any
Rho-GTPase (Figure 4-5A), indicating the requirement of the RGD-motif for
suilysin-induced host cell signalling.
Since macropore-formation is the final step in cytolysis, we speculated that the point-
mutations of suilysin might abolish macropore-formation. Therefore, we used the
fluorescent marker calcein to test cell membrane integrity. HEp-2 cells were
incubated with recombinant proteins (rSLY, rW461F or rSVD) at subcytolytic (100
ng/ml, Figure 4-5B) or lytic concentrations (400 ng/ml, Figure 4-5C) to induce large
channel formation in lipid bilayers. As intact cell membranes are impermeable for
calcein due to its size (diameter of 1.3 nm) macropore-formation (20 - 30 nm in size)
is required for incorporation of calcein by cells. As anticipated, rSLY was capable to
induce macropores in epithelial cells at lytic concentrations (Figure 4-5C), resulting in
a respective shift of the curve (cell population) on the x-axis. Both rW461F and rSVD
failed to form large pores. In accordance to these results, a homologous tryptophan-
mutant of pneumolysin formed large pores to significant lower degree and less
frequently compared to wild type pneumolysin analyzed by conductance
measurement of patch clamps (El Rachkidy et al., 2008; Korchev et al., 1998).
Furthermore, it has been found that rapid activation of RhoA and Rac1 in neuronal
cells by pneumolysin preceded the formation of lytic macropores (Iliev et al., 2007).
Likewise, the tryptophan-mutant of suilysin still retained Rac1-activation, but failed to
induce large channels. This suggests that cell-activation occurs before
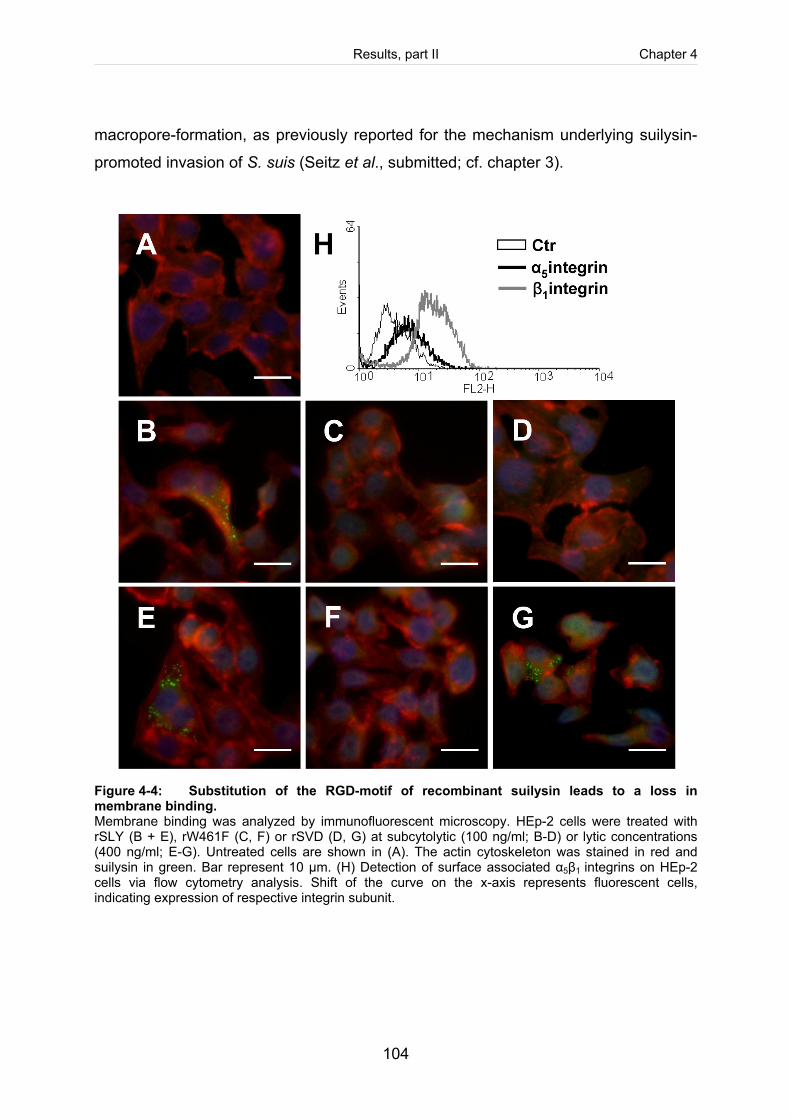
Results, part II Chapter 4
104
macropore-formation, as previously reported for the mechanism underlying suilysin-
promoted invasion of S. suis (Seitz et al., submitted; cf. chapter 3).
Figure 4-4: Substitution of the RGD-motif of recombinant suilysin leads to a loss in membrane binding. Membrane binding was analyzed by immunofluorescent microscopy. HEp-2 cells were treated with rSLY (B + E), rW461F (C, F) or rSVD (D, G) at subcytolytic (100 ng/ml; B-D) or lytic concentrations (400 ng/ml; E-G). Untreated cells are shown in (A). The actin cytoskeleton was stained in red and suilysin in green. Bar represent 10 µm. (H) Detection of surface associated α5β1 integrins on HEp-2 cells via flow cytometry analysis. Shift of the curve on the x-axis represents fluorescent cells, indicating expression of respective integrin subunit.
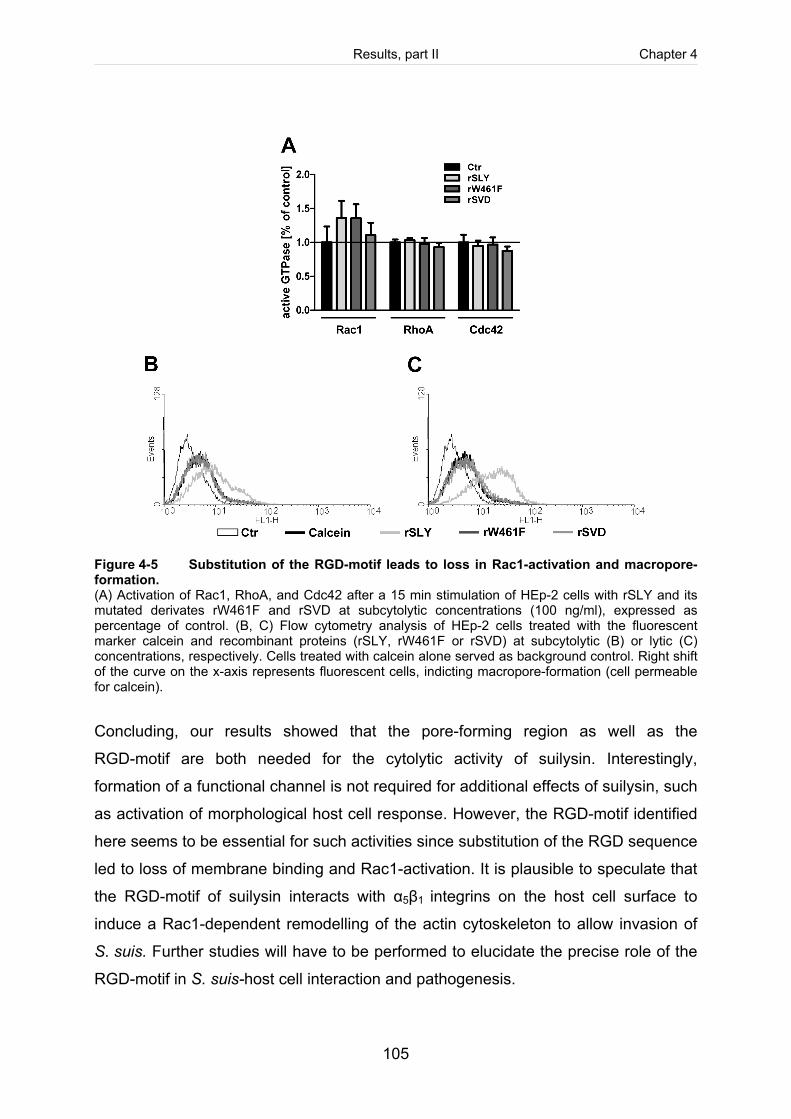
Results, part II Chapter 4
105
Figure 4-5 Substitution of the RGD-motif leads to loss in Rac1-activation and macropore-formation. (A) Activation of Rac1, RhoA, and Cdc42 after a 15 min stimulation of HEp-2 cells with rSLY and its mutated derivates rW461F and rSVD at subcytolytic concentrations (100 ng/ml), expressed as percentage of control. (B, C) Flow cytometry analysis of HEp-2 cells treated with the fluorescent marker calcein and recombinant proteins (rSLY, rW461F or rSVD) at subcytolytic (B) or lytic (C) concentrations, respectively. Cells treated with calcein alone served as background control. Right shift of the curve on the x-axis represents fluorescent cells, indicting macropore-formation (cell permeable for calcein).
Concluding, our results showed that the pore-forming region as well as the
RGD-motif are both needed for the cytolytic activity of suilysin. Interestingly,
formation of a functional channel is not required for additional effects of suilysin, such
as activation of morphological host cell response. However, the RGD-motif identified
here seems to be essential for such activities since substitution of the RGD sequence
led to loss of membrane binding and Rac1-activation. It is plausible to speculate that
the RGD-motif of suilysin interacts with α5β1 integrins on the host cell surface to
induce a Rac1-dependent remodelling of the actin cytoskeleton to allow invasion of
S. suis. Further studies will have to be performed to elucidate the precise role of the
RGD-motif in S. suis-host cell interaction and pathogenesis.

Results, part II Chapter 4
106
Experimental procedures
If not stated otherwise all materials were purchased from Sigma (Muenchen,
Germany).
Bacterial strain and growth conditions. Escherichia (E.) coli strains BL21, BL21
(DE3) and DH5α were used for molecular cloning and protein expression
experiments. E. coli were grown in Luria Bertani (LB) medium overnight under
aerobic conditions at 37°C with vigorous shaking. For heterologously expression of
suilysin Lactococcus lactis subsp. cremoris MG1363 (L. lactis) was used as
described recently (Baums et al., 2006).
DNA techniques. Routine molecular biology techniques including restriction
endonuclease digestion, DNA ligations, agarose gel electrophoresis, transformation
of E. coli and plasmid isolation were performed according to standard procedures
(Sambrook et al., 1989). Restriction enzymes were purchased from New England
Biolabs (Frankfurt am Main, Germany). Plasmid preparations were performed with
kits from Machery-Nagel (Dueren, Germany).
Construction and expression of recombinant wild type suilysin and its point-mutated derivates W461F and RGD-SVD. W461F and RGD-SVD substitutions
were constructed using site-directed-mutagenesis according to the instruction
manual of QuikChange® Site-Directed Mutagenesis Kit (Stratagene, Ja Jolla, CA) as
described in a previous study (Seitz et al., submitted; cf. chapter 3). As template DNA
pET45bslynew was used (Kock et al., 2009). For RGD replacement the
oligonucleotide primers encoding R124-G125-D126 to S124-V125-D126
slyRGDMutfor (CAGTATTGCGTCGGTAGATCTGACGCTTAG) and slyRGDMutrev
(CTAAGCGTCAGATCTACCGACGCAATACTG) were used. Mutation sites of each
suilysin derivate were sequenced using the primers slyseqfor
(GGATCATTCAGGTGCTTATG) and pET45seqrev (TGCTGGCGTTCAAATTTCGC).
Recombinant His-tagged suilysin (rSLY) was expressed in E. coli BL21

Results, part II Chapter 4
107
pET45bslynew (Kock et al., 2009). In accordance, recombinant point-mutated
suilysin W461F (rW461F) was expressed in E. coli BL21 pET45bslynewW461F as
described in a previous study (Seitz et al., submitted; cf. chapter 3) and recombinant
RGD-SVD (rSVD) in E. coli BL21 pET45bslynewRGD-SVD. Purification of all
recombinant proteins was verified by immunoblot analysis with anti-Penta His
antibodies (Qiagen, Hilden, Germany) as recommended and silver staining was used
to control the purity grade. The proteins were stored at -20°C. The wild type suilysin
was completely sequenced with standard oligonucleotid primers T7
(TAATACGACTCACTATAGGG) and T7 term (CTAGTTATTGCTCAGCGGT;
Eurofins MWG operon, Ebersberg, Germany).
Heterologous expression of SLY, W461F and SVD in L. lactis. Heterologous
expression in L. lactis was performed as described previously (Baums et al., 2006).
The sly-gene was amplified from chromosomal DNA of Streptococcus suis by PCR
with oligonucleotide primers slyPstI (TAGTCTGCAGCTCCTAGCCTCTGTGGCT AA)
and slyBamHIoptRBS (CAGAGGATCCAGGAGAAAACTTATGAGAAAAAG). After
digestion of the PCR product with BamHI and PstI the fragment containing the whole
sly-gene was cloned into the BamHI/PstI-digested shuttle vector pORI23 (Que et al.,
2000). The resulting plasmid was named pORI23-SLY and transferred in L. lactis as
described previously (Holo and Nes, 1989b). Transformants were screen by plating
on GM17 agar plates containing 5 µg/ml erythromycin. For heterologous expression
of W461F and SVD site-directed mutagenesis was performed as described above
using purified plasmid DNA pORI23sly and the respective oligonucleotid primers
slyTrp-Phefornew and slyTrp-Pherewnew as well as slyRGDMutfor and
slyRGDMutrev. Resulting constructs pORI23-W461F and pORI23-SVD were
electroporated in L. lactis.
Immunoblot analysis. Recombinant proteins were separated by SDS-polyacrylamid
gel electrophoresis with a 4% stacking and a 10% separating gel under denaturing
conditions and transferred to a PVDF-membrane (Serva, Heidelberg, Germany). For
Immunoblot analysis, membrane-blocking was performed overnight with 3% milk
powder in TBS with 0.5% Tween. Polyclonal antiserum raised against rSLY (Benga

Results, part II Chapter 4
108
et al., 2008) diluted 1:1,300 in 1% milk powder was used to detect either rSLY,
rW461F, rSVD or suilysin and its mutated derivates in culture supernatants of the
respective L. lactis strains (L. lactis pORI-SLY, L. lactis pORI-W461F and L. lactis
pORI-SVD). For detection of His-tagged recombinant proteins membranes were
blocked with 3% BSA in TBS with 0.5% Tween overnight and incubated with a
monoclonal anti-Penta His antibody (Qiagen) diluted 1:2,000 in 3% BSA. Membranes
were developed with horseradish peroxidase-conjugated anti-rabbit or anti-mouse
IgG antiserum diluted 1:10000 in 1% milk powder (Amersham, Freiburg, Germany)
followed by addition of SuperSignal® West Pico Chemiluminescent Substrate (Pierce,
Rockford, USA) according to the manufactures protocol.
HEp-2 epithelial cell culture. The human laryngeal epithelial cell line HEp-2 (ATCC
CCL 23) was used. Cells were maintained in Dulbecco’s modified Eagle’s medium
(DMEM, Gibco-Invitrogen, Groningen, The Netherlands) supplemented with 10%
fetal calf serum (FCS) and 5 mM glutamine at 37°C and 8% CO2. The cells were
subcultured every 2-3 days after detachment with 0.25% trypsin and 1 mM Na-EDTA
(trypsin-EDTA, Gibco-Invitrogen). For cellular cytotoxicity assay 0.2 x 105 cells were
seeded on a 96 well plate. For immunofluorescence 0.8 x 105 cells per well were
seeded on a 12 mm diameter glass cover slips placed in 24 well plates. The cells
were grown over-night and then used for experiments.
Determination of haemolytic activity and cytotoxicity. The haemolytic activity of
rSLY, rW461F and rSVD was determined by a haemolysis assay as described in a
previously (Seitz et al., submitted; cf. chapter 3; Takamatsu et al., 2001). Cytotoxicity
of recombinant proteins was detected by an LDH-release assay as described
previously (Seitz et al.; submitted; cf. chapter 3; Benga et al., 2004).
Briefly, L. lactis expressing either SLY, W461F or SVD (L. lactis pORI-SLY, -W461F
and –SVD), respectively, were grown in GM17 medium overnight, adjusted to an
optical density (OD600) of 0.02 in pre-warmed media and then grown to late
exponential growth phase (OD600 0.8). Subsequently bacteria were removed by
centrifugation and culture supernatants were concentrated and purified using Amicon
centrifugal filter devices 30 kDa (Millipore, Schwalbach/Ts., Germany) to remove

Results, part II Chapter 4
109
remaining medium components. Concentrated culture supernatants diluted 1:5 in
0.9% NaCl were used as test samples.
Immunofluorescent microscopy. Membrane binding of rSLY, rW461F and rSVD
was determined by immunofluorescent microscopy. HEp-2 cells were incubated with
the recombinant suilysin derivates at subcytolytic (100 ng/ml) or lytic (400 ng/ml)
concentrations for 30 min at 37°C. Cells were washed three times to remove
unbound suilysin, fixed with 2% parafolmaldehyde and permeabilised with
0.1% Triton X-100 in PBS (Bio-Rad, Muenchen, Germany). Blocking was performed
for 30 min at room temperature with PBS containing 10% FCS. Suilysin was stained
using a polyclonal antiserum raised against rSLY (Benga et al., 2008) diluted 1:1,300
in 1% milk powder for 1 h at room temperature followed by an incubation with
FITC-conjugated goat anti-rabbit antibody (1:1,000 in 1% FCS in PBS, Dianova,
Hamburg, Germany). F-actin was labelled with TRITC-conjugated Phalloidin (20
µg/ml, Invitrogen) for 40 min at room temperature. After final washing DAPI
(Invitrogen) was used for staining the nuclei. Antifading reagent DABCO (Sigma) was
used for sealing of the samples. Mounted samples were examined using inverted
immunofluorescence microscope Nikon Eclipse Ti-S equipped with a 40×, 0.6 S Plan
Fluor objective (Nikon, Duesseldorf, Germany) driven by NIS Elements software BR
3.2..
Detection of α5β1 integrin expression on HEp-2 cells. HEp-2 cell-suspension was
incubated with either a monoclonal anti-human CD29 antibody or a monoclonal anti-
human CD49e (Bio Legend) both used at a concentration of 500 ng per
1.5 x 106 cells (Bio Legend, Fell, Germany). All antibodies were diluted in blocking
buffer (PBS [pH 7.3] containing 3% BSA) and incubation was performed for 30 min at
room temperature with gently agitation. After three washing steps with blocking
buffer, cells were incubated with a TRITC-conjugated goat anti-mouse antibody
(1:1000, Dianova, Hamburg, Germany). Stained cells were washed three times, fixed
with 0.37% formaldehyde and resuspended in blocking buffer for subsequent
measurement. Flow cytometry measurement was performed as described above.

Results, part II Chapter 4
110
Fluorescent cells were counted at channel FL-1 and unstained cells severed as
background control. GLISA. HEp-2 cells were treated with rSLY, rW461F or rSVD at subcytolytic
concentration (100 ng/ml) for 15 min and rinsed twice with ice cold PBS to stop
incubation. Lysis of cells was achieved by addition of 1 ml of ice cold lysis buffer
(50 mM NaCl, 20 mM Tris-HCl, pH 7.4, 3 mM MgCl2, 1% Nonidet-P 40,
0.25% Triton X-100, 5 mM dithiothreitol, 100 µM PMSF). Cells were scraped off, and
the lysates were centrifuged at 14,000 rpm. The supernatant was used for a pull
down-based G-LISA (Cytoskeleton, CO, USA) to determine activation of Rac1,
RhoA, and Cdc42 according to the protocol by the supplier.
Cell-permeability and macropore-formation assay. Experiments were performed
in PBS containing 0.9 mM CaCl2/0.5 mM MgCl26H2O [pH 7.3] (Gibco-Invitrogen). Cell
permeability was determined after co-incubation of HEp-2 cells (4 x 105 cells/ml) with
recombinant proteins (rSLY, rW461F, and rSVD) at subcytolytic (100 ng/ml) or lytic
(400 ng/ml) concentrations (cf. cytotoxicity test) and the fluorescent marker calcein
(2 µg/ml, Sigma, Taufkirchen, Germany). Incubation was performed for 2 h at 37°C
on an end-over-end rotator. After two washing steps with PBS to remove extracellular
calcein, cells were fixed with 0.37% formaldehyde (for 10 min at room temperature
and then resuspended in 500 µl PBS for measurement of fluorescent cells using
FACScan® (Becton Dickinson, 488 nm Argon laser). Further analysis was performed
with the software WinMDI (version 2.9.). For each determination at least
10,000 events were measured. Initial analysis of fluorescent cells was carried out by
dot plot analysis (forward scatter [fsc] versus sideward scatter [ssc]) to define the cell
population of interest (data not shown). Subsequently, fluorescent cells were
detected at channel FL-1. Cells treated only with calcein served as background
control.
Acknowledgements This work was financially supported by the Deutsche Forschungsgemeinschaft
(DFG), Bonn, Germany (SFB 587).

Results, part II Chapter 4
111
References Baums, C. G., Kaim, U., Fulde, M., Ramachandran, G., Goethe, R., and Valentin-Weigand, P. (2006) Identification of a novel virulence determinant with serum opacification activity in Streptococcus suis. Infect Immun 74: 6154-6162.
Benga, L., Fulde, M., Neis, C., Goethe, R., and Valentin-Weigand, P. (2008) Polysaccharide capsule and suilysin contribute to extracellular survival of Streptococcus suis co-cultivated with primary porcine phagocytes. Vet Microbiol 132: 211-219.
Benga, L., Goethe, R., Rohde, M., and Valentin-Weigand, P. (2004) Non-encapsulated strains reveal novel insights in invasion and survival of Streptococcus suis in epithelial cells. Cellular Microbiology 6: 867-881.
Boulnois, G. J., Paton, J. C., Mitchell, T. J., and Andrew, P. W. (1991) Structure and function of pneumolysin, the multifunctional, thiol-activated toxin of Streptococcus pneumoniae. Mol Microbiol 5: 2611-2616.
Cue, D., Southern, S. O., Southern, P. J., Prabhakar, J., Lorelli, W., Smallheer, J. M., Mousa, S. A., and Cleary, P. P. (2000) A nonpeptide integrin antagonist can inhibit epithelial cell ingestion of Streptococcus pyogenes by blocking formation of integrin alpha 5beta 1-fibronectin-M1 protein complexes. Proc Natl Acad Sci U S A 97: 2858-2863.
Czajkowsky, D. M., Hotze, E. M., Shao, Z., and Tweten, R. K. (2004) Vertical collapse of a cytolysin prepore moves its transmembrane beta-hairpins to the membrane. EMBO J 23: 3206-3215.
El Rachkidy, R. G., Davies, N. W., and Andrew, P. W. (2008) Pneumolysin generates multiple conductance pores in the membrane of nucleated cells. Biochem Biophys Res Commun 368: 786-792.
Feldman, C., Mitchell, T. J., Andrew, P. W., Boulnois, G. J., Read, R. C., Todd, H. C., Cole, P. J., and Wilson, R. (1990) The effect of Streptococcus pneumoniae pneumolysin on human respiratory epithelium in vitro. Microb Pathog 9: 275-284.
Gekara, N. O. and Weiss, S. (2004) Lipid rafts clustering and signalling by listeriolysin O. Biochem Soc Trans 32: 712-714.
Giddings, K. S., Johnson, A. E., and Tweten, R. K. (2003) Redefining cholesterol's role in the mechanism of the cholesterol-dependent cytolysins. Proc Natl Acad Sci U S A 100: 11315- 11320.
Gottschalk, M., Segura, M., and Xu, J. (2007) Streptococcus suis infections in humans: the Chinese experience and the situation in North America. Anim Health Res Rev 8: 29-45.
Gottschalk, M., Xu, J., Calzas, C., and Segura, M. (2010) Streptococcus suis: a new emerging or an old neglected zoonotic pathogen? 25. Future Microbiol 5: 371-391. Hamzaoui, N., Kerneis, S., Caliot, E., and Pringault, E. (2004) Expression and distribution of beta1 integrins in in vitro-induced M cells: implications for Yersinia adhesion to Peyer's patch epithelium. Cell Microbiol 6: 817-828.
Holo, H. and Nes, I. F. (1989) High-Frequency Transformation, by Electroporation, of Lactococcus lactis subsp. cremoris Grown with Glycine in Osmotically Stabilized Media. Appl Environ Microbiol 55: 3119-3123.

Results, part II Chapter 4
112
Hotze, E. M. and Tweten, R. K. (2011) Membrane assembly of the cholesterol-dependent cytolysin pore complex. Biochim Biophys Acta.
Iliev, A. I., Djannatian, J. R., Nau, R., Mitchell, T. J., and Wouters, F. S. (2007) Cholesterol-dependent actin remodeling via RhoA and Rac1 activation by the Streptococcus pneumoniae toxin pneumolysin. Proc Natl Acad Sci U S A 104: 2897-2902.
Jacobs, A. A. C., Loeffen, P. L. W., vandenBerg, A. J. G., and Storm, P. K. (1994) Identification, Purification, and Characterization of A Thiol-Activated Hemolysin (Suilysin) of Streptococcus Suis. Infection and Immunity 62: 1742-1748.
Jonsson, K., Signas, C., Muller, H. P., and Lindberg, M. (1991) Two different genes encode fibronectin binding proteins in Staphylococcus aureus. The complete nucleotide sequence and characterization of the second gene. Eur J Biochem 202: 1041-1048.
Kock, C., Beineke, A., Seitz, M., Ganter, M., Waldmann, K. H., Valentin-Weigand, P., and Baums, C. G. (2009) Intranasal immunization with a live Streptococcus suis isogenic ofs mutant elicited suilysin-neutralization titers but failed to induce opsonizing antibodies and protection. Veterinary Immunology and Immunopathology 132: 135-145.
Korchev, Y. E., Bashford, C. L., Pederzolli, C., Pasternak, C. A., Morgan, P. J., Andrew, P. W., and Mitchell, T. J. (1998) A conserved tryptophan in pneumolysin is a determinant of the characteristics of channels formed by pneumolysin in cells and planar lipid bilayers. Biochem J 329 ( Pt 3): 571-577.
Lafont, F. and van der Goot, F. G. (2005) Bacterial invasion via lipid rafts. Cell Microbiol 7: 613-620.
Lalonde, M., Segura, M., Lacouture, S., and Gottschalk, M. (2000) Interactions between Streptococcus suis serotype 2 and different epithelial cell lines. Microbiology 146 ( Pt 8): 1913- 1921.
Michel, E., Reich, K. A., Favier, R., Berche, P., and Cossart, P. (1990) Attenuated mutants of the intracellular bacterium Listeria monocytogenes obtained by single amino acid substitutions in listeriolysin O. Mol Microbiol 4: 2167-2178.
Norton, P. M., Rolph, C., Ward, P. N., Bentley, R. W., and Leigh, J. A. (1999) Epithelial invasion and cell lysis by virulent strains of Streptococcus suis is enhanced by the presence of suilysin. Fems Immunology and Medical Microbiology 26: 25-35.
Polekhina, G., Giddings, K. S., Tweten, R. K., and Parker, M. W. (2005) Insights into the action of the superfamily of cholesterol-dependent cytolysins from studies of intermedilysin. Proc Natl Acad Sci U S A 102: 600-605.
Que, Y. A., Haefliger, J. A., Francioli, P., and Moreillon, P. (2000) Expression of Staphylococcus aureus clumping factor A in Lactococcus lactis subsp. cremoris using a new shuttle vector. Infect Immun 68: 3516-3522.
Ramachandran, R., Heuck, A. P., Tweten, R. K., and Johnson, A. E. (2002) Structural insights into the membrane-anchoring mechanism of a cholesterol-dependent cytolysin. Nat Struct Biol 9: 823- 827.
Rossjohn, J., Feil, S. C., McKinstry, W. J., Tweten, R. K., and Parker, M. W. (1997) Structure of a cholesterol-binding, thiol-activated cytolysin and a model of its membrane form. Cell 89: 685- 692.

