Embed Size (px)
Citation preview

Aus der Zahnklinik 2 – Zahnärztliche Prothetik
der Friedrich-Alexander-Universität Erlangen-Nürnberg
Direktor: Professor Dr. med. dent. Manfred Wichmann
Zur relativen Aktivierung der Kaumuskeln
beim Kauen von harter und weicher Nahrung
mit unwillkürlicher Kraftentfaltung.
Inaugural-Dissertation
zur Erlangung der Doktorwürde
der Medizinischen Fakultät
der
Friedrich-Alexander-Universität
Erlangen-Nürnberg
vorgelegt von
Sandra Fillmer
aus Immenstadt

Gedruckt mit der Erlaubnis der
Medizinischen Fakultät der Friedrich-Alexander-Universität
Erlangen Nürnberg
Dekan: Prof. Dr. med. Dr. h. c. Jürgen Schüttler
Referent: Prof. Dr. rer. nat. Peter Pröschel
Korreferent: Prof. Dr. med. dent. Manfred Wichmann
Tag der mündlichen Prüfung: 06.07.2011

Widmung Die vorliegende Arbeit widme ich meiner gesamten Familie, allen voran
meinem Opa der die Fertigstellung der Arbeit leider nicht mehr erleben
konnte.

Inhaltsverzeichnis
1. Zusammenfassung/Summary…………………………………………… 1
2. Einleitung und Problemstellung…………………………………............ 5
2.1. Zur motorischen Steuerung des Kauvorgangs………………... 5
2.2. Neuromuskuläre Schutzmechanismen…………………........... 6
2.3. Zum möglichen Einfluss der Bißkraft auf die
relative Aktivierung……………………………………………….. 9
2.4. Problemstellung…………………………………………………. 10
2.5. Zielsetzung………………………………………………………. 11
3. Material und Methode…………………………………………………… 12
3.1. Probanden und grundsätzliches Vorgehen…………………... 12
3.2. Registrierung der Muskelaktivitäten und der
Unterkieferbewegungen………………………………………… 13
3.3. Ablauf der Messungen…………………………………………. 14
3.4. Auswertung………………………………………………………. 15
4. Ergebnisse……………………………………………………………….. 19
4.1. Allgemein………………………………………………………… 19
4.2. Zeitlicher Verlauf der Muskelaktivitäten………………………. 20
4.3. Zeitlicher Verlauf der Aktivitätsverhältnisse………………….. 24
4.4. Zeitlicher Verlauf der minimalen interokklusalen
Distanzen………………………………………………………… 28
4.5. Korrelationen zwischen A/B-Verhältnissen und MIDs………. 29
4.6. Korrelationen zwischen A/B-Verhältnissen
und Muskelaktivitäten…………………………………………… 31
5. Diskussion…………………………………………………………………32
6. Schlussfolgerungen……………………………………………………... 39
7. Literaturverzeichnis……………………………………………………… 40
8. Abkürzungsverzeichnis…………………………………………………. 45
9. Danksagung……………………………………………………………… 46
10. Lebenslauf………………………………………………………………... 47

1
1 Zusammenfassung
Problemstellung und Zielsetzung
Neuere Untersuchungen zeigen, dass bei einer Abnahme der minimalen
interokklusalen Distanz (MID) in aufeinanderfolgenden Kauzyklen das
Arbeits/Balanceseiten Verhältnis (A/B-Verhältnis) der Masseteraktivitäten
zunimmt. Dieser Effekt, der "MID-abhängigen relativen Aktivierung" wurde
als Schutzstrategie gedeutet, die verhindern soll, dass sich Zähne auf der
Balanceseite beim Kauen destruktiv berühren. Diese logische Deutung ist
noch nicht hinreichend gesichert, da Änderungen des A/B-Verhältnisses
auch als Folge unterschiedlicher Kaukräfte beim Kauen unterschiedlich
harter Nahrung beobachtet wurden. Da eine Veränderung der MID beim
Kauen meist mit einer Veränderung der Kaukraft einhergeht, kann sich
theoretisch hinter der MID-abhängigen relativen Aktivierung tatsächlich eine
Abhängigkeit von der Kaukraft verbergen. Die Beantwortung dieser Frage ist
zum Verständnis neuromuskulärer Strategien, die für physiologische
Funktionabläufe bestimmend sind, wichtig. Ziel dieser Arbeit war es, die
These zu prüfen, dass die A/B-Verhältnisse der Mm. masseteres beim Kauen
primär von der MID abhängen und dass die Kaukraft nur über ihre mögliche
Assoziation zur MID mit den A/B-Verhältnissen korreliert ist.
Material und Methoden
Bei 24 funktionsgesunden männlichen Probanden wurden die Unterkiefer-
bewegungen sowie die elektrischen Aktivitäten der Mm. masseteres und
temporales bei rechts - und linksseitigem Kauen von "weichen" und "harten"
Gummibären gleicher Sorte und Charge aufgezeichnet. Für jede Person und
jeden Kauzyklus wurden aus den Unterkieferbewegungen die MIDs und aus
den r.m.s.-Elektromyogrammen die Aktivitätsamplituden und deren A/B-
Verhältnisse ermittelt. MIDs, Aktivitätsamplituden und A/B-Verhältnisse
wurden pro Kauzyklus über die Anzahl der Personen gemittelt. Die mittleren
A/B-Verhältnisse wurden einerseits mit den MIDs, andererseits mit den
Aktivitätsamplituden korreliert und die Beschaffenheit und Qualität der
Korrelationen miteinander verglichen.

2
Ergebnisse
Beim Kauen harter Nahrung waren die Muskelaktivitäten signifikant höher
und die A/B-Verhältnisse kleiner als mit weicher Nahrung. Gleichzeitig waren
bei harter Nahrung die MIDs signifikant größer. Mit beiden Konsistenzen
waren die A/B-Verhältnisse signifikant und eng mit den MIDs korreliert. Die
MID-Abhängigkeiten der A/B-Verhältnisse bei hartem und weichem Kaugut
gingen stetig ineinander über. Unabhängig vom Kaugut zeigten die A/B-
Verhältnisse eine signifikante und enge Korrelation zur MID, die durch eine
Potenzfunktion vom Typ 1/xa beschrieben werden konnte.
Im Gegensatz dazu ergaben sich zwischen den A/B-Verhältnissen und der
Muskelaktivität nur moderate und kaum signifikante Korrelationen.
Schlussfolgerungen
Die Ergebnisse sprechen für die zu prüfende Ausgangsthese, dass das A/B-
Verhältnis primär von der MID beeinflusst wird. Ein beobachteter grober
Zusammenhang zwischen der Kaukraft und dem A/B-Verhältnis ließ sich auf
die größeren MIDs zurückführen, die bei der härteren Nahrung auftraten.
Unter Brücksichtigung der experimentellen Einschränkungen bekräftigen die
Ergebnisse die Vermutung, dass die neuromuskuläre Steuerung in Form der
MID-abhängigen relativen Aktivierung primär eine Strategie zur Kontrolle von
Okklusalkontakten anwendet. Die in der Literatur postulierte Rekrutierung
zusätzlicher balanceseitiger Masseteraktivität bei erhöhtem Kraftbedarf
könnte eine Sekundärstrategie darstellen, die jedoch bei der Zerkleinerung
eines bestimmten Kauguts der MID-abhängigen relativen Aktivierung
untergeordnet ist. Um diese erstmalig getroffenen Aussagen zu erhärten
wäre es sinnvoll, das vorliegende Experiment durch erweiterte, modifizierte
Kaukraftvorgaben und Bolushärten zu wiederholen.

3
Summary
Problem and aims
Recent research indicated that with decreasing minimal interocclusal
distance of the jaw gape in sequent chewing strokes(MID) there results a
increasing ratio between the working- balancing- activity of the masseter
muscles . This effect of the “MID- depending relative activation” is suspected
as a protection strategy for inhibiting destructive tooth contacts on the
balancing side during chewing. This logic interpretation has not already been
saved enough, as changes in the working- balancing- ratio may also be a
result of different chewing forces during chewing food with different hard
properties. As a change of the MID during chewing is mostly accompanied by
a change of the bite forces, it could theoretically be possible that a
dependence of the bite forces of the MID-depending relative activation is
hiding. The reply of this question is important for the understanding of
neuromuscular strategies that set the tone for physiological function
procederes. This study aimed to prove the thesis that working- balancing-
ratios of the masseter muscles are primarily depending on the MID and
further that the chewing force is only correlated to working- balancing ratios
over its possible association to the MID.
Materials and methods
In 24 healthy male students, the jaw movements and the electric activities of
the masseter muscles and temporales muscles were recorded during
chewing soft and heard Gummi Bears on the right and on the left side. For
each person and each chewing stroke the MIDs were determined out of the
jaw movements - the activity amplitudes and its w-b ratios were detected out
of the r.m.s.- electromyograms. MIDs, activity amplitudes and w-b ratios were
averaged for each chewing stroke over the number of test persons. The
averaged w-b ratios were correlated on the one hand with the MIDs, on the
other hand with the activity amplitudes. The texture and the quality of the
correlations were compared.

4
Results
In chewing hard food the muscle activities were higher and the w-b- ratios
were lower than in chewing soft food. The MIDs were also significant larger
in chewing hard food. Both hard and soft food the w-b- ratios were significant
and strictly correlated to the MIDs. The MID- dependences of the w-b ratios
blended into each other in both cases. Irrespective from the kind of food the
w-b ratios showed a significant and tight correlation to the MID that could be
descriped with a power function type 1/xª.
In contrast there is only a moderate correlation between the w-b-ratios and
the muscle activity.
Conclusions
The results prove the entire thesis that the w-b- ratio is primarily influenced
by the MID. An observed rough context between the chewing force and the
w-b- ratio could be traced back to the bigger MIDs that appeared in the case
of chewing harder food. Considering the experimentally limits the results
support the suspicion that the neuromuscular control, in terms of the MID-
depending relative activation, primarily utilizes a strategy to control occlusal
contacts. Literature postulates an additional recrution of balancing- sided
masseter activity when more bite force is needed- this could be a secondary
strategy that is subordinate to the MID- depending relative activation when
hackling certain food. To corrobarate these results it would be suggestive to
repeat this experiment in an enlarged way, especially in regard to modified
chewing force tasks and different bolus properties.

5
2 Einleitung und Problemstellung
2.1 Zur neuromuskulären Steuerung des Kauvorgangs
Die Kenntnis der Strategien nach denen die motorischen Steuerzentren des
neuromuskulären Systems die Kräfte der Kaumuskeln auf die Zahnreihen
und das Kiefergelenk verteilen, ist für das Verständnis der Funktion des
Kauorgans von grundlegendem Interesse.
Nach gängiger Ansicht wird die Mastikation vom Zentralnervensystem (ZNS)
durch einen Reflexkreis gesteuert (8,19,42), der im Wesentlichen aus einem
Zusammenspiel von zentralen und peripheren Elementen besteht. Man
nimmt an, dass sich im ZNS, genauer im Hirnstamm, ein sog. zentraler
Mustergenerator befindet (19). Dieser besteht aus spezialisierten
Neuronenverbänden, die ein gleichbleibendes Aktivierungsmuster erzeugen
welches zu ebenso stereotypen zyklisch-rhythmischen Öffnungs- und
Schließbewegungen des Unterkiefers führt (8,19,29). Dieses
Bewegungsmuster muss allerdings an die sich ständig verändernden
Bedingungen bei der Mastikation angepasst werden. Diese resultieren aus
Volumen, - Textur, - und Lageänderungen des Speisebolus. Das
neuromuskuläre System muss daher die Aktivierung aller Kaumuskeln
fortgesetzt modifizieren um eine optimale Kraftübertragung auf Zahnreihen
und Kiefergelenke bei den variablen Belastungssituationen zu gewährleisten.
Zur Modifikation der stereotypen mastikatorischen Grundaktivierung nutzt
das ZNS zahlreiche "periphere" sensorische Informationen.
Hierzu gehören zum einen Informationen aus der Mundhöhle über Menge,
Konsistenz, Geschmack und Temperatur des Kauguts, die von Temperatur -,
Tast - und Chemosensoren generiert werden. Zum anderen werden
Informationen über die Kieferposition - und bewegung sowie über die Bißkraft
und die Kiefergelenkbelastung durch spezielle Mechanosensoren erfasst, die
für die Wahrnehmung von Vibration, Druck und Bewegung zuständig sind.
Dabei handelt es sich um Haut - und Schleimhautrezeptoren,
Muskelspindeln, parodontale Rezeptoren und Rezeptoren in der Kapsel des
Kiefergelenks (5,7,8,12,16,23,27,34,42). Alle diese Sensoren leiten ihre
afferenten Signale via Hirnstamm und Thalamus an den mastikatorischen

