Embed Size (px)
Citation preview

WhiB5, a Transcriptional Regulator That Contributes toMycobacterium tuberculosis Virulence and Reactivation
Stefano Casonato,a Axel Cervantes Sánchez,b Hirohito Haruki,c Monica Rengifo González,c Roberta Provvedi,d Elisa Dainese,a
Thomas Jaouen,e Susanne Gola,f Estela Bini,b Miguel Vicente,f Kai Johnsson,c Daniela Ghisotti,e Giorgio Palù,a
Rogelio Hernández-Pando,b and Riccardo Manganellia
Department of Histology, Microbiology and Medical Biotechnologies, University of Padua, Padua, Italya; Experimental Pathology Section, Department of Pathology,National Institute of Medical Sciences and Nutrition Salvador Zubiran, Mexico City, Mexicob; Institute of Chemical Sciences and Engineering, Laboratory of ProteinEngineering, École Polytechnique Fédérale de Lausanne, Lausanne, Switzerlandc; Department of Biology, University of Padua, Padua, Italyd; Department of BiomolecularSciences and Biotechnologies, University of Milan, Milan, Italye; and Centro Nacional de Biotecnología (CNB-CSIC), Madrid, Spainf
The proteins belonging to the WhiB superfamily are small global transcriptional regulators typical of actinomycetes. In this pa-per, we characterize the role of WhiB5, a Mycobacterium tuberculosis protein belonging to this superfamily. A null mutant wasconstructed in M. tuberculosis H37Rv and was shown to be attenuated during both progressive and chronic mouse infections.Mice infected with the mutant had smaller bacillary burdens in the lungs but a larger inflammatory response, suggesting a role ofWhiB5 in immunomodulation. Most interestingly, the whiB5 mutant was not able to resume growth after reactivation fromchronic infection, suggesting that WhiB5 controls the expression of genes involved in this process. The mutant was also moresensitive than the wild-type parental strain to S-nitrosoglutathione (GSNO) and was less metabolically active following pro-longed starvation, underscoring the importance of GSNO and starvation in development and maintenance of chronic infection.DNA microarray analysis identified 58 genes whose expression is influenced by WhiB5, including sigM, encoding an alternativesigma factor, and genes encoding the constituents of two type VII secretion systems, namely, ESX-2 and ESX-4.
Proteins belonging to the WhiB superfamily are typical of acti-nomycetes (36). They are small cytoplasmic proteins that con-
tain four conserved cysteine residues able to coordinate an Fe-Scluster (8). They were first described for Streptomyces as redox-sensing transcriptional regulators involved in sporulation, differ-entiation, and antibiotic production (8). The demonstration oftheir DNA-binding activity is very recent (33).
The Mycobacterium tuberculosis genome encodes 7 proteinsbelonging to this superfamily. All of them have a CXXC motifsimilar to that found in thioredoxins and other oxidoreductases,with the exception of WhiB5, which has a CXXXC motif (2).
M. tuberculosis WhiB-like proteins, with the exception ofWhiB2, were recently proposed to have protein disulfide reduc-tase activity (2), suggesting that they might represent bifunctionalproteins able to bind either DNA to regulate transcription or spe-cific target proteins to modify their activity by reducing specificintramolecular disulfides (15).
Of the seven WhiB-like proteins of M. tuberculosis, only four(WhiB1, WhiB2, WhiB3, and WhiB7) have been studied exten-sively. Almost nothing is known regarding the others (WhiB4,WhiB5, and WhiB6), beyond their expression profiles (16) andtheir protein disulfide reductase activity (2). WhiB1 was recentlyshown to be an essential protein that is able to repress its owntranscription by binding to the promoter region of its structuralgene (35). It was also shown to interact with GlgB [alpha(1,4)-glucan branching enzyme] and to reduce an intramolecular disul-fide bond present in this protein, suggesting a role in posttransla-tional regulation of GlgB (15).
WhiB2, also named WhmD, is encoded by an essential geneand is involved in cell division. A conditional mutant of this genein M. smegmatis was shown to produce filamentous cells undernonpermissive conditions (27, 28). Recently, both WhiB2 and a
WhiB2 homolog encoded by a mycobacteriophage were shown tobind the whiB2 promoter to inhibit its transcription (32).
WhiB3 was shown to interact with SigA, the principal sigmafactor of M. tuberculosis (37). It is able to respond to dormancysignals, including O2 and nitric oxide (NO), through its Fe-S clus-ter (34), and it represents an intracellular redox sensor able tointegrate environmental signals with the metabolic pathways re-sponsible for the biosynthesis of complex lipids such as poly- anddiacyltrehalose, sulfolipids, phthiocerol dimycocerosate, and tri-acylglycerol. Through this mechanism, WhiB3 contributes to themaintenance of intracellular redox homeostasis by channelingtoxic reducing equivalents into lipid anabolism (33).
Finally, WhiB7 confers low levels of resistance to several anti-bacterial drugs, and its structural gene is induced in their presence.Its overexpression was shown to induce the expression of genesinvolved in ribosomal protection and antibiotic efflux (23).
In this paper, we focus on M. tuberculosis WhiB5. An orthologof this protein is encoded in the chromosomes of all members ofthe M. tuberculosis complex as well as in those of several slow- andfast-growing mycobacteria, such as Mycobacterium ulcerans, My-cobacterium marinum, Mycobacterium avium, and Mycobacteriumkansasii. However, WhiB5 is not encoded in the genomes of My-
Received 14 December 2011 Returned for modification 1 February 2012Accepted 5 June 2012
Published ahead of print 25 June 2012
Editor: J. L. Flynn
Address correspondence to Riccardo Manganelli, [email protected].
Supplemental material for this article may be found at http://iai.asm.org/.
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/IAI.06328-11
3132 iai.asm.org Infection and Immunity p. 3132–3144 September 2012 Volume 80 Number 9
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.

cobacterium leprae and Mycobacterium smegmatis. We showedthat WhiB5 is involved in metabolic regulation during starvationand is necessary for full virulence during progressive infection andfor the ability of M. tuberculosis to reactivate after chronic infec-tion. Moreover, using DNA microarrays, we identified 58 genesbelonging to the WhiB5 regulon.
MATERIALS AND METHODSEthics statement. Animal studies were approved by the Institutional Eth-ics Committee of the National Institute of Medical Sciences and NutritionSalvador Zubiran in accordance with the Mexican national regulations onanimal care and experimentation (NOM 062-ZOO-1999).
Bacterial strains, media, and growth conditions. The strains usedand generated in this work are listed in Table 1. All experiments wereperformed with M. tuberculosis H37Rv. Bacteria were grown inMiddlebrook 7H9 broth or 7H10 agar medium supplemented with0.05% Tween 80, 0.2% glycerol, and 10% albumin-dextrose-NaCl(ADN) or oleic acid-albumin-dextrose-catalase (OADC) (Difco) at37°C. Liquid cultures were incubated in rolling bottles with gentlerotation. Plates were incubated at 37°C in sealed plastic bags. Esche-richia coli DH5� (Invitrogen) was grown in Luria broth (Difco) at37°C. Where necessary, streptomycin was added at 20 �g/ml, hygro-mycin (Hyg) was added at 50 �g/ml (M. tuberculosis) or 150 �g/ml (E.coli), or kanamycin was added at 20 �g/ml (M. tuberculosis) or 50�g/ml (E. coli). Pristinamycin I (Sanofi-Aventis, Paris, France) wasdissolved in dimethyl sulfoxide (DMSO) (50 mg/ml) and used at1 �g/ml.
DNA manipulations and electroporation. Plasmids used in this workare shown in Table 2. Recombinant DNA techniques were performedaccording to standard procedures, using E. coli DH5� as the initial host.DNA restriction and modifying enzymes were obtained from New Eng-land BioLabs and used according to the manufacturer’s suggestions. Prep-aration of electrocompetent cells and electroporation were performed as
previously described (20). Primers used in the study are shown in Table S1in the supplemental material.
Construction of an M. tuberculosis strain with an inducible copy ofwhiB5. Overexpression of whiB5 in M. tuberculosis was achieved using apristinamycin-inducible expression system based on the Streptomycespristinaespiralis ptr promoter and its pristinamycin-responsive negativerepressor, Pip (14). The whiB5 sequence was amplified from M. tubercu-losis H37Rv genomic DNA and cloned downstream of Pptr in the integra-tive vector pMYT769 to obtain pMYT790 (Table 2). The vector was thenintroduced into M. tuberculosis H37Rv. The efficiency of the system wasverified by reverse transcriptase PCR (RT-PCR) in the presence or ab-sence of induction by pristinamycin (1 �g/ml).
Construction of a whiB5 null mutant in M. tuberculosis. An M. tu-berculosis mutant lacking whiB5 was constructed using recombineering(39). For this purpose, a cassette conferring Hyg resistance flanked by twoDNA fragments of about 500 bp flanking the whiB5 gene was electropo-rated into an M. tuberculosis strain containing the plasmid pJV53, encod-ing phage recombinases (40) (Table 2), and transformants were selectedon Hyg. Two mutants were selected and analyzed by PCR. pJV53 wascured to recover the whiB5 mutant strain TB15. The mutant was thencomplemented by introducing the integrative plasmid pSC25, containingwhiB5 and its upstream region containing the native promoter. The com-plemented strain was named TB16.