Results, part II Chapter 4
113
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N Y.
Schwartz, M. A. and Shattil, S. J. (2000) Signaling networks linking integrins and rho family GTPases. Trends Biochem Sci 25: 388-391.
Scibelli, A., Matteoli, G., Roperto, S., Alimenti, E., Dipineto, L., Pavone, L. M., Della, Morte R., Menna, L. F., Fioretti, A., and Staiano, N. (2005) Flavoridin inhibits Yersinia enterocolitica uptake into fibronectin-adherent HeLa cells. FEMS Microbiol Lett 247: 51-57.
Scibelli, A., Roperto, S., Manna, L., Pavone, L. M., Tafuri, S., Della, Morte R., and Staiano, N. (2007) Engagement of integrins as a cellular route of invasion by bacterial pathogens. Vet J 173: 482- 491.
Seitz, M., Baums, CG, Gerhard, R, Just, I, Neis, C., Benga, L., Fulde, M., Nerlich, A, Rohde, M., Goethe, R., and Valentin-Weigand, P. Subcytolytic activity of suilysin promotes invasion of Streptococcus suis in HEp-2 epithelial cells by Rac-dependent activation of the actin cytoskeleton. Ref Type: submitted to Cellular Microbiology Sekino-Suzuki, N., Nakamura, M., Mitsui, K. I., and Ohno-Iwashita, Y. (1996) Contribution of individual tryptophan residues to the structure and activity of theta-toxin (perfringolysin O), a cholesterol- binding cytolysin. Eur J Biochem 241: 941-947.
Shepard, L. A., Shatursky, O., Johnson, A. E., and Tweten, R. K. (2000) The mechanism of pore assembly for a cholesterol-dependent cytolysin: formation of a large prepore complex precedes the insertion of the transmembrane beta-hairpins. Biochemistry 39: 10284-10293.
Shimada, Y., Maruya, M., Iwashita, S., and Ohno-Iwashita, Y. (2002) The C-terminal domain of perfringolysin O is an essential cholesterol-binding unit targeting to cholesterol-rich microdomains. Eur J Biochem 269: 6195-6203.
Takamatsu, D., Osaki, M., and Sekizaki, T. (2001) Thermosensitive suicide vectors for gene replacement in Streptococcus suis. Plasmid 46: 140-148.
Tang, J., Wang, C., Feng, Y., Yang, W., Song, H., Chen, Z., Yu, H., Pan, X., Zhou, X., Wang, H., Wu, B., Wang, H., Zhao, H., Lin, Y., Yue, J., Wu, Z., He, X., Gao, F., Khan, A. H., Wang, J., Zhao, G. P., Wang, Y., Wang, X., Chen, Z., and Gao, G. F. (2006) Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med 3: e151.
Tenenbaum, T., Essmann, F., Adam, R., Seibt, A., Janicke, R. U., Novotny, G. E., Galla, H. J., and Schroten, H. (2006) Cell death, caspase activation, and HMGB1 release of porcine choroid plexus epithelial cells during Streptococcus suis infection in vitro. Brain Res 1100: 1-12.
Tran, Van Nhieu, Caron, E., Hall, A., and Sansonetti, P. J. (1999) IpaC induces actin polymerization and filopodia formation during Shigella entry into epithelial cells. EMBO J 18: 3249-3262.
Trottier, S., Higgins, R., Brochu, G., and Gottschalk, M. (1991) A Case of Human Endocarditis Due to Streptococcus Suis in North-America. Reviews of Infectious Diseases 13: 1251-1252.
Tweten, R. K. (2005) Cholesterol-dependent cytolysins, a family of versatile pore-forming toxins. Infection and Immunity 73: 6199-6209.
Vanier, G., Segura, M., Friedl, P., Lacouture, S., and Gottschalk, M. (2004) Invasion of porcine brain microvascular endothelial cells by Streptococcus suis serotype 2. Infection and Immunity 72: 1441-1449.

Results, part II Chapter 4
114
Waheed, A. A., Shimada, Y., Heijnen, H. F., Nakamura, M., Inomata, M., Hayashi, M., Iwashita, S., Slot, J. W., and Ohno-Iwashita, Y. (2001) Selective binding of perfringolysin O derivative to cholesterol-rich membrane microdomains (rafts). Proc Natl Acad Sci U S A 98: 4926-4931.
Xu, L., Huang, B., Du, H., Zhang, X. C., Xu, J., Li, X., and Rao, Z. (2010a) Crystal structure of cytotoxin protein suilysin from Streptococcus suis. Protein Cell 1: 96-105.

Chapter 5
Results, part III:
Establishment of an intranasal CD1 mouse infection model for colonization and invasion of Streptococcus suis serotype 2
M. Seitz, A. Beineke, J. Seele, M. Fulde, P. Valentin-Weigand and C. G. Baums
Running title: Intranasal mouse infection model for S. suis
(Manuscript in preparation)


Results, part III Chapter 5
117
Abstract Streptococcus (S.) suis causes meningitis and various other diseases in pigs and
humans. Healthy piglets carrying virulent S. suis strains on their mucosal surfaces
are epidemiologically very important. The objective of this study was to establish an
intranasal mouse model for colonisation and invasion of the respiratory tract. CD1
mice were intranasally infected with a highly virulent S. suis serotype 2 strain under
different conditions of predisposition. Clinical, histological and bacteriological
examination revealed that invasion of host tissues occurred only in mice predisposed
intranasally with 1% acetic acid. This model is characterized by efficient colonization
as all mice carried S. suis on their respiratory mucosa 7 days post infection.
Furthermore, severe fibrinosuppurative or purulent necrotizing pneumonia associated
with S. suis was a common manifestation in this model. The intranasal S. suis model
was applied to investigate the relevance of suilysin in colonisation and invasion by
comparison of the wild type with its isogenic sly-mutant. Results suggested
attenuation in virulence but not in colonization of the suilysin mutant. In conclusion,
this study revealed the first intranasal mouse model to study colonization and
invasion of the respiratory tract by a highly virulent S. suis pathotype.

Results, part III Chapter 5
118
Introduction Streptococcus (S.) suis is a major swine pathogen worldwide, causing severe
diseases such as meningitis, septicaemia and bronchopneumonia (Higgins and
Gottschalk, 2005). It is also an important zoonotic agent. Humans might be infected
following contact with pigs or pork. Meningitis, septicaemia and the life-threatening
streptococcal toxic shock-like syndrome (STSS) are important manifestations of
S. suis infections in humans (Gottschalk et al., 2007; Tang et al., 2006).
Pigs and wild boars are considered the natural reservoir of S. suis (Baums et al.,
2007; Clifton-Hadley and Alexander, 1980; Higgins and Gottschalk, 2005). Different
mucosal surfaces might be colonized by S. suis. In weaning piglets, S. suis is among
other bacteria the most abundant colonizers of the upper respiratory and alimentary
tract (Baele et al., 2001; Lowe et al., 2011; O'Sullivan et al., 2011a; Su et al., 2008).
Healthy carriers of virulent S. suis strains play an important role in the epidemiology
of S. suis diseases in pig and humans (Arends et al., 1984; Ngo et al., 2011).
S. suis is characterized by a high diversity as reflected by the presence of at least 33
serotypes. Serotype 2 is worldwide the most prevalent among invasive isolates of
pigs and humans (Wei et al., 2009; Wisselink et al., 2000). The serotype is
determined by the polysaccharide capsule. The capsule protects the bacteria against
opsonophagocytosis and functions as an important virulence factor (Charland et al.,
1998; Smith et al., 1999). A number of other surface-associated factors have also
been demonstrated to contribute to pathogenicity of S. suis (Baums and Valentin-
Weigand, 2009). The pathogenesis of S. suis meningitis is not well understood, and
even less is known about the mechanisms employed by S. suis to colonize mucosa.
As a matter of fact, not a single factor of S. suis has been demonstrated to be crucial
for colonization.
Recently, a S. suis meningitis model was described in CD1 mouse, in which typical
histopathological lesions and inflammatory responses were recorded. Sudden death
of 20% of infected animals associated with high levels of systemic proinflammatory
cytokines was observed. Infected mice that survived developed clinical signs of
meningitis characterized by infiltrates of neutrophils and bacterial emboli

Results, part III Chapter 5
119
(Dominguez-Punaro et al., 2007). Similar to many other experimental infections of
mice, S. suis was applied intraperitoneally. As the upper respiratory tract and, in
particular, the tonsils are considered to be the port of entry for S. suis (Williams and
Blakemore, 1990), early steps in the pathogenesis of S. suis diseases cannot be
studied in the CD1 mouse model described by Dominguez-Punaro et al. (2007).
Thus, investigation of colonization of the respiratory mucosa requires a new murine
intranasal S. suis model. Here, we describe for the first time that S. suis serotype 2
colonizes and invades mice tissue efficiently after intranasal application following
predisposition. This work includes a description of an intranasal colonization and
invasion model for S. suis using CD1 mice. Furthermore, the model was applied to
identify suilysin-dependent phenotypes in vivo using encapsulated as well as
unencapsulated strains.

Results, part III Chapter 5
120
Results and Discussion To our knowledge, a mucosal infection model for S. suis in mice has not been
described. In this study we investigated whether a virulent S. suis serotype 2 strain
can infect CD1 mice after intranasal application. As healthy carrier animals play a
crucial role in the epidemiology of S. suis diseases, there is also a urgent need to
study colonization of mucosal surfaces. A major problem in swine is the restricted
availability of S. suis free piglets for such investigations. Therefore, the second
objective of this study was to introduce a mice model for S. suis colonization of
mucosal surfaces. For this, we compared different conditions of experimental
infection without and after predisposition with 1% acetic acid prior intranasal
infection.
Intranasal infection without predisposition. A dose of 5 x 109 CFU of S. suis strain
10 was applied intranasally to Crl:CD1 (ICR) mice, as this mouse strain has been
demonstrated to develop typical lesions of meningitis after intraperitoneal application
of S. suis (Dominguez-Punaro et al., 2007). Nine of 10 Crl:CD1 (ICR) mice did not
show any clinical signs of infection over an observation period of 5 or 12 days
(5 mice each, Table 5-2). The low morbidity in mice infected intranasally with a high
dose of S. suis serotype 2 is in accordance with the findings of Williams et al. (1988).
Apathy, continuing anorexia and a weight loss of more than 20% was registered in
one mouse starting on the 4th dpi (Table 5-2; Table 5-S1). Histophathological
screening revealed severe purulent meningitis and encephalitis associated with a
bacterial load of 4,000 CFU per mg brain tissue (Table 5-3 and Table 5-4).
Furthermore, the challenge strain was also reisolated in 3 of 10 mice in the TNL and
in 2 of 10 mice in the lungs on the 5th day post intranasal infection. The bacterial
load of the respiratory tract of these mice was below 100 CFU per μl TNL or mg
tissue, respectively. These results suggest that S. suis serotype 2 colonized the
respiratory tract and even caused meningitis, though only with low morbidity.

Results, part III Chapter 5
121
Intranasal infection model after predisposition with acetic acid. In piglets, S. suis
mucosal infection models have been described which include experimental
predisposition, such as infection with Bordetella bronchiseptica (Smith et al., 1996;
Smith et al., 1999) or local application of 1% acetic acid (Baums et al., 2006; Pallares
et al., 2003). Therefore we predisposed mice by nasal application of 12.5 µl 1%
acetic acid per nostril 1 h pre intranasal infectionem with S. suis. In this experiment,
morbidity was significantly higher in comparison to the previous experiment without
predisposition (P = 0.003; Table 5-2). Sixty-seven percent of mice received a
cumulative clinical index equal or above 3 and were, thus, classified as diseased
(Table 5-1 and Table 5-2). Six (Table 5-S1) out of 24 infected mice developed severe
clinical symptoms (anorexia, scrubby coat, rapid abdominal breathing, and apathy)
and were euthanized one day post infection for reasons of animal welfare (mortality
rate of 25%). Severe fibrinosuppurative or purulent necrotizing pneumonia as well as
severe infiltration of the splenic red pulp with neutrophils was diagnosed in these six
mice post mortem. S. suis was detected in the lungs of these six mice in high number
(up to 23,000 CFU per mg lung tissue; Table 5-4). Purulent meningitis and/or
encephalitis was found only in one of 24 infected mice. However, the challenge strain
was reisolated from the brain in 46% of these animals. The bacterial load in the brain
of these mice was below 150 CFU per mg brain tissue except in one mouse
(34,000 CFU per mg brain tissue). Severe fibrinosuppurative and even purulent
necrotizing rhinitis was a common finding in surviving mice. The challenge strain was
frequently detected in the lungs but also in other inner organs (Table 5-4).
Unexpectedly, in eight of 24 mice St. aureus was also detected in inner organs (see
number in brackets in Table 5-4 and Table 5-S1). A likely explanation for this finding
was that St. aureus colonizing the respiratory tract in mice might have invaded the
host after local application of 1% acetic acid. However, infection with St. aureus was
not a prerequisite for S. suis infection, since more than 50% of mice positive for
S. suis in the brain, lung, heart or any other organ were free of St. aureus in these
tissues. In this experiment, mice were sacrificed on the 3rd, 5th and 7th days post
infection to monitor the time course of early colonization and invasion as shown in
Table 5-4. The challenge strain was isolated from the upper respiratory tract (TNL
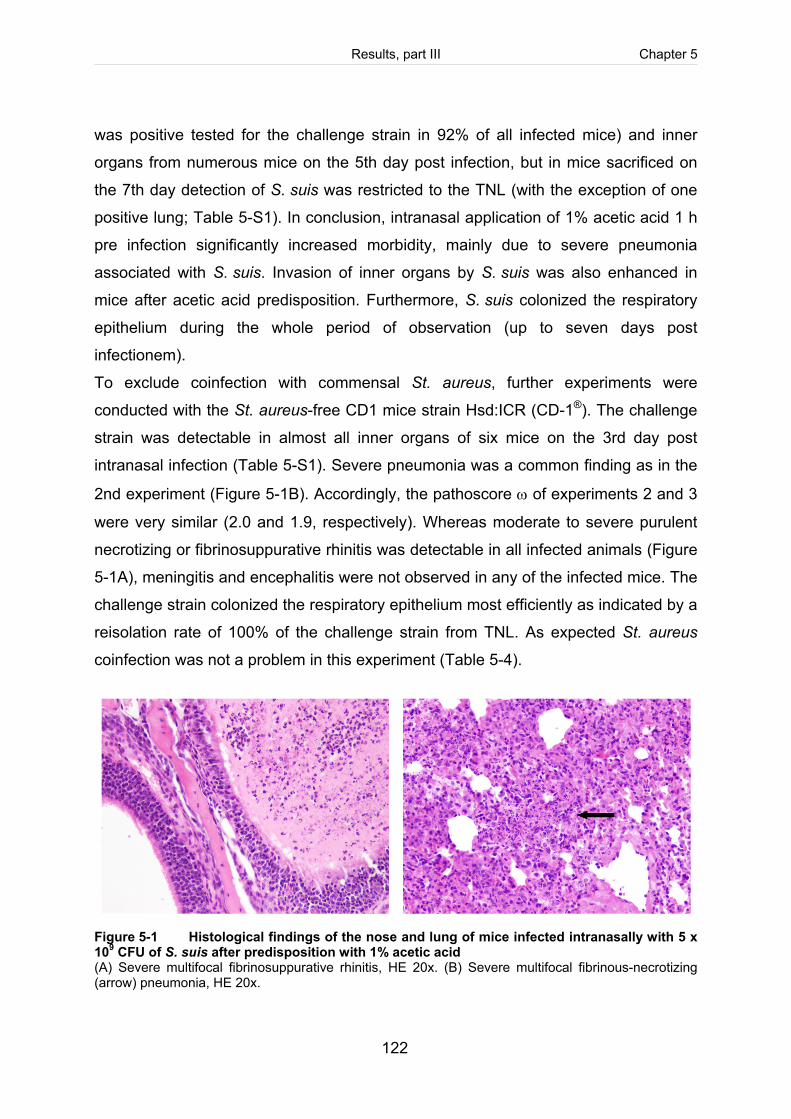
Results, part III Chapter 5
122
was positive tested for the challenge strain in 92% of all infected mice) and inner
organs from numerous mice on the 5th day post infection, but in mice sacrificed on
the 7th day detection of S. suis was restricted to the TNL (with the exception of one
positive lung; Table 5-S1). In conclusion, intranasal application of 1% acetic acid 1 h
pre infection significantly increased morbidity, mainly due to severe pneumonia
associated with S. suis. Invasion of inner organs by S. suis was also enhanced in
mice after acetic acid predisposition. Furthermore, S. suis colonized the respiratory
epithelium during the whole period of observation (up to seven days post
infectionem).
To exclude coinfection with commensal St. aureus, further experiments were
conducted with the St. aureus-free CD1 mice strain Hsd:ICR (CD-1®). The challenge
strain was detectable in almost all inner organs of six mice on the 3rd day post
intranasal infection (Table 5-S1). Severe pneumonia was a common finding as in the
2nd experiment (Figure 5-1B). Accordingly, the pathoscore ω of experiments 2 and 3
were very similar (2.0 and 1.9, respectively). Whereas moderate to severe purulent
necrotizing or fibrinosuppurative rhinitis was detectable in all infected animals (Figure
5-1A), meningitis and encephalitis were not observed in any of the infected mice. The
challenge strain colonized the respiratory epithelium most efficiently as indicated by a
reisolation rate of 100% of the challenge strain from TNL. As expected St. aureus
coinfection was not a problem in this experiment (Table 5-4).
Figure 5-1 Histological findings of the nose and lung of mice infected intranasally with 5 x 109 CFU of S. suis after predisposition with 1% acetic acid (A) Severe multifocal fibrinosuppurative rhinitis, HE 20x. (B) Severe multifocal fibrinous-necrotizing (arrow) pneumonia, HE 20x.

Results, part III Chapter 5
123
The results indicated that application of acetic acid prior to experimental S. suis
infection predisposed mice to rhinitis and pneumonia rather than to meningitis, the
most common pathology of S. suis infection in swine and humans. This is in contrast
to the effect of intranasal acetic acid predisposition in swine, leading mainly to
S. suis-associated pleuritis, peritonitis and meningitis (Baums et al., 2006; Pallares et
al., 2003) and the intraperitoneal model described by Dominguez-Punaro et al.
(2007) associated with a prevalence of meningitis in 40% of mice. However, as
demonstrated by bacteriology results, S. suis colonized the mucosal epithelium up to
seven days post infectionem and invaded into tissue after intranasal application and
acetic acid predisposition. Therefore, this model might be used to study colonization
and invasion of S. suis in mice, using bacteriological screening of different inner
organs as important read out parameter.
Table 5-1: Clinical score sheet for health monitoring of mice after infection with S. suis. Parameter 0 1 2
Body weight Constant or gain > 5% weight los > 20% weight loss
Coat Flat, glossy Rough, reduced grooming Scrubby, failure of grooming
Breathing Adequate, rhythmic Rapid, shallow Rapid, abdominal
Dehydrationa Normal skin elasticity
Moderately reduced skin elasticity Persisting skin fold, sunken eyes
Bearing Normal Moderately curved back Cowering, highly curved back
Eyes Normal Moderately protruding Highly swollen
Activity Normal active Depression Apathy, social isolation
Locomotion No anomaly Moderate incoordination Apraxia, stagger
a Elapsing of a drawn up dorsal skin fold

Results, part III Chapter 5
124
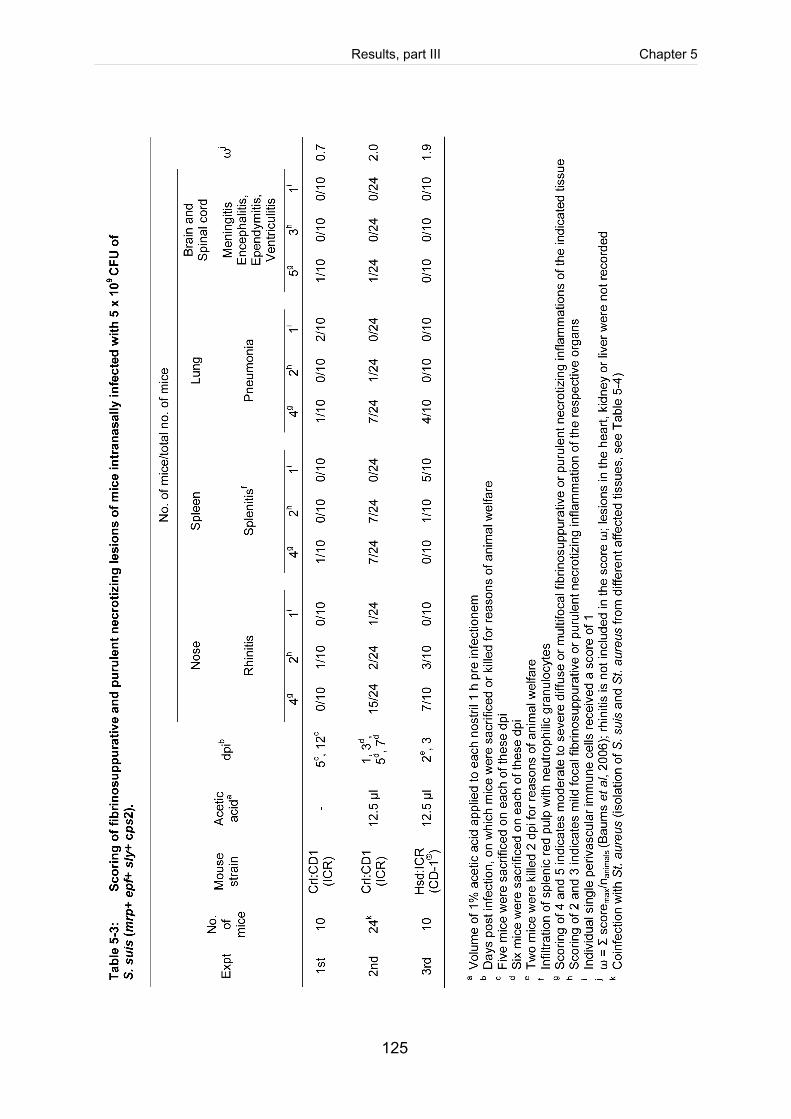
Results, part III Chapter 5
125

Results, part III Chapter 5
126

Results, part III Chapter 5
127
Evaluation of colonization and invasiveness of isogenic sly-mutants. The
intranasal mouse model with acetic acid predisposition was used to investigate the
contribution of suilysin to the invasiveness of S. suis. Six mice each were infected
either with the wild type strain 10 or its isogenic suilysin-deficient mutant (10Δsly).
None of the mice infected with 10Δsly developed clinical symptoms except for a
weight loss >5% one day post infectionem. In comparison, all mice infected with the
wild type strain 10 showed mild clinical symptoms of general sickness including
rough coat, moderate depression and persistent weight loss >5% (morbidity 100%).
No case of severe sickness with obvious neural dysfunction was seen (Table 5-5).
However, histopathological screening revealed one case of severe purulent
encephalitis located in the olfactory bulb in a mouse infected with the wild type strain
10 (Table 5-S1). In accordance with differences in morbidity between wild type and
10Δsly infected mice, the general pathoscore ω was substantially lower in mice
infected with the isogenic sly-mutant in comparison to the score of the wild type
infected group (0.5 versus 1.7, Table 5-6). Attenuation in virulence of the isogenic
sly-mutant is in accordance with results of experimental mouse infections reported by
Allen et al. (2001), who observed substantial suilysin-dependent differences in
mortality at high intraperitoneal doses. However, experimental infections of pigs with
suilysin-negative and wild type strains did not reveal an attenuated phenotype of the
sly-mutant (Allen et al., 2001; Lun et al., 2003).
In contrast to the two previous experiments with acetic acid predisposition cases of
pneumonia were not observed in wild type infected mice (Table 5-6). Furthermore,
the wild type strain was detectable only in the TNL but not in inner organs (Table
5-7). These differences might be related to differences in ages: Mice of experiments
2 and 3 were 4 weeks old at the time of infection, whereas mice of the 4th
experiment were 6 to 7 weeks old. Northworthy, Williams et al. (1988) also observed
that young mice were more susceptible to S. suis infection than old mice.
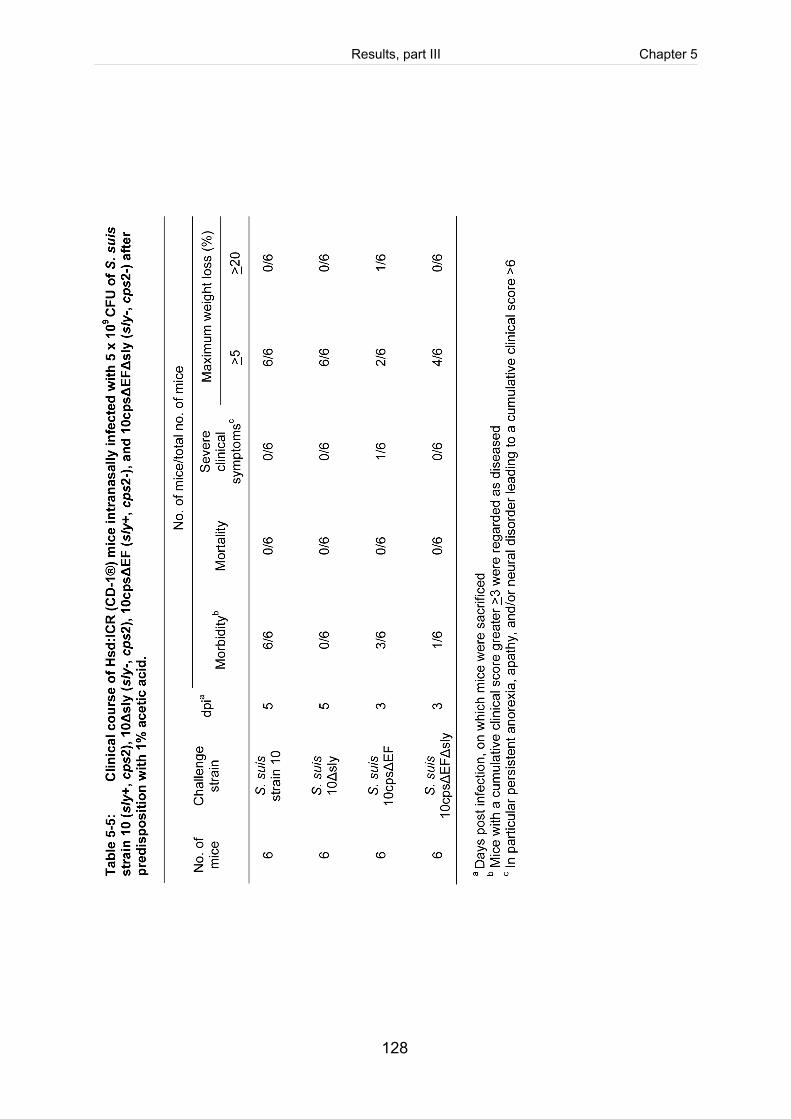
Results, part III Chapter 5
128

