Embed Size (px)
Citation preview

JOUtNAL OF BACnIUIOLOGY, Nov. 1967, P. 1531-1537Copyright 0 1"7 American Society for Microbiology
Dominant Mutation for Ethionine Resistancein Saccharomyces cerevisiae
K. D. SPENCE,' L. W. PARKS, Am S. K. SHAPIRODepartment of Microbiology, Oregon State University, Corvallis, Oregon 97331 and
Division ofBiological and Medical Research, Argonne National Laboratory,Argonne, Illinois 60439
Received for publication 30 August 1967
An ethionine-resistant mutant of Saccharomyces cerevisiae has been investigatedwhose mutation (etr2) confers resistance to the heterozygous diploid also containingthe sensitive allele, et". The mutation is apparently specific for reversal of ethionineinhibition. The principal difference between the sensitive et" strain and the mutantwas the latter's inability to concentrate large intracellular quantities of adenosyl-ethionine. Reduced incorporation of ethyl groups or ethionine in other cellularfractions of the mutant was also detected. The data show that the mutant has notlost the ability to form adenosylethionine. It is suggested that the mutant has anincreased ability to hydrolyze this sulfonium compound after it has been synthesized.It is possible that some of the ethionine is detoxified before it can participate in pro-tein or adenosylethionine synthesis. No mutant alteration in accumulation ofethionine from the medium was detected. In the presence of ethionine, the parentalstrain accumulated 25 times more adenosylethionine than did the mutant. However,with methionine, only twice as much adenosylmethionine was accumulated by theparental strain as by the mutant.
At least two genetically different types ofethionine-resistant mutants of Saccharomycescerevisiae have been isolated. The resistance ofthe majority of these mutants has been shown toresult from a single recessive mutation which alsoconferred resistance to p-fluorophenylalanine(13). The strains with the recessive mutationlacked the amino acid permease found in the wild-type organism. Nonspecificity in that permeationprocess in these clones was established. Appar-ently identical mutants have been isolated byothers (17). Similar mutants have also been de-scribed in Neurospora (1, 5).
In addition to those ethionine-resistant yeaststrains described above, a few mutants have beenisolated whose mutation is dominant in theheterozygous diploid with the wild-type allele.These organisms were of interest, since they ap-peared to have resulted from genetic alterationsinvolving enzymes of methionine metabolism.For this reason, the characteristics of growth andamino acid accumulation of these mutants wereinvestigated under conditions expected to char-acterize the physiological consequences of thegenetic alteration involved. The distribution of
1 Present address: Division of Biological and Medi-cal Research, Argonne National Laboratory, Ar-gonne, Ill. 60439.
labeled methyl and ethyl groups throughout sev-eral cellular fractions of the parental and mutantstrain was analyzed after exposure to labeledmethionine and ethionine.
MATERIALS AND METHODS
Chemicals and media. Adenosylmethionine andadenosylethionine were isolated from yeast cells (10,12). Methylthioadenosine and ethylthioadenosine wereobtained from acid hydrolysates of the parent sul-fonium compounds (7). All other chemicals wereobtained from commercial sources. Wickerham'ssynthetic medium (18), or modifications thereof, wereemployed in all experiments. Media included Wicker-ham's medium less methionine (WCLM) and Wicker-ham's minimal medium plus uracil (WM).
Organisms. S. cerevisiae strain 3701B, an ethionine-sensitive (ets) haploid auxotroph requiring uracil,served as the parental strain for selection of ethionine-resistant mutants. The ethionine-resistant mutantswere isolated by a technique reported previously (13).Two genetic types of ethionine-insensitive mutantshave thus far been discovered. The first type, termedetr (13), occurs in greater than 10-fold excess over asecond type, designated etr'. The representative strainof the latter type used in these studies was R3720(ura etrl).Growth in supplements. Growth responses of selected
haploid and diploid strains were determined on WMagar with no supplement and in the presence of either25 ,ug of DL-ethionine per ml or 50 tg of DL-p-fluoro-
1531
Vol. 94, No. $Printed In U.S.A.
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

SPENCE, PARKS, AND SHAPIRO
phenylalanine per ml. Growth was recorded after48 hr.
