Embed Size (px)
Citation preview

Aus der Bundesforschungsanstalt f u r Viruskrankheiten der Tiere, Tiibingen am Neckar
Zur Pathogenese der Schweinepesterkrankung von der Infektion bis zum Temperaturmaximum und iiber die
Serumrnakroglobuline
Von
W. MATTHAEUS und G. KORN
M i t 5 Abbildungen
(Eingegrngen am 11 . ] d i 1968)
In vorangegangenen Arbeiten untersuchten wir in bestimmten Phasen bei der Schweinepest(SP)-Erkrankung die klinischen Reaktionen, die Antikorper- (AK)-Bildung und die generellen Serumproteinveranderungen (20, 27, 28).
Das typische Kriterium der SP-Erkrankung war die Leukopenie bei steigender Temperatur mit einem erheblichen a,-Globulinanstieg und tief- greifenden Veranderungen im p-Globulinbereich. Die zuerst auftretenden neutralisierenden (neutr.) Aktivitaten wandern elektrophoretisch wie ein /je- Globulin und lassen sich bei Kombination von Gelfiltration und Ionenaus- tauschchromatografie des Serums als 0,-Makroglobulin (&M) nachweisen (28). Diese Makroimmunglobuline (IgM) interessieren insofern, als sie schon wah- rend des Temperaturanstiegs produziert werden, d. h. zu einem Zeitpunkt, in dem sich die fur die SP spezifischen Krankheitsvorgange abspielen. In ihrer Anwesenheit entwickeln sich z. B. die morphologisch erkennbaren geweblichen Veranderungen, wie solche der Gefaflwand oder perivaskulare lymphoide Infiltrate mit ihren Folgeerscheinungen sowie die Leukopenie und Anamie (27).
Die folgenden Untersuchungen befassen sich bei einer experimentellen Infektion mit vollvirulentem Schweinepestvirus durch subkutane Applikation und Kontakt mit den klinischen, serologischen und immunologischen Befunden der ersten Infektionstage bis zum Temperaturmaximum.
Material und Methoden 1 . Virusstamm. SP-Virus (SPV) 274 (15. Tag p. i.) v. 29. 5 . 64 vollviru-
lent. 2. Die Auftrennung der Serumproteine erfolgte in der Agarelektropho-
rese (25) und durch Gelfiltration (26, 28). Die Charakterisierung der Frak- tionen in der Immunoelektrophorese (26). Zur Bestimmung des absoluten Proteingehaltes im Serum diente die Biuretmethode.
3. Die Virus-Serumneutralisationsversuche wurden mit der HEIC- Methode (37) durchgefuhrt. Die Inkubation der Virus-Serumgemische erfolgte fur 1 Std. bei + 4' C und die Bestimmung des Neutralisationsindex (NI) rnit

Zur Pathogenese der Schweinepesterkrankung 313
der Virusverdunnungsmethode. Die virushaltigen Seren und die Fraktionen wurden 90 Min. bei + 56O C inaktiviert. Ein Normalserum und ein bekann- tes Immunserum (NI 1,6) liefen als Kontroll- bzw. als Vergleichsserum bei jedem Neutralisationsversuch mit. Ein NI von 0,6 und mehr kann nach unse- ren Erfahrungen als echte neutr. Wirksamkeit angesprochen werden.
4. Bei dem Agargelpriizipitationstest (AGPT) (36) wurden die Seren auf das Auftreten von prazipitierenden (praz.) AK (4) gegen ein positives Pankreasantigen gepriift.
5. Die Blutziihlungen erfolgten nach HALLMANN ( 1 l ) , die Furbung nach PAPPENHEIM.
6. Die Infektion der Versuchstiere geschah bei Nr. 301 (30 kg) und Nr. 303 (26 kg) subkutan (sk.). Die Tiere Nr . 302 (24 kg) und Nr . 304 (21 kg) wurden zur Kontaktinfektion (K) hinzugestellt. Alle Tiere wurden tfglich auf ihren klinischen und serologischen Status untersucht.
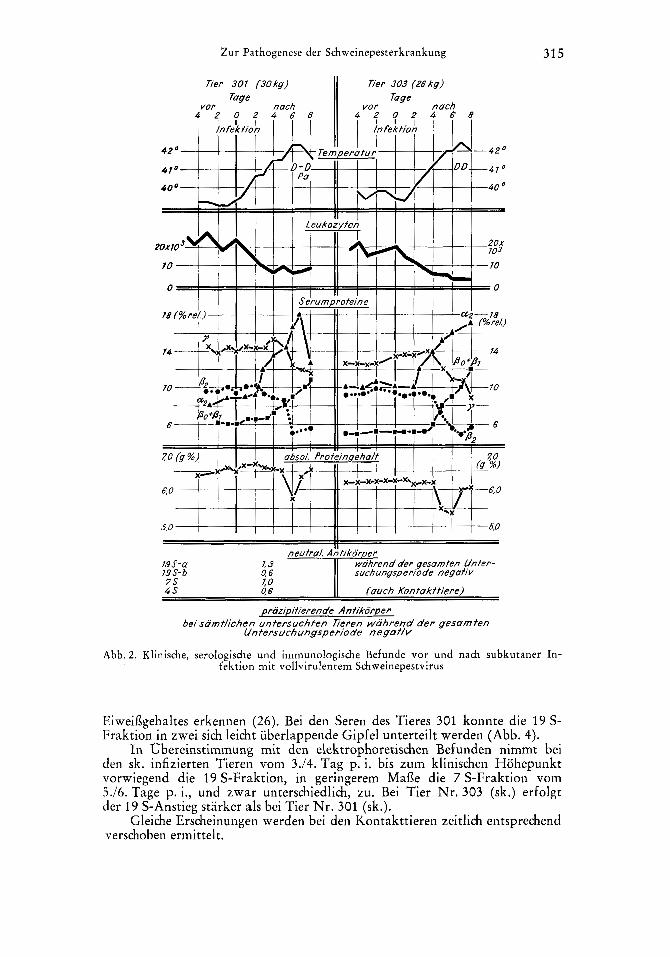
Befunde und Ergebnisse Klinische Befunde und Blutbilder. Die sk. mit vollvirulentem SPV infi-
zierten Tiere zeigten ab 2. Tag, die Kontaktkontrollen etwa 1 Woche spater (8./9. Tag) Temperaturerhohung und Leukozytenabfall. Die Temperaturen stiegen im Verlauf der Infektion auf uber 42O C. Mit Ausnahme des Tieres Nr. 301 (sk.) sanken bei allen anderen Tieren die Leukozyten auf Werte unter 4000 und wiesen bis zum Versuchsende eine standig fallende Tendenz auf. Hiervon waren anfanglich mehr die Lymphozyten, spater die Granulozyten starker betroffen. Die bei Tier Nr. 301 (sk.) ab 5. Tag p. i. beobachtete leichte Erhohung der Leukozyten war speziell auf eine Zunahme der neutrophilen Granulozyten zuruckzufuhren (Tab. 1 u. Abb. 2). Diese waren am 8. Tage p. i. so stark angestiegen, dafi sie etwa 81 o/o der Gesamtleukozyten aus- mach ten.