Bacterial two-hybrid assay. DNA fragments encoding the 4.2 domainof SigA (176 bp) (37), WhiB3 (329 bp), and WhiB5 (422 bp) were ampli-fied from M. tuberculosis H37Rv chromosomal DNA. The sigA fragmentwas cloned into pKT25, and whiB3 and whiB5 were cloned into pUT18c,to create 3= in-frame fusions with sequences encoding the Bordetella per-tussis CyaA T25 domain (pKT25::4.2sigA) and T18 domain (pUT18c::whiB3 and pUT18c::whiB5) (19). pKT25::4.2sigA was cotransformed witheither pUT18c::whiB3 or pUT18c::whiB5 into E. coli BTH101, and trans-formants were selected on solid medium at 28°C with appropriate selec-tion. The resulting strains were analyzed by a �-galactosidase assay. A
TABLE 1 Bacterial strains used in this work
Strain Genotype or description Source
E. coli strainDH5� fhuA2 �(argF-lacZ)U169 phoA glnV44 �80dlacZ�M15 gyrA96 recA1 relA1 endA1 thi-1 hsdR17 Invitrogen
M. tuberculosis strainsH37Rv PHRI collectiona
TB15 �37Rv derivative; �whiB5 hyg (whiB5 mutant) This workTB16 ��15 derivative containing pSC25; �whiB5::whiB5 hyg Kmr (whiB5-complemented strain) This workTB60 H37Rv derivative containing pMYT769; PfurA-pip int oriE Smr (strain containing empty
pristinamycin-inducible system)This work
TB84 H37Rv derivative containing pMYT790; Pptr-whiB5 PfurA-pip int oriE Smr (strain expressingwhiB5 from the pristinamycin-inducible system)
This work
TB141 H37Rv derivative containing pSC43 (strain expressing GFP from the whiB5 promoter region) This workTB142 TB15 derivative containing pSC43 (strain expressing GFP from the whiB5 promoter region) This work
a PHRI, Public Health Research Institute.
TABLE 2 Plasmids used in this work
Plasmid Description Source or reference
pMYT769 Integrative plasmid carrying the pristinamycin-dependent inducible system; Smr 14pMYT790 pMYT769 derivative in which whiB5 is under transcriptional control of the inducible ptr promoter; Smr This workpJV53 Recombineering plasmid for whiB5 mutant construction; Kmr 40pSC22 Plasmid containing the DNA allelic exchange substrate (AES) used for whiB5 gene replacement; Hygr This workpSC25 Integrative plasmid carrying whiB5 with its upstream region used to complement the whiB5 mutant;
Kmr
This work
pSC43 Derivative of the replicative plasmid pMV4-36 expressing gfp from the whiB5 promoter region; Hygr
Kmr Smr
This work
Role of WhiB5 in M. tuberculosis Virulence
September 2012 Volume 80 Number 9 iai.asm.org 3133
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.

single colony of each transformant was grown in 3 ml of LB mediumcontaining 0.5 mM IPTG (isopropyl-�-D-thiogalactopyranoside) for 16 hat 28°C. Overnight cultures were then diluted 1:5 in LB medium, and theoptical density at 600 nm (OD600) was measured (OD600 of approximately0.2 to 0.3). Two milliliters of diluted culture was centrifuged, resuspendedin 1 ml of buffer Z (70 mM Na2HPO4, 30 mM NaH2PO4, and 1 mMMgSO4), and incubated for 30 min on ice with 10 �l of toluene to perme-abilize cells. After addition of �-mercaptoethanol to a final concentrationof 100 mM, cells were incubated at 28°C in the presence of 0.2 ml of a 0.4%o-nitrophenyl-�-D-galactopyranoside (ONPG) solution. Reactions werestopped with 0.5 ml of 1 M Na2CO3, and the OD420 was measured aftercentrifugation. The enzymatic activity in Miller units was calculated withthe following formula: A {[OD420 (1.75�OD550)]�103}/OD600 �minof incubation � cell volume (ml).
Determination of growth inhibition by disk diffusion assay. M. tu-berculosis strains were grown to early exponential phase, and 100 �l ofculture containing 3 � 106 CFU was spread on 20-ml 7H10 plates. Paperdisks containing 10 �l of the following stock solutions were placed on topof the agar: 5 mM sodium nitroprusside, 0.5 M EDTA, 2 M diamide, 30%(wt/wt) hydrogen peroxide in water, 35 mM cumene hydroperoxide, and100% �-mercaptoethanol. The diameter of the inhibition zone was mea-sured after 15 days of incubation at 37°C.
Determination of GSNO resistance. M. tuberculosis strains weregrown in 7H9 broth to mid-log phase and then diluted to an OD600 ofapproximately 0.1. The bacterial suspensions were then treated with 1mM S-nitrosoglutathione (GSNO; Sigma) or left untreated. The OD600
was measured at different time points. Each day, an aliquot was takenfrom each sample, diluted, and plated on 7H10 plates for determination ofthe viable counts. All experiments were performed three times in tripli-cate.
Determination of metabolic activity during starvation. Culturesgrown for 7 days in 7H9 broth were pelleted, washed twice with phosphate-buffered saline (PBS), and then resuspended in PBS. The resulting sampleswere incubated without agitation at 37°C in sealed bottles. Metabolic activitywas determined at several time points by using a 2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide (XTT) assay (3). Briefly, 200�M XTT and 60 �M menadione were added to 250 �l of M. tuberculosissuspension and incubated for 40 min at 37°C before reading of the absor-bance at 450 nm.
RNA extraction and preparation of labeled cDNA for DNA microar-ray analysis. RNA extraction from in vitro-cultured M. tuberculosis wasperformed as previously described (20), and RNAs were analyzed using anAgilent Bioanalyser 2100 system (Agilent Technologies) according to themanufacturer’s instructions.
Fluorescently labeled cDNA copies of total RNA were prepared bydirect incorporation of fluorescent nucleotide analogues during a first-strand reverse transcription reaction (25).
Microarray hybridization and data analysis. M. tuberculosis oligoar-rays consisting of 4,295 70-mer oligonucleotides representing 3,924 openreading frames (ORFs) from M. tuberculosis strain H37Rv and 371 uniqueORFs from strain CDC 1551 that are not present in H37Rv were obtainedfrom the Center for Applied Genomics, International Center for PublicHealth (Newark, NJ).
Competitive hybridizations with equal amounts of purified Cy3- andCy5-labeled cDNA were performed in duplicate with both dye arrange-ments, as previously described (25).
Microarrays were scanned using an Agilent G2565CA microarrayscanner system (with Scan Control 8.1 software [Agilent Technologies]),and fluorescence intensities of the two channels at each spot were quan-tified using Agilent Feature Extraction 10.1 software (Agilent Technolo-gies).
Data were normalized with the Web-based tool DNMAD (diagnosisand normalization of spotted cDNA microarray data [http://dnmad.bioinfo.cnio.es/]) (41), using the print-tip lowess method after back-ground subtraction. Genes significantly differentially expressed were
identified using the significance analysis of microarrays (SAM) tool (38).Differentially expressed genes were defined by a q value of 0% and aminimum fold difference of �2-fold between the two samples. SAM ispart of the Tiger MultiExperiment Viewer package, version 4.6 (TMeV),available at http://www.tm4.org/mev/ (26).
RACE. Transcription start points (TSPs) of selected genes were deter-mined by rapid amplification of cDNA ends (RACE), using a 5=/3= RACEkit (Roche Applied Science) according to the manufacturer’s suggestions.Primer sequences are shown in Table S1 in the supplemental material.
Analysis of the whiB5 promoter region. The whiB5 promoter regionwas amplified from H37Rv genomic DNA and cloned into pCR-BluntII-TOPO (Invitrogen) to obtain pSC31. The fragment was then excised asa HindIII/NheI fragment and cloned upstream of the gfp gene inpMV4-36 (7) to obtain pSC43 (Table 2).
M. tuberculosis cells containing pSC43 were washed and resuspendedin lysis buffer (10 mM Tris-HCl [pH 8], 5 mM EDTA, 1� protease inhib-itor cocktail [Roche]) with an appropriate volume of 0.1-mm zirconiabeads. Cells were disrupted using a Mini-Bead Beater apparatus with threecycles of 30 s each at the highest speed. Samples were centrifuged for 2min, and the supernatants were recovered and filter sterilized. The fluo-rescence of cell extracts was measured with a Perkin Elmer LS50B spec-trophotometer with excitation at 480 nm and emission at 510 nm. Proteinconcentrations were determined using a Bradford protein assay kit (Bio-Rad). Fluorescence readings were then normalized for the protein con-centration and expressed as the relative fluorescence per mg of total pro-teins present in the extract.
Infection of THP-1-derived macrophages. THP-1 monocytes(American Type Culture Collection) were grown in suspension at 37°C in5% CO2 in bicarbonate-buffered RPMI (Gibco) supplemented with 10%(vol/vol) fetal bovine serum (FBS) (Gibco), 50 �mol/liter �-mercapto-ethanol, and 50 �g/ml gentamicin at a cell density of 0.2 � 106 to 1.0 �106/ml.
Cells were plated at a density of 7.5 � 104 cells/well in RPMI with 10%FBS, 50 �mol/liter �-mercaptoethanol, 50 �g/ml gentamicin, and 50ng/ml phorbol 12-myristate 13-acetate (PMA) (Sigma) for 24 h to becomefully differentiated macrophages before use in experiments. Differenti-ated THP-1 macrophages were washed extensively with serum-free RPMIand infected with M. tuberculosis in 96-well plates at a multiplicity ofinfection of 1:20 (CFU:macrophages) (22). After 90 min of incubation at37°C, the medium was removed and cells were washed twice with 100 �l ofwarm PBS to remove extracellular bacteria. The medium was replacedevery 48 h. Every 24 h for 8 days, starting from 90 min after the initialwashes, the medium was removed from three wells, and then intracellularbacteria were released by lysing the macrophages with 100 �l of 0.05%SDS. The suspensions obtained from the lysed macrophages were imme-diately diluted in 7H9 broth and plated to determine viable counts. About95% of macrophages remained viable during the entire experiment, asdetermined by trypan blue exclusion.
Mouse infection and RNA extraction from lung homogenates dur-ing experimental progressive and chronic tuberculosis infections. Toinduce progressive pulmonary tuberculosis, male BALB/c mice were anes-thetized, tracheas were exposed, and 2 � 105 viable bacilli were injected(17). Groups of 4 mice were killed at different time points after infection.
To induce chronic infection, 8-week-old B6D2F1 mice were anesthe-tized, and 4 � 103 bacilli were instilled intratracheally (4). Groups of 3mice in 2 different experiments were euthanized at different time points.
RNAs were extracted from the lungs of infected mice by use of a pre-viously described protocol (9), with some modifications. Briefly, lungsfrom three mice for each time point were perfused with 1 ml of TRIzol(Qiagen, Valencia, CA). The tissue was minced, transferred to a steriletube, and homogenized using a Multi-Gen 7 homogenizer (Pro Scientific)for three cycles in 30-s bursts at maximal speed. The homogenized sam-ples were centrifuged at 4,000 rpm for 10 min at 20°C, and the supernatantwas transferred to a new tube and immediately placed on ice. This super-natant contained eukaryotic RNA, whereas the pelleted material con-
Casonato et al.