Intracellular distribution ofradioactivityfrom labeledethionine and methionine. Prior to addition of labeledmethionine or ethionine, the strains were incubated inWCLM medium for 18 hr at 30 C. They were thencentrifuged, washed in distilled water, and redistrib-uted in fresh WCLM medium plus the labeled supple-ments. Samples were removed hourly from tubessupplemented with radioactive ethionine to ascertainthe accumulation kinetics of the analogue.The extraction series employing trichloroacetic acid
included the following steps. The cold 5% trichloro-acetic acid soluble fraction was extracted according tothe technique of Kemnpner and Cowie (2). This wasfollowed by extraction of the ethyl alcohol and hot10% trichloroacetic acid-soluble fractions accordingto the procedures of Schneider (11). Proteins wereextracted with 1.0 N NaOH and assayed by the methodof Lowry et al. (3). The content of the cold acid-soluble fraction was investigated by chromatographyafter extraction of the trichloroacetic acid with diethylether. The sterols were isolated from a duplicate set ofsamples according to the technique of Starr and Parks(14). The techniques for the preparation of neutralized1.5 N perchloric acid (PCA) extracts were describedpreviously (12). Radioactivity was assayed in aPackard Tri-Carb liquid scintillation spectrometer.
Chromatography. The methods of paper chromatog-raphy have been described (12). Whatman no. 1
paper was used in all cases in a solvent system of 60%n-butanol-15% acetic acid (v/v). The components ofthe cold trichloroacetic acid-soluble fractions werealso separated by high-voltage paper electrophoresis(200 ma, 2,000 v, 4 C, for 1 hr). The identification ofadenosylmethionine, adenosylethionine, methylthio-adenosine, ethylthioadenosine, ethionine, and methio-nine was confirmed by co-chromatography with sam-ples of the pure compounds. The presence of radio-active adenosylmethionine and adenosylethionine wasverified by hydrolysis at 100 C for 30 min at pH 2 to 4(7). Under these conditions, the radioactivity peaksshift from the sulfonium compounds to the hydrolysisproducts (7). Radioactive compounds were detectedon chromatograms with a Packard 7200 paper stripscanner or the compounds were eluted and assayed ina Packard Tri-Carb liquid scintillation spectrometer.
Genetic studies. Techniques for mating throughscoring of ascospores were the same as those reportedby Sorsoli et al. (13).
RESULTS
Genetics and inhibition. To elucidate the geneticcharacter of the isolate, the haploid ethionine-resistant strain R3720 (etr2) was mated with a
clone of complementary mating type which wasethionine-sensitive (etP). Strain 3701B (et) wasused in a control mating to give etl/et'. Theheterozygous and homozygous diploids weretested for analogue resistance. In this manner,the dominance or recessiveness of the mutationwith respect to ethionine could be determined. Inaddition, p-fluorophenylalanine inhibition was
monitored to investigate the possibility that themutation etr2 might reflect a general amino acidanalogue resistance rather than specific resistanceto ethionine alone. These results are shown inTable 1. The capacity for ethionine resistance isretained in the heterozygous diploid, ettm/etr2. Thisprotection does not extend to the unrelatedphenylalanine analogue, p-fluorophenylalanine.A 2:2 ratio for ethionine resistance in the heter-ozygous diploid was obtained in all scored asci,indicating that this is a single chromosomal al-teration.Net accumulation of ethionine-ethyl-1-14C. The
