Embed Size (px)
Citation preview

J Sci Food Agric 1995,68,465-469
Withn-Plot and Within-Plant Variation for Seed Content of Soya Bean Protease Inhbitors Stefan0 Marchetti,* Annalisa Giordano and Cristina Chiaba Dipartimento di Produzione Vegetale e Tecnologie Agrarie, Universita di Udine, Via delle Scienze 208, 33100 Udine, Italy (Received 12 August 1994; revised version received 24 October 1994; accepted 17 March 1995)
Abstract: Experiments were designed to determine the seed content of protease inhibitors in different plants of a plot or at different nodes along the main stem. The inhibitory activity of the samples was measured in uitro using sodium benzoyl-DL-arginine-p-nitroanihde as chromogenic substrate and bovine trypsin as target enzyme. Six soya bean (Glycine max Merr) strains were used, three capable of synthesizing the Kunitz trypsin inhibitor (L62-364, Williams 82, and 594-1) and three lacking this particular function (L81-4590, L81-4871, and L83- 4387). In all genotypes on trial, remarkable plant-to-plant variation was found, so that most strains had at least one part of the data distribution in common. Significant differences were also noted for antitryptic activities of seed samples collected at different positions on the main stem. Gradients interior to the plant varied according to the tested strain but those of Williams 82 and of its near isogenic line L81-4590 were very similar, indicating that the amount and type of variation depended on genotype and that changes in the inhibitory activity were attributable to variation in the synthesis of inhibitors other than Kunitz.
Key words: GIycine max, soya bean, protease inhibitors, antinutritional factors, epigenetic variation.
INTRODUCTION
inhibitors are the most important anti- tional factors in unprocessed soya bean seed. They
are able to form stoichiometric complexes with some digestive enzymes thus preventing their catalytic func- tions. Monogastric animals fed raw soya bean meal show pancreatic hypertrophy, excessive enzyme secre- tion and a metabolic disturbance in the utilisation of the sulphur amino acids (Green et a1 1973; Lyman et a1 1974). Compared with most other proteins, soya bean protease inhibitors are very stable to digestion and heat (Kakade 1974).
Several studies were made on the biochemical proper- ties and on the genetic control of the two major forms of soya bean protease inhibitors, namely the Kunitz trypsin inhibitor and the Bowman-Birk inhibitor family. The former is a 21 kDa protein (also designated as SBTI-A, ; Rackis et a1 1962) which strongly inhibits trypsin. Three variants of the protein are known, Ti", Tib, and Ti'; these are controlled by a codominant
* To whom correspondence should be addressed.
multiple allelic system at locus Ti (Singh et a1 1969; Hymowitz and Hadley 1972; Orf and Hymowitz 1977). In two soya bean accessions, namely PI 157.440 and PI 196.168, no band corresponding to the SBTI-A, protein could be detected; this condition appeared recessive to the alleles previously found (Orf and Hymowitz 1979).
The Bowman-Birk inhibitor family is composed of double-headed low-molecular-weight proteins (8 kDa). Screening of the soya bean germplasm collection led to the discovery of strains lacking some member of this family (Stahlhut and Hymowitz 1983) but, in contrast to the Kunitz inhibitor (Bernard and Hymowitz 1986), no introduction of these characters into nearly isogenic lines has been reported so far.
The biological significance of protease inhibitors is not perfectly known. According to Green and Ryan (1972), they could successfully be used to improve natural resistance in several plant species; actually, in some cases they have shown to act against insects and diseases (Steffens et a1 1978; Gatehouse et a1 1979; Hilder et a1 1987). However, in other experiments there was no evidence of a correlation between inhibitor content and degree of resistance (Sichkar et a1 1983;
465 J Sci Food Agric 00224142/95/%09.00 0 1995 SCI. Printed in Great Britain

466 S Marchetti, A Giordano, C Chiabd
Xavier-Filho et a1 1989); under these circumstances pro- tease inhibitors could simply represent storage proteins or inner elements for the control of proteolysis.
The selection of genotypes with a very low inhibitor content should contribute to clarification of the physio- logical role of these substances. It should be pointed out that, as far as the total antitryptic activity of raw flour is concerned, remarkable genetic variation was found in soya bean (Kakade et al 1972; Sichkar et a1 1983; Lorenzoni et al 1990). A highly significant geno- type x environment interaction was also observed but the magnitude of this did not seem to hamper the choice of parental strains.