3134 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.

tained the bacilli. To isolate the bacterial RNA, pellets were resuspended in1 ml of TRIzol and transferred to a 2-ml screw-cap tube containing zirco-nia and silica beads (1- and 0.1-mm diameter, respectively). Mycobacteriawere disrupted using a Mini-Bead Beater 8 apparatus (Biospec ProductsInc.), using three cycles of 30 s each at the highest speed. This material wasthen centrifuged for 1 min at maximum speed, and the supernatant wastransferred to a 2-ml screw-cap tube containing 300 �l of chloroform.After vigorous vortexing and centrifugation for 10 min at maximumspeed, the resulting supernatant, containing the M. tuberculosis RNA, wasprecipitated with a solution of glycogen, ammonium acetate, isopropanol,and ethanol. Finally, bacterial RNA was isolated using four cycles of pu-rification with an RNeasy column (Invitrogen), followed by extensiveDNase incubation to eliminate DNA contamination.
Mouse infection for determination of virulence and immunogenic-ity in experimental models of progressive and chronic infections. Themurine models described above were used to compare the levels of viru-lence and types of immune response induced by infection with the whiBmutant and the complemented and parental strains.
To induce acute progressive infection, 6- to 8-week-old BALB/cmale mice were distributed in three groups of 60 animals each andinfected with each of the different strains, using a high dose (2 � 105
cells) of viable bacilli to induce progressive disease. Ten mice fromeach group were left undisturbed to record survival from day 8 to day120 after infection. Six animals from each group were sacrificed byexsanguination 1, 3, 7, 14, 21, 28, 60, and 120 days after infection. Onelung lobe, the right or left, was perfused with 10% formaldehyde dis-solved in PBS and prepared for histopathological studies. The otherlobe was snap-frozen in liquid nitrogen and then stored at 70°C formicrobiological and immunological analysis.
To induce chronic infection, three groups of 50 female B6D2F1 miceeach were infected with either of the three strains, using a low-dose inoc-ulum (4 � 103 cells). Groups of 6 mice in 2 different experiments wereeuthanized 1, 3, and 5 months after infection. To induce reactivation, 10mice with stable chronic infection at 5 months of infection were treatedwith corticosterone (3 mg/liter dissolved in drinking water) and thensacrificed 1 month later. One lung was perfused with 10% formaldehydedissolved in PBS and prepared for histopathological studies. The otherlobe was snap-frozen in liquid nitrogen and then stored at 70°C formicrobiological and immunological analysis.
Preparation of lung tissue for histology and automated morpho-metry. For histology and automated morphometry analysis, one lobe ofthe lung was fixed by intratracheal perfusion with 10% formaldehyde for24 h, sectioned through the hilus, and embedded in paraffin. Sections (5�m thick) were stained with hematoxylin-eosin and used to determinethe percentage of the pulmonary area affected by pneumonia and thegranuloma area (in square micrometers), using an automated image an-alyzer (Q Win Leica, Milton Keynes, United Kingdom) (18). The sameparaffin-embedded material prepared for histopathological studies wasused to determine the local cytokine production by immunohistochem-istry. Lung sections from mice infected with either the mutant or parentalstrain in the model of progressive disease, taken at days 28, 60, and 120,were deparaffinized and maintained in 1� HCN buffer (HEPES, NaCl,and CaCl2). Sections were washed with 1� HCN plus 0.05% Tween 20,and the endogenous peroxidase activity was blocked with 6% H2O2 dis-solved in 1� PBS plus 0.1% sodium azide and incubated for 1 h. Afterblocking with normal swine serum, tissue sections were incubated withprimary antibodies overnight at 4°C at optimal dilutions, which had beendetermined previously. We used primary antibodies against tumor necro-sis factor alpha (TNF-�) (rabbit polyclonal IgG clone H-156; Santa CruzBiotechnology), gamma interferon (IFN- ) (goat polyclonal IgG cloneD-17; Santa Cruz Biotechnology), transforming growth factor beta(TGF-�) (rabbit polyclonal IgG; Santa Cruz Biotechnology), interleu-kin-4 (IL-4) (goat polyclonal IgG; Santa Cruz Biotechnology), and IL-10(goat polyclonal IgG clone M-18; Santa Cruz Biotechnology). Secondarybiotinylated antibodies (biotin–anti-rabbit IgG or biotin–anti-goat IgG)
were used to detect the binding of the primary antibodies. Finally, horse-radish peroxidase (HRP)-conjugated avidin and 3,3-diaminobenzidine(DAB)– hydrogen peroxide were used to develop the reaction. Tissue sec-tions were counterstained with hematoxylin. For morphometry, at leastthree random fields from each section for three different mice for eachtime point and each strain were studied. The total number of inflamma-tory cells, specifically in the area of pneumonia and in granulomas, wasquantified, and the percentage of cytokine-immunostained cells or posi-tive cells for each cytokine was determined using an automated imageanalyzer (Q Win Leica).
Determination of CFU in infected lungs. Right or left lungs from fourmice for each time point in two separate experiments were used for colonycounting (CFU). Lungs were homogenized with a Polytron homogenizer(Kinematica, Lucerne, Switzerland) in sterile 50-ml tubes containing 3 mlof isotonic saline. Four dilutions of each homogenate were spread ontoduplicate plates containing Bacto Middlebrook 7H10 agar enriched withOADC. Plates were incubated for 21 days prior to determination of CFU.
Real-time PCR analysis. Reverse transcription was performed withrandom primers and murine leucoblastoma virus retrotranscriptase(MULV RT) (Applied Biosystems). Five hundred nanograms of totalRNA was denatured at 98°C for 2 min and then chilled on ice and used toprepare 25 �l of annealing mixture (5.5 mM MgCl2, a 0.55 mM concen-tration of each deoxynucleoside triphosphate [dNTP], 0.25 mmol ran-dom hexamers, 32 U of MULV RT, 10 U of RNase inhibitor, and 1�reaction buffer [Applied Biosystems]). Samples were then incubated at25°C for 10 min, at 45°C for 50 min, and finally at 95°C for 5 min to allowannealing of the random hexamers. Quantitative PCR was performedwith SYBR green master mix (Applied Biosystems) as previously de-scribed (20). For bacterial mRNA analysis, results were normalized to theamount of sigA mRNA or 16S rRNA (21). RNA samples that had not beenreverse transcribed were included in all experiments to monitor DNAcontamination. For analysis of cytokines in lung homogenates, left orright lung lobes from three different mice per group in two differentexperiments were used to isolate mRNA by use of an RNeasy miniprep kit(Qiagen) according to the recommendations of the manufacturer. Qual-ity and quantity of RNA were evaluated through spectrophotometry andon agarose gels. The mRNA encoding glyceraldehyde 3-phosphate dehy-drogenase (G3PDH) was used as an internal control to normalize theexpression of the cytokine-expressing genes. Data are shown as numbersof copies of cytokine-specific mRNA/106 copies of G3PDH-specificmRNA (18).
Microarray data accession number. The data discussed in this publi-cation have been deposited in NCBI’s Gene Expression Omnibus (12) andare accessible through GEO Series accession number GSE30299 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?accGSE30299).
RESULTSA whiB5 null mutant shows normal growth on laboratory mediaand in macrophages. To study the physiological role of WhiB5,recombineering (40) was used to replace its structural gene with acassette conferring resistance to hygromycin. The resulting mu-tant strain (TB15) showed growth kinetics and colony morphol-ogy indistinguishable from those of the wild-type (wt) strain inMiddlebrook 7H9 or 7H10 medium (data not shown). Finally, theTB15 mutant was complemented by reintroducing the whiB5 genewith its upstream regulatory region at an ectopic locus of its chro-mosome, giving strain TB16.
M. tuberculosis is a facultative intracellular pathogen that cansurvive and grow inside macrophages. To investigate whetherWhiB5 was dispensable for intraphagosomal growth, we infectedTHP-1-derived macrophages with exponentially growing culturesof H37Rv and the whiB5 mutant TB15. As shown in Fig. 1, bothstrains were able to divide intracellularly during the entire courseof the experiment (6 days), with their CFU increasing about 100-
Role of WhiB5 in M. tuberculosis Virulence
September 2012 Volume 80 Number 9 iai.asm.org 3135
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.

fold. No difference between the two strains was observed, suggest-ing that WhiB5 is not required for intracellular growth in restingmacrophages.
