Embed Size (px)
Citation preview

DOI: 10.1126/science.1252215, 673 (2014);345 Science
et al.Rainer U. MeckenstockWater droplets in oil are microhabitats for microbial life
This copy is for your personal, non-commercial use only.
clicking here.colleagues, clients, or customers by , you can order high-quality copies for yourIf you wish to distribute this article to others
here.following the guidelines
can be obtained byPermission to republish or repurpose articles or portions of articles
): August 12, 2014 www.sciencemag.org (this information is current as of
The following resources related to this article are available online at
http://www.sciencemag.org/content/345/6197/673.full.htmlversion of this article at:
including high-resolution figures, can be found in the onlineUpdated information and services,
http://www.sciencemag.org/content/suppl/2014/08/06/345.6197.673.DC1.html can be found at: Supporting Online Material
http://www.sciencemag.org/content/345/6197/673.full.html#ref-list-1, 4 of which can be accessed free:cites 30 articlesThis article
http://www.sciencemag.org/cgi/collection/microbioMicrobiology
subject collections:This article appears in the following
registered trademark of AAAS. is aScience2014 by the American Association for the Advancement of Science; all rights reserved. The title
CopyrightAmerican Association for the Advancement of Science, 1200 New York Avenue NW, Washington, DC 20005. (print ISSN 0036-8075; online ISSN 1095-9203) is published weekly, except the last week in December, by theScience
on
Aug
ust 1
2, 2
014
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
on
Aug
ust 1
2, 2
014
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
on
Aug
ust 1
2, 2
014
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
on
Aug
ust 1
2, 2
014
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om
on
Aug
ust 1
2, 2
014
ww
w.s
cien
cem
ag.o
rgD
ownl
oade
d fr
om

R. Bellofatto, D. Berg, J. Bong, T. Christensen, A. Cox, P. Datta,D. Friedman, S. Gilson, J. Guan, S. Hall, R. Haring, C. Haymes,J. Ho, S. Iyer, J. Krawiecki, J. Kusnitz, J. Liu, J. B. Kuang,E. McQuinn, R. Mousalli, B. Nathanson, T. Nayak, D. Nguyen,H. Nguyen, T. Nguyen, N. Pass, K. Prasad, M. Ohmacht,C. Ortega-Otero, Z. Saliba, D. L. Satterfield, J.-s. Seo, B. Shaw,K. Shimohashi, K. Shiraishi, A. Shokoubakhsh, R. Singh, T. Takken,
F. Tsai, J. Tierno, K. Wecker, S.-y. Wang, and T. Wong. We thanktwo anonymous reviewers for their thoughtful comments.
SUPPLEMENTARY MATERIALS
www.sciencemag.org/content/345/6197/668/suppl/DC1Supplementary Text
Figs. S1 to S8Tables S1 and S2References (31–40)Movie S1
10 April 2014; accepted 20 June 201410.1126/science.1254642
OIL BIODEGRADATION
Water droplets in oil aremicrohabitats for microbial lifeRainer U. Meckenstock,1*† Frederick von Netzer,1 Christine Stumpp,1 Tillmann Lueders,1
Anne M. Himmelberg,1 Norbert Hertkorn,2 Philipp Schmitt-Kopplin,2 Mourad Harir,2
Riad Hosein,3 Shirin Haque,4 Dirk Schulze-Makuch5,6
Anaerobic microbial degradation of hydrocarbons, typically occurring at theoil-water transition zone, influences the quality of oil reservoirs. In Pitch Lake,Trinidad and Tobago—the world’s largest asphalt lake—we found that microorganismsare metabolically active in minuscule water droplets (1 to 3 microliters) entrapped in oil.Pyrotag sequencing of individual droplet microbiomes revealed complex methanogenicmicrobial communities actively degrading the oil into a diverse range of metabolites,as shown by nuclear magnetic resonance and Fourier transform ion cyclotron resonancemass spectrometry. High salinity and water-stable isotopes of the droplets indicate adeep subsurface origin. The 13.5% water content and the large surface area of thedroplets represent an underestimated potential for biodegradation of oil away from theoil-water transition zone.
