Embed Size (px)
Citation preview

JouRNAL OF BACTERIOLOGY, Sept. 1972, p. 649-657Copyright i 1972 American Society for Microbiology
Vol. 111, No. 3Printed in U.SA.
Biochemical and Genetic Aspects of NystatinResistance in Saccharomyces cerevisiae
MARTIN BARD
Department of Genetics, University of Sheffield, Sheffield, Great Britain
Received for publication 3 April 1972
Two phenotypically distinct sets of nystatin-resistant mutants were investi-gated. One set is resistant, respiratory competent, and requires no lipid forgrowth. The other set is more resistant, respiratory deficient, and lipid re-
quiring (unsaturated fatty acid or sterol). Both sets show altered sterol compo-sition as demonstrated by the Liebermann-Burchard colorimetric reaction, ul-traviolet spectrophotometry, and gas-liquid chromatography. Genetic analysisindicates that all nystatin-resistant mutants can be placed into one of six dis-tinct genetic groups. The phenotype's nystatin resistance, lipid requirement,and respiratory deficiency are recessive. There was one case of allelism formutants from different sets. Revertants of mutants which have the tripartitephenotype retain a residual level of nystatin resistance, but they are no longerlipid requiring or respiratory deficient. Growth studies in mutants which havethe tripartite phenotype reveal that the addition of ergosterol to the growthmedium results in decreased resistance to nystatin.
A relationship between the polyene anti-biotic, nystatin, and sterols in the cell mem-brane of sensitive organisms has been demon-strated by a number of investigators. For twoexcellent reviews on permeability alterationsand resulting cell death due to the administra-tion of this antibiotic see Lampen (15) andKinsky (13).The ready availability of nystatin-resistant
mutants in the yeast Saccharomyces cerevisiaeafforded an opportunity to correlate geneticchanges with alterations in sterol composition.In addition, a combined biochemical geneticapproach offered opportunities to observephenomena closely related to sterol productionsuch as mitochondrial formation and lipidmetabolism.Ahmed and Woods (2) identified three nys-
tatin-resistant genes. These are recessive andsubject to enhanced resistance by three domi-nant modifier genes. Some of these mutants donot synthesize ergosterol and/or an additionalsterol, 24(28)-dehydroergosterol (24). Resnickand Mortimer (20) reported a class of yeastlipid mutants which require the addition ofeither an unsaturated fatty acid or ergosterolto yeast extract-peptone-dextrose (YEPD)medium. These mutants, now designated ole2,ole3, and ole4, were also petite (respiratorydeficient), and preliminary genetic analysis
demonstrated that both the lipid requirementand petite phenotype segregated together (12).The findings that the ole2, 3, and 4 mutantsare also highly resistant to nystatin and thatall three phenotypes segregate together is animportant aspect of this investigation andstrengthens the evidence for the role of lipidsin the function and biogenesis of mitochondria.
Several investigators have observed relation-ships between lipogenesis and the productionof functional mitochondria in yeast. Sarachek(21), using a pantothenate-deficient strain,demonstrated that during the period of transi-tion from the logarithmic to the stationaryphase, cells grown in the absence of pantothen-ate, a metabolite essential in lipogenesis, be-come respiratory deficient. The addition ofexogenous acetate or unsaturated fatty acidsprevented the occurrence of respiratory defi-ciency, whereas the addition of saturated fattyacids and sterols did not. Parks and Starr (18)demonstrated that raising the temperature ofgrowing cells to 42 C, a condition conducive toa sharp reduction in sterol production, coin-cided with petite induction. Supplementationof the growth medium with ergosterol at ele-vated temperatures resulted in the inductionof only one-half as many petites.
This report is concerned with some of therelationships discussed above. In particular,
649
on October 3, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
on October 3, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
on October 3, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

J. BACTERIOL.
correlations are made between nystatin resist-ance and sterol pattems, and between sterolcomposition, lipid requirement, and respira-tory deficiency in mutant strains of yeast.
MATERIALS AND METHODSOrganisms. A wild-type strain of S. cerevisiae,
S288C, obtained from the Berkeley collection wasthe parent strain of all mutants used in this investi-gation. Nystatin-resistant mutants nyrl, 2, 5, 6, and15, as well as Resnick and Mortimer's (20) olel, 2, 3,and 4 mutants, were employed. In view of the exist-ence of mutants which are phenotypically similar tothe nyr set (2), no attempt is made to appropriategenetic symbols to this set. nyr mutants are referredto by their isolation numbers. This is not the case forthe ole set, wherein the numeral after the ole symboldoes refer to a genetic locus.
Because of the time lapse between the isolation ofthe nyr and ole sets (3 years), another wild-typestrain, X2180, derived as an "a" mating type segre-gant from a diploidized S288C culture was used asan internal control. This was necessary because dif-ferent wild-type S. cereuisiae strains differ substan-tially in amounts of ergosterol synthesized. Allgrowth was at 30 C.Media and genetic analysis. A YEPD culture
medium of 1% yeast extract, 2% peptone, 2% dex-trose, and 2% agar (solid) was the standard culturemedium with 1% Tween 80 (a polyoxyethylene deriv-ative of sorbitan monoleate) added as the source ofunsaturated fatty acid for olel, 2, 3, 4 mutants. Er-gosterol (Nutritional Biochemicals Corp., Cleveland,Ohio) was solubilized by adding 1.5% Tergitol (v/v,Union Carbide Institute, W. Va.) a nonionic deter-gent free of lipid. Various concentrations of thissterol from 10' M to 10-' M (solid) and 5 x 10-' Mto 7.5 x 10-6 M (liquid) were employed. An aqueousnystatin suspension (Grand Island Biological Co.,Long Island, N.Y.) was added to solid medium be-fore pouring at 49 C or less.
Minimal medium contained 0.67% yeast nitrogenbase (Difco), 2% dextrose, 1% Tween 80, and 2%agar. For the testing of petites, 0.5% ethanol re-placed dextrose as the carbon source.