Results, part III Chapter 5
129
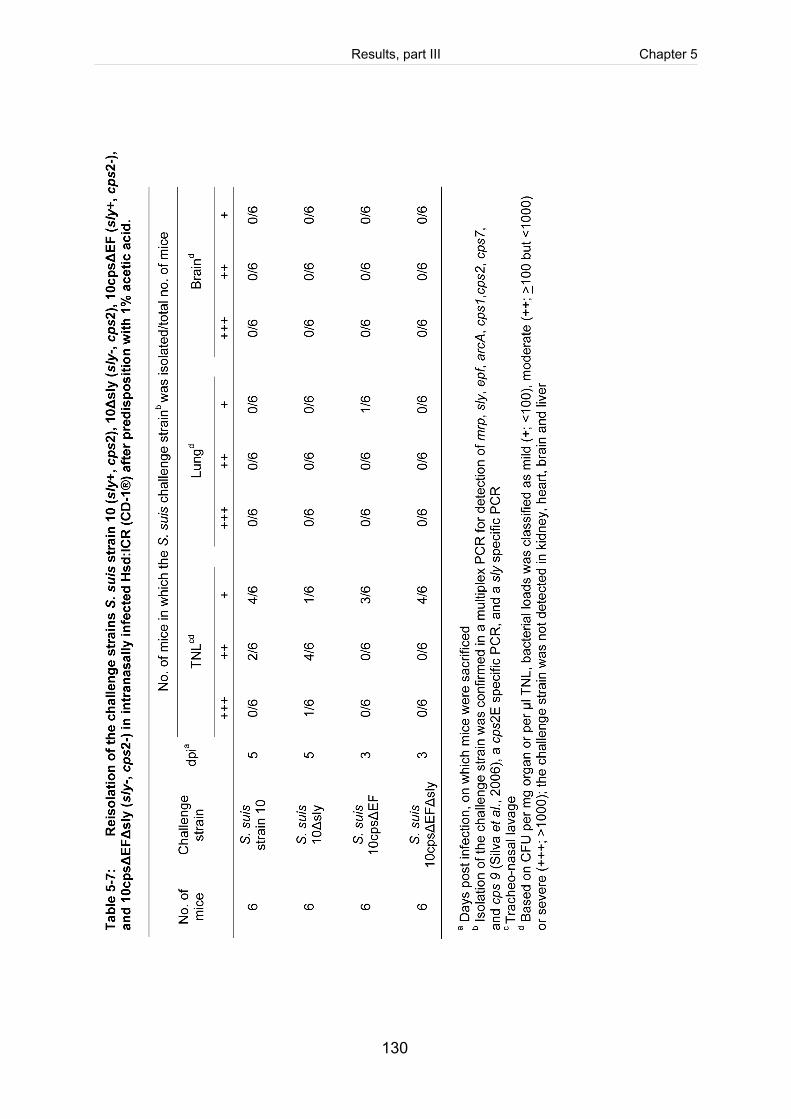
Results, part III Chapter 5
130

Results, part III Chapter 5
131
In conclusion, comparative evaluation of virulence of the wild type and the isogenic
sly-mutant in the intranasal infection model suggested attenuation of the sly-mutant.
Further experiments are necessary to clear why the wild type challenge strain was
not detectable in inner organs and why pathohistological lesions of inner organs were
relatively low, except for one case of severe encephalitis.
In vitro experiments with the respiratory epithelial cell line HEp-2 revealed a suilysin-
dependent invasive phenotype of the isogenic unencapsulated mutant 10cpsΔEF
(Seitz et al., submitted; cf. chapter 3). It is, however, not clear what role suilysin-
dependent invasion of host cells might play for virulence. Unencapsulated S. suis
strains are generally regarded as avirulent because of enhanced
opsonophagocytosis (Charland et al., 1998; Smith et al., 1999). However, Smith et al.
(1999) isolated the isogenic unencapsulated mutants 10cpsΔEF and 10cpsΔB from
CNS, serosae and joints after intranasal infections of gnotobiotic piglets, though
these piglets did not develop clinical signs of disease. Furthermore, unencapsulated
non-typeable stains have been isolated from diseased pigs (Silva et al., 2006).
Therefore, we used the intranasal mouse model to further investigate whether the
unencapsulated strain 10cpsΔEF also showed a suilysin-dependent invasive
phenotype by comparison with the double mutant 10cpsΔEFΔsly. In contrast to the
experiment with the encapsulated strains, mice were sacrificed on the 3rd and not on
the 5th day post infection to increase the probability of reisolation. However, neither
the unencapsulated mutant 10cpsΔEF nor the isogenic suilysin double mutant
(10cpsΔEFΔsly) was detectable in inner organs except for one reisolation for
10cpsΔEF from lung tissue (Table 5-6). Surprisingly, 50% morbidity (mild clinical
symptoms) were observed in mice infected with 10cpsΔEF, of which one mouse
(Table 5-S1) developed severe clinical symptoms (rapid abdominal breathing,
kyphosis, and showed a weight loss >20%; Table 5-5) associated with a mild
histiocytic interstitial pneumonia. The type of the latter lesion is atypical for S. suis
infections. In comparison, only one out of six mice infected with 10cpsΔEFΔsly
developed mild clinical symptoms, like shallow breathing and rough coat (16.7%
morbidity). Though the comparison of morbidity suggested a further suilysin-
dependent attenuation of the 10cpsΔEFΔsly double mutant, results have to be

Results, part III Chapter 5
132
interpreted with caution as the challenge strains were undetectable in inner organs.
As discussed for the wild type strain, further experiments with younger animals might
reveal different results.
The intranasal infection of mice with all different S. suis strains after acetic acid
predisposition resulted in severe rhinitis in at least 50% of each group (Table 5-6). To
avoid possible non-specific histopathological findings of the nose, rhinitis was
excluded from the general pathoscore ω and, therefore, differences in ω between
different strains are not due to rhinitis lesions (Table 5-5 and Table 5-6).
In all 4 groups at least 50% of mice were positive in the TNL for the respective
challenge strain. Quantification of bacteria obtained from TNL revealed no significant
difference between mice infected either with S. suis wild type strain 10 or its isogenic
sly-negative mutant strain 10Δsly (Table 5-7). Comparably, infection with either
10cpsΔEF or 10cpsΔEFΔsly showed no clear difference of the recovered bacterial
load of the upper respiratory tract, suggesting no deficiency in colonization of the
sly-negative strains 10Δsly and 10cpsΔEFΔsly.
In conclusion, an intranasal murine model for S. suis colonization and invasion was
established in this work. Severe purulent necrotizing pneumonia and rhinitis, rather
than meningitis, were common findings among infected mice. Furthermore,
preliminary results suggested attenuation in virulence of an isogenic sly-mutant. But
a suilysin-dependent phenotype with regard to early colonization was not recorded in
these experiments. However, this murine model is suitable to study invasion and
colonization of mucosal surfaces in mice in the future.

Results, part III Chapter 5
133
Material and Methods Bacterial strains and culture conditions. Streptococcus (S.) suis serotype 2 wild
type strain 10 was kindly provided by H. Smith (Lelystad, NL). This strain expresses
EF, MRP, SLY, FBPS and OFS. It has been used by different groups successfully for
mutagenesis and experimental intranasal infections of pigs (Baums et al., 2006;
Smith et al., 1999; Vecht et al., 1997). Strain 10cpsΔEF has been generated by
insertional mutagenesis of the genes cps2E and cps2F involved in biosynthesis of
the capsule and has been demonstrated to be severely attenuated in virulence, most
likely due to increased opsonophagocytosis (Smith et al., 1999). The corresponding
suilysin deficient mutants of wild type strain 10 (designed 10Δsly) and 10cpsΔEF
(designed 10cpsΔEFΔsly) were constructed as described previously (Benga et al.,
2008; Seitz et al., submitted; cf. chapter 3). Streptococci were grown on Columbia
agar supplemented with 7% sheep blood (Oxoid, Wesel, Germany) overnight under
aerobic conditions at 37°C.
Preparation of infection culture. For infection of mice, streptococci were grown in
Todd-Hewitt broth (THB, Difco, Detroit, USA) overnight at 37°C under aerobic
conditions, adjusted to an optical density (OD600) of 0.02 in pre-warmed media the
next day and then grown to late exponential growth phase (OD600 0.8). Streptococci
were harvested by centrifugation, resuspended in sterile PBS (pH 7.4) and adjusted
to the final concentration of 5 x 1011 CFU/ml for intranasal infection. Inoculum
concentrations were verified by platting 10-fold serial dilutions on Colombia agar
plates with 7% sheep blood after infection.
Intranasal infection of mice. Four (expt. 1 to 4) or six (expt. 5) week old specific-
pathogen-free female mice of the outbreed strain Crl:CD1 (ICR) were obtained from
Charles River Laboratories (Sulzfeld, Germany). Mice of the Staphylococcus (St.)
aureus free outbred strain Hsd:ICR (CD1®) were purchased from Harlan Laboratories
(AN Venray, The Netherlands). Animals were randomly divided into groups
consisting of five to six animals each. Mice were allowed to acclimate for one week

Results, part III Chapter 5
134
and cared for in accordance with the principles outlined in the European Convention
for the Protection of Vertebrate Animal Used for Experimental and Other Scientific
Purposes [European Treaty Series, no. 123:
http://conventions.coe.int/treaty/EN/Menuprincipal.htm; permit no.33.9-42502-04-
08/1589]. Before infection mice were anesthetized via inhalation of isofluran (IsoFlo®,
Albrecht, Germany). In experiment 2 to 5 mice were pre-treated with 12.5 µl 1%
acetic acid [pH 4.0] placed in each nostril 1 h prior intranasal infection. After a
controlled recovery phase and re-anaesthesia per isofluran inhalation, mice were
infected with 5 x 109 CFU of either S. suis wild type strain 10, 10Δsly, 10cpsΔEF or
10cpsΔEFΔsly. The dose was applied in two drops of 12.5 µl volume placed in front
of the nostrils.
Clinical score. Animals were clinically scored every 8 h. The health status was rated
using a clinical score sheet (Table 5-1), including weight development, clinical signs
of general sickness (rough coat, rapid breathing, dehydration), clinical signs
indicating meningitis (apathy, apraxia), and septicaemia (swollen eyes, depression),
developed on the basis of mouse clinical monitoring score described by the
Research office of the Australian National University
[http://www.anu.edu.au/ro/ORI/Animal/ AEEC001_MouseMonitoringSOP.doc]. A
cumulative score of 3 to 4 indicated mild clinical symptoms, a score of 5 to 6
moderate clinical symptoms and a score greater than 6 severe clinical symptoms, in
particular persistent anorexia, apathy, and/or neural disorder. Mice with a cumulative
score equal or greater than 3 were classified as diseased (calculation of morbidity).
In the case of severe weight loss (> 20%) and/or enduring severe clinical signs, mice
were euthanized for reasons of animal welfare by inhalation of CO2 and cervical
dislocation.
Histological screening. Immediately after euthanasia, necropsy was conducted and
the organs were aseptically removed and split for histological and bacteriological
screenings, including spleen, liver, kidney, heart, lung, brain, spinal cord, and nose.
The histological screenings were carried out as blind experiments. Findings were

Results, part III Chapter 5
135
scored as described for piglets (Baums et al., 2006). In contrast to piglets, in addition
to fibrinosuppurative lesions, purulent necrotizing lesions associated with the
challenge strain were scored as well (see results). Rhinitis was not included in the
general score ω designed to reflect lesions caused by invading streptococci.
Reisolation of S. suis strains from tissue and tracheo-nasal lavage (TNL). One
half of each organ was suspended in 5 ml PBS [pH 7.4] and weighed. All organs
were homogenized with an Ultra Turrax (IKA, Staufen, Germany). Ten-fold serial
dilutions of samples were plated on blood agar plates. Colony forming units (CFU)
were counted the next day after incubation at 37°C for 24 h and CFU per mg organ
was determined.
For sampling TNL the trachea was opened and a retrograde irrigation of the nasal
cavity with 300 µl PBS was collected. Number of typical α-haemolytic streptococci
per µl TNL was determined by serial plating on blood agar. Isolated α-haemolytic
streptococci were investigated in a S. suis multiplex PCR for the detection of mrp,
epf, sly, arcA, gdh, cps1, cps2, cps7, and cps9 (Silva et al., 2006). Isolates received
from mice challenged either with strain 10Δsly, 10cpsΔEF or 10cpsΔsly were
additionally tested in a cps2E-specific PCR (with oligonucleotide primers cps2Efor
(TTTCGCACTTTCAAGACGTG) and cps2Erev (GGACGGGTACCGACTAGACTC)
and sly-specific PCR using primers slyAgefor (TGTACCGGTGATTCCAAACAAGA
TATTAA) and slyAge3new (TTAACCGGTTACTCTATCACCTCATCCG). Based on
CFU per mg organ or per µl TNL, bacterial loads were classified as mild (+; <100),
moderate (++; >100 - <1000) or severe (+++; >1000).

Results, part III Chapter 5
136
Acknowledgements We thank Hilde Smith (DLO-Institute for Animal Science and Health, The
Netherlands) for providing strain 10 and the non-encapsulated isogenic mutant
10cpsΔEF. Oliver Goldmann (Helmholtz Centre for Infection Research,
Braunschweig, Germany) kindly introduced us to intranasal application techniques
and anaesthesia of mice.
This study was financially supported by the Deutsche Forschungsgemeinschaft
(DFG), Bonn, Germany (SFB587).

Results, part III Chapter 5
137
References Allen, A. G., Bolitho, S., Lindsay, H., Khan, S., Bryant, C., Norton, P., Ward, P., Leigh, J., Morgan, J., Riches, H., Eastty, S., and Maskell, D. (2001) Generation and characterization of a defined mutant of Streptococcus suis lacking suilysin. Infect Immun 69: 2732-2735.
Arends, J. P., Hartwig, N., Rudolphy, M., and Zanen, H. C. (1984) Carrier Rate of Streptococcus Suis Capsular Type-2 in Palatine Tonsils of Slaughtered Pigs. Journal of Clinical Microbiology 20: 945-947.
Baele, M., Chiers, K., Devriese, L. A., Smith, H. E., Wisselink, H. J., Vaneechoutte, M., and Haesebrouck, F. (2001) The gram-positive tonsillar and nasal flora of piglets before and after weaning. J Appl Microbiol 91: 997-1003.
Baums, C. G., Kaim, U., Fulde, M., Ramachandran, G., Goethe, R., and Valentin-Weigand, P. (2006) Identification of a novel virulence determinant with serum opacification activity in Streptococcus suis. Infect Immun 74: 6154-6162.
Baums, C. G. and Valentin-Weigand, P. (2009) Surface-associated and secreted factors of Streptococcus suis in epidemiology, pathogenesis and vaccine development. Anim Health Res Rev 10: 65-83.
Baums, C. G., Verkuhlen, G. J., Rehm, T., Silva, L. M., Beyerbach, M., Pohlmeyer, K., and Valentin- Weigand, P. (2007) Prevalence of Streptococcus suis genotypes in wild boars of Northwestern Germany. Appl Environ Microbiol 73: 711-717.
Benga, L., Fulde, M., Neis, C., Goethe, R., and Valentin-Weigand, P. (2008) Polysaccharide capsule and suilysin contribute to extracellular survival of Streptococcus suis co-cultivated with primary porcine phagocytes. Vet Microbiol 132: 211-219.
Charland, N., Harel, J., Kobisch, M., Lacasse, S., and Gottschalk, M. (1998) Streptococcus suis serotype 2 mutants deficient in capsular expression 9. Microbiology-Sgm 144: 325-332.
Clifton-Hadley, F. A. and Alexander, T. J. (12-7-1980) The carrier site and carrier rate of Streptococcus suis type II in pigs. Vet Rec 107: 40-41.
Dominguez-Punaro, M. C., Segura, M., Plante, M. M., Lacouture, S., Rivest, S., and Gottschalk, M. (2007) Streptococcus suis serotype 2, an important swine and human pathogen, induces strong systemic and cerebral inflammatory responses in a mouse model of infection. J Immunol 179: 1842-1854.
Gottschalk, M., Segura, M., and Xu, J. (2007) Streptococcus suis infections in humans: the Chinese experience and the situation in North America. Anim Health Res Rev 8: 29-45.
Higgins,R. and Gottschalk,M. (2005) Streptococcal diseases. In Diseases of Swine. pp. 769-783.
Lowe, B. A., Marsh, T. L., Isaacs-Cosgrove, N., Kirkwood, R. N., Kiupel, M., and Mulks, M. H. (2011) Microbial communities in the tonsils of healthy pigs. Vet Microbiol 147: 346-357.
Lun, S. C., Perez-Casal, J., Connor, W., and Willson, P. J. (2003) Role of suilysin in pathogenesis of Streptococcus suis capsular serotype 2. Microbial Pathogenesis 34: 27-37.

Results, part III Chapter 5
138
Ngo, T. H., Tran, T. B., Tran, T. T., Nguyen, V. D., Campbell, J., Pham, H. A., Huynh, H. T., Nguyen, V. V., Bryant, J. E., Tran, T. H., Farrar, J., and Schultsz, C. (2011) Slaughterhouse pigs are a major reservoir of Streptococcus suis serotype 2 capable of causing human infection in southern Vietnam. PLoS One 6: e17943.
O'Sullivan, T., Friendship, R., Blackwell, T., Pearl, D., McEwen, B., Carman, S., Slavic, D., and Dewey, C. (2011) Microbiological identification and analysis of swine tonsils collected from carcasses at slaughter. Can J Vet Res 75: 106-111.
Pallares, F. J., Halbur, P. G., Schmitt, C. S., Roth, J. A., Opriessnig, T., Thomas, P. J., Kinyon, J. M., Murphy, D., Frank, D. E., and Hoffman, L. J. (2003) Comparison of experimental models for Streptococcus suis infection of conventional pigs. Can J Vet Res 67: 225-228.
Seitz, M., Baums, CG, Gerhard, R, Just, I, Neis, C., Benga, L., Fulde, M., Nerlich, A, Rohde, M., Goethe, R., and Valentin-Weigand, P. Subcytolytic activity of suilysin promotes invasion of Streptococcus suis in HEp-2 epithelial cells by Rac-dependent activation of the actin cytoskeleton. Ref Type: submitted to Cellular Microbiology Silva, L. M., Baums, C. G., Rehm, T., Wisselink, H. J., Goethe, R., and Valentin-Weigand, P. (2006) Virulence-associated gene profiling of Streptococcus suis isolates by PCR. Vet Microbiol 115: 117-127.
Smith, H. E., Damman, M., van der Velde, J., Wagenaar, F., Wisselink, H. J., Stockhofe-Zurwieden, N., and Smits, M. A. (1999) Identification and characterization of the cps locus of Streptococcus suis serotype 2: the capsule protects against phagocytosis and is an important virulence factor. Infection and Immunity 67: 1750-1756.
Smith, H. E., Vecht, U., Wisselink, H. J., StockhofeZurwieden, N., Biermann, Y., and Smits, M. A. (1996) Mutants of Streptococcus suis types 1 and 2 impaired in expression of muramidase- released protein and extracellular protein induce disease in newborn germfree pigs. Infection and Immunity 64: 4409-4412.
Su, Y., Yao, W., Perez-Gutierrez, O. N., Smidt, H., and Zhu, W. Y. (2008) Changes in abundance of Lactobacillus spp. and Streptococcus suis in the stomach, jejunum and ileum of piglets after weaning. FEMS Microbiol Ecol 66: 546-555.
Tang, J., Wang, C., Feng, Y., Yang, W., Song, H., Chen, Z., Yu, H., Pan, X., Zhou, X., Wang, H., Wu, B., Wang, H., Zhao, H., Lin, Y., Yue, J., Wu, Z., He, X., Gao, F., Khan, A. H., Wang, J., Zhao, G. P., Wang, Y., Wang, X., Chen, Z., and Gao, G. F. (2006) Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med 3: e151.
Vecht, U., StockhofeZurwieden, N., Tetenburg, B. J., Wisselink, H. J., and Smith, H. E. (1997) Virulence of Streptococcus suis type 2 for mice and pigs appeared host-specific. Veterinary Microbiology 58: 53-60.
Wei, Z., Li, R., Zhang, A., He, H., Hua, Y., Xia, J., Cai, X., Chen, H., and Jin, M. (2009) Characterization of Streptococcus suis isolates from the diseased pigs in China between 2003 and 2007. Vet Microbiol 137: 196-201.
Williams, A. E. and Blakemore, W. F. (1990) Pathogenesis of meningitis caused by Streptococcus suis type 2. J Infect Dis 162: 474-481.
Williams, A. E., Blakemore, W. F., and Alexander, T. J. (1988) A murine model of Streptococcus suis type 2 meningitis in the pig. Res Vet Sci 45: 394-399.