Hence, the trypsin inhibitor content of soya bean seed can be modified through specific breeding programmes. Nevertheless, before starting such programmes, there is the need to clarify the amount of variation not attribut- able to genotype in order to specify the sampling and increase selection efficiency. The aim of this study was to provide information about the variability: (i) among plants reared in the same plot and sharing the same genotype, and (ii) within plants according to node posi- tion along the main stem.
EXPERIMENTAL
Soya bean strains
We used six soya bean (Glycine max Merr) strains, three of which provided the Ti" allele (L62-364, Williams 82, and 594-1) and the other three homozygous recessive (ti) at that locus (L81-4590, L81-4871, and L83-4387). The latter strains are nearly isogenic to Williams 82, Clark 63, and Amsoy 71, respectively; they were developed by backcrossing using PI 157.440 (ie Kum-du) as donor parent (Bernard and Hymowitz 1986). L62-364 is a semi-determinate Harosoy isoline derived from the combination with T117.
Except for 594-1, all strains were from the USDA soya bean germplasm collection; the morphological habit, the maturity group, and the pedigree of each are listed in Table 1 (for nearly isogenic lines, the pedigree of the recurrent parent is specified). The number of plants and nodes considered for analysis is also report- ed in Table 1. To study the influence of node position, seed was pooled from at least 20 plants; node position was determined according to Fehr and Caviness (1977).
Plants were grown on a dystric ferralic cambisol at Udine, north-eastern Italy. Seeds were planted on 3 June 1991 with a 0.75 m row spacing at a seeding rate of 20 m-' of row. Plots consisted of single-plant pro- genies allocated in three bordered 4 m rows; a cotyle- don fragment was taken from each plant of the middle row and used to verify the correct allelic composition at locus Ti through isoelectrofocusing (PhastGel IEF 3-9 in a Phastsystem, Pharmacia LKB; 2000 V, 2.5 mA, 3.5 W to a total of 410 Vh; staining with Coomassie R-250 from Sigma Chemical Co in 300 ml litre-' methanol). More than 20 genetic markers of morpho- logical type were also considered to exclude the pres- ence of off-types in the plots. At physiological maturity, plots were trimmed at the edges and plants were har- vested from the middle row. After threshing, the seed was freed from contaminants (wrinkled and mouldy seeds were also discarded), dried to constant weight in a ventilated oven at 35°C and stored at 2°C until analysis. In L62-364 and L83-4387, the protein content of the samples was estimated following AOAC (1984).
Trypsin inhibitor analysis
Soya bean flour was obtained by grinding seed in an analytical mill without raising the temperature. The determination of trypsin inhibitory activity was made according to Smith et al(1980), with some modifications (Lorenzoni et a1 1990). The activity of trypsin (bovine pancreas, 2 x crystallised, Sigma Chemical Co) was
TABLE 1 Stem termination, maturity group, pedigree notes and number of analysis performed at plant and node
level for each strain on trial
Strain Stem termination' Maturity group Pedigreeb N o of plants No of nodes
594- 1 dt2 L62-364 Dt, Williams 82 dt2
I 1 16 9 I1 2 17 10 111 3 20 12
L81-4590 dt2 111 3 39 10 L81-487 1 d t , IV 4 40 10 L83-4387 dt2 I1 5 39 11
a d t , , indeterminate; Dt , , semi-determinate. 1, Evans x Weber; 2, Mandarin Ottawa x A K Harrow; 3, Wayne x (Clark x Adams); 4, Lincoln x Richland; 5, Adams x Harosoy.

Variation in soya bean protease inhibitors 461
measured on raw flour extracts with the chromogenic substrate BAPNA (Na-benzoyl-DL-arginine-p-nitro- anilide; Sigma Chemical Co). A calibration curve was made by increasing the amount of the commercially purified Kunitz inhibitor (SBTI-A2 protein or type I S, Sigma Chemical Co) in the extraction buffer. Therefore, the antitryptic activity shown by a sample was expressed in terms of mg pure trypsin inhibited g-' flour (TI), irrespective of the allelic composition of the strain. For each flour, three independent extracts were used; different dilutions of the extracts were made in order to set the values well inside the calibration curve.
Statistical analysis
Data were submitted to one-way analysis of variance after having checked the normality of the distribution of data by means of Kolmogorov-Smirnov test; means of different plants or nodes were compared with Duncan's multiple range test at the probability level of P = 0.05. A comparison among genotypes was made using Student's t-test; prior to this, we verified the homo- geneity of variances with Bartlett's formula and also the lack of any correlation between variance (or standard deviation) and mean.