Das andere sk. infizierte Tier 303 zeigte am gleichen Tage nur 1000 Leu- kozyten, von denen 33 o/o intakte Lymphozyten, 58 O / o jedoch nekrobiotische Zellen waren. Diese konnten nur noch z. T. als Lympho- bzw. Granulozyten erkannt werden (Abb. I). Der Rest, der als Leukozyten gezahlten kernhaltigen Zellen, waren Normoblasten.
Serumproteinuntersuchungen Die allgemeinen Proteinveranderungen wahrend der Infektion wurden in1
Gesamtserum mittels der Agarelektrophorese verfolgt und nach dessen Auf- trennung durch Gelfiltration an Sephadex G-200, die davon betroffenen Serumeiweifikomponenten mit der Agar- und Immunoelektrophorese identifi- ziert und charakterisiert.
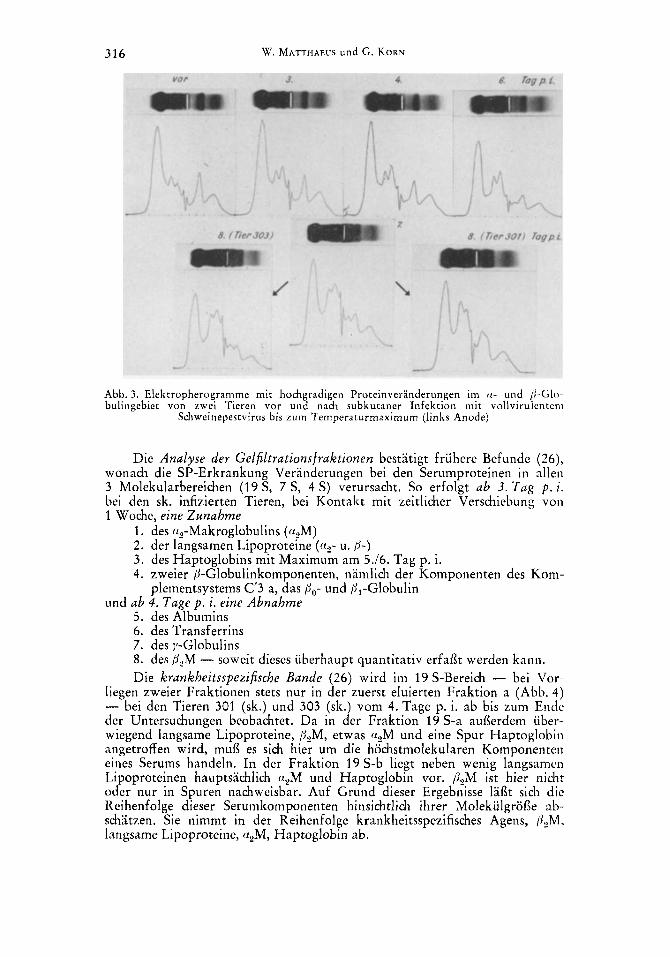
a) Agarelektrophorese. Einen Tag nach dem Temperaturanstieg und dern Leukozytenabfall, d. h. bei den sk. infizierten Tieren am 3. Tag p. i., bei den Kontakttieren wieder etwa um 1 Woche verschoben (9.110. Tag p. i.), steigt das Gesamt-(1-Globulin durch Zunahme des a,-Globulins an und bleibt bei 3 Tieren bis zum Totungstage vermehrt. Das eine sk. infizierte Tier (301) zeigt vom 7. zum 8. Tag p. i. einen heftigen a,-Globulinabfall. Ab 4. Infektionstag (sk.), mit einwochiger Verschiebung bei Kontaktinfektion, nehmen auch die anderen a-Globulinfraktionen zu. Zum gleichen Zeitpunkt setzen leichte Ver- anderungen im /%Globulingebiet ein, die in einer Zunahme des Po- und/oder /il-Globulins und einer /j,-Globulinabnahme resultieren und bei sk. Infektion Ende der 1. bzw. Anfang der 2. Infektionswoche - bei Kontakt 1 Woche spa- ter - ihren Hohepunkt erreichen (Abb. 3). Ab 4./5. Tag p. i. - bei den Kon- tnkttieren 6 Tage spater - wird eine geringe Verminderung des gesaniten

314 W. MATTHAEUS und G. KORN
Lymphozyten Granulozyten
Abb. 1. Blutleukozyten mit Kern- zerfall 8. Tag p. i. (Tier 303) Gesamtleukozytenzahl 1000/mm3. 33 O/a Lymphozyten, 58 "/o nekro- biotische Zellen, Rest Normo-
blasten
y-Globulingehaltes hauptsachlich durch Abnahme des yl-Globulins bis zum Totungstage beobachtet (Abb. 2).
b) Gelfiltration. Aus dem Kuruenverlauf, d. h. aus den Verschiebungen der Extinktionshohen der durch Gelfiltration aufgetrennten Seren lassen sich - unter der Voraussetzung streng eingehaltener Versuchsbedingungen - die in den einzelnen Molekulargewichtsbereichen (6 , 7) infektionsbedingten Ver- Znderungen hinsichtlich dieser Serumproteingruppen als auch ihres absoluten
Zur Pathogenese der Schweinepesterkrankung
?#$-a t 3 79 S- b 0 6
4s $6 7s 7,O
315
wahrend der gesamfen Unfer- suchungsperiode negativ
Couch Kontcrk f fiere)
Eiweiflgehaltes erkennen (26). Bei den Seren des Tieres 301 konnte die 19 S- Fraktion in zwei sich leicht uberlappende Gipfel unterteilt werden (Abb. 4).
In Obereinstimmung mit den elektrophoretischen Befunden nimmt bei dcn sk. infizierten Tieren vom 3./4. Tag p. i. bis zum klinischen Hohepunkt vorwiegend die 19 S-Fraktion, in geringerem Mafle die 7 S-Fraktion vom 5. /6 . Tage p. i., und zwar unterschiedlich, zu. Bei Tier Nr. 303 (sk.) erfolgt der 19 S-Anstieg starker als bei Tier Nr. 301 (sk.).
Gleiche Erscheinungen werden bei den Kontakttieren zeitlich entsprechend verschoben ermittelt.