Petroleum hydrocarbons are excellent elec-tron donors and carbon sources for mi-croorganisms; therefore, they are readilydegraded under oxic conditions. Albeit ki-netically slower, anaerobic degradation of
petroleum hydrocarbons also occurs with elec-tron acceptors such as sulfate, nitrate, and ferriciron or under methanogenesis (1). Methanogenicdegradation has been detected for oil reservoirs(2, 3), and although microorganisms are foundthroughout entire reservoirs (4), it is currentlyunderstood that the bulk of biodegradation pro-cesses are taking place at the oil-water transitionzone (4, 5). Because oil wells usually produce pres-surizedwater/oil suspensions containingdisturbedmicrobial communities, such samples provide lim-ited information on the habitat and processesin situ.Wehypothesized thatmicrobial life shouldbe possible in the oil body itself, within water en-closures containing active microbial communities.
We collected undisturbed oil samples fromPitch Lake, the world’s largest natural asphaltlake, in La Brea, Trinidad and Tobago (6) (fig.S1A). When oil samples were spread on alumi-num foil, small bubbles were visible beneath theoil surface. Many bubbles contained gas andcollapsed upon puncturing, whereas some con-tained entrapped water droplets of 1 to 3 ml.Sampling and microscopic inspection of singlewater droplets showed that they indeed harboredmicroorganisms, some of which were activelymotile under the microscope (fig. S1B). Thisobservation substantiated that microbial life canexist in such microliter-scale water droplets en-trained in oil, contrary to a previous hypothesisthat the low water activity would impose waterstress, making such environments too extreme formicrobial life (7). However, the water activity inthe droplets is much higher than in the sur-rounding oil. Furthermore, a dissolved ion analy-sis of bulk water showed that there were noobvious limitations for life caused by the lack ofessential nutrients such as ammonia (95 mg/l) orphosphate (5 mg/l).Bacterial communities of single water droplets
and of bulk oil showed a diverse composition(Fig. 1). Pyrotags were dominated by members ofthe orders Burkholderiales and Enterobacteriales.Other prominent lineages present in the waterdroplets were the Bacteriodales, Rhodospirillales,Sphingomonadales, and to a lesser extentThermotogales and Nitrosomonadales. The taxaidentified in the water droplets were typical foroil samples (e.g., the Burkholderiales) and largely