6
Cortex weiter, wo sie verarbeitet werden. Die Efferenzen ziehen vom Cortex
über den Thalamus und das Corpus striatum wieder zum Hirnstamm, und
gelangen über das Rückenmark zu den Kaumuskeln (8,19).
2.2 Neuromuskuläre Schutzmechanismen
Reflexe
Die Steuerung der mastikatorischen Bewegungen und Krafteinwirkungen
erfolgt nach den Idealvorstellungen der Funktionsdiagnostik so, dass die
Zahnreihen harmonisch und störungsfrei aufeinander gleiten und dass die
kraftaufnehmenden Strukturen nicht überlastet werden. Zu diesen Strukturen
gehören die Zahnreihen, das Kiefergelenk und die Kaumuskeln selbst, die ja
für jegliche Kraftproduktion ursächlich sind.
Okklusale Gleithindernisse in Form von Fehlkontakten wurden in der
Funktionslehre seit Jahrzehnten (und werden immer noch) als potentielle
Ursachen für Überlastungen der oralen Strukturen und damit auch für
Schäden der Zahnreihen oder des Kiefergelenks angesehen. Zur Reaktion
auf - und zur möglichen Adaptation an solche okklusalen Fehlkontakte besitzt
das neuromuskuläre System Schutzmechanismen, als deren zentrales
Element bislang die Reflextätigkeit angesehen wurde (8). Ein speziell für den
Bereich des Kausystems sehr bekannter Reflex ist der Mundöffnungsreflex.
Er sorgt dafür, dass die Kontraktion der Kaumuskeln und damit der
Kauvorgang sofort unterbrochen wird, wenn z.B. auf einen Kirschkern oder
eine Schrotkugel gebissen wird. Neben diesem zwar wichtigen aber selten
gebrauchten Schutzmechanismus gibt es andere Phänomene - sog.
exterozeptive Reflexe - von denen angenommen wird, dass sie eine
Schutzfunktion besitzen (8,18,42). Exterozeptive Reflexe werden
experimentell durch taktile Reize wie z.B. Beklopfen der Wangen,
Mundschleimhaut oder Zähne ausgelöst und führen zu kurzen Inhibitionen
der Kaumuskelaktivierung. Obwohl hierbei bislang keine adäquaten
Reduktionen der Bißkraft nachgewiesen wurden, nimmt man an, dass mit
Hilfe dieser Mechanismen insbesondere kleinere Störungen, die während
des Kauvorgangs auftreten, beherrscht werden können ohne den

7
funktionellen Ablauf zu unterbrechen. Diese Annahme basiert auf
Experimenten, die zumeist bei isometrischem Beißen mit geringen Bißkräften
und großen Bißsperrungen durchgeführt wurden. Da beim eigentlichen
Kauen jedoch sehr hohe Kräfte von ca. 200 bis 300 N (24) bei
Zahnabständen von wenigen Zehntel Millimetern (31,33) ausgeübt werden,
ist es nicht bekannt, in wie weit die exterozeptive Inhibition hierbei überhaupt
eine Rolle spielt. Einige neuere Untersuchungen deuten sogar darauf hin,
dass exterozeptive Reflexe mit enger werdendem Zahnabstand an
Wirksamkeit verlieren (18,40,42).
Die "bißsperrungs - bzw. MID - abhängige relative Aktivierung" als ein bisher
unbekannter Schutzmechanismus
Neuere Studien legen nahe, dass das neuromuskuläre System über einen
bisher nicht bekannten Schutzmechanismus verfügt, der dafür sorgt, dass
unerwünschte direkte Zahnkontakte, die beim Kauen auftreten, nicht zu
unphysiologischen Belastungen der Zähne oder des Kiefergelenks führen.
Dieser Schutzmechanismus ist vor allen Dingen zur Kontrolle der
Auswirkungen von sog. Balancekontakten von Bedeutung. Dies sind
Zahnkontakte, die beim einseitigen Kauen oder Zusammenbeißen auf der
Balanceseite (BS) auftreten können. Als Balanceseite bezeichnet man
diejenige Seite, die der sog. Arbeitsseite (AS) gegenüberliegt, auf der gekaut,
oder auf ein Testobjekt gebissen wird. Durch das komprimierte Kaugut oder
Testobjekt verbleibt auf der AS ein vertikaler Spalt zwischen den Zähnen, der
als "Bißsperrung" bezeichnet wird. Für die Bißsperrung beim Kauen
verwendet man auch synonym den Begriff "minimale interokklusale Distanz"
(MID) (30,31).
Unter der "bißsperrungs - bzw. MID - abhängigen relativen Aktivierung" der
Kaumuskeln, versteht man die Erscheinung, dass beim einseitigen Beißen
oder Kauen die wechselseitigen Verhältnisse der elektrischen Aktivitäten von
je zwei Elevatorenmuskeln mit der Bißsperrung bzw. MID assoziiert sind
(30,31). Dies wurde an den Arbeits/Balanceseiten-Verhältnissen (A/B-
Verhältnis) der beiden Masseter - bzw Temporalismuskeln und an den
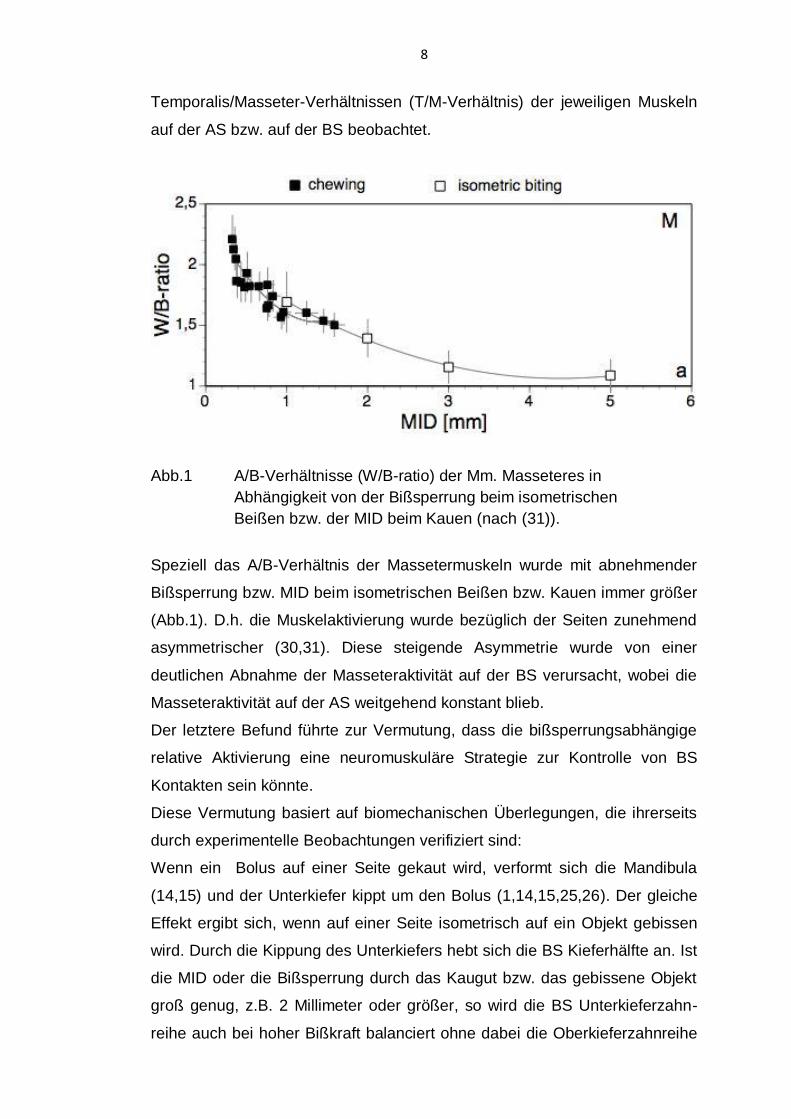
8
Temporalis/Masseter-Verhältnissen (T/M-Verhältnis) der jeweiligen Muskeln
auf der AS bzw. auf der BS beobachtet.
Abb.1 A/B-Verhältnisse (W/B-ratio) der Mm. Masseteres in
Abhängigkeit von der Bißsperrung beim isometrischen
Beißen bzw. der MID beim Kauen (nach (31)).
Speziell das A/B-Verhältnis der Massetermuskeln wurde mit abnehmender
Bißsperrung bzw. MID beim isometrischen Beißen bzw. Kauen immer größer
(Abb.1). D.h. die Muskelaktivierung wurde bezüglich der Seiten zunehmend
asymmetrischer (30,31). Diese steigende Asymmetrie wurde von einer
deutlichen Abnahme der Masseteraktivität auf der BS verursacht, wobei die
Masseteraktivität auf der AS weitgehend konstant blieb.
Der letztere Befund führte zur Vermutung, dass die bißsperrungsabhängige
relative Aktivierung eine neuromuskuläre Strategie zur Kontrolle von BS
Kontakten sein könnte.
Diese Vermutung basiert auf biomechanischen Überlegungen, die ihrerseits
durch experimentelle Beobachtungen verifiziert sind:
Wenn ein Bolus auf einer Seite gekaut wird, verformt sich die Mandibula
(14,15) und der Unterkiefer kippt um den Bolus (1,14,15,25,26). Der gleiche
Effekt ergibt sich, wenn auf einer Seite isometrisch auf ein Objekt gebissen
wird. Durch die Kippung des Unterkiefers hebt sich die BS Kieferhälfte an. Ist
die MID oder die Bißsperrung durch das Kaugut bzw. das gebissene Objekt
groß genug, z.B. 2 Millimeter oder größer, so wird die BS Unterkieferzahn-
reihe auch bei hoher Bißkraft balanciert ohne dabei die Oberkieferzahnreihe

9
zu berühren. Ist dagegen die MID oder die Bißsperrung kleiner als 2
Millimeter, so kann abhängig von der Bißkraft und den individuellen
anatomischen Verhältnissen die Hebung der BS Zahnreihe in die
Größenordnung der MID kommen, was dann zu Zahnkontakten (22,39) oder
einer Überlastung des Kiefergelenks auf der BS führen könnte (8,26).
Es ist biomechanisch logisch und durch eine aktuelle Untersuchung (39)
experimentell untermauert, dass in der beschriebenen Situation eine
Reduktion der BS Masseteraktivität die Kippung des Unterkiefers um den
Bolus vermindert, und so die Anhebung des Unterkiefers auf der
Balanceseite verringert. Somit kann ein Zusammenprall der Zähne von Ober-
und Unterkiefer auf der BS verhindert oder zumindest kontrolliert werden.
Dieser Mechanismus kann erklären, warum durch das Kauen normalerweise
keine Okklusionsschäden der Zähne oder Dysfunktionen des Kausystems
verursacht werden.
2.3 Zum möglichen Einfluß der Bißkraft auf die relative Aktivierung
Die beschriebene Sicherheitsstrategie beruht nach bisherigen Erkenntnissen
auf einer neuromuskulären Steuerung der relativen Aktivierung, die sich am
vertikalen Zahnabstand orientiert. Dies ist zwar durch die Assoziation
zwischen Bißsperrung bzw. MID und relativer Muskelaktivierung offenkundig,
es ist aber dennoch nicht klar, ob tatsächlich eine afferente Information über
den vertikalen Zahnabstand in den motorischen Steuerzentren verarbeitet
wird und ob diese Information von lagesensitiven Mechanorezeptoren
stammt.
Gewisse Zweifel an einem kausalen Zusammenhang zwischen Bißsperrung
und Muskelsteuerung ergeben sich aus der Tatsache, dass insbesondere bei
Kauvorgängen mit zunehmender Kompression der Nahrung nicht nur die
MID abnimmt, sondern gleichzeitig auch immer die Bißkraft zunimmt.
Dementsprechend wurden auch in mehreren Studien grobe
Zusammenhänge zwischen der Kaukraft und der relativen Muskelaktivierung
dokumentiert und entsprechend interpretiert:
W.L. Hylander et al. (9) analysierten die Funktion des M. masseter während
des Kauens bei Macaca fascicularis. Dabei verglichen die Autoren das A/B-

10
Verhältnis der Mm. masseteres beim Kauen harter Nahrung im Gegensatz zu
weicher Nahrung. Es zeigte sich, dass dieses Verhältnis beim Kauen harter
Nahrung symmetrischer war. Da gleichzeitig bei der härteren Nahrung
höhere Kaukräfte auftraten, wurde gefolgert, dass die Kaukraft die relative
Muskelaktivität beeinflusst (9,44). In einem ähnlichen Experiment, das aber
an Menschen durchgeführt wurde, verglichen Blanksma und van Eijden (3)
die EMG Amplituden beim Kauen von Gummibären mit denen beim Kauen
von Lakritze, welche einen höheren mechanischen Widerstand besitzt. Hier
zeigte sich beim Kauen der zäheren Nahrung eine höhere Muskelaktivität
was auch höhere Kaukräfte vermuten läßt. Parallel dazu war das A/B-
Verhältnis beim Kauen der zäheren Nahrung kleiner, d.h. symmetrischer als
beim Kauen der weicheren Nahrung. Daraus wurde geschlossen, dass der
M. masseter auf der BS verstärkt herangezogen wird, wenn mehr Bißkraft
erzeugt werden soll (3). Zu ähnlichen Ergebnissen kam schließlich auch eine
Studie von Piancino et al (28) in der ebenfalls Masseter A/B-Verhältnisse
beim Kauen zäher und weicher Nahrung verglichen wurden. Aus dem
symmetrischeren A/B-Verhältnis mit zäher Nahrung wurde auf eine mögliche
Schutzfunktion hinsichtlich des Kiefergelenks geschlossen.
Die genannten Studien interpretieren somit alle die Änderung der relativen
Aktivierung als Folge eines erhöhen Kraftbedarfs. In jeder dieser Arbeiten
kam zwar explizit oder implizit zum Ausdruck, dass die verschieden harten
Nahrungen auch mit unterschiedlichen Interokklusalabständen assoziiert
waren. Es wurden jedoch diesbezüglich keine weiteren Untersuchungen
angestellt.
2.4 Problemstellung
Die Frage, ob die zur Steuerung der relativen Kaumuskelaktivierung nötige
afferente periphere Information aus einer sensorischen Wahrnehmung der
Bißkraft oder aber der Bißsperrung resultiert, ist nicht nur von rein
wissenschaftlichem Interesse. Eine Beantwortung dieser Frage könnte auch
im Hinblick auf zahnärztliche Behandlungen von Bedeutung sein, bei denen
natürliche sensorische Informationsquellen z.B. bei der Extraktion von
Zähnen verloren gehen. Zur experimentellen Beantwortung der obigen

11
Frage müsste man die Bißkraft während des Kauens - also die sog.
"Kaukraft" - messen und diese zu den Aktivitätsverhältnissen in Beziehung
setzen. Hierfür müsste die Kaukraft intraoral zwischen den Zahnreihen
erfasst werden, was im Prinzip mit Aufnehmern geschehen könnte, die in
Zähne oder Implantate integriert sind. Wegen des großen methodischen und
finanziellen Aufwandes bei der Anwendung integrierter Sensoren (8,32,38)
empfiehlt es sich, zu einer pilotmäßigen Untersuchung der obigen
Fragestellung zunächst indirekte, aber einfachere Methoden anzuwenden:
Als Ersatz für die nur schwer zu messende Kaukraft kann das Oberflächen -
Elektromyogramm (EMG) verwendet werden, welches als wichtigstes
Hilfsmittel für das Studium von Krafteinwirkungen und neuromuskulären
Zusammenhängen allgemeine Verbreitung gefunden hat.
Es ist mehrfach belegt, dass die Elektromyogramme von Kaumuskeln mit der
Bisskraft beim isometrischen Beißen eng korreliert sind (2,6,17,20,43). Auch
für das Kauen wurden Assoziationen zwischen der Kaukraft und den EMGs
der einzelnen Kaumuskeln demonstriert (24,38).
2.5 Zielsetzung
Falls die neuromuskuläre Steuerung der relativen Muskelaktivierung kausal
von der Bißkraft abhängt und die mehrfach gezeigte Assoziation der relativen
Aktivierung mit der MID nur eine Folgekorrelation davon ist, dass die MID bei
steigender Bißkraft abnimmt, so sollte die relative Aktivierung eine
mindestens ebenso gute, wenn nicht bessere, Korrelation zur Bißkraft
aufweisen wie zur MID. In den zitierten Untersuchungen, die einen
Zusammenhang zwischen der relativen Aktivierung und der Bißkraft
demonstrierten (3,9,28,31,44) waren keine expliziten Korrelationen zwischen
diesen beiden Größen untersucht worden.
Ziel dieser Studie war es daher, zu überprüfen, ob die relative Aktivierung der
Kaumuskeln, ausgedrückt durch Muskelaktivitätsverhältnisse, beim Kauen
mit der Muskelaktivität ebensogut korreliert ist, wie mit der MID.