316 W. MATTHAEUS und G . KORN
A
Abb. 3. Elektropherogramme mit hochgradigen Proteinveranderungen im a- und P-Glo-- bulingebiet von zwei Tieren vor uiid nach subkutaner Infekt ion mit vollvirulenteni
Schweinepestvirus bis zum Temperaturmaximum (links Anode)
Die Analyse der Gelfiltrationsfraktionen bestatigt fruhere Befunde (26), wonach die SP-Erkrankung Veranderungen bei den Serumproteinen in allen 3 Molekularbereichen (19S, 7 S, 4 S) verursacht. So erfolgt a b 3 . T a g p . i. bei den sk. infizierten Tieren, bei Kontakt mit zeitlicher Verschiebung von 1 Woche, eine Z u n a h m e
1. des qMakroglobulins (cc,M) 2. der langsamen Lipoproteine ( ( I? - u. /j-) 3. des Haptoglobins mit Maximum am 5.16. Tag p. i. 4. zweier /j-Globulinkomponenten, namlich der Komponenten des Kom-
plementsystems C'3 a, das Pa- und /jl-Globulin und a b 4 . Tage p . i. eine A b n a h m e
5. des Albumins 6. des Transferrins 7. des y-Globulins 8. des &M - soweit dieses uberhaupt quantitativ erfaf3t werden kann. Die krankheitsspezifische Bande ( 2 6 ) wird im 19 S-Bereich - bei Vor-
liegen zweier Fraktionen stets nur in der zuerst eluierten Fraktion a (Abb. 4) - bei den Tieren 301 (sk.) und 303 (sk.) vom 4. Tage p. i. ab bis zurn Ende der Untersuchungen beobachtet. Da in der Fraktion 19 S-a auflerdem iiber- wiegend langsame Lipoproteine, &M, etwas u,M und eine Spur Haptoglobin angetroffen wird, mufl es sich hier um die hochstmolekularen Komponenten eiiies Serums handeln. In der Fraktion 19 S-b liegt neben wenig langsamen Lipoproteinen hauptsachlich rx,M und Haptoglobin vor. /j,M ist hier nicht oder nur in Spuren nachweisbar. Auf Grund dieser Ergebnisse lafit sich die Keihenfolge dieser Serumkomponenten hinsichtlich ihrer Molekiilgrofle ab- schatzen. Sie nimmt in der Reihenfolge krankheitsspezifisches Agens, P,M, langsame Lipoproteine, urM, Haptoglobin ab.

Zur Pathogenese der Shweinepesterkrankung 317
Als weitere auffallige Veranderung verschwindet wahrend der Infektion bei immunoelektrophoretischer Untersuchung in der 7 S-Fraktion der sk. infi- zierten Tiere ab 3./4. Tag p. i. bis zum Ende der Untersuchungen (8. Tag p. i.) eine vor dem Albumin gelagerte, lang gestreckte Bande (Praalbumin?) ganz oder teilweise (Abb. 5). Da diese Bande bei Verwendung von Kaninchenanti- seren, die sowohl gegen kalberserumhaltiges Kulturschweinepestvirus als auch gegen normales Schweineserum hergestellt wurden, gebildet wird, mui3 ange- nommen werden, dai3 diese Komponente als normaler Bestandteil des Schwei- neserums zu gelten hat.
Neutralisievende Antikovpev werden in den Gelfiltrationsfraktionen mit Sicherheit bei Tier 301 (sk.) ab 6 . Tag p. i. gefunden, wobei die hochmoleku- laren IgM in der Ultrazentrifuge mit 19 S sedimentieren (30) und gegenuber
7s I 4 s
19 s
a b
I
roo r, \ -
m I Eluat
Abb. 4. Elutionskurve eines Serums des Tieres 301 (sk.) bei der Gelfiltration an Sephadex G-200

318 W. MATTHAEUS und G. KORN
den niedermolekularen AK der 7 S-Gelfiltrationsfraktion vom Typ IgA vor- herrschen. AK vom Typ IgG konnten nicht nachgewiesen werden (28). Weder bei Tier Nr. 303 (sk.) noch den beiden Kontakttieren lieflen sich neutralisie- rende Aktivitaten ermitteln (Abb. 2).
Im Agavgelpriizipitationstest mit dem Pankreasantigen konnen bei kei- nem der infizierten Tiere, weder im Gesamtserum noch in den Fraktionen der Gelfiltration, praz. AK nachgewiesen werden.
c) Absoluter Proteingehalt des Serums. Der mit der Biuretmehtode taglich ermittelte absolute Serumproteingehalt der sk. infizierten Tiere zeigte bei fast gleichbleibender Tendenz bis zum 3. Tage p. i. am 5. und/oder am 6. Tage p. i. einen plotzlichen Abfall um etwa 0,8-1 ,O gO/u gegenuber den vorangegange- nen und folgenden Tagen. Am Tage des Minimums betrug der durchschnitt- liche absolute Proteingehalt 5,40 go/u.
Dieser Abfall des absoluten Proteingehaltes ging rnit einem ebenso plotz- lichen des Transferrins konform, der bei beiden sk. infizierten Tieren auch um 1 Tag verschoben einsetzte, aber gleichzeitig mit dem des absol. Proteingehal- tes erfolgte (Abb. 2). Diese Befunde wurden durch Analysen des Verlaufs der Gelfiltration und ihrer Fraktionen bestatigt. Danach handelte es sich um eiiie generelle Proteinabnahme in allen 3 Molekulargewichtsbereichen rnit Schwer- punkt auf dem 4 S-Bereikh, Hier wurde eine bevorzugte und relativ lebhafte Albumin- und Transferrinverminderung festgestellt.
Bei den Seren der Kontakttiere trat im absoluten Proteingehalt am 9./10. Tage nach Kontakt ein gleicher Ruckgang auf, dem ein ahnlicher des Transferrins wie oben beschrieben folgte.
Besprechung der Befunde Leukozyten
Bei den Leukopenien sind auch in vorliegenden Fallen alle Zellarten be- troffen (22, 27). Die Lymphozyten fallen zwar zahlenmaflig in den ersten Infektionstagen schneller ab, do& sind sie bei den moribunden Tieren in grofle- rer Zahl als die Granulozyten vorhanden. Wie die Blutausstrichpraparate von Tier Nr . 303 (sk.) (Tab. 1, Abb. 1) zeigen, gehen die Leukozyten im stromen- den Blut zugrunde. Nach LETTERER (48) hat die Steigerung der Zvtolyse im Blut von sensibilisierten Tieren und Menschen als morphisches Substrat den kugeligen segmentierten ,,Granulozyt", dessen Kern keine Strukturzeichnung mehr zeigt und dessen Zytoplasma ausgedehnte Vakuolisierung aufweist. Einen antileukozytaren Serumfaktor konnte PEHL (39) nicht nachweisen. So sieht er die Leukopenie sowie die oft vorhandene Anamie in erster Linie als Ausdruck einer Storung des hamatopoetischen Apparates gn, bei der die Leuko- und Erythropoese gleichermaflen betroffen sind. Er bewichnet diese Storung als symptomatische, reversible Panmyelose mit akuter Regenerationsstorung. Als Ursache einer derartigen Organsystemstorung fuhrt LETTERER (21) fur das Knochenmark ,,&were allergische Funktionsstorungen" an, die mit Bildungs- hemmung und Untergang seiner Zellen (Agranulozytose, Panmyelopathien) einhergehen. Mit derartigen Funktionsstorungen kann zwar der Abfall der Leukozyten erklart werden, nicht jedoch ihr Untergang im stromenden Blut. Fur diese Zellzerstorung ist in vivo moglicherweise eine direkte Viruswirkung auszuschlieflen, da das SP-Virus in Blutleukozytenkulturen (3, 23) ohne zyto- pathischen Eff ekt vermehrt wird. Die Ursache der Leukozytenzerstorung kann mit der Entwicklung der zellularen Immunitat (24) in Zusammenhang ge- bracht werden, worauf in einer weiteren Arbeit eingegangen wird.