consistent with an earlier characterization ofsolid samples from the Trinidad Pitch Lake (7)and also from a similar type of oil seep inCalifornia (8). A recent metagenomic investiga-tion of samples from 10 oil reservoirs also iden-tified similar taxa to be predominant (9). Therepeated detection of 16S ribosomal RNA genesequences of presumably aerobic, hydrocarbon-degrading populations such as the Burkholderialesin anoxic oil reservoirs might indicate heretoforeunrecognized physiological features of these taxa.The sequences clustering within the Enterobac-teriales in our study were of mostly unclassifiedaffiliation.Archaea were detectable in 7 of 12 analyzed
droplets and consisted almost exclusively of knownmethanogens within the Methanosarcinales andMethanomicrobiales. These taxa indicate thatacetotrophic and hydrogenotrophic methano-genesis play a role in the biodegradation of thepitch. In addition to these methanogenic archaea,halotolerant or halophilic Halobacteriales werealso present in all archaea-positive water drop-lets, albeit at lower relative abundance. Theseorganisms indicate a high salt origin of the waterdroplets, presumably from the formation waterof the reservoir. The presence of methanogenssuggests that methanogenesis was an importantterminal electron-accepting process in the waterdroplets. The emitted gas on Pitch Lake, however,was dominated by thermogenic methane, withonly a minor contribution of biogenic origin, asindicated by stable isotope ratios of carbon [–46.6 T0.2 per mil (‰)] and hydrogen (–169.4 T 4.6‰) ofmethane extracted from the oil and respectivereports in the literature (7, 10).To determine the origin of the water droplets,
we analyzed the dissolved ion composition aswell as water-stable isotopes of bulk water sam-ples from the oil. Both methods require watersamples of >100 ml and were therefore possiblefor water droplets extracted from the oil but notfrom single droplets. The water exhibited a near-neutral pH of 7.2 with salt concentrations similarto sea water (500 mM Na+, 536 mM Cl–); thesefindings exclude rainfall or fresh surface water asa direct source of the droplets. Furthermore, sur-face water cannot disperse into the heavy oilphase without severe mechanical shearing. Theelevated salt concentrations rather suggest a sub-surface origin of the dropletswith either seawateror brine influence, indicating that theymust havebeen entrapped in the oil already in the reservoiror during the ascent.A deep subsurface origin of the water droplets
was also strongly supported by stable isotopeanalysis of the bulk water phases (Fig. 2). The18O/16O and 2D/1H stable isotope ratios of thebulk water separated by gravitation and by
SCIENCE sciencemag.org 8 AUGUST 2014 • VOL 345 ISSUE 6197 673