12
3 Material und Methode
3.1 Probanden und grundsätzliches Vorgehen
Probanden
An der Untersuchung nahmen 24 männliche Studenten der Medizin oder
Zahnmedizin, mit einem mittleren Alter von 24 ± 3 Jahren freiwillig teil. Die
Probanden wiesen einen Biss der Angle Klasse I oder II auf, hatten keine
Zahnlücken und zeigten keine Symptome von kraniomandibulären
Dysfunktionen wie etwa Muskelbeschwerden, Kiefergelenkknacken oder
Kiefergelenksbeschwerden. Von den Versuchen ausgeschlossen waren
Personen mit Skelettanomalien wie z.B. Progenie, Prognathie oder Lang -
und Kurzgesichtssymptomatik. Alle Probanden wurden über den
Versuchsablauf aufgeklärt und gaben ihr mündliches Einverständnis. Das
experimentelle Protokoll war von der Ethikkommission der Medizinischen
Fakultät der Universität Erlangen-Nürnberg genehmigt worden.
Grundsätzliches Vorgehen
Jede Messsitzung bestand aus vier einzelnen Messungen. Bei den ersten
zwei Messungen kauten die Versuchspersonen jeweils einen industriell
standardisierten Gummibären (Goldbären, Haribo, Bonn, Deutschland) auf
der rechten - und auf der linken Kieferseite. Für die dritte und vierte Messung
wurden Gummibären der gleichen Charge, die zuvor gehärtet worden waren
in gleicher Weise auf der rechten, und dann auf der linken Seite gekaut.
Während der Kausequenzen, wurden die elektrischen Aktivitäten der Musculi
masseteres und temporales anteriores auf beiden Seiten, sowie die
Unterkieferbewegungen 30 Sekunden lang registriert. Zur Auswertung der
Messungen wurden aus den Elektromyogrammem die A/B-Verhältnisse und
die T/M-Verhältnisse gebildet. Aus den Aufzeichnungen der
Unterkieferbewegungen wurden die minimalen interokklusalen Distanzen
ermittelt. Die Aktivitätsverhältnisse wurden dann sowohl mit den MIDs als
auch mit den Muskelaktivitäten korreliert.

13
3.2 Registrierung der Muskelaktivitäten und der Unterkieferbewegungen
Die Summenaktionspotentiale der rechten und linken Mm. masseteres sowie
der rechten und linken Mm. temporales anteriores wurden mit Hilfe von
bipolaren Ag/AgCl Oberflächenelektroden (Noraxon, type 272, Scottsdale,
USA) abgeleitet. Der Durchmesser der sensitiven Elektrodenflächen betrug
10 mm, der Elektrodenabstand 20 mm. Zur richtigen Positionierung der
Elektroden, wurden die Probanden aufgefordert auf beiden Seiten ein
Gummibärchen zu kauen. Dabei wurden die Muskelbäuche beidseits
palpiert. Soweit dies möglich war, wurde die Längsachse der
Oberflächenelektroden nach dem Verlauf der Muskelfasern ausgerichtet. Um
den Hautwiderstand zu reduzieren, wurde die Haut an den Klebestellen
zuvor mit Cutasept®F (Bode Chemie, Hamburg) gereinigt und entfettet. Die
Elektroden waren bereits mit Kontaktgel versehen und selbstklebend. Eine
Referenzelektrode für die Erdung des Signals wurde bei jedem Probanden in
der Mitte der Stirn befestigt.
Die Rohelektromyogramme wurden verstärkt und im Bereich von 10 Hz bis 2
KHz gefiltert. Die EMG- Verstärker (Biovision, Wehrheim, Deutschland)
waren unmittelbar an den Elektroden angebracht. Sie hatten eine
Eingangsimpedanz von 10 GOhm , ein Aussteuerungsverhältnis von mehr
als 120 dB und ein Eigenrauschen von 1µV.
Zur Aufzeichnung der Unterkieferbewegungen aus denen später die MIDs
ermittelt werden sollten, wurde ein SIROGNATHOGRAPH verwendet. Dieser
registriert mit Hilfe eines magnetfeldsensitiven Kopfgestells die Bewegungen
eines kleinen Permanentmagneten, der an den unteren Schneidezähnen
befestigt wird. Hierzu wurde ein Autopolymerisat (Luxatemp, ESPE, Seefeld)
und ein provisorischer Befestigungszement (Temp Bond, KerrHawe, Bioggio)
verwendet. Um mögliche Artefakte durch das Erdmagnetfeld auszuschließen,
saßen die Probanden während des Versuches aufrecht in einem Stuhl, der
in Nord-Süd Richtung ausgerichtet war. Um Bewegungsartefakte zu
vermeiden, wurde der Rahmen des Sensors fest am Kopf des Probanden
fixiert, der zusätzlich durch eine Kopfstütze unterstützt wurde.
Die Bewegungssignale des Unterkiefers und die EMG-Signale wurden über
eine Verteilerbox einem 12-Bit A/D-Wandler Einschub (DAQ 6024, National

14
Instruments, Austin, Texas, USA) zugeleitet. Vom A/D-Wandler wurden die
Analogdaten mit einer Abtastfrequenz von 2KHz pro Kanal digitalisiert und
auf einem Laptop (Inspirion 8600, Dell, Austin, Texas, USA) abgespeichert.
Der zeitliche Ablauf der Messungen und die Datenerfassung wurden mit Hilfe
eines selbsterstellten Schaltbildes der Software DASYLab ® (measX,
Deutschland) kontrolliert und gesteuert.
3.3 Ablauf der Messungen
Vor dem Start der eigentlichen Messungen wurden die Verstärkungen der
EMG-Verstärker so eingestellt, dass keine Übersteuerungen auftraten. Dies
geschah während des Kauens von Gummibären, da hierbei in der Regel die
höchsten Muskelaktivitäten erzeugt werden. Traten während der Messungen
dennoch Übersteuerungen auf, so wurde die betreffende Registrierung mit
einer abgeschwächten Verstärkereinstellung wiederholt.
Jeder Proband führte in einer Sitzung die folgenden vier motorischen
Aktionen aus, die jeweils 30 Sekunden lang aufgezeichnet wurden:
1. Kauen eines "weichen" Gummibären auf der rechten Seite
2. Kauen eines "weichen" Gummibären auf der linken Seite
3. Kauen eines "harten" Gummibären auf der rechten Seite
4. Kauen eines "harten" Gummibären auf der linken Seite
Als weiche Gummibären sind hier und im Folgenden diejenigen gemeint, die
frisch der Packung entnommen wurden. Die "harten" Gummibären waren ca.
3 Stunden vor der Messung derselben Packung entnommen, und in einem
Gefrierfach gelagert worden. Die Probanden wurden instruiert, sowohl die
weichen, als auch die gehärteten Gummibären so zu kauen, wie es ihrer
Gewohnheit oder ihrem Empfinden entsprach. Bezüglich der Kaukraft
wurden keinerlei Vorgaben gemacht.
Vor Beginn jeder Messung wurde der Gummibär auf die Zunge manövriert
und die Zähne in die Interkuspitationsposition geführt. Der Unterkiefer wurde
in dieser Position locker gehalten, so dass die Zähne Kontakt hatten, aber

15
die Muskeln keine Aktivität erzeugten. In dieser Haltung wurde der
SIROGNATHOGRAPH auf Null abgeglichen und die Messung gestartet.
Im Anschluß an die hier genannten vier Kauaktionen wurden von den
Probanden noch weitere motorische Aktionen durchgeführt und registriert.
Hierbei handelte es sich um einseitiges isometrisches Beißen auf
Folienkraftaufnehmer, die Bißsperrungen von ca. 2 und 5 mm hervorriefen.
Die Auswertung dieser Experimente steht nicht in direktem Zusammenhang
zur vorliegenden Arbeit und bleibt einer anderen Fragestellung vorbehalten.
3.4 Auswertung
Verarbeitung der Rohelektromyogramme
Die auf der Festplatte des Meß-Laptops im DDF-Format abgespeicherten
Rohmyogramme wurden auf einen USB-Stick gezogen, auf ein Apple
PowerBook G4 übertragen und dort in die Rechen - und Analysesoftware
MATLAB® (The MathWorks, Natick, USA) importiert. Um die EMG-Daten
auswerten zu können, wurden sie in eine Form gebracht, die der r.m.s. (route
- mean - square) Darstellung des Elektromyogramms entsprach. Hierzu
wurden die fortlaufenden Zeitreihenwerte in allen EMG-Kanälen durch
Betragsbildung rechnerisch gleichgerichtet, um positive Amplituden zu
erhalten. Danach wurden die gleichgerichteten Aktivitätsverläufe mit der
Methode des gleitenden Durchschnitts geglättet. Hierzu wurde ein Fenster
mit einer Zeitbreite von 100 Punkten entsprechend 50 msec über die
gleichgerichteten EMG-Kurven gelegt. Die Messpunkte innerhalb des
Fensters wurden zu einem Punkt gemittelt, der dann jeweils einen
Datenpunkt der geglätteten EMG-Kurve darstellte. Sodann wurde das
Fenster um einen Messpunkt entlang der Zeitachse verschoben und die
Prozedur wiederholt.

16
Ermittlung der Aktivitätsamplituden, Aktivitätsverhältnisse und MIDs
Die weitere Auswertung fand ebenfalls in MATLAB® statt. Abb. 2 zeigt
beispielhaft die Zeitverläufe der geglätteten EMG - Signale der vier Muskeln,
sowie den Zeitverlauf des vertikalen Ausschlags der Unterkieferbewegung. In
den beiden oberen Feldern sieht man die Aktivität der rechten und linken
Massetermuskeln in mV während der 30 s dauernden Aufnahmeperiode.
Analog sind in den Feldern drei und vier die Aktivitäten der
Temporalismuskeln dargestellt. Die vertikale Bewegung des Unterkiefers ist
in der untersten Grafik ersichtlich.
Für jeden Probanden und jede Kauaktion wurden von MATLAB® aus den
geglätteten EMG - Zeitkurven (Abb.2) alle während der 30 Sekunden
Messzeit erzielten Aktivitätsmaxima bestimmt. Die zu jedem Kauakt
gehörende minimale interokklusale Distanz wurde als Abstand zwischen der
Null-Linie und dem höchsten Punkt des vertikalen Wegsignals bestimmt.
Aus den vier Akitivitätsmaxima eines jede Kauzyklus wurden die A/B-
Verhältnisse der Masseter- und Temporalismuskeln sowie die T/M-
Verhältnisse der Muskeln der AS und der BS ermittelt. Bei diesen
Verhältnissen handelt es sich um die Quotienten aus Arbeits- und
Balanceseitenaktivität der Mm. masseteres bzw. der anterioren Mm.
temporales (A/B-Verhältnisse) sowie die Quotienten aus Temporalis und
Masseteraktivität für Arbeits- und Balanceseite (T/M-Verhältnis).
Um Aktivitätsmaxima und Aktivtätsverhältnisse beim Kauen der harten und
weichen Gummibären unabhängig von interindividuellen Variationen
vergleichen zu können, wurden die Werte normiert. Hierzu wurde für jede
Person als Referenzwert die Aktivitsamplitude des arbeitsseitigen M.
masseter aus dem ersten Kauzyklus beim Kauen der weichen
Gummibärchen definiert und als 100% festgesetzt. Die absoluten Werte der
vier Aktivitätsmaxima eines jeden Kauzyklus beim Kauen sowohl der
weichen als auch der harten Gummibärchen wurden durch diesen
Referenzwert geteilt. Beim Kauen auf der rechten Seite wurde durch den
entsprechenden Bezugswert des rechten M. masseter, beim Kauen links
durch denjenigen des linken M. masseter geteilt.