Results, part III Chapter 5
139
Wisselink, H. J., Smith, H. E., Stockhofe-Zurwieden, N., Peperkamp, K., and Vecht, U. (2000) Distribution of capsular types and production of muramidase-released protein (MRP) and extracellular factor (EF) of Streptococcus suis strains isolated from diseased pigs in seven European countries. Vet Microbiol 74: 237-248.
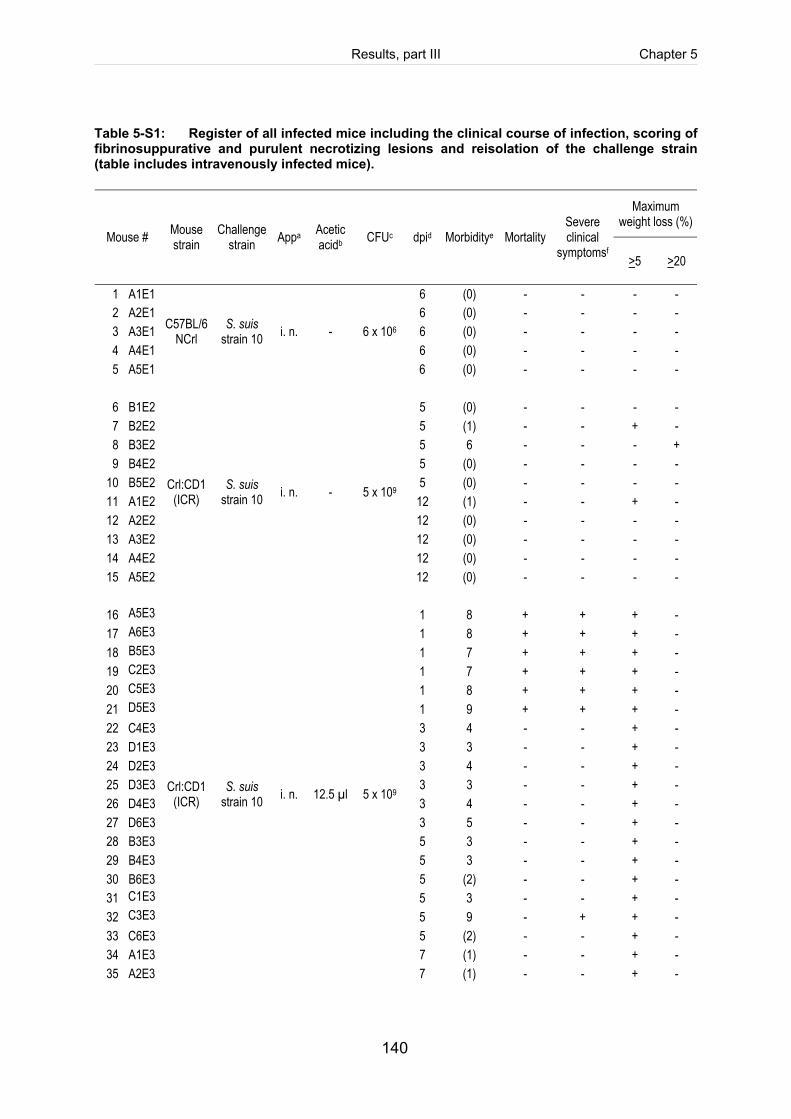
Results, part III Chapter 5
140
Table 5-S1: Register of all infected mice including the clinical course of infection, scoring of fibrinosuppurative and purulent necrotizing lesions and reisolation of the challenge strain (table includes intravenously infected mice).
Maximum weight loss (%)
Mouse # Mouse strain
Challenge strain Appa Acetic
acidb CFUc dpid Morbiditye Mortality Severe clinical
symptomsf >5 >20
1 A1E1 6 (0) - - - - 2 A2E1 6 (0) - - - - 3 A3E1 6 (0) - - - - 4 A4E1 6 (0) - - - - 5 A5E1
C57BL/6NCrl
S. suis strain 10 i. n. - 6 x 106
6 (0) - - - -
6 B1E2 5 (0) - - - - 7 B2E2 5 (1) - - + - 8 B3E2 5 6 - - - + 9 B4E2 5 (0) - - - -
10 B5E2 5 (0) - - - - 11 A1E2 12 (1) - - + - 12 A2E2 12 (0) - - - - 13 A3E2 12 (0) - - - - 14 A4E2 12 (0) - - - - 15 A5E2
Crl:CD1 (ICR)
S. suis strain 10 i. n. - 5 x 109
12 (0) - - - -
16 A5E3
1 8 + + + - 17 A6E3
1 8 + + + -
18 B5E3
1 7 + + + - 19 C2E3
1 7 + + + -
20 C5E3
1 8 + + + - 21 D5E3
1 9 + + + -
22 C4E3 3 4 - - + - 23 D1E3 3 3 - - + - 24 D2E3 3 4 - - + - 25 D3E3 3 3 - - + - 26 D4E3 3 4 - - + - 27 D6E3 3 5 - - + - 28 B3E3 5 3 - - + - 29 B4E3 5 3 - - + - 30 B6E3 5 (2) - - + - 31 C1E3
5 3 - - + -
32 C3E3
5 9 - + + - 33 C6E3 5 (2) - - + - 34 A1E3 7 (1) - - + - 35 A2E3
Crl:CD1 (ICR)
S. suis strain 10 i. n. 12.5 µl 5 x 109
7 (1) - - + -
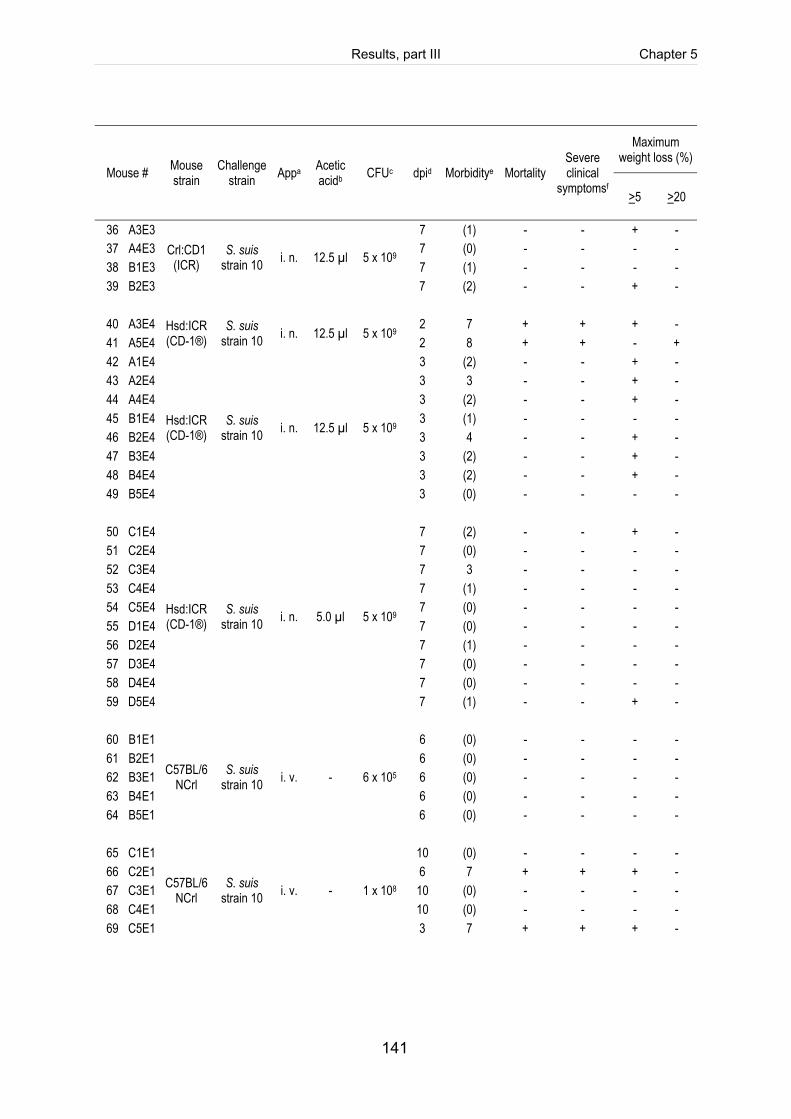
Results, part III Chapter 5
141
Maximum weight loss (%)
Mouse # Mouse strain
Challenge strain Appa Acetic
acidb CFUc dpid Morbiditye Mortality Severe clinical
symptomsf >5 >20
36 A3E3 7 (1) - - + - 37 A4E3 7 (0) - - - - 38 B1E3 7 (1) - - - - 39 B2E3
Crl:CD1 (ICR)
S. suis strain 10 i. n. 12.5 µl 5 x 109
7 (2) - - + -
40 A3E4 2 7 + + + - 41 A5E4
Hsd:ICR (CD-1®)
S. suis strain 10 i. n. 12.5 µl 5 x 109
2 8 + + - + 42 A1E4 3 (2) - - + - 43 A2E4 3 3 - - + - 44 A4E4 3 (2) - - + - 45 B1E4 3 (1) - - - - 46 B2E4 3 4 - - + - 47 B3E4 3 (2) - - + - 48 B4E4 3 (2) - - + - 49 B5E4
Hsd:ICR (CD-1®)
S. suis strain 10 i. n. 12.5 µl 5 x 109
3 (0) - - - -
50 C1E4 7 (2) - - + - 51 C2E4 7 (0) - - - - 52 C3E4 7 3 - - - - 53 C4E4 7 (1) - - - - 54 C5E4 7 (0) - - - - 55 D1E4 7 (0) - - - - 56 D2E4 7 (1) - - - - 57 D3E4 7 (0) - - - - 58 D4E4 7 (0) - - - - 59 D5E4
Hsd:ICR (CD-1®)
S. suis strain 10 i. n. 5.0 µl 5 x 109
7 (1) - - + -
60 B1E1 6 (0) - - - - 61 B2E1 6 (0) - - - - 62 B3E1 6 (0) - - - - 63 B4E1 6 (0) - - - - 64 B5E1
C57BL/6NCrl
S. suis strain 10 i. v. - 6 x 105
6 (0) - - - -
65 C1E1 10 (0) - - - - 66 C2E1 6 7 + + + - 67 C3E1 10 (0) - - - - 68 C4E1 10 (0) - - - - 69 C5E1
C57BL/6NCrl
S. suis strain 10 i. v. - 1 x 108
3 7 + + + -
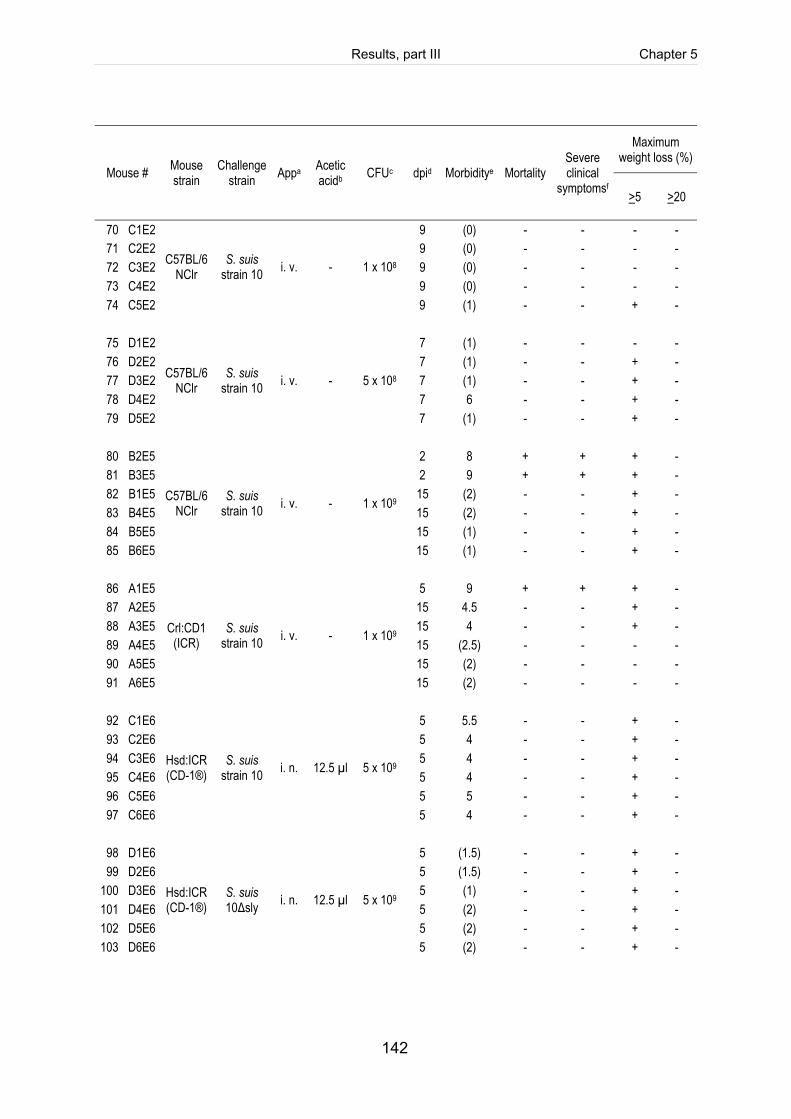
Results, part III Chapter 5
142
Maximum weight loss (%)
Mouse # Mouse strain
Challenge strain Appa Acetic
acidb CFUc dpid Morbiditye Mortality Severe clinical
symptomsf >5 >20
70 C1E2 9 (0) - - - - 71 C2E2 9 (0) - - - - 72 C3E2 9 (0) - - - - 73 C4E2 9 (0) - - - - 74 C5E2
C57BL/6NClr
S. suis strain 10 i. v. - 1 x 108
9 (1) - - + -
75 D1E2 7 (1) - - - - 76 D2E2 7 (1) - - + - 77 D3E2 7 (1) - - + - 78 D4E2 7 6 - - + - 79 D5E2
C57BL/6NClr
S. suis strain 10 i. v. - 5 x 108
7 (1) - - + -
80 B2E5 2 8 + + + - 81 B3E5 2 9 + + + - 82 B1E5 15 (2) - - + - 83 B4E5 15 (2) - - + - 84 B5E5 15 (1) - - + - 85 B6E5
C57BL/6NClr
S. suis strain 10 i. v. - 1 x 109
15 (1) - - + -
86 A1E5 5 9 + + + - 87 A2E5 15 4.5 - - + - 88 A3E5 15 4 - - + - 89 A4E5 15 (2.5) - - - - 90 A5E5 15 (2) - - - - 91 A6E5
Crl:CD1 (ICR)
S. suis strain 10 i. v. - 1 x 109
15 (2) - - - -
92 C1E6 5 5.5 - - + - 93 C2E6 5 4 - - + - 94 C3E6 5 4 - - + - 95 C4E6 5 4 - - + - 96 C5E6 5 5 - - + - 97 C6E6
Hsd:ICR (CD-1®)
S. suis strain 10 i. n. 12.5 µl 5 x 109
5 4 - - + -
98 D1E6 5 (1.5) - - + - 99 D2E6 5 (1.5) - - + -
100 D3E6 5 (1) - - + - 101 D4E6 5 (2) - - + - 102 D5E6 5 (2) - - + - 103 D6E6
Hsd:ICR (CD-1®)
S. suis 10Δsly i. n. 12.5 µl 5 x 109
5 (2) - - + -

Results, part III Chapter 5
143
Maximum weight loss (%)
Mouse # Mouse strain
Challenge strain Appa Acetic
acidb CFUc dpid Morbiditye Mortality Severe clinical
symptomsf >5 >20
104 A1E6 3 (2) - - - - 105 A2E6 3 3 - - + - 106 A3E6 3 3 - - + - 107 A4E6 3 9 - + - + 108 A5E6 3 (2) - - - - 109 A6E6
Hsd:ICR (CD-1®)
S. suis 10cpsΔE
F
i. n.
12.5 µl
5 x 109
3 (2.5) - - - -
110 B1E6 3 3 - - + - 111 B2E6 3 (1) - - + - 112 B3E6 3 (0) - - - - 113 B4E6 3 (2) - - + - 114 B5E6 3 (0) - - - - 115 B6E6
Hsd:ICR (CD-1®)
S. suis 10cpsΔE
FΔsly
i. n.
12.5 µl
5 x 109
3 (1) - - + -
a Intranasal (i. n.) or intravenous (i. v.) application of bacterial inoculum b Volume of 1% acetic acid applied to each nostril 1 h pre infectionem c Infection dose
d Days post infection, on which mice were sacrificed or killed for reasons of animal welfare e Mice with a cumulative clinical score greater >3 were regarded as diseased f In particular persistent anorexia, apathy, and/or neural disorder leading to a cumulative clinical score >6
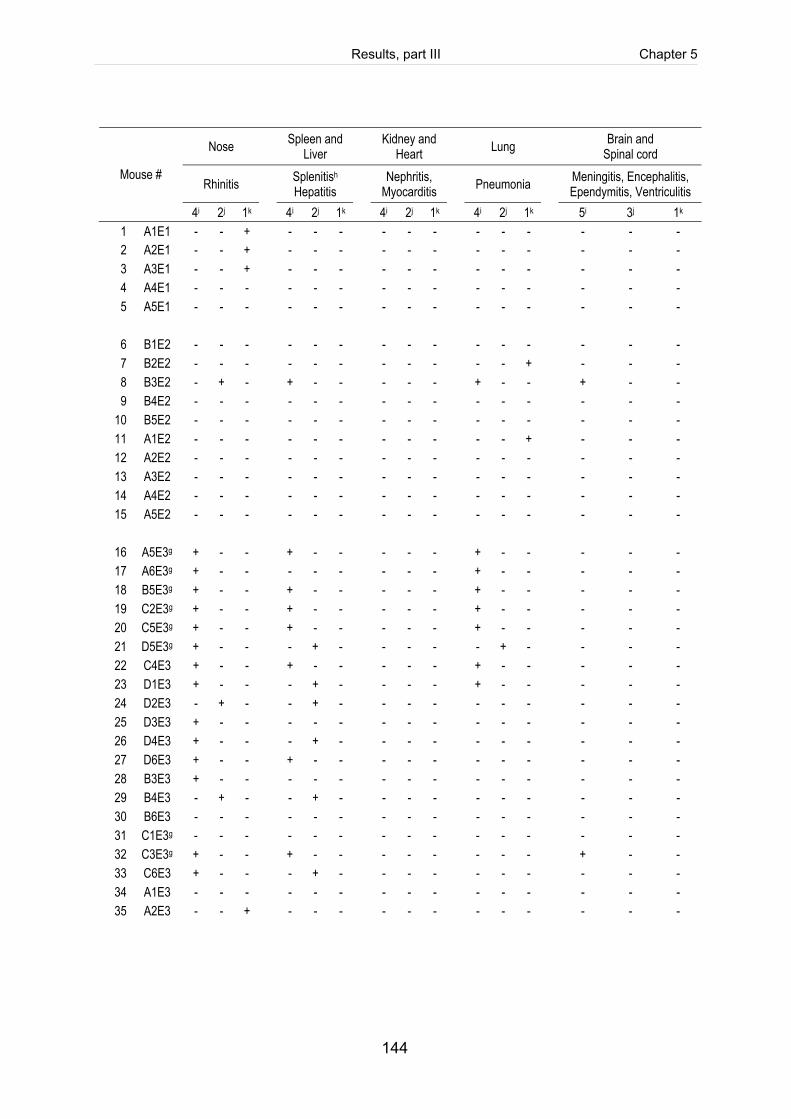
Results, part III Chapter 5
144
Nose Spleen and Liver Kidney and
Heart Lung Brain and Spinal cord
Rhinitis Splenitish Hepatitis Nephritis,
Myocarditis Pneumonia Meningitis, Encephalitis, Ependymitis, Ventriculitis
Mouse #
4i 2j 1k 4i 2j 1k 4i 2j 1k 4i 2j 1k 5i 3j 1k 1 A1E1 - - + - - - - - - - - - - - - 2 A2E1 - - + - - - - - - - - - - - - 3 A3E1 - - + - - - - - - - - - - - - 4 A4E1 - - - - - - - - - - - - - - - 5 A5E1 - - - - - - - - - - - - - - -
6 B1E2 - - - - - - - - - - - - - - - 7 B2E2 - - - - - - - - - - - + - - - 8 B3E2 - + - + - - - - - + - - + - - 9 B4E2 - - - - - - - - - - - - - - -
10 B5E2 - - - - - - - - - - - - - - - 11 A1E2 - - - - - - - - - - - + - - - 12 A2E2 - - - - - - - - - - - - - - - 13 A3E2 - - - - - - - - - - - - - - - 14 A4E2 - - - - - - - - - - - - - - - 15 A5E2 - - - - - - - - - - - - - - -
16 A5E3g + - - + - - - - - + - - - - - 17 A6E3g + - - - - - - - - + - - - - - 18 B5E3g + - - + - - - - - + - - - - - 19 C2E3g + - - + - - - - - + - - - - - 20 C5E3g + - - + - - - - - + - - - - - 21 D5E3g + - - - + - - - - - + - - - - 22 C4E3 + - - + - - - - - + - - - - - 23 D1E3 + - - - + - - - - + - - - - - 24 D2E3 - + - - + - - - - - - - - - - 25 D3E3 + - - - - - - - - - - - - - - 26 D4E3 + - - - + - - - - - - - - - - 27 D6E3 + - - + - - - - - - - - - - - 28 B3E3 + - - - - - - - - - - - - - - 29 B4E3 - + - - + - - - - - - - - - - 30 B6E3 - - - - - - - - - - - - - - - 31 C1E3g - - - - - - - - - - - - - - - 32 C3E3g + - - + - - - - - - - - + - - 33 C6E3 + - - - + - - - - - - - - - - 34 A1E3 - - - - - - - - - - - - - - - 35 A2E3 - - + - - - - - - - - - - - -