1Institute of Groundwater Ecology, Helmholtz ZentrumMünchen, 85764 Neuherberg, Germany. 2Research UnitBiogeochemistry, Helmholtz Zentrum München, 85764Neuherberg, Germany. 3Department of Chemistry, Faculty ofScience and Agriculture, University of the West Indies, St.Augustine Campus, Trinidad and Tobago. 4Department ofPhysics, Faculty of Science and Agriculture, Universityof the West Indies, St. Augustine Campus, Trinidad andTobago. 5School of the Environment, Washington StateUniversity, Pullman, WA 99164, USA. 6Center forAstronomy and Astrophysics, Technical University Berlin,10623 Berlin, Germany.*Present address: Biofilm Center, University Duisburg-Essen,45141 Essen, Germany. †Corresponding author. E-mail: [email protected]
RESEARCH | REPORTS
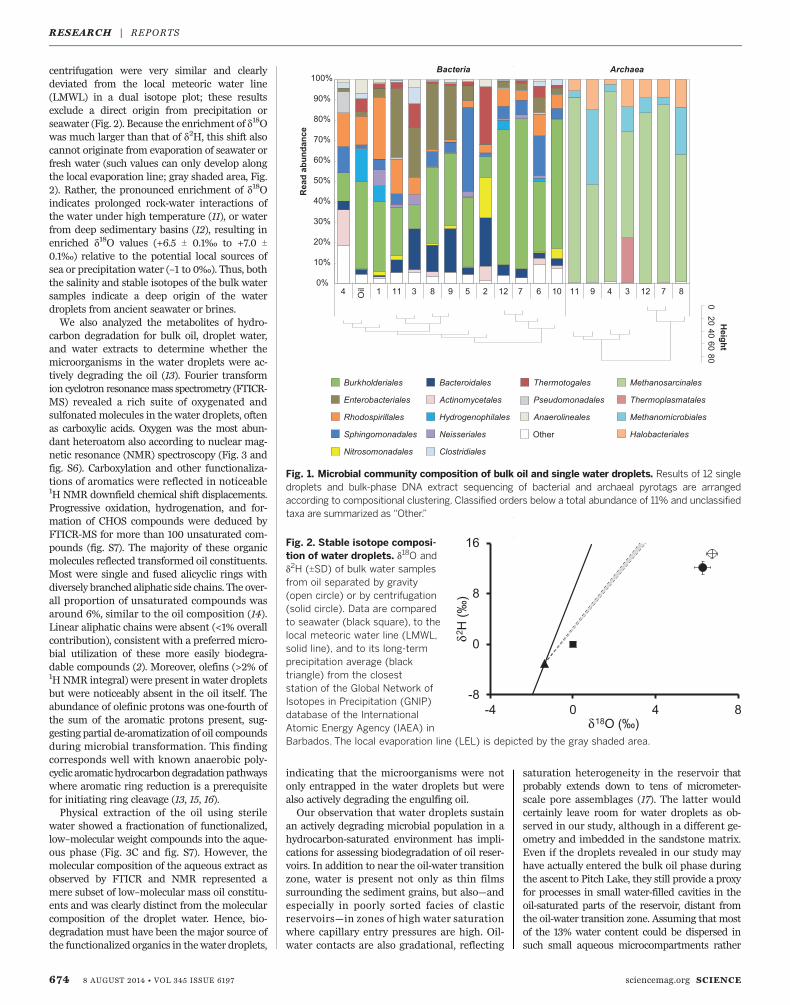
centrifugation were very similar and clearlydeviated from the local meteoric water line(LMWL) in a dual isotope plot; these resultsexclude a direct origin from precipitation orseawater (Fig. 2). Because the enrichment of d18Owas much larger than that of d2H, this shift alsocannot originate from evaporation of seawater orfresh water (such values can only develop alongthe local evaporation line; gray shaded area, Fig.2). Rather, the pronounced enrichment of d18Oindicates prolonged rock-water interactions ofthe water under high temperature (11), or waterfrom deep sedimentary basins (12), resulting inenriched d18O values (+6.5 T 0.1‰ to +7.0 T0.1‰) relative to the potential local sources ofsea or precipitation water (–1 to 0‰). Thus, boththe salinity and stable isotopes of the bulk watersamples indicate a deep origin of the waterdroplets from ancient seawater or brines.We also analyzed the metabolites of hydro-
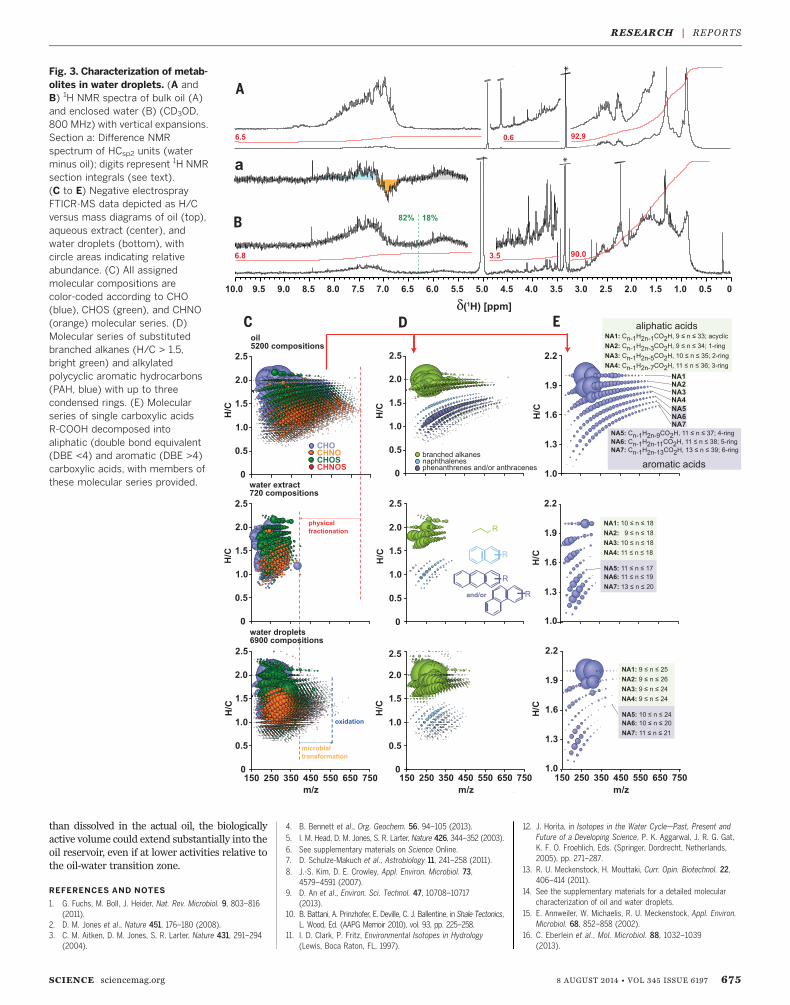
carbon degradation for bulk oil, droplet water,and water extracts to determine whether themicroorganisms in the water droplets were ac-tively degrading the oil (13). Fourier transformion cyclotron resonancemass spectrometry (FTICR-MS) revealed a rich suite of oxygenated andsulfonatedmolecules in the water droplets, oftenas carboxylic acids. Oxygen was the most abun-dant heteroatom also according to nuclear mag-netic resonance (NMR) spectroscopy (Fig. 3 andfig. S6). Carboxylation and other functionaliza-tions of aromatics were reflected in noticeable1H NMR downfield chemical shift displacements.Progressive oxidation, hydrogenation, and for-mation of CHOS compounds were deduced byFTICR-MS for more than 100 unsaturated com-pounds (fig. S7). The majority of these organicmolecules reflected transformed oil constituents.Most were single and fused alicyclic rings withdiversely branched aliphatic side chains. The over-all proportion of unsaturated compounds wasaround 6%, similar to the oil composition (14).Linear aliphatic chains were absent (<1% overallcontribution), consistent with a preferred micro-bial utilization of these more