17
Abb. 2: Beispiel für die in MATLAB® erstellten, geglätteten EMG -
Zeitverläufe (obere 4 Spuren), sowie den Zeitverlauf der
vertikalen Unterkieferbewegung (untere Spur) beim
rechtsseitigen Kauen eines weichen Gummibären.
Analog zu den Aktivitätsamplituden wurden auch die Aktivitätsverhältnisse
auf den Wert des jeweils ersten Kauzyklus beim Kauen der weichen
Gummibären normiert.
Somit ergab sich für jeden Probanden und jeden Kauzyklus ein Satz von acht
Aktivitätsmittelwerten (Masseter AS und BS sowie Temporalis AS und BS
jeweils beim Kauen der harten und weichen Gummibären), ein Satz von acht
Verhältnismittelwerten, (A/B-Verhältnis der Mm. masseteres und der Mm.
temporales, T/M-Verhältnis für die AS und die BS jeweils beim Kauen der
harten und weichen Gummibären) sowie ein Satz von zwei Mittelwerten der
MID für Kauen von harten und weichen Gummibären.

18
Für jeden der aufeinander folgenden Kauzyklen wurden schließlich die MID’s
und die normierten Aktivitäten und Aktivitätsverhältnisse über die Anzahl der
Probanden gemittelt, die den entsprechenden Zyklus während des Kauens
erreicht hatten.
Darstellung und statistische Auswertung
Die über alle Probanden gemittelten Werte der MIDs, Aktivitäten und
Aktivitätsverhältnisse werden als Funktion der fortlaufenden Kauzyklen in
Diagrammen dargestellt.
Des Weiteren werden die Aktivitätsverhältnisse sowohl zu den MIDs als auch
zu den Aktivitätsamplituden zugehörigen Kauzyklen in Beziehung gesetzt.
Um zu testen, wie jeder Parameter vom zeitlichen Verlauf des Kauvorgangs
abhängt, werden die Mittelwerte der Aktivitäten, Aktivitätsverhältnisse und
MIDs linearen - bzw. polynomischen Regressionen zweiten Grades
unterzogen. Eine Regression wird bei einer Irrtumswahrscheinlichkeit von p>
0,01 als signifikant betrachtet.

19
4 Ergebnisse
4.1 Allgemein
Die 24 Probanden führten im Durchschnitt 30 Kauzyklen mit den gehärteten
Gummibären und 29 mit den weichen Gummibären aus, wobei eine
Versuchsperson mehr als 60 Kauzyklen ausführte. Blieb diese
unberücksichtigt, reduzierte sich der Durchschnitt auf 27 Zyklen.
Die Ergebnisse werden auf die ersten 18 Kauzyklen beschränkt, da diese
Anzahl von allen 24 Probanden innerhalb der 30 s dauernden Messung
erreicht wurde.
Im ersten Teil werden sowohl die Aktivitätsverhältnisse und die Aktivitäten
der vier Elevatorenmuskeln als auch die Änderung der minimalen
interokklusalen Distanz im zeitlichen Verlauf dargestellt. Dabei werden in
einem Diagramm die Aktivitäten der Muskeln bzw. deren
Aktivitätsverhältnisse sowohl beim Kauen der weichen als auch der harten
Gummibären im Vergleich dargestellt. Die Beschreibung der Ergebnisse
bezieht sich immer auf die Entwicklung der prozentualen Werte im zeitlichen
Verlauf.
Im zweiten Teil der Ergebnisauswertung werden die Werte der
Aktivitätsverhältnisse auf der Arbeitsseite für die weichen und harten
Gummibären – diesmal getrennt für Kauen auf der linken und rechten Seite –
mit der MID korreliert.
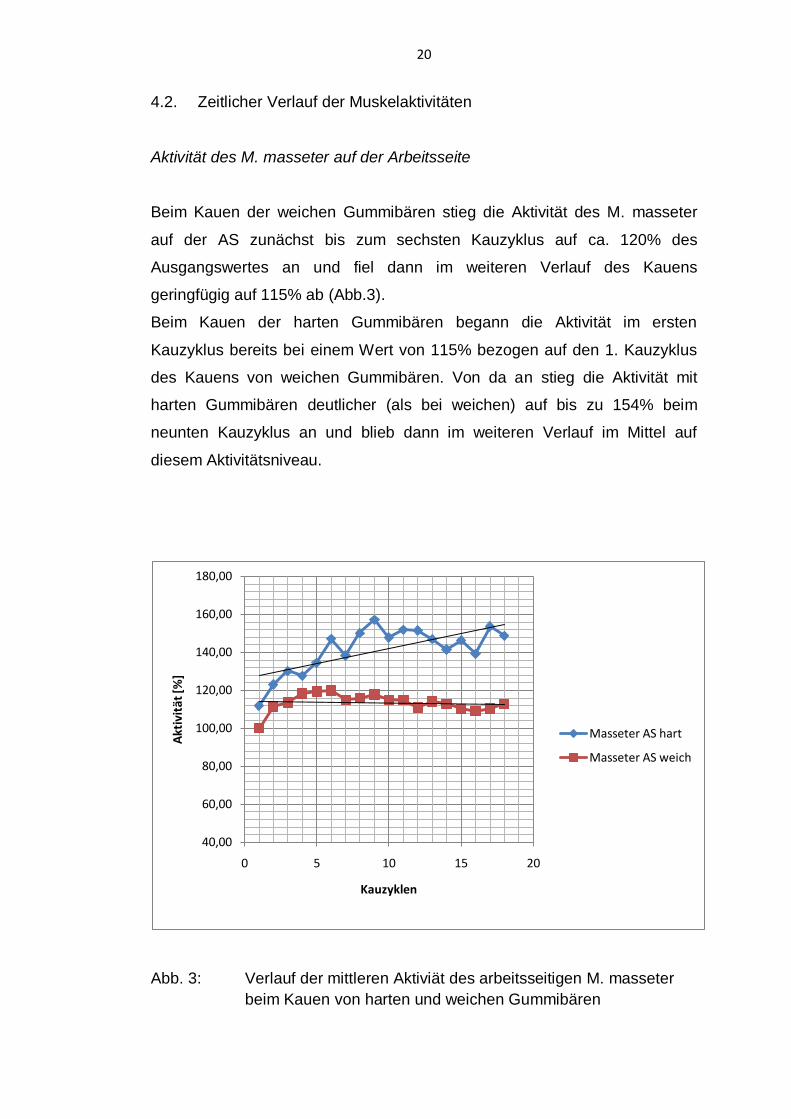
20
4.2. Zeitlicher Verlauf der Muskelaktivitäten
Aktivität des M. masseter auf der Arbeitsseite
Beim Kauen der weichen Gummibären stieg die Aktivität des M. masseter
auf der AS zunächst bis zum sechsten Kauzyklus auf ca. 120% des
Ausgangswertes an und fiel dann im weiteren Verlauf des Kauens
geringfügig auf 115% ab (Abb.3).
Beim Kauen der harten Gummibären begann die Aktivität im ersten
Kauzyklus bereits bei einem Wert von 115% bezogen auf den 1. Kauzyklus
des Kauens von weichen Gummibären. Von da an stieg die Aktivität mit
harten Gummibären deutlicher (als bei weichen) auf bis zu 154% beim
neunten Kauzyklus an und blieb dann im weiteren Verlauf im Mittel auf
diesem Aktivitätsniveau.
Abb. 3: Verlauf der mittleren Aktiviät des arbeitsseitigen M. masseter
beim Kauen von harten und weichen Gummibären
40,00
60,00
80,00
100,00
120,00
140,00
160,00
180,00
0 5 10 15 20
Akt
ivit
ät [%
]
Kauzyklen
Masseter AS hart
Masseter AS weich

21
Aktivität des M. masseter auf der Balanceseite
Die Aktivität des balanceseitigen M. masseter begann beim Kauen der
weichen Gummibären bei einem um ca. 15% niedrigeren Wert als die
Aktivität auf der Arbeitsseite (Abb.4). Im weiteren Verlauf nahm die
balanceseitige Aktivität dann linear ab bis auf ca. 65% des Referenzwertes
beim 18. Kauzyklus.
Die BS-Aktivität beim Kauen der harten Gummibären begann bei nahezu
100% des Referenzwertes und stieg bis zum neunten Zyklus auf 127% an.
Von da an fiel sie wieder leicht in Richtung des Anfangswertes, der bei 104
% lag.
Abb. 4: Verlauf der mittleren Aktiviät des balanceseitigen M. masseter
beim Kauen von harten und weichen Gummibären
40,00
60,00
80,00
100,00
120,00
140,00
160,00
180,00
0 5 10 15 20
Akt
ivit
ät [%
]
Kauzyklen
Masseter BS weich
Masseter BS hart

22
Aktivität des M. temporalis auf der Arbeitsseite
Die Aktivität des M. temporalis auf der Arbeitsseite stieg beim Kauen der
weichen Gummibären in den ersten beiden Zyklen auf 112% des
Anfangswertes, blieb für die nächsten 12 Kauzyklen ziemlich konstant in
diesem Bereich, um im fünfzehnten Zyklus auf 105% abzufallen (Abb. 5).
Im Vergleich zu dem weichen Bolus stieg die Aktivität des M. temporalis im
Verlauf des Kauens der harten Gummibären signifikant höher auf ca. 140%
des Anfangswertes beim neunten Kauzyklus an und pendelte bis zum 18.
Zyklus um dieses Niveau.
Abb. 5: Verlauf der mittleren Aktivität des arbeitsseitigen M. temporalis
beim Kauen von harten und weichen Gummibären.
40,00
60,00
80,00
100,00
120,00
140,00
160,00
180,00
0 5 10 15 20
Akt
ivit
ät [%
]
Kauzyklen
Temporalis AS hart
Temporalis AS weich
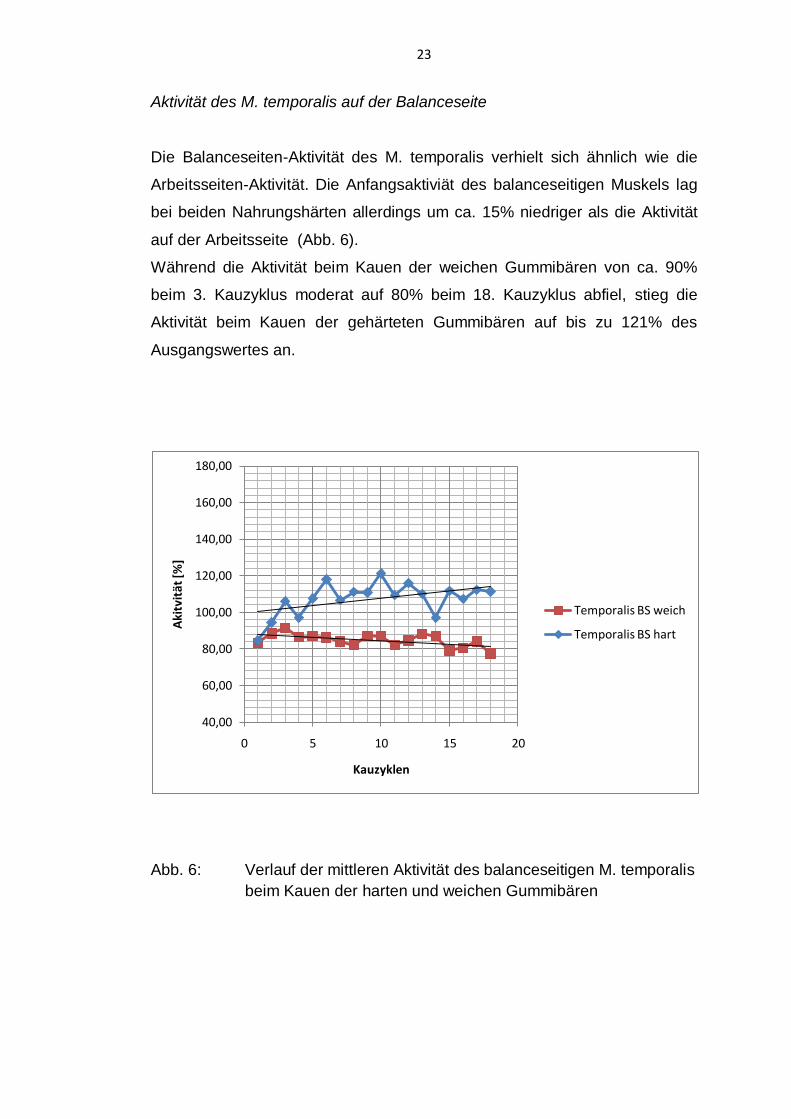
23
Aktivität des M. temporalis auf der Balanceseite
Die Balanceseiten-Aktivität des M. temporalis verhielt sich ähnlich wie die
Arbeitsseiten-Aktivität. Die Anfangsaktiviät des balanceseitigen Muskels lag
bei beiden Nahrungshärten allerdings um ca. 15% niedriger als die Aktivität
auf der Arbeitsseite (Abb. 6).
Während die Aktivität beim Kauen der weichen Gummibären von ca. 90%
beim 3. Kauzyklus moderat auf 80% beim 18. Kauzyklus abfiel, stieg die
Aktivität beim Kauen der gehärteten Gummibären auf bis zu 121% des
Ausgangswertes an.
Abb. 6: Verlauf der mittleren Aktivität des balanceseitigen M. temporalis
beim Kauen der harten und weichen Gummibären
40,00
60,00
80,00
100,00
120,00
140,00
160,00
180,00
0 5 10 15 20
Aki
tvit
ät [%
]
Kauzyklen
Temporalis BS weich
Temporalis BS hart