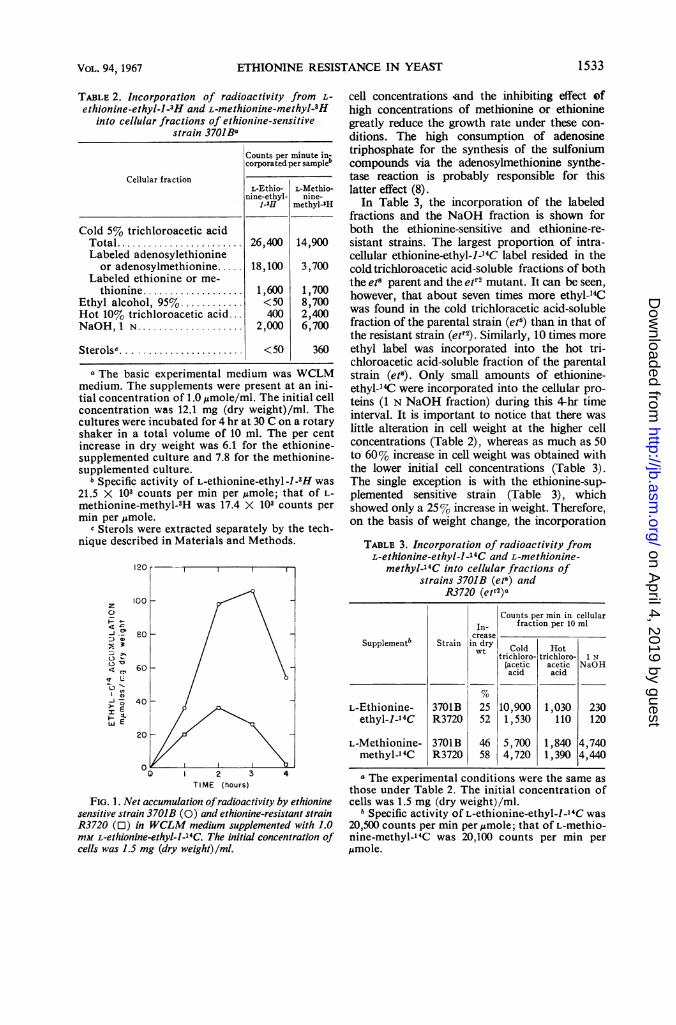
accumulation of exogenous ethionine-ethyl-1-14Cby both the et" and etr2 strains is shown in Fig. 1.After the 2nd hr, mutant R3720 had begun re-leasing ethyl-'4C label faster than it was beingaccumulated. This state in the parental strain,3701B, was delayed until the 3rd hr. By the end ofthe 4th hr, nearly all accumulated ethyl-14C hadreturned to the medium from the mutant, whereasrelatively large amounts still remained in theparental strain.
Intracellular distribution of radioactivity. Anexamination of the chemical fractions of the non-mutant parental strain, after growth in the pres-ence of equimolar amounts of ethionine-ethyl-l-3H and methionine-methyl-3H, showed that theradioactivity of the ethionine was incorporatedinto all except the ethyl alcohol and sterol frac-tions (Table 2). Detailed analysis of the cold 5%trichloroacetic acid-tractable materials showedthat most of the radioactivity in this pool, afterradioactive ethionine supplementation, was inadenosylethionine. With methionine-methyl-8H,a smaller proportion of the total counts extractedwith cold 5% trichloroacetic acid was in adenosyl-methionine. It should be pointed out that high
TABLE 1. Growth inhibition andgenetic character ofthe etr2 strain
Growtha Asci ratios(e$r2: el")Strain Markers .
3701B a ura eta + _ _R3720 a ura etrs + +3701B-ET48 a ura et' + -
a etaR3720-ET48 a ura etr2/ + + 14 0 0
ai et"
a The experiments were conducted on solid WMmedium. The cultures were incubated at 30 C for48 hr. Abbreviations: NS, no supplement; ETH,25 gg of DL-ethionine per ml; p-FPA, 50 ,ug of DL-p-fluorophenylalanine per ml.
1532 J. BAC-rERioL.
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
ETHIONINE RESISTANCE IN YEAST
TABLE 2. Incorporation of radioactivity from L-ethionine-ethyl-1-3H and L-methionine-methyl-3H
into cellular fractions of ethionine-sensitivestrain 3701Ba
Counts per minute in-corporated per sampleb
Cellular fractionL-EthiO- L-Methio-
nine-ethyl- nine-1-3H methyl-5H
Cold 5% trichloroacetic acidTotal .................. 26,400 14,900Labeled adenosylethionine
or adenosylmethionine..... 18,100 3,700Labeled ethionine or me-
thionine .................. 1,600 1,700Ethyl alcohol, 95% ............ < 50 8,700Hot 10% trichloroacetic acid 400 2,400NaOH, 1 N .................... 2,000 6, 700
Sterols ....................... <50 360
aThe basic experimental medium was WCLM
medium. The supplements were present at an ini-tial concentration of 1.0 Amole/ml. The initial cellconcentration was 12.1 mg (dry weight)/ml. Thecultures were incubated for 4 hr at 30 C on a rotaryshaker in a total volume of 10 ml. The per centincrease in dry weight was 6.1 for the ethionine-supplemented culture and 7.8 for the methionine-supplemented culture.
bSpecific activity of L-ethionine-ethyl-1-3H was21.5 X 103 counts per min per umole; that of L-
methionine-methyl-3H was 17.4 X 10' counts permin per Mumole.
c Sterols were extracted separately by the tech-nique described in Materials and Methods.