Hybridization was accomplished by mixing hap-loids of opposite mating types on YEPD plus Tween80 medium, and after 2 to 3 hr zygotes were isolatedby micromanipulation. After 24 hr of growth onYEPD plus Tween 80, diploids were sporulated bytransfer to a medium composed of 2% potassiumacetate, 0.22% yeast extract, 0.05% glucose, 2% agar,and the following additions per liter: L-leucine, 6mg; L-threonine, 300 mg; L-tryptophan, 2 mg; L-methionine, L-histidine, L-arginine, adenine, anduracil, 1 mg each; and L-lysine, 20 mg. After 3 to 5days of incubation, the cultures were examined forasci, and if well formed asci were abundant the cellwalls were digested with a 1: 25 dilution of Glusulase(a digestive enzyme from the snail crop, Endo Labo-ratories, Garden City, N.Y.). Dissected asci weretested for levels of resistance, respiratory compe-tency, and lipid requirements by replica plating.
Nystatin resistance determinations. Plates con-taining 25 to 800 units (U) of nystatin per ml werescored 24 hr after replica plating from YEPD plus1% Tween 80 medium lacking the antibiotic. Plateswere again scored after 36 hr, particularly for theslower growing ole2, 3, and 4 mutants. Scoring wasas follows. The designation "+" signifies confluentgrowth on nystatin plates for at least 50% of the de-terminations. The latter qualification is especiallyimportant for plates containing 25 U of nystatin perml, the minimum amount of antibiotic used. Higherconcentrations of nystatin gave more consistent re-sults. The notation "±" refers to nonconfluentgrowth such that only single colonies are seen ratherthan a uniform layer of cells. This same notation isalso used for a uniform layer of cells growing moreslowly than the same strain on a plate having alower concentration of nystatin. The designation "-'"means that the strain is nystatin sensitive andfails to grow. Segregants from dissected asci wereroutinely tested on several plates with varying con-centrations of nystatin to establish th,e resistancelevel of each of the four ascosporal colonies.
Extraction of nonsaponifiables and sterols.Yeast cultures grown for 48 to 96 hr were pelletedand washed once with distilled water before beingacid-saponified for 1 hr at 30 C in 0.1 N HCl (1).After an additional washing, pellets were subjectedto base-saponification at reflux temperatures for 90min in 15% methanolic KOH. The nonsaponifiablefraction was extracted with petroleum ether (bp, 30-60 C) and analyzed for ergosterol by either the Lie-bermann-Burchard colorimetric reaction or ultravi-olet (UV) spectrophotometry. For gas-liquid chro-matographic (GLC) analysis of sterols, batch cul-tures of yeast were grown, pelleted, washed, and sub-jected to total lipid extraction by ovemight stirringin chloroform-methanol (2: 1). The lipid extract,approximately 250 ml for each sample, was vacuumdried on a rotary evaporator and saponified for 3 hrat reflux temperatures in Florence flasks containing72 ml of absolute ethanol, 24 ml of ethyl ether, and 4ml of 10 N KOH. The nonsaponifiable fraction wasextracted with petroleum ether.
Colorimetry. The Liebermann-Burchard colori-metric reaction to detect M-sterols was carried outessentially according to the procedure of Stadtman(22). Upon the addition of Liebermann-Burchardreagent, lanosterol a Al-sterol, gives a clear yellowcolor (10).UV spectrophotometry. UV spectrophotometry
of ergosterol in absolute ethanol gives four absorptionbands at 262, 271, 282, and 293.5 nm (8). The pres-ence of ergosterol in each sample was determined bydissolving the nonsaponifiable fractions in absoluteethanol and scanning from 300 to 200 nm, with aBeckman DB spectrophotometer.
Chromatography. Preparative thin-layer chroma-tography for GLC analysis was accomplished byapplying nonsaponifiable samples to Silica Gel-Gplates activated at 110 C for 1 hr before use. Ergos-terol and lanosterol (Mann Research, New York,N.Y.) were applied to each plate as standards. Plateswere developed in a solvent system of hexane-di-
650 BARD
on October 3, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

NYSTATIN RESISTANCE IN S. CEREVISIAE
ethyl ether-acetic acid (80:20:1), and sterols weremade visible with an iodine (1% in methanol) spray.Sterols were eluted with hexane.GLC was performed on a Varian Aerograph model
1200. Two kinds of columns were employed. (i) Astainless-steel column (¼/8 inch by 6 ft) packed with3% neopentyl glycolsuccinate (NPGS) on 1"/%2O meshChromosorb W was operated at a column tempera-ture of 220 C and N, gas flow of 19.6 ml/min. (ii) Astainless-steel column (1/8 inch by 5 ft) packed withthe less polar 5% silicone gum rubber (SE-30) on 69%omesh Chromosorb W was operated at 225 C and N,gas flow of 32.4 ml/min. For both columns the de-tector temperature was 265 C, and the injector tem-perature was 255 C.
Silylation of sterols was accomplished by reactingbis(trimethylsilyl)trifluroacetamide (Regisil, RegisChemical, Chicago, Ill.) with sterol samples for 30min at 60 C in screw-cap test tubes in the dark.Columns were coated with silylating agent prior to
packing. Ergosterol purchased from Nutritional Bio-chemicals Corp. and Sigma Chemical Co. (St. Louis,Mo.) was used to determine a standard relative re-tention time. Both purchased products were isolatedfrom yeast. The retention times are relative to cho-lestane.Growth determination. Growth studies were car-
ried out with a Klett-Summerson photoelectric col-orimeter (red filter) in side-arm Erlenmeyer flasks.Doubling times were computed from areas of thegrowth curve of 30 to 100 Klett units, known to belinear with cell concentration (23).
RESULTS
Isolation and description of mutants. Awild-type strain of S. cerevisiae, S288C, wassubjected to 10 and 60 U of nystatin per ml onminimal medium plus 1% Tween 80 plates.Fifteen spontaneously produced nystatin-re-sistant mutants were isolated, five of whichwill be discussed in the present investigation(nyrl, 2, 5, 6 isolated as single colonies fromnystatin plates at 10 U/ml, and nyri5 isolatedas a single colony from a 60 U/ml plate).