24
4.3 Zeitlicher Verlauf der Aktivitätsverhältnisse
A/B-Verhältnis der Mm. masseteres
Im Verlauf des Kauens stieg das A/B-Verhältnis der Mm. masseteres bei
beiden Texturen signifikant und linear an (Abb. 7).
Beim Kauen der weichen Gummibären trat ein signifikant steilerer Anstieg bis
auf ca. 180% des Anfangswertes ein.
Dagegen stieg das A/B-Verhältnis beim Kauen der harten Gummibären nur
auf ca. 140% des Anfangswertes an.
Abb. 7: Verlauf des mittleren A/B-Verhältnisses der Aktivitäten der Mm.
masseteres beim Kauen von harten und weichen Gummibären
40,00
60,00
80,00
100,00
120,00
140,00
160,00
180,00
200,00
0 5 10 15 20
A/B
-Ver
häl
tnis
[%]
Kauzyklen
A/B Verhältnis Masseter hart
A/B Verhältnis Masseter weich
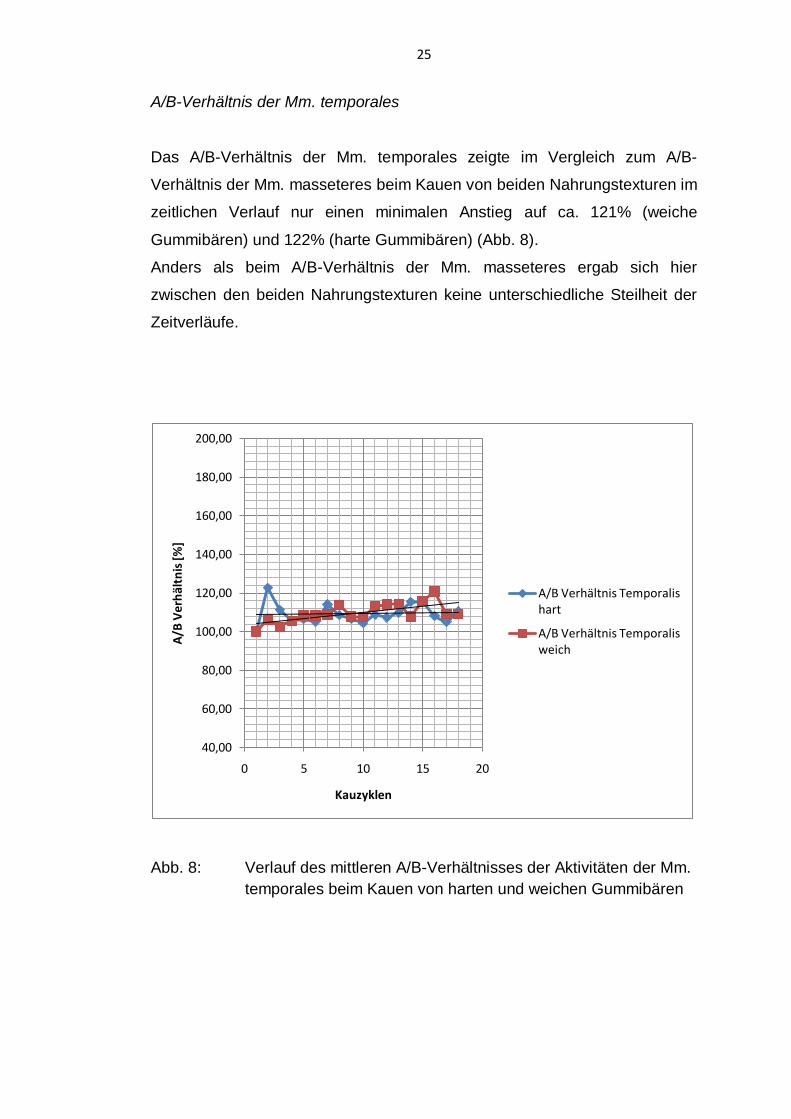
25
A/B-Verhältnis der Mm. temporales
Das A/B-Verhältnis der Mm. temporales zeigte im Vergleich zum A/B-
Verhältnis der Mm. masseteres beim Kauen von beiden Nahrungstexturen im
zeitlichen Verlauf nur einen minimalen Anstieg auf ca. 121% (weiche
Gummibären) und 122% (harte Gummibären) (Abb. 8).
Anders als beim A/B-Verhältnis der Mm. masseteres ergab sich hier
zwischen den beiden Nahrungstexturen keine unterschiedliche Steilheit der
Zeitverläufe.
Abb. 8: Verlauf des mittleren A/B-Verhältnisses der Aktivitäten der Mm.
temporales beim Kauen von harten und weichen Gummibären
40,00
60,00
80,00
100,00
120,00
140,00
160,00
180,00
200,00
0 5 10 15 20
A/B
Ver
häl
tnis
[%]
Kauzyklen
A/B Verhältnis Temporalis hart
A/B Verhältnis Temporalis weich
26
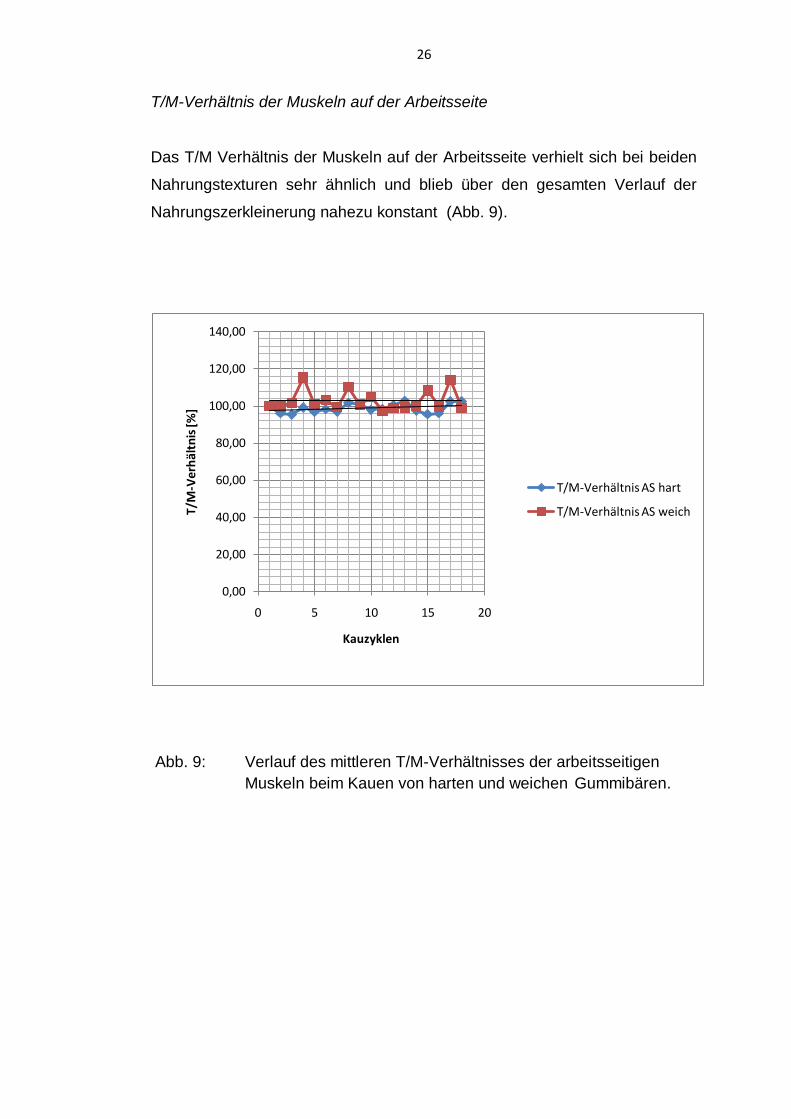
T/M-Verhältnis der Muskeln auf der Arbeitsseite
Das T/M Verhältnis der Muskeln auf der Arbeitsseite verhielt sich bei beiden
Nahrungstexturen sehr ähnlich und blieb über den gesamten Verlauf der
Nahrungszerkleinerung nahezu konstant (Abb. 9).
Abb. 9: Verlauf des mittleren T/M-Verhältnisses der arbeitsseitigen
Muskeln beim Kauen von harten und weichen Gummibären.
0,00
20,00
40,00
60,00
80,00
100,00
120,00
140,00
0 5 10 15 20
T/M
-Ver
häl
tnis
[%]
Kauzyklen
T/M-Verhältnis AS hart
T/M-Verhältnis AS weich
27
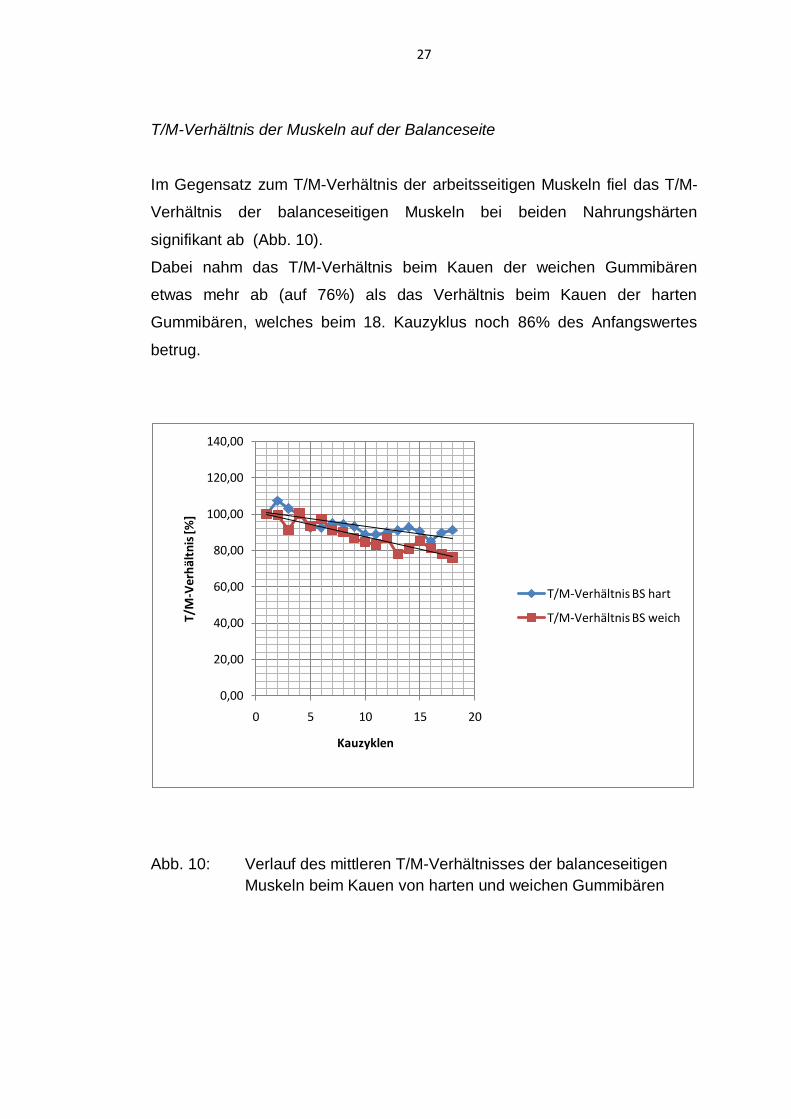
T/M-Verhältnis der Muskeln auf der Balanceseite
Im Gegensatz zum T/M-Verhältnis der arbeitsseitigen Muskeln fiel das T/M-
Verhältnis der balanceseitigen Muskeln bei beiden Nahrungshärten
signifikant ab (Abb. 10).
Dabei nahm das T/M-Verhältnis beim Kauen der weichen Gummibären
etwas mehr ab (auf 76%) als das Verhältnis beim Kauen der harten
Gummibären, welches beim 18. Kauzyklus noch 86% des Anfangswertes
betrug.
Abb. 10: Verlauf des mittleren T/M-Verhältnisses der balanceseitigen
Muskeln beim Kauen von harten und weichen Gummibären
0,00
20,00
40,00
60,00
80,00
100,00
120,00
140,00
0 5 10 15 20
T/M
-Ver
häl
tnis
[%]
Kauzyklen
T/M-Verhältnis BS hart
T/M-Verhältnis BS weich
28
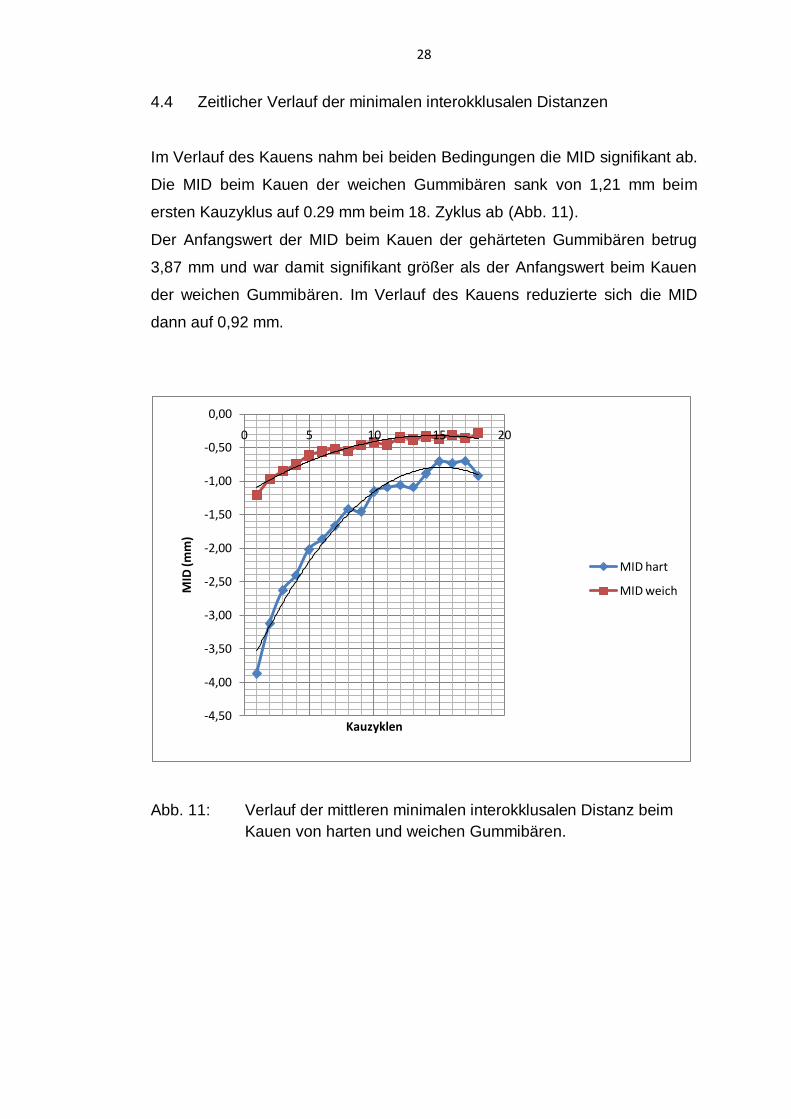
4.4 Zeitlicher Verlauf der minimalen interokklusalen Distanzen
Im Verlauf des Kauens nahm bei beiden Bedingungen die MID signifikant ab.
Die MID beim Kauen der weichen Gummibären sank von 1,21 mm beim
ersten Kauzyklus auf 0.29 mm beim 18. Zyklus ab (Abb. 11).
Der Anfangswert der MID beim Kauen der gehärteten Gummibären betrug
3,87 mm und war damit signifikant größer als der Anfangswert beim Kauen
der weichen Gummibären. Im Verlauf des Kauens reduzierte sich die MID
dann auf 0,92 mm.
Abb. 11: Verlauf der mittleren minimalen interokklusalen Distanz beim
Kauen von harten und weichen Gummibären.
-4,50
-4,00
-3,50
-3,00
-2,50
-2,00
-1,50
-1,00
-0,50
0,00
0 5 10 15 20
MID
(m
m)
Kauzyklen
MID hart
MID weich
29
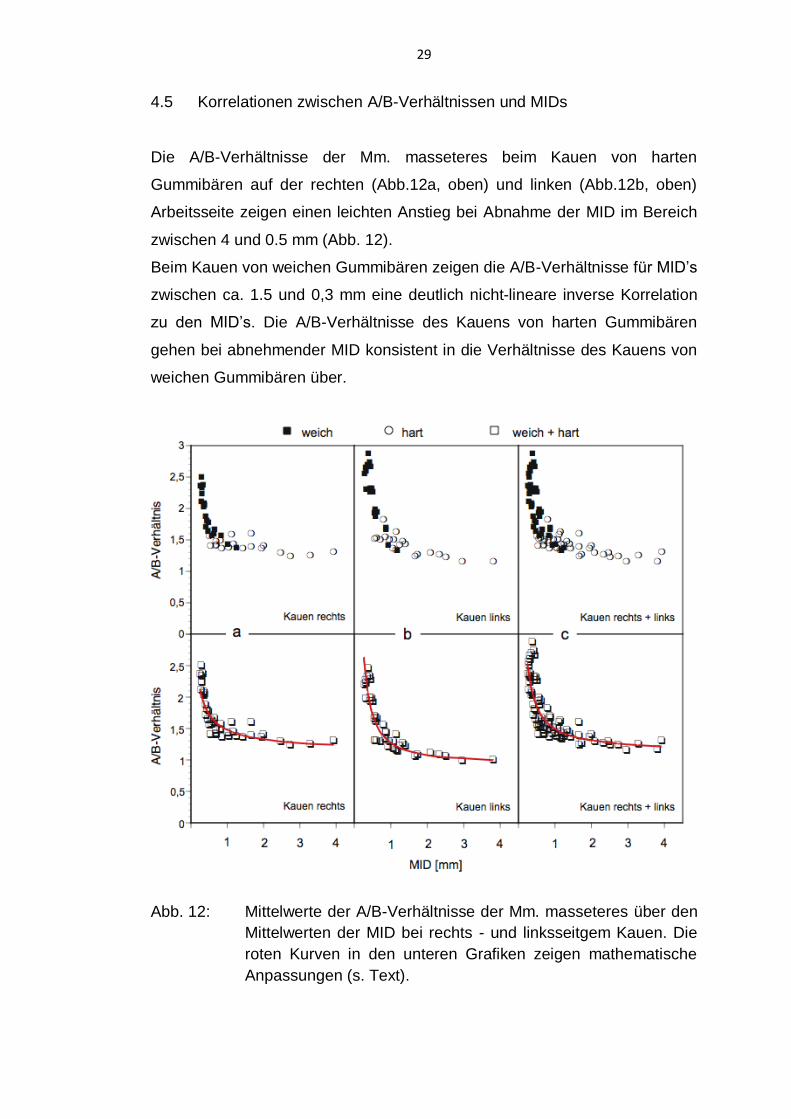
4.5 Korrelationen zwischen A/B-Verhältnissen und MIDs
Die A/B-Verhältnisse der Mm. masseteres beim Kauen von harten
Gummibären auf der rechten (Abb.12a, oben) und linken (Abb.12b, oben)
Arbeitsseite zeigen einen leichten Anstieg bei Abnahme der MID im Bereich
zwischen 4 und 0.5 mm (Abb. 12).
Beim Kauen von weichen Gummibären zeigen die A/B-Verhältnisse für MID’s
zwischen ca. 1.5 und 0,3 mm eine deutlich nicht-lineare inverse Korrelation
zu den MID’s. Die A/B-Verhältnisse des Kauens von harten Gummibären
gehen bei abnehmender MID konsistent in die Verhältnisse des Kauens von
weichen Gummibären über.
Abb. 12: Mittelwerte der A/B-Verhältnisse der Mm. masseteres über den
Mittelwerten der MID bei rechts - und linksseitgem Kauen. Die
roten Kurven in den unteren Grafiken zeigen mathematische
Anpassungen (s. Text).