120 _
100z
4~-
60
< rn 40
>
EA
0
0 2 3 4TIME (hours)
FIG. 1. Net accumulation ofradioactivity by ethioninesensitive strain 3701B (0) and ethionine-resistant strainR3720 (O) in WCLM medium supplemented with 1.0mM L-ethionine-ethyl-J-14C. The initial concentration ofcells was 1.5 mg (dry weight) lml.
cell concentrations and the inhibiting effect ofhigh concentrations of methionine or ethioninegreatly reduce the growth rate under these con-ditions. The high consumption of adenosinetriphosphate for the synthesis of the sulfoniumcompounds via the adenosylmethionine synthe-tase reaction is probably responsible for thislatter effect (8).
In Table 3, the incorporation of the labeledfractions and the NaOH fraction is shown forboth the ethionine-sensitive and ethionine-re-sistant strains. The largest proportion of intra-cellular ethionine-ethyl-1-'4C label resided in thecold trichloroacetic acid-soluble fractions of boththe et" parent and the etr2 mutant. It can be seen,however, that about seven times more ethyl-'4Cwas found in the cold trichloracetic acid-solublefraction of the parental strain (etB) than in that ofthe resistant strain (etr2). Similarly, 10 times moreethyl label was incorporated into the hot tri-chloroacetic acid-soluble fraction of the parentalstrain (et"). Only small amounts of ethionine-ethyl-'4C were incorporated into the cellular pro-teins (1 N NaOH fraction) during this 4-hr timeinterval. It is important to notice that there waslittle alteration in cell weight at the higher cellconcentrations (Table 2), whereas as much as 50to 60% increase in cell weight was obtained withthe lower initial cell concentrations (Table 3).The single exception is with the ethionine-sup-plemented sensitive strain (Table 3), whichshowed only a 25% increase in weight. Therefore,on the basis of weight change, the incorporation
TABLE 3. Incorporation of radioactivity fromL-ethionine-ethyl-J-'4C and L-methionine-
methyl-14C into cellular fractions ofstrains 3701B (et") and
R3720 (etra) a
Counts per min in cellularIn- fraction per 10 ml
creaseSupplementb Strain in dry Cold Hot
wt trichloro- trichloro- 1 N(acetic acetic NaOHacid acid
L-Ethionine- 3701B 25 10,900 1,030 230ethyl-1-'4C R3720 52 1,530 110 120
L-Methionine- 3701B 46 5,700 1,840 4,740methyl-l 4C R3720 58 4,720 1,390 4,440
a The experimental conditions were the same asthose under Table 2. The initial concentration ofcells was 1.5 mg (dry weight)/ml.
b Specific activity of L-ethionine-ethyl-l-_4C was20,500 counts per min per jAmole; that of L-methio-nine-methyl-_4C was 20,100 counts per min per,Amole.
1533VOL. 94, 1967
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
SPENCE, PARKS, AND SHAPIRO
of radioactivity from ethionine-ethyl-1-14C is fourtimes greater in the NaOH and 20 times greaterin the hot trichloroacetic acid fraction of the etstrain as compared with the etr2 strain. With thenatural amino acid, methionine, incorporationof label into both trichloroacetic acid fractions ofthe mutant was much closer to those values shownby the parental strain (Table 3). Incorporationof methionine-methyl-'4C into the protein frac-tions (1 N NaOH) was the same for both strains.
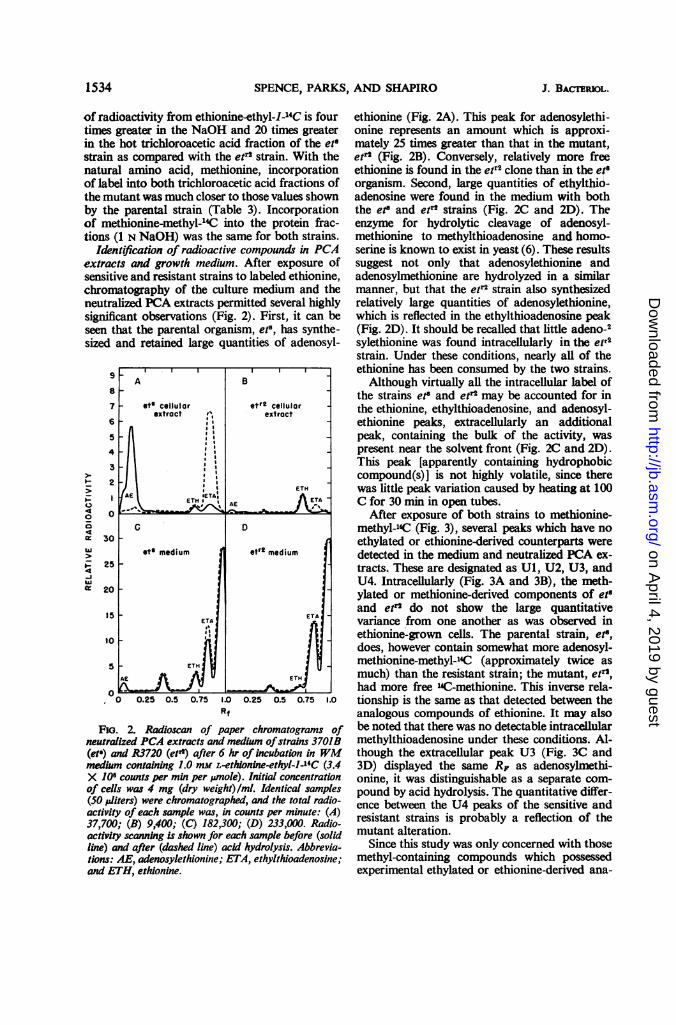
Identification of radioactive compounds in PCAextracts and growth medium. After exposure ofsensitive and resistant strains to labeled ethionine,chromatography of the culture medium and theneutralized PCA extracts permitted several highlysignificant observations (Fig. 2). First, it can beseen that the parental organism, et8, has synthe-sized and retained large quantities of adenosyl-