Resnick and Mortimer (20) used a YEPDplus Tween 80 medium to isolate the morephenotypically complex ole mutants. The olemutants employed throughout this investiga-tion are ole2-1 (KD46), ole2-2 (KD91), and thedouble mutant ole3o0e4 (KD18). In addition tobeing nystatin resistant, these mutants arepetite and require a lipid supplement, unlikethe nyr mutants which have resistance as theirsole phenotype.
Revertants at the ole2 locus were easily de-tected by their morphological appearance onsolid medium. ole2 colonies are always smalland retain a white liquidy appearance even onaged plates; in contrast, grande revertant colo-nies are larger and after several days of growth
on solid medium appear dry and amber-col-ored.
Table 1 records the range of resistance levelsof the ole and nyr mutants, two grande non-lipid-requiring revertants of ole2 (ole2-JR andole2-2R), and two genetically related wild-typestrains, S288C and X2180. The resistancelevels of olel (KD20 and KD115) are alsolisted. olel-1 and olel-2 are two grande allelesof a fatty acid desaturase mutant (12).The higher resistance values reported in this
investigation are largely due to the presence ofTween 80 in the medium (2). Tween 80 is asource of oleic acid for the lipid-requiringmutants, and, as a detergent, it solubilizesnystatin. Concomitantly, Tween 80 decreasesthe interaction of the antibiotic with the cellsurface (13). Therefore, with detergent effectstaken into consideration, Table 1 reveals therelative levels of resistance of the mutants.The most resistant mutants are ole2-1, ole2-2,and the double mutant ole301e4 which cangrow on plates of 800 U of nystatin per ml.The remaining mutants in order of decreasingresistance are: nyrl and nyri5 greater thannyr2, nyr5, nyr6, ole2-JR, and ole2-2R. Wildtypes S288C and X2180 and the fatty aciddesaturase mutants, olel-1 and olel-2, are sen-sitive.
TABLE 1. Range of nystatin resistance levels ofmutant and wild-type Saccharomyces cerevisiae
Resistance"Straina - - __
25C 50 75 100 400 600 800
Wild-type strainsS288C + - -X2180 + - -
nyr mutantsnyrl + + + + + 4: 4
nyr2 + + + 4 -
nyr5 + + + 4 -
nyr6 + + + 4- -
nyrl5 + + + + + 4 4
ole mutantsole2-1 + + + + + + +ole2-2 + + + + + + +ole3ole4 + + + + + + +ole2-2R + + + -ole2-1R + + : -
olel-1 + - -olel-2 + - -
aRecorded 24 hr after replica plating to YEPDplus 1% Tween 80 plates containing nystatin.
b +, Confluent growth for at least 50% of the de-terminations; -, spotted colonies without conflu-ence, or appreciable thinning out of the cell plaque;-, no growth.
c Units of nystatin per milliliter.
VOL. 111, 1972 651
on October 3, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
J. BACTERIOL.
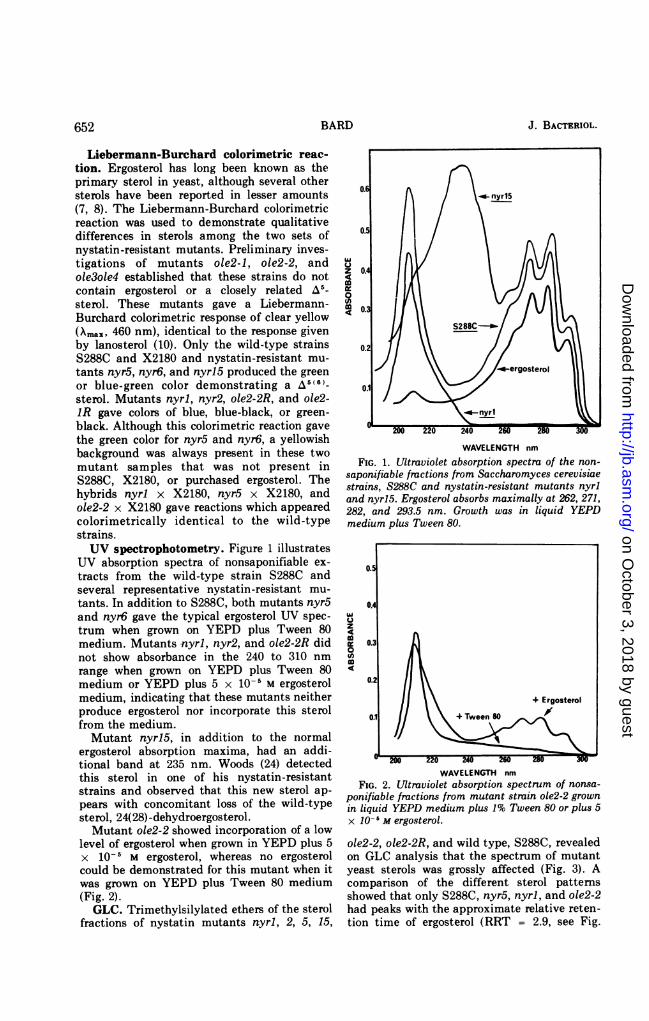
Liebermann-Burchard colorimetric reac-tion. Ergosterol has long been known as theprimary sterol in yeast, although several othersterols have been reported in lesser amounts(7, 8). The Liebermann-Burchard colorimetricreaction was used to demonstrate qualitativedifferences in sterols among the two sets ofnystatin-resistant mutants. Preliminary inves-tigations of mutants ole2-1, ole2-2, andole3ole4 established that these strains do notcontain ergosterol or a closely related A5-sterol. These mutants gave a Liebermann-Burchard colorimetric response of clear yellow(XA,, 460 nm), identical to the response givenby lanosterol (10). Only the wild-type strainsS288C and X2180 and nystatin-resistant mu-tants nyr5, nyr6, and nyrl5 produced the greenor blue-green color demonstrating a 5 (6)_sterol. Mutants nyrl, nyr2, ole2-2R, and ole2-IR gave colors of blue, blue-black, or green-black. Although this colorimetric reaction gavethe green color for nyr5 and nyr6, a yellowishbackground was always present in these twomutant samples that was not present inS288C, X2180, or purchased ergosterol. Thehybrids nyrl x X2180, nyr5 x X2180, andole2-2 x X2180 gave reactions which appearedcolorimetrically identical to the wild-typestrains.UV spectrophotometry. Figure 1 illustrates
UV absorption spectra of nonsaponifiable ex-tracts from the wild-type strain S288C andseveral representative nystatin-resistant mu-tants. In addition to S288C, both mutants nyr5and nyr6 gave the typical ergosterol UV spec-trum when grown on YEPD plus Tween 80medium. Mutants nyrl, nyr2, and ole2-2R didnot show absorbance in the 240 to 310 nmrange when grown on YEPD plus Tween 80medium or YEPD plus 5 x 10-5 M ergosterolmedium, indicating that these mutants neitherproduce ergosterol nor incorporate this sterolfrom the medium.Mutant nyr15, in addition to the normal
ergosterol absorption maxima, had an addi-tional band at 235 nm. Woods (24) detectedthis sterol in one of his nystatin-resistantstrains and observed that this new sterol ap-pears with concomitant loss of the wild-typesterol, 24(28)-dehydroergosterol.Mutant ole2-2 showed incorporation of a low
level of ergosterol when grown in YEPD plus 5x 10-5 M ergosterol, whereas no ergosterolcould be demonstrated for this mutant when itwas grown on YEPD plus Tween 80 medium(Fig. 2).GLC. Trimethylsilylated ethers of the sterol
fractions of nystatin mutants nyrl, 2, 5, 15,
uuz4c
4
WAVELENGTH nm
FIG. 1. Ultraviolet absorption spectra of the non-saponifiable fractions from Saccharomyces cerevisiaestrains, S288C and nystatin-resistant mutants nyrland nyr15. Ergosterol absorbs maximally at 262, 271,282, and 293.5 nm. Growth was in liquid YEPDmedium plus Tween 80.