30
Diese Aussagen gelten auch für die Zusammenlegung der Ergebnisse von
rechtsseitigem und linksseitigem Kauen (Abb.12c, oben).
Die drei unteren Grafiken der Abb.12 zeigen die gleichen Daten, wie die drei
oberen Grafiken, mit dem Unterschied, dass hier die Werte des Kauens von
harten und weichen Gummibären nicht unterschieden sind. An diese
Gesamtdarstellung wurden in Abb.12 unten durch Ausprobieren
mathematische Funktionen angepasst, die den Verlauf der Korrelation
beschreiben sollen. Als sehr gut passend erwies sich hier die allgemeine
Funktion
Gl.1 A/B-Verhältnis = a + eb MIDc
Der Parameter a charakterisiert den Grenzwert, gegen den das A/B-
Verhältnis bei zunehmender MID strebt und wurde mit dem Wert 1.1
angesetzt. Der Parameter eb ist ein Skalierungsfaktor und der Exponent c
bestimmt die Stärke der Krümmung mit der die Kurven bei sehr kleinen
MIDs nach oben biegen.
Für das rechtsseitige Kauen ergab sich A/B = 1.1 + e-0,949 x MID-0,723, für das
linksseitige Kauen ergab sich A/B = 1.1 + e-0,924 x MID-1,26 und für die
Zusammenlegung von rechts - und linksseitigem Kauen A/B = 1.1 + e-0,94 x
MID-0,96. Die Korrelationskoeffizienten (r-Werte) lagen mit 0,89, 0,95 und
0,89 relativ hoch.
Im Mittel über alle Kauzyklen ergab sich bei Zusammenlegung von rechts -
und linksseitigem Kauen für das weiche Kaugut ein A/B-Verhältnis von 2.06 ±
0.42 und für das harte Kaugut ein signifikant kleineres A/B-Verhältnis von
1.42 ± 0.11.
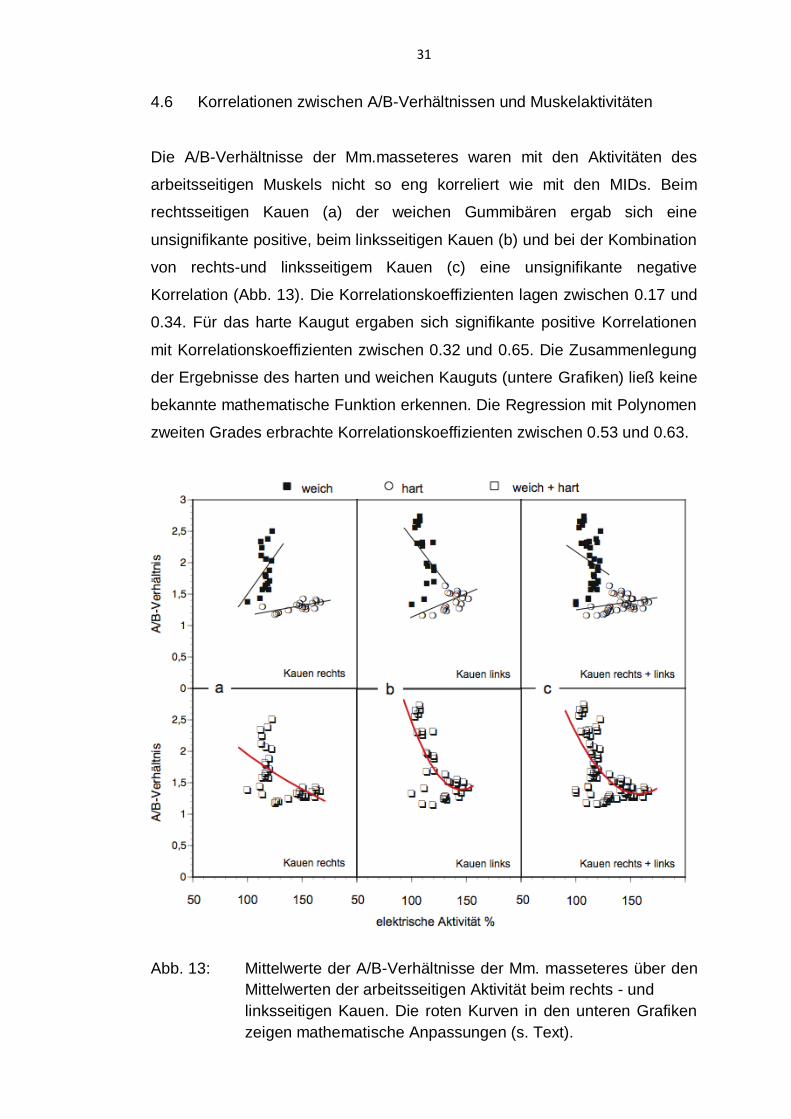
31
4.6 Korrelationen zwischen A/B-Verhältnissen und Muskelaktivitäten
Die A/B-Verhältnisse der Mm.masseteres waren mit den Aktivitäten des
arbeitsseitigen Muskels nicht so eng korreliert wie mit den MIDs. Beim
rechtsseitigen Kauen (a) der weichen Gummibären ergab sich eine
unsignifikante positive, beim linksseitigen Kauen (b) und bei der Kombination
von rechts-und linksseitigem Kauen (c) eine unsignifikante negative
Korrelation (Abb. 13). Die Korrelationskoeffizienten lagen zwischen 0.17 und
0.34. Für das harte Kaugut ergaben sich signifikante positive Korrelationen
mit Korrelationskoeffizienten zwischen 0.32 und 0.65. Die Zusammenlegung
der Ergebnisse des harten und weichen Kauguts (untere Grafiken) ließ keine
bekannte mathematische Funktion erkennen. Die Regression mit Polynomen
zweiten Grades erbrachte Korrelationskoeffizienten zwischen 0.53 und 0.63.
Abb. 13: Mittelwerte der A/B-Verhältnisse der Mm. masseteres über den
Mittelwerten der arbeitsseitigen Aktivität beim rechts - und
linksseitigen Kauen. Die roten Kurven in den unteren Grafiken
zeigen mathematische Anpassungen (s. Text).

32
5 Diskussion
Voraussetzungen
Ziel dieser Arbeit war es, einen Beitrag zur Klärung der Frage zu leisten, ob
beobachtete Korrelationen zwischen Aktivitätsverhältnissen der Kaumuskeln
und der Bißsperrung kausal von letzterer verursacht werden, oder ob dafür
eher die Kaukraft verantwortlich ist.
Anstoß und tieferer Hintergrund für diese Frage ist die Beobachtung, dass
besonders das A/B-Verhältnis der Mm. masseteres stark ansteigt, wenn die
MID im Verlauf der Nahrungszerkleinerung immer enger wird (13,31). Da der
Anstieg des Masseter-A/B-Verhältnisses als neuromuskuläre Strategie zum
Schutz der Zahnreihen vor Überlastung durch balanceseitige
Okklusalkontakte gedeutet wird (30,31), wäre es essentiell zu wissen,
welcher periphere sensorische Input den neuronalen Steuerzentren die
Annäherung des Unterkiefers an den Oberkiefer signalisiert. Offenkundig,
jedoch auch vordergründig, sprechen die Logik und bisherige experimentelle
Befunde dafür, dass die sensorische Information über eine Lageveränderung
des Unterkiefers von lagesensitiven Mechanorezeptoren geliefert wird, wofür
in erster Linie die Muskelspindeln des M. masseter in Frage kommen
(5,7,8,35). Diese Annahme ist jedoch nicht unumstritten, da sich bei der
Abnahme der MID im Verlauf der mastikatorischen Nahrungszerkleinerung
auch die Kaukraft verändern kann. Tatsächlich wurden beim Kauen härterer
Nahrung kleinere A/B-Verhältnisse gefunden als mit weicherer Nahrung. Da
härtere Nahrung mit größerer Kraft gekaut wird als weichere wurde
argumentiert (3,9), dass die neuromuskuläre Steuerung bei größerem
Bißkraftbedarf den balanceseitigen M. masseter stärker aktiviert und somit
eine symmetrischere relative Aktivierung erzeugt. Da aber härtere
Nahrungen größere Bißsperrungen implizieren, könnte sich hinter der
Korrelation zwischen Aktivitätsverhältnissen und Bißsperrung in Wirklichkeit
ein kausaler Einfluss der Bißkraft verbergen. In diesem Fall müsste bei einer
fortschreitenden Nahrungszerkleinerung insbesondere das A/B-Verhältnis
der Mm. masseteres mit der Kaukraft ebensogut korrelliert sein wie mit der
MID.