Characterization of whiB5 mutant immunopathogenicityduring experimental progressive tuberculosis. To characterizethe role of WhiB5 during experimental progressive tuberculosis,three groups of BALB/c mice (60 per group) were infected intra-tracheally with 2.5 � 105 CFU of either H37Rv, the whiB5 mutantTB15, or the complemented derivative TB16. Ten animals from
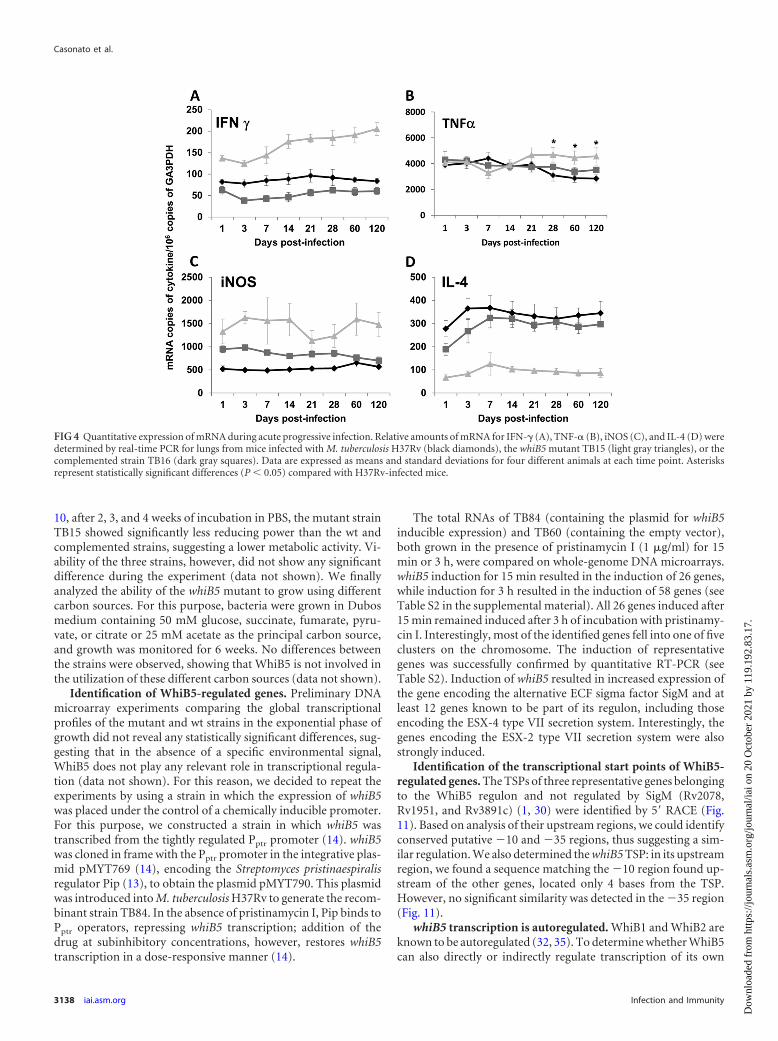
each group were left undisturbed until the end of the experiment(120 days). Among them, all of the animals infected with TB15survived, while seven of those inoculated with the parental straindied. However, only 3 of the 10 animals infected with the comple-mented strain TB16 died before the end of the experiment, indi-cating partial complementation (Fig. 2A). These data correlatedwell with the bacterial burdens in lung homogenates (Fig. 2B).After 1 week of infection and thereafter, a small but statisticallysignificant difference in bacterial loads was found in the lungs ofmice infected with the whiB5 mutant compared to those of theanimals infected with the parental or complemented strain. His-topathological analysis showed progressive pneumonia after 28days of infection with H37Rv, reaching its peak at day 120, when50% of the lung surface was affected. Although the lungs of miceinfected with the whiB5 mutant TB15 showed significantly smallerbacterial loads, the percentage of lung consolidation in these ani-mals was similar to that observed for mice infected with the pa-rental strain (Fig. 3A and B). The size of granulomas, which ap-peared 2 weeks after infection, was about 2-fold larger in miceinfected with the whiB5 mutant than in mice infected with the wtor complemented strain (Fig. 2C and 3A and B). These data sug-gested that the whiB5 mutant was able to induce a strong inflam-matory response. This was confirmed by the data shown in Fig. 4,which demonstrate higher expression levels of IFN- , TNF-�, andinducible nitric oxide synthase (iNOS) in the lungs of mice in-fected with the mutant strain than in those of mice infected withthe parental or complemented strain. Expression of IL-4, how-ever, was lower in the lungs of mice infected with the mutant
FIG 1 Growth of M. tuberculosis H37Rv and the whiB5 mutant in THP-1-derived macrophages. The results are expressed as CFU per well. The reportedvalues represent the averages and standard deviations obtained from threeparallel independent infections. The experiment was repeated twice, usingindependent bacterial inocula and THP-1 cultures. Black diamonds, H37Rv;gray squares, TB15.
FIG 2 Pathogenicity of the whiB5 mutant during acute progressive infection following intratracheal inoculation. (A) Survival after infection; (B) lung bacterialburdens; (C) sizes of granulomas in mice infected with M. tuberculosis H37Rv (black), the whiB5 mutant TB15 (light gray), or the complemented strain TB16(dark gray). Mice were sacrificed at 1, 3, 7, 14, 21, 28, 60, and 120 days postinfection. Asterisks represent statistically significant differences (P � 0.05) comparedwith H37Rv-infected mice.
Casonato et al.
3136 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.

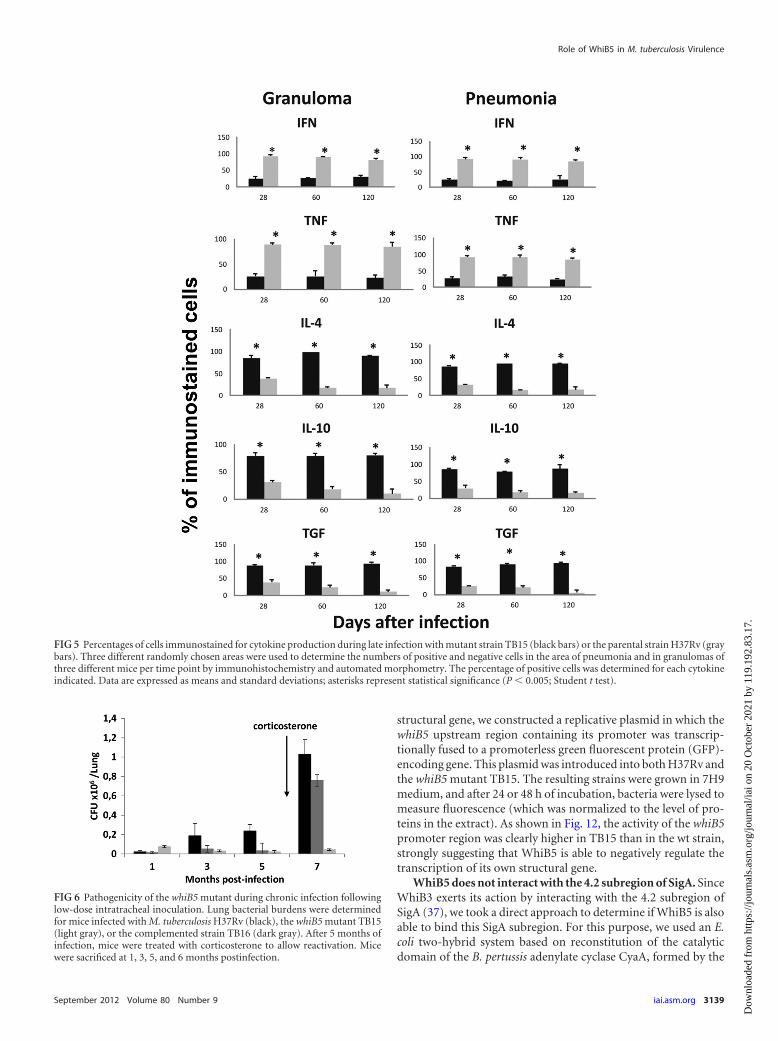
strain. These results were confirmed and extended at the proteinproduction level by use of immunohistochemistry and automatedmorphometry. Figure 3C to F show cells producing IFN- (C andD) or IL-10 (E and F) in granulomas of mice infected with thewhiB5 mutant TB15 (C and E) or the parental strain H37Rv (Dand F). Quantitative analysis of immunostained cells clearlyshowed significantly higher percentages of TNF-�- and IFN- -producing cells and smaller amounts of IL-4-, IL-10-, and TGF-�-producing cells in the lungs of mice infected with the mutantstrain than in those of mice infected with the parental strain. Thesituation was similar in the area of pneumonia and in granulomas(Fig. 5).
Characterization of whiB5 mutant immunopathogenicityduring chronic infection. The role of WhiB5 in M. tuberculosisvirulence was also investigated during experimental chronic tu-berculosis. Three groups of B6D2F1 mice (50 per group) wereinfected intratracheally with 4.0 � 103 CFU of either H37Rv, thewhiB5 mutant TB15, or the complemented strain TB16. As ex-pected, all animals were alive after 5 months of infection and had
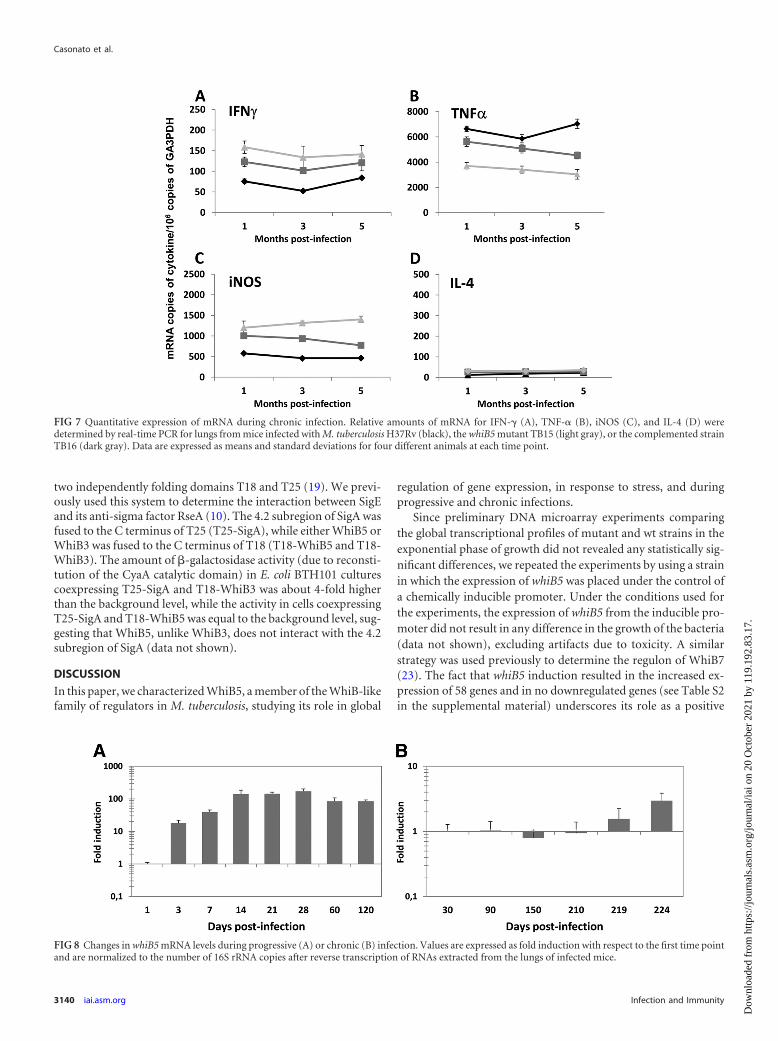
very small amounts of bacteria in their lungs (Fig. 6). While thebacterial burdens in the lungs of mice infected with the whiB5mutant TB15 were slightly lower than those detected in the lungsof mice infected with H37Rv or the complemented strain TB16,mice infected with the whiB5 mutant showed higher levels of cy-tokine mRNA (with the exception of that for TNF-�) (Fig. 7) andsimilar inflammation to that observed in mice infected with theparental or complemented strain (data not shown).