O.
Luz49co0Wco
0.4
0.3
0.2
- 200 zzu 2w 0 LOU 3W0
WAVELENGTH nmFIG. 2. Ultraviolet absorption spectrum of nonsa-
ponifiable fractions from mutant strain ole2-2 grownin liquid YEPD medium plus 1% Tween 80 or plus 5x 10- I M ergosterol.
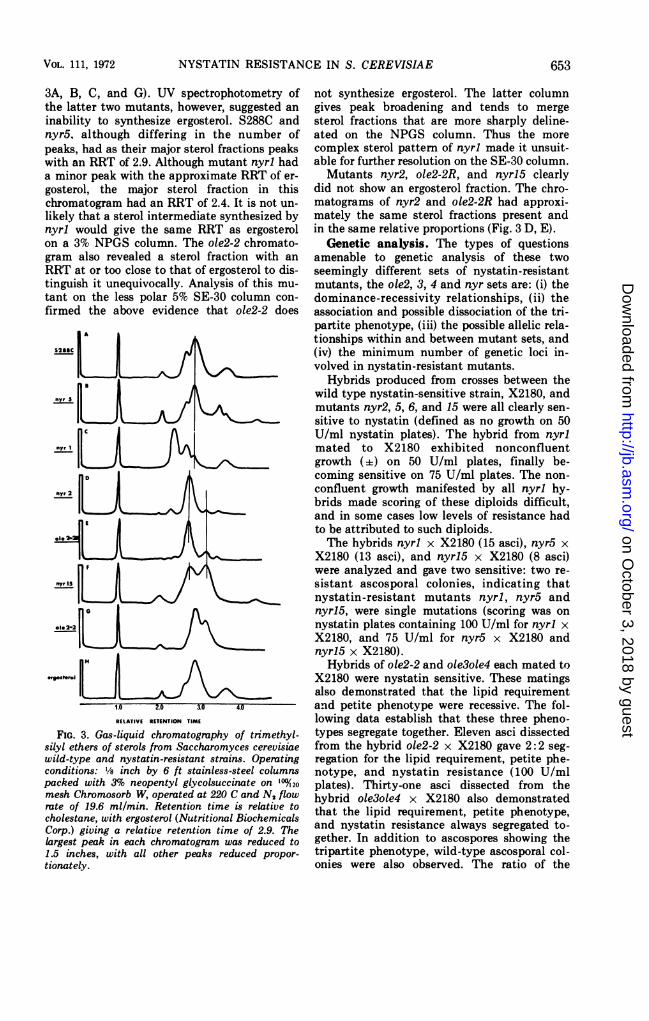
ole2-2, ole2-2R, and wild type, S288C, revealedon GLC analysis that the spectrum of mutantyeast sterols was grossly affected (Fig. 3). Acomparison of the different sterol patternsshowed that only S288C, nyr5, nyrl, and ole2-2had peaks with the approximate relative reten-tion time of ergosterol (RRT = 2.9, see Fig.
+ Ergosterol+TweenB
BARD652
on October 3, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
NYSTATIN RESISTANCE IN S. CEREVISIAE
3A, B, C, and G). UV spectrophotometry ofthe latter two mutants, however, suggested aninability to synthesize ergosterol. S288C andnyr5, although differing in the number ofpeaks, had as their major sterol fractions peakswith an RRT of 2.9. Although mutant nyrl hada minor peak with the approximate RRT of er-gosterol, the major sterol fraction in thischromatogram had an RRT of 2.4. It is not un-likely that a sterol intermediate synthesized bynyrl would give the same RRT as ergosterolon a 3% NPGS column. The ole2-2 chromato-gram also revealed a sterol fraction with anRRT at or too close to that of ergosterol to dis-tinguish it unequivocally. Analysis of this mu-tant on the less polar 5% SE-30 column con-firmed the above evidence that ole2-2 does
S2SSC LL
nyr 5
nyr 1
|
el*2-2
ergsterol
H
1.0 2.0 3.0 4.0
REtATIVE RETENTION TIME
FIG. 3. Gas-liquid chromatography of trimethyl-silyl ethers of sterols from Saccharomyces cerevisiaewild-type and nystatin-resistant strains. Operatingconditions: 1/8 inch by 6 ft stainless-steel columnspacked with 3%o neopentyl glycolsuccinate on 1'°20mesh Chromosorb W, operated at 220 C and N, flowrate of 19.6 ml/min. Retention time is relative tocholestane, with ergosterol (Nutritional BiochemicalsCorp.) giving a relative retention time of 2.9. Thelargest peak in each chromatogram was reduced to1.5 inches, with all other peaks reduced propor-tionately.
not synthesize ergosterol. The latter columngives peak broadening and tends to mergesterol fractions that are more sharply deline-ated on the NPGS column. Thus the morecomplex sterol pattem of nyrl made it unsuit-able for further resolution on the SE-30 column.Mutants nyr2, ole2-2R, and nyri5 clearly
did not show an ergosterol fraction. The chro-matograms of nyr2 and ole2-2R had approxi-mately the same sterol fractions present andin the same relative proportions (Fig. 3 D, E).