9
8
7
6
5
4
3
'- 2
I
4 0
4
t 30w
F 25-4-J
w
w- 20
15
10
5
,) E0 0.25 0.5 0.75 1.0 0.25 0.5 0.75 1.0
RF
FIG. 2. Radioscan of paper chromatograms ofneutralized PCA extracts and medium ofstrains 3701B(eta) and R3720 (etr) after 6 hr of incubation in WMmedium containing 1.0 mM L-ethionine-ethyl-l-'4C (3.4X 101 counts per min per unole). Initial concentrationof cells was 4 mg (dry weight)/ml. Identical samples(50 i,uiters) were chromatographed, and the total radio-activity of each sample was, in counts per minute: (A)37,700; (B) 9,400; (C) 182,300; (D) 233,000. Radio-activity scanning is shown for each sample before (solidline) and after (dashed line) acid hydrolysis. Abbrevia-tions: AE, adenosylethionine; ETA, ethylthioadenosine;and ETH, ethionine.
ethionine (Fig. 2A). This peak for adenosylethi-onine represents an amount which is approxi-mately 25 times greater than that in the mutant,etr2 (Fig. 2B). Conversely, relatively more freeethionine is found in the etr2 clone than in the et"organism. Second, large quantities of ethylthio-adenosine were found in the medium with boththe et and et"' strains (Fig. 2C and 2D). Theenzyme for hydrolytic cleavage of adenosyl-methionine to methylthioadenosine and homo-serine is known to exist in yeast (6). These resultssuggest not only that adenosylethionine andadenosylmethionine are hydrolyzed in a similarmanner, but that the etr2 strain also synthesizedrelatively large quantities of adenosylethionine,which is reflected in the ethylthioadenosine peak(Fig. 2D). It should be recalled that little adeno-2sylethionine was found intracellularly in the etr2strain. Under these conditions, nearly all of theethionine has been consumed by the two strains.
Although virtually all the intracellular label ofthe strains et" and etr2 may be accounted for inthe ethionine, ethylthioadenosine, and adenosyl-ethionine peaks, extracellularly an additionalpeak, containing the bulk of the activity, waspresent near the solvent front (Fig. 2C and 2D).This peak [apparently containing hydrophobiccompound(s)] is not highly volatile, since therewas little peak variation caused by heating at 100C for 30 min in open tubes.
After exposure of both strains to methionine-methyl-14C (Fig. 3), several peaks which have noethylated or ethionine-derived counterparts weredetected in the medium and neutralized PCA ex-tracts. These are designated as Ul, U2, U3, andU4. Intracellularly (Fig. 3A and 3B), the meth-ylated or methionine-derived components of ePand etr2 do not show the large quantitativevariance from one another as was observed inethionine-grown cells. The parental strain, et,does, however contain somewhat more adenosyl-methionine-methyl-14C (approximately twice asmuch) than the resistant strain; the mutant, et",had more free 14C-methionine. This inverse rela-tionship is the same as that detected between theanalogous compounds of ethionine. It may alsobe noted that there was no detectable intracellularmethylthioadenosine under these conditions. Al-though the extracellular peak U3 (Fig. 3C and3D) displayed the same RF as adenosylmethi-onine, it was distinguishable as a separate com-pound by acid hydrolysis. The quantitative differ-ence between the U4 peaks of the sensitive andresistant strains is probably a reflection of themutant alteration.