Wahrend bei den ubrigen Tieren die Leukozytenwerte bis unter 2000 ab- fallen, steigen diese bei Tier 301 (sk.) ab 5. Tag p. i. wieder an. Die Ursache
Zur Pathogenese der Schweinepesterkrankung 319
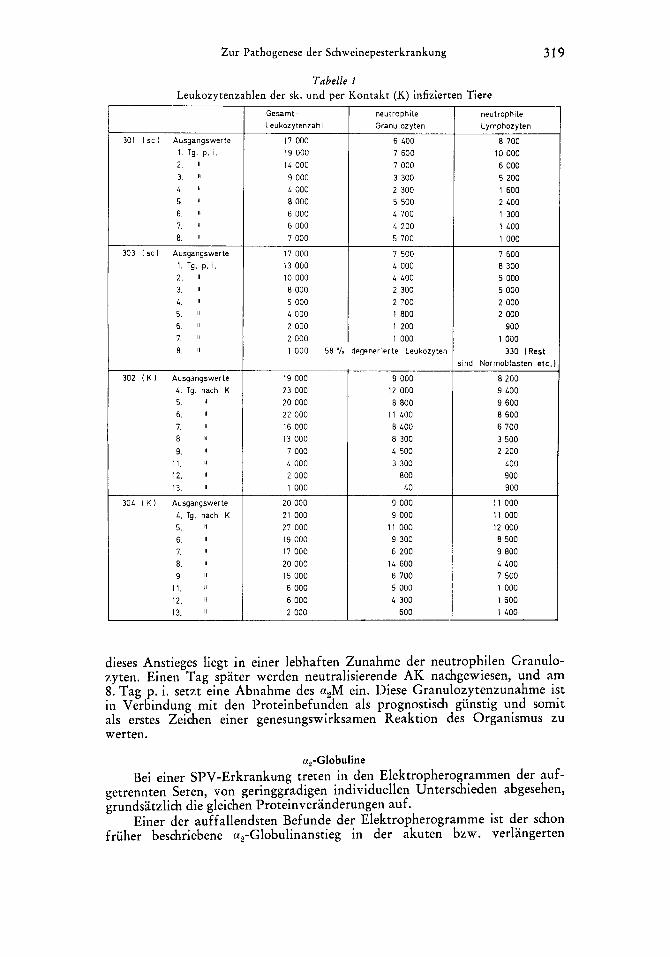
Tabelle 1 Leukozytenzahlen der sk. und per Kontakt (K) infizierten Tiere
301 I sc ) Ausgangswerte I . Tg. p. i. 2 11
3. If
L , 11
5. 11
6. 11
7. 8 1
8.
303 I sc 1 Ausgangswerte 1. Tg. p. i. 2. 11
3. 11
4. 11
5. 11
6. 11
7. 11
8. 11
302 ( K 1 Ausgangswerte L. Tg. nach K 5. 11
6. 11
7. 11
8. 11
9. 11
1 1 . 11
12. 11
13. 11
304 I K 1 Ausgangswerte 4 , Tg. nach K 5. 11
6. 11
7. 11
8. 11
9. 11
1 1 . 11
12. ‘1
13. 11
Gesarnt - Leukozytenzahl
17 000 19 000 1L 000 9 000 L 000 8 000 6 000 6 000 7 000
17 000 13 000 10 000 8 000 5 000 4 000 2 000 2 000
neutrophile Granulozyten
6 LOO 7 600 7 000 3 300 2 300 5 500 L 700 L 200 5 700
7 500 L 000 L LOO 2 300 2 700 I 800 I 200 1 000
1 000 58 % degenerierte Leukozyten
19 000 23 000 20 000 22 000 16 000 13 000 7 000 L 000 2 000 1 000
20 000 21 000 27 000 19 000 17 000 20 000 15 000 6 000 6 000 2 000
9 000 12 000 8 800
I 1 LOO 8 LOO 8 300 L 500 3 300
800 LO
9 000 9 000
1 1 000 9 300 6 200
14 600 6 700 5 000 L 300
600
neutrophile Lyrnphozyten
8 700 10 000 6 000 5 200 1 600 2 LOO 1 300 1 4 0 0 1 000
7 600 8 300 5 000 5 000 2 000 2 000
900 1 000
330 [Rest i ind Normoblasten etc.
8 200 9 LOO 9 600 8 600 6 700 3 500 2 200
LOO 900 900
1 1 000 1 1 000 12 000 8 500 9 800 L LOO 7 500 I 000 1600 1 LOO
dieses Anstieges liegt in einer lebhaften Zunahme der neutrophilen Granulo- zyten. Einen Tag spater werden neutralisierende AK nachgewiesen, und am 8. Tag p. i. setzt eine Abnahme des a,M ein. Diese Granulozytenzunahme ist in Verbindung mit den Proteinbefunden als prognostisch giinstig und somit als erstes Zeichen einer genesungswirksamen Reaktion des Organismus zu werten.
a,-Globuline Bei einer SPV-Erkrankung treten in den Elektropherogrammen der auf-
getrennten Seren, von geringgradigen individuellen Unterschieden abgesehen, grundsatzlich die gleichen Proteinveranderungen auf.
Einer der auffallendsten Befunde der Elektropherogramme ist der schon friiher beschriebene u,-Globulinanstieg in der akuten bzw. verlangerten

320 W. MATTHAEUS und G. KORN
Krankheitsphase. Nach WUHRMANN und MARKI (47) entspricht der (1,-Globu- linanstieg den Reaktionskonstellationen der akuten und chronischen Entzun- dung, und sie finden dazu eine Korrelation zu der Leukozytenvermehrung. In der leukozytaren Reaktion und den Bluteiweifiveranderungen sehen sie ver- schiedene Erscheinungsformen eines einheitlichen Geschehens. BERG und Mit- arbeiter (2) zeigen, dafi nicht die Zahl der Leukozyten, sondern ihre Aktivitat bezuglich der alkalischen Leukozytenphosphatase mit der a,-Globulinzunahme korreliert. Die alkalische beukozytenphosphatase greift in die De- und Trans- phosphorylierung sowie Hydrolyse und Synthese der Monoester der Phosphor- saure ein. Diskutiert wird auch die Beteiligung der alkal. Leukozytenphospha- tase an der eigentlichen Stoff wechsel( ,,intrinsic metabolic")-Aktivitat der Zelle, wie der Glykogenese oder der Nukleinsauresynthese (45).
In vorliegenden Untersuchungen wird der a,-Globulinanstieg bei der SP-Erkrankung im wesentlichen als ein Anstieg des a,M und Haptoglobins festgestellt. Als biologische Funktionen des u,M sind nach SCHULTZE und HEREMANS (41) folgende bekannt:
I . Eine fur selbstregulierende Prozesse (homeostatic processes) wichtige Schutzwirkung fur Trypsin und Plasmin (8, 9, 17, 18, 35) durch Komplex- bildung unter Hemmung, jedoch Erhaltung der enzymatischen Aktivitat;
2. Hemmung der Thrombinwirkung ohne Beeinflussung der Esterase- aktivitat (18 , 43);
3. Verhinderung eines tryptischen Abbaus des PIC-Globulins (Kompo- nente des Komplementsystems C'3 a) infolge Komplexbildung des Trypsins mit dem a,M (9);
4. Bindungsvermogen fur Insulin und andere Substanzen mit niedrigem Molekulargewicht (14);
5. Forderung des Oberlebens rontgenbestrahlter Mause tmd von hamato- poetischem Gewebe (12).
Schon fruher fanden wir den Genesungsprozefi von der rechtzeitigen Bil- dung von AK (27) abhangig, die bei verlangertem klinischem Hohepunkt ver- zogert auftraten. Der verlangerte klinische Hohepunkt war aber immer init einer a,-Globulinerhohung gekoppelt und, wie vorliegende Untersuchungen ergeben, mit einer solchen des a,M. Wahrend bei den drei moribunden Tieren, die am Ende der Untersuchungsperiode nur noch 2000 oder weniger Leuko- zyten und keine neutr. AK aufweisen, das u,M bis zur Totung erhoht vor- liegt, sinkt dieses beim Tier Nr. 301 (sk.) nach dem Anstieg der Leukozyten - speziell der neutrophilen Granulozyten - und dem Auftreten neutr. AK schlagartig ab.