RESULTS AND DISCUSSION
In all genotypes on trial, remarkable plant-to-plant variation was found in the seed content of protease inhibitors. In L8 1-4590 the average antitryptic activity was equal to 20.40 TI mg g-' flour but values fluctu- ated within a range of 11 mg TI g-' flour. In the remaining strains, ranges were even higher and at least partially overlapping (Fig 1).
The character showed a normal distribution in both Ti" and ti strains. The possible contribution of analyti- cal errors to this kind of distribution was found to be negligible; actually, in no instance was the coefficient of variation higher than 2.6%. With such
594-1 Williams 82
L62-364 L83-4387
L81-4871
L81-4590
a reliable
1 . I . I . I . I
10 20 30 40 50
mg TI g-1 flour
Fig 1. Range of antitryptic activity found for the strains on trial.
TABLE 2 Seed content of protease inhibitors (mg TI g- ' of flour) in dif-
ferent plants of 594-1
Plant no TI"
1 6 8
12 5 3 2
16 14 15 7
10 11 4
13 9
41.15a 40.10a 38.69b 37.94bc 3744~d 37.06cde 36.90cde 36.68def 35.90ef 35.60fg 3 4 4 g 32.79h 31.20i 3 1.03i 30.50ij 29.40j
Means with a letter in common are not statistically different at P = 0.05.
method of analysis, plants differed statistically in a number of cases. As an example, values recorded for 594-1 are given (Table 2).
On the other hand, when strains were compared among each other, significant differences were observed in a number of cases (Table 3). To allow this compari- son, it was necessary to transform original data into their logarithms because the within-strain mean square and the relative standard deviation were found posi- tively correlated with the mean ( r = 0.912, P = 0-05, and r = 0.934, P = 0.01, respectively).
Despite large differences being noted in the amount of seed yielded by individual plants, the seed content of protease inhibitors was always unaffected by this char- acter. For two strains showing the highest and lowest plant-to-plant variation (L83-4387 and L62-364, respectively), the protein content of the seed was also
TABLE 3
the six strains on trial Average values of antitryptic activity (mg TI g- of flour) in
Strain TI"
594- 1 3 5 4 a Williams 82 33,54a L62-364 29.90b L83-4387 28.60b L81-4871 24.30~ L81-4590 2 0 . m
Means with a letter in common are not statistically different at P = 0.05.
468 S Marchetti, A Giordano, C Chiabb
TABLE 4 Antitryptic activity (mg TI g-' of flour) found in seeds col-
lected at different nodes from 21 plants of L83-4387
Node position TI"
16 17 12 15 13 8
11 10 14 7 9
28.27a 27.51a 26.26b 25.63bc 25.50bc 24.50cd 24.50cd 23.87d 23.24d 23.11d 2 1.48e
@ Means with a letter in common are not statistically different at P = 0.05.
determined; the total antitryptic activity of raw flours was unrelated to percentage or to total protein (%protein x seed yield).
As to the amount of variation found within plants, it was expected that the seed content of protease inhibi- tors would remain fairly constant along the main stem of a plant; according to this hypothesis, variability ought to be very small if compared with that found among plants. On the contrary, differences among anti- tryptic activities found in seeds collected at different nodes were always significant (as an example, values recorded for L83-4387 are presented in Table 4). The within-plant mean square was similar in all genotypes and no correlation between variance and mean was observed.