After 5 months of chronic infection, 10 mice from each groupwere treated with corticosterone to induce immunosuppressionand infection reactivation. After 1 month of immunosuppression,all TB15-infected animals and 9 of 10 of the animals infected withthe parental or complemented strain were still alive. Survivorswere sacrificed, and the bacterial burden in the lung was deter-mined: animals infected with the wt or the complemented strainshowed a sharp increase in the number of CFU in the lungs, whilethe bacterial burden did not change significantly in whiB5 mu-tant-infected mice, suggesting that these animals were still able tocontrol the infection (Fig. 6). Taken together, these data clearlyindicate that WhiB5 contributes to the reactivation of chronictuberculosis in this model system.
Expression of whiB5 during progressive and chronic infec-tions. Given the role of WhiB5 in M. tuberculosis virulence andreactivation, we measured the transcription of its structural geneduring active and chronic infections with the wild-type strainH37Rv, using quantitative RT-PCR (with results expressed as thenumber of whiB5 cDNA copies/106 16S cDNA copies). As shownin Fig. 8A, whiB5 expression during progressive infection was at itsmaximum between day 14 and day 28. However, during chronicinfection, whiB5 showed low and stable expression throughoutthe experiment, with increased expression during reactivation(Fig. 8B).
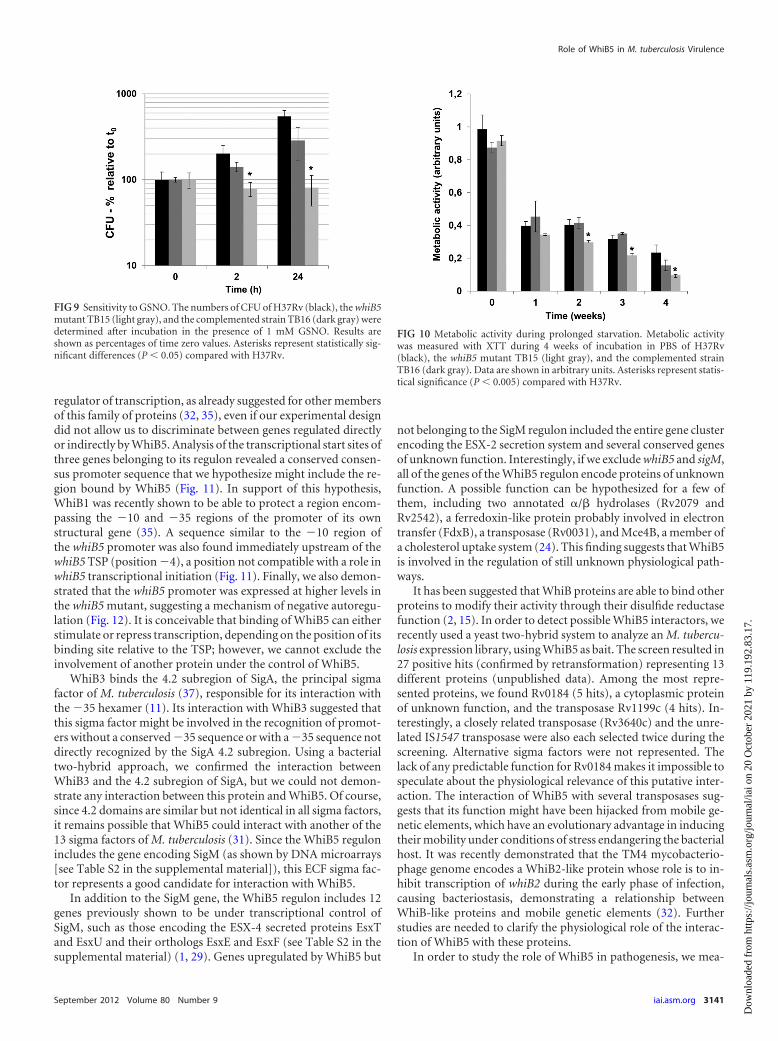
Sensitivity of the whiB5 mutant to environmental stress. Toinvestigate the specific role of WhiB5 in M. tuberculosis physiol-ogy, we tested the relative resistance of H37Rv and the whiB5mutant TB15 to different stresses mimicking the environmentsencountered by bacilli during active or chronic infection. Resis-tance to hydrogen peroxide, cumene hydroperoxide (an organicperoxide), sodium nitroprusside and acidified nitrate (both ofwhich are nitric oxide [NO] donors), EDTA, and �-mercaptoeth-anol was tested using a disk diffusion assay on agar plates or byexamining the killing curve for liquid medium. However, wecould not observe any differences in sensitivity to these six com-pounds among the strains tested (data not shown). Since previouswork had shown that whiB5 was slightly induced in the presence ofGSNO (6), we tested the sensitivity of the mutant strain to thiscompound. As shown in Fig. 9, during exposure to a GSNO con-centration of 1 mM, the wt strain was still able to proliferate, whilethe whiB5 mutant’s growth was arrested. The wt phenotype waspartially restored in complemented strain TB16.
Role of WhiB5 in the level of metabolic activity during star-vation. During chronic infection, it is hypothesized that bacilli areexposed to an environment that is poor of nutrients. In order todemonstrate whether WhiB5 might have a physiological role dur-ing starvation, the metabolic activity of the wt, mutant, and com-plemented strains was measured using XTT (a compound whichexhibits a color change when reduced to a formazan product by anactive electron transport chain) during prolonged incubation inPBS. This method was previously used to evaluate the metabolicactivity of M. avium in response to starvation (3). As shown in Fig.
FIG 3 Representative lung histopathology and immunohistochemistry ofBALB/c mice after 60 days of infection with the whiB5 mutant TB15 (leftpanels) or the parental strain (right panels). (A and B) Similar areas of pneu-monia were induced by either strain (asterisks), while larger granulomas (ar-rows) were produced by strain TB15. Granulomas from mice infected withTB15 (C) showed a larger amount of IFN- -immunostained cells than gran-ulomas from animal infected with the parental strain (D). However, granulo-mas in the lungs of mice infected with the mutant strain TB15 (E) exhibitedfewer IL-10-immunostained cells than granulomas of mice infected with theparental strain (F). Magnification, �100 (A and B) and �200 (C to F). PanelsA and B show H&E staining, and panels C to F show HRP-developedimmunohistochemistry.
Role of WhiB5 in M. tuberculosis Virulence
September 2012 Volume 80 Number 9 iai.asm.org 3137
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.
10, after 2, 3, and 4 weeks of incubation in PBS, the mutant strainTB15 showed significantly less reducing power than the wt andcomplemented strains, suggesting a lower metabolic activity. Vi-ability of the three strains, however, did not show any significantdifference during the experiment (data not shown). We finallyanalyzed the ability of the whiB5 mutant to grow using differentcarbon sources. For this purpose, bacteria were grown in Dubosmedium containing 50 mM glucose, succinate, fumarate, pyru-vate, or citrate or 25 mM acetate as the principal carbon source,and growth was monitored for 6 weeks. No differences betweenthe strains were observed, showing that WhiB5 is not involved inthe utilization of these different carbon sources (data not shown).
Identification of WhiB5-regulated genes. Preliminary DNAmicroarray experiments comparing the global transcriptionalprofiles of the mutant and wt strains in the exponential phase ofgrowth did not reveal any statistically significant differences, sug-gesting that in the absence of a specific environmental signal,WhiB5 does not play any relevant role in transcriptional regula-tion (data not shown). For this reason, we decided to repeat theexperiments by using a strain in which the expression of whiB5was placed under the control of a chemically inducible promoter.For this purpose, we constructed a strain in which whiB5 wastranscribed from the tightly regulated Pptr promoter (14). whiB5was cloned in frame with the Pptr promoter in the integrative plas-mid pMYT769 (14), encoding the Streptomyces pristinaespiralisregulator Pip (13), to obtain the plasmid pMYT790. This plasmidwas introduced into M. tuberculosis H37Rv to generate the recom-binant strain TB84. In the absence of pristinamycin I, Pip binds toPptr operators, repressing whiB5 transcription; addition of thedrug at subinhibitory concentrations, however, restores whiB5transcription in a dose-responsive manner (14).
The total RNAs of TB84 (containing the plasmid for whiB5inducible expression) and TB60 (containing the empty vector),both grown in the presence of pristinamycin I (1 �g/ml) for 15min or 3 h, were compared on whole-genome DNA microarrays.whiB5 induction for 15 min resulted in the induction of 26 genes,while induction for 3 h resulted in the induction of 58 genes (seeTable S2 in the supplemental material). All 26 genes induced after15 min remained induced after 3 h of incubation with pristinamy-cin I. Interestingly, most of the identified genes fell into one of fiveclusters on the chromosome. The induction of representativegenes was successfully confirmed by quantitative RT-PCR (seeTable S2). Induction of whiB5 resulted in increased expression ofthe gene encoding the alternative ECF sigma factor SigM and atleast 12 genes known to be part of its regulon, including thoseencoding the ESX-4 type VII secretion system. Interestingly, thegenes encoding the ESX-2 type VII secretion system were alsostrongly induced.