Die hier im Infektionsablauf in Erscheinung tretenden Unterschiede, d. h. bei Nr . 301 (sk.) die Regeneration der neutrophilen Granulozyten, der ver- zijgerte sprunghafte Abfall des Transferrins am 6 . Tage p. i. gegenuber dem 5. Tage p. i. Nr. 303 (sk.) und der plotzliche Ruckgang des a,M am 8. Tage p. i. wirken sich demnach vorteilhaft fur den Genesungsprozefi aus.
Auch in fruheren Untersuchungen (27) wurde mit beginnendem Heilungs- prozefi eine (1,-Globulinabnahme ermittelt. So zeigten in einer Oberlebens- gruppe vier von sechs Tieren einen vorubergehenden a,-Globulinanstieg und zwei uberhaupt keine u,-Glqbulin- und somit auch keine a,M-Veranderung. 11 schwerkranke Tiere, die spater verendeten, besafien dagegen aufier erhohter Temperatur und einer verlangerten Leukopenieperiode bis zum Totungstage einen erhohten (1,-Globulingehalt.
Aus unseren Befunden ist zu folgern, dafi dem u,M bei der SP-Erkran- kung eine Bedeutung zugesprochen werden mufi, doch bleiben Ursprung und Wirkungsweise vorerst ungeklart. Nach neueren Untersuchungen (1) an einem

Zur Pathogenese der Schweinepesterkrankung 321
vorgereinigten Praparat von u,M des Schweineserums uber seine Zusammen- setzung und Struktur werden etwa 20 O/o eines Dissoziationsproduktes neben Haptoglobin und Coeruloplasmin gefunden.
Unter dem Aspekt der a,-Globulinerhohung in der akuten und der ver- Iangerten akuten Phase der SP-Erkrankung sol1 die Vermutung von GRALHEER und PEHL (10) nicht unerwahnt bleiben, die dem SPV auf Grund seiner elek- trophoretischen Mobilitatsunterschiede im Serum und in der Gehirnflussigkeit cine Aviditat zum a,-Globulin zuschreiben. Obwohl nach unseren Unter- suchungen eine Beziehung zwischen dem u,-Globulin, insbesondere dem a,M, und dem SPV zu bestehen scheint, kann eine beiderseitige Adsorption ausge- schlossen werden, da gereinigte SPV-Praparate frei von a-Globulinkompo- nenten, das gleiche elektrophoretische Wanderungsverhalten und dieselbe Pra- zipitationslinie in der Immunoelektrophorese wie ungereinigte Praparate (29) besitzen. Welcher Art der beiderseitige Einflui3 sein konnte, bleibt z. Z. offen. Auffallend hierbei ist jedoch der schon erwahnte, bis in die verlangerte akute Phase erhohte u,M-Gehalt und der gleichzeitige Nachweis der krankheits- spezifischen Bande, die erstmalig am 4. Tag p. i. bei sk. Infektion nach begin- ncndem Serum-u,M-Anstieg, der am 3. Tag p. i. einsetzt, und wahrend der akuten und verlangerten akuten Phase zu beobachten ist.
Das wahrend der Infektion zunehmende Haptoglobin zieht sich bei der Gelfiltration vom absteigenden Ast der 19 S-Fraktion bis in die 7 S-Fraktion. Durch Nachweisreaktionen wurde dieses Eiweii3 als Haptoglobin identifiziert, was mit den Befunden von FIREMAN und Mitarbeiter ( 5 ) sowie WIRTH und Mitarbeiter (46) an Humanseren iibereinstimmt. AuBer uber die bekannte Bindungskapazitat fur Hamoglobin ist uber eine spezifische Funktion bei Infektionskrankheiten z. 2. nichts bekannt.
Der ab 3. Tag p. i. bei sk. Infektion beobachtete Anstieg der lungsumen Lipoproteine stimmt mit den Befunden von HANTSCHEL (13) uberein, der einen solchen am 5 . Tage p. i. findet. Lipoproteine besitzen Transportfunktio- lien fur Fette, Lipoide, fettlosliche Vitamine und Hormone (14), fungieren durch Elektronenanregung als Obertrager von Energie und anderen Reizen und verandern die Permeabilitat der Zellmembranen. Inwieweit diese Funk- tionen eine Virusinfektion, speziell die der SP-Erkrankung, beeinflussen, ist noch nicht bekannt.
8-Globuline Im Bereich der @-Globuline steht der starke Abfall des @,-Globulins ab
4. Tage p. i. (sk.) mit einer Transferrinabnahme (26) in Verbindung. Trans- ferrin dient dem Eisentransport ( 1 5 ) und enthalt etwa 1 O/o des Gesamtbestan- des an Eisen. Bei Eintritt eines Infektes fallt der Serumeisenspiegel ab und bleibt wahrend der Fieberperiode niedrig. Dies wird auch bei malignen Tumo- ren, schweren Allergien und Injektion von artfremden Eiweii3en und Giften beobachtet, wobei die Eisenabwanderung aus dem Blut haufig durch Hamoside- rinablagerung im RES nachweisbar (16) und auch bei der SP (31) beschrieben ist.
Als das Transferrin bildende Zellelemente werden bei der Ratte Zellen des peripheren Blutes, Knochenmarkzellen, Mastzellen und Makrophagen aus der Peritonealhohle und der Lunge sowie lymphoides Gewebe genannt. Letzte- res wird bei Primaten als Transferrin-synthetisierendes Gewebe nicht beob- achtet (44). Fur das Schwein liegen keine Untersuchungen vor, doch konnen ahnliche Orte angenommen werden. Bei der SP-Erkrankung werden Leuko- zyten als Zellen des peripheren Blutes (3, 23) sowie Lymphknoten (38), beides also fur die Transferrinbildung genannte Zellelemente bzw. Organe, als Orte

322 W. MATTHAEUS und G . KORN
der Virusvermehrung angesehen. Hier konnte somit die extreme Transferrin- abnahme mit konkurrierenden Prozessen erklart werden.
Der Abfall des Transferrins in der 4 S-Fraktion der Gelfiltration steht in Synergismus mit einer Verminderung des y-Globulins und einem Anstieg des /jo- und @,-Globulins der 7 S-Fraktion. Die Analysen dieser beiden 7 S-p- Globuline rnit der Immunoelektrophorese und Gelfiltration ergeben nach Ver- gleich mit den Ergebnissen an Humanseren, dai3 es sich hierbei um die Kompo- iienten B,A- und /jlC-Globulin des Komplementsystems C’3 a handelt.