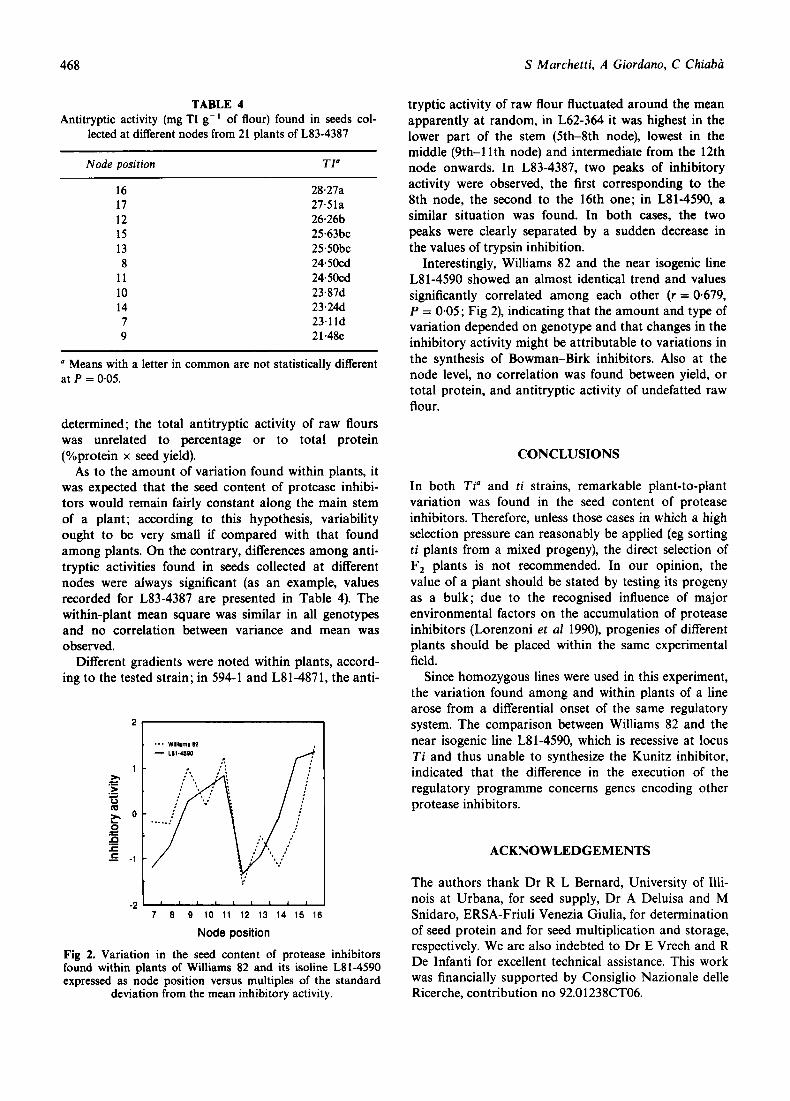
Different gradients were noted within plants, accord- ing to the tested strain; in 594-1 and L81-4871, the anti-
2
. _-. W i l l m a 12 - LB1-46W
-2 " " " " ' 7 8 9 10 11 12 13 14 15 16
Node position
Fig 2. Variation in the seed content of protease inhibitors found within plants of Williams 82 and its isoline L81-4590 expressed as node position versus multiples of the standard
deviation from the mean inhibitory activity.
tryptic activity of raw flour fluctuated around the mean apparently at random, in L62-364 it was highest in the lower part of the stem (5th-8th node), lowest in the middle (9th-1 lth node) and intermediate from the 12th node onwards. In L83-4387, two peaks of inhibitory activity were observed, the first corresponding to the 8th node, the second to the 16th one; in L81-4590, a similar situation was found. In both cases, the two peaks were clearly separated by a sudden decrease in the values of trypsin inhibition.
Interestingly, Williams 82 and the near isogenic line L8 1-4590 showed an almost identical trend and values significantly correlated among each other (r = 0.679, P = 0.05; Fig 2), indicating that the amount and type of variation depended on genotype and that changes in the inhibitory activity might be attributable to variations in the synthesis of Bowman-Birk inhibitors. Also at the node level, no correlation was found between yield, or total protein, and antitryptic activity of undefatted raw flour.
CONCLUSIONS
In both Ti" and ti strains, remarkable plant-to-plant variation was found in the seed content of protease inhibitors. Therefore, unless those cases in which a high selection pressure can reasonably be applied (eg sorting ti plants from a mixed progeny), the direct selection of F, plants is not recommended. In our opinion, the value of a plant should be stated by testing its progeny as a bulk; due to the recognised influence of major environmental factors on the accumulation of protease inhibitors (Lorenzoni et a1 1990), progenies of different plants should be placed within the same experimental field.
Since homozygous lines were used in this experiment, the variation found among and within plants of a line arose from a differential onset of the same regulatory system. The comparison between Williams 82 and the near isogenic line L81-4590, which is recessive at locus Ti and thus unable to synthesize the Kunitz inhibitor, indicated that the difference in the execution of the regulatory programme concerns genes encoding other protease inhibitors.
ACKNOWLEDGEMENTS
The authors thank Dr R L Bernard, University of Illi- nois at Urbana, for seed supply, Dr A Deluisa and M Snidaro, ERSA-Friuli Venezia Giulia, for determination of seed protein and for seed multiplication and storage, respectively. We are also indebted to Dr E Vrech and R De Infanti for excellent technical assistance. This work was financially supported by Consiglio Nazionale delle Ricerche, contribution no 92.01238CT06.