Identification of the transcriptional start points of WhiB5-regulated genes. The TSPs of three representative genes belongingto the WhiB5 regulon and not regulated by SigM (Rv2078,Rv1951, and Rv3891c) (1, 30) were identified by 5= RACE (Fig.11). Based on analysis of their upstream regions, we could identifyconserved putative 10 and 35 regions, thus suggesting a sim-ilar regulation. We also determined the whiB5 TSP: in its upstreamregion, we found a sequence matching the 10 region found up-stream of the other genes, located only 4 bases from the TSP.However, no significant similarity was detected in the 35 region(Fig. 11).
whiB5 transcription is autoregulated. WhiB1 and WhiB2 areknown to be autoregulated (32, 35). To determine whether WhiB5can also directly or indirectly regulate transcription of its own
FIG 4 Quantitative expression of mRNA during acute progressive infection. Relative amounts of mRNA for IFN- (A), TNF-� (B), iNOS (C), and IL-4 (D) weredetermined by real-time PCR for lungs from mice infected with M. tuberculosis H37Rv (black diamonds), the whiB5 mutant TB15 (light gray triangles), or thecomplemented strain TB16 (dark gray squares). Data are expressed as means and standard deviations for four different animals at each time point. Asterisksrepresent statistically significant differences (P � 0.05) compared with H37Rv-infected mice.
Casonato et al.
3138 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.
structural gene, we constructed a replicative plasmid in which thewhiB5 upstream region containing its promoter was transcrip-tionally fused to a promoterless green fluorescent protein (GFP)-encoding gene. This plasmid was introduced into both H37Rv andthe whiB5 mutant TB15. The resulting strains were grown in 7H9medium, and after 24 or 48 h of incubation, bacteria were lysed tomeasure fluorescence (which was normalized to the level of pro-teins in the extract). As shown in Fig. 12, the activity of the whiB5promoter region was clearly higher in TB15 than in the wt strain,strongly suggesting that WhiB5 is able to negatively regulate thetranscription of its own structural gene.
WhiB5 does not interact with the 4.2 subregion of SigA. SinceWhiB3 exerts its action by interacting with the 4.2 subregion ofSigA (37), we took a direct approach to determine if WhiB5 is alsoable to bind this SigA subregion. For this purpose, we used an E.coli two-hybrid system based on reconstitution of the catalyticdomain of the B. pertussis adenylate cyclase CyaA, formed by the
FIG 5 Percentages of cells immunostained for cytokine production during late infection with mutant strain TB15 (black bars) or the parental strain H37Rv (graybars). Three different randomly chosen areas were used to determine the numbers of positive and negative cells in the area of pneumonia and in granulomas ofthree different mice per time point by immunohistochemistry and automated morphometry. The percentage of positive cells was determined for each cytokineindicated. Data are expressed as means and standard deviations; asterisks represent statistical significance (P � 0.005; Student t test).
FIG 6 Pathogenicity of the whiB5 mutant during chronic infection followinglow-dose intratracheal inoculation. Lung bacterial burdens were determinedfor mice infected with M. tuberculosis H37Rv (black), the whiB5 mutant TB15(light gray), or the complemented strain TB16 (dark gray). After 5 months ofinfection, mice were treated with corticosterone to allow reactivation. Micewere sacrificed at 1, 3, 5, and 6 months postinfection.
Role of WhiB5 in M. tuberculosis Virulence
September 2012 Volume 80 Number 9 iai.asm.org 3139
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.
two independently folding domains T18 and T25 (19). We previ-ously used this system to determine the interaction between SigEand its anti-sigma factor RseA (10). The 4.2 subregion of SigA wasfused to the C terminus of T25 (T25-SigA), while either WhiB5 orWhiB3 was fused to the C terminus of T18 (T18-WhiB5 and T18-WhiB3). The amount of �-galactosidase activity (due to reconsti-tution of the CyaA catalytic domain) in E. coli BTH101 culturescoexpressing T25-SigA and T18-WhiB3 was about 4-fold higherthan the background level, while the activity in cells coexpressingT25-SigA and T18-WhiB5 was equal to the background level, sug-gesting that WhiB5, unlike WhiB3, does not interact with the 4.2subregion of SigA (data not shown).
DISCUSSION
In this paper, we characterized WhiB5, a member of the WhiB-likefamily of regulators in M. tuberculosis, studying its role in global
regulation of gene expression, in response to stress, and duringprogressive and chronic infections.
Since preliminary DNA microarray experiments comparingthe global transcriptional profiles of mutant and wt strains in theexponential phase of growth did not revealed any statistically sig-nificant differences, we repeated the experiments by using a strainin which the expression of whiB5 was placed under the control ofa chemically inducible promoter. Under the conditions used forthe experiments, the expression of whiB5 from the inducible pro-moter did not result in any difference in the growth of the bacteria(data not shown), excluding artifacts due to toxicity. A similarstrategy was used previously to determine the regulon of WhiB7(23). The fact that whiB5 induction resulted in the increased ex-pression of 58 genes and in no downregulated genes (see Table S2in the supplemental material) underscores its role as a positive
FIG 7 Quantitative expression of mRNA during chronic infection. Relative amounts of mRNA for IFN- (A), TNF-� (B), iNOS (C), and IL-4 (D) weredetermined by real-time PCR for lungs from mice infected with M. tuberculosis H37Rv (black), the whiB5 mutant TB15 (light gray), or the complemented strainTB16 (dark gray). Data are expressed as means and standard deviations for four different animals at each time point.
FIG 8 Changes in whiB5 mRNA levels during progressive (A) or chronic (B) infection. Values are expressed as fold induction with respect to the first time pointand are normalized to the number of 16S rRNA copies after reverse transcription of RNAs extracted from the lungs of infected mice.
Casonato et al.
3140 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.
regulator of transcription, as already suggested for other membersof this family of proteins (32, 35), even if our experimental designdid not allow us to discriminate between genes regulated directlyor indirectly by WhiB5. Analysis of the transcriptional start sites ofthree genes belonging to its regulon revealed a conserved consen-sus promoter sequence that we hypothesize might include the re-gion bound by WhiB5 (Fig. 11). In support of this hypothesis,WhiB1 was recently shown to be able to protect a region encom-passing the 10 and 35 regions of the promoter of its ownstructural gene (35). A sequence similar to the 10 region ofthe whiB5 promoter was also found immediately upstream of thewhiB5 TSP (position 4), a position not compatible with a role inwhiB5 transcriptional initiation (Fig. 11). Finally, we also demon-strated that the whiB5 promoter was expressed at higher levels inthe whiB5 mutant, suggesting a mechanism of negative autoregu-lation (Fig. 12). It is conceivable that binding of WhiB5 can eitherstimulate or repress transcription, depending on the position of itsbinding site relative to the TSP; however, we cannot exclude theinvolvement of another protein under the control of WhiB5.
WhiB3 binds the 4.2 subregion of SigA, the principal sigmafactor of M. tuberculosis (37), responsible for its interaction withthe 35 hexamer (11). Its interaction with WhiB3 suggested thatthis sigma factor might be involved in the recognition of promot-ers without a conserved 35 sequence or with a 35 sequence notdirectly recognized by the SigA 4.2 subregion. Using a bacterialtwo-hybrid approach, we confirmed the interaction betweenWhiB3 and the 4.2 subregion of SigA, but we could not demon-strate any interaction between this protein and WhiB5. Of course,since 4.2 domains are similar but not identical in all sigma factors,it remains possible that WhiB5 could interact with another of the13 sigma factors of M. tuberculosis (31). Since the WhiB5 regulonincludes the gene encoding SigM (as shown by DNA microarrays[see Table S2 in the supplemental material]), this ECF sigma fac-tor represents a good candidate for interaction with WhiB5.
In addition to the SigM gene, the WhiB5 regulon includes 12genes previously shown to be under transcriptional control ofSigM, such as those encoding the ESX-4 secreted proteins EsxTand EsxU and their orthologs EsxE and EsxF (see Table S2 in thesupplemental material) (1, 29). Genes upregulated by WhiB5 but
not belonging to the SigM regulon included the entire gene clusterencoding the ESX-2 secretion system and several conserved genesof unknown function. Interestingly, if we exclude whiB5 and sigM,all of the genes of the WhiB5 regulon encode proteins of unknownfunction. A possible function can be hypothesized for a few ofthem, including two annotated �/� hydrolases (Rv2079 andRv2542), a ferredoxin-like protein probably involved in electrontransfer (FdxB), a transposase (Rv0031), and Mce4B, a member ofa cholesterol uptake system (24). This finding suggests that WhiB5is involved in the regulation of still unknown physiological path-ways.
It has been suggested that WhiB proteins are able to bind otherproteins to modify their activity through their disulfide reductasefunction (2, 15). In order to detect possible WhiB5 interactors, werecently used a yeast two-hybrid system to analyze an M. tubercu-losis expression library, using WhiB5 as bait. The screen resulted in27 positive hits (confirmed by retransformation) representing 13different proteins (unpublished data). Among the most repre-sented proteins, we found Rv0184 (5 hits), a cytoplasmic proteinof unknown function, and the transposase Rv1199c (4 hits). In-terestingly, a closely related transposase (Rv3640c) and the unre-lated IS1547 transposase were also each selected twice during thescreening. Alternative sigma factors were not represented. Thelack of any predictable function for Rv0184 makes it impossible tospeculate about the physiological relevance of this putative inter-action. The interaction of WhiB5 with several transposases sug-gests that its function might have been hijacked from mobile ge-netic elements, which have an evolutionary advantage in inducingtheir mobility under conditions of stress endangering the bacterialhost. It was recently demonstrated that the TM4 mycobacterio-phage genome encodes a WhiB2-like protein whose role is to in-hibit transcription of whiB2 during the early phase of infection,causing bacteriostasis, demonstrating a relationship betweenWhiB-like proteins and mobile genetic elements (32). Furtherstudies are needed to clarify the physiological role of the interac-tion of WhiB5 with these proteins.
In order to study the role of WhiB5 in pathogenesis, we mea-
FIG 9 Sensitivity to GSNO. The numbers of CFU of H37Rv (black), the whiB5mutant TB15 (light gray), and the complemented strain TB16 (dark gray) weredetermined after incubation in the presence of 1 mM GSNO. Results areshown as percentages of time zero values. Asterisks represent statistically sig-nificant differences (P � 0.05) compared with H37Rv.
FIG 10 Metabolic activity during prolonged starvation. Metabolic activitywas measured with XTT during 4 weeks of incubation in PBS of H37Rv(black), the whiB5 mutant TB15 (light gray), and the complemented strainTB16 (dark gray). Data are shown in arbitrary units. Asterisks represent statis-tical significance (P � 0.005) compared with H37Rv.