Results, part III Chapter 5
145
Nose Spleen and Liver Kidney and
Heart Lung Brain and Spinal cord
Rhinitis Splenitish Hepatitis Nephritis,
Myocarditis Pneumonia Meningitis, Encephalitis, Ependymitis, Ventriculitis
Mouse #
4i 2j 1k 4i 2j 1k 4i 2j 1k 4i 2j 1k 5i 3j 1k 36 A3E3 - - - - + - - - - - - - - - - 37 A4E3 - - - - - - - - - - - - - - - 38 B1E3 - - - - - - - - - - - - - - - 39 B2E3 + - - - - - - - - - - - - - -
40 A3E4 + - - - - + - - - - - - - - - 41 A5E4 + - - - - - - - - - - - - - - 42 A1E4 + - - - + - - - - + - - - - - 43 A2E4 - + - - - - - - - - - - - - - 44 A4E4 + - - - - + - - - - - - - - - 45 B1E4 + - - - - - - - - - - - - - - 46 B2E4 + - - - - + - - - + - - - - - 47 B3E4 + - - - - + - - - + - - - - - 48 B4E4 - + - - - + - - - - - - - - - 49 B5E4 - + - - - - - - - + - - - - -
50 C1E4 - - - - - + - - - - - - - - - 51 C2E4 - - - - - + - - - - - - - - - 52 C3E4 - + - - - - - - - - - - - - - 53 C4E4 - - + - - - - - - - - - - - - 54 C5E4 - - - - - - - - - - - - - - - 55 D1E4 - - - - - - - - - - - - - - - 56 D2E4 + - - - - - - - - - - - - - - 57 D3E4 - - - - - - - - - - - - - - - 58 D4E4 - - - - - - - - - - - - - - - 59 D5E4 - - - - - - - - - - - - - - -
60 B1E1 - - - - - - - - - - - - 61 B2E1 - - - - - - - - - - - - 62 B3E1 - - - - - - - - - - - - 63 B4E1 - - - - - - - - - - - - 64 B5E1 - - - - - - - - - + - -
65 C1E1 - - - - - - - - - - - - 66 C2E1 - - - + - - - - - - + - 67 C3E1 - - - - - - - - - - - - 68 C4E1 - - - - - - - - - - - - 69 C5E1 + - - - - - + - - + - -
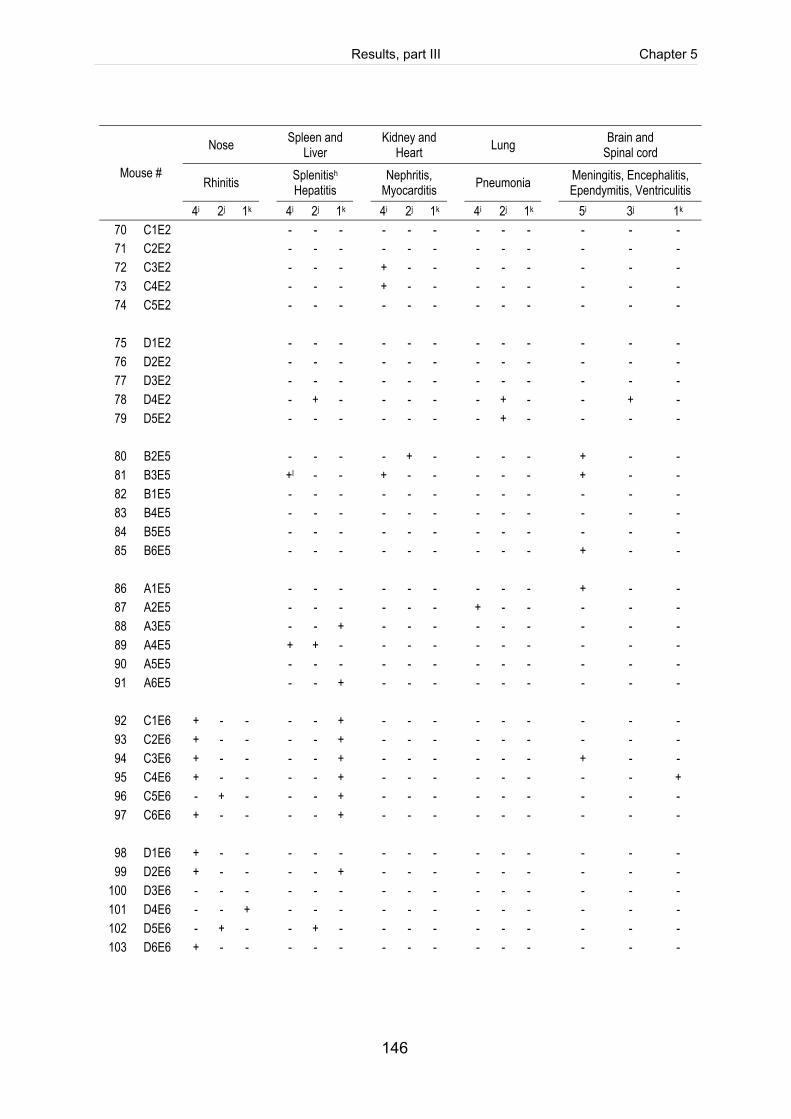
Results, part III Chapter 5
146
Nose Spleen and Liver Kidney and
Heart Lung Brain and Spinal cord
Rhinitis Splenitish Hepatitis Nephritis,
Myocarditis Pneumonia Meningitis, Encephalitis, Ependymitis, Ventriculitis
Mouse #
4i 2j 1k 4i 2j 1k 4i 2j 1k 4i 2j 1k 5i 3j 1k 70 C1E2 - - - - - - - - - - - - 71 C2E2 - - - - - - - - - - - - 72 C3E2 - - - + - - - - - - - - 73 C4E2 - - - + - - - - - - - - 74 C5E2 - - - - - - - - - - - -
75 D1E2 - - - - - - - - - - - - 76 D2E2 - - - - - - - - - - - - 77 D3E2 - - - - - - - - - - - - 78 D4E2 - + - - - - - + - - + - 79 D5E2 - - - - - - - + - - - -
80 B2E5 - - - - + - - - - + - - 81 B3E5 +l - - + - - - - - + - - 82 B1E5 - - - - - - - - - - - - 83 B4E5 - - - - - - - - - - - - 84 B5E5 - - - - - - - - - - - - 85 B6E5 - - - - - - - - - + - -
86 A1E5 - - - - - - - - - + - - 87 A2E5 - - - - - - + - - - - - 88 A3E5 - - + - - - - - - - - - 89 A4E5 + + - - - - - - - - - - 90 A5E5 - - - - - - - - - - - - 91 A6E5 - - + - - - - - - - - -
92 C1E6 + - - - - + - - - - - - - - - 93 C2E6 + - - - - + - - - - - - - - - 94 C3E6 + - - - - + - - - - - - + - - 95 C4E6 + - - - - + - - - - - - - - + 96 C5E6 - + - - - + - - - - - - - - - 97 C6E6 + - - - - + - - - - - - - - -
98 D1E6 + - - - - - - - - - - - - - - 99 D2E6 + - - - - + - - - - - - - - -
100 D3E6 - - - - - - - - - - - - - - - 101 D4E6 - - + - - - - - - - - - - - - 102 D5E6 - + - - + - - - - - - - - - - 103 D6E6 + - - - - - - - - - - - - - -
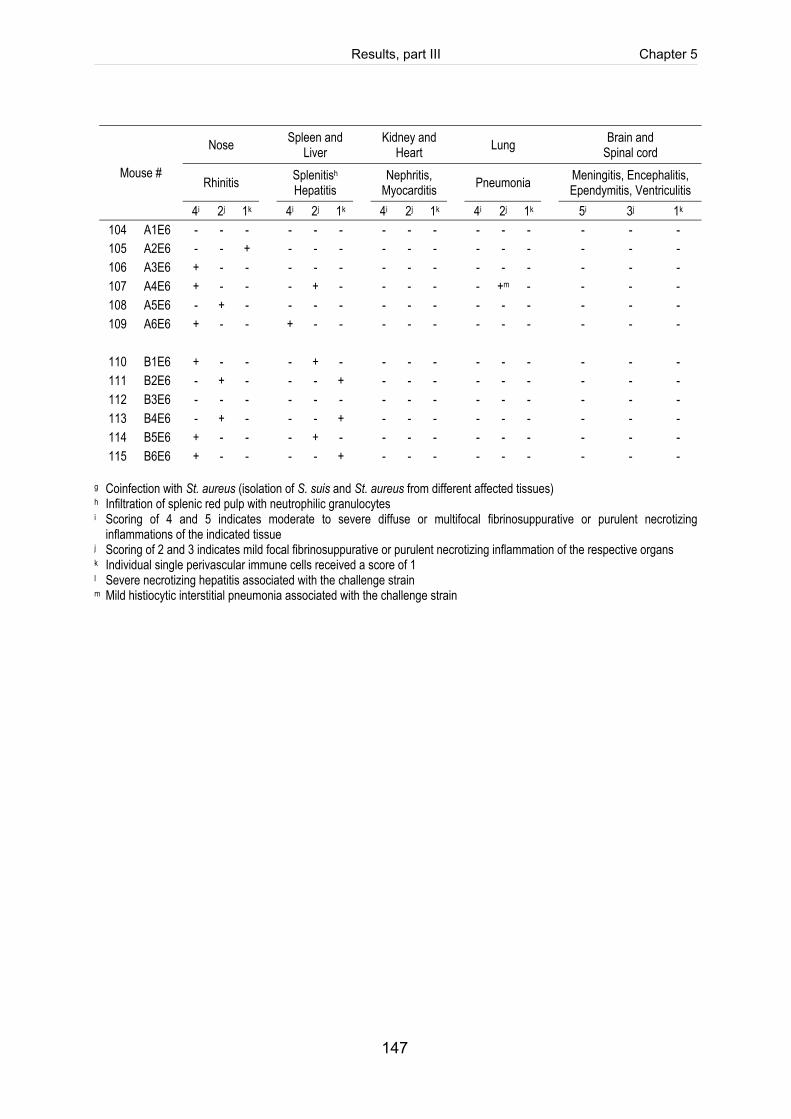
Results, part III Chapter 5
147
Nose Spleen and Liver Kidney and
Heart Lung Brain and Spinal cord
Rhinitis Splenitish Hepatitis Nephritis,
Myocarditis Pneumonia Meningitis, Encephalitis, Ependymitis, Ventriculitis
Mouse #
4i 2j 1k 4i 2j 1k 4i 2j 1k 4i 2j 1k 5i 3j 1k 104 A1E6 - - - - - - - - - - - - - - - 105 A2E6 - - + - - - - - - - - - - - - 106 A3E6 + - - - - - - - - - - - - - - 107 A4E6 + - - - + - - - - - +m - - - - 108 A5E6 - + - - - - - - - - - - - - - 109 A6E6 + - - + - - - - - - - - - - -
110 B1E6 + - - - + - - - - - - - - - - 111 B2E6 - + - - - + - - - - - - - - - 112 B3E6 - - - - - - - - - - - - - - - 113 B4E6 - + - - - + - - - - - - - - - 114 B5E6 + - - - + - - - - - - - - - - 115 B6E6 + - - - - + - - - - - - - - -
g Coinfection with St. aureus (isolation of S. suis and St. aureus from different affected tissues) h Infiltration of splenic red pulp with neutrophilic granulocytes i Scoring of 4 and 5 indicates moderate to severe diffuse or multifocal fibrinosuppurative or purulent necrotizing
inflammations of the indicated tissue j Scoring of 2 and 3 indicates mild focal fibrinosuppurative or purulent necrotizing inflammation of the respective organs
k Individual single perivascular immune cells received a score of 1 l Severe necrotizing hepatitis associated with the challenge strain m Mild histiocytic interstitial pneumonia associated with the challenge strain
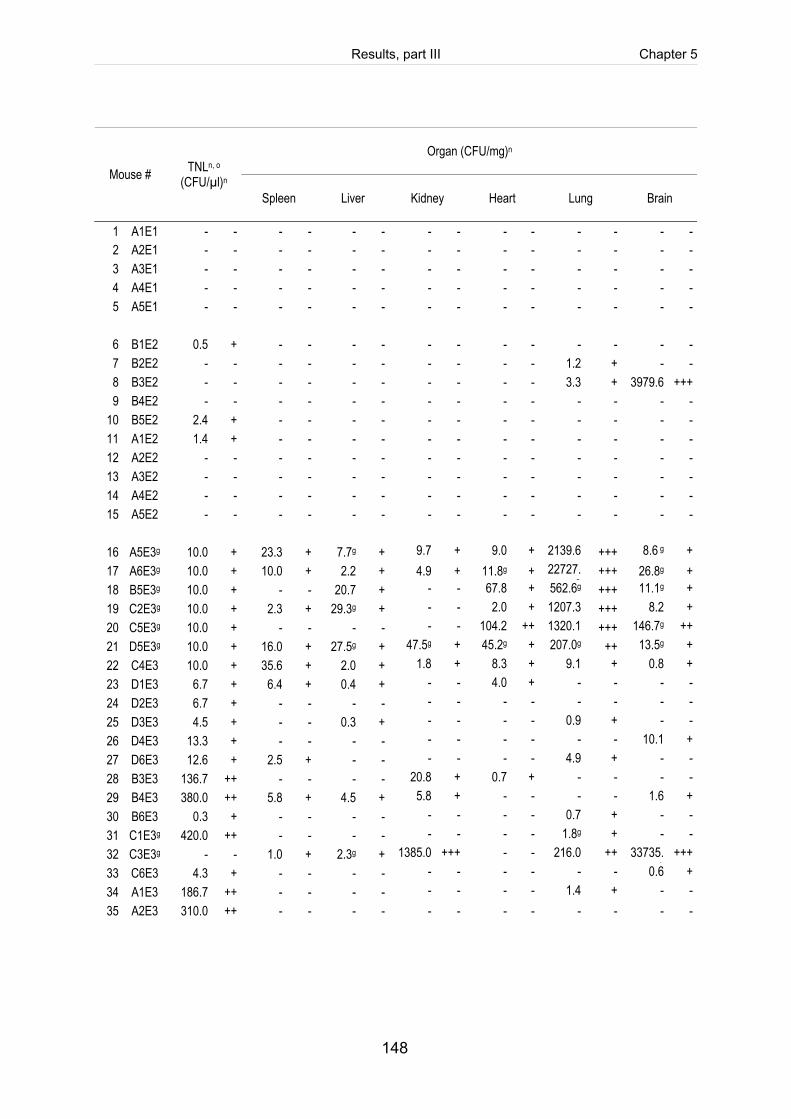
Results, part III Chapter 5
148
Organ (CFU/mg)n
Mouse # TNLn, o (CFU/µl)n
Spleen Liver Kidney Heart Lung Brain
1 A1E1 - - - - - - - - - - - - - - 2 A2E1 - - - - - - - - - - - - - - 3 A3E1 - - - - - - - - - - - - - - 4 A4E1 - - - - - - - - - - - - - - 5 A5E1 - - - - - - - - - - - - - -
6 B1E2 0.5 + - - - - - - - - - - - - 7 B2E2 - - - - - - - - - - 1.2 + - - 8 B3E2 - - - - - - - - - - 3.3 + 3979.6 +++ 9 B4E2 - - - - - - - - - - - - - -
10 B5E2 2.4 + - - - - - - - - - - - - 11 A1E2 1.4 + - - - - - - - - - - - - 12 A2E2 - - - - - - - - - - - - - - 13 A3E2 - - - - - - - - - - - - - - 14 A4E2 - - - - - - - - - - - - - - 15 A5E2 - - - - - - - - - - - - - -
16 A5E3g 10.0 + 23.3 + 7.7g + 9.7 + 9.0 + 2139.6
+++ 8.6 g +
17 A6E3g 10.0 + 10.0 + 2.2 + 4.9 + 11.8g + 22727.3
+++ 26.8g + 18 B5E3g 10.0 + - - 20.7 + - - 67.8 + 562.6g +++ 11.1g + 19 C2E3g 10.0 + 2.3 + 29.3g + - - 2.0 + 1207.3 +++ 8.2 + 20 C5E3g 10.0 + - - - - - - 104.2 ++ 1320.1
+++ 146.7g ++
21 D5E3g 10.0 + 16.0 + 27.5g + 47.5g + 45.2g + 207.0g ++ 13.5g + 22 C4E3 10.0 + 35.6 + 2.0 + 1.8 + 8.3 + 9.1 + 0.8 + 23 D1E3 6.7 + 6.4 + 0.4 + - - 4.0 + - - - - 24 D2E3 6.7 + - - - - - - - - - - - - 25 D3E3 4.5 + - - 0.3 + - - - - 0.9 + - - 26 D4E3 13.3 + - - - - - - - - - - 10.1 + 27 D6E3 12.6 + 2.5 + - - - - - - 4.9 + - - 28 B3E3 136.7 ++ - - - - 20.8 + 0.7 + - - - - 29 B4E3 380.0 ++ 5.8 + 4.5 + 5.8 + - - - - 1.6 + 30 B6E3 0.3 + - - - - - - - - 0.7 + - - 31 C1E3g 420.0 ++ - - - - - - - - 1.8g + - - 32 C3E3g - - 1.0 + 2.3g + 1385.0 +++ - - 216.0 ++ 33735.
9 +++
33 C6E3 4.3 + - - - - - - - - - - 0.6 + 34 A1E3 186.7 ++ - - - - - - - - 1.4 + - - 35 A2E3 310.0 ++ - - - - - - - - - - - -
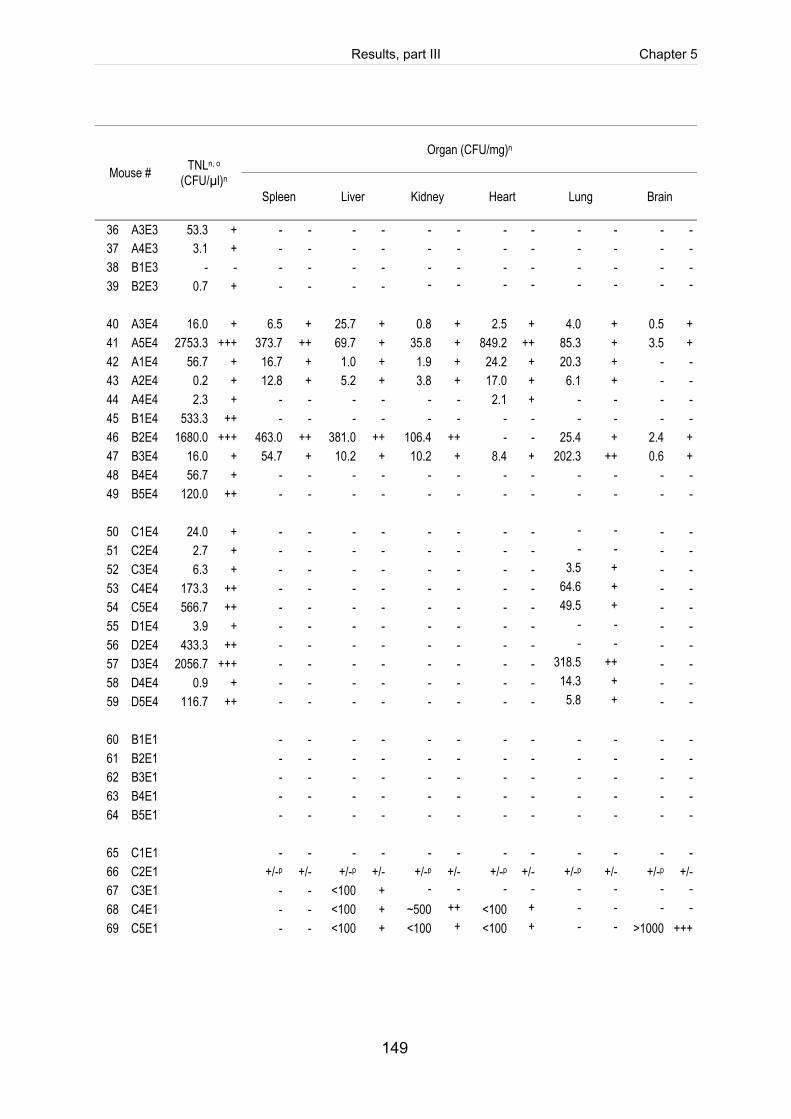
Results, part III Chapter 5
149
Organ (CFU/mg)n
Mouse # TNLn, o (CFU/µl)n
Spleen Liver Kidney Heart Lung Brain
36 A3E3 53.3 + - - - - - - - - - - - - 37 A4E3 3.1 + - - - - - - - - - - - - 38 B1E3 - - - - - - - - - - - - - - 39 B2E3 0.7 + - - - - - - - - - - - -
40 A3E4 16.0 + 6.5 + 25.7 + 0.8 + 2.5 + 4.0 + 0.5 + 41 A5E4 2753.3 +++ 373.7 ++ 69.7 + 35.8 + 849.2 ++ 85.3 + 3.5 + 42 A1E4 56.7 + 16.7 + 1.0 + 1.9 + 24.2 + 20.3 + - - 43 A2E4 0.2 + 12.8 + 5.2 + 3.8 + 17.0 + 6.1 + - - 44 A4E4 2.3 + - - - - - - 2.1 + - - - - 45 B1E4 533.3 ++ - - - - - - - - - - - - 46 B2E4 1680.0 +++ 463.0 ++ 381.0 ++ 106.4 ++ - - 25.4 + 2.4 + 47 B3E4 16.0 + 54.7 + 10.2 + 10.2 + 8.4 + 202.3 ++ 0.6 + 48 B4E4 56.7 + - - - - - - - - - - - - 49 B5E4 120.0 ++ - - - - - - - - - - - -
50 C1E4 24.0 + - - - - - - - - - - - - 51 C2E4 2.7 + - - - - - - - - - - - - 52 C3E4 6.3 + - - - - - - - - 3.5 + - - 53 C4E4 173.3 ++ - - - - - - - - 64.6 + - - 54 C5E4 566.7 ++ - - - - - - - - 49.5 + - - 55 D1E4 3.9 + - - - - - - - - - - - - 56 D2E4 433.3 ++ - - - - - - - - - - - - 57 D3E4 2056.7 +++ - - - - - - - - 318.5 ++ - - 58 D4E4 0.9 + - - - - - - - - 14.3 + - - 59 D5E4 116.7 ++ - - - - - - - - 5.8 + - -
60 B1E1 - - - - - - - - - - - - 61 B2E1 - - - - - - - - - - - - 62 B3E1 - - - - - - - - - - - - 63 B4E1 - - - - - - - - - - - - 64 B5E1 - - - - - - - - - - - -
65 C1E1 - - - - - - - - - - - - 66 C2E1 +/-p +/- +/-p +/- +/-p +/- +/-p +/- +/-p +/- +/-p +/- 67 C3E1 - - <100 + - - - - - - - - 68 C4E1 - - <100 + ~500 ++ <100 + - - - - 69 C5E1 - - <100 + <100 + <100 + - - >1000 +++
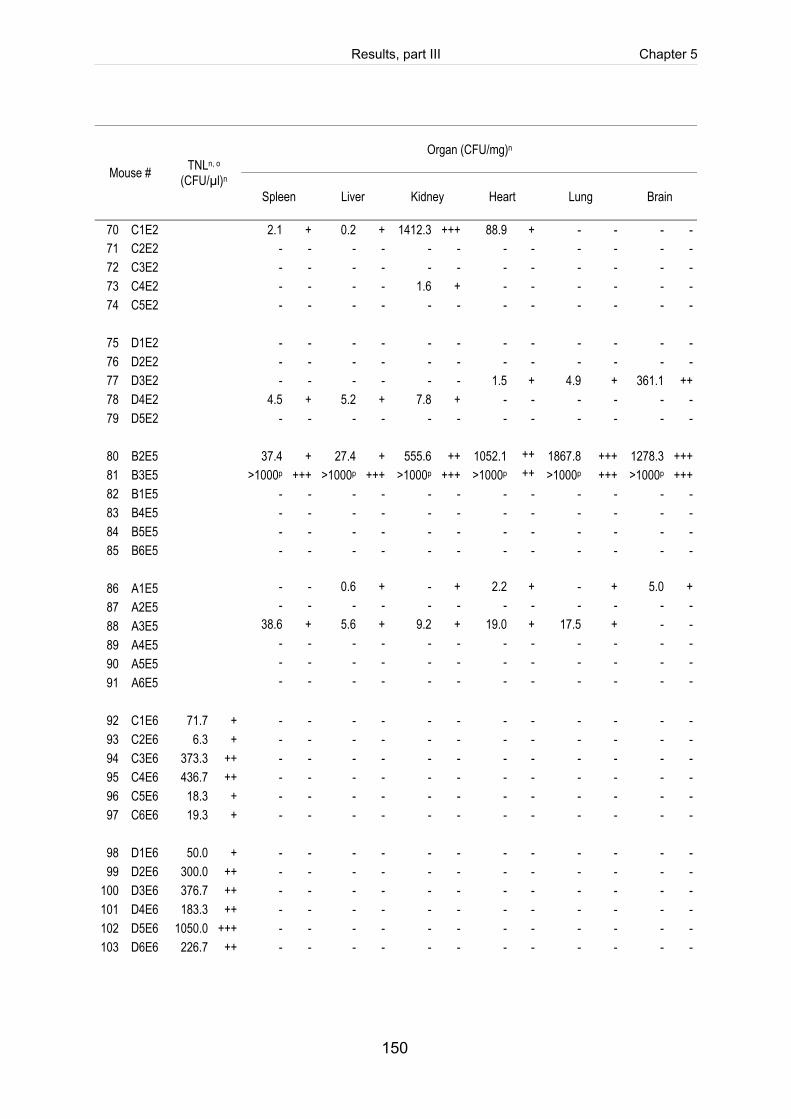
Results, part III Chapter 5
150
Organ (CFU/mg)n
Mouse # TNLn, o (CFU/µl)n
Spleen Liver Kidney Heart Lung Brain
70 C1E2 2.1 + 0.2 + 1412.3 +++ 88.9 + - - - - 71 C2E2 - - - - - - - - - - - - 72 C3E2 - - - - - - - - - - - - 73 C4E2 - - - - 1.6 + - - - - - - 74 C5E2 - - - - - - - - - - - -
75 D1E2 - - - - - - - - - - - - 76 D2E2 - - - - - - - - - - - - 77 D3E2 - - - - - - 1.5 + 4.9 + 361.1 ++ 78 D4E2 4.5 + 5.2 + 7.8 + - - - - - - 79 D5E2 - - - - - - - - - - - -
80 B2E5 37.4 + 27.4 + 555.6 ++ 1052.1 ++
1867.8 +++ 1278.3 +++
81 B3E5 >1000p +++ >1000p +++ >1000p +++ >1000p ++
>1000p +++ >1000p +++ 82 B1E5 - - - - - - - - - - - - 83 B4E5 - - - - - - - - - - - - 84 B5E5 - - - - - - - - - - - - 85 B6E5 - - - - - - - - - - - -
86 A1E5 - - 0.6 + - + 2.2 + - + 5.0 + 87 A2E5 - - - - - - - - - - - - 88 A3E5 38.6 + 5.6 + 9.2 + 19.0 + 17.5 + - - 89 A4E5 - - - - - - - - - - - - 90 A5E5 - - - - - - - - - - - - 91 A6E5 - - - - - - - - - - - -
92 C1E6 71.7 + - - - - - - - - - - - - 93 C2E6 6.3 + - - - - - - - - - - - - 94 C3E6 373.3 ++ - - - - - - - - - - - - 95 C4E6 436.7 ++ - - - - - - - - - - - - 96 C5E6 18.3 + - - - - - - - - - - - - 97 C6E6 19.3 + - - - - - - - - - - - -
98 D1E6 50.0 + - - - - - - - - - - - - 99 D2E6 300.0 ++ - - - - - - - - - - - -
100 D3E6 376.7 ++ - - - - - - - - - - - - 101 D4E6 183.3 ++ - - - - - - - - - - - - 102 D5E6 1050.0 +++ - - - - - - - - - - - - 103 D6E6 226.7 ++ - - - - - - - - - - - -
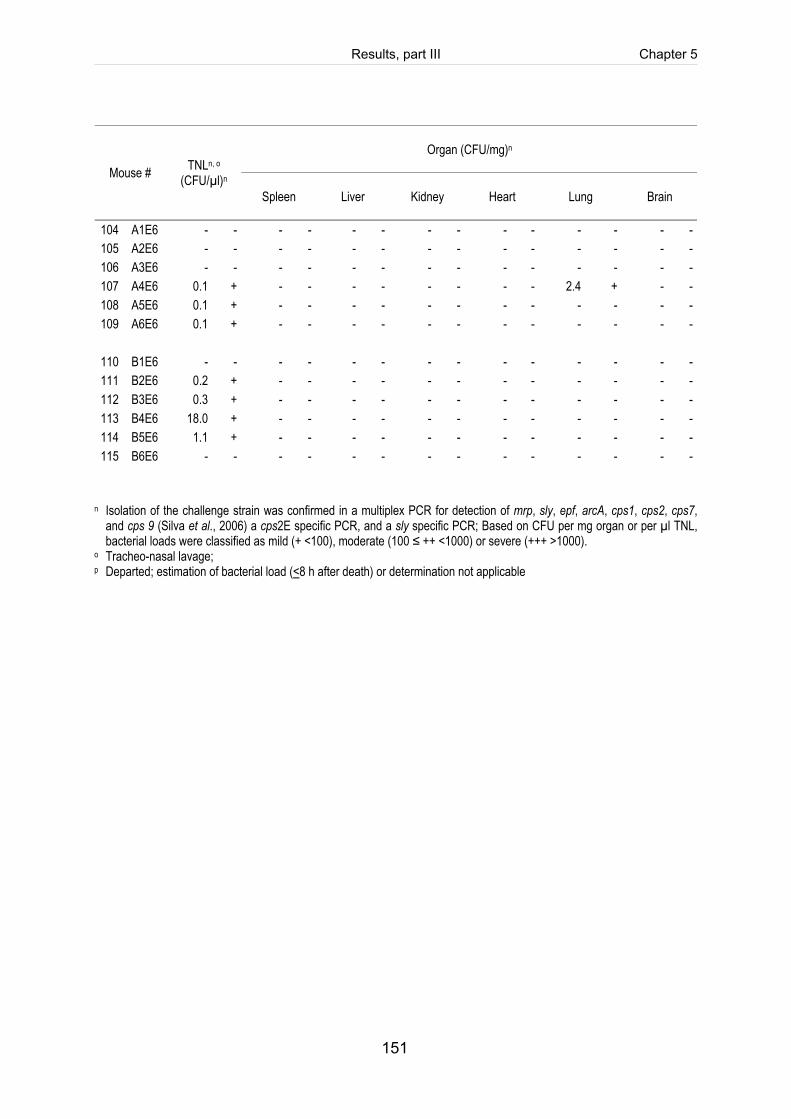
Results, part III Chapter 5
151
Organ (CFU/mg)n
Mouse # TNLn, o (CFU/µl)n
Spleen Liver Kidney Heart Lung Brain
104 A1E6 - - - - - - - - - - - - - - 105 A2E6 - - - - - - - - - - - - - - 106 A3E6 - - - - - - - - - - - - - - 107 A4E6 0.1 + - - - - - - - - 2.4 + - - 108 A5E6 0.1 + - - - - - - - - - - - - 109 A6E6 0.1 + - - - - - - - - - - - -
110 B1E6 - - - - - - - - - - - - - - 111 B2E6 0.2 + - - - - - - - - - - - - 112 B3E6 0.3 + - - - - - - - - - - - - 113 B4E6 18.0 + - - - - - - - - - - - - 114 B5E6 1.1 + - - - - - - - - - - - - 115 B6E6 - - - - - - - - - - - - - -
n Isolation of the challenge strain was confirmed in a multiplex PCR for detection of mrp, sly, epf, arcA, cps1, cps2, cps7,
and cps 9 (Silva et al., 2006) a cps2E specific PCR, and a sly specific PCR; Based on CFU per mg organ or per µl TNL, bacterial loads were classified as mild (+ <100), moderate (100 ≤ ++ <1000) or severe (+++ >1000).
o Tracheo-nasal lavage; p Departed; estimation of bacterial load (<8 h after death) or determination not applicable


Chapter 6
General discussion


General discussion Chapter 6
155
General discussion S. suis is considered as a major problem worldwide due to high economical losses in
swine husbandry. To reduce infection threat preventive antimicrobial medication and
early treatment with antibiotics in the case of clinical signs are the only treatment
options. Moreover, most currently available vaccines only provide homologous
protection. Therefore, understanding of pathogenesis and identification of bacterial
virulence (-associated) factors involved in colonisation, persistence, invasion and
immune evasion are urgently needed for better control and prevention measures.
Suilysin, the haemolysin of S. suis, has been identified many years ago (Jacobs et
al., 1994), but is still controversially discussed as a virulence factor of S. suis. In vivo
experiments using suilysin knock-out mutants demonstrated attenuation in virulence
in mice, but only slight attenuation was seen in piglets (Allen et al., 2001; Lun et al.,
2003). Furthermore, immunisation studies with a vaccine containing purified suilysin
completely protected mice against challenge with a highly virulent S. suis serotype 2
strain, but failed to fully protected piglets (Jacobs et al., 1994; Jacobs et al., 1996).
Nevertheless, the sly gene is prevalent in almost all highly virulent S. suis strains
isolated from diseased pigs and humans in Europe and Asia. Taken together suilysin
is currently considered to contribute to virulence, though the toxin seems not to be
essential for pathogenesis. At present, it remains unclear which biological activities
are relevant for virulence of S. suis. Suilysin belongs to the large family of
cholesterol-dependent pore-forming cytolysins (CDC), thus membrane damage
followed by cell lysis (Figure 6-1A) is the obvious but possibly not the only biological
effect which contributes to pathogenesis (Billigton et al., 2000).
As described in the general introduction, pathogenesis of S. suis includes three
crucial steps. Briefly, colonisation and invasion, dissemination within the bloodstream
and finally translocation into target tissues (Gottschalk and Segura, 2000). In
chapter 3 biological activities of suilysin which play a role in early stages of S. suis
pathogenesis were investigated. The first barrier of defence after the intranasal route
of infection is the mucosal epithelium lining of the upper respiratory tract. Although
S. suis is considered an extracellular pathogen, interaction with epithelial cells to

General discussion Chapter 6
156
breach the mucosal barrier is a crucial step. It is largely unknown which bacterial
factors and host cell components are involved in this process. The present study
showed that suilysin promotes invasion of an unencapsulated S. suis serotype 2
strain in vitro in a trigger-like uptake mechanism associated with membrane ruffles
(Figure 3-1). The polysaccharide capsule is assumed to interfere with the ability of
S. suis to interact with host tissue (Benga et al., 2004; Vanier et al., 2004). Since its
main function is protection against phagocytosis after entering the bloodstream, it
has been proposed that the capsule is down-regulated during colonisation of the
mucosal epithelium to allow adherence and invasion (Gottschalk and Segura, 2000;
Okamoto et al., 2004; Willenborg et al., 2011). In accordance, unencapsulated
S. suis stains showed a higher adherence and invasion rate in comparison to the well
encapsulated wild type strains (Benga et al., 2004). Furthermore, we found evidence
that the signal protein kinase Rac, belonging to the small family of Rho-GTPases, is
a key regulator in suilysin promoted uptake of S. suis (Figure 3-2). Recruitment of the
Rho family guanosine triphosphatases, namely Rac1, Rho, Cdc42, for bacterial
internalisation has been previously reported for several bacterial species (Agarwal
and Hammerschmidt, 2009; Burnham et al., 2007; Criss and Casanova, 2003; Finlay,
2005). Rho-GTPases can act as targets of bacterial toxins (Aktories et al., 2000) like
the cytotoxic necrotizing factors CNF1 and CNF2 from Escherichia coli (Schmidt and
Aktories, 2000) or the Salmonella outer protein E (Hardt et al., 1998). An interaction
of CDC with G-proteins has only been described for pneumolysin so far (Iliev et al.,
2007).
It still remains open whether these in vitro results reflect the in vivo situation of
S. suis pathogenesis. Therefore, an intranasal mouse infection model for colonisation
and invasion for S. suis was established (chapter 5). As asymptomatically colonized
carrier pigs play a key role in epidemiology of S. suis, a important step in
pathogenesis is sufficient colonisation of (respiratory) mucosal surfaces. Using an
intranasal instillation of a highly virulent S. suis serotype 2 strain after nasal
predisposition with acetic acid we were able to induce colonisation of the upper
respiratory tract (Table 5-4). We observed that invasive disease associated with
rhinitis, pneumonia and/or meningitis was only seen after pretreatment with acetic