Genetic analysis. The types of questionsamenable to genetic analysis of these twoseemingly different sets of nystatin-resistantmutants, the ole2, 3, 4 and nyr sets are: (i) thedominance-recessivity relationships, (ii) theassociation and possible dissociation of the tri-partite phenotype, (iii) the possible allelic rela-tionships within and between mutant sets, and(iv) the minimum number of genetic loci in-volved in nystatin-resistant mutants.
Hybrids produced from crosses between thewild type nystatin-sensitive strain, X2180, andmutants nyr2, 5, 6, and 15 were all clearly sen-sitive to nystatin (defined as no growth on 50U/ml nystatin plates). The hybrid from nyrlmated to X2180 exhibited nonconfluentgrowth ( on 50 U/ml plates, finally be-coming sensitive on 75 U/ml plates. The non-confluent growth manifested by all nyrl hy-brids made scoring of these diploids difficult,and in some cases low levels of resistance hadto be attributed to such diploids.The hybrids nyrl x X2180 (15 asci), nyr5 x
X2180 (13 asci), and nyriS x X2180 (8 asci)were analyzed and gave two sensitive: two re-sistant ascosporal colonies, indicating thatnystatin-resistant mutants nyrl, nyrS andnyr15, were single mutations (scoring was onnystatin plates containing 100 U/ml for nyrl xX2180, and 75 U/ml for nyr5 x X2180 andnyrl5 x X2180).
Hybrids of ole2-2 and ole3ole4 each mated toX2180 were nystatin sensitive. These matingsalso demonstrated that the lipid requirementand petite phenotype were recessive. The fol-lowing data establish that these three pheno-types segregate together. Eleven asci dissectedfrom the hybrid ole2-2 x X2180 gave 2:2 seg-regation for the lipid requirement, petite phe-notype, and nystatin resistance (100 U/mlplates). Thirty-one asci dissected from thehybrid ole3o0e4 x X2180 also demonstratedthat the lipid requirement, petite phenotype,and nystatin resistance always segregated to-gether. In addition to ascospores showing thetripartite phenotype, wild-type ascosporal col-onies were also observed. The ratio of the
VOL. 111, 1972 653
on October 3, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

J. BACTERIOL.
tetrad classes PD: NPD:T (parental ditype:non-parental ditype: tetratype) was 7:3:21which corresponds to 2:2, 0:4, and 1: 3 (scoredas grande:petite, sensitive:resistant; the pe-tite phenotype was scored with and without1% Tween 80 supplementation). The abovedata fit the 1PD: lNPD: 4T distribution oftetrad classes for two unlinked genes, with atleast one of the genes showing no centromerelinkage (19), and demonstrate that the sepa-rable single genetic lesions ole3 and ole4 dis-play the tripartite phenotype. In the combined42 asci analyzed, no segregant showed disso-ciation of the tripartite phenotype. This ac-cords with observations by Resnick and Mor-timer (20) and Keith et al. (12), who also failedto detect dissociation of the lipid requirementand petite phenotype.Grande revertants of ole2-1 and ole2-2 were
isolated and found to be non-lipid-requiringand less nystatin resistant than the parentalstrains (Table 1). Phenotypically, the revertedole mutants resemble the nyr set. Their solephenotype is a level of resistance that is morecomparable to several less resistant nyr mu-
tants (nyr2, 5, 6) than the highly resistant ole2,3, 4 set.The hybrid ole2-2R (revertant) x X2180 is
nystatin sensitive, demonstrating that the re-sistance phenotype of the revertant is reces-sive. Twenty-three asci analyzed from thiscross yielded 2:2 segregation for sensitivity:resistance when scored on 100 U/ml nystatinplates. All segregants were grande and, in ad-dition, sensitive on plates containing 150 and300 U of nystatin per ml. The original ole2-2mutant was not recovered, indicating that therevertant phenotype results from a reversemutation at or within a few map units of theoriginal locus rather than an unlinked modifieror suppressor gene.
Resistance levels of pairwise combinations ofnystatin-resistant mutants are listed in Table2. These crosses assess the allelic relationshipsamong mutants and provide the basis for esti-mating the number of genetic loci involved.Diploids nyr2 x ole2-2, nyr2 x ole2-2R, ole2-2x ole2-2R, and nyr5 x nyr6 demonstratelevels of resistance indicating the absence ofcomplementation. Segregation analysis ofthese diploids confirm that mutants ole2, ole2-2R, and nyr2 are allelic as well as mutantsnyr5 and nyr6. The diploids nyr2 x ole2-2,nyr2 x ole2-2R, and ole2-2 x ole2-2R are nei-ther petite, lipid requiring or highly nystatinresistant; the lower resistance levels of nyr2and ole2-2R are dominant in crosses to ole2-2.The phenotypic relationships within this latter
group illustrate that not only is the entire tri-partite phenotype recessive but also, pheno-typically, a low level of resistance is dominantto a higher level.The ambiguous results given by nyrl hybrids
obscured the complementation data. Whereasthe hybrid nyr2 x nyr5 was clearly nystatinsensitive on 50 U/ml plates, indicating twoindependent loci, the hybrid nyrl x nyr2 wasresistant on 50 U/ml plates, nonconfluent atslightly higher concentrations, and clearly sen-sitive above 100 U/ml. Dissected asci of nyrlcrossed to nyr2, 5, and 15 in each case revealeda 1:1:4 ratio indicative of two independentloci with at least one of the genes not linked toits centromere (Table 3).
ole3 and ole4 complement nyrl, nyr5, andnyri5 and therefore represent two additionalloci. Thus all nystatin-resistant mutants inthis investigation can be placed into the fol-lowing six separate genetic groups: (A) ole2-1,ole2-2, ole2-2R, nyr2; (B) ole3; (C) ole4; (D)nyrl; (E) nyr5, nyr6; (F) nyr15.Growth characteristics of mutant ole2-2.