Role of WhiB5 in M. tuberculosis Virulence
September 2012 Volume 80 Number 9 iai.asm.org 3141
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.
sured the expression of its structural gene during progressive andchronic infections. In the progressive disease model, when miceare infected with the H37Rv strain, the initial phase is dominatedby high-level production of Th1 cell cytokines that, together withhigh levels of TNF-� and iNOS, temporarily control the infection.Granulomas develop during this phase, with maximal activity atday 21 postinfection. Four weeks after infection, the expression ofTh1 cell cytokines, TNF-�, and iNOS starts to decline. Gradually,pneumonic areas prevail over granulomas. Pneumonia in coexis-tence with a large bacterial burden causes death. The highest levelof expression of whiB5 occurred from days 14 to 28 of infection,which corresponded to the higher activity of the protective immu-nity mediated by Th1 cytokines and activated macrophages (Fig.8A). This suggests that WhiB5 could be related to bacterial adap-tation to the high-stress environment produced by the immunesystem.
The expression of whiB5 was also measured in a murine modelof chronic infection. This model is characterized by small andstable bacillary loads without weight loss or death of the animals.Histological and immunological studies showed granulomas withhigh expression of TNF-�, IFN- , and iNOS (4). When cortico-sterone was supplied in drinking water to the infected animals,there was significant bacillary growth with development of lungconsolidation, representing reactivation. In contrast to latent in-fection in humans, in the murine model the bacilli are not in atruly dormant state because they can be recovered in axenic cul-ture (4). In this model, whiB5 in the wild-type strain showed lowand stable expression and exhibited increased expression duringreactivation (Fig. 8B).
To characterize the physiological functions of WhiB5, we con-structed a mutant of M. tuberculosis lacking the whiB5 gene. The
mutant did not show any relevant growth difference with respectto the wt in broth culture or after exposure of bacteria to severaldifferent stress conditions or to intracellular growth in restingmacrophages. However, we demonstrated that the mutant wasmore sensitive to GSNO than the wt parental strain (Fig. 9).GSNO is converted into glutathione (GSH) and NO from bacte-rial thioredoxin (5) and is important in M. tuberculosis killing byactivated macrophages (42). While its mechanism of actionagainst M. tuberculosis has not been elucidated fully, it might in-volve both the toxic action of NO and that of GSH. The latter isthought to provoke a redox imbalance involving the use of analternative thiol that mycobacteria use for regulating reduction oroxidation activity (i.e., mycothiol). Alternatively, GSH could exertits inhibitory effect after being converted to the penicillin-like de-rivative glutacillin, a �-lactam form of GSH (42). Our data show-ing sensitivity of our mutant to GSNO but not to two other NOdonors (sodium nitroprusside and acidified nitrate) suggest thatWhiB5 is involved in the response to the toxic action of GSH andnot to that of NO.
We also found that the whiB5 mutant produced less reduc-ing power (i.e., it is less metabolically active) than the wt pa-rental strain during long periods of nutrient starvation (Fig.10), even though this did not affect its viability, suggesting thatWhiB5 might be involved in sustaining the metabolic activityof M. tuberculosis during nutrient starvation. Both GSNO sen-sitivity and decreased reducing potential during starvationwere partially complemented by the reintroduction of a wtcopy of the gene, including its promoter, into an ectopic locusof the genome.
In all our experiments, the complemented strain showed onlypartial recovery of the wt phenotypes. To understand the reasonfor this finding, we measured whiB5 expression in both the wt andcomplemented strains but failed to show any relevant differencebetween the two strains. Of course, it is still possible that differ-ences in the fine-tuning of whiB5 expression, which might be dueto unrecognized regulatory regions far upstream of the promoter,are responsible for the only partial recovery of the wt phenotypes.
Finally, to determine the role of WhiB5 in virulence, the whiB5mutant was used to infect mice in progressive and chronic diseasemodels of infection. In both models, the whiB5 mutant was atten-uated. In the progressive disease model, animals infected with themutant strain showed longer survival with slightly smaller bacil-lary loads than those of mice infected with the parental or com-plemented strain (Fig. 2A and B). Moreover, mice infected withthe mutant strain showed more lung inflammatory consolidationthan those infected with the other strains, as well as larger granu-lomas (Fig. 2C) and higher expression levels of proinflammatorycytokines such as IFN- and TNF-�, along with less IL-4 produc-tion (Fig. 4), suggesting a larger protective immune response in
FIG 11 Map of the upstream regions of whiB5 and three genes belonging to its regulon. Transcriptional start points detected by 5= RACE are shown in bold,enlarged capital letters. Putative 10 and 35 regions are boxed, while conserved bases are underlined. Putative translational start codons are shown in bold.
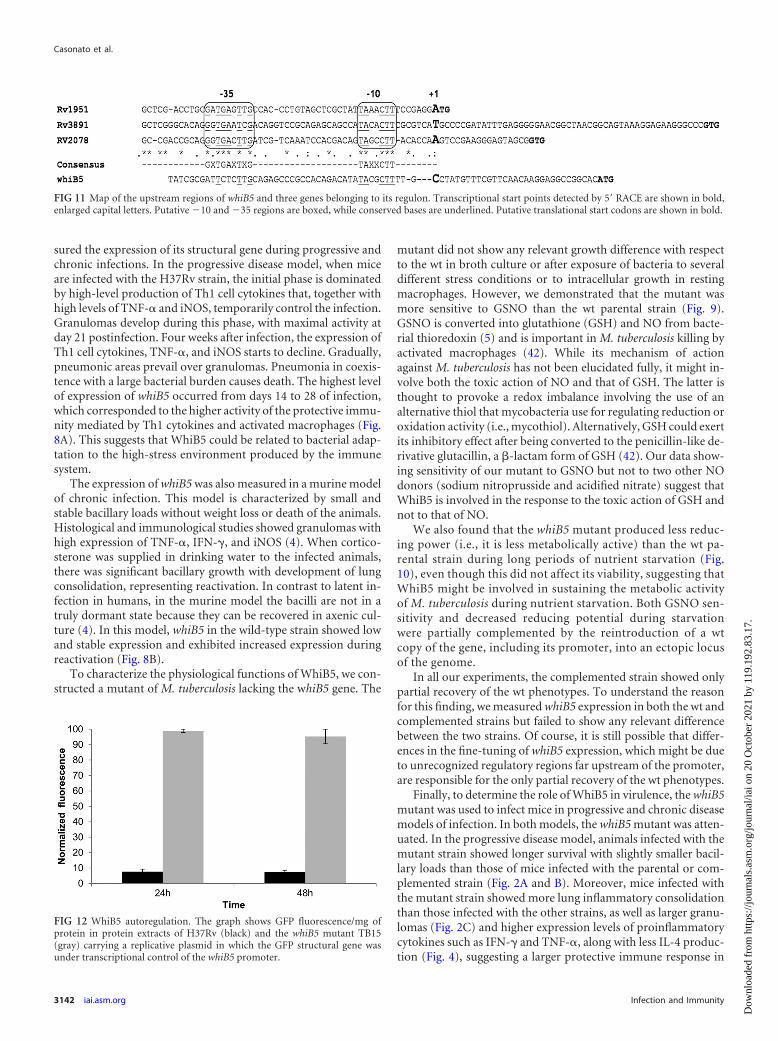
FIG 12 WhiB5 autoregulation. The graph shows GFP fluorescence/mg ofprotein in protein extracts of H37Rv (black) and the whiB5 mutant TB15(gray) carrying a replicative plasmid in which the GFP structural gene wasunder transcriptional control of the whiB5 promoter.
Casonato et al.
3142 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.

these animals. This cytokine profile was confirmed and extendedby immunohistochemistry and automated morphometry, whichshowed a significantly higher percentage of immunostained cellsreactive to the proinflammatory cytokines TNF-� and IFN- anda smaller amount of inflammatory cells immunoreactive to theanti-inflammatory cytokines IL-10, IL-4, and TGF-� in the lungsof mice infected with the mutant strain (Fig. 3 and 5). Thus, thesedata suggest that the lack of WhiB5 produces some antigenic mod-ifications which induce more inflammation and larger granulo-mas, constituted essentially of Th1 cells and activated macro-phages, in coexistence with smaller numbers of Th2 and Treg cells,as denoted by the lower percentages of IL-10- and TGF-immuno-stained lymphocytes, allowing more efficient clearance of the ba-cilli.
Since WhiB5 controls the expression of members of two typeVII secretion systems, we hypothesize that some antigens ex-ported by ESX-2 or ESX-4 could have an immunosuppressingfunction. However, the decreased virulence of the whiB5 mutantcannot be attributed solely to the downregulation of sigM, since ina previous study it was shown that a sigM mutant was not attenu-ated in mice (1).
Chronic infection induced by lower-level bacillus administra-tion resulted in smaller bacillary burdens in mice infected with themutant strain (Fig. 6), associated with more IFN- and iNOSexpression (Fig. 7), than in animals infected with the wt or com-plemented strain, as previously shown in the progressive infectionmodel. However, infection with the mutant strain resulted in lessTNF-� expression (Fig. 7), while it resulted in more expression ofthis cytokine in the progressive disease model. This discrepancybetween the two models of infection might result from the verysmall amount of bacteria infecting the lungs in the chronic infec-tion model or from the different genetic backgrounds of the miceused for these two models of infection. Interestingly, whenchronic infection was reactivated by corticosterone administra-tion, the mutant strain was not able to resume growth, while thelung bacterial burdens of mice infected with the wt or comple-mented strain increased significantly, suggesting that WhiB5might be involved with bacillary proliferation during reactivation(Fig. 6). To the best of our knowledge, this is the first M. tubercu-losis mutant shown to have this phenotype. This finding is partic-ularly interesting because the mechanism of reactivation of M.tuberculosis is still unknown. It is possible that the reduced meta-bolic activity of the whiB5 mutant under nutrient-limiting condi-tions, together with the presence of GSNO and GSH in macro-phages, does not allow the bacteria to resume growth afterpressure of the immune response is relieved by corticosterone,underscoring the importance of both the nutritional status andthe production of GSNO and/or GSH by macrophages in the es-tablishment and maintenance of chronic infection. Moreover,since WhiB5 controls the expression of two type VII secretionsystems (whose physiologic role is still completely unknown), it ispossible to hypothesize that some antigens exported by ESX-2 orESX-4 might be involved in immunomodulation or in quorum-sensing mechanisms necessary for reactivation. Based on thesefindings, we believe that further analysis of the whiB5 mutant’sbehavior and physiology during chronic infection might help toshed some light on the mechanism of reactivation of M. tubercu-losis.