easily biodegra-dable compounds (2). Moreover, olefins (>2% of1H NMR integral) were present in water dropletsbut were noticeably absent in the oil itself. Theabundance of olefinic protons was one-fourth ofthe sum of the aromatic protons present, sug-gesting partial de-aromatization of oil compoundsduring microbial transformation. This findingcorresponds well with known anaerobic poly-cyclic aromatic hydrocarbondegradationpathwayswhere aromatic ring reduction is a prerequisitefor initiating ring cleavage (13, 15, 16).Physical extraction of the oil using sterile
water showed a fractionation of functionalized,low–molecular weight compounds into the aque-ous phase (Fig. 3C and fig. S7). However, themolecular composition of the aqueous extract asobserved by FTICR and NMR represented amere subset of low–molecular mass oil constitu-ents and was clearly distinct from the molecularcomposition of the droplet water. Hence, bio-degradation must have been the major source ofthe functionalized organics in thewater droplets,
indicating that the microorganisms were notonly entrapped in the water droplets but werealso actively degrading the engulfing oil.Our observation that water droplets sustain
an actively degrading microbial population in ahydrocarbon-saturated environment has impli-cations for assessing biodegradation of oil reser-voirs. In addition to near the oil-water transitionzone, water is present not only as thin filmssurrounding the sediment grains, but also—andespecially in poorly sorted facies of clasticreservoirs—in zones of high water saturationwhere capillary entry pressures are high. Oil-water contacts are also gradational, reflecting
saturation heterogeneity in the reservoir thatprobably extends down to tens of micrometer-scale pore assemblages (17). The latter wouldcertainly leave room for water droplets as ob-served in our study, although in a different ge-ometry and imbedded in the sandstone matrix.Even if the droplets revealed in our study mayhave actually entered the bulk oil phase duringthe ascent to Pitch Lake, they still provide a proxyfor processes in small water-filled cavities in theoil-saturated parts of the reservoir, distant fromthe oil-water transition zone. Assuming that mostof the 13% water content could be dispersed insuch small aqueous microcompartments rather
674 8 AUGUST 2014 • VOL 345 ISSUE 6197 sciencemag.org SCIENCE
Fig. 1. Microbial community composition of bulk oil and single water droplets. Results of 12 singledroplets and bulk-phase DNA extract sequencing of bacterial and archaeal pyrotags are arrangedaccording to compositional clustering. Classified orders below a total abundance of 11% and unclassifiedtaxa are summarized as “Other.”