Experiments were carried out in an attempt toelucidate the relationship between nystatinresistance and growth rate of ole2-2 in YEPDplus lipid supplemented medium (Table 4).Several important observations can be madefrom this data. (i) Mutant ole2-2 supple-mented with two lipids, an unsaturated fattyacid and a sterol, has the doubling time of thewild-type strain S288C (1.5 hr). (ii) ole2-2 isresistant to 100 U of nystatin per ml in liquidTABLE 2. Nystatin resistance determinations of
hybrids
ResistancebHybrida
25C 50 75 100
nyrl x nyr2 + + 4
nyrl x nyr5 + 4 -
nyrl x nyrl5 + 4 4
nyrl x ole2-2R + - -
nyrl x ole2-2 + 4 4
nyr2 x nyr5 + - -
nyr2 x nyrl5 + _nyr2 x ole2-2 + + +nyr2 x ole2-2R + + +ole2-2 x oIe2-2R + + + 4
nyr5 x nyr6 + + + 4
nyr5 x nyrl5 + - -
aRecorded 24 hr after replica plating to YEPDplus Tween 80 plates containing nystatin.
b +, Confluent growth for at least 50% of the de-terminations; a, spotted colonies without conflu-ence, or appreciable thinning out of the cell plaque;-, no growth.
c Units of nystatin per milliliter.
654 BARD
on October 3, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
NYSTATIN RESISTANCE IN S. CEREVISIAE
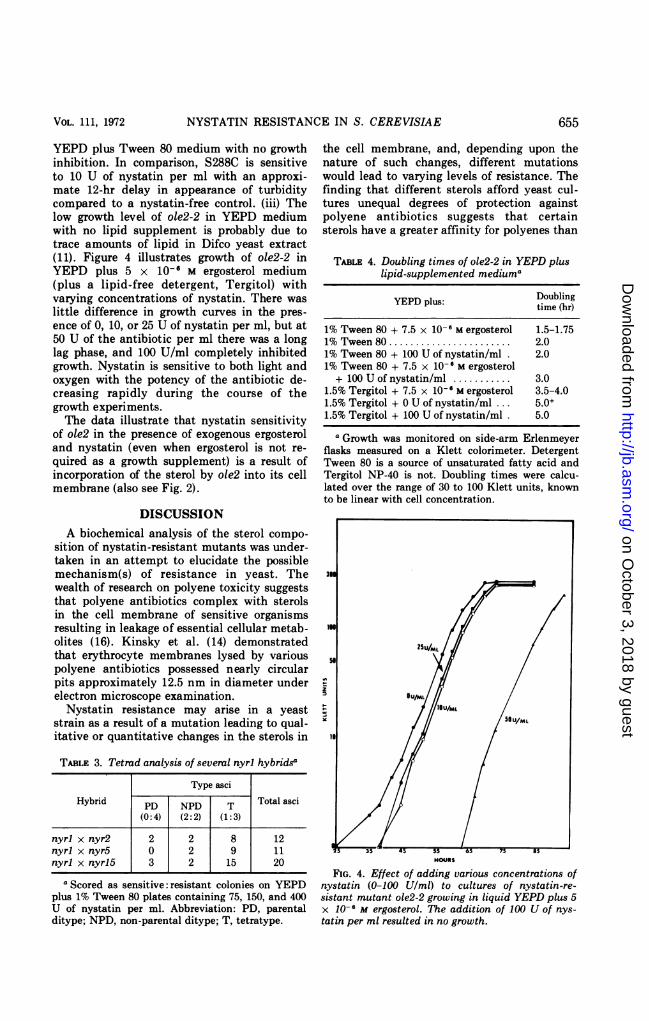
YEPD plus Tween 80 medium with no growthinhibition. In comparison, S288C is sensitiveto 10 U of nystatin per ml with an approxi-mate 12-hr delay in appearance of turbiditycompared to a nystatin-free control. (iii) Thelow growth level of ole2-2 in YEPD mediumwith no lipid supplement is probably due totrace amounts of lipid in Difco yeast extract(11). Figure 4 illustrates growth of ole2-2 inYEPD plus 5 x 10-6 M ergosterol medium(plus a lipid-free detergent, Tergitol) withvarying concentrations of nystatin. There waslittle difference in growth curves in the pres-ence of 0, 10, or 25 U of nystatin per ml, but at50 U of the antibiotic per ml there was a longlag phase, and 100 U/ml completely inhibitedgrowth. Nystatin is sensitive to both light andoxygen with the potency of the antibiotic de-creasing rapidly during the course of thegrowth experiments.The data illustrate that nystatin sensitivity
of ole2 in the presence of exogenous ergosteroland nystatin (even when ergosterol is not re-quired as a growth supplement) is a result ofincorporation of the sterol by ole2 into its cellmembrane (also see Fig. 2).
DISCUSSIONA biochemical analysis of the sterol compo-
sition of nystatin-resistant mutants was under-taken in an attempt to elucidate the possiblemechanism(s) of resistance in yeast. Thewealth of research on polyene toxicity suggeststhat polyene antibiotics complex with sterolsin the cell membrane of sensitive organismsresulting in leakage of essential cellular metab-olites (16). Kinsky et al. (14) demonstratedthat erythrocyte membranes lysed by variouspolyene antibiotics possessed nearly circularpits approximately 12.5 nm in diameter underelectron microscope examination.