33
Methodik
Um die Fragestellung direkt zu untersuchen müsste man die Bißkraft
während des Kauens - also die sog. "Kaukraft" - messen und diese mit den
Aktivitätsverhältnissen in Beziehung setzen. Für eine direkte Messung der
Kaukraft müsste diese intraoral zwischen den Zahnreihen erfasst werden.
Dies könnte im Prinzip mit Dynamometern oder mit Aufnehmern, die in
Zähne oder Implantate integriert sind geschehen. Aufgrund der
Mindestbauhöhen von Dynamometern (6,11,17,20,21,33,36,41,42,43) und
dem großen technischen Aufwand zur Anwendung integrierter Sensoren
(24,32,38) verursachen diese Methoden der Kraftmessung zum einen
erhebliche Bißsperrungen und zum anderen würden die physiologischen
Funktionsabläufe bei der Mastikation, die man eigentlich untersuchen will,
stark beeinträchtigt.
Zur Beantwortung der obigen Frage wurde deshalb als Ersatz für die
Kaukraft das EMG des M. masseter verwendet. Dies erscheint aus mehreren
Gründen gerechtfertigt:
Oberflächen-EMGs speziell der Kaumuskeln sind experimentell unkompliziert
und nicht-invasiv ableitbar und beeinträchtigen die natürliche Kaufunktion
nicht. In wissenschaftlichen Untersuchungen haben EMGs daher eine weite
Verbreitung gefunden.
Die Rechtfertigung für die Verwendung von EMGs als Ersatz für die Bißkraft
basiert primär auf zahlreichen Nachweisen stochastischer Beziehungen
zwischen EMG und Bißkraft bei isometrischen Beißvorgängen
(2,4,6,17,20,43). Auch in der vorliegenden Arbeit konnte dies durch die in 3.3
erwähnten, zusätzlichen motorischen Aktionen isometrischen Beißens auf
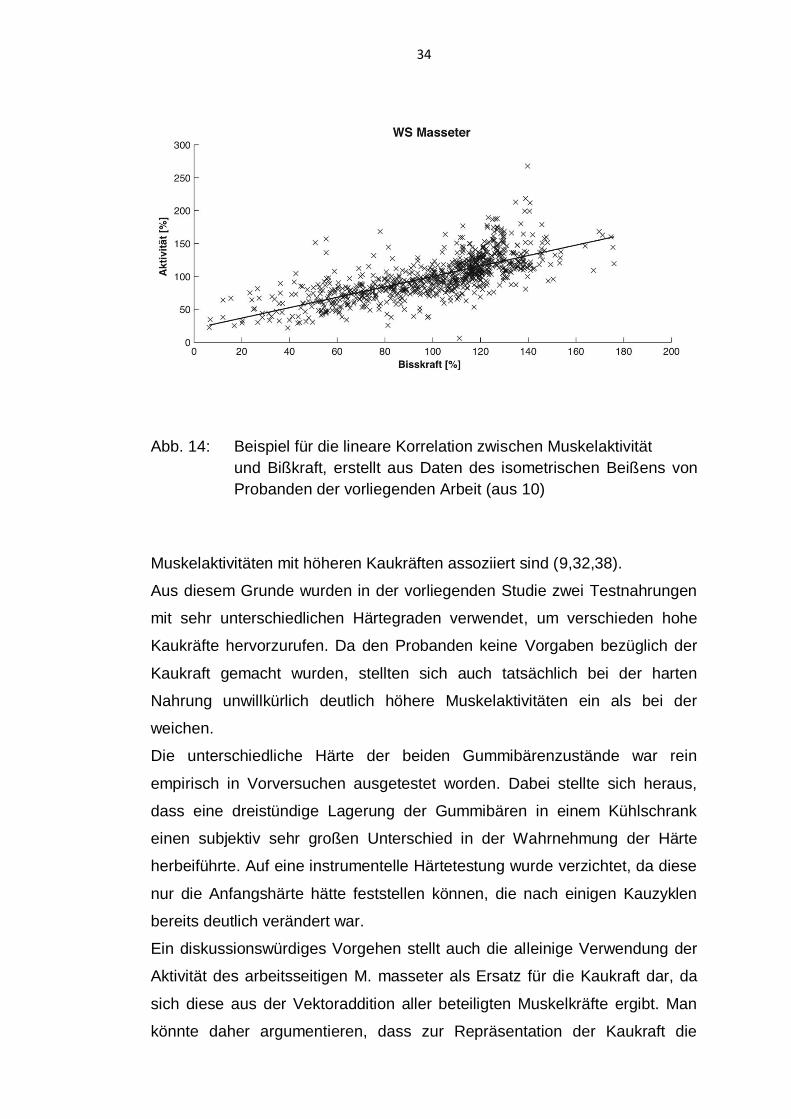
Folienkraftaufnehmer bestätigt werden (10). Abb.14 zeigt hierzu beispielhaft
eine Aktivitäts/Bißkraft Korrelation die an den Probanden der vorliegenden
Arbeit gewonnen wurde.
Aus den gut belegten Aktivitäts/Bißkraft Korrelationen beim isomentrischen
Beißen kann allerdings nicht ohne weiteres auf die Existenz ebensolcher
Korrelationen beim Kauen geschlossen werden und es wurden solche
Korrelationen auch noch nicht explizit nachgewiesen. Es wurde jedoch
gezeigt, dass im Mittel kleinere Muskelaktivitäten mit kleineren - und größere
34
Abb. 14: Beispiel für die lineare Korrelation zwischen Muskelaktivität
und Bißkraft, erstellt aus Daten des isometrischen Beißens von
Probanden der vorliegenden Arbeit (aus 10)
Muskelaktivitäten mit höheren Kaukräften assoziiert sind (9,32,38).
Aus diesem Grunde wurden in der vorliegenden Studie zwei Testnahrungen
mit sehr unterschiedlichen Härtegraden verwendet, um verschieden hohe
Kaukräfte hervorzurufen. Da den Probanden keine Vorgaben bezüglich der
Kaukraft gemacht wurden, stellten sich auch tatsächlich bei der harten
Nahrung unwillkürlich deutlich höhere Muskelaktivitäten ein als bei der
weichen.
Die unterschiedliche Härte der beiden Gummibärenzustände war rein
empirisch in Vorversuchen ausgetestet worden. Dabei stellte sich heraus,
dass eine dreistündige Lagerung der Gummibären in einem Kühlschrank
einen subjektiv sehr großen Unterschied in der Wahrnehmung der Härte
herbeiführte. Auf eine instrumentelle Härtetestung wurde verzichtet, da diese
nur die Anfangshärte hätte feststellen können, die nach einigen Kauzyklen
bereits deutlich verändert war.
Ein diskussionswürdiges Vorgehen stellt auch die alleinige Verwendung der
Aktivität des arbeitsseitigen M. masseter als Ersatz für die Kaukraft dar, da
sich diese aus der Vektoraddition aller beteiligten Muskelkräfte ergibt. Man
könnte daher argumentieren, dass zur Repräsentation der Kaukraft die

35
Aktivitäten aller vier betrachteten Muskeln addiert werden müssten, was in
vorangegangenen Arbeiten (38) schon praktiziert wurde. Dem ist allerdings
entgegen zu halten, dass beim isometrischen Beißen alle vier Muskeln in
gleicher Weise mit der Bißkraft korreliert waren (10), weshalb eine
Summenbildung keine andere Information erbracht hätte, als die Betrachtung
der Aktivität eines einzelnen Muskels. Dies gilt auch für die hier gemessenen
Aktivitäten während des Kauens (Abb. 3 - 6). Im Vorgriff auf die Diskussion
der Ergebnisse kann man festhalten, dass sich die Aktivitäten des
arbeitsseitigen M. masseter und der beiden Mm. temporales bei beiden
Nahrungshärten sehr ähnlich präsentierten, so dass auch ihre Summe keine
andere Information geboten hätte, als die Aktivität eines einzelnen Muskels.
Darüber hinaus liefert die Addition von Aktivitäten verschiedener Muskeln
keine sicheren Ergebnisse, da die Aktivitäten mit den Muskelkräften über
Faktoren korreliert sind, die für jeden Muskel unterschiedlich sein können
(11,37) und die nicht ermittelt werden können, da Muskelkräfte bisher nicht
direkt messbar sind.
Aus diesen Gründen kann die arbeitsseitigen Masseteraktivität als eine zwar
nicht perfekte, aber vertretbare Approximation der Bisskraft betrachtet
werden.
Ergebnisse
Zunächst bestätigen die Ergebnisse des Kauens von weichen Gummibären
die Aussagen einer vorangegangenen Arbeit (31). In den ersten Kauzyklen
stieg die arbeitsseitige Masseteraktivität leicht an um dann im weiteren
Verlauf leicht abzunehmen. Dagegen fiel die balanceseitige Aktivität von
Beginn des Kauens an nahezu linear ab. Dieses Verhalten (Abb.3,4) erklärt
den starken Anstieg des A/B-Verhältnisses (Abb.7) im Verlauf des Kauens.
Der gleichzeitige Rückgang der MID dokumentiert die zunehmende
Kompression des Kauguts. Die asymptotische Annäherung der MID mit
fortschreitendem Kauen an einen Wert von ca. 0.3 mm entspricht auch
früheren Ergebnissen (31) und kann bedeuten, dass auch bei erweichtem
Bolus eine von Null verschiedene interokklusale Distanz beibehalten wird.
Allerdings muss berücksichtigt werden, dass dieser Wert durch das

36
Rauschen des Sirognathographen und die Auswertung der Wegsignale
beeinflusst wird. Zur Ermittlung der MID wurden alle Weg/Zeit-Kurven so
korrigiert, dass der jeweils höchste (positivste bzw. am wenigsten negativste)
Messwert genau auf der Nulllinie lag. Dies wird grundsätzlich bei
Unterkieferbewegungsmessungen durchgeführt, um ein geringes Driften des
Signals über die Nulllinie hinaus auszugleichen. Diese Korrektur hat zur
Folge, dass das Rauschen in den negativen Wegbereich verschoben wird
und sich somit den MID-Werten doppelt so stark überlagert wie es eigentlich
in Wirklichkeit der Fall ist. Durch diesen Effekt wird den MID Messwerten ein
Biss von ca. 0.1 mm überlagert. Für die Ergebnisse spielt dies keine Rolle,
da hierdurch in der Korrelationsdarstellung der Abb.12 die Punktewolken
lediglich um diesen konstanten Wert (von 0.1 mm) zu größeren MIDs hin
verschoben werden. An der Korrelation zwischen A/B-Verhältnis und MID
ändert dies aber nichts. Es ist gegenwärtig nicht bekannt, ob eine MID von
0.3 mm unterschritten werden kann. Um diese Frage zu klären, müssten
Kauexperimente mit weicheren Testnahrungen durchgeführt werden. Die
Korrelation zwischen dem A/B-Verhältnis und der MID bestätigt ebenfalls
Ergebnisse einer vorangegangenen Arbeit (31).
Beim Kauen der harten Gummibären traten signifikant höhere Aktivitäten auf,
die sich auch im Verlauf des Kauens anders verhielten als beim Kauen der
weichen Gummibären. Da den Probanden hinsichtlich der Kaukraft keine
Vorgaben gemacht worden waren, spiegeln die Verläufe der Aktivitäten in
den Abb. 3 bis 6 die unwillkürliche Reaktion der neuromuskulären Steuerung
auf das harte Kaugut wieder. Anders als beim weichen Kaugut stieg hier die
Aktivität auf der AS im Verlauf des Kauens deutlich an, während sie auf der
BS zunächst leicht anstieg und dann eher gleich blieb. Daraus kann man
einerseits folgern, dass der zähere Bolus generell eine wesentlich höhere
Muskelaktivität und damit eine höhere Kaukraft hervorrief. Zum anderen
zeigen die Aktivitätsverläufe auf AS und BS, dass bei dem härteren Kaugut
der Muskel auf der BS deutlich stärker aktiviert ist als bei dem weichen
Kaugut. Analog hierzu waren die Mittelwerte der A/B-Verhältnisse der Mm.
masseteres bei hartem Kaugut signifikant kleiner als bei weichem Kaugut.
Dies alles spricht zunächst für die Vermutung von Hylander und van Eijden

37
(3,9), dass nämlich zur Erzielung einer höheren Kaukraft der balanceseitige
Muskel verstärkt herangezogen wird. Allerdings ist beim Kauen des harten
Bolus auch die MID von Beginn an wesentlich größer und nimmt bei
fortschreitender Nahrungszerkleinerung nicht auf den gleichen geringen Wert
von 0.3 mm ab, wie beim Kauen der weichen Testnahrung.
Eine Entscheidung darüber, ob nun die die Kaukraft oder aber die MID das
Verhalten der A/B-Verhältnisse kausal beeinflusst kann aus dem Vergleich
der Korrelationsdiagramme der Abb.12 und 13 abgeleitet werden.:
In Abb.12 steigen die A/B-Verhältnisse beim Kauen harter Nahrung mit
abnehmender MID stetig an und gehen konsistent in die A/B-Verhältnisse
des Kauens weicher Nahrung über, die ihrerseits mit abnehmender MID stark
ansteigen. Ungeachtet der Unterschiede zwischen den Nahrungstexturen
und der damit verbundenen Unterschiede im Aktivierungsverhalten (Abb.3,4)
zeigt die Abb.12c eine konsistente, hochsignifikante und starke Korrelation
zwischen dem A/B-Verhältnis und der MID. Die unlineare Form der
Korrelation legt den Schluss nahe, dass bei Unterschreiten einer bestimmten
MID von ca. 1 mm, die Empfindlichkeit der motorischen Steuerung der
Muskelaktivierung gegenüber einer weiteren Abnahme der MID drastisch
zunimmt. Mathematisch drückt sich dies im Exponenten c der MID in der
Regressionsgleichung (Gl.1) aus: je größer der Absolutbetrag von c ist, desto
schärfer wird der Knick, an dem die Regressionskurve nach oben (zu
größeren A/B-Verhältnissen hin) abbiegt. Obwohl Exponentialfunktionen
häufig grundlegende Zusammenhänge beschreiben, kann beim jetzigen
Stand des Wissens noch nicht beantwortet werden, welche neuronalen
physiologischen Mechanismen der Korrelation zu Grunde liegen. Die
Regressionsgleichung GL.1 muss daher zunächst lediglich als formales
Modell betrachtet werden, welches den Zusammenhang zwischen MID und
A/B-Verhältnis sehr gut beschreibt.
Anders als zur MID, zeigten die A/B-Verhältnisse zur Muskelaktivität des
arbeitsseitigen M. masseter weniger konsistente und weniger enge
Korrelationen (Abb.13). Für das weiche Kaugut ergab sich beim
rechtsseitigen Kauen eine ansteigende, für das linksseitige Kauen dagegen
eine absteigende Assoziation zwischen A/B-Verhältnis und Muskelaktivität.
Diese Korrelationen waren allerdings nicht signifikant und hatten sehr kleine

38
Korrelationskoeffizienten. Ursache für diese schlechten Korrelationen kann
sein, dass tatsächlich keine Korrelation besteht. Andererseits variierte die
Muskelaktivität beim Zerkleinern der weichen Gummibären nur sehr wenig,
so dass eine Korrelation möglicherweise nicht sichtbar werden konnte. Um
dies abzuklären wäre es nötig, durch geeignete Auswahl des Kauguts eine
größere Streuung der Muskelaktivitäten zu erreichen.
Letzteres ergab sich unwillkürlich beim Kauen der harten Gummibären, wo
sich zwar nicht sehr enge, aber zumindest signifikante Korrelationen
ergaben. Hier zeigte sich, dass mit zunehmender Aktivität und somit mit
zunehmender Kaukraft die A/B-Verhältnisse anstiegen, was im Grunde im
Widerspruch zu den in der Literatur getroffenen Aussagen (3,9) steht.
Dennoch lassen sich die Feststellungen von Hylander und van Eijden
widerspruchsfrei mit den hier erzielten Ergebnissen erklären, wenn man
berücksichtigt, dass im vorliegenden Versuch das härtere Kaugut mit einer
im Durchschnitt wesentlich größeren MID gekaut wurde, als das weiche
Kaugut.