Fig. 2. Stable isotope composi-tion of water droplets. d18O andd2H (TSD) of bulk water samplesfrom oil separated by gravity(open circle) or by centrifugation(solid circle). Data are comparedto seawater (black square), to thelocal meteoric water line (LMWL,solid line), and to its long-termprecipitation average (blacktriangle) from the closeststation of the Global Network ofIsotopes in Precipitation (GNIP)database of the InternationalAtomic Energy Agency (IAEA) inBarbados. The local evaporation line (LEL) is depicted by the gray shaded area.
RESEARCH | REPORTS
than dissolved in the actual oil, the biologicallyactive volume could extend substantially into theoil reservoir, even if at lower activities relative tothe oil-water transition zone.
REFERENCES AND NOTES
1. G. Fuchs, M. Boll, J. Heider, Nat. Rev. Microbiol. 9, 803–816(2011).
2. D. M. Jones et al., Nature 451, 176–180 (2008).3. C. M. Aitken, D. M. Jones, S. R. Larter, Nature 431, 291–294
(2004).
4. B. Bennett et al., Org. Geochem. 56, 94–105 (2013).5. I. M. Head, D. M. Jones, S. R. Larter, Nature 426, 344–352 (2003).6. See supplementary materials on Science Online.7. D. Schulze-Makuch et al., Astrobiology 11, 241–258 (2011).8. J.-S. Kim, D. E. Crowley, Appl. Environ. Microbiol. 73,
4579–4591 (2007).9. D. An et al., Environ. Sci. Technol. 47, 10708–10717
(2013).10. B. Battani, A. Prinzhofer, E. Deville, C. J. Ballentine, in Shale Tectonics,
L. Wood, Ed. (AAPG Memoir 2010), vol. 93, pp. 225–258.11. I. D. Clark, P. Fritz, Environmental Isotopes in Hydrology
(Lewis, Boca Raton, FL, 1997).
12. J. Horita, in Isotopes in the Water Cycle—Past, Present andFuture of a Developing Science, P. K. Aggarwal, J. R. G. Gat,K. F. O. Froehlich, Eds. (Springer, Dordrecht, Netherlands,2005), pp. 271–287.
13. R. U. Meckenstock, H. Mouttaki, Curr. Opin. Biotechnol. 22,406–414 (2011).
14. See the supplementary materials for a detailed molecularcharacterization of oil and water droplets.
15. E. Annweiler, W. Michaelis, R. U. Meckenstock, Appl. Environ.Microbiol. 68, 852–858 (2002).
16. C. Eberlein et al., Mol. Microbiol. 88, 1032–1039(2013).
SCIENCE sciencemag.org 8 AUGUST 2014 • VOL 345 ISSUE 6197 675
Fig. 3. Characterization of metab-olites in water droplets. (A andB) 1H NMR spectra of bulk oil (A)and enclosed water (B) (CD3OD,800 MHz) with vertical expansions.Section a: Difference NMRspectrum of HCsp2 units (waterminus oil); digits represent 1H NMRsection integrals (see text).(C to E) Negative electrosprayFTICR-MS data depicted as H/Cversus mass diagrams of oil (top),aqueous extract (center), andwater droplets (bottom), withcircle areas indicating relativeabundance. (C) All assignedmolecular compositions arecolor-coded according to CHO(blue), CHOS (green), and CHNO(orange) molecular series. (D)Molecular series of substitutedbranched alkanes (H/C > 1.5,bright green) and alkylatedpolycyclic aromatic hydrocarbons(PAH, blue) with up to threecondensed rings. (E) Molecularseries of single carboxylic acidsR-COOH decomposed intoaliphatic (double bond equivalent(DBE <4) and aromatic (DBE >4)carboxylic acids, with members ofthese molecular series provided.
RESEARCH | REPORTS

17. R. A. Schatzinger, J. F. Jordan, Reservoir Characterization: RecentAdvances (American Association of Petroleum Geologists, Tulsa,OK, 1999).