Nystatin resistance may arise in a yeaststrain as a result of a mutation leading to qual-itative or quantitative changes in the sterols in
TABLE 3. Tetrad analysis of several nyrl hybridsa
Type asciHybrid PD NPD T Total asci
(0:4) (2:2) (1:3)
nyrl x nyr2 2 2 8 12nyrl x nyr5 0 2 9 11nyrl x nyrl5 3 2 15 20
a Scored as sensitive: resistant colonies on YEPDplus 1% Tween 80 plates containing 75, 150, and 400U of nystatin per ml. Abbreviation: PD, parentalditype; NPD, non-parental ditype; T, tetratype.
the cell membrane, and, depending upon thenature of such changes, different mutationswould lead to varying levels of resistance. Thefinding that different sterols afford yeast cul-tures unequal degrees of protection againstpolyene antibiotics suggests that certainsterols have a greater affinity for polyenes than
TABLE 4. Doubling times of ole2-2 in YEPD pluslipid-supplemented mediuma
YEPD plus: Doublingtime (hr)
1% Tween 80 + 7.5 x 10-6 M ergosterol 1.5-1.751% Tween 80 ....................... 2.01% Tween 80 + 100 U of nystatin/ml 2.01% Tween 80 + 7.5 x 10-6 M ergosterol+ 100 U of nystatin/ml ........... 3.0
1.5% Tergitol + 7.5 x 10-6 M ergosterol 3.5-4.01.5% Tergitol + 0 U of nystatin/ml ... 5.0+1.5% Tergitol + 100 U of nystatin/ml 5.0
a Growth was monitored on side-arm Erlenmeyerflasks measured on a Klett colorimeter. DetergentTween 80 is a source of unsaturated fatty acid andTergitol NP-40 is not. Doubling times were calcu-lated over the range of 30 to 100 Klett units, knownto be linear with cell concentration.
HOURS
FIG. 4. Effect of adding various concentrations ofnystatin (0-100 U/ml) to cultures of nystatin-re-sistant mutant ole2-2 growing in liquid YEPD plus 5x 10-6 M ergosterol. The addition of 100 U of nys-tatin per ml resulted in no growth.
VOL. 111, 1972 655
on October 3, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

J. BACTERIOL.
others. For example, Gottlieb et al. (9) demon-strated that a cholesterol-filipin ratio of 1:4was more effective in reversing the in vitroinhibition of the antibiotic than was a lanos-terol-filipin ratio of 4: 1.
Mutations in which (i) ergosterol is synthe-sized in reduced amounts, (ii) ergosterol issynthesized but must compete for sterol sitesin the cell membrane with other more predom-inant sterols having less affinity for nystatin,or (iii) ergosterol is not synthesized but is re-placed in the cell membrane by one or moresterol intermediates, could lead to resistance.The above mechanisms assume that mutantstrains which do not synthesize ergosterol domake a sterol which meets the structural andfunctional requirements of the cell. A secondrequired assumption is that nystatin hasevolved as an antibiotic with a greater chem-ical affinity for sterols terminal in the meta-bolic pathway such as ergosterol and choles-terol than for sterols earlier in the pathway.The biochemical evidence relating nystatin
resistance in yeast to sterol synthesis revealsthat the above mechanisms may be operative.The strains under consideration here such asS288C, nyr5, and nyr6 most closely resembleone another in synthesizing ergosterol as theprimary sterol, although GLC chromatogramsof S288C and nyr5 vary in total number ofpeaks. nyrl may or may not synthesize ergos-terol as a minor sterol component. UV spectro-photometry suggests that nyrl5 synthesizes asterol having, in addition to the normal ergos-terol absorption spectrum, a band at 235 nm.This unidentified sterol differs from ergosterolmost probably by the formation of a conju-gated double bond system (8). Mutants nyr2and ole2-2R are allelic, and analysis of theirsterols indicates the absence of ergosterol.ole2-2 reveals a different sterol spectrum com-pared to the revertant ole2-2R or wild-typestrain.
Although six separate genes were identified,it cannot be assumed that each represents alesion in the ergosterol biosynthetic pathway.For example, at the ole2-nyr2 locus, the ole2allele may be responsible for the production ofa sterol early in the biosynthetic pathway re-sulting in a cell that is lipid requiring and pe-tite. The alternative nyr2 allele may produce asterol late in the pathway wherein the cell isnon-lipid-requiring and grande. If these alter-native alleles produce vastly different sterols,this locus may have regulatory rather than bio-synthetic functions.
It is interesting to note that wild-type, an-aerobically grown yeast deprived of lipid sup-
plement show a number of similarities to thelipid-requiring ole2, 3, and 4 mutants. (i) An-dreason and Stier (4, 5) demonstrated that theformer require an unsaturated fatty acid andergosterol for growth. (ii) Whereas the ole2, 3,4 mutants are petite and, therefore, lack func-tional mitochondria, mitochondrial function inanaerobically grown cells is repressed. (iii)Bulder (6) has demonstrated that in Schizosac-charomyces japonicus anaerobically growncells are hardly affected by the polyene anti-biotic, pimaricin, compared to aerobicallygrown cells. (Anaerobically grown cells haveone-twenty-fourth as much ergosterol as aero-bically grown cells.) His observation that re-sistance is concomitant with an inability to syn-thesize ergosterol helps to explain the highlevels of nystatin resistance manifested by theole2, 3, and 4 mutants. The Liebermann-Bur-chard colorimetric reaction suggests that thesemutants are blocked at or near lanosterol for-mation.
Keith et al. (12) established the inability ofmutants ole2-1, ole2-2, and ole3o0e4 to desat-urate palmitic and stearic acids given labeledacetate as precursor. They also found thatthese mutants required an unknown aminoacid or base in synthetic complete medium. Ihave now established that methionine is re-quired for growth. Parks (17) showed that themethyl group at the C-24 position of ergosterolis donated by S-adenosylmethionine, and inview of this it is suggested that mutations oc-curring just before or just after this methylationstep are related to the primary genetic lesionsin the ole2, 3, and 4 mutants. Although there isno clear correlation between the inability tosynthesize ergosterol and desaturate fattyacids, Akhtar et al. (3) suggest that the mecha-nism by which certain double bonds in sterolsare formed may be analogous to the way inwhich some long-chain fatty acids are desatu-rated. If regulation of lipid synthesis were de-pendent upon key desaturation steps, a lesionpreventing desaturation in the sterol pathway(whether in the ring or side chain) may lead to aconcomitant loss of desaturation of fatty acids.