General discussion Chapter 6
157
acid. This corresponds well to the situation of natural infection, where coinfections
with other pathogenic agents, such as PRRSV or Bordetella bronchiseptica, mucosal
irrigation or stress are predisposing factors for S. suis infections (Pallares et al.,
2003). In contrast to typical histopathological lesions for S. suis infection in swine,
which are characterized by fibrinosuppurative inflammation of the respective tissue
(Beineke et al., 2008; Williams and Blakemore, 1990), we observed purulent
necrotizing alteration of the respiratory apparatus (nose and lung) and the central
nervous system (CNS) in mice. Similar pathologies of the CNS were described for
mice after intraperitoneal challenge (Dominguez-Punaro et al., 2007; Vecht et al.,
1997). Presumably, induction of necrosis is directly linked to cytotoxic effects of
suilysin as observed for different cell types in vitro (Charland et al., 2000; Gottschalk
and Segura, 2000; Lalonde et al., 2000; Norton et al., 1999; Tenenbaum et al., 2005;
Vanier et al., 2004). Tenenbaum et al. (2006) found necrosis of porcine choroid
plexus epithelial cells after incubation with a sly-positive S. suis strain. A similar
distribution of S. suis and suilysin in affected brain tissues of experimental infected
pigs associated with massive infiltration of neutrophils and acute necrosis of neurons
was described by Zheng et al. (2009).
To further investigate the role of suilysin in early stages of S. suis pathogenesis in
vivo we applied this model to perform a comparative study using a highly virulent
serotype 2 strain, its isogenic sly-negative strain (10Δsly), as well as the respective
isogenic unencapsulated strains (10cpsΔEF and 10cpsΔEFΔsly). Preliminary results
suggested attenuation of sly-deficient strains indicated by reduced morbidity and less
histopathological changes (Table 5-5 and Table 5-6). In contrast, colonisation of the
respiratory epithelium by suilysin-negative strains was unaffected. Thus, suilysin
seems to be not critical for colonisation, but contributes to invasion of S. suis in vivo.
Nevertheless, the results are not conclusive because induction of invasive disease
such as meningitis or pneumonia failed in this trial except for two cases of meningitis
in the wild type infected group and rhinitis in all infected animals. For the closely
related pneumococci proven intranasal pneumonia and bacteraemia models in
mouse have been described previously (Berry et al., 1989; Medina, 2010; Saeland et
al., 2000). Intranasal challenge studies showed that pneumolysin deficient mutant

General discussion Chapter 6
158
strains were less virulent in mice indicated by a significant longer median survival
time, reduced apathetical alterations in the lung and diminished onset of bacteraemia
(Alexander et al., 1998; Berry et al., 1989; Jounblat et al., 2003). In contrast,
pneumolysin production was not required for successful nasopharyngeal colonization
(Rubins et al., 1998).
As described in chapter 3, we observed that suilysin effects did not require cytolytic
activities. Therefore, to further dissect functional domains involved in biological
activities of suilysin, we introduced point-mutations of the tryptophan-rich pore-
forming region (W461F) and the RGD-motif (SVD) within the sly gene (chapter 4).
Both mutations resulted in a loss of haemolytic and cytolytic activity (Figure 4-2). The
tryptophan-mutant had retained its ability to bind to the host-cell membrane and to
activate Rac. In contrast, an unmodified RGD-motif seems to be required for efficient
membrane binding (Figure 4-6) and more strikingly for activation of Rac1 (Figure
4-7). Therefore we hypothesized that the RGD-motif of suilysin interacts with
heterodimeric transmembrane cell surface receptors, namely α5β1 integrins, to
mediate suilysin-promoted activation of Rac1 and subsequent invasion of S. suis in
target cells. In addition, the proven cholesterol dependency of suilysin would be
explainable since integrins are most frequently expressed on cholesterol enriched
microdomains (lipid rafts) (Lafont and van der Goot, 2005). Nevertheless, it remains
hypothetical and the role of the first domain of suilysin and integrins in S. suis-host
cell interaction should be addressed in future studies.
Based on our results we propose the following model of suilysin-promoted invasion of
the upper respiratory tract by S. suis after colonisation of the mucosal surface as
illustrated by Figure 6-1B. Secreted suilysin binds to cholesterol enriched micro
domains (lipid rafts?) of the host cell membrane either via domain 4 and/or via an
interaction between α5β1 integrins and the RGD-motif of domain 1 (1). If applicable,
the prepore complex is formed and a downstream suilysin-mediated activation of
Rac1 (2) leads to induction of F-actin (3) and the formation of membrane ruffles.
Finally, S. suis is taken up by the cell (4). Suilysin-promoted invasion into host tissue
and intracellular persistence might be crucial for bacterial pathogenesis, since they
allow pathogens to escape the immune system for further dissemination and invasive
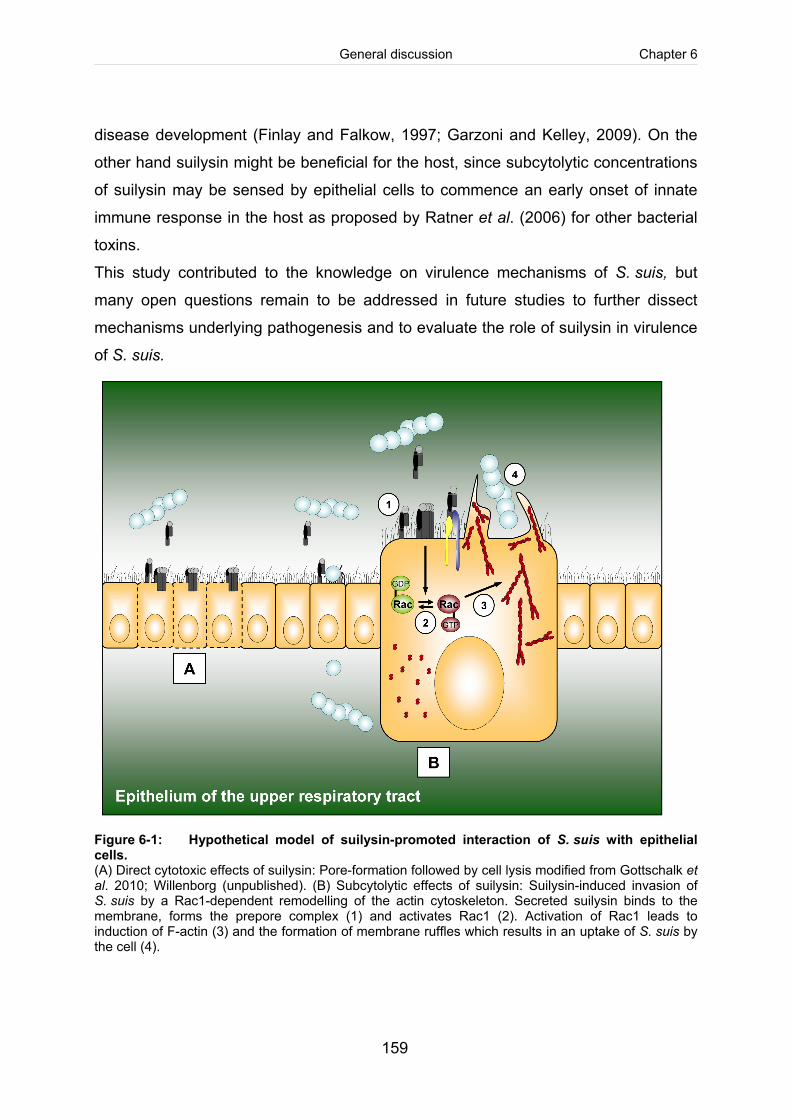
General discussion Chapter 6
159
disease development (Finlay and Falkow, 1997; Garzoni and Kelley, 2009). On the
other hand suilysin might be beneficial for the host, since subcytolytic concentrations
of suilysin may be sensed by epithelial cells to commence an early onset of innate
immune response in the host as proposed by Ratner et al. (2006) for other bacterial
toxins.
This study contributed to the knowledge on virulence mechanisms of S. suis, but
many open questions remain to be addressed in future studies to further dissect
mechanisms underlying pathogenesis and to evaluate the role of suilysin in virulence
of S. suis.
Figure 6-1: Hypothetical model of suilysin-promoted interaction of S. suis with epithelial cells. (A) Direct cytotoxic effects of suilysin: Pore-formation followed by cell lysis modified from Gottschalk et al. 2010; Willenborg (unpublished). (B) Subcytolytic effects of suilysin: Suilysin-induced invasion of S. suis by a Rac1-dependent remodelling of the actin cytoskeleton. Secreted suilysin binds to the membrane, forms the prepore complex (1) and activates Rac1 (2). Activation of Rac1 leads to induction of F-actin (3) and the formation of membrane ruffles which results in an uptake of S. suis by the cell (4).

General discussion Chapter 6
160
References Agarwal, V. and Hammerschmidt, S. (2009) Cdc42 and the phosphatidylinositol 3-kinase-Akt pathway are essential for PspC-mediated internalization of pneumococci by respiratory epithelial cells. J Biol Chem 284: 19427-19436.
Aktories, K., Schmidt, G., and Just, I. (2000) Rho GTPases as targets of bacterial protein toxins. Biol Chem 381: 421-426.
Alexander, J. E., Berry, A. M., Paton, J. C., Rubins, J. B., Andrew, P. W., and Mitchell, T. J. (1998) Amino acid changes affecting the activity of pneumolysin alter the behaviour of pneumococci in pneumonia. Microb Pathog 24: 167-174.
Allen, A. G., Bolitho, S., Lindsay, H., Khan, S., Bryant, C., Norton, P., Ward, P., Leigh, J., Morgan, J., Riches, H., Eastty, S., and Maskell, D. (2001) Generation and characterization of a defined mutant of Streptococcus suis lacking suilysin. Infect Immun 69: 2732-2735.
Beineke, A., Bennecke, K., Neis, C., Schroder, C., Waldmann, K. H., Baumgartner, W., Valentin- Weigand, P., and Baums, C. G. (2008) Comparative evaluation of virulence and pathology of Streptococcus suis serotypes 2 and 9 in experimentally infected growers. Vet Microbiol 128: 423-430.
Benga, L., Goethe, R., Rohde, M., and Valentin-Weigand, P. (2004) Non-encapsulated strains reveal novel insights in invasion and survival of Streptococcus suis in epithelial cells. Cellular Microbiology 6: 867-881.
Berry, A. M., Yother, J., Briles, D. E., Hansman, D., and Paton, J. C. (1989) Reduced virulence of a defined pneumolysin-negative mutant of Streptococcus pneumoniae. Infect Immun 57: 2037- 2042.
Billington, S. J., Jost, B. H., and Songer, J. G. (2000) Thiol-activated cytolysins: structure, function and role in pathogenesis. FEMS Microbiol Lett 182: 197-205.
Burnham, C. A., Shokoples, S. E., and Tyrrell, G. J. (2007) Rac1, RhoA, and Cdc42 participate in HeLa cell invasion by group B streptococcus. FEMS Microbiol Lett 272: 8-14.
Charland, N., Nizet, V., Rubens, C. E., Kim, K. S., Lacouture, S., and Gottschalk, M. (2000) Streptococcus suis serotype 2 interactions with human brain microvascular endothelial cells. Infect Immun 68: 637-643.
Clifton-Hadley, F. A. and Alexander, T. J. (1980) The carrier site and carrier rate of Streptococcus suis type II in pigs. Vet Rec 107: 40-41.
Criss, A. K. and Casanova, J. E. (2003) Coordinate regulation of Salmonella enterica serovar Typhimurium invasion of epithelial cells by the Arp2/3 complex and Rho GTPases. Infect Immun 71: 2885-2891.
Dominguez-Punaro, M. C., Segura, M., Plante, M. M., Lacouture, S., Rivest, S., and Gottschalk, M. (2007) Streptococcus suis serotype 2, an important swine and human pathogen, induces strong systemic and cerebral inflammatory responses in a mouse model of infection. J Immunol 179: 1842-1854.
Finlay, B. B. (2005) Bacterial virulence strategies that utilize Rho GTPases. Curr Top Microbiol Immunol 291: 1-10.

General discussion Chapter 6
161
Finlay, B. B. and Falkow, S. (1997) Common themes in microbial pathogenicity revisited. Microbiol Mol Biol Rev 61: 136-169.
Garzoni, C. and Kelley, W. L. (2009) Staphylococcus aureus: new evidence for intracellular persistence. Trends Microbiol 17: 59-65.
Gottschalk, M. and Segura, M. (2000) The pathogenesis of the meningitis caused by Streptococcus suis: the unresolved questions. Veterinary Microbiology 76: 259-272.
Hardt, W. D., Chen, L. M., Schuebel, K. E., Bustelo, X. R., and Galan, J. E. (1998) S. typhimurium encodes an activator of Rho GTPases that induces membrane ruffling and nuclear responses in host cells. Cell 93: 815-826.
Iliev, A. I., Djannatian, J. R., Nau, R., Mitchell, T. J., and Wouters, F. S. (2007) Cholesterol-dependent actin remodeling via RhoA and Rac1 activation by the Streptococcus pneumoniae toxin pneumolysin. Proc Natl Acad Sci U S A 104: 2897-2902.
Jacobs, A. A. C., Loeffen, P. L. W., vandenBerg, A. J. G., and Storm, P. K. (1994) Identification, Purification, and Characterization of A Thiol-Activated Hemolysin (Suilysin) of Streptococcus Suis. Infection and Immunity 62: 1742-1748.
Jacobs, A. A. C., vandenBerg, A. J. G., and Loeffen, P. L. W. (1996) Protection of experimentally infected pigs by suilysin, the thiol-activated haemolysin of Streptococcus suis. Veterinary Record 139: 225-228.
Jounblat, R., Kadioglu, A., Mitchell, T. J., and Andrew, P. W. (2003) Pneumococcal behavior and host responses during bronchopneumonia are affected differently by the cytolytic and complement- activating activities of pneumolysin. Infect Immun 71: 1813-1819.
Lafont, F. and van der Goot, F. G. (2005) Bacterial invasion via lipid rafts. Cell Microbiol 7: 613-620.
Lalonde, M., Segura, M., Lacouture, S., and Gottschalk, M. (2000) Interactions between Streptococcus suis serotype 2 and different epithelial cell lines. Microbiology 146 ( Pt 8): 1913- 1921.
Lun, S. C., Perez-Casal, J., Connor, W., and Willson, P. J. (2003) Role of suilysin in pathogenesis of Streptococcus suis capsular serotype 2. Microbial Pathogenesis 34: 27-37.
Medina, E. (2010) Murine model of pneumococcal pneumonia. Methods Mol Biol 602: 405-410.
Norton, P. M., Rolph, C., Ward, P. N., Bentley, R. W., and Leigh, J. A. (1999) Epithelial invasion and cell lysis by virulent strains of Streptococcus suis is enhanced by the presence of suilysin. Fems Immunology and Medical Microbiology 26: 25-35.
Okamoto, S., Kawabata, S., Terao, Y., Fujitaka, H., Okuno, Y., and Hamada, S. (2004) The Streptococcus pyogenes capsule is required for adhesion of bacteria to virus-infected alveolar epithelial cells and lethal bacterial-viral superinfection. Infect Immun 72: 6068-6075.
Pallares, F. J., Halbur, P. G., Schmitt, C. S., Roth, J. A., Opriessnig, T., Thomas, P. J., Kinyon, J. M., Murphy, D., Frank, D. E., and Hoffman, L. J. (2003) Comparison of experimental models for Streptococcus suis infection of conventional pigs. Can J Vet Res 67: 225-228.
Ratner, A. J., Hippe, K. R., Aguilar, J. L., Bender, M. H., Nelson, A. L., and Weiser, J. N. (2006) Epithelial cells are sensitive detectors of bacterial pore-forming toxins. J Biol Chem 281: 12994-12998.

General discussion Chapter 6
162
Rubins, J. B., Paddock, A. H., Charboneau, D., Berry, A. M., Paton, J. C., and Janoff, E. N. (1998) Pneumolysin in pneumococcal adherence and colonization. Microb Pathog 25: 337-342.
Saeland, E., Vidarsson, G., and Jonsdottir, I. (2000) Pneumococcal pneumonia and bacteremia model in mice for the analysis of protective antibodies. Microb Pathog 29: 81-91.
Schmidt, G. and Aktories, K. (2000) Rho GTPase-activating toxins: cytotoxic necrotizing factors and dermonecrotic toxin. Methods Enzymol 325: 125-136.
Tenenbaum, T., Adam, R., Eggelnpohler, I., Matalon, D., Seibt, A., GE, K. Novotny, Galla, H. J., and Schroten, H. (2005) Strain-dependent disruption of blood-cerebrospinal fluid barrier by Streptoccocus suis in vitro. FEMS Immunol Med Microbiol 44: 25-34.
Tenenbaum, T., Essmann, F., Adam, R., Seibt, A., Janicke, R. U., Novotny, G. E., Galla, H. J., and Schroten, H. (2006) Cell death, caspase activation, and HMGB1 release of porcine choroid plexus epithelial cells during Streptococcus suis infection in vitro. Brain Res 1100: 1-12.
Vanier, G., Segura, M., Friedl, P., Lacouture, S., and Gottschalk, M. (2004) Invasion of porcine brain microvascular endothelial cells by Streptococcus suis serotype 2. Infection and Immunity 72: 1441-1449.
Vecht, U., StockhofeZurwieden, N., Tetenburg, B. J., Wisselink, H. J., and Smith, H. E. (1997) Virulence of Streptococcus suis type 2 for mice and pigs appeared host-specific. Veterinary Microbiology 58: 53-60.
Willenborg, J., Fulde, M., de Greeff, A., Rohde, M., Smith, H. E., Valentin-Weigand, P., and Goethe, R. (2011) Role of glucose and CcpA in capsule expression and virulence of Streptococcus suis. Microbiology 157: 1823-1833.
Williams, A. E. and Blakemore, W. F. (1990) Pathogenesis of meningitis caused by Streptococcus suis type 2. J Infect Dis 162: 474-481.
Zheng, H., Punaro, M. C., Segura, M., Lachance, C., Rivest, S., Xu, J., Houde, M., and Gottschalk, M. (2011) Toll-like receptor 2 is partially involved in the activation of murine astrocytes by Streptococcus suis, an important zoonotic agent of meningitis. J Neuroimmunol 234: 71-83.

Chapter 7
Summary


Summary Chapter 7
165
Summary
Streptococcus (S.) suis is a swine pathogen of the upper respiratory tract causing
meningitis, arthritis and septicaemia. Due to high economical losses S. suis is
considered as an important pathogen in pig husbandry worldwide. Asymptomatically
colonized carrier-pigs are the most important source of infection. Moreover, S. suis is
a zoonotic agent associated with meningitis and the streptococcal toxic shock like
syndrome (STSS) in humans.
Suilysin is a secreted virulence-associated haemolysin of S. suis. It is present in
many S. suis strains and belongs to the family of Cholesterol-dependent pore-
forming cytolysins (CDC). At high concentrations these multifunctional toxins lyse
host cells. Additional biological activities of CDC such as the activation of host cells
or immune modulation, occur at subcytolytic concentrations. The role of suilysin in
pathogenesis of S. suis is still unclear.
The aim of this study was to analyse the role of suilysin in pathogen-host cell
interaction and to characterise the molecular mechanisms underling suilysin-
mediated biological effects.
Using an isogenic suilysin-negative mutant strain and the human respiratory
epithelial cell line HEp-2 a suilysin-dependent invasive phenotype of an
unencapsulated S. suis serotype 2 strain was identified in vitro. Subcytolytic
concentration of suilysin activated the small Rho-GTPase Rac1, which resulted in
formation of membrane ruffles and finally led to the uptake of S. suis by the cell.
Furthermore, point-mutation of the putative pore-forming region of the suilysin gene
abolished cytolytic activity, but activation of the G-protein Rac1 was unaffected. A
second putative functional region, a RGD-motif, was identified and also point-
mutated, resulting in loss of lytic activity, membrane binding and the ability to activate
Rac1. Thus, the RGD-motif is proposed to be involved in suilysin-mediated host cell-
activation. Integrins as possible interaction partners were identified on the surface of
HEp-2 cells, but further studies are needed to proof the precise role of integrins in
suilysin-mediated host cell-interaction.

Summary Chapter 7
166
After characterization of suilysin-dependent effects in vitro, an intranasal mouse
infection model for S. suis was established to investigate the role of suilysin in
colonisation and invasion of S. suis in vivo. After predisposition with acetic acid
followed by mucosal application of a S. suis serotype 2 strain we were able to induce
colonisation of the upper respiratory tract as well as invasive disease associated with
rhinitis, pneumonia and/or meningitis in mice. The suilysin-negative mutant strain
colonised the respiratory epithelium, but was slightly attenuated in virulence in
comparison to the wild type strain.
Taken together, these results show that subcytolytic activity of suilysin plays a role in
pathogen-host cell interaction. The established mouse model is suitable to study the
relevance of subcytolytic activities of suilysin in vivo in the future. The results of this
work improved our understanding of S. suis pathogenicity mechanisms.

Chapter 8
Zusammenfasung


Zusammenfassung Chapter 8
169
Zusammenfassung
Streptococcus (S.) suis ist ein Erreger im oberen Respirationstrakt des Schweins, der
vor allem Meningitiden, Arthritiden und Septikämien verursacht. Aufgrund von hohen
Verlusten in der Schweineproduktion sind S. suis-bedingte Erkrankungen weltweit
von großer Bedeutung. Epidemiologisch bedeutsam sind vor allem klinisch
inapparente Trägertiere, die als wichtigstes Reservoir für Neuinfektionen gelten.
Zudem besitzt S. suis ein zoonotisches Potential und spielt beim Menschen sowohl
eine wichtige Rolle als Meningitiserreger als auch als Verursacher des streptococcal
toxic shock like syndrom (STSS).
Zu den virulenzassoziierten Faktoren von S. suis gehört das Suilysin, welches ein
sezerniertes Hämolysin ist. Dieses tritt bei vielen Stämmen auf und zählt zur Familie
der cholesterol-dependent pore-forming cytolysins (CDC). Diese multifunktionalen
Toxine führen in hoher Konzentration zur Lyse von Wirtszellen. Neben diesen
lytischen Effekten sind auch biologische Aktivitäten im subzytoloytischen Bereich,
wie die Aktivierung von Wirtszellen oder immunmodulatorische Effekte, von
Bedeutung. Die genaue Rolle des Suilysins in der Pathogenese von S. suis ist bisher
nicht hinreichend geklärt.
Ziel dieser Arbeit war es daher, die Bedeutung von Suilysin in der Erreger-Wirtszell-
Interaktion und die molekularen Mechanismen Suilysin-vermittelter biologischer
Effekte näher zu untersuchen.
Mit Hilfe einer isogenen Suilysin-negativen Mutante konnte in in vitro
Untersuchungen an der humanen respiratorischen Epithelzelllinie HEp-2 ein Suilysin-
abhängiger invasiver Phänotyp eines unbekapselten S. suis-Serotyp-2-Stammes
gezeigt werden. Die durch subzytolytische Suilysin-Konzentration induzierte
Aktivierung der kleinen Rho-GTPase Rac1 vermittelte hierbei die Ausbildung von
membrane ruffles, wodurch S. suis in die Zelle aufgenommen wurde. Des Weiteren
konnte durch gezielte Punktmutagenese im Suilysin-Gen der putative Poren-bildende
Bereich ausgeschaltet werden, was zu einem Verlust der zytolytischen Aktivität
führte, nicht aber die G-Protein-aktivierende Eigenschaft des Suilysins beeinflusste.
Neben dem für die Porenbildung notwendigen Bereich im Suilysin-Gen, wurde eine

Zusammenfassung Chapter 8
170
weitere putative funktionale Domäne, ein RGD-Motiv, identifiziert und ebenfalls
punktmutiert. Der dadurch bedingte Verlust der zytolytischen Aktivität, der Bindung
des Suilysins an die Wirtszell-Membran und der Rac1-Aktivierung, weist auf eine
Beteiligung des RGD-Motivs an der Suilysin-vermittelten Interaktion mit Wirtszellen
hin. Als mögliche Interaktionspartner konnten Integrine auf der Zelloberfläche von
HEp-2-Zellen identifiziert werden, dennoch sind weitere funktionelle Studien
erforderlich, um die genaue Rolle von Integrinen in der Suilysin-vermittelten Zell-
Aktivierung aufzuklären.
Nach der Charakterisierung Suilysin-abhängiger Effekte im Zellkultursystem wurde
ein intranasales Mausinfektionsmodells für S. suis etabliert, um die Bedeutung des
Suilysins für die Kolonisation und Invasion von S. suis in vivo zu untersuchen. Nach
vorheriger Prädisposition mit Essigsäure war es möglich durch mukosale Applikation
des S. suis-Serotyp-2-Stammes eine Besiedlung des oberen Respirationstraktes
sowie das Auftreten von Rhinitis, Pneumonie und/oder Meningitis in der Maus zu
induzieren. Im Vergleich zum Wildtyp-Stamm erwies sich die Suilysin-negative
Mutante als geringgradig virulenzattenuiert, wohingegen keine Unterschiede in der
Kolonisation beobachtet wurden.
Zusammenfassend zeigen die Ergebnisse, dass Suilysin nicht nur zytolytische
Aktivitäten besitzt, sondern auch unter subzytolytischen Bedingungen eine
Bedeutung in der Erreger-Wirtszell-Interaktion hat. Die weitere Klärung der Relevanz
subzytolytischer Aktivitäten des Suilysins in vivo kann zukünftig durch das im
Rahmen dieses Projektes etablierten Mausmodels näher geklärt werden. Damit
wurde ein Beitrag zum besseren Verständnis von Pathogenitätsmechanismen von S.
suis geleistet.