Krankheitsspezifische Bande Die krankheitsspezifische Bande (26) mui3 als die hochstmolekulare Kom-
ponente des infektiosen Serums angesehen werden. Sie wird bei sk. Infektion ab 4. Tag p. i. beobachtet und im klinischen Hohepunkt auch von HANTSCHEI. (13) beschrieben. Uber ihre Identitat mit dem Virus kann eine endgultige Aus- sage nicht gemacht werden, doch sprechen weitere noch nicht veroffentlichte Untersuchungsergebnisse eher fur als gegen eine solche (29).
Praalbumin Fur die vor dem Albumin gelagerte, lang gestreckte Bande (Praalbu-
min?), die im klinischen Hohepunkt der SP-Krankheit verschwindet, gibt es augenblicklich keine Erklarung.
Absoluter Proteingehalt Im absoluten Proteingehalt des Serums tritt bei den schwerkranken Tieren
Nr. 301 (sk.) und 303 (sk.) am 5./6. Tag p. i. ein deutlicher Abfall auf, der niit dem Zeitpunkt einer starken Transferrinverminderung, einer y-Globulin- abnahme und dem Anstieg der P-Globulinkomponenten des Komplement- systems C’3 a zusammenfallt.
Der Literatur entnehmen wir, dai3 Plasmaproteine wie Albumin, die Iinmunglobuline (IgM, IgA, IgG), Praalbumin, u,M, Transferrin und Fibrino- gen generell in den Duodenal- und Jejunalsaften nachgewiesen wurden. Diese werden uber die interstitiellen Raume und die abfuhrenden Lymphgefai3e des Coriums z. T. ins Darmlumen verloren (41). Als Ursache eines vermehrten Austrittes nicht nur von Flussigkeit im Sinne einer Udembildung, sondern auch voli Serumbestandteilen aus den GefPi3en kommen die bei der SP-Erkrankung beschriebenen Permeabilitatsstorungen in Betracht. Diese treten individuell verschiedengradig auf und werden generell als Folge eines veranderten funk- tionellen Verhaltens der Gefafie angesehen, fur deren Ursache die ermittelte Lipoproteinzunahme u. a. nicht ausgeschlossen scheint. Des weiteren werden diese GefaGveranderungen als Folge hormonaler Storungen (33), von Schock- wirkung (42), eines gestorten Kreislaufs (40) bzw. als immunpathologischer Vorgang (19) diskutiert.
Die von MATTHIAS und SCHMIDT (32, 34) beschriebene Viskositatser- hohung des Blutplasmas bei der SP kurz vor dem Exitus steht unseres Er- achtens nicht mit den vorliegenden Befunden in Widerspruch.
Die bei steigendem (1,- und absinkendem @,-Globulin (Transferrin) ge- ringfugig abfallenden y-Globulinwerte aller Tiere deuten auf einen spezifi- schen 7-Globulinverbrauch bei der SP-Erkrankung hin, fur den vermutlich Antigen-Antikorper-Komplexbildungen und deren Ablagerung verantwort- lich sind.
Neutralisierende Antikorper Eine neutr. Aktivitat Lei3 sich nur in den Gelfiltrationsfraktionen des
Tieres 301 (sk.) nach dem Anstieg der neutrophilen Granulozyten und der /&-lB1-Globuline einen Tag vor dem Abfall des u,M nachweisen. In Uber-

Zur Pathogenese der Schweinepesterkrankung 323
einstimmung mit friiheren Angaben (28) wurden zu diesem Zeitpunkt vor- wiegend hochmolekulare AK vom IgM-Typ neben niedermolekularen AK vom IgA-Typ mit rl-Globulincharakter gebildet. Die neutr. Aktivitat der hochmolekularen AK fand sich bei diesem Tiere, bei dem die Seren in der Gel- filtration in 2 Subfraktionen des 19 S-Bereiches fraktionierbar waren, in der zuerst eluierten j3,M-reichen Fraktion 19 S-a. Keine oder eine nur geringe neutr. Aktivitat vies die wenig &M und vornehmlich a,M enthaltende Sqb- fraktion 19 S-b auf. Aus diesen Ergebnissen eht hervor, dai3 das a,M keibe
Die verschiedentlich in der 4 S-Fraktion beobachtete neutralisierende Aktivitat durfte auf einer geringen Verschmierung der AK der 7 S-Fraktion in die 4 S-Fraktion beruhen.
Der Nachweis der AK bei diesem Tiere deutet an, dai3 die beobachteten Vorgange in der aufgezahlten Reihenfolge einen Heilprozei3 einleiten, als deren erster Schritt der Anstieg der neutrophilen Granulozyten anzusehen ist. Die ersten Anzeichen von beginnenden Eiweiaveranderungen durch eine SP- Infektion treten stets bei den hochmolekularen Serumproteinen auf.
Fur geschickte technische Assistenz danken wir Frl. B. ZOETH und Herrn Ed SCHLOTTERER.
Diese Arbeit wurde rnit EWG-Mitteln gefordert.
Zusammenfassung Von zwei subkutan und zwei per Kontakt mit vollvirulentem Virus der
Schweinepest infizierten Tieren werden klinische, serologische und immunolo- gische Untersuchungen unter Verwendung der Agargel-, Immunoelektropho- rese und Gelfiltration durchgefuhrt. Die erhaltenen Serumfraktionen werden auf Eiweiflveranderungen, neutralisierende und prazipitierende Wirksamkeit sowie auf die ,,krankheitsspezifische“ Bande untersucht.
Nach einer Schweinepestinfektion beginnen die ersten Serumproteinver- anderungen bei den hochmolekularen Komponenten. Ein a,-Globulinanstieg, der dem Leukozytenabfall und Temperaturanstieg des 2./3. Tages folgt, be- ruht im wesentlichen auf einem solchen des a,-Makroglobulins. Dieses nimmt nach dem als erstes Genesungszeichen zu wertenden Wiederanstieg speziell der iieutrophilen Granulozyten und der Bildung der ersten neutralisierenden hoch- inolekularen Antikorper (IgM), aber vor einer 7-Globulinzunahme, wieder ab. Bei den p-Globulinen fallt die betrachtliche Transferrinabnahme und ihr Synergismus mit der Erhohung der Komponenten des Komplementsystem C’3 a sowie einer r-Globulinverminderung auf. Der Schwund der 7-Globu- line wird mit einer Antigen-Antikorperkomplexbildung und deren Abbau im Stoffwechsel bzw. ihrer Ablagerung im Gewebe erklart.
Auf einen Untergang von Blutleukozyten im stromenden Blut wird hin- gewiesen.
Die krankheitsspezifische Bande ist die hochstmolekulare Komponente des Serums. Sie ist bei den subkutan infizierten Tieren ab 4. Tag p. i. nachweisbar.
Summary The pathogenesis of swine fever from the time of infection until maximal pyrexia,
and observations on serum macroglobulin Clinical, xrological and immunological studies were made on two pigs
infected subcutaneously and two infected by contact with virulent swine fever virus. The techniques used were gel electrophoresis, immuno-electro- phoresis and gel filtration. Studies were made of the protein changes, neutralising and precipitating activity and of the “specific disease” band within the serum protein fractions.