ACKNOWLEDGMENTS
We thank G. Barthel, K. Hörmann, and M. Stöckl for technicalassistance and B. Horsefield for helpful discussions. Funding wasprovided by Helmholtz Zentrum München and the DFG priority
program SPP1319. Pyrotag sequencing data for communityanalyses are deposited at NCBI BioSample (www.ncbi.nlm.nih.gov/biosample) under accession number SRP036082. Original data fromFTICR-MS analyses are provided in the supplementary materials.
SUPPLEMENTARY MATERIALS
www.sciencemag.org/content/345/6197/673/suppl/DC1Materials and Methods
Supplementary TextFigs. S1 to S6Tables S1 to S5References (18–38)
13 February 2014; accepted 26 June 201410.1126/science.1252215
NITROGEN CYCLING
The environmental controls thatgovern the end product of bacterialnitrate respirationBeate Kraft,1* Halina E. Tegetmeyer,1,2 Ritin Sharma,3,4 Martin G. Klotz,5,6
Timothy G. Ferdelman,1 Robert L. Hettich,3,4 Jeanine S. Geelhoed,1,7 Marc Strous1,2,8†
In the biogeochemical nitrogen cycle, microbial respiration processes compete for nitrateas an electron acceptor. Denitrification converts nitrate into nitrogenous gas and thusremoves fixed nitrogen from the biosphere, whereas ammonification converts nitrate intoammonium, which is directly reusable by primary producers.We combined multiple parallellong-term incubations of marine microbial nitrate-respiring communities with isotopelabeling and metagenomics to unravel how specific environmental conditions select foreither process. Microbial generation time, supply of nitrite relative to nitrate, and thecarbon/nitrogen ratio were identified as key environmental controls that determinewhether nitrite will be reduced to nitrogenous gas or ammonium. Our results define themicrobial ecophysiology of a biogeochemical feedback loop that is key to global change,eutrophication, and wastewater treatment.
Currently,most fixednitrogen in thebiosphereoriginates from anthropogenic sources suchas the industrial production of fertilizerammonium. Uptake of fertilizer by crops isonly 17% efficient, and 1 to 5% of fertilizer
ammonium is converted biologically into nitrousoxide, a long-lived and powerful greenhouse gas(1). Microbial nitrification also converts a largeportion of the fertilizer ammonium to nitrate insoil, where it subsequently runs off into surfacewaters and contributes to eutrophication in coastalzones. Nitrate emissions are partially remediatedby denitrification in engineered environmentssuch as wastewater treatment plants. If the endproduct of microbial nitrate reduction could beinfluenced by tuning environmental conditions,this would yield substantial ecological and eco-
nomic benefits for both natural and engineeredsystems.Two microbial processes compete for nitrate
as an electron acceptor: denitrification (includ-ing anammox) and ammonification (includingdissimilatory nitrate reduction to ammonium).The carbon/nitrogen ratio (2–5), pH (5), nitriteversus nitrate concentration (5–7), soil sand con-tent (5), availability of fermentable carbon com-pounds (4, 8), temperature (7, 9), and sulfideconcentration (10–12) are potentially importantenvironmental controls on this competition. Thelack of consensus regarding which factors aremost important, and in which environments, islikely due to the complex and highly variablestructure of natural microbial communities.To unravel the selective forces behind the
ecological success of denitrification or ammoni-fication, we subjected natural communities tospecific environmental conditions in multipleparallel long-term incubations (13). The sourcecommunity was from coastal, sandy tidal flatsediments that make substantial contributionsto global denitrification and perform both de-nitrification and ammonification (12, 14). We didnot aim to reproduce the sediment communityin the laboratory; we simply used the sedimentas a highly microbially diverse inoculum to en-able the selection of optimally adapted nitrate-reducing communities. We performed 15 parallelanoxic incubations (Fig. 1 and table S1) withcontinuous substrate supply, which made it pos-