ACKNOWLEDGMENTSI gratefully acknowledge Alec D. Keith, under whose su-
pervision this investigation was carried out, and SeymourFogel for his helpful discussions.
LITERATURE CITED1. Adams, B. G., and L. W. Parks. 1967. Evidence for dual
physiological forms of ergosterol in Saccharomycescerevisiae. J. Cell Physiol. 70:161-168.
656 BARD
on October 3, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

NYSTATIN RESISTANCE IN S. CEREVISIAE
2. Ahmed, K. A., and R. A. Woods. 1967. A genetic anal-ysis of resistance to nystatin in Saccharomyces cere-visiae. Genet. Res. 9:179-193.
3. Akhtar, M., M. A. Parvez, and P. F. Hunt. 1969. Studieson the biosynthesis of the ergosterol side chain.Biochem. J. 113:727-732.
4. Andreason, A. A., and J. B. Stier. 1953. Anaerobic nutri-tion of Saccharomyces cerevisiae. I. Ergosterol re-quirement for growth in a defined medium. J. Cell.Comp. Physiol. 41:23-36.
5. Andreason, A. A., and J. B. Stier. 1954. Anaerobic nutri-tion of Saccharomyces cerevisiae. II. Unsaturatedfatty acid requirement for growth in a defined me-dium. J. Cell. Comp. Physiol. 43:271-282.
6. Bulder, C. J. E. A. 1971. Anaerobic growth, ergosterolcontent, and sensitivity to a polyene antibiotic of theyeast Schizosaccharomyces japonicus. Antonie vanLeeuwenhoek J. Microbiol. Serol. 37:353-358.
7. Eddy, A. A. 1958. Aspects of the chemical compositionof yeast, p. 203-207. In A. H. Cook (ed.), The chem-istry and biology of yeast. Academic Press Inc., NewYork.
8. Fieser, L. F., and M. Fieser. 1959. Steroids, p. 93. Rein-hold Publishing Corp., New York.
9. Gottlieb, D., H. F. Carter, J. H. Sloneker, and A. Am-mann. 1958. Protection of fungi against polyene anti-biotics by sterols. Science 128:361.
10. Goulston, G., L. J. Goad, and T. W. Goodwin. 1967.Sterol biosynthesis in fungi. Biochem. J. 102:15c-17c.
11. Jollow, D., G. M. Kellerman, and A. W. Linnane. 1968.The biogenesis of mitochondria. III. The lipid compo-sition of aerobically and anaerobically grown Sacchar-omyces cerevisiae as related to the membrane systemsof the cells. J. Cell Biol. 37:221-230.
12. Keith, A. D., M. R. Resnick, and A. B. Haley. 1969.Fatty acid desaturase mutants of Saccharomyces cere-visiae. J. Bacteriol. 98:415-420.
13. Kinsky, S. C. 1967. Polyene antibiotics, p. 122-141. InD. Gottlieb and P. D. Shaw (ed.), Antibiotics I.Springer Verlag, New York.
14. Kinsky, S. C., S. A. Luse, and L. L. M. van Deenen.1966. Interaction of polyene antibiotics with naturaland artificial membrane systems. Fed. Proc. 25:1503-1510.
15. Lampen, J. 0. 1966. Interference by polyenic antifungalantibiotics (especially nystatin and filipin) with spe-cific membrane functions. Symp. Soc. Gen. Micro-biol. 16:111-130.
16. Marini, F., P. Arnow, and J. 0. Lampen. 1961. The ef-fect of monovalent cations on the inhibition of yeastmetabolism by nystatin. J. Gen. Microbiol. 24:51-62.
17. Parks, L. W. 1958. S-adenosylmethionine and ergosterolsynthesis. J. Amer. Chem. Soc. 80:2023.
18. Parks, L. W., and P. R. Starr. 1963. A relationship be-tween ergosterol and respiratory competency in yeast.J. Cell. Comp. Physiol. 61:61-65.
19. Perkins, D. D. 1949. Biochemical mutants in the smutfungus Ustilago maydis. Genetics 34:607-626.
20. Resnick, M. A., and R. K. Mortimer. 1966. Unsaturatedfatty acid mutants of Saccharomyces cerevisiae. J.Bacteriol. 92:597-600.
21. Sarachek, A., and G. L. Fowler. 1961. Induction of herit-able respiratory deficiency in Saccharomyces by pan-tothenate starvation. Nature (London) 190:792-794.
22. Stadtman, T. 1957. Preparation and assay of cholesteroland ergosterol, p. 392-394. In S. P. Colowick and N.0. Kaplan (ed.), Methods in enzymology, vol. 3. Aca-demic Press Inc., New York.
23. Wisnieski, B. J., A. D. Keith, and M. R. Resnick. 1970.Double-bond requirement in a fatty acid desaturasemutant of Saccharomyces cerevisiae. J. Bacteriol.101:160-165.
24. Woods, R. A. 1971. Nystatin-resistant mutant of yeast:alterations in sterol content. J. Bacteriol. 108:69-73.
VOL. 11l , 1972 657
on October 3, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

ERRATA
Biochemical and Genetic Aspects of NystatinResistance in Saccharomyces cerevisiae
MARTIN BARDGenetics Department, University of California, Berkeley, California 94720
Volume 111, no. 3, p. 649. This paper was originally published with the following incorrect ad-dress: Department of Genetics, University of Sheffield, Sheffield, Great Britain.
Dr. Bard performed this project in partial satisfaction of the requirements for the degree of Doc-tor of Philosophy, in the Genetics Department of the University of California, Berkeley.
Purification and Properties of the AdenosineDiphosphate-Glucose and Uridine
Diphosphate-Glucose Pyrophosphorylases ofMycobacterium smegmatis: Inhibition and Activation
of the Adenosine Diphosphate-GlucosePyrophosphorylase
DAVID LAPP AND ALAN D. ELBEINDepartment of Biochemistry, The UJniversity of Texas Aledical School, San Antonio, Texas 78229
Volume 112, no. 1, p. 327. Abstract, line 15, change: "... . ADP-Glc pyrophosphorylase was in-hibited ...." to ". . . ADP-Glc pyrophosphorylase was activated. . . "





