Chapter 9
Literature


Literature Chapter 9
173
Literature
Agarwal, V. and Hammerschmidt, S. (2009) Cdc42 and the phosphatidylinositol 3-kinase-Akt pathway are essential for PspC-mediated internalization of pneumococci by respiratory epithelial cells. J Biol Chem 284: 19427-19436.
Aktories, K., Schmidt, G., and Just, I. (2000) Rho GTPases as targets of bacterial protein toxins. Biol Chem 381: 421-426.
Al Numani, D., Segura, M., Dore, M., and Gottschalk, M. (2003) Up-regulation of ICAM-1, CD11a/CD18 and CD11c/CD18 on human THP-1 monocytes stimulated by Streptococcus suis serotype 2. Clin Exp Immunol 133: 67-77.
Alcantara, R. B., Preheim, L. C., and Gentry-Nielsen, M. J. (2001) Pneumolysin-induced complement depletion during experimental pneumococcal bacteremia. Infect Immun 69: 3569-3575.
Alexander, J. E., Lock, R. A., Peeters, C. C., Poolman, J. T., Andrew, P. W., Mitchell, T. J., Hansman, D., and Paton, J. C. (1994) Immunization of mice with pneumolysin toxoid confers a significant degree of protection against at least nine serotypes of Streptococcus pneumoniae. Infect Immun 62: 5683-5688.
Allen, A. G., Bolitho, S., Lindsay, H., Khan, S., Bryant, C., Norton, P., Ward, P., Leigh, J., Morgan, J., Riches, H., Eastty, S., and Maskell, D. (2001) Generation and characterization of a defined mutant of Streptococcus suis lacking suilysin. Infect Immun 69: 2732-2735.
Alouf, J. E. (2000) Cholesterol-binding cytolytic protein toxins. Int J Med Microbiol 290: 351-356.
Amass, S. F., SanMiguel, P., and Clark, L. K. (1997) Demonstration of vertical transmission of Streptococcus suis in swine by genomic fingerprinting. J Clin Microbiol 35: 1595-1596.
Arends, J. P., Hartwig, N., Rudolphy, M., and Zanen, H. C. (1984) Carrier Rate of Streptococcus-Suis Capsular Type-2 in Palatine Tonsils of Slaughtered Pigs. Journal of Clinical Microbiology 20: 945-947.
Arends, J. P. and Zanen, H. C. (1988) Meningitis Caused by Streptococcus Suis in Humans. Reviews of Infectious Diseases 10: 131-137.
Baele, M., Chiers, K., Devriese, L. A., Smith, H. E., Wisselink, H. J., Vaneechoutte, M., and Haesebrouck, F. (2001) The gram-positive tonsillar and nasal flora of piglets before and after weaning. J Appl Microbiol 91: 997-1003.
Baums, C. G. and Valentin-Weigand, P. (2009) Surface-associated and secreted factors of Streptococcus suis in epidemiology, pathogenesis and vaccine development. Anim Health Res Rev 10: 65-83.
Baums, C. G., Verkuhlen, G. J., Rehm, T., Silva, L. M., Beyerbach, M., Pohlmeyer, K., and Valentin- Weigand, P. (2007) Prevalence of Streptococcus suis genotypes in wild boars of Northwestern Germany. Appl Environ Microbiol 73: 711-717.
Baums, C. G., Kaim, U., Fulde, M., Ramachandran, G., Goethe, R., and Valentin-Weigand, P. (2006) Identification of a novel virulence determinant with serum opacification activity in Streptococcus suis. Infect Immun 74: 6154-6162.

Literature Chapter 9
174
Beineke, A., Bennecke, K., Neis, C., Schroder, C., Waldmann, K. H., Baumgartner, W., Valentin- Weigand, P., and Baums, C. G. (2008) Comparative evaluation of virulence and pathology of Streptococcus suis serotypes 2 and 9 in experimentally infected growers. Vet Microbiol 128: 423-430.
Benga, L., Friedl, P., and Valentin-Weigand, P. (2005) Adherence of Streptococcus suis to porcine endothelial cells. J Vet Med B Infect Dis Vet Public Health 52: 392-395.
Benga, L., Fulde, M., Neis, C., Goethe, R., and Valentin-Weigand, P. (2008) Polysaccharide capsule and suilysin contribute to extracellular survival of Streptococcus suis co-cultivated with primary porcine phagocytes. Vet Microbiol 132: 211-219.
Benga, L., Goethe, R., Rohde, M., and Valentin-Weigand, P. (2004) Non-encapsulated strains reveal novel insights in invasion and survival of Streptococcus suis in epithelial cells. Cellular Microbiology 6: 867-881.
Berry, A. M., Yother, J., Briles, D. E., Hansman, D., and Paton, J. C. (1989) Reduced virulence of a defined pneumolysin-negative mutant of Streptococcus pneumoniae. Infect Immun 57: 2037- 2042.
Berthelot-Herault, F., Gottschalk, M., Labbe, A., Cariolet, R., and Kobisch, M. (2001) Experimental airborne transmission of Streptococcus suis capsular type 2 in pigs. Vet Microbiol 82: 69-80.
Berthelot-Herault, F., Morvan, H., Keribin, A. M., Gottschalk, M., and Kobisch, M. (2000) Production of Muraminidase-Released Protein (MRP), Extracellular Factor (EF) and Suilysin by field isolates of Streptococcus suis capsular types 2, 1/2, 9, 7 and 3 isolated from swine in France. Veterinary Research 31: 473-479.
Billington, S. J., Jost, B. H., Cuevas, W. A., Bright, K. R., and Songer, J. G. (1997) The Arcanobacterium (Actinomyces) pyogenes hemolysin, pyolysin, is a novel member of the thiol- activated cytolysin family. J Bacteriol 179: 6100-6106.
Billington, S. J., Jost, B. H., and Songer, J. G. (2000) Thiol-activated cytolysins: structure, function and role in pathogenesis. FEMS Microbiol Lett 182: 197-205.
Blume, V., Luque, I., Vela, A. I., Borge, C., Maldonado, A., Dominguez, L., Tarradas, C., and Fernandez-Garayzabal, J. F. (2009) Genetic and virulence-phenotype characterization of serotypes 2 and 9 of Streptococcus suis swine isolates. Int Microbiol 12: 161-166.
Bonifait, L., Grignon, L., and Grenier, D. (2008) Fibrinogen induces biofilm formation by Streptococcus suis and enhances its antibiotic resistance. Appl Environ Microbiol 74: 4969-4972.
Boulnois, G. J., Paton, J. C., Mitchell, T. J., and Andrew, P. W. (1991) Structure and function of pneumolysin, the multifunctional, thiol-activated toxin of Streptococcus pneumoniae. Mol Microbiol 5: 2611-2616.
Burnham, C. A., Shokoples, S. E., and Tyrrell, G. J. (2007) Rac1, RhoA, and Cdc42 participate in HeLa cell invasion by group B streptococcus. FEMS Microbiol Lett 272: 8-14.
Chabot-Roy, G., Willson, P., Segura, M., Lacouture, S., and Gottschalk, M. (2006) Phagocytosis and killing of Streptococcus suis by porcine neutrophils. Microb Pathog 41: 21-32.
Chanter, N., Jones, P. W., and Alexander, T. J. (1993) Meningitis in pigs caused by Streptococcus suis--a speculative review. Vet Microbiol 36: 39-55.

Literature Chapter 9
175
Charland, N., Harel, J., Kobisch, M., Lacasse, S., and Gottschalk, M. (1998) Streptococcus suis serotype 2 mutants deficient in capsular expression. Microbiology-Sgm 144: 325-332. Charland, N., Nizet, V., Rubens, C. E., Kim, K. S., Lacouture, S., and Gottschalk, M. (2000) Streptococcus suis serotype 2 interactions with human brain microvascular endothelial cells. Infect Immun 68: 637-643.
Chen, C., Tang, J., Dong, W., Wang, C., Feng, Y., Wang, J., Zheng, F., Pan, X., Liu, D., Li, M., Song, Y., Zhu, X., Sun, H., Feng, T., Guo, Z., Ju, A., Ge, J., Dong, Y., Sun, W., Jiang, Y., Wang, J., Yan, J., Yang, H., Wang, X., Gao, G. F., Yang, R., Wang, J., and Yu, J. (2007) A glimpse of streptococcal toxic shock syndrome from comparative genomics of S. suis 2 Chinese isolates. PLoS ONE 2: e315.
Clifton-Hadley, F. A. and Alexander, T. J. (1980) The carrier site and carrier rate of Streptococcus suis type II in pigs. Vet Rec 107: 40-41.
Clifton-Hadley, F. A., Alexander, T. J., and Enright, M. R. (1986) Monitoring herds for Streptococcus suis type 2: chance contamination of slaughter pigs. Vet Rec 118: 274.
Cockeran, R., Durandt, C., Feldman, C., Mitchell, T. J., and Anderson, R. (2002) Pneumolysin activates the synthesis and release of interleukin-8 by human neutrophils in vitro. J Infect Dis 186: 562-565.
Coue, M., Brenner, S. L., Spector, I., and Korn, E. D. (1987) Inhibition of actin polymerization by latrunculin A. FEBS Lett 213: 316-318.
Criss, A. K. and Casanova, J. E. (2003) Coordinate regulation of Salmonella enterica serovar Typhimurium invasion of epithelial cells by the Arp2/3 complex and Rho GTPases. Infect Immun 71: 2885-2891.
Cue, D., Southern, S. O., Southern, P. J., Prabhakar, J., Lorelli, W., Smallheer, J. M., Mousa, S. A., and Cleary, P. P. (2000) A nonpeptide integrin antagonist can inhibit epithelial cell ingestion of Streptococcus pyogenes by blocking formation of integrin alpha 5beta 1-fibronectin-M1 protein complexes. Proc Natl Acad Sci U S A 97: 2858-2863.
Czajkowsky, D. M., Hotze, E. M., Shao, Z., and Tweten, R. K. (2004) Vertical collapse of a cytolysin prepore moves its transmembrane beta-hairpins to the membrane. EMBO J 23: 3206-3215.
de Greeff, A., Buys, H., Verhaar, R., Dijkstra, J., van Alphen, L., and Smith, H. E. (2002) Contribution of fibronectin-binding protein to pathogenesis of Streptococcus suis serotype 2. Infect Immun 70: 1319-1325.
de Greeff, A., Wisselink, H. J., de Bree, F. M., Schultsz, C., Baums, C. G., Thi, H. N., Stockhofe- Zurwieden, N., and Smith, H. E. (2011) Genetic diversity of Streptococcus suis isolates as determined by comparative genome hybridization. BMC Microbiol 11: 161.
Dessing, M. C., Hirst, R. A., de Vos, A. F., and van der, Poll T. (2009) Role of Toll-like receptors 2 and 4 in pulmonary inflammation and injury induced by pneumolysin in mice. PLoS One 4: e7993.
Dominguez-Punaro, Mde L., Segura, M., Contreras, I., Lachance, C., Houde, M., Lecours, M. P., Olivier, M., and Gottschalk, M. (2010) In vitro characterization of the microglial inflammatory response to Streptococcus suis, an important emerging zoonotic agent of meningitis. Infect Immun 78: 5074-5085.

Literature Chapter 9
176
Dominguez-Punaro, M. C., Segura, M., Plante, M. M., Lacouture, S., Rivest, S., and Gottschalk, M. (2007) Streptococcus suis serotype 2, an important swine and human pathogen, induces strong systemic and cerebral inflammatory responses in a mouse model of infection. J Immunol 179: 1842-1854.
El Rachkidy, R. G., Davies, N. W., and Andrew, P. W. (2008) Pneumolysin generates multiple conductance pores in the membrane of nucleated cells. Biochem Biophys Res Commun 368: 786-792.
Enright, M. R., Alexander, T. J., and Clifton-Hadley, F. A. (1987) Role of houseflies (Musca domestica) in the epidemiology of Streptococcus suis type 2. Vet Rec 121: 132-133.
Fabisiak, M., Kita, J., Jedryczko, R., and Binek, M. (2005) Prevalence of the suilysin gene in Streptococcus suis strains isolated from diseased and healthy carrier pigs. Pol J Vet Sci 8: 141-145.
Feldman, C., Mitchell, T. J., Andrew, P. W., Boulnois, G. J., Read, R. C., Todd, H. C., Cole, P. J., and Wilson, R. (1990) The effect of Streptococcus pneumoniae pneumolysin on human respiratory epithelium in vitro. Microb Pathog 9: 275-284.
Finlay, B. B. (2005) Bacterial virulence strategies that utilize Rho GTPases. Curr Top Microbiol Immunol 291: 1-10.
Finlay, B. B. and Cossart, P. (1997) Exploitation of mammalian host cell functions by bacterial pathogens. Science 276: 718-725
Fittipaldi, N., Fuller, T. E., Teel, J. F., Wilson, T. L., Wolfram, T. J., Lowery, D. E., and Gottschalk, M. (2009) Serotype distribution and production of muramidase-released protein, extracellular factor and suilysin by field strains of Streptococcus suis isolated in the United States. Vet Microbiol 139: 310-317.
Fittipaldi, N., Gottschalk, M., Vanier, G., Daigle, F., and Harel, J. (2007) Use of selective capture of transcribed sequences to identify genes preferentially expressed by Streptococcus suis upon interaction with porcine brain microvascular endothelial cells. Appl Environ Microbiol 73: 4359- 4364.
Flanagan, J. J., Tweten, R. K., Johnson, A. E., and Heuck, A. P. (2009) Cholesterol exposure at the membrane surface is necessary and sufficient to trigger perfringolysin O binding. Biochemistry 48: 3977-3987.
Fongcom, A., Pruksakorn, S., Mongkol, R., Tharavichitkul, P., and Yoonim, N. (2001) Streptococcus suis infection in northern Thailand. J Med Assoc Thai 84: 1502-1508.
Garzoni, C. and Kelley, W. L. (2009) Staphylococcus aureus: new evidence for intracellular persistence. Trends Microbiol 17: 59-65.
Gekara, N. O. and Weiss, S. (2004) Lipid rafts clustering and signalling by listeriolysin O. Biochem Soc Trans 32: 712-714.
Gelber, S. E., Aguilar, J. L., Lewis, K. L., and Ratner, A. J. (2008) Functional and phylogenetic characterization of Vaginolysin, the human-specific cytolysin from Gardnerella vaginalis. J Bacteriol 190: 3896-3903.
Giddings, K. S., Johnson, A. E., and Tweten, R. K. (2003) Redefining cholesterol's role in the mechanism of the cholesterol-dependent cytolysins. Proc Natl Acad Sci U S A 100: 11315- 11320.

Literature Chapter 9
177
Giddings, K. S., Zhao, J., Sims, P. J., and Tweten, R. K. (2004) Human CD59 is a receptor for the cholesterol-dependent cytolysin intermedilysin. Nat Struct Mol Biol 11: 1173-1178.
Gottschalk, M., Lebrun, A., Wisselink, H., Dubreuil, J. D., Smith, H., and Vecht, U. (1998) Production of virulence-related proteins by Canadian strains of Streptococcus suis capsular type 2. Canadian Journal of Veterinary Research-Revue Canadienne de Recherche Veterinaire 62: 75-79.
Gottschalk, M., Petitbois, S., Higgins, R., and Jacques, M. (1991) Adherence of Streptococcus suis capsular type 2 to porcine lung sections. Can J Vet Res 55: 302-304.
Gottschalk, M. and Segura, M. (2000) The pathogenesis of the meningitis caused by Streptococcus suis: the unresolved questions. Veterinary Microbiology 76: 259-272.
Gottschalk, M., Segura, M., and Xu, J. (2007) Streptococcus suis infections in humans: the Chinese experience and the situation in North America. Anim Health Res Rev 8: 29-45.
Gottschalk, M., Xu, J., Calzas, C., and Segura, M. (2010) Streptococcus suis: a new emerging or an old neglected zoonotic pathogen? 25. Future Microbiol 5: 371-391.
Gottschalk, M. G., Lacouture, S., and Dubreuil, J. D. (1995) Characterization of Streptococcus suis capsular type 2 haemolysin. Microbiology 141 ( Pt 1): 189-195.
Greenwood, J., Steinman, L., and Zamvil, S. S. (2006) Statin therapy and autoimmune disease: from protein prenylation to immunomodulation. Nat Rev Immunol 6: 358-370.
Hamzaoui, N., Kerneis, S., Caliot, E., and Pringault, E. (2004) Expression and distribution of beta1 integrins in in vitro-induced M cells: implications for Yersinia adhesion to Peyer's patch epithelium. Cell Microbiol 6: 817-828.
Hall, A. (1998) Rho GTPases and the actin cytoskeleton. Science 279: 509-514.
Hardt, W. D., Chen, L. M., Schuebel, K. E., Bustelo, X. R., and Galan, J. E. (1998) S. typhimurium encodes an activator of Rho GTPases that induces membrane ruffling and nuclear responses in host cells. Cell 93: 815-826.
Herbert, D. and Todd, E. W. (1941) Purification and properties of a haemolysin produced by group A haemolytic streptococci (streptolysin O). Biochem J 35: 1124-1139.
Heuck, A. P., Hotze, E. M., Tweten, R. K., and Johnson, A. E. (2000) Mechanism of membrane insertion of a multimeric beta-barrel protein: perfringolysin O creates a pore using ordered and coupled conformational changes. Mol Cell 6: 1233-1242.
Higgins, R. and Gottschalk, M. (1990) An update on Streptococcus suis identification. J Vet Diagn Invest 2: 249-252.
Higgins,R. and Gottschalk,M. (2005) Streptococcal diseases. In Diseases of Swine. pp. 769-783.
Hoa, N. T., Chieu, T. T., Nghia, H. D., Mai, N. T., Anh, P. H., Wolbers, M., Baker, S., Campbell, J. I., Chau, N. V., Hien, T. T., Farrar, J., and Schultsz, C. (2011) The antimicrobial resistance patterns and associated determinants in Streptococcus suis isolated from humans in southern Vietnam, 1997-2008. BMC Infect Dis 11: 6.

Literature Chapter 9
178
Holo, H. and Nes, I. F. (1989) High-Frequency Transformation, by Electroporation, of Lactococcus lactis subsp. cremoris Grown with Glycine in Osmotically Stabilized Media. Appl Environ Microbiol 55: 3119-3123.
Hotze, E. M. and Tweten, R. K. (2011) Membrane assembly of the cholesterol-dependent cytolysin pore complex. Biochim Biophys Acta.
Huelsenbeck, J., Dreger, S., Gerhard, R., Barth, H., Just, I., Genth, H. (2007) Difference in the cytotoxic effects of toxin B from Clostridium difficile strain VPI 10463 and toxin B from variant Clostridium difficile strain 1470. Infect Immun 75(2): 801-9.
Iliev, A. I., Djannatian, J. R., Nau, R., Mitchell, T. J., and Wouters, F. S. (2007) Cholesterol-dependent actin remodeling via RhoA and Rac1 activation by the Streptococcus pneumoniae toxin pneumolysin. Proc Natl Acad Sci U S A 104: 2897-2902.
Iliev, A. I., Djannatian, J. R., Opazo, F., Gerber, J., Nau, R., Mitchell, T. J., and Wouters, F. S. (2009) Rapid microtubule bundling and stabilization by the Streptococcus pneumoniae neurotoxin pneumolysin in a cholesterol-dependent, non-lytic and Src-kinase dependent manner inhibits intracellular trafficking. Mol Microbiol 71: 461-477.
Jacobs, A. A. C., Loeffen, P. L. W., vandenBerg, A. J. G., and Storm, P. K. (1994) Identification, Purification, and Characterization of A Thiol-Activated Hemolysin (Suilysin) of Streptococcus Suis. Infection and Immunity 62: 1742-1748.
Jacobs, A. A. C., vandenBerg, A. J. G., and Loeffen, P. L. W. (1996) Protection of experimentally infected pigs by suilysin, the thiol-activated haemolysin of Streptococcus suis. Veterinary Record 139: 225-228.
Johnson, M. K., Geoffroy, C., and Alouf, J. E. (1980) Binding of cholesterol by sulfhydryl-activated cytolysins. Infect Immun 27: 97-101.
Jonsson, K., Signas, C., Muller, H. P., and Lindberg, M. (1991) Two different genes encode fibronectin binding proteins in Staphylococcus aureus. The complete nucleotide sequence and characterization of the second gene. Eur J Biochem 202: 1041-1048.
Jounblat, R., Kadioglu, A., Mitchell, T. J., and Andrew, P. W. (2003) Pneumococcal behavior and host responses during bronchopneumonia are affected differently by the cytolytic and complement- activating activities of pneumolysin. Infect Immun 71: 1813-1819.
Just, I., Selzer, J., Wilm, M., Eichel-Streiber, C., Mann, M., and Aktories, K. (1995) Glucosylation of Rho proteins by Clostridium difficile toxin B. Nature 375: 500-503.
Kim, D., Han, K., Oh, Y., Kim, C. H., Kang, I., Lee, J., Gottschalk, M., and Chae, C. (2010) Distribution of capsular serotypes and virulence markers of Streptococcus suis isolated from pigs with polyserositis in Korea. Can J Vet Res 74: 314-316.
King, S. J., Heath, P. J., Luque, I., Tarradas, C., Dowson, C. G., and Whatmore, A. M. (2001) Distribution and genetic diversity of suilysin in Streptococcus suis isolated from different diseases of pigs and characterization of the genetic basis of suilysin absence. Infection and Immunity 69: 7572-7582.
Kock, C., Beineke, A., Seitz, M., Ganter, M., Waldmann, K. H., Valentin-Weigand, P., and Baums, C. G. (2009) Intranasal immunization with a live Streptococcus suis isogenic ofs mutant elicited suilysin-neutralization titers but failed to induce opsonizing antibodies and protection. Veterinary Immunology and Immunopathology 132: 135-145.

Literature Chapter 9
179
Korchev, Y. E., Bashford, C. L., Pederzolli, C., Pasternak, C. A., Morgan, P. J., Andrew, P. W., and Mitchell, T. J. (1998) A conserved tryptophan in pneumolysin is a determinant of the characteristics of channels formed by pneumolysin in cells and planar lipid bilayers. Biochem J 329 ( Pt 3): 571-577.
Krawczyk-Balska, A. and Bielecki, J. (2005) Listeria monocytogenes listeriolysin O and phosphatidylinositol-specific phospholipase C affect adherence to epithelial cells. Can J Microbiol 51: 745-751.
Lafont, F. and van der Goot, F. G. (2005) Bacterial invasion via lipid rafts. Cell Microbiol 7: 613-620.
Lalonde, M., Segura, M., Lacouture, S., and Gottschalk, M. (2000) Interactions between Streptococcussuis serotype 2 and different epithelial cell lines. Microbiology 146 ( Pt 8): 1913- 1921.
Lecours, M. P., Gottschalk, M., Houde, M., Lemire, P., Fittipaldi, N., and Segura, M. (2011) Critical Role for Streptococcus suis Cell Wall Modifications and Suilysin in Resistance to Complement-Dependent Killing by Dendritic Cells. J Infect Dis 204: 919-929.
Logan, R. P., Robins, A., Turner, G. A., Cockayne, A., Borriello, S. P., and Hawkey, C. J. (1998) A novel flow cytometric assay for quantitating adherence of Helicobacter pylori to gastric epithelial cells. J Immunol Methods 213: 19-30.
Lowe, B. A., Marsh, T. L., Isaacs-Cosgrove, N., Kirkwood, R. N., Kiupel, M., and Mulks, M. H. (2011) Microbial communities in the tonsils of healthy pigs. Vet Microbiol 147: 346-357.
Lun, S. C., Perez-Casal, J., Connor, W., and Willson, P. J. (2003) Role of suilysin in pathogenesis of Streptococcus suis capsular serotype 2. Microbial Pathogenesis 34: 27-37.
Medina, E. (2010) Murine model of pneumococcal pneumonia. Methods Mol Biol 602: 405-410.
Mai, N. T., Hoa, N. T., Nga, T. V., Linh, le D., Chau, T. T., Sinh, D. X., Phu, N. H., Chuong, L. V., Diep, T. S., Campbell, J., Nghia, H. D., Minh, T. N., Chau, N. V., de Jong, M. D., Chinh, N. T., Hien, T. T., Farrar, J., and Schultsz, C. (2008) Streptococcus suis meningitis in adults in Vietnam. Clin Infect Dis 46: 659-667.
Malley, R., Henneke, P., Morse, S. C., Cieslewicz, M. J., Lipsitch, M., Thompson, C. M., Kurt-Jones, E., Paton, J. C., Wessels, M. R., and Golenbock, D. T. (2003) Recognition of pneumolysin by Toll-like receptor 4 confers resistance to pneumococcal infection. Proc Natl Acad Sci U S A 100: 1966-1971.
Mackay, D. J. and Hall, A. (1998) Rho GTPases. J Biol Chem 273: 20685-20688.
Messier, S., Lacouture, S., and Gottschalk, M. (2008) Distribution of Streptococcus suis capsular types from 2001 to 2007. Can Vet J 49: 461-462.
Michel, E., Reich, K. A., Favier, R., Berche, P., and Cossart, P. (1990) Attenuated mutants of the intracellular bacterium Listeria monocytogenes obtained by single amino acid substitutions in listeriolysin O. Mol Microbiol 4: 2167-2178.
Navacharoen, N., Chantharochavong, V., Hanprasertpong, C., Kangsanarak, J., and Lekagul, S. (2009) Hearing and vestibular loss in Streptococcus suis infection from swine and traditional raw pork exposure in northern Thailand. J Laryngol Otol 123: 857-862.