39
6 Schlußfolgerungen
Die vorliegende Arbeit zeigt, dass während der Zerkleinerung eines
bestimmten Kauguts bei unwillkürlichem, habituellem Kauen, das A/B-
Verhältnis der Mm. masseteres umso kräftiger ansteigt, je kleiner die MID in
aufeinanderfolgenden Kauzyklen wird. Die Korrelation zwischen A/B-
Verhältnis und MID ist unabhängig von der Konsistenz des Kauguts und von
der Kaukraft und läßt sich durch eine Potenzfunktion beschreiben. Während
das A/B-Verhältnis eng mit der MID korreliert, ist es unter den gegebenen
experimentellen Bedingungen nicht, oder nur schwach, mit der Kaukraft
assoziiert.
Im Vergleich zu weicher Nahrung wird beim Kauen härterer Nahrung die
Muskelaktivität von der neuromuskulären Steuerung erhöht und das A/B-
Verhältnis verkleinert. Dieser Effekt, der auch in der Literatur beschrieben ist,
bedeutet nicht, dass die Kaukraft das A/B-Verhältnis beeinflusst. Vielmehr
ließ sich im vorliegenden Fall die Verkleinerung des A/B-Verhältnisses auf
die größeren MIDs zurückführen, die bei der härteren Nahrung auftraten. Die
Ergebnisse legen daher den Schluss nahe, dass das A/B-Verhältnis primär
von der MID beeinflusst wird. Eine durchaus beobachtbare Beziehung
zwischen der Kaukraft und dem A/B-Verhältnis stellt eine Scheinkorrelation
dar, die auftritt, wenn unterschiedliche Kaukräfte mit unterschiedlichen MIDs
einhergehen. Um diese Schlussfolgerung zu erhärten, wäre es sinnvoll, das
vorliegende Experiment zu erweitern und zu modifizieren. Dazu wäre es
insbesondere interessant, durch geeignete Kaukraftvorgaben und
Bolushärten sowohl hohe, als auch niedrige Kaukräfte bei sehr kleinen MIDs
zu erzeugen.

40
7. Literaturverzeichnis
1 Baba K, Ai M, Mizutani H, Enosawa S. Influence of experimental
occlusal discrepancy on masticatory muscle activity during clenching.
J Oral Rehabil. 1996; 23: 55-60.
2 Bakke M, Michler L, Möller E. Occlusal control of mandibular
elevator muscles. Scand J Dent Res. 1992; 100(5): 284-291.
3 Blanksma NG, van Eijden TMGJ. Electomyographic heterogeneity in
the human temporalis and masseter muscle during static biting,
open\close excursions, and chewing.
J Dent Res. 1995; 74(6): 1318-1327.
4 Chen L, Pröschel PA, Morneburg TR. Influence of bite force on jaw
muscle activity ratios in subject-controlled unilateral isometric biting.
J Electromyogr Kinesiol. 2010; xxxxx
5 Goodwin GM, Luschei ES. Discharge of spindle afferents from jaw-
closing muscles during chewing in alert monkeys. J Neurophysiol
1975;38:560-571.
6 Hagberg C, Agerberg G, Hagberg M. Regression analysis of
electromyographic activity of masticatory muscles versus bite force.
Scand J Dent Res. 1985; 93:396-402
7 Hidaka O, Morimoto T, Kato T, Masuda Y, Inoue T, Takada K.
Behaviour of jaw muscle spindle afferents during cortically induced
rhythmic jaw closing movements in the anesthetized rabbit.
J Neurophysiol. 1999; 82: 2633-2640.
8 Hugger A., Türp J.C., Kerschbaum Th. Curriculum Orale
Physiologie. Quintessenz Verlags- GmbH 2006
9 Hylander WL, Johnson KR, Crompton AW. Muscle force
recruitment and biomechanical modeling: an analysis of masseter

41
muscle function during mastication in macaca fascicularis. Am J Phys
Anthropol. 1992; 88: 365-387.
10 Igler C. Zum Einfluss der Bisskraft auf
Kaumuskelaktivitätsverhältnisse beim einseitigen isometrischen
Beissen mit okklusionsnahen Zahnabständen, Dissertation, Erlangen,
2010
11 Iwasaki LR, Petsche PE, McCall Jr WD, Marx D, Nickel JC.
Neuromuscular objectives of the human masticatory apparatus during
static biting. Arch Oral Biol. 2003; 48: 767-777.
12 Johnsen SE, Trulsson M. Encoding of amplitude and rate of tooth
loads by human periodontal afferents from premolar and molar teeth.
J Neurophysiol. 2005; 93(4): 1889-1897.
13 Kimoto K, Fushima K, Tamaki K, Toyoda M, Sato S, Uchimura N.
Asymmetry of masticatory muscle activity during the closing phase of
mastication. Crani. 2000; 18(4): 257-263.
14 Korioth TW, Hannam AG. Deformation of the human mandible during
simulated tooth clenching. J Dent Res. 1994; 73(1): 56-66.
15 Kuboki T, Azuma Y, Orsini MG, Takenami Y, Yamashita A. Effects
of sustained unilateral molar clenching on the temporomandibular joint
space. Oral Surgery Oral Medicine Oral Pathologie. 1996; 82(6): 616-
624.
16 Lavigne G, Kim JS, Valiquette C, Lund JP. Evidence that
periodontal pressoreceptors provide positive feedback to jaw closing
muscles during mastication. J Neurophysiol. 1987; 58: 342-358.
17 Lindauer SJ, Gay T, Rendell J (1991) Electromyographic-force
characteristics in the assessment of oral function. J Dent Res.
70:1417-1421

42
18 Lobbezoo F, Sowman PF, Türker KS. Modulation of human
exteroceptive reflexes during simulated mastication. Clin Neurophysiol
2009;120:398-406
19 Lund JP. Mastication and its control by the brain stem. Critical
Reviews in Oral Biology and Medizin. 1990; 2(1): 33-64.
20 Manns A, Miralles R, Palazzi C. EMG, bite force, and elongation of
the Masseter muscle under isometric voluntary contractions and
variations of vertical dimension. J Prosthet Dent. 1979; 42(6): 674-682.
21 Mao J, Osborn JW. Direction of a bite force determines the pattern of
activity in jaw-closing muscles. J Dent Res. 1994; 73(5): 1112-1120.
22 Mohamed SE, Christensen LV, Harrison JD. Tooth contact patterns
and contractile activity of the elevator jaw muscles during mastication
of two different types of food. J Oral Rehabil. 1983; 10: 87-95.
23 Morimoto T, Inoue T, Masuda Y, Nagashima T. Sensory
components facilitating jaw-closing muscle activities in the rabbit.
Exp Brain Res. 1989; 76(2): 424-440.
24 Morneburg TR, Pröschel PA. In vivo forces on implants influenced by
occlusal scheme and food consistency. Int J Prosthodont 2003; 16:
481-486.
25 Okano N, Baba K, Ohyama T. The influence of altered occlusal
guidance on condylar displacement during submaximal clenching.
J Oral Rehabil. 2005; 32: 714-719.
26 Palla S, Gallo LM, Goessi D. Dynamic stereometry of the
temporomandibular joint. Orthod Craniofacial Res. 2003; 6(1): 37-47.
27 Pantanowitz L, Nalogh K. Significance of the juxtaoral organ (of
Chievitz). Head Neck. 2003; 127: 400-405.

43
28 Piancino MG, Bracco P, Vallelonga T, Merlo A, Farina D. Effect of
bolus hardness on the chewing pattern and activation of masticatory
muscles in subjects with normal dental occlusion. J Electromyogr
Kinesiol. 2008; 18(6): 931-937.
29 Proeschel P. An extensive classification of chewing patterns in the
frontal plane. J Craniomandibular Pract 1987;5:55-63
30 Pröschel PA, Jamal T, Morneburg TR. Motor control of jaw muscles
in chewing and in isometric biting with graded narrowing of jaw gape.
J Oral Rehabil. 2008; 35: 722-728.
31 Pröschel PA, Morneburg TR. Indications for jaw gape-related control
of relative muscle activation in sequent chewing strokes.
J Oral Rehabil. doi: 10.1111/j.1365-2842.2009.02036.x.
32 Pröschel PA, Morneburg TR. Task-dependence of activity/bite-force
relations and its impact on estimation of chewing force from EMG.
J Dent Res. 2002; 81(7): 464-468.
33 Pröschel PA, Raum J. Preconditions for estimation of masticatory
forces from dynamic EMG and isometric bite force-activity relations of
elevator muscles. Int J Prosthodont. 2001; 14(6): 563-569.
34 Proske U. What is the role of muscle receptors in proprioception?
Muscle Nerve. 2005; 31: 780-787
35 Proske U, Gandevia SC. The kinaesthetic senses. J Physiol 2009;
587:4139-4146.
36 Schindler HJ, Lenz J, Trüp JC, Schweizerhof K, Rues S. Small
unilateral jaw gap variations: equilibrium changes, co-contractions and
joint forces. J Oral Rehabil. 2009; 36: 710-718.

44
37 Schindler HJ, Rues S, Truep JC, Schweizerhof K, Lenz J. Activity
patterns of the masticatory muscles during feedback-controlled
simulated clenching activities. Eur J Oral Sci. 2005b; 113: 469-478.
38 Schindler HJ, Stengel E, Spiess WEL. Feedback control during
mastication of solid food textures – a clinical-experimental study.
J Prosthet Dent. 1998; 80: 330-336.
39 Schubert D. Zum Einfluss von okklusalen Balancekontakten auf die
relative Aktivierung der Kaumuskeln bei einseitigen Kau - und
Bisskraftapplikationen. Dissertation, Erlangen 2010
40 Slager GE, Otten E, Nagashima T, van Willigen JD. The riddle of
the large loss in bite force after fast jaw-closing movements. J Dent
Res 1998; 77: 1684-1693.
41 Throckmorton GS, Groshan GJ, Boyd SB. Muscle activity patterns
and control of temporomandibular jont loads. J Prosthet Dent. 1990;
63(6): 685-695.
42 Türker KS. Reflex control of human jaw muscles. Crit Rev Oral Biol
Med. 2002; 13(1): 85-104.
43 Van Eijden TM, Brugman P, Weijs WA, Oosting J (1990)
Coactivation of jaw muscles: recruitment order and level as a function
of bite force direction and magnitude. J Biomech. 1990; 23:475-485
44 Vinyard CJ, Wall CE, Williams SH, Johnson KR, Hylander WL.
masseter electromyography during chewing in ring-tailed lemurs
(lemur catta). Am J Phys Antropol 2006; 130:85-95.

45
8. Abkürzungsverzeichnis
AS Arbeitsseite (arbeitsseitig). Die Arbeitsseite ist diejenige
Seite auf der gekaut, auf ein Objekt gebissen oder
zu der der Unterkiefer, bei zahngeführten
Gleitbewegungen, hin bewegt wird.
BS Balanceseite (balanceseitig). Sie liegt der Arbeitsseite
gegenüber
A/B- Verhältnis Verhältnis der arbeitsseitigen zur balanceseitigen
Muskelaktivität
MID „Minimale interokklusale Distanz“

46
9. Danksagung
Herrn Prof. Dr. M. Wichmann möchte ich vielmals dafür danken, dass ich
diese Arbeit an der Zahnklinik 2 – Zahnärztliche Prothetik durchführen
konnte!
Sehr großen Dank gilt Herrn apl. Prof. Dr. P. Pröschel, der während des
gesamten Verlaufs der Promotion stets mit Rat und Tat zur Verfügung stand
und das Projekt hervorragend betreute.
Weiterhin bedanke ich mich bei Herrn Michael Igler, der mir mit Hilfe eines
neuen Computerprogramms die Auswertung der Daten deutlich erleichterte
und beschleunigte.
Zu guter Letzt möchte ich mich noch bei meinem Freund, Herrn Dr. med.
dent. Thomas Ebert bedanken, der mich tatkräftig mit seinem
zahnmedizinischen Wissen unterstützte.





