Since this study was only concerned with thosemethyl-containing compounds which possessedexperimental ethylated or ethionine-derived ana-
1534 J. BACTER1L.
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

ETHIONINE RESISTANCE IN YEAST
4
0
a4
I--
4
-j
8
7
6
5
4
3
2
0
35
30
25
20
5 I- U3
1U3 U4U
MTA MTA
0 0.25 0.5 0.75 1.0 0.25 0.5 0.75 1.0RF
FIG. 3. Radioscan of paper chromatograms ofneutralized PCA extracts and medium ofstrains 3701B(eta) and R3720 (etr') after 6 hr of incubation in WMmedium containing 1.0 mMu L-methionine-methyl-14C(2.4 X 106 counts per min per pAmole). Initial concentra-tion ofcells was 4 mg (dry weight) /ml. Identical samples(50 iditers) were chromatographed and the total radio-activity of each sample was, in counts per minute:(A) 22,600; (B) 19,300; (C) 136,000; and (D) 117,400.Radioactivity scanning is shown for each sample before(solid line) and (where there was alteration) after(dashed line) acid hydrolysis. Abbreviations: AM,adenosylmethionine; MTA, methylthioadenosine; MET,methionine; and U, unknown.
logues, methylated or methionine-derived constit-uents which had no such counterparts were notthoroughly investigated. It is important to see,however, that such compounds are synthesizedunder these conditions, although the ethionine-sensitive strain possessed sufficient selectivity toprevent the formation of their ethylated analogues(at least in detectable quantities) during the timeinterval employed.
DIscussIoN
Since the structures of methionine and ethi-onine differ only in the methyl or ethyl substit-uents, the growth inhibitory effects of ethioninemust be due to the organism's inability to sub-stitute ethyl groups for methyl groups in certain
critical reactions. Therefore, resistance to ethi-onine in yeast may be achieved either throughtotal exclusion of ethionine from the metabolismof the organism or through greater discriminationagainst ethyl over methyl groups in various in-dispensable cellular functions.The genetic and inhibition studies with the de-
scribed etr2 strain showed this organism to bedifferent from the etr strain described in a previouspaper (13). The latter mutation was found to berecessive in a heterozygous diploid, ets/etr. It wasshown that the etr mutation resulted in a non-specific alteration in the capacity of the mutant toaccumulate compounds supplied in the medium.As a result, the haploid etr strain was also re-sistant to the inhibitory effects of p-fluorophenyl-alanine. In contrast, the mutant (etr2) describedin this report, was sensitive to this phenylalanineanalogue. Unlike the etr mutation, the heter-ozygous diploid of etr2 with the wild-type allelewas resistant to ethionine. The segregation char-acteristics of the two markers were both 2:2 withrespect to et.These data suggest that the resistance of the
etr2 strain is due to a single chromosomal altera-tion which results in a specific reversal of the in-hibition by ethionine. The data show that thereis little, if any, alteration in the capacity of theetr2 mutant to accumulate either methionine orethionine, allowing one to conclude that this isnot a factor in etr2 resistance. This, of course,eliminates the possibility that ethionine is selec-tively excluded from the mutant cell and is con-sistent with the idea presented in previous reportswhich have supported the idea of nonspecificityin the amino acid concentrating system in S.cerevisiae (13). After the analogue enters the cellsof the et" or etr2 strains, it is quickly converted toother compounds. This would suggest thatethionine, as it arrives from the medium, wouldhave no more than a transient competitive effecton protein synthesis. This does not rule out thecontinuous regeneration of the analogue viatransethylation with adenosylethionine. Theethyl sulfonium derivative has been shown tofunction in a manner similar to adenosylmethi-onine when substituted in reaction mixtures con-taining the adenosylmethionine:homocysteinemethyltransferase and the appropriate methlyacceptor, homocysteine (7). One would expectethionine to be regenerated in this manner.Accumulation results suggested that the mecha-
nism of ethionine resistance involved degradationof ethionine to some product which was eitherinnocuous or could be easily released from thecell (Fig. 1). It was shown that the ethyl-14C ofthe labeled ethionine supplement was retained bythe et8 strain in larger quantities within the ex-
1535VOL. 94, 1967
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

SPENCE, PARKS, AND SHAPIRO