ACKNOWLEDGMENTS
We thank Issar Smith for critically reading the manuscript.This work was supported by the European Community’s Sixth Frame-
work Programme (contracts LSHP-CT-2006-037566 [Mycomancy] andLHSP-CT-2005-018923 [NM4TB]) and by Integrated Actions Italy-Spain.
REFERENCES1. Agarwal N, Woolwine SC, Tyagi S, Bishai WR. 2007. Characterization of
the Mycobacterium tuberculosis sigma factor SigM by assessment of viru-lence and identification of SigM-dependent genes. Infect. Immun. 75:452– 461.
2. Alam MS, Garg SK, Agrawal P. 2009. Studies on structural and func-tional divergence among seven WhiB proteins of Mycobacterium tubercu-losis H37Rv. FEBS J. 276:76 –93.
3. Archuleta RJ, Yvonne Hoppes P, Primm TP. 2005. Mycobacteriumavium enters a state of metabolic dormancy in response to starvation.Tuberculosis (Edinb.) 85:147–158.
4. Arriaga AK, Orozco EH, Aguilar LD, Rook GA, Hernandez Pando R.2002. Immunological and pathological comparative analysis between ex-perimental latent tuberculous infection and progressive pulmonary tu-berculosis. Clin. Exp. Immunol. 128:229 –237.
5. Attarian R, Bennie C, Bach H, Av-Gay Y. 2009. Glutathione disulfideand S-nitrosoglutathione detoxification by Mycobacterium tuberculosisthioredoxin system. FEBS Lett. 583:3215–3220.
6. Boshoff HI, et al. 2004. The transcriptional responses of Mycobacteriumtuberculosis to inhibitors of metabolism: novel insights into drug mecha-nisms of action. J. Biol. Chem. 279:40174 – 40184.
7. Delogu G, et al. 2004. Rv1818c-encoded PE_PGRS protein of Mycobac-terium tuberculosis is surface exposed and influences bacterial cell struc-ture. Mol. Microbiol. 52:725–733.
8. den Hengst CD, Buttner MJ. 2008. Redox control in actinobacteria.Biochim. Biophys. Acta 1780:1201–1216.
9. de Souza GA, et al. 2010. Using a label-free proteomics method to iden-tify differentially abundant proteins in closely related hypo- and hyper-virulent clinical Mycobacterium tuberculosis Beijing isolates. Mol. Cell.Proteomics 9:2414 –2423.
10. Dona V, et al. 2008. Evidence of complex transcriptional, translational,and posttranslational regulation of the extracytoplasmic function sigmafactor SigE in Mycobacterium tuberculosis. J. Bacteriol. 190:5963–5971.
11. Dove SL, Darst SA, Hochschild A. 2003. Region 4 of sigma as a target fortranscription regulation. Mol. Microbiol. 48:863– 874.
12. Edgar R, Domrachev M, Lash AE. 2002. Gene Expression Omnibus:NCBI gene expression and hybridization array data repository. NucleicAcids Res. 30:207–210.
13. Folcher M, et al. 2001. A transcriptional regulator of a pristinamycinresistance gene in Streptomyces coelicolor. J. Biol. Chem. 276:1479 –1485.
14. Forti F, Crosta A, Ghisotti D. 2009. Pristinamycin-inducible gene regu-lation in mycobacteria. J. Biotechnol. 140:270 –277.
15. Garg S, Alam MS, Bajpai R, Kishan KR, Agrawal P. 2009. Redox biologyof Mycobacterium tuberculosis H37Rv: protein-protein interaction be-tween GlgB and WhiB1 involves exchange of thiol-disulfide. BMCBiochem. 10:1. doi:10.1186/1471-2091-10-1.
16. Geiman DE, Raghunand TR, Agarwal N, Bishai WR. 2006. Differentialgene expression in response to exposure to antimycobacterial agents andother stress conditions among seven Mycobacterium tuberculosis whiB-likegenes. Antimicrob. Agents Chemother. 50:2836 –2841.
17. Hernandez-Pando R, et al. 1996. Correlation between the kinetics of Th1,Th2 cells and pathology in a murine model of experimental pulmonarytuberculosis. Immunology 89:26 –33.
18. Hernandez Pando R, Aguilar LD, Smith I, Manganelli R. 2010. Immu-nogenicity and protection induced by a Mycobacterium tuberculosis sigEmutant in a BALB/c mouse model of progressive pulmonary tuberculosis.Infect. Immun. 78:3168 –3176.
19. Karimova G, Pidoux J, Ullmann A, Ladant D. 1998. A bacterial two-hybrid system based on a reconstituted signal transduction pathway. Proc.Natl. Acad. Sci. U. S. A. 95:5752–5756.
20. Maciag A, et al. 2007. Global analysis of the Mycobacterium tuberculosisZur (FurB) regulon. J. Bacteriol. 189:730 –740.
21. Manganelli R, Dubnau E, Tyagi S, Kramer FR, Smith I. 1999. Differ-ential expression of 10 sigma factor genes in Mycobacterium tuberculosis.Mol. Microbiol. 31:715–724.
Role of WhiB5 in M. tuberculosis Virulence
September 2012 Volume 80 Number 9 iai.asm.org 3143
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.

22. Manganelli R, Voskuil MI, Schoolnik GK, Smith I. 2001. The Mycobac-terium tuberculosis ECF sigma factor SigE: role in global gene expressionand survival in macrophages. Mol. Microbiol. 41:423– 437.
23. Morris RP, et al. 2005. Ancestral antibiotic resistance in Mycobacteriumtuberculosis. Proc. Natl. Acad. Sci. U. S. A. 102:12200 –12205.
24. Pandey AK, Sassetti CM. 2008. Mycobacterial persistence requires theutilization of host cholesterol. Proc. Natl. Acad. Sci. U. S. A. 105:4376 –4380.
25. Provvedi R, Boldrin F, Falciani F, Palu G, Manganelli R. 2009. Globaltranscriptional response to vancomycin in Mycobacterium tuberculosis.Microbiology 155:1093–1102.
26. Provvedi R, Palù G, Manganelli R. 2008. Use of DNA microarrays tostudy global patterns of gene expression, p 95–110. In Parish T, Brown AC(ed), Mycobacteria protocols, 2nd ed. Humana Press, Totowa, NJ.
27. Raghunand TR, Bishai WR. 2006. Mapping essential domains of Myco-bacterium smegmatis WhmD: insights into WhiB structure and function. J.Bacteriol. 188:6966 – 6976.
28. Raghunand TR, Bishai WR. 2006. Mycobacterium smegmatis whmD andits homologue Mycobacterium tuberculosis whiB2 are functionally equiva-lent. Microbiology 152:2735–2747.
29. Raman S, et al. 2006. Mycobacterium tuberculosis SigM positively regu-lates Esx secreted protein and nonribosomal peptide synthetase genes anddown regulates virulence-associated surface lipid synthesis. J. Bacteriol.188:8460 – 8468.
30. Raman S, et al. 2001. The alternative sigma factor SigH regulates majorcomponents of oxidative and heat stress responses in Mycobacterium tu-berculosis. J. Bacteriol. 183:6119 – 6125.
31. Rodrigue S, Provvedi R, Jacques PE, Gaudreau L, Manganelli R. 2006.The sigma factors of Mycobacterium tuberculosis. FEMS Microbiol. Rev.30:926 –941.
32. Rybniker J, et al. 2010. Insights into the function of the WhiB-like protein
of mycobacteriophage TM4 —a transcriptional inhibitor of WhiB2. Mol.Microbiol. 77:642– 657.
33. Singh A, et al. 2009. Mycobacterium tuberculosis WhiB3 maintains redoxhomeostasis by regulating virulence lipid anabolism to modulate macro-phage response. PLoS Pathog. 5:e1000545.
34. Singh A, et al. 2007. Mycobacterium tuberculosis WhiB3 responds to O2
and nitric oxide via its [4Fe-4S] cluster and is essential for nutrient star-vation survival. Proc. Natl. Acad. Sci. U. S. A. 104:11562–11567.
35. Smith LJ, et al. 2010. Mycobacterium tuberculosis WhiB1 is an essentialDNA-binding protein with a nitric oxide-sensitive iron-sulfur cluster.Biochem. J. 432:417– 427.
36. Soliveri JA, Gomez J, Bishai WR, Chater KF. 2000. Multiple paralogousgenes related to the Streptomyces coelicolor developmental regulatory genewhiB are present in Streptomyces and other actinomycetes. Microbiology146:333–343.
37. Steyn AJ, et al. 2002. Mycobacterium tuberculosis WhiB3 interacts withRpoV to affect host survival but is dispensable for in vivo growth. Proc.Natl. Acad. Sci. U. S. A. 99:3147–3152.
38. Tusher VG, Tibshirani R, Chu G. 2001. Significance analysis of microar-rays applied to the ionizing radiation response. Proc. Natl. Acad. Sci.U. S. A. 98:5116 –5121.
39. van Kessel JC, Hatfull GF. 2008. Mycobacterial recombineering. Meth-ods Mol. Biol. 435:203–215.
40. van Kessel JC, Hatfull GF. 2007. Recombineering in Mycobacteriumtuberculosis. Nat. Methods 4:147–152.
41. Vaquerizas JM, et al. 2005. GEPAS, an experiment-oriented pipeline forthe analysis of microarray gene expression data. Nucleic Acids Res. 33:W616 –W620.
42. Venketaraman V, Dayaram YK, Talaue MT, Connell ND. 2005. Gluta-thione and nitrosoglutathione in macrophage defense against Mycobacte-rium tuberculosis. Infect. Immun. 73:1886 –1889.
Casonato et al.
3144 iai.asm.org Infection and Immunity
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/i
ai o
n 20
Oct
ober
202
1 by
119
.192
.83.
17.





