Literature Chapter 9
180
Ngo, T. H., Tran, T. B., Tran, T. T., Nguyen, V. D., Campbell, J., Pham, H. A., Huynh, H. T., Nguyen, V. V., Bryant, J. E., Tran, T. H., Farrar, J., and Schultsz, C. (2011) Slaughterhouse pigs are a major reservoir of Streptococcus suis serotype 2 capable of causing human infection in southern Vietnam. PLoS One 6: e17943.
Norton, P. M., Rolph, C., Ward, P. N., Bentley, R. W., and Leigh, J. A. (1999) Epithelial invasion and cell lysis by virulent strains of Streptococcus suis is enhanced by the presence of suilysin. Fems Immunology and Medical Microbiology 26: 25-35.
O'Sullivan, T., Friendship, R., Blackwell, T., Pearl, D., McEwen, B., Carman, S., Slavic, D., and Dewey, C. (2011) Microbiological identification and analysis of swine tonsils collected from carcasses at slaughter. Can J Vet Res 75: 106-111.
Okamoto, S., Kawabata, S., Terao, Y., Fujitaka, H., Okuno, Y., and Hamada, S. (2004) The Streptococcus pyogenes capsule is required for adhesion of bacteria to virus-infected alveolar epithelial cells and lethal bacterial-viral superinfection. Infect Immun 72: 6068-6075.
Okwumabua, O., Abdelmagid, O., and Chengappa, M. M. (1999) Hybridization analysis of the gene encoding a hemolysin (suilysin) of Streptococcus suis type 2: evidence for the absence of the gene in some isolates. FEMS Microbiol Lett 181: 113-121.
Orihuela, C. J., Gao, G., Francis, K. P., Yu, J., and Tuomanen, E. I. (2004) Tissue-specific contributions of pneumococcal virulence factors to pathogenesis. J Infect Dis 190: 1661-1669.
Padungtod, P., Tharavichitkul, P., Junya, S., Chaisowong, W., Kadohira, M., Makino, S., and Sthitmatee, N. (2010) Incidence and presence of virulence factors of Streptococcus suis infection in slaughtered pigs from Chiang Mai, Thailand. Southeast Asian J Trop Med Public Health 41: 1454-1461.
Pallares, F. J., Halbur, P. G., Schmitt, C. S., Roth, J. A., Opriessnig, T., Thomas, P. J., Kinyon, J. M., Murphy, D., Frank, D. E., and Hoffman, L. J. (2003) Comparison of experimental models for Streptococcus suis infection of conventional pigs. Can J Vet Res 67: 225-228.
Pan, X. Z., Ge, J. C., Li, M., Wu, B., Wang, C. J., Wang, J., Feng, Y. J., Yin, Z. M., Zheng, F., Cheng, G., Sun, W., Ji, H. F., Hu, D., Shi, P. J., Feng, X. D., Hao, X. N., Dong, R. P., Hu, F. Q., and Tang, J. Q. (2009) The Orphan Response Regulator CovR: a Globally Negative Modulator of Virulence in Streptococcus suis Serotype 2. Journal of Bacteriology 191: 2601-2612. Perch, B., Pedersen, K. B., and Henrichsen, J. (1983) Serology of capsulated streptococci pathogenic for pigs: six new serotypes of Streptococcus suis. J Clin Microbiol 17: 993-996.
Petersen, R., Hannerz, H., Tuchsen, F., and Egerton, J. R. (2011) Meningitis, sepsis and endocarditis among workers occupationally exposed to pigs. Occup Med (Lond).
Pinkney, M., Beachey, E., and Kehoe, M. (1989) The thiol-activated toxin streptolysin O does not require a thiol group for cytolytic activity. Infect Immun 57: 2553-2558.
Polekhina, G., Giddings, K. S., Tweten, R. K., and Parker, M. W. (2005) Insights into the action of the superfamily of cholesterol-dependent cytolysins from studies of intermedilysin. Proc Natl Acad Sci U S A 102: 600-605.
Princivalli, M. S., Palmieri, C., Magi, G., Vignaroli, C., Manzin, A., Camporese, A., Barocci, S., Magistrali, C., and Facinelli, B. (2009) Genetic diversity of Streptococcus suis clinical isolates from pigs and humans in Italy (2003-2007). Euro Surveill 14.

Literature Chapter 9
181
Que, Y. A., Haefliger, J. A., Francioli, P., and Moreillon, P. (2000) Expression of Staphylococcus aureus clumping factor A in Lactococcus lactis subsp. cremoris using a new shuttle vector. Infect Immun 68: 3516-3522.
Ramachandran, R., Tweten, R. K., and Johnson, A. E. (2004) Membrane-dependent conformational changes initiate cholesterol-dependent cytolysin oligomerization and intersubunit beta-strand alignment. Nat Struct Mol Biol 11: 697-705.
Ramachandran, R., Heuck, A. P., Tweten, R. K., and Johnson, A. E. (2002) Structural insights into the membrane-anchoring mechanism of a cholesterol-dependent cytolysin. Nat Struct Biol 9: 823- 827.
Rampersaud, R., Planet, P. J., Randis, T. M., Kulkarni, R., Aguilar, J. L., Lehrer, R. I., and Ratner, A. J. (2011) Inerolysin, a cholesterol-dependent cytolysin produced by Lactobacillus iners. J Bacteriol 193: 1034-1041.
Ratner, A. J., Hippe, K. R., Aguilar, J. L., Bender, M. H., Nelson, A. L., and Weiser, J. N. (2006) Epithelial cells are sensitive detectors of bacterial pore-forming toxins. J Biol Chem 281: 12994-12998.
Reid, T., Furuyashiki, T., Ishizaki, T., Watanabe, G., Watanabe, N., Fujisawa, K., Morii, N., Madaule, P., and Narumiya, S. (1996) Rhotekin, a new putative target for Rho bearing homology to a serine/threonine kinase, PKN, and rhophilin in the rho-binding domain. J Biol Chem 271: 13556-13560.
Ridley, A. J., Paterson, H. F., Johnston, C. L., Diekmann, D., and Hall, A. (1992) The small GTP- binding protein rac regulates growth factor-induced membrane ruffling. Cell 70: 401-410.
Robertson, I. D., Blackmore, D. K., Hampson, D. J., and Fu, Z. F. (1991) A longitudinal study of natural infection of piglets with Streptococcus suis types 1 and 2. Epidemiol Infect 107: 119- 126.
Rossjohn, J., Feil, S. C., McKinstry, W. J., Tweten, R. K., and Parker, M. W. (1997) Structure of a cholesterol-binding, thiol-activated cytolysin and a model of its membrane form. Cell 89: 685- 692.
Rossjohn, J., Gilbert, R. J., Crane, D., Morgan, P. J., Mitchell, T. J., Rowe, A. J., Andrew, P. W., Paton, J. C., Tweten, R. K., and Parker, M. W. (1998) The molecular mechanism of pneumolysin, a virulence factor from Streptococcus pneumoniae. J Mol Biol 284: 449-461.
Rossjohn, J., Polekhina, G., Feil, S. C., Morton, C. J., Tweten, R. K., and Parker, M. W. (2007) Structures of perfringolysin O suggest a pathway for activation of cholesterol-dependent cytolysins. J Mol Biol 367: 1227-1236.
Rubins, J. B., Paddock, A. H., Charboneau, D., Berry, A. M., Paton, J. C., and Janoff, E. N. (1998) Pneumolysin in pneumococcal adherence and colonization. Microb Pathog 25: 337-342.
Saeland, E., Vidarsson, G., and Jonsdottir, I. (2000) Pneumococcal pneumonia and bacteremia model in mice for the analysis of protective antibodies. Microb Pathog 29: 81-91.
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N Y.
Sanford, S. E. and Tilker, M. E. (1982) Streptococcus suis type II-associated diseases in swine: observations of a one-year study. J Am Vet Med Assoc 181: 673-676.

Literature Chapter 9
182
Saunders, F. K., Mitchell, T. J., Walker, J. A., Andrew, P. W., and Boulnois, G. J. (1989) Pneumolysin, the thiol-activated toxin of Streptococcus pneumoniae, does not require a thiol group for in vitro activity. Infect Immun 57: 2547-2552.
Schmidt, G. and Aktories, K. (2000) Rho GTPase-activating toxins: cytotoxic necrotizing factors and dermonecrotic toxin. Methods Enzymol 325: 125-136.
Schwartz, M. A. and Shattil, S. J. (2000) Signaling networks linking integrins and rho family GTPases. Trends Biochem Sci 25: 388-391.
Scibelli, A., Matteoli, G., Roperto, S., Alimenti, E., Dipineto, L., Pavone, L. M., Della, Morte R., Menna, L. F., Fioretti, A., and Staiano, N. (2005) Flavoridin inhibits Yersinia enterocolitica uptake into fibronectin-adherent HeLa cells. FEMS Microbiol Lett 247: 51-57.
Scibelli, A., Roperto, S., Manna, L., Pavone, L. M., Tafuri, S., Della, Morte R., and Staiano, N. (2007) Engagement of integrins as a cellular route of invasion by bacterial pathogens. Vet J 173: 482- 491.
Segers, R. P., Kenter, T., de Haan, L. A., and Jacobs, A. A. (1998) Characterisation of the gene encoding suilysin from Streptococcus suis and expression in field strains. FEMS Microbiol Lett 167: 255-261.
Segura, M. and Gottschalk, M. (2002) Streptococcus suis interactions with the murine macrophage cell line J774: adhesion and cytotoxicity. Infect Immun 70: 4312-4322.
Segura, M., Stankova, J., and Gottschalk, M. (1999) Heat-killed Streptococcus suis capsular type 2 strains stimulate tumor necrosis factor alpha and interleukin-6 production by murine macrophages. Infect Immun 67: 4646-4654.
Segura, M., Vadeboncoeur, N., and Gottschalk, M. (2002) CD14-dependent and -independent cytokine and chemokine production by human THP-1 monocytes stimulated by Streptococcus suis capsular type 2. Clin Exp Immunol 127: 243-254.
Segura, M., Vanier, G., Al Numani, D., Lacouture, S., Olivier, M., and Gottschalk, M. (2006) Proinflammatory cytokine and chemokine modulation by Streptococcus suis in a whole-blood culture system. FEMS Immunol Med Microbiol 47: 92-106.
Seitz, M., Baums, CG, Gerhard, R, Just, I, Neis, C., Benga, L., Fulde, M., Nerlich, A, Rohde, M., Goethe, R., and Valentin-Weigand, P. Subcytolytic activity of suilysin promotes invasion of Streptococcus suis in HEp-2 epithelial cells by Rac-dependent activation of the actin cytoskeleton. Ref Type: submitted to Cellular Microbiology Sekino-Suzuki, N., Nakamura, M., Mitsui, K. I., and Ohno-Iwashita, Y. (1996) Contribution of individual tryptophan residues to the structure and activity of theta-toxin (perfringolysin O), a cholesterol- binding cytolysin. Eur J Biochem 241: 941-947.
Shepard, L. A., Shatursky, O., Johnson, A. E., and Tweten, R. K. (2000) The mechanism of pore assembly for a cholesterol-dependent cytolysin: formation of a large prepore complex precedes the insertion of the transmembrane beta-hairpins. Biochemistry 39: 10284-10293.
Shimada, Y., Maruya, M., Iwashita, S., and Ohno-Iwashita, Y. (2002) The C-terminal domain of perfringolysin O is an essential cholesterol-binding unit targeting to cholesterol-rich microdomains. Eur J Biochem 269: 6195-6203.

Literature Chapter 9
183
Silva, L. M., Baums, C. G., Rehm, T., Wisselink, H. J., Goethe, R., and Valentin-Weigand, P. (2006) Virulence-associated gene profiling of Streptococcus suis isolates by PCR. Vet Microbiol 115: 117-127.
Smith, H. E., Damman, M., van der Velde, J., Wagenaar, F., Wisselink, H. J., Stockhofe-Zurwieden, N., and Smits, M. A. (1999) Identification and characterization of the cps locus of Streptococcus suis serotype 2: the capsule protects against phagocytosis and is an important virulence factor. Infect Immun 67: 1750-1756.
Smith, H. E., Vecht, U., Wisselink, H. J., StockhofeZurwieden, N., Biermann, Y., and Smits, M. A. (1996) Mutants of Streptococcus suis types 1 and 2 impaired in expression of muramidase- released protein and extracellular protein induce disease in newborn germfree pigs. Infection and Immunity 64: 4409-4412.
Su, Y., Yao, W., Perez-Gutierrez, O. N., Smidt, H., and Zhu, W. Y. (2008) Changes in abundance of Lactobacillus spp. and Streptococcus suis in the stomach, jejunum and ileum of piglets after weaning. FEMS Microbiol Ecol 66: 546-555.
Soltani, C. E., Hotze, E. M., Johnson, A. E., and Tweten, R. K. (2007) Specific protein-membrane contacts are required for prepore and pore assembly by a cholesterol-dependent cytolysin. J Biol Chem 282: 15709-15716.
Soltani, C. E., Hotze, E. M., Johnson, A. E., and Tweten, R. K. (2007b) Structural elements of the cholesterol-dependent cytolysins that are responsible for their cholesterol-sensitive membrane interactions. Proc Natl Acad Sci U S A 104: 20226-20231.
Sriskandan, S. and Slater, J. D. (2006) Invasive disease and toxic shock due to zoonotic Streptococcus suis: an emerging infection in the East? PLoS Med 3: e187.
Staats, J. J., Feder, I., Okwumabua, O., and Chengappa, M. M. (1997) Streptococcus suis: past and present. Vet Res Commun 21: 381-407.
Staats, J. J., Plattner, B. L., Stewart, G. C., and Chengappa, M. M. (1999) Presence of the Streptococcus suis suilysin gene and expression of MRP and EF correlates with high virulence in Streptococcus suis type 2 isolates. Veterinary Microbiology 70: 201-211.
Stachowiak, R., Wisniewski, J., Osinska, O., and Bielecki, J. (2009) Contribution of cysteine residue to the properties of Listeria monocytogenes listeriolysin O. Can J Microbiol 55: 1153-1159.
Strangmann, E., Froleke, H., and Kohse, K. P. (2002) Septic shock caused by Streptococcus suis: case report and investigation of a risk group. Int J Hyg Environ Health 205: 385-392.
Sukeno, A., Nagamune, H., Whiley, R. A., Jafar, S. I., Aduse-Opoku, J., Ohkura, K., Maeda, T., Hirota, K., Miyake, Y., and Kourai, H. (2005) Intermedilysin is essential for the invasion of hepatoma HepG2 cells by Streptococcus intermedius. Microbiol Immunol 49: 681-694.
Takamatsu, D., Osaki, M., and Sekizaki, T. (2002) Evidence for lateral transfer of the Suilysin gene region of Streptococcus suis. J Bacteriol 184: 2050-2057.
Takamatsu, D., Osaki, M., and Sekizaki, T. (2001) Thermosensitive suicide vectors for gene replacement in Streptococcus suis. Plasmid 46: 140-148.
Tang, J., Wang, C., Feng, Y., Yang, W., Song, H., Chen, Z., Yu, H., Pan, X., Zhou, X., Wang, H., Wu, B., Wang, H., Zhao, H., Lin, Y., Yue, J., Wu, Z., He, X., Gao, F., Khan, A. H., Wang, J., Zhao, G. P., Wang, Y., Wang, X., Chen, Z., and Gao, G. F. (2006) Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med 3: e151.

Literature Chapter 9
184
Tarradas, C., Borge, C., Arenas, A., Maldonado, A., Astorga, R., Miranda, A., and Luque, I. (2001) Suilysin production by Streptococcus suis strains isolated from diseased and healthy carrier pigs in Spain. Vet Rec 148: 183-184.
Tenenbaum, T., Adam, R., Eggelnpohler, I., Matalon, D., Seibt, A., GE, K. Novotny, Galla, H. J., and Schroten, H. (2005) Strain-dependent disruption of blood-cerebrospinal fluid barrier by Streptoccocus suis in vitro. FEMS Immunol Med Microbiol 44: 25-34.
Tenenbaum, T., Essmann, F., Adam, R., Seibt, A., Janicke, R. U., Novotny, G. E., Galla, H. J., and Schroten, H. (2006) Cell death, caspase activation, and HMGB1 release of porcine choroid plexus epithelial cells during Streptococcus suis infection in vitro. Brain Res 1100: 1-12.
Tenenbaum, T., Papandreou, T., Gellrich, D., Friedrichs, U., Seibt, A., Adam, R., Wewer, C., Galla, H. J., Schwerk, C., and Schroten, H. (2008) Polar bacterial invasion and translocation of Streptococcus suis across the blood-cerebrospinal fluid barrier in vitro. Cell Microbiol.
Tian, Y., Aarestrup, F. M., and Lu, C. P. (2004) Characterization of Streptococcus suis serotype 7 isolates from diseased pigs in Denmark. Vet Microbiol 103: 55-62.
Tomoyasu, T., Tabata, A., Hiroshima, R., Imaki, H., Masuda, S., Whiley, R. A., Aduse-Opoku, J., Kikuchi, K., Hiramatsu, K., and Nagamune, H. (2010) Role of catabolite control protein A in the regulation of intermedilysin production by Streptococcus intermedius. Infect Immun 78: 4012- 4021.
Tran, Van Nhieu, Caron, E., Hall, A., and Sansonetti, P. J. (1999) IpaC induces actin polymerization and filopodia formation during Shigella entry into epithelial cells. EMBO J 18: 3249-3262.
Trottier, S., Higgins, R., Brochu, G., and Gottschalk, M. (1991) A Case of Human Endocarditis Due to Streptococcus Suis in North-America. Reviews of Infectious Diseases 13: 1251-1252.
Tsuchiya, K., Kawamura, I., Takahashi, A., Nomura, T., Kohda, C., and Mitsuyama, M. (2005) Listeriolysin O-induced membrane permeation mediates persistent interleukin-6 production in Caco-2 cells during Listeria monocytogenes infection in vitro. Infect Immun 73: 3869-3877.
Tweten, R. K. (2005) Cholesterol-dependent cytolysins, a family of versatile pore-forming toxins. Infection and Immunity 73: 6199-6209.
Vadeboncoeur, C. and Pelletier, M. (1997) The phosphoenolpyruvate:sugar phosphotransferase system of oral streptococci and its role in the control of sugar metabolism. Fems Microbiology Reviews 19: 187-207. Vadeboncoeur, N., Segura, M., Al Numani, D., Vanier, G., and Gottschalk, M. (2003) Pro-inflammatory cytokine and chemokine release by human brain microvascular endothelial cells stimulated by Streptococcus suis serotype 2. FEMS Immunol Med Microbiol 35: 49-58.
Valentin-Weigand, P., Benkel, P., Rohde, M., and Chhatwal, G. S. (1996) Entry and intracellular survival of group B streptococci in J774 macrophages. Infect Immun 64: 2467-2473.
Vanier, G., Segura, M., Friedl, P., Lacouture, S., and Gottschalk, M. (2004) Invasion of porcine brain microvascular endothelial cells by Streptococcus suis serotype 2. Infection and Immunity 72: 1441-1449.
Vanier, G., Segura, M., and Gottschalk, M. (2007) Characterization of the invasion of porcine endothelial cells by Streptococcus suis serotype 2. Can J Vet Res 71: 81-89.

Literature Chapter 9
185
Vanier, G., Segura, M., Lecours, M. P., Grenier, D., and Gottschalk, M. (2008) Porcine brain microvascular endothelial cell-derived interleukin-8 is first induced and then degraded by Streptococcus suis. Microb Pathog.
Vanier, G., Fittipaldi, N., Slater, J. D., Dominguez-Punaro, Mde L., Rycroft, A. N., Segura, M., Maskell, D. J., and Gottschalk, M. (2009) New putative virulence factors of Streptococcus suis involved in invasion of porcine brain microvascular endothelial cells. Microb Pathog 46: 13-20.
Vecht, U., Wisselink, H. J., Jellema, M. L., and Smith, H. E. (1991) Identification of 2 Proteins Associated with Virulence of Streptococcus Suis Type-2. Infection and Immunity 59: 3156- 3162.
Vecht, U., StockhofeZurwieden, N., Tetenburg, B. J., Wisselink, H. J., and Smith, H. E. (1997) Virulence of Streptococcus suis type 2 for mice and pigs appeared host-specific. Veterinary Microbiology 58: 53-60.
Waheed, A. A., Shimada, Y., Heijnen, H. F., Nakamura, M., Inomata, M., Hayashi, M., Iwashita, S., Slot, J. W., and Ohno-Iwashita, Y. (2001) Selective binding of perfringolysin O derivative to cholesterol-rich membrane microdomains (rafts). Proc Natl Acad Sci U S A 98: 4926-4931.
Walker, J. A., Allen, R. L., Falmagne, P., Johnson, M. K., and Boulnois, G. J. (1987) Molecular cloning, characterization, and complete nucleotide sequence of the gene for pneumolysin, the sulfhydryl-activated toxin of Streptococcus pneumoniae. Infect Immun 55: 1184-1189.
Wangkaew, S., Chaiwarith, R., Tharavichitkul, P., and Supparatpinyo, K. (2006) Streptococcus suis infection: a series of 41 cases from Chiang Mai University Hospital. J Infect 52: 455-460.
Watson, K. C., Rose, T. P., and Kerr, E. J. (1972) Some factors influencing the effect of cholesterol on streptolysin O activity. J Clin Pathol 25: 885-891.
Wei, Z., Li, R., Zhang, A., He, H., Hua, Y., Xia, J., Cai, X., Chen, H., and Jin, M. (2009) Characterization of Streptococcus suis isolates from the diseased pigs in China between 2003 and 2007. Vet Microbiol 137: 196-201.
Weis, S. and Palmer, M. (2001) Streptolysin O: the C-terminal, tryptophan-rich domain carries functional sites for both membrane binding and self-interaction but not for stable oligomerization. Biochim Biophys Acta 1510: 292-299.
Wertheim, H. F., Nghia, H. D., Taylor, W., and Schultsz, C. (2009) Streptococcus suis: an emerging human pathogen. Clin Infect Dis 48: 617-625.
Willenborg, J., Fulde, M., de Greeff, A., Rohde, M., Smith, H. E., Valentin-Weigand, P., and Goethe, R. (2011) Role of glucose and CcpA in capsule expression and virulence of Streptococcus suis. Microbiology 157: 1823-1833.
Williams, A. E. and Blakemore, W. F. (1990) Pathogenesis of meningitis caused by Streptococcus suis type 2. J Infect Dis 162: 474-481.
Williams, A. E., Blakemore, W. F., and Alexander, T. J. (1988) A murine model of Streptococcus suis type 2 meningitis in the pig. Res Vet Sci 45: 394-399. Windsor, R. S. and Elliott, S. D. (1975) Streptococcal infection in young pigs. IV. An outbreak of streptococcal meningitis in weaned pigs. J Hyg (Lond) 75: 69-78.

Literature Chapter 9
186
Wisselink, H. J., Smith, H. E., Stockhofe-Zurwieden, N., Peperkamp, K., and Vecht, U. (2000) Distribution of capsular types and production of muramidase-released protein (MRP) and extracellular factor (EF) of Streptococcus suis strains isolated from diseased pigs in seven European countries. Veterinary Microbiology 74: 237-248.
Wu, T., Chang, H., Tan, C., Bei, W., and Chen, H. (2009) The orphan response regulator RevSC21 controls the attachment of Streptococcus suis serotype-2 to human laryngeal epithelial cells and the expression of virulence genes. FEMS Microbiol Lett 292: 170-181.
Wu, T., Zhao, Z., Zhang, L., Ma, H., Lu, K., Ren, W., Liu, Z., Chang, H., Bei, W., Qiu, Y., and Chen, H. (2011) Trigger factor of Streptococcus suis is involved in stress tolerance and virulence. Microb Pathog 51: 69-76.
Xiong, Y., Liu, Q., Qin, F. Y., Bai, Y., Zhu, W., Li, H. M., Guo, J. G., Qin, L., Pan, J., Long, J. M., and Chen, L. (2007) [Study on the molecular epidemiology of Streptococcus suis type 2 from healthy pigs in Guangxi]. Zhonghua Liu Xing Bing Xue Za Zhi 28: 593-596.
Xu, L., Huang, B., Du, H., Zhang, X. C., Xu, J., Li, X., and Rao, Z. (2010) Crystal structure of cytotoxin protein suilysin from Streptococcus suis. Protein Cell 1: 96-105.
Ye, C., Zheng, H., Zhang, J., Jing, H., Wang, L., Xiong, Y., Wang, W., Zhou, Z., Sun, Q., Luo, X., Du, H., Gottschalk, M., and Xu, J. (2009) Clinical, Experimental, and Genomic Differences between Intermediately Pathogenic, Highly Pathogenic, and Epidemic Streptococcus suis. J Infect Dis 199: 97-107.
Ye, C., Zhu, X., Jing, H., Du, H., Segura, M., Zheng, H., Kan, B., Wang, L., Bai, X., Zhou, Y., Cui, Z., Zhang, S., Jin, D., Sun, N., Luo, X., Zhang, J., Gong, Z., Wang, X., Wang, L., Sun, H., Li, Z., Sun, Q., Liu, H., Dong, B., Ke, C., Yuan, H., Wang, H., Tian, K., Wang, Y., Gottschalk, M., and Xu, J. (2006) Streptococcus suis sequence type 7 outbreak, Sichuan, China. Emerg Infect Dis 12: 1203-1208.
Yu, H., Jing, H., Chen, Z., Zheng, H., Zhu, X., Wang, H., Wang, S., Liu, L., Zu, R., Luo, L., Xiang, N., Liu, H., Liu, X., Shu, Y., Lee, S. S., Chuang, S. K., Wang, Y., Xu, J., and Yang, W. (2006) Human Streptococcus suis outbreak, Sichuan, China. Emerg Infect Dis 12: 914-920.
Zhao, Y., Liu, G., Li, S., Wang, M., Song, J., Wang, J., Tang, J., Li, M., and Hu, F. (2011) Role of a type IV-like secretion system of Streptococcus suis 2 in the development of streptococcal toxic shock syndrome. J Infect Dis 204: 274-281.
Zheng, H., Punaro, M. C., Segura, M., Lachance, C., Rivest, S., Xu, J., Houde, M., and Gottschalk, M. (2011) Toll-like receptor 2 is partially involved in the activation of murine astrocytes by Streptococcus suis, an important zoonotic agent of meningitis. J Neuroimmunol 234: 71-83.

Acknowledgments First and foremost, I would like to sincerely thank my supervisor, Prof. Dr. Peter Valentin-Weigand, for offering me the opportunity of performing this work, for his support and invaluable advice. I would like to thank my co-supervisors Prof. Dr. Sike Rautenschlein and PD Dr. Manfred Rohde for their interest in my work, their ideas and productive discussions. I would like to thank PD Dr. Christoph Baums for his competence and precious advises and Prof. Dr. Ralf Goethe for his excellent support and everyday supervision. Many thanks, to Prof. Dr. Ingo Just and PD Dr. Ralf Gerhard for good cooperation and expertise in G-proteins, PD Dr. Manfred Rohde for providing the wonderful electron micrographs, Prof. Dr. Andreas Beineke for the histopathological examination of endless mice organs and Dr. Oliver Goldmann for his assistance with the intranasal application techniques and anaesthesia of mice. I would like to thank all the past and present members of the Institute for Microbiology, for an excellent collegial and productive working atmosphere and for never getting tired of laughing with and at me: Jens Abel, Tina Basler, Tanja Bauer, Sabine Baumert, Laurentiu Benga, Nadine Büttner, Sabrina Diehl, Elke Eckelt, Marcus Fulde, Sabine Göbel, Nina Janze, Christoph Kock, Jochen Meens, Kristin Laarmann, Thorsten Meißner, Andreas Mietze, Franziska Müller, Christina Neis, Andreas Nerlich, Girish Ramachandran, Nantaporn Ruangkiattikul, Julian Sander, Franziska Schill, Jana Seele, Mathias Weigoldt, Yenehiwot Berhanu Weldearegay, Joerg Willenborg and Daniela Willms. Many thanks, to Jörg Merkel for helping me with all technical questions concerning computer and statistic trouble. Finally and most important, I would like to thank my parents, grandma and Jörg, for their constant support in all respects, for their love and encouragement all over the years, and allowing me to pursue my dreams.





