Variation in soya bean protease inhibitors 469
REFERENCES
AOAC 1984 Official Methods of Analysis (14th edn). Associ- ation of Oflicial Analytical Chemists, Washington, DC, USA.
Bernard R L, Hymowitz T 1986 Registration of L81-4590, L81-4871 and L83-4387 soybean germplasm lines lacking the Kunitz trypsin inhibitor. Crop Sci 26 650-651.
Fehr W R, Caviness C E 1977 Stages of soybean development. Iowa State Univ Coop Ext Serv, Agric and Home Econ Exp Stn Spec Rep no 80, Iowa, USA.
Gatehouse A M R, Gatehouse J A, Dobie P, Kilminster A M, Boulter D 1979 Biochemical basis of insect resistance in Vigna unguiculata. J Sci Food Agric 30 949-958.
Green G M, Olds B A, Matthews G, Lyman R L 1973 Protein as a regulator of pancreatic enzyme secretion in the rat. Proc SOC Exper Med Bioll42 1162-1 167.
Green T R, Ryan C A 1972 Wound-induced proteinase inhibi- tors in plant leaves: a possible defence mechanism against insects. Science 175 776-777.
Hilder V A, Gatehouse A M R, Sheerman S E, Barker R F, Boulter D 1987 A novel mechanism of insect resistance engineered into tobacco. Nature 330 160-163.
Hymowitz T, Hadley H H 1972 Inheritance of a trypsin inhib- itor variant in seed protein of soybeans. Crop Sci 12 197- 198.
Kakade M I 1974 Biochemical basis for the differences in plant protein utilization. J Agric Food Chem 22 550-555.
Kakade M I, Simons N R, Liener I E, Lambert J W 1972 Biochemical and nutritional assessment of different varieties of soybeans. J Agric Food Chem 20 87-90.
Lorenzoni C, Marchetti S, Bittolo M, Marzari R, Marocco A, Signor M, Snidaro M 1990 Trypsin inhibitor content in soybean (Glycine max (L.) Merr.) germplasm and com- mercial cultivars. Riu Agron 24 228-236.
Lyman R L, Olds B A, Green G M 1974 Chymotrypsinogen in the intestine of rats fed soybean trypsin inhibitor and its inability to suppress pancreatic enzyme secretions. J Nutr
Orf J H, Hymowitz T 1977 Inheritance of a second trypsin inhibitor variant in seed protein of soybeans. Crop Sci 17
Orf J H, Hymowitz T 1979 Inheritance of the absence of the Kunitz trypsin inhibitor in seed protein of soybeans. Crop Sci 19 107-109.
Rackis J J, Sesame H A, Mann R K, Anderson R L, Smith H K 1962 Soybean trypsin inhibitors: isolation, purification, and physical properties. Arch Biochem Biophys 98 471-478.
Sichkar V I, Levitsky A P, Grikun 0 A, Lobko V N, Marj'ushkin V F 1983 Activity of trypsin and chymotrypsin inhibitors of soybean forms with different resistance to soybean borer. Soybean Genetic Newsletter 10 123-125.
Singh L, Wilson C M, Hadley H H 1969 Genetic differences in soybean trypsin inhibitors separated by disc gel electro- phoresis. Crop Sci 9 489-491.
Smith C, Van Megen W, Twaalfhoven L, Hitchkock C 1980 The determination of trypsin inhibitor levels in foodstuffs. J Sci Food Agric 31 341-350.
Stahlhut R W, Hymowitz T 1983 Variation in the low molecu- lar weight proteinase inhibitors of soybean. Crop Sci 23
Steffens R, Fox F R, Kassel B 1978 Effect of trypsin inhibitors on growth and metamorphosis of corn borer larvae Ostrinia nubilalis Hubner. J Agric Food Chem 26 170-174.
Xavier-Filho J, Campos F A P, Ary M B, Silva C P, Carvalho M M M, Macedo M L R, Lemos F J A, Grant G 1989 Poor correlation between the levels of proteinase inhibitors found in the seeds of different cultivars of cowpea (Vigna unguiculata) and the resistance/susceptibility to predation by Callosobruchus maculatus. J Agric Food Chem 37 1139- 1143.
62 105-110.
81 1-813.
766-769.





