Neutralisationswirkung besitzt und diese nur f em j3,M (IgM) zukommt.
Zbl. Vet. Med., Reihe B, Bd. 16, H e f t 4 21

324 W. MATTHAEUS und G. KORN
After infection the first changes in serum proteins occur in the compo- nents of high molecular weight. A rise in alpha globulin, following the fall in leucocytes and the rise in temperature on the second to third days, involves mainly the alpha 2 macroglobulin. The subsequent fall is accom- panied by a rise in neutrophil leucocytes and the formation of the first neutralising high molecular antibodies (IgM) and these are seen as the first signs of recovery and take place before an increase in gamma globulin. Amoqg the beta globulins there occurs a considerable fall in transferrin which is synergistic with the rise in the components of the complement system C‘ 3 a as well as a fall in gamma globulin. This fall in gamma globulin is explained by the formation of an antigen-antibody complex and by the metabolic breakdown of the gamma globulin or its storage in the tissues.
A fall in circulating leucocytes is demonstrated. The disease-specific protein band comprises the high molecular compo-
nents of the serum and it can be shown to be present four days after sub- cutaneous infection.
Resume Sur la pathoghie de la peste porcine, de l’infection jusqu’au maximum
de la temperature, et sur les macroglobulines sCriques On prockde 21 des examens cliniques, skrologiques et immunologiques
sur deux animaux infectks par voie sous-cutanke et deux animaux infect& par contact avec le virus virulent de la peste porcine. On emploie les techni- ques de l’klectrophorkse en gklose, de l’immunoklectrophorkse et de la filtration en gel. On examine les modifications protkiques, l’activitk neu- tralisante et prkcipitante, de meme que la bande apkcifique de la maladie., dans les fractions skriques obtenues.
Aprks une infection de peste porcine, les premikres modifications des protkines skriques atteignent les composantes i poids molkculaire klevl.. Une augmentation des a,-globulines, suivant la chute des leucocytes et l’aug- mentation de tempkrature qui se produit au 2-3e jour, est conditionnke en fait par une augmentation des (1,-macroglobulines. Celles-ci diminuent i nouveau aprks une augmentation des granulocytes neutrophiles, que l’on peut interprkter comme premier signe de gukrison, et aprks la formation des premiers anticorps neutralisants, i poids molkculaire klevk (IgM), mais avant une augmentation des y-globulines. Chez les B-globulines, on observe Line diminution considerable des transfkrines, en synergie avec l’augmenta- tion des composantes du complkment C’3 a, de m2me qu’avec une diminu- tion des r-globulines. La diminution des r-globulines s’explique par la forma- tion d’un complexe antighne-anticorps et par leur dkgradation dans le mbta- bolisme, resp. leur stockage dans les tissus.
On signale une chute des leucocytes du sang circulant. La bande spkcifique de la maladie est la composante skrique au poids
molCculaire le plus Llevk. On peut dkji la mettre en kvidence chez les animaux infectks par voie sous-cutanke 4 jours p. i.
Resumen Sobre la patogenia de la peste porcina claisica desde la infecci6n hasta
el maiximo de temperatura y sobre las macroglobulinas sericas Partiendo de dos animales infectados por via subcuthea y otros dos por
contacto con virus completamente virulent0 de la peste porcina clisica se cfectuaron estudios clinicos, serol6gicos e inmunol6gicos utilizando la elec- troforesis sobre gel de agar y la inmunolbgica, asi como la filtraci6n a traves

325 Zur Pathogenese der Schweinepesterkrankung
de gel. Las fracciones skricas obtenidas se examinan en cuanto a alteraciones proteicas, actividades neutralizante y precipitante, y tambikn acerca de la banda xpatoespecifica>>.
Tras una infecci6n pestosa, las primeras alteraciones seroproteicas comien- zan en 10s componentes de peso molecular elevado. El aumento globulinico (lr , que sigue a1 descenso leucocitico y a1 ascenso de la temperatura del 2'/3er dia, se basa en eaencia en el correspondiente de la macroglobulina a2. Esta vuelve a disminuir despuks del nuevo aumento, que debe valorarge como primer sintoma de convalecencia, en especial de 10s granulocitos neuti6filos y de la formaci6n de 10s primeros anticuerpos neutralizantes de molkcula elevada (IgM), per0 antes de que aumente la globulina y. En las globulinas B, llama la atenci6n la disminuci6n transferrinica considerable y su sinergismo con el aumento de 10s componentes del sistema complementario C'3 a, y la disniinuci6n de la globulina y. La pkrdida de las globulinas y se explica con la formaci6n de un complejo antigeno-anticuerpos y su desintegracibn en el inetabolismo o bien su dep6sito en el tejido.
Se reflexiona sobre la destrucci6n de 10s leucocitos sanguineos en la sangre circulante.
La banda patoespecifica es el componente de peso molecular mis elevado del suer0 sanguineo. Se puede identificar a partir del 4' dia p. i. en 10s cerdos infectados por via subcutinea.
Literaturverzeihnis 1 . ARMAND, Y. J., et S. GUINAND, 1967: Composition et structure de l'a,-Macro-