perimental interval than by the etr2 mutant. Fromthe data obtained after chromatographic separa-tions, it was seen that the ethyl-P4C returning tothe medium was not ethionine but rather productscontaining this label. In fact, in both strains,nearly all the ethionine (intracellular and extra-cellular) disappeared after the 6-hr incubationinterval (Fig. 2). The most obvious difference be-tween the strains was discovered in the adeno-sylethionine peaks. The ability of S. cerevisiae toproduce large quantities of adenosylethionine iswell known (7). The difference between the abilityof et" and etr2 strains to maintain this sulfoniumcompound was large. The amount of '4C-adeno-sylethionine produced and maintained by theparental et8 was approximately 25 times greaterthat by the etr2 mutant. Two mechanisms areknown which yield methylthioadenosine (andpresumably ethylthioadenosine), and in bothcases the sulfonium compound is a precursor (9).The production of large quantities of ethylthio-adenosine by the etr2 strain indicates that thisorganism retains the ability to produce consider-able amounts of adenosylethionine.The nature of the ethyl-'4C distribution in the
intracellular and extracellular components maybe explained in two general ways. (i) ethionine isis converted to adenosylethionine which ishydrolyzed to ethylthioadenosine, and furtherdegraded or converted to the "solvent front"compound, or (ii) ethionine is converted toadenosylethionine, then ethylthioadenosine (asabove), but, in addition, the amino acid under-goes other reactions(s) leading to the "solventfront" component.The production of the "solvent front" compo-
nent and ethylthioadenosine are abilities whichare shared by both the et" and etr2 strains. Thefact that there is more adenosylethionine accumu-lated intracellularly by the ets strain, and lessextracellular ethylthioadenosine and "solventfront" compound, suggests that inhibition byethionine results from the inability of the ets strainto degrade adenosylethionine fast enough to ex-clude it from participating in one or moretransalkylation reactions. Conversely, one mustassume that the etr2 mutant has an increasedability to degrade adenosylethionine. It is stillpossible that some ethionine may be "detoxified"by reactions not involving adenosylethionine andethylthioadenosine as intermediates. In eithercase, there is a much higher level of adenosyl-ethionine in the eti9 cells, and, as the ethyl-'4Cdistribution experiments suggest (Tables 2 and 3),this undoubtedly results in the ethylation ofnormally methylated cellular components. Theabnormal ethylation of many cellular componentshas been reported (4, 15, 16).
For comparison with ethionine, the incorpora-tion of methionine-methyl-'4C was also mon-itored. It was seen that both the et" and etr2 strainsshowed similar distribution of the methyl-'4C. Inmost instances, however, incorporation or poolsizes, or both, were somewhat lower in the mutant(etrl). This shows that the mutation results in anincreased capacity for converting methionine (asethionine) to compounds which are not main-tained intracellularly. Furthermore, since themutation is donminant, it seems apparent that theetr2 mutation has resulted in an increased level ofcertain degradative enzymes rather than in a lossof enzyme function.The possibility that the resistant strain over-
comes ethionine inhibition by production of ab-normally large quantities of methionine has alsobeen considered. A number of experiments havebeen performed to investigate this possibility,but the results have not supported this idea. Forexample, two of three different etr2 mutants testedshow slightly increased levels of intracellularmethionine in unsupplemented synthetic media.One mutant possessed the same level as theparental strain, 3701B. In all instances, theselevels were low (2 to 4 m,umoles of methionine/mg of dry weight). Dilution of radioactivity byinternally produced methionine in the etr2 strainmight be expected to produce the effects shown inTable 2. Large quantities of internally synthesizedmethionine, if not present as the amino acid,must then either exist in the form of adenosyl-methionine or as the breakdown products ofmethionine or adenosylmethionine. Attemptshave failed to show any marked difference be-tween the parental et" and the mutant etr2 eitherin their rate of converting methionine to non-methionine compounds or in their levels ofadenosylmethionine synthetase (K. D. Spence,Ph.D. Thesis, Oregon State Univ., Corvallis,1965), an enzyme which is induced by methionine(8). These observations further support the ideathat the mutation alters the fate of the methionineor the methionine derivatives rather than increas-ing the synthesis of methionine.
ACKNOWLEDGMENTS
The authors thank D. J. Ehninger for her assistanceand Fritz Schlenk for his very helpful discussions.
This work was initiated during the tenure of U.S.Public Health Predoctoral Fellowship 5-Fl-GM-11,204-02 to K. D. Spence, and was supported in part byPublic Health Service Grant GM-06632 and by theU.S. Atomic Energy Commission.