sible to maintain the nitrate and/or nitrite con-centrations in the in situ micromolar range(<0.5 mM for nitrite, <10 mM for nitrate).Denitrification and ammonification have two
electron acceptors in common: nitrate and ni-trite. In theory, the outcome of the competitioncould be most easily explained by which of thesetwo compounds is supplied. In natural ecosys-tems, the relative supply of nitrate and nitrite iscontrolled by nitrification, a two-step process thatcan yield either compound as the end product. Inour experiments with nitrate as the terminalelectron acceptor, ammonification emerged asthe prevalent pathway, whereas supply of nitriteresulted in denitrification prevalence (Fig. 1). De-nitrification was always observed as the prevalentrespiratory pathway when nitrite was supplied,even in the presence of fermentable substrates andsulfide, at low pH or at a reduced copper concen-tration. Thus, the supply of nitrite or nitrate wasa key factor in the outcome of the competitionbetweendenitrification and ammonification. If ele-vated sulfide concentrations or changes in pH de-crease the rate of nitrite production relative to therate of nitrate production (15, 16), this would there-fore favor ammonification over denitrification,as was observed in some previous studies (5, 12).We further investigated the apparent success
of ammonification with nitrate as the electronacceptor in a 400-day chemostat incubation. In achemostat, the growth rate (or generation time)of the cultivated bacteria is controlled by theapplied dilution rate, enabling us to test whetherthis factor affected the outcome of the competi-tion in any way. The average in situ generationtime of the sampled community was estimatedat ~0.4 days, a value derived from ametagenomeof the tidal flat community (17). During the 400-day incubation, the generation time was variedbetween 1.0 and 3.4 days and the nitrite andnitrate concentrations always remained in thelowmicromolar range (<0.5 mMfor nitrite, <10 mMfor nitrate). The generation time strongly affectedthe outcome of the competition for nitrate (Fig. 2).As shown by mass balancing, denitrification pre-vailed at short generation times, whereas ammo-nification wasmost successful at long generationtimes, with a tipping point detected at a gen-eration time of ~1.7 days (Fig. 2). After 185 days,we increased the generation time in the chemo-stat from 1.0 to 1.7 days and ammonificationslowly became dominant; however, after day 230,denitrification regained prevalence. Ammonifica-tion only recovered after the generation time wasfurther increased to 3.4 days after day 240.To determine the mechanisms responsible
for the observed selective effect of microbialgeneration time, we characterized the selected
676 8 AUGUST 2014 • VOL 345 ISSUE 6197 sciencemag.org SCIENCE
1Max Planck Institute for Marine Microbiology, 28359Bremen, Germany. 2Institute for Genome Research andSystems Biology, Center for Biotechnology, University ofBielefeld, 33615 Bielefeld, Germany. 3UT-ORNL GraduateSchool of Genome Science and Technology, University ofTennessee, Knoxville, TN 37996, USA. 4Chemical ScienceDivision, Oak Ridge National Laboratory, Oak Ridge, TN37830, USA. 5Department of Biological Sciences, Universityof North Carolina, Charlotte, NC 28223, USA. 6State KeyLaboratory of Marine Environmental Science, XiamenUniversity, Xiamen 361005, China. 7NIOZ Royal NetherlandsInstitute for Sea Research, 4401NT Yerseke, Netherlands.8Department of Geoscience, University of Calgary, Calgary,Alberta T2N 1N4, Canada.*Present address: Department of Organismic and EvolutionaryBiology, Harvard University, Cambridge, MA 02138, USA.†Corresponding author. E-mail: [email protected]
RESEARCH | REPORTS


![Oil related microbiology.-Terje.ppt [Kompatibilitetsmodus] · Important microbial processes in oil production: Reservoir souringReservoir souring Microbial Influenced Corrosion (MIC)](https://img.pdfslide.net/doc/110x75/5cf82f5e88c993ea068b7135/oil-related-microbiology-terjeppt-kompatibilitetsmodus-important-microbial.jpg)


