globuline isolee du serum de porc. Biochim. Biophys. Acta 133, 289. 2. BERG, G., U. ROO- RAND, S. WITTE, H. GOTZ und D. BRESSEL, 1965: Uber die Beziehungen zwischen neutrophilen 1.eukozyten und Serumglobulinfraktionen. Clin. Chim. Acta 12, 282. 3. DUNNE, H. W., A. J. LUEDCKE, C. V. KEICH and J. F. HOKANSON, 1957: The in vitro growth df hog cholera virus in cells of peripheral blood. Am. J. vet. Res. 18, 502. 4. ERNST, H., 1962: Nachweis prazipitierender AK bei der SP mittels Agar-Diffusion. Deutsche Tierarztl. Wschr. 69, 474. 5. FIREMAN, P., W. E. VANNIER and H. C. GOODMAN, 1964: Immunoche- mica1 studies of human serum fractionated by gelfiltration with Sephadex G-200. Proc. Soc. Exp. Biol. Med. 115, 845. 6. FLODIN, .P, 1962: ,,Dextrangels and their application i n gel filtration", Pharmacia, Uppsala, Schweden. 7. FLODIN, P., and J. KILLANDER, 1962: Fractionation of human serum proteins by gel filtration. Biochim. Biophys. Acta 63, 403. 8. GANROT, P. O., 1967: Inhibition of Plasmin activity by a,M .Clin. Chim. Acta 16, 328. 9 . GANROT, P. O., 1966: a,-Anti-trypsin activity and different trypsin substrates. Chi. Chim. Acta 13, 518. 10. GRALHEER, H., und K. H. PEHL, 1956: Uber die Wanderung des im Blutserum und Liquor enthaltenen Schweinepestvirus bei der Papierelektro- phorese. Arch. exp. Vet. Med. 10, 279. 1 1 . NALLMANN, L., 1955: Klinische Chemie und Mikroskopie. Thierne Verlag, Stuttgart. 12. HANNA, M. G., P. NETTESHEIM, W. D. FISCHER, L. C. PETFRS and M. W. FRANCIS, 1967: Serum a-Globulinfraction: survival and recovery effect in irradiated mice. Science 157, 1458. 13. HANTSCHEL, H., 1965: Immunoelektrophoretische Untersuchungen an Seren von gesunden und pestkranken Schwei- nen. Arch. exp. Vet. Med. 19, 1323. 14. HEIDE, K., und H. HAUPT, 1964: Darstellung noch nicht therapeutisch angewandter Plasma roteine. Behringwerke-Mitt. 43, 161. 15. HEIL- MEYER, L., 1959: Allgem. Physiologie Jes Blutes; Handbuch der gesamten Hamatologie. 1 . 1 . 1957, § 1 . Urban-Schwarzenberg, Munchen-Berlin. 16. HEILMEYER, L., und J., 1959: Vorkommen und Funktion des Eisens im Organismus; Handbuch der gesamten Hamatolo- gie. 2, 2, 1 . 8 123. Urban-Schwarzenberg, Munchen-Berlin. 17. JAMES, K., F. B. TAYLOR .ind H. H. FUDENBERG, 1966: Trypsin stabilizers in human serum. The role of a,-macro- globulin. Clin. Chim. Acta 13, 359. 18. JAMES, K., F. B. TAYLOR and H. H. FUDENBERG, 1967: The effect of a,-Macroglobulin in human serum on trypsin, plasmin and thrombin activities. Biochini. Biophys. Acta 133, 374. 19. KORN, G., 1964: Pathogenese der SP als irnmunpathologischer Vorgang im Sinne der Allergielehre. Zbl. Vet. Med. B 11, 379. 20. KORN, G., und W. MATTHAEUS, 1967: Serumproteinveranderungen nach Infektion (Vaccination) mit dem lapinisierten Schweinepestvirusstamm SuiferidBehring. Zbl. Vet. Med. B 14, 539. 21. LETTERER, E., 1959: Allg. Pathologie. Thieme-Verlag, Stuttgart Seite 660. 22. LEWIS, P. A., and R. E. SHOPE, 1929: The blood in hog cholera. J. exp. Med. 50, 719. 23. LOAN, R. W., and D. P. GUSTAFSON, 1961 : Cultivation of Hog Cholera Virus in Subculturable Swine Buffy Coat Cells. Am. J. Vet. Res. 22, 741. 24. MACKANESS,
21"

326 W. MATTHAEUS und G. KORN
G. B., and R. V. BLANDEN, 1967: Cellular Immunity, in Progr. in Allerg . 11, 89; Karger- Verlag, Basel-New York. 25. MATTHAEUS, W., 1965 : Auftrennung t&r Serumproteine verschiedener Haussaugetiere in der Tragerelektrophorese. 1. und 2. Mitt. Verfahrenstechnik der Agar-Elektrophorese und Einflui der Pufferionenstarke auf den Trenneffekt. Zbl. Vet. Med. B, 12, 133, 189. 26. MATTHAEUS, W., 1967: Proteinveranderungen und imrnunoelek- trophoretische Untersuchungen bei der Schweinepesterkrankung. Zbl. Vet. Med. B, 14, 467. 27. MATTHAEUS, W., und G. KORN, 1966: Serumproteinveranderungen und klinische Symptome .bei verschiedenen Verlaufsformen der Schweinepest. Zbl. Vet. Med. B, 13, 558. 28. MATTH~EUS, W., und G. KORN, 1967: Die neutralisierenden Antikorper im Schwein nach experimentqller Infektion mit Schweinepestvirus. Zbl. Bakt. I. Orig. 204, 173. 29. MATTHAEUS, W., 1968 : Noch nicht abgeschlossene Untersuchungen. 30. MATTHAEUS, W., und K. STROHMAIER: noch unveroffentlicht. 31. MATTHIAS, D., und H. KLAUS, 1958: Pathologisch-physiologische Untersuchungen zum Hamoglobin- und Eisenstoffwechsel bei der Schweinepest. Arch. exp. Vet. Med. 12, 427. 32. MATTHIAS, D., und D. SCHMIDT, 1955: Die Viskositat des Blutplasmas bei gesunden und pestkranken Schweinen. Arch. exp. Vet. Med. 9, 534. 33. MATTHIAS, D., 1954: Das histologische Verhalten der Nebennieren bei der Schweinepest. Arch. exp. Vet. Med. 8, 226. 34. MATTHIAS, D., und D. SCHMIDT, 1957: Uber Veranderungen des Blutes bei Schweinepest. Arch. exp. Vet. Med. 11, 520. 35. MEHL, J. W., W. O'CONNELL and J. DE GROOT, 1964: Macroglobulin from human plasma which forms an enzymatically active compound with trypsin. Science 145, 821. 36. MOLNAR, J., 1956: Precipitationreaction with swine fever material. ref. Vet. Bull. 26, 834. 37. NISHIMURA, Y., U. SATO, T. HANAKI and K. NOBUTO, 1964: Studies in tissue culture of hog cholera virus. 11. Neutralisation test by HEIC-method. Jap. J. Vet. Sci. 26, 3, 133. 38. PEHL, K. H., und W. SCHULZE, 1958: Der Virusgehalt blutfreier Organe bei der Schweine est. Arch. exp. Vet. Med. 12, 861. 39. PEHL, K. H., 1959: Knochenmark und Pathogenese [ei einigen Viruskrankheiten der Haustiere. Arch. exp. Vet. Med. 8, 1054. 40. POTEL, K., und G. KORN, 1955: Zur Histologie des Digestionsapparates bei Schweinepest. Arch. exp. Vet. Med. 8, 626. 41. SCHULTZE, H. E., and J. F. HEREMANS, 1966: Molecular Biology of Human Proteins; Plasma Proteins. S. 205. Elsevier Publ. Comp., Arnsterdam- London-New York. 42. SCHULZ, L. CL., 1963: Die Bedeutung der Permeabilitatsstorun- gen bei Stoffwechselerkrankungen der Tiere. 17. Welttierarztekongrei. Bericht V. I. 333. 43. STEINBUCH, M., C. BLATRIX and F. JOSSO, 1967: a,-Macroglobulin as Progressive Antithrombin. Nature 216, 500. 44. zit. STECKER, V. J., and G. J. THORBECKE, 1967: Sites of Synthesis of Serum Proteins. I. Serum Proteins Produced by Macrophages in Vitro. J..Immunology 99, 643. 45. WILTSHAW, E., and W. C. MALONEY, 1955: Histochemical and Biochemical Studies on Leukocyte Alkaline Phospatase Activity. Blood 11, 1120. 46. WIRTH, K., U. ULLMANN, K. BRAND, K. HUTH und B. HESS, 1965: Analyse von Globulinen mit der Gelfiltration. Klin. Wschr. 43, 528. 47. WUHRMANN, F., MARKI zit. U. BERGER und K. HUMMEL, 1964: Einfuhrung in die Mikrobiologie und Imrnunologie. Urban- Schwarzenberg, Munchen-Berlin. * 48. LETTERER, E., 1967: Die Morphologie der immuno- pathischen Reaktionen; Handbuch der allgemeinen Pathologie VII/2, Springer Verlag, Berlin-Heidelberg-New York.
Anschrift der Verfasser : Bundesforschungsanstalt fur Viruskrankheiten der Tiere, 74 Tiibingen, Waldhauser Hohe.





