LrrERATURE CITED1. KAPPY, M. S., AND R. L. METZENBERG. 1965.
Studies on the basis of ethionine-resistance in
1536 J. BACTERIOL.
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

ETHIONINE RESISTANCE IN YEAST
Neurospura. Biochim. Biophys. Acta 107:425-433.
2. KEmPNER, E. S., AND D. B. CowiE. 1960. Metabolicpools and the utilization of amino acid analogsfor protein synthesis. Biochim. Biophys. Acta42:401-408.
3. LOWRY, 0. H., N. J. ROSEBROUGH, A. L. FARR, ANDR. J. RANDALL. 1951. Protein measurementwith the Folin phenol reagent. J. Biol. Chem.193:265-275.
4. MAW, G. A. 1966. Incorporation and distributionof ethionine-sulfur in the protein of ethionine-sensitive and ethionine-resistant yeasts. Arch.Biochem. Biophys. 115:291-301.
5. METZENBERG, R. L., M. S. KAPPY, AND J. W.PARSONS. 1964. Irreparable mutations andethionine resistance in Neurospora. Science145:1434-1435.
6. MUDD, S. H. 1959. Enzymatic cleavage of S-adenosylmethionine. J. Biol. Chem. 234:87-92.
7. PARKS, L. W. 1958. S-adenosylethionine andethionine inhibition. J. Biol. Chem. 232:169-176.
8. PIGG, C. J., W. A. SORSOLI, AND L. W. PARKS.1964. Induction of the methionine-activatingenzyme in Saccharomyces cerevisiae. J. Bac-teriol. 87:920-923.
9. SCHLENK, F. 1965. Biochemical and cytologicalstudies with sulfonium compounds, p. 48-70.In S. K. Shapiro and F. Schlenk [ed.], Trans-methylation and methionine biosynthesis.Univ. of Chicago Press, Chicago.
10. SCHLENK, F., C. R. ZYDEK, D. J. EHNINGER, ANDJ. L. DAINKO. 1965. The production of S-adenosyl-L-methionine and S-adenosyl-L-ethio-nine by yeast. Enzymologia 29:283-298.
11. SCHNEIDER, W. C. 1945. Phosphorus compoundsin animal tissues. I. Extraction and estimationof desoxypentose nucleic acid and of pentosenucleic acid. J. Biol. Chem. 161:293-303.
12. SHAPIRO, S. K., AND D. J. EHNINGER. 1966.Methods for the analysis and preparation ofadenosylmethionine and adenosylhomocysteine.Anal. Biochem. 15:323-333.
13. SORSOLI, W. A., K. D. SPENCE, AND L. W. PARKS.1964. Amino acid accumulation in ethionine-resistant Saccharomyces cerevisiae. J. Bacteriol.88:20-24.
14. STARR, P. R., AND L. W. PARKS. 1962. Effect oftemperature on sterol metabolism in yeast. J.Cellular Comp. Physiol. 59:107-110.
15. STEKOL, J. A. 1963. Biochemical basis for ethio-nine effects on tissues. Advan. Enzymol. 25 :369-393.
16. STEKOL, J. A. 1965. Formation and metabolism ofS-adenosyl derivatives of S-alkylhomocysteinsin the rat and mouse, p. 231-252. In S. K.Shapiro and F. Schlenk [ed.], Transmethylationand methionine biosynthesis. Univ. of ChicagoPress, Chicago.
17. SURDIN, Y., W. SLY, J. SIRE, A. M. BORDES, ANDH. DE ROBICHON-SZULMAJSTER. 1965. Pro-prietes et controle genetique du system d'accumulation des acides amines chez Saccharo-myces cerevisiae. Biochim. Biophys. Acta 107:546-566.
18. WICKERHAM, J. L. 1946. A critical evaluation ofthe nitrogen assimilation tests commonly usedin the classification of yeasts. J. Bacteriol. 52:293-301.
1537VOL. 94, 1967
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from




![First IKBKG Gene Mutation Study in Serbian Incontinentia ... · Incontinentia pigmenti (IP; Bloch-Sulzberg-er syndrome; MIM 308300) is a rare X-linked dominant genodermatosis [5]](https://img.pdfslide.net/doc/110x75/5f3bedf5651a4c1377610355/first-ikbkg-gene-mutation-study-in-serbian-incontinentia-incontinentia-pigmenti.jpg)
























